EVIDENCE IMPLICATING THE NATRIURETIC PEPTIDE SYSTEM IN THE ANTIHYPERTENSIVE EFFECT OF MODERATE ETHANOL CONSUlMPTION Pascal Guillaume Department of Physiology McGill University, Montréal, Canada A thesis submitted to the Faculty of Graduate Studies and Research in partial fulfilment of the requirements for the degree of Doctor of Philosop& Q 1997 by Pascal Guillaume

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EVIDENCE IMPLICATING THE NATRIURETIC PEPTIDE SYSTEM IN THE ANTIHYPERTENSIVE
EFFECT OF MODERATE ETHANOL CONSUlMPTION
Pascal Guillaume
Department of Physiology McGill University, Montréal, Canada
A thesis submitted to the Faculty of Graduate Studies and Research in partial fulfilment of the requirements
for the degree of Doctor of Philosop&
Q 1997 by Pascal Guillaume

National Library Bibliothéque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Street 395. rue Wellington Ottawa ON K1A ON4 ûttawaON KtAON4 Canada Canada
Your hie Vohe reterbnce
Our W Nme relerBi>CB
The author has granted a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or sell copies of this thesis in microfom, paper or electronic formats.
The author retains ownership of the copyright in ths thesis. Neither the thesis nor substantid extracts fiom it may be printed or otherwise reproduced without the author's pemiission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/tilm, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.
Canada

Voici donc un roman.

ABSTRACT
Chronic moderate ethanol (ETOH) consumption prevents the development of the age-
dependent increase m blood pressure (BP) in both humans and experimental animals. In the
present midies we proposed that the natriuretic peptide f a d y may be partially responsible
for this antihypenensive effect of ETOH. The natnuretic -stem consists of the atrial
natriuretic peptide (ANP), the brain natriuretic peptide (BNP). the C-type natriuretic peptide
(CNP) and the natriuretic peptide receptors (NPRs). The major function of the natriuretic
-stem is to decrease BP. Thus. the main objective of the present midies was to investigate
the interactions of acute and chronic ETOH administration with various components of the
natriuretic systern Acute studies: The injection of 1 and 2 g of ETOWkg B. W. in Sprague-
Dawley rats and 0.25 and 0.50 g of ETOWkg B.W. in hurnans resulted in a rapid transient
increase of circulating ANP levels. in the rats, this increase in plasma ANP levels was
associated with a rapid decrease of atrial ANP content and with a delayed increase in
ventricular ANP levels. Chronic studies: A 20°h v/v solution of alcohol was -+en to
spontaneously hypertensive (SHR) and Wistar-Kyoto (WKY) rats for 8 months. In both SHR
and WKY rats. chonic ETOH treatment decreased the BP. This lower BP in ETOH-treated
animals was associated with lower circulatmg ANP and BNP levels. whereas ETOH treatment
increased atrial ANP and BNP tissue contents. but not ANP and BNP mRNA. Furthemore.
chronic ETOH treatment reduced heart ventncular ANP content and ANP rnRNA while it
increased ventricular BNP content and BNP mRNA in SHR, but not in WKY rats. Chronic
ETOH reduced total natnuretic peptide binding sites (NPR-A and NPR-C) in the renal
elornerdi. Quantification of the receptor mbtypes demonstrated that this decrease was due Ci
to the dom-regdation of NPR-C. In the renal papilla. chronic ETOH treatrnent increased
natnuretic peptide binding sites (NPR-A). In addition to its effects on the penpheral
natnuretic synem chronic ETOH treatment altered the activity of the natnuretic system at
the level of the brain. Thus. chronic ETOH increased ANP and CNP levels in the
hypothalamus, pons and medulla of SHR rats. In WKY rats, ETOH had no effect on brain
ANP levels. but enhanced the concentration of CNP in the hypothalamus and medulla.
Chronic ETOH treatment also decreased the a&ty of NPR-C in some of the
circumventrîcular organs ofthe brain. in the nibfomical organ and cboroid plexus. but not in

the area postrerna.
These results demonstrate that both acute and chronic moderate ETOH administration
ahers the a c t ~ t y of vanous components of the natriuretic system Therefore. these ETOH-
induced modifications in cardiac. renal and brain natriuretic peptides and receptors may
contnbute to the antihypertensive effect of chronic moderate ETOH drinking.

RÉsUMÉ
La consommation continue d'éthanol (ETOH) en quantité modérée previent
l'augmentation de la pression sanguine liée à Fige autant chez l'animal que chez Iliumam.
Cette présente série d'études avançe l'hypothèse que la famiue des peptides natriurétiques
pourrait être en partie responsable de cet effet antihypertenseur de l'ETOK Le système
natriUrétique est composé du peptide natriurénque auriculaire (ANP), du peptide natriurétique
cérébral (BNP), du peptide natriurétique de type C (Cm) et des réce?teurs natriurétiques
(NPRs). Le rôle principal de ce système est de diminuer la pression sanguine. L'objectif
premier de la présente séries d'études est donc l'analyse des effets aigues et chroniques de
llalcool sur les différentes composantes du système natriurétique. Études a'gües: I L'injection
de 1 et 2 g d'ETOWkg de poid corporel chez le rat Sprague-Dawley et l'ingestion de 0.25 et
0.50 g d'ETOH/kg de poid corporei chez 1'buma.h provoque une rapide mais coune
augmentation des niveaux plasmatiques &ANP. Chez le rat, cette augmentation est associée
à une rapide diminution du contenu ANP de l'oreilette et à une plus lente augmentation du
contenu ANP du ventricule. Études chroniques: Une solution &ETOH à 20% viv est
administrée pendant 8 mois a des rats spontanément hypertendus ( S m ) et Wistar-Kyoto
(WKY). Chez les deux groupes de rats, la consommation contmue d'alcool diminue la
pression sanguine. Cette diminution de la pression sanguine se traduit par une diminution des
niveaux plasmatiques d'ANP et de BNP. Cependant, le contenu ANP et BNP de l'oreilette est
augmenté, bien que les niveaux auriculaires d'ARNrn ne soient pas modifiés. De plus, la
consommation continue d'ETOH réduit les niveaux ANP et ANP ARNm et augmente les
niveaux BNP et BNP ARNm du ventricule chez les rats SHR seulement. L'administration
d'alcool est également associée a une réduction du nombre total de récepteurs natriurétiques
(NPR-A et NPR-C) dans les glomérules rénaux. La quantification sélective des types de
récepteurs démontre que cette diminution est due principalement à la régulation négative des
récepteurs NPR-C. Dans la papille rénale, l'administration d'ETOH augmente le nombre total
de récepteurs natriurétiques (NPR-A). En plus de ces effets sur le système natriurétique
périphérique, la consommation contmue GETOH modifie également l ' a c t ~ t é du syçtème
natriurétique au niveau du cerveau. En effet, l'administration d'alcool augmente les niveaw
ANP et CNP de l!hypothalamus. du pons et de la médulla des rats SHR Chez les rats WKY.

la consommation d'ETOH n'a pas d'effet sur les niveaux ANP du cerveau mais augmente les
niveaux CNP de lliypothalarnus et de la rnédulla. Enfm. I'aftinité des récepteurs NPR-C des
organes circurnventriculaires du cerveau . t'organe subfomical et le choroid pexus mais non
la région postréma. est diminuée à la suite d'une consommation continue d'ETOH.
Ces résultats démontrent que l'administration aigue et chronique de quantitées modérées
d'alcool modifie les composantes principales du système natriurétique. De plus. la direction
de ces altérations produites par I'ETOH air les peptides et récepteurs natnurétiques du coeur.
du rem et du cerveau indique qu'elles peuvent contniuer à l'effet antihypertenseur des doses
modérées et chroniques d'alcool.

TABLE OF CONTENTS
Preface
Acknowledgements
Publications
Original contributions to knowledge
List of figures
List of tables
List of abbreviations
CHAPTER 1: GENERAL INTRODUCTION
1.1. ALCOHOL (ETOH)
1.1.1. HISTORICAL PERSPECTIVES
1.1.2. PHARMACOLOGY
1.1.3. ABSORPTION AND DlSTRIBUTION
1.1.4. EXCRETION AND METABOLISM
1.1.3.1. ETOH to acetaldehyde
a) .-l lcohol dehirdrogertnse (ADHI
b) Mcrosoninl ETOH-oxrdizrtlg -'stem ~~/EOS)
cl Cntnlclse
d) Noti-oxidative p a t h v q s
1.1 -3 -2. Acetaldehyde to acetate
a) A ldehyde dehydroger iase (A L DH)
1.1.5. MAJOR BIOLOGICAL ACTIONS OF ETOH
1.1 S. 1. Effects of ETOH on the central nervous sysrem
n) y-amfiiobii^;ric ocid (GA BA) receptors
b) iV-niethyl-D-aspartate ( N A D A ) receptors

j Oproidr
1.1.5 2. Effects of ETOH on the endocrine system
a) rlrgir wie vmopressiri /A 1 F)
b) Fivpothalamic-pi tir itaty-adred a i s
C) Nypothulani ic-p i hr 2 t a - g o a i s
e) Catecholumitre
J Etidothelitrni- a~rd platelet-derived wsoactive
factors
gl Others
1.1.5 -3. Effects of ETOH on the kidney
1.1.5.4. Effects of ETOH on the h e r
1.1.5.5. Effects of ETOH on the cardiovascular system
1.2. ALCOHOL AND BLOOD PRESSüRE
1.2.1. ACUTE ETOH C O N S W T I O N
1 . L l . l . Human studies
a) ETOH-iridirced decrecrse iri BP
b) E TOH-irldziced irrcrecrse irr BP
C) NO effect of ETOH oti BP
d) Factors irrflzmrci~rg the restrlts observed itr
htrnrntl stttdies
r ) Szmtmq-
1.2.1.2. Animal studies
a) E TOH-irrdtrced decrease ir r BP
b) No effect of ETOH or1 BP
C) Factors itrflireticitrg the results observed irl
nrrintnl st~rdies
d) Szrnrniary
1.2.1.3. Mechanisms

6) C'mocorzstrictiorz
C ) E TOH-irdziced itlcreme i ~ z HR
1.2.2. CHRONIC ETOH CONSUMPTION
1.2.2.1. Hirman studies
a) H e q ETOH cotrszimptzoti
b) Light a d nioderate ETOH cotrszinptzoti
C) Factors itifliieticing the resiilts observed iti
epidemiologicai stiidies
d) Szrnrnq
1 -2.2.2. Animal studies
CI) ETOH-itidziced decrease in BP
b) E TOH-it i h ced itrcreme itr BP
c) No effecct of ETOH ut1 BP
d) Factors itifliierici~ig the residts observeci U r
atrinial stzïdies
r ) Szmirmn-
1.2.2.3. Meclianisms
a) .\/cchnn~cms rc.rponrib/c_t;w rhc h ~ p ~ r r c n r I ~ L ' -!/l'cf r!t'
chronrc hc*crr-* E7'0tI consrtmprlon
b) .\ lcchtrnnms r"sponr h i c f i w rhc unrrb pcrrenr rrc qfl2cr r!/ '
chrrmrc / i g h ~ crnJ moricrcrrc. I!TOH conrrtmprion
1.3. NATRIURETIC PEPTIDE FAMILY
1.3.1. A-TYPE NATRIURETIC PEPTIDES
1.3.1.1. Atrial natriuretic peptide
a) Historical perspectives
b) Biuchemisrp atid release
C) Tissue distribiitiori
4 Ilfetcrbolisni

1.3.1.2. Urodiatin
1.3.1.3. Kterminal fragments of proANP
1.3.2. B-TYPE NATRIURETIC PEPTIDE
1 -3.2.1. Brain natriuretic peptide
0) Biocliemistg arld release
b) Tisszte disrrib~rtior~
C) .bfernbolisnt
1.3.3. C-TYPE NATRlURETIC PEPTIDE
1.32.1. C-type natriuretic peptide
a) Biochenzistn md refeme
C) hfetabolisnt
1.3.4. NATRIURETIC PEPTIDE RECEPTORS
1.3.4.1. Guanylate-cyclase receptors
1.3.4.2. Clearance receptor
1.3-4.3- Receptors for N-terminal fia_gnents of proANP
1.3 .-W. Tissue distribution
1.3.5. PHYSIOLOGICAL ACTIONS OF THE NATRIURETIC
PEPTIDES
1.3.5.1. Renal actions
1.3.5.2. Cardiovascular actions
1.3-5.3. Adrenal actions
1.3.5.1. Central newous system actions
1.3.5.5. Hormonal actions
1.3.5 -6. Pulmonary actions
1.4. ALCOHOL, BLOOD PRESSURE AND THE NATIUURETIC
PEPTIDE F.4MtLY
1 -4.1. ETOH AND NATMURETIC PEPTIDE FAMILY

1.4.2. WOREUNG HYPOTHESIS AND DWISION OF THE
THESIS
CHAPTER 2: ACUTE ETOH STUDIES
2.1, INCREASED PLASMA ATRIAL NATRIURETIC PEWIDE
.M'TER ACUTE iNJECTION OF ALCOAOL IN RATS
2.1.1. ABSTRACT
2.1.2. rNTRODUCTION
2.1.3. PdATERiALS AND METHODS
2.1.3.1. Animal treatments
2.1.3.2. Blood alcohol content
2.1.3.3. Tissue extraction
2 - 1 3 -4. Plasma preparation for hormonal measurements
2.1.2-5. Cornparison between direct and extracted assays
2. 1.3 - 6 . Radioirnrnunoassay procedures
2.1 -3.7. StatisticaI analysis
2.1.4. RESULTS
2.1.4.1. Eqeriment 1
2.1.1.2. Experiment 2
2.1.4.3. Evperirnent 3
2.1.3.4. Correlations
2.1 S. DISCUSSION
2.1.6. ACKNOWLEDGEMENTS
2.2. INCREASED PLASMA ATRIAL NATRIURETIC PEPTiDE
M'TER INGESTION OF LOW DOSES OF ETOH IN HUMANS
2.2.1. ABSTRACT
2-2.2. INTRODUCTION
2.2.3. MATERIALS AND METHODS
2.2.3.1 Subjects

2.2.3.2. Experimental design
2.2.3 -3. Estimation of blood aIcohol content
2.2.3 -4. Estimation of plasma ANP. AVP and cortisol
2.2.3.5. Statistical analysis
2.2.4. RESULTS
2.2.5. DISCUSSION
2.2.6. ACKNOWLEDGMENTS
CHAPTER 3: CKRONIC ETOH STUDIES
3.1. EFFECT OF CIERONIC ETOEI CONSUMITION ON THE
ATRLAL NATRlURETIC SYSTEM OF S m RATS
3.1.1. ABSTRACT
3.1.2. INTRODUCTION
3.1.3. MATERIALS AND METHODS
3.1.3.1. Treatments
3.1.3.2. Estimation of bIood ETOH
3.1.3.3- heparation of blood and tissue extracts for RiAs
3.1.3.4. RiAs
3.1.3.5. RNA extraction and hybridization
3.1.3.6. RT-l'CR
3.1.3-7. Statistical analysis
3.1.4. RESULTS
3.1.4.1. Effect of ETOH on BP and HR
3.1.1.2. .Effect of ETOH on the heart ANP system
3.1.4.3- Effect of ETOH on circulating hormone levels
3.1.5. DISCUSSION
3.1.6. ACKNOWLEDGEMENTS
3.2. EFFECT OF CEtRONIC ETOH CONSUMPTION ON EIEART
BRAIN NATRWRETIC PEPTIDE

3.2.1. ABSTRACT 166
3 2.2. INTRODUCTION 166
3.2.3. MATERlALS AND METHODS 167
3.2.4. RESULTS 171
3.2.4.1. Effect of age and ETOH on body weight. BP. HR 17 1
and total protein content in atrial and ventricdar
tissues
3.2.1.2. Effect of age on BNP
3.2.4.3. Effect of ETOH on BNP
3 - 2 - 4 4 Ventricular BNP mRNA by RT-PCR
3.2.5. DISCUSSION
3.2.6. ACKNOWLEDGEMENTS
3.3. RENAL ALTEUTIONS OF ATRIAL NATRiURETIC
PEPTIDE RECEPTOUS BY CFIRONIC MODERATE ETOH
TREATMENT
3.3.1. ABSTRACT
3.3.2. INTRODUCTION
3.3.3. METHODS
3.3.3.1. Animal treatments 1 1 -
J .J .J -2. Urine collection
3.3.3.3. Urinary cGMP excretion
3.3.3.4. ANP RIA
3.3.3.5. Blood aIcohol content
3 Z3.6. Preparation of membranes for cornpetitive binding
assay
3-3-3.7. Cornpetitive binding assay
3.3.3.8. Autoradiographic studies

3 -3.3.9. Statistics
3.3.4. RESULTS
3.3.4.1. BP and plasma ANP levels
3 -3.4-2. Urine analysis
3.3.4.3. Competitive binding studies
3 3.4.4. Autoradiography
3.3.4.5. Urinary cGMP excretion
3.3.5. DISCUSSION
3.3.6. ACKNOWLEDGEMENTS
3 A .QLTERATIONS IN B W LEVELS OF ATRIAL AND C-TYPE
NATRIURETIC PEPTIDES AFTER CEiRONIC MODER4TE
ETOH CONSUMPTION IN SHR RATS
3-41. ABSTRACT
3.4.2. INTRODUCTION
3-4.3. MATERIALS AND METHODS
3.4.3.1. Animal treatments
3.4.3.3. Tissue extraction
3.4.3.3. RIAS
3.4.3.4. Statistical analysis
3-44. RESULTS
3.4.4.1. Effect of age
3.4.4.2. Effect of strain
3.4.4.3. Effect of ETOH
3.4.5. DISCUSSION
3-46. ACKNOWLEDGEMENTS
3.5. CIRCUMVENTRICULAR ORGAN NATRZURETIC PEPTIDE
RECEPTORS FOLLOWING CEiROMC MODERATE ETOH
CONSUMPTION IN SBR AND WKY RATS

3.5.1. ABSTRACT
3.5.2. INTRODUCTION
3.5 -3. MATERIALS AND METHODS
3.5.3.1. Animal treatments
3.5.3.2. Autoradiographic studies
3.5.3.3. Statistical analysis
3.5.4. RESULTS
3.5.4.1. Subfomical organ and choroid plexus
3.5.4.2. Area postrema
3.5.5. DISCUSSION
3.5.6. ACKNOWLEDGEMENTS
CELAPTER 4: GENERAL DISCUSSION
4.1. EFFECT OF ETOH ON TEE BLOOD PRESSURE
4.2. EFFEC'ï OF ETOH ON PLASMA AND =ART
NATRIURETIC PEPTIDES
4.2.1. PLASMA
4.2.2. HEART ATRiA
3.2.3. HEART VENTRICLES
4.3. EFFECT OF ETOH ON RENAL NATRllTRETIC RECEPTORS
4.4. EFFECT OF ETOH ON BRAIN NATIUURETIC PEPTIDES
AND RECEPTORS
4.5. GENERAL SUMMARY AND FUTURE STUDIES
4.5.1. KEART AND VASCULATURE
4.5.2. KIDNJSY
4.5.3. BRArN
APPENDTX
REFERENCES

PREFACE
The present thesis consihg of four chapters. describes the effects of acute and chronic
moderate ETOH consurnption on the major aspects of the natriuretic peptide system. n i e
ETOH-induced alterations are fùrther analyzed as pan of the mechanism by which moderate
ETOH treatment prevents the age-dependent increase in BP. Chapter 1 includes a
comprehensive review of the Iiterature on ETOH on the effect of ETOH on BP. on the
natriuretic peptide family and on the interactions between natnuretic peptides and ETOH.
Chapters 2 and 3 comprise scientSc papers published. accepted or submitted for publication
on the general alterations in the natriuretic peptide system produced by acute and chronic
ETOH drinking. respectively. in cornpliance with the guidelines for thesis preparation
provided by the Faculty of Graduate Studies and Research. the following te.xt is reproduced
below:
'Candidates have the option of including, as part of the thesis, the text of a paper(s) submitted
or to be submitted for publication, or the clearly-duplicated text of a published paper(s). These
texts must be bound as an integral part of the thesis.
If this opfion is chosen, connecimg texts that provide logical bridges between the different papers
are mandatory. The thesis must be written in such a way that it is more than a mere collection of
manuscripts; in other words, results of a series of papers must be integrated.
The thesis must sfill con fm to dl other requirements of the 'Guidelines for Thesis Preparation'.
The thesis must indude: A Table of contents, an abstract in English and French, an introduction
which clearly States the rationale and objectives of the study, a comprehensive review of the
literature, a final conclusion and summary, and a thorough bibliography or reference k t .
Additional material must be provided where appropriate (e.g. in appendices) and in sufficient
detail to dlow a dear and precise judgement to be made of the importance and originality of the
research reported in the thesis.
In the case of manuscripts CO-authored by the candidate and others, the candidate is required
to make an expliat statement in the thesis as to who contributed to such work and to what extent.
Supervisors must attest to the accuracy of such statements at the doctoral oral defense. Since

the task of the exam'ners is made more difficult in these cases, it is in the candidate's interest to
make perfectly dear the responsibiliti es of al1 the authors of the CO-authored papers. Under no
circumstances can a co-author of any component of such a thesis serve as an examiner for that
thesis."
Chapter 2. Acute ETOH experiments. describes the effects of a single administration
of a low and moderate ETOH dose on cardiac and circulating ANP levels in rats (Section
2.1. ) and human volunteers (Section 2.2. ). The demonstration of a stimulatory effect of acute
ETOH treatment on plasma ANP levels in both human and animal studies indicated that the
naniuretic peptide system was sensitive to ETOH exposure. Furthermore. the direction of the
ETOH-induced changes in plasma ANP levels suggested the possibility of a depressor effect
of this hormonal syaem in ETOH-treated ind~duals . Therefore. we sou& to analyze the
long-terrn modifications in the major components of the natriuretic peptide synem during
regular rnoderate ETOH drinking. in order to investigate the role of this hormonal family in
mediating moderate ETOH's antiliypertensive effect.
in Chapter 3. Chronic ETOH experiments. the long-term administration of moderate
ETOH levels was found io prevent the age-dependent increase in BP in both spontaneously
hypertensive ( SHR) and Wistar-Kyoto (W) rats (Section 3.1. ). This antihyp ertensive effect
of ETOH was associated with specific alterations in cardiac and circulating ANP and BNP
levels and in cardiac ANP and BNP mRNA levels (Sections 3.1. and 3.2.). The ETOH
treatment also modified the natriuretic peptide binding sites in the liidney. thus affecting botli
ends of the endocrine synem (Section 3.3.). Furthermore. the contribution of the brain
natriuretic system was evaiuated following chronic ETOH treatment via natnuretic peptide
measurements in the major brain areas (Section 3.4. ) and natriuretic peptide binding sites in
the circumventncular organs ( Section 3 . 5 . ).
Thus. the major parameters of the natriuretic peptide system were analyzed followüq
chronic moderate ETOH treatment. Interestingly, all of these parameters were specifically
altered by ETOH m a direction supporthg a general role for this hormonal system in ETOHs
antihypertensive effect.

Fmaily. Chapter 4 contains a detailed discussion of the various ETOH-induced aiterations
m the natrhiretic peptide syaem and of the mechanisms by which these alterations are Lilcely
to contribute to the antihypertensive effect o f moderate ETOH drinkmg. A descrÏption of
future directions for studies investigating ETOH and the natriuretic peptide f a d y is also
included.

1 am particularly indebted to Dr. Chriaina Gianoulakis for her patience and guidance
throughout the course of my audies. The iittle 1 know about scieutifk writmg and thinking
is certainly due in large parts to her exceptional generosity. For her t h e &en in countless
scientific and general discussions. for her days spent correcthg and improving my
manuscnpts. for her enthusiasm and Irindness. 1 am extremely grateful.
1 am also thankfÙl to Dr. Jolanta Gutkowska for her advices and excellent knowledge of
science. for her support and encouragements throughout my doctoral midies. Her sugeestions
and criticisms were greatly appreciated.
1 wish to thank past and present -dents in the labs for their fnendship and their help:
Jean-Pascal De Waele and Neil Jamenski m Douglas Hospital: Salima Menari. Eric Morin. and
especially Sheila Emest (Pour l'amitié et les sentils coups de téléphone. pour l'opéra aussi ...)
in the Hôtel-Dieu.
1 am also appreciative of the excellent technical eqertise and assistance of Céline
Coderre and Nathalie Charron (Merci beaucoup!!). of Dr. Suheyla Mukaddam-Daher. Dr.
Marek Jankowski. Dr. Tham-Vinh Dam Sylvie Larocque and Diane Beaudry .
1 wish to express my gratitude to Ricardo Claudio for his assistance and patience with
the animals. His fnendship is certainly the gea t ea accomplishment of these studies (Merci
a toi aussi Sylvie ... ).
Fmaily. I am especially grateful to my family: My rnother. Monique. my father. Roland.
and my brother. Emmanuel. for their constant and unrelenting encouragements and supports
(Sans vous tous. je n'y serais peut-ètre pas amvé..!).
1 should also thank - with a toast - these daily glasses of wine -never more than two - wbich ailowed me to survive stress and maintained rny blood pressure to a relatively normal
level.. .

PUBLICATIONS
Guillaume P, Gutkowska J, Gianoulakis C ( 1994) Increased plasma atrial natriuretic peptide d e r acute injection of alcohol m rats. J P h m a c o l E r - nter 27 1 ; 1656- 1665.
Gianoulakis C, Guillaume P. De Waele JP, Angelogianni P ( 1995) Effect of stress and alcohol on the proopiornelanocortin/ Bendorphin system bz: Stress, gender, and alcohol-seekmg behaviour, Research rnonograph 29, Hunt WA. Zakhari S (ed.), NIH NIAAA. Bethesda. p. 145- 165.
GuiIIaume P. Jankowski M, Gutkowska J, Gianoulakis C ( 1996) Effect of chronic moderate ethanol consumption on heart brah natriuretic peptide. Eur J Phamtacol 3 16: 49-58.
Guillaume P, Jankowski M, Gianoulakis C, Gutkowska J ( 1996) Effect of chronic etbanol consumption on the atnal natriuretic system of spontaneously hypertensive rats (SHR). AIcoholExp Clin Res 20; 1653-1661.
Guillaume P, Dam TV, Gianoulakis C, Gutkowska J ( 1997) Renal alterations of atrial natriuretic peptide receptors by chronic moderate ethanol treatment. Am J Physioi272 (Rerial Physiof 41); F107-F116.
Gianodakis C, Guillaume P, Thavundayd J, Gutkowska J ( 1997) Increased plasma atriai natriuretic peptide afkr mgestion of low doses of ethanol in human. AIcohol C h Exp Res (in press).
Guillaume P. Gutkowska J, Gianoulakis C ( 1997) Alterations in brain levels of atrial and C- type natriuretic peptides after chronic moderate ethanol consumption in spontaneously hypenensive rats. Ezir J Phannocol (m press).
Guiilaume P, Gianoulakis C, Gudcowska J ( 1997) Circmentricular organ natriuretic peptide receptors following chronic moderate ethanol consumption in SHR and WKY rats. (manuscript in preparation).
ABSTRACTS-COMMUNICATIONS
mP, Gianoulakis C (1992) Alcohol-mduced secretion of atrial natriuretic factor (ANP) m rats: Possible implication of 8-endorphin. 2~~idAnrzuo~ meetirig of the Sociev for Neurmcierzce, Anaheim, USA.
urne P, Gutkowska J, Gianoulakis C ( 1993) Plasma atrial natriuretic factor after acute injection of alcohol in rats. 2e Jounzée de la Recherche de l'Hôtel-Dieu de Montréal. Montréal, Canada.

Guillaume P, Gianoulakis C ( 1993 ) Effect of iti vivo ethanol administration on the HPA-ais and pituitary D-endorphin. Xth R S ' sczetitrfic cot,feretice. Washington. USA.
Guillaume P. Gutkowska J. Gianoulakis C (1993) Acute and chronic effect of moderate ethanol consumption on the atrial natriuretic syaem Sernorar, 3th Pqchiatry Research dqi: Dozlglas Hospital Research Centre, Verdun. Canada.
Guillaume P. Gutkowska J. Gianoulakis C ( 1994) Plasma atrial natriuretic factor following acute mjection of ethano 1 in rat S. MIth it~teniatzoiral cotigress of Pharniacologr.. Mont réal. Canada.
Guillaume P, Gutkowska J. Gianoulakis C ( 1994) increased plasma atrial natriuretic peptide after acute injection of alcohol in rats. Firsr prce. Sémitraire de Recherche. Hôtel-Dieu de hfot~tréal, Montréal, Canada.
Guillaume P, Gutkowska J. Gianoulakis C ( 1994) Acute and chronic effects of ethanol on the atrial natriuretic peptide system. Sentitinr. Dcnrglas Hospital Research Ceirtre. Verdun. Canada.
G u i l l a e , Jankowski M. Gutkowska J. Gianoulakis C ( 1994) Effect of chronic moderate ethanol consumption on the atnal natriuretic peptide syaem in SHR and WKY rats. 3e Jozirtlée de la Recherche de l'Hôtel-Dieu de Motitréal. Montréal, Canada.
Guillaume P. Jankowski M. Gianoulakis C. Gutkowska J ( 1995) Effect of chronic ethanol consurnption on the atriai natriuretic system of spontaneously hypertensive rats (SHR). 6r Coitgrès de in Sucies rli<ébécoise d'&perret~;ioti artérielle. Québec. Canada.
Guillaume P, Gutkowska J. Gianoulakis C ( 1995) Acute and chronic effects of ethanol consumption on blood pressure and ANP syaem in S H R .rith Scret1tzfic rneetitigfor the Itlter-Atnerrcnti socrey of khpertet~sioti. Montréal. Canada.
Guillaume P. Gianoulakis C. lankowslii M. Gutkowska J ( 1995 ) Acute and chronic effects of ethanol connimption on blood pressure and ANP system in spontaneously hypertensiie rat S. 7th Eirropeati nzeetitig or1 Hypertetisiori. Milan. Italy .
Guillaume P, Dam TV. Trernblay J. Gianoulakis C. Gutkowska J ( 1995) Renal alterations of ANP receptors by chronic ethanol treatment in SHR and WKY rats. First Ezrropem cotferetice or1 Pharmacoiogy, Milan. Italy.
Guillaume P. Jankowski M. Gutkowska J. Gianoulakis C ( 1995) Effect of acute and chronic ethanol treatment on the heart ANP and BNP system First Atunial Physioiogy Resenrch dq, McGilI University. Montréal. Canada.

Guillaume P, Dam TV. Tremblay J. Gianoulakis C. Gutkowska J ( 1995) Renal alterations of atrial nahuretic peptide receptors by chronic moderate ethanol treatment. -le Jmtniée de la Recherche de l'hrotel-Dieu de Montréal. Montréal. Canada.
Guillaume P. Gutkowslia J. Gianoulakis C ( 1996) Effect of chronic ethanol consumption on brain levels of ANP. Sem imr. Doziglas Hospital Research Ceritre. Verdun. Canada.
Guillaume P. Jankowski M. Gutkowska J. Gianoulakis C ( 1996) Effect of chronic ethanol consump tion on heart brain natriuretic peptide. Semimr, bui A wual Physiologv Research d q McGill University. Canada.
Gutkowska J. Guillaume P. Jankowski M. Gianoulakis C ( 1996) Effect of alcohol on blood pressure and atrial natriuretic peptide system in spoataneously hypertensive rats. 16th Scietifrjic nreetirig of the hrtenzatiorral Sotie+* of Hyertemiori. Glasgow. Scotland.
vii

ORIGINAL CONTRIBUTION TO KNOWLEDGE
L in order to mvestigate the role of the natriuretic peptide family m the prevention of the age-
dependent increase in the blood pressure (BP) by moderate ethanol (ETOH) consumption.
the effect of acute and chronic ETOH treatment on the major components of the natriuretic
peptide syaems and their receptors are inveaigated in the heart and circulation. kidney and
brain. The chronic administration of moderate levels of ETOH resulted in a Iower BP and HR
m the ETOKtreated rats when compared to the water-treated animals. These results confirm
and ergand previous reports and are nipponing the existence of a unknown antihypertensive
mechanism associated with the alcohol consumption.
CI. Acute ingestion of moderate ETOH levels in rats and humans resulted in increased plasma
ANP levels. In the rats. this mcrease is associated with a rapid decrease in atnal ANP content
and a delayed increase in ventncular ANP content. suggeaing the release of ANP fiom the
heart. These observations indicate that the heart natriuretic peptide system is stimulated by
the ETOH exposure.
[IL Chronic administration of moderate ETOH levels resulted in Iower plasma ANP and BNP
levels. Nevertheless. atrial and ventricular ANP and BNP mRNA activities are not reduced
by ETOH. despite the lower BP. Rather. increased atrial ANP and BNP contents and
increased ventncular BNP levels are observed in ETOH-treated rats. Moreover. the chronic
moderate ETOH treatment prevented the development of ventricular hypertrophy.
Considering the pattern of food and Buid intake during the iight and dark phases of the daily
cycle. these results indicate that the heart natnuretic system in the ETOH-treated rats is
mamtamed at an higher potential actMty by a dinerent rnechanism than in the water-treated
rats, suggeaing the possibility of a chronic stimulation of the heart oatnuretic -stem by
ETOH.
W . Chronic admirisration of moderate ETOH levels resulted in a reduction of the clearance

receptors (NPR-C ) m the renal glomeruli. Furthemore. chronic ETOH treatment mcreased
natriuretic peptide binding sites in the renal papiua. Excretory cGMP levels were either
elevated or unfhaaged foIlowing the ETOH administration despite the Iower BP. suggesting
a firnctional effect of the ETOH-induced natriuretic receptor alterations. These observations
suggest enhanced renal a c t ~ t y of the natriuretic system in ETOH-treated rats.
V. Cbronic administration of moderate ETOH levels resulted m increased or unchanged ANP
and CNP levels in the hypothalamus. pons and medulla. despite the lower BP. Moreover.
there was a reduction in the afEin.ity of NPR-C in the subfornical organ and choroid plexus.
but not in the area postrema following the alcohol treatment. These results suggen some
stimulations of the brain natriuretic peptides by the ETOH administration.
VI. In general. the direction of these cardiac. renal and brain alterations in the natriuretic
peptide system foster the hypothesis that the natnuretic peptides and receptors may mediate.
at least in part. the antihypenensive effect of moderate ETOH consumption.

LIST OF FIGURES
Chapter I
Figure 1.1.1. Schematic representation of gastric and hepatic alcohol metabolimi during acute and chronic ETOH consumption
F i e 3 1 Schematic representation of the transcription. translation and processing of the natriuretic peptide f a d y
Figure 1.3.2. Schematic representation of the structure of the natriuretic peptide farnily in rats
Figure 1 -3.3. Diagram of the natnuretic peptide receptors (NPR)
F e 1 3 4 Physiological actions of the natriuretic peptides
Figure 1 3 . 5 . Schematic diagram s u r n r n a ~ n g the ( 1 ) tubular. (2) hemodynamic. and ( 3 ) hormonal effects of the natnuretic peptide family on the kidney
Figure 1.3.6. Schematic diagram summarin'ng the effects o f the natriuretic peptide family on the heart and the vasculature
Figure 1.3.7. Schematic diagram m r i z i n g the effects of the natriuretic peptide family on the adrenal gland
Figure 1.3.8. Schematic diagram nimmarizuig the effiects of the natnuretic peptide famiIy on the brain
Chapter 2
Figure 2.1.1. Effect of the i.p. injection of morphine on the circulating ANP levels using either extracted (A) or unextracted (B) plasma for the RLA
Figure 2.1.2. Changes in blood alcohol content (BAC) with tirne after the i.p. injection of 1 or 2 g of ETOWkg B.W. (40% v/v solution)
Figure 2.1.3. Changes m plasma ANP (A). fi-endorphin (B) and corticosterone (C) content with t h e after the i.p. injection o f 1 or 2 g of ETOWlrg B.W. (JO0/0 v/v solution) or the equivalent amount of saline

Figure 2.1.4.
Figure 2.1.5.
Figure 2.1.6.
Figure 2.1.7.
Figure 2.2.1.
Figure 2.2.2.
Figure 2.2.3.
Figure 2.2.4.
Figure 2.2.5.
Figure 3.1.1.
Figure 3.1.2.
Figure 3.1.;.
Variations of ANP fiom basal levels in the plasrna (A). nght atrium (B). left atrium (C) and ventricles (D) after the i-p. injection of 2 g of ETOWkg B.W. (40% v/V solution) or the equivalent amount of saline
Correlation between the ETOH-induced changes in the plasma ANP and O-endorphin (i3-EP) contents
Correlations between the blood alcohol content (BAC) and the plasma ANP (A) and O-endorphin (B ) contents
Correlations between the increase in plasma ANP levels and the corresponding decrease in right atrial (A) and left atrial (B) ANP content at 1 5 min post-ETOH
Changes in blood alcohol content following ingestion of 0.25 and 0.50 g ETOH/kg B.W.
Hean rate foUowing (A) the placebo, (B) the 0.25 and (C) the 0.50 g ETOWkg B.W. drmks. Systolic and diastolic BP following (D) the placebo. (E) the 0.25 and (F) the 0.50 g ETOWkg B.W. drinks
Effect of(A) the placebo. (B) the 0.25 and (C) the 0.50 g ETOWkg B. W. drinks on the plasma ANP content
Effea of (A) the placebo. (B) the 0.25 and (C) the 0.50 g ETOHkg B.W. drinlis on the plasma content of vasopressin
Effect of(A) the placebo. ( B ) the 0.25 and (C) the 0.50 g ETOHkg B.W. drinks on the plasma cortisol content
Chapter 3
Progression of blood pressure dunng 8 months of water or 20% viv ETOH consurnption in WKY rats and SHR
Progression of heart rate during 8 months of water or 20°/o v/v ETOH consumption in WKY rats and SHR
Effect of ETOH treatment on the right atrial ANP system (content. concentration and mRNA) in WKY rats and SHR

Figure 3.1.4. Effect of ETOH treatment on the left atrial ANP system (content. 153 concentration and mRNA) m WKY rats and SHR
Figure 3.1.5.
Figure 3.1.6.
Figure 3.1.7.
Figure 3.2.1.
Figure 3.2.2.
Figure 3.2.3.
Figure 3.2.4.
Figure 3.2.5.
Figure 3.2.6.
Figure 3.2.7.
Figure 3.2.8.
Figure 3.3.1.
Effect of ETOH treatment on the ventncular ANP system (content. concentration and mRNA) in WKY rats and SHR
Autoradiograms of a representative Northem blot for atrial (A) and ventricular (B ) ANP mRNA m WKY and SHR rats after 8 months of water (H) or 20% vlv ETOH treatment (E)
Relative quantification of ANP mRNA in rat ventricles by RT-PCR
Circulating BNP levels in WKY rats and SHR at 7 weeks of age and afier 8 rnonths of water or 20% vlv ETOH consumption
Effect of age on nght atrial (A-B), lefi atnal ( G D ) and ventricular (E-F) heart BNP content and concentration
Effect of water or chronic ETOH treatment on the right atrial BNP system (total content (A). concentration (B) and mRNA (C)) in WKY rats and SHR
Effect of water or chronic ETOH treatment on the left atrial BNP system (total content (A), concentration (B) and mRNA (C)) in WKY rats and SKR
Autoradiogram of a representative Northem blot of atrial BNP mRNA in WKY and SHR rats after 8 months of water (H) or 20% vlv ETOH treatment (E)
Effect of water or chronic ETOH treatment on the ventricular BNP system (total content (A), concentration (B) and mRNA (C)) in WKY rats and SHR
Autoradiogram of a representative Northem blot of ventricular BNP mRNA in WKY and SHR rats after 8 months of water (H) or 20°h V/V ETOH treatment (E)
Ventncular BNP mRNA by RT-PCR in adult SHR and WKY rats
Urine volume and osmolanty in SHR and WKY rats d e r 7% months of ETOH or water treatment

Figure 3.3.2.
Figure 3.3.3.
Figure 5.3.5.
Figure 3.3-6.
Figure 3.41.
Figure 3.42.
Figure 3.43.
Figure 3.4.4.
Urine sodium concentration (A), sodium excretion (B). potassium concentration (C) and potassium excretion (D) m SHR and WKY rats afier 7% months of ETOH or water treatment
Cornpetitive inhiôition of '"EANP binding by increasing concentrations of unlabelled ANP and cANF to the glomemlar membranes in SHR and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afler 8 months of ETOH or water treatment )
Competitve inhibition of '"1-ANP binding by increasing concentrations of unlabeiied ANP to the papillary (CM) membranes in SHR and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afier 8 rnonths of ETOH or water treatment)
Autoradiographs of bmdmg of 50 pM '%ANP m the kidneys of adult SHR and WKY rats, afier 8 months of ETOH or water treatment
Quantification by densitometry of the displacement of total "'1-ANP bmding by 10" M unlabeiied cANF in the kidneys of adult SHR and WKY rats. after 8 months of ETOH or water treatment
Unnary excretion of cGMP (nM1day) measured from the urine collected during the iight phase of the daily cycle following 7 ' 2
months of water or ETOH treatment in SHR and WKY rats
Standard RIA curve of CNP
ANP content (ng) and concentration (@mg protein) in the hypothalamus (HYPO). pons and meduiia (MED) of 7 and 38 week- old SHR and WKY rats
CNP content (ng) and concentration (@mg protein) in the hypothalamus (HYPO). pons and medulla (MED) of 7 and 38 week- old SHR and WKY rats
ANP content (ng ) and concentration (ng/mg protein) after 8 months of water or ETOH (20% v/v) treatment m the hypothalamus (HYPO). pons and medulla (MED) of 7 and 38 week-old SHR and WKY rats

Figure 3 - 4 5
Figure 3.5.1.
Figure 3.5.2.
Figure 4.2.1.
Figure 4.2.2.
Figure 4.3.1.
Figure 4.3.2.
Figure 4.41.
Figure 4.4.2.
Figure 4.5.1.
CNP content (ng) and concentration (ng/mg protem) after 8 months of water or ETOH (20% v h ) treatment m the hypothalamus (HYPO). pons and medulla (MED) of 7 and 38 week-old SHR and WKY rats
Average cornpetition bbding curves of "1-ANP in adult (38 weeks- old) SHR and WKY rats subfomical organ (SFO). after 8 months of water o r ETOH (20% VI\;) treatment
Average cornpetition binding c w e s of ' 2 5 ~ - A N P in adult (38 weeks- old) SHR and WKY rats choroid plexus (CP). afier 8 months of water o r ETOH (20% v h ) treatment
Average cornpetition binding curves of "'EANP in adult (38 weeks- old) SHR and WKY rats area postrerna (AP). after 8 months of water or ETOH (20% v/v) treatment
Chapter 4
Schernatic representation of the natriuretic peptide syaem ( ANP and BNP) in the heart
Schematic representation of the possible ETOH-induced changes in circulating and heart natriuretic peptides during the Light and dark phases of the daily cycle
Schematic representation of the renal natriuretic system
Schematic representation of the ETOH-induced changes in renal natnuretic peptide receptors
Schematic representation of the natriuretic peptide system in the central nervous system (CNS)
Schematic representation of the ETOH-induced changes in brain natriuretic peptides and receptors
Schematic diagram s u m m a ~ n g the effects of chronic moderate ETOH consumption on the natriuretic peptide system and the mechanisms by which these modifications may mediate part of alcohol's antihypertensive effect

LIST OF TABLES
Chapter 1
Table 1.1.1
Table 1.1 .2.
Table 1.2.1.
Table 1.2.2.
Table 1.2.3.
Table 1.2.4.
Table 2.1.1.
Table 2.1.2.
Table 2.1.3.
Table 2.2.1.
Table 3.1.1.
Table 3.1 .2.
Table 3.2.1.
Table 3.2.2.
ETOKinduced alterations in the a c t ~ t y of the central nervous system (CNS)
ETOH-induced alterations in the activity of the endocrine system
Human studies on the acute effect of ETOH on the BP and the E-R
Animal studies on the acute effect of ETOH on the BP and the HR
Hurnan studies on the chronic effect of ETOH on the BP
Animal studies on the chronic eEect of ETOH on the BP
Chapter 2
Effeas of ETOH (2 &kg B.W.) on the plasma levels of B-endorphin. corticosterone. ACTH, aldosterone and AVP
Effects of ETOH (2 @hg B.W.) on blood pressure and heart rate
Correlations between the various parameters
Urine output and urine sodium and potassium excretion (rnean = SEM) following ingestion of O. 0.25 and 0.50 g ETOH/kg B.W.
Chapter 3
Blood alcohol, ANP. AVP. corticosterone and aldosterone in SHR and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afier 8 months of ETOH or water treatment)
Rotein levels in the cardiac compartments of WKY and SHR rats
Body weight and da* liquid consumption in WKY and SHR rats during the 8 months of treatment
Systolic blood pressure and heart rate in WKY and SHR rats during the 8 months of treatment

Table 3.2.3. Protein levels in the cardiac compartments of WKY and SHR rats 1 74
Table 3.3.1. Blood pressure (BP). body weight (BW). liquid connimption (LC) 198 and circulating ANP levels in SHR and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afier 8 months of ETOH or water treatment)
Table 3 -3.2. Kinetic parameters for glomerular natriuretic receptors in SHR and 202 WKY rats at age 7 weeks (before ETOH treatment) and at age 3s weeks (afier 8 months of ETOH or water treatment)
Table 3.3.3- Kinetic parameters for papillary (IM) oatnuretic receptors in SHR 706 and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afler 8 months of ETOH or water treatment)
Table 3.4.1. Blood pressure (BP), body weight (BW) and daily liquid connimption 227 (LC) m SHR and WKY rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (afier 8 months of ETOH or water treatment)
Table 3.5.1. Natnuretic receptor cliaracteristics (B,, and K.J in the SFO. CP and 21 1 AP afler 8 months ofwater or ETOH treatment in WKY and SHR rats

AD: angiotensin II
AC: adenylate cyclase
ACTH: adrenocorticotropic hormone
ADH: alcohol dehydrogenase
ALD: alcoholic liver disease
ALDH: aldehyde dehydrogenase
AMPA: a-amino-3-hydroxy- 5-rnethyl-4-isoxazole p roionic acid
ANOVA: analysis of variance
ANP: atrial natnuretic peptide
ANP,,.: long acting sodium stimulator
.&NP,,,,: vesse1 dilator
.&NP,,-,,: kahuretic stimulator
AP: area postrema
-4V3V: anteroventral third ventricie
AVP: arginine vasopressin
BAC: blood alcohol content
bFGF: basic fibrobast growth factor
BNP: brain natnuretic peptide
BP: blood pressure
B.W.: body weight
CAMP: 3'. 5' -cyclic adenosine monophosphate
cANF: des-[GlnHh. Seri". Gly"". Leu"'. Gly""]ANP,,2.,,,
cGMP: Y. 5' -cyclic guanosine moaophosphate
CCIF: congestive heart failure
CNP: C-type natriuretic peptide
CNS: central nervous system

CO: cardiac output
CP: choroid ple.nis
CSF: cerebrospmal fluid
EAA: excitatory aminoacid
EC F: extracellular fluid
EDRF: endothelium-derived relaxhg factor
ETOH: ethanol
GABA: y-arninobutync acid
GC: guanylate cyclase
GFR: glomedar filtration rate
G 1: gastrointestinal
HPA: hypothalamic-pituitary-adrenal
APG: hypothalamic-pituitary-gonadal
HPT: hypothalamic-pituitary-thyroid
i.c.v.: intracerebroventricular
IMC D: inner medullary collecting duct
i.p.: intrap entoneal
LHRH: luteinking hormone releasing hormone
mRNA: messenger ribonucleic acid
M EOS: microsomaI ethanol-o'cidizing system
NAD: nicotinamide-adenine dinucleotide
NMDA: N-methyl-D-aspartate
NO: nitric oxide
NPR: natriuretic peptide receptor
OVLT: organum vasculosum of the laminae terminalis
PGE,: prostaglandin EI
PNS: penpberal nervous system
POMC: proopiomelanocortin
RAAS: renin-angiot ensin-aldosterone system

RIA: radioimmunoassay
RT-PCR: reverse transcriptase polymerase chah reaction
SFO: subfornical organ
SHR: spontaneously hypertenske rats
SMC: smooth muscle celIs
T, : triiodothyronine
T,: thyroxine
TGF-II: transfonning growth factor-D
TNF-a : tumor necrosis factor- a
UD: urodilatin
VR: venous return
WKY: Wistar-Kyoto rats

CEIAPTER 1
GENERAL INTRODUCTION

1.1. ALCOHOL (ETOA)
1 .l. 1. RISTORICAL PERSPECTIVES
The discovery of alcohol dates back from the early steps of history. when coUected h i t s
and plants were lefl unattended in the sun. This natural fermentation of fi-uits and plants
produced a liquid (beer. wine) whose taste was pleasant and whose consumption promoted
leisure and conversation. It soon became part of the daily Life of early civilizations. in rituals.
m bonding. in politics. in medicine. Later. the Arabs discovered that they codd increase the
alcohol concentration of their liquids through alcohol distillation (liquor). intereaingly. the
word "alcohol' is also of Arabic origin. meaning "he ly divided spirit".
Yet. its detrimental effects when consumed in excess were also rapidly noticed since the
oldest code of laws. the code of Harnrnurabi of Babylooia. aiready regimented its
consumption. With the industrialization and the mass production of the XiXth century.
alcoliolic beverages became increasingly commercialized and the problems associated with
excessive drinking started to be considered seriously.
There is therefore a delicate balance between a positive. therapeutic or social use of
alcohol and a aegative. excessive or unhealthy alcohol consumption. As a consequence of this
dual action of alcohol. the cultures of the XXth century have oscillated between two
extremes. fiom the complete prohibition of alcohol m the United-States to the complete lasity
in alcohol consumption in France or Italy. A better approach. in my opinion. would be to
recognized both the beneficial effects of light alcohol drinking and the detnmental
consequences of heavy alcohol consumption.
1.1.2. PAARMACOLOGY
The chemical structure of alcohols consists of an hydroxyl group (OH) attached to a
chah of saturated carbons of vanous lengths. Although different types of alcohol e'cist. ethyl
alcohol or ethanol (ETOH) is the intoxïcating agent present in au alcoholic beverages. It is
a relatively simple molecule formed by a two carbon chah: CH,-CH,-OH. The molecular

composition of ETOH is both hydrophillic and hydrophobic. The polar hydroql group
renders ETOH misci'ble m all proportions with water. On the other hand. the non-polar CH,-
C H - portion makes ETOH soluble in lipid-dissoking fluids. Nevertheless. ETOH is poorly
soluble m Lipids themselves since its pamtion coefficient between the lipid and aqueous phases
is around 0.1 (molkg membrane)/(molll water). meaning that its concentration in tissue
lipids is always 10°/o of its concentration in body waters (Seeman et al.. 1972).
This characteristic of ETOH suggeas a rather weak dmg. Due to its strong preference
for the aqueous phase. the ETOH concentration in the blood must be high enough to force
ETOH into the ce11 membranes in order to achieve any effect on the various tissues of the
body (Goldstem. 1992). Paradoxically. this low potency of ETOH as a dmg is the main cause
of the ETOH-related health problems. associated with chronic heaw alcohol consumption.
n i e physical mtercalation of ETOH mto the Lipid membrane fits the Meyer-Overton concept.
which correlates the potency of an anesthetic with its iipid solubility (Meyer. 1937: Seeman
et al.. 1972: McCreely and Hunt. 19%). Non-saturable ETOH binding to ce11 membranes has
also been demonstrated. suggeaing the absence of specific receptors (Seeman et al.. 1972).
The nature of the ETOH interaction with the ce11 membrane is therefore considered similar
to that of a weak anesthetic. ETOH dissohies physically into the phospholipid membrane to
produce expansion and disordering. associated with elevated fluidity (Seeman et al.. 1974:
Chm and Goldstem. 1977; Taraschi and Rubin. 1985; Hunt. 1993 ). Most of the p hysiological
actions oT ETOH result from these non-specific interactions and from the secondary
modifications m ionic channels (Messing et al.. 1986). honnonal/neurotransminer systems and
their receptors ( V a k e ~ s et al.. 1989: White et al.. 1990). G-proteins and second messengers
(Stenstrom et al.. 1986: Diamond et al.. 1987: Chamess et al., 1988). and gene evpression
characteristics (Wilke et ai., 1994).
The blood alcohol content (BAC) depends on the dose administered. the route of alcohol
into the vascular system and the rate of its metabolism and excretion.
1.1.3. ABSORPTION AND DISTRIBUTION
The major route by whicli ETOH reaches the circulation is through the gastrointestinal

(CI) tract. In rare occasions ETOH may also be absorbed through pulmonary tissues (Leaer
and Greenberg. 195 1). The skin however is a structure quite impermeable to allow ETOH
a s i o n (Bowers er al., 1942).
Within the GI tract. ETOH is absorbed by passive diffusion from the small intestine and.
to a d e r extent fiom the stomch (Li 1980: Brick et al.. 1988). Several factors may affect
the rate of absorption. The diffusion of ETOH into the blood is directly related to its
concentration. Stornach emptying tirne is also important because of the high proportion of
ETOH abçorbed fiom mteainal tissues. The passive difision of ETOH fiom the GI tract to
the vaxulature is also regulated by the diffusion gradient. modified by the blood flow or the
BAC (Berggren and Goldberg. 1940). Another major factor is the presence of food in the
stomach known to delay both ETOH diffusion and stomach emptying (Melanby er a/.. 19 19:
Kalant et al.. 197 1). It is of importanc.z to note that longer ETOH transit tirnes in the nomach
and slower absorption rates are associated with lower final BAC values. because of the
significant contribution of the gastric and hepatic fira-pass ETOH metabolism (see section
1.1.4. ) (Goldstein, 1992).
Once into the cuculation. ETOH is distributed throughout the aqueous compartments
of the body. Because of their rich blood supply. the brain. liver. kidneys and lungs equilibrate
faster with BAC than less vascularized tissues (Pohorecky and Brick. 1988).
1.1 .4. EXCRETION .AND METABOLISM
A d l fiaction of mgested ETOH is eliminated mtact through the kidneys. the lungs and
the skin. The rea is metabolized. first to acetaldehyde and then to acetate.
1.1.4.1. ET0 H to acetaldehvde
al d kohol dehydrogetiase
CH,CH,OH + NAD' =s CH,CHO + NADH + W tETOI1) ( ci~litrildCh'.dL')
The major enzymatic pathway for the oxidation of ETOH to acetaldehyde involves the
enzyme alcohol dehydrogenase (ADH). ADH is a cytosolic zinc-containing enzyme which
requires nicotinamide-adenine dinucleotide (NAD) as a coenzyme to activate the reaction

(Von Wartburg et al.. 1964; Ehrig et al.. 1990). ADH is expressed m the liver and on the
gastnc walls (Smith et al., 1972; Ehrig et al., 1990) and is characterized by its Iow Michaelis
constants a, which suggest an actMty even with very low ETOH concentrations (Lieber,
1994).
Even though the ADH m o l d e s mediate most of ETOH oxidation, the rat e-limiting nep
m the reaction is usu* the regaieration of NAD' f?om NADH During alcohol mtoxication.
the hepatocytes use the majority of their available dehydrogenases to reoxidize NADY often
m spite of theh mability to maintah redox homeostasis. This results in the altered metabolism
of various other compounds, such as Eits, steroids and carbohydrates (Lundquist er ai-. 1962:
Forsander, 1966).
During acute alcohol consumption, ETOH is metabolized by gastric and hepatic ADH
(figure 1.1.1. ). FM-pas metabolisn reduces the bioavailability of ETON both directly fiorn
the stomach and during its first passage through the liver (JuIhunen et al.. 1985; Lim er al..
1993). During chronic heavy alcohol consumption or in alcoholics, the gaaric first-pass
metabolism is reduced due to the decreased activity of the gaanc ADH (Di Padova et al..
1987). in contrast to gastric ADH, the a c t ~ t y of the h e r ADH system is not sigdicantly
altered by chronic ETOH consumption, until the appearance of serious h e r disease.
6) Micrmonrai etha~rol-oxidizi~rg system MEOS)
CH,CH,OH + NADPH + H* + 0, =, CH,CHO + NADP+ + 2H,O (ETOH) (acddebyde)
A second enzymatic pathway for the oxidation of ETOH, independent of ADH, has been
identified in liver microsornes (Lieber and De Carli, 1968, 1970). It involves a family of
ETOH-inducible P-450 enzymes (P-450 2E 1 is the major one induced by ETOH) and the
presence of oxygen and NADPH (Ohnishi and Lieber, 1977). The Y, of MEOS for ETOH
is about one order of magnitude higher than the Y, of ADH, meaning that at low ETOH
concentrations the hepatic ADH pathway is responsible for most of ETOH metabolism (figure
1.1.1 .). However, during ETOH intoxication, a significant portion of ETOH may be
catabolized by MEOS.
Similarly, t6e long-term use of alcohol in both experimental animais and alcoholics bas
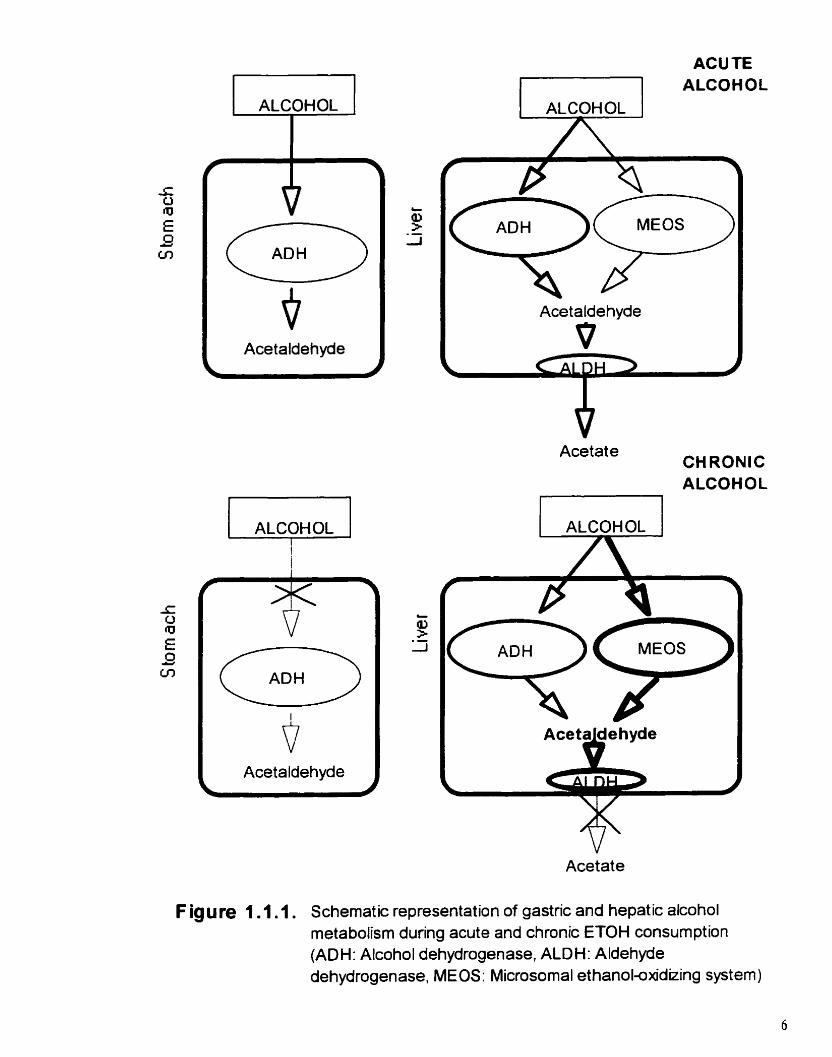
ALCOHOL '7
Acetaldehyde
Acetaldehyde
ACUTE 7 1 ALCOHOL 1 ALCOHOL 1
Acetaldehyde
Acetate CH RONlC ALCOHOL
ALCOHOL ,
f
Aceta dehyde + ?
Acetate
F ig u fe 1 .1.1. Schematic representation of gastric and hepatic alcohol metabolism during acute and chronic ETOH consumption (ADH: Alcohol dehydrogenase, ALDH: Aldehyde dehydrogenase, MEOS: Microsomal ethanoi~xid~ing system)

been associated with a sigdicant induction of P-450 2E 1 and with elevated levels of P-150
2E 1 transnipts (Tnitsumi et a/. . 1989: Diehl et ai.. 199 1 a: Takahashi et ai.. 1 993 ). Therefore.
during chronic ETOH connimptioa. a subaantial portion of ETOH metabolism is achieved
through the MEOS (figure 1.1.1. ).
CH3CH20H + H20Z * CHICHO + 2Hr0 (ErOEI) c r i ~ ~ & h ' . & )
A third minor enzymatic pathway has been described for the oxidation of ETOH. Hepatic
catalase has been s h o w to metabolize ETOH iti vitro in the presence of hydrogen peroxide
(H201) (Keilin and Hartree. 1945). However. this pathway is unlikely to contniute much to
ETOH metabolism since the rate-lirniting step is the formation of HZ02 . hdeed. in the liver
HZOZ is generated m significantly lower levels than expected based on the content of hepatic
catalase (Bovens et al.. 1972). Furthemore. indirect estimations of the Y, for ETOH by the
catalase reaction has reponed high values. m the same range as those reported for the MEOS
(Oshino et al.. 1 973 ).
d) iVor1-oxïdative patlnvqs
The production of fatty acid ethyl esters fiom ETOH by the action of the enzyme fatty
acid ethyl ester synthase has been reported (Mogelson and Lange. 1984). This non-oxidative
metabolism of ETOH is present in ogans lacking the traditional ox ida t~e ETOH pathways.
such as the brain. the pancreas and the heart. and has been implicated in alcohol-related
mjuries (Laposata and Lange. 1986: Bora and Lange. 1993 ). Interestingiy. the actkity of the
brain fatty acid ethyl ester synthase is increased in alcoholics (Laposata et ni.. 1987).
1.1.4.2 Acetaldehvde to acetate
CH,CHO + NAD+ + H,O =s CH3COOH + NADH + W ( ~~xtdd&> dC) ( ~ L X ~ L . )
The major consequence of the oxidation of ETOH to acetaldehyde is to convert a rather

weak drug to a toxic one. Acetaldehyde has been s h o w to form protein adducts with liver
microsomal proteins such as collagen (Baraona et ai.. 1993) and with cuculating protems
such as hemogiobin (Stevens er ai.. 198 1 ) and Lipoproteins (Wehr et al.. 1993). resulting in
antibody production. alterations in protein release and modifications in enzymatic activities
(Hoemer et al.. 1986: Solomon 1987: Tuma et al.. 1990). Acetaldehyde has also been s h o w
to promote lipid peroxidation and f?ee radical-mediated to'ricity (Müller and Sies. 1982).
Furthemore. acetaldehyde accumulation is associated with autonomic and cardiovascular
effects such as the "flush syndrome". hyperventilation and nausea. Therefore. under normal
conditioos. the rapid oxidation of acetaldehyde to acetate by hepatic ALDK in the presence
of NAD'. maintains acetaldehyde levels in very low concentrations (figure 1.1.1. ).
Long-term ETOH consumption renilts in the significant reduction of ALDH actMty
(figure 1.1.1. ) (Hasumura er ai.. 1975). This lower capacity of acetaldehyde oxidation.
associated with the enhanced production of acetaldehyde fiom MEOS. leads to bcreased
circulatmg and t h e acetaldehyde levels and the possibility of chronic toxicity (Pikkarainen
er nl., 198 1 : Nomura and Lieber. 198 1 ).
t .1.5. iMAJOR BIOLOGICAL ACTIONS OF ETOH
The incorporation of alcohol into the cell membrane of the cells of vanous tissues
produces a cohon of physiological effects. Among the physiological systems greatly affected
by alcohol are: the central nervous system (CNS). the endocrine system the kidney. the liver
and the cardiovascular qaem The majority of the behavioral effects of ETOH are rnediated
through ETOH-induced alterations in the a c t ~ t y of the CNS. Various components of the
cardiovascular system nich as the contractile properties of the heart or the blood pressure.
are also xnodulated by alcohol consumption. Because of its importance in ETOH metabolisrn
the h e r is greatly affected by ETOH. especially during long-tem consumption. The diuretic
effects of alcohol are mediated in pan through the kidney. Finally. a ournber of hormooal
syaerns. such as arginine vasopressin (AVP), prolactin. testosterone or the hypothalamic-
pituitary-adrenal (HPA) a i s , are also rnodified by the presence of ETOH.

