Bio1.J. Linn. SOC., 1,pp. 135-148. With 1 plate and4figures April 1969 Evolution and speciation in a tropical high mountain flora OLOV HEDBERG Institute of Systematic Botany, University of Uppsala, S weh Few ecosystems provide better opportunities for the study of evolution and speciation than those inhabiting the uppermost parts of the high east African mountains. These mountains are mainly of volcanic origin and lie widely scattered across the wide plateaux of east Africa, several of them reaching altitudes between 3500 and 6000 m. Their vegetation deviates very much from that of the intervening lower country, displaying a marked zonation with a montane forest belt, a (subalpine) ericaceous belt, and an afroalpine belt. The flora of the latter, the afroalpine flora, is of exceptional interest in this connection. The afroalpine flora is famous for its large numbers of geographically vicarious taxa-its Giant Senecios and Giant Lobelias are as renowned as the finches of the Galapagos Islands. Ecologically, the afroalpine biota is indeed also an island biota-the high mountain summits protrude as isolated temperate islands above the warm surrounding plains. These mountains have evidently stood isolated from each other since their origin. Pleistocene climatic changes have certainly modified their vegetation zonation to a considerable extent, but direct contacts between their afroalpine enclaves during the Pleistocene or earlier seem most im- probable. These enclaves must therefore have been isolated from each other and from other high mountain areas for avery long time, and dispersal of plants between them must presumably have occurred mainly by long distance transport, possibly facilitated by cyclones. Some 80% of the afroalpine species of vascular plants are endemic to the high mountains of tropical east Africa and Ethiopia. Vicarious taxa occur of different status. In some cases one species occurs on all or most of the east African mountains with a vicariad in other parts of the world, as exemplifiedby Subularia monticola (afroalpine) and S. aquatica (circumboreal). In other cases each of two species is conlined to one group of mountains, as in the species pair Lobelia wollastonii (Virunga Volcanoes and Ruwenzori) and L. telekii (Elgon, Aberdare, M t Kenya). Finally, there are several groups of vicarious taxa where each taxon is confined as a rule to a single mountain, as in the Lobelia deckenii group with six cognate species. Numerous similar cases occur among spiders and insects. The differentiation of these vicarious species is assumed to have occurred through natural selection in connection with genetic drift, acting upon geographically isolated and originally very small random samples of the gene pools concerned. The amount of differentiation between different mountain populations differs considerably between different groups-numerous inter- mediate stages exist between morphologically indistinguishable populations and full-fledged vicarious species. The rate of evolutionary change seems to differ considerably between different genera and families. Whether internal barriers to interbreeding exist between these vicarious taxa is in most cases unknown. No less interesting than the geographically vicarious taxa mentioned above are some cases of altitudinal vicariism. Some afroalpine species appear to have evolved from afromontane forest species through progressive adaptations favouring survival in the inhospitable afroalpine climate. The most remarkable examples are provided by the strangely specialized Giant Senecios and Giant Lobelias. CONTENTS Introduction . . 136 Endemism in the afroalpine flora . . . 136 Age of isolation of the montane enclaves . . 136 Age of isolation of afroalpine enclaves . . 141 Degree of differentiation in various groups . . 143 Genetic systems, pollination biology, polyploidy . . 144 Mode of evolution . . 144 References . . 147 PAGE

Evolução e Especiação Em Uma Montanha Tropical
Dec 03, 2015
O artigo discorre sobre os processos de adaptação e evolução de floras montanas tropicais
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Bio1.J. Linn. SOC., 1,pp. 135-148. With 1 plate and4figures
April 1969
Evolution and speciation in a tropical high mountain flora
OLOV HEDBERG Institute of Systematic Botany, University of Uppsala, S w e h
Few ecosystems provide better opportunities for the study of evolution and speciation than those inhabiting the uppermost parts of the high east African mountains. These mountains are mainly of volcanic origin and lie widely scattered across the wide plateaux of east Africa, several of them reaching altitudes between 3500 and 6000 m. Their vegetation deviates very much from that of the intervening lower country, displaying a marked zonation with a montane forest belt, a (subalpine) ericaceous belt, and an afroalpine belt. The flora of the latter, the afroalpine flora, is of exceptional interest in this connection.
The afroalpine flora is famous for its large numbers of geographically vicarious taxa-its Giant Senecios and Giant Lobelias are as renowned as the finches of the Galapagos Islands. Ecologically, the afroalpine biota is indeed also an island biota-the high mountain summits protrude as isolated temperate islands above the warm surrounding plains. These mountains have evidently stood isolated from each other since their origin. Pleistocene climatic changes have certainly modified their vegetation zonation to a considerable extent, but direct contacts between their afroalpine enclaves during the Pleistocene or earlier seem most im- probable. These enclaves must therefore have been isolated from each other and from other high mountain areas for avery long time, and dispersal of plants between them must presumably have occurred mainly by long distance transport, possibly facilitated by cyclones.
Some 80% of the afroalpine species of vascular plants are endemic to the high mountains of tropical east Africa and Ethiopia. Vicarious taxa occur of different status. In some cases one species occurs on all or most of the east African mountains with a vicariad in other parts of the world, as exemplified by Subularia monticola (afroalpine) and S . aquatica (circumboreal). In other cases each of two species is conlined to one group of mountains, as in the species pair Lobelia wollastonii (Virunga Volcanoes and Ruwenzori) and L. telekii (Elgon, Aberdare, M t Kenya). Finally, there are several groups of vicarious taxa where each taxon is confined as a rule to a single mountain, as in the Lobelia deckenii group with six cognate species. Numerous similar cases occur among spiders and insects.
The differentiation of these vicarious species is assumed to have occurred through natural selection in connection with genetic drift, acting upon geographically isolated and originally very small random samples of the gene pools concerned. The amount of differentiation between different mountain populations differs considerably between different groups-numerous inter- mediate stages exist between morphologically indistinguishable populations and full-fledged vicarious species. The rate of evolutionary change seems to differ considerably between different genera and families. Whether internal barriers to interbreeding exist between these vicarious taxa is in most cases unknown.
No less interesting than the geographically vicarious taxa mentioned above are some cases of altitudinal vicariism. Some afroalpine species appear to have evolved from afromontane forest species through progressive adaptations favouring survival in the inhospitable afroalpine climate. The most remarkable examples are provided by the strangely specialized Giant Senecios and Giant Lobelias.
