This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Experimental and computational study of inter- and intra- species specificityof gonadotropins for various gonadotropin receptors
Joseph Aizen a, Noga Kowalsman b, Makito Kobayashi c, Lian Hollander a, Young Chang Sohn d,Goro Yoshizaki e, Masha Y. Niv b,f, Berta Levavi-Sivan a,⇑a The Robert H. Smith Faculty of Agriculture, Food and Environment, Department of Animal Sciences, The Hebrew University of Jerusalem, Rehovot 76100, Israelb The Robert H. Smith Faculty of Agriculture, Food and Environment, The Institute of Biochemistry, Food Science and Nutrition, The Hebrew University of Jerusalem,Rehovot 76100, Israelc Department of Life Sciences, International Christian University, Osawa, Mitaka, Tokyo 181-8585, Japand Department of Marine Molecular Biotechnology, Gangneung-Wonju National University, Gangneung 210-702, South Koreae Department of Marine Biosciences, Tokyo University of Marine Science and Technology, Minato, Tokyo 108-8477, Japanf The Fritz Haber Research Center for Molecular Dynamics, The Hebrew University of Jerusalem, Jerusalem 91904, Israel
a r t i c l e i n f o
Article history:Received 25 March 2012Received in revised form 20 August 2012Accepted 21 August 2012Available online 30 August 2012
Keywords:LHFSHMolecular modelingG-protein-coupled receptor (GPCR)Signal transductionGlycoprotein hormone receptor
a b s t r a c t
The gonadotropins follicle-stimulating hormone (FSH) and luteinizing hormone (LH) and their receptorsplay critical roles in vertebrate reproduction. In order to study intra- and interspecies ligand promiscuityof gonadotropins, COS-7 cells were transiently transfected with one of the gonadotropin receptor genes,FSHR or LHR, and tested for activation by gonadotropins from representative fish orders: Aquilliformes(eel; e), Salmoniformes (trout; tr), and Perciformes (tilapia; ta), and of mammalian origin: porcine (p),bovine (b) and human (h).
The study reveals complex relations between the gonadotropin hormones and their receptors. Eachgonadotropin activated its own cognate receptor. However, taLHR was also activated by hCG and eLHRwas activated by hFSH, hCG, and trFSH. For FSHR, the only cross-reactivity detected was for hFSHR, whichwas activated by pFSH and bFSH. These findings are of great interest and applicability in the context ofactivation of various GTHRs by their ligands and by ligands from other vertebrates. Analysis of the three-dimensional models of the structures highlights the importance of residues outside of the currentlyestablished hormone-receptor interface region. In addition, the interface residues in taFSHR and theeffect of exon duplication, which causes an insert in the LRR domain, are suggested to affect the interac-tion and binding of taFSH.
� 2012 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
The gonadotropins (GTHs) follicle-stimulating hormone (FSH)and luteinizing hormone (LH) are key regulators of vertebratereproduction. Together with thyroid-stimulating hormone (TSH)and chorionic gonadotropin (CG), FSH and LH belong to the glyco-protein hormone family whose members bear carbohydrate moie-
ties that influence their biosynthesis, secretion, half life andbiological potency (Pierce and Parsons, 1981). Each of these hor-mones is comprised of a common a subunit noncovalently boundto a variable b subunit that confers the receptor-binding specificityfor each dimeric ligand (Pierce and Parsons, 1981). In fish as well,two distinct GTHs (referred to as FSH and LH) have been discoveredand the cDNA of the corresponding specific subunits have beencloned. The availability of molecular cloning techniques has al-lowed isolation of the genes coding for the GTH subunits in 56 fishspecies representing 14 teleost orders (Levavi-Sivan et al., 2010).
The temporal pattern of GTH secretion in fish suggests that FSHand LH are secreted differentially during the reproductive cycle.FSH is considered to regulate early phases of gametogenesis, suchas vitellogenesis and spermatogenesis, whereas LH is involved inthe final maturation processes, such as oocyte maturation and ovu-lation in the female, and spermiation and milt production in themale (Levavi-Sivan et al., 2010). As in other vertebrates, oncereleased into the circulation, GTHs bind to membrane-bound
0303-7207/$ - see front matter � 2012 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.mce.2012.08.013
Abbreviations: 7TMD, seven-transmembrane domain; b, bovine; CG, chorionicgonadotropin; e, Japanese eel (Anguilla japonica); ECD, extracellular domain; FSH,follicle-stimulating hormone; FSHR, FSH receptor; GPCR, G-protein-coupled recep-tor; GTH, gonadotropins; GTHR, gonadotropin receptor; h, human; LH, luteinizinghormone; LHR, LH receptor; LRR, leucine-rich repeat; Luc, luciferase; p, porcine;PKA, protein kinase A; ta, tilapia (Oreochromis niloticus); TPA, 12-O-tetradecanoyl-phorbol-13-acetate; tr, Manchurian trout (Brachymystax lenok); TSH, thyroid-stimulating hormone.⇑ Corresponding author. Address: Department of Animal Sciences, The Robert H.
Smith Faculty of Agriculture, Food and Environment, The Hebrew University, P.O.Box 12, Rehovot 76100, Israel. Tel.: +972 8 9489988; fax: +972 8 9489307.
E-mail address: [email protected] (B. Levavi-Sivan).
Molecular and Cellular Endocrinology 364 (2012) 89–100
Contents lists available at SciVerse ScienceDirect
Molecular and Cellular Endocrinology
journal homepage: www.elsevier .com/locate /mce

Author's personal copy
receptors expressed on the somatic cells of the gonads. The pres-ence of two distinct GTHRs in a single fish species was confirmedby the molecular cloning of their two distinct cDNAs in several fishspecies belonging to different taxa (reviewed in Levavi-Sivan et al.,2010).
The FSH and LH receptors are G-protein-coupled receptors(GPCRs) of the rhodopsin-like receptor family (family A) (Gether,2000). These GPCRs consist of a large extracellular domain (ECD)characterized by multiple leucine-rich repeats (LRRs) that are crit-ical for specific hormone binding, a seven-transmembrane domain(7TMD), and a short intracellular C terminus (Vassart et al., 2004).The LRR-containing ectodomain of GTHRs is responsible for thebinding specificity of their specific ligands, which translates intoactivation of the rhodopsin-like serpentine domain, itself responsi-ble for transducing the signal within the cell, mainly via activationof the stimulatory G proteins (Vassart et al., 2004; Karges et al.,2005). Fish GTHRs follow this general structure, but the ECDs offish FSHRs contain remarkable differences compared to mamma-lian FSHRs (Levavi-Sivan et al., 2010). Integration of the molecularphylogeny of glycoprotein hormone receptors with the analysis ofsyntenic loci of vertebrate orthologs, and subsequent characteriza-tion of the physical maps with the phylogeny of flanking genes, im-plies that vertebrate glycoprotein hormone receptors haveundergone a divergent evolution (Chauvigne et al., 2010).
Ligand selectivity and binding specificity of mammalian GTHRsto their respective agonists are well defined (Costagliola et al.,2005). Experimental evidence has shown the importance of severalresidues for the hormones’ binding to, and selectivity for theirrespective receptors. For example: in human GTHs, the region be-tween the Cys residues in positions 10 and 12 in the b subunit, the‘‘seatbelt’’ region, was found to be important for selective bindingof the hormone (Campbell et al., 1991). In hFSHR, residues in LRRs1, 7 and 8 were found to be involved in hFSH binding (reviewed by(Bogerd, 2007)). Residues important for binding between hCG andhLHR were identified in LRR3 and LRR6 regions in hLHR (Bogerdet al., 2005; Bogerd, 2007).
In contrast to the mammalian receptors, in some teleosts, FSHRand LHR can be cross-activated by their ligands and therefore, tel-eost receptor specificity is unclear (Levavi-Sivan et al., 2010). Stud-ies on GTHRs from representatives of the piscine ordersSiluriformes (Kumar et al., 2001a; Kumar et al., 2001b; Vischerand Bogerd, 2003) and Cypriniformes (Kwok et al., 2005; Soet al., 2005) have indicated that FSHRs show a preference for FSHbut also respond to LH, whereas LHRs respond specifically to LH.However, in the sea bass, each GTHR is exclusively activated byits corresponding hormone (Rocha et al., 2007). In trout, purifiedGTHs from pituitaries specifically activate their cognate receptor(Sambroni et al., 2007). Studies on receptors from a salmonid fish,the amago salmon (Oba et al., 1999a, 1999b), concluded that LHR,but not FSHR, responds to both GTHs.
What are the molecular determinants that dictate the level ofinteraction specificity? A structural view of the interactions islikely to provide some clues. Indeed, crystal structures of GTHs,and in particular the crystallization of hFSH/hFSHR–ECD complex(PDB entry 1XWD; Fan and Hendrickson, 2005), have paved theway to increasingly accurate modeling of GTHs and their receptorsand to revealing their intermolecular interactions (Caltabiano et al.,2008). Several reports have used molecular modeling to under-stand the effect of naturally occurring mutations in hCG and hLHRrelated to syndromes in the human reproductive system (Leunget al., 2006; De Leener et al., 2008). Mutations in GTHRs and struc-ture–function information can be accessed via databases (Van Dur-me et al., 2006; Kleinau et al., 2007; Kreuchwig et al., 2011).Structural modeling in combination with mutagenesis experi-ments provided a better understanding of hormone/receptor–ECDinteractions and specificities (Bhowmick et al., 1996; Liu and Dias,
1996; Miguel et al., 2008; Angelova et al., 2010; Puett et al., 2007)and signal transduction by the GPCR domain of GTHRs (Puett et al.,2007; Angelova et al., 2008; Puett et al., 2010; Angelova et al.,2011).
