MOLECULAR AND CELLULAR BIOLOGY, Feb. 1988, p. 595-604 Vol. 8, No. 2 0270-7306/88/020595-10$02.00/0 Copyright © 1988, American Society for Microbiology Expansions and Contractions of the Genetic Map Relative to the Physical Map of Yeast Chromosome III LORRAINE S. SYMINGTON AND THOMAS D. PETES* Department of Molecular Genetics and Cell Biology, University of Chicago, 920 East 58th Street, Chicago, Illinois 60637 Received 10 August 1987/Accepted 3 November 1987 To examine the relationship between genetic and physical chromosome maps, we constructed a diploid strain of the yeast Saccharomyces cerevisiae heterozygous for 12 restriction site mutations within a 23-kilobase (5-centimorgan) interval of chromosome m. Crossovers were not uniformly distributed along the chromosome, one interval containing significantly more and one interval significantly fewer crossovers than expected. One-third of these crossovers occurred within 6 kilobases of the centromere. Approximately half of the exchanges were associated with gene conversion events. The minimum length of gene conversion tracts varied from 4 base pairs to more than 12 kilobases, and these tracts were nonuniformly distributed along the chromosome. We conclude that the chromosomal sequence or structure has a dramatic effect on meiotic recombination. In general, one expects to find that the frequency of recombination between two genes should be proportional to the physical distance between the two markers. Deviations from this expectation have been observed, however, and are usually attributed to one of two factors: the proximity of chromosomal structural elements (such as the centromere) to the genetic interval, or the presence (or absence) of sequence-specific "hotspots" for recombination within the genetic interval. The clearest example of an effect of chromosome structure on recombination of adjacent sequences is the effect of the centromere on meiotic exchange in Drosophila melano- gaster. The genetic map is contracted relative to the physical map at the centromere (6). Several experiments have indi- cated the possibility of a similar centromeric repression of meiotic exchange in fungi. First, in Aspergillus nidulans and Saccharomyces cerevisiae, there is relatively more mitotic than meiotic recombination near the centromere (23, 31). The second type of experiment concerned the physical analysis of cloned centromeric sequences of S. cerevisiae (3, 8, 13). Two short conserved sequences (8 and 25 base pairs [bp]) flanking an A+T-rich spacer of 70 to 100 bp are sufficient for centromere function. In some cases, the phys- ical distance between a cloned centromere and a selectable gene was known, and therefore a correlation between ge- netic and physical distance near the centromere could be established. Although the ratio between centimorgans (cM) and kilobases (kb) for some of these centromeric regions was lower than the average for the genome (37), for others the ratio was about the same. A third type of experiment involved moving the centromere of yeast chromosome III to a new position within the same chromosome. In such yeast strains, Lambie and Roeder (20) found that meiotic recom- bination increased in an interval near the original position of the centromere, whereas recombination in the interval into which the centromere had been introduced decreased. In addition to effects of structural elements on recombi- nation, there are sites that appear to stimulate homologous recombination locally (reviewed by Whitehouse (41]). For example, the 8-bp Chi site (36) enhances homologous recom- * Corresponding author. bination in Escherichia coli by acting as a recognition sequence for the RecBC enzyme (32). In fungi, a number of sites (cog in Neurospora, M26 in Schizosaccharomyces pombe, and YS17 in Sordaria brevicolis) have been identi- fied that stimulate meiotic recombination (1, 12, 22). In addition, there appear to be regions of the yeast chromo- some that are unusually hot (4) or cold (21) for meiotic crossovers. The mechanism of action of these sites has not yet been determined. Recombination events in eucaryotes may be either recip- rocal or nonreciprocal (gene conversions). In certain orga- nisms, meiotic gene conversion events can be detected at a single heterozygous locus (A representing one allele and a representing the other) as a departure from 2A:2a segrega- tion, resulting in either 3A.la or 1A:3a tetrads. Gene con- version events therefore represent the nonreciprocal transfer of information between homologous genes. Since conversion events are frequently associated with reciprocal exchange of flanking markers (16), most current models of recombihation invoke the conversion event as an intermediate in the process of forming a reciprocal exchange (15, 25, 39). The meiotic conversion frequencies for different genes and different alleles vary considerably in a single organism (9, 10, 41). Some of this variation may be the result of the location of the allele in relation to recombination-stimulating sites, such as those described above. Within a single gene, the frequency of gene conversion often varies as a function of the distance from one end of the gene. This polarity is usually explained by postulating the presence of an initiating site for gene conversion near one end of the gene. Recent studies in S. cerevisiae with the rRNA gene (19) suggest an association between the initiation of transcription and mi- totic recombination. Estimates of the amount of DNA transferred during a meiotic gene conversion event in S. cerevisiae are largely based on coconversion frequencies. Both Fogel et al. (9, 10) and S. R. Judd (personal communication) concluded that conversion tracts were usually several hundred base pairs in length. In addition, Fogel et al. found that meiotic conver- sion tracts were usually continuous. Although the results of most other studies in fungal systems are consistent with the conclusion that meiotic conversion tracts are generally less 595

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR AND CELLULAR BIOLOGY, Feb. 1988, p. 595-604 Vol. 8, No. 20270-7306/88/020595-10$02.00/0Copyright © 1988, American Society for Microbiology
Expansions and Contractions of the Genetic Map Relative to thePhysical Map of Yeast Chromosome III
LORRAINE S. SYMINGTON AND THOMAS D. PETES*Department of Molecular Genetics and Cell Biology, University of Chicago, 920 East 58th Street, Chicago, Illinois 60637
Received 10 August 1987/Accepted 3 November 1987
To examine the relationship between genetic and physical chromosome maps, we constructed a diploid strainof the yeast Saccharomyces cerevisiae heterozygous for 12 restriction site mutations within a 23-kilobase(5-centimorgan) interval ofchromosome m. Crossovers were not uniformly distributed along the chromosome,one interval containing significantly more and one interval significantly fewer crossovers than expected.One-third of these crossovers occurred within 6 kilobases of the centromere. Approximately half of theexchanges were associated with gene conversion events. The minimum length of gene conversion tracts variedfrom 4 base pairs to more than 12 kilobases, and these tracts were nonuniformly distributed along thechromosome. We conclude that the chromosomal sequence or structure has a dramatic effect on meioticrecombination.
In general, one expects to find that the frequency ofrecombination between two genes should be proportional tothe physical distance between the two markers. Deviationsfrom this expectation have been observed, however, and areusually attributed to one of two factors: the proximity ofchromosomal structural elements (such as the centromere)to the genetic interval, or the presence (or absence) ofsequence-specific "hotspots" for recombination within thegenetic interval.The clearest example of an effect ofchromosome structure
on recombination of adjacent sequences is the effect of thecentromere on meiotic exchange in Drosophila melano-gaster. The genetic map is contracted relative to the physicalmap at the centromere (6). Several experiments have indi-cated the possibility of a similar centromeric repression ofmeiotic exchange in fungi. First, in Aspergillus nidulans andSaccharomyces cerevisiae, there is relatively more mitoticthan meiotic recombination near the centromere (23, 31).The second type of experiment concerned the physicalanalysis of cloned centromeric sequences of S. cerevisiae (3,8, 13). Two short conserved sequences (8 and 25 base pairs[bp]) flanking an A+T-rich spacer of 70 to 100 bp aresufficient for centromere function. In some cases, the phys-ical distance between a cloned centromere and a selectablegene was known, and therefore a correlation between ge-netic and physical distance near the centromere could beestablished. Although the ratio between centimorgans (cM)and kilobases (kb) for some of these centromeric regions waslower than the average for the genome (37), for others theratio was about the same. A third type of experimentinvolved moving the centromere of yeast chromosome III toa new position within the same chromosome. In such yeaststrains, Lambie and Roeder (20) found that meiotic recom-bination increased in an interval near the original position ofthe centromere, whereas recombination in the interval intowhich the centromere had been introduced decreased.