CONTENTS
Introduction . . 136 Endemism in the afroalpine flora . . . 136 Age of isolation of the montane enclaves . . 136 Age of isolation of afroalpine enclaves . . 141 Degree of differentiation in various groups . . 143 Genetic systems, pollination biology, polyploidy . . 144 Mode of evolution . . 144 References . . 147
PAGE

136 OLOV HEDBERC
INTRODUCTION
Few ecosystems provide better opportunities for the study of evolution and speciation than those inhabiting the uppermost parts of the high east African mountains. The latter are mainly of volcanic origin and lie scattered across the wide plateaux of east Africa, several of them reaching altitudes between 3500 and 6000 m. The most im- portant are the Virunga Volcanoes, Ruwenzori, Elgon, Aberdare, Mt Kenya, Kili- manjaro and Mt Meru. Their vegetation deviates profoundly from that of the inter- vening lower country, displaying a marked zonation with a montane forest belt, a (sub- alpine) ericaceous belt, and an afroalpine belt (Hedberg, 1951). The flora of the latter, the afroalpine flora, is of special interest in this connection.
The climate of the afroalpine belt is rather harsh, with 'summer every day and winter every night', and its flora is poor in species. This afroalpine flora is, however, extremely interesting because of the striking ecological adaptations displayed by many species (Hedberg, 1964) and its large amount of geographically vicarious taxa. Its Giant Senecios and Giant Lobelias are as renowned in this respect as the finches of the Galapagos Islands. Ecologically, the afroalpine biota is indeed also an island biota-the high mountain summits protrude as isolated temperate islands above the warm surrounding plains.
ENDEMISM IN THE AFROALPINE FLORA
Some 80% of the afroalpine species of vascular plants are endemic to the high moun- tains of east Africa and Ethiopia (Hedberg, 1961). Vicarious taxa occur of different status. In some cases one species occurs on all or most of the east African mountains with a vicariad in other parts of the world, as exemplified by Subularia monticola" (afroalpine) and S. aquaticu (circumboreal). In other cases each of two species is con- fined to one group of mountains, as in the species pair Lobelia wollastonii (Virunga Volcanoes and Ruwenzori) and L. telekii (Elgon, Aberdare, Mt Kenya). Finally, there are several groups of vicarious taxa where each taxon is confined as a rule to a single mountain, as in the Lobelia deckenii group with six cognate species. Numerous similar examples occur among spiders and insects.
The vicarious species of the afroalpine flora have evidently arisen under the influence of geographical isolation. The mountains harbouring them are of unequal ages (Miocene to late Pleistocene) and must have stood isolated from each other since their origin (cf. Hedberg, 1961). Their afroalpine enclaves are at present very efficiently isolated from each other. In order to investigate how long this isolation may have lasted we must mobilize the evidence available concerning the Pleistocene climatic and vegetational history of east Africa. A considerable part of this paper must therefore be devoted to evaluation of such evidence.
AGE OF ISOLATION OF THE MONTANE ENCLAVES
That considerable climatic changes have occurred in east Africa during the Pleisto- cene is evidenced by traces of extensive earlier glaciations-terminal moraines have
+ The botanical nomenclature used in this paper follows Hedberg (1957) to which publication the reader is referred for authors' names and full references.

EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 137
been found down to 1740 m on Ruwenzori, c. 3500 m on Elgon, c. 2900 m on Mt Kenya, and c. 3600 m on Kilimanjaro (cf. Gregory, 1894; Mackinder, 1900; Meyer, 1900; Nilsson, 1932; Hedberg, 1951; Baker in Coe, 1967; Livingstone, 1967). The retreat of the glaciers on Ruwenzori has been shown by radiocarbon dating to be contemporaneous with deglaciation in Europe (Livingstone, 1962, 1967) and hence with glacial retreat also in North America (Flint & Deevey, 1951), Patagonia (Auer, 1958), and New Zealand (Gage, 1961). According to Osmaston (1967) the lowering of the firn-line on Ruwenzori during the glacial maximum could be accounted for by a decrease of about 4" C below the present mean temperature, whereas on Kilimanjaro the corresponding decrease was estimated at about 6". Additional evidence for world- wide changes of climate has been supplied by oceanographic studies from the floor of the Atlantic, suggesting that the temperature of its surface water was about 5" lower during the last glaciation than at present (Emiliani, 1958). Other estimates of reduced mean temperature during the last Glacial are quoted in Moreau (1963 : 399) ; see also van Zinderen Bakker (1967b).
Studies of ancient shorelines along some of the large east African lakes have indicated a much higher water level during some earlier 'pluvial' epochs (cf. Nilsson, 1932, 1940, 1949). These Pluvials were at first believed to have been contemporaneous with the glaciations on the mountains, but recent investigations have shown them to have been at least partly out of phase-some of them were furthermore evidently caused by tectonic movements rather than by climatic shifts. The last major lake phase of Lake Victoria seems to have occurred between 13,000 and 3000years ago (Kendall in van Zin- deren Bakker, 1 9 6 7 ~ : 98)-at about the same time as a Pluvial prevailed in central Sahara, allowing the southwards extension of the Mediterranean flora into the moun- tains of this area (QuCzel, 1963).
The method of pollen analysis, which has been so helpful in elucidating the vegeta- tion history of Boreal countries, has been only recently attemptedineast Africa (Hedberg, 1955), and even today only a few pollen diagrams are available from this part of the world (Osmaston, 1958 ; van Zinderen Bakker, 1964; Coetzee, 1964,1967 ; Livingstone, 1967; Morrison, 1968, etc.). These diagrams certainly indicate the occurrence of important vegetational changes during and after the last glaciation, but their interpreta- tion is by no means easy (cf. Morrison, 1966; Livingstone, 1967).
