1 23 European Journal of Applied Physiology ISSN 1439-6319 Eur J Appl Physiol DOI 10.1007/s00421-012-2374-0 Exercise thermoregulatory responses following a 28-day sleep-high train-low regimen Stylianos N. Kounalakis, Ola Eiken & Igor B. Mekjavic

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
European Journal of AppliedPhysiology ISSN 1439-6319 Eur J Appl PhysiolDOI 10.1007/s00421-012-2374-0
Exercise thermoregulatory responsesfollowing a 28-day sleep-high train-lowregimen
Stylianos N. Kounalakis, Ola Eiken &Igor B. Mekjavic

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL ARTICLE
Exercise thermoregulatory responses following a 28-daysleep-high train-low regimen
Stylianos N. Kounalakis • Ola Eiken •
Igor B. Mekjavic
Received: 3 November 2011 / Accepted: 27 February 2012
� Springer-Verlag 2012
Abstract The potentiated exercise-sweating rate observed
during acute hypoxia is diminished after a sleep-high train-
low (SH-TL) regimen. We tested the hypothesis that this
attenuation of the sweating response after SH-TL is com-
pensated for by an increase in heat loss via vasodilatation.
Nine male subjects participated in a 28-day SH-TL regimen.
Before (pre-), and after (post-) the SH-TL protocol, they
performed an _VO2peak test under normoxia and hypoxia.
Additionally, pre- and post-SH-TL they completed three
30-min constant-work rate trials on a cycle ergometer. In one
trial, the subjects inspired room air while exercising at 50 %
of normoxic _VO2peak (CT). In the remaining trials, subjects
exercised in hypoxia (FIO2 12.5 %), either at the same
absolute (HAT) or relative (50 % of hypoxic _VO2peak)
work rate (HRT) as in CT. Despite similar exercise core
temperature responses between pre- and post-SH-TL trials,
sweating rate was potentiated in HAT pre-SH-TL [CT:
1.97 (0.42); HRT: 1.86 (0.31); HAT: 2.55 (0.53) mg
cm-2 min-1; p \ 0.05]. Post-SH-TL exercise sweating rate
was increased for CT, and remained unchanged in HRT and
HAT [CT: 2.42 (0.76); HRT: 2.01 (0.33); HAT: 2.59
(0.30) mg cm-2 min-1]. Pre-SH-TL, the forearm-fingertip
skin temperature difference (Tskf-f) was higher in HAT than
in CT and HRT by *3.5�C (p \ 0.05). The inter-condition
differences in Tskf-f were diminished post-SH-TL. In con-
clusion, the decrease in sweating rate during hypoxic exer-
cise, following a SH-TL regimen, was countered by an
increase in vasodilatation (reduced Tskf-f), whereas SH-TL
enhanced the sweating response during normoxic exercise.
The mechanisms underlying these SH-TL-induced altera-
tions in thermoregulatory responses remain to be settled.
Keywords Near infrared spectroscopy � Sweating �Aerobic training � Altitude acclimatization �Relative work rate � Absolute work rate
Introduction
The exercise sweating response is initiated by thermal
factors, namely, the elevation in core (Tc) and/or skin (Tsk)
temperatures, and by non-thermal factors such as acute
hypoxia and muscle ischemia (Mekjavic and Eiken 2006).
For example, during steady-state cycling at a given exter-
nal workload and at similar Tc and Tsk, sweating rate
is potentiated by local ischemia in the working muscles
(Eiken and Mekjavic 2004; Kacin et al. 2005) and by
hypoxemia (Kacin et al. 2007). During exercise with local
ischemia, the reduction in blood flow to the working
muscles is accompanied by an attenuated cutaneous vaso-
dilatation, which is compensated for by an exaggerated
sweating response (Kacin et al. 2005); a phenomenon that
would appear to serve the whole-body heat balance.
Likewise, at the same absolute work rate, acute hypoxia
enhances evaporative over conductive heat loss despite no
alterations in Tc (Greenleaf et al. 1969).
Communicated by George Havenith.
S. N. Kounalakis � I. B. Mekjavic
Department of Automation, Biocybernetics and Robotics,
Jozef Stefan Institute, Ljubljana, Slovenia
S. N. Kounalakis (&)
Human Performance-Rehabilitation Laboratory,
Faculty of Physical and Cultural Education,
Evelpidon Hellenic Military University, Vari, Greece
e-mail: [email protected]
O. Eiken
Department of Environmental Physiology, School of Technology
and Health, Royal Institute of Technology, Stockholm, Sweden
123
Eur J Appl Physiol
DOI 10.1007/s00421-012-2374-0
Author's personal copy

Prolonged interventions, such as aerobic training (Rob-
erts et al. 1977) and altitude acclimatization also modify
the sweating response. When such interventions are con-
current, their effects may be conflicting. For example, the
popular intervention to enhance working capacity in ath-
letes is the sleep-high train-low (SH-TL) regimen. The two
components of the SH-TL regimen, which may influence
the exercise thermoregulatory responses, are hypoxic
acclimatization (SH) and aerobic training (TL). Acclima-
tization to hypoxia would presumably reduce the magni-
tude of the response observed during acute hypoxia,
namely potentiation of sweating at the expense of con-
ductive heat loss (Eiken and Mekjavic 2004; Shibasaki
et al. 2001). In contrast, the training component of SH-TL
enhances the magnitude and gain of the sweating response
(Nadel et al. 1974; Roberts et al. 1977) and shifts the core
temperature threshold for onset of sweating and vasodila-
tation toward lower Tc levels (Roberts et al. 1977). Pre-
viously, we reported that the augmented sweating response
during hypoxic compared to normoxic exercise conducted
at identical absolute workloads, was no longer evident after
SH-TL (Kacin et al. 2007).
The present study was designed to investigate the
combined role of hypoxic acclimatization and aerobic
training on the exercise sweating and vasomotor responses,
with the concomitant exploration of the oxygenation levels
and blood volume in the working muscles and cerebral
area. The primary aims were to confirm previous findings
(Kacin et al. 2007) of potentiation of the sweating rate
during acute hypoxia, and of an attenuation of the exag-
gerated sweating responses following a 28-day SH-TL
regimen. The secondary aims were to investigate whether
the SH-TL-induced attenuation of the sweating response
during hypoxic exercise would be accompanied by changes
in cutaneous vasomotor tone, and hence in the provisions
for convective heat loss, and also to shed some light on the
mechanisms underlying such SH-TL-induced changes in
thermoregulatory responses by investigating regional
changes in muscle blood volume and oxygenation during
exercise. Thus, both endurance training and hypoxic
acclimatization may alter the exercise responses for blood-
flow distribution and oxygenation of cerebral and con-
tracting muscle tissues (Ainslie et al. 2008; Wang et al.
2010), factors that, as mentioned, are known to affect heat-
dissipation mechanisms (Greenleaf et al. 1969; Eiken and
Mekjavic 2004; Kacin et al. 2005, 2007).
Methods
Nine healthy, male subjects with an average (SD) age of 24
(3) years and stature of 179 (5) cm participated in the
study. All subjects were aerobically fit, but none were
endurance athletes. They were all accustomed to cycle
exercise as well as exercise testing on cycle ergometer.