1.1.5.1. Effects of ETOA on the central nervous svstem lCNSl
ETOH is classified as a general neurodepressant dmg. Nevertheless. low levels of alcohol
have been found to produce excitation. suggesting a biphasic effect (Hunt. 1993). At low
BAC (below 50 mgdl). the cognitive h c t i o n s of the cerebral cortex are mostly afTected.
resuitmg in reduced mbibition. decreased tension and general euphona. Higher BAC (between
50 and 150 m m ) impair motor coordination of cerebellar ori_gin. whereas BACS above 200
mg/dl greatly depress consciousneçs. respiration and cardiovascular regulations. With chronic
heavy ETOH consumption. tolerance to these effects may appear. Furthemore. when long-
tenn ETOH drinking is abruptly aopped. a state of general hyperexcitability called "alcohol
withdrawal syndrome" may occur indicating addiction and physical dependence and
characterized by convulsions. seinire. tremulousness and delirium tremens. These various
effects of alcohol are mediated by ETOH-induced aiterations in the actMty of a number of
neurotranmitter systems such a s y-amhobutyric acid (GABA). glutamate or opioids. A brief
description of these alterations and their physiological si_guificance is outlined (table 1 . 1 . 1 . ).
a) y -aminobzc~*rrc acrd (GA BA) receptors
GABA is the major Uihibitory neurotransrnitter in the brain. Its actions are mediated via
two classes of receptors. GABA, and GABA,. GABA, is responsible for mon of the
inhibitory effect of GABA through the activation of the chloride channel associated with the
receptor, leadmg to an ~ U Y of Cl- ions which hyperpolarizes the affected neuronal cells and
therefore decreases the e'rcitability of the neurons (Olsen and Tobin. 1990).
Acute ETOH consumption has been impiicated with the potentiation of GABA, activity
and with the facilitation ofthe association between GABA and its major receptor (Suzdac et
al., 1986: Deitrich et a/. , 1989; Leidenheimer and Hams. 1992; Hunt, 1993). The resulting
neuronal hyperpolarization and lower firing rate in those tissues rnay evlain in part the
intoxicating and depressant eEects associated with high BAC.
In contrast. long-term ETOH exposure is associated with a reduction of the ETOH-
mediated enhancement of GABA, actMty. suggesting tolerance to its acute effect ( M a n and
Harris, 1987: Morrow er al. 1988). Aiterations in the gene expression of GABA, have been
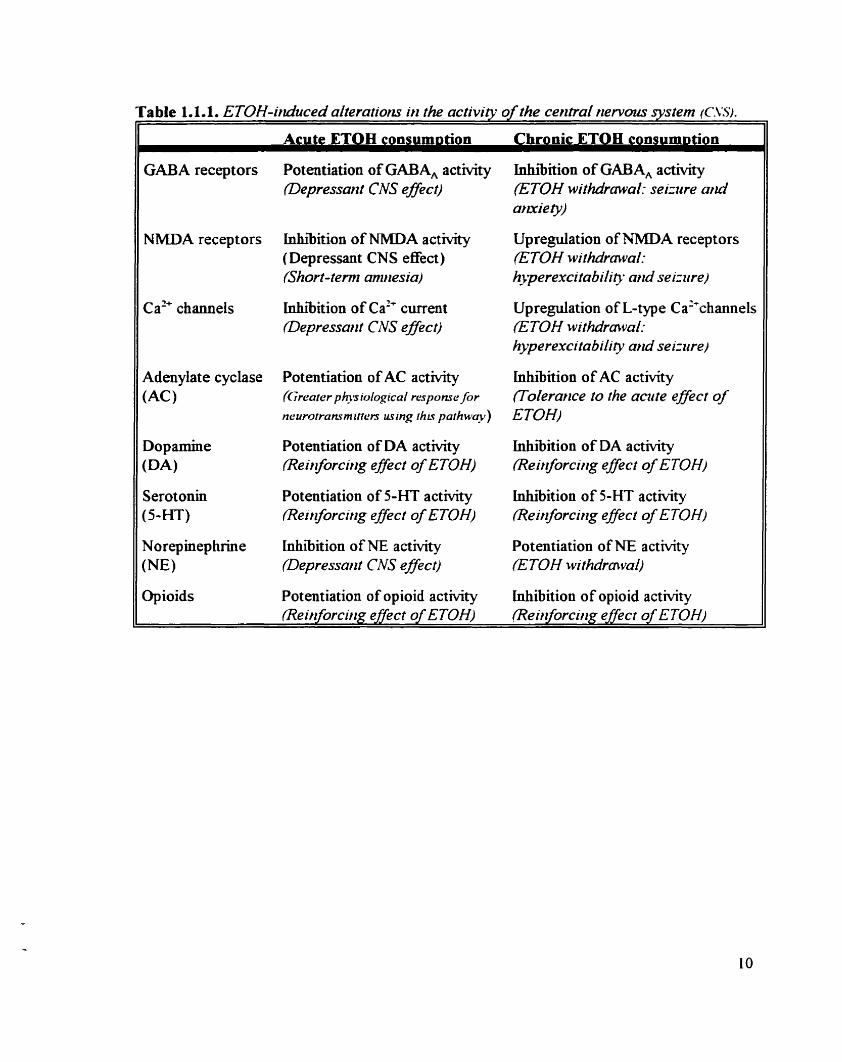
Table 1.1.1. ETOH-irrduced alteratiorzs irr the actzvitv o f the central rrervous systern (C-ï.%
Acute ETOE consum~tion Chronic ETOB coosum~tion
GABA receptors
NMDA receptors
Ca" channels
Adenylate cyclase (AC)
Opioids
Potentiation of GABA, activity (Depressant CNS effec)
lnhr'bition of NMDA activity (Depressant CNS effect) {Short-term amrresia)
inhibition of Ca" curent Oepressarrt C M e ffec t)
Potentiation of AC activity {Grearer p b s iological response fir
neurorransmirrers using rhrs parhway)
Potentiation of DA a c t ~ t y lReirrfocirig effect of ETOH)
Potentiation of 5-HT actMty Reirforcirzg eflect of ETOH)
inhibition of NE a c t ~ t y (Depressarit CNS effect)
Potentiation of opioid activity
Inhibition of GABA, actMty ETOH withdrawal: seizzue arui arzxiev)
Upregulation of NMDA receptors (ETOH withdrmval: h3perexcitability a d seizztre)
Upregulation of L-type Ca2'channels ETOH withdrmvai: hyperexcitabiiity arui seizztre)
Inhibition of AC activÏty (Tolermce to the amte eflecr of ETOH)
lnhiibition of DA actMty meit florcirzg e ffect of E TOH)
inhibition of 5-HT a c t ~ t y (Reirforcirrg effecr of ETOH)
Potentiation of NE activity ETOH rvithdmvai)
Luhibition of opioid activity

postuiated (Morrow et al.. 199 1 ). To sorne extent, these chronic m o ~ c a t i o n s m GABAergic
actMty may provide an explanation for the seizures and anxiety observed during ETOH
withdrawal (Hunt. 1993 ).
b) iV-nie thvl- D-asparta t e N34DA) receptors
Glutamate is one of the major excitatory neurotransmitters in the brain and has been
show to interact maHily with 3 types of receptors: NMDA kainate and a-amino-3-hydroxy-
5-methyl-4-isoxazole proionic acid (AMPA). Like GABA,. these receptors are ionotropic
since the): are coupled to an ion channel permeable to various cations. Kainate and AMPA
receptors are believed to mediate fast neuronal excitation through Na'. whereas the NMDA
receptor is mvoived m slower excitatory responses through Ca". NMDA receptor actMty and
inward calcium flunes have been implicated with newotransminer release. learning and
memory (Collingridge and Leiter. 1989: Cotman et al.. 1989).
Acute intoxicating levels of ETOH are reported to depress NMDA receptor functions
and to mhibit NMDA-activated Ca2+ currents (Lovinger et al.. 1989: H o b et al.. 1989:
White et al.. 1990). The iower intracellular Ca2+ IeveIs decrease neuronal activity and may
account for the sedative effect of alcohol. Similarly. the ETOKmediated inhibition of NMDA
activity in hippocampal cells may explain the "short-term amnesia" observed ofien afier
excessive drinking (Diamond and Messing. 1994). In contrast. lower levels of ETOH have
been associated with enhanced NMDA receptor functions. suggesting a biphasic effect of
alcohol ( Lima-Landman and Albuquerque. 1989).
Long-term ETOH treatment induces the upregulation of NMDA receptors. in order to
cornpensate for the chronic reduction of CaL' infiuxes by the continuous presence of alcohol
(Michaelis et al.. 1978. I W O : lorio et al.. 1992). This overexpression of NMDA receptors
produces a state of excessive glutamate and excitatory amino acids (EAA) activation.
conmbutsig to the hyperexcitabdity and seipire production during alcohol withdrawal (Grant
et al., 1990: Nutt and Peters, 1994).

cl C bllage-dependent calcitin~ (Ca'-) charnels
ETOH does not only disrupt receptor-mediated ion channels but also voltage-gated ion
channels. 'The L-type ~ a " channel is particularly sensitive to ETOH. Much &e for NMDA
receptor functions acute ETOH administration mhiits Ca2+ current through this channel
(Leslie et a/.. 1983: Treistman el ni.. 199 1). resulting in reduced byaptic release of
neurotransmitters. whereas adaptive compensations occur with chronic alcohol consumption.
producing the upregulation of L-type Ca" channels. This upregulation of L-type Ca'+
channels contniutes to a number of ETOHs withdrawal syndromes, such as seizures and
overexcitability (Messing et al.. 1 986: Little. 1 99 1 ).
d) S e c o d nzesserrger gstenrr (Adeqvlate cydase)
The incorporation of ETOH into neuronal ceil membranes has been implicated with
alterations of the adenosine 3'. 5'-cyclic monophosphate (CAMP) second messenger system
(Hofban and TabakoE 1990). Acute ETOH exposure stimulates adenylate cyclase (AC) Ma
the enhanced activity of stimulatory G-proteins. resulting in greater physiological responses
to neurotransmitters using this pathway. such as monoamines (Rabin and Molinoff. 198 1 :
Leithm and TabakofE 1984). In contraa. chronic ETOH consumption appears to reduce the
efficiency of the CAMP system and therefore of the translation between the neurotranmitter
signal and the biological effect. probably via heterologous desensitization (Israel et al.. 1972:
Gordon er al.. 1986). This is an example of tolerance to ETOH following its prolonged
exposure.
e) h./otrmtirres
n i e major monoamine neurotransmitters are dopamine. serotonin and norepinephnne.
Acute ETOH consumption is associated with elevated dopamine levels in the nucleus
accumbens, probably because of the ETOH-mediated mcreased activity of serotonin receptors
(Di Chiara and Imperato. 1988: Lovinger. 199 1 ). In contrast. lower brain levels of dopamine
and serotonin have been obsewed following chronic ETOH conaimption, suggesting adaptive
compensations (Wu et al.. 1986: Wang et al.. 1993). The implication of serotonin and

dopamine in the reinforcing effect of alcohol is suggeaed by lower brain concentrations of
both monoamines and by elevated serotonin receptor numbers in ETOH-prefening arains of
rats. predisposing these animals to alcohol drinking (Murphy et al.. 1987: McBride et al..
IWO: Hunt. 1993).
Acute ETOH consumption is associated with lower brain levels of norepmephrine (Alan
et a/.. 1987: Patel and Pohorecky. 1989). During chronic ETOH drlliking. norepinephnne
release is increased in the brain. possibly because of the dom-regdation of the inhibitory
presynaptic a*-adrenoceptors located on norepinephric neurons (Nutt et al.. 1988: Wang et
O!.. 1993). This overac t~ ty of the CNS norepinephnne system has been implicated with the
alcohol withdrawal syndrome (Hawley et al.. 1985 ).
Opioidî
Alcohol preference may also depend on the a c t ~ t y of the opioid system (Gianoulaliis.
1993). Various opioid receptor antagonids have been reported to reduce voluntary ETOH
htake (Hyytia et al.. 1993 ). Moreover. although acute ETOH consumption increases plasma
and CSF Bendorphin leveis (Borg et al-. 1982: Gianoulakis and Barcomb. 1987: Gianoulakis
et ai.. 1996). chronic ETOH exposure, such as in alcohol addicts, is associated with reduced
central 0-endorphin levels. suggesting a desensitization of the opioid syaem and the
possibility of opioid-dependent ETOH addiction and dependence (Gienazzani et al.. 1982:
Hutchison et d.. (988).
Although most short-terrn and moderate ETOH consumptions produce only reversible
changes m neurophysiological functions. prolonged heavy ETOH drinlimg has been implicated
with various more permanent brain Iesions. such as Wernicke's encephalopathy and
Korsakoff s psychosis (Tuk. 1 992: Chames. 1993 ). The former is characterized by confiision.
ataxia and oculornotor abnonnalities. The latter is a sequelae of Wemicke's encephalopathy
and consists of short-term memory anmesia. n i e exact role of alcohol in the etiolow of those
diseases is unclear. However. chronic ETOH consumption-associated malnutrition and
especiaiiy thiamine deficiency have been positively h k e d with the development of these

neurological disorders (Blass et ai.. 1977: Victor er al. . 1989).
1 . l S . Z . Effects of ETOH on the endocrine system
A number of ETOH-mediated manifestations such as diwesis, tolerance. withdrawal and
even liver abnormalities. have been associated with specific ETOH-induced alterations in the
a c t ~ t y of some endocrine hormonal systems (table 1.1.2.).
a) ArgirNrie vasopressir~ (A 17')
Acute ETOH consumption induces diuresis (Murray. 1932). This effect has been
attnbuted. at leaa in part. to the ETOH-mediated inhibition of AVP release from the
posterior pituitary gland (Eisenhofer and Johnson. 1982: Leppaluoto er ai.. 1992). More
specificaUy. AVP suppression by ETOH appears to be limited to the ascending dope of BAC.
whereas an increase in plasma AVP content is observed during the descending BAC levels
(Linkola et al.. 1978: Eisenhofer and Johnson. 1982). This biphasic AVP response may be
due to a secondary increase in osmolarity. suppressing the direct ETOH inhibition of AVP.
In contrast. stimulations of AVP release have been observed with chronic ETOH
connunption to cornpensate for the continuous presence of alcohol. redting in water
retention during alcohol withdrawal (Eisenhofer et ni.. 198% Emsley et al.. 1987: Taivainen
er al.. 1995). interestingly. many peptides of the brain. including AVP. are important for the
acquisition and retention of certain learned behaviours and for the etiology of convulsive
disorders (Kasting et al. . 1980: Cicero. 198 1 ). The dernonaration that both the tolerance to
the hypothermie effects of alcohol and the duration of the ETOH withdrawal seinires are
prolonged in ACT-treated rats suggests that elevated AVP levels may also participate in the
developrnent of tolerance and physical dependence (HotEnau et al.. 1978: Rigter et al. . 1 980).
b) Hypothaianric-piti~iraq-adred IHPA) ais
Acute ETOH drinking increases circulahg cortisol (humans) / corticosterone (rodents)
levels (Ellis et al.. 1966: Ienkins and Connelly. 1968). This elevatioo appears to be under
hypothalamic and pituitaiy controL smce plasma ACTH levels are also increased after ETOH
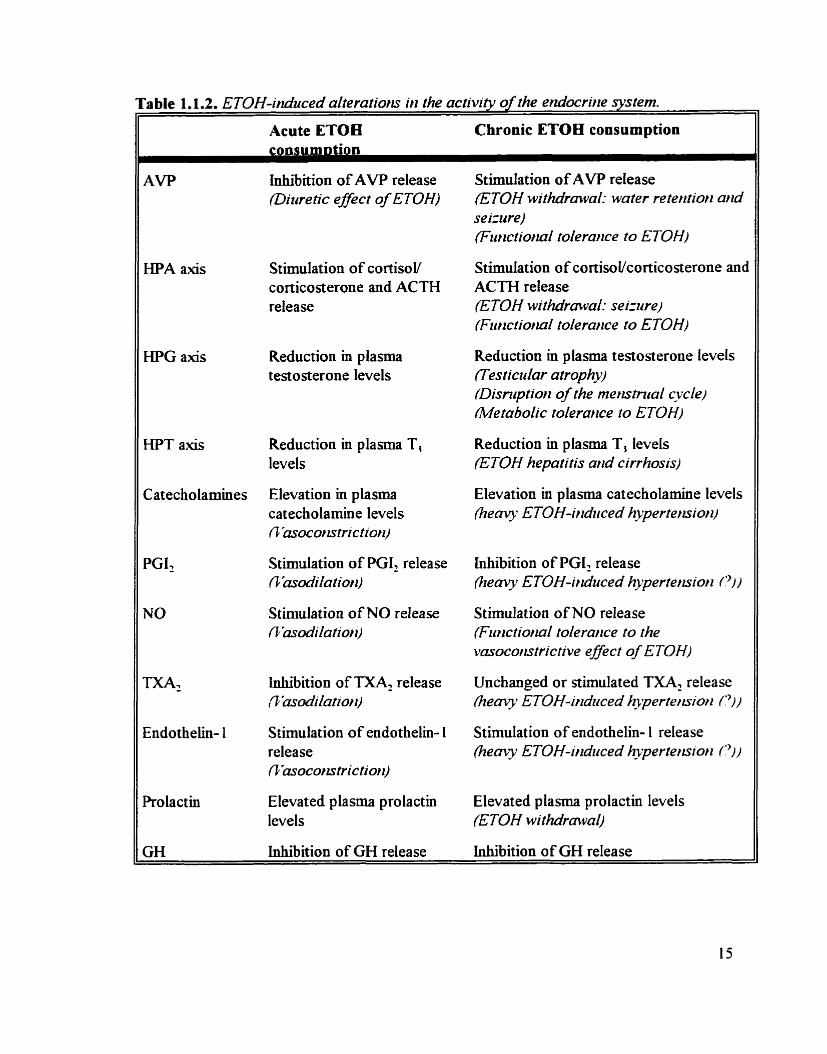
Acute ETOB Chronic ETOH consumption
AVP
I-IPA axis
HPG a i s
HPT a i s
Catecholamines
ml
Endothelin- 1
Prolactin
GH
inhibition of AVP release (Ditiretic effect of ETOH)
Stimulation of cortisol/ corticosterone and ACTH release
Reduction m plasma testosterone levels
Reduction in plasma T, levels
Elevation in plasma cat ectiolamine levels 0 'asocorzstrictiorl)
Stimulation of PGI, release f i 'mdilatiorr)
Stimulation o f NO release flrasodilutiorl)
lnhiition of TXAl release ,~'asodilutiott)
Stimulation of endothelin- 1 release flhsocortstrictio?l)
Elevated plasma prolactin levels
Inhidition of GH release
Stimulation of AVP release (ETOH withdrawal: wuter reterrtiorl arld se izure) (Fzir~ctiorral toferance to ETOH)
Stimulation of cortisoUcorticosterone and ACTH release ETOH tvithdrawal: seiztrre) (Ftrrzctiorral tolerurlce to ETOH)
Reduction m plasma testost erone tevels flesrictdar arrophy) (Disruptiorr of the rner~striral cycle) Metabofic toferailce to E TOH)
Reduction in plasma T, levels (ETOH hepatitis and cirrhosis)
Elevation in plasma catecholamine levels (heavy E TOH-iridiiced hypertemiort)
inhibition of PGI, release fieavy ETOH-iruiuced hyperterzsiorz P))
Stimulation of NO release (Fwlctiorraf tolerurtce to the vasocorntrictive effect of ETOH)
Unchanged or stimulated TXA: release (heavy E TOH- irzdziced hijoertemioti PI/
Stimulation of endothelin- 1 release k e m y ETOH-induced hyperterrsiori PI)
Elevated plasma prolactin levels (ETOH withdrawal)

consumption (Rivier et al.. 1984: Redei et al.. 1986).
Excessive long-term use of ETOH is also associated with high plasma levels of cortisol
(humans) / corticosterone (rodents). leadmg to various clinical features descnied as "pseudo-
Cushing's syndrome" (Frajria and Angeli. 1977: Van ThieL 1983). hcreased
CoibsoVcorticosterone levels may also be related to the development of phytical dependence
to ETOH. since adrenalectomy markedly decreased the severity of the alcohol withdrawaI
seinires m rats (Sze et al.. 1974). The HPA a i s may also participate in functional tolerance
to ETOH since the administration of corticosterone antagonists attenuated the development
of the tolerance to the hypothermie effect of chronic ETOH drinking in rats (Sze. 1977:
Tabakoff and Yanai. 1979).
Both acute and chronic ETOH administration reduce circulating testosterone levels
(Gordon et al.. 1976; Cicero and Badger. 1977: Lester and Van =el. 1977). Elevated
hepatic catabolism of testosterone (Rubm er a/.. 1976). increased arornatase a c t ~ t y (Gordon
et al.. 1979a). decreased testosterone biosynthesis by the testis (Gordon et al.. 1980: Cicero
and B e l 1980) and diininished lutemiMg hormone releasing hormone (LHRH) release from
the hypothalamus (Chapm et al.. 1980: Cicero. 198 1 ) have ail been implicated with this effect.
Longtemi ETOH drinking is associated with testicular atrophy in males and disruption
ofthe menstrual cycle in females (Van Thiel et al.. 1977: Cobbs et al.. 1978). The increased
conversion of androgens to estrogens also produces " feminizationt' in some long-tenn
alcoholics (Gordon et ni.. 1979b). Fiiially. ETOH-mediated hypogonadism Las been
implicated m the metabolic tolerance to alcohol. indeed. the levels of hepatic ADH are under
the negative control of testosterone. so that with the suppression of testosterone synthesis by
ETOH. there is a reduction in this inhibitory control and a si&cant increase in the
subsequent rnetabolism of alcohol. leading to metabolic tolerance (Rachamin et al.. 1980:
Cicero. 1982).

d) &pothalamic-pinrita~~thyroid (Wu mis
Cûculating triiodothyronine (T,) levels are often lower with acute and chronic ETOH
constmptioa especially in alcoholics with h e r diseases (Bexmudez et al., 1975; Israel er al.,
1979). The most ~ e l y mechanism invohred appears to be a direct effect of ETOH on the
conversion of thyroxine (T,) to T, in the liver (Israel er al.. 1979). Long-term ETOH
alterations of the thyroid hormone may play a role in the etioloa of alcobolic hepatitis and
cirrhosis since the hypermetabolic state induced in the h e r by chronic h e a y ETOH
administration is eliminated in thyroidectomized animals (Bernstein et al., 1975 ).
e) Cntechoiantirle
Plasma catecholamine levels are generaily elevated following acute and chroaic ETOH
consumption (Guaza and Borrell. 1983 : Ireland et al.. 1984: Howes and Reid. 1985 : Howes
et ni.. 1986). interestingly. these ETOH-induced alterations in cuculating catecholamine
levek appear to remit prirnarily fiom reductions m epinephrine and norepinephrine clearance.
rather than fiom mcreased catecholamine release (Howe and Reid 1985 : Howes et a/. . 1986).
The substantial pressor effects of catecholamine may also explain in part the hypertension
associated with chronic heavy ETOH consumption (see section 1.2.2.3.).
f) Etldotheliirni- arrd plorelet-cierived vnsoactive factors
Tlie production and release of prostacyclin (PGI,). a local vasodepressor agent Iocated
m endothelial celis. is increased foilowing acute ETOH administration (Karanian et ni.. 1985:
Mehta er al.. 1987: Karanian and Salem, 1988). in contrast. its release is significantly
decreased following chronic Iieaky ETOH consumption or in alcoholics (Foraermann and
Feuerstain. 1987: Karanian and Salem, 1988).
The secretion of nitric oade (NO). another vasodepressor factor produced by endothelial
cells. is also increased afier acute alcohol drinking in several vascular beds. such as in
pulmonary arieries (Greenberg et al.. 1993 ). However. the ETOH-induced contraction
observed in certain vessels. such as in the aorta. has been s h o w to r e d t fiom the partial
suppression of endothelium-dependent vasorelaxation, çuggesting an inhibitory effect of

ETOH on NO release in these vessels (Hatake et al.. 1989: Wang and Fang. 1993: Hatake
et al., 1993). Nevertheles a reduction of the ETOH-induced contraction has been obsewed
m these vessels following chronic ETOH consumption. mediated by a secondary increase in
NO production and release fiom endothelial ceils and suggesting functional tolerance to the
vasoactive effects of ETOH (Knych. 1992: Hatake et al.. 1994).
Acute ETOH consumption sijpficantly lowers the production and release of
thromboxane A, (TXAZ). a vasoconstnctor agent produced by the platelet (Miai l idis et 01..
1983: Mehta er al.. 1987: Karanian and Salem 1988). TXAl is unchanged or slightly
increased following chronic ETOH treatment (Forstermano and Feuerstain. 1987: Karanian
and Salem, 1988).
Both acute and chronic ETOH connimption have been associated with elevated levels
of endothelin- 1. a local vasoconstrictor factor located in endothelial cells (Kawano et al..
199 1 : Nanji er al.. 1 994).
The balance of these opposhg effects of ETOH on the various vascular beds may explain
the presence of either vasodilation and vasoconstriction following acute and chronic ETOH
consumption (see section 1.2.1.3. and 1.2.2.3. )
g) Others
Although conflictmg, aiterations m the actMty of additional hormonal systerns by ETOH
have been observed. Elevated plasma prolactin levels have been described in alcoholics
(Linclholm et al.. 1978). The negative correlation between prolactin levels and the severity
of alcohol withdrawal rnay suggest a role for prolactin in the withdrawal symptoms
(Pohorecki and Brick 1988). The release of growth hormone (GH) is reduced both by acute
and chronic ETOH consumption (Gandai et al.. 1978: Redmond. 198 1).
1.1.5.3. Effects of ETOH on the kidnev
The modifications m water and electrolytes balance by alcohol drinking are also mediated
by direct effects of ETOH on the kidneys. Acute ETOH consumption produces diuresis. ai
least during the ascending portion of the BAC (Jones. 1990). Sodium excretion is initially

unaffected but is later iocreased to restore plasma ion levels altered by the initial water
exmetion (Ponticelli and Montagnino. 1979). Althou& the diuretic effect of ETOH is due in
part to the inhibition of AVP release. direct modifications m renal hemodynamics may also
occur. such as elevated giornedar filtration rate (GFR) (Kmck and Krecke. 1965). The
delayed natriuretic effect may also depend on GFR but appears to r e d t mainly fiom reduced
Na' reabsorption by the nephron. probably because of the inhibition of basolateral N a * K -
ATPase punip activity and the reduced gradient for Na+ reabmrption (Israel and Kalant. 1963:
Parenti et al.. 199 1 : Rothman et al.. 1992).
Chronic ETOH consumption is associated with adaptive alterations to compensate for
the repentnie acute effect of aicohol driuking. Tbe water retention and s i w c a n t antidiuresis
observed in alcoholics and chronic ETOH animal models may result 6om the elevation in
AVP release (Taivamen et al., 1995). Sodium retention is generally noted probably because
of the secondary mcreased activity of Na+/K+-ATPase with chronic ETOH. resulting in lower
mtracellular Na+ concentrations and elevated gradients for Na' reabsorption by the nephron
(Beard and Knon. 1968: Ponticelli and Montagnho. 1979: Rodrigo et al.. 199 1 ).
Hypokalemia is sometimes associated with alcoholism mainly because of deficient nutrition
(Wadstein and Ohlin. 1979). The general stimulation of Na*/K'-ATPase actMty rnay also
increase intracellular K' and hyperpolarize the cells. leading to lower circulating K- levels.
Hypokalemia lias been implicated with the development of deiirium tremens (Wadstein and
Ohlin. 1979). Plasma rnagnesium levels are also decreased by chronic ETOH consumption
and hypomagnesemia may be important for withdrawal symptoms such as seinire (Nielsen.
1965).
Renal dysfunctions are usually reversible with cessation of druiliing. although renal
papillan, necrosis rnay develop in alcoholics with liver diseases (Edmondson er al.. 1966: De
Marchi et al., 1993).
ï.l.S.4. Effects of ETOA on the liver
The central role of the h e r in the metabolism of ETOH predisposes this organ to the
toxic effects of oxidative enzymes and ETOH metabolites. However, long-term heavy ETOH

intoxication is needed for the appearance of hepatotoxicity since both hurnan and animal
studies have s h o w that chronic moderate ETOH connunption is not associated with any
manifestation of alcoholic h e r diseases (ALD) (Van de Wiel et al.. 1990: Savolainen er al..
1993). ALD is characterized by three specific histopathologic stages: fatty liver. hepatitis and
Gbrosis/cirrhosis (Crabb. 1993). Malnutrition is a conmbuting factor. but ALD rnay ail1 occur
in the absence of nutritional deficiencies (Lieber, 1994).
The oxidations of ETOH and acetaldehyde by ADH and ALDH use NAD+ as a cofactor.
With the chronic presence of large amounts of ETOH elevated levels of reducing agents such
as NADH are produced by the liver. overwheiming the hepatic capacity for NAD'
regeoeration (redo't mechanism) and leadmg to various metabolic disorders. lncreased
NADWNAD' ratio is associated with trigiyceride accumulation in the liver and the
development of fàtty liver and hypoglycemia (Salaspuro et ai.. 198 1 : Jauhonen er ai.. 1982:
Lieber. 1994). The increased MEOS actMty in alcoholics not only accelerates the oxidation
of ETOH. but also the metabolism of vanous other microsoma1 substrates. generating on
occasion metabolites more toxic than the precursors. nich as superoxide radicals (Dai et ai..
1993).
Acetaldehyde is responsiile for several hepatotoxic effects of ETOH. Lipid pero'adation
caused by acetaldehyde may damage hepatic tissue (Müller and Sies. 1982). Acetaldehyde-
protein adducts may also delay protein secretion into the plasma. resulting in hepatocyte
enlargement (Baraona et al.. 1977). modify enzymatic a c t ~ t y (Solomon. 1 987) and produce
immune reactions (Hoemer et ol.. 1986: Niemela et al.. 1987). Finally. acetaldehyde rnay
stimulate the collagen accumulation observed during the various stages of ALD (Mann et al..
1979; Moskage et al., IWO).
1.1.5.5. Effects of ETOH on the cardiovascular svstem
The two major components of the cardiovascular syaem the blood vessels and the heart
itself are both greatly affected by ETOH consumption. Because of the controversial nature
of the data obtained fiom studies mvestigating the interaction of alcohol with the vasculature.
m tenns of both blood pressure and vascular disorders. and because of their importance for

the description of the thesis hypothesk, the ETOH-blood vessels interactions wili be discussed
extensively m the section of Alcohol and Blood Pressure (section 1.2.). Consequently, the
present section wiil review only the modifications mduced by acute and chronic ETOH
drinking on the cardiac tissue.
The acute exposure of the heart tissue to ETOH reduces cardiac contractility (Wong,
1973; HoMlitz and Atkins, 1974; Urbano-Marquez et al.. 1989; Pagala et al., 1995). The
strength of the myocardium contraction is a weil defined process depending on the electrical
excitation of the cellular membrane and the subsequent activation of the mtracellular
contractile protems. Briefly, the openhg of the fast Na' channels leads to mtracellular
depolarization and to the openhg of the voltagedependent Ca" channels fiom the
cytoplasmic and sarcoplasmic reticulum membranes. Extracellular and aored Ca" rushes into
the cardiac cells, giving nse to a transient mcrease in cytosoiic Ca2+ levels. This, in tum,
activates rnyonlament contractioq via the bmdmg of Ca2+ to troponin C. The activation of the
contractile protem is terminated by the active transport of Ca2' out of the cytosol through
ATP-dependent pumps and Na+/Ca2' exchangers (Thomas et al.. 1994). The physical
intercalation of ETOH into cardiac cell membranes reduces the peak amplitude of the
cytosolic Ca" mcrease, probably via several modifications in the excitation-contraction
process (Thomas et aL. 1989). Reductions m the CaL' currents 60m the cytoplasmic and
sarcoplasmic reticulum membranes (Mongo et al.. 1990) and alterations in ionic pump
activity, such as the inhibition of the Na', K'-ATPase a c t ~ t y (Williams et ai.. 1975), have
been iniplicated with the lower cardiac contractiiity. Lower Ca2+ norage m the sarcoplasmic
retic* probably by the reduction m the activity of the Ca" pump (Thomas et ai.. 1994).
and a decreased sensitivity of the myofilaments to CaL' (Damiger et al., 199 1; Rozanski et
al.. 1992) may dso contniute to the lower contractility induced by acute ETOH.
Chronic ETOH exposure is associateci with adaptive modi6cations in cardiac morphology
to compensate for the cardiodepressant action of repeated alcohol intoxication. Gradua1
mcrease m total heart mass and dilation of the ventricles are noted (Capasso et al., 1992). The
Can channel den* is Hicreased, refiecting m n d a r y changes and tolerance to the inhiibitory
effects of acute ETOH exposure (Littleton, 1988). Components of the alcohol degradation

produas, such as acetaldehyde and fatty acid ethyl esters, which are elevated after chroaic
ETOH consumption, may dso depress cardiac contractility (Williams et al.. 1975: Lange,
1991). Eventuaily m humans, after at least a decade of heavy ETOH consumption, permanent
damages in the har t tissue may occur. Cardiomyopathy, the moa common long-tem ETOH-
related disorder, is characterized by cardiomegaly, dilation of the left ventricle and low-output
congestive heait fadure (Sheehy, 1992; Rubm and Urbano-Marquez, 1994; Reedy and
Richardson, 1994). It is a Sgdicant cause of cardiac morbidity and mortaiity. Chronic heavy
ETOH consumption is also associated with arrhythmias and in certain instances with sudden
death (Regan, 1990).
1.2. ALCOHOL AND BLOOD PRESSURE
1.2.1. ACLJTE ETOEI CONSUMPTION
1.2.1.1. Auman studies
In the last 25 years, a number of studies have mvestigated the acute hemodynarnic effects
of ETOH m human subjects (Table 1.2.1 .). However, a consensus regarding the direction. if
any, of the ETOH-induced change in BP has been difncult to obtain.
a) ETOH-ir2duced decreme irl BP
A majority of studies particuiarly in recent years, have reported a depressor effect of
acute ETOH connimption (in 38% of the midies mvestigating acute ETOH administration
and BP) (see table 1.2.1.. part A). in generai, these studies demonstrated a slow but long-
lasting hypotensive effect of acute ETOH administration fiom 30 minutes to 8 hours
following the alcohol ingestion. Furthemore, the BP r e a c t ~ t y to various stimuli mch as
exercise, was also sigiuficantly reduced in ETOH-treated mdividuals. However, HR values
were mcreased in most studies.
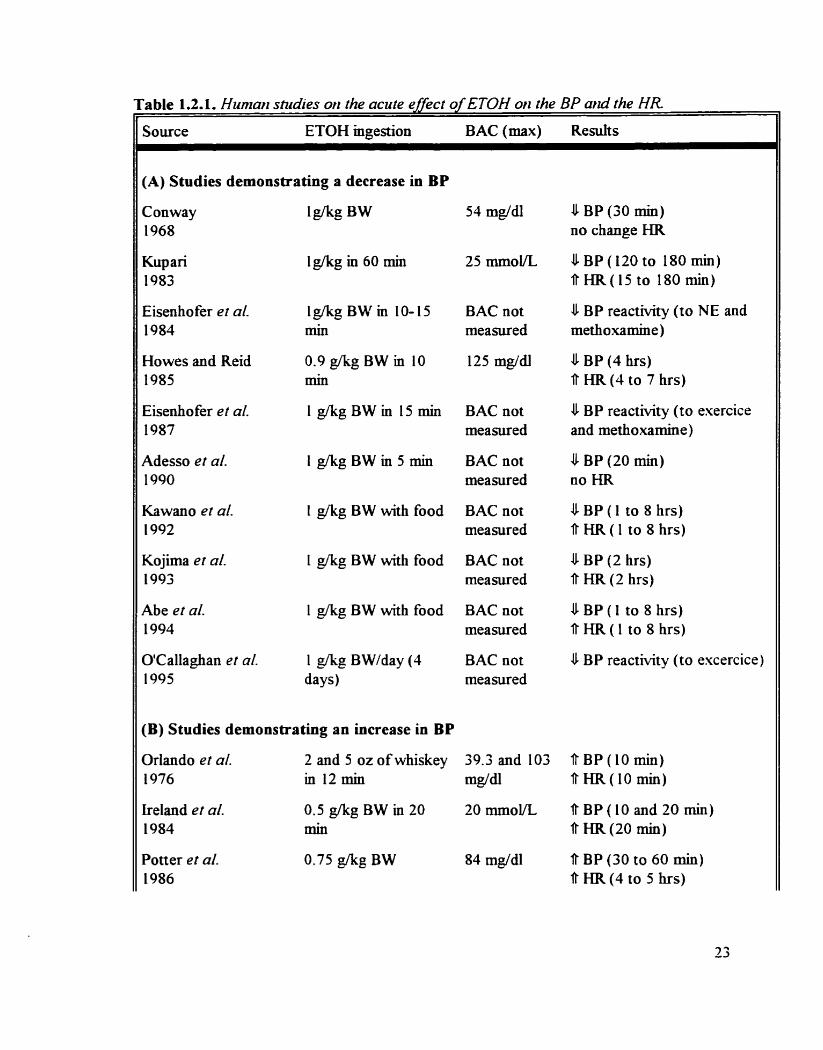
Source ETOH ingestion BAC (max) Results
Table 1.2.1, Hzintari studies on the mute effecr of ETOH on the BP artd the HR
B
1
(A) Studies demonstrating a decrease in BP
Conway 1968
Kupari 1983
Eisenhofer et al. 1984
Howes and Reid 1985
Eisenhofer et al. 1987
Adesso et al. 1990
Kawano et al. 1992
Kojima et al. 1993
Abe et al. 1994
lg/kg in 60 min 25 mmoVL
lg/kgBWmlO-15 BACnot min measured
0.9 g/kg BW in 10 125 mg/d min
1 g/kg BW in 15 min BAC not measured
l g k g B W i n 5 r n i n BACnot rneasured
1 g k g BW with food BAC not measured
1 g/kg BW with food BAC not measured
1 g k g BW with food BAC not measured
1 g/kg BW/day (4 BAC not d a ~ s ) measured
(B) Studies demonstrating an increase in BP
Orlando et al. 2 and 5 oz of whiskey 39.3 and 103 1976 in 12 min mg/dl
Ireland et al. 0.5 g/kg BW in 20 20 mmol/L 1984 min
Potter et al. 0.75 g/kg BW 84 mg/dl 1986
4 BP (30 min) no change HR
uBP(12O to 180 min) h HR(l5 to 180 min)
4 BP r e a c t ~ t y (to NE and methoxamine)
4 BP (4 hrs) h HR (4 to 7 hrs)
4 BP reactMty (to exercice and methoxarnine)
4 BP (20 min) no HR
4 BP ( 1 to 8 hrs) h HR ( 1 to 8 hrs)
4 BP (2 hrs) h KR (2 hrs)
4 BP ( 1 to 8 hrs) fi HR ( 1 to 8 hrs)
4 BP r e a c t ~ t y (to exercice)
fi BP(10 min) fi HR ( 10 min)
h BP ( 10 and 20 min) h HR (20 min)
h BP (30 to 60 min) fi HR (4 to 5 hrs)

Maheswaran et al. -1 dday 199 1
( C ) Siudies demonstrating no change in BP
R S e r al. 1969
Gould et al. 1971
Zostér and SeIlers 1977
Giles et al. 1982
Eisenhofer et al. 1983
Howes and Reid 1985
Potter et al. 1986
Stott et al. 1987
Made et al. 1993
Koskinen et ai. 1994
O'Caiiaghan et ai. 1995
6 oz of whiskey in 10 min
2 oz of whiskey in 10 min
0.3 and 0.6 g/kg B W in 10 min
180 cc of volka in 5- 15 min
0.5 and 1 g/kg B W in 10-15 min
Igkg in 60 min
0.9 g/kg BW in 10 min
1.3 g/kg BW in 60 min
0.5 g/kg BW in 10 min
1 g/kg BW in 30 min
BAC not measured
100 mg/d
BAC not measured
36.6 and 85.3 mg/dl
130 mg/dl
50 and 80 mg/a
25 mmol/L
125 mg/dl
84 mg/dl
99 rng/dl
62 mg/dl
18.9 mg/dl (breath)
BAC not davs) measured
no change BP fi HR (30 to 90 min)
no change BP no HR
no change BP 11 HR ( 15 and 45 min)
no change BP lt HR (30 to 240 min)
no change BP r e a c t ~ t y (to NE)
no change BP ( 15 to 90 min) 'IIHR(15 to 180 min)
no change BP ( I O min to 3 hrs) 'II HR (4 to 7 hrs)
no change BP ( 1 to 5 hrs) ft HR (4 to 5 hrs)
no change BP ( 1 hr to 2 days) R HR(1 to 9 hrs)
no change BP ft HR (25 and 40 min)
no change BP (3 hrs) 1 HR variability
no change BP

b) ETOH-irzcizrced irzcrease in BP
hcreased BP values after acute ETOH consumption have been reponed in a few studies
(m 16% of the studies mvestigating acute ETOH adtninistration and BP) (see table 1.7.1. pan
B). In general . these studies observed a rapid ETOH-induced hypertensive effect in the fira
30 minutes following the alcohol mgestion. Furthemore. short-lasting pressor effects of acute
ETOH administration are also noticed in heavy alcoholics. HR values were also elevated in
these studies.
C) No eDct of ETOH orz BP
Several experiments. particularly the older ones. were unable to find any m o ~ c a t i o n s
in the BP values d e r ETOH administration (m -16% of the studies investigating acute ETOH
administration and BP) (see table 1.2.1.. part C). In generaL the doses of ETOH adrninistered
in these studies were lower than m the other experiments investigating acute ETOH
conçumption and BP. Nevertheless. ETOH-mediated increases in HR va lus were noticed in
most studies.
d) Factors rrzflzrertcrng the restr lts obsen'ed nt htrmm stzrdies (A cute ETON)
A nurnber of factors have prevented a straight-fonvard comparative analysis between
studies and lirnited the value of the proposed final eEect of acute ETOH on the BP.
1 ) Previous ETOH consumption : One of the major limitation in human e~~erirnents is the
impossibility to control previous habits of alcohol consumption. Early nudies did not even
report the average ETOH consurnption of their subjects (Conway. 1968). Others simply
mixed ail the drinkmg habits of the patients together. observùig the combined effect of ETOH
m excessive. rnoderate and non-drinkers (Ra et al.. 1969: Gould et nl., 197 1 ; Ireland et al..
1984: Howes and Reid. 1985: Potter et ai., 1986: Kawano et n i . 1992: Kojima et al.. 1993:
Abe er ai.. 1994). Only a few studies screened specifically for low or moderate alcohol
consumption (Zsotér and SeIIers. 1977; Giles et al.. 1982; Kupari. 1983; Maule et al.. 1993:
Koskmen et al.. 1994: OCallaghan er al.. 1995).
2 ) Dose of ETOH: Another difncdty is the alcoholic beverage itself. The quantity. volume

and concentration of ETOH are dBerent in almost every experiment. The BAC may be
considered a relatively good parameter in order to standardize most of the results: its absence
in about a third of the experiments is therefore unfortunate.
3 ) hiration of ETOH drinkmg: The duration of the alcohol consumption may also affect the
way ETOH mteracts with the BP. This parameter varied from 5 minutes in the experiment of
Adesso et al. ( 1990) to 60 minutes in the experiment of Kupari ( 1983).
4) Resence of food: The administration of ETOH with food (ffiwano et al.. 1992) or in a
faaing state (Orlando et al.. 1976) may also affect the outcorne. since the presence of food
in the stomach delay ETOH diffusion and stomach emptying. increasing the first-pass ETOH
metabo lism (Kalant et ai., 1 97 1 ).
5 ) PathophysioloPical conditions: Various clinical conditions with potentially detrimental
consequences on BP sensdMty to ETOH. such as hypertension (Kojima er al.. 1993: Abe et
al.. 1994). heart disease (Conway. 1968: Gould et aL. 197 1 ) or angina pectoris (Orlando er
al.. 1976) have been evaluated without proper controls.
5) Aee of the subiects: Alcohol phannacokinetics are altered by the biological changes
associated wati a& (Pohorecky and Brick. 1988). This may alter the effect of acute ETOH
mgestion on the BP in the vanous studies. since the average age of the subjects ranged fiom
20 years (Adesso et a/.. 1990) to 57 years (Conway. 1968).
6) Time-course of BP measurements: Some e~~er imen t s measured the BP values minutes
afier the ETOH administration (Conway. 1968: Zsotér and Sellers. 1977: treland et al..
1984), whereas others descnied the BP alterations m terms of hours (Howes and Reid. 1985:
Kawano et al.. 1992: Koskinen et al.. 1994) and even days (Stott et ai-. 1987: Abe et al..
1994). Therefore. the initial a d o r delayed effect of ETOH administration on the BP may be
missed in some of these studies.
7) Office versus ambulatorv BP: The stress and the discodort of office BP may have been
responsble for some of the short-laaing pressor effects of ETOH. but have been elirninated
in recent years with the development of ambulatory BP (Kawano et al.. 1992: Abe et al..
1994; 0'Callagha.n er ai., 1995).
8) Isovolumetic versus isocaloric conditions: The type of control used in each audy may also

affect the r e d t . Isovolumetri~ conditions are used m most of the experiments (Zsotér and
Stellers. 1977: Ireland et aL. 1984: Adesso et al.. 1990: Koskinen et al.. 1994). although the
calonc component of alcohol is best accounted for with isocaloric controls (Stott et al.. 1987:
Made et al., 1993 : Abe et ai., 1994).
e) Srrmman
Because of the dinerent methodological procedures and the numerous factors that rnay
influence the resuits observed in human studies. it is dBcult to conclude on the exact effect
of acute ETOH consumption on the BP. Nevertheless. a tentative general description of the
effect of ETOH m human aibjeas is possible. indeed. rnoderate ETOH consumption appears
to produce longlasting reductions in the BP. Such reductions rnay be seen a s early as 20 to
30 minutes &er the ETOH administration (Conway, 1968: Adesso et ni.. 1990). but are most
probably significantly difEerent around the first hour and up to 8 hours following the alcohol
mgestion (Kupari. 1983: Howes and Reid. 1985: Kawano et ai.. 1992: Kojima et ai.. 1993:
Abe et a(. . 1994). hterestingly. a rapid and short-lasting pressor effect of ETOH rnay exist
m the firn minutes following alcohol consumption (Orlando et al.. 1976: Lreland et ai-. 1984:
Potter et ai.. 1986). The absence of this rapid BP increase durinp ambulatory BP
meawements rnay suggest an effect of stress and discodort rather than alcohol itself (Abe
et ai. 1994: O'Callaghan et al.. 1995). The caloric load of ETOH rnay also eqlain tliis fast
mcrease m BP m experiments using isovolumetric controls (Ireland et ai.. 1984: Potter et ni..
1986). The possibility of a secondary withdrawal reaction rnay also exist. especially in audies
where the subjects are regular ETOH drinkers (3 and more drinkdday) (Potter et al.. 1986).
Lower quantities or longer time of ETOH administration are not associated wkh sipificant
modifications on the BP (Zostér and Sellers. 1977; Stott et ai.. 1987; Made et al., 1993 ). In
contrast. almost al1 experiments have reported a significant increase in the E-R lasting from
a few minutes to a few hours following the ETOH ingestion.
1.2.1.2. Animal studies
There have been comparatively few animals studies on the acute effect of ETOH on the

BP (Table 1.2.2.). Surprisingly. the direction of the ETOH-induced change is Iess
controversial.
CI) E TOH-irrdztced decrease itt BP
Moa of the animal audies have demonstrated a depressor effect of acute ETOH
connimption on the BP (in 6706 of the studies investigating acute ETOH administration and
BP) (see table 1.2.2.. part A). in generaL the ETOH-induced hypotensive effect was noticed
m studies using gastric mtubation as their route of alcohol administration. Furtherrnore. these
studies demonstrated a slow but long-lasting hypotensive effect of ETOH from 30 minutes
to 4 hours. The BP reactivity to vanous stimuli. nich as stress. was also reduced in ETOH-
treated animals. However, the HR values were elevated in most studies.
6) No effect of E TOH mi BP
A few experiments reported no significant variation in the BP following ETOH or
acetaldehyde administration (in 33% of the nudies investigating acute ETOH administration
and BP) (see table 1.2.2.. part B). in general. these studies used i.p. administration of low
levels of ETOH. Nevertheless. the IiR values were elevated in certain studies.
C) Factors itrjltretrcitrg the reszdts obsened itr mimai stirdies (Acirte ETOH)
I ) Previous ETOH exposure: The main advantage of animal versus human studies is the
coqlete control over the conditions. pan and present. of the e'cperimental subject S. ETOH-
naive animals enable to audy the initial acute effects of alcohol. without the possibility of
secondary modifications produced by years of regular alco bol consumption.
2 ) Dose of ETOH: Nevertheless. the concentration and volume of the ETOH &&en in any
study may ail1 result in dserent BAC levels and therefore in different effects on the BP
( Roine et ai.. 1993).
3) Route of ETOH administration: Comparative analysis is further complicated by the route
of ETOH administration in animal studies. either i.p. (Sparrow et al.. 1987). i. t: (Gulati er
ai.. 1989; Tabuzchi and Pang, 1992) or through gastric intubation (Malinowska et al.. 1989:

Source Species and ETOH BAC Results
Table 1.2.2. Animal studies on the amte eflect of ETOH ori the BP ard the HR
I
1
A) Studies demonstrating a decrease in BP
Guiati et al. 1989
Tabrizchi and Pang 1992
Sparrow et al. 1987
Malinowska et al. 1989
Thompson and Adams 1994
Brackett et al. 1994
and 90%) (Cats)
2.4 and 4.8 i-v. mg/min/kg BW ( Sprague-Dawley rats)
[bl Z.P. admirlistmtior
1.p. ( Sprague-Dawley rats)
2 g k g BW gmtric iritztbatiotl
(Wistar rats)
5 gfkg BW gastric itltn batzoti (Sprague- Dawley rats)
2,4 and 6 g/kg BW gmtric itrtu batioti ( Sprague- Dawley rats)
BAC not measured
BAC not measured
BAC not measured
BAC not measured
BAC not rneasured
63, IO3 and 22 1
mg/ dl
4 BP (30 min) 8 HR (30 min)
6 BP (12 min) (4.8 m a l 3 BW) no change HR (12 min)
4 BP reactMty (to strtss) fi H R ( 5 to 120 min)
8 BP (30 to 60 min) fi HR(l5 to 60 min)
8 BP (60 to 230 min) h HR (60 to 240 min)
4 BP (60 to 120 min) (4 and6 g/kg BW) f i HR (for 4 hrs)
B) Studies demonstrating no change in BP
[a 1 i. p. adnzirlstratiorz

Sparrow et al. 0.08 to 6.3 g/kg BW BAC not 1987 i-p. measured
( Sprague-Dawley rats)
Pawlak et al. 3.6 and 12 mgkg BAC not 1993 B W i. p. acetaldehyde meanired
( Wistar rats)
[b 1 i. v. adnz fizistrarioit
Abdel- Rahman 0.1.0.5andlg/kg 71.8 1994 BW i.v. ( SHR. WKY mg/dl(O.5
and Sprague-Dawley g/kg BW)
no change BP ( 5 to 130 mm) h HR ( 5 to 120 min)
no change BP ( 15 to 60 mm) no change HR ( 15 to 60 min )
no change BP (90 min) fi' HR
I rats)

Brackett et ai., 1994).
4) Different specieh: Inter-species variability and strah differences in the cardiovascular
response to acute ETOH consumption may also produce confusion Hi the direction and
magnitude of the BP m o ~ c a t i o n s .
5) Human versus animal studies: Although comparative analysis between animal and human
studies are possible. a direct extrapolation of results obtained on laboratory animals to human
physiology and pathophysiology is hazardous. For example. alcohol metabolism is faner in
rats than in humans (40 vernis 18 mgdl per hour). reducing the effect of a particular BAC
level in rats compared to the effect observed in humans (York 1982: GiIl et al.. 1986).
Nevertheless. animal experiments remain useful models of ETOH consumption and
alcoholism by allowing the nudy of parameters not accessible on human subjects.
d) S z i m n i q
in generai these studies support a slow and long-lasting depressor effect of acute ETOH
consumption in animal experiments. The short-Iasting mcrease in the BP observed
immediately afier ETOH administration in human studies is absent fiom evperiments
performed on rats and cats. The direct recording of BP through a catheter in several studies
may prevent the stress and discornfort caused by the irnmobilition or manipulation of the
animals (Sparrow et al.. 1987: Mahowska et al.. 1989: Thoinpson and Adams. 1994).
Smaller ETOH doses are not associated with any modification in BP values (Tabrizchi and
Pang. 1992: Abdel-Rahman. 1994). In contrast. the HR is elevated in most experiments.
1.2.1.3. Mechanisms
Because of the short-term duration of the acute ETOH effect on the BP. vascular flow
and resistance rather thm the renal characteristics have been investigated. Non-specific
association of ETOH with vascular smootli muscle cells and secondary modifications in ion
cliannels and pumps. as well as ETOH-induced modifications in local and endocrine
vasoactive factors have been postulated. Depending on the vascular bed and the doses of
ETOH @en, vasodilation or vasoconsûiction have been O bserved ( Altura and Altura. 1 9 82).

Low ETOH concentration increases the blood flow in mesenteric. splanchnic. coronary
and portal vessek (Fewings et al., 1966: Carmichael et al.. 1988: Maule et ai.. 1 993 ; Pirwitz
et al.. 1995). Si@cant reductions in total peripheral resistance (TPR) during the
hypotensive phase of acute ETOH administration have also suggested a general peripheral
vasodilation (Kupan. 1983: Kawano et al.. 1992: Abe et ai.. 1994).
1 ) Direct vasodilatory effect of ETOH: A portion of the ETOH-mediated relaxation of
precapillary çphrinters. artenoies and muscdar venules is proposed to result fiom a direct
action of alcohol on smooth muscle cells (Altura and Altura. 1982).
2) Ca2'levels: Lower mtraceilular levels of C a . probabiy via the reduction of intracellular Na-
levels. have been demonstrated with acute ETOH administration and may decrease the
contractile a c t ~ t y of smooth muscle ceiis (Blauaein, 1977: Turlapaty et al.. 1979: Kojirna
et al.. 1993 ).
3 ) Inhibition of ~ressor hormonal vasoconstriction: Moderate amounts of ETOH have also
been shown to reduce the vasoconariction produced by hormonal factors. such as
catecholarnine or AVP (Edgarian and Altura. 1976: Altura and Altura. 1 983 ). Similarly. the
production of local vasoconstrictor agents. such as thromboxane A, (TXA) fiom the
platelets. is reduced foUovhg light ETOH consumption (Mikhailidis et al.. 1983 ). In contrast.
the ETOH-induced mhiition of AVP release has been demonstrated not to contribute much
to the initial decrease m BP d e r ETOH consumption. but may be imponant in sustainin_o the
hypotensive effect by srirnulating diuresis (Cowley and Liard. 1988: Kawano et ni.. 1 992).
4) Stimulation of hormonal vasodilation: The release of local vasodepressor agents. such a s
prostacyclin (PGL) and nitric oxide (NO) fiom the endothelial celis. is increased foilowing
acute consumption of low levels of ETOH (Karanian et al.. 1985: Greenberg et al.. 1993 ).
6) C'mocor lstrictiorr
ETOH is associated with direct vasoconstriction in cerebral. intrapulrnonary and renal
arteries (Altura and Altura. 1982: Toda et al.. 1983).
1 ) Inhibition of hormonal vasodilation: There is the possibility that elevated levels of ETOH

reduce the vasodilation produced by acetylchohe and ATP. although low ETOH
concentrations may potentiate the vasorela?tation caused by these agents (Criscione et ai..
1989). The suppression by hi& ETOH concentration of endothelium-dependent
vasorelaxation. possibly via NO. has also been posnilated in rat aorta (Hatake et ai.. 1989:
Wang and Panp 1993: Hatake et al.. 1993).
2) Stimulation ofhomonal vasoconstriction: The potentiation by heavy ETOH consumption
of the pressor effects of vasoconstrictor agents such as catecliolamines may also occur
(Edganan and Altura. 1976: Talesnik et al., 1980). In several studies. there is an
augmentation in circulating catecholamine levels following acute ETOH administration.
probabiy due to an mcreased release from the adrenal medulla and sympathetic nerves as weli
as to a reduction in catecholarnine clearance (Eisenhofer et al., 1983; Ireland et al., 1981:
Howes and Reid 198% Grassi et ai.. 1989). However. this increase may be secondary to the
hypotensive effect of ETOH since an inverse relationship has been found between the a c t ~ t y
of catecholamine and the extent of the BP reduction (Kawano et a/., 1992). Sirnilarly. the
actMty of the RAAS appears to be unchanged following a single exposure to ETOH. or only
slightly mcreased as a compensatory mechanism to the depressor effect of alcobol (Porter et
ai., 1986: Stott et al., 1987: Kawano et al.. 1992).
3 ) Ca"leve1s: n i e ETOH-induced vasoconstriction rnay ultimately depend on intracellular
Ca" levels. There is some reports that suggest that Ca" levels are increased in certain
vascular beds following acute ETOH admmistration through the inhibition of Na'.K'-ATPase
a c t ~ t y ( Arkwright et ai.. 1984).
C) E TOH-ir zdzt ced irrcrease irr HR
The ETOH-mduced tachycardia obsewed m most studies rnay represent a reflex response
to compensate for the reduction in BP. both fiom initial vasodilation and fiorn delayed
diuresis, thus explainhg the prolonged effect of ETOH on HR a c t ~ t y (Stott et ai.. 1987:
Malinowslia et al.. 1989; Kawano et al.. 1992). It may also be secondary to the decreased
myocardial contractility ( Urbano-Marquez er al.. 1989).

1.2.2. CBRONIC ETOB CONSUMPTION
1.2.2.1. Human studie~
There have been 60 or so epidemiological and experimental studies investigating the
relationship between chronic alcohol consumption and BP since 19 15 (Table 1.2.3. ).
Remarkably. this issue is stiU very controversial. Most of the experiments rneasured the
average systolic and diastolic BP for several speci6c groups of ETOH drinkers. ranghg fiom
occasional to heaw ETOH coonimption. ûne ofthe early problem was the various and often
misleadmg ways the alcohol consurnption was reported in these specific groups. either as o r
ml. g or ?h of ETOH. complicating comparative analysis (Turner. 1990). Therefore. for
standardkation of this chapter. the levels of alcohol consumption m humans are aIi expressed
m standard druiks per day. As a d e . 1 drink is dehed as the equivalent of log of ETOH. or
about 1 glas of wine. one c m of beer or a single measure of spirits. Thus low. moderate and
heavy ETOH con~uniption usuaiiy refer to less than 1 drink/day. between 2 and 3 drinkslday
and a bove 4 drinlidday respectively.
O) Heap E TOH cotuirnlptiori
With the exception of two eqeriments (Baghurt and Dwyer. 198 1 : Coates et al.. 1985 ).
heaw ETOH consumption has always been associated with elevated BP and hypertension
(Table 1.2.3.). The £ira evidence of a pressor efFect of large quantities of alcohol was
published by Camille Lian m 19 15 wbo reported a linear mcrease in systolic BP among French
s e ~ c e m a i with mcreasing amounts of ETOH consumed daily. interestingiy. the group with
the lowea average consumption of alcohol. descnbed as "sobres" (sober). still managed to
dnnk 1 liter of wine/day! Subsequently. from the late 1960s to 1995. a great number of
epidemiological midies observed significant elevations in BP following heacy ETOH
consumption. such as Gyntelberg and Meyer ( 1974) ( 2 5 drinksiday). Klatsky et al. ( 1977)
( r 5 drinkskiay) or Trevisan er al.. ( 1987) ( r 7 drinkdday). Sirnilarly, experimental studies on
heavy alcoholics revealed sigruficant mcreases in their BP compared to moderate drinkers and
non-drinkers (Saunders et al.. 198 1 : Ibsen et ni.. 198 1 : Potter and Beevers. 1984: Puddey et
al.. 1985: Seppa et al.. 1994).