Both van Zinderen Bakker (1964) and Coetzee (1964,1967) interpret their diagrams as indicating a considerable downwards shift, amounting to some 1000-1100 m, of the vegetation belts of Mt Kenya and Cherangani during the maximum of the last glacia- tion (the 'Mt Kenya hypothermal'), which was calculated by Coetzee (1967: 88) to imply a decrease in annual mean temperatures by some 5*1"-8*8"C. Similarly Mor- rison (1968) concluded that near the site investigated by him at Muchoa in south-west Uganda the vegetation at the same time must have approached that at present occurring in the ericaceous belt, from which he calculated for the relevant epoch an annual mean temperature 5" C lower than today. In my opinion, however, the evidence for these con- clusions is insufficient, Occurrence of afroalpine vegetation has been deduced mainly by high pollen percentages of grasses and Compositae, and of ericaceous belt vegeta- tion largely by grasses plus Artemisiu, Stoebe and Cltzortia together with Ericaceae. None of these genera or families are, however, by any means restricted to the afroalpine
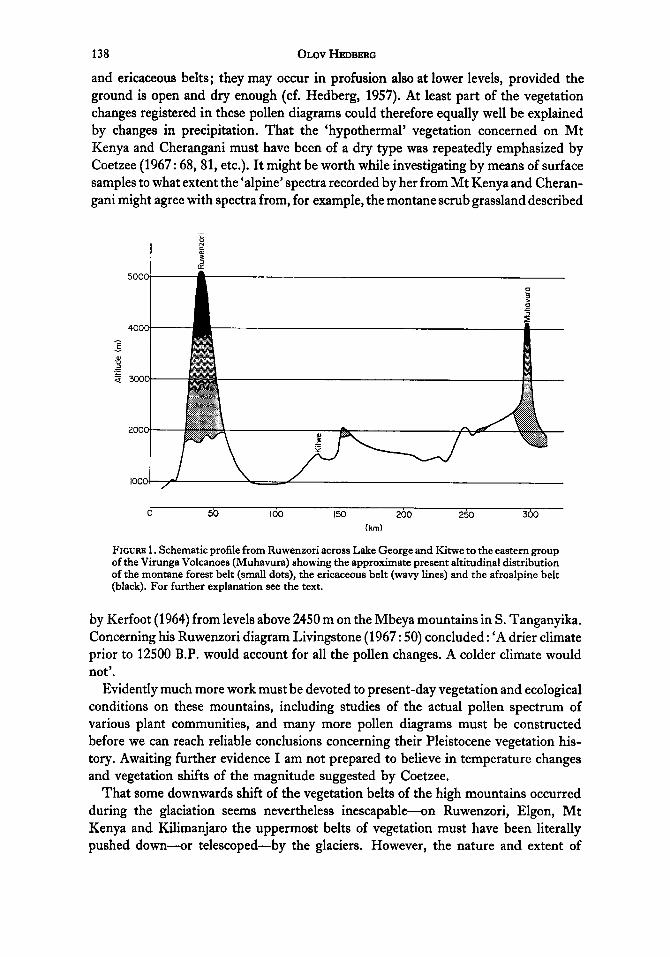
138 OLOV HEDBERG
and ericaceous belts; they may occur in profusion also at lower levels, provided the ground is open and dry enough (cf. Hedberg, 1957). At least part of the vegetation changes registered in these pollen diagrams could therefore equally well be explained by changes in precipitation. That the ‘hypothermal’ vegetation concerned on Mt Kenya and Cherangani must have been of a dry type was repeatedly emphasized by Coetzee (1967: 68, 81, etc.). It might be worth while investigating by means of surface samples to what extent the ‘alpine’ spectra recorded by her from Mt Kenya and Cheran- gani might agree with spectra from, for example, the montane scrub grassland described
0 50 I00 150 200 250 300 (km)
FIGURE 1. Schematic profile from Ruwenzori across Lake George and Kitwe to the eastern group of the Virunga Volcanoes (Muhavura) showing the approximate present altitudinal distribution of the montane forest belt (small dots), the ericaceous belt (wavy lines) and the afroalpine belt (black). For further explanation see the text.
by Kerfoot (1964) from levels above 2450 m on the Mbeya mountains in S. Tanganyika. Concerning his Ruwenzori diagram Livingstone (1967 : 50) concluded : ‘A drier climate prior to 12500 B.P. would account for all the pollen changes. A colder climate would not’.
Evidently much more work must be devoted to present-day vegetation and ecological conditions on these mountains, including studies of the actual pollen spectrum of various plant communities, and many more pollen diagrams must be constructed before we can reach reliable conclusions concerning their Pleistocene vegetation his- tory. Awaiting further evidence I am not prepared to believe in temperature changes and vegetation shifts of the magnitude suggested by Coetzee.
That some downwards shift of the vegetation belts of the high mountains occurred during the glaciation seems nevertheless inescapable-on Ruwenzori, Elgon, Mt Kenya and Kilimanjaro the uppermost belts of vegetation must have been literally pushed down-or telescoped-by the glaciers. However, the nature and extent of
EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 139
this shift remains to be established. It should also be remembered that below the firn- line glacier tongues keep in the main to valley bottoms, leaving the intervening ridges ice-free up to high level (cf. Nilsson, 1932), and that dense afroalpine vegetation may grow close to the side of a glacier (Hedberg, 1964: Fig. 9; 1968: 186). To what extent the montane forest belt was also lowered is hardly possible to deduce from the palaeoclimatical and pollen-analytical evidence available. Its lower limit on most mountains has been pushed up in historical time by exploitation for agriculture (cf. Hedberg, 1951).
What possibilities may then have existed for contacts between the biotas of the different mountains during the glaciations? The profile in Fig. 1 demonstrates the
f 500
400 - E - al 3 +.
2 300 ._
200
0 100 200 300 400 500 600 (km)
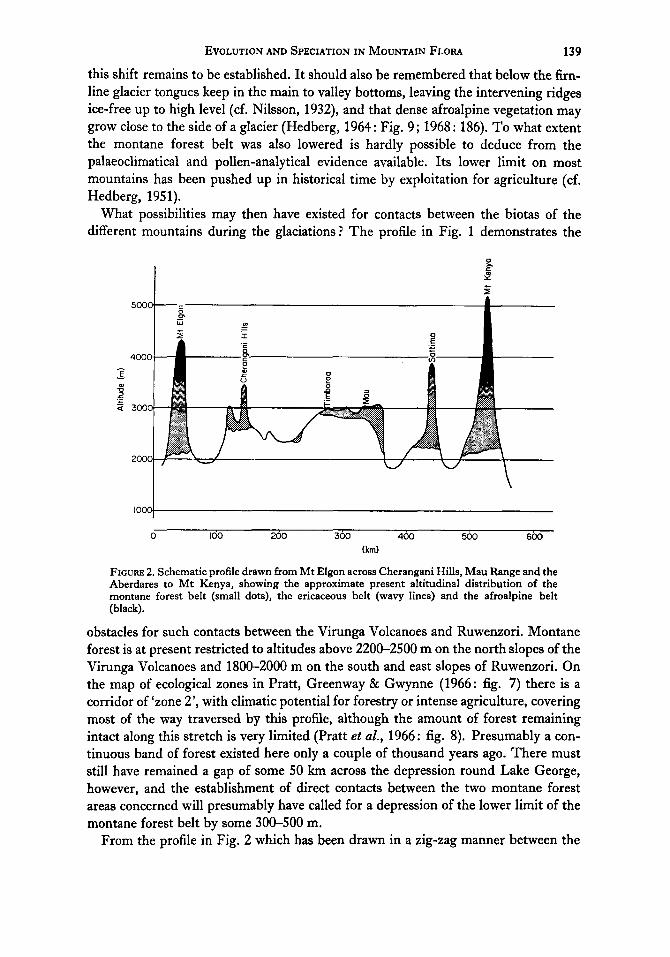
FIGURE 2. Schematic profile drawn from Mt Elgon across Cherangani Hills, Mau Range and the Aberdares to Mt Kenya, showing the approximate present altitudinal distribution of the montane forest belt (small dots), the ericaceous belt (wavy lines) and the afroalpine belt (black).
obstacles for such contacts between the Virunga Volcanoes and Ruwenzori. Montane forest is at present restricted to altitudes above 2200-2500 m on the north slopes of the Virunga Volcanoes and 1800-2000 m on the south and east slopes of Ruwenzori. On the map of ecological zones in Pratt, Greenway & Gwynne (1966: fig. 7) there is a corridor of ‘zone 2’, with climatic potential for forestry or intense agriculture, covering most of the way traversed by this profile, although the amount of forest remaining intact along this stretch is very limited (Pratt et al., 1966: fig. 8). Presumably a con- tinuous band of forest existed here only a couple of thousand years ago. There must still have remained a gap of some 50 km across the depression round Lake George, however, and the establishment of direct contacts between the two montane forest areas concerned will presumably have called for a depression of the lower limit of the montane forest belt by some 300-500 m.