In addition to effects of structural elements on recombi-nation, there are sites that appear to stimulate homologousrecombination locally (reviewed by Whitehouse (41]). Forexample, the 8-bp Chi site (36) enhances homologous recom-
* Corresponding author.
bination in Escherichia coli by acting as a recognitionsequence for the RecBC enzyme (32). In fungi, a number ofsites (cog in Neurospora, M26 in Schizosaccharomycespombe, and YS17 in Sordaria brevicolis) have been identi-fied that stimulate meiotic recombination (1, 12, 22). Inaddition, there appear to be regions of the yeast chromo-some that are unusually hot (4) or cold (21) for meioticcrossovers. The mechanism of action of these sites has notyet been determined.Recombination events in eucaryotes may be either recip-
rocal or nonreciprocal (gene conversions). In certain orga-nisms, meiotic gene conversion events can be detected at asingle heterozygous locus (A representing one allele and arepresenting the other) as a departure from 2A:2a segrega-tion, resulting in either 3A.la or 1A:3a tetrads. Gene con-version events therefore represent the nonreciprocal transferof information between homologous genes. Since conversionevents are frequently associated with reciprocal exchange offlanking markers (16), most current models of recombihationinvoke the conversion event as an intermediate in theprocess of forming a reciprocal exchange (15, 25, 39).The meiotic conversion frequencies for different genes and
different alleles vary considerably in a single organism (9, 10,41). Some of this variation may be the result of the locationof the allele in relation to recombination-stimulating sites,such as those described above. Within a single gene, thefrequency of gene conversion often varies as a function ofthe distance from one end of the gene. This polarity isusually explained by postulating the presence of an initiatingsite for gene conversion near one end of the gene. Recentstudies in S. cerevisiae with the rRNA gene (19) suggest anassociation between the initiation of transcription and mi-totic recombination.
Estimates of the amount of DNA transferred during ameiotic gene conversion event in S. cerevisiae are largelybased on coconversion frequencies. Both Fogel et al. (9, 10)and S. R. Judd (personal communication) concluded thatconversion tracts were usually several hundred base pairs inlength. In addition, Fogel et al. found that meiotic conver-sion tracts were usually continuous. Although the results ofmost other studies in fungal systems are consistent with theconclusion that meiotic conversion tracts are generally less
595

596 SYMINGTON AND PETES
than 1 kb in length (summarized by Orr-Weaver and Szostak[29]), DiCaprio and Hastings (5) and Fogel et al. (11)reported occasional coconversion of markers in differentgenes.
In most previous studies on meiotic recombination, thenumber of heterozygous markers within one chromosomalregion is small and the markers represent mutations inexpressed genes. In addition, the amount of DNA sequenceheterogeneity between the interacting chromosomes otherthan the heterozygous genetic markers themselves is un-known. As described below, we used recombinant DNAprocedures to construct a diploid strain that was heterozy-gous for 12 restriction site mutations within a 23-kb, 5-cMregion of chromosome III. These mutations were con-structed in vitro in cloned fragments of DNA, which werethen reinserted into a diploid yeast strain. Thus, the physicallocation and nature of these markers were precisely known.We identified tetrads with a crossover in this genetic intervaland used Southern analysis to characterize the position ofthe exchange; a similar approach has been used by Borts andHaber (personal communication) to investigate meiotic ex-changes within pBR322 sequences integrated in the yeastchromosome between duplicated mating type loci. We foundthat the frequency of both crossovers and gene conversionevents was different in different regions of the chromosome.
MATERIALS AND METHODS
Media and growth conditions. Media for yeast growthwere prepared as described by Sherman et al. (34). YPDmedium (1% yeast extract, 2% dextrose, 2% Bacto-peptone[Difco Laboratories], and 3% agar for plates) was used fornonselective growth. Nutritional markers were scored onsynthetic medium lacking one amino acid (2% dextrose,1.7% yeast nitrogen base [Difco], 0.5% ammonium sulfate,3% agar). Medium containing 5-fluoro-orotate (5-FOA) wasused to select ura3 mutant strains and was made by supple-menting synthetic complete medium to 0.5 mM uracil and0.88% 5-FOA (2). For sporulation, diploid cells were grownto log phase in YPA (1% yeast extract, 2% Bacto-peptone,
1% potassium acetate) and then transferred to SM (2%potassium acetate plus any required amino acids). Yeaststrains were grown vegetatively at 32°C and sporulated atroom temperature.
E. coli strains were grown at 37°C in LB medium (1%Bacto-tryptone [Difco], 0.5% yeast extract, 0.5% NaCl plus1.5% agar for plates). For growth of plasmid-containingstrains, the medium was supplemented with 50 ,ug of ampi-cillin per ml.
Strains. A complete list of yeast strains is given in Table 1.Most of the strains were derived from XJ24-24a (37); thisstrain was also used as a source ofDNA for construction ofthe plasmids used in our study (28). The strain LS1 was aspontaneous ura3 mutant strain isolated from XJ24-24a bygrowth on 5-FOA plates. The haploid strain LS1 was madeinto a diploid (LS2) by using a plasmid (YpC50-HO, pro-vided by I. Herskowitz) that contained a cloned gene thatcatalyzed mating type switching. Since the LS2 strain spor-ulated poorly, we obtained a spore from this cross (LS2-la)and crossed it to a haploid strain (SJR14) derived from adifferent genetic background. The resulting diploid (LS3)sporulated well. We isolated a spore (LS3-17c) from a tetradderived from this diploid in which no crossing-over hadoccurred between LEU2 and the centromere; such tetradscould be identified because they showed first-division segre-gation relative to the heterozygous centromere-linked TRPJgene (26). Thus, strain LS3-17c should contain the samesequences between LEU2 and the centromere as XJ24-24a.Most of the other strains in the study were constructed by
transformation. The haploid LS18 was derived from LS2-laby transformation with a BamHI restriction fragment de-rived from plasmid pLS48 (Table 2). This plasmid contains10 different filled-in restriction sites in two fused BamHIfragments (C2G and D8B, Fig. 1) derived from chromosomeIII; in addition, the fragment contains an insertion of thegene URA3, which was used to select transformants. Toensure that the sequences on the transforming fragmentreplaced the wild-type sequences (33), we analyzed thetransformed strain by Southern analysis. The haploid LS20was derived by transformation of LS2-la with a BamHI
TABLE 1. Yeast strainsRelevant genotype or description (reference)Strain
XJ24-24a .... a trpl-J arg4-17 tyr7-1 ade6 MAL2 (37)LS1 .... a trpl-J arg4-17 tyr7-1 ade6 MAL2 ura3LS2.... a trpl-larg4-17 tvr7-1 ade6 MAL2 ura3
a trpl-J arg4-17 tyr7-1 ade6 MAL2 ura3LS2-la ...... a trpl-J arg4-17 tyr7-1 ade6 MAL2 ura3SJR14 .... a leu2-3,112 his3AJ ura3-52 can]LS3.... Diploid formed by crossing SJR14 with LS2-laLS3-17c.... a ura3 canlLS18.... LS2-la transformed to Ura+ with BamHI fragment of pLS48a; contains restriction site changes in C2G and D8B
fragmentsLS20.... LS2-la transformed to Ura+ with BamHI fragment of pLS37a; contains restriction site changes in G4B fragmentLS24.... Diploid formed by crossing LS3-17c with LS20LS25....U..ra- derivative of LS24LS25-70d .... a Ieu2 ade6 ura3 can]; contains G4B site changesLS36.... Diploid formed by crossing LS18 with LS25-70d; heterozygous for restriction site changes in fragments C2G, D8B
and G4BLS42.... Ura- derivative of LS36LS45.... Derivative of LS42 that lacks mutant ura3 insertion in TylbLS46.... Ura+ diploid derived from LS45 by transformation with plasmid pLS55a; contains fill-in of ClaI site opposite of
centromere from other changesLS47.... Ura- derivative of LS46; contains changes in CG2, D8B, and G4B fragments in addition to ClaI change described
for LS46a Plasmids are described in Table 2.b Other details concerning the construction of this strain are given in the text.
MOL. CELL. BIOL.

