ECOPHYSIOLOGY Evidence of threshold temperatures for xylogenesis in conifers at high altitudes Sergio Rossi Annie Deslauriers Tommaso Anfodillo Vinicio Carraro Received: 23 June 2006 / Accepted: 17 November 2006 / Published online: 13 December 2006 ȑ Springer-Verlag 2006 Abstract Temperature is the most important factor affecting growth at high altitudes. As trees use much of the allocated carbon gained from photosynthesis to produce branches and stems, information on the timing and dynamics of secondary wood growth is crucial to assessing temperature thresholds for xylogenesis. We have carried out histological analyses to determine cambial activity and xylem cell differentiation in conifers growing at the treeline on the eastern Alps in two sites during 2002–2004 with the aim of linking the growth process with temperature and, consequently, of defining thresholds for xylogenesis. Cambial activity occurred from May to July–August and cell differen- tiation from May–June to September–October. The earliest start of radial enlargement was observed in stone pine in mid-May, while Norway spruce was the last species to begin tracheid differentiation. The duration of wood formation varied from 90 to 137 days, depending on year and site, with no differ- ence between species. Longer durations were observed in trees on the south-facing site because of the earlier onset and later ending of cell production and differ- entiation. The threshold temperatures at which xylo- genesis had a 0.5 probability of being active were calculated by logistic regressions. Xylogenesis was active when the mean daily air temperature was 5.6–8.5ŶC and mean stem temperature was 7.2–9ŶC. The similar thresholds among all trees suggested the existence of thermal limits in wood formation that correspond with temperatures of 6–8ŶC that are sup- posed to limit growth at the treeline. Different soil temperature thresholds between sites indicated that soil temperature may not be the main factor limiting xylogenesis. This study represents the first attempt to define a threshold through comparative assessment of xylem growth and tissue temperatures in stem meristems at high altitudes. Keywords Alps Cambial activity Cell differentiation Treeline Tree ring Introduction High-altitude forests have been studied extensively in order to gain an understanding of why trees cannot grow above a certain altitude (Ko ¨ rner 2003). Given that these ecotones are strongly temperature-limited, they have recently assumed additional relevance as potential indicators of climate change (Beniston et al. 1997; Theurillat and Guisan 2001; Pisaric et al. 2003). The physiological determinant of treeline position at a global scale is still uncertain despite several hypotheses having been put forward (Tranquillini 1979; Sveinbjo ¨ rnsson 2000; Smith et al. 2003). The hypothesis best supported by experimental data asserts that low temperatures limit the production of new cells by meristems irrespective of photoassimilate abundance (growth limitation hypothesis; Ko ¨ rner 1998). Deslau- riers and Morin (2005) found that tracheid production rate varies with the seasonal dynamics of minimum temperature. Cambial activity becomes particularly Communicated by Hermann Heilmeier. S. Rossi (&) A. Deslauriers T. Anfodillo V. Carraro Treeline Ecology Research Unit, Dipartimento TeSAF, Universita ` degli Studi di Padova, viale dell’Universita ` 16, 35020 Legnaro, PD, Italy e-mail: [email protected] 123 Oecologia (2007) 152:1–12 DOI 10.1007/s00442-006-0625-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ECOPHYSIOLOGY
Evidence of threshold temperatures for xylogenesis in conifersat high altitudes
Sergio Rossi Æ Annie Deslauriers Æ Tommaso Anfodillo ÆVinicio Carraro
Received: 23 June 2006 / Accepted: 17 November 2006 / Published online: 13 December 2006� Springer-Verlag 2006
Abstract Temperature is the most important factor
affecting growth at high altitudes. As trees use much of
the allocated carbon gained from photosynthesis to
produce branches and stems, information on the timing
and dynamics of secondary wood growth is crucial to
assessing temperature thresholds for xylogenesis. We
have carried out histological analyses to determine
cambial activity and xylem cell differentiation in
conifers growing at the treeline on the eastern Alps in
two sites during 2002–2004 with the aim of linking the
growth process with temperature and, consequently, of
defining thresholds for xylogenesis. Cambial activity
occurred from May to July–August and cell differen-
tiation from May–June to September–October. The
earliest start of radial enlargement was observed in
stone pine in mid-May, while Norway spruce was the
last species to begin tracheid differentiation. The
duration of wood formation varied from 90 to
137 days, depending on year and site, with no differ-
ence between species. Longer durations were observed
in trees on the south-facing site because of the earlier
onset and later ending of cell production and differ-
entiation. The threshold temperatures at which xylo-
genesis had a 0.5 probability of being active were
calculated by logistic regressions. Xylogenesis was
active when the mean daily air temperature was
5.6–8.5�C and mean stem temperature was 7.2–9�C.
The similar thresholds among all trees suggested the
existence of thermal limits in wood formation that
correspond with temperatures of 6–8�C that are sup-
posed to limit growth at the treeline. Different soil
temperature thresholds between sites indicated that
soil temperature may not be the main factor limiting
xylogenesis. This study represents the first attempt
to define a threshold through comparative assessment
of xylem growth and tissue temperatures in stem
meristems at high altitudes.
Keywords Alps � Cambial activity �Cell differentiation � Treeline � Tree ring
Introduction
High-altitude forests have been studied extensively in
order to gain an understanding of why trees cannot
grow above a certain altitude (Korner 2003). Given
that these ecotones are strongly temperature-limited,
they have recently assumed additional relevance as
potential indicators of climate change (Beniston et al.
1997; Theurillat and Guisan 2001; Pisaric et al. 2003).
The physiological determinant of treeline position at a
global scale is still uncertain despite several hypotheses
having been put forward (Tranquillini 1979;
Sveinbjornsson 2000; Smith et al. 2003). The hypothesis
best supported by experimental data asserts that low
temperatures limit the production of new cells by
meristems irrespective of photoassimilate abundance
(growth limitation hypothesis; Korner 1998). Deslau-
riers and Morin (2005) found that tracheid production
rate varies with the seasonal dynamics of minimum
temperature. Cambial activity becomes particularly
Communicated by Hermann Heilmeier.
S. Rossi (&) � A. Deslauriers � T. Anfodillo �V. CarraroTreeline Ecology Research Unit, Dipartimento TeSAF,Universita degli Studi di Padova, viale dell’Universita 16,35020 Legnaro, PD, Italye-mail: [email protected]
123
Oecologia (2007) 152:1–12
DOI 10.1007/s00442-006-0625-7

limited after the young plants have emerged from the
herbaceous-shrub layer where the generally warmer
temperatures are more favourable to growth (Grace
and Norton 1990; Hattenschwiler and Korner 1995).
