JMB—MS 424 Cust. Ref. No. RH 91/94 [SGML] J. Mol. Biol. (1995) 248, 79–96 Evolution of the Phycobiliproteins Kirk E. Apt*, Jackie L. Collier and Arthur R. Grossman Carnegie Institute of Amino acid sequence alignments and phylogenetic analyses have been used Washington, Department of to examine the relationships among 100 phycobiliprotein sequences. The alignments revealed a number of highly conserved amino acid residues that Plant Biology, 290 Panama St., Stanford, CA 94305 are involved in chromophore attachment and conformation, a-b interactions and phycobilisome assembly. The phylogenetic analysis confirmed that the U.S.A. phycobiliprotein subfamilies, previously classified by their biochemical and spectroscopic properties, also formed coherent evolutionary groups. The a and b subunits formed two distinct evolutionary lines that originate from a common ancestor. The pattern of divergence among the a subfamilies was identical to that of the b subfamilies, strongly suggesting that the a and b subunits of each phycobiliprotein type have coevolved. The phylogenetic data support a monophyletic separation of the eukaryotic sequences from the extant cyanobacterial sequences. The eukaryotic phycoerythrins appeared more closely related to the marine Synechococcus phycoerythrins than to the other cyanobacterial phycoery- thrins. The cryptophyte phycobiliproteins formed a monophyletic group within the rhodophyte lineage. In conjunction with other phylogenetic markers, the analysis of additional phycobiliprotein sequences may help to further resolve the relationships among phycobiliprotein-containing organisms. Keywords: allophycocyanin, cyanobacteria; phycocyanin; phycoerythrin; *Corresponding author red algae Introduction Phycobiliproteins are a family of light-harvesting macromolecules that function as components of the photosynthetic apparatus in cyanobacteria and several groups of eukaryotic algae including the red algae, cryptomonads and glaucophytes (reviewed by MacColl & Guard-Friar, 1987; Zuber, 1987; Glazer, 1987, 1989; Tandeau de Marsac et al ., 1990; Bryant, 1991; Holzwarth, 1991; Mo ¨rschel, 1991; Grossman et al ., 1993; Sidler, 1994). Phycobiliproteins may comprise 60% of the soluble protein in these cells, and their main function is to trap light energy in the 495 to 650nm wavelength range and transfer it to chlorophyll a of the photosynthetic reaction centers. Phycobiliproteins can be divided into three major groups based on their spectral properties (Table 1): phycoerythrin (PE), A max = 560 nm, phycocyanin (PC), A max = 620 nm, and allophycocyanin (AP), A max = 650 nm. Each of the different phyco- biliproteins is composed of two non-identical polypeptide subunits (a and b), which contain one or more covalently linked open-chain tetrapyrrole chromophores. The chromophores, known as phyco- bilins, are covalently attached to the phyco- biliproteins by either one or occasionally two cysteinyl thioether linkages. There are several structurally distinct phycobilin chromophores with different spectroscopic properties. When covalently attached to phycobiliproteins (Table 1) the chro- mophores phycourobilin (PUB), phycobiliviolin (PXB), phycoerythrobilin (PEB) and phycocyanobilin (PCB) have absorbance maxima at approximately 498 nm, 568 nm, 535 to 567 nm and 620 to 660 nm, respectively. Cryptomonad phycobiliproteins have additional novel chromophore types (Wedemayer et al ., 1991, 1992). The phycobiliproteins are further divided into a number of specific types (Table 1) that are characterized by different numbers, combi- nations and positions of the chromophores. The 3-D structure of the phycobiliproteins (Schirmer et al ., 1985, 1986, 1987; Duerring et al ., 1990, Abbreviations used: AP-B, allophycocyanin-B; AP, allophycocyanin; AP-18, b-allophycocyanin-like polypeptide; CCA, complementary chromatic adap- tation; LCM, high molecular weight core-membrane linker polypeptide containing the TEA phycobiliprotein domain; PC, phycocyanin; PE, phycoerythrin; PEB, phycoerythrobilin; PEC, phycoerythrocyanin; PUB, phycourobilin; PXB, phycoviolibilin; RCI, rescaled consistency index; TEA, high molecular weight terminal energy acceptor phycobiliprotein domain. 0022–2836/95/160079–18 $08.00/0 7 1995 Academic Press Limited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JMB—MS 424 Cust. Ref. No. RH 91/94 [SGML]
J. Mol. Biol. (1995) 248, 79–96
Evolution of the Phycobiliproteins
Kirk E. Apt*, Jackie L. Collier and Arthur R. Grossman
Carnegie Institute of Amino acid sequence alignments and phylogenetic analyses have been usedWashington, Department of to examine the relationships among 100 phycobiliprotein sequences. The
alignments revealed a number of highly conserved amino acid residues thatPlant Biology, 290 PanamaSt., Stanford, CA 94305 are involved in chromophore attachment and conformation, a-b interactions
and phycobilisome assembly. The phylogenetic analysis confirmed that theU.S.A.phycobiliprotein subfamilies, previously classified by their biochemical andspectroscopic properties, also formed coherent evolutionary groups. The aand b subunits formed two distinct evolutionary lines that originate from acommon ancestor. The pattern of divergence among the a subfamilies wasidentical to that of the b subfamilies, strongly suggesting that the a and bsubunits of each phycobiliprotein type have coevolved.
The phylogenetic data support a monophyletic separation of theeukaryotic sequences from the extant cyanobacterial sequences. Theeukaryotic phycoerythrins appeared more closely related to the marineSynechococcus phycoerythrins than to the other cyanobacterial phycoery-thrins. The cryptophyte phycobiliproteins formed a monophyletic groupwithin the rhodophyte lineage. In conjunction with other phylogeneticmarkers, the analysis of additional phycobiliprotein sequences may help tofurther resolve the relationships among phycobiliprotein-containingorganisms.
Keywords: allophycocyanin, cyanobacteria; phycocyanin; phycoerythrin;*Corresponding author red algae
Introduction
Phycobiliproteins are a family of light-harvestingmacromolecules that function as components of thephotosynthetic apparatus in cyanobacteria andseveral groups of eukaryotic algae including the redalgae, cryptomonads and glaucophytes (reviewed byMacColl & Guard-Friar, 1987; Zuber, 1987; Glazer,1987, 1989; Tandeau de Marsac et al., 1990; Bryant,1991; Holzwarth, 1991; Morschel, 1991; Grossmanet al., 1993; Sidler, 1994). Phycobiliproteins maycomprise 60% of the soluble protein in these cells,and their main function is to trap light energy in the495 to 650 nm wavelength range and transfer it tochlorophyll a of the photosynthetic reaction centers.
Phycobiliproteins can be divided into three major
groups based on their spectral properties (Table 1):phycoerythrin (PE), Amax = 560 nm, phycocyanin(PC), Amax = 620 nm, and allophycocyanin (AP),Amax = 650 nm. Each of the different phyco-biliproteins is composed of two non-identicalpolypeptide subunits (a and b), which contain one ormore covalently linked open-chain tetrapyrrolechromophores. The chromophores, known as phyco-bilins, are covalently attached to the phyco-biliproteins by either one or occasionally twocysteinyl thioether linkages. There are severalstructurally distinct phycobilin chromophores withdifferent spectroscopic properties. When covalentlyattached to phycobiliproteins (Table 1) the chro-mophores phycourobilin (PUB), phycobiliviolin(PXB), phycoerythrobilin (PEB) and phycocyanobilin(PCB) have absorbance maxima at approximately498 nm, 568 nm, 535 to 567 nm and 620 to 660 nm,respectively. Cryptomonad phycobiliproteins haveadditional novel chromophore types (Wedemayeret al., 1991, 1992). The phycobiliproteins are furtherdivided into a number of specific types (Table 1) thatare characterized by different numbers, combi-nations and positions of the chromophores.
The 3-D structure of the phycobiliproteins(Schirmer et al., 1985, 1986, 1987; Duerring et al., 1990,
Abbreviations used: AP-B, allophycocyanin-B; AP,allophycocyanin; AP-18, b-allophycocyanin-likepolypeptide; CCA, complementary chromatic adap-tation; LCM, high molecular weight core-membranelinker polypeptide containing the TEA phycobiliproteindomain; PC, phycocyanin; PE, phycoerythrin; PEB,phycoerythrobilin; PEC, phycoerythrocyanin; PUB,phycourobilin; PXB, phycoviolibilin; RCI, rescaledconsistency index; TEA, high molecular weight terminalenergy acceptor phycobiliprotein domain.
0022–2836/95/160079–18 $08.00/0 7 1995 Academic Press Limited
JMB—MS 424
Phycobiliprotein Evolution80
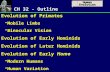
Table 1Subunit type and chromophore composition for phycobiliproteins with known amino acidsequencesPhycobiliprotein Subunit Chromophore position and typegroup abbr. 51–62 78 88 128 146 178
AllophycocyaninsTEA PCBaAP PCBbAP PCB
aAP-B PCBbAP-18 PCB
PhycocyaninsaPC PCBbPC PCB PCBaR-PC PCBbR-PC PCB PEBaR-PCII PEBbR-PCII PCB PEBaPEC PXBbPEC PCB PCB
PhycoerythrinsaPE PEB PEBbPE PEB PEB PEB
WH8020 aPEI PEB PEBWH8020 bPEI PEB PEB PEBWH8020 aPEII PUB PEB PEBWH8020 bPEII PUB PEB PEBWH8103 aPEII PUB PUB PUBWH8103 bPEII PUB PEB PEB
aB-PE PEB PEBbB-PE PEB PEB PEBaR-PE PEB PEBbR-PE PUB PEB PEB
Absorption maximum for chromophores: PCB 620–660 nm, PEB 535 to 567 nm, PXB 568 nm, and PUB498 nm.
