Biodiversity and Conservation 4, 382-394 (1995) Experimental release of an Iberian lynx (Lynx pardinus) ALEJANDRO RODRIGUEZ*, LUIS BARRIOS and MIGUEL DELIBES Estacirn Biolrgica de Dohana, Conse]o Superior de lnvestigaciones CientiJicas, Avda. Maria Luisa s/n, 41013 SeviUa, Spain Received 17 August 1994; revised and accepted 20 October 1994 Reintroduction to the wild of threatened species has become a modern additional justification for captive propagation. This conservation procedure is costly, and both economic resources and the absence of optimal conditions in the field limit the IUCN recommendations for reintroduction to a small proportion of potential candidate species. Furthermore reintroduction attempts often fail. In carnivores, reintroduction failure is attributed to unsuitable adaptation in the field by captive-reared animals, due to their lack of hunting skills, their tendency to leave the target area, their inadequate interaction with conspecifics or their excessive confidence in humans. This list of causes is based on very few studies of carnivore adaptation after reintroduction. In very rare and endangered species, monitoring individual case-histories is the only way to evaluate reintroduction success. We report a successful experimental release of an Iberian lynx (Lynx pardinus) which grew up in captivity. Careful feeding-training and avoidance of human contact during the captive phase, as well as good habitat quality and correct interaction with other wild lynx in the release site, seem to account for the observed success. Permanence of the lynx within the release area might be related to the availability of territory vacancies in the receiving population. Our results allow some optimism for future reintroductions of this endangered species in areas where it has become extinct recently. Keywords: experimental release: pre-release training; adaptation to the wild of captive animals: Lynx pardinus. Introduction Reintroduction and restocking, as defined by the IUCN (1987), are important tools for conservation, either for reinstalling extinct populations or for reinforcing weak ones. Careful reintroduction projects are costly (Kleiman et al., 1991) and, beside the availability of economic resources, their success depends on a set of biological (status and knowledge of the species' biology), environmental and bio-political requirements (Kleiman et al., 1994). These preconditions are rarely satisfied, leading to a low rate of recommendation for reintroduction programmes (4.5% of the 418 threatened species covered by the IUCN/Species Survival Commission Action Plans; Stuart, 1991). Although many reintroductions have been made success has been achieved only in a few cases. For example, Beck and colleagues (1994) document 145 reintroduction projects, only for conservation purposes, where captive-born animals (mainly birds and mammals) have been involved, but the success rate was only the 11%, success being defined as the *To whom correspondence should be addressed. 0960-3115 © 1995 Chapman & Hall

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biodiversity and Conservation 4, 382-394 (1995)
Experimental release of an Iberian lynx (Lynx pardinus) A L E J A N D R O R O D R I G U E Z * , LUIS B A R R I O S and M I G U E L D E L I B E S Estacirn Biolrgica de Dohana, Conse]o Superior de lnvestigaciones CientiJicas, Avda. Maria Luisa s/n, 41013 SeviUa, Spain
Received 17 August 1994; revised and accepted 20 October 1994
Reintroduction to the wild of threatened species has become a modern additional justification for captive propagation. This conservation procedure is costly, and both economic resources and the absence of optimal conditions in the field limit the IUCN recommendations for reintroduction to a small proportion of potential candidate species. Furthermore reintroduction attempts often fail. In carnivores, reintroduction failure is attributed to unsuitable adaptation in the field by captive-reared animals, due to their lack of hunting skills, their tendency to leave the target area, their inadequate interaction with conspecifics or their excessive confidence in humans. This list of causes is based on very few studies of carnivore adaptation after reintroduction. In very rare and endangered species, monitoring individual case-histories is the only way to evaluate reintroduction success. We report a successful experimental release of an Iberian lynx (Lynx pardinus) which grew up in captivity. Careful feeding-training and avoidance of human contact during the captive phase, as well as good habitat quality and correct interaction with other wild lynx in the release site, seem to account for the observed success. Permanence of the lynx within the release area might be related to the availability of territory vacancies in the receiving population. Our results allow some optimism for future reintroductions of this endangered species in areas where it has become extinct recently.
Keywords: experimental release: pre-release training; adaptation to the wild of captive animals: Lynx pardinus.
Introduction
Reintroduction and restocking, as defined by the IUCN (1987), are important tools for conservation, either for reinstalling extinct populations or for reinforcing weak ones. Careful reintroduction projects are costly (Kleiman et al., 1991) and, beside the availability of economic resources, their success depends on a set of biological (status and knowledge of the species' biology), environmental and bio-political requirements (Kleiman et al., 1994). These preconditions are rarely satisfied, leading to a low rate of recommendat ion for reintroduction programmes (4.5% of the 418 threatened species covered by the IUCN/Species Survival Commission Action Plans; Stuart, 1991). Although many reintroductions have been made success has been achieved only in a few cases. For example, Beck and colleagues (1994) document 145 reintroduction projects, only for conservation purposes, where captive-born animals (mainly birds and mammals) have been involved, but the success rate was only the 11%, success being defined as the
*To whom correspondence should be addressed.
