Evidence that progestins play an important role in spermiation and pheromone production in male sea lamprey (Petromyzon marinus) Mara Beth Bryan a,1 , Yu-Wen Chung-Davidson a , Jianfeng Ren b,2 , Stephen Bowman a , Alexander P. Scott c , Mar Huertas a , Michael Patrick Connolly d , Weiming Li a,⇑ a Department of Fisheries and Wildlife, Michigan State University, 13 Natural Resources Building, 480 Wilson Road, East Lansing, MI 48824, USA b College of Fisheries and Life Sciences, Shanghai Ocean University, 999 Huchenghuan Road, Shanghai 201306, China c The Centre for Environment, Fisheries and Aquaculture Science, Barrack Road, Weymouth, Dorset DT4 8UB, UK d Department of Biochemistry and Molecular Biology, Michigan State University, 212 Biochemistry Building, 603 Wilson Road, East Lansing, MI 48824, USA article info Article history: Received 10 April 2014 Revised 30 December 2014 Accepted 14 January 2015 Available online 23 January 2015 Keywords: Progestin Progesterone Lamprey Steroid receptor Pheromone production abstract Progestins (progestogens, C 21 steroids) have been shown to regulate key physiological activities for repro- duction in both sexes in all classes of vertebrates except for Agnathans. Progesterone (P) and 15a-hydrox- yprogesterone (15a-P) have been detected in sea lamprey (Petromyzon marinus) plasma, but the expression patterns and functions of putative progestin receptor genes have not yet been investigated. The first objective of this study was to determine the differences in mRNA expression levels of nuclear progestin receptor (nPR) and the membrane receptor adaptor protein ‘progesterone receptor membrane component 1’ (pgrmc1) in putative target tissues in males at different life stages, with and without lam- prey GnRH-I and -III treatment. The second objective was to demonstrate the function of progestins by implanting prespermiating males (PSM) with time-release pellets of P and measuring the latency to the onset of spermiation and plasma concentrations of sex pheromones and steroids. The third objective was to measure the binding affinity of P in the nuclear and membrane fractions of the target tissues. Expression levels of nPR and pgrmc1 differed between life stages and tissues, and in some cases were dif- ferentially responsive to lamprey GnRH-I and -III. Increases in nPR and pgrmc1 gene expressions were cor- related to the late stages of sexual maturation in males. The highest expression levels of these genes were found in the liver and gill of spermiating males. These organs are, respectively, the site of production and release of the sex pheromone 3 keto-petromyzonol sulfate (3kPZS). The hypothesis that pheromone pro- duction may be under hormonal control was tested in vivo by implanting PSM with time-release pellets of P. Concentrations of 3kPZS in plasma after 1 week were 50-fold higher than in controls or in males that had been implanted with androstenedione, supporting the hypothesis that P is responsible for regulating the production of the sex pheromone. P treatment also accelerated the onset of spermiation. Saturation and Scatchard analyses of the target tissues showed that both nuclear and membrane fractions bound P with high affinity and low capacity (K D 0.53 pmol/g testis and 0.22 pmol/g testis, and B max 1.8 and 5.7 nM, respectively), similar to the characteristics of nPR and mPR in other fish. The fact that a high proportion of P was also converted in vivo to 15a-P means that it is not yet possible to determine which of these two steroids is the natural ligand in the sea lamprey. Ó 2015 Elsevier Inc. All rights reserved. 1. Introduction The sea lamprey (Petromyzon marinus) is one of the earliest extant vertebrates for which the hypothalamic–pituitary–gonadal (HPG) axis has been shown to coordinate and control reproduction (Sower et al., 2006). Research regarding lamprey steroids and receptors can yield insights into the evolution of steroids as hor- monal signals that have significant roles in development and reproduction (Bryan et al., 2008). Thus far, partial sequences of genes for only three nuclear steroid receptors have been amplified in sea lamprey (Thornton, 2001), which is half of the number observed in gnathostome vertebrates. The three genes are for an estrogen receptor (nER), a corticoid receptor (nCR), and a progestin receptor (nPR; Thornton, 2001). Additionally, the lamprey genome http://dx.doi.org/10.1016/j.ygcen.2015.01.008 0016-6480/Ó 2015 Elsevier Inc. All rights reserved. ⇑ Corresponding author. Fax: +1 517 432 1699. E-mail address: [email protected] (W. Li). 1 Present address: Energy Biosciences Institute, University of California, Berkeley, CA 94720, USA. 2 Visiting Michigan State University while carrying out this work. General and Comparative Endocrinology 212 (2015) 17–27 Contents lists available at ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

General and Comparative Endocrinology 212 (2015) 17–27
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Evidence that progestins play an important role in spermiation andpheromone production in male sea lamprey (Petromyzon marinus)
http://dx.doi.org/10.1016/j.ygcen.2015.01.0080016-6480/� 2015 Elsevier Inc. All rights reserved.
⇑ Corresponding author. Fax: +1 517 432 1699.E-mail address: [email protected] (W. Li).
1 Present address: Energy Biosciences Institute, University of California, Berkeley,CA 94720, USA.
2 Visiting Michigan State University while carrying out this work.
Mara Beth Bryan a,1, Yu-Wen Chung-Davidson a, Jianfeng Ren b,2, Stephen Bowman a, Alexander P. Scott c,Mar Huertas a, Michael Patrick Connolly d, Weiming Li a,⇑a Department of Fisheries and Wildlife, Michigan State University, 13 Natural Resources Building, 480 Wilson Road, East Lansing, MI 48824, USAb College of Fisheries and Life Sciences, Shanghai Ocean University, 999 Huchenghuan Road, Shanghai 201306, Chinac The Centre for Environment, Fisheries and Aquaculture Science, Barrack Road, Weymouth, Dorset DT4 8UB, UKd Department of Biochemistry and Molecular Biology, Michigan State University, 212 Biochemistry Building, 603 Wilson Road, East Lansing, MI 48824, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 April 2014Revised 30 December 2014Accepted 14 January 2015Available online 23 January 2015
Keywords:ProgestinProgesteroneLampreySteroid receptorPheromone production
Progestins (progestogens, C21 steroids) have been shown to regulate key physiological activities for repro-duction in both sexes in all classes of vertebrates except for Agnathans. Progesterone (P) and 15a-hydrox-yprogesterone (15a-P) have been detected in sea lamprey (Petromyzon marinus) plasma, but theexpression patterns and functions of putative progestin receptor genes have not yet been investigated.The first objective of this study was to determine the differences in mRNA expression levels of nuclearprogestin receptor (nPR) and the membrane receptor adaptor protein ‘progesterone receptor membranecomponent 1’ (pgrmc1) in putative target tissues in males at different life stages, with and without lam-prey GnRH-I and -III treatment. The second objective was to demonstrate the function of progestins byimplanting prespermiating males (PSM) with time-release pellets of P and measuring the latency tothe onset of spermiation and plasma concentrations of sex pheromones and steroids. The third objectivewas to measure the binding affinity of P in the nuclear and membrane fractions of the target tissues.Expression levels of nPR and pgrmc1 differed between life stages and tissues, and in some cases were dif-ferentially responsive to lamprey GnRH-I and -III. Increases in nPR and pgrmc1 gene expressions were cor-related to the late stages of sexual maturation in males. The highest expression levels of these genes werefound in the liver and gill of spermiating males. These organs are, respectively, the site of production andrelease of the sex pheromone 3 keto-petromyzonol sulfate (3kPZS). The hypothesis that pheromone pro-duction may be under hormonal control was tested in vivo by implanting PSM with time-release pellets ofP. Concentrations of 3kPZS in plasma after 1 week were 50-fold higher than in controls or in males thathad been implanted with androstenedione, supporting the hypothesis that P is responsible for regulatingthe production of the sex pheromone. P treatment also accelerated the onset of spermiation. Saturationand Scatchard analyses of the target tissues showed that both nuclear and membrane fractions bound Pwith high affinity and low capacity (KD 0.53 pmol/g testis and 0.22 pmol/g testis, and Bmax 1.8 and 5.7 nM,respectively), similar to the characteristics of nPR and mPR in other fish. The fact that a high proportion ofP was also converted in vivo to 15a-P means that it is not yet possible to determine which of these twosteroids is the natural ligand in the sea lamprey.
� 2015 Elsevier Inc. All rights reserved.
1. Introduction
The sea lamprey (Petromyzon marinus) is one of the earliestextant vertebrates for which the hypothalamic–pituitary–gonadal
(HPG) axis has been shown to coordinate and control reproduction(Sower et al., 2006). Research regarding lamprey steroids andreceptors can yield insights into the evolution of steroids as hor-monal signals that have significant roles in development andreproduction (Bryan et al., 2008). Thus far, partial sequences ofgenes for only three nuclear steroid receptors have been amplifiedin sea lamprey (Thornton, 2001), which is half of the numberobserved in gnathostome vertebrates. The three genes are for anestrogen receptor (nER), a corticoid receptor (nCR), and a progestinreceptor (nPR; Thornton, 2001). Additionally, the lamprey genome

18 M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27
contains DNA sequences similar to the ‘fast-acting’ membrane pro-gestin receptor (mPR) which has five variants, a, b, c, d, and e, inhigher vertebrates (Zhu et al., 2003a,b; Tang et al., 2005; Karteriset al., 2006; Pang and Thomas, 2011; Pang et al., 2013), and a pro-tein termed the ‘progesterone receptor membrane component 1’(PGRMC1; Falkenstein et al., 1996; Mourot et al., 2006; Cahill,2007; Thomas, 2008; Thomas et al., 2014). Until recently, this latterprotein has been the subject of much controversy (Cahill, 2007). Atthe time the present study was carried out (2005–2006), PGRMC1was accepted as a bona fide membrane receptor for P. However, it isnow (Thomas et al., 2014) accepted that this protein has moderatesteroid specificity for P and may have higher affinity for testoster-one and cortisol in Chinese hamster ovary cells (Meyer et al., 1996).Nevertheless, despite not being a membrane receptor itself, itspresence has been shown to be essential for the actual mPR func-tions. It has therefore been termed an ‘adaptor’ protein (Thomaset al., 2014). It must be pointed that the functionality of the estro-gen membrane receptor (and probably membrane receptors for allsorts of other compounds) is also dependent on the presence ofPGRMC1 (Thomas et al., 2014). In other words, increased expres-sion of the PGRMC1 gene is not a direct proof that this increaseis necessarily related to the activity of P.