CLUSTERED MEIOTIC EXCHANGES IN S. CEREVISIAE 597
TABLE 2. Recombinant plasmidsDescriptiona (reference)Plasmid
G4B.. 7.2-kb BamHI fragment containing the LEU2 gene from chromosome III inserted into the BamHI site of YIp5 (28)D8B.. 10.5-kb BamHI fragment (located centromere-proximal to G4B) inserted into BamHI site of YIp5 (28)C2G.. 8.6-kb BamHI fragment containing centromere of chromosome III inserted into BamHI site of YIp5 (28)A2C.. 3.5-kb BamHI fragment contiguous with C2G inserted into BamHI site of YIp5 (28)pLS31.. XhoI- (PvuI+) derivative of G4BpLS32.. BstEII- (MaeII+) derivative of pLS31pLS33.. Bcll- (ClaI+) derivative of pLS32pLS34.. Sal- (PvuI+) derivative of pLS33pLS35.. 1.1-kb HindlIl fragment containing URA3 gene inserted in HindlIl site of pLS33 within the Ty elementpLS36.. 1.1-kb BamHI fragment containing the URA3 gene (17) inserted into BclI site of pLS32pLS37.. 1.1-kb Hindlll fragment containing the URA3 gene inserted into the HindIII site of pLS32 within the Ty elementpLS38.. Asp718- (SnaBI+) derivative of pLS37pLS41.. Xhol- (PvuI+) derivative of D8BpLS42.. Spel- (AluI+) derivative of pLS41pLS43.. BglIIl- (ClaI+) derivative of pLS42pLS44.. BglII2- (ClaI+) derivative of pLS43pLS45.. BglII3- (ClaI+) derivative of pLS44pLS46.. Insertion of 8.6-kb C2G BamHI fragment from pLS54 into one of BamHI sites of pLS45 so that the orientation of
the fragments is the same as in chromosome IIIpLS47.. BamHI- derivative of pLS46; the site deleted was between the C2G and D8B fragmentspLS48.. 1.1-kb Hindlll fragment containing URA3 gene inserted into HindlIl site within yeast DNA insert of pLS47pLS51.. MluI- (BssHII+) derivative of C2GpLS52.. BglII4- (ClaI+) derivative of pLS51pLS53.. Bcll- (ClaI+) derivative of PLS52pLS54.. Xbal- (AluI+) derivative of pLS53pLS55.. Clal- (NruI+) derivative of A2Ca Restriction that were deleted by filling in the single-strand gap are indicated with a - superscript; the new sites formed by the fill-in are indicated by a +
superscript.
fragment derived from plasmid pLS37 (Table 2). This frag-ment contains three different filled-in restriction sites de-rived from the BamHI fragment G4B and a wild-type URA3gene (Fig. 1). The diploid LS24 was constructed by crossingLS3-17c with LS20. A Ura- derivative of LS24 (LS25) wasisolated on 5-FOA plates; this derivative retained a mutantinsertion of ura3 within the G4B BamHI fragment. A sporederived from LS25 (LS25-70d) that contained the filled-inrestriction sites in the G4B fragment was crossed to LS18 tocreate a diploid strain (LS36) that was heterozygous for all12 restriction site polymorphisms. A Ura- derivative ofLS36 was isolated on 5-FOA plates; this derivative (LS42)was shown to lack the URA3+ insertion derived from theLS18 parental strain (presumably as the result of a mitoticgene conversion event involving the homologous chromo-some), although the mutant ura3 insertion derived fromLS25-70d was retained. To remove this mutant insertion, wetransformed LS42 with a wild-type URA3 gene (derivedfrom the plasmid MB1068 [17]), and then a Ura- derivative(LS45) of the strain was selected. To insert an additionalrestriction site polymorphism in LS45 on the other side ofthe centromere, we used the two-step transplacement tech-nique (42), first transforming LS45 with plasmid pLS55. Thisplasmid contains a filled-in restriction site in a BamHIfragment from the other side of the centromere as well as awild-type URA3 gene. The resulting transformed diploid(LS46) was plated on 5-FOA medium to isolate a diploidstrain (LS47) that retained all restriction site polymorphismsbut had lost the URA3 insertion. The arrangement of therestriction site polymorphisms for the diploid strains LS42,LS45, and LS47 is shown in Fig. 2.When we attempted to fill in the SalI, BclI, and Asp718
sites within the G4B BamHI fragment, we found that haploidstrains containing these changes could not be obtained; thisregion presumably contains an essential gene. Haploidstrains containing a filled-in XbaI site in the C2G BamHI
fragment grew extremely slowly and were not used in ourstudy.
E. coli RK1448 (38) was used as a host in most of thecloning experiments. Since some of the mutations generatedrestriction sites that could only be cleaved when unmethyl-ated, E. coli MM161 (from M. Marinus) was used as a hostfor some cloning experiments.DNA isolation and manipulations. DNA was isolated from
5-ml stationary-phase cultures of S. cerevisiae (34). For bothsmall-scale and large-scale plasmid DNA isolations, thealkaline lysis procedure was used (24); large-scale isolationswere further purified by CsCl-ethidium bromide densitygradients. A complete list of plasmids used for this study isgiven in Table 2. Standard cloning procedures were per-formed as described in Maniatis et al. (24). Restrictionenzymes were purchased from New England Biolabs, Inter-national Biotechnologies Inc., and Boerhinger-Mannheim.E. coli DNA polymerase, the Klenow fragment of DNApolymerase, and T4 DNA ligase were purchased from NewEngland Biolabs. 32P-labeled dATP was bought from Amer-
BG4B
B
BSXBS SaBcAiG1I I 1 11 1 1-mk lks j
Tyl-17 LEU2 ?
DSB B C2G B A2CB
X G2 G3 V-I4MXbI I 1 G11 1
ars CEN ars
HI1Kb
FIG. 1. Restriction map of the LEU2 to CEN region of chromo-some III. Abbreviations: A, Asp718; B, BamHI; Bc, Bcll; Bs,BstEII; C, ClaI; G, BglII; M, MluI; S, SpeI; Sa, Sail; X, XhoI; Xb,XbaI. The BgIII sites are designated Gl to G4 from LEU2 to CEN3.Symbols: ?, an essential function mapped within the G4B region; 0,position of CEN3 sequences; O, approximate locations of the twoars elements within the C2G region (30). Several of the restrictionsites shown in this figure were determined by Newlon et al. (28).
VOL. 8, 1988

598 SYMINGTON AND PETES
LS42
ura3
nsX Bs
IIIsIIliI
S G1 X G2 G3 BBcG4 MLS45
Bs X Bs
11 II liii1S G1 X G2 G3 BBeG4M
LS47
Bs X Bs
S G1 X G2 G3 BBcG4M C
FIG. 2. Locations of the wild-type restriction sites within strainsLS42, LS45, and LS47. Abbreviations and symbols are as for Fig. 1.
sham Corp. Southern filters were prepared by using Gene-screen (New England Nuclear Corp.) or Hybond nylonmembranes (Amersham Corp.) as specified by the manufac-turers. Hybridizations were done by methods recommendedfor these membranes with probes of specific activity of about5 x 107 cpm/,Ig in the presence of 5% dextran sulfate.
Yeast transformation. Transformation was performed bythe spheroplast technique (14). For transformations involv-ing linear DNA fragments (transplacements [33, 42]), weadded 10 to 30 ,g of DNA to 200 ,ul of spheroplasts (106spheroplasts per ,lI). Following nutritional selection, trans-formants were screened by Southern analysis (35) to identifythose yeast strains with the restriction pattern diagnostic ofthe desired substitution.