0960-3115 © 1995 Chapman & Hall

Experimental release of an Iberian lynx 383
maintenance of a self-sustainable population. Reintroduction has usually been undertaken in extreme cases of species closeness to extinction (e.g. the Arabian oryx Oryx leucoryx, Stanley Price, 1989; and the golden lion tamarin, Leontopithecus rosalia, Kleiman et al., 1986), or when species have been extirpated from most of their original ranges (e.g. the peregrine falcon Falco peregrinus in the United States or the European lynx Lynx lynx in western and central Europe; Campbell, 1980; Breitenmoser and Breitenmoser-Wiirsten, 1990). Wild populations of such threatened species are so small that extracting individuals for translocation is often inadvisable (Stanley Price, 1991). This lack of a suitable wild source of animals is an especially common problem in carnivores (Yalden, 1993). Then, attention is drawn to animals born in captivity: along with their genetic reservoir function, it is considered that the final objective of captive propagation is species reintroduction to the wild (Seal, 1986; Foose, 1989; Chivers, 1991; Wilson and Stanley Price, 1994).
The deficient adaptation to wild conditions is considered as one of the main reasons of the reintroduction failure (Foose, 1987; Stanley Price, 1989; Kleiman et al., 1994). Among carnivores, unsuitable adaptation in the field by captive individuals is due to five main reasons (see Yalden, 1993). First, they are unable to hunt their natural prey, probably because of shortcomings in the learning of complex behaviour when captive (Kleiman, 1980; Ginsberg, 1994); one reported consequence is a switch to hunting domestic prey, increasing mortality caused by conflict with humans (Breitenmoser and Hailer, 1987). Second, they do not always establish a population in the target area. Some animals may move away from the release site, as occurs with translocated wild animals both in suitable (Pettifer, 1981; Slough, 1989) and unsuitable (McArthur, 1981) release habitats; a direct management implication is an increase in expected mortality if wandering animals go beyond reserve boundaries. Third, reintroduced individuals may home, as translocated wild animals can do (e.g. Miller and Ballard, 1982; Fritts et al., 1984). Fourth, they do not interact properly with conspeciflcs. For example, Mills (1991) observed that reintroduced cheetah (Acinonyx jubatus) males fought with resident cheetahs and then left the area. Severe injuries were also reported by Pettifer (1981) for a reintroduced cheetah after fighting with residents. Fifth, they do not fear humans, so released animals are more vulnerable to hunting or road deaths than their wild conspecifics; as a consequence, for example, captive red wolves (Canis rufus) suitable for reintroduction are separated as much as possible from humans and are taught to be afraid of them (Moore and Smith, 1991).
As Rahbek (1993) states, there is a great gap between the theory and practice of reintroduction of endangered species. Specific reports on mammal adaptation monitored after reintroduction are scarce (Kleiman, 1989; Lindburg, 1992; notable exceptions are reported by Kleiman et aL, 1986, and Stanley Price, 1989). However, monitoring released animals seems to be the only way to evaluate the factors affecting reintroduction success or failure (Stanley Price, 1989; May, 1991). In very rare and threatened species, individual case histories should be of a great value in evaluating the effect of such factors (Lindburg, 1992).
In a recent study, Rodriguez and Delibes (1992) show that the world population of the Iberian lynx (Lynx pardinus) is only about 1100 adult individuals, distributed among nine isolated populations. The Iberian lynx has been considered as the most threatened carnivore in Europe (Mallinson, 1978), and one of the most rare cats of the world (IUCN, in preparation). Here we report the results of an experimental release of a wounded young lynx, which grew up and recovered in captivity.

384 Rodriguez et al.
The ease history
On 6 July 1991, a wounded male kitten Iberian lynx (2.0 kg) was found on a road near Cardefia, a locality within the Eastern Sierra Morena (southern Spain) nucleus, which is considered currently as the most healthy population in the range of the Iberian lynx (Rodriguez and Delibes, 1992). The lynx was anaesthetized with 0.5mg atropine and 50 mg ketamine for veterinary examination. Superficial wounds were found on the right foreleg and head, and radiographs showed a fracture of the proximal epiphysis of the left femur. Injuries were cleaned and closed with absorbable sutures, and the fracture was reduced under radioscopy. Before recovery from anaesthesia, a serum-diluted mixture of antibiotics (40mg gentamicin and 50mg netilmicin) and a vitamin complex were administered. On the following days the same treatment was applied with the food.