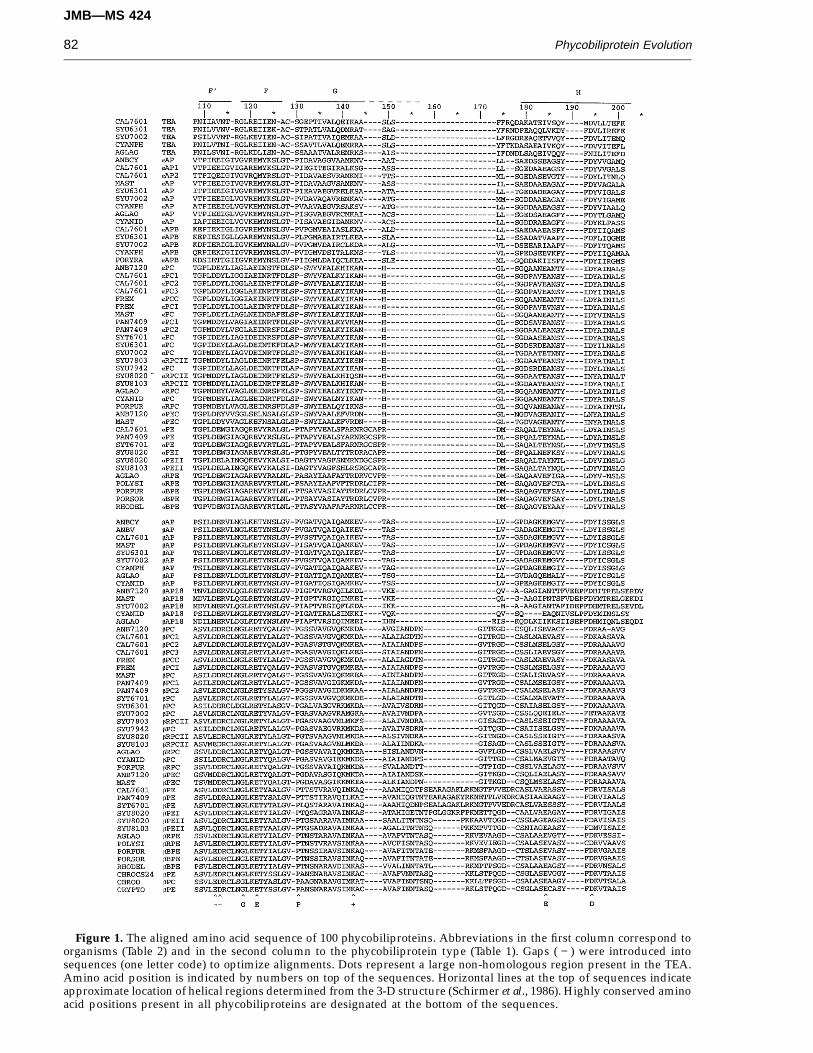
1991; Ficner et al., 1992; Ficner & Huber, 1993) revealsimilar tertiary structures for the a and b subunits.Each of the phycobiliprotein subunits has ninea-helical domains (marked in Figure 1) separated byirregular loops. The X and E helices of one subunitand the Y and A helices of the other subunit areinvolved in extensive interactions and help stabilizethe ab heterodimer (referred to as a monomer, byconvention). The phycobiliprotein trimers ((ab)3) arecomposed of three identical ab monomers arrangedaround a 3-fold symmetry axis. The hexameric ((ab)6)aggregates are composed of two trimers associatedin a face to face arrangement. The trimers andhexamers are disc-like structures, which possess acentral cavity. The F' helix of the a subunit and theF' and F helices of the b subunit protrude into thiscentral cavity with the b subunit F helix being themore prominent.
Phycobiliprotein discs (trimers and hexamers) areassembled with the aid of linker polypeptides intophycobilisomes, which are organized into twodistinct structural domains, the core and the rods.The core, composed of discs of AP, forms a physicalconnection with the outer surface of the thylakoidmembranes and is often associated with photosys-tem II. Radiating from the core is a series of rods thatare composed of stacked phycobiliprotein discs.Discs proximal to the core are of PC, while thosedistal to the core are of PE (or PEC in somecyanobacteria). Measurements of energy transferhave shown that light energy absorbed by PE is
sequentially transferred to PC, then AP and finallyis passed primarily through the phycobilisometerminal energy acceptor (TEA, the phycobiliprotein-like domain of the large ‘‘anchor’’ polypeptide orLCM) to chlorophyll a. This transfer of energy occurswith an efficiency approaching 100% in vivo(reviewed by Glazer, 1989).
Linker polypeptides are believed to bind in thecentral cavity of the trimers and hexamers. Differentlinker polypeptides are specifically associated witheach type of phycobiliprotein hexamer. Theyfunction to stabilize the phycobilisome and optimizeits absorbance and energy transfer characteristics.
It has long been recognized that there arenumerous similarities among the different phyco-biliprotein subunits, leading to the proposal that thedifferent subunits belong to a protein family that hasevolved from a single ancestral protein (Glazer et al.,1976; Glazer, 1980). This study was conducted toprovide a detailed examination of the evolutionaryrelationships within and among the phycobiliproteingroups utilizing sequence data from 100 differentphycobiliprotein subunits (Table 2).
Results
The alignments
The alignment of 100 phycobiliprotein subunitsis given in Figure 1. These alignments consider

JMB—MS 424
Phycobiliprotein Evolution 81
Figure 1(A).
JMB—MS 424
Phycobiliprotein Evolution82
Figure 1. The aligned amino acid sequence of 100 phycobiliproteins. Abbreviations in the first column correspond toorganisms (Table 2) and in the second column to the phycobiliprotein type (Table 1). Gaps ( − ) were introduced intosequences (one letter code) to optimize alignments. Dots represent a large non-homologous region present in the TEA.Amino acid position is indicated by numbers on top of the sequences. Horizontal lines at the top of sequences indicateapproximate location of helical regions determined from the 3-D structure (Schirmer et al., 1986). Highly conserved aminoacid positions present in all phycobiliproteins are designated at the bottom of the sequences.

JMB—MS 424 PAGE 83
Tabl
e2
The
list
ofkn
own
phyc
obili
prot
ein
subu
nit
sequ
ence
sA
glao
=A
glao
tham
nion
negl
ectu
ma
Pan7
409
=P
seud
anab
aena
PCC
7409
TE
A(A
pt&
Gro
ssm
an,1
993b
)aP
C1
(Dub
bs&
Bry
ant,
1993
)aA
P(A
pt&
Gro
ssm
an,1
993a
)bP
C1
(Dub
bs&
Bry
ant,
1993
)bA
P(A
pt&
Gro
ssm
an,1
993a
)aP
C2
(Dub
bs&
Bry
ant,
1993
)bA
P18
(Apt
&G
ross
man
,199
3c)
bPC
2(D
ubbs
&B
ryan
t,19
93)
aRPC
(Apt
&G
ross
man
,199
3a)
aPE
(Dub
bs&
Byr
ant,
1987
;Dub
bs&
Bry
ant,
1991
)bR
PC(A
pt&
Gro
ssm
an,1
993a
)bP
E(D
ubbs
&B
ryan
t,19
91)
aRPE
(Apt
&G
ross
man
,199
3a)
Poly
si=
Poly
siph
onia
bold
iia
bRPE
(Apt
&G
ross
man
,199
3a)
aRPE
(Roe
ll&
Mor
se,1
993)
Anb
v=
Ana
baen
ava
rabi
lis(A
naba
ena
PCC
7118
)bR
PE(R
oell
&M
orse
,199
3)bA
P(D
elan
geet
al.,
1981
)Po
rpur
=Po
rphy
ridi
umpu
rpur
eum
(P.c
ruen
tum
)a
Anb
cy=
Ana
baen
acy
lindr
ica
aRPC
(Duc
ret
etal
.,19
94)
aAP
(Min
ami
etal
.,19
85)
bRPC
(Duc
ret
etal
.,19
94)
bAP
(Min
ami
etal
.,19
85)
aBPE
(Sid
ler
etal
.,19
89)
Anb
7120
=A
naba
ena
PCC
7120
bBPE
(Sid
ler
etal
.,19
89)
bAP1
8(L
.War
ner,
per
sona
lco
mm
unic
atio
n)Po
rsor
=Po
rphy
ridi
umso
rdid
uma
aPC
(Bel
knap
&H
asel
korn
,198
7)aB
PE(F
icne
ret
al.,
1992
)bP
C(B
elkn
ap&
Has
elko
rn,1
987)
bBPE
(Fic
ner
etal
.,19
92)
aPE
C(S
wan
son
etal
.,19
92)
Pory
ra=
Porp
hyra
purp
urea
a
bPE
C(S
wan
son
etal
.,19
92)
aAPB
(M.R
eith
,per
sona
lco
mm
unic
atio
n)C
al76
01=
Cal
othr
ixPC
C76
01R
hode
l=R
hode
llavi
olac
eaa
TE
A(H
oum
ard
etal
.,19
90)
aBPE
(Ber
nard
etal
.,19
92)
aAP1
(Hou
mar
det
al.,
1988
b)bB
PE(B
erna
rdet
al.,
1992
)aA
P2(H
oum
ard
etal
.,19
88b)
Syt6
701
=Sy
nech
ocys
tis
PCC
6701
bAP
(Hou
mar
det
al.,
1988
b)aP
C(A
nder
son
&G
ross
man
,199
0a)
aAPB
(Hou
mar
det
al.,
1988
a)bP
C(A
nder
son
&G
ross
man
,199
0a)
aPC
1(M
azel
etal
.,19
88)
aPE
(And
erso
n&
Gro
ssm
an,1
990b
)aP
C2
(Cap
uano
etal
.,19
88)
bPE
(And
erso
n&
Gro
ssm
an,1
990b
)aP
C3
(Maz
el&
Mar
liere
1989
)Sy
t680
3=
Syne
choc
ysti
sPC
C68
03bP
C1
(Maz
elet
al.,
1988
)aA
P(S
uet
al.,
1992
)b
bPC
2(C
apua
noet
al.,
1988
)bA
P(S
uet
al.,
1992
)b
bPC
3(M
azel
&M
arlie
re,1
989)
Syt6
714
=Sy
nech
ocys
tis
PCC
6714
aPE
(Maz
elet
al.,
1986
;Sid
ler
etal
.,19
86)
aAP
(Dim
agno
&H
asel
korn
,199
3)b
bPE
(Maz
elet
al.,
1986
;Sid
ler
etal
.,19
86)
bAP
(Dim
agno
&H
asel
korn
,199
3)b
Chr
oo=
Chr
oom
onas
sp.a
Syu6
301
=Sy
nech
ococ
cus
PCC
6301
(Ana
cyst
isni
dula
nsU
TE
X62
5)aP
C1
(Sid
ler
etal
.,19
87;S
idle
ret
al.,
1990
)bT
EA
(Cap
uano
etal
.,19
91)
aPC
2(S
idle
ret
al.,
1987
;Sid
ler
etal
.,19
90)b
aAP
(Hou
mar
det
al.,
1986
)bP
C(S
idle
ret
al.,
1987
;Sid
ler
etal
.,19
90)
aAPB
(Sut
eret
al.,
1987
)C
hroc
s24
=C
hroo
mon
asC
S24a
bAP
(Hou
mar
det
al.,
1986
)aP
E3
(Jen
kins
etal
.,19
90)b
aPC
(Wal
shet
al.,
1980
;Lin
d,1
988)
bPE
(God
ovac
-Zim
mer
man
net
al.,
1992
)bP
C(F
reid
enre
ich
etal
.,19
78;L
ind
,198
8)C
rypt
o=
Cry
ptom
onas
Fa
Syu7
002
=Sy
nech
ococ
cus
PCC
7002
(Agm
enel
lum
quad
rupl
icat
um)
bPE
(Rei
th&
Dou
glas
,199
0)T
EA
(Bry
ant,
1991
)C
yani
d=
Cya
nidi
umca
ldar
ium
aaA
P(B
ryan
t,19
91)
cont
inue
dov
erle
af

JMB—MS 424 PAGE 84
Tabl
e2—
cont
inue
daA
P(O
ffne
r&
Trox
ler,
1983
)aA
PB(B
ryan
t,19
91)
bAP
(Off
ner
&Tr
oxle
r,19
83;L
iu&
Trox
ler,
1993
)bA
P(B
ryan
t,19
91)
bAP1
8(V
alen
tin
etal
.,19
92)
bAP1
8(B
ryan
t,19
91)
aPC
(Off
ner
etal
.,19
81)
aPC
(de
Lor
imer
etal
.,19
84;P
ilot
&Fo
x,19
84)
bPC
(Tro
xler
etal
.,19
81)
bPC
(de
Lor
imer
etal
.,19
84;P
ilot
&Fo
x,19
84)
Cya
nph
=C
yano
phor
apa
rado
xaa
Syu7
803
=Sy
nech
ococ
cus
WH
7803
TE
A(B
ryan
t,19
88)
aRPC
II(W
ilson
etal
.,19
91)
aAP
(Bry
ant
etal
.,19
85)
bRPC
II(W
ilson
etal
.,19
91)
bAP
(Bry
ant
etal
.,19
85)
aPE
I(N
ewm
anet
al.,
1994
)b
bAP
(Bry
ant
etal
.,19
85)
bPE
I(N
ewm
anet
al.,
1994
)b
aAPB
(Mic
halo
wsk
iet
al.,
1990
)Sy
u794
2=
Syne
choc
occu
sPC
C79
42(A
nacy
stis
nidu
lans
R2)
Frem
=Fr
emye
lladi
plos
ipho
nFD
33aP
C(L
auet
al.,
1987
a)aP
Cc
(Con
ley
etal
.,19
88)
bPC
(Lau
etal
.,19
87b)
aPC
i(C
onle
yet
al.,
1988
)Sy
u802
0=
Syne
choc
occu
sW
H80
20bP
Cc
(Con
ley
etal
.,19
88)
aRPC
II(d
eL
orim
ier
etal
.,19
93)
bPC
i(C
onle
yet
al.,
1988
)bR
PCII
(de
Lor
imie
ret
al.,
1993
)M
ast=
Mas
tigo
clad
usla
min
osus
(Fis
cher
ella
PCC
7603
)aP
EI
(Wilb
anks
&G
laze
r,19
93)
aAP
(Sid
ler
etal
.,19
81)
bPE
I(W
ilban
ks&
Gla
zer,
1993
)bA
P(S
idle
ret
al.,
1981
)aP
EII
(Wilb
anks
etal
.,19
91)
bAP1
8(R
umbe
liet
al.,
1987
)bP
EII
(Wilb
anks
etal
.,19
91)
aPC
(Fra
nket
al.,
1978
)Sy
u810
3=
Syne
choc
occu
sW
H81
03bP
C(F
rank
etal
.,19
78;S
chir
mer
etal
.,19
87)
aRPC
II(d
eL
orim
ier
etal
.,19
93)
aPE
C(F
uglis
talle
ret
al.,
1983
;Ebe
rlei
n&
Kuf
er,1
990)
bRPC
II(d
eL
orim
ier
etal
.,19
93)
bPE
C(F
uglis
talle
ret
al.,
1983
;Ebe
rlei
n&
Kuf
er,1
990)
aPE
II(d
eL
orim
ier
etal
.,19
92)
bPE
II(d
eL
orim
ier
etal
.,19
92)
Des
igna
ted
abbr
evia
tion
sfo
ror
gani
sms
are
used
inFi
gure
s1,
4to
6.Sy
nony
ms
are
plac
edin
pare
nthe
ses
afte
rth
ecu
rren
tly
reco
gniz
edor
gani
smal
nam
e.O
rgan
ism
sm
arke
dby
(a)
are
euka
ryot
ic,a
llot
hers
are
prok
aryo
tic.