From the profile in Fig. 2 which has been drawn in a zig-zag manner between the
140 OLOV HEDBBRC
mountains concerned, and from the maps in Pratt et al. (1966), it appears that before the destruction of so much forest by man there would presumably have been direct contacts between the montane forests of Elgon, Cherangani and the Mau Range. The climatic lower limit of the montane forest belt on Aberdare and Mt Kenya is certainly a good deal lower than its actual limit, and direct contacts between these montane forests and those of Mau, Cherangani and Elgon may well have existed here under present climatic conditions before the intervention by agricultural man. The obstacles to similar contacts between the montane forest ecosystems of Elgon and Ruwenzori are considerably larger, because of a long gap of intervening lowland below 1200 m altitude. Fairly recent direct contacts between all or most of the now isolated areas of montane forest in east Africa were nevertheless deemed necessary by Moreau (1963) to explain the present distribution of montane birds and other organisms.
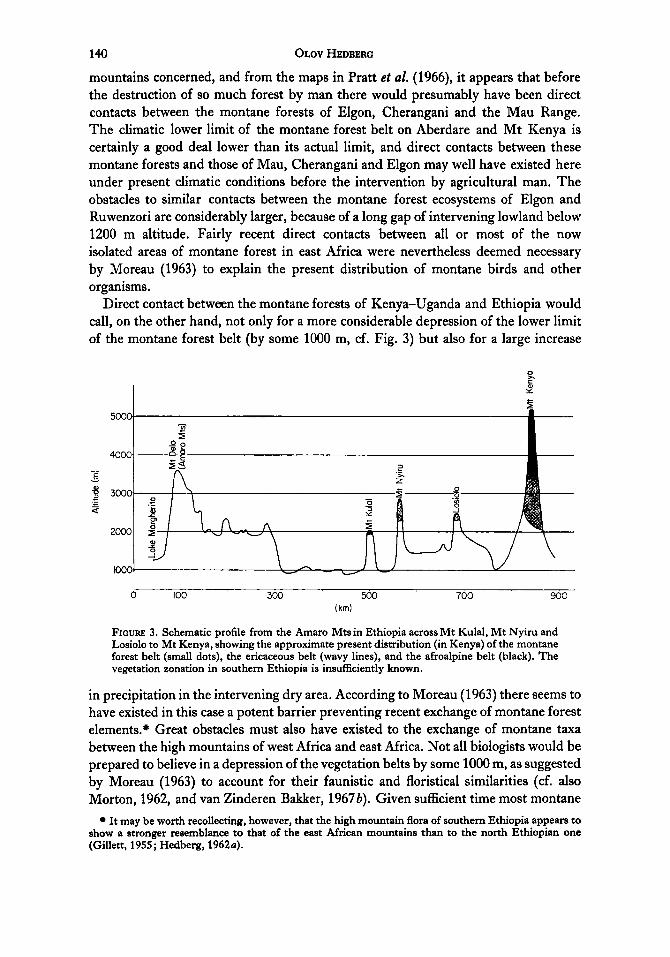
Direct contact between the montane forests of Kenya-Uganda and Ethiopia would call, on the other hand, not only for a more considerable depression of the lower limit of the montane forest belt (by some 1000 m, cf. Fig. 3) but also for a large increase
0 100 300 500 700 900 (km)
FIGURE 3. Schematic profile from the Amaro Mts in Ethiopia across Mt Kulal, Mt Nyiru and Losiolo to Mt Kenya, showing the approximate present distribution (in Kenya) of the montane forest belt (small dots), the ericaceous belt (wavy lines), and the afroalpine belt (black). The vegetation zonation in southern Ethiopia is insufficiently known.
in precipitation in the intervening dry area. According to Moreau (1963) there seems to have existed in this case a potent barrier preventing recent exchange of montane forest elements." Great obstacles must also have existed to the exchange of montane taxa between the high mountains of west Africa and east Africa. Not all biologists would be prepared to believe in a depression of the vegetation belts by some 1000 m, as suggested by Moreau (1963) to account for their faunistic and floristical similarities (cf. also Morton, 1962, and van Zinderen Bakker, 1967b). Given sufficient time most montane
It may be worth recollecting, however, that the high mountain flora of southern Ethiopia appears to show a stronger resemblance to that of the east African mountains than to the north Ethiopian one (Gillett, 1955; Hedberg, 19620).

EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 141
forest species of animals and plants should, I believe, be able to pass even a fairly wide gap without depending upon continuous distribution of the whole ecosystem. Time has indeed been available here on quite a different scale from that in the formerly glaciated parts of Europe, where dispersal of plants and animals has nevertheless been remarkably efficient (cf. Webb, 1966).
One potent vector of transport which has received little attention although it has been available in tropical Africa is cyclones. Admittedly, cyclones are rare in east Africa-there are only two on record, with an interval of 80 years (Sansom, 1953). During the time of, say, one million years such a frequency would, however, by no means be insignificant. I can find no better way of illustrating their potentialities for the dispersal of plant diaspores, birds and insects than to quote the report on the Lindi cyclone, which struck the coast of southern Tanganyika near Lindi on 15 April, 1952, destroying, inter aZia, most of the township of Lindi, and uprooting forest trees and sisal plants over large areas. Before it crossed the coastline the centre of the cyclone passed a small ship a few miles off the coast, the captain of which stated in his report that in the calm storm centre birds and insects were ‘falling all over the ship. Many were dead and all were dazed’. Commenting on this the meteorological author said: ‘Their condition might be explained by the strain of their buffeting by the storm; but it is at least equally plausible to suggest that they had been swept up to great heights in the inner ring of the storm and fell through the subsiding air of the central calm.’ (Sansom, 1953 : 9.) A cyclone like this will certainly have a large ‘collecting area’ for air masses containing wind-blown organisms and diaspores, and when these are swept up to great height in the central whirl they may evidently be transported over considerable dis- tances before they are released. The track of the present cyclone traversed much of southern Tanganyika (Sansom, 1953).