Table 1.2.3. Hzrmari snrdies or1 the chrotiic effecrs of ETOH oti the BP (the range of the
Source Number of Age Results
(A) Studies demonstrating a U- or J-shaped curve
[a J Laver dme of ETON measwed: < I dril~Wdav
Harburg et al. 1980
Criqui et al. 1981
Gordon and Kannel 1983
Harlan et al. 1984
Klastsky et al. 1986
Weissfeld et al. 1988
Marmot et al. 1994
GiUrnan et al. 1995
MacMahon et al. 1984
[b] Laoer dase ETOH rneaszrreci: f drirzWdqv
5550 25-64 J-shaped curve m males ( 1 to >3 drinkdday ) U-shaped cuve in females
doses of daiiy ETOH corrrumptior2 memured h i each stuc& is irrdicated iri parenthesis).
I m
1
1
J-shaped curve in males (0.04 to 7.3 drinkdda y) U-shaped curve in females
J-shaped c w e in males (< 1 to >3 drinksiday ) U-shaped curve in females
J-shaped curve in males (< 1 to >3 drinksiday ) U-shaped c w e in females
U-shaped curve in males (< 1 to >3 drinkdda y ) U-shaped curve in females
S-shaped curve in females ( < 1 to >6 drinkdday )
I-shaped and threshold curves in males ( < O S to >2 drinkdday) U- and J-shaped curves in fernales
J-shaped curve in fernales (< 1 to > 10 drinkdday )
J-shaped c w e in males (< l to >3 drinkdd a y) J-shaped curve in females
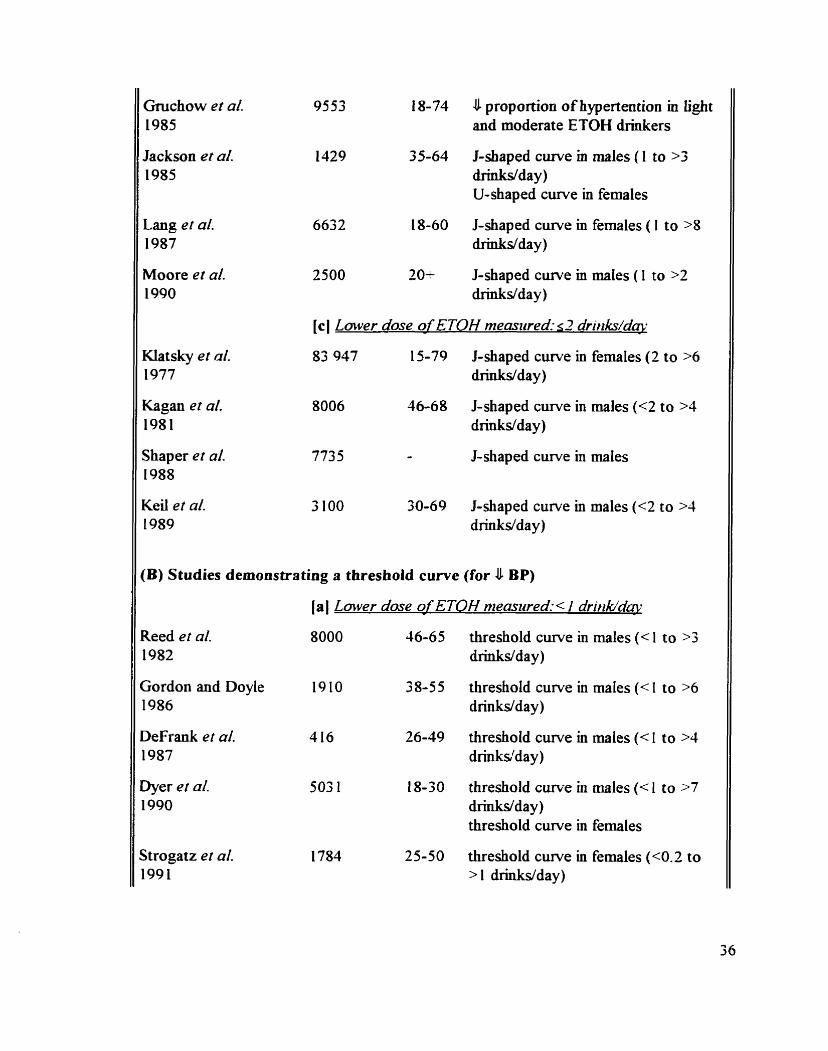
Gruchow er al. 9553 18-74 Vproportionofhypertentionintight 1985 and moderate ETOH drinkers
Jackson et al. 1985
Lang et ai. 1987
Moore er al. 1990
Kagan et ai. 1981
Shaper er al. 1988
Keil et al. 1989
1429 35-64 J-shaped curve m m i e s ( 1 to >3 drinkdda y) U-shaped curve in females
6632 18-60 J-shaped curve in fernaIes ( 1 to >8 drinkdday )
2500 ZO+ J-shaped curve in males ( 1 to >2 drinks/day)
[c 1 Laver dose of E TOH rnemzired: < 2 drirzks/dai:
83 947 15-79 J-shaped curve in females (2 to >6 drinkdday )
8006 46-68 J-shaped curve in males (<2 to >4 drinkdday)
773 5 - J-shaped c w e in males
3 100 30-69 J-shaped curve in males (<2 to >4 drinkdda y)
II (B) Studies detnonstrating a threshold eurve (for 11 BP)
[a 1 Lower dose cf ETOH nreaswed: < 1 dritiWdav
Reed et ai. 8000 46-65 threshold curve in males (< 1 to >3 1982 drinkdda y )
Gordon and Doyle 1910 3 8-5 5 threshold curve in maIes (< 1 to >6 1986 driokslday )
DeFrank et al. 4 16 26-49 threshold curve in males (<l to >4 1987 drinkdday )
Dyer et al. 1990
503 1 18-30 threshold c w e in males (< 1 to >7 drinkdday ) threshold curve in females II ~;-lpatz et al. 1784 25-50 threshold cuve in females (<O. 1 to > 1 drinkdday )
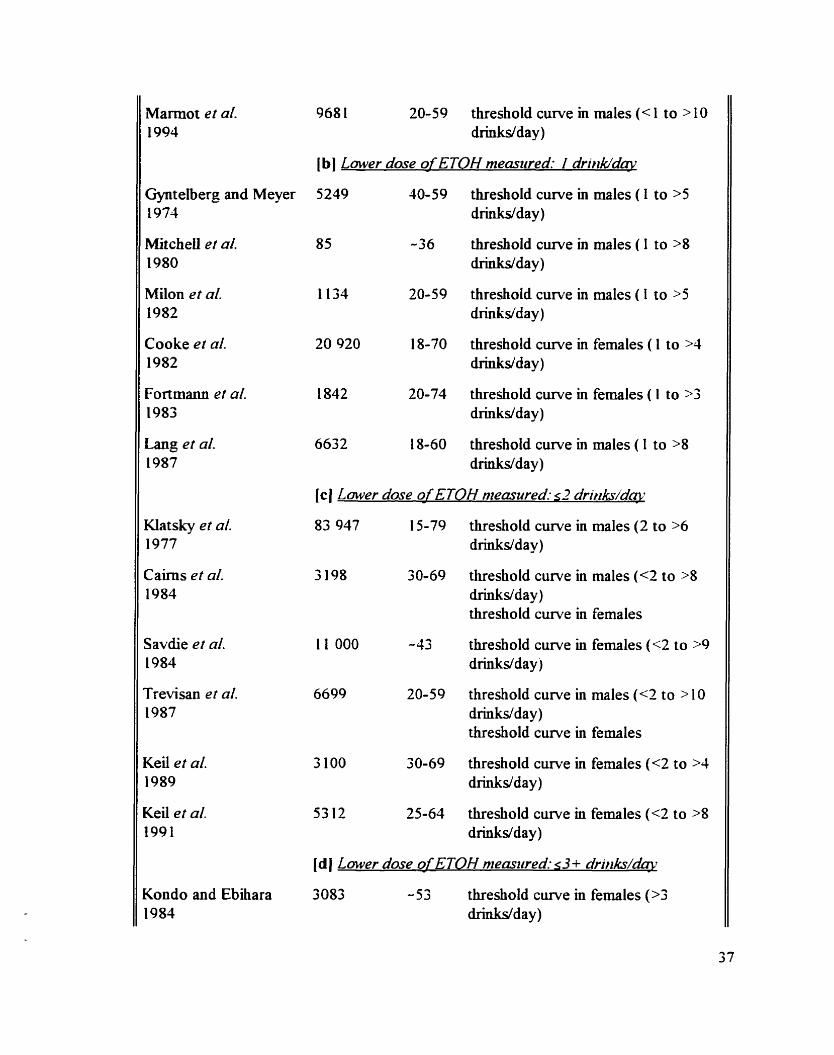
Marmot et ai. 1994
Gyntelberg and Meyer 1974
Mitchell et al. 1980
Milon er al. 1982
Cooke et al. 1982
Fortmann et al. 1983
Lang et al. 1987
Klatsky et al. 1977
Cairns et al. 1984
Savdie et al. 1984
ïrevisan et ni. 1987
Keil er al. 1989
Keil et al. i991
Kondo and Ebihara 1984
968 1 20-59 threshold curve in males (< 1 to > 10 drinkdday )
[b] Laver dose qf ETOH rneaîured: I dritik/d-
5249 40-59 threshold c w e in males ( 1 to >5 drinkdda y )
85 -36 threshold curve in males ( 1 to >8 drinkdda y)
1134 20-59 threshold curve in males ( 1 to >5 drinkdday )
20 920 18-70 threshold c u v e in females ( 1 to >4 drinkdday )
1842 20-74 threshold curve in females ( 1 to >3 drinkdday )
6632 18-60 threshold curve in males ( 1 to >8 drinkdda y)
[c 1 Lower dase of ETOH measztred 52 dritcksh"d
83 947 15-79 threshold curve in males (2 to >6 drinkdda y)
3198 30-69 threshold cuve in males (<2 to >8 drinkdda y) threshold curve in females
11 O00 -43 threshold curve in females (<2 to >9 drinkdda y j
6699 20-59 threshold curve in males (<2 to > 1 O drinkdday ) threshold curve in females
3 100 30-69 threshold cuve in females (<2 to >4 dnnkdday)
53 12 25-64 threshold curve in females (<2 to >8 drinkdda y)
[d 1 Lwer dose of ETON nlemwed: 53 + dritzksldaÿ
3083 - 5 3 threshold curve in females (>3 drinks/day)
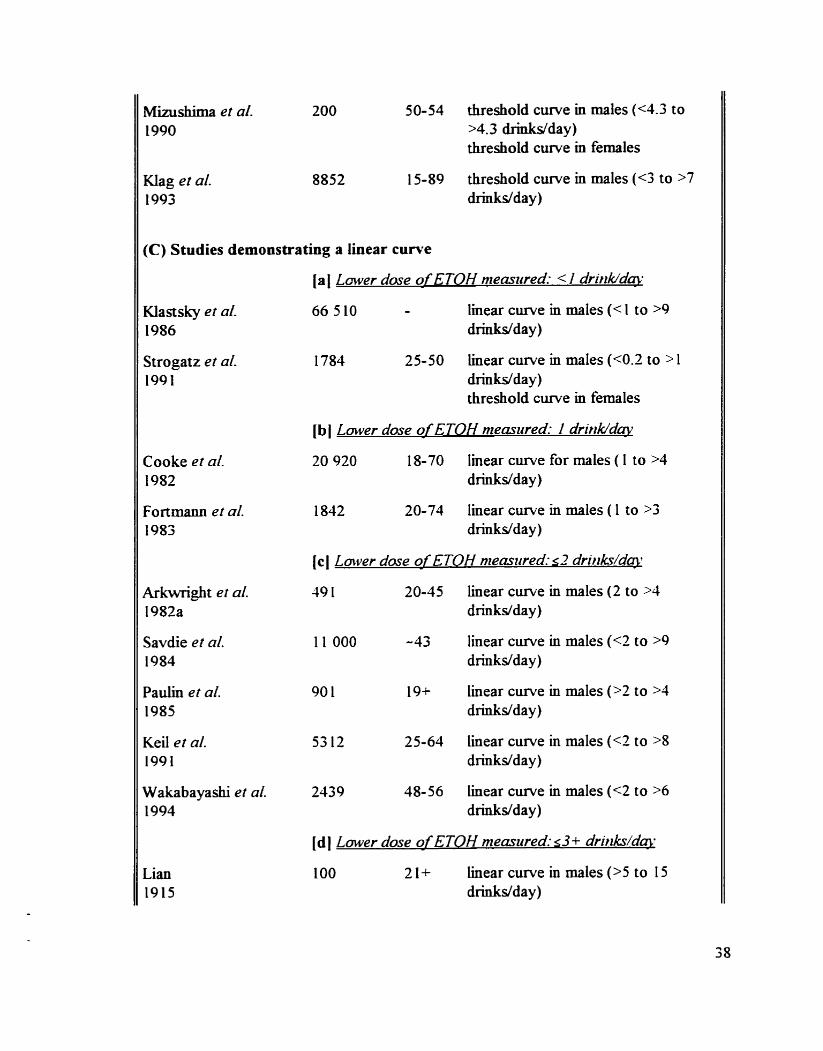
Mishima et al. 200 1990
Klag et al.
5 0- 54 threshold cuve m males (c4.3 to >4.3 drinkdday) threshold curve in females
15-89 threshold cuve in males (<3 to >7 1 9 6 drinkdda y )
(C) Studies demonstrating a Iinear curve
[a 1 Luwer dose of E TOH rneasztred: < i drir&dày
Strogatz et al. 199 1
Cooke et al. 1982
Fortmann et al. 1983
Arkwright et al. 1982a
Savdie et ni. 1984
Paulin et al. 1985
Keil et al. 199 1
Wakabayashi et al. 1994
66 5 10 - linear curve in males ( 4 to drinkdda y)
1784 25-50 linear curve in males ( < O 2 to > 1 drinkdday ) threshold curve in females
[b 1 Lmver dose of ETOH nieasured: I drirzWd~v
20 920 18-70 linear curve for males ( 1 to >4 drinkdday )
1842 20-74 linear curve in males ( 1 to >3 drinkdday )
[c 1 Laver dose of ETOH nzeasured: 57 dfitrks/da
49 1 20-45 linear cuve in males (2 to >4 drinkdda y)
1 1 O00 -43 h e a r curve in males (<2 to >9 drinkdda y)
90 1 19+ h e a r curve in males (>2 to >4 drinkdday )
53 12 25-64 Linear curve in males (<3 to >8 drinkdday )
2439 48-56 Linear curve in males (<2 to >6 drinkdda y)
[ d 1 Lower dose of E TOH memureci: 53 + dri>ih/dm
1 O0 2 1 + linear curve in males (>5 to 1 5 drinkdda y)
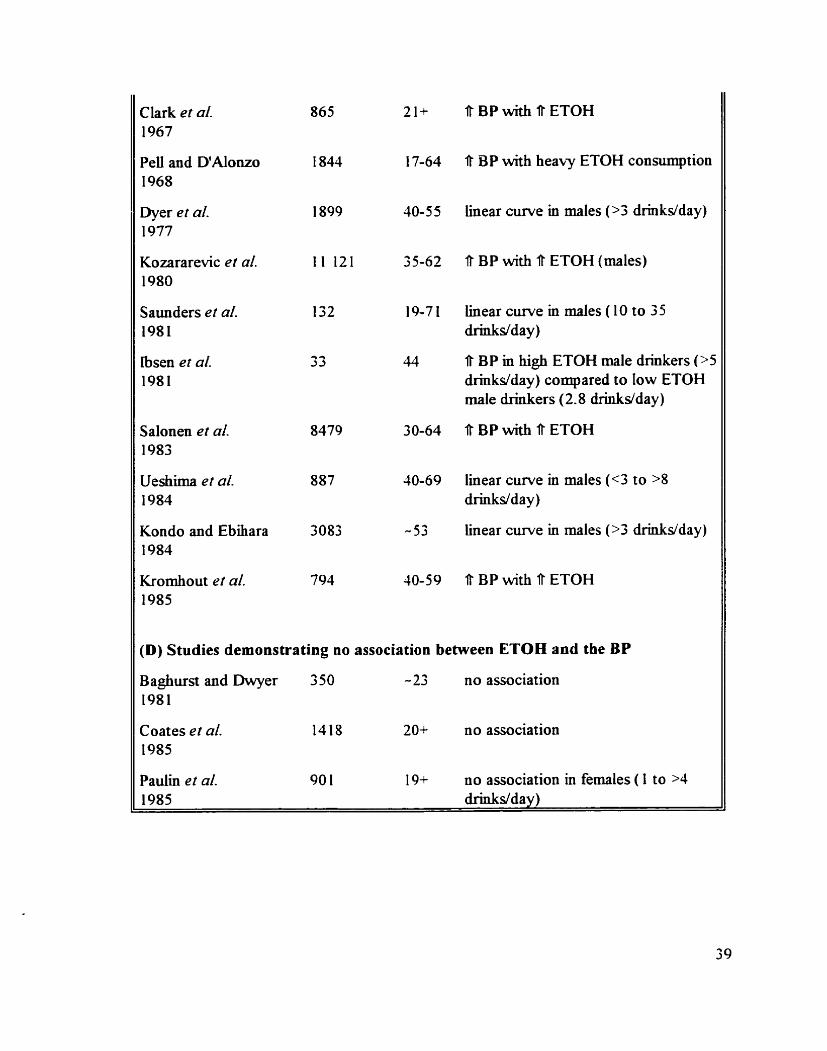
Clark et al. 1967
Peu and D'Alonzo 1968
Dyer et al. 1977
Kozararevic et al. 1980
Saunders et al. 1981
Ibsen et al. 1981
Salonen et al. 1983
UeshUna et al. 1984
Kondo and Ebihara 1984
Krornhout et al. 1985
17-64 T BP with heaky ETOH conçumption
40-55 linear cume in males (>3 drinkdday)
35-62 h BP with fF ETOH (males)
19-71 l inearcweinmales( lOto34 drinkslda y)
44 h BP in high ETOH male drinkers (>5 drinkdday ) compared to low ETOH male drinkers (2.8 drinks/day )
30-64 ft BP with h ETOH
40-69 ünear curve in males (<3 to '8 drinkdday )
-53 Linear c w e in males (>3 drinkdday)
(D) Studies demonstrating no association between ETOR and the BP
Bagburst and m e r 3 50 -23 no association 1981
Coates et al. 1418 20+ no association 1985
Paul i et al. 90 1 19+ no association in females ( l to >4 1985 drinkdday )

b) Light and nzoderate E ?'OH corrsurnprio~
The controversy lies with the effect of Light and moderate alcohol consumption on the
BP. This area of the ETOH-B P relationship is important to consider since it represents the
alcohol consumption of the ovenvhelmmg majority ofthe population (Kaplan. 199 1 : Harburg
et al.. 1994). Vanous patterns of BP progression with increasing ETOH connimption have
been descnied (Gleibemian and Harburg 1986: Criqui et al.. 1986: MacMahon et al.. 1 987).
1 ) U- or J-shaped curve: In quite a few number of nudies. the existence of a U- or J-shaped
cuve has been reported. suggeshng that light and moderate ETOH drinkers have sigmficantly
lower BP than both heavy and non-drinkers (see table 1.2.3.. part A). in the experirnent of
Harburg et ai. ( 1980) for example. male and female subjects with m~des t ETOH consumption
ranging from 0.04 to 1 and fiom 0.04 to 3.2 drllilidday respectively were associated with
reduced systolic BP when compared to non-drinkers. Heavy ETOH consumption (27
drinkdday) was still associated with hypertension. interestingly. the range and extent of the
ETOH-induced hypotensive etfect has been consistently geater in female than in male
subjects.
2) Threshold-sha~ed curve: There are also several studies where ETOH consumption below
a certain level has no sigdicant effect on the BP. suggesting a threshold-shaped curve with
increasing quantity of alcohol. in these studies. although there is no significant hypo- or
hypeitensive effect of light and moderate ETOH consumption. heavy alcohol drinlcing above
a certain quantity is associated with hypertension (see table 1 .LX. part B). For esample.
Trevisan et ai. (1987) noted no significant effect of ETOH on the BP below alcohol
consumption ofabout 5 W d a y . Above these levels. elevations in BP were reported. The
lefi-end portion of the threshold-shaped curve has been consiaently longer in female
compared to male subjects.
3 ) Lmear-shaped cuve: A smaller number of experiments have proposed a linear association
between ETOH and the BP. suggesting a significant BP increase with the lowest quantity of
reported ETOH (see table 1.2.3.. part C). In the epidemiological study of Ueshirna et a/.
( 1984) for example. a linear increase in systolic BP is noted from i3 to 28 drinkdday.

c) Factors irifz<erici~ig the restrlts observed iri epidemiological shrdies
There are a number of factors to consider when investigating the comparative value of
these audies. particularly when lookuig at the low end of the ETOH-BP relationship.
1 ) Low spectrum of ETOH consumotion: T O some extent. the shape of this relationship
depends primady on the number and values of the lowest categories of ETOH consumptions
&en in each experiments. Indeed. most of the Linear-shaped curves are reponed in studies C
where the lowest group of aicohol drinkers consumed 52 drinkdday (Savdie et al.. 1984: Keil
et al.. 199 1 : Wakabayashi et al.. 1994) or even s3 drinkdday (Lian. 19 15: Dyer et al.. 1977:
Sauoders et al.. 198 1 : Arkwright et al.. 1982a: Ueshima et al.. 1984). Any effect of light and
moderate ETOH consumption is loa in such e'rperiments. In fact. only a handfùl
epidemiological studies specifically recorded more than 1 dose of alcohol consumption below
2 drinkdday (Harburg et al.. 1980: Criqui et al.. 198 1 : Gordon and KanneL 1983: Harlan et
al.. 1984: Weissfeld et al.. 1988: Strogatz et al.. 199 1 : Marmot et al.. 1994: Gillman et a/..
1995). Ofthese experiments only one failed to observe a .J- or U-shaped relationship for the
BP (Strogatz et al.. 1991).
2) Non-drinker catepory: Nwertheless the scientific community has been reluctant to accept
the possibility of an hypotensive effect of chronic light and moderate ETOH consumption.
One of the major concem is the possible heterogeneity of the non-drinkers categoiy. It has
been suggeaed that previously heavy ETOH drinkers and ex-drinkers who were forced to
stop drinkmg because of developmg health problems are also present in this category. so that
the average BP is higher than it should have been if Me-long non-drinkers were the only
members of this eroup (Shaper. 1990: Beilin and Puddey. 1992). Likewise. it has been
postulated that socioeconomical factors could explain the lower BP with light ETOH
consumption. since wine drinkers who represent the majority of light and moderate alcohol
drinkers tend to be fiom higher social class than non-druikers (Colslier and Wallace. 1989:
Burke et al.. 1995). However. J-shaped cuwes were still observed in epidemiological nudies
with life-long non-drinkers. Similarly. the exclusion of ETOH abstainers because of poor
health or the consideration of the social level throughout the ETOH consumption spectrum
did not modify the pattern of the BP alterations, suggesting a distinct effect of the lower

ETOH levels on the BP (Marmot and Bninner. 199 1 : Cullen, 1993; Kem 1993 ).
3 ) Self-reporting: There is a tendency to underestimate the quantity of ETOH consumed in
epidemiological self-reports particularly m the lower leveis of ETOH consumption (Redman
et al.. 1987). This tendency may increase the range of the depressor action of alcohol and
limit M e r the value of a number of linear- and threshold-shaped experiments with hi@
initial ETOH consumption.
4) Contnîuting factors: The age of the subjects is likely to mod@ the relationship between
ETOH and the BP (Lang et al.. 1987: Weissfeld et ai., 1988: Keil et al.. 199 1: Gillman et al.,
1995). Likewise. the lipid content (Salonen et al.. 1983: Gruchow et d. 1985: Minishima
et al.. 1990). the race (Klatsky et aL. 1986: Dyer et al.. 1990). the smoking habits (Savdie
et al.. 1984: Keil et al.. 199 1). the physical actMty (Reed et aL. 1982: Keil et al.. 199 I ) or
the presence of ilInesses such as hypertension (Potter and Beevers. 1984) have al1 been
demonarated to contribute differently to the s ens i t~ ty of the ETOH-BP curve. Sex-
dependent modifications in this relationship have also been noticed in almoçt every
eqerirnent. Indeed there has been a longer and deeper hypotensive portion of the ETOKBP
spectmm in female cornpared to male subjects (Harburg et ni.. 1980: Weissfeld et al.. 1988).
5) Pattern of ETOH connimption: The pattern of the ETOH consumption rnay influence the
BP. since binge rather than replar drinking is associated with elevated BP ( Seppa et O/. . 1994).
6) Time-course of ETOH measurement: The time from the last aicoholic drink may also
~ e a t l y m o d e the BP values. since the experirnent may measure either the depressor = component of the repeated acute ETOH consumption or a secondary withdrawai reaction
(Howes et al., 1986: Abe et al.. 1994).
7) Type of alcoholic beveraq: distinct components of the various ETOH beverages. such as
wine. beer or spirits. may daerently alter the ETOKBP relationship.
Interestingly. the existence of a U- or J-shaped cuwe has been achowledged for the
effect of alcohol on coronary heart disease and mortality (Veenstra. 199 1 : Renaud and De

LorgeriL 1992; Coate. 1993: Gronbaek et a/.. 1994). It is therefore iikely that the effect of
ETOH on cardiovascular components such as the BP is also biphasic in nature. with hypo-
and hypertensive actions of low and heavy ETOH consumption. respectively.
1.2.2.2. Animal studies
Much like for the acute studies. there have been fewer animal experiments on the chronic
effects of ETOH on the BP (Table 1.2.4. ). The issue remams controversial.
a) ETOH-iridztced decrease itl BP
Vanous studies have demonstrated an antihypertensive effect of chronic moderate ETOH
consumption (m 50% ofthe çtudies mvestigating chronic ETOH administration and BP) (see
table 1.2.1.. part A). in general. these studies reponed a slow and progressive
antihypertensive effect of ETOH in spontaneously hypertensive ( S m ) and Wistar-Kyoto
(WKY) rats that needed at least 12 weeks to reach çignificance. Most studies used ETOH
rnixed in water in moderate concentration (20% v/v).
b) ETOH-i~rd~tced increase iti BP
Despite the experirnental evidence. acceptance of the possibility of a depressor effect of
chronic moderate ETOH consumption by the scientific comrnunity has been diicult. This is
probably due to several reports suggeaing a pressor effect of chronic ETOH administration
(m 42% ofthe snidies investigating chronic ETOH administration and BP) (see table 1.2.4..
part B). In general. these studies reported a fast hypertensive effect of ETOH in Sprague-
Dawley and Wistar rats that need between 1 to 4 weeks to reach sigtüficance. Al1 studies used
ETOH mixed in water in low to moderate concentrations (5 to 20% vk) .
c,i !Vo effect of ETOH oti BP
In one study. the administration of a low concentration of ETOH ( 10% v/v) for 1 1 weeks was
not associated with significant modifications in the BP of SHR and WKY rats (see table
1.2.4.. part C).
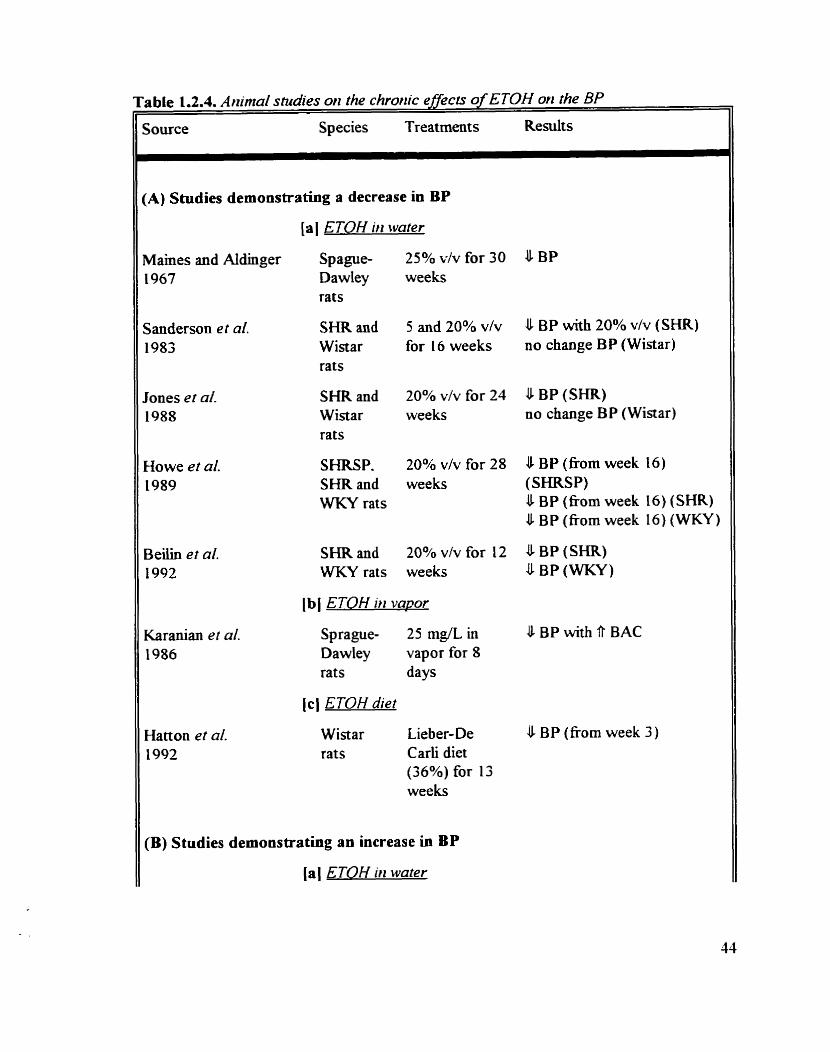
tource Species Treatments R e d t s
A) Studies demonstrating a decrease in BP
[al ETOH iri water
vlames and Aldinger Spague- 25% v/v for 30 8 BP
$anderson et ai. 1983
loues et al. 1988
Kowe et al. 1989
Beilin et al. I99Z
Karanian et ai. 1986
Hatton er al. 1992
Dawley weeks rats
SHR and 5 and 20% v/v Wistar for 16 weeks rats
SHRand 20%v/vfor24 Wistar weeks rats
SHRSP. 20% v/v for 28 SKR and weeks WKY rats
SHRand 20°/0v/vfor12 WKY rats weeks
[bl ETOH i ~ i vapor
Sprague- 25 mg/L in Dawley vapor for 8 rats days
Wistar Lieber- De rats Carli diet
(36%) for 13 weeks
8 BP with 20% vlv (Sm) no change BP (Wistar)
8 BP (SHR) no change BP (Wistar)
4 BP (fiom week 16) (SHRSP) 4 BP (fkom week 16) (SHR) & BP (fiom week 16) (WKY)
8 BP (SHR) 8 BP (WKY)
8 BP with h BAC
6 BP (fiom week 3)
(B) Studies demonstrating an increase in BP
[a ( ETOH iri water

Chan and Sutter 1983
Chan et al. 1985
Abdel-Rahman and Wooles 1987
Strickland and Wooles 1989
Hsieh et al. 1992
Vasdev et a/. 1993
Wist ar rats
W ist ar rats
Sp rague- Dawley and W ist ar rats
Sp rague- Dawley rats
Wistar rats
WKY rats
20% v/v for 12 T BP (Eom week 1) weeks
20% v/v for 12 f l BP (6om week 4) weeks (stressed and unstressed rats)
20% vlv for 12 h BP (fiom week 6) weeks
20% vlv for 22 ft BP (Eom week 4) weeks
15% v/v for 4 f i BP (fiorn week 1 ) weeks
5% vlv for 14 fi BP (fiom week 1 ) weeks
(C) Studies demonstrating no change in BP
[a 1 E TOH iti ivater
Khetarpal and SHR and 10% v/v for 1 1 no change in BP (Sm) Volicer WKYrats weeks no change in BP (WKY) 1979

d) Factors Nzfrz~etzcitzg the reszihs obserced in attinzai shcdies (Chrotzic E TUH)
1 ) Dehvdration: It has been argued that the antihypertensive effect of chronic moderate
ETOH consumption may not be the r e d t of alcohol per se but of secondary dehydration
because of the rat aversion to ETOH. Smail but significant reductions in fluid consumption
have been observed m a few studies (Khetarpal and Volicer. 1979: Howe et ai.. 1989: Beilin
et ai.. 1992). However. the administration of water for 1 dayiweek to offset any dehydration
caused by ETOH stiil resuited in the prevention of the age-dependent hypertension (Jones et
al., 1988). Moreover. the hematocrit was unaltered in ETOH-treated animals. suggesting no
modiications in plasma volume (Sanderson et ni.. 1983: Beilin et al.. 1992).
2) Nutritional deficit: The presence of nutritional deficit is unlü<eiy since most studies reported
no change in the body weight of the ETOH-treated animals (Khetarpal and Volicer. 1979:
Sanderson et ai.. 1983: Jones et d.. 1988: Howe et al.. 1989: Hanon et a!.. 1992). Sirnilarly.
cardiac mass was unaKected by the ETOH treatment ( Howe et al.. 1 989). Finally. in a study
where nutritional deficits were specifically avoided by the administration of a Lieber-De Cadi
ETOH diet. a significant antihypertensive effect of alcohol was still observed (Hatton et ai..
1992).
3 ) Stress: The surprisingly rapid and extremely significant increase in the BP afier less than
1 (Hsieh et ai., 1992; Vasdev et al.. 1993) or 4 (Chan and Sutter. 1983: Chan et ai.. 1985:
Strickland and Wooles. 1989) weeks of ETOH administration suggen a stress response to
the novel liquid mixture rather than a direct pressor effect of ETOH. The result of Vasdev et
al. ( 1993) with 5% viv ETOH is questionable, particularly the huge 20 (week 1 ) to 80 (week
I I ) mm HG BP increase. considering the low level of ETOH &en (tested with no success
by Khetarpal and Volicer in 1979) and the slow and subtle modikations in BP reported in
every other experiment.
4) Time-course of BP measurement: The timing of the BP measurement may be critical for
the pressor and depressor effect of chronic ETOH consumption. much like for repeated acute
ETOH administration when the initial hypotensive response is foliowed by a secondary
hypertensive reaction (Abe et aL. 1994). Similarly. Stricklmd and Wooles ( 1989) reported
that afiemoon BP calculation is associated with some hypotensive eEects. whereas moming

recordmg is hypertensive. suggesting a direct relationshrp between BAC levels and BP values.
5) Genetics: Genetic predispositions may also e.uplain some of the pressor actions of chronic
ETOH drinking. The antihypertensive effect of chronic ETOH consumption is stronger in
SHR rats (Jones et al., 1988; Howe et ai.. 1989: Beilin et al., 1992). in contrast, the
hypertensive or the lack ofhypotenske effect of chronic ETOH consumption is noted almost
esclusively in Wistar rats (Chan and Sutter. 1983: Sanderson et al.. 1983: Chan et al.. 1985:
Jones et al.. 1988: Hsieh et al.. 1992). uiterestingly. even the group of Chan casts doubts on
the possibility to produce a pressor effect with a difrent ntain of rats (Chan et al.. 1985).
e) Sunimary
A progressive general antihypertensive action of chronic moderate ETOH consumption
has been demonstrated in rats. although some pressor response to ETOH may e i a .
depending on the timing of the BP recording and the strain of animal used.
1.2.2.3. iVlechanisms
A few hypotheses have been proposed to explain both the pressor and depressor
components of chronic ETOH consumption. but the exact mechanisms remain unclear.
a) Mecht iisnis resporrrible for rhe bpertetsive eefjecr of chrotiic h e m ~ E TO ff cotrrmprioii
1 ) Withdrawal : The well documented pressor effect of replar large amounts of ETOH may
be the result of a secondary withdrawal reaction (Clark and Friedman. 1985: King er al..
199 1 ). It is possible that in regular heaw drinkers the repeated hypotensive effect observed
d e r each acute alcohol administration is followed by a compensatory hypertensivc reaction.
probably due to secondary alterations in vascular membrane charactenstics. In the long m.
wch a repetition of "intemittent withdrawal reactions" may produce a sustained state of
hypertension in alcoholics (Melville. 198 1 ). Partial validation of this hypothesis has been
obtahed recently from the group of Abe in short-term chronic experiments. Indeed. a single
administration of moderate ETOH m human subjects produced ody a direct depressor effect
measured by ambulatory BP. In contrast. repeated alcohol administration for a weeli

produced a secondaq pressor response. aiggesting the initiation of a short-term withdrawal
reaction (Abe et ai.. 1994). in most cases. lowering the alcohol consumption would reduce
the BP m alcoholic patients. demonstrating the reversibility of ETOH-induced hypertension
(Maheswaran et al., 1992: Ueshima et ai.. 1993).
2) Calcium: Cellular modifications in Ca" content have been W e d with alcohol-induced
hypertension (Somlyo and Souûyo. 1994). indirect evidence has shown that the pressor effect
of acute (Potter and Beevers 1984) and chronic (Arkwright et ai.. i982b: Hsieh et al.. 1992)
alcohol consumption may be due to the increased accumulation of Ca" in smooth muscle
ceils which causes an increase in the vascular tone. Exactly how this shift of Ca" from
extracelhdar to mtracellular space is achieved is uncertain. However. a nurnber of mechanisms
have been proposed mcluding (a) the inhibition of Ca2' ATPase activity (Sun. 1979). (b) the
mcreased Na' permeability . (c) the inhibition of NaT/K'-ATPase a c t ~ t y (Blaustein. 1977:
Arkwight et al.. 1984: Coca et ni.. 1992) and (d) the upregulation of voltage-activated Ca2'
channels (Littleton. 1988) foiiowing ETOH mtake and the intercalation of ETOH hto cellular
membranes.
3 ) Pressor hormones: Several pressor hormones. nich as catecholamine. cortisol and renin.
have been suggested to contribute to the high BP associated with chronic heavy ETOH
consumption. It has been proposed that ETOH could mcrease both theù levels and potentiate
dieu pressor action on the vasculature. Of particular interest is the hypothesis proposing that
the repetitive ETOH-mediated increases in circulating norepinephrine levels and
norepinephrine-dependent vasoconstriction may lead to a slow pressor effect of catecholamine
in long-terni alcohol drinkers (Aitura and Ahna. 1982; Ireland et ai.. 1984: Chan et al.. 1985:
Criscione et ai.. 1989). n i e elevation in norepinephnne niay also reflect enhanced actMty of
the sympathetic nervous system by ETOH exposure (Malhotra et al.. 1985: Beilin and
Puddey, 1992: R a n h et al., 1995). However. nich increases in plasma catecholamine levels
and sympathetic a c t ~ t y have been absent f?om other studies investigating chronic ETOH
consumption (Ibsen et ai.. 198 1 : Arkwright er al.. I98Za: Poaer and Beevers. 1 984: Hatton
et ai,. 1992). Similarly. inconsistent results have been obtained for cortisol and the renin-
angiotensin-aldosterone system (RAAS), so that the sigdicance of these modifications. if

any. seems linnted (Arkwright et ai.. 1982b: Potter and Beevers. 1984: Vasdev et ai.. 1993 ).
4) Others: A genetic predisposition to hi& BP and hi& alcohol consumption may esia in
certain i n d ~ d u a l s (Myrhed. 1974). The salt present in alcoholic beverages. particularly in
beers. has also been proposed as a mediator of the alcohol-induced hypertension (Puddey et
al.. 1986). ETOH may also be indirectly related to high BP in some cases since psychological
factors. such as stress, may predispose an i n d ~ d u a i to hypertension and alcohol drinliing
(Harburg et ai.. 1973: McQueen and Cellentano. 1982). In nich cases. alcohol is a way to
cope with detrimental psychological conditions rather than the cause of elevated BP.
b) Mechariisnts respomi b le for the arttihypertercsive e ffecr of chrorttc light a d moderute
ET0H coratrrnptiort
I ) Lipoproteins: The major hypothesis for the beneficial effect of low and moderate ETOH
consumption on BP and cardiovascular diseases involves modifications by ETOH of the
various Lipoprotein subfiactions such as hi&-density lipoprotein (HDL). low-density
lipoprotein (LDL) and very low-density lipoprotein (VLDL). HDL is imp licated in "reverse
cholesterol transport" fiom the vascular walls to the liver and is important in preventing
cholesteroi-derived v a d r damage (Gordon et al.. 1977: Gordon et al.. 1989). Ln contrast.
LDL has been positivety associated with hypertension and coronary heart disease (Castelli and
Anderson. 1986; Sachinidis et al.. 1990).
Early epidemiological and experimental audies have observed significant increases in
HDL levels after chronic alcohol connimption. even in low doses (CasteIli et ai.. 1977:
Barboriak et ni.. 1979; AUen and Adena. 1985: Langer et al.. 1992). Interesiingly. lower
threshold levels have been found in female subjects. explainhg perhaps to some estent the
greater incidence of U-shaped curves in women compared to men (Weidner et al.. 199 1 ).
However. the analysis of the two HDL nibfiactions. HDL, and D L , . have indicated that
modest ETOH consumption mcreases mody the levels of HDL-,, whereas HDL, is augmented
prirnady following chronic heavy ETOH consumption or in alcoholics (Haskeli er al.. 1981:
Moore and Pearson, 1986: Diehl et ai.. 1988). This is of importance since HDL was initially
believed to produce moa of the HDL beneficial effects on BP and cardiovascular disease

(Miller et aL. 198 1 : Ballantnie et a[. 1982). Fortwiately. in recent years. HDL, has also been
impiicated m these beneficial eEects ( H a 6 e r et al.. 1985: Langer et al. 1992). Furthemore.
various recent experiments have reported elevations in both HDL, and HDL, levels with light
and moderate alcohol consumption. confirming the hypothesis that HDL may indeed explain
parts of the hypotensive action of iow to moderate alcohol consumption (Miller et ai.. 1 988:
Hojnacki et ai.. 1988: Stampfer et al.. 199 1: Razai et al.. 1992: Rossouw et al.. 1992:
Gaziano et al.. 1993). A relative reduction in the LDUHDL ratio has also been found with
moderate. but not heavy. ETOH consumption in monkeys (Hojnacki et ai.. 1988) and with
the phenotic compounds of red wines (Frankel et al.. 1993).
2) Hemostasis and Thrombosis: Antithrombotic effects of alcohol have also been linked vliith
the beneficial actions of chronic moderate ETOH consumption and especially with the
reduction of coronary artery disease. Long-term consumption of modest amounts of alcohol
have been associated with lower plasma fibrinogen levels and with increased fibrinolytic
a c t ~ t y . assessed by endogenous tissue-type plasminogen activator (t-PA) (Maede et al..
1 979: Veenstra et ai.. 1990: Ridker et al.. 1 994). A reduction in platelet aggregation has also
been demonstrated with chronic ETOH drinking (Elmer et al.. 1984: Pikaar et al.. 1987:
Rubin and Rand. 1991). The hypothesis tliat alcohol interferes with hemostasis and
thrombosis is funher supported by studies indicatmg that acute and short-term chronic alcohol
coosumption decreased TXAz release fiom platelets and increased PGI, synthesis f?om
endothelial cells (Mikhailidis et al., 1983: Landolfi and Steiner. 1984: James and Walsh.
1985).
3) Others: The non-specifk association of ETOH with the cellular membranes produces
widespread modifications m basic cell physiology and therefore m the communication between
cells and between organs. Not-surpnsingly (section 1.1.5.2, ). most aspects of endocrine
physiology. such as hormone production. release. receptor Ievels or second messenger
systems. are altered by alcohol and the various homeostatic systems regulated by these
endocrine agents. such as the BP. are likely to be modified accordingly. Several very general
homonal systems with vascular. rend and central componeats nich as RAAS, catecholamine
or AVP, have been associated with pressor responses and positive regdation of BP

homeostasis. Accordin& experimental studies have tried to link such systems with the hi@
ETOH-induced hypertension (section 1.2.2.3. pan 3). The absence of a seneral and
widespread hormonal family with hypotensive actions and a negative regulation of BP
homeostasis prevented early investigations on the humoral contribution to the beneficial
effects of light and moderate ETOH connimption. Nevertheless. a few reports have noted the
attenuation of the pressor effect of acute ETOH in certain vascular beds foilowing chronic
ETOH chking, mediated by the increased actMty of local vasodilating factors. such as NO
and PGIz(Karanian et aL. 1985: Knych. 1992: Hatake er aL. 1994). However. in the lan 15
years the discovery of an expanding family of natriuretic and vasorelaxant peptides opened
a new avenue to investigate the mechanisms responsible for the hypotensive effects of low
ETOH consumption. Much like the hypothesis of a slow pressor effect of epinephrine
following heavy ETOH consumption. a slow hypotensive action of "natnuretic peptides"
mediated by vascular. central and renal modifications may exia in chronic moderate ETOH
drinkers. leading to a sustain antihypertensive effect. The report by Colantonio et al. ( 1 99 1 )
of a significant mcrease in circulating atrial natriuretic peptide (ANP) levels. the fin member
of this natriuretic peptide family. after acute administration of modest amounts of ETOH
further demonstrated the need of designing eqeriments to survey the possible implication of
the natriuretic peptide system in the cardiovascular effects of acute and chronic ETOH
consumption.
1.3. NATFUURETIC PEPTLDE FAMILY
During the paa 15 years. a new hormonal system for the negative regulation of total
body fluid and BP homeostasis has been identified and characterized. Three major members
have been isolated. The first two. the A- and B-type natriuretic peptides (ANP and BNP) are
located mainly m the heart tissue. whereas the third one, the C-type natriuretic peptide (CNP).
is found mainly in the CNS. Their major biological activities include natriuresis. diuresis.
vasorelaxation and the mhiiition of several pressor hormones. These physiological effects are

mediated by the association of the natriuretic peptides with three specific natriuretic peptide
receptors. Interactions of the natriuretic peptides with the &st two natriuretic peptide
receptors (NPR-A and NPR-B) stimuiate the production and accumulation of guanosine Y.5'-
monophosphate (cGMP). cGMP appears to be the second messenger system of the natriuretic
peptide farnily. Interactions of the natriuretic peptides with the third natriuretic peptide
receptor (NPR-C) does not stimulate the production of cGMP. Therefore. NPR-C is moaiy
considered to be a clearance receptor.
1.3.1. A-TYPE NATRIURETIC PEPTIDES
1.3.1.1. Atrial natriuretic peptide
a) Historrcal perspecrives
Modifications in body fluid volumes and therefore in BP have long been known to
control the secretion of sodium and water by the kidney (Starling. 1909). However. the
humoral aspect of this "pressure-natriuresis" rnechanisrn (Guyton. 1980) escaped investigation
until relatively recently. in 1956. Kisch noted the presence of granules in atrial cardiocytes.
an obsenration that wodd open a new concept. the heart as an endocrine organ (Braunward.
1964). These granules were later better described by Jarnieson and Palade ( 1964). who
suggested for them a secretory role. In the next decade. more specific expenments were
performed. These studies indicated that atrial distention produced diuresis. whereas it
decreased circulatory renin and AVP levels (Brennan et al.. 197 1 ). In 1976. Marie and
colleagues obsewed that atnal granulanty was increased by salt and water load. This
"suggested a relationship between atnal specific granules and the replation of water-
electrolyte balance ..." (De Bolci, 1979). The crincal last step was made in 198 1 when De Bold
et al. ( 198 1 ) demonstrated that the mjection of atnal extracts in aneçthetized rats led to rapid
natrkesis (30-fold mcrease) and diuresis ( IO-fold increase) and to a sigruficant fa11 in the BP.
The atrial natriuretic peptide (ANP) was found.

b) Biochemisby and release
The ANP gene consists of 3 exons (codmg regions) and 2 mirons (interverhg sequences)
(Greenbug et al., 1984; Nemer et al.. 1984; Gardner et ai.. 1988). In bumaas and rats, the
first exon encodes for the 24 aminoacid signal peptide and the £ira 20 aminoacid of the
prohormone. The second exon encodes for the remainder of proANP, with the exception of
the laa (in humans) or three laa (in rats) residues, coded by the third exon (figure 1.3.1.).
Translation of ANP mRNA produces the aaual 152 aminoacid preproANP precursor (Cwie
et al., 1983; Oikawa et al. , 1984). The removal of the 24 aminoacid signal sequence and the
two carboxyl (C)tterminal ar@e ( k g ) residues fomd in rats, but not in humans, @es nse
to the 126 aminoacid proANP (Flynn et al.. 1983; Atlas et al.. 1984). The prohomone was
iater found to be the major storage form of the peptide m atrial granules (Bloch et al.. 1985:
Thiiauit et al., 1987), m contrast to most other hormones which are stored m their active
fom. It is believed that membrane-bound enzymes cleave the prohormone either upon or
immediately afier secretion (Thiibault et al., 1986; Sei et al., 1992). The final proteolytic
processing is made behiveen arginine (Arg), and serine (Ser), forming the mature C-terminal
28 aminoacid ANPw931, identified in the circulation (Gutkowska er al.. 1984; Tanaka et al..
1984; Theiss et al.. 1987) (figure 1.3.2.). Even though ANPw-12, is the main C-terminal
peptide f o m processed fiom proANP, cleavages of proANP at other atypical sites have been
detected in other tissues. For example, amino (Nkelongated forms of ANP are found in the
kidney (ANP,,,,) (see section 1.3.1.2.) (Schulz-happe et al.. 1988) and the testis (ANP,,
2 6 ) (Pandey and Orgebm-Cria . 199 1 ), while shorter processed forms of ANP have been
identzed in the brain ( A N P l o ~ l x and ANPI,-,,) (Shiono et al.. 1986; Ueda el al., 1987).
A common characteristic of the C-terminal natriuretic peptides is the presence of a
-hide bridge, which is essential for biological aaMty (Misono et al.. 1984). Interestmgly,
the structural similarity between most of the natriuretic peptides resides withm the 17
aminoacid ring structure formed by the disulphide bridge. in fact, among the various known
natriuretic homones (ANP, BNP and CNP), 1 1 aminoacid of the ring structure are identical
(Koller and Goeddel, 1992). Among species, the ANP amhoacid sequence is also remarkably
well conserved. Betweeo humans and rats for example, mature ANP is identical, with the

5ut
ANP mRNA
PreproANP
fragment
Exon 1 Exon 2 Exon 3
BNP gene I Sut
BNP mRNA V
PrepraBNP ! I -26
ProBNP
N-terminal fragment B N P-45
CNP gene
CNP
Exon 1 Exon 2 I
mRNA Y
PreproCNP 1 I -23
ProCNP I
N-terminal fragment 1 1 O3
CNP-53
F ig ure 1.3.1 . Schematic representation of the transcription, translation and processing of the natriuretic peptide family
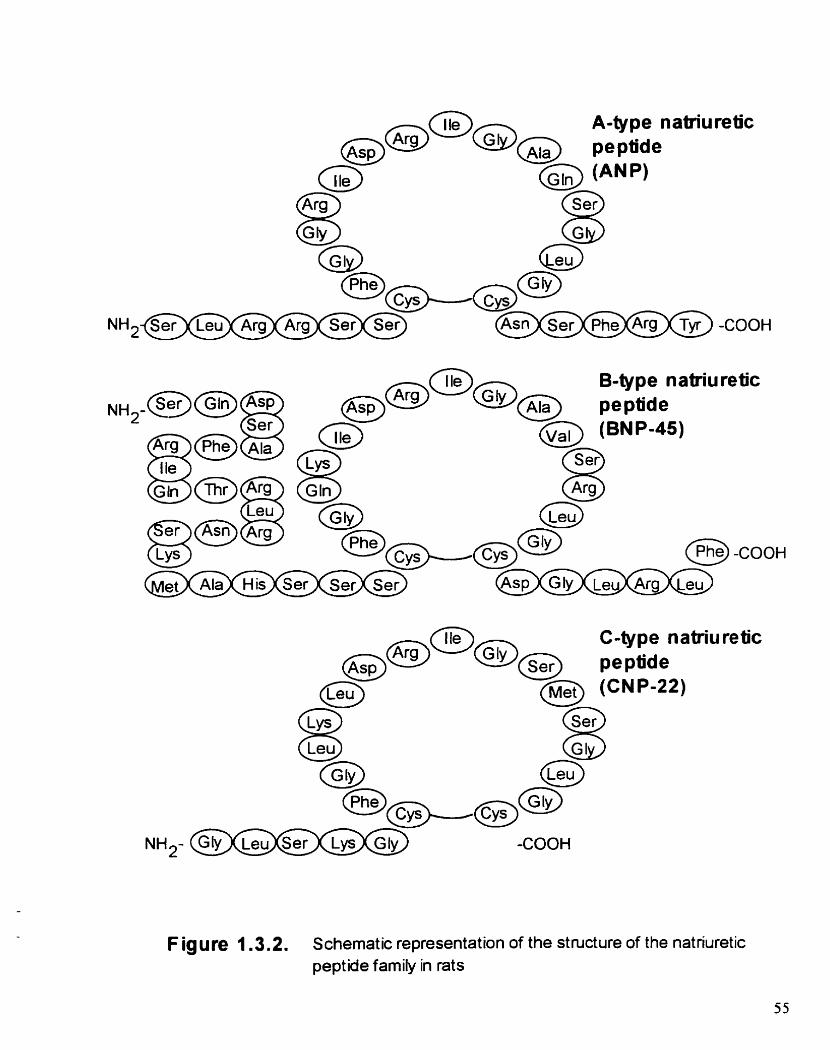
A-type natriuretic
B-type natriuretic
F ig ure 1 3.2. Schematic representation of the stmcture of the natriuretic peptide famiîy in rats

exception of one substitution at position 1 10. fkom isoleucine (Ile) (humans) to methionine
(Met) (rats) (Kangawa and Matsuo, 1984).
The major stimulus for the atrial release of ANP appears to be atnal stretch (Haass et al..
1987: Greenwald et al.. 1989: Ruskoaho. 1992: Espmer. 1994). Therefore. factors promotmg
atnal distention by volume expansion. such as head-out water immersion (Katsube et al..
1985: Anderson et al.. 1986). changes in body posture (Solomon er al.. 1986). increased
heart rate (Schielbmger and Lmden, 1986) or elevated dietary sodium intake (Richards et al..
1986) increase circulating ANP levels. Similarly. plasma ANP levels are also elevated in
pathophysiological conditions associated with increased atrial filling pressure nich as
congestive hean failure (Cm) (Burnett et al.. 1986: Tsunoda et al.. 1986) or hypertension
(Gutkowska et al.. I986a: Morü et al.. 1986a: Sagneila et al.. 1986: Hollister and Luagami.
199 1 ; Naruse et al.. 1994). Other stimuli have been associated with cardiac ANP release.
indeed, vasoconstrictor hormones such as AVP (Itoh et al., 1987: houe et ni.. 1988 ).
angiotensin II (Diets 1988 ) or endotheiin (Schielbinger and Greening, 1992) are found to
affect ANP production and release. Glucocorticoids (Gardner et al.. l986a). testosterone
(Deng and Kaufkaa 1993 ) and thyroid hormone (T,) (Argentin et al.. 1987) accelerate ANP
secretion. Opioids (such as O-endorphin) are also possibly involved in the regulation of
cardiac ANP release (Voilmar et al., 1987: Widera er al.. 1992).
C) Tissue distribzrtiorr
As implied by its name. ANP was first isolated fiom atrial tissue (Flynn et al.. 1983).
Early radioimmunoassays demonstrated higher total ANP content in the right compared to
the left atrium of rats (Gutkowska et al.. 1984; Gutkowska et al,. l987a). As was previously
discussed. ANP is stored m the atrial granules in its higher molecular weight form (proANP)
and is released upon atrial aretch. Most of the ANP in circulation originates fiom the hean
atria. However. ANP immunoreact~ty and transcripts have also been found although in
srnailer quantities, in several extra-atrial tissues (Gutkowska and Nemer. 1989).
Heart ventricular tissue has been reported to contain ANP mRNA and proANP (Nemer et
a[. 1986). Morphological (absence of granules) and secretory differences between the atria

and the ventricles have however suggested that contrary to the atria. m the ventricles ANP
is not stored in granules but it is released in a constitutive rnanner (Bloch et al., 1986). In
view of the relative large size of the ventricles and the conthuous secretion of ventricular
ANP. the ventricles contniute substantially to circulatmg ANP levels. This contribution is
particularly important in pathophysiological conditions associated with ventricular
hyperirophy. such as congestive heart failure (Cantin et ai., 1988) or hypertension ( Arai rr
ai.. 1988: Ogawa et al.. 199 1 ). in such cases. the ventricular tissue is considered as the major
source of plasma ANP levels.
ANP. proANP and ANP mRNA are also present in the lungs (Sakamoto et ai.. 1986:
Gutkowska et al.. 1987~: Gutkowka et al.. 1989). Within the respiratory syaem ANP has
been localized by various techniques in the respiratory epithelium (Gutkowska and Nemer.
1989), pleural fluid (Vesely et al.. 1989) and pulmonary vems ( SpringalI et al.. 1 988). Marked
elevations in plasma ANP levels and in lung ANP content and transcnpt are observed in
cardiomyopathie hamsters. which exhibit decreased atrial ANP production (Currie et ai-.
1987: Gutkowska et al.. 1989). These observations suggest that as was observed for the heart
ventricles. under certain pathophysiological conditions. the lmg may enhance the production
and secretion of ANP. contributhg to the elevated plasma ANP levels in the presence of
diminished a c t ~ t y of the heart ANP system. Furthemore. a paracrine role for ANP in
pulmonary homeostasis has been proposed (Perreault and Gutkowska. 1995).
Extensive localization of ANP immunoreact~ty has been identified in the central (CNS)
and peripheral (PNS) newous systems (Kawata et al.. 1985: Standaert et al.. 1986). n i e
highest concentrations of ANP (Morii et ai., 1985) and ANP mRNA (Gundlach and Knobe.
1992) are found in the hypothalamus. and particularly in areas involved in the replation of
fluids and BP. such as the anteroventral third ventricle region (AV3V) (Ku and Zhanp 1994).
Other brain areas where immunoreactive ANP has been detected include the midbrain.
thalams. septum, spmal cord oi£àctory bulbs and mucosa. pons and medda (Kawata et a/-.
1985 ; Gutkowska and Marcinkiewicz, 1989; Gutkowska et al., 199 1 ). ANP is also seen in
the cerebrospmal fluid (Manuno er al.. 1987). Atypical processing of proANP in the CNS is
suggested by the isolation of shorter 24 and 25 aminoacid ANPs (Ueda er al.. 1987). In the

PNS. ANP has been detected in the autonomie system, panicularly in the m a t h e t i c and
parasympathetic ganglia (Debinski et al.. 1986).
Both ANP and ANP mRNA are found in the antenor pituitary (Gardner er ai.. 1986b:
Gutkowska and Cantïn. 1988). Local modulation of pituitary hormones by ANP has been
posnilated. in contraa. only mature ANP is present m the posterior puititary. suggesting an
exogenous production. possibly fiom the hypothalamus (Gutkowska et al.. 1987b).
The chromaffm ceüs ofthe adrenal meduila have been reported to possess both proANP
and ANP (McKenzie er al.. 1985: ûng et al.. 1987). The possibility of local ANP rynthesis
is also hypothesized in the vasculanire. notably in the aortic arch (Gardner et al.. 1987). the
gonads (Gutkowska et al.. 1993) and the GI tract (Vuolteenaho et ai., 1988). because of the
presence of the hi@ (proANP) and low (ANP) molecular weight forms.
Small quantities of ANP (Figueroa et al.. 1990) and ANP mRNA (Greenward et al..
1992) are found throughout the renal nephron. The added presence of an elongated N-
terminal form of ANP (ANP,,,) (see section 1.3.1-2.) in the kidney indicates a local role for
the A-type natriuretic peptides in this organ (Goetz 199 1 ).
d) Me rabolisnt
ANP has a shon half-life in plasma, ranging fiom Iess than a minute in rats (Katsube er
ai.. 1986: Krieter and Trapain. 1989) to less than 5 minutes in humans (Gnadinger et ol..
i 986: Biollaz et of.. 1987). The rapid removal of ANP fiom the circulation has been shown
to occur primarily in the kidney (Needleman et al.. 1989) with lesser contributions of otlier
ogans. such as the her. lungs and mtestine (Crozier et al.. 1986: Krieter and Trapain. 1989:
Gerbes and Vollmar, 1990). Two major processes. one proteolytic and the other receptor-
mediated have been reported in ANP clearance. Proteolytic degradation is rnoaly the result
of peptide cleavage wittiin the ring of ANP by endopeptidase 21.1 1. yielding inactive products
(Stephenson and Kenny. 1987: Seymour et al.. 1991 : Kanazawa et ai.. 1992). Endopeptidase
24.1 1 is a membrane-bound enzyme disaiiuted primady in rend glornenili and on the luminal
side of the proximal tubules (Olins et ai.. 1987: Erdos and SkidgeL 1989; Landry et al..
1993). It is also eqressed in vascuiar tissues (Johnson et al.. 1990: Dussaule et ab. 1993 ).

Receptor-mediated clearance of ANP is beiieved to be through NPR-C (see section
1.3.1.4.3.). the so-called "clearance receptor" (Maack et ai.. 1987). Upon ANP binding. the
receptor- ligand complex is mtemalized and ANP is degraded by lysosornal hydrolysis
(Nussenzveig et ai.. 1990: Maack et ai.. 1993 ). The abundance of NPR-C in the vasculature.
kidney. b r a h lungs heart and adrenals M e r supports for a specific role of this receptor in
ANP metaboiism (Nakao et ai.. 1992: Maack et ai.. 1993 ).
1.3.1.2. Urodilatin (UD)
Lately. the atypical processing of proANP in the kidney has attracted attention. Schulz-
Knappe et al. m 1988. isolated from human urine a 32 aminoacid peptide who tumed out to
be a 4 aminoacid N-terminal extension of mature ANP. ANP,,_,,, - .. - or urodilatin (UD) as it
is now called - is absent Ciom the circulation ( D r u m e r et ai.. 199 1 ). It has been postulated
that the distal tubular nephron is the major. if not the only. site of synthesis for UD in the
body (Goetz 199 1 : Forssmann. 1995). Low levels of gene expression (Greenward et ai..
1992) and proANP (Ritter et ai.. 1992) have been identified in this tissue. suggeaing local
synthesis. Therefore. UD may be released fi-om the cells of the distal tubule and interact with
the luminally-situated natriuretic receptors (see section 1.2.3.1.) of the inner medulla
collecting ducts (IMCD) (Gunning and Brenner. 1992: Valentin et al.. 1993). Aiternatively.
the lridneys may represent only a distinct processing site for exogenous proANP. although the
absence ofthe ANP precursor m the circulation renders this hypothesis unlikely (Gutkowslia
et al.. 1984; Forssmann. 1995). Experimental evidence has demonstrated Iittle or no storage
space in renal tubular cells. suggeaing a constitutive rather than regulated mode of UD
release (Ritter et al.. 199 1 ). The regulation of UD secretion is still unclear. Initially. a neural
control was pormlated (Emmeluth et ni.. 1992). However. the demonstration of unaltered UD
release in denervated kidneys suggested the presence of an humoral regulation as well
(Emmeluth et al.. 1993 ). The clearance of UD is done mostly through urine excretion. In
contraa to ANP. üD appears to be quite resistant to proteolytic degradation by
endopepidase 24.1 1 (Gagelmann et al.. 1988). Recently, a paracrine role has been attributed
to CID for the tubular diuretic and natriuretic effects previously ascribed solely to circulating

1.3.1 3 . N-terminal frarmen ts of proANP
It is a common occurrence m endocrinology that several different hormones may
originate ficm a single prohormone. As a example of this, proopiomelanocortm (POMC) is
the precursor of at Ieast 4 known peptides, includhg adrenocorticotropic hormone (ACTH)
and B-endorphin (Mains et al.. 1977; Eipper and Mains, 1980). Recent aidence has
suggested a nmilar muhihormonal role for the ANP precursor (Vesely, 1995). Indeed 3
distinct N-terminal peptides are derived from the first 98 aminoacid of proANP, nameiy 1 )
ANPI-, or "long acting sodium stimulator", 2) ANP,,,, or "vesse1 dilator" and 3) ANP,,,
or "kaliuretic stimulator" (Vesely et al.. 1987; Martin et al.. 1990). The 3 new peptides lack
the characteristic 17 aminoacid ring structure associated with other natriuretic peptides
(YandIe, 1 994). Nevertheless, N-teminal ANP molecules have been found t O produce similar
natriuretic, diuretic and BP lowering effects as C-terminal ANP (Martm et al., 1990: Vesely
et al.. 1994). Whereas the hal maturation step for ANP is believed to occur on or just afier
Rs release fiom hem atria, for N-terminal ANP molecuies a M e r proteolytic process may
occur within the circuiation to form the 3 N-terminal peptides (Vesely, 1995). Specific RIAS
toward various portions of N-terminal ANP molecules have demonstrated that ANP,.,,,I
ANP,,, and ANP,,, indeed circulate as distinct peptides (Winters et al.. 1988: Gower et
al., 1994). The release ofthe N-terminal ANPs is reguiated Ma the same stimuli as ANP. such
as atrial stretch and rapid heart beat (Ngo et al., 1989; Dietz et al.. 199 1). The tissue
distnibution of N-terminal ANPs is dl under investigation but should parallel to some exqent
the tissue distribution of ANP. Thus, the atrial tissue is certainiy their major site of spthens
(Vesely et al., 1992). ANP,,, and ANP,,,, have been localized in the kidney, particuiarly in
the p r o d tubule @mirez et al.. 1992; Saba et al.. 1993). Immunofluorescent studies have
also demonstrated the presence of the same two peptides m cholinergie neurons of the CNS
(Veseiy et al., 1992). in the circulation, ANP,,, and AN&,,, are found m concentrations 10
to 24 times higher than that of ANP, whereas plasma Ievels are approxirnatively 3
times higher than those of ANP (Winters et al.. 1988; Vesely et al.. 1994). Ln addition, the

vanous half-hes cf the N-terminal peptides are about 15 times longer than the half-live of
ANP (Achemian et ai.. 1992), a situation accounting in part for their much geater
concentration in circulation than that of the classical natrîuretic peptides (ANP. BNP and
CNP) (Vesely. 1995). The exact clearance mechanism is not clear. although it appears to be
independent corn clearance receptors (Vesely. 1995). Much aiil has to be leamed fiom N-
terminal natriuretic peptides. but their elevated plasma levels and longer half-lives may
suggea prolonged effects. in contrast to the acute nature of the C-terminal natriuretic
peptides responses.
1 .3.t. B-TYPE NATRrURETIC PEPTIDE
1.3.3.1. Brain natriuretic ~ept ide
a) Biochentistp artd releuse
The BNP gene consists of 3 exons and 2 introns (Roy and Flynn, 1990: Steinhelper.
1993). Like ANP. moa of mature BNP is encoded by the second exon (figure 1.3.1.)
(Seilhamer et al.. 1989). In addition. the BNP gene contains an ATTTA motif in the 3'-
untranslated region. a motif absent from the ANP gene. This sequence has been implicated
wRh mRNA instability and suggests a differential transcriptional reepiation between the ANP
and BNP genes (Davidson and Struthers. 1994: Ogawa et al.. . 1995). Post-transcriptional
dserences have also been noted between the two natriuretic peptides. There is a lower
structural homology for BNP and proBNP between species than for ANP and ANP
precursors. indicating interspecies diversity (Lang et al.. 1992: Datidson and Struthers.
1994). In human cardiac tissue. BNP mRNA produces a 134 aminoacid precursor which
becomes. after the removal of the 26 aminoacid signal peptide. the 108 aminoacid proBNP
(Sudoh et al.. 1989: Seilhamer e t al.. 1989). A shorter coding sequence for the second exon
of rat BNP gene gives rise to a 95 aminoacid BNP precursor for this specie (Roy et aL.
1990). A final proteot)ac cleavage imrnediateiy after an Arg-X-X-Arg sequence produces the
C-terminal mature BNP fom with the characteristic 17 aminoacid ring structure of natriuretic
peptides (Steinhelper et al.. 1993). Interestingiy. there is a greater sequence h o m o l o ~

between ANP and BNP than between the BNPs of various species (Yandle. 1994).
Consequently. a 32 aminoacid f o m of BNP circulates in humans (Sudoh et al . 1989). a 45
aminoacid form m rats (Aburaya et al.. 1989a) (figure 1.3.2. ) and a 26 aminoacid form in pigs
(Aburaya et ai.. 1989b). To date. no experiments have investigated the proBNP portion of
the peptide. although its greater interspecies sequence bomology than the C-teminal BNP
(Stemhelper er ai.. 1993) and the elevated plasma lwels of the N-terminal proBNP (Tateyama
et al.. 1992: Yandle et ai.. 1993 ) may suggest same biological fùnctions. much like for the
newly identified proANPs.
BNP was originaily isolated fiom the porcine brain (Sudoh ft al.. 1988). However.
subsequent experiments have indicated that the major site of BNP synthesis. in hurnans. pigs
and rats is the heart rather than the brain (Minamino et al., 1988: Saito et al.. 1989:
Mukoyama er al.. 199 1 ). Withui the heart itself. the ventricles rather than the atria appear to
be the main site ofBNP production. Although about 100-fold higher concentrations of BNP
are found in atrial tissue. 6096 of the total BNP secretion is still observed after the
experimental removal of the atna. comparatively to onIy 50h for ANP (Ogawa et ai.. 199 1 ).
Pathophysiological conditions associated with hypertrophied ventricles tùrther accelerate the
synthesis and secretion of BNP fiom this tissue (Kohno et ai.. 1 992a: Dagnino er ai.. 1992:
Yokota et ai.. 1993). The scarcity of ventricular granules suggeas a constitutive mechanism
of secretion for BM? much like for ventricular ANP (Lang et al.. 1992). in the cardiac
atrium BNP is colocalized with ANP in granules (Lida et al.. 1990: Hasegawa et al.. 199 1 :
Thibault et al.. 1992). However. the storape foms of BNP in atrial tissue Vary amongst
species. fiom mature BNP in rats (Thibault et ai.. 1992) to a misture of BNP and proBNP
in humans (Takeyama et ai.. 1990) or mainly proBNP in pigs (Minamino et d.. 1988).
Because of the ventricular l o c ~ t i o n of BNP within the heart. its mechanisrn of release
depends more on left ventricular end-diastolic pressure than on atrial stretch (Nakamura et
ai.. 1992: Richards et al.. 1993 ). Thus. stimuli favouring elevated BP (Lang et ai.. 199 1 :
Yokota et al.. 1993) or supraventricular tachycardia (Kohno et al.. 1992b) have been
demonstrated to increase heart BNP release. Humoral factors such as endothelin- 1. but not
AVP or angiotensin iI . have also been shown to modulate BNP secretion (Hirata et ai., 1990;

Horio et al.. 1992). Nevertheles the avial portion of BNP colocalized with ANP m granules
is still repiated by atrial distension (Kohno et al.. 1992b).
6) Tisstre distribzrtiotr
As previously discussed the cardiac tissue and particularly the heart ventricles appear to
be the most prominent site of BNP synthesis and release in the body (Mukoyama et al. .
199 1 ). Whereas BNP is present in the porcine brai.. mainly in the bramstem the spinal cord
and the hypothalamus (Ueda et al.. 1988), ody smail amounts of this natnuretic peptide have
been detected in the rat and human brain. with the exception of the spinal cord where
si@cant levels are found (Ogawa et al.. 1990: Nakao et al.. 1990: Hira et al.. 199 1 ). BNP
and proBNP (also termed "aldosterone secretion inhiiitory factor or ASIF") have been
localized m adrenal chromaffin cells and proposed to locally modulate aldosterone secretion
(Nguyen et ai.. 1989a.b). The lung and the aorta have also been found to contain the BNP
gene (Dagnino et al.. 199 1 ). ûther peripheral tissues (e.g. GI tract) have been shown to
possess BNP imrnunoreacthdy. but local biosynthesis is still uncenain (Sharkey et ai.. 199 1 ).
in the circulation. BNP levels are usually about 50h of those of ANP (Takeyama et al.. 1992).
although they may equal or surpass ANP levels in pathophysiological conditions such as heart
failure. largely because of ventncular hypenrophy (Mukoyama et al.. 199 1 ).
C) Metabolisnt
Although the structural similanty between the A- and B-type natnuretic peptides suggest
similar metabolic pathways. a longer haü-live for BNP than for ANP has been reported iii
plasma (Holmes et a/.. 1993). Resumably. this ditference is the result of the lower affuiity of
BNP to the clearance receptors (Mukoyama et ai.. 199 1: Suga et al.. l992a). It may also
resuit fiom a slower hydrolysis of BNP tban ANP by the enzymatic pathway of endopeptidase
24.1 1 (Kenny et al.. 1993).