Subu
nits
mar
ked
by(b
)w
ere
not
used
inth
ean
alys
is.

JMB—MS 424
Phycobiliprotein Evolution 85
Figure 2. Highly conserved amino acid positions in phycobiliproteins and their possible functions. Residues markedby stars (*) are 100% conserved in all phycobiliprotein types, including TEA proteins. Numbers in parentheses refer tothe corresponding residue position for PC from Schirmer et al. (1986).
information obtained from the X-ray crystalstructure of Mastigocladus laminosus PC (Schirmeret al., 1987), which places an insertion of twoamino acids at positions 74 and 75 for aPC (betweenthe B and E helices) and a large insertion fromamino acids 150 to 173 for bPC (between the G andH helices). The other phycobiliproteins werealigned with respect to these sequences. Theresulting alignment had several gaps or insertionsthat are different from those previously reported.For instance, two insertions for bPE at amino acids158 to 169 and 176 to 177 have previously beentreated as a single insertion (Sidler et al., 1986;Wilbanks et al., 1991). A gap from residues 189 to192 in the middle of the H helix was added toaccommodate the C terminus of the bAP18 se-quences. An insertion was also placed in the aPE
sequence at the end of the G helix at residues 145to 148. This insertion includes the cysteine of thesecond chromophore attachment site. A gap was
introduced in the TEA phycobiliprotein domainfrom amino acids 62 to 70, between the B and Ehelix (Houmard et al., 1990; Capuano et al., 1991).The positions of gaps and insertions in thealignments presented in Figure 1 should beconsidered tentative until further knowledge of thethree-dimensional structure of other phyco-biliproteins has been resolved. The a subunits ofthe cryptomonad phycobiliprotein sequences couldnot be aligned since they are highly divergent fromthe other a subunits and their three-dimensionalstructures have not been resolved.
These alignments reveal a number of residuesthat are highly conserved in all phycobiliproteinsubunits (Figure 2). One group of conservedresidues is critical for the absorption and transferof light energy (Schirmer et al., 1986). Residue 88(cysteine) is the chromophore attachment sitepresent in all phycobiliproteins except for the highmolecular weight TEA, in which the chromophore
JMB—MS 424
Phycobiliprotein Evolution86
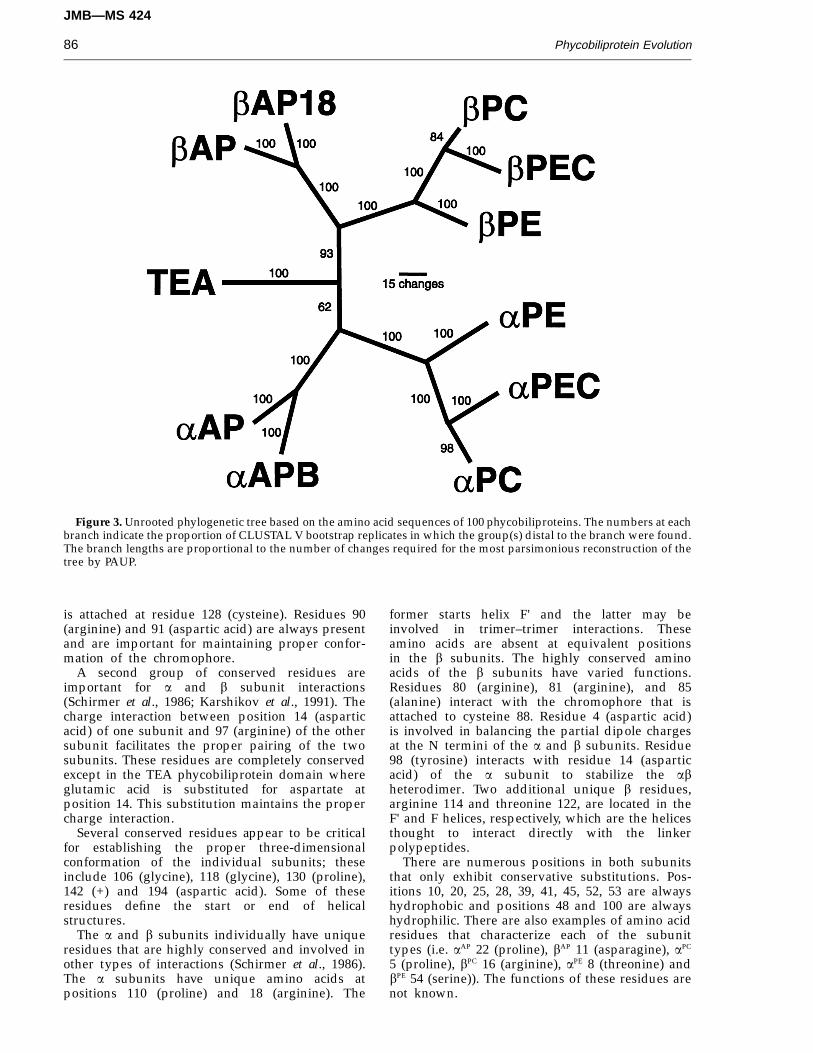
Figure 3. Unrooted phylogenetic tree based on the amino acid sequences of 100 phycobiliproteins. The numbers at eachbranch indicate the proportion of CLUSTAL V bootstrap replicates in which the group(s) distal to the branch were found.The branch lengths are proportional to the number of changes required for the most parsimonious reconstruction of thetree by PAUP.
is attached at residue 128 (cysteine). Residues 90(arginine) and 91 (aspartic acid) are always presentand are important for maintaining proper confor-mation of the chromophore.
A second group of conserved residues areimportant for a and b subunit interactions(Schirmer et al., 1986; Karshikov et al., 1991). Thecharge interaction between position 14 (asparticacid) of one subunit and 97 (arginine) of the othersubunit facilitates the proper pairing of the twosubunits. These residues are completely conservedexcept in the TEA phycobiliprotein domain whereglutamic acid is substituted for aspartate atposition 14. This substitution maintains the propercharge interaction.
Several conserved residues appear to be criticalfor establishing the proper three-dimensionalconformation of the individual subunits; theseinclude 106 (glycine), 118 (glycine), 130 (proline),142 (+) and 194 (aspartic acid). Some of theseresidues define the start or end of helicalstructures.
The a and b subunits individually have uniqueresidues that are highly conserved and involved inother types of interactions (Schirmer et al., 1986).The a subunits have unique amino acids atpositions 110 (proline) and 18 (arginine). The
former starts helix F' and the latter may beinvolved in trimer–trimer interactions. Theseamino acids are absent at equivalent positionsin the b subunits. The highly conserved aminoacids of the b subunits have varied functions.Residues 80 (arginine), 81 (arginine), and 85(alanine) interact with the chromophore that isattached to cysteine 88. Residue 4 (aspartic acid)is involved in balancing the partial dipole chargesat the N termini of the a and b subunits. Residue98 (tyrosine) interacts with residue 14 (asparticacid) of the a subunit to stabilize the abheterodimer. Two additional unique b residues,arginine 114 and threonine 122, are located in theF' and F helices, respectively, which are the helicesthought to interact directly with the linkerpolypeptides.
There are numerous positions in both subunitsthat only exhibit conservative substitutions. Pos-itions 10, 20, 25, 28, 39, 41, 45, 52, 53 are alwayshydrophobic and positions 48 and 100 are alwayshydrophilic. There are also examples of amino acidresidues that characterize each of the subunittypes (i.e. aAP 22 (proline), bAP 11 (asparagine), aPC
5 (proline), bPC 16 (arginine), aPE 8 (threonine) andbPE 54 (serine)). The functions of these residues arenot known.

JMB—MS 424
Phycobiliprotein Evolution 87
Figure 4. Strict consensus trees of the allophycocyanins. (a) aAP and aAPB (B) subunits: (b) bAP and bAP18 (18) subunits:(c) TEA. The numbers at each branch indicate the proportion of CLUSTAL V bootstrap replicates in which the group(s)distal to the branch were found. The branch lengths are proportional to the number of changes required for themost parsimonious reconstruction of the tree by PAUP. See Table 2 for abbreviations.