At least two progestins, P and 15a-P have been identified inlamprey plasma, and the formation of the latter from the formerhas been demonstrated in vitro in testis tissue (Kime and Rafter,1981; Kime and Callard, 1982; Lowartz et al., 2003, 2004; Bryanet al., 2004). Several studies also showed that circulating concen-trations of these steroids, especially 15a-P, increased significantlyin response to injections of lamprey GnRH-I and -III (Deragon andSower, 1994; Gazourian et al., 2000; Bryan et al., 2004; Younget al., 2007). The facts that P and 15a-P are made by the testes ofspermiating (i.e., ripe-running) males, are released into the plasma,and are stimulated by lamprey GnRH-I and -III, strongly suggestthat progestins act as hormones in lampreys. However, further evi-dence is required, including the demonstration of the presence ofreceptor(s) for the steroids in putative target tissues and theadministration of the steroid to live fish causing physiological orbehavioral changes (or both).
The classic method for demonstrating the presence of steroidreceptors is to check the binding of tissue-derived proteins toradioactively-labeled steroids. We have used this approach todemonstrate the existence of a receptor for androstenedione (Ad)in the lamprey testis and kidney (Bryan et al., 2007). However,we were unable to obtain pure radiolabelled 15a-P. Thus, in thepresent study, we have only used radiolabelled P for the bindingassays.
Another approach for investigating the role of steroid receptorsis to investigate changes in the expression of their genes. In thepresent study, we have done this for nPR and pgrmc1, but not,unfortunately for mPR (as at the time these experiments were car-ried out in 2005–2006, the sequences for the various variants ofthis protein had not yet been identified in the lamprey genome).Our approach relies not just on the demonstration that mRNAsfor these proteins are present in lamprey, but that they occur in tis-sues that are the likely targets for progestin action, and that theirlevel of expression is influenced by reproductive stage and activity.
Receptor levels in tissues are known to change markedly withstage of maturation – as a mechanism for increasing or decreasingthe physiological response to hormones (Pasmanik and Callard,1988), and steroid receptor levels have been shown to change inresponse to gonadotropins (Thomas et al., 1997, 2001, 2005). Inchoosing which reproductive stages and which tissues to investi-gate for gene expressions, we turned to the literature on both lam-preys and teleosts. It is well-established that PSM injected withlamprey GnRH-III reach maturity faster (Deragon and Sower,1994) and that PSM injected with lamprey GnRH-I and -III also
have higher levels of 15a-P (Bryan et al., 2004; Young et al.,2007) and P (Deragon and Sower, 1994; Gazourian et al., 2000).Similarly in many teleosts, it has been shown that injection withgonadotropins or GnRH-agonists accelerates maturity and bringsabout a rise in progestin levels (Ueda et al., 1985; Mylonas et al.,1997; Vermeirssen et al., 1998). In teleosts, the progestins, in turn,have been shown to have direct effects on milt production includ-ing the cation content of seminal fluid, pH, fluidity and spermcapacitation (see review by Scott et al., 2010; Ueda et al., 1985;Miura et al., 1995). Furthermore, both nuclear and membrane pro-gestin receptors have been shown to be present in teleost testesand sperm (Thomas, 2000, 2003, 2008). Based on these observa-tions, we hypothesize that the lamprey testis is not just a sourceof progestins, but also a likely target organ for them.
In male sea lamprey, it is well established that production andrelease of 3-keto-petromyzonol sulfate (3kPZS), a powerfulfemale-attracting pheromone, is initiated at the onset of spermia-tion (Li et al., 2002; Siefkes et al., 2003; Yun et al., 2003) at essen-tially the same time that 15a-P levels are at their highest. Thus, wehypothesize that P and/or 15a-P might be responsible for stimulat-ing the synthesis or release of 3kPZS (or both). In other words, theliver, where the precursor of 3kPZS is synthesized (Brant et al.,2013; Li et al., 2002; Siefkes et al., 2003) and the gills, where the3kPZS is released (Siefkes et al., 2003; Brant et al., 2013), mightbe putative target organs for progestins.
To summarize, our goals in the present study were to character-ize progestin functions in male sea lamprey. The first objective wasto determine the differences in mRNA expression levels of nPR andpgrmc1 in putative target tissues in males at different life stages,with and without lamprey GnRH-I and -III treatment. The secondobjective was to demonstrate the function of progestins byimplanting PSM with time-release pellets of P and measuring thelatency to the onset of spermiation and plasma concentrations of3kPZS. The third objective was to measure the binding affinity ofP in the nuclear and membrane fractions of the target tissues.
2. Materials and methods
2.1. Animals and reagents
All experiments involving animals were approved by the Mich-igan State University Institutional Animal Use and Care Committee.PSMs were collected during upstream spawning migrations by U.S.Fish and Wildlife Service personnel in May 2005, and had massesof 231.2 ± 9.9 g and lengths of 48.8 ± 0.7 cm. Animals were heldat the Hammond Bay Biological Station (U.S. Geological Survey,Great Lakes Science Center, Millersburg, MI, USA) in tanks contain-ing approximately 160 l of continuous-flow water (2 l/min) fromLake Huron for at least 2 days prior to treatment applications. Toobtain spermiating males (SM), animals were held in tanks untilmilt was expressed from the cloaca upon gentle pressure. SMhad masses of 238.2 ± 11.3 g and lengths of 47.9 ± 0.8 cm. Temper-ature during acclimation and experimental periods was main-tained at 16 �C (± 1 �C). Parasitic-phase lamprey were obtainedfrom fisherman by USGS personnel, transported to Michigan StateUniversity (East Lansing, MI, USA) and held at temperatures of10 �C or less until sampling. Small parasitic-phase lamprey (SP)were collected in October 2005 and had masses of 100.0 ± 7.0 gand lengths of 40.0 ± 1.2 cm. Large parasitic-phase lamprey (LP)were collected in January 2006 and had masses of 246.6 ± 13.1 gand lengths of 50.8 ± 0.9 cm. All lamprey were anesthetized withMS-222 prior to experimental treatment. All reagents wereobtained from Sigma (St. Louis, MO, USA) unless otherwise noted.Lamprey GnRH-I (Sherwood et al., 1986) and -III (Sower et al.,1993) were synthesized by BAChem Peptides (King of Prussia, PA,USA).

Fig. 2. Neighbor-joining tree of vertebrate progesterone receptor membranecomponent 1 (PGRMC1) and 2 (PGRMC2) proteins. Values next to the branchesrepresent the percentage of bootstrap support from 1000 replicates. The tree isdrawn to scale, with branch lengths in the same units as those of the evolutionarydistances used to infer the phylogenetic tree. The PGRMC1 and PGMRC2 proteinsequences of human (Homo sapiens), mouse (Mus musculus), chicken (Gallus gallus),frog (Xenopus tropicalis), zebrafish (Danio rerio) and rainbow trout (Oncorhynchusmykiss) were downloaded from NCBI. The protein GenBank accession numbers werelisted in Table 1. Neighbor-joining tree was constructed based upon thesealignments using MEGA4 (Tamura et al., 2007) and the evolutionary distanceswere computed using the JTT matrix-based method (Jones et al., 1992). Thephylogenetic tree supports that the identity of this receptor in sea lamprey isorthologous to the PGRMC1 in other vertebrates.
M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27 19
2.2. Tissue collection
Only adults were used in lamprey GnRH-injection experiments.Both types of lamprey GnRH were dissolved in 0.9% saline andinjected intraperitoneally. Saline was used as a control (n = 8). Ani-mals were given two serial injections of 50 lg/kg, 24 h apart, asdescribed previously (Sower, 1989; Deragon and Sower, 1994;Gazourian et al., 2000; Bryan et al., 2004; Young et al., 2004). Bloodsamples (0.8–1.0 ml) were collected 24 h after the second injectionfrom the caudal vein using heparinized syringes, processed toobtain plasma, and stored at �20 �C until use. Tissues werecollected at 24 h after the second injection. Tissue samples forreal-time quantitative PCR (whole brain, gill, liver, and testes) werefrozen in liquid nitrogen, and stored at �80 �C until use. SP and LPwere given two serial injections of 0.9% saline, 24 h apart, andblood and tissues were collected as described above but at 6 h afterthe second injection. Sex was determined surgically during tissuecollection, and only samples from male parasitic-phase lampreywere included in the analyses.