This leads to a progressive decrease in longitudinal
annual growth when a plant approaches the ‘‘tree
habitus’’, i.e., more that 3 m in height. Because trees
growing at the treeline appear to be limited by pro-
cesses involving tissue formation, a basic knowledge of
intra-annual stem growth is fundamental to determin-
ing a reliable temperature threshold directly based on
cambial activity processes.
Within a global perspective, treeline position seems
to coincide with a mean temperature in the growing
season ranging between 6 and 7�C (Korner 2003),
suggesting that growth processes might be strongly
limited below this threshold. In situ measurements at
the northern treeline revealed that growth occurred
when the average daily temperature was above 5�C
(Holtmeier 1997; Schmitt et al. 2004), and no cam-
bial activity was observed at a temperature of 2�C
(Philipson et al. 1971) or with soil temperatures below
3–5�C (Turner and Streule 1983; Shonenberger and
Frey 1988; Korner 2003). Grace (1989) proposed stem
growth at the polar treeline with temperatures above
7�C, while Kramer and Kozlowski (1979) reported that
plants tended to become dormant at temperatures
below 10�C. Malyseve (1993) found that temperatures
of between 0 and 5�C during the growing season were
more effective than temperatures exceeding 10�C in
delimiting the arctic forest boundary in northern Asia .
Comparative studies on temperature limits to cell
growth processes therefore suggest that the critical
temperature range is between 0 and 10�C (Korner
1998).
Relationships between climate and wood produc-
tion, in terms of tree-ring width or density, have been
widely studied at the treeline in the Alps (Gindl et al.
2000; Carrer and Urbinati 2001; Motta and Nola 2001).
A number of studies have revealed positive effects of
June–July temperature on tree-ring width in Pinus
cembra and Larix decidua (Urbinati et al. 1998; Carrer
and Urbinati 2004; Oberhuber 2004), but no reliable
conclusion could be reached about the period of wood
production at the treeline. Carrer et al. (1998) mea-
sured stem growth with band dendrometers and found
that intra-annual variations in stem size indicated that
tree-ring formation in L. decidua and Picea abies at the
treeline lasts 50–60 days, from mid-June to the begin-
ning of August. These results provided experimental
support for the proposed relationships between sum-
mer temperature and tree-ring width. However, the
radius changes recorded by dendrometers also take
into account variations in stem water content (Zweifel
et al. 2000; Deslauriers et al. 2006) and, consequently,
are only an indirect assessment of the xylogenetic
activity. Tranquillini (1979) reported cell production in
the cambium occurring between mid-June and mid-
July and cell wall formation persisting until October at
1950 m a.s.l. in the Tyrol (Austria) in L. decidua
seedlings. However, studies of xylem cell formation are
still required on tree-shaped plants in order gain a
better understanding of xylogenesis at the upper forest
limits.
The few analyses carried out to date which take cell
production and xylogenesis in high-altitude forests into
account have provided very little information on
cambial activity, duration of xylem cell development
and the effect of climate on treeline trees over a short
time scale. Data are therefore needed on periods of
intra-annual xylem cell formation and threshold tem-
peratures for cambial activity. The aim of the investi-
gation reported here was to assess the timing and
dynamics of cell production and differentiation in the
stems of high-altitude conifer species in order to link
the growth processes with air, soil and meristem tem-
peratures and, consequently, define thresholds of
cambial activity. Histological analyses of developing
xylem cells were carried out on samples collected
within a very short time scale (7 days), thereby pro-
viding a precise definition of intra-annual tree-ring
formation during 2002–2004 at two sites in the eastern
Italian Alps.
Materials and methods
Study sites
The study area was located near the Cinque Torri
mountain group (Cortina d’Ampezzo, Belluno prov-
ince), at a high altitude above the upper limit of the
closed forest, in the eastern Italian Alps (46�27¢N,
12�08¢E). Two sites (5T-S and 5T-N) with uneven-aged
trees were selected on the two opposite slopes of the
mountains. 5T-S was located at 2080 m a.s.l. on a
south-facing slope, within a mixed open forest con-
sisting of groups of 5–15 trees of larch (Larix decidua
Mill.) and stone pine (Pinus cembra L.) and the occa-
sional Norway spruce [Picea abies (L.) Karst.]; as such,
this site corresponds to the timberline ecotone defini-
tion (Korner 1998). The forest originated from a col-
onisation of abandoned pastures and areas felled
during the first World War. 5T-N was located at a
distance of 500 m from 5T-S, on the north-facing slope
at 2130 m a.s.l. This site corresponds to a treeline
2 Oecologia (2007) 152:1–12
123
ecotone, as trees were very sparse and tree height
above the site decreased abruptly, with only isolated
larches and pines.
Temperature monitoring
Two standard weather stations were installed to mea-
sure air, soil and stem temperatures as a means of
monitoring microclimate variations at the sites. These
stations were positioned between the groups of trees at
site 5T-S and between the sparse trees at site 5T-N.