The evolutionary trees
Overall tree
Phylogenetic analyses of the sequences presentedin Figure 1, by either distance matrix (CLUSTAL V)or parsimony (PROTPARS) methods, revealed thateach of the phycobiliprotein subfamilies previouslyidentified by spectral characteristics (summarizedby MacColl & Guard-Friar, 1987) formed a coherentgroup in a high percentage of bootstrap replicates.The minimum possible length for the overall tree(min) is 1129 steps, and its maximum possible length(max) is 8141 steps. The shortest family of equallyparsimonious trees found by PROTPARS was 2659steps in length (rescaled consistency index(RCI) = 0.332), and the longest tree (a CLUSTAL Vbootstrap consensus tree) was 2693 steps(RCI = 0.326). The tree presented in Figure 3 reflectsan overall consensus of all the trees found. The aPE
and bPE subfamilies each contain only one distincttype of phycobiliprotein polypeptide, while the aAP
plus aAPB, bAP plus bAP18, aPC plus aPEC and bPC plus bPEC
subfamilies each contain two distinct but closelyrelated types of phycobiliprotein polypeptides. Forthese subfamilies, the rootings shown in Figure 3were chosen for clarity of presentation, but sincealternative rootings of each subfamily are possible,this presentation may not reflect the true roots of thesubfamily trees. This tree is based solely on sequenceinformation, but is also consistent with insertion/deletion and chromophore differences among thedifferent phycobiliprotein subunits.
A distinct split in the overall tree separates the afrom the b subunits. The two halves of the diagrammirror each other, with near identical branchingpatterns and similar branch lengths. The first branchin both the a and b lines divides the subunits of thecore (AP) from those of the rods (PC, PE, PEC). Onthe AP branches, bAP18 diverges from bAP and aAPB
from aAP. On the rod phycobiliprotein branches,there is a sequential divergence of PE and then PECfrom PC. The TEA phycobiliprotein-like domain isclose to the center of the tree, but CLUSTAL Vbootstrapping places it with the a subunits 93% of thetime to the exclusion of the b subunits.
Allophycocyanin subtrees
The aAP subfamily (Figure 4(a)) shows a distinctseparation into the aAPB and aAP forms. A separationof eukaryotic from cyanobacterial sequences ispreserved in both parts of this strict consensus of two410-step trees and five 412-step trees (min = 308steps, max = 601; consensus is 426 steps long withRCI = 0.432). The eukaryotic versus cyanobacterialdichotomy was found in 69% (aAP forms) and 75%(aAPB forms) of PAUP bootstrap replications.
The bAP subfamily tree (Figure 4(b)) is the strictconsensus of 12 trees, 360 to 363 steps in length(min = 282, max = 577; consensus is 393 steps longwith RCI = 0.379). The bAP18 forms are stronglyseparated from the bAP forms. The bAP18 cyanobacte-rial sequences were grouped to the exclusion of thebAP18 eukaryotic sequences in 87% of PAUP bootstrapreplicates of the bAP subfamily.

JMB—MS 424
Phycobiliprotein Evolution88
Figure 5. Strict consensus trees of the phycocyanins. (a) aPC and aPEC subunits: (b) bPC and bPEC subunits. The numbersat each branch indicate the proportion of CLUSTAL V bootstrap replicates in which the group(s) distal to the branch werefound. The branch lengths are proportional to the number of changes required for the most parsimonious reconstructionof the tree by PAUP.
For the TEA phycobiliprotein-like domain sub-family (Figure 4(c)), the strict consensus of two trees168 steps long is shown (min = 160, max = 179;consensus is 171 steps long with RCI = 0.394). Theseparation between eukaryotic and cyanobacterialforms was found in 94% of PAUP bootstrapreplications of the TEA subfamily alone.
Phycocyanin subtrees
The tree shown in Figure 5(a) is the strictconsensus of ten aPC subfamily trees, 385 to 388 stepsin length (min = 243, max = 666; consensus is 470steps long with RCI = 0.24). This tree retains theseparation of aPC from aPEC forms. Some closephylogenetic relationships (e.g. between Fremyellaand Calothrix or between Synechococcus strains 6301and 7942) are supported by PAUP bootstrapreplications of the aPC subfamily alone. The marinecyanobacterial sequences (Synechococcus strains7803, 8020, 8103) formed a tight group, and Anabaena7120 appeared to be closely related to Mastigocladus.
The strict consensus of 13 bPC subfamily trees, 424
to 429 steps long (min = 266, max = 691; consensus is490 steps long with RCI = 0.257), shows a verysimilar pattern to the aPC subtree (Figure 5(b)). Somegroupings of lower bootstrap support were lost, andthe eukaryotic sequences were grouped to theexclusion of the cyanobacterial sequences in 73% ofPAUP bootstrap replicates of the bPC sequences alone.
Phycoerythrin subtrees
Only one aPE subfamily topology was found(Figure 6(a)). It is 220 steps long with an RCI of 0.643(min = 178, max = 383), and shows a separation ofthe eukaryotic from the cyanobacterial sequences in71% of PAUP bootstrap replications of this subfamilyalone. The marine cyanobacteria (Syn 8020, Syn 8103)were placed between the freshwater cyanobacteriaand the eukaryotic sequences.
A strict consensus of four bPE subfamily trees, 297to 299 steps long (min = 219, max = 475; consensus is306 steps long with RCI = 0.472) is very similar to theaPE tree (Figure 6(b)). The eukaryotic and cyanobac-terial sequences were clearly separated in 97% of
Figure 6. Strict consensus trees of the phycoerythrins. (a) aPE subunits: (b) bPE subunits. The numbers at each branchindicate the proportion of CLUSTAL V bootstrap replicates in which the group(s) distal to the branch were found. Thebranch lengths are proportional to the number of changes required for the most parsimonious reconstruction of the treeby PAUP.
JMB—MS 424
Phycobiliprotein Evolution 89
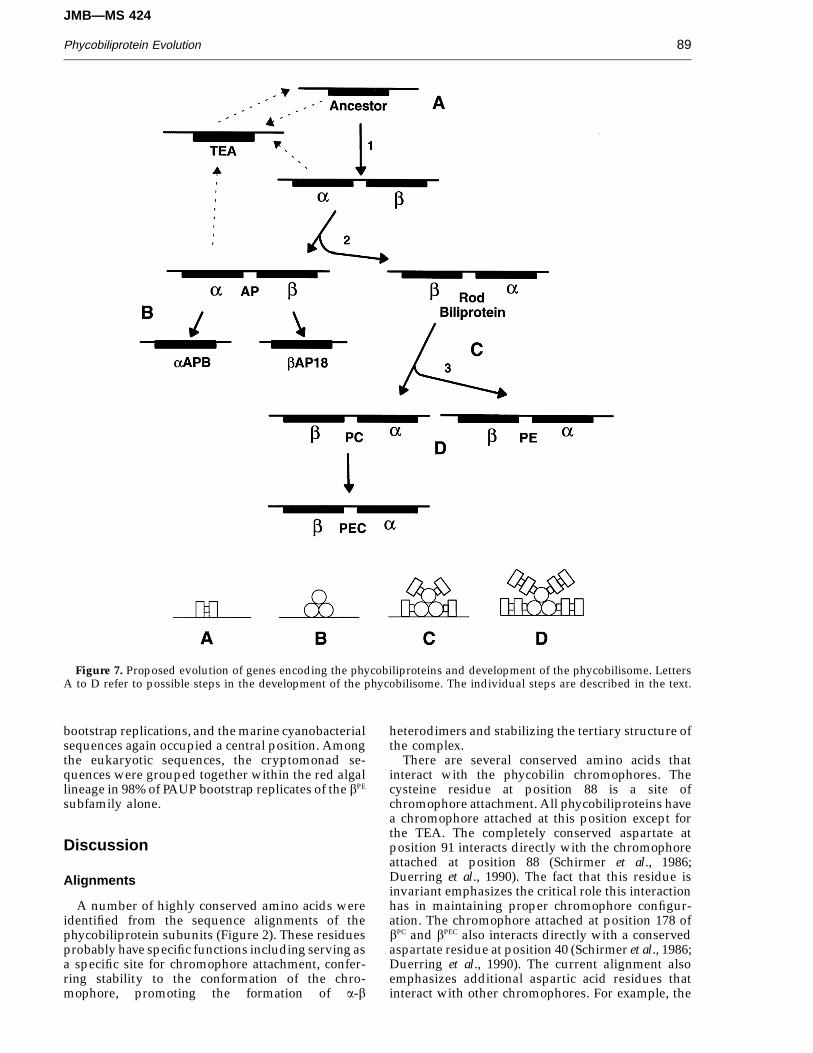
Figure 7. Proposed evolution of genes encoding the phycobiliproteins and development of the phycobilisome. LettersA to D refer to possible steps in the development of the phycobilisome. The individual steps are described in the text.
bootstrap replications, and the marine cyanobacterialsequences again occupied a central position. Amongthe eukaryotic sequences, the cryptomonad se-quences were grouped together within the red algallineage in 98% of PAUP bootstrap replicates of the bPE
subfamily alone.
Discussion
Alignments
A number of highly conserved amino acids wereidentified from the sequence alignments of thephycobiliprotein subunits (Figure 2). These residuesprobably have specific functions including serving asa specific site for chromophore attachment, confer-ring stability to the conformation of the chro-mophore, promoting the formation of a-b
heterodimers and stabilizing the tertiary structure ofthe complex.
There are several conserved amino acids thatinteract with the phycobilin chromophores. Thecysteine residue at position 88 is a site ofchromophore attachment. All phycobiliproteins havea chromophore attached at this position except forthe TEA. The completely conserved aspartate atposition 91 interacts directly with the chromophoreattached at position 88 (Schirmer et al., 1986;Duerring et al., 1990). The fact that this residue isinvariant emphasizes the critical role this interactionhas in maintaining proper chromophore configur-ation. The chromophore attached at position 178 ofbPC and bPEC also interacts directly with a conservedaspartate residue at position 40 (Schirmer et al., 1986;Duerring et al., 1990). The current alignment alsoemphasizes additional aspartic acid residues thatinteract with other chromophores. For example, the

JMB—MS 424
Phycobiliprotein Evolution90
aPE subunit has a chromophore attached at position146 that interacts with a conserved aspartic acidresidue at position 174. The double linkedchromophore of bPE is attached at positions 51 and 62and interacts with a unique aspartic acid at position55 (Ficner et al., 1992). The alignment also predictsthat the additional chromophore at position 78 of aPEII
may interact with a unique aspartic acid residue atposition 130. The alignment of the TEA with thephycobiliprotein subunits suggests that its chro-mophore (attached at position 128) also interacts withthe conserved residues at positions 90 (arginine) and91 (aspartic acid). In fact, it has been suggested thatthe chromophore of the TEA probably lies in thesame ‘‘pocket’’ as the chromophore attached toresidue 88 (cysteine) of the other phycobiliproteinsubunits, even though the primary sequence of theattachment site is different (Bryant, 1988; Houmardet al., 1990; Capuano et al., 1991). The difference in thechromophore attachment site of the TEA may berelated to its role as a terminal energy acceptor to PSII(reviewed by Glazer, 1989).