Before enrolling in the study, each subject was familiarized
with the _VO2peak test and abstained from altitude exposures
[500 m above sea level for at least 1 month before the
experiments. Subjects’ participation was approved by a
physician. Following familiarization with the protocol and
instrumentation, subjects gave their written consent to
participate in the study. The protocol was approved by the
National Committee for Medical Ethics at the Ministry of
Health (Republic of Slovenia).
Study outline
All subjects conducted a normoxic and a hypoxic _VO2peak
test, and three constant-workload exercise tests, before
(pre-) and after (post-) the 28-day SH-TL regimen, during
which the subjects were exposed to normobaric hypoxia for
at least 9 h every night; ambient partial pressure of oxygen
corresponded approximately to that of an altitude above sea
level of 2,800 m (1st week), 3,000 m (2nd week) and
3,200 m (last 2 weeks) (for details see below). During
daytime, all subjects underwent 1 h training sessions in
room air on a cycle ergometer, five times per week. The
SH-TL regimen was conducted at the Olympic Sport
Centre Planica (Ratece, Slovenia) situated at an altitude of
900 m above sea level. The daily training sessions were
thus conducted at an altitude of 900 m. During the evening
and night-time hours, subjects were sequestered on one
floor of the Olympic Centre in which the oxygen levels
were reduced to simulate the partial pressure of oxygen at
the desired altitude.
Peak work rate tests
Subjects’ pre- and post-SH-TL _VO2peak was determined on
an electrically braked cycle ergometer (ERG 900S, Schil-
ler, Switzerland) twice: on one occasion under normobaric
normoxic and the other under normobaric hypoxic [fraction
of inspired O2 (FIO2) 12.5 %] conditions. All tests were
conducted in a laboratory situated at sea level (Orthopaedic
Hospital Valdoltra, Ankaran, Slovenia). Subjects were
transported from the Olympic Sport Centre Planica to the
laboratory on the day of the test, and returned to the
Olympic Sport Centre upon completion of the tests. Each
subject performed an incremental work rate (30 W min-1)
protocol to exhaustion. Baseline values were measured
during a 5-min period of rest preceding the exercise bout.
The initial workload was 60 W and the target pedalling rate
70–80 rpm. The criteria to achieve maximal _VO2 were the
observation of a plateau in _VO2 during the last 15 s of the
trial and the subject’s inability to maintain the required
Eur J Appl Physiol
123
Author's personal copy

cadence. Work rate was interpolated between stages with
the formula: W = work rate of last stage com-
pleted ? [(work rate increment) 9 (time into current
stage/duration of the stage in seconds)] in order to define
the 50 % of peak workload (Wpeak) that should be used for
the constant-work rate tests. Peak workload was calculated
with the same formula assuming that the first 15 s of the
stage were completed.
Constant work rate tests
The three 30-min constant-work rate tests were conducted
on a cycle ergometer (Monark 849-E, Sweden) at the same
time of the day in a counterbalanced order pre- and post-
SH-TL. In these tests, subjects breathed either normobaric
normoxic air or a normobaric hypoxic gas mixture
(inspired fraction of oxygen, FIO2 12.5 %). In the norm-
oxic trial [control test (CT)], the subjects inspired room air
while exercising at 50 % of Wpeak attained in the normoxic_VO2peak test. In the remaining two trials, subjects exercised
in hypoxia either at the same absolute work rate as in CT
[hypoxic absolute work rate test (HAT)] or at 50 % of that
attained in the hypoxic _VO2peak test [hypoxic relative work
rate test (HRT)].
Training
Exercise training was conducted once per day, for 1 h,
5 days per week on a cycle ergometer (Monark 849-E,
Sweden). During each training session, heart rate (HR) was
monitored and stored continuously with a telemetry system
(Hosant, Italy). The work rate was adjusted to maintain HR
at a level observed at a work rate equivalent to 50 % of
normoxic Wpeak before training. The training was con-
ducted under normoxia in the hypoxic facility (b-Cat B.V.,
The Netherlands) of the Olympic Sports Centre Planica.
Analytical methods and equipment
Cardiorespiratory responses during the _VO2peak tests were
obtained at 10-sec intervals with a metabolic cart (Schiller
A-T 104, Switzerland). Subjects breathed through a
respiratory valve that permitted the inhalation of either
atmospheric air, or a decompressed hypoxic mixture from a
200-litre Douglas bag reservoir. During the normoxic trials
subjects inspired ambient air, whereas in the hypoxic trials
they inspired a premixed hypoxic mixture (12.5 % O2,
87.5 % N2). The hypoxic mixture was decompressed and
humidified, by passing it through a small bath of water, and
then accumulated in the Douglas bag. The O2 and CO2
analyzers of the metabolic cart were calibrated with two
different gas mixtures and the volume measurement device
with a 3-litre syringe, before each test, and in accordance
with the manufacturer’s recommendations. In the constant
load trials, minute oxygen uptake and carbon dioxide
elimination were determined from analyses of mixed
expired gases and inspired minute ventilation ( _VE). _VE was
measured with a turbine ventilation module (K520, KL
Engineering Co., CA, USA). The expiratory side was
connected via respiratory hosing to a plexiglas mixing box,
from which a continuous 0.5 l min-1 sample of expired
gases was drawn and analyzed for O2 and CO2 content
using a gas analyzer (1.440D, Servomex Ltd., England).
Core temperature (Tc) was measured by telemetry using
a gastrointestinal pill (Minimitter, USA). Mean skin sur-
face temperature ( �Tsk) was calculated as an arithmetic
average of temperatures (unweighted) measured at four
sites (forearm, chest, thigh and calf) using skin thermistors
(MSR 12, Switzerland). �Tsk can be used as an index of
convective and/or conductive heat loss from the body core
to the skin surface. The ventral fingertip temperature was
also measured to allow the calculation of the difference in
skin temperature between forearm and fingertip (Tskf-f).
Forearm-fingertip skin temperature difference is considered
a valid index of cutaneous vasomotor tone; high values of
Tskf-f indicate vasoconstriction (House and Tipton 2002;
Rubinstein and Sessler 1990).
Local sweating rate ( _msw) was measured with a venti-
lated capsule placed on the forehead. Forced evaporation of
sweat under the capsule (surface area 4.8 cm2) was
achieved by a constant flow of air (1 l min-1) through the
capsule. _msw was estimated from continuous measurements
of the difference between the temperature and the humidity
of inflowing and outflowing air. Air temperature was
measured with thermistors (LM35, National Semiconduc-
tor Corp., Santa Clara, CA, USA) and the relative humidity
with capacitance hygrometers (Valvo air humidity sensor,
Valvo-Philips GmbH, Hamburg, Germany).
Heart rate was monitored with a telemetry system (Polar
Electro Inc., Lake Success NY, USA), and capillary oxy-
haemoglobin saturation (SpO2) using an oximeter (Nellcor
BCI 3110, USA) with the sensor placed on the left index or
middle finger. Subjects were instructed to relax their hands
during measurement of SpO2. The oximeter is accurate to
±2 units across the range of 70–100 % and demonstrates
acceptable resilience to motion artifacts (Langton and
Hanning 1990). Local and general ratings of perceived
exertion (RPEs) were obtained every 5 min using a Borg
scale (6–20).