AGE OF ISOLATION OF AFROALPINE ENCLAVES
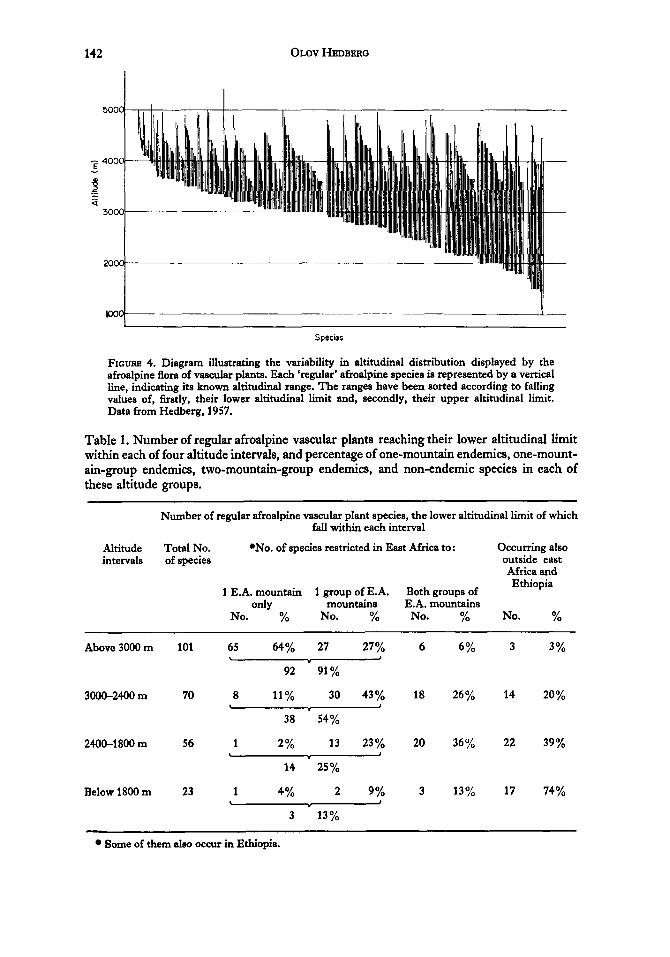
While direct migration of forest elements between at least some of the relevant mountains seems plausible in the recent past the possibilities for similar dispersal of the afroalpine flora would at first sight appear very limited. Both from phytogeographical and ecological points of view however, this flora is rather heterogeneous (Hedberg, 1961, 1965). By definition it comprises all species known to occur in the afroalpine belt. Even when some accidental afroalpines are left out, the altitudinal range of the remaining species shows a very wide variation (Fig. 4). Some are entirely confined to the afroalpine belt, others occur also in the ericaceous belt, and many occur even far down in the montane forest belt. The possibilities for inter-mountain dispersal of these species must be assumed to be largely dependant upon their altitudinal range-those which regularly occur in the lower part of the montane forest belt should have found much greater opportunities for dispersal during glacial or pluvial epochs than those which are confined to higher levels. This assumption is strongly supported by analysis of the percentage of endemics in various altitudinal groups (Table 1). That group of species which do not occur below 3000 m consists mainly of endemics-only 3 % of them occur outside the mountain areas of east Africa and Ethiopia. The lower down the mountain the species descend, the lower becomes the percentage of endemics, especially
142 OLOV HEDBERC
Y
Species
FIGURE 4. Diagram illustrating the variability in altitudinal distribution displayed by the afroalpine flora of vascular plants. Each ‘regular’ afroalpine species is represented by a vertical l i e , indicating its known altitudinal range. The ranges have been sorted according to falling values of, firstly, their lower altitudinal limit and, secondly, their upper altitudinal limit. Data from Hedberg, 1957.
Table 1. Number of regular afroalpine vascular plants reaching their lower altitudinal limit within each of four altitude intervals, and percentage of one-mountain endemics, one-mount- ah-group endemics, two-mountain-group endemics, and non-endemic species in each of these altitude groups.
Number of regular afroalpine vascular plant species, the lower altitudinal limit of which fall within each interval
Altitude Total No. +No. of species restricted in East Africa to: Occurring also intervals of species outside east
Africa and Ethiopia
1 E.A. mountain 1 group of E.A. Both groups of onlv mountains E.A. mountains
NO. % No. % No. % No. %
Above 3000 m 101 65 64% 27 27% 6 6% 3 3% \ ,
92 91%
3000-2400 m 70 8 11% 30 43% 18 26% 14 20% , I
38 54%
2400-1 800 m 56 1 2% 13 23% 20 36% 22 39% \ I
14 25%
Below 1800m 23 1 4% 2 9% 3 13% 17 74%
3 13% ~
Some of them also occur in Ethiopia.

EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 143
of one-mountain endemics. These results indicate that the ecological islands formed by the upper parts of the high mountains, particularly their afroalpine (and ericaceous) belt(s) have been effectively isolated from each other much longer than their zones of montane forest. The possibilities for earlier continuous distribution areas of these ‘qualified’ afroalpines are so slight that their migration between the mountains must be assumed to have taken place mainly by independent long-distance dispersal (Hedberg, 1961)” The vectors of dispersal between the mountains for such afroalpine vascular plants are evidently beyond observation or experimental study, but also in this case I would like to suggest cyclones as a plausible agent.
Establishment of an afroalpine vascular plant species on a new mountain may have been facilitated in many cases by the occurrence of vast lava fields and ash screes, created by volcanic eruptions and acting as ‘agar plates’ for alien diaspores of ecologi- cally adapted species. The patches of open soil which are common in stabilized vegetation in the upper part of the afroalpine belt may not be so favourable in this respect, since most seedlings on these are often eliminated by solifluction (Hedberg, 1964: 33).
DEGREE OF DIFFERENTIATION IN VARIOUS GROUPS
Detailed morphological studies of the afroalpine flora have demonstrated that the degree of differentiation between different mountain populations differs considerably in different groups of plants (Hedberg 1957: 374; 1958: 187). Some species occur on many or all of the high mountains concerned without displaying any perceptible morphological differentiation between their isolated mountain populations, e.g. Sagina afroalpina, Arabis alpina and Subularia monticola. In other cases different mountain populations of one species show statistical differences but their variation ranges overlap so much as to preclude taxonomic distinction, e.g. in Alchemilla john- stonii, Veronica glandulosa and Valeriana kilimandscharica (concerning the latter see Kokwaro, 1968). In yet other cases different mountain populations considered to be of common ancestry present distinctly discontinuous variation ranges, making taxo- nomic separation possible, as in the Philippia trimera group, the Lobelia deckenii group, and the species pair Romulea congoensis-R. keniensis. The results of this comparison indicate that the speed of evolutionary diversification has been comparatively low in the afroalpine representatives of the families Juncaceae, Caryophyllaceae, Cruciferae, Crassulaceae, Labiatae and Rubiaceae, and comparatively rapid in the genera Al- chemilla, Philippia, the Giant Lobelias, and in some genera of the Compositae, notably Helichrysum and Senecio (cf. Hedberg, 1957: 376). This generalization holds good also is the comparison is restricted to such taxa which have not been found below 2400 m, and which consequently would seem to have had small chances of migrating between the mountains by other means than long-distance dispersal of diaspores.