1-3.3. C-TYPE NATRILJRETIC PEPTIDE
1.3.3.1. C - t y ~ e natriuretic peptide
a) Biochernktry and releare
The CNP gene is composed of at lean 2 exons and 1 intron (figure 1.3.1 .) (Tawaragi et
01.. 1990. 199 1 : Ogawa et ai.. 1992). Exon 1 codes for a 23 aminoacid signal peptide and for
the hst 7 aminoacid of proCNP, the rest of which is encoded by evon 2 (Yandle. 1994). As
for BNP. several ATTTA motifs have been identified fiom the 3'-untranslated region of rat
cDNA. suggesting mRNA denabilkation (Kojima et al.. IWO). The cleavage of the signal
peptide fiom the 126 aminoacid preproCNP results in the production of the 1 O3 aminoacid
proCNP. interehgly, CNP precursors have the highest interspecies sequence homology of
al1 natriuretic peptides (above 9 6 O i o h o m o l o ~ between rat. pig and human proCNPs) (Kojima
et ni.. IWO: Tawaragi et al.. 199 1 ). As for the A- and B-type natriuretic peptides. the mature
fonns of CNP. a 22 aminoacid (CNP-22) and a 53 aminoacid form (CNP-53). are contained
in the C-terminal end of the prohormone. CNP-53 appears to be the predominant form in
mon tissues (Yandle. 1993). The ha1 cleavage is believed to occur immediately afler an Are-
X-X-Arg sequace 8 proCNP for CNP- 5 3 and on a dibasic Lys-Lys sequence. direct- f?om
proCNP or from CNP-53. for the CNP-22 (Yandle. 1994). Both biologicaily active forms of
CNP exhibit the characteristic 17 aminoacid ring structure and disulphide bridge. but lack the
C-terminal Mig extension present in ANP and BNP (figure 1.3.2.) (Sudoh et nl.. 1990). As
for BNP. the production of additional biologically active peptides fiom the N-terminal portion
of proCNP awaits investigation.
CNP was originally discovered in the porcine brain (Sudob er al.. 1990). in contrast to
ANP and BNP which are located prùnarily within the heart. CNP is virtually absent fiom the
cardiac tissue and located almost exclusively in the CNS (Komatsu et al.. 199 1: Minamino
et ai.. 199 1 ). A role for CNP as a neurotransmitter has been postulated. The regdation of
CNP synthesis and release, particularly in the CNS, remains unclear (Espiner. 1994).

b) Tissue distributiorl
CNP is the most prevalent of the 3 natriuretic peptides in the brain ( M m a e o et ai..
199 1 : Togashi et al.. 199 1 ). Within the CNS. its regional distribution cliffers From that of
ANP. Species variations have also been observed. in the rat. CNP immunoreact~ty is the
highest in the cerebellum foilowed by the hypothalamus. thalamus. septum and olfactov
buibs (Komatsu et al.. 199 1 ). whereas in the human brain the highea CNP concentration is
found in the hypothalamus. followed by the rnidbrain and the thalamus (Komatsu et al..
199 1 ). CNP mRNA has also been detected in the CNS. notably in the anteroventral preoptic
area (AV3V) and the arcuate nucleus ofthe hypothalamus (Herman et a/.. 1993). Some CNP
immunoreact~ty has also been detected m other areas of the nervous system such as the
spinal cord (Ueda er al.. 199 1 ). the anterior and posterior pituitary gland (Komatsu er al..
199 1 ) and the CSF (Kaneko er CIL, 1993).
CNP immunoreact~ty (Suga et al.. 1 992b: Stingo et al.. 1992: Heublein et al.. 1992)
and gene expression (Komatsu et al.. 1992a) have been reponed in vascular endothelial cells.
providing the concept of a "vascular natriuretic peptide system". The modulation of CNP
secretion by V ~ ~ O U S growth factors and cytokines. nich as transfonning growtth factor4
(TGF-B). tumor necrosis factor-a (TNF-a). basic fibroblast growth factor (bFGF).
mterteukin- 1 and thrombin. suggests a role for the CNP-mediated vasorelalration and growth
inhibition in vascular remodelling ( Suga et al., 1 992b. 1 993 ).
Smaller concentrations of CNP have been found in the kidney (Komatsu et al.. 199 1 :
Suzuki et al.. 1993). The recent identification of CNP trmscript in various nephron segments
by PCR experiments suggests the possibility of a local role for the C-type natriuretic peptide
in this tissue (Terada et al., 1994).
The presence of CNP within the heart itself remains controversial. The low
concentrations of atrial and ventricular immunoreactive CNP detected by early eqeriments
have been shown to be due to the cross-react~ty with the high cardiac concentrations of
ANP and BNP (Minamho et al.. 199 1 ). However. the recent demonstration of CNP mRNA
gene expression m the hean atria and ventricles may indicate a small paracrine role for CNP
m the heart (Vollmar et ai.. 1993). particularly in pathophysiological conditions (Wei et al..

i 993 ).
CNP and CNP mRNA have also been detected m the adrenal chromaffin cells (Babinski
eral., 1991).
c) ~letuboiisnz
Very low CNP levels are present in the plasma. in agreement with its central rather than
peripheral localization m the body (Mmamino et ni.. 199 1 : Stingo et al.. 1992). Nevertheless.
it has been s h o w that of the 3 natriuretic peptides. CNP is the best substrate for
endopeptidase 24.1 1 (Kenny et ai.. 1993). Moreover. the binding atfinity of CNP to the
clearance receptor. although lower than that of ANP. is higher than the binding affinity of
BNP (Suga et al.. 1992a).
1.3.4. NATRHJRETIC PEPTIDE RECEPTORS
1.3.41. Guanvlate-cvclase (CC1 receotors (NPR-A and NPR-B)
The guanylate-cyclase (GC) receptors consist of an extracellular igand-binding region.
a single traosmembrane domain and two mtraceliular regioos. a kinase homology domain and
a GC catalytic domain (figure 1.3.3.) ( KoUer and Goeddel, 1992: Nakao et ni.. 1992: Maack
er ai.. 1993). The gene coding for the GC receptors contain 22 exons and 2 1 introns in the
rat (Yamaguchi et al.. 1990). The fùn 6 exons encode for the extracellular portion of the
receptors. Exoo 7 represents the membrane-spanning region. The intracellular portion of the
receptors is separated in two. with exons 8 to 15 encoding for a protein-kinase like domain
and exons 16 to 22 for the GC domain. The transcription of the GC receptor genes results in
the production of two distinct giycoprotein receptors structurally M a r . a 1057 aminoacid
NPR-A and a 1047 aminoacid NPR-B (Schulz et al.. 1989; Chang et al.. 1989: Chinkers c f
al., 1989). The second messager guanilate Y. 5'-monophosphate (cGMP) is produced upon
peptide bmding to the extracellular region by the activation of the GC catalytic domain (Kuno
et al.. 1986: Shmg et ni., 1988). Most of the pbysiological actMties of the natriuretic peptides
are believed to occur via the production and cytoplasrnic accumulation of cGMP. The kinase
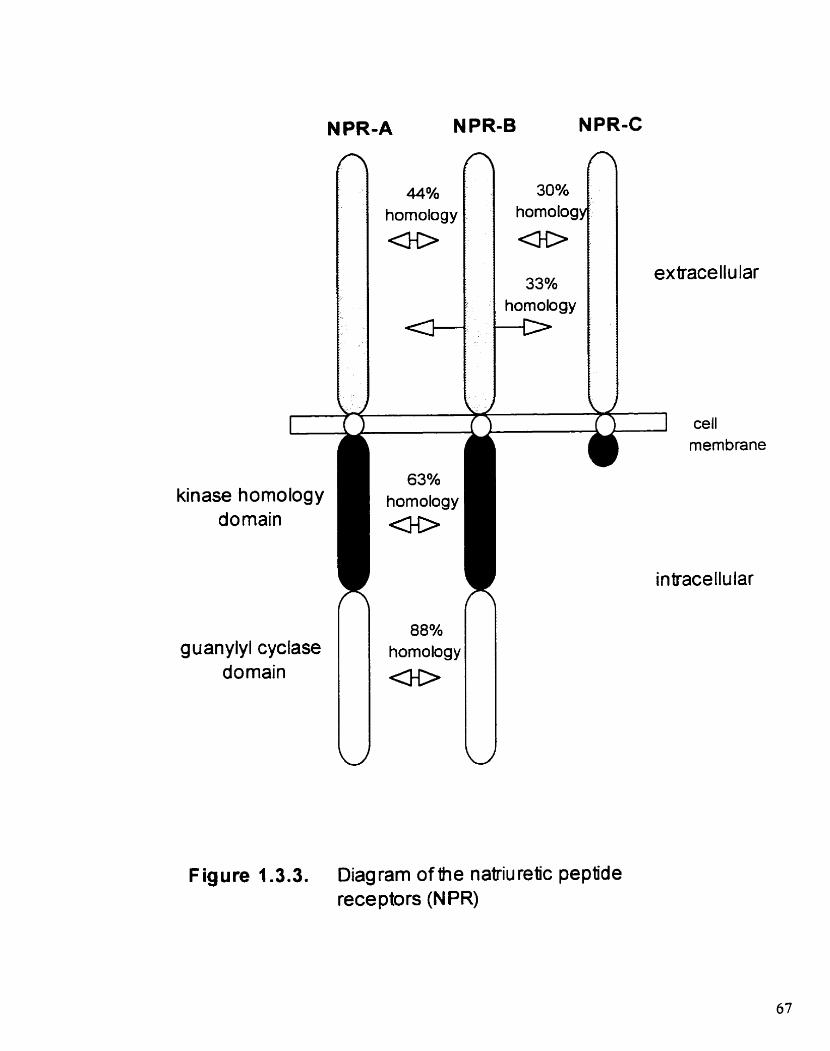
kinase homology domain
g uanylyl cyclase domain
N PR-A N PR-B NPR-C
44% homology
a-D
88% homology
0-D
30% homology
a-P 33%
homology
-I>
extracellular
cell membrane
Figure 1.3.3. Diagramofthenatriureticpeptide receptors (NPR)

homology domain may be important in transducing binding information to cGMP production
(Chinicers and Garbers. 1989: Koller et al., 1992). A negative regulation of cGMP by this
spec%c area has been postulated. since the deletion of the kinase homolou domam results
in constitutive production of the second messenger (Larose et al.. 199 1 : Chinliers et al..
1991).
The aminoacid sequence between the two receptors presents a 44% homolou in the
extracellular region. 63% homology in the kinase homology domain and 880h homolog in
the GC region (Chang et ai., 1989). The GC receptors also have about a 30% sequence
homology with the extraceMar peptide-buidhg domain of NPR-C. the third and stnicturally
different receptor of the natriuretic peptides (Fuller et al.. 1 988).
The relative selectkity of the GC receptors to the various natriuretic peptides lias been
assessed ni ce11 membrane preparations fiom various tissues (Suga et al.. 1992a). The NPR-A
receptor bmds most effectively ANP and BNP. However. BNP is usually 1 O-times kss potent
than ANP m stimulahg cGMP production by this receptor (Schulz et ai.. 1989). In contrast.
CNP bas Little effect on the cGMP production by the NPR-A receptor. suggesting the
following rank order of selectivity: ANPzBNPKNP (Suga er al.. 1992a). The NPR-B
receptor recognizes especially CNP. suggesting CNP>ANPzBNP as a rank order of
selectivity (Koller et ai.. 199 1 ). Thus. ANP and CNP are postulated to be the major ligands
for NPR-A and NPR-B respectively. As yet. no specific receptor for BNP has been identified.
1.3.4.2. Clearance receotor (NPR-Cl
The mature NPR-C receptor. structuraily vety cliffereut fiom the GC NPR-A and NPR-B
receptors. exists predomioantly as a homodimer and consists of an extracellular peptide-
bmdmg domaIn. a single transmembrane area and a small(37 aminoacid) intracellular region
(figure 1 - 3 2 . ) (Fuller et al., 1988: Uchida et al.. 1989). The NPR-C gene contains 8 e'rons
and 7 introns (Lowe et ai., 1990; Porter et ai., 1990; Sahelci et al.. 199 1). Exons 1 to 6
encode for the extracellular domain, whereas exons 7 and 8 code for the transmembrane-
spanning region and the intracellukir area respectively. The translation of the mRNA gives rise
to a 537 aminoacid NPR-C precursor in bovine tissue (Fuller er al., 1988). In contrast to

NPR-A and NPR-B. this receptor lacks the cytoplasmic GC and kinase homology domams.
Eariy experiments faiied to noticed a si@cant association between peptide binding to this
receptor and any physiologicd effect of the natriuretic peptides (Schenk et ai.. 198% Leitman
et ai.. 1986). Furthemore. the demonstration of NPR-C intemaikation upon ligand binding
and the subseguent lysosomal degradation of the ligand itself (Nussemeig et ai.. 1990:
Maack et al.. 1993). led Maack et ai.. m 1987. to propose a role for NPR-C in natriuretic
peptide clearance. This so-called "clearance receptor" then may function as a b d e r system
to prevent sudden variations in natnuretic peptide levels and BP. However. other Imes of
evidence indicate a biological role for the NPR-C receptor (Anand-Srivastava and Trachte.
1993 : Smyth and Keenan. 1994). Indeed recent experimeots suggest that NPR-C may inhibit
adenylase cyclase (AC)-mediated adenosine 3'. 5'-monophosphate (CAMP) production
( Anand-Scrivastava et al.. 1990: Tseng et cri.. 1990: Drewett er ai.. 1992). activate the
phosphomositol pathway (Hirata et al.. 1989) and mediate parts of the antiproliferative action
of the natnuretic peptides (Levin. 1993 ).
In contrast to both GC NPR-A and NPR-B receptors. NPR-C does not have stringent
requirements for peptide binding. It exhibits hi& a f i i t y for ANP. BNP. CNP and even for
vanous tnmcated. ring-deleted and linear biologically inactive ANP analogs (Leitman et al..
1986: Maack et ai.. 1987: Olins et nf.. 1988: Bovy et oi., 1989). Its rank order of specificity
for the major natriuretic peptides is ANP>CNP>BNP (Suga et al.. 1992a).
1.3.4.3. R e c e ~ tors for N-terminal fra~ments of D~oANP
Recent experiments have postulated the existence of separate and distinct receptors for
the newly identified peptides produced fiom the processing of the N-terminal fragments of
the proANP molecule (see section 1.3.1.3) (Vesely et ai.. 1990: Gwining et al.. 1992: Zeidel.
1995). However. the characterization or the molecdar cloning of these receptors have not
yet been reported.
1.3.1.4. Tissue distribution
Both GC (NPR-A and NPR-B) and clearance (WR-C) receptors are present in renal

tissues (Napier er al.. 1984: De Léan et al.. 1985: Brown and Zuo. 1992). Within the
nephron. the glomenilus and huer medda collectmg ducts (LMCD) have the hi&ea
concentration of natriuretic peptide binding sites (Mattin er al.. 1989: Brown and Zuo. 1992).
The quantification of the receptor subtypes with des-(Glnl". Ser"'. Gl y "". Leu"'.
Gly"" ]ANF,u2-,2, (or cANF). a tmca ted and ring deleted ANP analog with NPR-C
s p d c i t y (Maack et al.. 1987). demonsh-ated a &ed receptor population in the glomenilus
with 50 to 80% of the receptors being of the NPR-C type. and a homogenous GC receptor
population (NPR-A or NPR-B) in the iMCD (Martin er al.. 1989: Nugiozeh er al.. 1990).
Although the messages for NPR-A NPR-B and NPR-C have been detected throughout the
kidney (TaUenco-Melnyk er al., 1992: Greenwald et al.. 1992: Canaan-Kuhl et ai.. 1992:
Teada et al.. 1994). no expression of NPR-B mRNA bas been reponed yet. suggesting the
presence of mature NPR-A and NPR-C. but not NPR-B. in renal tissues (Brown and Zuo.
1992: Luk et al.. 1994). Kidney vascular segments. such as the vasa recta. and renal
meduilary interstitial cells have also been s h o w to possess nattiuretic peptide receptors
(Bianchi er al., 1987; Fontoura et al., 1990).
The vascular tissues primanly express the NPR-C receptor (Leitman et al.. 1986: Cahill
et al.. 1990). Nevertheless. the message for both GC and clearance receptors have been
detected in vascular cells. although the subtype of GC receptor varies with the cell type. since
only NPR-A or NPR-B are present in aortic endothelial and smooth muscle cells. respectively
(Suga er al.. 1992b: KatAchi et ai.. 1992). The NPR-C receptor is also localized in platelets
(Anand-Scrivastava et al., 199 1 ; Schif in et ai.. 199 1 ).
NPR-A. NPR-B and NPR-C mRNAs are expressed in cardiac tissues (Nunez er ai..
1992; Rutherford et al.. 1992). The expression ofthe NPR-C receptor appears to be the mon
abundant. whereas the prevalence of NPR-B mRNA seems lower than that of the other two
receptors (Nunez er al., 1992).
In the CNS. the NPRs are mainly localized in the circumventricular organs. such as the
subfomical organ (SFO). area postrema (AP). choroid plexus (CP) and organum vasculosum
of the lamina terminalis (OVLT) (Quuion et al.. 1986: Mantyh er al., 1987: Brown and
Czaniecki 1990a). The quantification of the receptor subtypes has revealed the presence of

on& GC receptors in the SFO and AP (NPR-A ancilor NPR-B) (Himeno et al.. 1992: Konrad
et al.. 1992a.b; Brown and Zuo. 1993). but of the mixture of GC and clearance receptors in
the CP (Himeno et al.. 1992: Zorad et al.. 1993 ). Lower NPRs concentrations are expressed
within the brain. but Sumners and Tang. in 1992. postulated that NPR-A and NPR-B
predorninate in fibroblan and neuronal cells. respectively. NPRs have also been described in
the olfactory bulb (Gutkowska et al.. 1991), spinal cord (Sirnonnet et al.. 1989). cerebral
microvessels (Whitson et al., 1991) and on the blood side of the BBB (Ettnish, 1992).
Similady. natriuretic peptide binding sites have been demonstrated in the anterior. but not
posterior. pituitary (Von Schroeder et al.. 1985: Pang et al.. 199 1 ).
in the adrenal gland, NPRs are present in the glomenilosa layer of the cortex (Meloche
et al.. 1986: Takayanagi et ai.. 1987). Both NPR-A and NPR-C are expressed in the adrenal
cottes. aithough the proportion of each receptor subtype varies between species (Schiffrin er
al., 1985; Meloche et al., 1986; Ohashi et al., 1988).
Various experiments have demonstrated the preponderance of NPR-C receptors in lune
membranes. in similar proportion than those found in the glornerulus and vasculature
(Leitman et al., 1987: Uchida et ai.. 1989: Abe et al., 1995). Other organs have also shown
natriuretic peptide bhdmg sites. such as the thyroid gland (Tseng et al.. 1990). testis (Pandey
et al.. 1986). liver (TalIerico-Melnyk et al.. 1992) and uterus (Itoh et al.. 1994: Dos Reis er
al., 1995).
1.3.5. PAYSIOLOGICAL ACTIONS OF THE NATRIURETIC PEPTiDES
It is generally accepted that the natriuretic peptide family plays a sipficant role in 1 )
body fluid and 2) cardiovascular homeostasis (figure 1.3.4 ). The p hysiological regulation and
control of these two major functions by the natriuretic hotmones involves the interaction of
the natriuretic peptides with various organs and hormonal systems throughout the body.
1.3.5.1. Renal actions
The natxiuretic and diuretic properties of the natriuretic peptides have been reco+pked
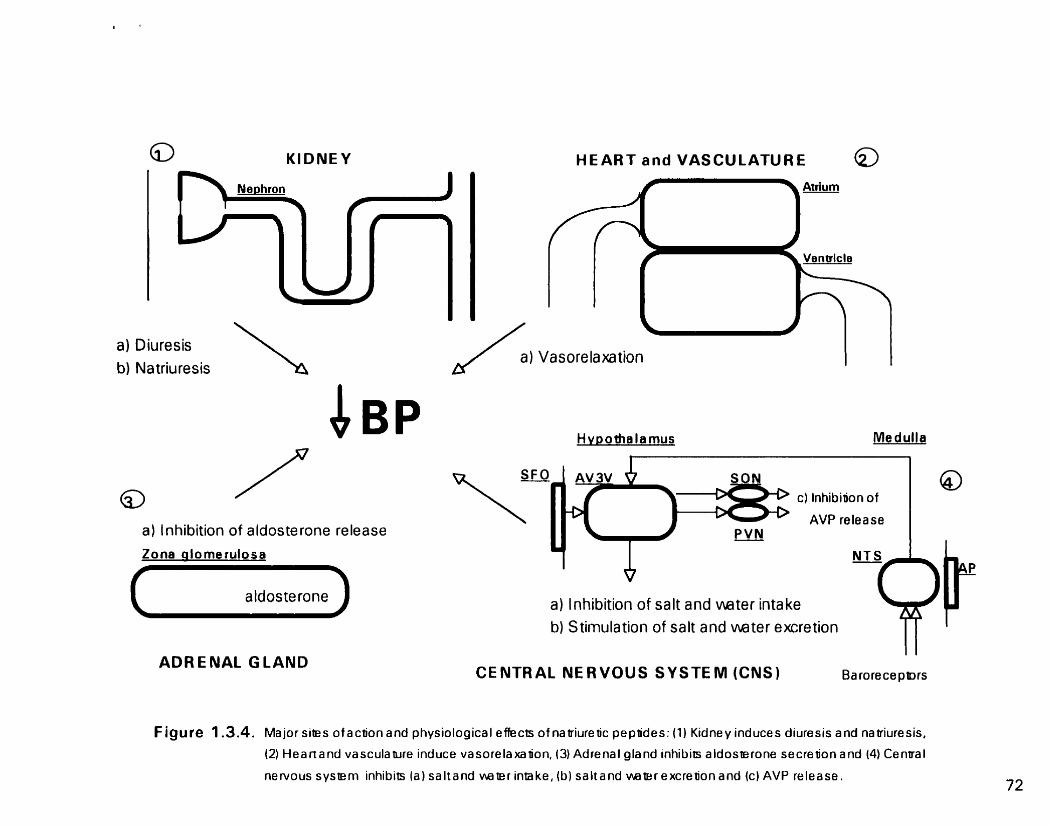
a) Diuresis b) Natriuresis
a) l nhibition of aldoste rone release Zona qlomerulosa
aldos te rone u ADRENAL GLAND
H E A R T and VASCULATURE 0
i - ty~od~alarnus Medulla
AVP release I
a) Inhibition of salt and m t e r intake b) Stimulation of salt and w t e r excretion
CENTRAL NERVOUS S Y S T E M (CNS) Barorecepnrs
Figure 1.3.4. Major s i s s ofaction and physiological effeco; ofnatriuretic peptides: (1) Kidney induces diuresis and naaiuresis,
(2) Heanand vasculahire induce vasorelaxanon, (3) Adrenal gland i n h i b i ~ aldosterone secretion and (4) Cenaal
netvous system inhibits (a) saltand viater intake, (b) saltand vm&r excretion and (c) AVP release.

early (De Bold et al.. 198 1 : Maack et ai.. 1984; Weidmann et al.. 1986: Zeidel and Brenner.
1987: Hoimes et ni.. 1993). in addition. elevations in potassium magnesium chloride and
phosphate excretion have also been observed followkg ANP administration (Maack et ai-.
1984: Cody et al., 1986). Altliough not completely elucidated. tubular. hemodynamic and
hormonal contriiutions have been postdated to explam the natriuretic and diuretic properties
of the natriuretic peptides (figure 1.3.5. ).
( 1 ) A direct effect of ANP. as well as of BNP and UD. on tubular sodium reabsorption
has been reported (Murray et ab. 1985: Blaine. 1990). ln the IMCD. the C-terminal
oaaiuretic peptides have been shown to inhibit sodium reuptake by regdathg the amiloride-
semitnie sodium channels. via NPR-A receptors and cGMP production (Sonnenberg et al..
1986: Zeidel et al.. 1988; Zeidel 1993 ). interestiogly. the N-terminal ANP molecules have
also been show to produce natriuresis. but mdependently From cGMP production. This effect
is mediated by the mhibition of the Na+/K+-ATPase actMty- probably through enhanced PGE,
actMty (Gunning et al. , 1 992; Zeidel- 1995 ).
(2) A modest mcrease of the glomemlar filtration rate (GFR) has been demonstrated by
mcreases of both plasma ANP and BNP levels (Atlas er al.. 1984: Maack et al.. 1 983). This
effect is most likely due to an eievation in the gIomemlar capillary hydraulic pressure induced
by the relaxation and constriction of the afferent and efferent anenoles. respectively (Marin-
Grez et al.. 1986: Veldkamp et al.. 1988: Kimura et ai.. 1990: Lanese et ai.. 199 1 ). The
relaxation of the glomerdar mesangial cells by ANP may also esplain. at leaçt in part. the
effect of the natriuretic peptides on the G R by expanding the capilIary surface area available
for filtration and thus mcreasing the filtration coefficient (Singhal et ai.. 1989). However. the
correlatioii between GFR and natriuresis is ail1 incompletely understood. since some
expedents have demoostrated natriuretic effects of ANP without sigdicant modications
in the GFR (Salazar er ai., 1986: Blaine. IWO). Another intrarenal hemodparnic process
possibly important in natriuresis consias of the natriuretic peptide-mediated increases in
medullary blood flow. particularly Ma the vasa recta (Kiberd et ai.. 1987). This vasodilation
is believed to rnodiSi the medullary tonicity and pressure gradients between the vasa recta and
coilechg ducts. leading to a reduced passive reabsorption of sodium and wat er by the tubular
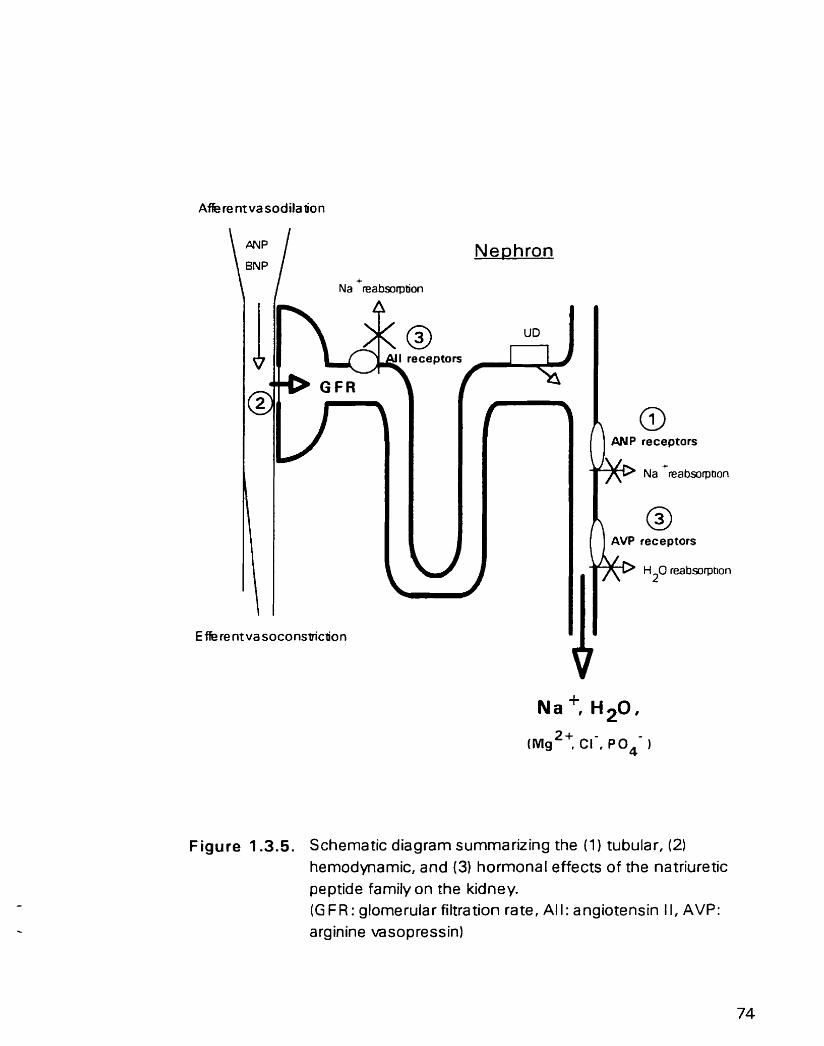
INP
I, m 2 u
Efierentvasoconsmction
) ANP receptors
O receptors
2+ (Mg , CI-, ~ 0 ~ -
Figure 1.3.5. Schematic diagram summarizing the (1) tubular, (2) hemodynamic, and (3) hormonal effects of the natriuretic peptide farnily on the kidney. (G FR : glomerular filtration rate, AI 1: angiotensin I 1, AVP: arginine vasopressin)

structures and to sodium secretion tiom the mterstitnim into the tubular fluid (renal rnedullary
washout) (Takezawa et ai.. 1987: Davis and Briggs, 1987: Zeidel et al.. 1993).
(3) Intrarenal interactions between the natnuretic peptide family and other hormonal
systems implicated in body fluid homeostasis have also been noted. Thus. in the cortical
collecting ducts. ANP. BNP and üD have an inhibitory action on the antidiuretic effects of
AVP (Zeidel et al.. 1987: Espmer. 1994). In addition. inhibition of the antinatnuretic effects
of angiotensin II at the level of the proximal rubules (Harris and Skinner. 1990) and of
aldosterone in the kidney cytosol (Honuchi et ai.. 1 989) by ANP have been demonstrated.
E.utrarenal interactions between the natnuretic peptides and vanous hormonal systems have
also been observed and are discussed m the foliowing sections (sections 1-3-53. to 1.3.5-5.).
1.3.5.2. Cardiovascular actions
The significant BP reducing effects of the natriuretic peptides have been documented in
both experimental a-1s and hurnans (figure 1.5.6.) (Maack et al., 1984: Volpe et al.. 1981:
Richards et ai., 1985: Tikkanen et al., 1985: Weidmann et al., 1986).
( 1 ) hi vitro studies iiidicated that this hypotensive effect may be due to a direct effect of
ANP or BNP on the vascular smooth muscle cells. producing relaxation (Cume et al.. 1983:
Garcia et al.. 1985: Bolli et al.. 1987). The vasorelaxant effects of the natriuretic peptides are
usually more pronounced in larger arteries than in smaIIer vessels. and appear to be associated
with cGMP accumulation (Winquia et al., 1984: Faison er al.. 1985). In recent years. the
identification of CNP in endotheliai cells and NPR-B in smooth muscle cells have also
suggested the existence of a local "vascuiar natriuretic peptide system" with vasodilatory and
grow-th inhibitory properties (Komatsu et nL. 1 W2a: Ogawa, 1995 ).
(2) Acute in vivo experiments have associated the hypotensive response to the natriuretic
peptides with a reduction in the cardiac output (CO). which may be the result of a Iower
venous retum (VR) (Breuhaus et ai.. 1985: k I I e et ai.. 1992). The reduced VR is probably
due to the ANP-mediated decrease m blood volume, secondary to the shift in the extracellular
fluid (ECF) fiom the vascular to the interstitial space (Almeida et al.. 1986; Trippodo and
Barbee. 1987: Groban et al.. 1990) and to the increased resistance to the VR (Chien et al..
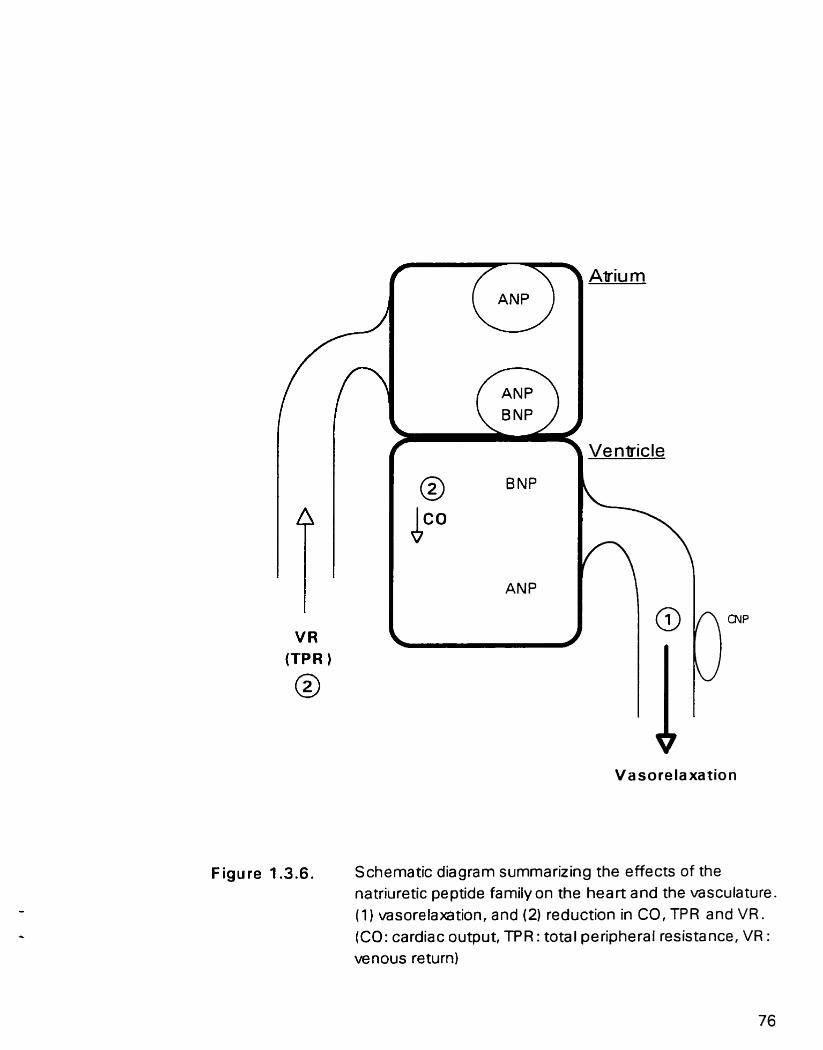
BNP
ANP
Atriu m
Vasorelaxation
Figure 1.3.6. Schematic diagram summarizing the effects of the natriuretic peptide family on the heart and the vasculature. (1) vasorelaxation, and (2) reduction in CO, TPR and VR. (CO: cardiac output, TPR : total peripheral resistance, VR : venous return)

1987; Christensen, 1993). With long-terni ANP administration, the initial decrease in CO is
no longer observed, but the continued hypotensive response appears to result fiom a
secondary decrease in total peripheral resistance (TPR) (Parkes et al., 1988; Charles et al..
1993). The exact mechanisms by wfüch chronic ANP Uifiision lower TPR are unclear, but
ANP-induced inhibition of angiotensin II or endothelin-dependent vasoconstriction has been
suggested (Kohno et al.. 199 1).
An action ofthe natriuretic peptides on baroreceptors has also been postulated because
of the absence of reflex tachycardia after the lower BP and CO resultmg from ANP infùsions
(Schulz et al., 1988).
1.3.5.3. Adrenal actions
( 1 ) Natriuretic peptides have been s h o w to inhibit aldosterone release (figure 1.3.7.)
(Maack et al.. 1984; Jennhgs et al.. 1990). Although this inhi'bition is mediated in part by the
ANP-induced suppression of the action of various aldosterone secretagogues nich as
angiotensin II (Cuneo et al., 1987), potassium (Clark er al.. 1992) or ACTH (Agdera.
1987), a direct effect on the adrenal gland has also been demonstrated (Chartier et al.. 1984:
Higuchi et al., 1986; Rosenberg et al.. 1 989). The mhiitioa of the adrenal zona glomedosa
aldosterone production by ANP appears to be exerted primarily at the early aeps of
steroidogenesis, duriog the formation of pregnenolone (Racz et al.. 1985: ElIiott and
ûoodfiiend, 1986) and also to some extent during the final conversion of corticosterone to
aldosterone (Campbell et al., 1985). In contrast to most other effects of the natriuretic
peptides, the exact mechanism of action for this inhibition remains controversial. None of
cGMP analogs affect sigiilficantly the aldosterone production (Matsuoka er al.. 1987;
Gan& er al., 1989; Gangdy, 1992). Therefore. a biological role has been suggested for the
NPR-C in the adrenal corte- even though some reports aill argue for a contniution of the
GC recepton (Bahr et ai., 1993). The possiiility of a local "adrenal natriuretic peptide
systern" bas also been suggested foUowing the demonstration of ANP, BNP and CNP
transcripts m adrenal chromfi celk (Ong et al., 1987; Nguyen et al., 1989a; Babinski et al.,
199 1).
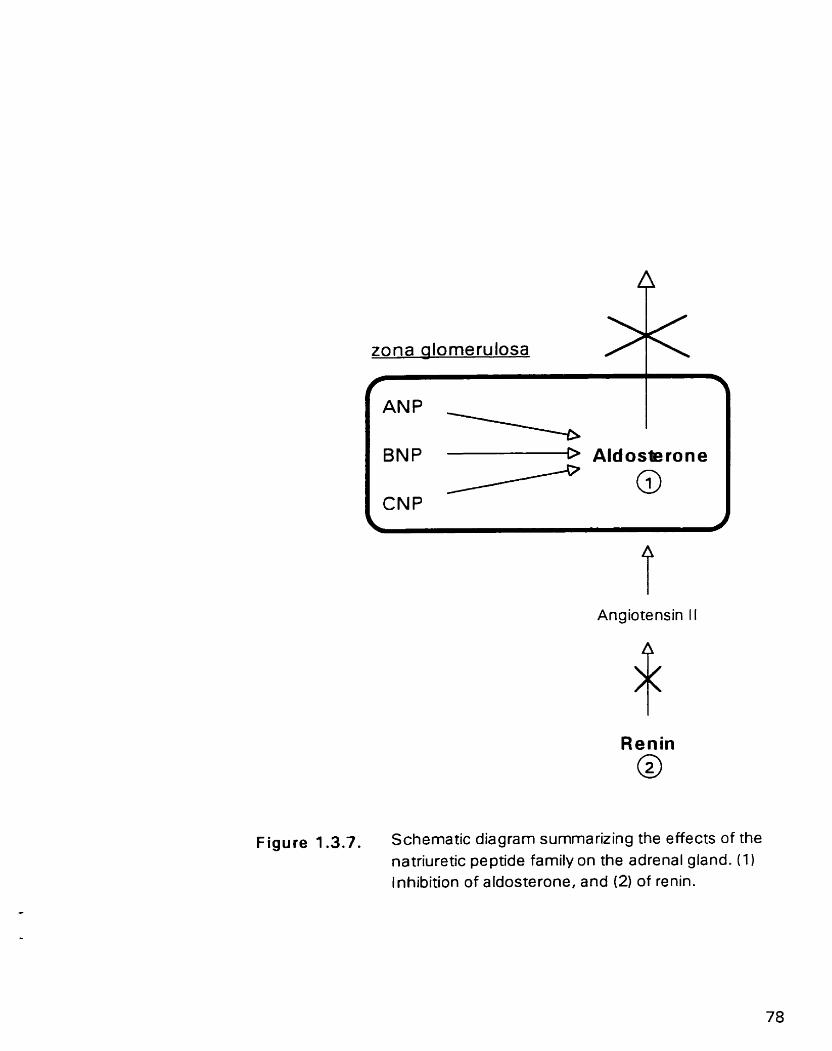
zona alornerulosa
1 BNP Aldosderone
Angiotensin II
Renin
O
Figure 1.3.7. Schematic diagram summarizing the effects of the natriuretic peptide family on the adrenal gland. (1 1 l nhibition of aldosterone, and (2) of renin.

(2) ANP and BNP also inhibit renin release fiom the renal juxtaglomerular cells (Maacli
et al.. 1984: K u t z et al.. 1986). The renin suppression by the natriuretic peptides seems to
be cGMP dependent (Kurtz et al.. 1986).
1.3.5.4. Central nervous svstem KNS) actions
Recently. a major role for the central component of the natriuretic peptide system in
cardiovascular and k i d homeostasis has beai postulated ( h u r a et al.. 1992). The penpheral
stimuli received by the bram natriuretic system such as plasma ANP binding to the
circumventncular organ receptors or the neuronal signais fiom the baroreceptors. are believed
to produce a cohort of CNS responses that rnay include three components: the modulation
of £luid and salt intake. the modulation of fluid and salt excretion and the modulation of other
honnonal syaems implicated in fluid and salt homeostasis (figure 13.8.).
( 1 ) Intracerebroventricular (i.c. v. ) ANP injections have been shown to inhibit water
mtake during angiotensin Il -induced drinking or in water-deprived rats (Antues-Rodrigues
er al.. 1985: Nakamura et al.. 1985: Katsuura et al.. 1986: Lappe er ai.. 1986). Similarly. an
inhibition of salt appetite is reported after central administration of ANP in rats. rabbits and
sheeps ( Antunes-Rodrigues et al.. 1986: Ta jan et al.. 1988: Weisinger et al.. 1992). The
exact location for these antidipsogenic and antiadipsianic actions of natriuretic peptides is
uriclear. but it may mvofve the AV3 V area. an hypothalamic structure important in body fluid
homeostasis (Buggy and Bealer. 1987: Ku and B a n g 1994). The implication o f the SFO has
also been dernonstrated. since ANP microinjections into the subfomical organ attenuate
angiotensin 11 -induced drinking (Ehrlich and Fitts 1990) and inhibit the actkity of
angiotensùi II -sensitive neurons projecting to the AV3V area (Hattori et ai.. 1988: Shibata
et al.. 1992).
(2) Central mfùsions of ANP have been associated with diuresis and. in some instances.
natriuresis (Fhs er al.. 198% Israel and Barbeila. 1986: Shoji er al.. 1987). Although part of
the water excretion appears to result Eom the ANP-mediated modulation of central AVP
(Standaert et al.. 1987). a direct effect of the brain natriuretic system on renal diuresis has
also been suggested (Shoji et al., 1987). Again. the AV3V area seems important in the central
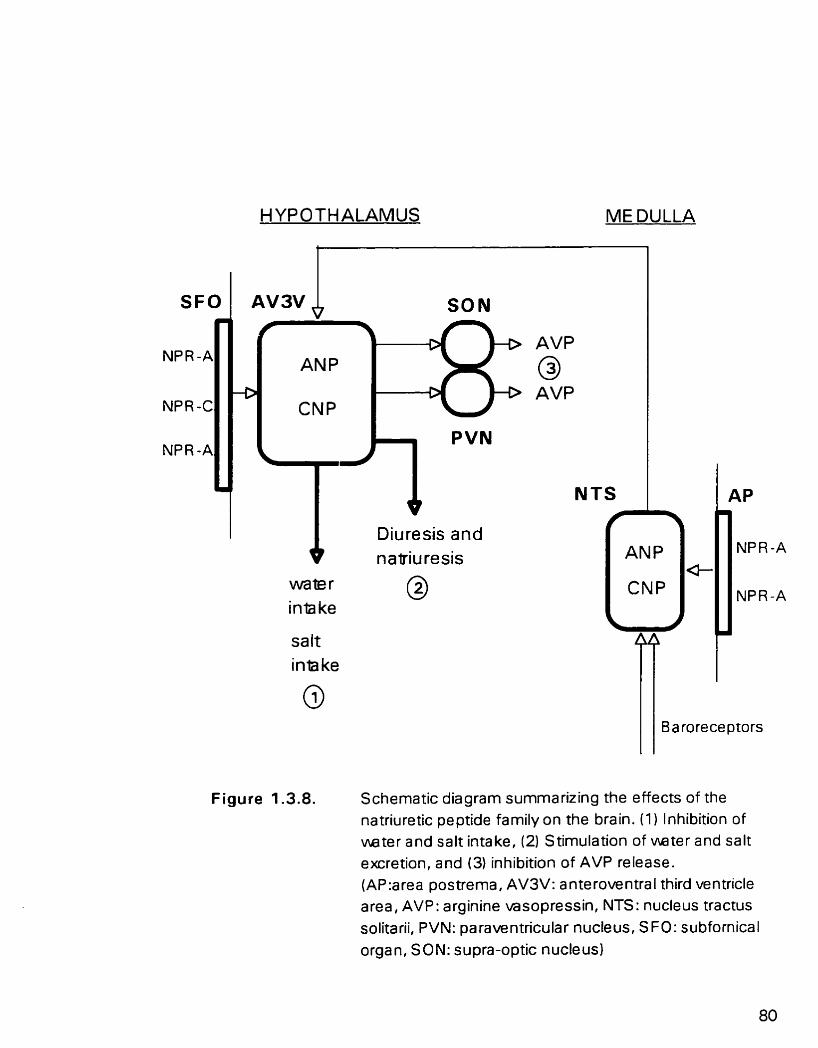
HYPOTHALAMUS ME DULLA
NPR
NPR
NPR
CNP
SON
PVN I
I C Diuresis and nalriuresis
Figure 1.3.8.
NTS
I I Ba roreceptors
Schematic diagram summarizing the effects of the natriuretic peptide family on the brain. (1 ) Inhibition of -ter and salt intake, (2) Stimulation of m t e r and salt excretion, and (3) inhibition of AVP release. (AP:area postrema, AV3V: a nteroventrai third ventricle area, AVP: arginine vasopressin, NTS: nucleus tractus solitarii, PVN: paraventricular nucleus, S FO: subfornical organ, SON: supra-optic nucleus)

control of water and sodium handling by the kidney (Imura et al.. 1992). Some reports have
demonstrated a depressor action of i.c. v. ANP mjections. especially during central angiotensin
[I -mediated hypertension. but the issue remains controversial (Itoh et al.. 1986a: Castro et
al.. 1987). The hypothalamus may be the site of action for this central hypotensive effect of
natriuretic peptides. In addition. the meduila oblongata. particularly around the NTS and the
nucleus arnbiguous. may be important. since ANP microinjections in these areas have
produced significant reductions in BP (McKitnck and Calaresu. 1988: Levin et al.. 1989:
Ermirio et a', 199 1).
(3) Several hormonal and neurotransmitter systems are modulated by the central
administration of ANP. The release of AVP fiom the supraoptic and paraventricular nuclei
m mhibited by central infusion of natriuretic peptides (Crandall and Gregg, 1986: Samson et
al.. 1987: Standaert et al.. 1987). Basal AVP release 6om the posterior pituitary is also
reduced foilowing I.C. v. ANP administration (Obana et al.. 1985: Januszewicz et al.. 1985).
An inhibitory action of centrally-administrated ANP on dopaminergic (Nakao et al.. 1986).
noradrenergic (Vatta et al.. 1992) and angiotensin II -sensitive neurons (Hattori et al.. 1985)
has also been demonstrated. In the cases where appropriate studies were perfomed. these
central effects ofthe natriuretic peptides appeared to be mediated by cGMP (Giridhar et al..
1992).
A role for ANP m CSF formation. particularly in the choroid plexus. lias been suggested
(Tsutsurni et al.. 1987: Steardo and Nathanson. 1987).
1.3.5.5. )iormonal actions
Enhanced aaivay of the natriuretic peptide system has been associated with the inhibition
of the secretion of ACTH (King and Baertschi 1989: Dayanilhi and Antoni 1990: Fink et al..
199 1 ), prolactin (Samson and Bianchi 1 988) and luteinking hormone secretion (Samson et
al., 1988: Franci et al.. 1990; Samson et al.. 1993 ), but not of thyroid-stirnulating hormone
(Franci et al.. 1992). in contrast. acute exposure to ANP has been shown to enhance
testosterone secretion (Mukhopadhyay et al.. 1986) and to inhibit progesterone production
(Pandey et al., 1985).

The naaniretic peptides have been shown to inhibit most of the angiotensm U actions ( at
the levels of the vasculature. brain. kidney and adrenals). suggesting the concept of
physiological antagonism between the angiotensin and natriuretic peptide systems
(Mendelsohn et ai., 1987: Espiner. 1994).
1.3.5.6. Pulmonarv actions
Bronchorelaxation (Potvin and Vanna. 1989). regdation of lung pemeability and thus
prevention of pulmonary edema (Lofton et ai.. 1990) and a role m &actant production (Ishii
et ai.. 1989) have been suggested for ANP in pulmonary tissues. The mechanim, of action of
the natriuretic peptides at the level of the lungs is still unclear. There is some evidence that
production of cGMP is needed.
1.4. ALCOHOL (ETOH), BLOOD PRESSURE (BP) AND THE
NATRKRETIC PEPTIDE FAMILY
1 .-LI, ETHANOL (ETOH) .AND THE NATRCZTRETIC PEPTIDE FAMILY
in 199 1. when this project was initiated. there was only one published report on the
mteractions between alcohol and the natriuretic peptide system. The group of Colantonio er
al. (1991) had investigated the effect of a single ETOH dnnk ( 1 mVkg B.W.) on the
circulahg ANP levels m healthy human males. They noticed a rapid increase in plasma ANP
levels following the ETOH consumption. lasting one hour. in quasi-isovolurnetric conditions
(0.1 L of urine on average). They concluded on a possible role of ANP on the ETOH-induced
diuresis. probably through the Hihibition of AVP release as wggested by the opposite changes
in circulating ANP and AVP levels.
Since then. four other publications with divergent methodologies have proposed various
and often different associations behhieen acute ETOH consumption and ANP. comp licating
the issue. Leppiiluoto et al. ( 1992) observed a significant decrease in senun ANP levels 2
hours d e r a 1.5 g/kg B.W. ETOH drink in seven healthy males. However, the lack of blood

analysis early afier the ETOH consumption may renilt in missing the rapid and short-lived
ANP increasr noted by Colantonio et al. ( 199 1). Moreover. the rapid diuresis obsewed
during the fust hour (0.9 L on average) and the corresponding lower plasma volume should
rapidly counteract any initial stimulating effect of ETOH on the ANP system. Intereaingly.
the lower ANP levels are associated with a secondary increase in plasma AVP contents.
confirming the possibility of a link between the two hormonal syaems. The same year.
Hpynen er al. ( 1992) reported the absence of any eBect of alcohol ( 1 b@kg B.W.) on the
circulating ANP levels of ten healthy male volunteers. However. the water loading of the
subjects prior to the ETOH experiment and the corresponding increase in plasma volume
mua have sigdïcantly augmented plasma ANP content and limited the possibility or the
e.xtent of a further ANP increase by ETOH. Variations in circulating AVP levels were also
absent fiom this study (Hynynen et al.. 1992). In addition. the siguikant diuresis produced
even before the alcohol was administered could limit any subsequent potentiation of the ANP
system by ETOH. In 1994. Ekman and colleagues noticed the inhibition of the noctumal
increase Ui plasma ANP levels more than 8 hours afier the administration of a IgAg B.W.
ETOH drink in nme heaithy subjects. However. in this study the firn blood sample was taken
two hours afier ingestion of the ETOH drink lirniting the value of the experiment in
mvestigating the early rapid changes on the release of ANP. However. they noticed that the
content of the proANP molecules was not affected by the ETOH treatrnent. suggestin_e an
mcreased peripheral elunination of ANP rather than the inhibition of ANP release following
the ETOH drink to explain the observed result.
While our studies were in progress. the first midy on the interaction of ETOH with the
ANP system using experirnental animals was published (Wigle et al.. 1993a. 1995). Animal
experiments are important since they allow an accurate control of a number of parameters.
such as previous alcohol habits. heterogeneity of the subjects. food and water/ETOH intake.
etc. that may affect the outcome of the mteraction between ETOH and the natriuretic peptide
system and that cannot be controlled in human studies. Acute heavy ETOH consumption ( 5
e/kg B.W.) was administered through gastric gavage. Because of the hi& volumes of water C
@en to control rats, plasma ANP levels progressively increased during the experiment. In

contran. no modification was observed in circulatmg ANP levels after the ETOH
administration. Both atrial and ventricular ANP contents foilowed the same progressive
changes m the two groups of rats. This may be the result of the mhibition of gastnc emptying
and of the presence of secondary gastnc irritation. by such swere alcohol intoxication, rather
than of a direct eEect of ETOH itselfon the ANP system hdeed, the extremely slow and
progressive mcrease m BAC suggests a partial suppression of the normal rate of ETOH
absorption by the GI tract. Lower doses of alcohol should be used in fiiture experiments in
order to mimic the BAC c w e observed in human subjects after acute ETOH consumption.
The effects of chronic ETOH exposure on the natriuretic peptide system are even less
clear. This issue was investigated for the fïrst and ody tirne m 1993 following a 6 weeks
administration of a 20% vlv ETOH solution, using the Sprague-Dawley rats (Wigle et al..
1993b,c). No modification m the BP was noted, which is not surprishg considering the slow
and progressive nature of the antihypertensive effect of moderate ETOH consumption in rats
(see section 1.2.2.2.). Plasma and atrial ANP levels were unaffected by the ETOH treatment.
but ventricular ANP content was significantly elevated in ETOH-treated rats (Wigle er al..
1993b). Intereçtingiy, circulahg BNP Ievels were also augmented in these animais (Wigle et
al., 1993~). in order to really analyze the possible contribution of the natnuretic peptide
system in the prevention of the age-dependent increase m BP by chronic rnoderate ETOH
drinking, it may be necessary to investigate this systern after the antihypertensive effect of
ETOH has been established (after at least 6 months) and not only during the early stages of
the treatment (after 6 weeks). Furthemore, since this prevention of the age-dependent
increase m BP is more evident in the hypertensive strains of rats, such as S m experiments
should be designed to compare the effects of ETOH m the hypertensive rats versus the
normot ensive controls.
In recent years, ANP has also been implicated with the attenuation of delirium tremens
and convulsions during ETOH withdrawal (Bezzegh et al.. 199 1; Kovacs. 1993).
1.4.2. WORKLNG EiYPOTKESIS AND DlVISION OF TEE TaESlS
in the previous sections, the evidence for the antihypertensive ef5ects of acute and

chronic moderate ETOH consumption has been rwiewed. A few mechanisms, nich as
lipoproteins or hemostatic processes, have been descnbed to explain this effect of alcohoL but
a conviucing hypothesk is stin lachg. It has been suggested that ETOH may alter the actMty
of a number of physiological systems which rnay then mediate its antihypertensive effects. A
new and expandmg fandy of natriuretic peptides with widespread BP lowering and
vasorelaxant effects has also been discussed. Recent evidence of some alterations m the
a+ of this system by ETOH just opened the possiibility of a new mediator for several of
the ETOH-mduced effects on the cardiovascular system
InteresMgly, alcohol and the naviuretic peptide f d y have several nmilar effects on
the kictney, the cardiovascular system and the brah. They both have some depressor effects
on various brain functions. They both produce transient antihypertensive effects and lower
cardiac contractiiity. They both mduce diuresis and natriuresis. Since a number of studies have
demonstratecl that ETOH alters the activity of a number of hormonal syaems such as AVP.
it is possible that the activity of various componaits of the natriuretic peptide system may also
be affected by alcohoL It is surprising then that the implication of the natriuretic peptide
family on some of the effects of alcohol had not been investigated pnor to 1991. For
example, it has been postdated that the dimesis caused by ETOH is mediated by the inhibition
of AVP (Eisenhofer and Johnson, 1982; Leppaluoto et al., 1 992). Interestmgly, AVP release
kom the hypothalamus is also suppressed by AM> (Januszewicz et al., 1986a; Crandall and
Gregg, 1986; Standaen et al., 1987). Thus, it is possible that the inhiiitory effect of ETOH
on AVP release is mediated by a aimulatory effect of ETOH on ANP secretion. Then. some
of the cardiovascular and rend effects of ETOH previously attnbuted to AM> may be
mediated rather by the natriuretic peptide family. Similady, such modifications in the
natriuretic peptide system may produce, during long-tenu moderate ETOH administration.
permanent antihypertensive effects that may explain, at leaa m part, the U-shaped BP c w e
and the prevention of the age-dependent mcrease in BP observed m human subjects and
experirneatal animals following chronic moderate ETOH consumption.
Therefore, the present series of experiments were undertaken to mvestigate the following
hyp othesis:

Acute and chronic alcohol exposure alters the activity of distinct
components of the natriuretic system (natriuretic peptides and natriuretic
receptors). These alterations may mediate some of the effects of ETOH on the
cardiovascular system.
(1) ACUTE ETOH STUDiES: Chapter 2 describes our audies investigating the eEects
of acute low and moderate ETOH consutnption on cardiac and circulating ANP levels in rats
(Section 2.1. ) and human volunteers (Section 2.2.).
(a) A n h l studies (Section 2.1.): Male LongEvans rats receive a 1 or 2 g of ETOWkg
BW through i-p. injection. Circulating and cardiac ANP levels are measured at various time
points foiiowing the ETOH injection.
(b) Human studies (Section 2.2.): Normotensive i n d ~ d u a l s receive a 0.25 or 0.50 g of
ETOWkg BW in unsweeteaed orange juice. Plasma ANP levels are measured for 3 hours
following the ETOH ingestion.
These studies should d o w the determination of any acute and/or short-lasting activation
of the natriuretic peptide syaem. The demonstration of a stimulatory effect of acute ETOH
treatment on plasma ANP levels should indicate whether the natnuretic peptide syaem is
sensitive to alcohol exposure. in addition. any stimulatory effect of ETOH on the circulating
ANP levels may suggen the possiiility of a sipficant antihypertensive effect of this hormonal
system during chronic moderate ETOH drinking.
(LI) CHROMC ETOH STUDIES: Chapter 3 describes our studies surveying the effects
of chronic moderate ETOH consumption on the a c t ~ t y of distinct components of the
naaiuretic system (natrituetic peptides and natriuretic receptors). Male Wistar- Kyoto ( WKY )
and spontaneously hypertensive ( S m ) rats receive a 20% vlv solution of ETOH for 8
months The effect of chronic moderate ETOH consumption on the age-dependent increase
m the BP is descnied in section 3.1. The studies of Chapter 3 are performed on some of the
major sites of natriuretic peptides synthesis and action. such as the heart. liidney and brain.

following long-term moderate alcohol consumption.
(a) Cardiac natriuretic -stem (Sections 3.1. and XZ.1 : The heart is the central
component of the ANP and BNP systems. It may directly alter the BP. both fiom
modifications in cardiac contractility or hormonal release. It is also sensitive to ETOH.
Circulating ANP (chapter 3.1) and BNP (chapter 3.2) levels are measured following chronic
moderate ETOH consumption. Modifications in atrial and ventricular ANP. BNP. ANP
mRNA and BNP mRNA are also mvestigated following the alcohol treatment.
(b) Renal natnuretic svstem (Section 3.3.): The kidney is associated with the
dwelopment of hypertension. It mediates some of the major effects of the natriuretic peptides
through specific receptors. Its functions are altered by ETOH exposure.
Modifkations m glomenilar and papillary natriuretic peptide receptor characteristics are
investigated following chronic moderate ETOH consumption.
(c) Brain natriuretic wstem (Sections 3.1. and 3.5.): The brain is the main target for
several of alcohol-induced effects. In recent years. its importance in the natriuretic peptide
family physiology has been demonstrated. It also controls hdirectly the BP via hormonal
sensors in circumventncular organs and through neuronal inputs via baroreceptors.
Modifications in ANP and CNP levels in the major brain areas (chapter 3.4.) and
alterations in natriuretic peptide receptor characteristics in the circumventncular organs
(chapter 3.5.) are investigated following the chrooic administration of moderate amounts of
ETOH.
Thus, the present studies describe the general modifications in the actMty of the
natriuretic peptide system following chronic moderate ETOH consumption. This work
repreçents the kst s w e y on the existence and direction of such alterations and proposes the
hypothesis that this homonai system may mediate parts of the antihypertensive effect usually
observed following clironic moderate ETOH consumption.