2.3. Real-time quantitative PCR
All primers and amplicons were designed using Primer Expresssoftware (Applied Biosystems; Branchburg, NJ, USA). Primers andamplicons for nPR were designed based on NCBI Accession NumberAY028458, described by Thornton (2001). To design primers andamplicons for pgrmc1 (Fig. 1), a search was done on the tracearchive (Libants et al., 2009) and later confirmed on the sea lam-prey genome assembly (Smith et al., 2013). The identity of theputative lamprey pgrmc1 was confirmed by BLASTN and TBLASTXsearches of the NCBI Database (nr) and alignment with well-char-acterized pgrmc1 genes (Fig. 2). A phylogenetic tree was con-structed using neighbor-joining method based on thesealignments using MEGA4 (Tamura et al., 2007), and the result sup-port the identity of this receptor as the ortholog to pgrmc1. Theevolutionary distances were computed using the JTT matrix-basedmethod (Jones et al., 1992). All positions containing gaps and miss-ing data were eliminated from the dataset (complete deletionoption). From the most recent transcriptome assembly, severalmPRs were found (a/b: full length; c: partial sequence; d: fulllength; e: partial sequence), but no pgrmc2 was present (unpub-lished data).
For nPR, the forward primer was 50 CCG TCG GCC GTG TTA TTC30, the reverse primer was 50 CAT GAA CTA CAT CCG AGA GCT CAA30, the probe was TaqMan� MGB 50 TGC TGA TGA CCA CCT T 30 witha 6-FAM fluorescent label at the 50 end (with 30 MGB quencher),and the cDNA standard was 50 TAATACGACTCACTATAGGCATGAAC-TACATCCGAGAGCTCAACAAGGTGGTCATCAGCAAG AATAACACGGCCGACGGTCTATAGTGTCACCTAAAT 30. For pgrmc1, the forwardprimer was 50 CAG CAG ATC GTG GAG GAG TTC 30, the reverse
Fig. 1. ClustalW alignment of sea lamprey progestin receptor membrane component 1Accession No. NP_001117916).
primer was 50 CTT CCT CCT CGC CGA TTG T 30, the probe was Taq-Man� MGB 50 CCG CCA TCG TGT TCT 30 with a 6-FAM fluorescentlabel at the 50 end (with 30 MGB quencher), and the cDNA standardwas 50 TAATACGACTCACTATAGGCAGCAGATCGTGGAGGAGTTCG-GAGGCC GCCATCGTGTTCT CTGTGACAATCGGCGAGGAGGAAGTC-TATAGTGTCACCTAAAT 30.
Real-time quantitative PCR followed the method described by(Chung-Davidson et al., 2013a,b,c). Internal standard 40S ribo-somal RNA was confirmed to have no change among the treatmentgroups (data not shown). Data analysis was performed usingSequence Detection Systems v2.3 (Applied Biosystems). Statisticalanalysis was performed in two ways for pgrmc1 and nPR for eachtissue. To analyze whether the number of transcripts per ng of totalRNA differed by stage, a one-way ANOVA was performed, and wasfollowed by Fisher’s LSD tests if significant results (p < 0.05) wereobtained. To analyze the effect of injections of lamprey GnRH onthe number of transcripts per ng of total RNA, a two-way ANOVAwas performed in which both the stage of the animal and the treat-ment were independent variables. Fisher’s LSD tests were per-formed if significant results were obtained.
(PGRMC1, NCBI Accession No. KJ648627) to Oncorhynchus mykiss PGRMC1 (NCBI

20 M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27
2.4. In vivo functional experiments
To test the effect of progestins on male reproductive physiology,migratory lamprey were collected in traps on the St. Marys River(MI, USA) in July 2006 and transported to the Hammond Bay Bio-logical Station. Sixty-four fish were divided into eight tanks andacclimated at 16 ± 1 �C for 5 days. Two tanks were used per treat-ment: low-dose (15 mg) P pellet, high-dose (150 mg) P pellet, pla-cebo, and no implant. The 21-day release pellets formulated forfish were obtained from Innovative Research of America (Sarasota,FL, USA). Each fish was checked for spermiation every 2–4 days bygently pressing on its abdomen. Blood samples were taken weeklyto confirm steroid release and test for conversion to 15a-P usingRIA (Bryan et al., 2004). Plasma samples were also used to measurecirculating concentrations of the male lamprey pheromone usingELISA (Bryan et al., 2007). Measurements of the dorsal rope(Chung-Davidson et al., 2013a) were taken weekly using calipers,and differences between treatment groups were tested using atwo-way ANOVA, with time and treatment as the independentvariables, followed by Fisher’s LSD. Expressed fluids were checkedunder a microscope to determine presence of sperm. Upon reach-ing spermiation, animals were removed from the tanks, to reducethe potentially confounding effects of SM releasing pheromonesinto the tank (Brant et al., 2013; Chung-Davidson et al., 2013b,c).Differences between treatment groups in time to reach maturationwere tested with survival analysis using the Mantel–Cox test.
Table 1GenBank accession number and predicated length of PGRMCs proteins used forphylogenetic analysis.
Protein name Species GenBank No. Length
2.5. Binding experiments
Nuclear and membrane fractions of brain, gill, liver and testistissues (0.4 g each) were freshly extracted from PSM and SM usingNE-PER Nuclear and Cytoplasmic Extraction Reagents (Pierce Bio-technology/Thermo Scientific, Rockford, IL, USA) and BioVisionMembrane Protein Extraction Kit (BioVision Inc., Milpitas, CA,USA) according to the manufacturers’ instructions. The protein pel-let was resuspended in 3.2 ml Hepes buffer for binding assays.
To measure total binding in the target tissues, 100 ll proteinextract was incubated with 5 nM of 3H-P for 2 h at 4 �C. For bothnuclear and membrane binding assays, free steroids wereextracted by adding 1 ml of charcoal-dextran, centrifuged at2500�g for 15 min and the radioactivity of the supernatant weremeasured. To measure the binding affinity and capacity of nuclearand membrane fractions, testis was chosen and the method fol-lowed Huertas et al. (2014). Briefly, 150 ll of testis nuclear ormembrane protein extracts were incubated with 150 ll of increas-ing concentrations (0.1–10 nM) of 3H-P with (non-specific binding,BNS) or without (total binding, BT) 1.5 mM ‘cold’ P for 2 h at 4 �C.Specific binding (Bs) was determined by subtracting BNS from BT.Free steroids (F) were calculated by subtracting BT from totalamount of 3H-P. Charcoal separation method was used for bothnuclear and membrane fractions. The dissociation constant (Kd)and binding capacity (Bmax) were calculated by Scatchard plotand saturation curves (X-axis: Bs, Y-axis: F) using the Ligand Bind-ing Module of SigmaPlot software (Systat Software Inc., San Jose,CA, USA).
PGRMC2 Homo sapiens NP_006311 247PGRMC2 Mus musculus NP_081834 217PGRMC2 Gallus gallus NP_001006441 195PGRMC2 Xenopus tropicalis XP_002932101 194PGRMC2 Danio rerio NP_998269 201PGRMC1 Homo sapiens NP_006658 195PGRMC1 Mus musculus NP_058063 195PGRMC1 Gallus gallus NP_001258868 192PGRMC1 Xenopus tropicalis NP_001006842 177PGRMC1 Danio rerio NP_001007393 179PGRMC1 Oncorhynchus mykiss NP_001117916 181PGRMC1 Petromyzon marinus KJ648627 183
3. Results
3.1. Sea lamprey PGRMC1
The full-length sequence of sea lamprey PGRMC1 was identifiedfrom sea lamprey genome and transcriptome databases (Smithet al., 2013), and aligned to rainbow trout PGRMC1 (Fig. 1). A phy-logenetic analysis showed that the sea lamprey PGRMC1 was closeto the known vertebrate PGRMC1s (Fig. 2 and Table 1). Potential
protein motifs of sea lamprey PGRMC1 were predicted by the Scan-Site ‘‘MotifScan’’ module function (Obenauer et al., 2003). The 183amino acid sea lamprey sequence contains one SH2 targetsequence, one tyrosine kinase site, and one basophilic and two aci-dophilic serine/threonine kinase sites (Table 2), slightly differentfrom human PGRMC1, which contains a cytochrome b5 domain,two SH2 target sequences, one SH3 target sequence, one tyrosinekinase site, two acidophilic serine/threonine kinase sites, and bind-ing sites for ERK1 and PDK1 (Cahill, 2007).
3.2. Real-time quantitative PCR
In lamprey testis, there were differences between life stages inexpression levels of pgrmc1 (p < 0.0001; ANOVA; Fig. 3A), with PSMand SM having higher levels than LP (p = 0.0054 for PSM, p = 0.0243for SM; Fisher’s LSD). Following injections of lamprey GnRHs,expression levels of pgrmc1 did not change in PSM, but increasedin SM (lamprey GnRH-I, p = 0.0089; lamprey GnRH-III, p = 0.0260;Fig. 3B). Differences in nPR expression levels were also observedbetween life stages (p < 0.0001; ANOVA; Fig. 3C), with SM havinghigher expression levels than any other group (p = 0.0133 forPSM, p = 0.0019 for LP, p = 0.0093 for SP; Fisher’s LSD), and PSMhaving higher expression levels than LP or SP (p < 0.0001 for LPand SP, Fisher’s LSD). Levels of nPR did not change in PSM afterlamprey GnRH injection, and increased in SM in response to lam-prey GnRH-I only (p = 0.0264; Fig. 3D).