The lower percentage of endemics among those species occurring down to 2400 m altitude may not necessarily imply that the latter have had access to more continuous migration routes between the mountains. The explanation may at least partly be that such species can utilize a much larger area on each mountain-both for the production of diaspores on one mountain and for successful reception of them on another-than those species restricted to areas above 3000 m.

144 OLOV HEDBERC
GENETIC SYSTEMS, POLLINATION BIOLOGY, POLYPLOIDY
In the earlier part of this paper I have stressed that the afroalpine flora offers beautiful examples of evolutionary differentiation under the influence of geographical isolation, and I have tried to analyse the probable extent of isolation involved. It may now be time to devote some attention to what is known of the mode of evolution in this flora.
The genetic systems of afroalpine vascular plants have been very little studied. Sexual propagation seems to be much more common than apomixis, although apomixis (probably facultative) occurs in at least some species of Alchemilla (Hjelmqvist, 1956 ; Hedberg, 1957 : 281). A considerable proportion of the sexually reproducing afroalpine plants nevertheless have the capacity of reproducing also by asexual means, such as branching of the rhizome or caudex (Carex spp., Haplosciadium,Haplocarpha, etc.), or by basal adventitious shoots (as in most tufted, cushion-forming or tussock-forming plants, e.g. most grasses, Sagina spp., Alchemilla spp., Valerianu kilimandscharica, and in most Giant Lobelias (cf. Hedberg, 1957 : Plate 6A). Several sexually reproducing species are probably largely autogamous, e.g. Montia fontana, Subularia monticola, Sibthorpia europaea, and several grasses, but detailed knowledge is lacking.
The pollination biology is unknown for most afroalpines (cf. Hedberg, 1964: 38). Ornithogamy occurs in at least the Giant Lobelias and in Proteas, both of which are regularly visited by the sunbird Nectarinia johnstonii; the Giant Senecios also are said to be often visited by the same sunbirds (Hedberg, 1964). Although numerous species of insects occur in the afroalpine belt, e.g. on flowering specimens of Helichrysum and Euryops, etc. (Salt, 1954; Hedberg, 1964; Coe, 1967) no detailed observations of insect pollination are known to me. Wind pollination is certainly important in grasses, sedges, Alchemilla spp., and some Ericaceae.
The role of polyploidy for speciation seems to be relatively limited in the afroalpine flora. Differences in ploidy between different mountain populations of one species are indicated only for Deschampsia jlexuosa, although more than one chromosome number from one and the same species has been encountered also in a few other cases (Antho- xanthum, Pentaschistis, Poa, Anagallis and Senecio-cf. Hedberg, 1957).
MODE OF EVOLUTION
Most mountain populations of at least such ‘qualified’ afroalpines which do not occur below the 3000 m level are likely to have originated from one or a few diaspores brought by long-distance dispersal. From the outset each population of this kind will represent a very small random sample of the gene pool of the parent species. As long as the popula- tion was small and perhaps fluctuating in sue, evolution may be expected to have been comparatively rapid and genetic drift may easily have occurred. A probable example of the latter is provided by the vicarious species of the Lobelia deckenii group, which differ mainly in some minute details of splitting of the corolla and pubescence of the bracts, sepals, petals and anthers, for which it is difficult to visualize any selective advantage (cf. Plate 1 : A, B, D). The main pollinator for all of them seems to be the same species of Nectarink.
In other cases the differentiation into distinct taxa seems to have been aided by dif- ferential selection, as in the vicarious species pair Lobelia wollastonii-L. telekii. These

EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 145
two differ, inter alia, in the height of the stem carrying the inflorescence, in the length and pubescence of the bracts, the size of the flowers, etc. Their differences seem to be well adjusted to the ecological differences between the almost permanently wet afro- alpine environment on Ruwenzori and the Virunga Volcanoes, where the former species often occurs in dense Dendrosenecio forest, and the much drier afroalpine vegetation on Elgon, Aberdare and Mt Kenya, where Lobelia telekii occurs as a rule in open Alchemilla scrub (cf. Hedberg, 1964: Figs 46-48 and Plate 1 : C here).
Beautiful examples of geographically and altitudinally vicarious taxa are furnished by the Giant Lobelias and Giant Senecios, which also display some remarkable ecolo- gical adaptations to the inhospitable afroalpine climate (Hedberg, 1964). The former are represented in the montane forest belt by a few species with lax and sometimes branched stems, considered by Hauman (1933) to approach the ancestral forms of all the relevant species. Their afroalpine representatives are more robust with dense leaf rosettes and compact inflorescences, creating ameliorated microclimates of their own(seeillustrationsinHedberg, 1964). Thesespeciesfallnaturallyinto three groups (the Lobelia deckenii group, the L. telekii group, and the L. lanuriensis group), each of which is represented by at most one species on each of the high mountains. This indicates, of course, that those groups are older than the differentiation between the species within each of them, and that hybridization between the groups is insignificant. Only one, sterile, inter-group hybrid is known to me, viz. L. keniensis x L. telekii (see Hed- berg, 1957: 188, 334).