Tetrad analysis. Tetrad dissection was performed as de-scribed by Sherman et al. (34). Those tetrads in which acrossover had occurred between LEU2 and the centromerewere identified by looking for second-division segregationbetween LEU2 and the centromere-linked heterozygousmarkers TRPJ and ARG4 (26). In DNA samples derivedfrom spores of the diploid LS42 (27 tetrads), all 12 polymor-phic sites were analyzed. In samples derived from LS45 (41tetrads), in most cases all sites except the XhoI site centro-mere-distal to the LEU2 gene were examined. In samplesderived from LS47 (24 tetrads), all sites centromere-prox-imal to the LEU2 mutation were analyzed. Restrictiondigests of DNA with BstEII, XhoI, and SpeI (after transferto Hybond) were hybridized to a probe containing all of theG4B region except Tyl-17 (Fig. 1); filters containing DNAtreated with BglII, BclI, MIuI, and XhoI-BamHI were hy-bridized to the plasmid probe pLS47, which contained DNAderived from the D8B and C2G sequences (Fig. 1).
RESULTS
Construction of plasmids containing restriction site muta-tions. A gene bank constructed from a circular derivative ofchromosome III was used as a source of cloned DNAfragments for this study (28); the bank was prepared byinserting BamHI fragments of yeast DNA into the BamHIsite of the yeast vector Ylp5. Three plasmids (C2G, D8B,and G4B) were identified as containing contiguous BamHIfragments from chromosome III (28). The order of the
fragments (centromere to telomere) was C2G-D8B-G4B(Fig. 1). The C2G fragment contains the centromere ofchromosome III and two ars elements (putative origins ofreplication); the G4B fragment contains a gene involved inleucine biosynthesis (LEU2), a tRNA3LeU gene, and a por-tion of a Ty element (28, 30, 40).We constructed restriction site mutations within the
cloned plasmids by cleaving them with a restriction enzymethat left recessed 3' ends. These ends were filled in withDNA polymerase (Klenow fragment), and the cleaved plas-mid was religated. The ligated plasmids contain a small (4 to5 bp) insert at the recognition site of the enzyme used in theinitial digestion. For this study, we used restriction enzymesthat had sites which, when filled in, produced a site recog-nized by a different restriction enzyme. For example, whena BgIII site (AGATCT) is filled in, the new site (AGATCGATCT), although no longer recognized by BglII, is recog-nized by the ClaI enzyme (ATCGAT recognition sequence).By using this procedure, we could be sure that the appropri-ate sequence changes had been made without sequencingeach mutation. Successive restriction site alterations weremade within each of the three plasmids until at least fourmutations per plasmid were obtained (Table 2).
Construction of yeast strains. Three closely related diploidyeast strains, LS42, LS45, and LS47, were used in thesestudies. The construction of these strains is described indetail above and outlined in Table 1. In brief, we trans-formed yeast strains with plasmids (Table 2) derived fromchromosome III that contained changes in certain restrictionsites. The DNA fragments containing these changes replacedthe wild-type sequences on chromosome III. By variousgenetic crosses, we constructed diploid strains that wereheterozygous for these changes. One of the changes was inthe gene LEU2. The differences among the three strains aresummarized in Fig. 2. The results obtained with all strainswere very similar and have been pooled.
Analysis of meiotic recombination between LEU2 and thecentromere. The basic plan of these experiments was simple.The diploid strains described above were induced to undergomeiosis. We identified tetrads that had a crossover betweenLEU2 and the centromere and mapped the position of thecrossover by doing Southern analysis of DNA isolated fromcultures of each of the four spores.
Tetrads that had a crossover between LEU2 and thecentromere were identified by comparing the segregationpatterns of the heterozygous LEU2 and TRPI mutations(Fig. 3). The TRPJ gene is very tightly linked to thecentromere of chromosome IV, and therefore the wild-typeTRPI allele will almost always segregate away from themutant trpl allele at the first meiotic division (26). The LEU2gene is less tightly linked to the centromere of chromosomeIII. If there is no crossing-over between either gene and theirrespective centromeres, then one should obtain only twoclasses of tetrads (both representing first-division segrega-tion of LEU2), those that segregate 2 Trp+ Leu+ to 2 Trp-Leu- spores and those that segregate 2 Trp+ Leu- to 2 Trp-Leu+ spores. If there is a single crossover between LEU2and the centromere (or between TRPI and its centromere),the genotypes for all four spores should be different (te-tratype segregation): 1 Trp+ Leu+:1 Trp+ Leu-:1 Trp-Leu+:1 Trp- Leu-; this type of tetrad represents a second-division segregation. Since the TRPI gene is 0.5 cM from thecentromere and the LEU2 gene is about 5 cM from thecentromere, we expected most of the tetratypes wouldrepresent crossovers between LEU2 and the centromere.This expectation was supported by the physical analysis of
MOL. CELL. BIOL.

CLUSTERED MEIOTIC EXCHANGES IN S. CEREVISIAE
- - *-0
I I\ i\b.
T T t Ct
- 11
L TXFIG. 3. Genetic screen for crossing-over between LEU2 and
CEN3. The TRPI gene is very tightly linked to CEN4 and rarelyexhibits crossing over. LEU2 is less tightly linked to CEN3. (a)When no crossovers occur between LEU2 and CEN3, the Leu' andLeu- chromosomes segregate from each other at the first meioticdivision, resulting in one of two classes of tetrads, either 2 Trp+Leu+:2 Trp- Leu- or 2 Trp+ Leu-:2 Trp- Leu+. (b) When acrossover occurs between LEU2 and the centromere, the wild-typeand mutant alleles segregate at the second meiotic division, resultingin the formation of a tetratype tetrad (four spores of differentgenotype, Trp+ Leu+:Trp+ Leu-:Trp- Leu+:Trp- Leu-).
the recombinant spores described below. As described be-low, we also analyzed segregation of LEU2 relative toARG4, a second centromere-linked marker in the strain.Of the tetrads derived from LS42, LS45, and LS47, 12.7%
showed tetratype segregation between LEU2 and TRPI (26of 200 for LS42, 41 of 320 for LS45, and 24 of 197 for LS47).Since this frequency of tetratype asci is very similar to thatobserved for the diploid strain LS25 (12 of 98, 12.2%), whichhas no heterozygous restriction sites between LEU2 and thecentromere, we conclude that the presence of these sitesdoes not greatly perturb the frequency of meiotic recombi-nation on chromosome III.
In this study, we examined DNA from 91 tetratype tetradsderived from the diploids LS42, LS45, and LS47. As dis-cussed below, 82 of these tetrads represent crossoversbetween LEU2 and the centromere; 9 of the tetrads areeither crossovers between LEU2 and the centromere orTRPI and the centromere. In recombinant tetrads, DNAsamples from all four spores were examined by Southernanalysis. We found that the tetrads could be grouped intothree classes. In class 1 tetrads (44 tetrads), all restrictionsite markers segregated 2:2. In addition, in most class 1tetrads, two of the four spores contained restriction sites inthe parental configurations and the other two spores con-tained the restriction patterns expected as the result of asingle reciprocal crossover. In class 2 tetrads (30 tetrads),one or more of the heterozygous restriction sites showedconversion (3:1 or 1:3 patterns of segregation), and thepattern of restriction sites observed in the four spores wasconsistent with a single crossover adjacent to the conversiontract. Class 3 tetrads (17 tetrads) appeared to be the result of
multiple recombination events. This class included tetrads inwhich the conversion tract was separated from the positionof the crossover and those in which multiple crossoversoccurred. In addition to examining tetrads that had cross-overs between LEU2 and the centromere, we examined 11tetrads in which the LEU2 gene had shown conversion and19 control tetrads that did not show either conversion ofLEU2 or tetratype segregation between LEU2 and TRPJ.Both the experimental and control tetrads are discussed indetail below.