Most studies have performed molecular modeling of humanGTHs and receptors, but a small number of studies have modeledstructures of other species, e.g. rat FSHR and LHR, as summarizedin the SSFA-GPHR and GRIS servers (Kleinau et al., 2007; Van Dur-me et al., 2006), combinations of a and b subunits for horse anddonkey CG and FSH (Chopineau et al., 2004), duck FSH (Shenet al., 2006), horse TSHR (Royer et al., 2008) and the LH and FSHb subunits of snakehead fish (Chatterjee et al., 2005).
Recent progress in the isolation of fish GTH cDNAs has enabledthe production of a variety of species-specific recombinant GTHs(Levavi-Sivan et al., 2008). The availability of recombinant GTHsfrom different fish species, in parallel to the cloning of various pis-cine GTHRs and modeling of possible ligand–receptor interactions,has provided an opportunity for further analysis of the connectionsbetween these important ligands and their receptors.
In the current study, we used ligand–receptor structural model-ing and recombinant GTHs from fish species representing three tel-eost orders, to study the molecular basis of ligand selectivity. Theavailable biodiversity of hormones and receptors on the one hand,and cross-species reactivity on the other, provide the motivationfor understanding hormone specificity as well as naturally occur-ring mutations that might reveal key determinants of selectivity.
2. Materials and methods
2.1. Recombinant GTH production and purification
2.1.1. Tilapia (Oreochromis niloticus)Using the methylotrophic yeast Pichia pastoris, recombinant
tilapia (ta) LH and FSH were produced as a biologically active,single-chain polypeptide as described previously ((Kasuto andLevavi-Sivan, 2005) and (Aizen et al., 2007a), respectively). Briefly,the taLHb (GenBank Accession No. AAP49576) or taFSHb (GenBankAccession No. AAP49575) and a (GenBank Accession No.AAP49577) mature protein-coding sequences were joined to forma fusion gene that encodes a ‘‘tethered’’ polypeptide in whichone of the b chains forms the N-terminal part and the a chainforms the C-terminal part. A ‘‘linker’’ sequence of six amino acids(three Gly–Ser pairs) was placed between the a and b chains to as-sist in the chimerization of the subunits, and a six-His tail wasplaced at the end of the b subunit to enable purification of the re-combinant protein. Recombinant taLHba and taFSHba were able tostimulate the release of 11-ketotestosterone from the testes, or therelease of estradiol from the ovaries (Kasuto and Levavi-Sivan,2005; Aizen et al., 2007a; Levavi-Sivan et al., 2008).
2.1.2. Japanese eel (Anguilla japonica)Using baculovirus, recombinant Japanese eel (e) FSH and LH
were produced in silkworm larvae as a biologically active, single-chain polypeptide (Kobayashi et al., 2010). Biological activities ofrecombinant eFSH and eLH were confirmed by in vitro oocyte mat-uration and in vivo spermatogenesis (Kobayashi et al., 2010).
2.1.3. Manchurian trout (Brachymystax lenok)Using baculovirus, recombinant Manchurian trout (tr) FSH and
LH were produced in silkworm larvae as a biologically active, sin-gle-chain polypeptide (Kobayashi et al., 2006; Ko et al., 2007). Bio-logical activities of recombinant trFSH and trLH were confirmed byin vitro receptor activation, sex steroid production, and in vivoovarian development and milt production (Ko et al., 2007).
90 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100

Author's personal copy
All recombinant GTHs were dissolved in PBS at a concentrationof 1 lg/ll, aliquoted and kept at –80 �C until use.
2.2. Materials
Dulbecco’s modified Eagle’s medium (DMEM), trypsin–EDTAand antibiotics were purchased from Biological Industries (BetHaEmek, Israel). Fetal bovine serum (FBS) was purchased fromInvitrogen. BSA, human (h) CG, bovine (b) FSH and porcine (p)FSH were purchased from Sigma (Ness Ziona, Israel). FuGENE 6was purchased from Roche (Israel). As recombinant hFSH we usedGonal-F (Merck Serono, Israel).
2.3. Constructs and luciferase reporter gene assay
Since both FSHR and LHCGR signaling are predominantly med-iated through the cAMP/protein kinase A (PKA) pathway to pro-mote steroid production, we used this signaling pathway for thecomparison between the different GTHs. To activate the cAMP/PKA signal-transduction pathways, we used a sensitive luciferase(LUC) reporter gene assay where LUC is transcriptionally regulatedby a cAMP response element (CRE; Invitrogen). The pCRE-LUC plas-mid codes for a CRE upstream of the LUC reporter gene. The spec-ificity of the response is confirmed when the CRE-LUC reportersystem is shown to be significantly activated by forskolin, an aden-ylate cyclase activator, but not by 12-O-tetradecanoylphorbol-13-acetate (TPA), a protein kinase C activator (Biran et al., 2008).
An in vitro LUC reporter assay for GTHR transactivation was ap-plied. The assay was based upon a COS-7 cell line transiently trans-fected with a DNA construct containing one of the GTHR genes, andan additional construct containing the LUC reporter gene. In eachanalytical run, three dose–response curves divided over three 96-well plates were prepared, with approximately 3.4 � 104 cells perwell in culture medium. Each dose–response curve consisted of ablank (assay medium only) and 11 serial dilutions to obtain con-centrations of 500.0–0.1 ng recombinant GTH per well. Plates wereincubated in a CO2 incubator (37 �C, 5% CO2), and LUC activity wasmeasured as luminescence counts using a TriStar LB 941 platereader (Berthold Technologies, Germany) and analyzed by Mikro-win 2000 version 4.4 software (Mikrotec, Germany). To comparethe biological activities of the different test items, their half-max-imal effective concentrations were calculated using Prism software(version 4.03; GraphPad, San Diego, CA, USA) and compared with areference standard.
The cDNAs encoding eFSHR and eLHR were kindly donated byDr. Y. Kazeto (Division of Marine Life Sciences, Research Facultyof Fisheries Science, Hokkaido University, Japan); hFSHR was ob-tained from the Missouri S&T cDNA Resource Center(www.cdna.org). hLHR was kindly donated by Dr. T. Gudermann(Walther-Straub-Institute of Pharmacology and Toxicology, Lud-wig-Maximilians University of Munich, Germany).
The open reading frames of tilapia and trout LHR and FSHR weresubcloned by PCR, according to known sequences from GenBank:rainbow trout (Onchorynchus mykiss) FSH and LH (AF439405 andAF439404, respectively); Nile tilapia (Oreochromis niloticus) FSHand LH (AB041763 and AB041762, respectively). Total RNA was ex-tracted from fish testes and ovaries using Total RNA Isolation Re-agent (Trizol) and treated with DNase using the RNase-FreeDNase kit (Qiagen) following the manufacturer’s instructions.PCR primers were designed to cover the beginning and end ofthe receptors (Supplemental Table S1). The primers were used asa template for the Advantage 2 PCR System (Clontech), accordingto the manufacturer’s instructions. The PCR products were ana-lyzed by 1.5% agarose gel electrophoresis and stained with ethi-dium bromide. A PCR product of the predicted size for thedesired receptor was obtained and cloned into pGEMT Easy Vector
(Promega). The clones were sequenced by the Weizmann InstituteSequencing Unit (Rehovot, Israel). The desired receptors werecloned in pcDNA3.1 expression vector (Zeo-; Invitrogen) undercontrol of the CMV promoter.
2.4. Cell culture and transient transfection of cells
Transient transfection, cell procedures and stimulation proto-cols were generally performed as described previously (Levavi-Si-van et al., 2005; Biran et al., 2008). Briefly, COS-7 cells weregrown in DMEM supplemented with 10% FBS, 1% glutamine,100 U/ml penicillin, and 100 mg/ml streptomycin (Biological Indus-tries) under 5% CO2 until confluent. Co-transfection of the desiredreceptors (at 3 lg/plate), a reporter plasmid (at 3 lg/plate for FSHsand 0.3 lg/plate for LHs), and pCMV-b-galactosidase (at 1 lg/plate)was carried out with FuGENE 6. The cells were serum-starved for36 h, stimulated with various agent concentrations for 6 h, and thenharvested and analyzed. Lysates prepared from the harvested cellswere assayed for both LUC and b-galactosidase activity, the latterused as an internal standard to normalize the LUC activity directedby the test plasmid as described previously (Levavi-Sivan et al.,2005). Experiments were repeated a minimum of three times fromindependent transfections, each performed in triplicate. COS-7 cellstransfected with a vector without GTHR cDNA showed no change inLUC activity in any experimental group (data not shown).
2.5. Sequence alignment and analysis
The sequences of the GTHs and their receptors were extractedfrom the ExPASy server using the UniProtKB database (Gasteigeret al., 2003; Wu et al., 2006) (see Supplementary Table S2). Toinvestigate the binding between the GTHs and GTHRs, we usedonly the ECD part of the receptors in our sequence and structuralanalyses.
Multiple sequence alignments were performed using thePROMALS3D server (http://prodata.swmed.edu/promals3d/pro-mals3d.php) with default parameters (Pei et al., 2008) or ClustalW(http://www.ebi.ac.uk/Tools/msa/clustalw2/) (Larkin et al., 2007)The ClustalW multiple sequence alignments were used for calculat-ing conservation scores. Sequence identities between human andpig or human and bovine FSH a and b subunits were calculated bythe EMBOSS Stretcher tool (http://www.ebi.ac.uk/Tools/psa/em-boss_stretcher/) (Myers and Miller, 1988) which is part of theEMBOSS package (Rice et al., 2000). EMBOSS Stretcher calculatesan optimal global alignment of two sequences. The calculationwas performed for the protein only, without the signal peptide.For identification of subtype-specific sites in multiple sequencealignments, the servers multi-Harmony (http://www.ibi.vu.nl/pro-grams/shmrwww/) (Brandt et al., 2010) and SDPpred, which pre-dicts amino acid residues that determine differences in functionalspecificities of homologous proteins (http://bioinf.fbb.msu.ru/SDP-pred/) (Kalinina et al., 2004), were used.