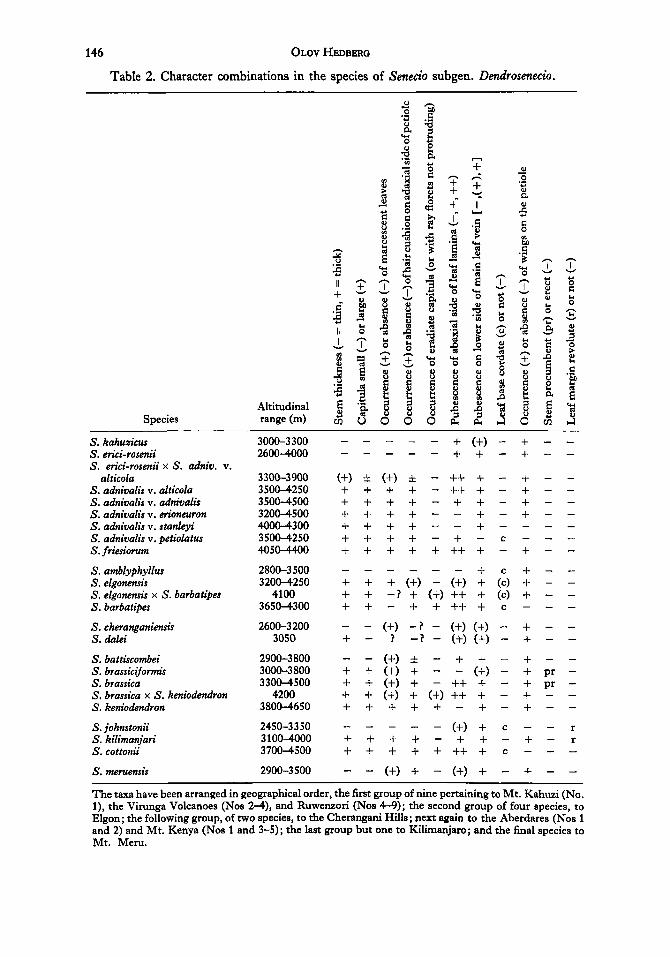
The taxonomic interrelations between the Giant Senecios are more complicated. On each mountain harbouring more than one species there is a succession of altitudi- nally more or less vicarious species, so that the low-level ones are thin-stemmed with open, few-leaved leaf rosettes, thin, caducous and sparsely pubescent leaves, and small capitula with long ray-florets, etc., whereas high-level species tend to have shorter and more robust stems, dense leaf rosettes closing up at night and formed by thick leaves, which often are densely pubescent below and marcescent, large capitula without or with small ray-florets, etc. (cf. Hauman, 1935). Superimposed upon this pattern there occurs variation in some other characters of less obvious ecological importance such as the occurrence of revolute leaf margins, cordate or attenuate leaf bases, winged or unwinged petioles, and a very remarkable type of procumbent stem (see Table 2). The character combinations accounted for in this table would seem to indicate that, for instance, the three species known from Kilimanjaro are more closely related to one another than, e.g., S. johnstonii to any of the other thin-stemmed forest species (S. eyicirosenii, S. amblyphyllus, and S. battiscombei). AS indicated in the table hybridiza- tion is known or inferred in several cases (cf. Hedberg, 1957: 227, 356), and extensive introgression between two species seems to occur on the Virunga Volcanoes (Hedberg, 1957: 369). The variation pattern in the collective species S. adnivalis on Ruwenzori is insufficiently known, but at least within this species the variation appears so continuous as to suggest hybridization. Gene exchange between different species on one mountain Seems plausible in several cases
The variation pattern displayed by Dendrosenecio might be explained by parallel adaptation of (upper) montane forest taxa to afroalpine conditions, complicated by occasional long distance dissemination of diaspores. The closely related species pair
10
146 OLOV HBDBERC
Table 2. Character combinations in the species of Senecio subgen. Dendrosenecio.
S. kahuzicus S. erici-rosenii S. erici-rosenii x S. adniv. v.
S. adnivalis v. alticola S . adnivalis v. adnivalis S. adnivalis v. erioneuron S. adnivalis v. stanleyi S . adnivalis v. petiolatus S . friesiorum
S. amblyphylh S. elgonensis S. elgonensis x S. barbatipes S. barbatipes
S . cheranganiensis s. &lei
S. battiscombei S. brassiciformis S. brassica S. brassica x S. keniodendron S. keniodendron
S . johnstonii S. kilimanjari S. cottonii
S. meruensis
alticola
3000-3300 2600-4000
3300-3900 3500-4250 3500-4500 3200-4500 4000-4300 3500-4250 4050-4400
2800-3 500 3200-4250 4100
3650-4300 2600-3 200
3050
2900-3 800 3000-3800 3300-4500 4200
3800-4650
2450-3350 3 1 OO-4000 3700-4500
2900-3500
- - - -
(+I z t + + + + + + + + + + + + - - + + + + + + - - + - - - + + + + + + + + - - + + + + - -
The taxa have been arranged in geographical order, the first group of nine pertaining to Mt. Kahuzi (No. l), the Virunga Volcanoes (Nos 24), and Ruwenzon (Nos 4-9); the second group of four species, to Elgon; the following group, of two species, to the Cherangani Hills; next again to the Aberdares (Nos 1 and 2) and Mt. Kenya (Nos 1 and 3-5); the last group but one to Kilimanjaro; and the final species to Mt. Meru.

EVOLUTION AND SPECIATION IN MOUNTAIN FLORA 147
Senecio brassica and brassicijiimis must obviously represent a case of such inter- mountain dispersal, and S. kahuxints on M t Kahuzi is so strikingly intermediate between S. erici-rosenii and S. adnivalis var. alticola that it might easily be visualized as a recent introduction from the hybrid population between the latter occurring on the Virunga Volcanoes.
In conclusion I would like to comment upon the meaning of the term speciation as used in the title of my contribution. To a zoologist, speciation would in most cases imply the formation of internal barriers to interbreeding between two or more morpho- logically distinguishable population systems. Botanists are on the whole less demanding in their definitions-to most of us speciation means simply the formation of genetically or spatially isolated and morphologically distinguishable population systems-or, alas, groups of specimens. This may appear very primitive to many of you, but for the larger part of the plant kingdom there is no way around it at present. To my knowledge only one investigation has been made of the compatibility of different populations of an afroalpine species, and that is my own study of Arabis alpina (Hedberg, 1962b). There is certainly a wide scope for experimental taxonomy in the afroalpine flora, but the task is not easy. The Giant Senecios I have just been discussing are, for instance, very slow-growing with irregular flowering periods perhaps 10-20 years apart, and very difficult to keep in cultivation. For some time to come we shall therefore presumably have to be satisfied with the sort of data I have presented in this paper.
REFERENCES
AUER, V., 1958, The Pleistocene of Fuego-Patagonia. Part 11: The history of the flora and vegetation.
CARCASSON, R. H., 1964. A preliminary survey of the zoogeography of African butterflies. E. Afr. Wild
COE, M. J., 1967. The ecology of the Alpine zone of Mount Kenya. Monographiue biol. 17: I-VIII,
COETZEE, J. A., 1964. Evidence for a considerable depression of the vegetation belts during the upper
COETZEE, J. A., 1967. Pollen analytical studies in east and southern Africa. In E. M. van Zinderen Bakker
EMILIANII, C., 1958. Ancient temperatures. Scient. A m . Feb. 1958: 54-63. FLINT, R. F. & DEEVEY, E. S., JR., 1951. Radiocarbon dating of late-Pleistocene events. Am.J. Sci. 249:
GAGE, M., 1961. New Zealand glaciations and the duration of the P1eistocene.J. Glaciol. 3: 940-943. GILLETT, J. B., 1955. The relation between the highland floras of Ethopia and British East Africa.
GREGORY, J. W., 1894. Contributions to the geology of British East Africa. Part I. The glacial geology
HAUMAN, L., 1933. Les ‘Lobelia’ g6ants des montagnes du Congo belge. Mkm. Inst. r. colon. belge, 1933:
HAUMAN, L., 1935. Les ‘Senecio’ arborescents du Congo. RewueZool. Bot. afr. 28: 1-76. HEDBERG, O., 1951. Vegetation belts of the East African mountains. Suensk bot. Tidskr. 45: 140-202. HEDBERG, O., 1955. A pollen-analytical reconnaissance in Tropical East Africa. Oikos, 5: 137-166. HEDBERG, O., 1957. Afroalpine vascular plants. A taxonomic revision. Symb. bot. upsal. 15 (1). HEDBERG, O., 1958. The taxonomic treatment of vicarious taxa. In 0. Hedberg (ed.), Systematics of to-
HEDBERG, O., 1961. The phytogeographical position of the afroalpine flora. Rec. Adw. Botany, 1: 914-
HEDBERG, O., 1 9 6 2 ~ . Mountain plants from southern Ethiopia, collected by Dr. John Eriksson, Ark.