Class 1 tetrads: single crossovers. Of 91 tetrads, 44 weresingle crossovers, unassociated with gene conversion forany of the heterozygous restriction sites. Three types ofclass 1 tetrads were observed. The most common type (22tetrads) had two spores that contained the parental config-uration of restriction sites (one resembling each parent) andtwo spores with the complementary recombinant pattern ofrestriction sites (Fig. 4).The second type of class 1 tetrad (13 representatives) had
all 12 heterozygous restriction sites segregating 2:2, and allspores had the sites in the parental configurations. In addi-tion, in these tetrads, the TRPI and ARG4 genes showedfirst-division segregation (ditype tetrads). Since the LEU2gene showed second-division segregation with respect toboth TRPI and ARG4 (which is also centromere linked), thesimplest explanation of these tetrads is that recombinationhas occurred between the most centromere-proximal restric-tion site (MluI) and the centromere. This conclusion issupported by a second argument: all tetrads of this typederived from LS47 (three of three) had a crossover betweenthe MluI site and the restriction site marker on the other sideof the centromere from LEU2. Since this restriction sitemarker (which was about 1 kb from the centromere) showedfirst-division segregation with TRPJ, we conclude that thesetetrads have a crossover between the MIuI site and thecentromere.The third type of class 1 tetrad (nine tetrads) had the
parental configuration of markers for all 12 heterozygousrestriction sites between LEU2 and the centromere. Inaddition, these tetrads were tetratype for TRPI and ARG4.This type of tetrad may represent a crossover between TRPJand its centromere. Since the ARG4 gene shows approxi-mately 25% second-division segregation (27), we cannot ruleout the alternative possibility that these tetrads representtwo events, a crossover between LEU2 and its centromereproximal to the MluI site (as described above) and a secondcrossover between ARG4 and its centromere. Since wecannot resolve these possibilities, we cannot include thesedata as representing crossovers between LEU2 and thecentromere.
Thus, of the 82 tetrads containing a crossover betweenLEU2 and the centromere, 35 (43%) represent single recip-rocal crossovers. The positions of these exchanges areshown in Fig. 5a.
Class 2 tetrads: single crossovers associated with geneconversion of adjacent markers. Since gene conversionevents are associated with reciprocal recombination of flank-ing markers and are assumed to be an intermediate in thegeneration of a reciprocal exchange (10, 29, 41), it is notsurprising that many of the tetrads that had a crossoverbetween LEU2 and the centromere were associated withconversion events of adjacent markers. These conversionevents were detected as restriction site markers that segre-gated either 3 wild-type restriction sites:1 filled-in site or 1wild-type site:3 filled-in sites (Fig. 4). We included in thisclass only those tetrads in which the conversion tract was
a.
Tr Tt t
LfL I I
VOL. 8, 1988 599
600 SYMINGTON AND PETES
a)a b c d
b)a b c d
C)a b c d
_ -
Sm l9.lKb_w :-14.9a10.
_0 - 8.6_ ~~~~6.3
4.2
a B4.2X6.3 B8.6 B3 aBB3 a B X B Bb B X B B b B X B B b B X 14.9 B
B 19.1Kb B c B X B Bc
dB d B X 9B B 10.5 B B
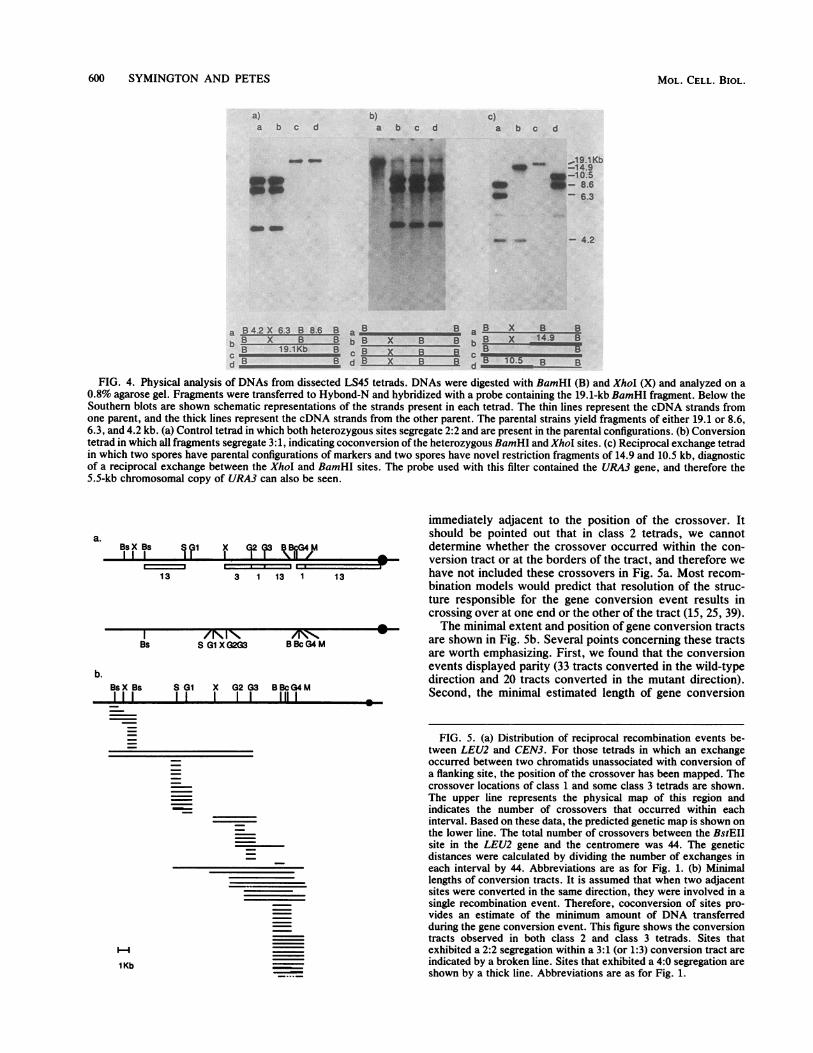
FIG. 4. Physical analysis of DNAs from dissected LS45 tetrads. DNAs were digested with BamHI (B) and XhoI (X) and analyzed on a0.8% agarose gel. Fragments were transferred to Hybond-N and hybridized with a probe containing the 19.1-kb BamHI fragment. Below theSouthern blots are shown schematic representations of the strands present in each tetrad. The thin lines represent the cDNA strands fromone parent, and the thick lines represent the cDNA strands from the other parent. The parental strains yield fragments of either 19.1 or 8.6,6.3, and 4.2 kb. (a) Control tetrad in which both heterozygous sites segregate 2:2 and are present in the parental configurations. (b) Conversiontetrad in which all fragments segregate 3: 1, indicating coconversion of the heterozygous BamHI and XhoI sites. (c) Reciprocal exchange tetradin which two spores have parental configurations of markers and two spores have novel restriction fragments of 14.9 and 10.5 kb, diagnosticof a reciprocal exchange between the XhoI and BamHI sites. The probe used with this filter contained the URA3 gene, and therefore the5.5-kb chromosomal copy of URA3 can also be seen.
a.BsXBs I X GI92 ?3 kBrI
13 3 1 13 1 13
I /N IN mN,Bs S G1 XG2G3 BBcG4 M
b.BsX Bs S G1 X G2 3 BBcG4MI II I I I I lIIII
immediately adjacent to the position of the crossover. Itshould be pointed out that in class 2 tetrads, we cannotdetermine whether the crossover occurred within the con-version tract or at the borders of the tract, and therefore wehave not included these crossovers in Fig. 5a. Most recom-bination models would predict that resolution of the struc-ture responsible for the gene conversion event results incrossing over at one end or the other of the tract (15, 25, 39).The minimal extent and position of gene conversion tracts
are shown in Fig. 5b. Several points concerning these tractsare worth emphasizing. First, we found that the conversionevents displayed parity (33 tracts converted in the wild-typedirection and 20 tracts converted in the mutant direction).Second, the minimal estimated length of gene conversion
FIG. 5. (a) Distribution of reciprocal recombination events be-tween LEU2 and CEN3. For those tetrads in which an exchangeoccurred between two chromatids unassociated with conversion ofa flanking site, the position of the crossover has been mapped. Thecrossover locations of class 1 and some class 3 tetrads are shown.The upper line represents the physical map of this region andindicates the number of crossovers that occurred within eachinterval. Based on these data, the predicted genetic map is shown onthe lower line. The total number of crossovers between the BstEIIsite in the LEU2 gene and the centromere was 44. The geneticdistances were calculated by dividing the number of exchanges ineach interval by 44. Abbreviations are as for Fig. 1. (b) Minimallengths of conversion tracts. It is assumed that when two adjacentsites were converted in the same direction, they were involved in asingle recombination event. Therefore, coconversion of sites pro-vides an estimate of the minimum amount of DNA transferredduring the gene conversion event. This figure shows the conversiontracts observed in both class 2 and class 3 tetrads. Sites thatexhibited a 2:2 segregation within a 3:1 (or 1:3) conversion tract areindicated by a broken line. Sites that exhibited a 4:0 segregation areshown by a thick line. Abbreviations are as for Fig. 1.