In lamprey brain, there were no significant differences inpgrmc1 mRNA expression levels between life stages (Fig. 4A). Injec-tions of lamprey GnRH-I and -III resulted in higher pgrmc1 expres-sion levels in PSM (p = 0.0296 and p = 0.0306, respectively), but didnot affect pgrmc1 expression levels in SM (Fig. 4B). There were dif-ferences in brain nPR expression levels between life stages(p = 0.0210), with SM having higher expression levels than PSMand SP (Fig. 4C). Injections of lamprey GnRH-I and -III resulted inhigher nPR expression levels in PSM (p = 0.0059 and p = 0.0059,respectively), but did not affect nPR expression levels in the brainof SM (Fig. 4D).
In lamprey gills, SM had higher expression levels of pgrmc1 thanany other life stage (p < 0.0001; Fig. 5A), and expression levelsincreased after lamprey GnRH injection in both PSM (lampreyGnRH-I, p = 0.0256; lamprey GnRH-III, p < 0.0001) and SM (lam-prey GnRH-I, p = 0.0020; lamprey GnRH-III, p = 0.0082; Fig. 5B).In PSM, lamprey GnRH-III increased expression levels more thanlamprey GnRH-I (p = 0.0043). Differences were also observedbetween life stages for nPR expression levels in the gills, with SMhaving much higher levels than any other life stage (p < 0.0001;Fig. 5C). Injection of neither type of lamprey GnRH had a significanteffect on nPR expression levels in PSM and SM, although there wasa trend toward an increase in SM (p < 0.1000; Fig. 5D).
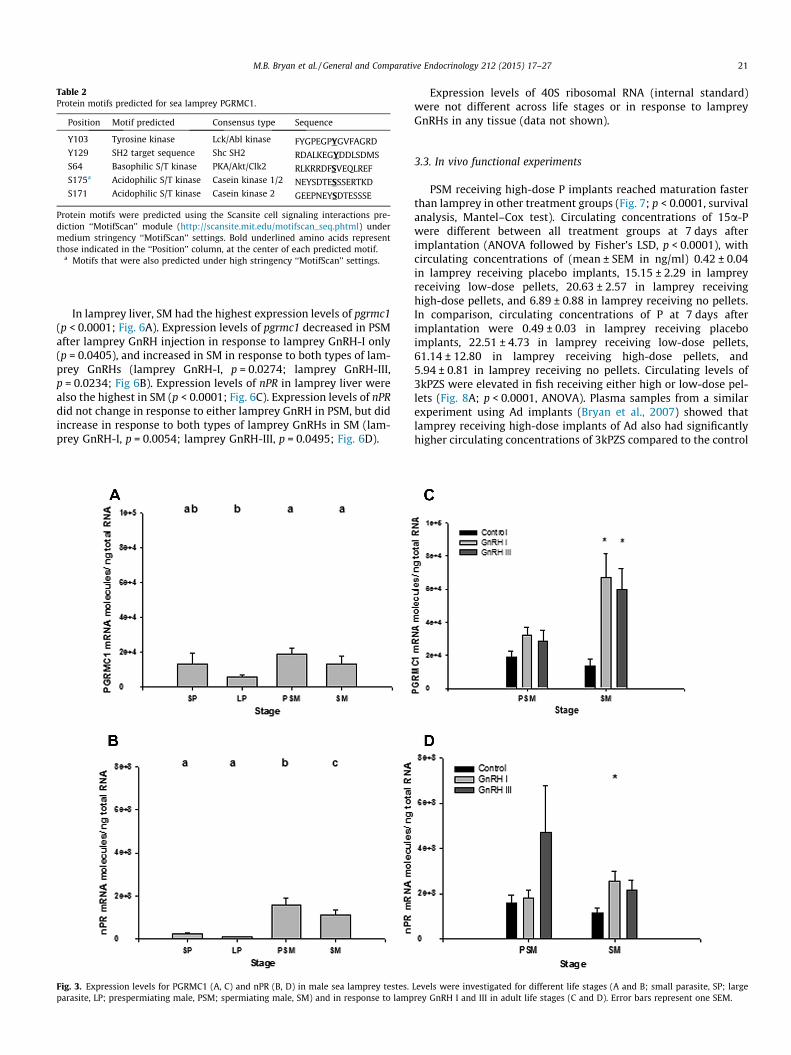
Table 2Protein motifs predicted for sea lamprey PGRMC1.
Position Motif predicted Consensus type Sequence
Y103 Tyrosine kinase Lck/Abl kinase FYGPEGPYGVFAGRDY129 SH2 target sequence Shc SH2 RDALKEGYDDLSDMSS64 Basophilic S/T kinase PKA/Akt/Clk2 RLKRRDFSVEQLREFS175a Acidophilic S/T kinase Casein kinase 1/2 NEYSDTESSSERTKDS171 Acidophilic S/T kinase Casein kinase 2 GEEPNEYSDTESSSE
Protein motifs were predicted using the Scansite cell signaling interactions pre-diction ‘‘MotifScan’’ module (http://scansite.mit.edu/motifscan_seq.phtml) undermedium stringency ‘‘MotifScan’’ settings. Bold underlined amino acids representthose indicated in the ‘‘Position’’ column, at the center of each predicted motif.
a Motifs that were also predicted under high stringency ‘‘MotifScan’’ settings.
M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27 21
In lamprey liver, SM had the highest expression levels of pgrmc1(p < 0.0001; Fig. 6A). Expression levels of pgrmc1 decreased in PSMafter lamprey GnRH injection in response to lamprey GnRH-I only(p = 0.0405), and increased in SM in response to both types of lam-prey GnRHs (lamprey GnRH-I, p = 0.0274; lamprey GnRH-III,p = 0.0234; Fig 6B). Expression levels of nPR in lamprey liver werealso the highest in SM (p < 0.0001; Fig. 6C). Expression levels of nPRdid not change in response to either lamprey GnRH in PSM, but didincrease in response to both types of lamprey GnRHs in SM (lam-prey GnRH-I, p = 0.0054; lamprey GnRH-III, p = 0.0495; Fig. 6D).
Fig. 3. Expression levels for PGRMC1 (A, C) and nPR (B, D) in male sea lamprey testes.parasite, LP; prespermiating male, PSM; spermiating male, SM) and in response to lamp
Expression levels of 40S ribosomal RNA (internal standard)were not different across life stages or in response to lampreyGnRHs in any tissue (data not shown).
3.3. In vivo functional experiments
PSM receiving high-dose P implants reached maturation fasterthan lamprey in other treatment groups (Fig. 7; p < 0.0001, survivalanalysis, Mantel–Cox test). Circulating concentrations of 15a-Pwere different between all treatment groups at 7 days afterimplantation (ANOVA followed by Fisher’s LSD, p < 0.0001), withcirculating concentrations of (mean ± SEM in ng/ml) 0.42 ± 0.04in lamprey receiving placebo implants, 15.15 ± 2.29 in lampreyreceiving low-dose pellets, 20.63 ± 2.57 in lamprey receivinghigh-dose pellets, and 6.89 ± 0.88 in lamprey receiving no pellets.In comparison, circulating concentrations of P at 7 days afterimplantation were 0.49 ± 0.03 in lamprey receiving placeboimplants, 22.51 ± 4.73 in lamprey receiving low-dose pellets,61.14 ± 12.80 in lamprey receiving high-dose pellets, and5.94 ± 0.81 in lamprey receiving no pellets. Circulating levels of3kPZS were elevated in fish receiving either high or low-dose pel-lets (Fig. 8A; p < 0.0001, ANOVA). Plasma samples from a similarexperiment using Ad implants (Bryan et al., 2007) showed thatlamprey receiving high-dose implants of Ad also had significantlyhigher circulating concentrations of 3kPZS compared to the control
Levels were investigated for different life stages (A and B; small parasite, SP; largerey GnRH I and III in adult life stages (C and D). Error bars represent one SEM.
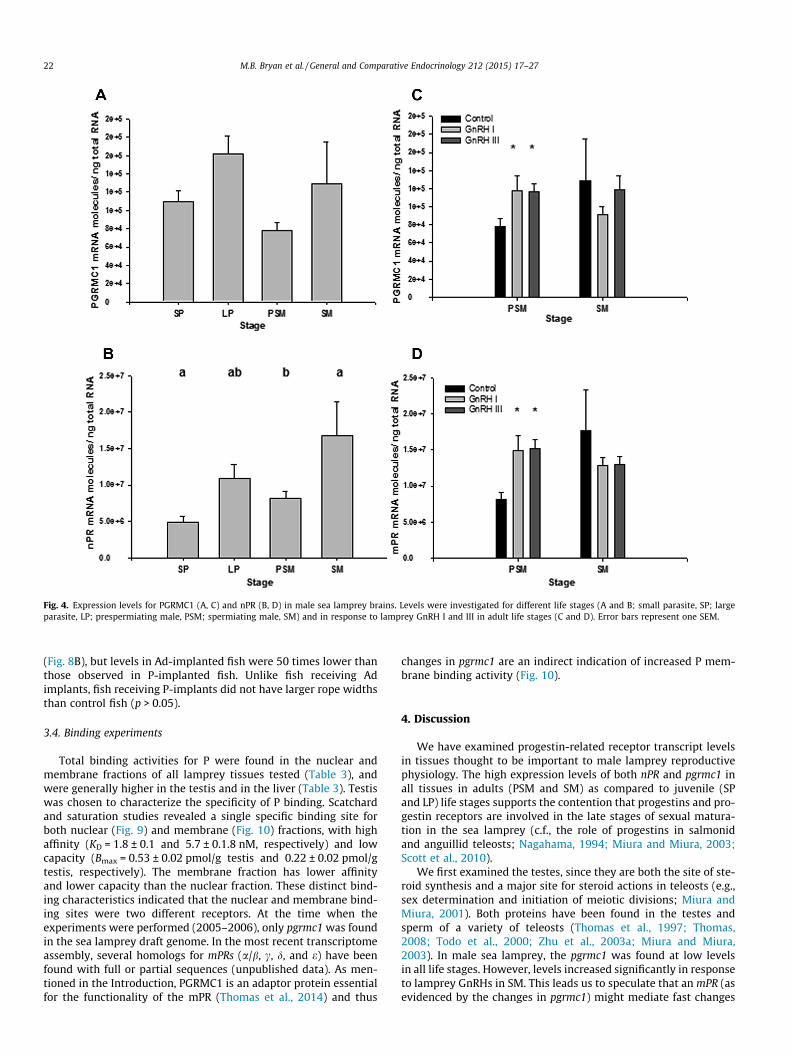
Fig. 4. Expression levels for PGRMC1 (A, C) and nPR (B, D) in male sea lamprey brains. Levels were investigated for different life stages (A and B; small parasite, SP; largeparasite, LP; prespermiating male, PSM; spermiating male, SM) and in response to lamprey GnRH I and III in adult life stages (C and D). Error bars represent one SEM.