Cerebral (Cox) and muscle (Mox) oxygenations were
monitored at rest and during exercise by a continuous-wave
near infrared spectroscopy (NIRS; Oxymon MKII Artinis,
The Netherlands). The theory, limitations and the reli-
ability of the measurements during incremental exercise
Eur J Appl Physiol
123
Author's personal copy

with the specific NIRS device have been detailed previ-
ously (Subudhi et al. 2008). Briefly, the NIR light consisted
of two wavelengths (780 and 850 nm) and the micromolar
changes in tissue oxygenation (oxygenated D[HbO2] and
deoxygenated D[HHb] haemoglobin) can be calculated
with the known differential path length factors (DPF) of
4.95 and 5.93 for the muscle (Duncan et al. 1995) and
cerebral tissue (Van der zee et al. 1992), respectively.
Differential path length factors is given by the L/Lo, where
Lo is the distance (cm) between light entering and exiting
the tissue, and L is the average distance travelled by each
photon. In addition, total haemoglobin (D[HbT]), which is
the sum of D[HbO2] and D[HHb] can be used as an index
of regional blood volume (Van Beekvelt et al. 2001). Two
pairs of NIRS probes were used to detect micromolar
changes in the brain and muscle during rest and exercise.
The cerebral probe was positioned on the forehead, at the
level of the left frontal cortex, and the muscle probe above
the vastus lateralis, 15 cm above the proximal line of the
patella and 5 cm lateral to the midline of the thigh. Probes
were kept in place with a plastic spacer (adjusted optode
distance up to 5 cm), which was stabilized on the shaved
and cleaned skin with a double-sided adhesive tape.
Adhesive tape was also used to prevent external light from
entering the measuring area. Skinfolds were taken in the
sagittal plane between emitter and receiver to account for
skin and adipose thickness. Because the exact DPFs were
unknown, cerebral and muscle measurements were nor-
malized to reflect changes from the onset of the exercise
protocol (arbitrarily defined as 0 lL). Therefore, the direct
comparison between cerebral and muscle oxygenation with
the respective data from pulse oximetry was not possible.
Near infrared spectroscopy data were recorded at 50 Hz
and stored for further analysis. All technical considerations
(probe position and stabilization) outlined in reports from
previous studies using the same NIRS device (Subudhi
et al. 2007, 2008) were taken into account, ensuring high
reliability of the NIRS measurements (Subudhi et al. 2007).
Lastly, we used a sufficient optode distance (*4 cm) to
minimize the influence of skin blood flow (Owen-Reece
et al. 1996), a factor that could underestimate the relative
extent of tissue deoxygenation, but could not alter the final
conclusions.
Normobaric hypoxia in five bedrooms and one living
room (total area of *100 m2) was achieved with an oxy-
gen dilution system (b-cat, The Netherlands), based on the
Vacuum-Pressure Swing Adsorption (VPSA) principle.
The oxygen levels in each room were monitored and
recorded continuously with oxygen sensors (Rae PGM-
1100, USA). The hypoxic stimulus during the SH-TL
period was adjusted to maintain SpO2 values between 85
and 90 % during sleep. Each subject was requested to
either wear, or have in close proximity, a personal clip-on
type of oxygen analyzer with an audible alarm, that was
activated in the event that the oxygen level decreased
below the pre-set level. Heart rate and finger-pulse oxim-
etry (Nonin, 3100, USA) were recorded continuously every
night.
Fasted, resting blood samples were drawn in the morn-
ing hours pre- and post-SH-TL. Blood samples were
assayed for haemoglobin, haematocrit, and reticulocytes
using the cytochemical impedance method (Pentra120;
Horiba ABX Diagnostics).
The environmental conditions were kept similar
between pre- and post-SH-TL trials. Namely, environ-
mental temperature ranged from 21.0 (0.4) �C in HAT
before, to 21.9 (0.6) �C in CT after SH-TL, and the relative
humidity from 47 (3) % in HRT before, to 51 (5) % in
HAT after the SH-TL.
Data analysis
A 2-way ANOVA (Condition, Pre–Post) was used to define
the effects of hypoxic relative and absolute work rate and the
SH-TL intervention. A Tukey post-hoc test was employed to
assign the specific differences in the analysis of variance in
case of a significant main effect. Rating of perceived exertion
was analyzed using the non-parametric Friedman test. The
threshold core temperature for sweating and the slopes
relating sweating to the increase in core temperature during
constant-work rate exercise were determined with the least
squares linear regression analysis. The reported values are
means from the individual linear regression analysis, which
were calculated for the linear part of each individual plot.
Values were presented as means (SD) unless indicated
otherwise. Significance level was set at 0.05.
Results
Haematology
Blood haemoglobin [mean (SD) pre-SH-TL: 144 (14) g l-1;
post-SH-TL: 144 (7) g l-1] and haematocrit [pre-SH-TL: 42
(3) %; post-SH-TL: 44 (2) %] remained unchanged after the
SH-TL regimen, but there was a tendency, albeit not sig-
nificant, of an increase in erythrocytes [pre-SH-TL: 5.0 (0.4)
1012 l-1; post-SH-TL: 5.1 (0.4) 1012 l-1] during this period
(p = 0.09).
Resting HR and SpO2 levels
The 5-min resting HR (obtained with the subjects seated
idle on the bike) was significantly reduced after the SH-TL
regimen. Specifically, HR was 84 (8), 86 (13) and 83 (13)
for the pre-SH-TL, and 76 (10), 76 (11) and 78 (6) for the
Eur J Appl Physiol
123
Author's personal copy

post-SH-TL control (CT), hypoxic relative (HRT), and
hypoxic absolute (HAT) trials, respectively. The 5-min
mean SpO2 resting values before each trial were 5 % units
higher in the post-SH-TL hypoxic conditions. Namely, pre-
SH-TL SpO2 values were 97 (2), 88 (3) and 88 (4) %,
whereas post-SH-TL values were 98 (2), 93 (2) and 93
(2) % for the CT, HRT, and HAT trials, respectively.
_VO2peak and Wpeak
Sleep-high train-low improved (p = 0.03) normoxic_VO2peak by 5.5 % from 60.1 (8.5) to 63.5 (8.6) ml kg-1
min-1, but did not significantly affect hypoxic _VO2peak.
Sleep-high train-low improved Wpeak by 11.9 % (p \ 0.01)
in normoxia and by 8.5 % in hypoxia (p = 0.02), with
Wpeak pre- and post-SH-TL amounting to 311 (31) and 348
(36) W, respectively, in normoxia and to 270 (26) and 293
(23) W, respectively, in hypoxia.
Constant work rate exercise
The constant-work rate exercise data were only analyzed
up to the 25th minute of cycling, due to a progressive drop-
out of the subjects after this time point. Each subject
exercised at the same relative intensity pre- and post-SH-
TL. This intensity was set at *50 % of the Wpeak for the
CT and HRT trials, and at *60 % in the HAT trial
(Table 1). The work rate applied during the post-SH-TL
trials was 25–30 W higher so that the same relative
intensity was achieved in each condition (Table 1).