RECOVERY
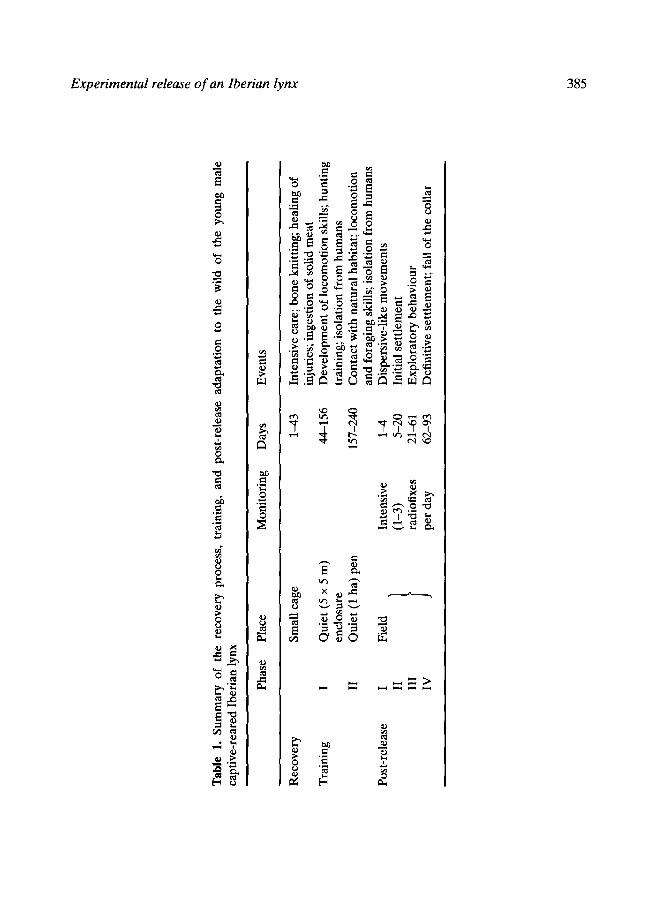
In order to assure correct knitting of the bones, the lynx was put in a small cage, with padded walls, which reduced its movements, especially jumping. The cage was placed in a dark and quiet room, and the lynx was visited once a day for feeding and medication. In the early days the lynx was only able to eat soft food (triturated meat), but small pieces of meat were gradually accepted. This period lasted 43 days, sufficient time for the femur to knit completely (Table 1).
TRAINING: FIRST PHASE
The lynx was moved to a 5 × 5 m outdoor enclosure, provided with trunks and ledges at different heights, in a quiet environment. The extremity affected by the fracture seemed fairly weak and the animal clearly limped, as the other wounds healed up. Food consisted of dead prey treated with vitamins and aminoacids. Food was set progressively at places of difficult access to encourage exercise. Fifteen days after its arrival in the enclosure, the first live prey (a rabbit) was provided. The lynx took 4 days to chase the rabbit; thereafter it was able to chase living prey regularly. It also frequented several ledges, though it preferred one at a height of 2.5 m. About 45 days after the beginning of exercises, limping was nearly imperceptible.
TRAINING: SECOND PHASE
After 112 days in the small enclosure (Table 1) the animal was weighed (4.9kg) and provided with a radiocollar. Then it was moved (about 200 km) to an enclosure (1 ha) with natural vegetation where wild rabbit density was first increased by releasing 100 individuals and supplying vegetables. Thus, the lynx could hide within dense vegetation and it was forced to search and eat wild prey under seminatural conditions. Its physical condition was monitored each few days by observation, and both rabbit remains and analysis of its faeces indicated regular successful predation. The purposes of this phase were to exercise the animal's limbs and to improve its foraging behaviour. Human contact and other disturbance were reduced to a minimum. This period lasted 83 days (Table 1).
Tab
le 1
. S
umm
ary
of t
he r
ecov
ery
proc
ess,
tra
inin
g, a
nd p
ost-
rele
ase
adap
tati
on t
o th
e w
ild
of t
he y
oung
mal
e ca
ptiv
e-re
ared
Ibe
rian
lyn
x
t~
Phas
e Pl
ace
Mon
itor
ing
Day
s E
vent
s
Rec
over
y
Tra
inin
g
Post
-rel
ease
Smal
l cag
e 1-
43
I Q
uiet
(5
x 5
m)
44-1
56
encl
osur
e II
Q
uiet
(1
ha)
pen
157-
240
I Fi
eld
Inte
nsiv
e 1-
4 II
]
(1-3
) 5-
20
III
I ra
diof
ixes
21
-61
IV
per
day
62-9
3
Inte
nsiv
e ca
re;
bone
kni
ttin
g; h
eali
ng o
f in
juri
es; i
nges
tion
of
soli
d m
eat
Dev
elop
men
t of
loco
mot
ion
skill
s; h
unti
ng
trai
ning
; iso
lati
on f
rom
hum
ans
Con
tact
wit
h na
tura
l ha
bita
t; l
ocom
otio
n an
d fo
ragi
ng s
kills
; is
olat
ion
from
hum
ans
Dis
pers
ive-
like
mov
emen
ts
Init
ial s
ettl
emen
t E
xplo
rato
ry b
ehav
iour
D
efin
itiv
e se
ttle
men
t; f
all o
f th
e co
llar
0o

386 Rodriguez et al.
RELEASE
Movements and range On 2 March 1992, the lynx (then estimated to be about 1 year old) was recaptured, weighed (6.0 kg) and provided with a new radiocollar, furnished with an activity sensor. A portion of the collar was made of light rubber, whose resistance decreased with time, so that the collar should fall when the animal's neck reached the size of the collar perimeter or before. Weight was within the normal values described for juveniles (individuals aged between 0.5 and 1.0 year; Beltr~in and Delibes, 1993). The lynx was released in a pine stand 8.7 km east to the point where it was runover 9 months before. Within such habitat, dense shrub cover is available only locally. However, at a greater scale the main habitat types are scrubland (Quercus, Cistus, Rosmarinus, Phillyrea) and grassland with scattered trees (Quercus rotundifolia). The border between the two habitats is sharp.