The ExPASy server provides details on the signal peptide forhGTHs and hGTHRs, porcine and bovine FSH and eGTHs. To identifythe signal peptide in the other sequences, the server Signal-3L(http://www.csbio.sjtu.edu.cn/bioinf/Signal-3L/) was used. Signal-3L is an automated method for predicting signal peptide sequencesand their cleavage sites in eukaryotic and bacterial protein se-quences (Shen and Chou, 2007). Secondary structure was predictedvia the PSIPRED Protein Structure Prediction Server (http://bio-inf.cs.ucl.ac.uk/psipred/) (Jones, 1999; Bryson et al., 2005).
2.6. Structural models of the GTHs and their receptors
Models of the GTH and GTHR proteins were prepared via theI-TASSER server (http://zhanglab.ccmb.med.umich.edu/I-TASSER/)
J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100 91

Author's personal copy
(Zhang, 2007; Roy et al., 2010). For the receptor models we direc-ted the I-TASSER server to use the structure of the human FSH/FSHR–ECD complex (PDB entry 1XWD (Fan and Hendrickson,2005)) as one of the templates. For the ligand models, we used de-fault parameters with no predefined template.
Visualization and superposition of the models were performedusing Discovery Studio 2.5 (Accelrys Inc.). To create GTH/GTHR–ECD complex models, sequence alignment-based superpositionwas used taking the 1XWD structure as template. In addition tocomplexes for each species, we created models of the complexesof mixed species: pFSH/hFSHR, bFSH/hFSHR, hCG/taLHR hFSH/eLHR, hCG/eLHR, and trLH/eLHR. These models were subjected tominimization of 3000 Steepest-Descent steps using Discovery Stu-dio 2.5 minimization protocol using an all-atom general purposeCHARMm Momany and Rone force-field (Momany and Rone,1992), which is the Discovery Studio 2.5 default force field.
We used the known interface residues of the hFSH–hFSHR–ECDstructure (1XWD) in order to extract the list of residues found atthe interface of the modeled GTH/GTHR–ECD complexes. Toidentify the interface residues in the modeled GTH and GTHRswe used the multiple sequence alignments and the structuralalignment of the models. For each GTH and GTHR, residues thatwere found in the equivalent positions to the interface residuesof 1XWD structure were considered as ‘‘interface residues’’ for thatspecific GTH or GTHR. Note, that the numbers of the residues werefer to are based on the whole sequence as extracted fromUniProt.
2.7. Electrostatic and charge calculation
Electrostatic calculation of the GTHs and their receptors wasconducted using DelPhi (Sitkoff et al., 1994) electrostatic calcula-tion module as implemented in the Discovery Studio 2.5 packageusing default parameters. Delphi solves the Poisson–Boltzmannequation using the finite-difference techniques on a cubic lattice.
All the residues in human FSH that face the FSHR in 1XWD com-plex structure were manually extracted (see Supplemental TableS3). For each of these residues the overall charge was calculatedby summing the partial charge of its atoms using the electrostaticcalculation module. The resulting list of residues and their chargeare presented in Supplemental Table S3.
2.8. Statistical analysis
In order to be able to compare the relative activities of all re-combinant hormones used, we set the maximal response of LHand FSH to their respective cognate receptors to 100%. Resultsare expressed as percentage of this value. All data are presentedas mean ± SEM. Ligand concentrations that induce half-maximalstimulation (i.e. EC50 values) were determined by fitting the CRE-LUC reporter gene activity to sigmoidal dose–response curvesusing GraphPad PRISM.
3. Results
3.1. GTH specificity to their receptors
The first aim of this research was to characterize intra- andinterspecies ligand cross-reactivity of GTHs from various species.We studied the effects of the recombinant GTHs of human, tilapia,trout and eel on their own cognate and complementary receptors(FSHR and LHR), and on the GTHRs of the other species. Ligand-in-duced activation of fish GTHRs was studied by transient transfec-tion assay in COS-7 cells expressing one of the receptors.
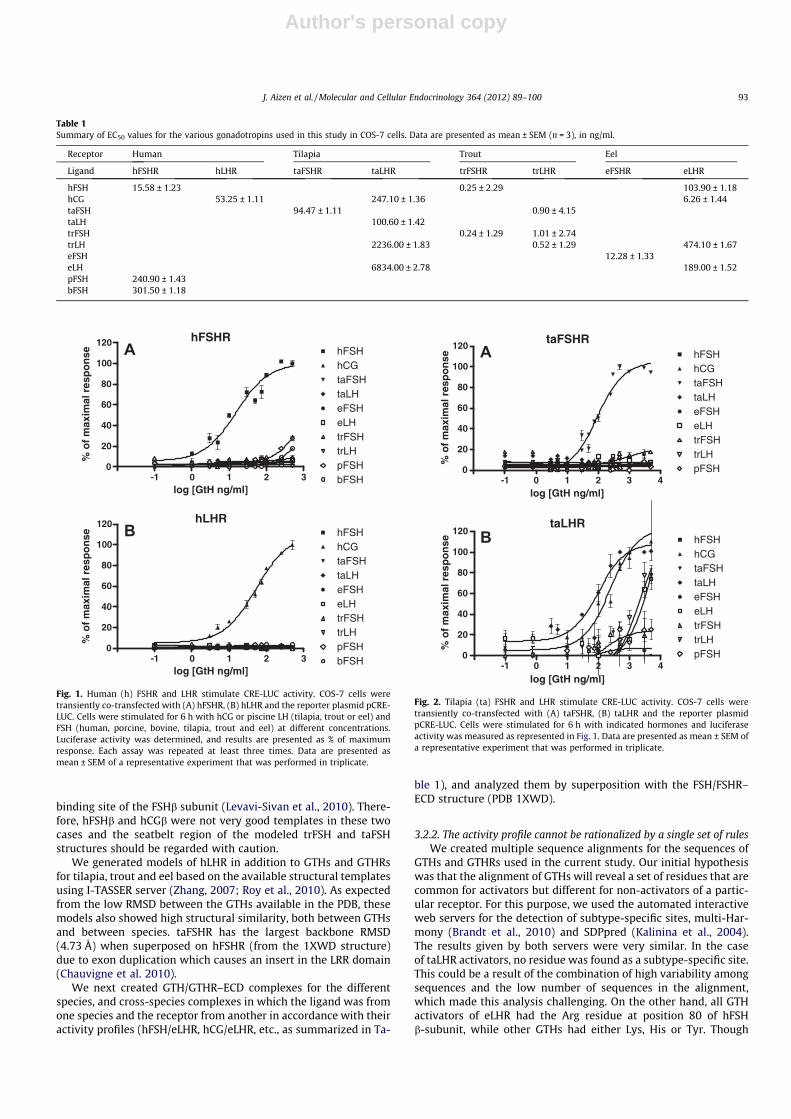
Each hGTH was able to activate its own cognate receptor. hFSHwas most efficient at activating hFSHR (minimal effective dose3 ng/ml; Table 1), while pFSH and bFSH had much lower potenciesand minor effects (27.87% ± 2.06 and 18.12% ± 1.26 of maximalactivation, respectively; Table 1). Of all GTHs tested in this study,only hCG could activate hLHR (EC50 = 53.25 ± 1.11 ng/ml), (Table1, Fig. 1B).
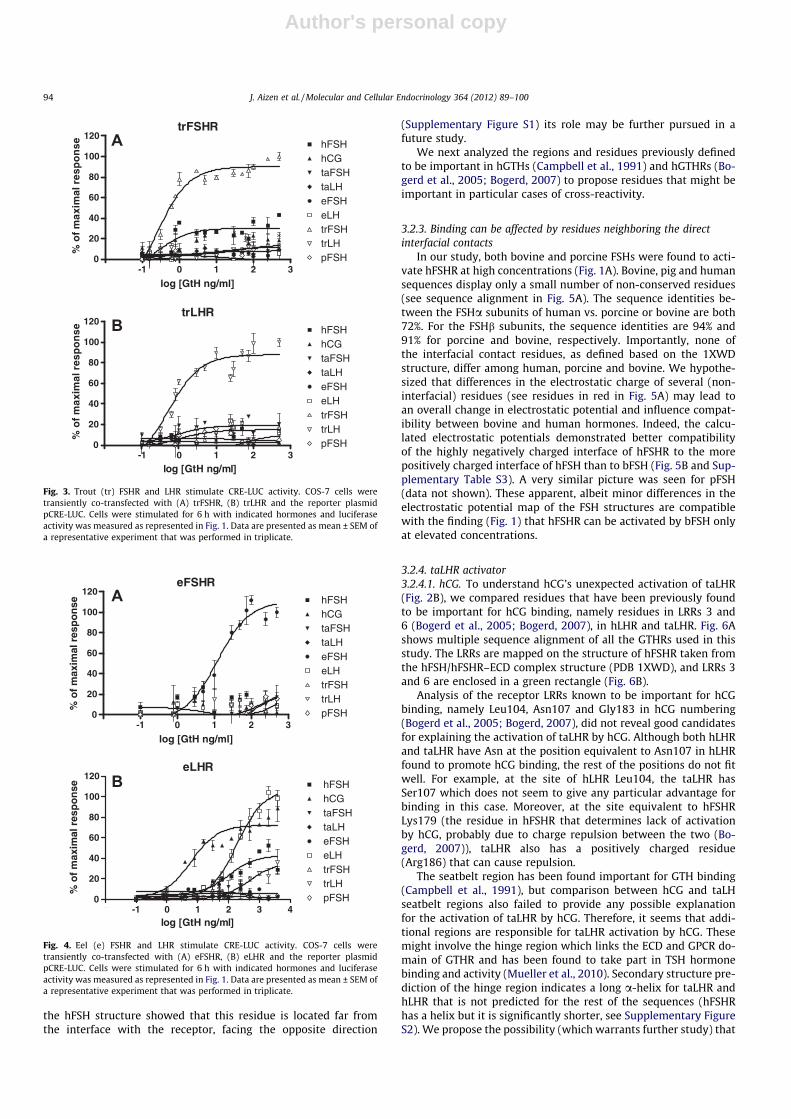
Recombinant taFSH significantly stimulated CRE-LUC activity inCOS-7 cells expressing taFSHR in a dose-responsive manner, buthad no effect on COS-7 cells expressing taLHR at any of the concen-trations tested (Fig. 2). Surprisingly, although taLHR was most effi-ciently activated by its own cognate ligand, taLH, it wasequipotently activated by hCG (EC50 = 247.10 ± 1.36 ng/ml and100.60 ± 1.42 ng/ml, respectively; Fig. 2B, Table 1). LH of less-evolved fish (eel and trout) stimulated CRE-LUC activity of taLHRwith very high EC50 values (Fig. 2B, Table 1). trFSHR was efficientlyactivated by trFSH, at a minimal effective dose of less than 1 ng/mland EC50 of 0.24 ± 1.29 ng/ml (Fig. 3A, Table 1). This receptor wasalso activated by hFSH, but only to a low extent. trLHR was acti-vated by recombinant trLH (minimal effective dose 0.5 ng/ml andEC50 value of 0.52 ± 1.29 ng/ml), and to a very low extent also bytaFSH and hFSH (Fig. 3B, Table 1). eFSH produced a concentra-tion-dependent increase in the activation of eFSHR, with a minimaleffective dose of 5 ng/ml and EC50 value 12.28 ± 1.33 ng/ml(Fig. 4A, Table 1). Interestingly, eLHR was activated not only byeLH (EC50 = 189.00 ± 1.52 ng/ml), but also by hCG (EC50 = 6.26 ±1.44 ng/ml), hFSH (EC50 = 103.90 ± 1.18 ng/ml) and even trLH(EC50 = 474.10 ± 1.7 ng/ml; Fig. 4B, Table 1).