_VO2 and _VE
The mean _VO2 during the trials was *200–300 ml min-1
higher post-SH-TL, however this increment was not sig-
nificant. Both pre- and post-SH-TL, _VO2 were lower
(p \ 0.05) in HRT than in CT and HAT whereas there was
no difference in _VO2 between CT and HAT (Table 1).
Minute ventilation was significantly increased post-SH-TL
for all conditions (Table 1). In both testing periods _VE was
higher in HAT compared with CT and HRT by *12 and
18 l min-1 pre- and post-SH-TL, respectively (Table 1).
HR, SpO2 and RPE
Heart rate was higher in HAT than in CT and HRT both
pre- and post-SH-TL, but no significant differences were
observed between CT and HRT (Table 1). The SpO2 levels
were always lower (p \ 0.05) in the hypoxic trials but SH-
TL elevated (p \ 0.05) SpO2 both in the HAT and HRT
trials (Table 1). Local and general RPEs were higher
(p \ 0.05) during HAT than in CT and HRT, but in any
given exercise condition RPEs were unchanged by SH-TL
(Table 1).
Tc, �Tsk and Tskf-f
The mean core temperature at rest and during exercise and
the relative increase in Tc above resting values, are pre-
sented in Table 2 and Fig. 1, respectively. Core tempera-
ture at rest (baseline) and during the last 5 min of exercise
Table 1 Mean (SD) cardiorespiratory responses and median (range)
local and general rate of perceived exertion (RPE) after 25 min of
steady-state cycling before and after the SH-TL for the control trial
(CT; exercise in normoxia at 50 % of normoxic Wpeak), the hypoxic
relative trial (HRT; exercise in hypoxia at 50 % of hypoxic Wpeak)
and the hypoxic absolute trial (HAT; exercise in hypoxia at the same
absolute WR as in CT) (n = 9)
Pre-SH-TL Post-SH-TL
CT HRT HAT CT HRT HAT
WR (Watts) 149 (7) 127 (6) 149 (7) 180 (9) 153 (8) 180 (9)
% of Wpeak 50 (1) 50 (1) 60 (1)b,c 51.8 (2) 52.0 (3) 61.4 (2)b,c
HR (b min-1) 156 (6) 161 (6) 171 (9)b,c 154 (9) 158 (5) 167 (9)b,c
_VO2 (l min-1) 2.18 (0.4) 1.68 (0.3)b 2.01 (0.4)c 2.39 (0.2) 2.03 (0.4)b 2.25 (0.3)c
_VE (l min-1) 45 (8) 49 (9) 60 (8)b,c 59 (9)a 61 (9)a 78 (11)a,b,c
SpO2 (%) 97.4 (1.1) 73.3 (4.9)b 72.9 (4.1)b 96.6 (1.0) 77.7 (4.1)a,b 77.3 (4.3)a,b
RPE local 12 (7–15) 14 (8–17) 16 (10–19)b 12 (7–15) 14 (10–18) 15 (10–18)b
RPE general 12 (6–14) 14 (9–16) 16 (10–19)b 12 (7–15) 13 (7–16) 15 (10–19)b
_VO2 oxygen consumption, _VE minute ventilation, HR heart rate, SpO2 blood oxygen saturationa Significant difference compared to values before SH-TLb Significant difference compared to CTc Significant difference compared to HRT, p \ 0.05
Eur J Appl Physiol
123
Author's personal copy
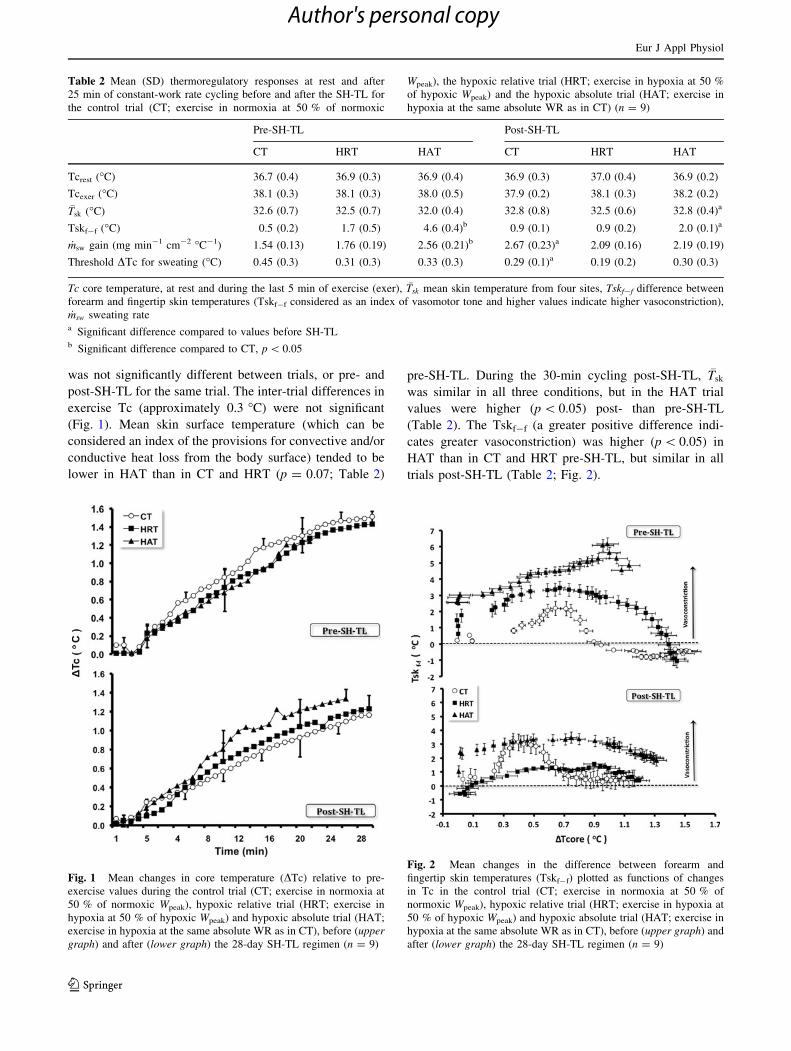
was not significantly different between trials, or pre- and
post-SH-TL for the same trial. The inter-trial differences in
exercise Tc (approximately 0.3 �C) were not significant
(Fig. 1). Mean skin surface temperature (which can be
considered an index of the provisions for convective and/or
conductive heat loss from the body surface) tended to be
lower in HAT than in CT and HRT (p = 0.07; Table 2)
pre-SH-TL. During the 30-min cycling post-SH-TL, �Tsk
was similar in all three conditions, but in the HAT trial
values were higher (p \ 0.05) post- than pre-SH-TL
(Table 2). The Tskf-f (a greater positive difference indi-
cates greater vasoconstriction) was higher (p \ 0.05) in
HAT than in CT and HRT pre-SH-TL, but similar in all
trials post-SH-TL (Table 2; Fig. 2).