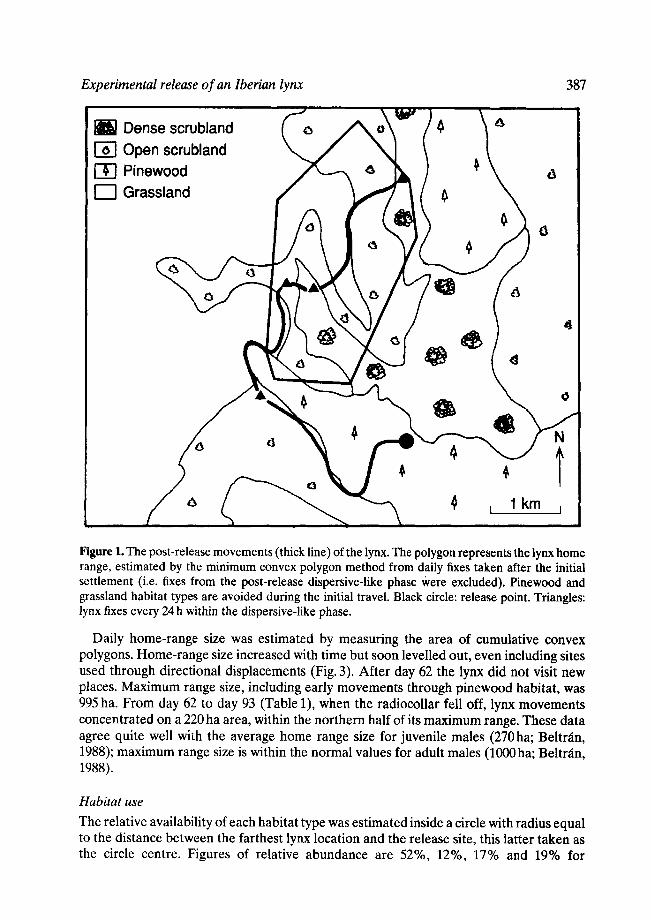
The lynx was followed intensively during the first 3 days (Table 1). From the release site it moved to the west through the pinewood as far as a dense patch of riparian vegetation. It remained there for the rest of the daylight period but the sensor indicated continuous activity. This site was only 300 m from the release point. Early in the night it moved along the stream, still through the pinewood, to the border between pines and scrubland (Fig. 1 ). It followed this border to a dense scrub patch where it was found in the morning of the second day. The lynx remained there during the daytime; and the next night it contacted the grassland habitat but avoided entering it. It skirted northwards along the edge between grassland and scrubland. In the morning of the third day the lynx was near this edge, inside the scrubland, where it showed some activity during the day and the following night. This patch was the first area with high rabbit density it encountered, and probably it stopped for foraging. Next day, still following the border between pastures and scrubland, it reached the area where it finally established itself. This site is about 4 km north of the release site (Fig. 1). The lynx travelled a minimum distance of 8.1 km and took 4 days to cover the distance. During this period it was active 64% of diurnal and 50% of nocturnal monitoring time. Activity level was about twice that reported for juvenile male lynx in Dofiana (SW Spain; Belmin, 1988). Thus, displacement was fairly slow and activity time relatively high, suggesting, in addition to prey searching, a rather detailed habitat exploration.
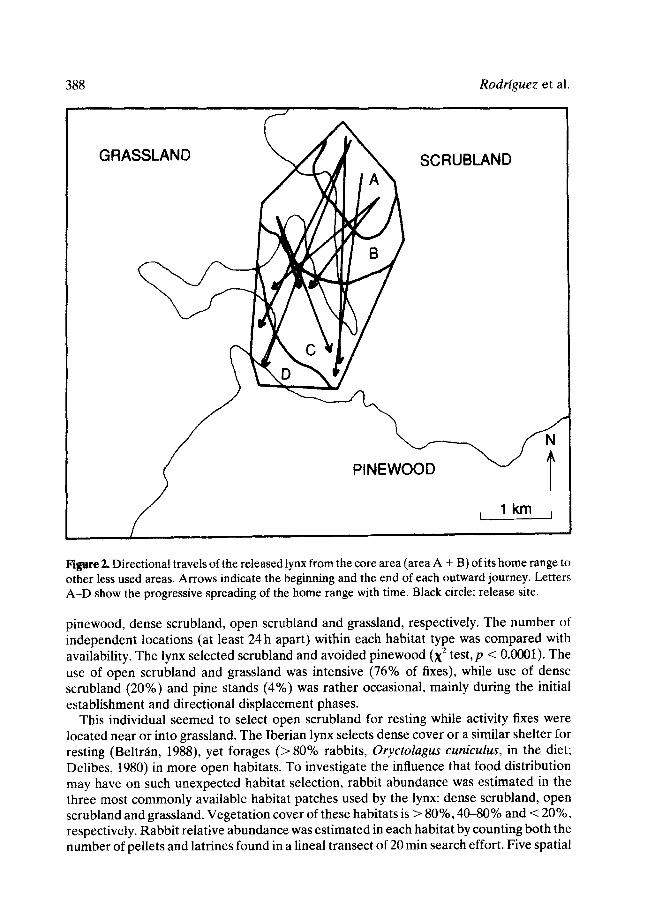
From the fourth day on after release, lynx location was determined at least once a day. From day 4 to day 20 (Table 1), fixes concentrated within a small area of scrubland (about 80ha), near the grassland border (Fig. 2, area A). From day 21 to day 61 the pattern of movements consisted of nine long and fast directional travels whose start and end were in the A + B area (Fig. 2). After the first directional displacement the lynx enlarged his home range to area B, i.e. it also used this area for daily movements and for diurnal resting. Habitat characteristics of area B are similar to those of area A. The A + B area (Fig. 2) is defined as the core area of the lynx home range because (1) it contains 79% of all fixes, and (2) daily movements within it are rather short and slow as compared with those detected in other parts of the home range.