3.2. Rationalization of the activation profiles using modeling andbioinformatics analysis
To rationalize the activation profile of GTHRs by the different li-gands, the sequences of the GTHs and GTHRs, as well as the struc-tural and physicochemical details of the ligand–receptorinteractions were analyzed. In particular, we were intrigued bythe activation of hFSHR by high concentrations of porcine and bo-vine FSH (Fig. 1). We also wondered what enables activation of taL-HR by hCG in addition to the cognate ligand (Fig. 2), and whatenables activation of eLHR, not only by eLH as expected, but alsoby trLH, hFSH and hCG (Fig. 4).
3.2.1. Structures and models of GTHs and their receptorsWe began by inspecting the existing structures of representa-
tive hGTHs and their receptors. The PDB database contains severalhGTH structures: hCGa (PDB entries 1E9J, 1HD4, 1DZ7 and 1HCV(Spinelli et al., 1996; Erbel et al., 1999; Erbel et al., 2000)), hCG(PDB entries 1QFW, 1HRP, 1HCN (Lapthorn et al., 1994; Wuet al., 1994; Tegoni et al., 1999)), hFSH (PDB 1FL7 (Fox et al.,2001)), hFSH bound to hFSHR–ECD (PDB 1XWD (Fan and Hendrick-son, 2005)) and the TSH binding domain of hTSHR (PDB 3GO4(Sanders et al., 2007)). To compare these available structures westructurally superimposed them using the Discovery Studio 2.5superposition by sequence alignment protocol. By superposingthe individual structures onto the units in the hFSH/hFSHR–ECDcomplex, we found high similarity between the different structuresof the GTH subunits and receptors, with backbone RMSD (rootmean square deviation) ranging from 1.6 to 2.31 Å. The high simi-larity between the structures and the sequence similarity betweenpiscine sequences and sequences of the structural templates, led usto conclude that in most cases, we can use the available crystalstructures (in particular the hFSH/hFSHR–ECD complex and thehCG structures) as templates for the fish GTHs and GTHRs. Note,however, that in trFSHb and taFSHb, the third Cys residue is miss-ing, leading to structural changes in the area between the 10th and12th Cys residues (the seatbelt region) which is a major part of the
92 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100
Author's personal copy
binding site of the FSHb subunit (Levavi-Sivan et al., 2010). There-fore, hFSHb and hCGb were not very good templates in these twocases and the seatbelt region of the modeled trFSH and taFSHstructures should be regarded with caution.
We generated models of hLHR in addition to GTHs and GTHRsfor tilapia, trout and eel based on the available structural templatesusing I-TASSER server (Zhang, 2007; Roy et al., 2010). As expectedfrom the low RMSD between the GTHs available in the PDB, thesemodels also showed high structural similarity, both between GTHsand between species. taFSHR has the largest backbone RMSD(4.73 Å) when superposed on hFSHR (from the 1XWD structure)due to exon duplication which causes an insert in the LRR domain(Chauvigne et al. 2010).
We next created GTH/GTHR–ECD complexes for the differentspecies, and cross-species complexes in which the ligand was fromone species and the receptor from another in accordance with theiractivity profiles (hFSH/eLHR, hCG/eLHR, etc., as summarized in Ta-
ble 1), and analyzed them by superposition with the FSH/FSHR–ECD structure (PDB 1XWD).
3.2.2. The activity profile cannot be rationalized by a single set of rulesWe created multiple sequence alignments for the sequences of
GTHs and GTHRs used in the current study. Our initial hypothesiswas that the alignment of GTHs will reveal a set of residues that arecommon for activators but different for non-activators of a partic-ular receptor. For this purpose, we used the automated interactiveweb servers for the detection of subtype-specific sites, multi-Har-mony (Brandt et al., 2010) and SDPpred (Kalinina et al., 2004).The results given by both servers were very similar. In the caseof taLHR activators, no residue was found as a subtype-specific site.This could be a result of the combination of high variability amongsequences and the low number of sequences in the alignment,which made this analysis challenging. On the other hand, all GTHactivators of eLHR had the Arg residue at position 80 of hFSHb-subunit, while other GTHs had either Lys, His or Tyr. Though
Table 1Summary of EC50 values for the various gonadotropins used in this study in COS-7 cells. Data are presented as mean ± SEM (n = 3), in ng/ml.
Receptor Human Tilapia Trout Eel
Ligand hFSHR hLHR taFSHR taLHR trFSHR trLHR eFSHR eLHR
hFSH 15.58 ± 1.23 0.25 ± 2.29 103.90 ± 1.18hCG 53.25 ± 1.11 247.10 ± 1.36 6.26 ± 1.44taFSH 94.47 ± 1.11 0.90 ± 4.15taLH 100.60 ± 1.42trFSH 0.24 ± 1.29 1.01 ± 2.74trLH 2236.00 ± 1.83 0.52 ± 1.29 474.10 ± 1.67eFSH 12.28 ± 1.33eLH 6834.00 ± 2.78 189.00 ± 1.52pFSH 240.90 ± 1.43bFSH 301.50 ± 1.18
hFSHR
-2 -1 0 1 2 30
20
40
60
80
100
120hFSH
pFSHbFSH
taFSHtaLH
eLH
trLHtrFSH
hCG
eFSH
A
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
hLHR
-2 -1 0 1 2 30
20
40
60
80
100
120
hCGhFSH
pFSHbFSH
taLHtaFSH
eLHeFSH
trLHtrFSH
B
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
Fig. 1. Human (h) FSHR and LHR stimulate CRE-LUC activity. COS-7 cells weretransiently co-transfected with (A) hFSHR, (B) hLHR and the reporter plasmid pCRE-LUC. Cells were stimulated for 6 h with hCG or piscine LH (tilapia, trout or eel) andFSH (human, porcine, bovine, tilapia, trout and eel) at different concentrations.Luciferase activity was determined, and results are presented as % of maximumresponse. Each assay was repeated at least three times. Data are presented asmean ± SEM of a representative experiment that was performed in triplicate.
taFSHR
-2 -1 0 1 2 3 40
20
40
60
80
100
120
eLHeFSH
trLHtrFSH
pFSH
hCGtaFSHtaLH
hFSHA
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
taLHR
-2 -1 0 1 2 3 40
20
40
60
80
100
120
eLHeFSH
trLHtrFSH
pFSH
hCGtaFSHtaLH
hFSHB
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
Fig. 2. Tilapia (ta) FSHR and LHR stimulate CRE-LUC activity. COS-7 cells weretransiently co-transfected with (A) taFSHR, (B) taLHR and the reporter plasmidpCRE-LUC. Cells were stimulated for 6 h with indicated hormones and luciferaseactivity was measured as represented in Fig. 1. Data are presented as mean ± SEM ofa representative experiment that was performed in triplicate.
J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100 93
Author's personal copy
the hFSH structure showed that this residue is located far fromthe interface with the receptor, facing the opposite direction
(Supplementary Figure S1) its role may be further pursued in afuture study.
We next analyzed the regions and residues previously definedto be important in hGTHs (Campbell et al., 1991) and hGTHRs (Bo-gerd et al., 2005; Bogerd, 2007) to propose residues that might beimportant in particular cases of cross-reactivity.
3.2.3. Binding can be affected by residues neighboring the directinterfacial contacts
In our study, both bovine and porcine FSHs were found to acti-vate hFSHR at high concentrations (Fig. 1A). Bovine, pig and humansequences display only a small number of non-conserved residues(see sequence alignment in Fig. 5A). The sequence identities be-tween the FSHa subunits of human vs. porcine or bovine are both72%. For the FSHb subunits, the sequence identities are 94% and91% for porcine and bovine, respectively. Importantly, none ofthe interfacial contact residues, as defined based on the 1XWDstructure, differ among human, porcine and bovine. We hypothe-sized that differences in the electrostatic charge of several (non-interfacial) residues (see residues in red in Fig. 5A) may lead toan overall change in electrostatic potential and influence compat-ibility between bovine and human hormones. Indeed, the calcu-lated electrostatic potentials demonstrated better compatibilityof the highly negatively charged interface of hFSHR to the morepositively charged interface of hFSH than to bFSH (Fig. 5B and Sup-plementary Table S3). A very similar picture was seen for pFSH(data not shown). These apparent, albeit minor differences in theelectrostatic potential map of the FSH structures are compatiblewith the finding (Fig. 1) that hFSHR can be activated by bFSH onlyat elevated concentrations.