22 M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27
(Fig. 8B), but levels in Ad-implanted fish were 50 times lower thanthose observed in P-implanted fish. Unlike fish receiving Adimplants, fish receiving P-implants did not have larger rope widthsthan control fish (p > 0.05).
3.4. Binding experiments
Total binding activities for P were found in the nuclear andmembrane fractions of all lamprey tissues tested (Table 3), andwere generally higher in the testis and in the liver (Table 3). Testiswas chosen to characterize the specificity of P binding. Scatchardand saturation studies revealed a single specific binding site forboth nuclear (Fig. 9) and membrane (Fig. 10) fractions, with highaffinity (KD = 1.8 ± 0.1 and 5.7 ± 0.1.8 nM, respectively) and lowcapacity (Bmax = 0.53 ± 0.02 pmol/g testis and 0.22 ± 0.02 pmol/gtestis, respectively). The membrane fraction has lower affinityand lower capacity than the nuclear fraction. These distinct bind-ing characteristics indicated that the nuclear and membrane bind-ing sites were two different receptors. At the time when theexperiments were performed (2005–2006), only pgrmc1 was foundin the sea lamprey draft genome. In the most recent transcriptomeassembly, several homologs for mPRs (a/b, c, d, and e) have beenfound with full or partial sequences (unpublished data). As men-tioned in the Introduction, PGRMC1 is an adaptor protein essentialfor the functionality of the mPR (Thomas et al., 2014) and thus
changes in pgrmc1 are an indirect indication of increased P mem-brane binding activity (Fig. 10).
4. Discussion
We have examined progestin-related receptor transcript levelsin tissues thought to be important to male lamprey reproductivephysiology. The high expression levels of both nPR and pgrmc1 inall tissues in adults (PSM and SM) as compared to juvenile (SPand LP) life stages supports the contention that progestins and pro-gestin receptors are involved in the late stages of sexual matura-tion in the sea lamprey (c.f., the role of progestins in salmonidand anguillid teleosts; Nagahama, 1994; Miura and Miura, 2003;Scott et al., 2010).
We first examined the testes, since they are both the site of ste-roid synthesis and a major site for steroid actions in teleosts (e.g.,sex determination and initiation of meiotic divisions; Miura andMiura, 2001). Both proteins have been found in the testes andsperm of a variety of teleosts (Thomas et al., 1997; Thomas,2008; Todo et al., 2000; Zhu et al., 2003a; Miura and Miura,2003). In male sea lamprey, the pgrmc1 was found at low levelsin all life stages. However, levels increased significantly in responseto lamprey GnRHs in SM. This leads us to speculate that an mPR (asevidenced by the changes in pgrmc1) might mediate fast changes

Fig. 5. Expression levels of mRNA for PGRMC1 (A, C) and nPR (B, D) in male sea lamprey gills. Levels were investigated for different life stages (A and B; small parasite, SP;large parasite, LP; prespermiating male, PSM; spermiating male, SM) and in response to lamprey GnRH I and III in adult life stages (C and D). Error bars represent one SEM.
M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27 23
needed in the gonads of SM for the release and activation of sperm(e.g., the increase in pH necessary for sperm activation; Miura andMiura, 2003;), particularly as pgrmc1 has been shown to be neces-sary for the acrosome reaction in mammals (Losel et al., 2005).
In the testis, nPR was much higher in PSM and SM than in thesexually immature parasitic phase lamprey, thus indicating animportant role in the adult phase. Surprisingly, levels of nPR onlyincreased in response to lamprey GnRH-I in SM. The fact that somenPR could be found in the testis of parasitic male lamprey mayindicate a role in initiation of the first meiotic division, as previ-ously suggested for the progestin 17,20b-dihydroxpregn-4-en-3-one (17,20b-DHP) in the eel (Anguilla japonica; Miura et al., 2006).
Studies were also carried out on the brain, a common target tis-sue for sex steroids that regulate reproductive behaviors (Brintonet al., 2008; Pasmanik and Callard, 1988; Zhu et al., 2003a). Sex ste-roids often have feedback effects on hypothalamic and pituitaryhormones, and the feedback effects are mediated by steroid recep-tors. It has been demonstrated in Atlantic croaker (Micropogonundulatus) that mPR in the brain mediates a negative feedback ofprogestins on GnRH release (Thomas et al., 2004). Both nPR andpgrmc1 were found in the lamprey brain, and their transcript levelsincreased in response to lamprey GnRHs in PSM only.
The highest levels of nPR were observed in the liver and gills inSM, thus lending support to our hypothesis that progestins areassociated with sea lamprey pheromone production and release.The highest levels of pgrmc1 were also found in the same organs
again providing evidence (albeit indirectly) that a membranereceptor for P is involved. The sea lamprey pheromone is synthe-sized in liver (Brant et al., 2013), modified and released by special-ized cells in male gills (Brant et al., 2013; Siefkes et al., 2003). Sincethis change occurs simultaneously with the onset of spermiation, itis likely that the same steroid responsible for spermiation is alsoresponsible for pheromone synthesis and release. In addition, Preception is involved in cholesterol regulation (Cahill, 2007), andbile acids such as sea lamprey pheromones are synthesized fromcholesterol (Elliott and Hyde, 1971; Javitt, 1994; Russell andSetchell, 1992; Salen and Shefer, 1983). Furthermore, the high levelof serum bile acids in patients with intrahepatic cholestasis ofpregnancy was associated with high levels of serum and urinarymono- and disulphated P metabolites (Abu-Hayyeh et al., 2013;Laatikainen and Karjalainen, 1973; Meng et al., 1997; Reyes andSjovaall, 2000; Sjovall and Sjovall, 1970). It is thus perhaps no coin-cidence that we found the highest levels of both pgrmc1 and nPR inthe liver of spermiating male sea lamprey, which are able to makemilligram quantities of the sex pheromone (Li et al., 2002).
Binding activities for P were found in the nuclear and mem-brane fractions in all lamprey tissues tested (Table 3), suggesteda widespread action of progestin signaling in lamprey by classicaland non-classical hormone actions. Nuclear binding in lampreyhas similar affinity to that of the zebrafish (Kd = 2 nM) and seatrout(Kd = 1.8, Bmax = 0.63 pmol/g ovary) for their cognate progestinligand 17,20b-DHP (Hanna et al., 2010; Pinter and Thomas,
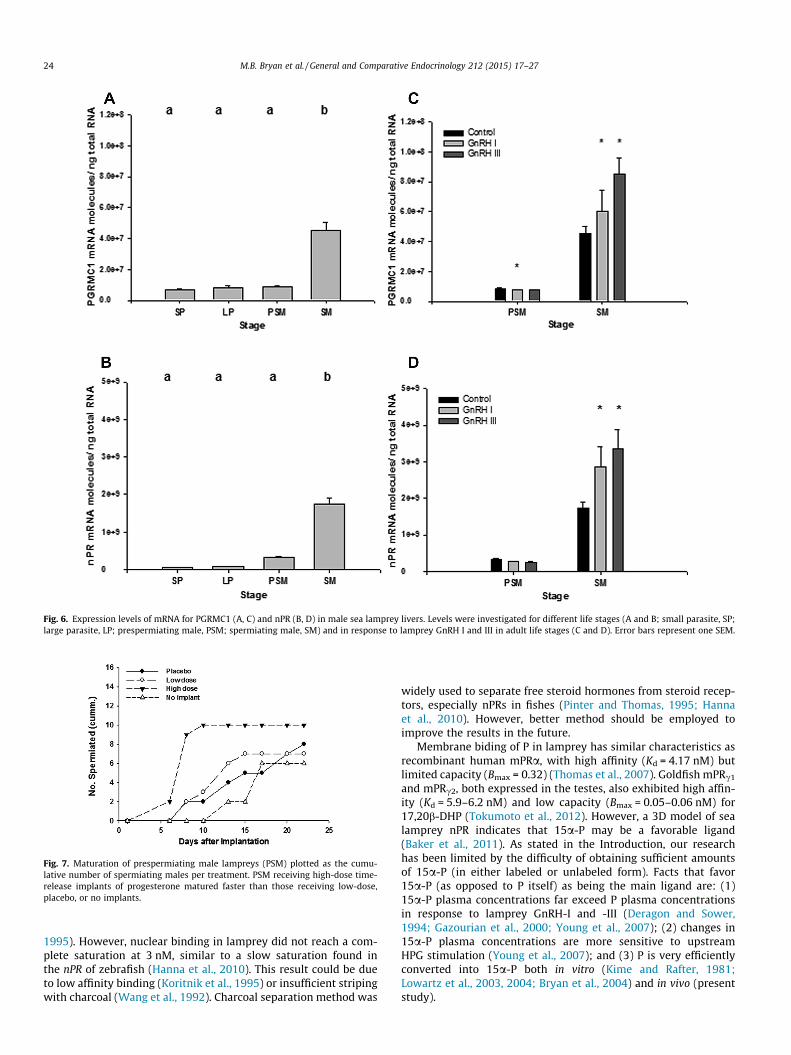
Fig. 6. Expression levels of mRNA for PGRMC1 (A, C) and nPR (B, D) in male sea lamprey livers. Levels were investigated for different life stages (A and B; small parasite, SP;large parasite, LP; prespermiating male, PSM; spermiating male, SM) and in response to lamprey GnRH I and III in adult life stages (C and D). Error bars represent one SEM.