Temperatures were measured at intervals of 1 min and
recorded as averages every 15 min by means of CR10X
dataloggers (Campbell Scientific Corporation). Air
temperature was measured at 2 m. Soil temperature
was measured by six sensors per site which were placed
in the soil at a depth of about 15 cm. Stem temperature
was measured on two selected trees per species at 5T-S
and on three pines and one spruce at 5T-N. Stem
sensors were placed facing south, at a height of 1.3 m,
beneath the bark close to the cambial zone and pro-
tected by insulating shields. Daily mean, minimum and
maximum air, stem and soil temperatures were later
calculated with the time series obtained from the 96
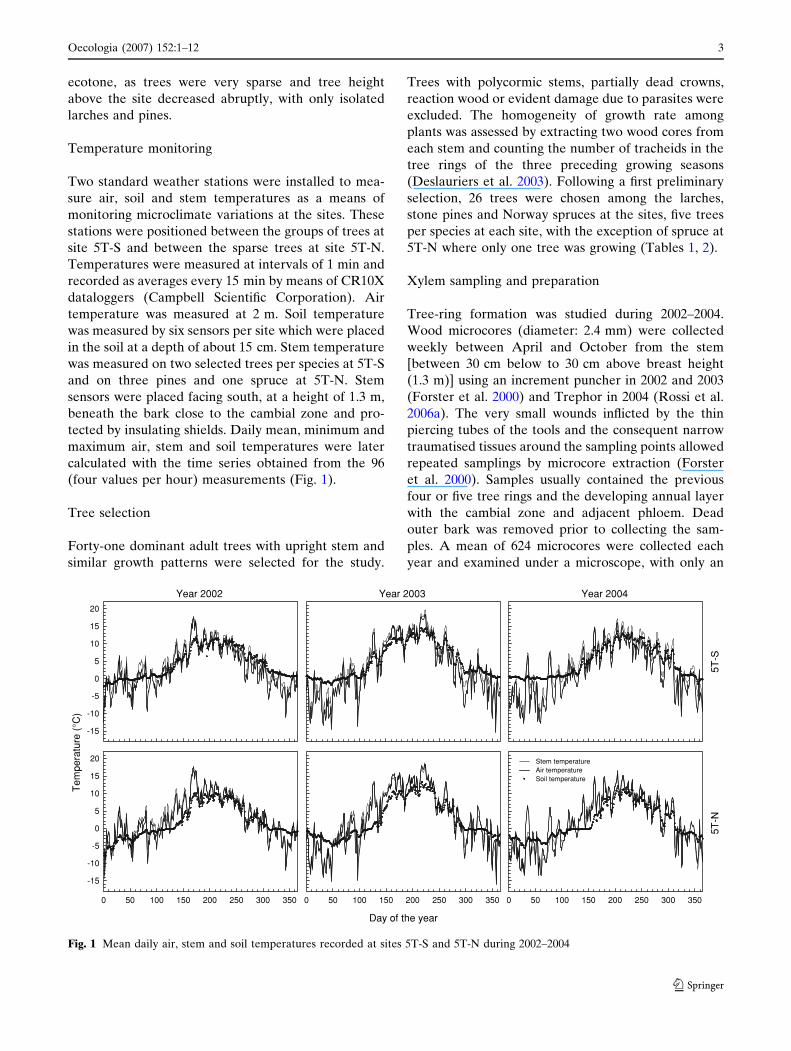
(four values per hour) measurements (Fig. 1).
Tree selection
Forty-one dominant adult trees with upright stem and
similar growth patterns were selected for the study.
Trees with polycormic stems, partially dead crowns,
reaction wood or evident damage due to parasites were
excluded. The homogeneity of growth rate among
plants was assessed by extracting two wood cores from
each stem and counting the number of tracheids in the
tree rings of the three preceding growing seasons
(Deslauriers et al. 2003). Following a first preliminary
selection, 26 trees were chosen among the larches,
stone pines and Norway spruces at the sites, five trees
per species at each site, with the exception of spruce at
5T-N where only one tree was growing (Tables 1, 2).
Xylem sampling and preparation
Tree-ring formation was studied during 2002–2004.
Wood microcores (diameter: 2.4 mm) were collected
weekly between April and October from the stem
[between 30 cm below to 30 cm above breast height
(1.3 m)] using an increment puncher in 2002 and 2003
(Forster et al. 2000) and Trephor in 2004 (Rossi et al.
2006a). The very small wounds inflicted by the thin
piercing tubes of the tools and the consequent narrow
traumatised tissues around the sampling points allowed
repeated samplings by microcore extraction (Forster
et al. 2000). Samples usually contained the previous
four or five tree rings and the developing annual layer
with the cambial zone and adjacent phloem. Dead
outer bark was removed prior to collecting the sam-
ples. A mean of 624 microcores were collected each
year and examined under a microscope, with only an
2002raeY
51-
01-
5-
0
5
01
51
02
3002raeY 4002raeY
053003052002051001050
Tem
pera
ture
(°C
)
51-
01-
5-
0
5
01
51
02
raeyehtfoyaD
053003052002051001050
5T-S
5T-N
053003052002051001050
erutarepmetmetSerutarepmetriAerutarepmetlioS
Fig. 1 Mean daily air, stem and soil temperatures recorded at sites 5T-S and 5T-N during 2002–2004
Oecologia (2007) 152:1–12 3
123

infrequent detection of isolated resin ducts. Wood
samples were always taken at least 5 cm apart to avoid
getting resin ducts on adjacent cores, which is a com-
mon reaction to disturbance in conifers (Deslauriers
et al. 2003). As only 13 sections in 3 years showed
tangentially oriented clusters of resin ducts in the
developing tree ring, there was clearly only incidental
spreading of the disturbance reaction of the xylem to
the wound to adjacent samples.
The microcores were placed in Eppendorf microtu-
bes with an ethanol solution (50% in water) and stored
at 5�C to avoid tissue deterioration. Microcores were
oriented by marking the transversal side with a pencil
under a stereo-microscope at a magnification of 10–
20·, dehydrated through a successive series of ethanol
and D-limonene and embedded in paraffin (Rossi et al.
2006a). Transverse sections were cut (6–10 lm in
thickness) from the samples with a rotary microtome.
Xylem analysis
Sections were stained for 10 min with cresyl violet
acetate (0.16% in water) and observed within 20 min
under visible and polarized light at a magnification of
400–500· to differentiate the developing xylem cells. In
each sample, the radial number of cells in the cambial
zone, radial enlargement phase, cell-wall thickening
phase and mature cells were counted along three radial
files. In cross-section, cambial cells were characterized
by thin cell walls and small radial diameters (Rossi
et al. 2006b). During cell enlargement, the tracheids
were composed of a protoplast still enclosed in the thin
primary wall but with a radial diameter at least twice
that of a cambial cell. Deformed rows of tracheids were
frequently observed in this phase due to the enlarge-
ment process occurring despite strong compression
between xylem tissues and bark. Observations under
polarized light discriminated between enlarging and
cell-wall thickening tracheids. Because of the
arrangement of cellulose microfibrils, the developing
secondary walls glistened when observed under polar-
ized light. However, no glistening was observed in the
enlargement zones where the cells still only consisted
of a primary wall (Abe et al. 1997). The progress of
cell-wall lignification was followed with cresyl violet
acetate stain, which reacted with the lignin (Rossi et al.
2006b), changing from violet to blue with increasing
lignification. The colour change over the whole cell
wall revealed the end of lignification and maturation of
the tracheid (Gricar et al. 2005).
The cell number in the three files was averaged for
each tree and used to assess onset, duration and ending
of xylem growth. Xylem formation was considered to
have begun in the spring when at least one horizontal
row of cells was observed in the enlarging phase. In
late summer, when no further cell was observed to be
undergoing wall thickening and lignification, xylem
formation was considered to be complete.