3.2.4. taLHR activator3.2.4.1. hCG. To understand hCG’s unexpected activation of taLHR(Fig. 2B), we compared residues that have been previously foundto be important for hCG binding, namely residues in LRRs 3 and6 (Bogerd et al., 2005; Bogerd, 2007), in hLHR and taLHR. Fig. 6Ashows multiple sequence alignment of all the GTHRs used in thisstudy. The LRRs are mapped on the structure of hFSHR taken fromthe hFSH/hFSHR–ECD complex structure (PDB 1XWD), and LRRs 3and 6 are enclosed in a green rectangle (Fig. 6B).
Analysis of the receptor LRRs known to be important for hCGbinding, namely Leu104, Asn107 and Gly183 in hCG numbering(Bogerd et al., 2005; Bogerd, 2007), did not reveal good candidatesfor explaining the activation of taLHR by hCG. Although both hLHRand taLHR have Asn at the position equivalent to Asn107 in hLHRfound to promote hCG binding, the rest of the positions do not fitwell. For example, at the site of hLHR Leu104, the taLHR hasSer107 which does not seem to give any particular advantage forbinding in this case. Moreover, at the site equivalent to hFSHRLys179 (the residue in hFSHR that determines lack of activationby hCG, probably due to charge repulsion between the two (Bo-gerd, 2007)), taLHR also has a positively charged residue(Arg186) that can cause repulsion.
The seatbelt region has been found important for GTH binding(Campbell et al., 1991), but comparison between hCG and taLHseatbelt regions also failed to provide any possible explanationfor the activation of taLHR by hCG. Therefore, it seems that addi-tional regions are responsible for taLHR activation by hCG. Thesemight involve the hinge region which links the ECD and GPCR do-main of GTHR and has been found to take part in TSH hormonebinding and activity (Mueller et al., 2010). Secondary structure pre-diction of the hinge region indicates a long a-helix for taLHR andhLHR that is not predicted for the rest of the sequences (hFSHRhas a helix but it is significantly shorter, see Supplementary FigureS2). We propose the possibility (which warrants further study) that
trFSHR
-2 -1 0 1 2 30
20
40
60
80
100
120
trLHtrFSHeLHeFSHtaLHtaFSHhCGhFSH
pFSH
A
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
trLHR
-2 -1 0 1 2 30
20
40
60
80
100
120
trLHtrFSHeLHeFSHtaLHtaFSHhCGhFSH
pFSH
log [GtH ng/ml]
B
% o
f m
axim
al r
esp
on
se
Fig. 3. Trout (tr) FSHR and LHR stimulate CRE-LUC activity. COS-7 cells weretransiently co-transfected with (A) trFSHR, (B) trLHR and the reporter plasmidpCRE-LUC. Cells were stimulated for 6 h with indicated hormones and luciferaseactivity was measured as represented in Fig. 1. Data are presented as mean ± SEM ofa representative experiment that was performed in triplicate.
eFSHR
-2 -1 0 1 2 30
20
40
60
80
100
120
eLHeFSH
trLHtrFSH
taLHtaFSHhCGhFSH
pFSH
A
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
eLHR
-2 -1 0 1 2 3 40
20
40
60
80
100
120hFSHhCGtaFSHtaLHeFSHeLHtrFSHtrLHpFSH
B
log [GtH ng/ml]
% o
f m
axim
al r
esp
on
se
Fig. 4. Eel (e) FSHR and LHR stimulate CRE-LUC activity. COS-7 cells weretransiently co-transfected with (A) eFSHR, (B) eLHR and the reporter plasmidpCRE-LUC. Cells were stimulated for 6 h with indicated hormones and luciferaseactivity was measured as represented in Fig. 1. Data are presented as mean ± SEM ofa representative experiment that was performed in triplicate.
94 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100

Author's personal copy
the putative helix in the hinge region is important for binding andactivation, and allows the activation of taLHR by hCG.
3.2.5. eLHR activators3.2.5.1. hFSH. To investigate hFSH activation of eLHR (cross-typeand cross-species activation, see Fig. 4B), all of the residues inthe receptor previously found (Bogerd, 2007) to be important forhFSH binding (see sequences of LRRs 1, 7 and 8 that are enclosedin a light blue rectangle in Fig. 6A and the surface patches in lightblue in Fig. 6B) were compared with their equivalent residues ineLHR.
Previous studies have shown that the size of hFSHR Leu55 isimportant for binding hFSH and that having a Tyr residue at thatposition (as it is in hLHR) disables binding of hFSH (Bogerd,2007). The eLHR has a His residue at the equivalent position. Ourmodels predict that the His residue in the eel receptor is still smallenough to allow binding of hFSH to the receptor (Fig. 7). Our mod-
els also showed the steric hindrance created by the Tyr (Tyr58) res-idue in the complex hFSH/hLHR (Fig. 7). Note that minimization ofthe models and/or changing rotamers of Tyr58 did not resolve thisincompatibility.
In addition, eLHR has an Arg residue at position 61 which isequivalent to hFSHR Arg52, found to also be involved in hFSH bind-ing, albeit to a lesser extent. These similarities provide testablehypotheses for molecular determinants responsible for the activa-tion of eLHR by hFSH. For example, we predict that mutation ofHis64 in eLHR to a Tyr residue will impair the activation of eLHRby hFSH, whereas mutation to Ala should allow binding of hFSHto eLHR and possibly eLHR activation.
3.2.5.2. trLH. An interesting case of cross-species activation of eLHRis that by trLH (Fig. 4B). Inspection of the multiple sequence align-ment of the receptors (Fig. 6) did not provide any obvious explana-tion for this activation. We thus looked for a possible explanation
Fig. 5. Sequence and structure comparisons of human, porcine and bovine FSH. (A) Sequence alignment of human, porcine and bovine FSH a and b subunits. The conservationof the residues is shown by: ‘⁄’ (identical residues in all sequences), ‘:’ (highly conserved column), ‘.’ (weakly conserved column). Interface residues according to the PDB1XWD structure are highlighted in yellow. Residues that differ between human and porcine or bovine sequences that have a different electrostatic charge are colored red. Thesignal peptide is in gray. (B) Better fit between the electrostatic potential on the interface of human FSH and human FSHR. The electrostatic potential on the surface of humanand bovine FSH and human FSHR is shown on the surface representation of the molecules. The electrostatic scale ranges from �3 (red) to 3 (blue) kT/E.
J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100 95

Author's personal copy
FSHR
FSHβ
FSHα
A
B
Fig. 6. LRRs involved in either hFSH or hCG binding. (A) Sequence alignment of GTHRs used in this work. The signal peptide is colored in gray. The repeated b-sheet (LRR)structures are indicated by colored rectangles and numbered according to human (h) FSHR. b segments that were found to be involved in hFSH and hCG responsiveness are inblue and green, respectively. Residues of interest are colored in the alignment text and numbered according to hFSHR. Note that tilapia (ta) FSHR has an insert after LRR1, andthere may therefore be a difference in the LRR numbering in this case. (B) Top view of the hFSH/hFSHR complex from structure 1XWD mapping the LRRs. The receptor is insurface representation in gray. LRRs that were found to be involved in hFSH or hCG binding are in blue and green, respectively. Other LRRs are in yellow. FSH is shown as aribbon: magenta for the a subunit and blue for the b subunit.(For interpretation of the references to colour in this figure legend, the reader is referred to the web version ofthis article.)
96 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100

Author's personal copy
in the sequences of the GTHs with an emphasis on the ‘‘seatbelt’’region. We noted that trLH and eLH are very similar in the seatbeltsequence (Fig. 8). In fact, with 13 identical residues and 3 addi-tional similar residues (out of a total 18 residues), these two se-quences were the closest among all GTH ‘‘seatbelt’’ sequencesused in this study. The high similarity between eLH and trLH inthe seatbelt area might explain activation of eLHR by trLH.
3.2.5.3. hCG. Cross-species activation of eLHR by hCG (Fig. 4B) canbe rationalized by the similarity of LRR residues important for hLH/hCG binding (Bogerd et al., 2005; Bogerd, 2007) (LRRs 3 and 6 high-lighted in green in Fig. 6): hLHR and eLHR have identical residuesat these positions: Leu104 vs. Leu110, Asn107 vs. Asn113 respec-tively. In addition, eLHR contains a Gly189 residue (Gly183 inhLHR) in this specificity-determining position. These similaritiesmay contribute to the ability of hCG to activate eLHR, and interest-
ingly may explain the known observation of a single injection ofhCG is sufficient to induce complete spermatogenesis from sper-matogonial proliferation through to spermiogenesis (Miura et al.,1991).
4. Discussion
Fish have undergone rapid molecular evolution (Albertson andKocher, 1999; Holland, 1999; Prince and Pickett, 2002; Kocher,2003), which makes them an excellent model for the study of spe-cies specificity of GTHs to their cognate and diverse receptors. Weused GTHs belonging to three teleost orders, from Anguilliformes,representative of less evolved teleosts, to Salmoniformes, and final-ly the relatively modern Perciformes. All of the recombinant GTHsused in this study were previously used in different biological as-says and were found to stimulate estradiol and 17a,20b-dihy-droxy-4-pregnen-3-one (DHP) secretion from carp ovaries.Moreover, steroid secretion differed according to the type of GTHand the ovarian developmental state. The recombinant gonadotro-pin from less-evolved fish (eel and trout) were more efficient ineliciting estradiol secretion, while in mature ovarian fragmentsthe DHP secretion was higher in response to recombinant LHs thanto recombinant FSHs (Aizen et al., 2012). Moreover, biologicalactivities of recombinant taLH and taFSH have been confirmed bysteroid secretion from tilapia gonads (Kasuto and Levavi-Sivan,2005; Aizen et al., 2007a; Levavi-Sivan et al., 2008), biologicalactivities of eFSH and eLH by in vitro oocyte maturation andin vivo spermatogenesis (Kobayashi et al., 2010), and biologicalactivities of trFSH and trLH by in vitro receptor activation, sex ste-roid production, and in vivo ovarian development and milt produc-tion (Ko et al., 2007).