Fig. 7. Maturation of prespermiating male lampreys (PSM) plotted as the cumu-lative number of spermiating males per treatment. PSM receiving high-dose time-release implants of progesterone matured faster than those receiving low-dose,placebo, or no implants.
24 M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27
1995). However, nuclear binding in lamprey did not reach a com-plete saturation at 3 nM, similar to a slow saturation found inthe nPR of zebrafish (Hanna et al., 2010). This result could be dueto low affinity binding (Koritnik et al., 1995) or insufficient stripingwith charcoal (Wang et al., 1992). Charcoal separation method was
widely used to separate free steroid hormones from steroid recep-tors, especially nPRs in fishes (Pinter and Thomas, 1995; Hannaet al., 2010). However, better method should be employed toimprove the results in the future.
Membrane biding of P in lamprey has similar characteristics asrecombinant human mPRa, with high affinity (Kd = 4.17 nM) butlimited capacity (Bmax = 0.32) (Thomas et al., 2007). Goldfish mPRc1
and mPRc2, both expressed in the testes, also exhibited high affin-ity (Kd = 5.9–6.2 nM) and low capacity (Bmax = 0.05–0.06 nM) for17,20b-DHP (Tokumoto et al., 2012). However, a 3D model of sealamprey nPR indicates that 15a-P may be a favorable ligand(Baker et al., 2011). As stated in the Introduction, our researchhas been limited by the difficulty of obtaining sufficient amountsof 15a-P (in either labeled or unlabeled form). Facts that favor15a-P (as opposed to P itself) as being the main ligand are: (1)15a-P plasma concentrations far exceed P plasma concentrationsin response to lamprey GnRH-I and -III (Deragon and Sower,1994; Gazourian et al., 2000; Young et al., 2007); (2) changes in15a-P plasma concentrations are more sensitive to upstreamHPG stimulation (Young et al., 2007); and (3) P is very efficientlyconverted into 15a-P both in vitro (Kime and Rafter, 1981;Lowartz et al., 2003, 2004; Bryan et al., 2004) and in vivo (presentstudy).
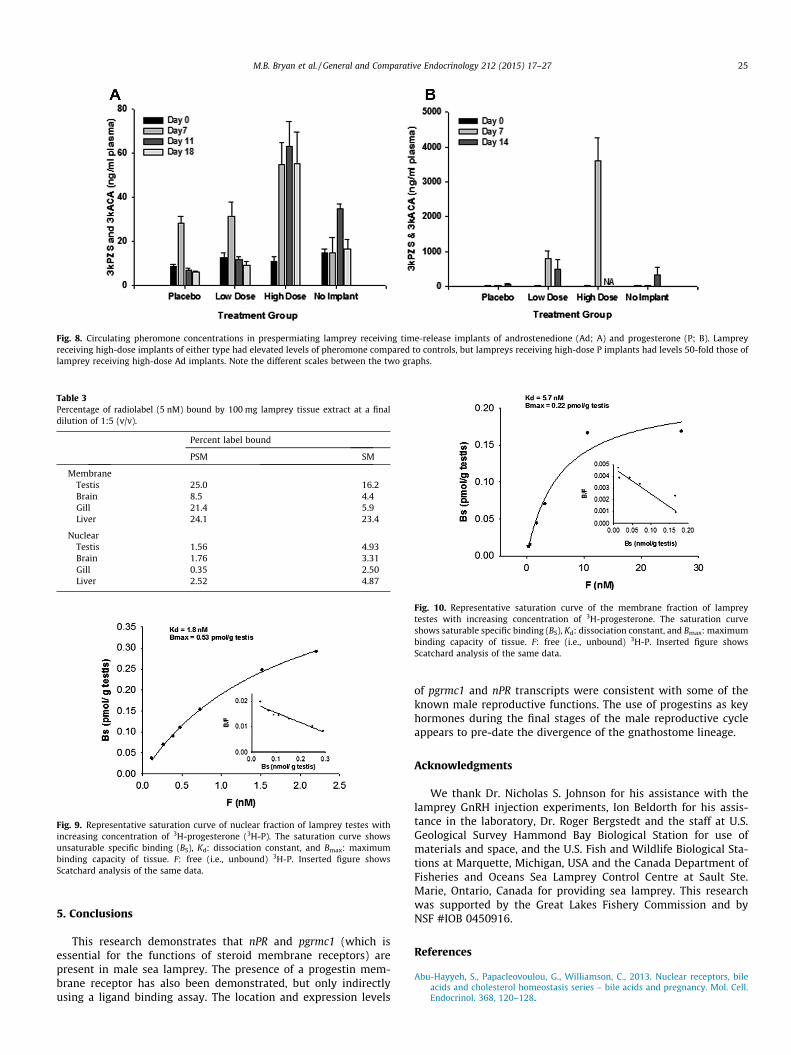
Fig. 8. Circulating pheromone concentrations in prespermiating lamprey receiving time-release implants of androstenedione (Ad; A) and progesterone (P; B). Lampreyreceiving high-dose implants of either type had elevated levels of pheromone compared to controls, but lampreys receiving high-dose P implants had levels 50-fold those oflamprey receiving high-dose Ad implants. Note the different scales between the two graphs.
Table 3Percentage of radiolabel (5 nM) bound by 100 mg lamprey tissue extract at a finaldilution of 1:5 (v/v).
Percent label bound
PSM SM
MembraneTestis 25.0 16.2Brain 8.5 4.4Gill 21.4 5.9Liver 24.1 23.4
NuclearTestis 1.56 4.93Brain 1.76 3.31Gill 0.35 2.50Liver 2.52 4.87
Fig. 9. Representative saturation curve of nuclear fraction of lamprey testes withincreasing concentration of 3H-progesterone (3H-P). The saturation curve showsunsaturable specific binding (BS), Kd: dissociation constant, and Bmax: maximumbinding capacity of tissue. F: free (i.e., unbound) 3H-P. Inserted figure showsScatchard analysis of the same data.
Fig. 10. Representative saturation curve of the membrane fraction of lampreytestes with increasing concentration of 3H-progesterone. The saturation curveshows saturable specific binding (BS), Kd: dissociation constant, and Bmax: maximumbinding capacity of tissue. F: free (i.e., unbound) 3H-P. Inserted figure showsScatchard analysis of the same data.
M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27 25
5. Conclusions
This research demonstrates that nPR and pgrmc1 (which isessential for the functions of steroid membrane receptors) arepresent in male sea lamprey. The presence of a progestin mem-brane receptor has also been demonstrated, but only indirectlyusing a ligand binding assay. The location and expression levels
of pgrmc1 and nPR transcripts were consistent with some of theknown male reproductive functions. The use of progestins as keyhormones during the final stages of the male reproductive cycleappears to pre-date the divergence of the gnathostome lineage.
Acknowledgments
We thank Dr. Nicholas S. Johnson for his assistance with thelamprey GnRH injection experiments, Ion Beldorth for his assis-tance in the laboratory, Dr. Roger Bergstedt and the staff at U.S.Geological Survey Hammond Bay Biological Station for use ofmaterials and space, and the U.S. Fish and Wildlife Biological Sta-tions at Marquette, Michigan, USA and the Canada Department ofFisheries and Oceans Sea Lamprey Control Centre at Sault Ste.Marie, Ontario, Canada for providing sea lamprey. This researchwas supported by the Great Lakes Fishery Commission and byNSF #IOB 0450916.
References
Abu-Hayyeh, S., Papacleovoulou, G., Williamson, C., 2013. Nuclear receptors, bileacids and cholesterol homeostasis series – bile acids and pregnancy. Mol. Cell.Endocrinol. 368, 120–128.

26 M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27
Baker, M.E., Asnaashari, P., Chang, D.J., McDonnell, S., 2011. 3D models of lampreyprogesterone receptor complexed with progesterone, 7a-hydroxy-progesteroneand 15a-hydroxy-progesterone. Steroids 76, 169–176.
Brant, C.O., Chung-Davidson, Y.-W., Li, K., Scott, A.M., Li, W., 2013. Biosynthesis andrelease of pheromonal bile salts in mature male sea lamprey. BMC Biochem. 14,30.
Brinton, R.D., Thompson, R.F., Foy, M.R., Baudry, M., Wang, J., Finch, C.E., Morgan,T.E., Pike, C.J., Mack, W.J., Stanczyk, F.Z., Nilsen, J., 2008. Progesterone receptors:form and function in brain. Front. Neuroendocrinol. 29, 313–339.
Bryan, M.B., Scott, A.P., Li, W., 2007. The sea lamprey (Petromyzon marinus) has areceptor for androstenedione. Biol. Reprod. 77, 688–696.