Based upon the conclusions derived fromsequence comparisons and the crystal structure, theacquisition of new chromophore attachment siteswould require at least two specific amino acidchanges. A cysteine residue would be needed tocovalently bind the chromophore and an asparticacid residue establishes the extended conformationand influences the dipole moment of the chro-mophore. A third requirement may be thedevelopment of a specific enzymatic activity thatcatalyzes chromophore attachment (Fairchild et al.,1992; Fairchild & Glazer, 1994).
Energy absorbed by the different chromophores ofthe phycobiliprotein hexamers is efficiently trans-ferred to the b subunit 88 chromophore (Schirmeret al., 1987). Unique characteristics of the b subunitshould be responsible for this preferential energytransfer. The b subunits contain a conservedhydrophobic residue (alanine) at position 85, alongwith two arginine residues at positions 80 and 81 thatinteract directly with the chromophore at position 88(Schirmer et al., 1986). The a subunits typically havea hydrophilic residue (usually lysine or threonineand never alanine) at position 85 and rarely anarginine at positions 80 or 81. Hence, conservation ofspecific residues in the b subunit that interact withthe chromophore attached to residue 88 (Cys) may besufficient to create a chromophore environment thatensures the unidirectional transfer of energy to theb subunit. Additional modifications of the b subunitsalso favor unidirectional energy transfer betweenphycobiliprotein hexamers. Methylation of as-paragine 73, which occurs in most of the b subunitsand is absent in the a subunits (Klotz & Glazer, 1987),increases the efficiency of unidirectional energytransfer within the phycobilisome (Swanson &Glazer, 1990; Thomas et al., 1993). However, thismodification does not occur in all b subunits,suggesting that other mechanisms also assistunidirectional energy transfer.
A second set of conserved residues are involved in
a-b dimerization. The alignments reveal two aminoacids, aspartate 14 and arginine 97 that are conservedin both a and b subunits. These residues form theprincipal charge interactions that stabilize a-bheterodimer formation (Schirmer et al., 1986;Karshikov et al., 1991). The b subunits also containtwo unique residues that are involved in a-b dimerformation. Residue 98 (tyrosine) interacts with the asubunit residue at position 14 (aspartic acid), and aunique aspartate at position 4 of the b subunit helpsto balance the partial dipole charge at the N terminiof both the a and b subunits, which are in directcontact with each other. Both interactions stabilizethe ab association (Schirmer et al., 1986). Theaspartate at position 4 may also play a role inpreventing b-b homodimerization by destabilizingthe interaction of the N termini of identical bsubunits. The net ( + ) charge of the N terminus of thea subunits may similarly help prevent a-ainteractions. Finally several conserved hydrophobicresidues in the X, Y, A and E helices are also knownto mediate a-b interactions (Schirmer et al., 1986).Additional unique residues found in each subunitprobably are also responsible for assuring that thecorrect subunits form proper ab heterodimers.
The conserved nature of phycobiliprotein se-quences further suggests that the three-dimensionalstructures of the different phycobiliproteins aresimilar. Hence, the known structures of C-PC, PECand B-PE should help predict structures of othertypes of phycobiliproteins.
Evolution of the phycobiliproteins
The overall phycobiliprotein tree (Figure 3)suggests that all of the phycobiliprotein subfamilies,including the TEA phycobiliprotein-like domain,originated from a common ancestral protein.Schirmer et al. (1985) and Pastore & Lesk (1990) haveproposed, based on similarity in three-dimensionalstructure, that a globin-like protein was the ancestorof the phycobiliproteins. However, the low aminoacid sequence similarity between the globins and thephycobiliproteins makes this suggestion difficult toassess by a sequence-based analysis, and it alsoprevents the use of the globins as an outgroup forrooting the overall phycobiliprotein tree. Thesymmetry of the overall tree may suggest that its rootlies near the node connecting the TEA to the rest ofthe tree. The two distinct and symmetrical lines ofdescent in the tree indicate that the close physical andenergetic interactions between the a and b subunitshave required specific co-evolutionary adaptationswithin each class of phycobiliprotein.
A hypothetical outline of early phycobiliproteinevolution is presented in Figure 7. Since a numberof amino acid residues involved in chromophoreattachment and stability, ab interactions and higherorder assembly (summarized in Figure 2) are highlyconserved in all modern phycobiliproteins, wepropose that the ancestral phycobiliprotein polypep-tide also contained these residues. The phyco-

JMB—MS 424
Phycobiliprotein Evolution 91
biliprotein ancestor probably possessed a single bilinchromophore, a three-dimensional structure like thatof modern phycobiliproteins, and the residuesnecessary to form homodimers, trimers, andhexamers. Since residues that are proposed to interactwith linker polypeptides are also conserved, theancestral phycobiliprotein probably associated witha precursor of the linker polypeptides. The twoprotein types probably co-evolved from a very earlystage. A possible ancestral complex is represented asa short rod in Figure 7A. This complex may have beenassociated with the thylakoid membrane, where ittransferred excitation energy to the photosyntheticreaction centers. Energy transfer within homodimerswould not be unidirectional, so energy transfer wouldhave been relatively inefficient.
The topology of the overall phycobiliprotein treesuggests that extant phycobiliproteins arose by aseries of duplications and divergences from theancestral protein. The origin of the TEA is uncertain,and several possibilities are presented in Figure 7(broken lines). If the TEA arose directly from the sameancestor as the other phycobiliproteins, the greatersimilarity of the TEA to a rather than to b subunitsmight reflect ancestral characteristics from which theb subunits have diverged, or convergent evolutionamong the a subunits and the TEA, since the TEApairs with the bAP18 subunit in current phycobilisomemodels (Glazer, 1987). Alternatively, the TEA mighthave arisen from the duplication of an ancestral asubunit at a later stage of phycobiliprotein evolution.It also seems likely that at a later stage of evolutionmultiple linker domains fused with the TEA to formthe complete high molecular weight linker polypep-tide.
A duplication of the ancestral gene generated a pairof tandem phycobiliprotein genes (Figure 7, step 1).This gene pair diverged and co-evolved with oneanother as the two subunits developed specialfunctions within a hexameric structure. Cotranscrip-tion of a tandem gene set might have helped tomaintain the 1:1 stoichiometry necessary forheterodimer formation. Virtually all a and b subunitsare found as cotranscribed tandem gene sets,emphasizing the importance of cotranscription. Thespecialization of the a and b subunits was a criticalstep in phycobiliprotein evolution. The differences inchromophore ‘‘environments’’ between the twosubunits helped establish a complex with uni-directional energy flow. This is a major driving forcein the evolution of phycobiliprotein subunits (Huber,1989). Unique amino acids in the b subunit may playa role in ensuring thataandb subunits efficiently pairand that the energy flow is from a to b . With thepositions of the a and b subunit fixed within thephycobiliprotein hexamer, the b subunit couldfurther specialize to develop binding sites for specificlinker polypeptides, which might further improveunidirectional energy flow and also facilitate theformation of assemblages of hexamers.
A duplication of the ancestral heterodimer pairthen gave rise to two separate lines of descent,generating the core (AP) and the rod (PC and PE)
phycobiliproteins (Figure 7, step 2). In one of theselines, the order of the genes was reversed, perhapsduring duplication.
The AP subunits were in turn duplicatedindependently of one another, giving rise to the aAPB
and bAP18 core components. By this stage of evolutionthe TEA had fused with multiple linker domains toform the complete high molecular weight core-mem-brane linker polypeptide (LCM). The LCM serves acritical role to stabilize and attach the AP subunits tothe thylakoid membrane along with being a terminalenergy acceptor. Along with the LCM polypeptide, thefour AP phycobiliproteins may have formed astructure closely resembling a modern phycobil-isome core (Figure 7B). The development of the corewith specialized subunits is important for efficientenergy transfer at the interface between thephycobiliproteins and the chlorophylls of thephotosynthetic reaction centers. The rod precursorheterodimer pair duplicated again to form the PC andPE subfamilies (Figure 7, step 3). These gene setscoevolved with the rod-core (Figure 7C) and rodlinker polypeptides, giving rise to the basic modernphycobilisome structure (Figure 7D).
The changes in the spectral characteristics of thephycobiliproteins along the proposed lines of descentfrom AP to PE are summarized in Table 1. There is anincrease in the total number of chromophores foundon each subunit, which increases the total lightabsorption capacity per protein. There is also achange in the types of chromophores presentresulting in a broadening of light absorbance with anincreasing capacity to absorb shorter wavelengths.The extreme example of adaptation to shorterwavelength light absorption is found in PEII. Thisform of PE has the majority of chromophores as PUB,which have their peak absorbance at approximately498 nm. PEII is found in marine cyanobacteria that aregenerally exposed only to blue-green light, and thehigh levels of PUB may be an adaptation to this lightregime (Wood, 1985). Hence, the evolution of thephycobiliproteins appears to have been drivenpredominantly by two factors. First, it was crucial todevelop highly efficient unidirectional energy flow(Huber, 1989). Second, it was advantageous to obtainincreased light harvesting capacity for shorterwavelengths of light.
The evolutionary scheme proposed here varies inseveral ways from two previous proposals (Troxleret al., 1981; Zuber, 1987). Those proposals involvedmore or less separate a and b evolutionary lines, butrequired that either the a or b subunits developindependently more than once. A recently publishedphycobiliprotein phylogenetic tree (Ducret et al.,1994) has some similarities to the overall treepresented here, except that the TEA is placedbetween the aAP and rod a subunits. A putative rootfor the tree is attached between the TEA and the aPC
branch, implying a closer relationship of the aAP to theb subunits than to the other a subunits. Again thismodel requires two independent origins of the asubunits. The analysis presented here does notsupport these proposals. Fuglistaller et al. (1985) and

JMB—MS 424
Phycobiliprotein Evolution92
Zuber (1987) have also proposed an evolutionaryrelationship between the phycobiliproteins and thelinker polypeptides based on a low level of aminoacid sequence similarity. We have compared thesequences of all known linker polypeptides withthose of the phycobiliproteins and find no evidenceto support a relationship between the two groups ofproteins (data not shown).
Evolution of the organisms
As described above, the most recent commonancestor of the extant cyanobacteria probablypossessed a phycobilisome composed of the LCM,aAPB,bAP18, and theaandb subunits of AP, PC, and PE, alongwith appropriate linker polypeptides. Hence, ifenough phylogenetically informative changes arepresent, analysis of any of these sequences shouldreveal the historical relationships among thephycobiliprotein-containing organisms. However, ashas also been observed in cyanobacterial molecularphylogenies based on other data (Giovannoni et al.,1988; Wilmotte et al., 1992; Wood & Townsend, 1990),many relationships have been difficult to resolve witha high degree of reliability. Part of this difficulty mayresult from the potential age of cyanobacteria, whichhas been suggested to be 3.5 billion years (reviewedby Awramik, 1992). Nonetheless, there are severalaspects of the relationships among phycobiliprotein-containing organisms that can be evaluated with thedata that are currently available.