Pharmacological characterization of piscine and human GTHRsrevealed that every ligand enhanced luciferase expression throughits own cognate receptor. We also found several cases of cross-spe-cies activation. Porcine and bovine FSHs activated hFSHR. hCG en-hanced luciferase expression not only through its own cognatereceptor but also through the tilapia and eel LHRs. This resultwas not surprising since hCG is broadly employed in spawninginduction trials of many fish species in culture today, including fishfrom different species and teleost orders (reviewed by (Zohar andMylonas, 2001)). eLHR had the broadest activation spectrum,responding not only to eLH but also to hCG and trLH.
Previous studies on the specificity of piscine GTHRs have re-ported that in catfish, zebrafish, coho salmon, Atlantic salmonand eel, LHRs are specific for their respective LHs, whereas theFSHRs, although showing a clear preference for FSH, respond toLH as well (Yan et al., 1992; Miwa et al., 1994; Kumar et al.,2001a; Vischer and Bogerd, 2003; Kwok et al., 2005; So et al.,2005; Kazeto et al., 2008; Andersson et al., 2009).
In contrast, in another Salmoniformes species, the amago sal-mon, when an extremely high GTH concentration (5 lg/ml) wasused, LHR was found to respond to both LH and FSH, while FSHRresponded only to FSH (Oba et al., 1999a, 1999b). However, in an-other study using GTHRs of the same species and recombinanttrout GTHs, both receptors were found to be ligand-selective (Koet al., 2007). In agreement with our findings, in eel, each recombi-nant GTH activated its own cognate receptor in a dose-dependentmanner (Kazeto et al., 2008). When using concentrations of 100and 200 ng/ml, LH succeeded in activating eFSHR 1.5-fold (Kazetoet al., 2008), and 1.2-fold in this study. However, the higher eLHconcentration failed to induce luciferase expression througheFSHR, suggesting that this effect is minor and not significant.Combination of our results with these studies, suggest the follow-ing general pattern: the larger but still limited functional selectiv-ity of the FSHRs for both GTHs might depend on the fish species or
Fig. 7. No steric hindrance in hFSH/eLHR complex. The figure shows complex ofhuman (h) FSH/hFSHR (top) and the minimized models of hFSH/eel (e) LHR(middle) and hFSH/hLHR (bottom). The hFSH and the receptors are in ribbonrepresentation with the receptors in gray and the a and b subunits of hFSH inmagenta and blue, respectively. bTyr121 is depicted in CPK representation as arethe receptor residues from LRR3 that interact with it. In the bottom panel, the areaof clash is circled in orange. The inserts to the left show the interacting residues assticks with the shortest distance between them. Note that in hFSH/hLHR, theshortest distance is less than the minimal distance of the H-bond (2.6 Å), and thereis a clash between hFSH L117 and hLHR Y58.
J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100 97

Author's personal copy
taxon, whereas teleost LHRs, like their mammalian counterparts,seem to be specific for their corresponding ligand.
An evaluation of the physiological consequences and bindingcharacteristics of GTHs to their receptors in fish may provide anexplanation for why evolutionary pressure generated, in some spe-cies, a LHR that is selective for LH, and a FSHR that can bind bothLH and FSH. Using specific ELISAs, it was found that plasma taFSHand taLH peak concomitantly during vitellogenesis and final oocytematuration (Aizen et al., 2007b). The high receptor–ligand specific-ity for taGTHs, as shown in the current study, is in agreement withthe physiology of their secretion. The simultaneous presence ofboth GTHs in the circulation during different stages of reproduc-tion may be an evolutionary force favoring selective GTHRs, sinceat each point in time the ovary has several generations of follicles,each at a different stage of development and probably requiring adifferent gonadotropin. In eel, on the other hand, the cross-reactiv-ity of eLHR exists both within the species and with human ligands.Interestingly, the eel is the most ancient and least evolved speciestested here. We propose that because it is the least evolved speciesits receptors may fit ligands of other more evolved species becauseit holds the base sequence of their receptors. However, this was notthe case for eFSHR which was not activated by any ligands otherthan the cognate FSH.
Structural modeling provides a powerful tool for analyzing andpredicting ligand–receptor binding and activation (Niv et al., 2006;Brockhoff et al., 2010; Yarnitzky et al., 2010). Here we used struc-tural modeling and sequence analysis to rationalize the molecularbasis of cross-type hormone specificity and cross-species activity.Previous studies have explored and identified regions in GTHsand their receptors that are important for binding selectivity, suchas the ‘‘seatbelt’’ region of the hGTHs (Campbell et al., 1991) andspecific LRRs and residues in hLHR and hFSHR (Bogerd et al.,
2005; Bogerd, 2007). Investigating these regions in the human, tila-pia, trout and eel GTHs and GTHRs resulted in several residues thatcould partially explain the observed activity profile. For example,our models could explain why hFSH activates eLHR but not hLHR,as they showed steric interference in the hFSH/hLHR complex dueto the size of the hLHR Tyr58 which does not fit the small cavitycreated by the hFSHbLeu117, bTyr121 and aArg66 residues, inaccordance with previous studies (Bogerd, 2007; Fan and Hend-rickson, 2005) while the eLHR His64 is still small enough to enablethe binding of hFSH.
We could not find a rationalization for hCG activation of taLHRby analyzing regions and residues in the GTH and GTHR–ECDwhich have been previously found to be important for GTH/GTHRbinding. We suspect that there are additional regions in the GTHRsthat may participate in binding and/or activation by the GTHs thatmay explain the activity profile seen for taLHR. One such candidateis the long hinge region that links the ECD to the GPCR domain ofthe receptors. Involvement of the hinge region in receptor activa-tion has been shown for TSH (Mueller et al., 2010). There is noavailable structure of any GTHR hinge region. In fact, most of it ispredicted to be unstructured. Further studies of this region, as wellas of Arg80 highlighted by multiple sequence analysis in the cur-rent study, are likely to shed more light on GTH interactions withtheir receptors.
Our analysis of the activity profile of bFSH and pFSH with hFSHRshowed that all of the established interfacial residues (as definedby and derived from the FSH/FSHR–ECD crystal structure) in hFSH,bFSH and pFSH were the identical, but a few residues near theinterface differed between the species, causing a change in theelectrostatic map of the GTHs. We suggest that this difference isat least partially responsible for the significant lowering of hFSHRactivation by bFSH and pFSH, similarly to the suggestion of
Fig. 8. Multiple sequence alignment of the hormones’ b subunits. The figure shows an alignment of GTH b subunits for the species used in this work. The Cys residues are inpink and numbered. The sequences between Cys10 and Cys12 (the seatbelt region) are enclosed in an orange rectangle. The signal peptide is in gray.
98 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100

Author's personal copy
Angelova et al. (2010) who examined the role of equivalent hGTHRinterface residues that bind to the a subunit of the GTH in the FSH/FSHR complex crystal structure and LH/LHR and TSH/TSHR models.
To the best of our knowledge, we present the first attempt topharmacolologyically and structurally analyze the interactions be-tween GTHs and GTHRs for tilapia, trout and eel. These studiesopen the venue for studying the interface residues in taFSHR andthe effect of exon duplication, which causes an insert in the LRRdomain, on the interaction and binding of taFSH. The finding thattaLHR and eLHR are activated by hCG and eLHR is activated alsoby hFSH is of great interest and applicability in the context of acti-vation of various GTHRs by their ligands and by ligands from othervertebrates.
Acknowledgments
We would like to thank Dr. Y. Kazeto (Division of Marine LifeSciences, Research Faculty of Fisheries Science, Hokkaido Univer-sity, Japan) for the sequence of the Japanese eel LH and FSH recep-tors, and to Dr. T. Gudermann (Walther-Straub-Institute ofPharmacology and Toxicology, Ludwig-Maximilians University ofMunich, Germany) for the hLHR. This research was partially fundedby United States-Israel Binational Science Foundation Grant2005096, by Niedersachsen-Israel research fund (Grant ZN2448).and by the Nutrigenomics and Functional Foods Research Centerat The Robert H. Smith Faculty of Agriculture, Food and Environ-ment, The Hebrew University of Jerusalem.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.mce.2012.08.013.
References
Aizen, J., Kasuto, H., Golan, M., Zakay, H., Levavi-Sivan, B., 2007a. Expression andcharacterization of biologically active recombinant tilapia FSH:immunohistochemistry, stimulation by GnRH and effect on steroid secretion.Biol. Reprod. 764, 692–700.
Aizen, J., Kasuto, H., Levavi-Sivan, B., 2007b. Development of specific enzyme-linkedimmunosorbent assay for determining LH and FSH levels in tilapia, usingrecombinant gonadotropins. Gen. Comp. Endocrinol. 153, 323–332.
Aizen, J., Kobayashi, M., Selicharova, I., Sohn, Y.C., Yoshizaki, G., Levavi-Sivan, B.,2012. Steroidogenic response of carp ovaries to piscine FSH and LH depends onthe reproductive phase. Gen. Comp. Endocrinol. 1781, 28–36.
Albertson, R.C., Kocher, T.D., 1999. Shape differences in the trophic apparatus of twoLake Malawi cichlid species and their hybrid progeny - A landmark-basedmorphometric approach. Am. Zool. 395, 115A-115A.
Andersson, E., Nijenhuis, W., Male, R., Swanson, P., Bogerd, J., Taranger, G.L., Schulz,R.W., 2009. Pharmacological characterization, localization and quantification ofexpression of gonadotropin receptors in Atlantic salmon (Salmo salar L.) ovaries.Gen. Comp. Endocrinol. 1633, 329–339.