Bryan, M.B., Scott, A.P., Li, W., 2008. Sex steroids and their receptors in lampreys.Steroids 73, 1–12.
Bryan, M.B., Scott, A.P., Young, B.A., Cerny, I., Li, W., 2004. 15a-Hydroxyprogesteronein male sea lampreys, Petromyzon marinus L. Steroids 69, 473–481.
Cahill, M.A., 2007. Progesterone receptor membrane component 1: an integrativereview. J. Steroid Biochem. Mol. Biol. 105, 16–36.
Chung-Davidson, Y.-W., Priess, M.C., Yeh, C.-Y., Brant, C.O., Johnson, N.S., Li, K.,Nanlohy, K.G., Bryan, M.B., Brown, C.T., Choi, J., Li, W., 2013a. A thermogenicsecondary sexual character in male sea lamprey. J. Exp. Biol. 216,2702–2712.
Chung-Davidson, Y.-W., Wang, H., Bryan, M.B., Wu, H., Johnson, N.S., Li, W., 2013b.An anti-steroidogenic inhibitory primer pheromone in male sea lamprey(Petromyzon marinus). Gen. Comp. Endocrinol. 189, 24–31.
Chung-Davidson, Y.-W., Wang, H., Siefkes, M.J., Bryan, M.B., Wu, H., Johnson, N.S., Li,W., 2013c. Pheromonal bile acid 3-ketopetromyzonol sulfate primes theneuroendocrine system in sea lamprey. BMC Neurosci. 14, 11.
Deragon, K.L., Sower, S.A., 1994. Effects of lamprey gonadotropin-releasinghormone-III on steroidogenesis and spermiation in male sea lampreys. Gen.Comp. Endocrinol. 95, 363–367.
Elliott, W.H., Hyde, P.M., 1971. Metabolic pathways of bile acid synthesis. Am. J.Med. 51, 568–579.
Falkenstein, E., Meyer, C., Eisen, C., Scriba, P.C., Wehling, M., 1996. Full-length cDNAsequence of a progesterone membrane-binding protein from porcine vascularsmooth muscle cells. Biochem. Biophys. Res. Commun. 229, 86–89.
Gazourian, L., Evans, E.L., Hanson, L., Chase, C.F., Sower, S.A., 2000. The effects oflamprey GnRH-I, -III and analogs on steroidogenesis in the sea lamprey(Petromyzon marinus). Aquaculture 188, 147–165.
Hanna, R.N., Daly, S.C.J., Pang, Y., Anglade, I., Kah, O., Thomas, P., Zhu, Y., 2010.Characterization and expression of the nuclear progestin receptor in zebrafishgonads and brain. Biol. Reprod. 82, 112–122.
Huertas, M., Almeida, O.G., Canario, A.V.M., Hubbard, P.C., 2014. Tilapia maleurinary pheromone stimulates female reproductive axis. Gen. Comp.Endocrinol. 196, 106–111.
Javitt, N.B., 1994. Bile acid synthesis from cholesterol: regulatory and auxiliarypathways. FASEB J. 8, 1308–1311.
Jones, D.T., Taylor, W.R., Thornton, J.M., 1992. The rapid generation of mutation datamatrices from protein sequences. Comput. Appl. Biosci. 8, 275–282.
Karteris, E., Zervou, S., Pang, Y., Dong, J., Hillhouse, E.W., Randeva, H.S., Thomas, P.,2006. Progesterone signaling in human myometrium through two novelmembrane G protein-coupled receptors: potential role in functionalprogesterone withdrawal at term. Mol. Endocrinol. 29, 1519–1534.
Kime, D.E., Callard, C.V., 1982. Formation of 15a-hydroxylated androgens by thetestis and other tissues of the sea lamprey, Petromyzon marinus, in vitro. GeneralGen. Comp. Endocrinol. 4, 267–270.
Kime, D.E., Rafter, J.J., 1981. Biosynthesis of 15-hydroxylated steroids by gonads ofthe river lamprey, Lampetra fluviatilis, in vitro. Gen. Comp. Endocrinol. 44, 69–76.
Koritnik, D.R., Marschke, K.B., Koshy, A., 1995. Characterization of a hepatic proteinin nonhuman primates that binds mibolerone but not dihydrotestosterone ormethyltrienolone. Steroids 60, 759–767.
Laatikainen, T., Karjalainen, O., 1973. Excretion of progesterone metabolites in urineand bile of pregnant women with intrahepatic cholestasis. J. Steroid Biochem. 4,641–648.
Li, W., Scott, A.P., Siefkes, M.J., Yan, H.G., Liu, Q., Yun, S.S., Gage, D.A., 2002. Bile acidsecreted by male sea lamprey that acts as a sex pheromone. Science 296, 138–141.
Libants, S.V., Carr, K., Wu, H., Teeter, J.H., Chung-Davidson, Y.-W., Zhang, Z.,Wilkerson, C., Li, W., 2009. The sea lamprey Petromyzon marinus genome revealsthe early origin of several chemosensory receptor families in the vertebratelineage. BMC Evol. Biol. 9, 180.
Losel, R., Breiter, S., Seyfert, M., Wehling, M., Falkenstein, E., 2005. Classic and non-classic progesterone receptors are both expressed in human spermatozoa.Horm. Metab. Res. 37, 10–14.
Lowartz, S.M., Petkam, R., Renaud, R., Beamish, F.W.H., Kime, D.E., Raeside, J.,Leatherland, J.F., 2003. Blood steroid profile and in vitro steroidogenesis byovarian follicles and testis fragments of adult sea lamprey,Petromyzon marinus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 134,365–376.
Lowartz, S.M., Renaud, R.L., Beamish, F.W.H., Leatherland, J.F., 2004. Evidence for15a- and 7a-hydroxylase activity in gonadal tissue of the early-life stages of sealampreys, Petromyzon marinus. Comp. Biochem. Physiol. B Biochem. Mol. Biol.138, 119–127.
Meng, L.J., Reyes, H., Palma, J., Hernandez, I., Ribalta, J., Sjovall, J., 1997. Profiles ofbile acids and progesterone metabolites in the urine and serum of women withintrahepatic cholestasis of pregnancy. J. Hepatol. 27, 346–357.
Meyer, C., Schmid, R., Scriba, P.C., Wehling, M., 1996. Purification and partialsequence of high affinity progesterone-binding site(s) from porcine livermembranes. Eur. J. Biochem. 239, 726–731.
Miura, T., Kasugai, T., Nagahama, Y., Yamauchi, K., 1995. Acquisition of potential forsperm motility in vitro in Japanese eel Anguilla japonica. Fisher. Sci. 61, 533–534.
Miura, T., Higuchi, M., Ozaki, Y., Ohta, T., Miura, C., 2006. Progestogen is an essentialfactor for the initiation of the meiosis in spermatogenetic cells of the eel. PNAS103, 7333–7338.
Miura, T., Miura, C., 2001. Japanese eel: a model for analysis of spermatogenesis.Zool. Sci. 18, 1055–1063.
Miura, T., Miura, C., 2003. Molecular control mechanisms of fish spermatogenesis.Fish Physiol. Biochem. 28, 181–186.
Mourot, B., Nguyen, T., Fostier, P., Bobe, J., 2006. Two unrelated putative membrane-bound progestin receptors, progesterone membrane receptor component 1(PGRMC1) and membrane progestin receptor (mPR) beta, are expressed in therainbow trout oocyte and exhibit similar ovarian expression patterns. Reprod.Biol. Endocrinol. 4, 6.
Mylonas, C.C., Scott, A.P., Vermeirssen, E.L., Zohar, Y., 1997. Changes in plasmagonadotropin II and sex steroid hormones, and sperm production of striped bassafter treatment with controlled-release gonadotropin-releasing hormoneagonist-delivery systems. Biol. Reprod. 57, 669–675.
Nagahama, Y., 1994. Endocrine regulation of gametogenesis in fish. Int. J. Dev. Biol.38, 217–229.
Obenauer, J.C., Cantley, L.C., Yaffe, M.B., 2003. Scansite 2.0: proteome-wideprediction of cell signaling interactions using short sequence motifs. NucleicAcids Res. 31, 3635–3641.
Pang, Y., Dong, J., Thomas, P., 2013. Characterization, neurosteroid binding and braindistribution of human membrane progesterone receptors d and e (mPRd andmPRe) and mPRd involvement in neurosteroid inhibition of apoptosis.Endocrinology 154, 283–295.
Pang, Y., Thomas, P., 2011. Progesterone signals through membrane progesteronereceptors (mPRs) in MDA-MB-468 and mPR-transfected MDA-MB-231 breastcancer cells which lack full-length and N-terminally truncated isoforms of thenuclear progesterone receptor. Steroids 76, 921–928.
Pasmanik, M., Callard, G., 1988. A high abundance androgen receptor in goldfishbrain: characteristics and seasonal changes. Endocrinology 123, 1162–1171.
Pinter, J., Thomas, P., 1995. Characterization of a progestogen receptor in the ovaryof the spotted sea-trout, Cynoscion nebulosus. Biol. Reprod. 52, 667–675.
Reyes, H., Sjovaall, J., 2000. Bile acids and progesterone metabolites in intrahepaticcholestasis of pregnancy. Ann. Med. 32, 94–106.
Russell, D.W., Setchell, K.D.R., 1992. Bile acid biosynthesis. Biochemistry 31, 4737–4749.
Salen, G., Shefer, S., 1983. Bile acid synthesis. Ann. Rev. Physiol. 45, 679–685.Scott, A.P., Sumpter, J.P., Stacey, N.E., 2010. The role of the maturation-inducing
steroid, 17,20b-dihydroxypregn-4-en-3-one, in male fish: a review. J. Fish Biol.76, 183–224.