Table 2 Mean (SD) thermoregulatory responses at rest and after
25 min of constant-work rate cycling before and after the SH-TL for
the control trial (CT; exercise in normoxia at 50 % of normoxic
Wpeak), the hypoxic relative trial (HRT; exercise in hypoxia at 50 %
of hypoxic Wpeak) and the hypoxic absolute trial (HAT; exercise in
hypoxia at the same absolute WR as in CT) (n = 9)
Pre-SH-TL Post-SH-TL
CT HRT HAT CT HRT HAT
Tcrest (�C) 36.7 (0.4) 36.9 (0.3) 36.9 (0.4) 36.9 (0.3) 37.0 (0.4) 36.9 (0.2)
Tcexer (�C) 38.1 (0.3) 38.1 (0.3) 38.0 (0.5) 37.9 (0.2) 38.1 (0.3) 38.2 (0.2)
�Tsk (�C) 32.6 (0.7) 32.5 (0.7) 32.0 (0.4) 32.8 (0.8) 32.5 (0.6) 32.8 (0.4)a
Tskf-f (�C) 0.5 (0.2) 1.7 (0.5) 4.6 (0.4)b 0.9 (0.1) 0.9 (0.2) 2.0 (0.1)a
_msw gain (mg min-1 cm-2 �C-1) 1.54 (0.13) 1.76 (0.19) 2.56 (0.21)b 2.67 (0.23)a 2.09 (0.16) 2.19 (0.19)
Threshold DSc for sweating (�C) 0.45 (0.3) 0.31 (0.3) 0.33 (0.3) 0.29 (0.1)a 0.19 (0.2) 0.30 (0.3)
Tc core temperature, at rest and during the last 5 min of exercise (exer), �Tsk mean skin temperature from four sites, Tskf-f difference between
forearm and fingertip skin temperatures (Tskf-f considered as an index of vasomotor tone and higher values indicate higher vasoconstriction),
_msw sweating ratea Significant difference compared to values before SH-TLb Significant difference compared to CT, p \ 0.05
Fig. 1 Mean changes in core temperature (DTc) relative to pre-
exercise values during the control trial (CT; exercise in normoxia at
50 % of normoxic Wpeak), hypoxic relative trial (HRT; exercise in
hypoxia at 50 % of hypoxic Wpeak) and hypoxic absolute trial (HAT;
exercise in hypoxia at the same absolute WR as in CT), before (uppergraph) and after (lower graph) the 28-day SH-TL regimen (n = 9)
Fig. 2 Mean changes in the difference between forearm and
fingertip skin temperatures (Tskf-f) plotted as functions of changes
in Tc in the control trial (CT; exercise in normoxia at 50 % of
normoxic Wpeak), hypoxic relative trial (HRT; exercise in hypoxia at
50 % of hypoxic Wpeak) and hypoxic absolute trial (HAT; exercise in
hypoxia at the same absolute WR as in CT), before (upper graph) and
after (lower graph) the 28-day SH-TL regimen (n = 9)
Eur J Appl Physiol
123
Author's personal copy
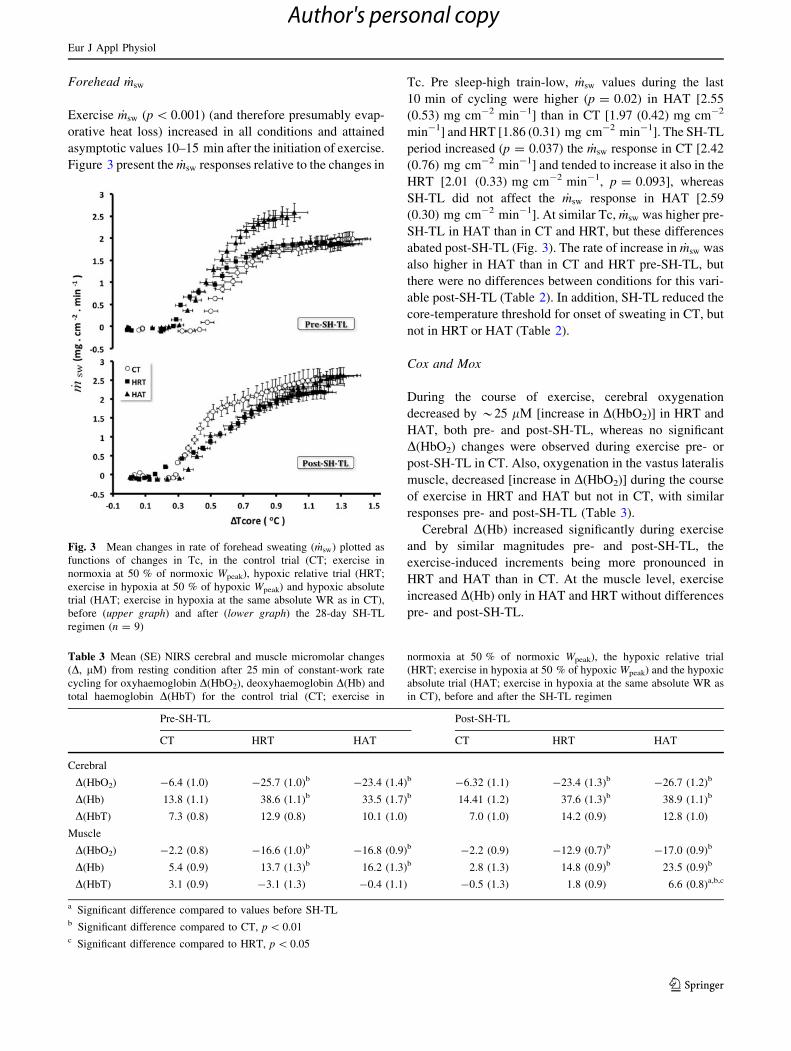
Forehead _msw
Exercise _msw (p \ 0.001) (and therefore presumably evap-
orative heat loss) increased in all conditions and attained
asymptotic values 10–15 min after the initiation of exercise.
Figure 3 present the _msw responses relative to the changes in
Tc. Pre sleep-high train-low, _msw values during the last
10 min of cycling were higher (p = 0.02) in HAT [2.55
(0.53) mg cm-2 min-1] than in CT [1.97 (0.42) mg cm-2
min-1] and HRT [1.86 (0.31) mg cm-2 min-1]. The SH-TL
period increased (p = 0.037) the _msw response in CT [2.42
(0.76) mg cm-2 min-1] and tended to increase it also in the
HRT [2.01 (0.33) mg cm-2 min-1, p = 0.093], whereas
SH-TL did not affect the _msw response in HAT [2.59
(0.30) mg cm-2 min-1]. At similar Tc, _msw was higher pre-
SH-TL in HAT than in CT and HRT, but these differences
abated post-SH-TL (Fig. 3). The rate of increase in _msw was
also higher in HAT than in CT and HRT pre-SH-TL, but
there were no differences between conditions for this vari-
able post-SH-TL (Table 2). In addition, SH-TL reduced the
core-temperature threshold for onset of sweating in CT, but
not in HRT or HAT (Table 2).
Cox and Mox
During the course of exercise, cerebral oxygenation
decreased by *25 lL [increase in D(HbO2)] in HRT and
HAT, both pre- and post-SH-TL, whereas no significant
D(HbO2) changes were observed during exercise pre- or
post-SH-TL in CT. Also, oxygenation in the vastus lateralis
muscle, decreased [increase in D(HbO2)] during the course
of exercise in HRT and HAT but not in CT, with similar
responses pre- and post-SH-TL (Table 3).