Since the distribution of PE among phyco-biliprotein-containing organisms does not matcheither morphological or molecular phylogenies ofcyanobacteria (Rippka et al., 1979; Bryant, 1982;Giovannoni et al., 1988), the PE gene set has probablybeen lost independently from a number of cyano-bacterial lines. In some strains lacking PE (i.e.Mastigocladus and Anabaena), a late-arising dupli-cation of the PC genes generated PEC, which hasevolved toward the spectral characteristics of PE. Theevolution of PEC in these strains could be analogousto the evolution of PE in the ancestor of extant cyano-bacteria. The close relationship between the PECssuggests that they share a common origin. A commonancestor for the PEC-containing organisms (Mastigo-cladus and Anabaena) is also supported by the highdegree of similarity between their PC sequences(Figure 4). Other late-arising gene duplications insome cyanobacteria have produced multiple special-ized PC gene sets that can be expressed differentiallyin response to environmental conditions (reviewedby Grossman et al., 1993; Tandeau de Marsac &Houmard, 1993). For example, in cyanobacteria (i.e.Calothrix†, Fremyella†, and Pseudanabaena) exhibitingcomplementary chromatic adaptation (CCA) theexpression of the PE and two PC gene sets variesdepending on light quality. In some CCA organismsthe expression of a third distinct PC gene set is
dependent upon the availability of sulfur in theenvironment. Although these CCA strains did notgroup together reliably in this analysis, they havelikely derived the CCA phenotype from a commonancestor.
The marine Synechococcus (WH7803, WH8020, andWH8103) tend to form a separate group from theother cyanobacteria. A similar result was obtained byUrbach et al. (1992) and Wood & Townsend (1990). Inboth of the aPE and bPE subfamilies the marineSynechococcus were placed between the eukaryoticalgae and the remaining cyanobacteria. Consideringthat red algal plastids are believed to have evolvedfrom the endosymbiosis of a cyanobacterium(reviewed by Howe et al., 1992; Lewin, 1993), thisresult suggests that extant marine cyanobacteria aremore closely related to the plastid precursor of redalgae than the other cyanobacteria. Phylogeneticanalysis of additional sequences from the phyco-biliprotein-containing organisms would be useful tofurther test this possibility.
The eukaryotic phycobiliproteins of the Rhodo-phyta, Cryptophyta and Glaucophyta were consist-ently grouped together, to the exclusion of thecyanobacterial sequences, although not always at ahigh level of significance. If, as these data suggest, theeukaryotic phycobiliproteins form a true mono-phyletic group, phycobiliprotein-containing plastidswere probably derived from a common ancestralsymbiont. This is consistent with the work of Palenik& Haselkorn (1991) who examined 16 S ribosomalsequences and found that plastids formed a mono-phyletic group when compared to a diverse group ofcyanobacteria.
The b subunits of the cryptomonad phyco-biliproteins form a well defined group that is nestedwithin the rhodophyte bPE suggesting that a red algalbPE was the ancestor to all cryptomonad phyco-biliprotein b subunits. This is consistent with theproposal that the plastids of cryptomonads areremnants of a red algal endosymbiont (Douglas et al.,1990). The phycobiliproteins of cryptomonad algaeare not organized into typical phycobilisomes(Ludwig & Gibbs, 1989), and the a subunits bear norecognizable sequence similarity to other phyco-biliprotein subunits (Sidler et al., 1987, 1990). Thedivergence of the aPE gene may have resulted fromevolutionary pressures following transfer to thenuclear (or nucleomorph) genome (Jenkins et al.,1990) and the loss of the other phycobiliproteins.
Analysis of phycobiliprotein sequences, in con-junction with the crystal structure of phyco-biliprotein hexamers, may help to clarify the patternsand pathways of protein evolution. In combinationwith other sequence data, they may also extend ourunderstanding of the evolution of the cyanobacteria,rhodophytes, and cryptophytes.
Materials and MethodsThe phycobiliprotein sequences were obtained from the
sources listed in Table 2. The amino acid sequences werealigned using CLUSTAL V (Higgins et al., 1992), followed
† Calothrix PCC7601 and Fremyella diplosiphon FD33have probably originated from a common isolate, butthe exact history is not clear.

JMB—MS 424
Phycobiliprotein Evolution 93
by optimization of the alignments using structuralsimilarities derived from the 3-D structures of PC(Schirmer et al., 1986, 1987) and PE (Ficner et al., 1992). Ingeneral, refinements to the CLUSTAL V alignmentproduced only minor changes in the tree topologies of thephycobiliprotein subfamilies. Three methods were used toproduce unrooted dendrograms, or trees, based on thisalignment. CLUSTAL V was used to produce trees of theentire data set by the neighbor-joining (a distance matrix)method. The inclusion or exclusion of positions containinggaps in any sequence had little effect on overall treetopology, suggesting that insertion/deletion events werecongruent with other sequence changes. Two sets of 1000bootstrap replications (excluding gaps) were used in theCLUSTAL V analysis presented here. Unrooted trees of theentire data set were also produced using the parsimonymethod of PROTPARS from PHYLIP 3.4 (Felsentein, 1991)with randomized sequence input order. Each of threePHYLIP trials produced a different group of equal-lengthtrees that varied only in the topology and rooting withineach subfamily. Individual subfamilies of phyco-biliproteins were then further analyzed using the programPAUP Version 3.1 (Swofford, 1993). Gaps were treated asmissing data and 1000 bootstrap replications of eachsubtree were performed using the heuristic tree-bisection/resection search method.
All of the complete trees produced by CLUSTAL V andPROTPARS revealed the same relationships among thesubfamilies. However, the relationships within eachsubfamily were less clearly resolved, and a number ofdifferent topologies and rootings onto the overall tree weregenerally found for each subfamily tree. Therefore, all ofthe topologies obtained for each subfamily by CLUSTAL V,PROTPARS, and PAUP were analyzed, and all those within1% (two to four steps) of the shortest tree were used toproduce a strict consensus tree. In general, only branchespoorly supported (<50%) by bootstrap analysis were notretained in the consensus trees. For each tree, the value ofthe rescaled consistency index (RCI; Swofford 1993) isgiven as a measure of the degree to which the data fit thetree. An RCI of 1.0 indicates that the minimum possiblenumber of changes for each character will fit the tree, whilean RCI of 0 would indicate that all the characters mustundergo the maximum possible number of changes inorder to fit the tree. Also given are the lengths of theshortest (min) and longest (max) theoretically possibletrees, as well as the lengths of the trees used in buildingthe consensus tree and the length of the consensus treeitself. Consensus trees will always be longer than fullyresolved bifurcating trees from which they are derived.
Acknowledgements
The authors thank Glenn Ford for providing excellentcomputer assistance. We also thank Drs Michael Reith andLaura Warner who generously provided unpublishedsequences. This work was partially supported by NSFgrants DMB-8906066 to K.A. and MCB-9202386 to A.R.G.We thank the Carnegie Institution for additional support.This article is C.I.W.-D.P.B. publication number 1222.
ReferencesAnderson, L. K. & Grossman, A. R. (1990a). Genes for
phycocyanin subunits in Synechocystis sp. strain PCC6701 and assembly mutant UV16. J. Bacteriol. 172,1289–1296.
Anderson, L. K. & Grossman, A. R. (1990b). Structure andlight-regulated expression of phycoerythrin genes inwild-type and phycobilisome assembly mutants ofSynechocystis sp. strain PCC 6701. J. Bacteriol. 172,1297–1305.
Apt, K. E. & Grossman, A. R. (1993a). Characterization andtranscript analysis of the major phycobiliproteingenes from Aglaothamnion neglectum (Rhodophyta).Plant Mol. Biol. 21, 27–38.
Apt, K. E. & Grossman, A. R. (1993b). Genes encodingphycobilisome linker polypeptides on the plastidgenome of Aglaothamnion neglectum (Rhodophyta).Photosyn. Res. 35, 235–245.
Apt, K. E. & Grossman, A. R. (1993c). The phycobilisomeb18 subunit gene of allophycocyanin is located on theplastid genome in Aglaothamnion neglectum (Rhodo-phyta) and cotranscribed with an unidentified openreading frame. J. Phycol. 29, 716–718.
Awramik, S. M. (1992). The oldest records of photosyn-thesis. Photosyn. Res. 33, 75–89.
Belknap, W. R. & Haselkorn, R. (1987). Cloning andlight-regulation of expression of the phycocyaninoperon of the cyanobacterium Anabaena. EMBO J. 6,871–884.
Bernard, C., Thomas, J. C., Mazel, D., Mousseau, A.,Castets, A. M., Tandeau de Marsac, N. & Dubacq, J. P.(1992). Characterization of the genes encodingphycoerythrin in the red alga Rhodella violacea:evidence for a splitting of the rpeB gene by an intron.Proc. Natl. Acad. Sci., U.S.A. 89, 9564–9568.
Bryant, D. A. (1982). Phycoerythrocyanin and phyco-erythrin: properties and occurence in cyanobacteria.J. Gen. Microbiol. 128, 835–844.
Bryant, D. A. (1988). Genetic analysis of phycobilisomebiosynthesis, assembly, structure, and function inthe cyanobacterium Synechococcus sp. PCC7002. InLight-energy Transduction in Photosynthesis: HigherPlants and Bacterial Models (Stevens, S. E. & Bryant,D. A., eds), pp. 62–90. American Society of PlantPhysiologists, Rockville, MD.
Bryant, D. A. (1991). Cyanobacterial phycobilisomes;progress towards a complete structural and functionalanalysis via molecular genetics. In The PhotosyntheticApparatus: Molecular Biology and Operation (Bogorad,L. & Vasil, K., eds), pp. 257–300. Academic Press Inc,New York.
Bryant, D. A., de Lorimer, R., Lambert, D. H., Dubbs, J. M.,Stirewalt, V. L., Stevens, Jr, S. E., Porter, R. D., Tam, J.& Jay, E. (1985). Molecular cloning and nucleotidesequence of the a and b subunits of allophycocyaninfrom the cyanelle genome of Cyanophora paradoxa. Proc.Natl. Acad. Sci., U.S.A. 82, 3242–3246.
Capuano, V., Mazel, D., Tandeau de Marsac, N. &Houmard, J. (1988). Complete nucleotide sequence ofthe red-light specific set of phycocyanin genes fromthe cyanobacterium Calothrix PCC 7601. Nucl. Acids.Res. 16, 1626.
Capuano, V., Braux, A. S., Tandeau de Marsac, N. &Houmard, J. (1991). The ‘‘anchor polypeptide’’ ofcyanobacterial phycobilisomes, molecular character-ization of the Synechococcus sp. PCC 6301 apcE gene.J. Biol. Chem. 266, 7239–7247.
Conley, P. B., Lemaux, P. G. & Grossman, A. R. (1988).Molecular characterization and evolution of se-quences encoding light harvesting components in thechromatically adapting cyanobacterium Fremyelladiplosiphon. J. Mol. Biol. 199, 447–465.
de Lorimier, R., Bryant, D. A., Porter, R. D., Liu, W. Y., Jay,E. & Stevens, S. E. Jr (1984). Genes for the a and b

JMB—MS 424
Phycobiliprotein Evolution94
subunits of phycocyanin. Proc. Natl. Acad. Sci., U.S.A.81, 7946–7950.
de Lorimier, R., Chen, C. J. & Glazer, A. N. (1992). Sequencecomparison of two highly homologous phycoery-thrins differing in bilin composition. Plant Mol. Biol.20, 353–356.
de Lorimier, R. M., Wilbanks, S. M. & Glazer, A. N. (1993).Genes of the R-phycocyanin II locus of marineSynechococcus spp. and comparison of protein-chromophore interactions in phycocyanins differingin bilin composition. Plant Mol. Biol. 21, 225–237.