CHAPTER 2
ACUTE ETOH STUDIES

Section 2.1.
INCREASED PLASMA ATRIAL NATRIURETIC PEPTIDE
AFTER ACUTE INJECTION OF ALCOHOL IN RATS
P. GuiUaume, J. Gutkowska and C. Gianoulakis
The Journal of Pharmacology and Experimentd Therapeutics
(J Pharmacol Exp Ther 27 1 : 1656- 1665. 19941
Contribution by CO-authors: Dr. J. Gutkowska and Dr. C . Gianoulakis were my CO-
supenisors.
Acknowledeements: R Claudio was the animal technician responsible for the weCbeing of
the mimals. D. Beaudry performed some of the O-endorph iodinations.

2.1. INCREASED PLASMA ATRIAL NATRIURETIC PEPTIDE
AFTER ACUTE INJECTION OF ALCOHOL IN RATS
2.1.1. ABSTRACT
A number of mechanisms may be involved in the protective effect of low ethanol
(ETOH) consumption on the development of the age-dependent hypertension in both
human and experimental animals. I t was the objective of the present studies to test the
hypothesis that acute administration of low doses of ETOH would increase the plasma
content of Atrial Natriuretic Peptide (ANP). an hormone known to decrease blood
pressure. Plasma ANP levels were signiticantly increased within 15 minutes following the
i .p. injection of 1 or Zg ETOH/ kg B.W.. The increase in plasma ANP was more
pronounced and longer lasting following the i.p. injection of 2 than ig ETOHkg
B. W. .This increase in plasma A N P level was associated with a rapid decrease of atrial
ANP. but not of ventricular A N P which on the contrary was significantly elevated at 120
minutes post-injection. It has been suggested that opioids could play a significant role in
controlling ANP release. In fact. circulating levels of Il-endorphin were also rapidly
increased following the ETOH injection. with a timecourse pattern similar to that of ANP.
Furthermore. a highl y positive correlation was found be tween the ET0 H- induced changes
of plasma ANP and R-endorphin contents. S ignificant increases in plasma corticosterone
and adrenoconicotropic hormone. but not aldosterone contents. were observed follow ing
the i.p. injection of Zg ETOH/kg B.W.. while plasma arginine vasopressin levels were
significantly decreased at 15 but not at 170 minutes post-ethanol. There was no significant
elevation in blood pressure during the 120 minutes experimental period. although a small
tachycardia did develop in the ETOH-treated animals. Thus. acute in v i w administration
of ETOH increased plasma ANP content in a dose-dependent manner and rnay play a role
in the "protective" effect of low ETOH consumption in the development of the age-
dependent hypertension.

2.1.2. INTRODUCTION
A number of epidemiological studies have revealed a positive association between
h igh ethanol ( ETOH) consurnption and hypertension (Gleiberman and Harburg. 1986).
However. the effect on blood pressure of low ETOH consumption is less clear
( MacMahon. 1987). Indeed. several investigations have reported a significant decrease in
blood pressure with low regular consumption of alcoholic beverages ( 1 to 3 drinks per
day). while others reported no significant changes in blood pressure ( MacMahon. 1987).
Moreover. animal s tud ies have demonstrated the prevention or delay in the development
of the age dependent hypertension with chronic low ETOH ingestion (Howe et ai.. 1989).
The mechanisms by which low ETOH consurnption rnay prevent the age dependent
increase in blood pressure are still not clear. One such mechanism may involve the Atrial
Naniuretic Peptide (ANP). a 28 arnino acids peptide synthesized and released mostly from
the hem atria (De Bold eral., 1981. Needleman ef al., 1989. Nakao et al.. 1993). This
hormone has a definite role in body tluid homeostasis (Blaine. 1990). I t s major effect is
to lower b i d pressure through increased diuresis. natriuresis and vascular relaxation. and
a decrease in the secretion of several pressor hormones (Inagami et al.. 1987). Among the
mechanisms modulating the release of ANP are changes in atrial stretch (due to changes
in blood volume). in blood pressure. in osmolarity. or in the concentration of opioid
peptides (Blaine. 1990. Pesonen et al., 1990).
Recently. several studies have been published on the effects of acute ETOH on the
A N P system in man. However. the interactions between the two are still unclear since
either an increase (Colantonio et ai.. 199 1 ). or a decrease (Leppaluoto et al.. 1992) or no
change (Hynynen et al., 1992) in the plasma ANP levels were found following acute
ETOH administration. This variability of the acute effects of ETOH on the A N P system
in human could be attributed to various factors, such as dose of ETOH used, nutritional
status. previous ETOH exposure. genetic factors. etc. (Beilin et Puddey. 1992). Therefore.
it would be interesting to study the effects of ETOH on the ANP systern using an
experimental animal mode1 such as the rat. where al1 the confounding factors (nutritional
status. previous ETOH exposure. genetic factors. etc.) could be thoroughly controlled.

Acute administration of ETOH has been reported to decrease plasma levels of
arginine vasopressin (AVP) (Pohorecky and Brick. 1988). Interestingly. ANP has also
been shown to suppress AVP release from the supra-optic nucleus (SON) (Clark et al..
1991). Similady. acute ETOH is known to increase plasma concentrations of
catecholamine (Zgombick et al.. 1986). corticosterone (Ellis. 1966). and 8-endorphin. a
potent endogenous opioid peptide (Gianoulakis. 1989). 1 ntzrestingl y. it has been reported
that p-opioid receptor antagonists decrease ANP secretion (Gutkowska er al.. l986b.
Widera et al.. 1992). while p-opioid receptor agonists increase ANP secretion (Vollmar
et al.. 1987). One might therefore hypothesize that one of the rnechanisrns by which
ETOH rnay enhance the release of ANP is through irs effects on the release of endogenous
opioid peptides. such as 8-endorphin. which binds with about equal aftïnity to both p and
8 recep tors (Charness. 1989). Additional mechanisms could include changes in plasma
osmolarity and release of other hormones. such as epinephrine. by ETOH intake
( MacMahon. 198% Colantonio et al., 199 1 )
Stress has been reported to increase the release of ANP (Horky et al.. 1985).
Therefore. it is important to distinguish between the stress mediated and ETOH specific
ef'fects on the activity of the ANP system. Since both ETOH and stress have been shown
to increase the release of O-endorphin. adrenocorticotropic hormone (ACTH ) and
conicosterone. changes in the plasma content of these hormones would indicate both the
effectiveness of the ETOH dose administered in the present studies. as well as the presence
or absence of stress due to the necessary handling and manipulations of the animals during
the course of the experiment. Furthermore. estimation of the plasma 8-endorphin levels
(a potent endogenous opioid peptide which could interact with the p-opioid receptors and
increase the release of ANP) would allow us to investigate the possibility that the ETOH-
induced changes in the plasma B-endorphin content may mediate. at least in part. the
effects of ETOH on the release of ANP. by estimating the correlation between the ETOH-
induced changes in plasma A N P and R-endorphin contents.
The aim of the present studies was to test the hypothesis that acute administration of
a low to moderate dose of ETOH in the rat would induce a rapid increase in plasma ANP

levels. A positive correlation between moderate ETOH intake and increased activity of the
A N P system might help to explain. at least in part. the prevention or delay of the age
dependent increase in blood pressure by chronic low ETOH consumption observed both
in human and experimental animals (Howe et al., 1989). The content of ANP in the hean
aria and ventricles was also rneasured and correlated with the changes in the plasma ANP
content. In addition to ANP. the changes in plasma O-endorphin. corticosterone. ACTH.
aldosterone and AVP were rneasured.
2.1.3. MATERIALS AND METHODS
2.1 .3.1. -al Treatrnents
Male Long-Evans rats (purchased from Charles River Breeding laboratories. S t-
Constant. Québec. Canada) weighing 100-300 grams were used in this study . A seven days
period was allowed for acclimatization of the animals before the initiation of the
experirnents. Al1 animals were handled twice daily to reduce stress levels during the
experiment.
EXPERlMENT 1 : The objective of the studies in experiment 1 was to perform a time
course study on the effect of ETOH on plasma ANP content. Rats were lightly
anaesthetized with "Metofane" ( Methoxytlurane. inhalation anaesthetic. MTC Canberra
Packard). and a siiastic catheter (0.035 in 1. D.. Dow Corning) was inserted into the letl
jugular vein. The catheter was tunnelled subcutaneously to the back of the neck and the
disral end was plugged by a süiinless steel pin after being tïlled with heparinized saline (70
pllml). The animals were then placed in individual cages and allowed at least a two days
post-surgery rest. On the day of the experiment. at time O. an initial blood sample (0.3 ml)
was withdrawn through the catheter to calculate initial hormone levels. prior to any
treatrnent. Then. the animals were randoml y assig ned to the alcohol treatment groups or
to the saline treatrnent. control group. The animals in the ETOH groups were injected
intraperitoneally (i.p.) with I or Ig ETOH/kg B.W. (40% vlv solution in saline). The
animals in the control group were injected i.p. with the equivalent volume of saline.

Additional blood samples. of 0.4 ml each. were withdrawn through the catheter at 15. 30.
60 and 170 minutes for estimation of the blood alcohol content and levels of ANP. fi-
endorphin and corticosterone. Following each withdrawal of 0.4 ml blood. an equal
volume of 0.9% saline was injected through the catheter to replace the withdrawn blood.
EXPERIMENT 2: The objective of the studies in experiment ? was to investigate the
effects of stress and of the temporary changes in blood volume due to serial withdrawal
of blood samples in experiment 1 (although an equivalent amount of sa1 ine was injected).
Thus. in experiment 2. animals did not have a catheter. but they were sacri ficed at 15 and
120 minutes following the administration of Zg ETOH/kg B.W.. the dose which as
demonstrated in experiment 1 induced a pronounced and long lasting increase of plasma
ANP levels. or the equivalent volume of saline. Thus. 35 male Long-Evans rats were
randomly assigned to 5 treatment groups (5 animals per group). Animals in group 1 and
group 7 were injected i.p. with 2g ETOH/kg B.W. and were sacrificed at 15 and 120
minutes post-ETOH injection respectively . Animals in group 3 and group 4 were injecred
i.p. with the equivalent volume of saline. and were sacrificed at 15 and 110 minutes
respectively. To estimate the hormone levels at tirne O (prior to administration of the
ETOH or saline solution), animals in group 5 received no injection and were sacrificed
within 70 seconds following their removal from the home cage. Immediatel y following
sacrifice, tmnk blood was collected for estimation of the blood alcohol content (BAC) and
the concentration of ANP. fi-endorphin. corticosterone. ACTH. aldosterone and A V P in
the plasma. In addition, the atria and ventricles were dissected to study the effects of
ETOH on the tissue ANP content.
EXPERIMENT 3: The objective of the studies of experiment 3 was to investigate the
e&ct of ETOH on blood pressure (BP) and heart rate (HR) . Thus. rats were placed in a
ratrainer and the BP and HR were recorded regularly on a polygraph (Grass Instruments.
Quincey, MASS) prior to and at specific intervals following the i.p. injection of either Ig
ETOH/kg B. W. or the equivalent volume of saline (for a 2 hours period). using the tail-

cuff method (Pfeffer et al.. 197 1 ). The dose of 2g ETOHIkg B. W. was chosen because.
as demonstrated in experiment 1. it induced a pronounced and long lasting increase in
plasma ANP levels.
2.1.3.23lood Alcohol C o n t a
50 pl of whole blood from each blood sample was deproteinized with 450 pl of ice-
cold trichloroacetic acid (6.15% w/v. Sigma) for estimation of the blooci ETOH levels
using the dehydrogenase enzymatic method (Hawkins et al.. 1966).
2.1.3.3. m u e Extraction
To estimate the content of ANP. ventricles as well as the left and right atria were
carefully removed and placed separately in ice-cold 0.1 N HCl. Tissues were rapidly
boiled for 5 minutes and then cooled on ice. Atria were homogenized with a
microultrasonic cell disrupter (Kondes. Vineland. N. J. ). whereas ventricles were
homogenized using a Polyuon. An aliquot was taken to measure the protein content using
the Bradford's method (Bradford. 1976). and the remaining homogenate was centrifuged
for 15 minutes at 4°C and supernatants were stored frozen at -75°C for estimation of the
ANP content using a specific RIA (Gutkowska et al.. 1984).
2.1.3.4. ~ r m o n for homoml memuremen&
Blood samples were collected in chilled eppendorf tubes (experiment 1 ) or 15 ml
conical centrifuge tubes (experiment 2) and proteases inhibitors were added to a final
concentration of ethylenediamine-retra-acetate (EDTA) ( 1 mglml), phenylmethylsulfonyl
tluoride (PMSF) (40 pl ImMIml) and Pepstatin A (20 pl 0.5 mM1ml). and were
immediately centrifuged at 4°C. The plasma was stored frozen at -75°C for subsequent
RIAS. for estimation of the ANP. AVP.B-endorphin. ACTH. corticosterone and
aldosterone content.

2.1.3.5. Cornparison between direct and e-ed ANP
Due to the scarcity of blood in each time interval of experiment 1. unextracted plasma
was used for estimation of the plasma ANP content (direct ANP RIA). Therefore. in order
to evaluate the sensitivity of the direct ANP assay. we injected i.p. 1 groups of rats (4 in
each group) with either saline or morphine. a known stimulus of ANP release (Gutkowska
et al.. 1986b) and sacrificed the animals 15 minutes following the injection. Trunk blood
was collected. and both direct and extracted ANP RIA were performed. As it is clearly
shown in Figure 1.1.1 .. though the absolute values of plasma ANP content are different.
both methods indicated a similar change in plasma ANP levels following morphine
injection. Thus. the direct assay is suitable to investigate the changes in plasma ANP
content following ETOH administration.
2.1.3.6. Radioimmunoassav (RIA) ~rocedura . . Determination of atrial (final dilution. 1: 15000). ventricular ( 1 : 1000) and plasma (50
or 100 ANP content was done using a direct second-antibody RIA, developed from che
method of Gutkowska et al.. 1984. The antiserum (produced by the immunization
procedure of Gutk~wska et al.. 1984. final dilution 1:30000) showed a good cross-
reactivity with other fragments of the pro-peptide. but less than 5% cross-reactivity with
oxidized ANP. The iodination of the peptide. using lactoperoxydase. was done as
described elsewhere (Gutkowska et al.. 1 %Va). 0.75 pghube was the minimum quantity
of ANP detecrable with this RIA. The intra- and inter-assay coefficients of variation were
3 .2 and 8.5% respectively.
Estimation of plasma content of B-endorphin-like irnmunoreactivity was performed
as described previously (Bourassa et al.. 1978) using an antiserum (final dilution. 1 :3OOOO)
specific to the C-terminal portion of B-endorphin( 1-3 1 ). This antiserum shows a 100 %
cross-reactivity with 8-1 ipotropin and w ith the non-acetylated and a-N-acetyl forms of O-
endorphin(1-3 1). 70% cross-reactivity with Bzndorphin(1-27). but no cross-reactivity with
ACTH. a and fi-melanotropin and a and gamma-endorphin (Gianoulakis. 1987).
Iodination of standard B-endorphin(1-31) to be used as tracer was donc using the

1 Saline Morphine
NON-EXTRACTE D
Figure 2.1.1. Effectofthei.p.injectionof morphineonthecirculatingANP levels using either extracted (A) or unextracted (B) plasma for the RIA. Values are expressed as Means *SEM (n=4). Levels of signlicance refer to the drfference between the treatments; ""P<O.OOI.

chloramine T method (Bourassa et al.. 1978). 10 pg/tube was the smallest quantity of R-
endorphin detectable with this RIA. Intra- and inter-assay coefficients of variation were
2.8 and 8.3 % respectively.
Plasma conicosterone levels were measured in ETOH extracted plasma samples. as
previousl y described (Krey et al.. 1975). using a specific antiserum (83- 163. Endocrine
Science. Tarzana. CA) which showed a small cross-reactivity with desoxycorticosterone
( ~ 4 % ) but not with cortisol. ['H 1-corticosterone was purchased from New Eng land Nuclear
(101 Cihnmol). The minimum level of detection with this assay was 10 pg/tube. The intra-
and inter-assay coefficients of variation were 10.4 and 14.4 % respectively .
Plasma ACTH was rneasured with a sensitive RIA (Orth, 1979). The antiserum
(Peninsula laboratories. final dilution. 1 :3OOûO) cross-reacted 1 0 % with ACTH( 1-39) and
ACTH(1-24) but less than 1 % with O-endorphin, a and 8-melanotropin. and a and B-
lipotropin. Iodinated ACTH( 1-34) was purchased from Incstar (Steel Water. MIN). The
minimal detectable level of plasma ACTH was about 0.3 pg/tube. Intra- and inter-assay
coefficients of variation were 5.4 and 10.7 % respectively.
Plasma aldosterone was determine in ETOH extracted samples using a sensitive RIA
previously described (Mayes er al., l97O). This antiserum (A3-375. Endocrine Science.
Tarzana. CA) shows a 0.04% cross-reactivity with adrenosterone and a 0.01% cross-
reactivity with corticosterone. ['H I-aldosterone was purchased from New England Nuclear
(100 Cilmmol). 10 pg/tube was the lowest amount of the steroid detectable using this
assay. The intra- and inter-assay coefficients of variation were 11.2 and 15.1 %
respect ive1 y.
Estimation of plasma AVP levels was performed in I ml of acetone extracted plasma
using a second-antibody RIA (Stowsky et al.. 1974). Iodination of the peptide was done
by the chloramine T method. The antiserum (Peninsula Laboratories. final dilution
1 :500ûû) showed a 3 % cross-reactivity with Lysn-vasopressin but no cross-reactivity w ith
ANP. 0.2 pglml was the smallest quantity of AVP detectable with this RIA. Intra- and
inter-assay coefficients of variation were 4 and 1 1 % respectively.

. . 2.1.3.7. Statacal
The data are presented as the mean S . E. M. The significance of difference among
the various groups was evaluated by a two-way analysis of variance (ANOVA). with time
as the first independent variable and trestment as the second independent variable.
followed by the Neuman-Keuls multiple cornparison test. Correlat ions were calculated
through the Pearson's r test. A P value of 50.05 was considered significant.
2.1.4. RESULTS
2.1.4.1. Ex~eriment 1
Injection of ETOH as a 40 % vlv solution resulted in a rapid increase in the blood
alcohol content (BAC). reaching maximum levels within l5 minutes following the i.p.
injection of Ig ETOH/kg B. W. (95.7 + 2.6 mgldl) and within 30 minutes following the
i.p. injection of 3g ETOHkg B. W. (705 + 1 1 mgldl) (Figure 2.1.2. ). The i.p. injection
of ETOH. but not of saline. induced a fast increase in the plasma ANP content. reaching
maximum concentrations within 15 minutes post-injection for both lg ETOHIkg B. W.
(43.4 + 3.6 pg1100 pl) and 2g ETOHIkg B.W. (63.0 f 3.7 pgllûû p l ) (Figure 2.1.3A.).
Following this initial increase. the plasma ANP content started to decl ine slowly toward
the basal levels. At 120 minutes post-injection. the ANP concentration in the plasma of the
rats injected with the Ig ETOH dose was still significantly higher than the plasma ANP
content prior to the ETOH injection. as well as the ANP content in the plasma of the
saline-injected animals (Figure 1.1.3A.). However. the ANP concentration in the plasma
of the rats injected with the lg ETOH dose was not significantly different frorn tliat in the
saline-injected animals at 30. 60 and 120 minutes post-injection. A two-way ANOVA with
time as the first independent variable and treatment as the second independent variable
indicated a significant effect of time (F,.,, =7 1-09; PrO.O 1) and treatment (F1.,, =MAO:
P~0 .0 1). and a significant interaction of time and treatment (F,,,=9.47: PcO.0 1). The i.p.
injection of both 1 and Zg ETOHIkg B.W.. but not of saline. induced an increase in the
plasma content of Bendophin (Figure 2.1.3 B. ) and corticosterone (Figure 1.1.3C. ). For
B-endorphin. significant differences due to time (F,.,, = 2 2 2 1 : P5O.O 1) . treatment
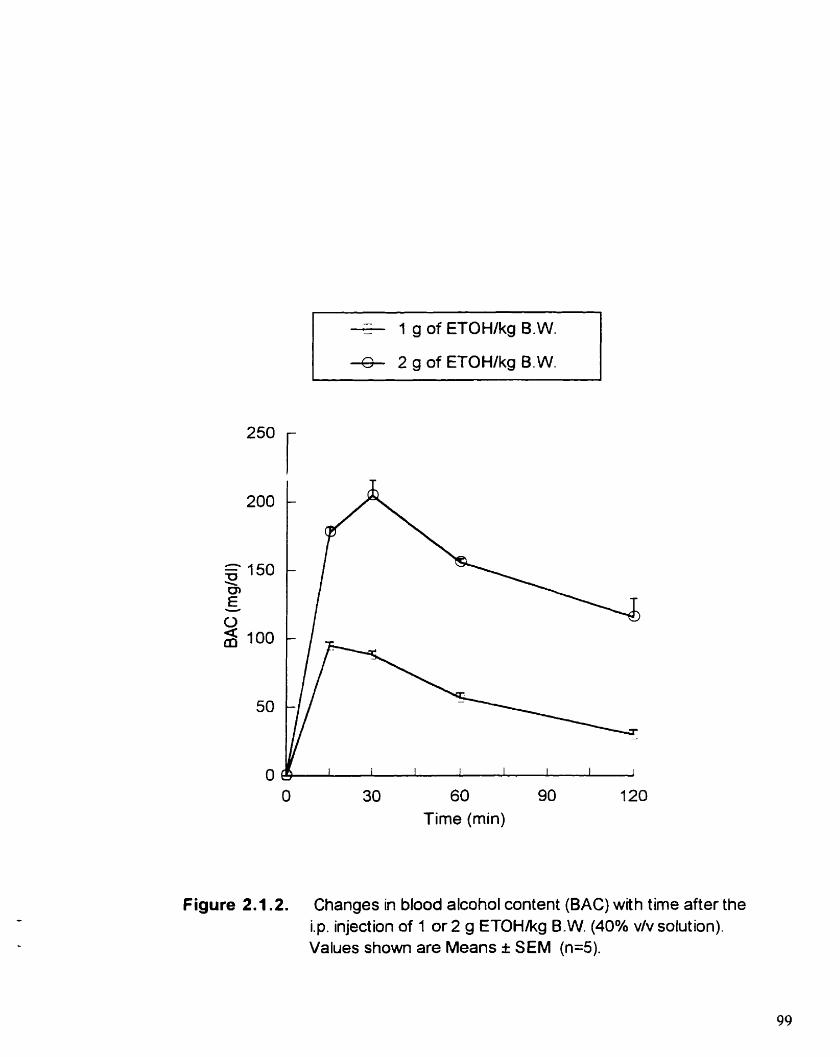
- -- - 1 1gofETOHlkgB.W.
++ 2 g of ETOHlkg B.W.
O 30 60 90 120 Time (min)
Figure 2.1.2. Changes in blood alcohol content (BAC) with time after the i.p. injection of 1 or 2 g ETOHkg B.W. (40% vfv solut ion). Values shown are Means I SEM (n=5).
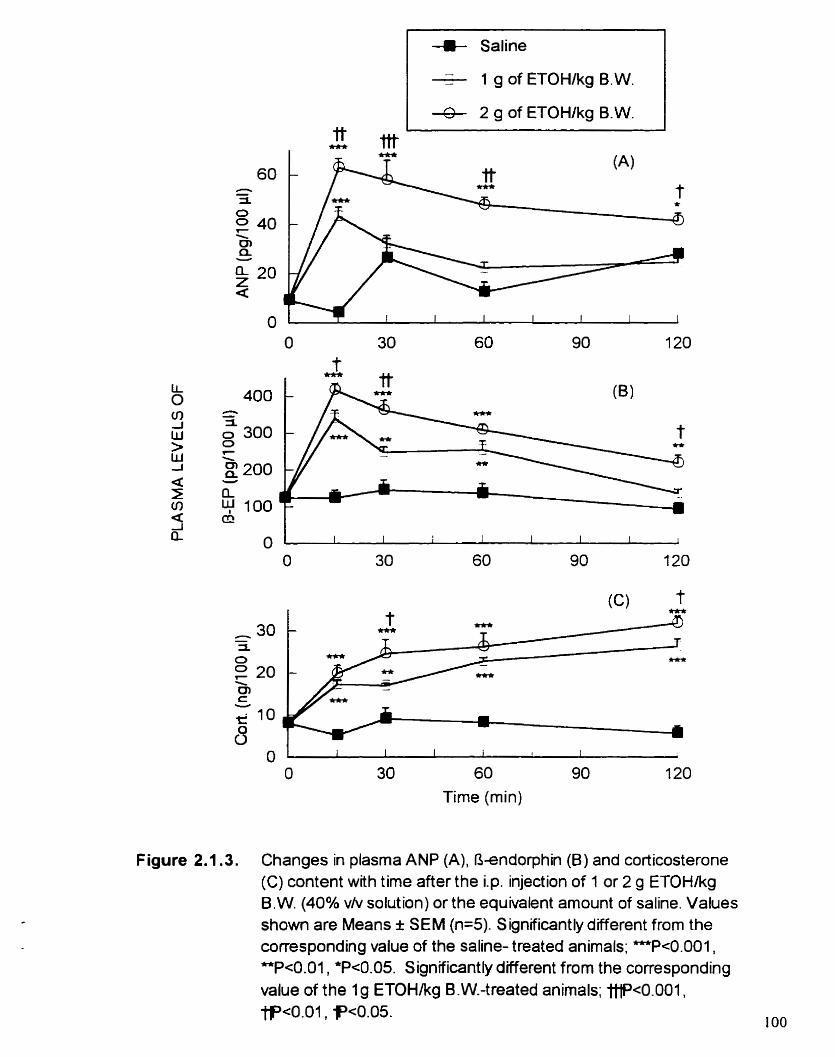
+ Saline
- 1 1gofETOHlkgB.W.
+ 2 g of ETOHlkg B.W.
o 1 I 1 I i 1 1 I
1
O 30 60 90 120 Time (min)
Figure 2.1 3. Changes in plasma ANP (A), Rendorphin (B) and corticosterone (C) content with time after the i.p. injection of 1 or 2 g ETOHlkg B.W. (40% vhr solution) or the equivalent amount of saline. Values shown are Means i SEM (n=5). Significantly drfferent from the corresponding value of the saline- treated animals; "P~0.00I1 "P<0.01, 'Pc0.05. Significantiydifferent from the corresponding value of the 1 g ETOHkg B.W.-treated animals; ~ ~ 0 . 0 0 1 , tp<O.OI, p<0.05.

(F,., =48.76: Pc0.01) and time by treatment interaction (F,.,=5.03: Pdl.01) were
observed. Similarly. for corticosterone. significant effects of time (F,.,, = 19.04: P4.0 1).
ueatrnent (F:.,, =76.O 1 : Pg0.0 1 ) and time by treatment interaction (F,., =6.56: PcO.0 1 )
were present.
2.1.4.2. -riment 2
Figure 4 presents the % change from basal levels (time O) in the concentration of
ANP in the plasma (Figure 7.1.4A.). the right (Figure 2-1-48.) and left (Figure 1 . 1 AC. )
atria and in the ventricles (Figure 2.1.4D.) of animals sacrificed at 15 and 120 minutes
following the i.p. injection of Zg ETOHIkg B.W. or saline. In agreement with the results
of experiment 1. the i.p. injection of 2g ETOHIkg B. W. induced an increase in the plasma
ANP content at 15 and 120 minutes post-injection. while i.p. injection of saline did not
induce statistically signiiïcant changes in the plasma ANP content either at 15 or at 120
minutes post-injection. The increase in the plasma ANP content was associated with a
decrease in the ANP content in the right and the lefi atria at both 15 and 120 minutes post-
ETOH injection. In the right atria. the ANP content changed from 6.90 + 0.45 pglrng
protein prior to ETOH to 4.41 + 0.33 and 5.61 f 0.5 1 pglmg protein at 15 and 120
minutes post-ETOH respective1 y. In the left atria. the i.p. ETOH injection decreased the
ANP content tiom 4.92 f 0.2 1 pglmg protein prior to the ETOH injection to 3.00 f 0.2 1
pglmg protein at 15 minutes and 3.27 f 0.15 pglmg protein at 120 minutes post-ETOH.
The i.p. injection of the equivalent volume of saline did not alter significantly the content
of A N P either in the right atria (6.90 + 0.45 pglmg protein at O minutes. 6.17 f 0.15
pglmg protein at 15 minutes and 6.42 + 0.24 pglmg protein at 120 minutes) or the left
atria (4.92 f 0.2 1 pglmg protein at O minutes. 4.74 0.30 pglmg protein at 15 minutes
and 4.1 1 + 0.42 pglmg protein at 120 minutes). Contrary to the atria. i.p. injection of
ETOH induced a slow increase in the tissue ANP content in the ventricles. from 306.8 f
15.5 ng/mg protein at time O (prior to the ETOH injection) to 355.6 + 3 1 .O nglmg protein
at 15 minutes and 651.4 & 19.9 nglmg protein at 120 minutes post-injection. The i.p.
injection of saline did not induce a significant change in the ventricular ANP content
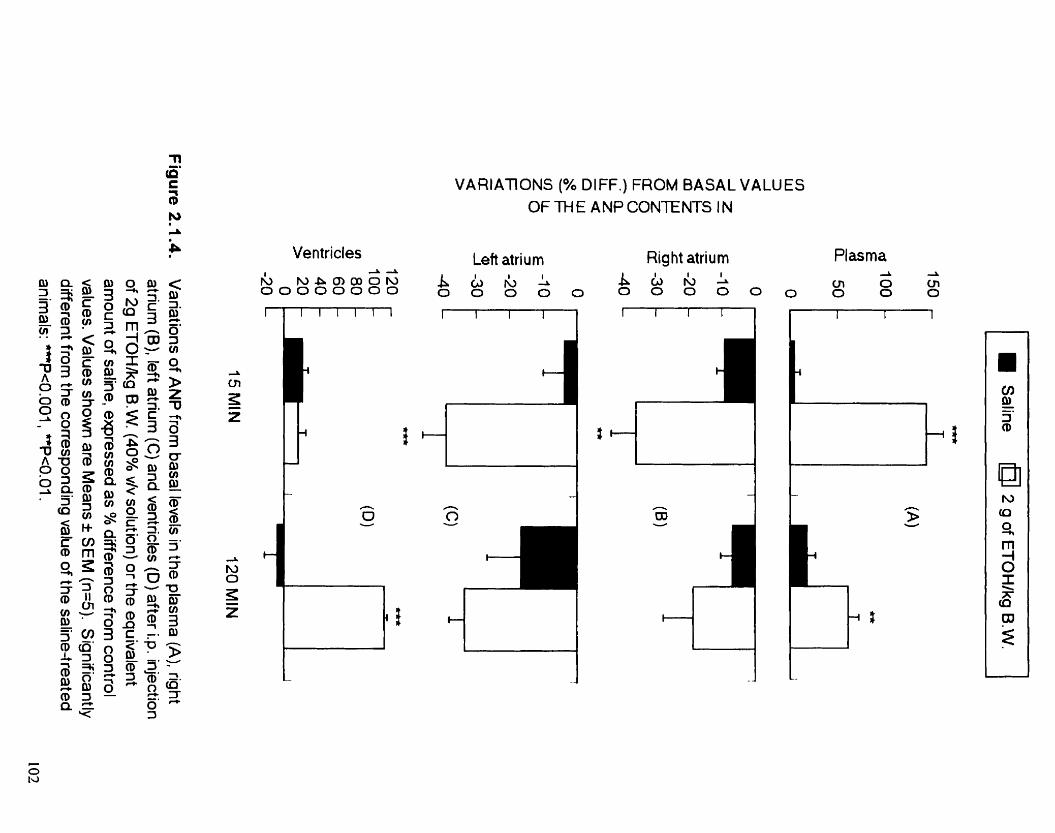
Ventricles
VARIATIONS (% DIFF.) FROM BASAL VALUES OF THE ANP CONTENTS IN
Left atrium Rig ht atrium Plasma

(370.6 + 14.2 ng/mg protein at 15 minutes and 184.6 + 41.1 nglmg protein at 120
minutes post-saline injection).
Table 2.1.1. shows the concentration of B-endorphin. corticosterone. ACTH.
aldosterone and AVP in the plasma of animals sacrificed without any injection (tirne 0) and
of animals sacrificed at 15 and 120 minutes following the i.p. injection of Ig ETOHikg
B.W.. or the equivalent volume of saline. ETOH induced an initial 3 fold increase in
plasma B-endorphin content at 15 minutes post-injection. At 2 hours post-ETOH. the
plasma O-endorphin levels were still 2 times higher than the basal levels. No significant
change in plasma B-endorphin content was observed following the i.p. injection of saline.
Although plasma ACTH levels presented a 6 fold increase over basal at 15 minutes and
only a 3 fold increase at 120 minutes post-ETOH injection. plasma corticosterone
presented a significant increase at 15 minutes (4 times higher than basal) and at 120
minutes post-injection (7 times higher than basal). As shown for R-endorphin. sa1 ine
treatment did not significantl y al ter the plasma ACTH and corricosterone content. In
contrat to ANP. fi-endorphin. ACTH and corticosterone. the plasma aldosterone levels
remained unaffected by either ETOH or saline treatments. at both 15 and 120 minutes
post-injection. Finally. plasma AVP levels were significantly decreased in the ETOH-
treated rats at 15 minutes cornpareci to both conuol and saline-treated animals. while at 170
minutes post-injection the plasma AVP levels were back to control values.
2.1.4.3.
The effects of the i.p. injection of Zg ETOHlkg B.W. or saline on the blood pressure
and hem rate are shown on Table 2.1 2. No significant variation in the blood pressure was
observed. However. a significant difference did developed in the heart rate between the
2 treatrnent groups. Within 15 minutes. heart rate values were significantly higher in the
ETOH-treated animals. and remained elevated for the remainder of the experiment .
2.1.4.4.
Since similar patterns of changes in the plasma R-endorphin and ANP levels were
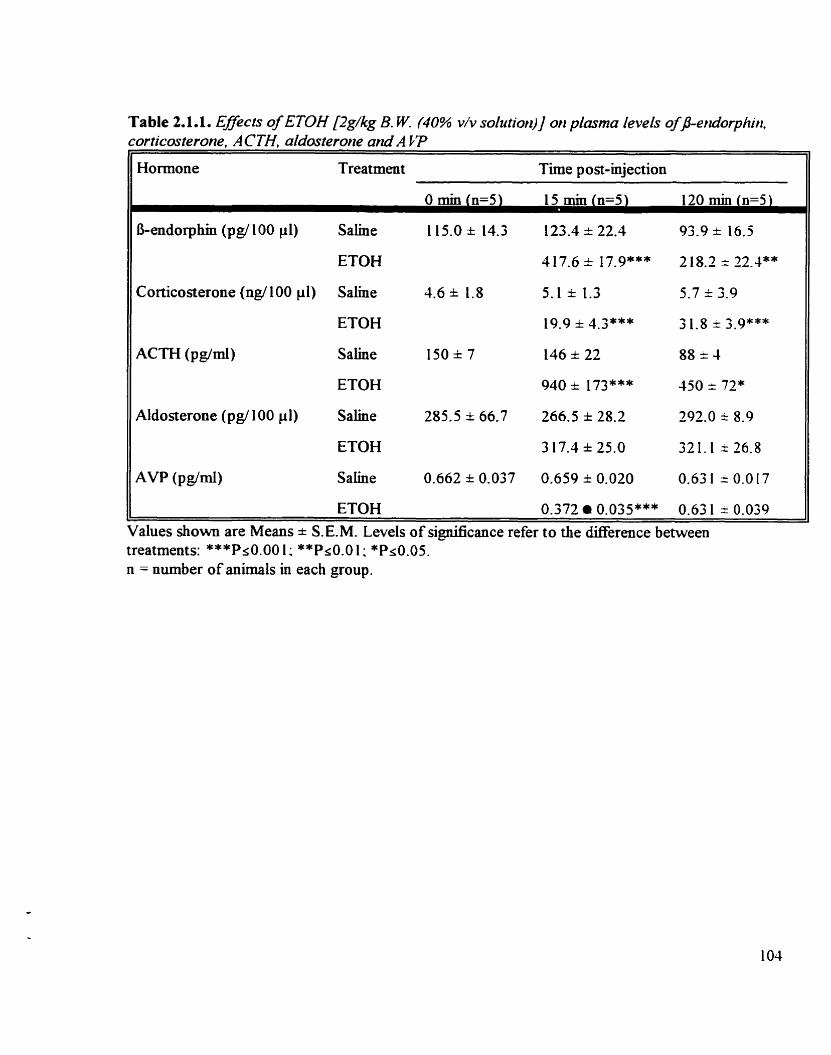
Table 2.1.1. Effects of ETOH [2g/kg B. W. (40% v/v solutzoti)] ori p l m a Ievels 00-eruiorphirz, cortzcmterone. A CTH. ido os ter or te ami A C.'P II Hormone
-
Treatment Time post-injection
Il-endorp hin (pg/ 1 O0 p 1) Saline
ETOH
Corticosterone (ng/ 1 O0 pl) Saline
ETOH
ACTH (pg/mU Saline
ETOH
Aldosterone (pg1100 pl) Saline
ETOH
AVP (pg/ml) Saline
ETOH 0.372 0.035*** 0.63 1 = 0.039 Values s h o w are Means * S.E.M. Levels of sigdicance refer to the ciifference between treatrnents: ***PsO.OO 1 : **Ps0.0 1 ; *Ps0.05. n = number of animals in each group.
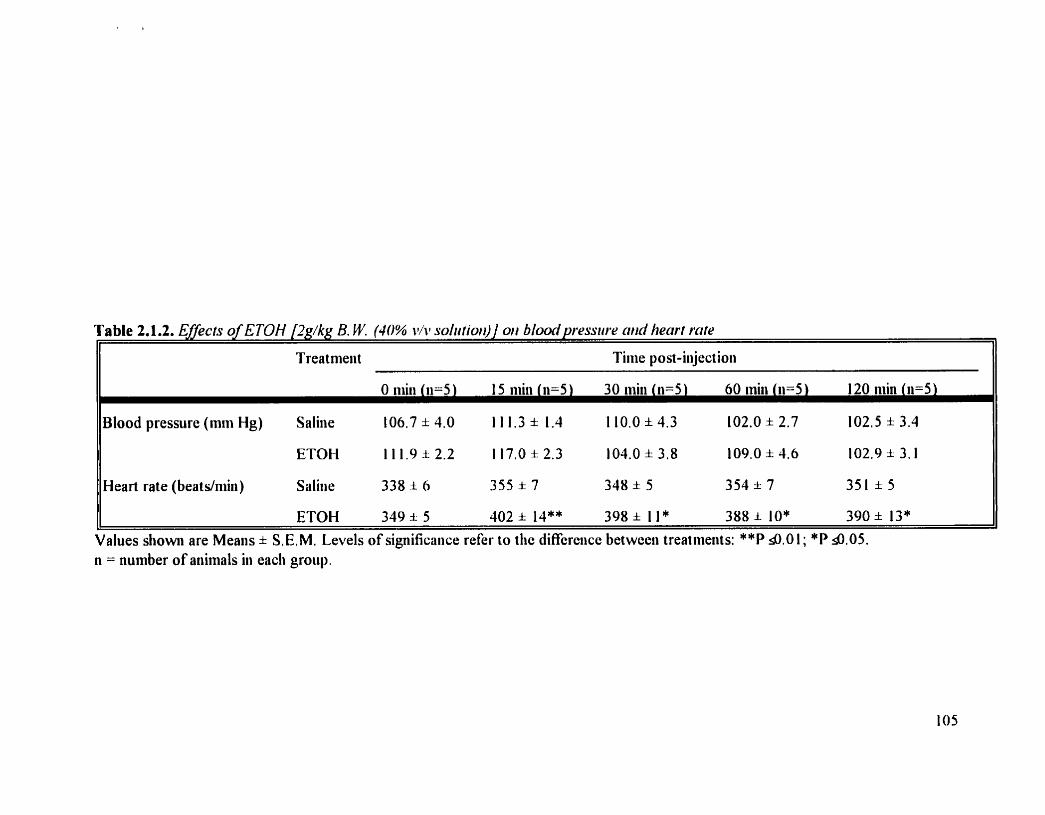
Table 2.1.2. Efiects ofETOH [&/kg B. W. (40% v/v sol~~tiotJJ ot, bloodpessrrtv otid hemt torite
Treatriieiit Tiiiie post-iiijectioii
ll~lood pressure (iniii Hg) Saline 106.7 * 4.0 I l 1.3 * 1.4 110.0 4.3 102.0 * 2.7 102.5 * 3.4
ll~eart rate (beatslniiii) Saline 338 k 6 3 5 5 * 7 348 * 5 354 11: 7 351 k 5
ETOH 349 i 5 402 k 14** 398 k 1 l * 388 k IO* 390k 13* Values show are Meaiis -t S.E. M. Levels of sigiiificaiice refer to the differeiice betweeii treatineiits: **P 4.0 1 ; *P d.05. ti = riumber of atiinials iii eacli group.

observed following the i.p. injection of ETOH. the presence of a positive correlation
between the ETOH-induced changes in the plasma 0-endorphin and ANP contents was
investigated. A positive and highly significant correlation (P<0.001) was found between
the 3 hormones (Figure 2.1.5.). Sirnilarly. positive correlations were also found between
ANP and BAC (Pc0.001) and between &-endorphin and BAC (P~0.01) respectively
(Figure 2.1.6A. and 2.1.68.). Moreover. a signitïcant inverse correlation (Figure 2.1.7. )
was observed between the decrease in atrial ANP content and the increase in plasma ANP
levels ( P ~ 0 . 0 1 ) at 15 minutes post-ETOH administration. Table 2.1.3. presents the
possible correlations between the parameters measured in the present studies. Interest ing l y.
an inverse correlation was also found between plasma ANP and plasma AVP levels. as
well as between BAC and plasma AVP.
2.1.5. DISCUSSION
The present investigations examined the acute in vivo e k t of the i.p. injection of
a moderate dose of ETOH on the circulating ANP levels as well as on the atrial and
ventricular ANP content. I t is important to note that due to the small arnount of blood
removed in the tirne-course study. a direct RIA for ANP was performed. using unextracted
plasma. This explains the somewhat higher plasma levels of ANP in the present study
compared to other publications. However. the major objective of this investigation was to
study the relative differences in the plasma ANP levels between the ETOH and saline
treated groups of animals. The direct plasma assay has been used previously in several
studies with reliable resulü. showing good correlations between the direct and the
extracted plasma assays (Steinhelper et al.. 1 990. B id mon et al., 199 1 ). Moreover . in the
present studies. the suitability of the direct assay was evaluated by estimating the plasma
A N P levels following the i.p. injection of morphine. which is known to increase ANP
release (Gutkowska et al., l986b). us ing unextracted (direct assay) and ex tracted plasma.
Indeed. the percent increase of plasma A N P levels using the direct assay was similar to
that obtained using a plasma extraction step prior to the RIA (Figure 2.1. L . ) . supporting

O 20 40 60 80 Plasma ANP (pg1100 pl)
Figure 2.1.5. Correlation between the ethanokinduced changes in the plasma ANP and Bendorphin (R-EP) contents. Values correspond to individual animais (n=75).
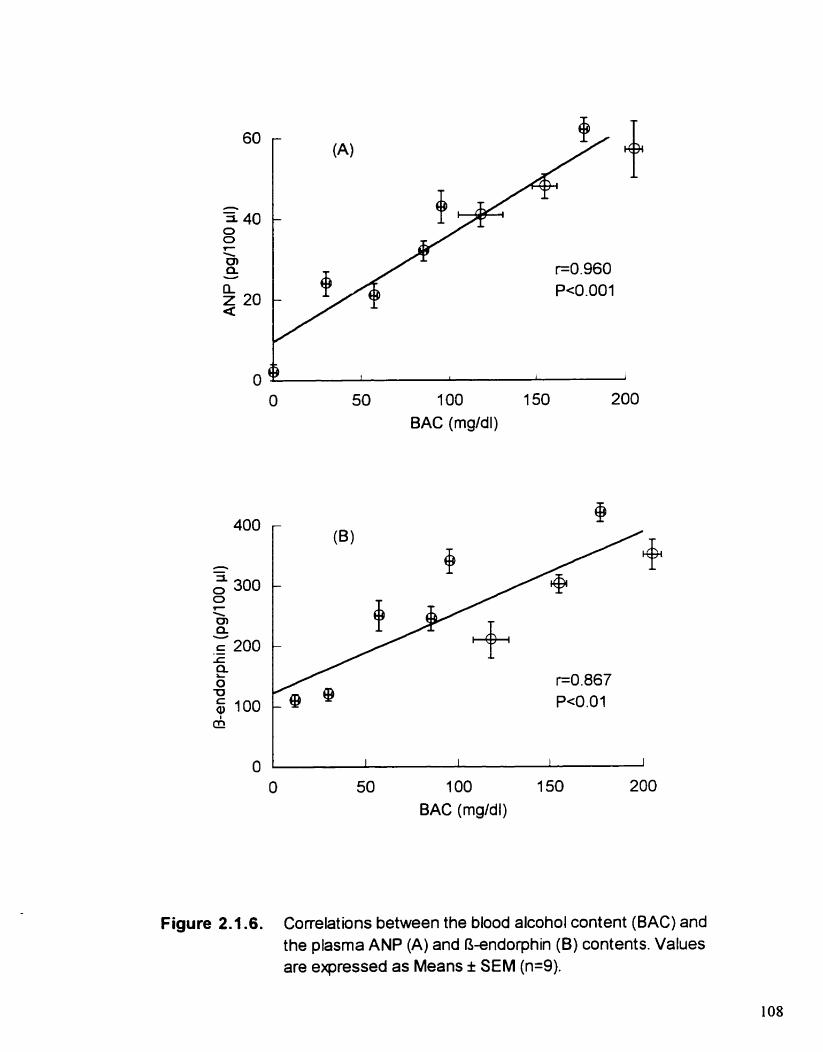
O 50 100 150 200 BAC (mgldl)
O 50 1 O0 150 200 BAC (mgldl)
Figure 2.1.6. Correlations between the blood alcohol content (BAC) and the plasma ANP (A) and Rendorphin (B) contents. Values are expressed as Means I SEM (n=9).

Plasma (pg1100 pl)
Plasma (pg1100 pl)
Figure 2.1.7. Correlations between the increase in plasma ANP levels and the corresponding decrease in right atrial (A) and left atrial (B) ANP content at 15 minutes post-ETOH. Values correspond to individual animais (n=20).
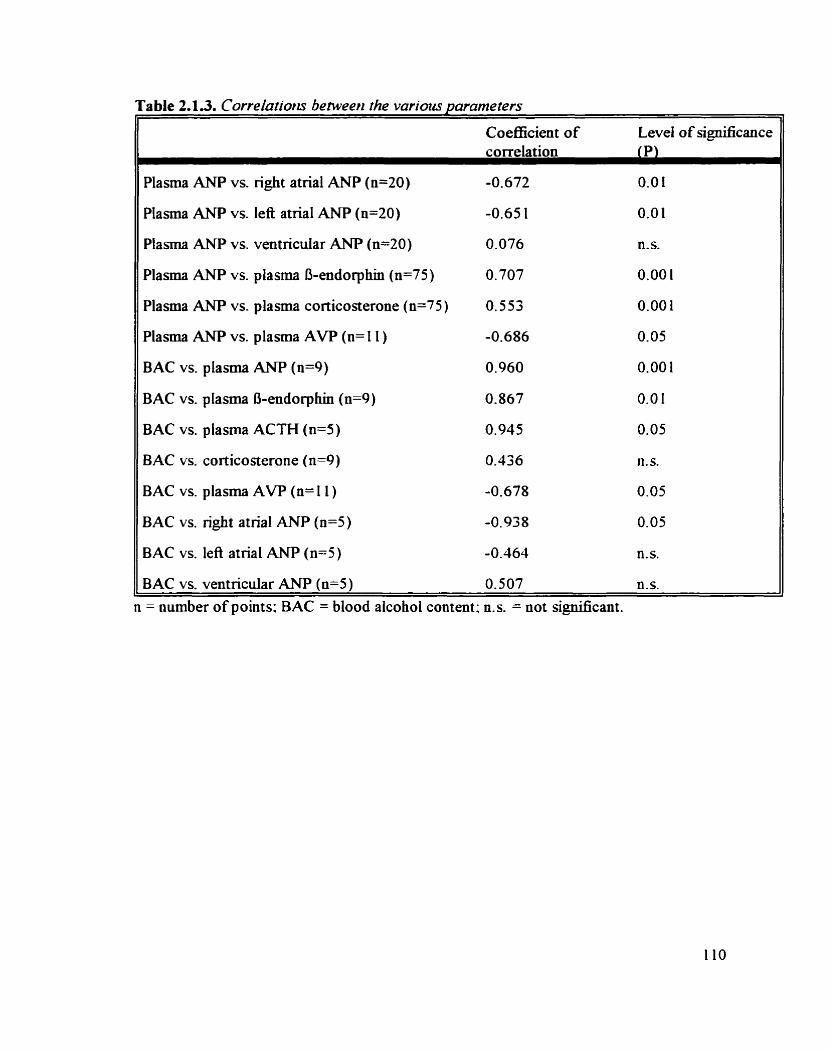
Table 2.1.3. Correlations betweerr the variais riarameters
Coefficient of Level o f sigdïcance
Plasma ANP vs. right atnal ANP ( ~ 2 0 )
Plasma ANP vs. left atnal ANP (n=20)
Plasma ANP vs. ventricular ANP (n=20)
Plasma ANP vs. plasma B-endorphin (n=75)
Plasma ANP vs. plasma corticosterone (n=75)
Plasma ANP vs. plasma AVP (n= 1 1 )
BAC vs. plasma ANP (n=9)
BAC vs. pIasma B-endorphin (n=9)
BAC vs. plasma ACTH (n=5)
BAC vs. corticosterone (n=9)
BAC vs. plasma AVP (n= 1 1 )
BAC vs. nght atnal ANP (n=5)
BAC vs. lefi atnal ANP (n=5)
BAC vs. ventncular ANP (n=5) 0.507 n. S.
n = number of points: BAC = blood alcohol content: n.s. = not significant.

the suitability of the direct plasma RIA for comparing changes in plasma ANP
concentrations following drug treatmenü or physiological conditions.
Results indicated that i.p. injection of 1 or 2g ETOHlkg B. W. induced an increase
in the circulating ANP levels in a dose-dependent manner. Funhermore. the ETOH-
mediated increase in plasma ANP levels was longer lasting following the 2 than the Ig
ETOH/kg B. W. dose. Since rats are rnetabolizing alcohol more quickly than humans
(Roine et al.. 1991). the high BAC obtained i n the present studies does not have the same
et'fect at the level of consciousness in the rat as it would have in humans. hdeed. BACs
that may produce coma or stupor in most humans subjects. in the rats produce some motor
incoordination but the animals remain fully conscious. as was observed in the present
studies and was reported by others investigators (Sparrow et al.. 1987). Therefore. in
many studies using experimental anirnals. doses up to 2g ETOHlkg B.W. (and BACs up
to 200 mg/dl) are considered moderate with respect to the effects of ETOH on
coordination and other behavioral responses (Subramanian et al.. 1990. Kettunen et al..
1993).
Acute ETOH has also been shown to increase plasma corticosterone (Ellis. 1966) and
ACTH (Rivier et al.. 1984) content. although this effect seems to depend on the ETOH
dose used and the time post-injection. Stress due to manipulation of the anirnals did not
seem to account for the observed increase in the plasma ANP levels. Indeed. the stable
baseline in plasma ACTH and corticosterone levels of the saline-treated anirnals was a
good indication that there was no activation of the hypothalam ic-h ypop hyseal-adrenal axis.
due to stress induced by the handling of the experirnental anirnals.
The increare in plasma ANP levels at both 15 and 120 minutes following the 2g ETOH/kg
B.W. dose was associated with a decrease in the atrial ANP content. in the atria. ANP is
known to be stored in its higher molecular weight form in the ce11 granules (Lang er al..
1992). The decreased atrial ANP content is therefore consistent with an increased
secretion, at a tirne when the increase in the rate of ANP biosynthesis is not yet
contributing to the tissue ANP content. Moreover. a significant inverse correlation was
observed between the changes in atrial ANP content and the changes in the circulating

ANP levels. In the ventricles. opposite to the atria. ANP is released in a constitutive
rnanner (Lang et al.. 1992) and there is no. or very little storage of ANP. Thus. an
increase in the rate of ANP biosynthesis would be associated with increased tissue content.
as was observed at 2 hours post-ETOH administration in the present studies and could
indicate an increased contribution of ventricular ANP in the plasma. Alternatively. the
increased ANP content in the ventricles could be due to an inhibitory effect of ETOH on
the ANP release by the ventricles. Thus. further studies estimating changes in the ANP
mRNA levels in this tissue at various intervals post-ETOH administration are needed to
understand better the effects of ETOH on the ventricular ANP system.
The ETOH-induced increase in plasma ANP levels is consistent with the tïndings of
Colantonio et al. (1991) in humans. where a drink of 1 g ETOHIkg B.W. induced a
significant increase in the plasma ANP content for the 2 hours of the study. In contrasr.
Leppaluoto et al. ( 1992) administering 1.5 g ETOHIkg B. W. and Hynynen et al. ( 1992)
administering 1 g ETOHIkg B.W. reported either a decrease. or no effect of ETOH. on
the plasma A N P content. The hypothesis put forward to explain the ETOH-ANP
interaction States that alcohol rapidly increases plasma osmolarity . which in turn increases
the release of ANP from the hem atria (Kurnik et al., 1991). Subsequently. the slower
ETOH-mediated dehydration and diuresis. induced at least in part by the ETOH-mediated
inhibition of AVP release (Pohorecky and Brick. 1988 and this report) decreases plasma
volume and atrial stretch. thus lowering A N P release. Any effect preventing the
hyperosmolarity observed following ETOH administration m ight prevent the increase in
ANP release. It is possible then that the initial volume-loading (prior to ETOH
administration) of the patients in the experiment of H ynynen et al. ( 1992) elevared plasma
ANP levels and masked any effect of the subsequent ETOH administration on further
release of the natriuretic peptide. Similarly. higher doses of ETOH. like those used in the
studies of Leppaluoto et al. (1992). might lead to a more rapid and more consistent
diuresis. an effect which will rapidly override the initial ETOH-induced hyperosmolarity.
Thus. lower doses of ETOH and initial isovolumetric conditions may be necessary to
demonstrate the ETOH-induced increase in plasma ANP levels.

Recently. Wigle et al. ( l993a). in the only reponed study on the effects of acute
ETOH administration on ANP using an animal model (Sprague-Dawley rats). reported a
lack of effect of ETOH on plasma ANP levels for 2 hours following the administration of
a high ETOH dose (5g/kg B. W.) through gastric gavage. while animals receiving an equal
volume of water presented a signifiant increase in plasma ANP levels. Interestingly. both
ETOH and water treated animals presented a decrease in the ventricular ANP content and
an increase in the atriai ANP content. A possible explanation for these changes could be
that the high amount of liquid administered in this particular study resulted in a water-
loading effecr. with a rapid decrease in the ventricular ANP content and a slower increase
in the atrial ANP content of both the ETOH and water treated animals. indicating
increased ANP release and availability to cope with this water-loading situation (Wigle et
al.. l993a). Knowing that circulating ANP is mostly derived from the h e m (Nakao et al..
1993). it is surprising that despite the similar changes in heart ANP contents. the ETOH-
given rats exhibited lower plasma ANP levels than their controls. However. it is possible
that the tremendous diuresis usually associated witli a very high dose of ETOH (and
therefore decreased plasma volume) may explain the decrease or rather. the absence of
increase. in plasma ANP levels. Moreover. such high doses of ETOH and the route of
administration may cause gastric irritation and increase sympathetic activi ty. therefore
interfering with the interactions of the drug with various physiological systems.
Furthermore. some investigators (Colantonio et al.. 199 1 ) have proposed an indirect
effect of ETOH on the activity of the ANP system. mediated by its effects on various
hormonal and neurotransmitter systems. among which could be the endogenous opioid
peptide systems. Experimental evidence has indicated that opioids may play a significant
role in controlling ANP release (Gutkowska et al.. 1986b. Widera et al.. 1997). Indeed.
p-opioid receptor agonists. like fentanyl. have been shown to increase ANP release
(Vollrnar et al.. 1987. Pesonen et al.. 1990). while naloxone (an opioid receptor
antagonist) has been reported to significantiy suppress the increase in plasma ANP levels
follow ing water-immersion in human (W idera n al., 1992). Moreover. a'-adrenerg ic
agonists. such as clonidine. also stimulate ANP, via activation of central opioid receptors

(Pan and Gutkowska. 1988). Interestingly. naloxone also suppressed the water immersion-
induced decrease in blood pressure. suggesting a definite role of the endogenous opioid
system in the conuol of ANP and its subsequent effect on blood pressure. Since ETOH has
long been shown to induce a rapid increase in plasma O-endorphin in both humans and
experimental animals (Gianoulakis. 1989). it was interesting to determine whether a
correlation between the ETOH-induced changes of plasma ANP and fi-endorphin could be
detected in the present study. Indeed. a strong. positive association (r=0.707. Pc0.001)
was found between the effects of ETOH on plasma ANP and O-endorphin levels.
Therefore. the increased plasma levels of A N P following injection of a low doses of
ETOH may represent the additive effect of the ETOH-induced increase in both osmolarity
and plasma levels of endogenous opioid peptides. such as O-endorphin. The presence of
an indirect effect of ETOH on the heart A N P system is further supported by the
observation that exposure of cultured myocytes to ETOH did not alter the rate of ANP
release (Wigle et al.. 1993a).
The effect of acute ETOH on the blood pressure is still not fully understood. Several
studies (Eisenhofer et ai.. 1984. Howes et al.. 1985. Eisenhofer et al.. 1987. Sparrow et
ai.. i987. Maiinowska et al.. 1989. Adesso et al.. 1990. Kawano et al., 1992) have
reported a decrease. wh ile others (Ireland et al.. 1984. Howes et ai.. 1986. Potter et al..
1986. Maheswaran et ai.. 1991) have shown an increase or no change (Stott et al.. 1987)
in blood pressure following ETOH ingestion. The renin-ang iotensin-aldosterone system
is not believed to play an important role in the acute effects of ETOH on blood pressure.
and any changes observed in the activity of the renin-ang iotensin-aldosterone sys tem may
be secondary to the variations in sympathetic activity and blood pressure (Stott et al..
1987). The absence of ETOH-induced changes in the plasma levels of aldosterone. which
was also seen in the present studies. provide further support to this hypothesis. The
elevated h a r t rate. the only consistent observation in the various investigations (also seen
in the present studies). may be due to : (a) a reflex mechanism to maintain blood pressure
(Kawano et al.. 1992): (b) a decrease in blood volume (Stott et al.. 1987): ( c ) ETOH
metabolites. such as acetaldehyde and their mediated secondary increase in catecholarn ine

(Potter et al., 1986); and ( d ) CNS-mediated effects (Malinowska et al,, 1989).
Several hypothesis have been put forward to explain the effects of ETOH on the
blood pressure. Various confounding factors. like the dose of ETOH. its duration of
exposure. genetic predispositions. nutritional status. timing of measurement fol low ing the
ETOH administration. or the route of ETOH administration rnay play an important role
and must be taken into account when considering the general effect of ETOH on various
physiological systems (Beilin et Puddey. 1992). At the vascular level. ETOH could cause
vasoconstriction by decreasing the clearance of Norepinephrine (Howes et al.. 1986). by
a translocation of Ca" from plasma to smooth muscle cells (Potter et al.. 1986) or by
decreasing the vasorelaxation induced by Acetylcholine and thus affecting the release of
Endothelium-Derived Relaxing Factor (EDRF) from the endothelial cells (Hatake et al..
1993. Criscione el al.. 1989). On the other hand. ETOH may also cause vasodilation by
a direct effect on arterioles. capillaries and venules (Altura and Altura. 1981) or by an
inhibitory eftèct on the a-adrenoceptors induced vasoconstriction ( Eisenhofer et al. 1984).
I t has also been proposed that the rapid increase in blood pressure. seen in sorne studies
following acute ETOH exposure. could be simply due to the caloric load of ETOH. similar
to chat observed after a glucose load (Stott et al.. 1987). More recently. Kawano et al.
(1992) suggested a biphasic effect of ETOH in man. with a transient increase in blood
pressure observed within the first hour. followed by a sustained hypotension due CO
peripheral vasod i lation (associated w ith increased Cardiac Output and decreased Total
Peripheral Raistance) for up to 8 hours. In this model. sympathetic activity is initiated by
the vasodilatory effects of ETOH. and prevents either directly. or through an action on
renin, or by increasing the heart rate. the development of a more severe hypotension.
From our study. it seems that ANP could be added to chis growing number of hormonal
factors implicated in the complex action of ETOH on the cardiovascular system. Consistent
with the mode1 of Kawano et al.. (1992). an acute elevation of circulating ANP could help
modulate the effects of ETOH on the vasculature and the kidneys. and could prevent the
initial activation of counterbalancing systems by iü inhibition of sympathetic activity and

its suppression of renin and aldosterone release. The body could therefore achieve a
s igni ficant decrease in blood pressure. before the stimulus for compensatory actions is
strong enough to override the initial hypotensive effect of ANP. Again. the elevated heart
rate, observed in the present studies. is seen as the result of those compensatory
rnechanisms . In addition. extra airial ANP systems may also play an important role in the ETOH
induced diuresis and blood pressure changes. For example. ANP is known to suppress the
release of AVP from the supra-optic nucleus (SON) (Clark et al.. 1991) and an increase
in central ANP levels could be responsible. at least in part. for the ETOH mediated
decrease in AVP release reported previousl y (Gutkowska and Nemer. 1989) and observed
in the present studies. Whether the decrease in circulating AVP levels following the ETOH
administration is the result of a direct effect of ETOH or is mediated by the increase in
central and peripheral ANP activity rernains to be evaluated.
H igh ETOH drinking is associated with an increased prevalence of hypertension
(MacMahon. 1987). On the other hand. low doses of ETOH are associated with
signi tkant lower blood pressures as indicated by a number of epidemiological studies
demonstrating a non4 inear association between increased ETOH consump t ion and blood
pressure (the J or U shaped curves) (MacMahon. 1987). Interestingly. a similar non-linear
association is now well documented between alcohol consumption and cardiovascular
diseases (Marmot and Brunner. 199 1 ). Variations in Lipoproteins (HDL and LDL)
(Criqui. 1986. Hojnacki et al. 1988) or an attenuation of the vasoconstrictor responses to
Norepinephrine (Criscione et al., 1989) with low doses of ETOH. are a few proposed
rnechanisms for this protective effect of low ETOH consumption on hypertension and
cardiovascular diseases. The present investigations suggest that ANP could be an additional
rnechanism involved in the protective effects of chronic low ETOH consumption on the
blood pressure. Chronic studies are presently under way to study this hypothesis.
In conclusion. acute ETOH has been shown to significantly and rapidly increase
plasma ANP levels, associated with a decrease of the atrial A N P content. This increase in
circulating ANP levels could be mediated in part by the ETOH-induced increase in the

plasma content of endogenous opioid peptides, such as fi-endorphin. S tudies of chronic
ethanol treatment as well as evaluation of the effects of ETOH on the extraatrial natriuretic
peptide system. are necessary before any conclusions could be drawn on the possible
importance of ANP in the prevention of the age-dependent hypertension observed with
regular low alcohol consumption.
2.1.6. ACKNOWLEDGEMENTS
The authors wish to ihank Mr Ricardo Claudio and Mme Diane Beaudry for technical
assistance. as well as Dr Dominique Walker for the generous donation of the ACTH
tracer. This work was supported by a grant of MRC of Canada (MT-10337). by the
Canadian Heart and Stroke Foundation (J.G.) and by the Alcohol Research Program ar
Douglas Hospital ( C G . ) . P.G. is a recipient of a scholarship from the "Fonds pour la
formation de Chercheurs et ['Aide à la Recherche" (FCAR).

Section 2.2.
INCREASED PLASMA ATRIAL NATRIURETIC PEPTIDE
AFTER INGESTION OF LOW DOSES OF ETHANOL IN HUMANS
C. Gianodakis, P. Guillaume, J. Thavundayil and J. Gutkowska
Alcoholism: Clinicd and Experimental Rerearch
Wcohol Clin Grp Res, in press, 1997)
Contribution by CO-authors: 1 was in charge of the AVP radioimmunoassay, the urine volume
and urine electrolytes measurements. I took an active part in the writing and editmg of the
manuscript.