1Kb
MOL. CELL. BIOL.

CLUSTERED MEIOTIC EXCHANGES IN S. CEREVISIAE
tracts varied considerably, from 4 bp (the size of eachmutation) to as long as 12 kb. Third, although a few geneconversion tracts were discontinuous (2:2 segregation of aheterozygous site flanked by other sites segregating 3:1 or1:3), most gene conversion events were continuous. Fourth,different regions of the chromosome had different frequen-cies of conversion. For example, there were 26 conversionevents involving the heterozygous BamHI site and only 6events involving the XhoI site. There were frequent cocon-version events involving the contiguous BamHI, BclI, BgIII(4), and MluI sites. These conversion tracts sometimesextended as far as the SpeI site, suggesting that there may bean initiation site for conversion near the MluI site fromwhich events extend for variable distances towards theLEU2 gene. Such polarity gradients for conversion havebeen observed within some genes (reviewed by Orr-Weaverand Szostak [29]). Fifth, the endpoints of conversion tractscoincided with the hotspots for simple crossovers that werenot associated with conversion events, suggesting that con-version-associated crossovers might occur at the same posi-tions.
Class 3 tetrads: multiple recombination events. Of the 82tetrads that had a crossover between LEU2 and the centro-mere, 17 were complex. In these tetrads, there were eithermore than one crossover or more than one gene conversion
event. We also included in this class those tetrads in whichthe crossover was separated from the conversion tract by asite exhibiting 2:2 segregation. The class 3 events are shownin Fig. 6, as are the number and type of recombinationevents required to explain the recombinant patterns. Theassumptions used in determining the number of requiredrecombination events were that both gene conversion andcrossing-over occur after DNA synthesis and that geneconversion is the result of mismatch repair of asymmetricheteroduplexes (25). In addition, we allowed for formation of"patchy" heteroduplexes but only if a region of 2:2 segre-gation was flanked on both sides by regions of gene conver-sion. As one example of this type of analysis, we can obtainthe pattern of recombination tracts found in tetrad LS42-6 byallowing one crossover and two nonadjacent gene conver-sion events (Fig. 6). One of the unusual features of tetradLS45-225 (found also in one of the tetrads that had a geneconversion event at LEU2 [described below], one of thecontrol tetrads, and one of the class 2 tetrads) is that one ofthe restriction site markers segregated 4:0. Although 4:0segregations are usually explained as the result of mitoticrecombination events (10), our results suggest that some ofthese events may be the result of multiple meiotic recombi-nation events, since the 4:0 tracts were often associated with3:1 or 1:3 conversion tracts. It should also be pointed out
LS42-6(CO+2NAC)
===XXXXXXXXX
XXXX XXXX XX Xxxxxxxxxxxxx
LS42-34(CO+NAC)
xxxxxxxxxxxxXXXXXXXXXXXX= ==XX=XXXXXX
LS42-117(2CO+1 CACO)
XXX ==XXXXXXX= ='XXXXX===3
LS42-139(CO+NAC)xxxxxxxxxxxx-XXXXXXXXXXX. =.X X=X X XX X
LS42-235(2CO+CACO)XXXXXXXXX-X-=-xx==XXXXXX= =S=====X=XXXXXXXXXX-X=
LS42-344(CO+NAC)XXXXXXXX, - .-===== = =XXXX
xxxxxxxxxxxx
LS45-12(CO+CACO).--XXXXXXXX-XXX--XXXXXXXxxxxxxxxxxxx
LS45-23(CO+CACO)
=====XXXX-s.=XX==xxxxx-XXXXXXXXXXXXxxxxxxxxxxxx
LS45-140(CO+NAC)===XXXXXXXXX
xxxxxxxxxxxxXXXXXXXXXXX-
LS45-150(CACO+NAC)===XXXXXXXXXXXXXXXXXXX.-xxxxx-------
LS45-225(CACO+2NAC)-==XXXXXXXXX===X==XXXXXXxxxxxxxxxxxxXXXXX==-..s
LS47-55(CACO+NAC)xxxxxxxxxxxxx
XXXXXXXXM=====
LS47-72(CACO+NAC)
X....,. ==
.= = ==XXXXXXXXXXXXXX.X='==
LS47-229(CO+NAC)-----xxxxxxxx
XXXXXXXXXXXXXXXXXXXXX=====
LS47-237(CO+NAC)
w==XXXXXXX
xxxxxxxxxxxxx
XXXXXXsX = = ==
LS42-337(3CO+2NAC)XXX ..X=-=-=xxxxxxxxxxx-=====X==X===
LS45-216(2CACO)
...XXXXX= ===X---xxxxx--==xxxxxxxxxxxxXXXXXXXX XX X
FIG. 6. Recombination patterns in tetrads with multiple events. Each line of X's or ='s represents the pattern for the 12 heterozygousrestriction sites in one spore; a parental (nonrecombinant) pattern would be two lines with X's only and two lines with ='s only. The leftmostsite is centromere-distal. Combinations of crossovers and gene conversion events required to give the recombinant tetrads are indicated inparentheses (CO, simple crossover; CACO, conversion-associated crossover; NAC, nonadjacent conversion; 2CO and 3CO, double or triplecrossover, respectively.
VOL. 8, 1988 601

602 SYMINGTON AND PETES
that the patterns found in LS45-225 and similar tetrads can
be explained in a number of different ways. In particular,assumptions about the mechanism and timing of gene con-
version affect the conclusions. Whatever recombinationevents generate class 3 tetrads, it is clear that meioticrecombination patterns are often unexpectedly complex.LEU2 conversion tetrads. In the LS42, LS45, and LS47
strains, the LEU2 gene showed conversion in 3.4% of tetrads(Fig. 5b). We analyzed 11 such tetrads in order to examinethe frequency of coconversion of flanking sites. As ex-
pected, in all such tetrads, the BstEII site that marked theleu2 mutation had converted. Two tetrads had converted thetwo distal markers as well as five proximal markers, result-ing in conversion tracts of at least 11.5 kb. Of the 11conversion tetrads examined, 3 were complex, involvingeither more than one conversion tract or a nonadjacentcrossover. Thus, the frequency of complex events among
LEU2 conversion tetrads was about the same as in crossovertetrads.
Control tetrads. In addition to studying tetrads in which a
crossover occurred between LEU2 and the centromere, we
also examined tetrads in which no apparent crossover hadoccurred (LEU2 and TRPI showing first-division segrega-
tion). Of 19 tetrads, 15 had the parental restriction patternfor all spores. Three had gene conversion events, two tetradswith single converted sites and a third with a long conversion
tract (4:0 segregation for three of the four centromere-proximal restriction sites, 3:1 segregation for the MluI site).One tetrad had a two- or four-strand double crossover. Thus,approximately 16% of tetrads that did not have a second-division segregation for LEU2 had a detectable gene conver-sion event. Since about 13% (91 of 717)of unselected tetradshad a crossover between LEU2 and the centromere andsince about half of meiotic gene conversion events are notassociated with reciprocal recombination, this result was notunexpected; the difference between the expected value of13% of unselected tetrads with a conversion event without acrossover and the observed value of 16% was not statisti-cally significant (contingency chi-squared value of 0.001).