HEDBERG, O., 19623. Intercontinental crosses in Arubis alpina L. Curyologiu, 15: 253-260. HEDBERG, O., 1964. Features of afroalpine plant ecology. Actu phytogeogr. w c . 49: 1-144.
Ann. Acad. Sci. Fennicae (A), 111 Geol.-Geogr. 50: 1-239.
LifeJ. 2: 122-157.
1-136.
Pleistocene on the east African mountains. Nature, Lond. 204: 564-566.
(ed.), Palueoecology of Africa, 3: I-XII: 1-146, Figs 1-22, PIS 1-8. Cape Town: A. A. Balkema.
257-300.
Webbia, 11: 459469.
of Mount Kenya.Q.Jlgeo1. SOC. Lond. 50 (1894): 515-530.
1-52.
day. Uppsala Univ. ibsskr. 1958: 6,186-194.
919.
Bot. (2), 4 (13): 421-135.

148 OLOV HEDBERC HEDBERG, O., 1965. Afroalpine flora elements. Webbia, 19: 519-529. HEDBERG, O., 1968. Taxonomic and ecological studies on the afroalpine flora of Mt Kenya. Hochgebirgs-
HJELMQVIST, H., 1956. The embryology of some African Alchemilla species. Bot. Notiser, 109: 21-32. KERFOOT, O., 1964. A preliminary account of the vegetation of the Mbeya Range, Tanganyika. Kirkia, 4:
KOKWARO, J. O., 1968. Valerianaceae in Milne-Redhead, E. & Polhill, R. M. (eds), Flora of Tropical
LIVINCSTONE, D. A., 1962. Age of deglaciation in the Ruwenzori Range, Uganda. Nature, Lond. 194:
LIVINGSTONE, D. A., 1967. Postglacial vegetation of the Ruwenzori Mountains in equatorial Africa.
forschung, 1.171-194.
191-206.
East Africa.
859-860.
Ecol. Monoer. 37: 25-52. MACKINDER, k. J., 1900. A journey to the summit of Mount Kenya, British East Africa. Geogrl J. 15
(1900): 453-486. MEYER,-H., 1900. Der Kilimandjaro. Reisen und Studien. Berlin. MOREAU, R. E., 1963. Vicissitudes of the African biomes in the late Pleistocene. Proc. 2001. SOC. Lond.
MORRISON, M. E. S., 1966. Low-latitude vegetation history with special reference to Africa. Proc. int.
MORRISON, M. E. S., 1968. Vegetation and climate in the uplands of south-westem Uganda during the
MORTON, J. K., 1962. The upland floras of West Africa their composition, distribution and significance
NILSSON, E., 1932. Quaternary glaciations and pluvial lakes in British East Africa. Geogr. Annlr, 13
NILSSON, E., 1940. Ancient changes of climate in British East Africa and Abyssinia. Georg. Annlr, 22:
NILSSON, E., 1949. The pluvials of east Africa. Geogr. Annlr, 31: 204-211. OSMASTON, H. A., 1958. Pollen analysis in the study of the past vegetation and climate of Ruwenzori
and its neighbourhood. B.Sc. thesis, Oxford. OSMASTON, H. A., 1967. The sequence of glaciations in the Ruwenzori and their correlation with glacia-
tions of other mountains in East Africa and Ethiopia. In E. M. van ZinderenBakker (ed.), Palaeoecology of Africa, 2: 26-28.
PRATT, D. J., GREENWAY, P. J. & GWYNNE, M. D., 1966. A classification of east African rangeland, with an appendix on terminology. J. appl. Ecol. 3: 369-382.
Q U ~ E L , P., 1963. De I’application de techniques palynologiques ?i un territoire dbsertique. Paldoclimato- logie du quartenaire rdcent au Sahara. Changes of climate. Proc. Rome Symp. organized by U N E S C O
141: 395-421.
Symp. World clim. 8000-0 B.C., 142-148. Royal Meteorological Society, London.
later Pleistocene period. I. Muchoya Swamp, Kigezi District. J. Ecol. 56: 363-384.
in relation to climate changes. C.ru. I V e B n u . Plhi2re A.E.T.F.A.T., 391-409.
(1931): 241-348.
1-79.
&f W M O , 243-249. SALT, G., 1954. A contribution to the ecology of upper Kilimanjaro. J. Ecol. 42: 375423. SANSOM, H. W., 1953. The Lindi Cyclone 15 April 1952. Mem. E. Afr. met. Dep. 3 : 1,l-16, Figs 1-5. WEBB, D. A., 1966. Dispersal and establishment: what do we really know I In J. G. Hawkes (ed.), Repro-
ZINDEREN BAKKER, E. M. VAN, 1964. A pollen diagram from equatorial Africa, Cherangani, Kenya.
ZINDEREN BAKKER, E.M., VAN, 1967a. Palaeoecology of Africa, 2 (1964-1965). Cape Town: A. A. Balkema ZINDEREN BAKKER, E. M. VAN, 1967 b. Upper Pleistocene and Holocene stratigraphy and ecology on the
basis of vegetation changes in Sub-Saharan Africa. In Bishop, W. W. & Clark, J. D. (ed.), Background to Evolution in Africa. Chicago: Univ. Press.
ductive Biology and Taxonomy of Vascular Plants. B.S.B.I. Conference Reports, No. 9.
Geologie Mijnb. 43 : 123-1 28.
EXPLANATION OF PLATE
PLATE 1
A. Lobelia deckenii, flowering specimen. Tanzania, Kilimanjaro, S. slope above Mweka, in the lower part of the ericaceous belt, c. 3000 m. The flowers are barely visible between the wide ovate bracts. Photo: 0. Hedberg, Nov. 1967. B. Lobelia keniensis, flowering specimen. Kenya, Mt. Kenya, Teleki Valley, in the afroalpine belt, C . 4200 m. Photo: 0. Hedberg, July 1948. Lobelia keniensis and L. deckenii are closely related vicarious species (cf. text). C. Lobelia telekii, flowering specimen. The flowers are hidden between the long, hairy bracts. Kenya, Mt. Kenya, Teleki Valley, in the afroalpine belt, C. 4200 m. Photo 0. Hedberg, July 1948. D. Lobelia keniensis, young flowering specimen photographed early in the morning. The leaves have just started to unfold from their tightly folded night position (cf. Hedberg, 1964). Note the snow cover on the ground. Kenya, Mt. Kenya, Teleki Valley, in the afroalpine belt,c.4200m. Photo: 0. Hedberg, July 1948.

Related Documents