The inter-journey periods within the core area were rather short. Distance, direction, target area, dates and duration of these movements are shown in Table 2. Seven directional travels reached a narrow valley covered with dense scrubland (the area C, Fig. 2), and the other two were directed to the limit with the pine stand where the lynx was released, namely the area D (Fig. 2). All directional travels were mainly directed to the south and rarely reached the southern pinewood habitat strip.
Experimental release of an Iberian lynx 387
Dense scrubland rE] open scrubland i - ~ Pinewood I I Grassland
0
4
t 1 km i
O
Figure 1. The post-release movements (thick line) of the lynx. The polygon represents the lynx home range, estimated by the minimum convex polygon method from daily fixes taken after the initial settlement (i.e. fixes from the post-release dispersive-like phase ~vere excluded). Pinewood and grassland habitat types are avoided during the initial travel. Black circle: release point. Triangles: lynx fixes every 24 h within the dispersive-like phase.
Daily home-range size was estimated by measuring the area of cumulative convex polygons. Home-range size increased with time but soon levelled out, even including sites used through directional displacements (Fig. 3). After day 62 the lynx did not visit new places. Maximum range size, including early movements through pinewood habitat, was 995 ha. From day 62 to day 93 (Table 1), when the radiocollar fell off, lynx movements concentrated on a 220 ha area, within the northern half of its maximum range. These data agree quite well with the average home range size for juvenile males (270ha; Beltr~in, 1988); maximum range size is within the normal values for adult males (1000 ha; Beltr~in, 1988).
Habitat use
The relative availability of each habitat type was estimated inside a circle with radius equal to the distance between the farthest lynx location and the release site, this latter taken as the circle centre. Figures of relative abundance are 52%, 12%, 17% and 19% for
388 Rodriguez et al.
GRASSLAND SCRUBLAND
PINEWOOD
1 km
Figure 2, Directional travels of the released lynx from the core area (area A + B) of its home range to other less used areas. Arrows indicate the beginning and the end of each outward journey, letters A-D show the progressive spreading of the home range with time. Black circle: release site.
pinewood, dense scrubland, open scrubland and grassland, respectively. The number of independent locations (at least 24 h apart) within each habitat type was compared with availability. The lynx selected scrubland and avoided pinewood (X 2 test, p < 0.0001). The use of open scrubland and grassland was intensive (76% of fixes), while use of dense scrubland (20%) and pine stands (4%) was rather occasional, mainly during the initial establishment and directional displacement phases.
This individual seemed to select open scrubland for resting while activity fixes were located near or into grassland. The Iberian lynx selects dense cover or a similar shelter for resting (Beltrfin, 1988), yet forages (> 80% rabbits, Oryctolagus cuniculus, in the diet: Delibes, 1980) in more open habitats. To investigate the influence that food distribution may have on such unexpected habitat selection, rabbit abundance was estimated in the three most commonly available habitat patches used by the lynx: dense scrubland, open scrubland and grassland. Vegetation cover of these habitats is > 80%, 40-80% and < 20%, respectively. Rabbit relative abundance was estimated in each habitat by counting both the number of pellets and latrines found in a lineal transect of 20 min search effort. Five spatial

Tab
le 2
. Dat
e of
sta
rt,
dura
tion
(da
ys u
ntil
retu
rn t
o th
e ho
me
rang
e co
re a
rea)
, m
inim
um d
ista
nce
trav
elle
d (k
m),
and
are
a vi
site
d (s
ee F
ig. 2
) du
ring
the
nin
e di
rect
iona
l jou
rney
s m
ade
by th
e re
leas
ed l
ynx
Dir
ecti
onal
tra
vel
1 2
3 4
5 6
7 8
9
Dat
e of
sta
rt
24 M
arch
2 A
pril
5
Apr
il
9 A
pril
D
irec
tion
S
SE
S
E
SW
S
Dur
atio
n (d
ays)
2
2 4
2 D
ista
nce
7.6
5.5
6.2
4.0
Vis
ited
area
D
C
C
C
13
Ap
fill
5A
p~
1
7A
pri
12
7A
pri
l 1
May
S
W
SW
S
SW
S
1 1
1~
1
~5
3.