Angelova, K., de Jonge, H., Granneman, J.C.M., Puett, D., Bogerd, J., 2010. Functionaldifferences of invariant and highly conserved residues in the extracellulardomain of the glycoprotein hormone receptors. J. Biol. Chem. 28545, 34813–34827.
Angelova, K., Fanelli, F., Puett, D., 2008. Contributions of intracellular loops 2 and 3of the lutropin receptor in gs coupling. Mol. Endocrinol. 221, 126–138.
Angelova, K., Felline, A., Lee, M., Patel, M., Puett, D., Fanelli, F., 2011. Conservedamino acids participate in the structure networks deputed to intramolecularcommunication in the lutropin receptor. Cell. Mol. Life Sci. 687.
Bhowmick, N., Huang, J.N., Puett, D., Isaacs, N.W., Lapthorn, A.J., 1996.Determination of residues important in hormone binding to the extracellulardomain of the luteinizing hormone chorionic gonadotropin receptor by site-directed mutagenesis and modeling. Mol. Endocrinol. 109, 1147–1159.
Biran, J., Ben-Dor, S., Levavi-Sivan, B., 2008. Molecular identification and functionalcharacterization of the kisspeptin/kisspeptin receptor system in lowervertebrates. Biol. Reprod. 794, 776–786.
Bogerd, J., 2007. Ligand-selective determinants in gonadotropin receptors. Mol. Cell.Endocrinol. 260, 144–152.
Bogerd, J., Granneman, J.C.M., Schulz, R.W., Vischer, H.F., 2005. Fish FSH receptorsbind LH: how to make the human FSH receptor to be more fishy? Gen. Comp.Endocrinol. 1421–2, 34–43.
Brandt, B.W., Feenstra, K.A., Heringa, J., 2010. Multi-Harmony: detecting functionalspecificity from sequence alignment. Nucleic Acids Res. 38, W35–W40.
Brockhoff, A., Behrens, M., Niv, M.Y., Meyerhof, W., 2010. Structural requirements ofbitter taste receptor activation. Proc. Natl. Acad. Sci. USA. 10724, 11110–11115.
Bryson, K., McGuffin, L.J., Marsden, R.L., Ward, J.J., Sodhi, J.S., Jones, D.T., 2005.Protein structure prediction servers at university college london. Nucleic AcidsRes. 33, W36–W38.
Caltabiano, G., Campillo, M., De Leener, A., Smits, G., Vassart, G., Costagliola, S.,Pardo, L., 2008. The specificity of binding of glycoprotein hormones to theirreceptors. Cell. Mol. Life Sci. 6516, 2484–2492.
Campbell, R.K., Deanemig, D.M., Moyle, W.R., 1991. Conversion of humanchoriogonadotropin into a follitropin by protein engineering. Proc. Natl. Acad.Sci. USA. 883, 760–764.
Chatterjee, A., Shen, S.T., Yu, J.Y.L., 2005. Molecular cloning of cDNAs and structuralmodel analysis of two gonadotropin beta-subunits of snakehead fish (Channamaculata). Gen. Comp. Endocrinol. 1433, 278–286.
Chauvigne, F., Tingaud-Sequeira, A., Agulleiro, M.J., Calusinska, M., Gomez, A., Finn,R.N., Cerda, J., 2010. Functional and evolutionary analysis of flatfishgonadotropin receptors reveals cladal- and lineage-level divergence of theteleost glycoprotein receptor family. Biol. Reprod. 826, 1088–1102.
Chopineau, M., Martinat, N., Gibrat, J.F., Galet, C., Lecompte, F., Foulon-Gauze, F.,Pourchet, C., Guillou, F., Combarnous, Y., 2004. Identification of amino-acids inthe a-subunit first and third loops that are crucial for the heterospecific follicle-stimulating hormone activity of equid luteinizing hormone/choriogonadotropin. Eur. J. Endocrinol. 1506, 877–884.
Costagliola, S., Urizar, E., Mendive, F., Vassart, G., 2005. Specificity and promiscuityof gonadotropin receptors. Reproduction 1303, 275–281.
De Leener, A., Caltabiano, G., Erkan, S., Idil, M., Vassart, G., Pardo, L., Costagliola, S.,2008. Identification of the first germline mutation in the extracellular domain ofthe follitroplin receptor responsible for spontaneous ovarian hyperstimulationsyndrome. Hum. Mutat. 291, 91–98.
Erbel, P.J.A., Karimi-Nejad, Y., De Beer, T., Boelens, R., Kamerling, J.P., Vliegenthart,J.F.G., 1999. Solution structure of the alpha-subunit of human chorionicgonadotropin. Eur. J. Biochem. 2602, 490–498.
Erbel, P.J.A., Karimi-Nejad, Y., van Kuik, J.A., Boelens, R., Kamerling, J.P., Vliegenthart,J.F.G., 2000. Effects of the N-linked glycans on the 3D structure of the free alpha-subunit of human chorionic gonadotropin. Biochemistry 3920, 6012–6021.
Fan, Q.R., Hendrickson, W.A., 2005. Structure of human follicle-stimulatinghormone in complex with its receptor. Nature 4337023, 269–277.
Fox, K.M., Dias, J.A., Van Roey, P., 2001. Three-dimensional structure of humanfollicle-stimulating hormone. Mol. Endocrinol. 153, 378–389.
Gasteiger, E., Gattiker, A., Hoogland, C., Ivanyi, I., Appel, R.D., Bairoch, A., 2003.ExPASy: the proteomics server for in-depth protein knowledge and analysis.Nucleic Acids Res. 3113, 3784–3788.
Gether, U., 2000. Uncovering molecular mechanisms involved in activation of Gprotein-coupled receptors. Endocr. Rev. 211, 90–113.
Holland, P.W.H., 1999. Gene duplication: past, present and future. Semin. Cell Dev.Biol. 105, 541–547.
Jones, D.T., 1999. Protein secondary structure prediction based on position-specificscoring matrices. J. Mol. Biol. 2922, 195–202.
Kalinina, O.V., Mironov, A.A., Gelfand, M.S., Rakhmaninova, A.B., 2004. Automatedselection of positions determining functional specificity of proteins bycomparative analysis of orthologous groups in protein families. Protein Sci.132, 443–456.
Karges, B., Gidenne, S., Aumas, C., Haddad, F., Kelly, P.A., Milgrom, E., de Roux, N.,2005. Zero-length cross-linking reveals that tight interactions between theextracellular and transmembrane domains of the luteinizing hormone receptorpersist during receptor activation. J. Mol. Endocrinol. 198, 2086–2098.
Kasuto, H., Levavi-Sivan, B., 2005. Production of biologically active tethered tilapiaLHba by the methylotrophic yeast Pichia pastoris. Gen. Comp. Endocrinol. 1403,222–232.
Kazeto, Y., Kohara, M., Miura, T., Miura, C., Yamaguchi, S., Trant, J.M., Adachi, S.,Yamauchi, K., 2008. Japanese eel follicle-stimulating hormone (FSH) andluteinizing hormone (LH): production of biologically active recombinant FSHand LH by Drosophila S2 cells and their differential actions on the reproductivebiology. Biol. Reprod. 795, 938–946.
Kleinau, G., Brehm, M., Wiedemann, U., Labudde, D., Leser, U., Krause, G., 2007.Implications for molecular mechanisms of glycoprotein hormone receptorsusing a new sequence-structure-function analysis resource. Mol. Endocrinol.212, 574–580.
Ko, H., Park, W., Kim, D.-J., Kobayashi, M., Sohn, Y.C., 2007. Biological activities ofrecombinant Manchurian trout FSH and LH: their receptor specificity,steroidogenic and vitellogenic potencies. J. Mol. Endocrinol. 381, 99–111.
Kobayashi, M., Hayakawa, Y., Park, W., Banba, A., Yoshizaki, G., Kumamaru, K.,Kagawa, H., Kaki, H., Nagaya, H., Sohn, Y.C., 2010. Production of recombinantJapanese eel gonadotropins by baculovirus in silkworm larvae. Gen. Comp.Endocrinol. 1673, 379–386.
Kobayashi, M., Morita, T., Ikeguchi, K., Yoshizaki, G., Suzuki, T., Watabe, S., 2006. Invivo biological activity of recombinant goldfish gonadotropins produced bybaculovirus in silkwonn larvae. Aquaculture 2561–4, 433–442.
Kocher, T.D., 2003. Evolutionary biology: fractious phylogenies. Nature 423, 489–491.
Kreuchwig, A., Kleinau, G., Kreuchwig, F., Worth, C.L., Krause, G., 2011. Researchresource. Update and extension of a glycoprotein hormone receptors webapplication. Mol. Endocrinol. 254, 707–712.
Kumar, S., Ijiri, S., Trant, J.M., 2001a. Molecular biology of the channel catfishgonadotropin receptors: 2. Complementary DNA cloning, functional expression,
J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100 99

Author's personal copy
and seasonal gene expression of the follicle-stimulating hormone receptor. Biol.Reprod. 653, 710–717.
Kumar, S., Ijiri, S., Trant, J.M., 2001b. Molecular biology of channel catfishgonadotropin receptors: 1. Cloning of a functional luteinizing hormonereceptor and preovulatory induction of gene expression. Biol. Reprod. 643,1010–1018.
Kwok, H.-F., So, W.-K., Wang, Y., Ge, W., 2005. Zebrafish gonadotropins and theirreceptors: I. cloning and characterization of zebrafish follicle-stimulatinghormone and luteinizing hormone receptors- evidence for their distinctfunctions in follicle development. Biol. Reprod. 726, 1370–1381.
Lapthorn, A.J., Harris, D.C., Littlejohn, A., Lustbader, J.W., Canfield, R.E., Machin, K.J.,Morgan, F.J., Isaacs, N.W., 1994. Crystal structure of human chorionic-gonadotropin. Nature 369, 455–461.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J.,Higgins, D.G., 2007. Clustal W and Clustal X version 2.0. Bioinformatics 2321,2947–2948.