Statistical analyses
Analysis of variance (ANOVA) was used to compare
results (onset, duration and ending of xylogenesis, final
number of cells produced) between sites and species.
Responses among years were considered as within-
subject effects in repeated measures ANOVA [GLM
Table 1 Characteristics of the sampled trees at sites 5T-S and 5T-N in terms of mean and standard deviation of tree heights, diametersand ages
Species Height (m) Diameter (cm) Age (year)
5T-S 5T-N 5T-S 5T-N 5T-S 5T-N
Larix decidua 11.6 ± 1.0 6.4 ± 0.7 26.8 ± 4.8 15.4 ± 1.6 60.6 ± 18.7 63.8 ± 9.2Pinus cembra 10.1 ± 0.7 6.7 ± 1.5 29.2 ± 8.6 20.4 ± 5.2 60.4 ± 7.9 56.0 ± 13.4Picea abies 10.4 ± 0.6 8.0 22.4 ± 6.2 22.8 45.0 ± 8.7 74.0
Table 2 June–September mean air, stem and soil temperatures during 2002–2004 at sites 5T-S and 5T-N
Year Air temperature (�C) Stem temperature (�C) Soil temperature (�C)
5T-S 5T-N 5T-S 5T-N 5T-S 5T-N
2002 8.9 (2.8)a 9.1 (2.7) 10.2 (4.5) 9.6 (3.3) 9.3 (4.2) 7.3 (1.8)2003 11.4 (2.9) 11.6 (2.8) 12.8 (4.8) 11.7 (3.3) 11.1 (4.9) 9.3 (2.4)2004 9.0 (2.0) 9.2 (1.9) 11.1 (4.3) 9.7 (1.7) 9.8 (4.6) 7.0 (2.6)
a Annual mean temperatures are reported in parentheses. All values were calculated from daily means
4 Oecologia (2007) 152:1–12
123

procedure in SAS (1999)]. The hypothesis of com-
pound symmetry of the variance-covariance matrix
(Mauchly’s criterion) and sphericity (Huynh-Feldt
condition) were tested to check the assumptions for
using repeated measures ANOVA (Potvin et al. 1990).
The ANOVA model residuals were also examined
graphically and by performing the Shapiro-Wilk sta-
tistic [UNIVARIATE procedure in SAS (1999)] for
outliers and evidence of non-normality (Quinn and
Keough 2002).
Logistic regressions [LOGISTIC procedure in SAS
(1999)] were used to calculate the probability of xylo-
genesis being active at a given temperature where
binary responses were coded as non-active (value zero)
or active (value 1). The logistic regression takes the
general form:
LogitðpxÞ ¼ lnpx
1� px
� �¼ b0 þ b1xj
where px is the probability of xylogenesis being active,
xj is the temperature on a given day j, b0 and b1 are the
intercept and slope of the logit regression (Quinn and
Keough 2002). Temperatures thresholds (x) were cal-
culated when the probability of xylogenesis being ac-
tive was 0.5, i.e. when Logit(p) = 0 and x = –b0/b1.
Therefore, for a temperature above x, the wood for-
mation was more likely to be active than non-active.
Model verification included v2 of the likelihood ratio,
Wald’s v2 for regression parameter and goodness of fit
and Hosmer-Lemeshow’s C for eventual lack of fit
(Quinn and Keough 2002). For each tree, site and year,
the model was fitted with the respective temperature
series (mean, minimum and maximum air, stem and
soil temperatures). Only ten of 702 models were ex-
cluded because of a lack of fit. Temperature thresholds
were then compared between sites and species using
univariate ANOVA models [GLM procedure in SAS
(1999)].
Results
Air, stem and soil temperature
The area around the Cinque Torri has a typical alpine
climate of cold winters and cool summers, with mean
daily air temperatures reaching 17–20�C during 2002–
2004. In the three study years, the mean annual air
temperature was between 1.9 and 2.9�C and the mean
June–September air temperature was between 8.9 and
11.6�C, with higher values recorded in 2003 (Table 2).
The mean daily air temperature was very similar at
sites 5T-S and 5T-N, with differences restricted to
0.2�C during the warmest months (Fig. 1). However,
the main differences between sites were observed in
the day/night temperatures. Warmer daytime air tem-
peratures and colder temperatures during the night
were recorded at site 5T-S; consequently, daily tem-
perature excursions were higher at the south-facing
site. Stem temperature was coupled to air temperature,
with higher values collected on trees at site 5T-S.
However, stem temperature was closer to air temper-
ature from April to September while, in the colder
months, air temperature data with wider amplitudes
and higher temperature excursions were recorded.
Major differences were observed between the two sites
for annual and June–September soil temperatures
(Table 2). During sampling in May, snowmelts were
observed to occur earlier at site 5T-S. Soil tempera-
tures recorded in late spring also confirmed these
observations: during the snowmelt at site 5T-N, the
soil temperature fluctuated around 0�C and the soil
only began to warm when the snow had disappeared,
10–15 days after site 5T-S (Fig. 1).
Cambial activity
Similar annual dynamics were observed in the cambial
zones of the three tree species (Fig. 2). In spring and
autumn, when there was no cell production, the dor-
mant cambium consisted of six to eight cells in close
proximity to each other, as shown by the horizontal
dotted line in Fig. 2. In May, the number of cells in the
cambial zone had increased to 12–14, indicating the
onset of cell division in the cambium. Some variations
among trees were observed in terms of cambial cell
numbers, especially during the spring and early sum-
mer. Cambial activity was considered to have begun
when the number of cambial cells exceeded the mini-
mum number of dormant cells and the standard devi-
ation did not cross the horizontal dotted line (Fig. 2).
The earliest increases in the number of cambial cells
were observed in pine at the beginning of May; this was
followed 1–2 weeks later in larch and spruce. At site
5T-N, wide cambial zones were always observed
1–2 weeks later than at site 5T-S, with the exception of
pine in 2002 and spruce in 2004. However, the first
samplings in 2002 began in mid-May when pine was
already showing wide cambial zones, thereby pre-
venting a precise assessment of the differences
between the two sites for this species. The earliest
cambial activity onset was observed for the three
species in 2003, when very high temperatures were also
recorded in April–May (Fig. 1).