DeLange, R. J., Williams, L. C. & Glazer, A. N. (1981). Theamino acid sequence of the b subunit of allophyco-cyanin. J. Biol. Chem. 256, 9558–9566.
Dimagno, L. & Haselkorn, R. (1993). Isolation andcharacterization of the genes encoding allophyco-cyanin subunits and two linker proteins fromSynechocystis 6714. Plant Mol. Biol. 21, 835–845.
Douglas, S., Murphy, C., Spencer, D. & Gray, M. (1990).Cryptomonad algae are evolutionary chimeras of twophylogenetically distinct unicellular eukaryotes.Nature (London), 350, 148–151.
Dubbs, J. M. & Bryant, D. A. (1987). Organization of thegenes encoding phycoerythrin and the two differen-tially expressed phycocyanins in the cyanobacteirumPseudanabaena PCC 7409. In Progress in PhotosynthesisResearch (Biggens, J., ed.), vol. 4, pp. 765–768.Martinus Nijhoff Publ, Dordrecht.
Dubbs, J. M. & Bryant, D. A. (1991). Molecular cloning andtranscriptional analysis of the cpeBA operon from thecyanobacterium Pseudanabaena species PCC 7409. Mol.Microbiol. 5, 3073–3085.
Dubbs, J. M. & Bryant, D. A. (1993). Organization andtranscription of the genes encoding two differentiallyexpressed phycocyanins in the cyanobacteriumPseudoanabaena sp. PCC 7409. Photosyn. Res. 36,169–183.
Ducret, A., Sidler, W., Frank, G. & Zuber, H. (1994). Thecomplete amino acid sequence of R-phycocyanin I aand b subunits from the red alga Porphyridiumcruentum. Structural and phylogenetic relationships ofthe phycocyanins and the phycobiliprotein families.Eur. J. Biochem. 221, 563–580.
Duerring, M., Huber, R., Bode, W., Ruembeli, R. & Zuber,H. (1990). Refined three-dimensional structure ofphycoerythrocyanin from the cyanobacteriumMastigocladus laminosus at 2.7 A. J. Mol. Biol. 211,633–644.
Duerring, M., Schmidt, G. B. & Huber, R. (1991).Isolation, crystallization, crystal structure analysisand refinement of constitutive C-phycocyanin fromthe chromatically adapting cyanobacteria Fremyelladiplosiphon at 1.66 A resolution. J. Mol. Biol. 217,577–592.
Eberlein, M. & Kufer, W. (1990). Genes encoding bothsubunits of phycoerythrocyanin, a light-harvestingbiliprotein from the cyanobacterium Mastigocladuslaminosus. Gene, 94, 133–136.
Fairchild, C. D. & Glazer, A. N. (1994). Oligomericstructure, enzyme kinetics, and substrate specificityof the phycocyanin a subunit phycocyanobilin lyase.J. Biol. Chem. 269, 8686–8694.
Fairchild, C. D., Zhao, J., Zhou, J., Colson, S. E., Bryant,D. A. & Glazer, A. N. (1992). Phycocyanin a subunitphycocyanobilin lyase. Proc. Natl. Acad. Sci., U.S.A. 89,7017–7021.
Felsenstein, J. (1991). PHYLIP 3.4, University of Washing-ton.
Ficner, R. & Huber, R. (1993). Refined crystal structure of
phycoerythrin from Porphyridium cruentum at 0.23-nmresolution and localization of the g subunit. Eur. J.Biochem. 218, 103–106.
Ficner, R., Lobeck, K., Schmidt, G. & Huber, R. (1992).Isolation, crystallization, crystal structure analysisand refinement of B-phycoerythrin from the red algaPorphyridium sordidum at 2.2 A resolution. J. Mol. Biol.228, 935-950.
Frank, G., Sidler, W., Widmer, H. & Zuber, H. (1978). Thecomplete amino acid sequence of both subunits ofC-phycocyanin from the cyanobacterium Mastigo-cladus laminosus. Hoppe-Seyler’s Physiol. Chem. 359,1491–1507.
Freidenreich, P., Apell, G. S. & Glazer, A. N. (1978).Structural studies of phycobiliproteins II. C-phyco-cyanin: amino acid sequence of the b subunit. Specificcleavage of the a subunit. J. Biol. Chem. 253, 212–219.
Fuglistaller, P., Suter, F. & Zuber, H. (1983). The completeamino-acid sequence of both subunits of phyco-erythrocyanin from the thermophilic cyanobacteriumMastigocladus laminosus. Hoppe-Seyler’s Physiol. Chem.364, 691–712.
Fuglistaller, P., Suter, F. & Zuber, H. (1985). Linkerpolypeptides of the phycobilisome from the cyano-bacterium Mastigocladus laminosus: amino-acid se-quences and relationships. Biol. Chem. Hoppe-Seyler,366, 993–1001.
Giovannoni, S. J., Turner, S., Olsen, G. J., Barns, S., Lane,D. J. & Rice, N. R. (1988). Evolutionary relationshipsamong cyanobacteria and green chloroplasts.J. Bacteriol. 170, 3584–3592.
Glazer, A. N. (1980). Structure and evolution ofphotosynthetic accessory pigment systems withspecial reference to phycobiliproteins. In The Evolutionof Protein Structure and Function (Sigman, D. S. &Brazier, M. A. B., eds), pp. 221–244. Academic Press,New York.
Glazer, A. N. (1987). Phycobilisomes: assembly andattachment. In The Cyanobacteria (Fay, P. & Van Baalen,C., eds.), pp. 69–94. Elsevier Biomedical, Amsterdam.
Glazer, A. N. (1989). Light guides. Directional energytransfer in a photosynthetic antenna. J. Biol. Chem. 264,1–4.
Glazer, A. N., Apell, G. S., Hixson, C. S., Bryant, D. A.,Rimon, S. & Brown, D. M. (1976). Biliproteins ofcyanobacteria and rhodophyta: homologous family ofphotosynthetic accessory pigments. Proc. Natl. Acad.Sci., U.S.A. 73, 428–431.
Godovac-Zimmerman, J., Sheil, M., Wrench, P. M. & Hiller,R. G. (1992). Amino acid sequence of the b-subunit ofphycoerythrin from the cryptophyte algae ChroomonasCS24. Biochim. Biophys. Acta, 1120, 117–121.
Grossman, A. R., Schafer, M. R., Chaing, G. G. & Collier,J. L. (1993). The phycobilisome, a light-harvestingcomplex responsive to environmental conditions.Microbiol. Rev. 57, 725–749.
Higgins, D. G., Bleasby, A. J. & Fuchs, R. (1992). CLUSTALV: improved software for multiple sequence align-ment. CABIOS, 8, 189–191.
Holzwarth, A. R. (1991). Structure-function relationshipsand energy transfer in phycobiliprotein antennae.Physiol. Plant, 83, 518–528.
Houmard, J., Mazel, D., Moquet, C., Bryant, D. A. &Tandeau de Marsac, N. (1986). Organization andnucleotide sequences of genes encoding core com-ponents of the phycobilisomes from Synechococcus6301. Mol. Gen. Genet. 205, 404–410.
Houmard, J., Capuano, V., Coursin, T. & Tandeau deMarsac, N. (1988a). Allophycocyanin B, a terminal

JMB—MS 424
Phycobiliprotein Evolution 95
energy acceptor in cyanobacterial phycobilisomes.Mol. Microbiol. 2, 101–107.
Houmard, J., Capuano, V., Coursin, T. & Tandeau deMarsac, N. (1988b). Genes encoding core componentsof the phycobilisome in the cyanobacterium Calothrixsp. strain PCC 7601: occurance of a multigene family.J. Bacteriol. 170, 5512–5521.
Houmard, J., Capuano, V., Colombano, M. V., Coursin, T.& Tandeau de Marsac, N. (1990). Molecularcharacterization of the terminal energy acceptor ofcyanobacterial phycobilisomes. Proc. Natl. Acad. Sci.,U.S.A. 87, 2152–2156.
Howe, C. J., Beanland, T. J., Larkum, A. W. D. & Lockhart,P. J. (1992). Plastid origins. Trends Ecol. Evol. 7, 378–383.
Huber, R. (1989). A structural basis of light energy electrontransfer in biology. EMBO J. 8, 2125–2147.
Jenkins, J., Hiller, R. G., Speirs, J. & Godovac-Zimmerman,J. (1990). A genomic clone encoding a cryptophytephycoerythrin a-subunit. Evidence for three asubunits and an N-terminal membrane transitsequence. FEBS Letters, 273, 191–194.
Karshikov, A., Duerring, M. & Huber, R. (1991). Role ofelectrostatic interaction in the stability of the hexamerof constitutive phycocyanin from Fremyella diplosiphon.Protein Eng. 4, 681–690.
Klotz, A. V. & Glazer, A. N. (1987). g-N-Methylasparaginein phycobiliproteins, occurrence, location and biosyn-thesis. J. Biol. Chem. 262, 17350–17355.
Lau, R. H., Alvarado-Urbina, G. & Lau, P. C. K. (1987a).Phycocyanin a-subunit gene of Anacystis nidulans R2:cloning, nucleotide sequencing and expression inEscherichia coli. Gene, 52, 21–29.
Lau, P. C. K., Condie, J. A., Alvarado-Urbina, G. & Lau, R.(1987b). Nucleotide sequence of phycocyanin b-sub-unit gene of cyanobacterium Anacystis nidulans strainR2. Nucl. Acids Res. 15, 2394.
Lewin, R. A. (ed.) (1993). Origins of Plastids: Symbiogenesis,Prochlorophytes and the Origins of Chloroplasts.Chapman and Hall, New York.
Lind, L. K. (1988). Cloning and characterization of genes forlight-harvesting antenna polypeptides in Synechococ-cus PCC 6301. PhD Thesis, University of Umea.
Liu, B. & Troxler, R. (1993). A Cyanidium caldariumallophycocyanin b subunit gene. Plant Physiol. 103,293–294.
Ludwig, M. & Gibbs, S. P. (1989). Localization ofphycoerythrin at the lumenal surface of the thylakoidmembrane in Rhodomonas lens. J. Cell. Biol. 108,875–884.
MacColl, R. & Guard-Friar, D. (1987). Phycobiliproteins,CRC Press, Boca Raton.
Mazel, D. & Marliere, P. (1989). Adaptive eradication ofmethionine and cysteine from the cyanobacteriallight-harvesting proteins. Nature (London), 341,245–248.
Mazel, D., Guglielmi, G., Houmard, J., Sidler, W., Bryant,D. A. & Tandeau de Marsac, N. (1986). Green lightinduces transcription of the phycoerythrin operon inthe cyanobacteirum Calothrix 7601. Nucl. Acids Res. 14,8279–8290.
Mazel, D., Houmard, J. & Tandeau de Marsac, N. (1988). Amultigene family in Calothrix sp PCC 7601 encodesphycocyanin, the major component of the cyanobacte-rial light-harvesting antenna. Mol. Gen. Genet. 211,296–304.