Cerebral D(Hb) increased significantly during exercise
and by similar magnitudes pre- and post-SH-TL, the
exercise-induced increments being more pronounced in
HRT and HAT than in CT. At the muscle level, exercise
increased D(Hb) only in HAT and HRT without differences
pre- and post-SH-TL.
Fig. 3 Mean changes in rate of forehead sweating ( _msw) plotted as
functions of changes in Tc, in the control trial (CT; exercise in
normoxia at 50 % of normoxic Wpeak), hypoxic relative trial (HRT;
exercise in hypoxia at 50 % of hypoxic Wpeak) and hypoxic absolute
trial (HAT; exercise in hypoxia at the same absolute WR as in CT),
before (upper graph) and after (lower graph) the 28-day SH-TL
regimen (n = 9)
Table 3 Mean (SE) NIRS cerebral and muscle micromolar changes
(D, lM) from resting condition after 25 min of constant-work rate
cycling for oxyhaemoglobin D(HbO2), deoxyhaemoglobin D(Hb) and
total haemoglobin D(HbT) for the control trial (CT; exercise in
normoxia at 50 % of normoxic Wpeak), the hypoxic relative trial
(HRT; exercise in hypoxia at 50 % of hypoxic Wpeak) and the hypoxic
absolute trial (HAT; exercise in hypoxia at the same absolute WR as
in CT), before and after the SH-TL regimen
Pre-SH-TL Post-SH-TL
CT HRT HAT CT HRT HAT
Cerebral
D(HbO2) -6.4 (1.0) -25.7 (1.0)b -23.4 (1.4)b -6.32 (1.1) -23.4 (1.3)b -26.7 (1.2)b
D(Hb) 13.8 (1.1) 38.6 (1.1)b 33.5 (1.7)b 14.41 (1.2) 37.6 (1.3)b 38.9 (1.1)b
D(HbT) 7.3 (0.8) 12.9 (0.8) 10.1 (1.0) 7.0 (1.0) 14.2 (0.9) 12.8 (1.0)
Muscle
D(HbO2) -2.2 (0.8) -16.6 (1.0)b -16.8 (0.9)b -2.2 (0.9) -12.9 (0.7)b -17.0 (0.9)b
D(Hb) 5.4 (0.9) 13.7 (1.3)b 16.2 (1.3)b 2.8 (1.3) 14.8 (0.9)b 23.5 (0.9)b
D(HbT) 3.1 (0.9) -3.1 (1.3) -0.4 (1.1) -0.5 (1.3) 1.8 (0.9) 6.6 (0.8)a,b,c
a Significant difference compared to values before SH-TLb Significant difference compared to CT, p \ 0.01c Significant difference compared to HRT, p \ 0.05
Eur J Appl Physiol
123
Author's personal copy

There was a tendency (p = 0.07) for an exercise-
induced increase in cerebral D(HbT) in HRT and HAT
compared to in CT pre-, as well as post-SH-TL. Exercise
increased D(HbT) in the muscle (p = 0.04) only in one
condition: HAT post-SH-TL (Table 3).
Discussion
Sleep-high train-low is a training strategy to acclimatize
individuals to altitude. It has been demonstrated to enhance
aerobic performance not only at altitude, but also at sea
level. It is for the later reason that it is commonly incor-
porated in athletes’ training schedules. The present study
investigated the effect of a SH-TL regimen on exercise
temperature regulation. Prior to SH-TL, during exercise at
a given external (absolute) work rate, and for similar core
and skin temperatures, hypoxia potentiated the sweating
response. This acute hypoxia-induced potentiation of
sweating rate was diminished after the SH-TL period. In
acute hypoxia, the augmented sweating, and hence provi-
sions for evaporative heat loss, occurred in concert with a
reduced cutaneous vasodilatation. Thus, SH-TL reversed
the augmentation of the sweating response observed in
acute hypoxia but increased the provisions for conductive
heat loss. Sleep-high train-low improved the normoxic
exercise sweating response, possibly by increasing the
secretion capacity and sensitivity of the sweat glands, as a
consequence of the aerobic training (Nadel et al. 1974;
Roberts et al. 1977; Buono and Sjoholm 1988).
Effects of the SH-TL regimen on exercise heat
dissipation mechanisms
That the pre-SH-TL exercise sweating response was greater
in the hypoxic than the normoxic condition, at a given
absolute work rate, does not necessarily imply that the
physiological pathways responsible for this increase were
triggered by oxygen deficiency per se. Presumably, the
exaggerated sweating response was secondary to the
greater relative work rate imposed by hypoxia (Bocqueraz
et al. 2004; Eiken and Mekjavic 2004; Kacin et al. 2007).
This hypoxia-induced augmentation of sweating rate was
accompanied by higher exercise values for HR, _VE, RPE,
DCox, DMox, TSkf-f in HAT than in CT, suggesting
increased sympathetic response resulting from greater
influence of central command and muscle metabosensitive
drive in HAT. Indeed, central command and muscle me-
taboreflex drive have previously been implicated as causal
factors in the exacerbation of sweating during exercise
(Eiken and Mekjavic 2004; Kondo et al. 1999; Shibasaki
et al. 2003). Furthermore, in agreement with previous
findings (Kacin et al. 2007), the present pre-SH-TL exac-
erbation of the exercise sweating response in HAT was
abated after a SH-TL regimen, as evidenced by the similar
post-SH-TL values of _msw, in CT, HAT and HRT for
identical changes in Tc (Fig. 3). Even though our data does
not allow us to discern the mechanisms underlying this SH-
TL-induced abatement of the exercise sweating response in
hypoxia, it appears likely that it was associated with the
acclimatization to hypoxia rather than with the aerobic
training. Thus, it appears from cross-sectional studies
comparing trained and untrained individuals (Amano et al.
2011; Buono and Sjoholm 1988; Tankersley et al. 1991;
Yamazaki et al. 1994; Yanagimoto et al. 2002) and from
longitudinal training studies (Nadel et al. 1974; Roberts
et al. 1977) that endurance training augments the exercise
sweating response. This notion is compatible with our
finding that SH-TL increased the sweating response during
normoxic exercise, both by lowering the core temperature
threshold for onset of sweating and by increasing the gain
of the sweating response (Table 2 and Fig. 3). Considering
that SH-TL did not alter the normoxic exercise responses
for Cox, Mox, RPE, _VO2, _VE, HR, SpO2, �Tsk or Tc, it
appears likely that the augmentation of the sweating
response was a result of peripheral adaptation of the sweat
glands rather than of increased sympathetic drive. Like-
wise, Buono and Sjoholm (1988) attributed their finding
that the in vivo secretory activity of sweat glands is greater
in aerobically trained than in sedentary subjects, to train-
ing-induced peripheral adaptations.