Oecologia (2007) 152:1–12 5
123

Once annual activity had finished and the cambium
had stopped dividing, the number of cells in the cam-
bial zone fell to a minimum value that corresponded to
quiescence conditions of the meristems. High vari-
ability was also observed at the end of cell production.
Halts in cambial activity were observed from mid-July
L. decidua
468
1012141618
P. abies P. cembra
468
1012141618
468
1012141618
sllecl
aib
mac
fo
re
bm
uN
468
1012141618
468
1012141618
Day of the year
100 150 200 250 300 100 150 200 250 300100 150 200 250 300468
1012141618
20
02
ra
eY
30
02
ra
eY
40
02
ra
eY
Fig. 2 Numbers of cells in the cambial zones of Larix decidua,Picea abies and Pinus cembra during 2002–2004 at sites 5T-S(black dots) and 5T-N (white dots). Error bars and horizontaldotted line indicate the standard deviations among trees in terms
of the number of dormant cambial cells. Periods of cambialactivity, when error bars do not cross the horizontal dotted line,are highlighted in grey
6 Oecologia (2007) 152:1–12
123

for pine in 2004 (5T-N) to the end of August for spruce
in 5T-S. Cell division ended earlier in trees at site 5T-N
than in trees at site 5T-S, with the exception of larch in
2002. Delays in the onset and ending of cambial
activity led to an average reduction of 30% in the
overall period for cell production between sites 5T-S
and 5T-N. However, a very short period of cambial
activity was observed in 2002 for pines growing at site
5T-S in which there was a rapid decline in the number
of cambial cells, from 14.2 on 28 May (day 148 of the
year) to 10.6 on 25 June (day 176).
Cell differentiation
Onset, duration and ending of cell differentiation were
computed in days of the year for each tree. The aver-
ages are reported in Fig. 3, where the bars correspond
to the mean value and error bars to the standard
deviation between the five trees.
The onset of radial enlargement occurred between
mid-May and mid-June and was significantly different
between the three species (ANOVA, F = 23.20,
P < 0.0001) (Fig. 3a–c). The earliest start of radial
enlargement was observed every year in pine tracheids
at both sites. Larch tracheids began cell differentiation
4–14 days after pine and 2–11 days before spruce, with
the exception of 2002 when tracheid enlargement was
observed to occur earlier in spruce than in larch. In all
species, the onset of cell differentiation occurred 3–
10 days earlier at the south-facing site (ANOVA,
F = 14.90, P < 0.01). Significant differences among
years were detected by repeated measurements
(ANOVA, F = 16.57, P < 0.0001). The first tracheids
were observed in cell enlargement 3–13 days later in
2002 and 2004 than in 2003.
Xylogenesis was considered to be concluded when
cells were no longer observed in the process of radial
enlargement, wall thickening or lignification. Cell dif-
ferentiation ended from early September to mid-Octo-
ber (Fig. 3d–f). The range of variation for the
completion of xylogenesis was greater than that for
onset of cell differentiation (44 days for end of cell dif-
ferentiation versus 30 days for the onset of cell
enlargement). Significant variations in the ending of cell
differentiation were observed among species (ANOVA,
F =14.79, P < 0.001), with average differences of 8 days
between pine and larch and 2 weeks between pine and
spruce. Spruce were the last to complete lignification,
except in 2003 when immature cells were observed in
larch until the end of September. Cell differentiation
ended earliest at site 5T-N for all species (ANOVA,
F = 17.03, P < 0.001), with larches showing the greatest
differences between sites 5T-S and 5T-N. The repeated
measurements revealed a significant difference among
the 3 years (ANOVA, F = 66.14, P < 0.0001), with xy-
logenesis ending much later in 2004 – after 1 October
and 22 September in 5T-S and 5T-N, respectively.
The duration of xylogenesis – i.e., the time required
to complete cell differentiation for all the tracheids
forming the tree ring – varied between 90 and 137 days
(Fig. 3g–i), with no difference detected between species
(ANOVA, F = 0.16, P > 0.05). Conversely, there were
significant differences between sites (ANOVA,
F = 23.03, P < 0.001), with longer durations of tree-ring
formation estimated for site 5T-S, where 123.5 days
were required to complete cell differentiation. At site
5T-N, the average period between the onset of cell
enlargement and ending of lignification was 107 days.
Larger reductions were observed for larch in all 3 years
and for pine in 2002. Significant variations among the
3 years were detected by repeated measurements
(ANOVA, F = 23.49, P < 0.0001). In 2004, the average
duration of tree-ring formation was 124 days versus 113
estimated in 2003. Shorter periods were calculated in
2002 for completion of the tree ring, with 108 days of
cell differentiation.
Interaction effects between sites and species were
also tested, but the results were not significant
(P > 0.05), thus indicating that site effects on onset,
duration and ending of xylogenesis were independent
of species.
Xylem cells in the tree ring
At the end of the growing season, we found different
numbers of cells in the tree ring of the three species
(ANOVA, F = 8.82, P < 0.01), as reported in Fig. 4.
The highest numbers of tracheids were observed in
spruce, with values ranging from 45 (in 2002, 5T-N) to
76 (in 2002, 5T-S), while the number of xylem cells in
larch varied between 32 (in 2002, 5T-N) and 51 (in 2003,
5T-S). Fewer cells were produced in trees at site 5T-N
than at 5T-S (ANOVA, F = 6.58, P < 0.05), with
15–30% fewer tracheids in the tree ring (57 cells pro-
duced in 5T-S versus 45 cells in 5T-N overall for the
three species). The final number of cells varied signifi-
cantly between years (ANOVA, F = 16.77, P < 0.0001),
with the fewest tracheids observed in 2002. There
was no evident pattern in 2003 and 2004. The largest
numbers of cells were produced by spruce in 2004 and
by larch and pine in 2003.
Threshold temperatures
The threshold temperature at which there was a 0.5
probability of active xylogenesis was calculated and
Oecologia (2007) 152:1–12 7
123

reported as an average for each species and site
(Fig. 5). For air temperature, the ranges of thresholds
when both sites were considered were 1.7–4.8, 5.6–8.5
and 10.9–13.3�C for the minimum, mean and maximum
temperatures, respectively. The minimum, mean and
maximum stem temperatures at which there was a 0.5
probability of active xylogenesis were higher than the
air temperature threshold, being 2.6–4.2, 7.2–9 and
17.2–22�C (Fig. 5). The standard deviations associated
with the mean values of stem temperature thresholds
were low, indicating a very slight variation between
trees, especially for the minimum and mean stem
temperature. Lower thresholds were found for mini-
mum (0.2–4.5�C), mean (2.6–7.5�C) and maximum
(8.3–12.6�C) soil temperatures. No difference was
found between species and sites (ANOVA, P > 0.05),
with the exception of minimum air temperature
(ANOVA, F = 1.84, P < 0.05) and all soil temperature
series (ANOVA, P < 0.01).