Michalowski, C. B., Bohnert, H. J. & Loffelhardt, W. (1990).A novel allophycocyanin gene (apcD) from Cyanophoraparadoxa cyanelles. Nucl. Acids Res. 18, 2186.
Minami, Y., Yamada, F., Hase, T., Matsubara, H.,
Murakami, A., Fujita, Y., Takao, T. & Shimonishi, T.(1985). Amino acid sequences of allophycocyanin a-and b-subunits isolated from Anabaena cylindrica.Presence of an unknown derivative of aspartic acid inthe b-subunit. FEBS Letters, 191, 216–220.
Morschel, E. (1991). The light-harvesting antennae ofcyanobacteria and red algae. Photosynthetica, 25,137–144.
Newman, J., Mann, N. H. & Carr, N. G. (1994).Organization and transcription of the class IPhycoerythrin genes of the marine cyanobacteriumSynechococcus sp. WH7803. Plant Mol. Biol. 24,679–683.
Offner, G. D. & Troxler, R. F. (1983). Primary structure ofallophycocyanin from the unicellular rhodophyte,Cyanidium caldarium, the complete amino acidsequences of the a and b subunits. J. Biol. Chem. 258,9931–9940.
Offner, G. D., Brown-Mason, A. S., Ehrhardt, M. M. &Troxler, R. F. (1981). Primary structure of phycocyaninfrom the unicellular rhodophyte Cyanidium caldarium.I. Complete amino acid sequence of the a subunit.J. Biol. Chem. 256, 12167–12175.
Palenik, B. & Haselkorn, R. (1991). Multiple evolutionaryorgins of the prochlorophytes, the chlorophyllb-containing prokaryotes. Nature (London), 355,265–267.
Pastore, A. & Lesk, A. M. (1990). Comparison of thestructure of globins and phycocyanins: Evidence forevolutionary relationship. Proteins: Struct. Funct.Genet. 8, 133–155.
Pilot, T. J. & Fox, L. (1984). Cloning and sequencing of thegenes encoding the a and b subunits of C-phycocyaninfrom the cyanobacterium Agmenellum quadruplicatum.Proc. Natl. Acad. Sci., U.S.A. 81, 6983–6987.
Reith, M. & Douglas, S. (1990). Localizaiton ofb-phycoerythrin to the thylakoid lumen of Cryp-tomonas F does not involve a signal peptide. Plant Mol.Biol. 15, 585–592.
Rippka, R., Deruelles, J., Waterbury, J. B., Herdman, M. &Stanier, Y. (1979). Generic assignments, strain historiesand properties of pure cultures of cyanobacteria.J. Gen. Microbiol. 111, 1–61.
Roell, M. & Morse, D. (1993). Organization, expression andnucleotide sequence of the operon encoding R-phyco-erythrin a and b subunits from a macrophytic red alga.Plant Mol. Biol. 21, 47–58.
Rumbeli, R., Wirth, M., Suter, F. & Zuber, H. (1987). Thephycobiliprotein b16.2 of the allophycocyanin core fromthe cyanobacteria Mastigocladus laminosus. Character-ization and complete amino-acid sequence. Biol. Chem.Hoppe-Seyler, 368, 1–9.
Schirmer, T., Bode, W., Huber, R., Sidler, W. & Zuber, H.(1985). X-ray crystallographic structure of thelight-harvesting biliprotein C-phycocyanin from thethermophilic cyanobacterium Mastigocladus laminosusand its resemblance to globin structures. J. Mol. Biol.184, 257–277.
Schirmer, T., Huber, R., Schneider, M., Bode, W., Miller, M.& Hackert, M. L. (1986). Crystal structure analysis andrefinement at 2.5 A of hexameric C-phycocyanin fromthe cyanobacterium Agmenellum quadruplicatum. Themolecular model and its implications for lightharvesting. J. Mol. Biol. 188, 651–676.
Schirmer, T., Bode, W. & Huber, R. (1987). Refinedthree-dimensional structures of two cyanobacterialC-phycocyanins of 2.1 and 2.5 A resolution. Acommon principle of phycobilin-protein interaction.J. Mol. Biol. 196, 677–695.

JMB—MS 424
Phycobiliprotein Evolution96
Sidler, W. (1994). Phycobilisome and phycobiliproteinstructures. In The Biology of Cyanobacteria (Bryant, D.A., ed.), in the press, Kluwer Academic Publ,Dordrecht.
Sidler, W., Gysi, J., Isker, E. & Zuber, H. (1981). Thecomplete amino acid sequence of both subunits ofallophycocyanin, a light harvesting protein-pigmentcomplex from the cyanobacterium Mastigocladuslaminosus. Hoppe-Seyler’s Z. Physiol. Chem. 362,611–628.
Sidler, W. A., Kumpf, B., Rudiger, W. & Zuber, H. (1986).The complete amino acid sequence of C-phycoery-thrin from the cyanobacterium Fremyella diplosiphon.Biol. Chem. Hoppe-Seyler, 367, 627–642.
Sidler, W. A., Nutt, H., Kumpf, B. & Zuber, H. (1987).Structural principles of crytomonad biliproteins andcyanobacterial phycobilisomes. In Progress in Photo-synthesis Research (Biggens, J., ed.), vol. 2, pp. 153–156.Martinus Nijhoff Publ. Dordrecht.
Sidler, W., Kumpf, B., Suter, F., Klotz, A. V., Glazer, A. N.& Zuber, H. (1989). The complete amino-acidsequence of the a and b subunits of B-phycoerythrinfrom the rhodophytan alga Porphyridium cruentum.Biol. Chem. Hoppe-Seyler, 370, 115–124.
Sidler, W., Nutt, H., Kumpf, B., Frank, G., Suter, F., Brenzel,A., Wehrmeyer, W. & Zuber, H. (1990). The completeamino acid sequence and the phylogenetic origin ofphycocyanin-645 from the cyptophytan algaChroomonas sp. Biol. Chem. Hoppe-Seyler, 371, 537–547.
Su, X., Fraenkel, P. G. & Bogorad, L. (1992). Excitationenergy transfer from phycocyanin to chlorophyll in anapcA-defective mutant of Synechocystis PCC6803.J. Biol. Chem. 267, 22944–22950.
Suter, F., Fuglistaller, P., Lundell, D. J., Glazer, A. N. &Zuber, H. (1987). Amino acid sequences of a-allophy-cocyanin B from Synechococcus 6301 and Mastigocladuslaminosus. FEBS Letters, 217, 279–282.
Swanson, R. V. & Glazer, A. N. (1990). Phycobiliproteinmethylation. Effect of the g-N-methylasparagineresidue on energy transfer in phycocyanin and thephycobilisome. J. Mol. Biol. 214, 787–796.
Swanson, R. V., de Lorimier, R. & Glazer, A. N. (1992).Genes encoding the phycobilisome rod substructureare clustered on the Anabaena chromosome: character-ization of the phycoerythrocyanin operon. J. Bacteriol.174, 2640–2647.
Swofford, D. L. (1993). PAUP: Phylogenetic analysis usingparsimony, Version 3.1.1. Computer program dis-tributed by the Illinois Natural History Survey,Champaign, Illinois.
Tandeau de Marsac, N. & Houmard, J. (1993). Adaptationsof cyanobacteria to environmental stimuli, new stepstowards molecular mechanisms. FEBS Rev. 104,119–189.
Tandeau de Marsac, N., Mazel, D., Capuano, V., Damerval,T. & Houmard, J. (1990). Genetic analysis of thecyanobacterial light-harvesting antenna complex. InMolecular Biology of Membrane-bound Complexes inPhototrophic Bacteria (Drews, G. & Daves, E. A., eds),pp. 143–154. Plenum Press, New York.
Thomas, B. A., Bricker, T. M. & Klotz, A. V. (1993).Post-translational methylation of phycobilisomes andoxygen evolution efficiency in cyanobacteria. Biochim.Biophys. Acta, 1143, 104–108.
Troxler, R. F., Ehrhardt, M. M., Brown-Mason, A. S. &Offner, G. (1981). Primary structure of phycocyaninfrom the unicellular rhodophyte Cyanidium caldariumII. Complete amino acid sequence of the b subunit.J. Biol. Chem. 256, 12176–12184.
Urbach, E., Rober, D. L. & Chisholm, S. W. (1992). Multipleevolutionary orgins of prochlorophytes within thecyanobacterial radiation. Nature (London), 355, 267–270.
Valentin, K., Maid, U., Emich, A. & Zetsche, K. (1992).Organization and expression of a phycobiliproteingene cluster from the unicellular red alga Cyanidiumcaldarium. Plant Mol. Biol. 20, 267–276.
Walsh, R. G., Wingfield, P., Glazer, A. N. & DeLange, R. J.(1980). Amino acid sequence of C-phycocyanin alphasubunit from Synechococcus 6301 (Anacystis nidulans).Fed. Proc. Fed. Amer. Soc. Exp. Biol. 39, 1998.
Wedemayer, G. J., Wemmer, D. E. & Glazer, A. N. (1991).Phycobilins of cryptophycean algae. Structures ofnovel bilins with acryloyl substituents from phyco-erythrin 566. J. Biol. Chem. 266, 4731–4741.
Wedemayer, G. J., Kidd, D. G., Wemmer, D. E. & Glazer,A. N. (1992). Phycobilins of cryptophycean algae.Occurrence of dihydrobiliverden and mesobiliverdinin cryptomonad biliproteins. J. Biol. Chem. 267,7315–7331.
Wilbanks, S. M. & Glazer, A. N. (1993). Rod structure ofa phycoerythrin II-containing phycobilisome. I.Organization and sequence of the gene clusterencoding the major phycobiliprotein rod componentsin the genome of marine Synechococcus sp. WH8020.J. Biol. Chem. 268, 1226–1235.
Wilbanks, S. M., de Lorimier, R. & Glazer, A. N. (1991).Phycoerythrins of marine unicellular cyanobacteria.III. Sequence of a class II phycoerythrin. J. Biol. Chem.266, 9535–9539.
Wilmotte, A., Turner, S., de Peer, Y. V. & Pace, N. R. (1992).Taxonomic study of marine oscillatoriacean strains(cyanobacteria) with narrow trichomes. II. Nucleotidesequence analysis of the 16S ribosomal RNA. J. Phycol.28, 828–838.
Wilson, W. H., Newman, J., Mann, N. H. & Carr, N. G.(1991). Cloning and sequence analysis of thephycocyanin genes of the marine cyanobacteriumSynechococcus sp. WH7803. Plant Mol. Biol. 17,931–933.
Wood, A. M. (1985). Adaptation of photosyntheticapparatus of marine ultraphytoplankton to naturallight fields. Nature (London), 316, 253–255.
Wood, A. M. & Townsend, D. (1990). DNA polymorphismwithin the WH7803 serogroup of marine Synechococ-cus spp. (cyanobacteria). J. Phycol. 26, 576–585.
Zuber, H. (1987). The structure of light-harvestingpigment-protein complexes, In The Light Reactions(Barber, J., ed.), pp. 197–259. Elsevier Biomedical,Amsterdam.
Edited by R. Huber
(Received 19 September 1994; accepted 17 January 1995)
Related Documents