The question arises then as to what mechanism might
underlie the SH-TL-induced reversal of the initial hypoxic
exaggeration of the exercise sweating response, occurring
presumably despite improved peripheral provisions for
sweat secretion. Possibly, a reduced sudomotor drive dur-
ing hypoxic exercise may have been a consequence of SH-
TL-induced improvement of blood flow in the working
muscles resulting in reduced intramuscular accumulation
of metabolic end products and hence in reduced activation
of muscle metabosensitive afferents (Kondo et al. 1999;
Eiken and Mekjavic 2004); SH-TL increased D(HbT) in
the vastus lateralis, and hence presumably leg muscle blood
flow (Van Beekvelt et al. 2001), during the HAT. Notably,
judging from the increase in �Tsk and decrease in TSkf-f
(indices of cutaneous vasodilatation) it appeared that SH-
TL also increased cutaneous blood flow during HAT. The
notion that both the subsided sweating response and
increased vasodilatory response during HAT after SH-TL
were secondary to the increased muscle blood flow is
supported by previous findings that graded ischemia in the
working leg muscles increases sweating and reduces
cutaneous vasodilatation (Kacin et al. 2005). Another fac-
tor that may have contributed to the SH-TL-induced
Eur J Appl Physiol
123
Author's personal copy

reversation of the exaggerated exercise sweating response
observed during acute exposure to hypoxia is the con-
comitant increase in the exercise hyperpnea. In agreement
with previous studies (Townsend et al. 2005) present
results showed that a main acclimatization effect of SH-TL
is an increased hypoxic ventilatory drive resulting in
improved oxyhaemoglobin saturation, whereas the regimen
appears to have marginal effects on haematological vari-
ables (Ashenden et al. 1999; Gore and Hopkins 2005).
Thus, the SH-TL-dependent increase in arterial oxygen
content during HAT may have reduced the sympathetic
outflow induced by hypoxia in peripheral, carotid chemo-
receptors and in oxygen sensitive cells within the brain
stem (Saito et al. 1988; Stickland et al. 2008; Xie et al.
2001).
Regulation of core temperature
During exercise, the increase in core temperature is a result
of heat production and heat dissipation. The former is a
function of the external load, and the later presumably
modified by the relative work rate (Gonzalez-Alonso et al.
2008). Thus, in the present study, the CT and HAT trials
incorporate the same external work rate, but the HAT trial
induced a greater relative work rate. Similar Tc responses
would therefore be anticipated simply on the basis of the
heat produced. However, since the heat-dissipation
responses are predominantly related to factors associated
with the relative work rate, HAT exercise would be
expected to initiate a greater heat loss. The manner in
which heat is lost via different pathways appears not to be
uniform during hypoxic exercise. Namely, the expected
potentiation of heat loss is only evident in the sweating
response, whereas the decreased vasodilatation reflects a
reduction in the conductive and/or convective heat loss.
The overall heat loss was not determined in the present
study, but presumably it was maintained at similar levels.
This would, to a degree, explain the lack of any difference
in the exercise Tc response during exercise at different
absolute work rates. A novel finding of the present study is
therefore that the factors associated with the relative work
rate do not have the same effect on the heat-loss pathways,
and that the process of acclimatization to hypoxia reduces
this effect. It is unlikely, that central thermoregulatory
mechanisms regulate the magnitude of heat loss, such that
enhancement of heat loss in one pathway (i.e., evaporative)
is compensated by a reduction in another (i.e., conductive).
Nevertheless, in the present study the overall effect of the
balance in the evaporative and conductive heat loss resul-
ted in similar exercise-induced increment in Tc.
From a practical perspective, the reduced sweating rate
during exercise in hypoxia after a SH-TL regimen might be
beneficial for alpinists, reducing any problems with alti-
tude-induced dehydration. Moreover, it appears that ath-
letes who have undertaken a SH-TL regimen to improve
endurance performance at sea level will not risk suffering
from reduced sweating rate since the normal increase in
exercise sweating rate following aerobic training seems to
be preserved also after SH-TL, provided the exercise bout
is executed under normoxic conditions.
Methodological limitations
The design of the present study, allows for analyses of the
combined effects of hypoxic acclimatization and aerobic
training, but does not permit one to distinguish the indi-
vidual contributions from these two components of the SH-
TL regimen. Addition of another condition, for instance a
sleep-low train-low condition, would make it possible to
single out effects contributed by the aerobic training per se.
Other delimitations of the present study include the lack of
direct measurement of cutaneous blood flow and the single
site for determining sweating rate. Skin temperature
between forearm and fingertip was used as an index of
vasoconstriction; although it does not necessarily reflect
overall cutaneous blood flow it has been shown to provide
a robust and reliable qualitative index of cutaneous vaso-
motor responses (House and Tipton 2002, Rubinstein and
Sessler 1990). Sweat secretion was only measured on the
forehead. Considering the substantial regional differences
in sweating rate, more sites of measurements are warranted
to make general assumptions regarding sweating responses
(Havenith and van Middendorp 1990; Kondo et al. 1998;
Machado-Moreira et al. 2008). Finally, to make it possible
to generalize the present findings, it is necessary to also
perform experiments in female subjects.
Thus in summary it appears that, a 28-day SH-TL reg-
imen can reduce the hypoxic exacerbation of the exercise
sweating response via higher convective heat loss and
improve the sweating response during normoxic exercise.
As regards mechanisms underlying these SH-TL-induced
changes in exercise heat dissipation, the present data on
blood-volume distribution and oxygenation of the cerebral
and exercising muscle tissues do not permit firm conclu-
sions, but are compatible with the notion that improved
muscle blood flow may play a role. Whether these SH-TL
responses are attributable to the effect of hypoxic accli-
matization or the aerobic training remains to be settled.
Acknowledgments We would like to thank all the subjects for their
cooperation and patience. We are grateful to Bogomir Vrhovec and
Mojca Amon for their valuable assistance. This study was supported
by a grant from the Slovenian Research Agency (ARRS grant no. L7-
2413) to Igor B. Mekjavic, and from b-Cat B.V. (The Netherlands) to
Igor B. Mekjavic and Ola Eiken.
Eur J Appl Physiol
123
Author's personal copy

Conflict of interest The authors state that there is no personal
conflict of interest in the present study.
References
Ainslie PN, Hamlin M, Hellemans J, Rasmussen P, Ogoh S (2008)
Cerebral hypoperfusion during hypoxic exercise following two
different hypoxic exposures: independence from changes in
dynamic autoregulation and reactivity. Am J Physiol Regul
Integr Comp Physiol 295:R1613–R1622
Amano T, Ichinose M, Koga S, Inoue Y, Nishiyasu T, Kondo N
(2011) Sweating responses and the muscle metaboreflex under
mildly hyperthermic conditions in sprinters and distance runners.
J Appl Physiol 111:524–529
Ashenden MJ, Gore CJ, Dobson GP, Hahn AG (1999) ‘‘Live high,
train low’’ does not change the total haemoglobin mass of male
endurance athletes sleeping at a simulated altitude of 3,000 m
for 23 nights. Eur J Appl Physiol Occup Physiol 80:479–484
Bocqueraz O, Koulmann N, Guigas B, Jimenez C, Melin B (2004)
Fluid-regulatory hormone responses during cycling exercise in
acute hypobaric hypoxia. Med Sci Sports Exerc 36:1730–1736
Buono MJ, Sjoholm NT (1988) Effect of physical training on
peripheral sweat production. J Appl Physiol 65:811–814
Duncan A, Meek JH, Clemence M, Elwell CE, Tyszczuk L, Cope M,
Delpy DT (1995) Optical pathlength measurements on adult
head, calf and forearm and the head of the newborn infant using
phase resolved optical spectroscopy. Phys Med Biol 40:295–304
Eiken O, Mekjavic IB (2004) Ischaemia in working muscles
potentiates the exercise-induced sweating response in man. Acta
Physiol Scand 181:305–311
Gonzalez-Alonso J, Eiken O, Mekjavic IB (2008) A critical core
temperature and the significance of absolute and relative work
rate. In: Groeller Herbert, Taylor NAS (eds) Physiological bases
of human performance during work and exercise. Elsevier,
Sydney, pp 481–485
Gore CJ, Hopkins WG (2005) Counterpoint: positive effects of
intermittent hypoxia (live high:train low) on exercise perfor-
mance are not mediated primarily by augmented red cell volume.