Discussion
The growth limitation hypothesis attempted to explain
the existence of cold treelines throughout the world by
suggesting that cell formation (i.e. cell division and
differentiation) could not occur below a minimum
temperature threshold; if it did occur, however, there
would be an abrupt slow down (Korner 1998).
According to several authors (Korner 1998, 2003), this
minimum temperature should range between 0 and
10�C. According to Korner (2003), mean air and stem
threshold temperatures of 6–8�C define treeline posi-
tions worldwide . However, these thresholds were only
P. abies
noitamrof
gnir-eertfognidn
E)r aey
ehtfosyad(
200
220
240
260
280
300
2002 2003 2004
fonoitaru
D)syad(
noitamrof
gnir-eert 60
80
100
120
140
160
Year
2002 2003 2004 2002 2003 2004
L. decidua
gnir-eertfotesnO
)raeyehtfo
syad(noita
mrof 100
120
140
160
180P. cembra
5T-S5T-N
a b c
d e f
g h i
Fig. 3 Onset (a–c), ending (d–f) and overall duration (g–i) of tree-ring formation for L. decidua, P. abies and P. cembra at sites 5T-Sand 5T-N during 2002–2004. Error bars indicate standard deviation among trees
8 Oecologia (2007) 152:1–12
123

calculated on an annual basis. Our histological analyses
performed on at a weekly scale showed that xylogen-
esis was active when the minimum daily air tempera-
ture was above 2–4�C and the minimum stem
temperature was higher than 4�C. The converging
temperature thresholds among sites and species in the
three study years confirmed the existence of thermal
limits to stem growth, thus also explaining the shorter
periods of xylogenesis observed at the north-facing
site.
Cambium is a sink for non-structural carbohydrates,
and cambial activity requires a continuous supply of
energy in the form of sucrose which, for the first cells to
be formed, is extracted from the storage tissues or
L. decidua
0
20
40
60
80
100
5T-S
5T-N
P. abies
sllec fo rebmu
Ngnir eert eht ni
0
20
40
60
80
100
P. cembra
Year
2002 2003 20040
20
40
60
80
100
Fig. 4 Final number of xylem cells produced by L. decidua, P.abies and P. cembra during 2002–2004 at sites 5T-S and 5T-N.Error bars indicate the standard deviation among trees
L. decidua
)C°(
erutarepmet
riA
0
2
4
6
8
10
12
14
5T-S 5T-N
)C°(
erutarepmetlio
S
0
2
4
6
8
10
)C°(
erutarepmet
metS
0
4
8
12
16
20
P. abies
Site
5T-S 5T-N
P. cembra
5T-S 5T-N
Fig. 5 Threshold minimum (black dots), mean (white dots) andmaximum (grey dots) temperatures corresponding with the 0.5-probability of active xylogenesis for L. decidua, P. abies and P.cembra estimated during 2002–2004 at sites 5T-S and 5T-N.Error bars indicate the standard deviation among trees
Oecologia (2007) 152:1–12 9
123

produced by photosynthesis (Hansen and Beck 1990,
1994; Oribe et al. 2003). During cell maturation, trees
assign a large amount of carbon obtained from pho-
tosynthesis to the deposition of cellulose microfibrils in
order to provide the developing cells with secondary
walls (Hansen et al. 1997). The estimated mean tem-
perature of 6–8�C seems to be the threshold limiting
the demand for photo-assimilates by the metabolic
processes involved in cell growth. Since shoot exten-
sion was also inhibited by air temperatures lower than
6–8�C (James et al. 1994), and 5% of maximum rate of
root growth occurred at a soil temperature of 6�C
(Turner and Streule 1983 in Shonenberger and Frey
1988), a critical mean temperature of between 6 and
8�C does exist, affecting growth processes in all parts of
the tree (shoots, stem and roots). The temperature-
limited growth mechanisms at the cell level are,
therefore, also expected to be the same in all plant
meristems.
The onset of cell division occurred early in the
season, when air and stem temperatures were still low,
and it ended at the end of July or, at the latest, in
August, when stem temperatures achieved maximum
values (12–15�C; compare Figs. 1 and 2). The high
spring temperatures in 2003 induced an earlier
resumption of cell production in the cambium and a
consequent earlier onset of xylem cell differentiation.
Shorter durations of cambial activity were observed in
trees at the north-facing site (5T-N), where cell pro-
duction lasted for 6–11 weeks. These results are con-
sistent with the 6-week-long active period required to
maintain long-term growth by Pinus sylvestris near the
treeline in northern Finland (Schmitt et al. 2004). Al-
though the conclusion of cell division indicated the end
of tracheid production, wood formation continued
until the autumn through enlargement, wall formation
and lignification of the newly produced tracheids. The
differentiation processes are dynamic phases with
durations in earlywood and latewood (Rossi et al.
2006b). The large amount of woody material deposi-
tion that initiates the formation of thicker cell walls in
latewood tracheids is related to a longer duration ra-
ther than higher rate of secondary wall formation
(Uggla et al. 2001). In the species studied here, the
latest latewood tracheids remained in the maturation
phase for up to 40–60 days, as reported for the same
sites by Rossi et al. (2006b); by subtracting the Julian
days, this corresponded to the end of differentiation
(Fig. 3d–f) and conclusion of cambial activity (Fig. 2).
Trees living in cold climates concentrate xylem cell
formation early in the season, synchronizing cambial
activity with photoperiod and culminating wood pro-
duction at the end of June when the day length is
maximum (Rossi et al. 2006c). By avoiding high cell
production rates during late summer, plants guarantee
newly formed tracheids enough time before winter to
complete differentiation, particularly cell-wall forma-
tion and lignification in latewood.