2.2. INCREASED PLASMA ATRIAL NATRIURETIC PEPTIDE
AFIXR INGESTION OF LOW DOSES OF ETHANOL IN
HUMANS
2.2.1. ABSTRACT
Previous studies have demonstrated that in human, acute consurnption of high doses
of ethanol (ETOH). administered in a large quantity of fluid. with or without volume-
loading. induced either a decrease or an increase in the plasma content of atrial natriuretic
peptide (ANP). a substance which has a hypotensive effect. The objective of the present
investigations was <O examine the effect of low doses of ETOH (0: 0.75: and 0.50 g
ETOH per kg. B. W.) administered to 6 normotensive individuals. in a small volume of
fluid without prior volume-loading. Prior to and at various intervals following
administration of the placebo or ETOH drinks the heart rate and blood pressure (BP) were
measured and blood samples were taken for estimation of the plasma ANP. arginine
vasopressin (AVP) and cortisol contents. Results indicated small changes in BP and heart
rate following ingestion of either the placebo or ETOH drinks. On the other hand a
signifiant increase in the plasma ANP content was observed at 15 min following ingestion
of both the 0.15 and 0.50 g ETOH per kg B.W. doses. but not following the placebo
drink. The plasma ANP levels were still elevated at 45 min post ETOH intake. bur had
returned to basal levels at 120 min after the ETOH drink. Interestingly. it was noticed that
the higher dose of ETOH (0.50 g) did not induce a higher plasma ANP concentration than
the Iower dose (0.25 g) of ETOH. however the plasma ANP content remained elevated for
a longer period. Furthermore. the increase in plasma ANP content was not due to ETOH
or stress induced increases in the plasma AVP and cortisol contents. since the plasma
concentration of these hormones remained either at basal or below basal levels for the
duration of the experiment. In conclusion. ingestion of low amounts of ETOH equivalent
to 1 or 2 standard drinks induced an increase in plasma A N P content.

2.2.2. INTRODUCTION
A number of epidemiological studies have indicated that though high (more than 1
dr i n ks per day ) ethanol ( ETOH) consump t ion induces hypertension. low to moderate
ETOH consumption ( 1-3 drinks per day) not only does not cause hypertension. but it
seems to prevent or delay the development of the age dependent increase in blood pressure
(BP) (Klatsky et al.. 1977: Gleiberman and Harburg. 1986: MacMahon. 1987). Thus. it
was observed that the BP of non-drinkers was higher than the BP of low ETOH drinkers
matched forage. sex. obesity etc (MacMahon. 1987). Since ETOH does not have specitïc
receptors to interact with. the mechanism by which it induces its antihypertensive effect
is not known. In general. it is accepted that ETOH alters the activity of a number of
neurotransmitter and hormonal systems which in turn mediate a number of its effects. I t
is reasonable then to suggest that low ETOH intake inhibits the activity of substances
having a pressor effect and/or stimulates the activity of substances having antihypertensive
effect. The mechanisms or physiolog ical systems mediating this antihypertensive effect of
ETOH are not known. One such mechanism may be the inhibition of the vasoconstrictor
effect of norepinephrine by small but not by high concentrations of ETOH (Criscione et
al.. 1989). Another suggested mechanism may be associated with the effect of ETOH on
the concentration of plasma Very Low Density Lipoproteins (VLDLs) and High Density
Lipoproteins (HDLs) (Criqui. 1986). A trial Natriuretic Peptide ( ANP) is a 28-am ino acid
peptide which has been shown to have a hypotensive effect (Cantin and Genest. 1987).
Though ANP was originaily isolated from the heart atria (De Bold et al.. 1981). its
presence has also been demonstrated in the heart ventricles. and in a nurnber of neuronal
and endocrine tissues such as the brain. pituitary gland. ganglia of the autonornic nervous
system and the lungs (Samson. 1985: Cantin and Genest. 1987: Gurkowska et al.. 1982:
Gutkowska and Nemer. 1989). ANP has been shown to decrease the BP by inducing
vasorelaxation, by increasing diuresis and natriuresis and decreasing the activity of a
number of pressor hormones and /or neurotransmitters such as vasopressin. aldosterone.
angiotensin I I . renin and norepinephrine (Cantin and Genest. 1987: Gutkowska and
Nemer. 1989). Thus, it has been hypothesized that part of the antihypertensive effect of

low ETOH consumption may be mediated by the activation of the ANP system (Colantonio
et al., 1991). It is possible that following low ETOH consumption ANP is released into
the circulation inducing a transient hypotensive effect. In contrat. when the ETOH
consumption is high. in addition to ANP various pressor systems are activated. such as
catecholarnines. which evennially may lead to the development of hypertension in chronic
alcoholics. However, the published reports on the effect of acute ETOH ingestion on the
ANP system in human have indicated controversial results. since an increase (Colantonio
et al.. 199 1 ). a decrease (Leppaluoto et al.. 1992) or no change (Hynynen ef al.. 1991)
in plasma ANP levels have been reported following ETOH intake. These different effects
of ETOH on plasma ANP content rnay be due to differences in the experirnental conditions
between the various studies. such as differences in the dose of ETOH administered. the
volume in which die ETOH was administered. previous loading of the subjects. time when
the study was performed and the nutritional state of the subjects.
Interestingly. al1 the studies investigating the effect of ETOH on plasma ANP content
have used doses of ETOH. ranging from 0.5 to 1.5 g ETOH / kg B.W. . adrninistered in
large volumes of fluid (from 500 to 750 ml). wirh (Hynynen et al.. 1992) or without
(Colantonio et al.. 1991 : Leppaluoto et al.. 1992: Ekman et al.. 1994) previous volume-
loading of the subjects. Volume loading could itself trigger ANP release (Lang et al..
1985). In fact presently. there are no published studies investigating the effect of low doses
of ETOH administered in smdl volumes of tluid. Thus. it was the objective of the present
investigations to examine the effect of low doses of ETOH (0. 0.15. and 0.50 g ETOH per
kg B.W.) administered in a mal1 volume of tluid. on the circulating ANP levels at various
intervals post-drink. in 6 normotensive individuals. These doses of ETOH were chosen
because when taken by a 60-70 kg B.W. individual. they correspond to one (0.15 g
ETOH) and two (0.50 g ETOH) standard drinks. which the epidemiological studies have
shown to have a protective effect on the age dependent development of hypertension. A
standard drink is equivalent to approximately 14 g of ETOH. In addition to plasma ANP.
the plasma content of vasopressin (AVP) and cortisol were estimated. Furthermore. the
BP. heart rate and urine excretion were recorded during the 3 hrs of the experimental

per iod .
2.2.3. MATERIALS AND METHODS
2.2.3.1.
Six (4 males and 2 fernales) Caucasian normotensive individuals. 15-30 years old
with body weight between 59 to 73 kg (65 + 2) participated in the study. All subjects
were tested for excess alcohol consumption using the Michigan Alcohol ism Screening Test
(MAST) (Selzer. 197 1 ), and a drinking behaviour interview (Shelton et al.. 1969). The
subjects consumed alcohol socially. but none of the subjects was an alcoholic at the time
of testing. In addition. al1 subjects underwent a complete rnedical examination. lndividuals
under chronic medication suffering from diabetes or hypertension. or having liver. brain.
kidney. or other chronic diseases as well as subjects with high ETOH consumption were
excluded from the study. Furthermore. the subjecü were not taking any over the counter
med icat ion or steroidal antiintlammatory drugs. The fernale subjecü were not on birtli
control medication and were tested during the fol1 icular phase of the reproductive cycle.
The investigations were approved by the Douglas Hospital H urnan Research Comm ittee.
and were conducted in accordance with the guidelines proposed in the declaration of
Helsinki. Informed consent was obtained from al1 subjects.
2.2.3.2. merimental Design
Al1 subjects abstained from alcohol for 48 hr prior to testing. Subjects participated
in three testing sessions. In the tïrst testing session subjects received a placebo drink
consisting of unsweetened orange juice and tonic water (2 parts orange juice 1 part tonic
water). The g las was dipped in ETOH to provide the smell and taste of ETOH. On the
second and third testing sessions. the subjects were randomly given either 0.25 g ETOH
per kg B. W. (as a mixture of 1 part 95 % ETOH 5 parts unsweetened orange juice). or
0.50 g ETOH per kg B. W.. (as a mixture of 1 part 95 % ETOH 2 parts unsweetened
orange juice). The total volume of each drink was between 1 16- 145 ml. The exact volume

depended on the subject's body weight. However. for the same subject the same volume
of tluid was ingested in ail 3 testing sessions. There was a minimum of one week interval
between testing sessions.
On the day of testing. subjects arrived at the research unit early in the morning
without having breakfast. A catheter was placed in the lefi arrn vein at 8 a m . The subjects
were allowed to rest for 1 hour. To avoid the feeling of nausea. often occurring following
ETOH ingestion with an empty stomach. which couid act as a stressor and release
hormones known to influence the ANP secretion (cortisol. epinephrine. norepinephrine).
following placement of the catheter the subjects were given a light breakfast consisting of
a toast without butter or marmalade. They were allowed to have 100 ml of water with their
breakfast, but not tea or coffee and were not allowed to smoke for the duration of the
testing session. 60 min afier placing the i.v. catheter. a 10 ml blood sample was drawn and
BP and hem rate were measured. Then. the subjects were given the placebo or the
appropriate ETOH drink. which they were requested to drink within 5 min. Additional 10
ml blood samples were drawn at 15. 45. 110 and 180 min post drink. These time intervals
were chosen based on the blood ETOH content following ETOH ingestion which rises
sharply within 15 min. reaches maximum within 45 to 60 min and then decreases
gradually. so that at 3 hr post drink the alcohol in blood is low but still measurable.
Following the drawing of the blood samples. BP and heart rate were measured while the
subject was lying in bed. During the intervals between blood sampling. the subjects were
allowed to get up. sit in the living room of the research unit, watch television. read and
converse with research unit personnel. The subjects collected the urine for estimation of
the total volume excreted during the 3 hrs of the experirnental period. as well as Na' and
K' excretion. The content of Na' and K' in urine was measured direcily by a tlame
p hotometer (Instrumentation Laboratory Autoanalyser. 1 L943).
2.2.3.3. Estimation of Blood Alcohol Content
To estirnate the blood alcohol content, 750 pl of whole blood were rnixed with 1 .O
ml of 6 % ice cold perchloric acid and centrifuged at 4°C. The supernatant was kept

frozen at -75°C till the time of estimation of the blood alcohol content using the alcohol
dehydrogenase enzymatic method (Hawkins et al.. 1966).
2.2.3.4. Estimation of Plasmê ANP. AVP and Cortisol Contents
10 ml of blood were drawn and placed in ice-chilled EDTA vacutainer tubes
containing 100 ui of 10 mM pepstatin A (Sigma Chernical Co. St. Louis MO. USA. No
P-4265) to prevent degradation of ANP and AVP. Blood samples were centrifuged at 4°C
and the plasma was stored at -75 "C until the tirne of the assay.
The ANP content in the plasma was estimated using a sensitive RIA as described
previously (Gutkowska et al.. 1985). The antibody for ANP was raised in the laboratory
of Dr. Gutkowska (Gutkowska et al.. 1985). Prior to RIA. ANP peptides were extracted
from 1 ml of plasma using C-18 Sep-Pak Cartridges (Waters Scientific Mississauga
Ontario. Canada). The iodination of ANP, .:, was perforined using the lactoperoxidase
rnethod (Gutkowska et ai.. 1987a). The minimum quantity of ANP detectable with this
RIA was 0.12 fmoles/tube. AI1 samples were analyzed in the sarne assay to avoid the
between assays variation. The intra-assay coefficient of variation was 6.8 % .
Estimation of plasma AVP content was performed in 1 ml of acetone-extracted
plasma (Skowsky er al.. 1974). The antisemm was purchased from Peninsula Laboratories
(Belmont CA. USA) and was used at a final dilution of 1:50000. I r presented a 3 % cross-
reactivity with Lys,-vasopressin and no cross-reactivity with ANP. AVP was iodinated
using the chloramine T method. The sensitivity of the antiserum was 0.2 pg/ml. Al1
samples were analyzed in the same assay to avoid the between the assays variation. The
intra-assay coefficient of variation was 4 % .
For estimation of the plasma cortisol levels. plasma was extracted in absolute ETOH
and cortisol was determined by RIA according to previously pub1 ished procedure (Krey
et al., 1975). using a cortisol antiserurn (F3-3 14: Endocrine Sciences. Tarzana. CA. USA)
at a final dilution of 1 : 1000. The intra-assay coefficient of variation was 9 % . Of the
physiologically occurring steroids tested for cross-reactivity only cortisone and
desoxycortisol presented a signifiant cross-reactivity with the antibody. 4.7 % and 7.5 %

respect ive1 y. The contribution of these cross-reactions in normal human plasma
determinations is small due to the ordinarily low concentrations of desoxycortisol and
cortisone relative to cortisol. Total cortisol (bound plus free) was determined by this
method.
. . 2.2.3.5. Statistical A-
Statistical evaluation of hormone content and other physiological measures was done
by 2 way analysis of variance (ANOVA) for repeated measures across time and dose with
the various sample times as the B n t variable and treatment (placebo. 0.15 g and 0.50 g
ETOHlkg B.W.) as the second variable. The ANOVA was followed by the Neuman-Keuls
multiple comparison test to determine the significance of difference between the different
treatments and time intervals of testing. A value of p < 0.05 was considered signitïcant.
2.2.4. RESULTS
in Figure L I . 1.. the changes in blood alcohol content with time after ingestion of the
ETOH drinks are shown. The concentration of ETOH in the circulation increased sharpl y
within the first 15 min and was still high at 45 and 120 min. The ETOH was cleared from
the blood slowly so that it was still detectable at 180 min after the ETOH drink (Fig.
7.7.1.). In Figure 2.2.2.. the changes in heart rate (Fig. 2.2.2A. B and C) and BP (Fig.
1.1.2D. E and F) following the placebo and the alcohol drinks are shown. There was no
significant change in the heart rate following the placebo as well as the 0 . 3 and 0.50 g
ETOHIkg B.W. drinks. Small changes in both systolic and diastolic pressure were
observed during the three hour experimental period. Statistical analysis (two way ANOVA)
indicated a significant effect of time on both systolic [F4.,,,=6.32: p < 0.001 1 and diastolic
[F,,,= 10.28: p < 0.01 1 BP. Thus. systolic BP was decreased following ingestion of the
placebo, 0.25g and OSOg ETOH/kg B.W. drinks. Post-hoc analysis using the Neuman-
Keul's multiple comparison test indicated a significantly lower systolic BP at 60 min
following the placebo drink and at 120 and 180 min following the 0.15 g ETOH drink

.+ 0.25 g ETOHIkg B.W.
+ 0.50 g ETOHIkg B.W.
Time of drink (min)
Figure 2.2.1. Changes in blood alcohol content folloning ingestion of 0.25 and 0.50 g ETOHIkg B.W.
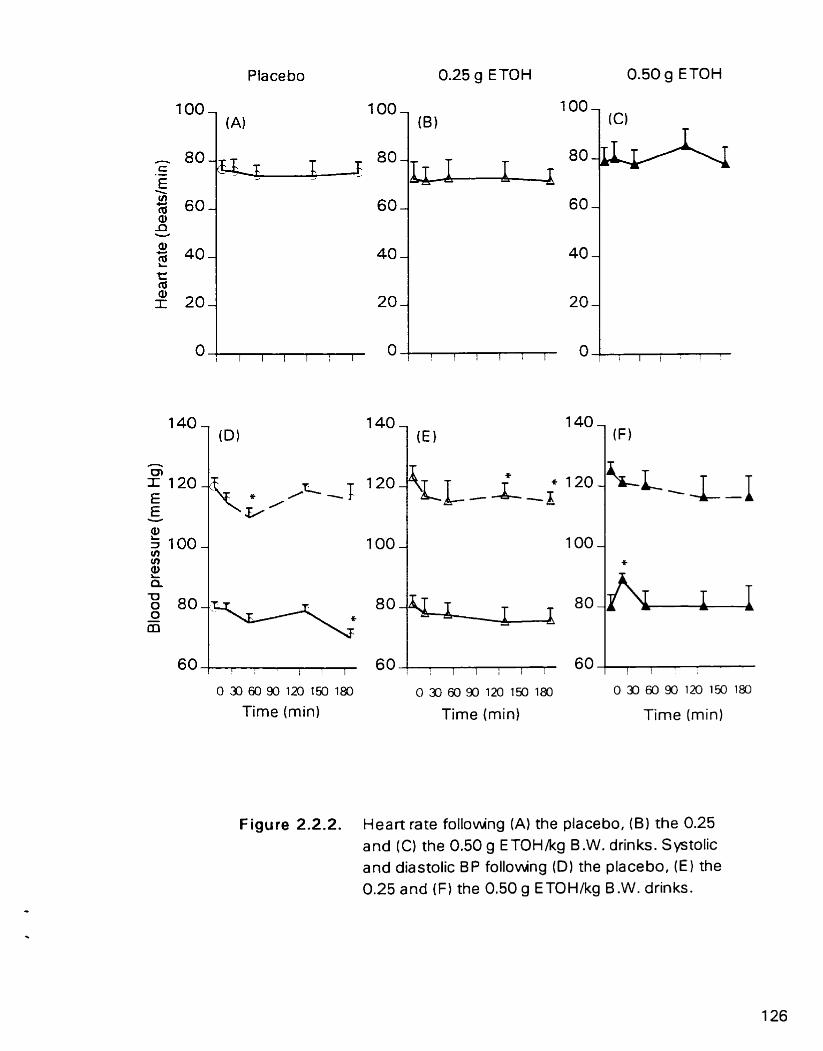
Pta ce bo 0.25 g E TOH 0.50 g ETOH
O a 60 90 120 150 180
Time (min)
I /
I I I 1 60 0 O 30 60 90 120 150 180 O 30 60 90 120 150 180
Time (min) Time (min)
Figure 2.2.2. Heart rate follodng (A) the placebo, (B) the 0.25 and (C) the 0.50 g ETOHkg B.W. drinks. Systolic and diastolic BP folloming (D) the placebo, (E) the 0.25 and (F) the 0.50 g ETOHIkg B.W. drinks.

cornparecf to the corresponding systolic BP at time O min. Following ingestion of the 0.50
g ETOH drink a gradual decrease in systolic BP was also observed which however did not
reach a level of significance at any of the time points that it was cested. The diastolic BP
also presented a small decrease with time following the placebo and the 0.15 g ETOH/kg
B.W. drinks. At 180 min following the placebo drink the diastolic BP was signitïcantly
lower than at time O min. Following the 0.15 g ETOH drink the gradual decrease in
diastolic BP did not reach a level of significance. Interestingly. at 15 min following the
0.5 g ETOH drink there was a short lasting. but statistically significant increase in the
diastolic BP.
The ef'fect of ETOH on the plasma ANP content is shown in Figure 2.7.3A.B and
C. Both 0.25 and 0.50 g ETOH per kg B.W. induced an increase in plasma ANP content.
As shown on Figure 2 . L 3 . . the plasma ANP content was elevated at 15 and 45 min but
had returned to basal levels by 120 min post-ETOH ingestion. Statistical analysis (two way
ANOVA for repeated measures) indicated a signiticant effect of time [F,.,, = 8: p < 0.00 1 1.
However. there was no significant effect of treatment and no significant interaction
between treatment and time factors. Post-hoc analysis using the Neuman-Keul's multiple
comparison test indicated that at 15 min following the 0.75 g alcohol drink and at 15 and
45 min following the 0.5 g alcohol drink the plasma ANP content was significantly higher
than the basal ANP levels ( levels at time O) (p <0.01]. This increase in the concentration
of plasma ANP with time was a specific effect of ETOH since no increase was observed
following the placebo drink. In fact. a small decrease with time was observed in the
plasma ANP content following the placebo drink, which however did not reach a level of
significance. Furthermore. it was also noticed that the high dose of ETOH (0.5 g
ETOHfkg B.W.) did not induce a higher plasma ANP concentration than the low dose of
ETOH (0.25 g ETOH/kg B.W.) even though blood alcohol content was about 2 times
higher. However. the plasma ANP levels remained elevated for a longer period follow ing
the high ETOH dose.
Figure 2.2.4A. B and C demonstrate the effect of ETOH ingestion on the plasma
AVP content. Statistical analysis indicated a significant effect of time [F,.,(, = 2.5:
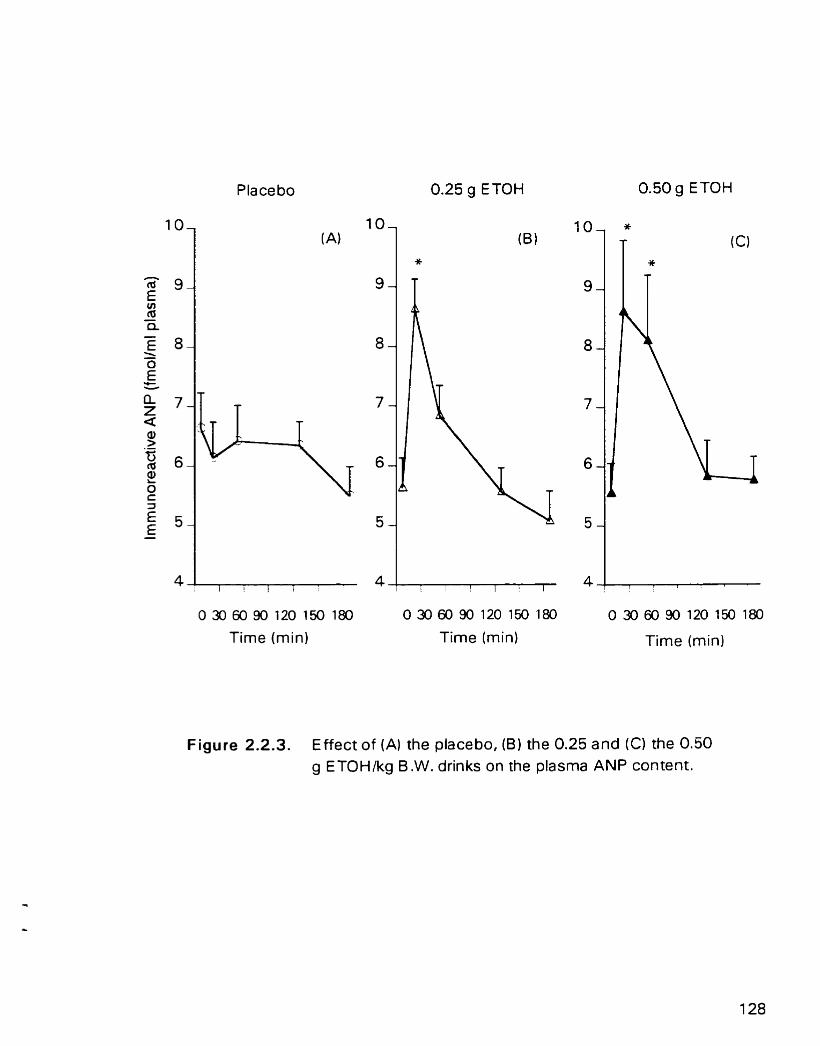
Placebo
r
3 30 60 90 120 1 5 0 180
Time (min)
l I ! I ! I
O 30 60 90 120 150 180
Time (min)
O 30 Hl 90 120 150 180
Time (min)
Figure 2.2.3. Effect of (A) the placebo, (B) the 0.25 and (C) the 0.50 g ETOHlkg B.W. drinks on the plasma ANP content.

Placebo
O 30 60 90 120 150 180
Time (min)
O 30 60 90 120 150 180 O 30 60 90 120 150 180
Time (min) Time (min)
Figure 2.2.4. Effect of (A) the placebo, (BI the 0.25 and (C) the 0.50 g ETOH/kg B.W. drinks on the plasma content of vasopressin.

p <0.05]. Post-hoc analysis using the Neuman-Keul's multiple comparison test indicated
that following either the placebo or the 0.25 g ETOH drinks there was no significant
change in the plasma AVP levels. while following the 0.5 g ETOH dose there was a
gradual decrease in the plasma AVP content. In fact the plasma AVP levels at 110 and 180
min post-drink were significantly different from those at O (prior to the drink) and at 15
min post-drink (p < 0.05. Neuman-Keuls multiple comparison test). As expected. this
decrease in plasma AVP content following the G.5 g ETOH dose was associated with a
higher urine excretion to 365 f 52 ml. compared to 230 + 17 ml and 265 + 38 ml
following the placebo drink and the 0.25 g ETOH dose respectively. Estimation of Na+
and K' contents in the urine collected during the 3 hr of the experimental period indicated
a signiticant increase of NaT excretion following the 0.25 g ETOH drink (p < 0.05) but
not following the 0.50 g ETOH drink . On the other hand. a small but significant
(p < 0.05) decrease in the excretion of K+ in the urine was observed following the 0.50 g
but not the 0.25 g ETOH drink (Table 2.2.1.).
Figure 3.1.5A. B and C indicate that ETOH ingestion did not induce an increase
above the basal levels in the plasma content of immunoreactive (ir)-cortisol . Statistical
analysis indicated a significant effect of time on the plasma cortisol content [F4.,,=?.81:
p < 0.051. A gradual decrease with time was observed as expected from the circadian
rhythm of circulating cortisol in humans. Post-hoc analysis using the Neuman-Keul's
multiple comparison test indicated that the plasma cortisol content at time O (prior to
administration of the drinks) was significantly higher than the plasma cortisol content at
60. 110 and 180 min following the placebo drink (p<0.05) and 110 and 180 minutes
following the 0.35 g ETOH drink (p < 0.05). while following the 0.50 g ETOH drink the
time dependent decrease in the plasma cortisol content did not reach a level of
significance. Thus. ingestion of the 0.5 g ETOH/kg B.W. drink did not induce an increase
in the plasma cortisol levels. However. it prevented the decrease in the plasma cortisol
levels that was observed with time following ingestion of either the placebo. or the 0.25
g ETOHkg B. W.
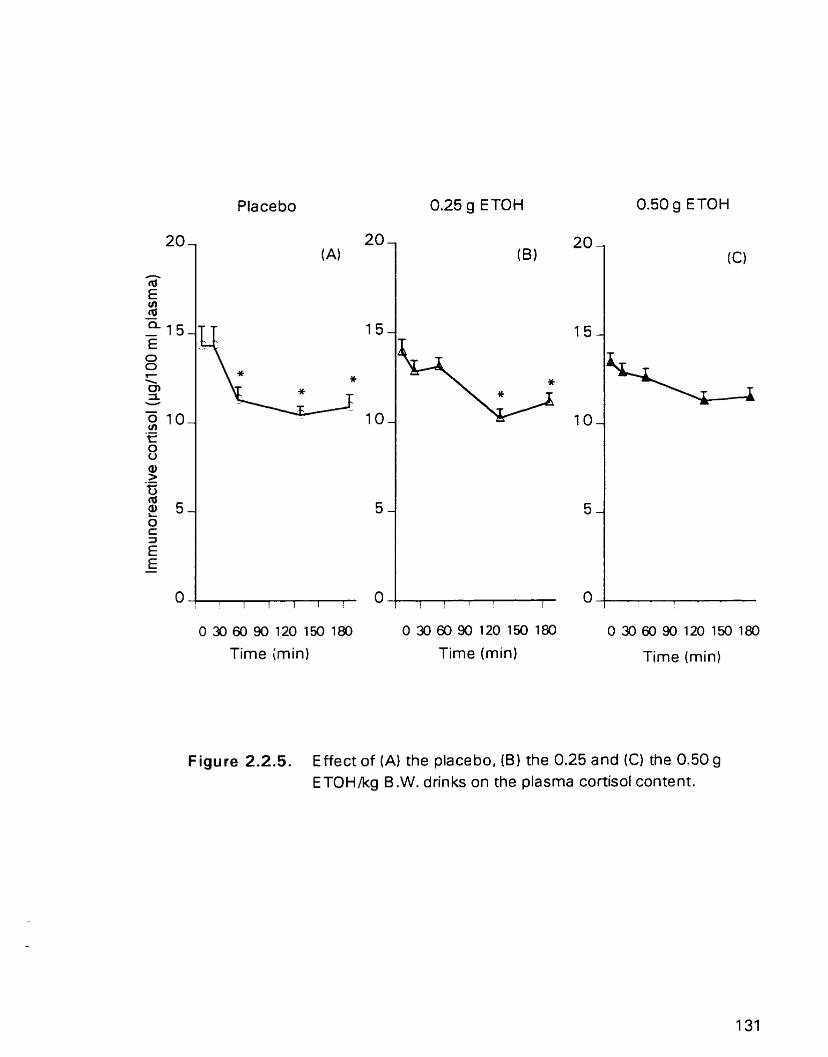
Placebo
O 30 60 90 120 150 180
Time (min)
O 30 60 90 120 150 180
Time (min) O 30 60 90 120 150 180
Time (min)
Figure 2.2.5. Effect of (A) the placebo, (B) the 0.25 and (C) the 0.50 g ETOHIkg B.W. drinks on the plasma cortisol content.

Table 2.2.1. Uritie output orid urzrze sodilirn and potassium excretiori heari * SEhl)
f o l h i ~ i g irrgestion of O. 0.25 und 0.50 g ETOWkg B. W.
Treatment Urine volume Na' (mmoVhr) K' (rnmoh)
[[ 0.50 2 ETOH 365 k 52* 4.4 1 + 1.36 2.10 * 0.61* 11 * Significant clifference fiom the placebo ( O g ETOH); p 50.05.
** Sigpficant Merence fkom the 0.50 g ETOH dose; p 50.05.
Neurnan-Keuls multiple cornparison test.

2.2.5. DISCUSSION
Both experimental (Howe et al.. 1989) and epidemiological (MacMahon. 1987)
smdies have shown that low to moderace ETOH consumption ( 1 to 2 drinks per day) delays
or prevents die age dependent increase in BP. The mechanism by which ETOH exerts this
antihypertensive effect is not clear .
The present studies indicated a small transient increase of diastolic but not systol ic
BP within 15 min following ingestion of the 0.50 g ETOH dose. A dissociation between
the effect of aicohol on the systolic and diastolic BP has been previously reponed (Criqui.
1986). It was proposed that ETOH's effect on systolic and diastolic BP may provide a clue
to the underl y ing pressor (or depressor) mechanism b y wh ic h alcohol modifies
cardiovascular homeostasis. Indeed. elevation in systolic BP may imply a greater effect of
ETOH on the cardiac output or tluid volume. whereas elevations in diastolic BP may
suggest moditkations in vascular resistance (Ireland et al.. 1984: Puddey et al.. 1985).
Thus. the initial and rapid increase in diastolic BP following the higher dose of alcohol
may represent sorne short-lived vasoconstriction produced by ETOH. Previous studies
investigating the acute eiiects of ETOH on BP have yielded conflicting results. Thus
following high ETOH intake (intoxicating doses) both a decrease (Conway. 1968) and no
change (Riff et al.. 1969) in BP have been reponed. Likewise. ingestion of ETOH at doses
below the intoxicating levels induced no change in BP (Gould et al., 1971). increase in BP
(Orlando et al.. 1976) or immediate increase followed by a decrease in BP (Ireland et al..
1984). In another study the contribution of (a) expectancy. (b) tluid volume and ( c )
intoxicating doses of ETOH on the BP changes following an ETOH drink were
investigated (Adesso et ai.. 1990). Surprisingly. results indicated that neither expectancy
nor tluid volume had a signitkant effect on BP. On the other hand. ETOH induced an
initial decrease in the BP followed by an increase toward the basal levels during the
detoxification period. This increase in BP during the detoxification period may be
attributed to ETOH withdrawal.
Estimation of the blood alcohol content following ETOH ingestion indicated that i t
remained well below the intoxicating levels. Furthermore, it was noticed that the dose of

0.5 g ETOH/kg B. W.. did not induce a larger increase in ANP content than the dose of
0.25 g ETOH/kg B.W.. indicating that large doses of ETOH may not be more effective
in stimulating the release of ANP than srnall doses of ETOH. However. following the 0.50
g dose of ETOH the plasma ANP content remained elevated for a longer period post-drink
than following the 0.25 g ETOH dose. suggesting a longer lasting effect of the higher
dose. A similar increase in plasma ANP content has been reported following
administration of 1 .O ml ETOH /kg B. W. in a total volume of 750 ml (Colantonio et al..
1991). Interestingly. data from this study demonstrated that the maximum increase in
plasma ANP content was obtained within the tïrst IO minutes post-ETOH when the blood
alcohol content was about 40 mg/dl. As blood ETOH content increased to intoxicating
values the plasma ANP content started to decrease. clearly indicating that maximum
response of the ANP system to ETOH is obtained with low concentrations of ETOH. in
agreement with the resulu of the present study . In addition. administration of the placebo
drink. consisting of two parts orange juice one part tonic water at a final volume equal to
that of the alcohol drinks did not alter the plasma ANP content. indicating that (a) the
volume of the fluid ingested was not high enough to induce sufficient increase in plasma
volume and suetching of the atria to stimulate the release of ANP and (b) the increase in
plasma ANP concentration following the ETOH drink was a specific effect of ETOH. and
was nor due to orange juice.
However. studies by other investigators have failed to observe an ETOH-induced
increase in plasma ANP content (Leppiiluoto et ai.. 1992: Hynynen et al.. 1992: Ekman
et ai., 1994). For example in a recent study when doses of 0. 0.50 and 1 g ETOH per kg
/ B.W. were administered in 500 ml of tluid at 18:OO there was no increase in plasma
ANP content. while a dose dependent inhibition of the nocturnal peak (occurring at
midnight) of plasma ANP was observed (Ekman et ai., 1994). In these studies the earliest
measurement of plasma ANP was at 2 hr after initiation of drinking. Since both the present
and previous (Colantonio et al.. l99 1 ) studies indicate that the increase in plasma ANP
content occun fast following drinking and is short lasting, returning to basal levels withi n
2-3 hrs post-drink. even if there was an initial increase in plasma ANP content. it could

not have been detected in the snidies by Ekman et al ( 1994). In another study ( Leppaluoto
er al.. 1992) a decrease in plasma ANP content was observed at 30 min and was
maintained for at least 2 hr, following ingestion of 1. 5 g ETOHfkg B. W. administered
in 500 ml tluid at 18:M) hr. On the other hand. there was no effect on plasma A N P content
following administration of 1 g ETOH /kg B.W. in volumes ranging from 415-560 ml at
18:00 hr to subjects who were slightly volume-loaded prior to ETOH administration
(Hynynen et al.. 1992). The differences in the results of those studies frorn those of the
present studies and of Colantonio et al (1991) rnay be attributed to a number of factors
such as the rime of day when the ETOH was administered. sarnpling time intervals post
ETOH ingestion and prior volume-loading of the subjects. For exarnple, volume-loading
may stimulate the heart ANP system prior to the administration of ETOH. so that a
subsequent stimulation by ETOH rnay not induce a further increase in ANP release. In
addition. rapid urine excretion usuall y associated wi th volume-loading and the
corresponding reduction in plasma volume. as was observed in the study by Leppaluoto
et al. (1992). may also counteract any stimulatory effect of ETOH on ANP release.
The present studies also indicated that the 0.50 g ETOH dose induced a gradua1
decrease in plasma AVP content and increased diuresis. This decrease in the plasma AVP
content and increased diuresis observed following the 0.50 g ETOH drink. can not be due
to die volume of the tluid ingested since it was not observed following either the placebo
or the lower dose of ETOH (0.25 g ETOH / kg B.W.) which were administered in the
saine volume of fluid. Indeed. the three drinks given to each subject were isovolumetric
but not isoosmotic. In fact. ETOH ingestion increases plasma osmolality. Interestingly.
it has been shown that increase in plasma osmolality stimulates the release of A N P and
increases plasma ANP content (Arjamaa and Vuolteenaho. 1985). Thus. one of the
mechanisms by which ETOH increases ANP release could be the increase in plasma
osmolality with increasing alcohol concentration in the blood. The absence of a decrease
in AVP secretion following the low dose of ETOH may be explained by the low
concentration of ETOH in circulation achieved with the 0.25 g dose. as was previously
reported (Ekman et al., i 994). Most of the diuretic effect associated with acute ETOH

consumption is attributed to the inhibition of AVP release from the posterior pituitary
(Eisenhofer and Johnson. 1982: Goldsmith and Dodge. 1985). Interestingl y, ANP has also
been reponed to suppress hypothalamic AVP (Januszewicz et al.. 1986: Colantonio et al..
1991: Ekman et al.. 1994). in the present studies. the increase in circulating ANP levels
by the 0.5 g ETOH drink is associated with a decrease in plasma AVP levels and increased
diuresis. It is conceivable therefore. that the inhibition of AVP release following ETOH
administration is mediated by the observed stimulatory effect of ETOH on ANP release.
The decrease in AVP and increase in ANP release would lead to increased diuresis as was
observed in the present (table 2.2.1 .) and other studies (Potter et al., 1986: Colantonio ef
al.. 1991: Hynynen et al.. 1991: Ekman et al.. 1994). Estimation of the content of
electrolytes (Na' and K') excreted in the urine during the three hour of experimental
period indicated a small increase in Na' excretion following ingestion of the 0.75 but not
of the 0.50 g ETOH / kg B.W. drink (table 2.2.1.). Other studies using ETOH doses
higher than 0.50 g ETOH also did not observe signitïcant changes in urine Na' and K-
concentrations (Potter et al.. 1986: Colantonio et al.. 199 1 : Hynynen et al.. 1992: Ekman
et al.. 1994). The use of specific ANP and AVP antagonists would help further detïne the
possible associations between these two hormonal systems in the ET0 H- i nduced di ures is . although a major problem for studies using ANP antagonists is the low potency and
specificity of the currently known ANP antagonists (Anand-Srivastava and Trachte. 1993).
Among the factors known to increase the release of A N P is stress (Horky er al..
1985). Therefore. it is possible that activation of the Hypothalamic-Pituitary- Adrenal
(HPA) mis. either by stress associated with the experimental procedure or by the ETOH
administration. could induce an increase in the ANP release. However. it seems that this
was not the case in the present studies since estimation of plasma cortisol content indicated
no increase in plasma cortisol levels following either the placebo or ETOH drinks and thus
no activation of the HPA-axis. confirrning the absence of stress associated with the
experimental procedure. Interestingl y. a number of studies have demonstrated activation
of the HPA axis associated with an increase in plasma cortisol levels following
administration of moderate to high doses of ETOH (Mendelson and Stein. 1966:

Mendelson et al.. 197 1 : Valimaki et al., 1984: Schuckit et al.. 1987. 1988) but not of
low doses of ETOH (Schuckit et al.. 1987. 1988) to humans. Since in the present studies
the doses of ETOH used are considered low (equivalent to one or two standard drinks).
the absence of a stimulating effect of ETOH on the HPA axis is in agreement with the
pub 1 is hed reports.
A number of mechanisms have been proposed to explain the antihypertensive effect
of low to rnoderate ETOH consumption. These include ETOH-induced increase of the
HDL content in circulation (Criqui. 1986) and the inhibitory effect of ETOH on the
activity of substances known to have a pressor effect. For example. low but not high
concentrations of ETOH induce an inhibitory effect on the vasoconstrictor activity of
norepinephrine (Kalsmer. 1970: Altura et al.. 1983: Criscione et al.. 1989). ETOH
consumption has also been shown to increase the secret ion of nitric oxide (NO) in several
vascular beds. such as in the pulmonary arteries (Greenberg et al.. 1993). NO is a
vasodilator produced by the endothelial cells.
The present studies indicated that an additional mechanisrn may be implicated in the
antihypertensive effect of low ETOH consumption. involving the A N P systern. ANP. a
hormone released from the hem atria. decreases BP by inducing vasorelaxation. increas i ng
diuresis. and decreasing the release and/or activity of a number of pressor hormones
(Chartier et al.. 1984: Yasujima et al.. 1985. 1986: lnagami et al.. 1987). lntravenous
administration of excess ANP antibodies induced a decrease in diuresis and natriuresis. an
increase in plasma renin but had no effect on plasma aldosterone and BP (Chartier et al..
1984). Furthermore. infusion of low doses of A N P suppressed the hyperrensive effect of
norepinephrine. epinephrine and angiotensin I I (Yasujima et al.. 1986: Inagami et al..
1987). Thus. it seems that ANP rnay act as a buffering system that protects the
cardiovascular system against a sudden increase of BP induced by sudden activation of the
noradrenergic or the renin-angiotensin systems (Inagami et al.. 1987). Therefore. it is
reasonable to propose that agents stimulating the release of ANP may provide a protection
against the activation of systems known to exert a hypertensive effect.

It is possible then that the protective effect of low and moderate doses of ETOH on
the age-dependent increase in BP. is partially due to its ability to activate the hem ANP
system. In addition. ETOH rnay alter the activity of other renal and /or cardiovascular
systems. such as AVP. and thus modi@ diuresis and BP. Supporting the stimulatory effect
of acute ETOH exposure on the hem ANP system are animal studies demonstrating a dose
dependent increase in plasma ANP content (section 2.1 .). This increase in plasma ANP
content is fast. reaches maximum levels within 15 min following an i.p. injection of
ETOH (as observed in the present studies) and is associated with a decrease in the ANP
content in the heart atria.
Therefore. it rnay be hypothesized that frequent activation of the heart A N P system
by low daily alcohol ingestion ( 1 to 2 drinks per day) will stimulate the release of ANP
and eventually rnay increase the rate of ANP synthesis and the content of specific ANP
mRNA. Conrrary to the beneficial effect of chronic low to moderate ETOH consumption
on BP. following chronic heavy use of ETOH the increased activity of the heart ANP rnay
not be sufficient to counteract the hypertensive effects of ETOH. such as increased
sympathetic activity , increased sensitivity of blood vessels to pressor substances and
intermittent ETOH withdrawal reactions and eventually hypertension will develop. in this
case. it rnay be expected that the ETOH-induced hypertension will be associated with high
circulating ANP levels as has been reported for other conditions of hypertension (Imada
er al.. 1985: Nozuri et al., 1986). Therefore. ETOH administration rnay induce both
h ypertensive and antihypertensive effects. At low ETOH concentrations. the net impact
of the antihypertensive effects rnay be dominant. while at high ETOH concentrations rhe
net impact of hypertensive effects rnay predom inate.
In summary. the present studies indicated that ingestion of low amounts of ETOH.
equivalent to I or 3 standard drinks. but not of a placebo drink increased the plasma ANP
content in norrnotensive subjects. This increase in the release of ANP by low doses of
ETOH rnay be partially responsible for the antihypertensive effects of low to moderate
daily ETOH consumption reported in a number of epidemioiogical (human) and
experimental (animal) studies.

2.2.6. ACKNOWLEDGEMENTS
The authors wish to thank the nursing staff of the research unit at Douglas Hospital
Research Centre. This study was supported by gram MT- 10337 and MT- 1 1674 from the
MRCC and grants from the Kidney Foundation and the Canadian Heart and Stroke
Foundation to Gutkowska J. and by the Alcohol Research Program at Douglas Hospital.
Guillaume P. is a recipient of a scholarship from the "Fond pour la Formation de
Chercheurs et l'Aide a la Recherche (FCAR) du Québec"

CBAPTER 3
CHRONIC ETOH STUDIES

Section 3.1.
EFFECT OF CHRONIC ETHANOL CONSUMPTION ON THE ATRIAL
NATRIURETIC SYSTEM OF SPONTANEOUSLY HYPERTENSIVE
RATS
P. Guiilaume, M. Jankowski, C. Gianoulakis and J. Gutkowska
Alcohoüsm: C h c d and Experimentai Research
Wcohol Clin Eirp Res 20: 1653- 166 1, 1996)
Contribution bv CO-authors: Dr. M. Jankowski introduced me to the technical aspects of
Northem blottmg and was responsible for the RT-PCR midies. Dr. C. Gianoulakis and Dr.
J. Gutkowska were my CO-supervisors.
Achowled~ements: - R Claudio was the animal technician. C. Coderre. N. Charron and S.
Laroque performed some of the ANP or AVP iodinations.

3.1. EFFECT OF CHRONIC ETHANOL CONSUMPTION ON THE
ATRIAL NATRIURETIC SYSTEM OF SPONTANEOUSLY
HYPERTENSIVE RATS ( S H R )
3.1.1. ABSTRACT
There is a lot of discussion on the effects of ethanol (ETOH) on the blood pressure
(BP). It has been suggested that chronic moderate ETOH consumption prevents the
development of agedependent hypertension in humans and spontaneously hypertensive rats
(SHR). However. the mechanism mediating this effect is unknown. In the present studies.
we hypothesized the implication of Atrial Natriuretic Peptide (ANP). a BP-lowering
hormone. on the antihypertensive effect of moderate ETOH consumption. A 10% viv
solution of alcohol was given as drinking iluid to SHR and normotensive Wistar-Kyoto
(WKY) rats for up to 32 weeks. This treatment prevented. at least in part. the age-
dependent increase of BP in SHR and WKY rats. The lower BP was associated with
s igni ficantl y lower levels of circulating atrial natriuretic peptide ( ANP) in both groups.
After chronic ETOH administration. total ANP content and concentration were higher in
the left and right atria of SHR and WKY rats than in water-treated controls. Despite the
ETOH- induced increase in atrial A N P content. there was no signitïcant change in atrial
ANP mRNA . suggesting decreased atrial release. Chronic ETOH treatment significantl y
reduced ANP mRNA in the ventricles of S H R but not of WKY rats. Correspondingly.
ventricular ANP content and concentration were lowered by ETOH in SHR only. Chronic
ETOH administration induced a signifiant increase of plasma arginine vasopress in ( AVP)
and a signitkant decrease of plasma aldosterone in S H R but not in WKY rats.
Thus. chronic ETOH treatment prevented the age-dependent elevation of BP in both
SHR and WKY rats. and altered the activity of heart ANP as well as of the aldosterone
and AVP systems.

3.1.2. INTRODUCTION
A positive correlation between chronic high ethanol (ETOH) consumption and
hypertension has long been recognized in humans (Lian. 1915). However. it appears that
aithough this relationship is linear for 3 or more standard drinks per day ( 1 standard drink
corresponds to 8-10 grams of ETOH). a different kind of association is seen with low or
moderate ETOH consumption ( MacMahon. 1987). Thus. a number of epidemiological
studies have reponed a U- or J-shaped curve for blood pressure (BP) with increased ETOH
drinking. suggesting that individuals consuming 1-3 drinks per day had lower or similar
BP as abstinent individuals and significantly lower BP than heavy (5 or more drinkdday )
drinkers (Gleiberman and Harburg. 1986: MacMahon. 1987). Interestingly. a similar
effect has been seen in animal studies where chronic moderate ETOH administration
prevented or delayed the development of age-dependent hypertension. particularly in rat
populations known to become hypertensive (SHR and SHRSP) (Sanderson et al.. 1983:
Jones et ai., 1988: Howe et al., f 989: Beil in er ai., I992).
The mechanisms by which chronic moderace ETOH administration prevents the
development of hypertension are still not known. One such mechanism could involve the
atrial natriuretic peptide (ANP).
Atrial natriuretic peptide (ANP). a 18-amino acid peptide synthesized mostly in heart
atria (De Bold et al.. 198 1 ) . is known to lower BP through several mechanisrns. including
natr iuresis. diuresis. vasodilation. and by decreasing the secretion of various pressor
hormones. such as renin-angiotensin I I . aldosterone. norepinephrine and arginine
vasopressin (AVP) (Needleman er al.. 1989: Blaine. 1990: Nakao et al.. 1993). Earlier
(Section 2.1.). we demonstrated that acute injection of moderate ETOH doses in rats
induced a rapid increase in plasma ANP levels. and this elevation was associated with a
reduction of right and left atrial ANP content. In contras. the acute ingestion of a large
dose of ETOH in Sprague-Dawley rats has been associated with no increase in circulating
ANP levels (Wigle er al.. 1993a). Recently. a short-term chronic ETOH study (6 weeks)
using Sprague-Dawley rats was also published by Wigle et al. ( 1993b. 1993c) who noted
that. although there were no changes in plasma ANP levels. plasma brain natriuretic

peptide (BNP) and ventricular ANP contents were significantly elevated. After 6 weeks
of ETOH administration however. BP values in the ETOH-treated and control animals
were not significantly diffèrent (Howe et al.. 1989). Therefore. it would be interesting to
examine the effect of chronic treatment with moderate amounts of ETOH for a duration
sufficient to produce a significant difference in BP between both groups.
Using Wistar-Kyoto (WKY) rats and SHR. the present studies investigated the effect
of chronic moderate ETOH consumption for 31 weeks (a duration suficient to demonstrate
prevention of the age-dependent increase in BP) (Howe et al.. 1989) on circulating ANP
levels, as well as on ANP content and ANP mRNA in h a r t atria and ventricles. BP. heart
rate (HR). body weight (BW) and fluid consumption were evaluated on a regular bais.
In addition to ANP. the plasma leveis of AVP. corticosterone and aidosterone were also
estimated to provide an indication on the implication of other hormonal systems in the
ancihypertensive effect of chronic moderate ETOH consumption and to contribute to a better
understanding of the interactions between the various endocrine systerns.
3.1.3. MATERIALS AND METHODS
3 . 1 . 3 . 1 . T r e e
Male WKY rats and SHR. approximately 6 weeks old. were purchased from Charles
River Breeding Farms (St-Constant. Québec. Canada). Upon reception. the animals were
randomly assigned to 1 of 4 groups (Group 1 : WKY-H,O ( n = 15): Group 2: WKY-ETOH
( n = 15): Group 3: SHR-H,O ( n = 15) and Group 4: SHR-ETOH ( n = 15)). After a 7-day
acclimatization period. ETOH was gradualiy added to the drinking water of Groups 2 and
4. up io a 30% v/v solution in 15 days (5 % v/v for 5 days. 10% v/v for 5 days. 15 % v/v
for 5 days and 20% vlv subsequently. as described by Howe et al.. 1989). AI1 animals had
free access to pelleted rat chow (Purina. Richmond. VA) and tluids (water or ETOH
solution) for the duration of 8 months.
BP and HR were rneasured every 1 weeks and BW every week. Liquid consumprion
was recorded daily. BP and HU were evaluated by the tail-cuff method (Pfeffer er al..
1%' 1). After 8 months, a11 rats were killed by decapitation. Trunk blood was collected.

and atria and ventricles were removed carefully for subsequent estimation of A N P and
ANP mRNA content. To investigate the effect of age. a third group of animals ( 1 1 WKY.
II SHR) was sacrificed prior to the initiation of ETOH treatment. at about 7 weeks of age.
3.1.3.2, Estimation of blood ETOH
From fresh blood. 50-pl samples were deproteinized with 450 pl of ice-cold
trichloroacetic acid (6.35 w/v) for the measurement of blood ETOH levels at the tirne of
death. using the dehydrogenase enzymatic method (Hawkins et ai.. 1966).
. . 3.1-3.3. Prepgration of blood and ti.ssue e x t w for R a d i o i m m u n ~ v s (RIAS)
Trunk blood was collected in chilled 15 ml conical centrifuge tubes containing
protease inhibitors to the following final concentrations: 1 mg/rnl ethylenediamine-tetra-
acetate (EDTA). 10 mM phenylmethylsulfonyl fluoride (PMSF. P7626. Sigma Chemical
Co.. St-Louis. MO) and 5 m M pepstatin A (P4265. Sigma) . The tubes were imrnediately
centrifuged at 4°C and the plasma stored at -75°C for subsequent RIAS.
The ventricles as well as the left and right heart atria were excised and placed in
separate tubes containing ice-cold 0.1 N HCI. They were rapidl y boiled for 5 minutes and
then cooled on ice. The tissues were homogenized subsequently in a Polytron. and an
aliquot was taken for the estimation of protein content using Bradford's method (Bradford.
1976). The remaining homogenates were centrifuged for 15 minutes at 4°C and the
supernatants stored frozen at -75°C for the measurernent of ANP content.
3.1.3.4. RIAs
Atrial (final dilution. 1: 15 000). ventricular (final dilution. 1: 1 000) and plasma ( 100
p l ) ANP content was quantified by a direct second-antibody RIA (Gutkowska et al.. 1984:
Gutkowska et al., l987a). The val idity of the direct assay for the investigation of the
relative differences between various treatments has been previously demonstrated (Bidmon
et ai., 1991 and Section 2.1 .). AVP levels were estimated in acetone-extracted plasma
samples. also using a second-antibody RIA (Skowsky et al.. 1974). Corticosterone (Krey

et al.. 1975) and aldosterone (Mayes et al.. 1970) levels were measured by sensitive RI As
in ETOH-extracted plasma samples.
* * . 3.1.3.5. RNA extraction and hvbnd-
For estimation of ANP mRNA. the atria and ventricles tiom 5 animals per group
were careful [y dissected. frozen irnmed iatel y in isopentane. and stored singl y at -75 OC
until total R N A extraction.
RNA from cardiac tissues was isolated according to the acid guanidinium-thiocyanate-
phenol-chloroforrn methoci. as described elsewhere (Chomczynski and Sacchi. 1987). The
RNA concentration in each sample was determined by UV absorbance at 260 nm. Ten
pg of ventricular or two pg of atrial RNA were subjected to electrophoresis through 1.5 %
agarose gels containing 0.22 M formaldehyde and transfered ont0 nylon membranes
(Hybond N + . Amersham International PLC. UK) by capillary blotting. Immobilized RNA
samples were hybridized with randoml y primed a3'P-cDNA probes corresponding to ANP
and a-tubulin mRNA sequences. A random priming kit (Gibco. BRL. Bethesda. MD) and
a3'P-dCTP (3000 Ci/mmol. Amersham. Ar1 ington Heights. 1 L) were used for label1 ing
the probes. The Pst 1-digested 660 bp fragment from plasmid clone PN- 1-1 lserved as the
ANP probe (gift from Dr. Mona Nemer. Institut de Recherche Clinique de Montréal). A
550-bp fragment of a-tubulin cDNA probe. generated by reverse transcription (RT) of rat
atrial RNA and amplified by polymerase chain reaction (PCR). was used to monitor the
amount of RNA in each sarnple (Dagnino et al.. 1992). The blots were prehybridized in
buffer containing 120 mM Tris. 8 mM EDTA. 0.6 M NaCI. 0.01% sodium
pyrophosphate. 0.02% SDS, and 0.01 % heparin (H7005. Sigma). pH 7.4. Hybridization
was carried out in the same buffer for 16 hours with heparin content increased to 0.03 %
and supplemented with 10% of dextran sulfate as well as 10" dpm/ml of the labelled probe.
After hybridization. the membranes were washed in 2 X SSC/ 1 % SDS ( 10 minutes at
room temperature) and exposed on a Phosphorimager. Radioactive bands were measured
with Image-Quant software (Molecular Dynamics, Sunnyvale. CA). The tissue
concentrations of ANP mRNA reported in the text were normalized to the a-tubulin

rnRNA concentration in each sample to correct for differences in the RNA content applied.
3.1.3.6- RT-PCR
a) First strand cDNA qwthesis and quant~fication reaction product
Total RNA sarnples were treated with RQI DNAse (Promega. Fisher Scientific.
Canada) to eliminate any contamination with genomic DNA. Then. total RNA (2 pg) was
annealed with 0.5 pg of random hexamer and reverse transcribed in 50 mM Tris-HCI
buffer (pH 8.3). 7 rnM KCI. 3 rnM MgCl,. 10 mM DTT. 0.5 mM dNTP. 40 units
RNAsin (Pharmacia) and 200 units Moloney muiine leukemia virus reverse transcriptase
(BRL Laboratories, Gaithersburg. MD. USA). The reaction was left to incubate for 1 hour
at 37°C. A 3-pl aliquot of the RT reaction product was denaturated at 95 O C for 5 minutes
and then electrophoresed on 1 % agarose gel. After gel dry ing on autoradiographic films.
counts (over 400 bp in length) were integrated for each sample using Image-Quant
software. Input of cDNA sarnples in each PCR was normalized according to Irnage-Quant
rneasurements of the radioactive RT reaction product (Kolls et al.. 1993).
b) PCR ampiifkarion of ANP transcript
PCRs were conducted according to previously described procedures ( Dagnino er al..
1991: Dagnino et al.. 1992: Jankowski et al.. 1996). The number of cycles for the
exponential amplification of PCRs and the concentration of genomic DNA were
standarized in separate experirnents (Jankowski et al.. 1996). The following prirners were
used for PCR amplification: ANP forward. 5'-CAGCATGGGCTCCTTCTCCA-3'. and
ANP reverse. 5'-GTCAATCCTACCCCCGAAGCAGCT-3'. cDNA (and DNAse-treated
RNA samples used as negative control) was subjected to PCR amplification of 25 cycles
for 1 minute, at 94°C. 1 minute at 6 1 "C and 3 minutes at 73 O C . After separation on 1.5 %
agarose gels. products arising from arnplitication of cDNA (lower band) or genomic DNA
(upper band) yielded 430 bp or 541 bp fragments respectively. The bands from each
sarnple were measured by 1 mage-Quant software. and the cDNA am pl ificat ion products
were normalized to the arnount of genomic fragments (100%). as described previously
(Jankowski et al., 1996).

. . 3.1.3.7. -cal Audysis
The data are presented as Mean f SEM. The significance of differences among the
various groups was evaluated by two-way analysis of variance (ANOVA). with time as
the first independent variable and treatment as the second independent variable. fol lowed
by the Neuman-Keuls multiple comparison test. A P value of L 0.05 was considered to be
signit-cant.
3.1.4. RESULTS
3.1.4.1. Effect of ethanol on the hlpod m u r e and heart rate
Variations in the BP of SHR and WKY rats with age and treatment are shown in Fig.
3.1.1. Two-way ANOVA with time as the first independent variable and treatment as the
second independent variable indicated a significant effect of rime (F,.,, = D . S 6 . Pr0.0 1 )
and treatment (F,.,,=336.66. P~0.01) with a significant interaction of time and treatrnent
(F,,,,, =37.63. Pi0.0 1) in SHR. Similarly. a significant effect of time (F,.,, = 18.23.
P~0 .0 1 ) and treatment (F,.,,, =?O9.X. Pr0.01) with time-treatment interaction
(F, . - ,,,=23.36. P50.01) was noted in WKY rats. A rapid BP increase in both water- and
ETOH-ueated SHR was observed during the First half of the experimenr ( = 17 weeks) but
subsequently levelled off at around 200 mm Hg ( l? to 31" week). Meanwhile. WKY rats
experienced a slow and gradua1 elevation of BP values with age. The effect of ETOH
treatment in both SHR and WKY was seen as a slower rate of increase in BP compared
to corresponding water-treated controls. Thus. the BP rise in rats drinking ETOH was
s igni ficantly lower than in their H,O-treated control counterparts. In fact. a highl y
significant difference in BP between SHR-ETOH and SHR-H20 was observed from the
32"' week onward (P~O.ûû1). while in WKY rats. this significant difference was apparent
only from the 25" week of treatment (P50.05). After 32 weeks. the BP average was 189
+ 4 and 162 + 3 mm Hg ( n = 15) (p10.001) in SHR-H,O and SHR-ETOH (Groups 3 and
4) respectively. and 131 + 2 and 112 + 2 mm Hg ( n = 15) ( ~ ~ 0 . 0 5 ) in WKY-H,O and
WKY-ETOH rats (Groups 1 and 2) respectively.
A similar association was demonstratecl for HR (Fig. 3.1 2.). Again. H R values were
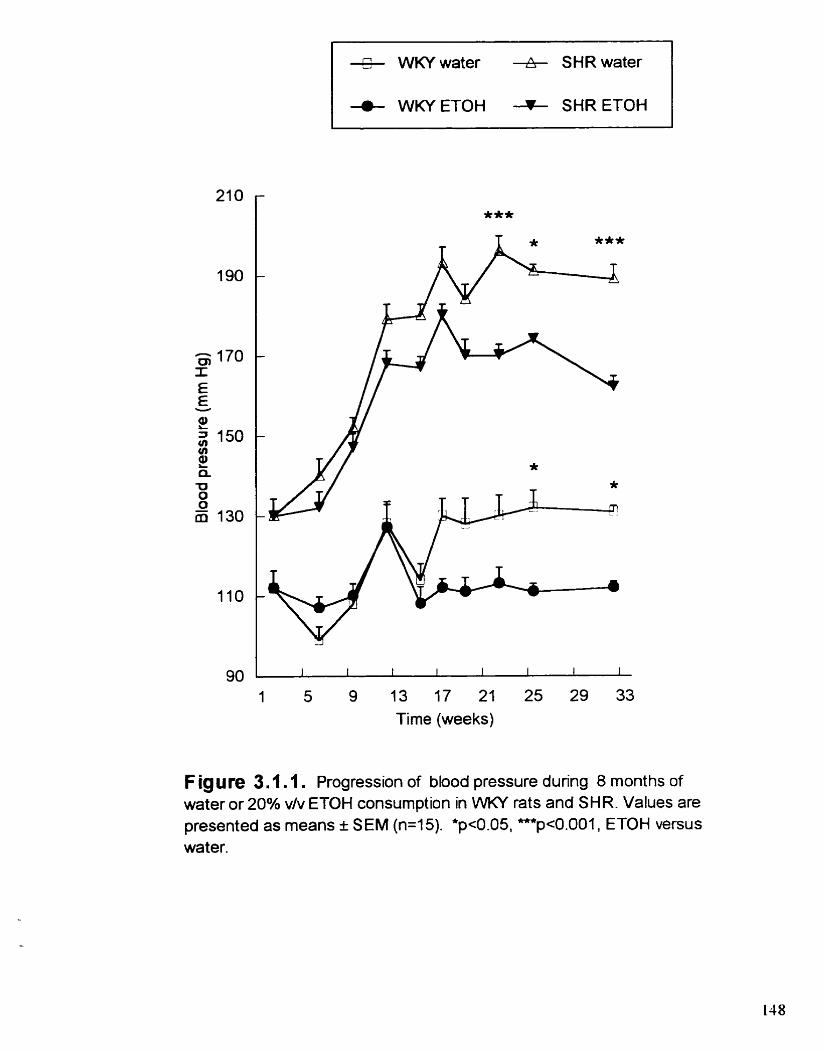
+ WKY water + SHR water
+ WKY ETOH + SHR ETOH
I 5 9 13 17 21 25 29 33 Time (weeks)
F Ïg ure 3.1.1. Progression of blood pressure during 8 months of water or 20% v/v ETOH consumption in WKY rats and SHR. Values are presented as means I SEM (n=15). *p<0.05, "pc0.001, ETOH versus water.