8 6.
8 6.
2 6.
2 7.
6 C
D
C
C
C
OO

390 Rodriguez et al.
h a
600
500
400
300
200
100
0
directional travels included
I
core area
i i i I
10 20 30 40 50 60 70 80 90
days after release
Figure 3. The size of the lynx home range against the number of days after release. The home range core area is defined as the area A + B (see Fig. 2).
random replicates were made within each habitat. Temporal replicates were considered unnecessary because of the short lynx monitoring period (spring season) and the high persistence of rabbit pellets (Simonetti, 1989), that reduces variability of pellet number over time. Pellet and latrine abundance did not differ between open scrubland and grassland. However, each of these habitats contained more latrines (X 2 test, p < 0.02) and more pellets (p < 0.00t) than dense scrubland habitat. Moreover, rabbits are rare in pine woodland. So, the lynx seemed to select the best food conditions, once shelter requirements are satisfied.
Discussion
The five problems pointed out for general carnivore reintroductions seemed to be overcome in the case presented here. The survival of this released lynx is the best evidence of its ability to hunt. We assume that it preyed mainly upon rabbits, because of their abundance and the relative scarcity of suitable alternative prey. We also think that the training period with live rabbits may determine the subsequent lynx hunting success (e.g. through formation of an appropriate prey search image; Dawkins, 1971 ). This result may question, in some instances, the rules that, in some countries, prohibit the improvement of hunting skills of captive animals by giving them live prey (Yalden, 1993; Shepherdson, 1994).
No attempt at homing behaviour was detected. Furthermore, there was a rather high degree of fidelity to the reintroduction area, as a home range was established within 10 km of the release point. When released the estimated age of the lynx was about 1 year. At this age most lynx are subadult (see Fritts and Sealander, 1978). So, one might expect a typical transient pattern of movements, with irregular displacements and less preferred habitat use (Ferreras, 1994). Conversely, initial dispersive-like movements were shorter than those reported in previous studies (up to 30 km; Beltrfin, 1988: Ferreras, 1994) and time to settlement was also shorter (4 days vs several months for males in the Dofiana area:

Experimental release o fan Iberian lynx 391
Beltr~in, 1988; Ferreras, 1994). Resource quality within its home range can be considered excellent (rather than poor, as predicted for transient individuals; Rolley, 1987) according to the standards of food and cover reported for the species (Delibes, 1989; Rodrfguez and Delibes, 1990; Palomares et al., 1991).
A strong intrasexual competition for the highest quality habitats seems to exist (Delibes, 1989). Because we assume that the density of lynx within the release area is high (Rodriguez and Delibes, 1992), such fast settlement can be viewed as the filling of a resident male vacancy, as no study reports tolerance of resident males to dispersal males (Beltr~in, 1988). This hypothesis is supported by the observed small home-range size (about half of the size reported for adult males; Beltr~in, 1988), in agreement with predicted values for relatively derlse populations (Rolley, 1987). Therefore, settlement of reintroduced lynx may depend on the availability of vacancies near the release area. The absence of exploratory travels northwards from the lynx range may also correspond with the presence there of a resident mine range. There were no signs of abnormal interactions among the released individual and other resident lynx.
Although observed habitat selection agrees quite well with lynx preferences as previously defined, because space use of resident population is unknown, we cannot distinguish between two alternative mechanisms of range selection, one based on habitat preferences, the other determined by avoidance of resident-occupied space. However, it is unlikely that resident individuals defend the worst habitat type (Delibes, 1989), i.e. pine stands. So, in this case, habitat preference-mediated movements may guide an initial search phase, while further settlement may depend on distribution of resident conspecifics of the same sex. Directional travels can be seen as exploratory behaviour to gain information about neighbouring space occupancy.
Two farms are located in the northwestern border of the lynx home range. No attack on poultry was detected, and human presence seems to be avoided, as the western limit of its home range runs parallel (at several hundred metres) to a line of isolated farms.
It may be worthwhile to note, as illustrated in this paper, that the study of the establishment process of reintroduced animals can be very useful in understanding the genesis of home range after dispersal in carnivores.
A reintroduction can be considered successful if the population reinforced or created resists extinction (Beck et aL, 1994). At the individual level, this implies reproduction of released animals. From this point of view we cannot yet state that the present attempt is successful because lynx contact was lost before mating was detected. However, lynx establishment in a stable range within a dense population suggest that future mating opportunities are likely to occur.
The relative success of this reintroduction cannot be wholly attributed to the chance of finding a vacant territory. The procedure has met a number of those criteria that Griffith and colleagues (1989) associate with successful reintroductions: suitability and protection of habitat (the area is within a natural park), as well as care and development of pre-release physical and behavioural skills of the released animal (Box, 1991). Our results suggest that recovered young wild lynx and perhaps, those born in captivity, may be good candidates for future reintroduction, in order to extend the species range to areas where extinction has occurred recently (Rodriguez and Delibes, 1990), after adequate habitat management. New case studies are needed to evaluate the efficiency of reintroduction techniques for this species.