J Appl Physiol 99:2055–2057 Discussion 2057–2058
Greenleaf JE, Greenleaf J, Card DH, Saltin B (1969) Exercise-
temperature regulation in man during acute exposure to simu-
lated altitude. J Appl Physiol 26:290–296
Havenith G, van Middendorp H (1990) The relative influence of
physical fitness, acclimatization state, anthropometric measures
and gender on individual reactions to heat stress. Eur J Appl
Physiol Occup Physiol 61(5–6):419–427
House JR, Tipton MJ (2002) Using skin temperature gradients or skin
heat flux measurements to determine thresholds of vasoconstric-
tion and vasodilatation. Eur J Appl Physiol 88:141–145
Kacin A, Golja P, Eiken O, Tipton MJ, Gorjanc J, Mekjavic IB (2005)
Human temperature regulation during cycling with moderate leg
ischaemia. Eur J Appl Physiol 95:213–220
Kacin A, Golja P, Eiken O, Tipton MJ, Mekjavic IB (2007) The
influence of acute and 23 days of intermittent hypoxic exposures
on the exercise-induced forehead sweating response. Eur J Appl
Physiol 99:557–566
Kondo N, Takano S, Aoki K, Shibasaki M, Tominaga H, Inoue Y
(1998) Regional differences in the effect of exercise intensity on
thermoregulatory sweating and cutaneous vasodilation. Acta
Physiol Scand 164:71–78
Kondo N, Tominaga H, Shibasaki M, Aoki K, Koga S, Nishiyasu T
(1999) Modulation of the thermoregulatory sweating response to
mild hyperthermia during activation of the muscle metaboreflex
in humans. J Physiol 515(Pt 2):591–598
Langton JA, Hanning CD (1990) Effect of motion artefact on pulse
oximeters: evaluation of four instruments and finger probes. Br J
Anaesth 65:564–570
Machado-Moreira CA, Smith FM, van den Heuvel AM, Mekjavic IB,
Taylor NA (2008) Sweat secretion from the torso during
passively-induced and exercise-related hyperthermia. Eur J Appl
Physiol 104:265–270
Mekjavic IB, Eiken O (2006) Contribution of thermal and nonthermal
factors to the regulation of body temperature in humans. J Appl
Physiol 100:2065–2072
Nadel ER, Pandolf KB, Roberts MF, Stolwijk JA (1974) Mechanisms
of thermal acclimation to exercise and heat. J Appl Physiol
37:515–520
Owen-Reece H, Elwell CE, Wyatt JS, Delpy DT (1996) The effect of
scalp ischaemia on measurement of cerebral blood volume by
near-infrared spectroscopy. Physiol Meas 17:279–286
Roberts MF, Wenger CB, Stolwijk JA, Nadel ER (1977) Skin blood
flow and sweating changes following exercise training and heat
acclimation. J Appl Physiol 43:133–137
Rubinstein EH, Sessler DI (1990) Skin-surface temperature gradients
correlate with fingertip blood flow in humans. Anesthesiology
73:541–545
Saito M, Mano T, Iwase S, Koga K, Abe H, Yamazaki Y (1988)
Responses in muscle sympathetic activity to acute hypoxia in
humans. J Appl Physiol 65:1548–1552
Shibasaki M, Kondo N, Crandall CG (2001) Evidence for metabo-
receptor stimulation of sweating in normothermic and heat-
stressed humans. J Physiol 534:605–611
Shibasaki M, Secher NH, Selmer C, Kondo N, Crandall CG (2003)
Central command is capable of modulating sweating from non-
glabrous human skin. J Physiol 553:999–1004
Stickland MK, Morgan BJ, Dempsey JA (2008) Carotid chemore-
ceptor modulation of sympathetic vasoconstrictor outflow during
exercise in healthy humans. J Physiol 586:1743–1754
Subudhi AW, Dimmen AC, Roach RC (2007) Effects of acute
hypoxia on cerebral and muscle oxygenation during incremental
exercise. J Appl Physiol 103:177–183
Subudhi AW, Lorenz MC, Fulco CS, Roach RC (2008) Cerebrovas-
cular responses to incremental exercise during hypobaric
hypoxia: effect of oxygenation on maximal performance. Am J
Physiol Heart Circ Physiol 294:H164–H171
Tankersley CG, Smolander J, Kenney WL, Fortney SM (1991)
Sweating and skin blood flow during exercise: effects of age and
maximal oxygen uptake. J Appl Physiol 71:236–242
Townsend NE, Gore CJ, Hahn AG, Aughey RJ, Clark SA, Kinsman
TA, McKenna MJ, Hawley JA, Chow CM (2005) Hypoxic
ventilatory response is correlated with increased submaximal
exercise ventilation after live high, train low. Eur J Appl Physiol
94:207–215
Van Beekvelt MC, Colier WN, Wevers RA, Van Engelen BG (2001)
Performance of near-infrared spectroscopy in measuring local O2
consumption and blood flow in skeletal muscle. J Appl Physiol
90:511–519
van der Zee P, Cope M, Arridge SR, Essenpreis M, Potter LA,
Edwards AD, Wyatt JS, McCormick DC, Roth SC, Reynolds EO
et al (1992) Experimentally measured optical path lengths for the
adult head, calf and forearm and the head of the newborn infant
as a function of inter optode spacing. Adv Exp Med Biol
316:143–153
Wang JS, Wu MH, Mao TY, Fu TC, Hsu CC (2010) Effects of
normoxic and hypoxic exercise regimens on cardiac, muscular
and cerebral hemodynamics suppressed by severe hypoxia in
humans. J Appl Physiol 109:219–229
Eur J Appl Physiol
123
Author's personal copy

Xie A, Skatrud JB, Puleo DS, Morgan BJ (2001) Exposure to hypoxia
produces long-lasting sympathetic activation in humans. J Appl
Physiol 91:1555–1562
Yamazaki F, Fujii N, Sone R, Ikegami H (1994) Mechanisms of
potentiation in sweating induced by long-term physical training.
Eur J Appl Physiol Occup Physiol 69:228–232
Yanagimoto S, Aoki K, Horikawa N, Shibasaki M, Inoue Y,
Nishiyasu T, Kondo N (2002) Sweating response in physically
trained men to sustained handgrip exercise in mildly hyperther-
mic condition. Acta Physiol Scand 174:31–39
Eur J Appl Physiol
123
Author's personal copy
Related Documents