- - WKY water . SHR water
+ WKY ETOH + SHR ETOH
1 5 9 13 17 21 25 29 33 Time (weeks)
F igure 3.1.2. Progression of heart rate during 8 months of \Rater or 20% v h ETOH consumption in WKY rats and SHR. Values are presented as means *SEM (n =IO). 4.05, ETOH versus -ter.

significantly higher in SHR than in WKY rats. In both strains. HR declined gradually
during the fint 15 weeks of treatment. followed by a slow increase to initial values by the
3 P week. The effect of ETOH in both SHR and WKY was again a slower HR increase
so that by the end of this study there was a significant difference in HR between ETOH-
and H20-treated SHR. Thus. by the end of the experimental period. HR values were 441
+ 7 and 404 + 9 bears/min (n= 10) (ps0.05) in SHR-H,O and SHR-ETOH (Groups 3 and
4) respectively. and 379 + 8 and 363 + 7 beats/min ( n = 10) in WKY-H,O and WKY-
ETOH rats (Groups 1 and 2) respectively.
No signitïcant BW difference w u observed between ETOH- and water-treated
animals of both strains. Similarly. no siginificant variation between the liquid consumption
of ETOH- and water-treated SHR and WKY rats was noted. The average volume of tluids
ingested was around 50 ml/day. ETOH-treated animals consumed around 7.5 g of
ETOH/day .
3.1.4.2. Effect of ethanol on the heart ANP svstem
Plasma ANP levels were significantly lower in ETOH-treated rats than in their age-
matched controls (Table 3.1.1 .). Interestingly. there was no significant difference in
plasma A N P values between ETOH-treated adults and young 7-weeb-old animals
(p =0.795 and p =0.70 1 for WKY and SHR rats. respectively). Again. as observed for BP.
the variations in circulating ANP levels between the H,O- and ETOH-treated groups were
greater in SHR than in WKY rats. SHR-ETOH rats presented a 50 % decrease compared
to their water-treated counterparts wh ile WKY-ETOH showed a 34 % reduction (Table
3.1.1.).
The next 3 figures show corresponding tissue A N P variations in the right (Fig.
3.1.3.) and left (Fig. 3.1.4.) atria and ventricles (Fig. 3.1 S.) in terms of total ANP
content. concentration (pg/mg proteins) and m RNA levels. Au toradiograms of
representative Northern blots for atrial and ventricular ANP mRNA are shown on Fig.
3.1 A. Both ETOH- and water-treated SHR and WKY rats presented d a r levels of total
protein content in the atna (Table 3.1.2.). Since in most cases the changes in total protein
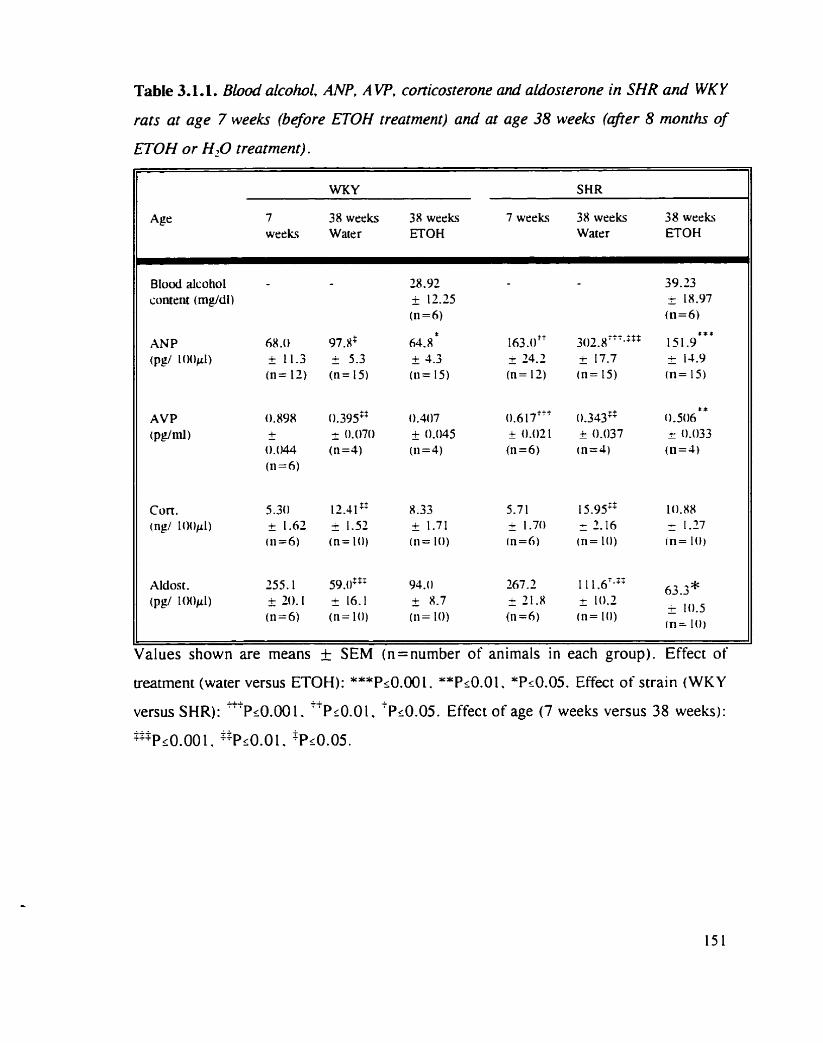
Table 3.1.1. Blood alcohol, ANP, A VP, corticosterone and aldosterone in SHR and WK Y
rats ut age 7 weeks (before ET0H treatment) and at age 38 weeks (Mer 8 months of
ETOH or H20 treatment).
WKY SHR
4.F 7 38 weeks 38 weeks 7 week. 38 weeks 38 weeks weeks Water ETOH Water ETOH
Blod alcohol content (me/dI)
12.41" 8.33 5 1.52 5 1.71 ( n = 10) (II= IO)
59.0'$' 94.0 + 16.1 & 8.7 (n = I O ) (n= IO)
307 8'''.3$ -. i: 17.7 (n= 15)
0.343'$ 4 0.037 in=4)
1 5.95" - - 2.16 In= 10)
1 1 1.6'."
I 10.2 cri= IO)
39-23 t - 18.97 (n=6)
151.9"' 5 14.9 tn= 15)
0.506'* * 0.033 (n=4)
10.88 - - 1-27 cn= I O )
633* 2 10.5 (n= I O )
Values shown are + SEM (n=number in each group). Effect of
treatment (water versus ETOH): ***P~0.00 1 . **P.0.0 1. *P<0.05. Effect of strain (WKY t-L= . . --
versus SHR): ' ''Pr0.001. ' 'PrO.01. 'P~0.05. Effect of age (7 weeks versus 38 weeks): . . . --- . . -- - "'P<O.OO 1. "P<O.O 1. 'P~0.05.
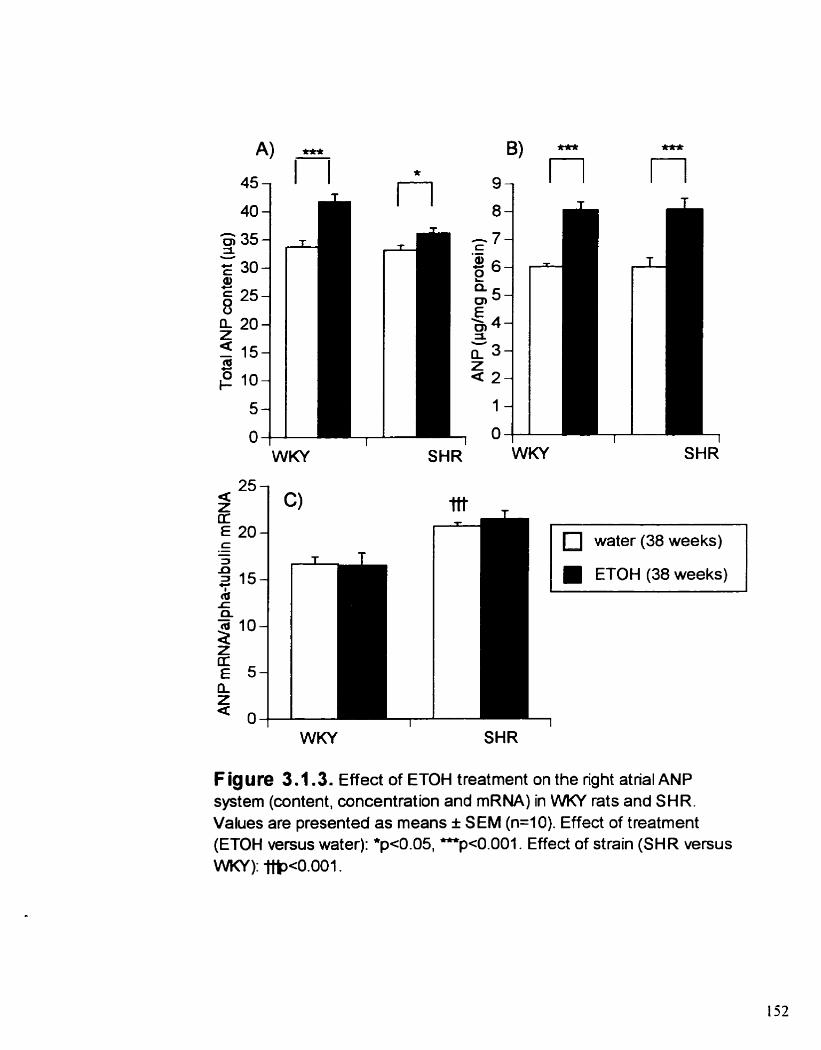
SHR WKY
water (38 weeks)
ETOH (38 weeks)
SHR
F 3.1.3. Effect of ETOH treatment on the right atrial ANP system (content, concentration and mRNA) in W rats and SHR. Values are presented as means I SEM (n=10). Effect of treatment (ETOH venus water): *p<0.05, "p<0.001. Effect of strain (SHR versus W): ttp<O.OOl.
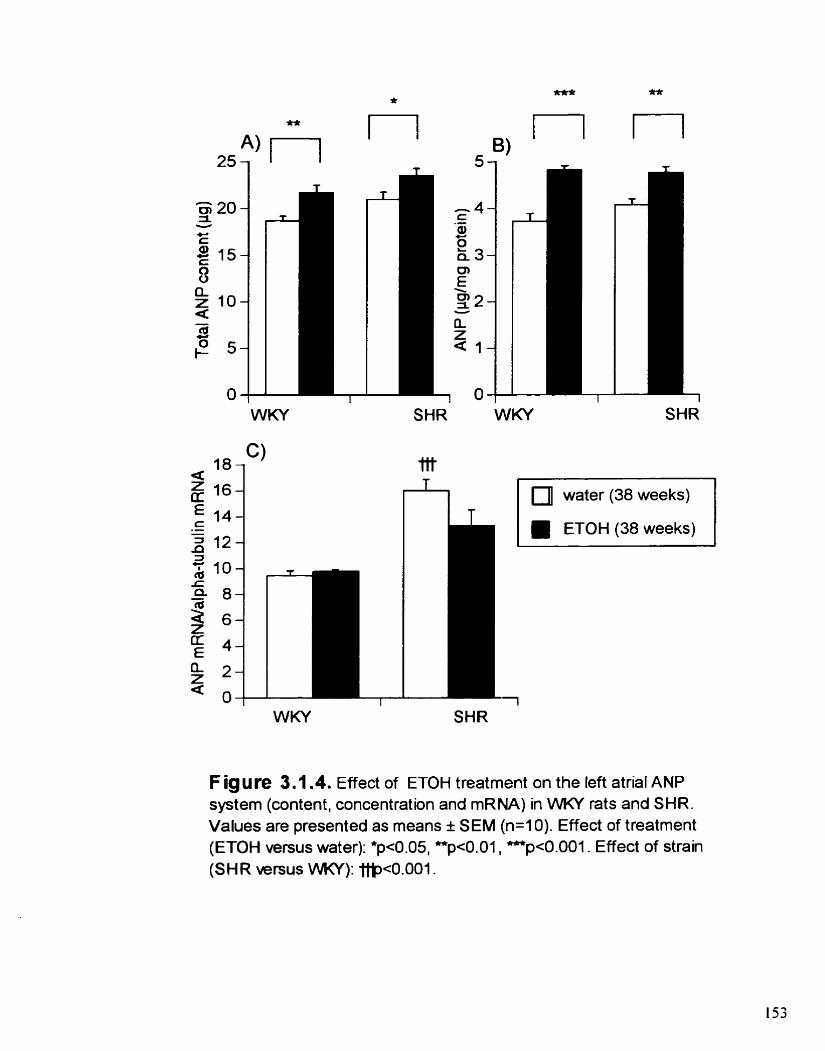
SHR WKY SHR
1 water (38 weeks)
SHR
F ig Urê 3.1.4. Effect of ETOH treatment on the left atrial ANP system (content, concentration and mRNA) in WKY rats and SHR. Values are presented as means i SEM (n=10). Effect of treatment (ETOH verçus water): Pc0.05, "pc0.01, -p<0.001. Effect of strain (SHR versus W): m<0.001.
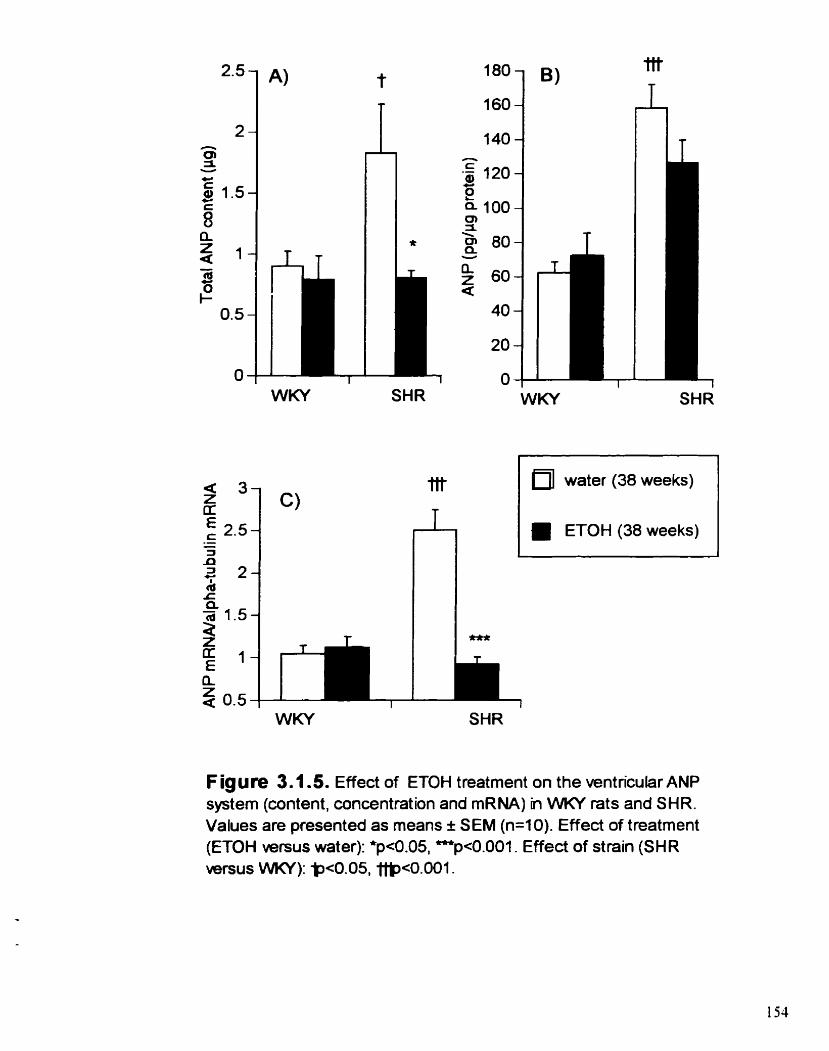
SHR I I
WKY SHR
SHR
water (38 weeks)
ETOH (38 weeks)
Figure 3.1.5. Effect of ETOH treatment on the ventricular ANP system (content, concentration and mRNA) in WKY rats and SHR. Values are presented as means I SEM (n=10). Effect of treatment (ETOH venus water): *p<0.05, "pc0.001. Effect of strain (SHR venus W): *<0.05, m<0.001.
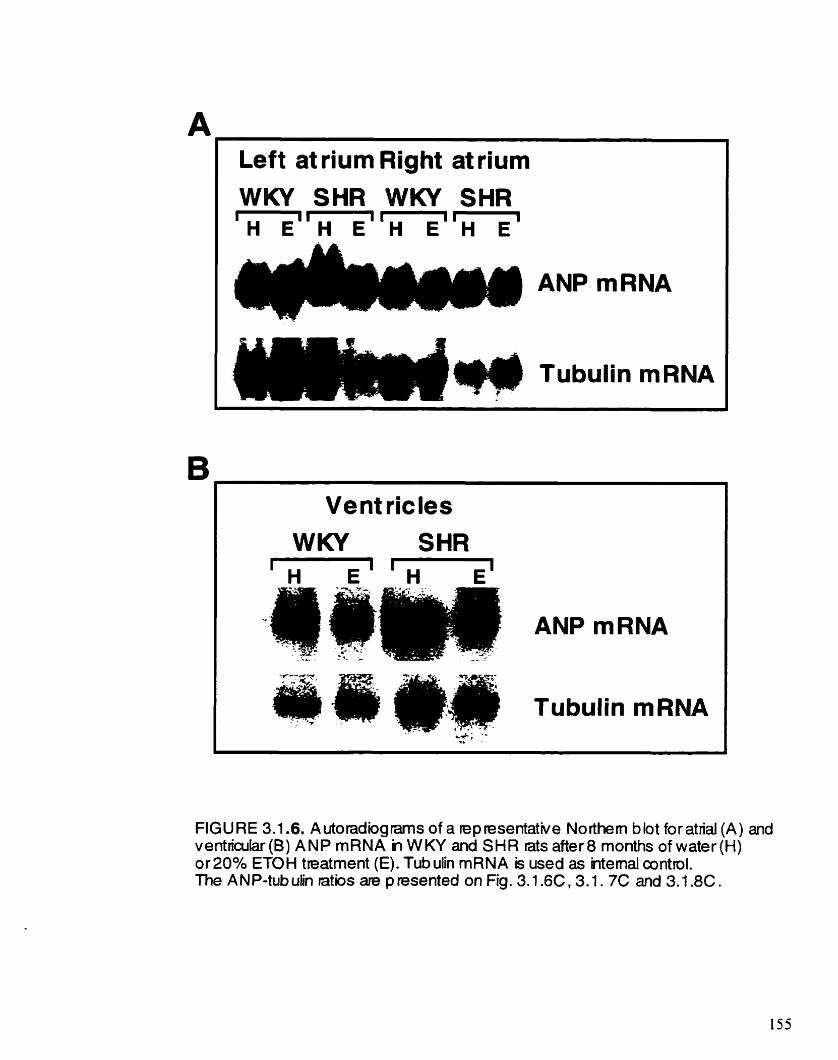
Left atrium Right atrium
WKY SHR WKY SHR
ANP mRNA
Tubulin mRNA
Vent ric les WKY SHR
ANP mRNA
Tubulin mRNA
FIGURE 3.1.6. Automdiogmms of a representative Nodhern blot foratnal (A) and ventricular(6) ANP mRNA in WKY and SHR mts after 8 rnonths of water(H) or 20% ETOH treatment (E). Tubulin mRNA is used as interna1 contiol. The ANP-tub UR ratios are p resented on Fig. 3.1.6C, 3.1.7C and 3.1.8C.
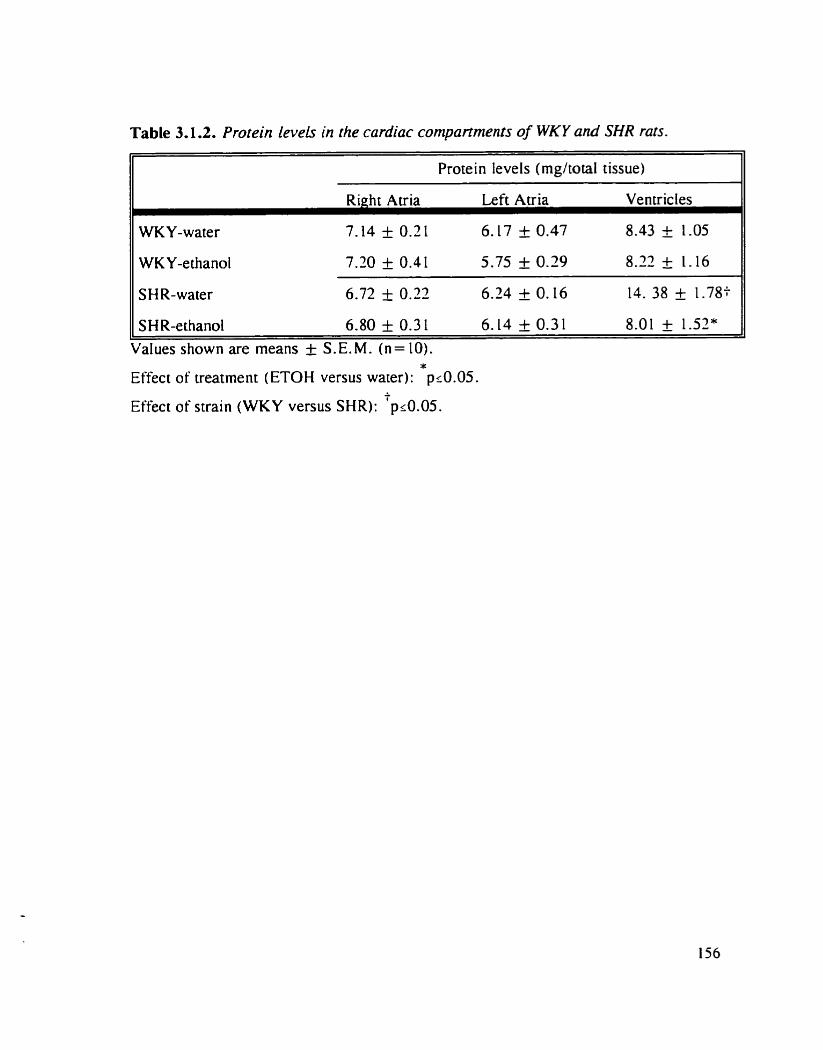
Table 3.1 -2. Protein levek in the cardiuc compartmenîs of WK Y and SHR rats.
II Protein levels (mghotal tissue)
Right Atria Left Atria Ventricles
II WKY -water 7.14 + 0.21 6.17 t 0.47 8.43 + 1.05
1 S H R-ethanol 6.80 * 0.3 1 6.14 & 0.3 1 8.01 & l X * Values shown are means & S.E.M. (n= 10).
* Effect of treatment (ETOH versus water): pc0.05.
Effect of main (WKY versus SHR): 'p0.05.

content represent the changes m the total mas of the tissue and are usually proportional to
the changes in tissue weight (Deshaies et al.. 1975: Szabo et nf., 1979), this result suggens
no modification by ETOH on the atrial weight. Chronic ETOH administration resulted in
significantly higher ANP concentrations in the right and left atria of SHR and WKY
animals. Correspondingly. total ANP content was significantly higher in ETOH- versus
H,O-treated SHR and WKY rats . Northern bloc analysis revealed that ETOH had little
effect on the atrial ANP mRNA content of either WKY or SHR (Fig. 3.1.3C.. 3.1 .K.
and 3.1.6A.). although ANP mRNA was significantly higher in SHR compared to WKY
rats.
The ventricles of the SHR presented significantly higher total protein content than those
of WKY rats mdicahg ventricular hypertrophy (Table 3.1.2. ). However. ETOH prevented
the development of this hypertrophy in SHR as indicated by the reduction of the total
ventricular protein content in SHR-ETOH rats to levels observed for the total ventncuiar
protein content of WKY rats (Table 3.1.2.). Accordingly . the ventricles (Fig. 3.1 -5. ) of
H,O-treated SHR presented significantly higher total ANP content and ANP concentration
than those of water-treated WKY rats. This tïnding was further supported by a significant
elevation of ANP mRNA in hypertensive compared to normotensive animals. [n S H R .
chronic ETOH treatrnent resulted in a significant decrease of total A N P content. but not
of ventricular ANP concentration (Fig. 3. MA. and 3.1.58.). There was no sigr'f rcant
difference in the toal content and concentration of venuicular ANP between the water- and
ETOH-treated WKY rats. Correspondingly . in Northtrn Blot studies. ventricular ANP
mRNA was significantly decreased by long-term ETOH administration in SHR but not
in WKY rats (Fig. 3.1.5C. and 3.1.68.). In fact. ETOH-treated SHR presented total
ventricular ANP content and ANP mRNA values which were similar to those of water-
and ETOH-treated WKY. RT-PCR analysis was performed on ventricular tissue as an
additional control of the northern blot results (Fig. 3.1.7.). PCR amplification of cDNA
and genomic DNA gave products predicted from gene sequences. No PC R amplitïcation
was obtained when cDNA was replaced by total RNA samples. The abundance of the A N P
transcript was estimated by cornparing the hybridization signal of the cDNA PC R products
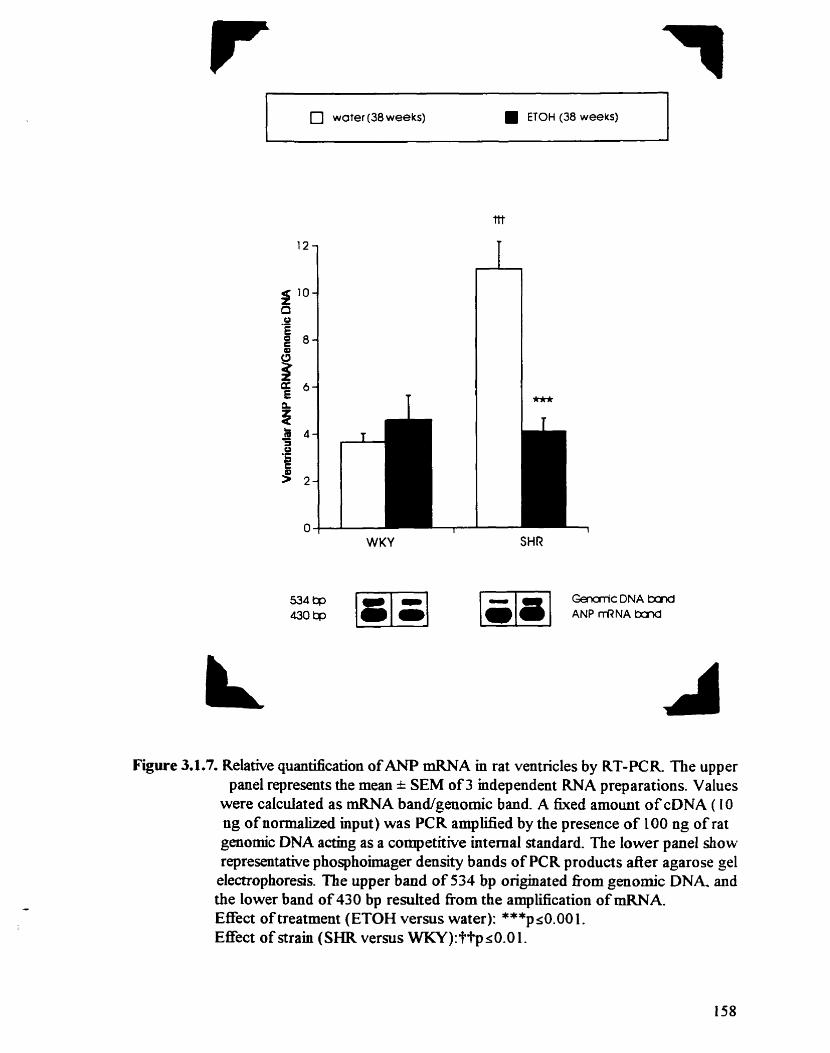
13 water (38 weeks) ETOH (38 weeks)
WKY SHR
Figure 3.1.7. Relative quantification of ANP mRNA in rat ventricles by RT-PCR The upper panel represents the mean * SEM of 3 mdependent RNA preparations. Values
were calculated as mRNA band/genomic band. A fixed amount of cDNA ( 10 ng of normalized input) was PCR amplified by the presence of 100 ng of rat gaiornic DNA acting as a cornpetitive intemal standard. The lower panel show representative phosphoimager d e n e bands of PCR products after agarose gel electrophoresis. The upper band of 534 bp originated nom genomic DNA. and the lower band of 430 bp resulted fiom the amplincation of mRNk Effect of treatment (ETOH versus water): ***p 50.00 1. Effect of strain (SHR versus WKY):Wp 50.0 1.

to the signal arising from the genornic template ( %). RT-PCR studies revealed a similar
significant decrease of ANP mRNA for SHR-ETOH only. without any modification in
WKY-ETOH rats (Fig. 3.1.7.). Again. the ANP mRNA level in SHR-ETOH was similar
to that in WKY-H.0 and WKY-ETOH.
3.1.4.3. Effect o f e t b o l on c i r m hormone leveh
Table 3.1.1. shows the blood alcohol . ANP. AVP, corticosterone and aldosterone
contenrs in the animals of the two strains. both before and after the 8 months experiment.
The animals were sacritïced during the light phase of the daily cycle (between 9 AM and
12) at a time when they are not active and are not consuming ETOH. Accordingly. the
blood alcohol content in ETOH-treated rats was generally low. although large variations
occurred. Plasma AVP levels were signitïcantly higher in the SHR-ETOH than in the
SHR-H,O group (pr0.01). but not in WKY-ETOH compared to the WKY-H,O group.
Circulating AVP levels were significantly decreased with age (psO.01) in both S H R and
WKY rats but. interestingly. significantly lower plasma AVP levels (pr0.01) were found
in young 7 weeks old S H R animais compared to age-matched WKY rats. Plasma
corticosterone levels had a tendency to be lower with ETOH but this never reached
significance. In contrat. plasma aldosterone levels were signitïcantl y lower in the S H R-
ETOH compared to the SHR-HIO group (pc0.05). but not between the ETOH and water
WKY groups.
3.1 S. DISCUSSION
The present studies demonstrated that chronic moderate ETOH consumption
prevented the age-dependent increase of BP in both hypertensive (SHR) and normotensive
(WKY) rats. The ETOH treatment was also associated with significant alterations in the
activity of the ANP system.
Since the pioneering work of Lian in 19 15. it has been well-documented that there
is a direct association between high ETOH consumption and hypertension. However. in
the last 15 years. with the publication of more extensive epidemiological studies. this

association has become less clearcut. especially with low and rnoderate ETOH
consumption (Gleiberman and Harburg. 1986: MacMahon. 1987). Indeed. it has been
repetitively reported that subjecn with moderate ETOH consumption (berween 1 and 3
standard drinks per day) had lower systolic blood pressure than both heavy drinkers and
non-drinkers (known as the U- or J-shaped curve) (MacMahon. 1987). Although some
investigators (Beilin and Puddey, 1992) still view this non-linear relationship as an artifact
(incorrect screening of subjects. etc.). a similar association between ETOH consumption
and cardiovascular diseases is now recognized (Marmot and Brunner . 1 99 1 ).
In animal models. where various confounding factors such as previous ETOH
exposure or nutritional status are easier to control. it was initially thought that ETOH at
moderate doses would either augment (Chan and Sutter. 1983) or have no significant effect
on BP (Khetarpal and Volicer. 1979). However. recent studies have shown that ETOH
in concentrations up to 70 % vlv may protect against the age-dependent increase in BP.
especially in hypertensive animals such as S H R and SHRSP (Sanderson et al.. 1983:
Jones et al.. 1988: Howe et al., 1989: Beilin et al.. 1992). Indeed. the present report
demonstrates a protective effect of ETOH on the age-dependent elevation of BP in both
hypertensive (SHR) and normotensive (WKY) rats. confirming previous reports
(Sanderson et al., 1983: Jones et al.. 1988: Howe et al., 1989: Beilin et al., 1992).
Interestingly. in WKY rats. ETOH appeared to completely prevent any age-dependent rise
in BP. Thus. this beneficiai effect of moderate alcohol consumption could be important not
only for hypertensive but also for normotensive individuals. However. i t is noteworth y that
the protective action of rnoderate ETOH consumption on the development of hypertension
is a slow process. requiring 23 weeks in SHR and 25 weeks in WKY rats to reach
statistical signi ticance.
The ETOH-induced decrease in BP could be due to dehydration and reduced blood
volume. However. this is unlikely since: a) the animals consumed equal volumes of water
and ETOH solution. b) no growth retardation was evident in ETOH-treated rats. as would
be expected if they suffered from chronic dehydration. c) plasma volumes have been
reporteci to remain unaltered w ith chronic administration of moderate amounts of alcohol

(Beilin m al.. 1992). and d) in studies where once a week water replaced ETOH solution
as the drinking fluid. to offset chronic dehydration. the BP-lowering effect of chronic
ETOH was not modified (Jones et al., 1988).
The mechanisms of this s d l e d "protective" action of alcohol are not known. In the
present experiments. we investigated the hypothesis that the A N P system may be
responsible. at least in part. for the antihypertensive action of ETOH. Indeed. A N P is
known to decrease BP (Needleman et ai., 1989: Nakao et al.. 19930. and in our
preliminary experiments we have observed that in humm. plasma ANP levels are elevated
by acute administration of low and moderate ETOH doses and that it thus may counteract
some of ETOH's pressor effects (Section 2 . 2 . ) . This was further affirmed in rats which
presented a rapid transient rise in plasma ANP levels following acute ETOH rreatment
(Section 2.1. ). 1 t thus appears important to investigate the impact of chronic ETOH
administration on ANP at a time when its antihypertensive effect is well-developed. The
previously pub1 ished chronic ETOH s tud ies have investigated the variations of natr iuret ic
peptides in normotensive (Sprague-Dawley ) rats follow ing a 6-week period of ETOH
ueatment (Wigle et al.. 1993b.c). at which time the protective effect of ETOH on BP has
not been developed yer.
Plasma ANP levels have been reported to be elevated in various models of
hypertension. in SHR (Morii et al., 1985: Gutkowska et al.. 1986a: Ruskoaho and
Leppaluoto. 1988). DOCA-sa1 t (Itoh et al.. 199 1 : Kohno et al.. 1992a) and one-kidney .
one-clip ( 1 K IC) hypertension (Garcia et al.. 1987). In the present studies. we have also
found significantly higher plasma ANP values in hypertensive animals. Circulating A N P
levels were measured by direct RIA. as described previously (Bidmon et al.. 1991 and
Section 2.1 .). This elevation appears to be secondary to the rise in BP and is therefore
believed to be. in both animal models and in humans (Hollister and Inagami. 1991). a
compensatory mechanism to prevent a furrher increase of BP. Similarly. in the present
studies. the significantly lower circulating ANP values in ETOH-treated rats (which
showed lower BP levels) could be the resul t of a diminished demand for counterbalancing

mechanisms.
In the present experirnents. moderate ETOH consumption elevated ANP
concentration and content of both the left and right atria. without altering ANP mRNA.
It can therefore be argued that the observed lower circulating ANP levels are due to
decreased ANP release From both atria. The increased ANP accumulation in atria and the
finding that atrial ANP mRNA is not decreased. but maintained at the sarne level in
ETOH-treated animais suggest also that the system is not down-regulated by long-term
ETOH use. Interestingly. higher atrial ANP mRNA and total ANP contents have been
observed in ETOH-treated (6 weeks of 20% v/v alcohol) Sprague-Dawley rats during high
BAC levels (Wigle et al.. 1993b). This rnay explain the absence of lower atrial ANP
mRNA levels during low BAC levels. despite the lower BP. Ir is possible also that when
an i mals treated chronical ly wi th moderate amounts of alcohol are exposed to stressful
situations or conditions which would increase BP (i.e.. via heightened sympathetic
activity). the atria rapidly release the larger quantities of stored ANP. thus counteracting
the hypertensive effects and preventing the development of hypertension. If this is true.
decreased plasma ANP concentrations could be expected under the normal non-stress
conditions and low BAC levels of the present studies as direct consequence of the ETOH-
induced fa11 in BP. Further experiments are needed to test this hypothesis.
Initially. it was thought that ANP was localized exclusively in heart atria (De Bold
et al.. 198 1). However. scientists have now demonstrated that ventricles synthesize and
release natriuretic hormones such as ANP and BNP. especiall y under pathophys iological
conditions (Arai et al.. 1988: Gutkowska and Nerner. 1989: Ogawa et al.. 199 1 ). 1 ndeed.
it appears that a shift in ANP synthesis and release from the atria to the ventricles takes
place with hypertension. leading to a significant increase in the proportion of circulating
ANP derived boom hypertrophied ventricular tissue (Ogawa et al. , 199 1 : Dagnino et al. .
1992). The same effect was seen in the present studies where adult S H R showed higher
ventricular ANP content and ANP mRNA than adult WKY. Interestingly. chronic ETOH
treatment prevented the development of the ventricular hypertrophy in SHR. as indicated
by the significant decrease in total ventricular protein content. total ventricular ANP

content and ventricular ANP mRNA compared to water-treated SHR, Since the BP
elevations in these animals are markedly reduced. bringing them closer to normotensives.
the shift in ANP production from atria to ventricles is thus likely to occur to a lesser
extent. This marked difference between WKY and S H R at the level of ventricular ANP
is likely to explain the greater decrease in circulating ANP observed in S HR compared
to WKY following ETOH treatment. Whereas the mild. albeit signitïcant decl ine of plasma
A N P in WKY-ETOH rats appears to be the result of atrial alterations only (decreased
release). a funher mechanism (reduced activity of the ventricular ANP system) seems to
be present in SHR. This would explain the greater effect of ETOH treatment on the ANP
system in SHR. Knowing that the ventricles have little or no storage space (Lang et al..
1992). in contrast to the atria. and therefore that ANP is released constitutively . a 3-fold
decrease of ANP mRNA in SHR will be reflected directly in plasma ANP levels. Of
course. in such a scherne. prevention of the age-dependent progression of hypertension by
ETOH is still the cause of the different modifications in the cardiac ANP systern of S H R
and WKY rats.
In fact. the important question is whether this general decrease in plasma and
ventricular ANP observed after chronic moderate ETOH consumption is secondary to the
effect of ETOH on BP? It is indeed possible that initially (as seen in acute studies for
example. Chapter 1). the activity of the ANP system is elevated by ETOH and this
increase is sufficient to mediate some of the long-term actions of ETOH on BP. The
observed results may therefore be compensatory to the repeated stimulatory effect of acute
ETOH consumption on the ANP system. such that its sensitivity is elevated in ETOH-
treated rats. Sirnilar compensatory reactions to a repeated stimulation by ETOH have been
demonstrated for several other physiolog ical systems. such as AVP. N M D A receptors.
GABA, receptors. calcium channels or Na'/K+-ATPase activity (Diamond and Messing.
1994; Nua and Peters. 1994). On the other hand. the reported ETOH effect on the heart
A N P system may also simply be secondary to iü impact on BP.
Acute ETOH consurnption mduces diuresis through the inhibition of AVP release
(Eisenhofer and Johnson, 1982). In contrast, chronic ETOH consumption is associated with

some stimulations of the AVP systern to compensate for the continuous presence of a1cohoL
resuiting m the possibility of some water retention (Eisenhofer et al.. 1985: Taivainen et a/. . 1995). Furthemore. ANP is known to suppress AVP release from the supra-optic nucleus
(Januszewicz et al.. 198613: Clark et ai.. 199 1) . Therefore. the significant elevation of
circulating AVP in the SHR-ETOH group may indicate a heightened activity of this
hormone with chronic ETOH treatment because of its lower inhibition by the significant
fail in plasma ANP. More studies are needed on the effects of ETOH on the interactions
between these two hormonal systems and on the role of these interactions in cardiovascular
regulation. Circulating aldosterone Ievels were also significantly lower in the SHR-ETOH
group. indicating a decreased activity of the renin-angiotensin-aldosterone system (RAAS)
in hypertensive anirnals chronically consuming alcohol. A similar lowering of plasma
aldosterone levels was observed in normotensive Sprague-Dawley rats by Wigle er al.
(1993b). but not in the WKY controls used in the present studies. Genetic differences
could explain this discrepancy. Indeed. there are genetic differences in the renin gene of
WKY and SHR (Kreutz er al.. 1993). I t would be interesting to Further investigate the
activity of the RAAS system in S H R as a possible rnechanism underlying the protective
effect of moderate ETOH administration.
In conclusion. the present investigations have demonstrated that the prevention of the
age-dependent increase in BP by chronic moderate ETOH consumption is associated with
significant aiterations of the heart ANP system. Indeed. circulating ANP levels are lower
fol low ing long-term chronic ETOH treatment and the development of ETOH-induced
protection against elevated BP. Atrial ANP concentration is slightly augmented without
significant variation of atrial ANP mRNA. which could suggest decreased atrial A N P
release. Ventricular ANP content and ANP mRNA are significantly lower in the SHR-
ETOH group. suggesting a possible reduction of ANP release from the ventricles.
Further experiments are needed during ETOH intoxication and with other components of
the natriuretic peptide system. such as B N P levels (Wigle et al.. 1993~). renal ANP
receptor density and central ANP and C-type natriuretic peptide. to understand the
interactions between ETOH and this system and to ascertain its possible importance in the

so-cal led " protective " effect of chronic rnoderate alcohol consumption observed in both
humans and experimental animals.
3.1.6. ACKNOWLEDGEMENTS
The authors thank Mr. Ricardo Claudio whose help in the care of animais made this
work so much easier. Special thanks are also given to Céline Coderre. Nathalie Charron
and Sylvie Laroque for their excellent technical assistance. This work was supported by
gram tiom Medical Research Council of Canada (MT- 10337 and MT- 1 1674). Canadian
Heart and Stroke Foundation. Caiiadian Kidney Foundation (J.G.) and by the Alcohol
Research program at Douglas Hospital (C.G.). P.G. is recipient of a studentship from the
"Fonds pour la formation de chercheurs et l'aide à la recherche" (FCAR).

Section 3.2.
EFFEXT OF CHRONIC MODERATE ETHANOL CONSWPTION ON
HEART BRAIN NATRIURETIC PEPTIDE
P. Guillaume, M. Jankowski, J. Gutkowska and C. Gianouiakis
European Journal of Pharmacology
(Eur ] Pharmacol, 3 16: 49-58, 1996)
Contniution bv CO-authors: Dr. M. Jankowski introduced me to the technical aspects of
Northem blotthg and was responsile for the RT-PCR studies. Dr. J. Gutkowska and Dr. C.
Gianouiakis were my CO-supervisors.
Acknowledgements: R Claudio was the animal technician. C. Coderre and N. Charron
performed the BNP iodinations.

3.2. EFFECI' OF cRRONIC MODERATE ETHANOL CONSUMPTION
ON HEART BRAIN NATFUURETIC PEPTIDE
3.2.1. ABSTRACT
There is experimental evidence indicating that chronic moderate ethanol consumption
delays the age-dependent mcrease m blood pressure. Smce the brain natriuretic peptide (BNP)
is a potent hypotensive hormone. the effect of chronic ethanol treatment on the heart BNP
systern was mvestigated ushg spontaneously hypertensive ( S m ) and Wistar- Kyoto ( WKY )
rats. Chronic moderate ethanol consumption resulted in significantly lower circulating BNP
IeveIs for both SHR (206.9 * 18.5 versus 306.9 i 28.1 pg/rni, n= 12. Ps0.05) and WKY rats
( 13 1.3 * 20.7 versus 220.6 * 25.0 p g / d n= 12. Ps0.05). Lefi and right atrial BNP content
and concentration in WKY rats and tefi atrial BNP content and concentration m SHR rats
were aupented by the ethanol treatment. but not atrial BNP mRNA. in ventricular tissue.
alcohol had no effect on total BNP content of either SHR or WKY rats. but it induced a
significant elevation in ventncular BNP concentration (pg/mg protein) and BNP rnRNA in
SHR but not WKY rats. Thus. chronic ethanol treatment resulted in specsc alterations in
the a c t ~ t y of the heart BNP systern.
3.2.2. INTRODUCTION
Brain natriuretic peptide (BNP). originaliy discovered in porcine brain (Sudoh et al..
1988) is now considered the second member of the natriuretic peptide family. the k a being
atrial natriuretic peptide (ANP) (Davidson and Struthers. 1994. Nakao et al.. 1993 ). BNP is
an NH2 elongated version of ANP with hi& sequence homology in its ring anicture (Lang
et ni.. 1992). However. in contraa to other natriuretic hormones. its aminoacid sequence
displays important interspecies diversity (Davidson and Struthers. 1994). In the rat. the -15
aminoacid BNP molecule is believed to be as potent as ANP in its ability to lower blood
pressure and to cause natriuresis, diuresis and vasorelaxation (Holmes et al.. 1993: Lang et
aL. 1992). Ahhough BNP is colocalized with ANP in atrial granules (Thibadt et al., 1992).
the major site of BNP synthesis and secretion in the body is the ventncle (Kohno et al..

l9Wa: Ogawa et al.. 199 1 ). Moreover. it appears that changes in the a c t ~ t y of ventricular
BNP correlate better with the progression of hypertension as well as other pathophysiologicd
conditions, such as congestive heart failure. than changes in the a a ~ t y of the ANP syaem
(Naruse et al.. 1994: Lang et al.. 1992: Kohno et al.. 1992~).
It has been reported by Howe et ni. ( 1989) and confirmed by us (Section 3.1.) that
chronic moderate ethanol consumption prevents or delays the age-dependent increase in blood
pressure in both nonnotensive Wktar-Kyoto ( WKY) and spontaneously hypertensive ( SHR)
rats. The mechanimis underlying this effect are ni1 unknown but the activation of one or
several uiembers of the natriuretic peptide family by chronic ethanol is an attractive
hypothesis. Furthermore. circulating B NP levels were recent ly found t O be elevat ed following
a short chronic ethanol treatmeot (six weeks) (Wigle et al.. 1993~). However. since the
prevention of the age-dependent increase in blood pressure by chronic moderate ethanol
consumption is a progressive effect. observed only afier 22 weeks of alcohol administration
(Section 3.1 .), it was the objective of the present studies to investigate the changes of the
plasma and heart BNP levels foliowing chroaic ethanol treatment for a period sufncient to
demonstrate significantky lower blood pressure values in the ethanol-treated compared to the
ethanol-naive rats. Circdating, atrial and ventricular BNP levels were measured by
radioimmunoassay before and after the chronic administration of ethanol. Contents of BNP
rnRNA were also evaiuated in the atria and ventricles of SHR and WKY rats.
3.2.3. MATERIALS AND METHODS
A total of 72. 6 weeks-old. rats (36 male SHR and 36 male WKY rats) were purchased
fiom Charles River Breeding Laboratones (St-Constant, Québec. Canada). Following one
week acclirnatization period, the animals from each strain were randomly assigned to one of
the following 3 groups: 1 ) the initial age control group; 2) the water-treated control poup
and 3) the ethanol-treated group. in the initial age control group. 12 (7 weeks-old) SHR and
WKY rats were sacrificed to d o w estimation of the eEect of age on the BNP system. In the
water-treated control group, 12 SHR (SHR-water) and 12 WKY (WKY-water) rats were
given fiee access to drinking water for the 8 months of the experimental period. In the

ethanol-treated group, 12 SHR ( SHR-ethanol) and 1 2 WKY ( WKY-ethanol) rats were @en
6ee access to an ethanol solution that was gradudy increased to 20% viv in 15 days (5% viv
for 5 days 10% v/v for 5 days. 15% viv for 5 days and 20% v/v subsequently) (Howe et ai..
1989). The ethanol treatment laaed 8 months. AU animals were maintamed on a 12 h dark
and 12 h Li& cycle (lights on at 6:00 a.m) and _&en fiee access to pellet chow (Purina.
Richmond. Va) during the experirnental penod. The blood pressure and the heart rate were
measured monthly by the tail-cuff method (ffeffer et ai.. 197 1 ). At the end of the
experirnental penod the animals were sacrificeci. between 9:00 to 1 200 a. m.. during the light
cycle.
T h blood was collected in chilled 15 ml conical centrifùge tubes containing 1 mglm1
ethylenediaminetetraacetate (EDTA). 10 mM phenylmethylnilfonyl fluoride ( Sigma Chemical
Co.. P-7626) and 5 mM pepstatm A (Sigma Chemical Co.. P-4265). A 50 pl aliquot of whole
blood was placed m 450 pl of ice-cold trichloroacetic acid (6.25Y0 w/v. Sigma) for estimation
of the blood aicohol content at the tirne of death using the dehydrogenase enzymatic rnethod
(Hawkins et al.. 1966). The remaining blood was centrifuged and the plasma was stored
frozen at -75°C until assayed for BNP content. From each group. 6 animals were sacrificed.
the ventricles. lefi and nght atna were dissected and placed in separate tubes containing ice
cold 0.1 N HCI. The tissues were boiled for 5 min and homogenized using a Polytron
(Kmernatica GMBK Luzem S h r l a n d ) . Protems were estimated by the Bradford's method
(Bradford. 1976). The atria and ventricles fiom the remaining 6 animais per goups were
dissected. immediately fiozen in isopentane and aored at -75°C for estimation of BNP
rnRNA.
Plasma BNP was extracted fiom 2 ml plasma using C,, Sep-Pali cartridges (Millipore.
Milford, Ma) and was measured using a second-antibody radioimrnunoassay (RLA) ( Itoh er
ai.. 1989). Bnefly. the iodination of BNP was doue by the lactoperoxidase method. as
demibed elsewhere for ANP (Gutkowska et ai.. l987a). The standard curve was prepared
by serial dilution of synthetic rat BNP (fiom 12.2 to 3 125 pg/ml). The standards and
unknowns were incubated for 48 h at 4°C with an antibody specific for the rat BNP
(Peuinsula Laboratories, h a 1 dilution 1 :8000). This antibody reacted 100% with rat BNP-( 1-

45 ) and cross-reacted 100% with rat BNP-( 1-32), but had no cross-reactivity with rat or
human ANP. porcine or human BNP. or with arginine vasopressin. angiotensin II and
endothelin-1. %BNP (6000 cpm) was then added and incubated for 24 h at 4OC. To
separate bound fiom 6ee. the second-antiiody (GARGG. 1 5 0 dilution) and the n o m l rabbit
serum (NRS. 135 dilution) were added and lef€ to mcubate for 4 h at room temperature.
FinaUy. 6.25% polyethylengiycol was added prior to a 20 min (3000 g) centrifugation. the
supematants were aspirateci and the pellets counted in a gammacounter (Canberra Packard).
The sensitMty of this assay is 24 pdtube. The &a- and interassay coefficients of variation
were 5.8 and 9.5%. respectively. Resdts were expressed as pgml plasma.
Atrial and venaicular homogenates were ccentrifuged, the supematants were appropnately
diluted and the BNP content was measured directly using the same RIA procedure descnbed
for estimation of plasma BNP levels. Results were expressed a s pg/tissue (total content) and
@mg protein (concentration).
The RNA extraction Iiom the atrial and ventricular tissues was done by the acid
_müinidinium thiocyanate phenol chlorofonn method (Chomczynski and Sacchi. 1987). Total
RNA ( 10 yg) was separated by electrophoresis ( 1 SOh agarose gel containing 0.22 M
formaldehyde) and blot transfemed to a nylon membrane. B lots were su bsequently h ybndized
with random primed [a-"Pl-labelled cDNA probes corresponding to BNP and a-tubulin
mRNA sequences. Random priming kit (Gibco. BRL. Bethesda. MD) and [a-"PI-~CTP
(3000 Ciimmol) (Amersham. Arhgton Heights. IL) were used for labelling the probes. As
BNP probe, a 347 bp fiagment covering sequences in the 5'- untranslated region of the rat
BNP cDNA and in the 3'- sequence of the second exon of BNP gene was used (Dagnino et
al.. 1992). This fragment was generated by reverse transcription of rat ventncular RNA. As
a-tubulin probe. a 550 bp fragment of a-tubulin cDNA was generated by reverse
transcription of rat atrial RNA and amplified by the polymerase chah reaction (PCR)
(Dapino er al.. 1992). Membranes were hybrichd in b a e r containing 120 rnM Tns. 8 rnM
EDTA, 0.6 M NaCL 0.0 1% Na pyrophosphate. 0.02% sodium dodecyl sulfate (SDS). 0.03%
heparin (Sigma Chernical Co, St-Louis, MO. H7005), pH 7.4 and supplemented with 10%
dextran sulphate and IO6 dpdrnl of radioactive labelled probe. Washing was doue in 2 X

sodium chloride-sodium citrate buffer (SSC)/ 1% SDS ( I O min at room temperature). 0.1 X
SSC1O.O 1 % SDS ( 15-30 min at 65OC) and 0.1 X SSC ( 10 min at room temperature). The
inteosity of the hybridization signals were subsequently quantitated by densitomet~y using the
PhospliorIrnager (Molecular Dpamics. Sunnyvale. CA). In preliminary midies. we used both
a-tubulin and GAPDH probes to control for the hybridization signal. Our results indicated
that there was no sigiilficant effect of ethanol on either a-tubulin or GAPDH mRNAs. Thus.
to correct for ditferences in the amount of total mRNA applied on the electrophoresis geL a-
tubulin was used as intemal control and the results are expressed as the ratio of BNP/a-
tubulin rnRNAs.
RT-PCR was conducted according to previously described procedures (Dagnino et ai-.
1992). Normalized input of cDNA with 60 ng of liver @NA. 0.2 rnM dNTPs. 10 pCi "P
(ha1 concentration) and 40 pmol of fonvard and reverse primers correspondhg to sequences
in the 1 and 2 exon ofthe BNP gene. were added for amplification with 2.5 units of Thernzzrs
aqziuticzcs (taq) DNA polymerase ( Perkin-Elmer-Cetus. Nonvalk CT. USA). The following
primers were used for the PCR ampiifkation reaction: BNP fonvard. 5'-
CCATCGCAGCTGCCTGGCCCATCACTTCTG-3'. BNP reverse. 5'-
GACTGCGCCGATCCGGTC-3'. PCR amplification was camed out with 25 cycles of 1 min
at 94'C. 1 min at 6 I°C and 3 min at 72°C. Amplification of the cDNA and the genomic DNA
templates yielded respectively a 347-bp and a 546-bp fragment. M e r separation on 1.5
agarose gels BNP cDNA (lower band) and genomic cDNA (upper band) were measured on
Phosphorlrnager (Molecular Dynamics. Sunnyvale. CA) and BNP cDNA amplification
products were normaliied to the amount of genomic fragments.
Data are presented as means 5 S.E.M. The significance of difference among the various
groups was calculated by a two-way analysis of variance (ANOVA). This analysis was
followed by the Neuman-Keuls multiple cornparison test. A P value of sO.05 was considered
si+Pnificant.

33.4.1. Effert of age and ethanol on bodv weight. blood oressure. heart rate and total
grotein content in atrial and ventricular tissues.
SHR and WKY rats received a 20% v/v solution of ethanol for 8 months (32 weeks).
Controls of both grains had fiee access to normal water. Table 3.2.1. shows the changes in
body weight and daily fluid intake at various steps during the experirnental period. No
sipificant Merences were observed between the body weights of the water- vernis the
ethanol-treated animals. or between the daily fluid mtake of the water- versus the ethanol-
treated rats. For each strain of rats. a two-way ANOVA with the treatment as the fira
independent variable and time as the second independent variable was perfomed for the
blood pressure and heart rate (Table 3.2.2.). For the blood pressure. s i ~ c a n t effect of
treatment (F( l.l62)= 17-92. Ps0.00 1 ), time (F(8.162)=34.50. Ps0.00 1 ) and a significant
interaction oftreatment with time (F(8.162p4.95. Ps0.00 1 ) are demonstrated for SHR rats.
Sirnilarly. significant effects of treatment (F( l.l62)=J.O 1. Ps0.05). time (F(8.162)=4.77.
Pr0.00 1 ) and treatment by time interaction (F(8.162)=3.53. PiO.0 1 ) are present for WKY
animals. hdeed. chronic moderate ethanol consumption resulted in significantly lower BP
values from the 6' (in SKR) and 7' (in WKY) month oftreatment onward. as compared to
age-matched water-treated controls (Table 3.2.2.). For the heart rate. the two-way ANOVA
indicates a significant effect of treatment (F( J.l62)= 19.8 1. Ps0.00 1 ) and time
(F(8.162)=8.64. Ps0.00 1 ). but no significant interaction between treatment and time
(F(8.162)= 1.85. P=0.08) for SKR rats. A significant effect of treatment (F( 1.162)=4.50.
PsO.05) and time (F(8.162)=19.67. Pi0.00 1). but no significant interaction of treatment by
time (F(8,162)= 1 .O4. P=OA l ) is also observed in WKY animals. Chronic moderate ethanol
consumption significantly lowered beart rate values during the 7m month of treatment in WKY
rats and during the 8" month of treatment in SHR animals (Table 3 2 . 2 . ).
The total protein content in the atria and ventricles of SHR and WKY rats is s h o w on
Table 3-22 , in most cases. changes in total protein content represent changes in the total
mass of the tissue and are usually proportional to changes m tissue weight ( Szabo et al.. 1979:
Deshaies et al.. 1975). With age, both SHR and WKY rats presented similar increases in the
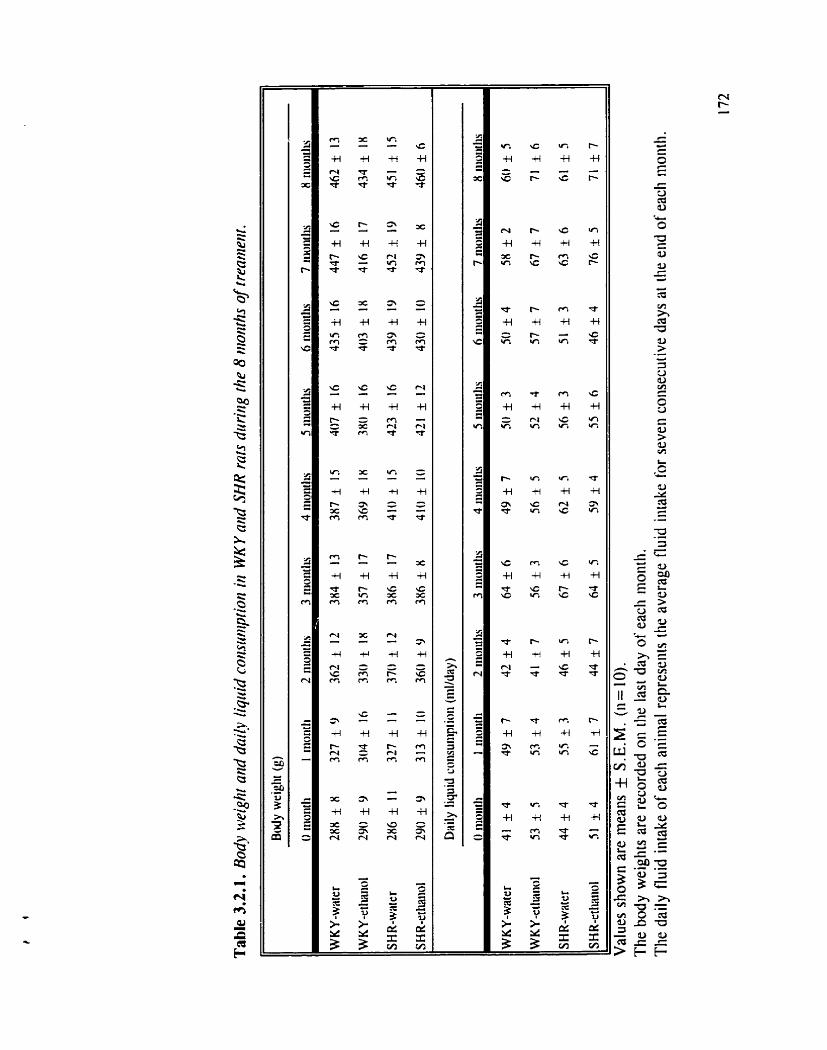
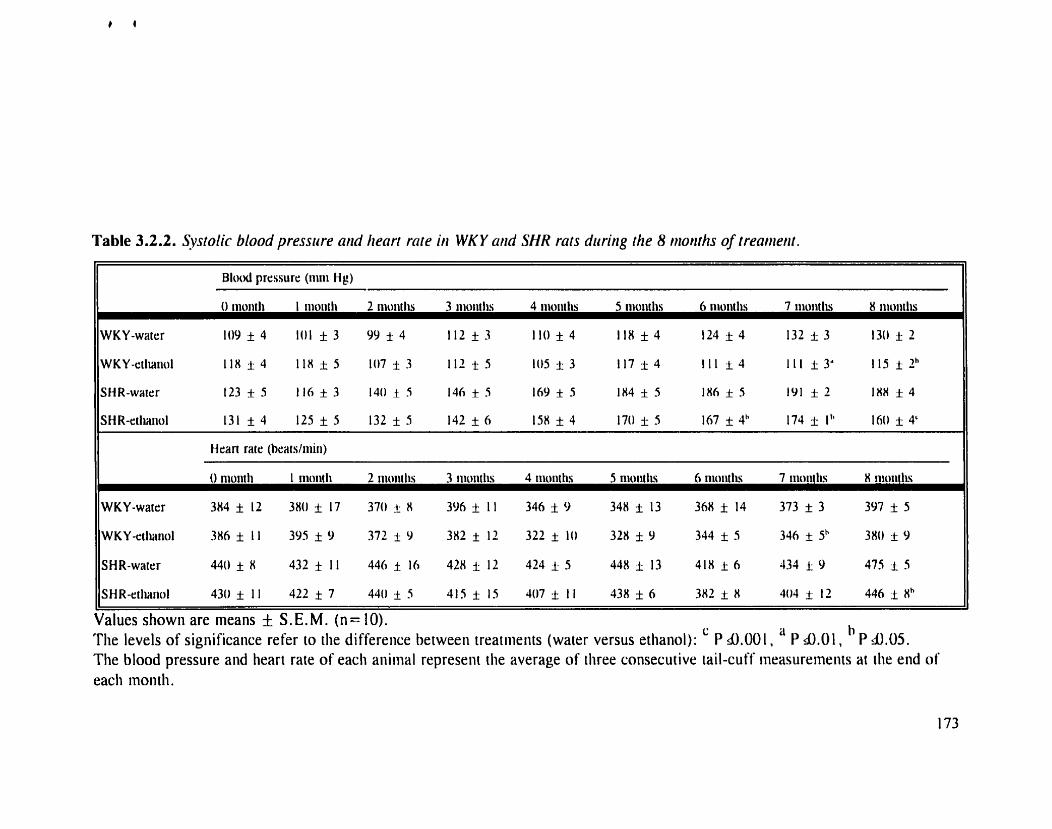
Table 3.2.2. Systolic blood pressrire attd heart rote iti WKY attd SHR rots diirittg the 8 mnths of treametrt.
BlotKi pressure (iiiiii Hg)
Hran rate (beats/iiiiii)
- -
I
1
1
4 .
I 1
7
(
l
Values shown are means f S . E . M . ( n = 10). C h The levels of significaiice refer to the differeiice betweeii treatiiieiits (water versus ethaiiol): P 4.00 1. 9 4.0 1 , P iU.05.
WKY-water 3 X 4 f I 2 3 8 0 f 1 7 3 7 0 3 3 3 9 6 k 1 1 3 4 6 1 9 3 4 8 f 1 3 3 6 8 1 1 4 3 7 3 f 3 397 f 5
WKY -eilüiiiol 3 H 6 f l I 3 9 5 f 9 3 7 2 i : 9 3 8 2 1 1 2 3 2 2 1 1 0 3 2 X f 9 344 f 5 346 I Sh 380 f 9
SHR-water 4 4 0 1 8 4 3 2 f 1 1 4 4 6 f I 6 4 2 8 k 1 2 4 2 4 k 5 448 f 13 418 1: 6 434 k 9 475 k 5
SHR-etli;iriol 4 3 O f I I 4 2 2 1 7 4 4 0 k 5 4 1 S f I S 4 0 7 k 1 1 33H f 6 382 f X 404 f 12 446 f Xh
The blood pressure and heari rate of eacli aiiiinal represent the average of tliree consecutive tail-cuff ineasuremeiits ai the end of eacli inontli.