Leung, M.Y.K., Steinbach, P.J., Bear, D., Baxendale, V., Fechner, P.Y., Rennert, O.M.,Chan, W.Y., 2006. Biological effect of a novel mutation in the third leucine-richrepeat of human luteinizing hormone receptor. Mol. Endocrinol. 2010, 2493–2503.
Levavi-Sivan, B., Aizen, J., Avitan, A., 2005. Cloning, characterization and expressionof the D2 dopamine receptor from the tilapia pituitary. Mol. Cell. Endocrinol.2361–2, 17–30.
Levavi-Sivan, B., Bogerd, J., Mananos, E.L., Gomez, A., Lareyre, J.J., 2010. Perspectiveson fish gonadotropins and their receptors. Gen. Comp. Endocrinol. 1653, 412–437.
Levavi-Sivan, B., Golan, M., Aizen, J., Elizur, A., 2008. Fish recombinantgonadotropins. CYBIUM Int. J. Ichthyol 32, 17–21.
Liu, C., Dias, J.A., 1996. Long loop residues 33–58 in the human glycoproteinhormone common alpha subunit contain structural components for subunitheterodimerization and human follitropin-receptor binding. Arch. Biochem.Biophys. 3291, 127–135.
Miguel, R.N., Sanders, J., Chirgadze, D.Y., Blundell, T.L., Furmaniak, J., Smith, B.R.,2008. FSH and TSH binding to their respective receptors: similarities,differences and implication for glycoprotein hormone specificity. J. Mol.Endocrinol. 413–4, 145–164.
Miura, T., Yamauchi, K., Takahashi, H., Nagahama, Y., 1991. Human chorionic-gonadotropin induces all stages of spermatogenesis in vitro in the maleJapanese eel (Anguilla japonica). Dev. Biol. 1461, 258–262.
Miwa, S., Yan, L.G., Swanson, P., 1994. Localization of two gonadotropin receptors inthe salmon gonad by in vitro ligand autoradiography. Biol. Reprod. 503, 629–642.
Momany, F.A., Rone, R., 1992. Validation of the general-purpose quanta(R) 3.2 /charmm(R) force-field. J. Comput. Chem. 13, 888–900.
Mueller, S., Jaeschke, H., Gunther, R., Paschke, R., 2010. The hinge region: animportant receptor component for GPHR function. Trends Endocrinol. Metab.212, 111–122.
Myers, E.W., Miller, W., 1988. Optimal alignments in linear space. Comput. Appl.Biosci. 41, 11–17.
Niv, M.Y., Skrabanek, L., Filizola, M., Weinstein, H., 2006. Modeling activated statesof GPCRs: the rhodopsin template. J. Comput. Aided Mol. Des. 207–8, 437–448.
Oba, Y., Hirai, T., Yoshiura, Y., Yoshikuni, M., Kawauchi, H., Nagahama, Y., 1999a.Cloning, functional characterization, and expression of a gonadotropin receptorcDNA in the ovary and testis of amago salmon (Oncorhynchus rhodurus).Biochem. Biophys. Res. Commun. 2632, 584–590.
Oba, Y., Hirai, T., Yoshiura, Y., Yoshikuni, M., Kawauchi, H., Nagahama, Y., 1999b. Theduality of fish gonadotropin receptors: cloning and functional characterizationof a second gonadotropin receptor cDNA expressed in the ovary and testis ofamago salmon (Oncorhynchus rhodurus). Biochem. Biophys. Res. Commun. 2652,366–371.
Pei, J., Kim, B.H., Grishin, N.V., 2008. PROMALS3D: a tool for multiple proteinsequence and structure alignments. Nucleic Acids Res. 367, 2295–2300.
Pierce, J.G., Parsons, T.F., 1981. Glycoprotein hormones: structure and function.Annu. Rev. Biochem. 50, 465–495.
Prince, V.E., Pickett, F.B., 2002. Splitting pairs: the diverging fates of duplicatedgenes. Nat. Rev. Genet. 311, 827–837.
Puett, D., Angelova, K., da Costa, M.R., Warrenfeltz, S.W., Fanelli, F., 2010. Theluteinizing hormone receptor: insights into structure–function relationships
and hormone-receptor-mediated changes in gene expression in ovarian cancercells. Mol. Cell. Endocrinol. 3291–2, 47–55.
Puett, D., Li, Y., DeMars, G., Angelova, K., Fanelli, F., 2007. A functionaltransmembrane complex: the luteinizing hormone receptor with boundligand and G protein. Mol. Cell. Endocrinol. 260, 126–136.
Rice, P., Longden, I., Bleasby, A., 2000. EMBOSS: the European molecular biologyopen software suite. Trends Genet. 166, 276–277.
Rocha, A., Gomez, A., Zanuy, S., Cerda-Reverter, J.M., Carrillo, M., 2007. Molecularcharacterization of two sea bass gonadotropin receptors: cDNA cloning,expression analysis, and functional activity. Mol. Cell. Endocrinol. 2721–2,63–76.
Roy, A., Kucukural, A., Zhang, Y., 2010. I-TASSER: a unified platform for automatedprotein structure and function prediction. Nat. Protoc. 54, 725–738.
Royer, J., Lefevre-Minisini, A., Caltabiano, G., Lacombe, T., Malthiery, Y., Savagner, F.,Pardo, L., Rodien, P., 2008. The cloned equine thyrotropin receptor ishypersensitive to human chorionic gonadotropin; identification of threeresidues in the extracellular domain involved in ligand specificity.Endocrinology 14910, 5088–5096.
Sambroni, E., Le Gac, F., Breton, B., Lareyre, J.J., 2007. Functional specificity of therainbow trout (Oncorhynchus mykiss) gonadotropin receptors as assayed in amammalian cell line. J. Endocrinol. 1952, 213–228.
Sanders, J., Chirgadze, D.Y., Sanders, P., Baker, S., Sullivan, A., Bhardwaja, A., Bolton,J., Reeve, M., Nakatake, N., Evans, M., Richards, T., Powell, M., Miguel, R.N.,Blundell, T.L., Furmaniak, J., Smith, B.R., 2007. Crystal structure of the TSHreceptor in complex with a thyroid-stimulating autoantibody. Thyroid 175,395–410.
Shen, H.B., Chou, K.C., 2007. Signal-3L: A 3-layer approach for predicting signalpeptides. Biochem. Biophys. Res. Commun. 363, 297–303.
Shen, S.T., Cheng, Y.S., Shen, T.Y., Yu, J.Y.L., 2006. Molecular cloning of follicle-stimulating hormone (FSH) b subunit cDNA from duck pituitary. Gen. Comp.Endocrinol. 1483, 388–394.
Sitkoff, D., Sharp, K.A., Honig, B., 1994. Accurate calculation of hydration free-energies using macroscopic solvent models. J. Phys. Chem. 987, 1978–1988.
So, W.K., Kwok, H.F., Ge, W., 2005. Zebrafish gonadotropins and their receptors: II.Cloning and characterization of zebrafish follicle-stimulating hormone andluteinizing hormone subunits–their spatial-temporal expression patterns andreceptor specificity. Biol. Reprod. 726, 1382–1396.
Spinelli, S., Frenken, L., Bourgeois, D., deRon, L., Bos, W., Verrips, T., Anguille, C.,Cambillau, C., Tegoni, M., 1996. The crystal structure of a llama heavy chainvariable domain. Nat. Struct. Biol. 39, 752–757.
Tegoni, M., Spinelli, S., Verhoeyen, M., Davis, P., Cambillau, C., 1999. Crystalstructure of a ternary complex between human chorionic gonadotropin (hCG)and two Fv fragments specific for the a and b-subunits. J. Mol. Biol. 2895, 1375–1385.
Van Durme, J., Horn, F., Costagliola, S., Vriend, G., Vassart, G., 2006. GRIS:glycoprotein-hormone receptor information system. Mol. Endocrinol. 209,2247–2255.
Vassart, G., Pardo, L., Costagliola, S., 2004. A molecular dissection of the glycoproteinhormone receptors. Trends Biochem. Sci. 293, 119–126.
Vischer, H.F., Bogerd, J., 2003. Cloning and functional characterization of a gonadalluteinizing hormone receptor complementary DNA from the African catfish(Clarias gariepinus). Biol. Reprod. 681, 262–271.
Wu, C.H., Apweiler, R., Bairoch, A., Natale, D.A., Barker, W.C., Boeckmann, B., Ferro,S., Gasteiger, E., Huang, H., Lopez, R., Magrane, M., Martin, M.J., Mazumder, R.,O’Donovan, C., Redaschi, N., Suzek, B., 2006. The universal protein resource(UniProt): an expanding universe of protein information. Nucleic Acids Res. 34,D187–191.
Wu, H., Lustbader, J.W., Liu, Y., Canfield, R.E., Hendrickson, W.A., 1994. Structure ofhuman chorionic-gonadotropin at 2.6 angstrom resolution from MAD analysisof the selenomethionyl protein. Structure 26, 545–558.
Yan, L., Swanson, P., Dickhoff, W.W., 1992. A two-receptor model for salmongonadotropins (GTH I and GTH II). Biol. Reprod. 473, 418–427.
Yarnitzky, T., Levit, A., Niv, M.Y., 2010. Homology modeling of G-protein-coupledreceptors with X-ray structures on the rise. Curr. Opin. Drug Discov. Devel. 133,317–325.
Zhang, Y., 2007. Template-based modeling and free modeling by I-TASSER in CASP7.Proteins 69, 108–117.
Zohar, Y., Mylonas, C.C., 2001. Endocrine manipulations of spawning in culturedfish: from hormones to genes. Aquaculture 1971–4, 99–136.
100 J. Aizen et al. / Molecular and Cellular Endocrinology 364 (2012) 89–100
Related Documents