In larch, stone pine and Norway spruce, overall
wood formation lasted for a period varying from 100 to
140 days between mid-May and the beginning of
October. Korner and Paulsen (2004) also estimated an
(unspecified) growing period of 129–139 days for the
eastern Alps by counting the days when the soil tem-
perature at a depth of 10 cm exceeded 3.2�C, which in
the spring corresponded to a weekly mean canopy
temperature of 0�C. Between the south and north-
facing sites, Differences of 1–3 weeks in the active
period of cambium have been estimated between the
south- and north-facing sites, based on rapid declines in
height and root seasonal growth patterns observed
across altitudinal gradients in treeline ecotones (Han-
sen-Bristow 1986; Stevens and Fox 1991; Sveinbjorns-
son 2000). This reduction led to differences of 5–
25 days in the duration of wood formation, and these
differenceswere in turn related to delays of 3–10 days
in the onset of differentiation and advances of 3–
20 days in the ending of cell-wall lignification. A de-
layed onset of cell enlargement found in trees at the
north-facing site (5T-N) could not be related to dif-
ferences in air temperatures given the similarity be-
tween sites. On the contrary, greater differences
between sites were observed in the stems, with tem-
peratures being up to 1.4�C warmer at the southern site
(5T-S), and in the soil during late spring. Other studies
have reported that where temperature strongly limits
tree radial growth, the thawing of the upper soil layer
must have started before radial growth is able to
commence (Graumlich and Brubaker 1986; Cairns and
Malanson 1998; Vaganov et al. 1999). Cold soils inhibit
root activity and water uptake, thus delaying turgor
increase in the vacuoles and enlargement of the newly-
produced tracheids in the stem. However, our results
showed different soil threshold temperatures between
sites, thus indicating that soil temperature could not be
the main limiting factor affecting xylogenesis. In terms
of stem temperature, converging thresholds were ob-
served for the two sites, demonstrating that a minimum
threshold of 3–4�C has to be reached to sustain xylo-
genesis. As stem sensors measure cambium tempera-
ture, where the xylogenesis processes occur, this result
demonstrates the direct effect of temperature on the
metabolic activities of wood formation (Grace 1988;
James et al. 1994).
In the three study years, differences of 15–18 days
were estimated in the duration of secondary stem
10 Oecologia (2007) 152:1–12
123

growth, with a variability of up to 15% in complete cell
maturation. Delays in the onset of radial growth did
not necessarily correspond to corresponding delays in
the ending of growth. Higher variability was observed
at the conclusion of differentiation: about 10 ± 8 days
at the start of xylogenesis versus 22 ± 14 days at the
end of lignification at both sites. Wood formation is a
complex process with several differentiation phases.
Cells produced in spring and early summer must pass
through several of these phases before reaching phys-
iological maturity. Moreover, a higher cell production
during cambial activity leads to an increased number of
developing tracheids (Ford et al. 1978) and, conse-
quently, prolonged cell maturation later in the season
(Gricar et al. 2005).
Conclusion
The results of our study reveal that in larch, stone
pine and Norway spruce, wood formation occurred
when certain threshold temperatures were reached.
Although the timing and duration of xylogenesis
varied among these species, sites and years, air and
stem temperature thresholds were stable for all of the
trees studied, ranging from 5.6 to 8.5�C and from 7.2
to 9�C, respectively. These results correspond to the
supposed temperatures limiting growth at the treeline
and thus provide strong evidence that temperature is
a critical factor controlling xylem cell production and
differentiation at high altitudes. This study represents
the first attempt to define a threshold through com-
parative assessment of xylem growth and tissue tem-
peratures in stem meristems of trees growing at high
altitudes.
Acknowledgments This work was funded by the MAXY 2004(CPDA045152) and MIUR-PRIN 2005 (2005072877). The au-thors wish to thank C. Filoso, F. Fontanella, M. Gardin, L. Ma-rini, M. Mazzaro and R. Menardi for their technical support andthe Regole of Cortina d’Ampezzo for permitting the study ontheir property. Special thanks are extended to M. Carrer and C.Korner for their recommendations on the manuscript.
References
Abe H, Funada R, Ohtani J, Fukazawa K (1997) Changes in thearrangement of cellulose microfibrils associated with thecessation of cell expansion in tracheids. Trees 11:328–332
Beniston M, Diaz HF, Bradley RS (1997) Climatic change athigh elevation sites: an overview. Clim Change 36:233–251
Cairns DM, Malanson GP (1998) Environmental variablesinfluencing the carbon balance at the alpine treeline: amodeling approach. J Veg Sci 9:679–692
Carrer M, Urbinati C (2001) Spatial analysis of structural andtree-ring related parameters in a timberline forest in theItalian Alps. J Veg Sci 12:643–652
Carrer M, Urbinati C (2004) Age-dependent tree-ring growthresponses to climate in Larix decidua and Pinus cembra.Ecology 85:730–740
Carrer M, Anfodillo T, Urbinati C, Carraro V (1998) High-altitude forest sensitivity to global warming: results fromlong-term and short-term analyses in the Eastern ItalianAlps. In: Beninston M, Innes JL (eds) The impacts ofclimate variability on forests. Springer, Berlin HeidelbergNew York, pp 171–189
Deslauriers A, Morin H (2005) Intra-annual tracheid productionin balsam fir stems and the effect of meteorologicalvariables. Trees 19:402–408
Deslauriers A, Morin H, Begin Y (2003) Cellular phenology ofannual ring formation of Abies balsamea in the Quebecboreal forest (Canada). Can J For Res 33:190–200
Deslauriers A, Rossi S, Anfodillo T (2006) Dendrometer andintra-annual tree growth: what kind of information can beinferred? Dendrochronologia (in press)
Ford ED, Robards AW, Piney MD (1978) Influence of environ-mental factors on cell production and differentiation in theearlywood of Picea sitchensis. Ann Bot 42:683–692
Forster T, Schweingruber FH, Denneler B (2000) Incrementpuncher: a tool for extracting small cores of wood and barkfrom living trees. IAWA J 21:169–180