392 Rodriguez et al.
Acknowledgements
We are grateful to R. Laffitte, J. Aldama, and the staff of the Direcci6n Provincial de la Agencia de Medio Ambiente de C6rdoba for care and veterinary assistance of the lynx. We thank also the wardens of the Parque Natural de Cardefia y Montoro for their help with radio-tracking. This research was supported by the Consejo Superior de Investigaciones Cientificas (CSIC) and the Instituto Nacional para la Conservaci6n de la Naturaleza. Personal support to AR and LB was provided by grants from the Junta de Andalucia and CSIC, respectively. D.W. Yalden and an anonymous referee kindly made useful comments on the manuscript.
References
Beck, B.B., Rapaport, L.G., Stanley Price, M.R. and Wilson, A.C. (1994) Reintroduction of captive-born animals. In Creative Conservation: Interactive management of wild and captive animals (P.J.S. Olney, G.M. Mace and A.T.C. Feistner, eds) pp. 265--86. London: Chapman and Hall.
Beltr~tn, J.F. (1988) Ecologia y Conducta Espacio-temporal del Lince Ib6rico (Lynx pardina T.) en el Parque National de Dofiana. Ph.D. Thesis. Universidad de Sevilla.
Beltr~in, J.F. and Delibes, M. (1993) Physical characteristics of Iberian lynxes (Lynx pardinus) from Dofiana, southwestern Spain. J. Mature. 74, 852--62.
Box, H.O. (1991) Training for life after release: simian primates as examples. Syrup. zool. Soc. Lond. 62, 111-23.
Breitenmoser, U. and Breitenmoser-Wttrsten, C. (1990) Status, Conservation Needs and Reintroduction of the Lynx (Lynx lynx) in Europe. Strasbourg: Council of Europe.
Breitenmoser, U. and Hailer, H. (1987) La r6introduction du lynx (Lynx lynx L. 1758): une appr6ciation apr~s 15 ans d'experience en Suisse. Ciconia 11, 119-30.
Campbell, S. (1980) Is reintroduction a realistic goal? In Conservation Biology. An evolutionary- ecological perspective (M.E. Soul~ and B.A. Wilcox, eds) pp. 263-9. Sunderland: Sinauer.
Chivers, D,J. (1991) Guidelines for re-introductions: procedures and problems. Syrup. zool. Soc. Lond. 62, 89-99.
Dawkins, M. (1971) Perceptual changes in chicks: another look at the 'search image' concept, Anita. Behav. 19, 566-74.
Delibes, M, (1980) Feeding ecology of the Spanish lynx in the Coto Dofiana. Acta Theriol. 25, 309-24. Delibes, M. (1989) Factors regulating a natural population of Iberian lynxes. In Reintroduction of
Predators in Protected Areas pp. 96--9. Assessorato Pianificazione Territoriale Settore Parchi Naturali, Regione Piemonte,Torino.
Ferreras, P. (1994) Patrones de Dispersi6n del Lince Ib6rico en Dofiana e Implicaciones para su Conservaci6n. Ph.D. Thesis. Universidad Aut6noma de Madrid.
Foose, T.J. (1987) Species survival plans and overall management strategies. In Tigers of the World (R.L. Tilson and U.S. Seal, eds) pp. 304-16. Park Ridge: Noyes Publications.
Foose, T.J. (1989) Species survival plans: the role of captive propagation in conservation strategies. In Conservation Biology and the Black-footed Ferret (U.S. Seal, E.T. Thorne, M.A. Bogan and S.H. Anderson, eds) pp. 210-22. Dexter: Yale University Press.
Fritts, S.H., Paul, W.G. and Mech, L.D. (1984) Movements of translocated wolves in Minnesota. J. Wildl. Manage. 48, 709-21.
Fritts, S.H. and Sealander, J.A. (1978) Reproductive biology and population characteristics of bobcats (Lynx rufus) in Arkansas. J. Mature. 59, 347-53.
Ginsberg, J.R. (1994) Captive breeding, reintroduction and the conservation of canids. In Creative Conservation: Interactive management of wild and captive animals (P.J.S. Olney, G.M. Mace and A.T.C. Feistner, eds) pp. 365--83. London: Chapman and Hall.

Experimental release o f an Iberian lynx 393
Griffith, B., Scott, J.M., Carpenter, J.W. and Reed, C. (1989) Translocation as a species conservation tool: status and strategy. Science 245, 477-80.
IUCN (1987) The IUCN Position Statement on Translocation of Living Organisms: Introductions, re-introductions and restocking. Gland: IUCN.
Kleiman, D.G. (1980) The sociobiology of captive propagation. In Conservation Biology. An evolutionary-ecologicalperspective (M.E. Soul6 and B.A. Wilcox, eds) pp. 243--61. Sunderland: Sinauer.