DISCUSSION
Our results indicate that the frequency and type of meioticrecombination events are a complex function of the structureand/or sequence of the chromosome. Our data can besummarized as follows: the density of crossover events isnot uniform along the chromosome, crossover events are
frequently associated with conversion events, meiotic con-
version tracts in S. cerevisiae can be long (at least 12 kb) andare usually continuous, and complex recombination eventsare fairly common. These conclusions will be discussed indetail below.The simple crossovers are concentrated in three regions in
the LEU2 to centromere III interval (Fig. Sa). One can
calculate the expected number of crossovers in each physi-cal interval (assuming a uniform distribution of exchanges)by multiplying 44 (the total number of crossovers) by thefraction of the total distance (between LEU2 and the centro-mere) represented by each interval. We found that the regionbetween the heterozygous BamHI and BglII (3) restrictionsites had significantly more (chi-squaredvalue of 9.3) and theregion between the XhoI and SpeI sites had significantlyfewer (chi-squared value of 8.5) crossovers than expectedfrom a uniform distribution. It should be emphasized that wedo not know whether the nonuniform distribution of recomn-bination events is the result of the structure of the chromo-
A(a).
(b)
(c)
(d)
(e)
B(a)
(b)
(c)
MOL;CELL. BIOL.
TTZX.(d)
c
(a)
(b)
(C)
(d)
-f--h-I.rDc:
-~~ZC
FIG. 7. Models for recombination. (A) Holliday model for re-combination. (a) Strands of the same polarity are nicked at homol-ogoIs positions. (b) The broken strands unwind and exchange toproduce a symmetrical heteroduplex. The crossed-strand structureis known as the Holliday junction or Holliday intermediate. (c)Nicks are repaired, and the junction may branch-migrate to enlargethe region of hybrid DNA on both duplexes.(d) The intermediate isresolved by cleavageof the crossed strands. Depending on whichstrands are cleaved, exchange of flanking markers may result. (e)Mismatch correction of the heteroduplex generates conversion to
'IL

CLUSTERED MEIOTIC EXCHANGES IN S. CEREVISIAE 603
some, the sequence of the chromosome, or both factors. Forexample, it is possible that the certain regions have morerecombination events than other regions because the meioticchromosome is folded in a way that affects the susceptibilityof the chromosome to the recombination machinery. Alter-natively, the distribution of the recombination events may beprimarily the result of differences in DNA sequence. By thisinterpretation, the hotspots for meiotic recombination maybe analogous to Chi sites in E. coli (36).
One-third of the simple crossovers occurred within 6 kb ofthe centromere. This frequency of exchange is approxi-mately the average frequency for the LEU2 to CEN3 inter-val, indicating that there is not a strong repression of meioticrecombination by the centromere. We observed in relatedstudies that a marker 1 kb from CEN3 also recombined atrates comparable to the average genomic value of 0.37cM/kb (unpublished data). Lambie and Roeder (20) sug-gested that the yeast centromere represses recombination,since they found that moving the chromosome III centro-mere to a new position on the same chromosome increasedrecombination in the region from which the centromere wasremoved and decreased recombination at the new position.Our results indicate either that this repression is relativelyweak or that moving the centromere has a global effect onrecombination within the chromosome that is not directlyrelated to centromeric repression on exchange.As expected from previous studies (for example, reference
16), crossovers between LEU2 and the centromere werefrequently associated with gene conversion events at adja-cent restriction sites. The mechanism by which gene conver-sion occurs and the association with reciprocal recombina-tion are explained differently by different models ofrecombination. In the Holliday model (15), gene conversionresults from the repair of mismatches in symmetric hetero-duplexes (Fig. 7A). In the Meselson-Radding model (25),gene conversion is the result of mismatch repair of a single(asymmetric) heteroduplex (Fig. 7B). In the model ofSzostak et al. (39), most conversion events result from gaprepair of a double-strand break of the DNA (Fig. 7C). In allmodels, the structure formed during conversion is resolvedinto two chromosomes (either recombined or parental) by
either wild-type or mutant information. (B) Meselson-Raddingmodel. (a) Recombination is initiated by formation of a single-strandnick; the resulting free DNA end serves as a primer for DNA repairsynthesis, displacing a single strand. (b) The displaced strandinvades a homologous duplex to form a D-loop. (c) The D-loop is
degraded, and the asymmetric heteroduplex enlarges by DNAsynthesis on the donor chromatid coupled with degradation on therecipient duplex. (d) Ligation of the nicks produces a Hollidayjunction that may branch-migrate to generate symmetrical hetero-duplex adjacent to the region of asymmetrical heteroduplex. Reso-lution of the Holliday junction yields either crossover or noncross-over products as described for (A) above. Mismatch correction ofthe asymmetrical heteroduplex yields either gene conversion (3:1) orrestoration (2:2) events. (C) Double-strand-break repair model (39).(a) A double-strand break is made in one duplex, which is thenenlarged to a gap with 3' overhangs by the action of nucleases. (b)One 3' end invades a homologous duplex, displacing a D-loop. (c)The D-loop enlarges by repair synthesis until the other 3' end can
pair with complementary sequences on the displaced strand. (d)Repair synthesis from the other 3' end repairs the gap. Ligation ofthe nicks results in the formation of two Holliday junctions that maybranch-migrate. Resolution of both junctions in the same sense(both inner or both outer strands) leads to noncrossover products;whereas resolution in the opposite sense generates flanking markerexchange.
cleavage of the intermediates at the end of the conversiontract.The distribution of gene conversion events, like the dis-
tribution of crossovers, is not uniform along the chromo-some. The length of the conversion tracts was longer thanexpected on the basis of most previous genetic studiesinvolving coconversion of mutant alleles within single genes(10), although there are two reports of conversion tracts thatextended over large genetic distances (5, 11). Althoughalmost all of the observed conversion tracts were continuous(as in previous studies [10]), we found a few tracts in whicha site that segregated 2:2 was flanked by sites that weresegregating 3:1 or 1:3 (Fig. 5b). Such discontinuous tractscan be explained either as two independent gene conversionevents within one region or as a patchy repair of a hetero-duplex involving multiple sites (18).About 22% of the tetrads that had a crossover between
LEU2 and the centromere appeared to have more than onerecombination event (class 3 tetrads). Since 20% of thecontrol tetrads (those that did not have a crossover betweenLEU2 and the centromere) had either gene conversion ormultiple crossover events, it is likely that many of the class3 tetrads can be explained as representing two independentrecombination events. Some of the tetrads, however, ap-peared to be too complex to represent independent recom-bination events. For example, tetrad LS42-337 (Fig. 6)required at least three recombination events other than theprimary single crossover. Such tetrads can be explained intwo ways. First, recombination events within a chromo-somal region, although representing different regions ofheteroduplex formation and resolution, may be concerted;such concerted recombination events have been suggestedpreviously (summarized by Whitehouse [41] and Fink andPetes [7]). Second, some recombination events may be theresult of a different mechanism than repair and resolution ofasymmetric heteroduplexes formed in G2 (25). For example,patchy repair of symmetric heteroduplexes formed in Glcould explain any pattern of class 3 tetrads. Whatevermechanisms are involved in formation of the class 3 pat-terns, it is clear that recombination events that appeared tobe simple crossovers between LEU2 and the centromere bygenetic criteria (second-division segregation) are often verycomplex. In studies of yeast meiotic recombination in plas-mid sequences inserted between duplicated MAT loci, Bortsand Haber (personal communication) detected similar fre-quencies of complex events.
In summary, genetic distance in S. cerevisiae is not simplyproportional to physical distance. The frequency of recom-bination events, both crossovers and gene conversions,varies along the length of the chromosome.
ACKNOWLEDGMENTSWe thank Carol Newlon for yeast strain XJ24-24a and the
plasmids used in constructing the restriction site changes. We thankM. Fitzgerald-Hayes, S. Fogel, S. Jinks-Robertson, M. Kupiec, S.Liebman, C. Newlon, S. Roeder, L. Sandler, F. Stahl, G. Smith, A.Stapleton, and A. Vincent for useful discussions and/or commentson the manuscript and Michael White for technical assistance.The research was supported by Public Health Service research
grant GM24110 from the National Institutes of Health. L.S.S. was aSpecial Fellow of the Leukemia Society of America.