Gindl W, Grabner M, Wimmer R (2000) The influence oftemperature on latewood lignin content in treeline Norwayspruce compared with maximum density and ring width.Trees 14:409–414
Grace J (1988) Temperature as a determinant of plant produc-tivity. In: Long SP, Woodward FI (eds) Plants and temper-ature. Cambridge University Press, Cambridge, pp 91–107
Grace J (1989) Tree lines. Philos Trans R Soc Lond B324:233–245
Grace J, Norton DA (1990) Climate and growth of Pinussylvestris at its upper altitudinal limit in Scotland: evidencefrom tree growth-rings. J Ecol 78:601–610
Graumlich LJ, Brubaker LB (1986) Reconstruction of annualtemperature (1590–1979) for Longmire, Washington, de-rived from tree-rings. Quat Res 25:223–234
Gricar J, Cufar K, Oven P, Schmitt U (2005) Differentiation ofterminal latewood tracheids in silver fir trees during autumn.Ann Bot 95:959–965
Hansen-Bristow K (1986) Influence of increasing elevation ongrowth characteristics at timberline. Can J Bot 64:2517–2523
Hansen J, Beck E (1990) The fate and path of assimilationproducts in the stem of 8-year-old Scots pine (Pinussylvestris L.) trees. Trees 4:16–21
Hansen J, Beck E (1994) Seasonal changes in the utilization andturnover of assimilation products in 8-year-old Scots pine(Pinus sylvestris L.) trees. Trees 8:172–182
Hansen J, Turk R, Vogg G, Heim R, Beck E (1997) Conifercarbohydrate physiology: updating classical views. In: Ren-nenberg H, Eschrich W, Ziegler H (eds) Trees: contribu-tions to modern tree physiology. Backhuys Publishers,Leiden, pp 97–108
Hattenschwiler S, Korner C (1995) Responses to recent cli-matewarming of Pinus sylvestris and Pinus cembra withintheir montane transition zone in the Swiss Alps. J Veg Sci6:357–368
Holtmeier F-K (1997) Timberlines: research in Europe andNorth America. In: Loven L, Salmela S (eds) Pallastunturisymposium. Finnish Forest Research Institute, Finland, pp23–36
James JC, Grace J, Hoad SP (1994) Growth and photosynthesisof Pinus sylvestris at its altitudinal limit in Scotland. J Ecol82:297–306
Oecologia (2007) 152:1–12 11
123

Korner C (1998) A re-assessment of high elevation treelinepositions and their explanation. Oecologia 115:445–459
Korner C (2003) Alpine plant life: functional plant ecology ofhigh mountain ecosystems, 2nd edn. Springer, Berlin Hei-delberg New York
Korner C, Paulsen J (2004) A world-wide study of high altitudetreeline temperatures. J Biogeogr 31:713–732
Kramer PJ, Kozlowski TT (1979) Physiology of woody plants.Academic, New York
Malyshev L (1993) Levels of the upper forest boundary innorthern Asia. Vegetatio 109:175–186
Motta R, Nola P (2001) Growth trends and dynamics in sub-alpine forest stands in the Varaita valley (Piedmont, Italy)and their relationships with human activities and globalchange. J Veg Sci 12:219–230
Oberhuber W (2004) Influence of climate on radial growth ofPinus cembra within the alpine timberline ecotone. TreePhysiol 24:291–301
Oribe Y, Funada R, Kubo T (2003) Relationships betweencambial activity, cell differentiation and the localisation ofstarch in storage tissues around the cambium in locallyheated stems of Abies sachalinensis (Schmidt) Masters.Trees 17:185–192
Philipson WR, Ward JM, Butterfield BG (1971) The vascularcambium: its development and activity. Chapman & Hall,London
Pisaric MFJ, Holt C, Szeicz JM, Karst T, Smol JP (2003)Holocene treeline dynamics in the mountains of northeast-ern British Columbia, Canada, inferred from fossil pollenand stomata. Holocene 13:161–173
Potvin C, Lechowicz MJ, Tardif S (1990) The statistical analysisof ecophysiological response curves obtained from experi-ments involving repeated measures. Ecology 71:1389–1400
Quinn GP, Keough MJ (2002) Experimental design and dataanalysis for biologists. Cambridge University Press, Cambridge
Rossi S, Anfodillo T, Menardi R (2006a) Trephor: a new tool forsampling microcores from tree stems. IAWA J 27:89–97
Rossi S, Deslauriers A, Anfodillo T (2006b) Assessment of cambialactivity and xylogenesis by microsampling tree species: anexample at the Alpine timberline. IAWA J 27:383–394
Rossi S, Deslauriers A, Anfodillo T, Morin H, Saracino A, MottaR, Borghetti M (2006c) Conifers in cold environments
synchronize maximum growth rate of tree-ring formationwith day length. New Phytol 169:279–290
SAS (1999) SAS version 8.02. SAS Institute, Cary, N.C.Schmitt U, Jalkanen R, Eckstein D (2004) Cambium dynamics of
Pinus sylvestris and Betula spp. in the northern boreal forestin Finland. Silva Fenn 38:167–178
Shonenberger W, Frey W (1988) Untersuchungen zur Okologieund Technik der Hochlagenaufforstung. For-schungsergebnisse aus dem Lawinenanrissgebiet Stillberg.Schweiz Z Forstwes 139:735–820
Smith WK, Germino MJ, Hancock TE, Johnson DM (2003)Another perspective on altitudinal limits of alpine timber-lines. Tree Physiol 23:1101–1112
Stevens GC, Fox JF (1991) The causes of treeline. Annu RevEcol Evol Syst 22:177–191
Sveinbjornsson B (2000) North American and European tree-lines: external forces and internal processes controllingposition. Ambio 29:388–395
Theurillat JP, Guisan A (2001) Potential impact of climatechange on vegetation in the European Alps: a review. ClimChange 50:77–109
Tranquillini W (1979) Physiological ecology of the alpinetimberline. Springer, Berlin Heidelberg New York
Turner H, Streule A (1983) Wurzelwachstum und Sprossent-wicklung junger Koniferen im Klimastress der alpinenWaldgrenze, mit Berucksichtigung von Mikroklima, Photo-synthese und Stoffproduktion. In: Bohm W, Kutschera L,Lichtenegger E (eds) Wurzelokologie und Ihre Nutzanwen-dung. Irding, Gumpenstein, pp 617–635
Uggla C, Magel E, Moritz T, Sundberg B (2001) Function anddynamics of auxin and carbohydrates during earlywood/latewood transition in Scots pine. Plant Physiol 125:2029–2039
Urbinati C, Carrer M, Sudiro S (1998) Dendroclimatic responsevariability of Pinus cembra L. in upper timberline forests ofItalian Eastern Alps. Dendrochronologia 15:101–117
Vaganov EA, Hughes MK, Kirdyanov AV, Schweingruber FH,Silkin PP (1999) Influence of snowfall and melt timing ontree growth in subarctic Eurasia. Nature 400:149–151
Zweifel R, Item H, Hasler R (2000) Stem radius changes andtheir relation to stored water in stems of young Norwayspruce trees. Trees 15:50–57
12 Oecologia (2007) 152:1–12
123
Related Documents