Sherwood, N.M., Sower, S.A., Marshak, D.R., Fraser, B.A., Brownstein, M.J., 1986.Primary structure of gonadotropin-releasing hormone from lamprey brain. J.Biol. Chem. 261, 4812–4819.
Siefkes, M.J., Scott, A.P., Zielinski, B., Yun, S.S., Li, W., 2003. Male sea lampreys,Petromyzon marinus L., excrete a sex pheromone from gill epithelia. Biol.Reprod. 69, 125–132.
Sjovall, J., Sjovall, K., 1970. Steroid sulphates in plasma from pregnant women withpruritus and elevated plasma bile acid levels. Ann. Clin. Res. 2, 321–337.
Smith, J.J., Kuraku, S., Holt, C., Sauka-Spengler, T., Jiang, N., Campbell, M.S., Yandell,M.D., Manousaki, T., Meyer, A., Bloom, O.E., Morgan, J.R., Buxbaum, J.D.,Sachidanandam, R., Sims, C., Garrett, A.S., Cook, M., Krumlauf, R., Wiedemann,L.M., Sower, S.A., Decatur, W.A., Hall, J.A., Amemiya, C.T., Saha, N.R., Buckley,K.M., Rast, J.P., Das, S., Hirano, M., McCurley, N., Guo, P., Rohner, N., Tabin, C.J.,Piccinelli, P., Elgar, G., Ruffier, M., Aken, B.L., Searle, S.M.J., Muffato, M.,Pignatelli, M., Herrero, J., Jones, M., Brown, C.T., Chung-Davidson, Y.-W.,Nanlohy, K.G., Libants, S.V., Yeh, C.-Y., McCauley, D.W., Langeland, J.A., Pancer,Z., Fritzsch, B., de Jong, P.J., Zhu, B., Fulton, L.L., Theising, B., Flicek, P., Bronner,M., Warren, W.C., Clifton, S.W., Wilson, R.K., Li, W., 2013. The lamprey genome:illuminating vertebrate origins. Nat. Gen. 45, 415–421.
Sower, S.A., 1989. Effects of lamprey gonadotropin-releasing hormone and analogson steroidogenesis and spermiation in male sea lampreys. Fish Physiol.Biochem. 7, 101–107.
Sower, S.A., Chiang, Y.-C., Lovas, S., Conlon, J.M., 1993. Primary structure andbiological activity of a third gonadotropin-releasing hormone from lampreybrain. Endocrinology 261, 4812–4819.
Sower, S.A., Moriyama, S., Kasahara, M., Takahashi, A., Nozaki, M., Uchida, K.,Dahlstrom, J.M., Kawauchi, H., 2006. Identification of sea lamprey GTHbeta-likecDNA and its evolutionary implications. Gen. Comp. Endocrinol. 148, 22–32.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: molecular evolutionarygenetics analysis (MEGA) software version 4.0. Mo. Biol. Evol. 24, 1596–1599.
Tang, Y.T., Hu, T., Arterburn, M., Boyle, B., Bright, J.M., Emtage, P.C., Funk, W.D., 2005.PAQR proteins: a novel membrane receptor family defined by an ancient 7-transmembrane pass motif. J. Mol. Evol. 61, 372–380.
Thomas, P., 2000. Nuclear and membrane steroid receptors and their functions inteleost gonads. In: Norber, B., Kjesbu, O.S., Taranger, G.L., Andersson, E.,Stefasson, S.O. (Eds.), Proceedings of the Sixth International Symposium onthe Reproductive Physiology of Fish. Institute of Marine Research andUniversity of Bergen, Bergen, Norway, pp. 149–156.
Thomas, P., 2003. Rapid, nongenomic steroid actions initiated at the cell surface:lessons from studies with fish. Fish Physiol. Biochem. 28, 3–12.

M.B. Bryan et al. / General and Comparative Endocrinology 212 (2015) 17–27 27
Thomas, P., 2008. Characteristics of membrane progestin receptor alpha (mPRalpha)and progesterone membrane receptor component 1 (PGRMC1) and their rolesin mediating rapid progestin actions. Front. Neuroendocrinol. 29, 292–312.
Thomas, P., Breckenridge-Miller, D., Detweiler, C., 1997. Binding characteristics andregulation of the 17,20b,21-trihydroxy-4-pregnen-3-one (20b-S) receptor ontesticular and sperm plasma membranes of spotted seatrout (Cynoscionnebulosus). Fish Physiol. Biochem. 17, 109–116.
Thomas, P., Pang, Y., Dong, J., 2014. Enhancement of cell surface expression andreceptor functions of membrane progestin receptor a (mPRa) by progesteronereceptor membrane component 1 (PGRMC1): evidence for a role of PGRMC1 asan adaptor protein for steroid receptors. Endocrinology 155, 1107–1119.
Thomas, P., Pang, Y., Dong, J., Groenen, P., Kelder, J., de Vlieg, J., Zhu, Y., Tubbs, C.,2007. Steroid and G protein binding characteristics of the seatrout and humanprogestin membrane receptor a subtypes and their evolutionary origins.Endocrinology 148, 705–718.
Thomas, P., Pang, Y., Zhu, Y., Detweiler, C., Doughty, K., 2004. Multiple rapidprogestogen actions and progestogen membrane receptor subtypes in fish.Steroids 69, 567–573.
Thomas, P., Pinter, J., Das, S., 2001. Upregulation of the maturation-inducing steroidmembrane receptor in spotted seatrout ovaries by gonadotropin during oocytematuration and its physiological significance. Biol. Reprod. 64, 21–29.
Thomas, P., Tubbs, C., Detweiler, C., Das, S., Ford, L., Breckenridge-Miller, D., 2005.Binding characteristics, hormonal regulation and identity of the spermmembrane progestogen receptor in Atlantic croaker. Steroids 70, 427–433.
Thornton, J.W., 2001. Evolution of vertebrate steroid receptors from an ancestralestrogen receptor by ligand exploitation and serial genome expansions. PNAS98, 5671–5676.
Todo, T., Ikeuchi, T., Kobayashi, T., Kajiura-Kobayashi, H., Suzuki, K., Yoshikuni, M.,Yamauchi, K., Nagahama, Y., 2000. Characterization of a testicular 17 alpha,20beta-dihydroxy-4-pregnen-3-one (a spermiation-inducing steroid in fish)receptor from a teleost, Japanese eel (Anguilla japonica). FEBS Lett. 465, 12–17.
Tokumoto, T., Tokumoto, M., Oshima, T., Shimizuguchi, K., Fukuda, T., Sugita, E.,Suzuki, M., Sakae, Y., Akiyama, Y., Nakayama, R., Roy, S.R., Rahman, M.S., Pang,
Y.F., Dong, J., Thomas, P., 2012. Characterization of multiple membraneprogestin receptor (mPR) subtypes from the goldfish ovary and their roles inthe induction of oocyte maturation. Gen. Comp. Endocrinol. 177, 168–176.
Ueda, H., Kambegawa, A., Nagahama, Y., 1985. Involvement of gonadotrophin andsteroid hormones in spermiation in the amago salmon, Oncorhynchus rhodurus,and goldfish, Carassius auratus. Gen. Comp. Endocrinol. 59, 24–30.
Vermeirssen, E.L., Scott, A.P., Mylonas, C.C., Zohar, Y., 1998. Gonadotrophin-releasing hormone agonist stimulates milt fluidity and plasma concentrationsof 17,20beta-dihydroxylated and 5beta-reduced, 3alpha-hydroxylated C21steroids in male plaice (Pleuronectes platessa). Gen. Comp. Endocrinol. 112,163–177.
Wang, J.-X., Yamamura, H.I., Wang, W., Roeske, W.R., 1992. The use of the filtrationtechnique in in vitro radioligand binding assays for membrane-bound andsolubilized receptors. In: Hulme, E.C. (Ed.), Receptor–Ligand Interactions, aPractical Approach. IRL Press, Oxford, pp. 213–234.
Young, B.A., Bryan, M.B., Glenn, J.R., Yun, S.S., Scott, A.P., Li, W., 2007. Dose-responserelationship of 15a-hydroxylated sex steroids to gonadotropin-releasinghormones and pituitary extract in male sea lampreys (Petromyzon marinus).Gen. Comp. Endocrinol. 151, 108–115.
Young, B.A., Bryan, M.B., Sower, S.A., Scott, A.P., Li, W., 2004. 15a-Hydroxytestosterone induction by GnRH-I and GnRH-III in Atlantic and GreatLakes sea lamprey (Petromyzon marinus L.). Gen. Comp. Endocrinol. 136, 276–281.
Yun, S.-S., Scott, A.P., Li, W., 2003. Pheromones of the male sea lamprey, Petromyzonmarinus L.: structural studies on a new compound, 3-keto allocholic acid, and 3-keto petromyzonol sulfate. Steroids 68, 297–304.
Zhu, Y., Bond, J., Thomas, P., 2003a. Identification, classification, and partialcharacterization of genes in humans and other vertebrates homologous to afish membrane progestogen receptor. PNAS 100, 2237–2242.
Zhu, Y., Rice, C.D., Pang, Y., Pace, M., Thomas, P., 2003b. Cloning, expression, andcharacterization of a membrane progestogen receptor and evidence it is anintermediary in meiotic maturation of fish oocytes. PNAS 100, 2231–2236.
Related Documents