Kleiman, D.G. (1989) Reintroduction of captive mammals for conservation: guidelines for reintroducing endangered species into the wild. BioSci. 39, 152-61.
Kleiman, D.G., Stanley Price, M.R. and Beck, B.B. (1994) Criteria for reintroductions. In Creative Conservation: Interactive management of wild and captive animals (P.J.S. Olney, G.M. Mace and A.T.C. Feistner, eds) pp. 287-303. London: Chapman and Hall.
Kleiman, D.G., Beck, B.B., Dietz, J.M. and Dietz, L.A. (1991) Costs of a re-introduction and criteria for success: accounting and accountability in the golden lion tamarin conservation program. Syrup. zool. Soc. Lond. 62, 125--42.
Kleiman, D.G., Beck, B.B., Dietz, J.M., Dietz, L.A., Ballou, J.D. and Coimbra-Filho, A.F. (1986) Conservation program for the golden lion tamarin: captive research and management, ecological studies, educational strategies, and reintroduction. In Primates: the road to self-sustaining populations (K. Benirschke, ed.) pp. 959--79. New York: Springer Verlag.
Lindburg, D.G. (1992) Are wildlife reintroductions worth the cost? Zoo Biol. 11, 1-2. Mallinson, J. (1978) The Shadow of Extinction. London: Macmillan. May, R.M. (1991) The role of ecological theory in planning re-introduction of endangered species.
Syrup. zool. Soc. Lond. 62, 145-63. McArthur, K.L. (1981) Factors contributing to effectiveness of black bear transplants. J. Wildl.
Manage. 45, 102-10. Miller, S.D. and Ballard, W.B. (1982) Homing of transplanted Alaskan brown bears. J. Wildl.
Manage. 46, 869-76. Mills, M.G.L. (1991) Conservation management of large carnivores in Africa. Koedoe 34, 81-90. Moore, D.E. and Smith, R. (1991) The red wolf as a model for carnivore re-introductions. Symp.
zool. Soc. Lond. 62, 263-78. Palomares, F., Rodrfguez, A., Laffitte, R. and Delibes, M. (1991) The status and distribution of the
Iberian lynx Felis pardina (Temminck) in Coto Dofiana area, SW Spain. Biol. Conserv. 57, 159-69.
Pettifer, H.L. (1981) The experimental release of captive-bred cheetah (Acinonyx jubatus) into the natural environment. In Worldwide Furbearer Conference Proceedings (J.A. Chapman and D. Pursley, eds) pp. 1001-24. Falls Church, VA: R.R. Donnelley and Sons Co.
Rahbek, C. (1993) Captive breeding - a useful tool in the preservation of biodiversity? Biodiv. Conserv. 2, 426-37.
Rodrfguez, A. and Delibes, M. (1990) El Lince Ibdrico (Lynx pardina) en Espaha. Distribuci6n y problemas de conservaci6n. Madrid: Instituto National para la Conservaci6n de la Naturaleza.
Rodrlguez, A. and Delibes, M. (1992) Current range and status of the Iberian lynx Felis pardina Temminck, 1824. Biol. Conserv. 61, 189-96.
Rolley, R.E. (1987) Bobcat. In Wild Furbearer Management and Conservation in North America (M. Novak, J.A. Baker, M.E. Obbard and B. Malloch, eds) pp. 670--81. Toronto: Ministry of Natural Resources.
Seal, U.S. (1986) Goals of captive propagation programmes for the conservation of endangered species. Int. Zoo Yb. 24/25, 174-9.
Shepherdson, D. (1994) The role of environmental enrichment in the captive breeding and reintroduction of endangered species. In Creative Conservation: Interactive management of wild and captive animals (P.J.S. Olney, G.M. Mace and A.T.C. Feistner, eds) pp. 167-77. London: Chapman and Hall.

394 Rodriguez et al.
Simonetti, J.A. (1989) Tasas de defecaci6n y descomposici6n de fecas de Oryctolagus cuniculus en Chile central. Medio Ambiente (Chile) 10, 92-5.
Slough, B.G. (1989) Movements and habitat use by transplanted martens in the Yukon territory. J. Wildl. Manage. 53, 991-7.
Stanley Price, M.R. (1989) Animal Re-introductions: The Arabian orvx in Oman. Cambridge: Cambridge University Press.
Stanley Price, M.R. (1991) A review of mammal re-introductions and the role of the Re-introduction Specialist Group of IUCN/SSC. Syrup. zool. Soc. Lond. 62, 9-25.
Stuart, S.N. (1991) Re-introductions: to what extent are they needed? Syrup. zool. So¢. Lond. 62, 27-37.
Wilson, A.C. and Stanley Price, M.R. (1994) Reintroduction as a reason for captive breeding. In Creative Conservation: Interactive management of wild and captive animals (P.J.S. Olney, G.M. Mace and A.T.C. Feistner, eds) pp. 243~4. London: Chapman and Hall.
Yalden, D.W. (1993) The problems of reintroducing carnivores. Symp. zooL Soc. Lond. 65. 289-306.
Related Documents