LITERATURE CITED1. Angel, T., B. Austin, and D. G. Catcheside. 1970. Regulation of
recombination at the his3 locus in Neurospora crassa. Aust. J.Biol. Sci. 23:1229-1240.
2. Boeke, J. D., F. Lacroute, and G. R. Fink. 1984. A positive
VOL. 8, 1988

604 SYMINGTON AND PETES
selection for mutants lacking orotidine-5'-phosphate decarbox-ylase activity in yeast: 5-fluoro-orotic acid resistance. Mol. Gen.Genet. 197:345-346.
3. Clarke, L., and J. Carbon. 1980. Isolation of a yeast centromereand construction of functional small circular chromosomes.Nature (London) 287:504-509.
4. Coleman, K. G., H. Y. Steensma, D. B. Kabak, and J. R. Pringle.1986. Molecular cloning of chromosome I DNA from Saccha-romyces cerevisiae: isolation and characterization of the CDC24gene and adjacent regions of the chromosome. Mol. Cell. Biol.6:4516-4525.
5. DiCaprio, L., and P. J. Hastings. 1976. Gene conversion andintrageneic recombination at the sup6 locus and the surroundingregion in Saccharomyces cerevisiae. Genetics 84:697-721.
6. Dobzhansky, T. 1930. Translocations involving the third andfourth chromosomes of Drosophila melanogaster. Genetics 15:347-399.
7. Fink, G. R., and T. D. Petes. 1984. Gene conversion in theabsence of reciprocal recombination. Nature (London) 310:728-729.
8. Fitzgerald-Hayes, M., L. Clarke, and J. Carbon. 1982. Nucleo-tide sequence comparisons and functional analysis of yeastcentromere DNAs. Cell 29:235-244.
9. Fogel, S., R. Mortimer, K. Lusnak, and F. Tavares. 1979.Meiotic gene conversion: a signal of the basic recombinationevent in yeast. Cold Spring Harbor Symp. Quant. Biol. 43:1325-1341.
10. Fogel, S., R. K. Mortiner, and K. Lusnak. 1981. Mechanisms ofgene conversion, p. 289-339. In J. N. Strathern, E. W. Jones,and J. R. Broach (ed.), The molecular biology of the yeastSaccharomyces, vol. 1. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
11. Fogel, S., R. K. Mortimer, and K. Lusnak. 1983. Meiotic geneconversion in yeast: molecular and experimental perspectives,p. 65-107. In J. F. T. Spencer, D. M. Spencer, and A. R. W.Smith (ed.), Yeast genetics. Springer-Verlag, New York.
12. Gutz, M. 1971. Site-specific induction of recombination inSchizosaccharomyces pombe. Genetics 69:317-337.
13. Hieter, P., D. Pridmore, J. H. Hegemann, M. Thomas, R. W.Davies, and P. Philippsen. 1985. Functional selection and anal-ysis of yeast centromeric DNA. Cell 42:913-921.
14. Hinnen, A., J. R. Hicks, and G. R. Fink. 1978. Transformationof yeast. Proc. Natl. Acad. Sci. USA 75:1929-1933.
15. Holliday, R. 1964. A mechanism for gene conversion in fungi.Genet. Res. 5:282-304.
16. Hurst, D. D., S. Fogel, and R. K. Mortimer. 1972. Conversion-associated recombination in yeast. Proc. Natl. Acad. Sci. USA69:101-105.
17. Jinks-Robertson, S., and T. D. Petes. 1985. High frequencymeiotic gene conversion between repeated genes on nonhomol-ogous chromosomes in yeast. Proc. Natl. Acad. Sci. USA82:3350-3354.
18. Kalogeropoulos, A., and J.-L. Rossignol. 1980. Evidence forindependent mismatch corrections along the same hybrid DNAtract during meiotic recombination in Ascobolus. Heredity45:263-270.
19. Keil, R., and G. S. Roeder. 1984. cis-Acting recombination-stimulating activity in a fragment of the ribosomal DNA of S.cerevisiae. Cell 39:377-386.
20. Lambie, E. J., and G. S. Roeder. 1986. Repression of meioticcrossing over by a centromere (CEN3) in Saccharomyces cere-visiae. Genetics 114:769-789.
21. Larkin, J. C., and J. L. Woolford. 1984. Isolation and charac-
terization of the CR Y1 gene: a yeast ribosomal protein gene.Nucleic Acids Res. 11:403-420.
22. MacDonald, M., and H. Whitehouse. 1979. A buff spore colormutant in Sordaria brevicollis showing high frequency conver-sion. I. Characteristics of the mutant. Genet. Res. 34:87-119.
23. Malone, R. E., J. E. Golin, and M. S. Esposito. 1980. Mitoticversus meiotic recombination in Saccharomyces cerevisiae.Curr. Genet. 1:241-248.
24. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
25. Meselson, M., and C. Radding. 1975. A general model forgenetic recombination. Proc. Natl. Acad. Sci. USA 72:358-361.
26. Mortimer, R. K., and D. Schild. 1981. Genetic mapping inSaccharomyces cerevisiae, p. 11-26. In J. N. Strathern, E. W.Jones, and J. R. Broach (ed.), The molecular biology of theyeast Saccharomyces, vol. 1. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
27. Mortimer, R. K., and D. Schild. 1985. Genetic map of Saccha-romyces cerevisiae. Microbiol. Rev. 49:181-212.
28. Newlon, C., R. P. Green, K. J. Hardeman, K. E. Kim, L. R.Lipchitz, P. G. PalzkilH, S. Synn, and S. T. Woody. 1986. Struc-ture and organization of yeast chromosome III. UCLA Symp.Mol. Cell. Biol. New Series 33:211-223.
29. Orr-Weaver, T., and J. Szostak. 1985. Fungal recombination.Microbiol. Rev. 49:33-58.
30. Palzkill, T. G., S. G. Oliver, and C. S. Newlon. 1986. DNAsequence analysis of ars elements from chromosome III ofSaccharomyces cerevisiae: identification of a new conservedsequence. Nucleic Acids Res. 14:6247-6263.
31. Pontecorvo, G., and E. Kafer. 1958. Genetic analysis based onmitotic recombination. Adv. Genet. 9:71-104.
32. Ponticeili, A. S., D. W. Schultz, A. F. Taylor, and G. R. Smith.1985. Chi-dependent DNA strand cleavage by RecBC enzyme.Cell 41:145-151.
33. Rothstein, R. J. 1983. One-step gene disruption in yeast. Meth-ods Enzymol. 101:202-211.
34. Sherman, F., G. R. Fink, and J. B. Hicks. 1983. Methods inyeast genetics. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
35. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
36. Stahl, F. W., and M. M. Stahl. 1977. Recombination pathwayspecificity of Chi. Genetics 94:235-248.
37. Strathern, J. N., C. Newlon, I. Herskowitz, and J. B. Hicks.1979. Isolation of a circular derivative of yeast chromosome III:implications for the mechanism of mating type interconversion.Cell 18:309-319.
38. Symington, L. S., L. Fogarty, and R. D. Kolodner. 1983. Geneticrecombination of homologous plasmids catalyzed by cell-freeextracts of Saccharomyces cerevisiae. Cell 35:805-813.
39. Szostak, J., T. Orr-Weaver, R. Rothstein, and F. Stahl. 1983.The double-strand break repair model for recombination. Cell33:25-35.
40. Warmington, J., R. Anwar, C. Newlon, R. Waring, R. WayneDavies, K. I. Indge, and S. Oliver. 1986. A "hotspot" for Tytransposition on the left arm of yeast chromosome III. NucleicAcids Res. 14:3475-3485.
41. Whitehouse, H. 1982. Genetic recombination: understanding themechanisms. John Wiley & Sons, Inc., New York.
42. Winston, F., F. Chumley, and G. R. Fink. 1983. Eviction andtransplacement of mutant genes in yeast. Methods Enzymol.101:211-228.
MOL. CELL. BIOL.
Related Documents