Table 3 .LS. Protein fevels in the cardiac compartments of WKY and SHR rats.
Protein levels (mgltotal tissue)
Right Atria Left Atria Ventricles
WKY-ini tial 1.28 & O. 17 1.00 f 0.19 4.40 0.40
WKY-water 7.08 I 0. lga 6.19 I 0 . 5 8 ~ 8.54 + 1.12 b
WK Y-ethanol 7.12 + 0.42 5.24 f 0.17 8.19 _+ 1.26
SHR-initial 1.50 & 0.13 0.86 + 0.07 4.49 & 0.38
SHR-water 6.51 O. 19" 6.36 + O. 17" 14. 13 f 1 .82"~
S H R-ethanol 6.66 0.27 6.10 f 0.34 7.88 + 1.40'
Values shown are means & S.E.M. (n=6) . a h Effect of age (initial versus water): Pi0.001. C ~ r ~ . ~ l . Pc0.05. d Effect of strain (WKY versus SHR): P50.05.
Effect of treatment (water versus erhanol): ' ~ 1 0 . 0 5 .

total protem content m the atria, suggesting similar mcrease m the atrial weight. However.
even though at 7 weeks of age the total protem content m the ventricles of SHR and WKY
rats is Smilar, at 38 weeks of age the ventricles of the SHR rats presented significantly higher
total protein content compared to age-mtched WKY rats, mdicating that agkg mduced a
larger increase in the ventricular tissue weight of the SHR animals. Chronic moderate ethanol
consumption had no signifïcant effect on the total protem content m the atria and ventncles
of the WKY rats and in the atria of SHR animais. However, ethanol prevented the age-
induced mcrease in ventricuiar total protein content in SHR rats, suggestmg the prevention
of the ventricular hypertrophy usually observed with age m this strain.
3.2.4.2. Effect of age on BNP
Young (7 weebold) and aduh (38 weeks-old) SHR rats have sigiilficantly higher plasma
BNP values than WKY animals of the same age (3 32.3 p g h l 16.9 versus 194.9 pg/ml*
25.5, n= 12, Ps0.0 1, for 7 weeks-old anirnals and 306.9 p g / d 28.1 versus 220.6 pg/ml*
25.0, n=12, PsO.05, for 38 weeks-old rats) (Fig. 3.2.1.). Total BNP content (pg) is
significantly increased with age in both the nght and the lefi atria of SHR and WKY rats (Fig.
3.2.2A. and C.), while the concentration (&mg protem) is significantly decreased (Fig.
3.2.2B. and D.). As observed for the atria, total ventricular BNP content is increased with age
in both strains of animais (Fig. 3.2.28.). However, contrary to the atna, ventricular BNP
concentration is also augmented in the adult compared to young h l s (Fig. 3.2.2F. ).
Companson of the heart BNP content between the SHR and WKY rats of the same age
mdicated: (a) 7 week-old SHR rats presented a higher total BNP content in the nght atria. but
not in the lef€ atria and the ventncles, compared to 7 week-old WKY rats. No significant
difference is obsewed in the concentration of BNP in the right and left atna or in the
ventricles between the 7 week-old SHR and WKY rats. (b) At 38 weeks of age, the total BNP
content is significantiy higher m the right and lefi atria and in the ventricles of SHR compared
to WKY rats. However, this significant Merence in the total BNP content is not associated
with a Pgnificant Merence m the BNP concentration (&mg protein) in either the nght and
left atria or the ventricles of the 38 week-old SHR versus WKY rats.
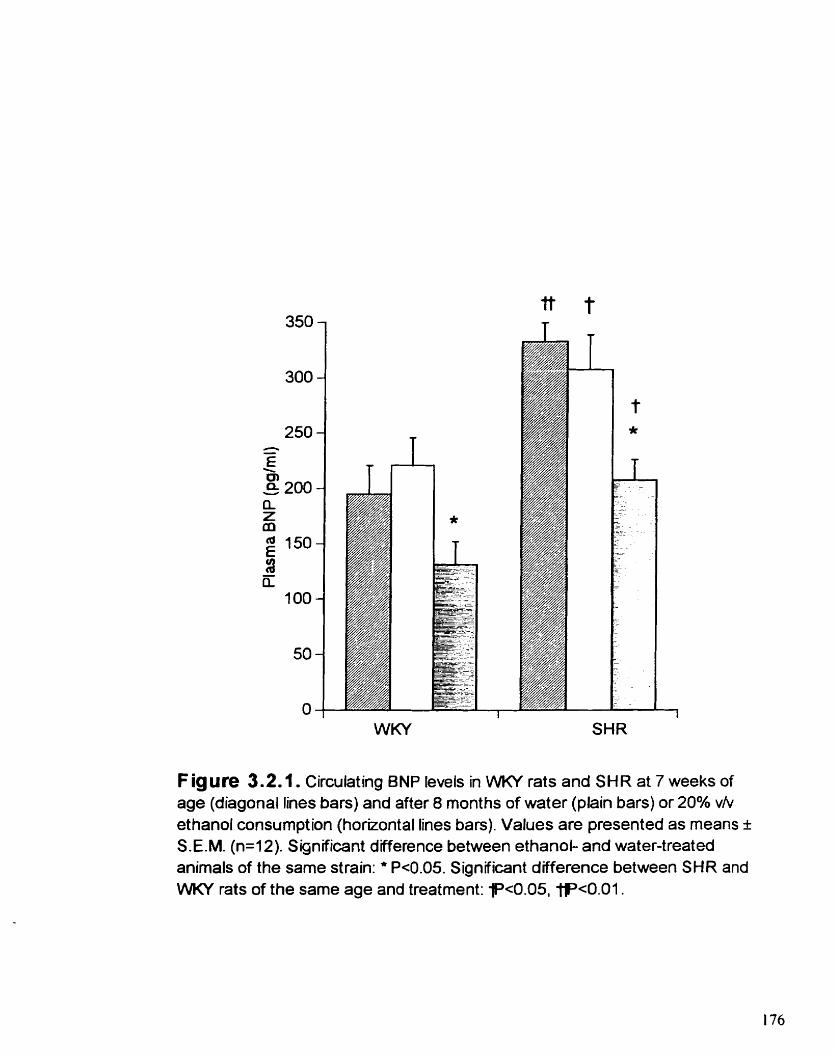
WKY SHR
F ig urê 3 .2.1. Circulating BNP levels in WKY rats and SHR at 7 weeks of age (diagonal lines bars) and after 8 months of water (plain bars) or 20% v/v ethanol consurnption (horizontal lines bars). Values are presented as means i S.E.M. (n=12). Significant difTerence between ethanol- and water-treated animals of the same strain: * PcO.05. Significant difference between SHR and WKY rats of the same age and treatment: P<0.05, W<O.O1.
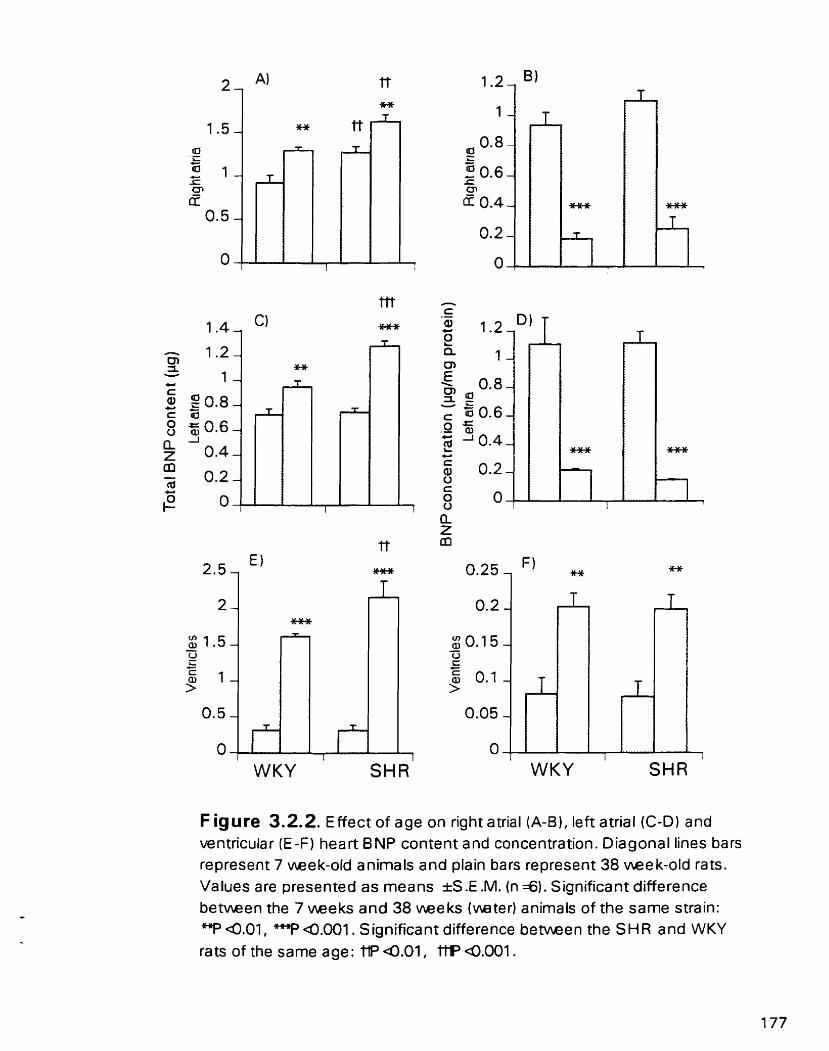
a wKY SHR SHR WKY
F igure 3.2.2. Effect of age on right atrial (A-B), left atrial (C-Dl and ventricular (E-F) heart BNP content and concentration. Diagonal lines bars represent 7 wek-o ld animals and plain bars represent 38 wek -o l d rats. Values are presented as means +S .E .M. (n =6). Significant difference be-en the 7 w e k s and 38 w e k s (nater) animals of the same strain: +(P 4.01, 4.001. Significant difference b e t w e n the SHR and WKY rats of the same age: ttP 4.01. ttP 4.001.

3.2.4.3. Effect of ethanol on BNP
At the time of sacrifice, the blood alcohol content detected was very low for both strains
(29.97 * 11.24 and 38.27 =: 17.82 mg/dL n=6. in WKY-ethanol and SHR-ethanoL
respectively). This low blood alcohol content is due to the fact that the animals are sacrificed
d u ~ g the light cycle (between 9:00 and 12:OO a m ) wben they are not actively consmhg
ethanoL Fig. 3.2.1. demonstrates that the circulating BNP levels are significantly lower in the
ethanol-treated rats of both strains. Plasma BNP is estimated to be 306.9 * 28.1 and 206.9
= 18.5 p g / d (n= 12. Pi0.05) in SHR-water and SHR-ethanol respectively. a 33% decrease.
and 220.6 * 25.0 and 13 1.3 * 20.7 p g M ( 0 4 2 . Ps0.05) in WKY-water and WKY-ethanol.
a 30% decrease.
The ethanol-induced changes m the total content (pg) and concentration (pumg protein)
of BNP. as well as BNP mRNA in the hean right and lefi atria and ventricles are s h o w on
the figures 3.2.3. to 3.2.7. WKY rats have increased right atrial BNP content and
concentration following chronic moderate ethanol consumption (Fig. 3.2.3. ). whereas right
atrial BNP levels in SHR are sirnilar following the 8 rnonths of either water or ethanol
administration. In lefi atnal tissue. SHR and WKY rats have sigruficantly elevated BNP levels.
both m ternis oftotal content (pg) and concentration (@mg protein) (Fig. 3.2.1.). However.
the contents of BNP mRNA m both atria are unchanged by the ethanol treatment (Fig. 3.2.5. ).
The ethanol treatment prevented the development of ventncular hypertrophy in SHR rats. as
mdicaîed by the Iower total protein content in the ventricles of ethanol- compared to water-
treated SHR animals (Table 3.2.3.). The chronic ethanol consumption also significantly
increased both ventricular BNP concentration (n=6, P=0.05) and ventricular BNP mRNA
(n=6. Ps0.05) in SHR rats (Fig. 3.2.6. and 3 -2.7. ). In contrast, total ventricular BNP content
and concentration and ventricular BNP mRNA were not affected by the ethanol treatment in
normotensive WKY animals (Fig. 3.2.6. and 3-2.7.)-
3.2.44. Ventricular BNP mRNA bv RT-PCR
To confirm the ethanol-induced increase in ventricular BNP mRNA content in the SHR
but not WKY rats, which was demonstrated by Northem blot analysis, the content of BNP
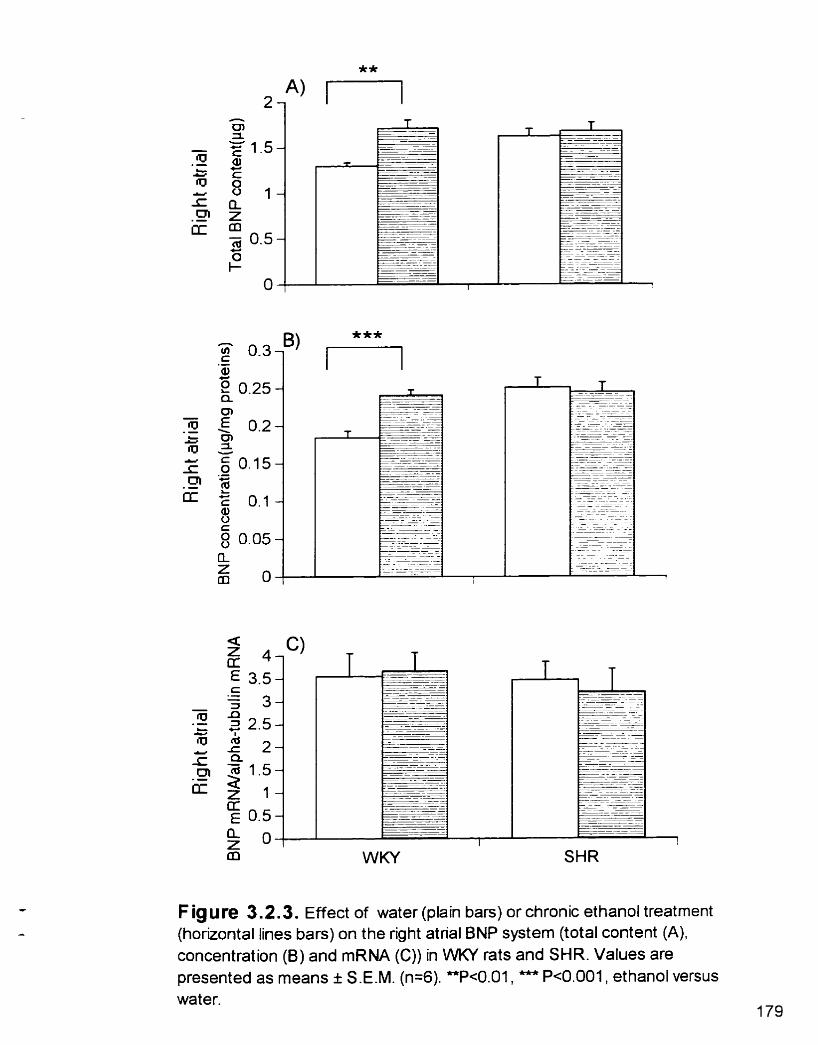
i
SHR
F igufê 3.2.3. Effect of water (plain bars) or chronic ethanol treatment (horizontal lines bars) on the right atrial BNP system (total content (A), concentration (B) and mRNA (C)) in WKY rats and SHR. Values are presented as means r S.E.M. (n=6). *P<0.01, " P<0.001, ethanol versus water.
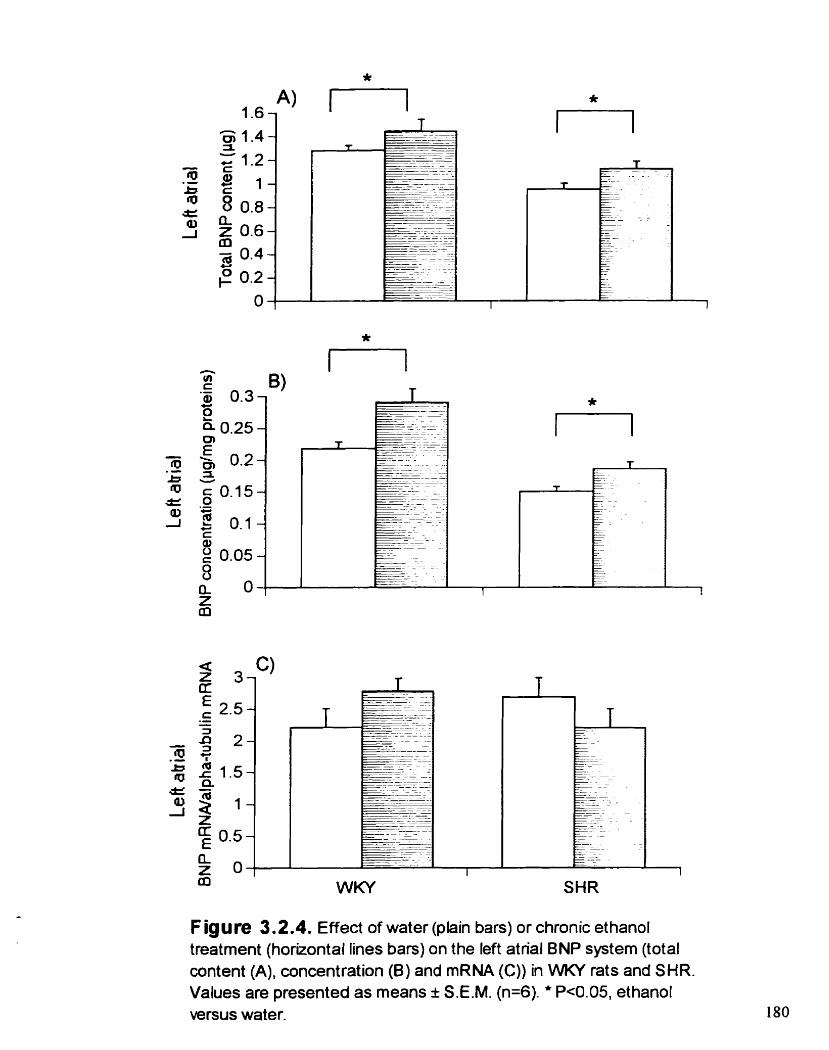
SHR
F igure 3.2.4. Effect of water (plain bars) or chronic ethanol treatment (horizontal Iines ban) on the left atnal BNP system (total content (A), concentration (6) and mRNA (C)) in WKY rats and SHR. Values are presented as means k SEM. (n=6). * Pc0.05, ethanol versus water.
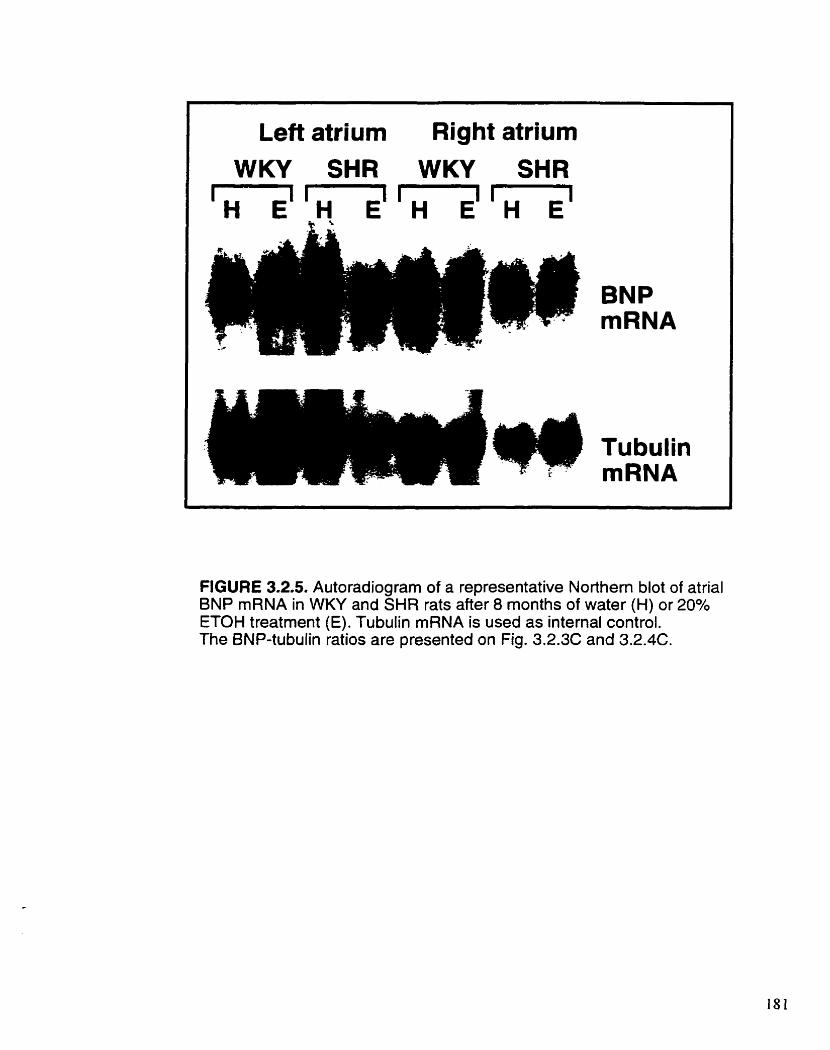
Left atrium Right atrium WKY SHR WKY SHR nnnn H E H E H E H E
BNP mRNA
Tubulin mRNA
FIGURE 3.2.5. Autoradiogram of a representative Northem blot of atrial BNP mRNA in WKY and SHR rats after 8 months of water (H) or 20% ETOH treatment (E). Tubulin mRNA is used as interna1 control. The BNP-tubulin ratios are presented on Fig. 3.2.3C and 3.2.4C.
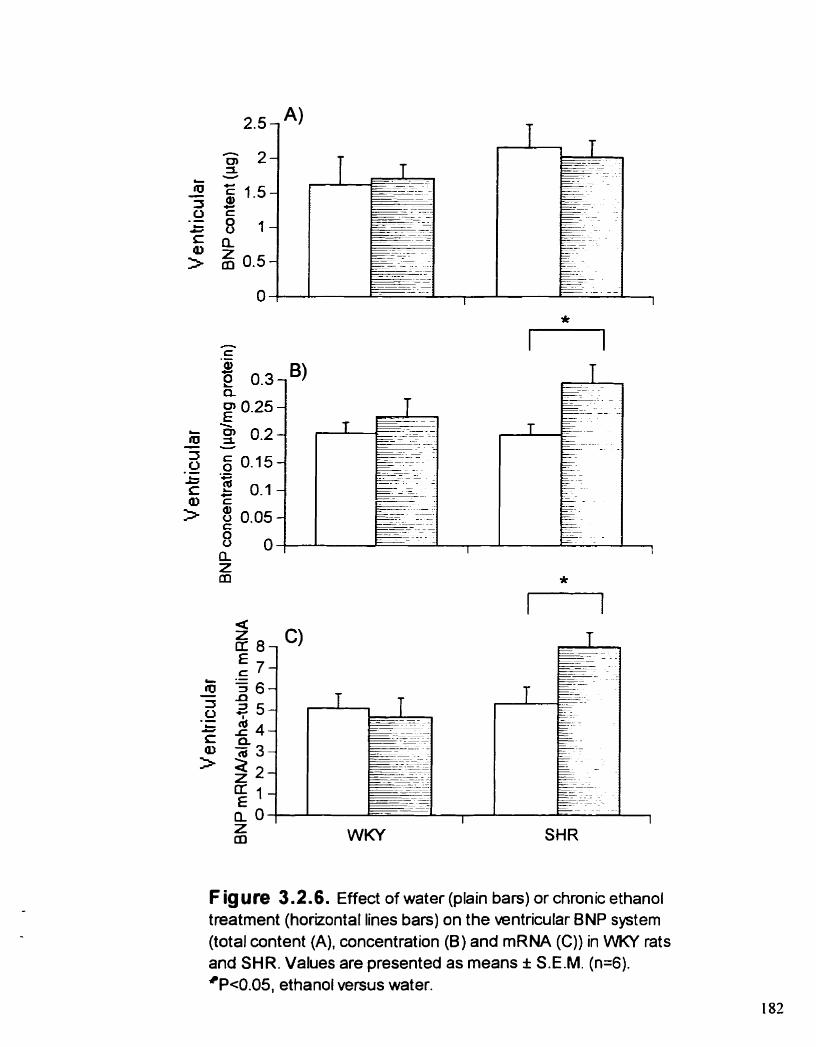
SHR
F igufe 3.2.6. Effect of water (plain bars) or chronic ethanol treatment (horizontal lines bars) on the ventricular BNP system (total content (A), concentration (B) and mRNA (C)) in WKY rats and SHR. Values are presented as means I S.E.M. (n=6). +Pc0.05, ethanol venus water.

WKY SHR
BNP mRNA
Tubulin mRNA
FIGURE 3.2.7. Autoradiogram of a representative Northem blot of ventricular BNP mRNA in WKY and SHR rats after 8 months of water (H) or 20% ETOH treatment (E). Tubulin mRNA is used as intemal control. The BNP-tubulin ratios are presented on Fig. 3.2.6C.

mRNA in the ventricies of water- and ethanol-treated SHR and WKY rats was also estimated
by RT-PCR Results Eom the RT-PCR analysis demonstrated that long-term ethanol
treatment induced a sigdicant hcrease in the BNP mRNA content in the ventricles of SHR
rats, a 9 1% mcrease fkom the SHR-water group (Eg. 3.2.8. ). In agreement with the Nonhem
blot analysis, chronic ethanol treatment failed to mduce a signifïcant mcrease of the BNP
mRNA content m the ventricles of WKY rats.
3.2.5 DISCUSSION
Aithough the hypertensbe effect of the long-term consumption of hi& quantities of
alcohol has been weil documenteci (Riddey et ai.. 1985; Lian, 1915), the effect of chronic low
to moderate ethanol consumption remams controversial (MacMahon, 1987). indeed it has
been previousty reported by Howe et al. (1989) and confirmed by us that chronic
administration of moderate quantities of ethano1 delays or even prevents the age-dependent
increase in blood pressure not only in hypenensive animals, but in nonnotensive animals as
weil.
Since BNP has hypotensive properties, the objective of the present studies was to
mvestigate the effect of chronic moderate ethano1 consumption on the heart BNP system The
present audies demonstrated that chronic ethanol treatment induced a decrease in circulating
BNP levels of both SHR and WKY rats. The low plasma BNP levels are associated with
elevated aaial BNP content and concentration, without any alterations in atrial BNP mRNA.
In the ventricles, the BNP concentration and the BNP mRNA content are s iwcant ly
increased by the ethanol treatment m SHR, but not WKY rats.
Chronic moderate ethanol conçumption significantiy decreased plasma BNP levels and
significantly increased the content and concentration of BNP in the right and lefi atna of
WKY animals and in the lefi atria of SHR rats. It has been demonstrated that BNP is
cosecreted with ANP by the atrial tissue (Thibault et al . 1992; Iida et al.. 1990; Aburaya et
al.. 1989~). Indeed, two types of atnal granules have been reported , type 1 with ANP alone.
and type II with both ANP and BNP (Hasegawa et ai., 199 1). This may explain the similar
variations of ANP (Section 3.1. ) and BNP m the atrial tissue following chronic
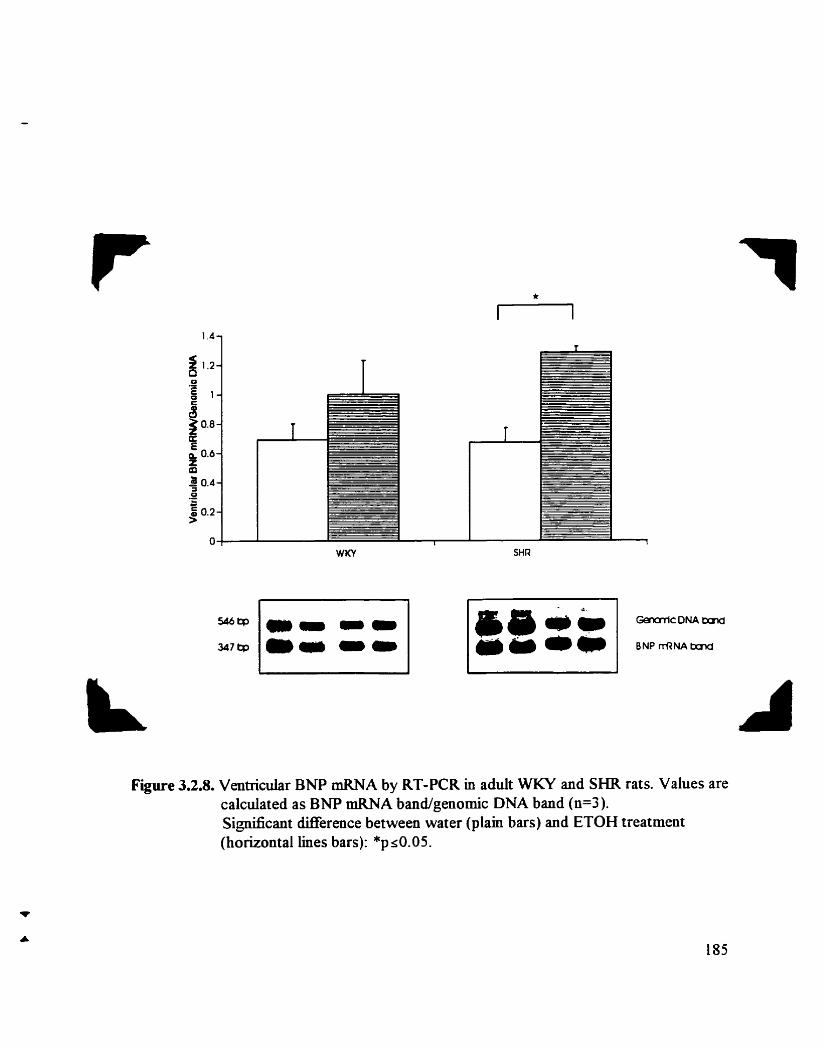
SHR
1 Gencrric DNA Pcnd
BNP M?NA bnd
Figure 3.2.8. Venaicular BNP mRNA by RT-PCR in adult WKY and SHR rats. Values are calculated as BNP mRNA band/genornic DNA band (n=3). Siguficant difference between water (plain bars) and ETOH treatment (horizontal lines bars): *p ~0.05.

ethanol treatment. A differential regulation of lefi and right rat atrial cardiocytes has also been
reported (Marie et al.. 1976). It is therefore possible that ethanol may have a different effect
on the right than lefl atrial cardiocytes. as observed in SHR rats where lefi atnal BNP content
is increased by ethanol while right atrial BNP levels remain unchanged. The reason for this
strain ciifference in right atrial BNP content is unclear. but since numerous genetic and
hormonal deviations f?om normal rats have been observed in SHR anirnals (Kreutz et al.,
1992: Lang et a!.. 198 1 : Nagaoka and Lovenbeg. 1977). it may be possible that the BNP
system and its s e n s i t ~ t y to ethanol are also dif5erent in the hypertensive strain. The lower
circulating BNP levels rnay be secondary to the hemodynamic changes induced by the ethanol
treatment. However, &ce atrial BNP mRNA contents in ethanol-treated rats remained at the
same lwek as those of the water-treated controls. which displayed sigdicantly higher blood
pressure and plasma BNP levels. it may be suggested that the ethanol treatment maintained
the atnal BNP synthesis at the same level as that of the water-treated rats by a different
rnechanism than secondary hemodynamic changes due to reduced blood pressure. This
rnechanism may be either a direct or indirect effect of ethanol. Thus. it seems that in water-
treated controls the BNP a c t ~ t y is stimulated by the higher blood pressure. whereas in
ethanol-treated rats the BNP activity is stimulated by ethanol. It is possible then that when the
blood alcohol content is hi& (immediately afier drinkhg) there is an increase in atnal BNP
release. whereas in the absence or with tow levels of aIcohol, the release of BNP is not
stimuiated and BNP accumulation m the atria occurs. Indeed at the time of sacrifice (between
9:00 to 12:00 a.m), the blood alcohol content was very low for both strains of rats.
In contrast to ANP, in the heart the major site of BNP synthesis and release is the
ventricle (Lang et al.. 1992: Ogawa et a/.. 1991). Therefore. the BNP concentration in
circulation depends more on the ventncular BNP release which is controlled by the blood
pressure and the ventricular load than on atnal distention pressure (Richards et al.. 1993).
Ventncular granules are also practicaiiy nonexistent. forcing BNP to be released in a
constitutive rnanner (Davidson and Struthers. 1994). Moreover. it appears that in
pathophysiological conditions, Wte congestive heart failure or hypertension. a shifl occurs in
the production ofnatriuretic peptides (BNP, but also ANP) fiom atria to ventricles (Lang et

al., 1992). In consequence, the release of BNP (and ANP) fkom the hypertrophied ventricles
of SHR rats is PgniScantly higher than f?om the ventricles of WKY rats (Kohno et ai., 1994;
Yokota et al.. 1993; Kinnunen et ai., 1993; Roy et al., 1992; Kohno et al., 1992a; Yokota
et al., 1990; and this section).
In the present studies, chronic ethanol consumption augmented significantly BNP
concentration and BNP mRNA m the ventricular tissue of Sm but not WKY rats,
suggesting an mcrease in the rate of BNP synthesis by the ventricles. Interestingly, chronic
moderate ethanol connunption prevented ventricular hypertrophy m the SHR rats. Smce
ventricular hypertrophy is produced by high blood pressure and cardiac overload (Kinnunen
et ai., 1993; Kohno et al.. 1992a), the absence of elevated blood pressure in ethanol-treated
SHR rats may have prevented the development of the hypertrophy. Therefore, the elevated
ventricuiar BNP mRNA levels and BNP concentration, but not totaI BNP content, following
chronic alcohol treatment suggeas a direct mmulatory effect of ethanol on the ventricular
BNP n/stem of SHR rats, despite a lower ventricular mass and thus a lower basal ventricular
total BNP content than the water-treated SHR rats. Again, genetic differences between SHR
and WKY rats may explain this variation in ventricdar BNP s e n s i t ~ t y to chronic ethanol
consumption. Although the effects of acute moderate ethanol administration on the BNP
system are dl unhown, a recent study reported a significant mcrease in circulating BNP
levek (with no variations m ANP or heart BNP content and BNP mRNA) follovhg a short-
term (6 weeks) chronic ethanol treatment in Sprague-Dawley rats (Wigle er al., 1993~).
mdicathg an mcreased actiMty of the BNP system as was also suggested in the p resent midies
by the mcreased content of ventricular BNP mRNA in SHR rats. However, even though both
venûicular BNP mRNA and BNP concentration are increased in SHR rats following ethanol
treatment, plasna BNP content is decreased. The reason for the discrepancy between plasma
BNP Iwels and venaicular BNP concentration imd mRNA is unclear. It may be expected that
the lower blood pressure in ethanol-treated rats should have led to a decrease in ventricular
BNP synthesis and release and eventually to lower ventricular BNP mRNA content. inaead.
chronic ethanol treatment prevented the expected decrease in ventricular BNP a c t ~ t y in
WKY rats and even enhanced the potential BNP activity m SHR rats. Therefore, it is possible

that a transient mcrease in plasma BNP levels is produced after each ethanol drink during the
period of elevated blood alcohol content. so that the activity of the ventricular BNP system
is maintained (WKY rats) or even enhanced (SHR rats). Aiternatively. it has been proposed
that BNP mRNA is more unstable than ANP mRNA requiring lager amounts to maintain
basal tissue levels and secretion by the ventncles (Lang et al.. 1992). Thus. one would need
to ver@ (by pulse-chase experiments) whether this increase in ventricular BNP mRNA in
ethanol-treated SHR rats reflects substmtial elevations m BNP synthesis and release.
interestin&. acute and short-tem chronic ethanol experiment s have rep oned significant
increases in plasma BNP and ANP levels (Chapter 2 and Wigle et al.. 19932). In chronic
ethmol experiments, the blood pressure and circulating ANP (Section 3.1.) and BNP levels
are decreased. Nevertheless. the potential biosynthetic a c t ~ t y of the atrial and ventricular
natriuretic BNP system appears to be maintained or even enhanced cornpared to the potential
biosynthetic actMty of the correspondig water-treated controls. despite the decrease in
blood pressure observed following moderate ethanol consumption. This maintenance of the
potential biosynthetic BNP a c t ~ t y in the atria and ventncles of the ethanol-treated rats may
be due to the repetitive stimulatory effects of the daily ethanol consumption. Future audies
estimating plasma BNP levels following acute ethanol administration in rats chronically
exposed to moderate amounts of alcohol or the determination of circulating BNP levels at
various mtervals during the chronic ethanol treatment should provide a better insight on the
exact pattern of changes in the activity of the hean BNP s y a e a its possible correlation with
the changes m the blood pressure and the possible contribution of the natriuretic peptides in
the prevention of the age-dependent hypertension by moderate ethanol consumption.
Conversely. it is possible that these ethanol-induced increases in plasma BNP and ANP levels
following acute and short-term chronic ethanol exposure (Cliapter 2 and Wigle et a/. . 19%)
rnay have no long-term effects on the blood pressure. in such a case. the lower plasma levels
of BNP and ANP in rats treated chronically with ethanol couid represent only a seconda-
response to the ethanol-induced "hypotensivet' effect, which could have been mediated by
other physiological systerns, such as the ethanol-induced changes in lipoproteins (Hojnaclii
et al., 1988).

In sumrnary. the present experiments have demonstrated that chronic ethanol exposure
for 8 months alters significantly the heart %NP levels in both normotensive and hypertensÏve
rats. Circulatmg BNP levels are lowered by ethanol in both strains. Moreover. BNP mRNA
leveis are signrficant~ elevated in the ventricles but not atna of ethanol-treated SHR but not
WKY rats. Further studies should be performed to determine the significance of the ethanol-
induced changes in the a c t ~ t y of the heart BNP system on the prevention of the age-
dependent increase in blood pressure by low to moderate chronic ethanol coosumption.
3.2.6. ACKNOWLEDGEMENTS
The authors wish to express their appreciation to the excellent technical assistance of Ms
Céline Coderre and Nathalie Charron. Many thanks are also due to Mr Ricardo Claudio for
the excellent care of the animals. P. G. is a recipient of a "Fond pour la formation de
chercheurs et l'aide à la recherche (FCAR)" scholarship. This work was also supported by
gants fiom MRC CANADA (MT 10337 and MT 1 1674). by the Canadian Heart and Stroke
Foundation (J.G.) and by the Alcohol Research Program at Douglas Hospital (C.G. )

Section 3.3.
RENAL, ALTERATIONS OF ATRIAL NGTRIURETIC PEPTIDE
RECEPTORS BY CHRONIC MODERATE ETHANOL CONSUMPTION
P. Guillaume, T.V. Dam, C. Gianodakis and J. Gutkowska
Arnerican Journal of Physiology
(Am Physiol272: Rend Physiol. 4 1 : F107-F116, 1997)
Contribution bv CO-authors: Dr. T.V. Dam mtroduced me to the technical aspects of
autoradiography. Dr. C . Gianoulakis and Dr. J. Gutkowska were my CO-supervisors.
Acknowledgements: R Claudio was the animal technician. C. Coderre and N. Charron
perfomed the ANP and cGMP iodinations.

3.3. RENAL ALTERATIONS OF ATRIAL NATRIüRETIC PEPTIDE
RECEPTORS BY CHRONIC MODERATE ETHANOL TREATMENT
3.3.1. ABSTRACT
Previous studies have shown that chronic moderate ethanol (ETOH) consumption
prevents the age-dependent increase in blood pressure. However. the physiological syaems
mediating ETOHs antihypertensive effects are not knom. The objective of the present
studies was to investigate the effects of chronic (8 rnonths) moderate ETOH consumption on
renal natriuretic receptors of hypertensive (SHR) and normotensive (WKY) rats. using
cornpetitive binding assay and autoradiographic techniques. In the renal giomeruli. the
manmal bmdmg capacity ( B a of the heterogeneous ANP receptor population (NPR-A and
NPR-C) was significantly lower in ETOH-treated SHR and WKY rats compared to water-
treated controls. Quantification of receptor subtypes showed that this decrease was pnrnarily
the result of NPR-C down-regdation. The apparent dissociation constant (&) was also
decreased by the ETOH treatment. In the rend papilla. the B,, of the homogeneous receptor
population (NPR-A) was significantly elevated by long-terrn ETOH consumption in both
strains compared to water-treated controls. However. the Y, was unaltered by the ETOH
administration. Thus. ETOH treatment induced specific alteratioos in renal natriuretic
receptors which may play a role in the "protective" effect of moderate ETOH consumption
on the age-dependent increase in blood pressure.
3.3.2. [NTRO DUCTION
Chronic low to moderate ethanol (ETOH) consumption has been reported to prevent or
delay the age-dependent increase in the blood pressure (BP) in hypertensive (SHR) and
normotensive (WKY) rats (Howe et al., 1989 and Section 5 .1 .), and to lower systolic blood
pressure (BP) in h u m s (MacMahon, 1987). However, the mechanisrns mediating this
"beneficial" e f f i of low to moderate ETOH consumption on BP remah unknown. Following
the demonstration of elevated circulating levels of atnal natriuretic peptide (ANP) with acute
moderate ETOH administration. the natriuretic peptide family has been proposed as one of

the possible mediators of this effect (Section 2.1 .). ANP is part of a new farnily of hormonal
peptides implicated m the regdation of water and sodium homeostasis. and in the control of
BP (Gunning and Brenner. 1992). Specificaify. ANP. a 28 ammoacid peptide of cardiac
ori* has been s h o w to decrease BP by vasorelaxation. diuresis and natriuresis (Gunning
and Brenner. 1992). Additional natriuretic peptides have recently been characterized: brain
aatriuretic peptide (BNP) and C-type nafriuretic peptide (CNP) (Gunninp and Brenner. 1992).
ANP and BNP are mainly synthesized and release in the circulation by the hean. whereas
C N P is moaly found in the braio.
The physiological actions of the natriuretic peptides are mediated by 3 specific natriuretic
receptors (Koller and GoeddeL 1992). The ka two natriuretic peptide receptors (NPR-A and
NPR-B) are single transmembrane proteins with a guanylyl cyclase (GC) region in the
intraceiiular domain. producing cGMP as their second messager. ANP preferably binds to
NPR-A. whiie NPR-B appears to be the natural receptor for the C-type natriuretic peptide
(CNP) (Koller and GoeddeL 1992). The third natriuretic receptor. NPR-C. is an homodimer
protein with a very short intraceiluiar domain. Its abundance and its lower specificity to the
various natriuretic peptides prompted scientists to consider this receptor as part of a buffer
system to clear natriuretic hormones from plasma (Maack er al.. 1987). Yet. recent
association of NPR-C with CAMP has raised doubts about its sole function as a clearance
receptor (Anand-Srivastava and Trachte. 1993 ).
The presence of both NPR-A and NPR-C has been demonstrated on the epithelial
membrane of rend glomenrli, whereas only NPR-A was reported in the renal papilla (Manin
et al., 1989: Brown and Zuo. 1992). The presence of both NPR-A and NPR-C receptors in
the kidney confirms its importance m mediating the diuretic. natriuretic and hypotensive effect
of ANP. Furthemore. ETOH. a weak diuretic. is h o w n to alter the fluidity of renal
membranes and to mod* the a c t ~ t y of the Na'K+-ATPase pump in kidney tubules
(Goldstein. 1987). It is therefore possible that chronic ETOH conntmption is also causing
modifications in renal natriuretic receptor characteristics. These alterations rnay explain. at
least m part. the prevention ofthe age-dependent increase in BP by chronic moderate ETOH
conçumption.

The ami of the present studies was to investigate the effect of moderate chronic ETOH
consumption for 8 months on the natriuretic receptors in the renal glomenili and inner
meduila of spontaneously hypertensive (SHR) and Wistar-Kyoto (WKY) rats. using
competitive bmding assays and autoradiographic techniques. In addition. the effects of ETOH
treatment on urine volume. osmolarity and Na' and K* content were measured at the end of
the experimental penod. The duration of the ETOH treatment was based on previous studies
demonstrating the presence of a ~ ~ c a n t antihypenensive effect of ETOH after 6 (Sm)
and 7 (WKY) months of alcohol consumption.
3.3.3. METHODS
3.3.3.1. Animai tre-
48 male SHR and 48 male WKY rars. aged 6 weeks. were used in the present studies
(purchased from Charles River Breeding Laboratories. St-Constant. Québec. Canada).
After a 7 days acclimatization period. the animals from each strain were randomly
assigned to one of the following 3 groups: Grou? 1 : 18 animals from each strain were
given drinking water containing moderate amounts of ETOH (WKY-ETOH and SHR-
ETOH). ETOH was gradually added ro the drinking water. as previously described by
Howe et al. ( 1989). Brie tly . rats received 5 % v/v for 5 days. 10 % vlv for 5 days. 15 %
vlv for 5 days and then 20 % v/v for the remaining of the 8 rnonth experimental period.
During that time. access to pellet chow (Purina. Richmond. VA) and liquid was ad
libitum. Grou? 2: 18 animals from each strain were given free access to food and water
(WKY -H,O and SHR-H,O). . G r o u : To characterize the renal natriuretic receptors of
SHR and WKY rats at the beginning of the study. 12 rats from each strain were sacrified
at 7 weeks of age.
Body weight and blood pressure were measured at the beginning of the study (7
weeks-old) and every month during the 8 months experimental period. The tail-cuff
method was used to record BP.

3.3.3.2.
7% months into the experiment. 6 rats per group were randomly selected to be placed
in metabolic cages. Urine collection was monitored on a 12 hours basis corresponding to
the light and dark cycle of the room. Animals were given free access to ETOH or water
throughout the 3 days of the experiment. while food was given only during the dark cycle
to prevent any contamination of the urine during the light cycle. Urine volumes were
measured directl y and the urine samples collected during the 1 ig ht cycle were subsequentl y
kept frozen for estimation of the Na+/K + concentration and osmolaliry. Na' and K ' were
meassured directl y by an Instrumentation Laboratory autoanalyser. Osmolality was
est imated using 50 pl-frozen samples in an osmorneter (W ide-Range Osmometer.
Advanced Instruments. Needham heights. MA).
3.3.3.3. Ilrinw cGMP excretion
Excreted cGMP levels in the urine samples collected during the light cycle were
determined by radioirnmunoassay as previously described (Tremblay er al.. 1993).
. . 3.3.3.4. ANP rad]-av (RI&
Trunk blood was collected in chilled 15 ml conical centrifuge tubes containing
protease inhibitors to the following final concentrations: 5 mM pepstatin A (P4265. Sigma
chemical CO.. St-Louis. MO), 10 m M phenylmethylsuifonyl tluoride (PMSF) (P7626.
Sigma) and 1 mg/ml ethylened iam ine-tetra-acetate (EDTA). The blood samples were
immediately centrifuged at 4°C and the plasma stored at -75°C for ANP measurements.
Plasma ANP levels were measured by a direct RIA (Gutkowska et al.. 1984). The
validity of the direct assay for the investigation of the relative differences between various
treatments has been previously demonstrated (Section 2. I .).
3.3.3.5.Jllood &oh01 content (BAC)
A 50 pl aliquot of whole blood was placed in 450 pl of ice-cold trichloroacetic acid
(6.25 % w/v, Sigma) for estimation of the BAC at the time of death. using the

dehydrogenase enzymatic method (Hawkins et al.. 1966).
. . . 3.3.3.6. Pre-n of membranes for competitive bind-
After 8 months of treatment. rats were sacrificed by decapitation and their kidneys
were rapidly excised and decapsulated. The kidneys of 12 rats/group were randomly
chosen to be analyzed by competitive binding assay. while the kidneys of the rernaining
6 rats/group were left for autoradiographic studies. The kidneys of 12 young (7 weeks-
old). ETOH-naive animals were also prepared for competitive binding assays. Each kidney
was cut longitudinaily and the inner medulla (IM) was removed and kept frozen ar -75°C
until assayed. The ourer medulla was discarded and the remaining cortical tissue was
minced in a tissue grinder. Subsequently . glomeruli were isolated according to the graded
sieving method of Misra (1972). slightly modified. Brietly. the cortical mesh was
suspended in 0.9% saline and filtered through a 100-pm nylon sieve. Then. the collected
material was passed successively through a 50-pm and a 100-pm sieves. Finally. a last
sieve of 150 -pm was used to collect the glorneruli preparation. This preparation was then
washed in 0.05 M Tris-HCI buffer. pH 7.4 (washing buffer). before being centrifugated
( 180 rpm for 3 minutes) and resuspended in the same buffer. The solution was kept at -
75°C until used.
The glornerular solutions were homogenized with a Polytron (Kinematica GMBH.
Luzern. Switzerland). The homogenates were centrifuged at l5OOOg (4°C) for 30 minutes
and were then washed twice with 0.05 M Tris-HCI buffer. pH 7.4 (washing buffer). The
pellets were then resuspended in 1 ml of the same buffer. Protein concentrations were
measured using a modified version of the Bradford's method. using bovine serurn albumin
(BSA) as standard. Specitïc binding of ANP increased linearly with increasing protein
concentrations between 25pg and 50 pg . Accordingl y. the membrane preparat ions were
diluted to a 35 pg150 pl concentration (well within the linear range) with the competitive
binding assay buffer which consisted of 0.05 M Tris-HCI. pH 7.4. 0.1 % Bacitracine
(Sigma chemical Co., B-0125). 0.5 mM phenylmethylsulfonyl fluoride (PMSF) (Sigma
chemical Co., P-7626). 5 mM MgCl, (Fisher, Fair Lawn. N1. M-33), 0.493 % MnCI,

(Sigma chemical Co.. M-3634). 0.037% EDTA and 0.4% BSA.
Renal inner medullary tissues were homogenized in washing buffer with a Polytron
(Kinematica GMBH. Luzern, Switzerland). Homogenates were centrifuged at 4000g (4°C)
for 70 minutes. Supernatants were subsequently centrifuged at 14000g (4°C) for 45
minutes. Pellers were resuspended in 2 ml of washing buffer. homogenized and centrifuged
again at 14000g (4°C) for 45 minutes. Pellets were finally resuspended in 500 ml of the
sarne buffer and an aliquot was taken for measurements of proteins. As observed for the
glomerular membranes. specific binding of ANP increased linearl y with increasing protein
concentration between 25pg and 50 pg. Aceordingly, the membrane preparations were
diluted to a 30 pg/50 pl concentration with the membrane buffer.
. . . . 3.3.3.7. -ive bind-
The competitive binding assays were carried out according to the method described
by Gutkowska et al. (1988). Binding as a function of time (room temperature) showed
bind ing equil ibrium after I hours. Displacement studies were therefore performed in
duplicates at room temperature for 3 hours with the following components added in
succession on ice: competitive binding assay buffer. standards ( 100 pl). '?-AN P (70000
cpm/50 pl. specific activity: 1000 Ci/mmol) and glomerular or IM membranes (50 pl ).
The standards were ANP,,.,,, ( 10"' M to M) and des-[CiIn"" Seri''. Gly"". Leu"'.
GlylYANPl,,2.,21 (or cANF) ( IO-" M to IO-' M). a ring-deleted ANP analog which binds
specifical l y to NPR-C receptors (Peninsula Laboratories) (Maack et al.. 1987).
Displacement with these unlabelled natriuretic peptides al lowed the d ifferentiation between
the receptor subtypes.
After the 3 hours incubation period. the reactions were stopped by dilution with 3 ml of
cold 0.05 M Tris-HCI buffer and filtered through 1 % polyethylenimine (PEI) (Sigma
chemical Co., P-3 143)-treated Whatman GF/C filters. Filters were washed twice with 3
ml of 0.05 M Tris-HCI buffer. allowed to dry and counted in a gammacounter (Canberra
Packard) .

3.3.3.8. AutorridiograDhi-
The kidneys of 6 ratslgroup were rapidly decapsulated. frozen intact in isopentane
and kept at -75°C until used. 20-pm-thick cryostat sections were prepared and thaw-
mounted ont0 poiy-L-lycine treated slides for autoradiography as described elsewhere
(Brown and Zuo. 1992: Mukaddarn-Daher et al.. 1995). Briefly. kidney sections were
preincubated for 15 minutes at 25°C in 50 m M Tris-HCI buffer, pH 7.4. containing O. 1 %
pol yethylenimine to reduce the non-specific binding of 1'51-ANP. Sections were then
incubated at 25°C with 50 m M HCL buffer. pH 7.4. containing 150 mM NaCI. 10 m M
MgCl?, 40 pglml Bacitracin. 0.5% BSA and 50 pM "'1-ANP. The cornpetitive inhibition
of total l5I-ANP binding to the membranes by unlabelled peptides was examined for each
animal by coincubating with various concentrations of unlabelled cANF ( 10-". IO-' . 10.'
and 10h M) at room ternperature for one hour. The binding observed in the presence of 10-
W was considered the non-specific binding (NSB). The incubation was stopped by
washing the slides in 4 successive dishes containing 50 mM Tris-HCl. pH 7.1 (4°C).
followed by a final dip in water ro wash the salts. Sections were left to dry overnight and
then cxposed for 5 days on Phosphor-sensitive screens. The intensity of the signals were
subsequently quantitated by densitometry using a Phosphor Imager (Molecular Dynamics.
Sunnyvale. CA). Relevant kidney sections were transferred to Maclntosh and processed
there for better quality of the irnaging.
3.3.3.9. Statisîia
For each group. the maximum binding capacities (BmJ and apparent dissociation
constants (K3) were calculated from the cornpetitive binding assays in both glornerul i and
IM using the LIGAND iterative model-fitting computer program. Results are presented
as Mean + SEM. Comparisons between groups were done by two-way ANOVAs with the
strain (WKY or SHR) as the first independent variable and the treatrnent (HLO or ETOH)
or the age (7 weeks-old or 38 weeks-old) as the second independent variable. followed by
the Neurnan-Keuls multiple cornparison test. A p value of 50.05 was considered
significant.

3.3.4. RESULTS
3.3.4.1. BP and ~ l a s m a ANP l e v e l ~
The chronic moderate ETOH consumption si@cantly prevented (m WKY rats) or
delayed (in SHR rats) the age-dependent increase in BP (Table 3.3.1. ). A two-way ANOVA
with the strain and the treatment as the two mdependent variables indicated a significant effect
of the strain (F,,,=307.80, ps0.001) and the treatment (F1,,=49.25, ps0.001), but no
significant strain by treatment mteraction (F1.,,=O. 10, p=0.75) on the BP.
The age-dependent increase in circulating ANP levels was also prevented by the ETOH
treatment m both SHR and WKY rats. Indeed, the plasma ANP levels were 54 and 28%
lower m the ETOH- versus the Hetreated SHR and WKY rats, respectively (Table 3 2.1. ).
At the tMe of sacrifice (between 9:00 and 12:OO AM), the blood alcohol content (BAC) was
28.9 * 12.2 and 39.2 * 18.9 mg/dl in WKY-ETOH and SHR-ETOH, respectively. This low
BAC is explained by the fàct that the animais were sacrificed during the light cycle, when they
are not actively eating and drinking and thus not consurning ETOH.
3.3.4.2. Urine analvsis
On the 36& week of age, urine was collected for 3 consecutive days fkom six animals of
each experimental group. Long-term alcohol drinkmg induced a sigmficant decrease in the
amount of urine coUected (Figure 3 -3.1 A. ). Urine volumes are measured as 8.4 * 0.6 and 4.6
* 0.6 d d a y (n=6, p 50.00 1) in WKY-H,O and WKY-ETOH respectively, and as 6.8 * 0.6
and 4.6 * 0.6 (n=6, poO.05) in SHR-H,O and SHR-ETOH respectively. No si&cant
dinereoce between strains is noted. Urine of chronicaily ETOH-fed animals is also more
concentrated, as demonstrated by the signiticantly elevated osmolarity values (Figure
3.3.1 B. ). Nevertheless, the daily fluid intake and the body weight (Table 3.3.1. ) are not
difTerent between the water- and ETOH-treated rats of both arains, suggeaing no
dehydration or volume depletion m alcohol-fed animals.
Urine sodium concentration is sigdicantly increased in ETOH-treated animals of both
strains (40.2 * 6.4 versus 8 1.1 * 8.3 mM, n=6, p sO.0 1, m WKY rats, and 35.0 5.1 vernis
55.1 k 6.2 mM, n=6, ps0.05, in SHR rats) (Figure 3.3.2AJ. Interestingly. the sodium
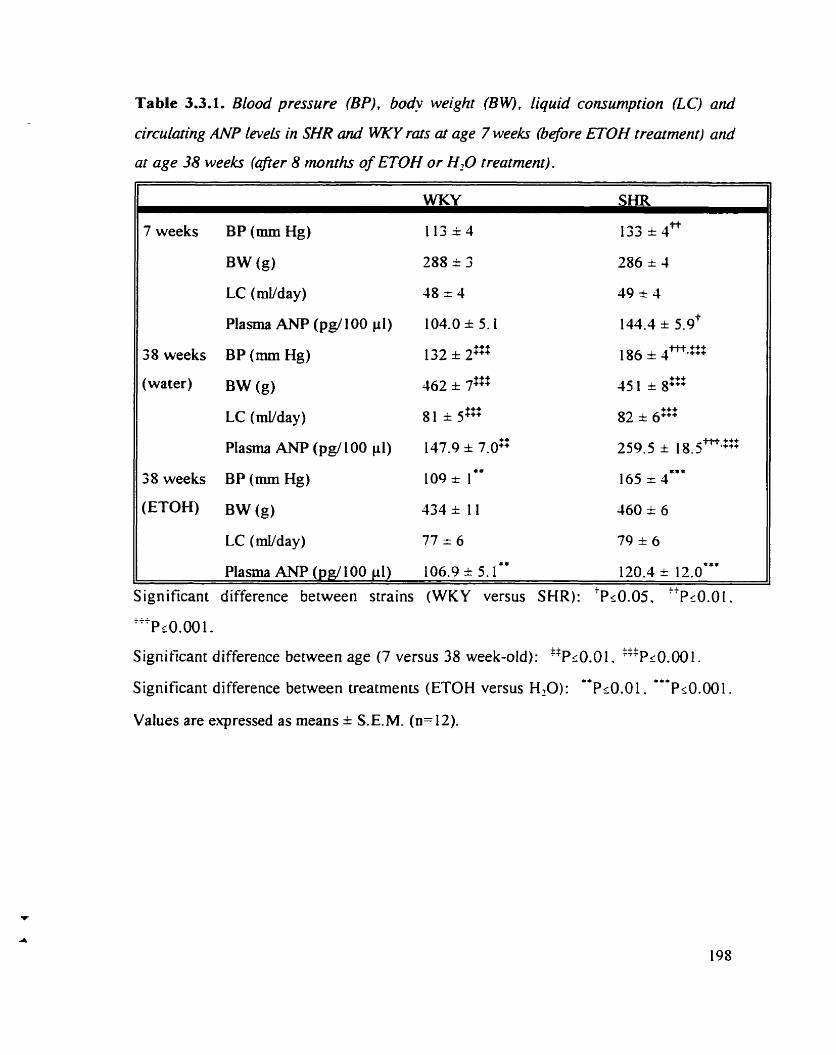
Table 3.3.1. Blood pressure (BP). body weight (B W). liquid consumption (ZC) and
circulnrng ANP levels in SHR and MY nus ut age 7 weekr &@ore ETOH trearmenr) and
at age 38 weeks (Mer 8 m n i h of ETOH or H,O treatment).
7 weeks BP (mm Hg) 1 1 3 2 4 133 * 4" BW (g) 288 i= 3 286 * 4
LC ( d d a y ) 48*4 49 I 4
PlasmaANP(pg/lOOyl) 104.0k5.1 144.4 * 5.9'
38 weeks BP (mm Hg) 132 k 2::j 186 * 4w.$$:
(water) BW(g) 462 k ît:: 45 1 * 8$z$
LC (mVday) 81 * St:: 82 i 6:::
PlasmaANP(pK/lOOpl) 147.9k7.0:: 259.5 k 1 8Srn*:::
Xweeks BP(rnmHg) 109 * 1 * * 165 * 4***
(ETOH) BW(g) 434 5 I I 460 i 6
LC ( d d a y ) 77 i 6 79 i 6
PlasmaANP(pg/lOOy1) 106.9*5.1" 120.4 * 12.0*** . . - -
Significant difference between strains (WKY versus SHR): 'P~0.05. "Pc0.01. . . . --- * * * P50.001.
. . . S ignificant difference between age (7 versus 38 week-old): ' i ~ c ~ . ~ 1. "'Pc0.00 1.
La **a
Significant difference between treatments (ETOH versus H20): Pc0.01. Pc0.001.
Values are expressed as means * S.E. M. (n= 12).
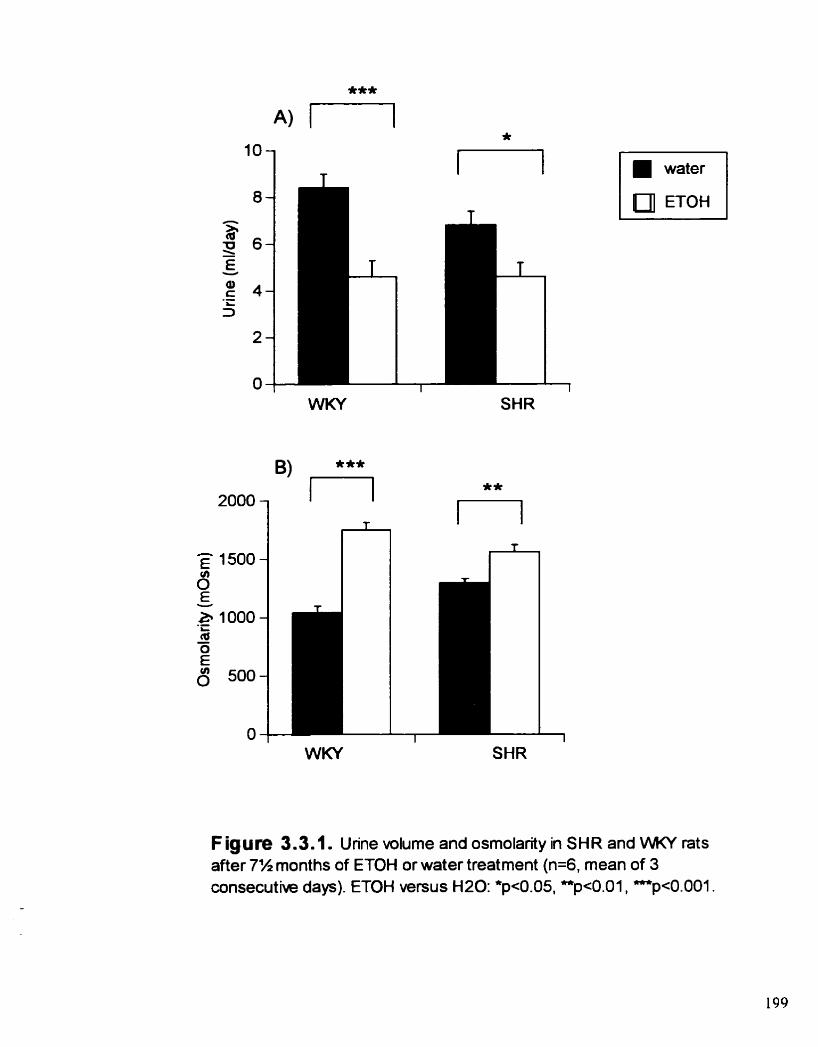
WKY SHR
WKY SHR
1 water 1
F igure 3.3.1. Urine volume and osmolarity in SHR and W rats after 7% months of ETOH or water treatment (n=6, mean of 3 consecutNe days). ETOH versus H20: *p<0.05, pc0 .01 , -p<0.001.

1 water UI] ETOH
I
SHR SHR WKY
T
WKY SHR SHR
F igure 3.3.2. Urine sodium concentration (A), sodium excretion (8). potassium concentration (C) and potassium excretion (D) in SHR and WKY rats after 7Xmonths of ETOH or meter treatment (n 4, mean of 3 consecutive days). ETOH versus H20: P 4.05, 4.01, -p 4.001. WKY versus SHR: mQ.01.

excretion is also increased by ETOH, even with the reduction m total urine volume (Figure
3 -3 -2B.). In contrast, urine potassium concentration is not different between the water and
ETOH groups of the same grain (Figure 3.3.2C.). However, K' concentration is higher in
ETOH-naive SHR (90.6 * 2.3 m . , n=6) compared to ETOH-naive WKY rats (75.1 * 2.6
mM, n=6) (po0.01). Because of the reduced urine volume, the potassium excretion is lower
in ETOH-treated rats (Figure 3.3.2D. ).
3.3.4.3. Comaetitive bindine studies
The density and afEnity of natriuretic receptors are evaluated by cornpetitive inhibition
of "1-ANP binding to the membrane receptors by mcreasing concentrations of unlabelled
ANP or CM.
a) Glomemlur nutriirretic receptors:
The kmetic parameters obtasied from the cornpetitive binding curves are s h o w on Table
3.3.2. To ailow the differentiation between guanylyl cyclase and "clearance" receptors.
IabeUed ANP is displaced by increasing concentrations of cold CM. This displacement
represents between 50 and 90% of the total glomedar natriuretic binding sites in WKY and
SHR rats, so that the majority of the natriuretic receptors in renal glornerular membranes are
of the NPR-C subtype.
Strain effect :
Both young (7 week-old) and adult (38 week-old) ETOH-naive SHR rats exhibit lower
total glomerular natriuretic bmding sites when compared to ETOKnaive WKY rats of the
same age, as observed by the lower displacement of '"1-ANP by unlabelled ANP (Figure
3.3.3A. and C.). The B, for glomerular NPR-C receptors obtained from the inhibition of
'%WP binding by cANF is also sigiuncantly lower m SHR compared to WKY rats of the
same age (Table 3.3.2.). In contrast, the density of the guanylyl cyclase receptor subtype
@PR-A), obtained by subtracting NPR-C receptor number fiom total glomerular binding, is
not Sgiuficantly different between the two strains of rats (Table 3.3.2. ) (Kollenda et aL . 1 990 :
Mukaddam-Daher et al., 1995).
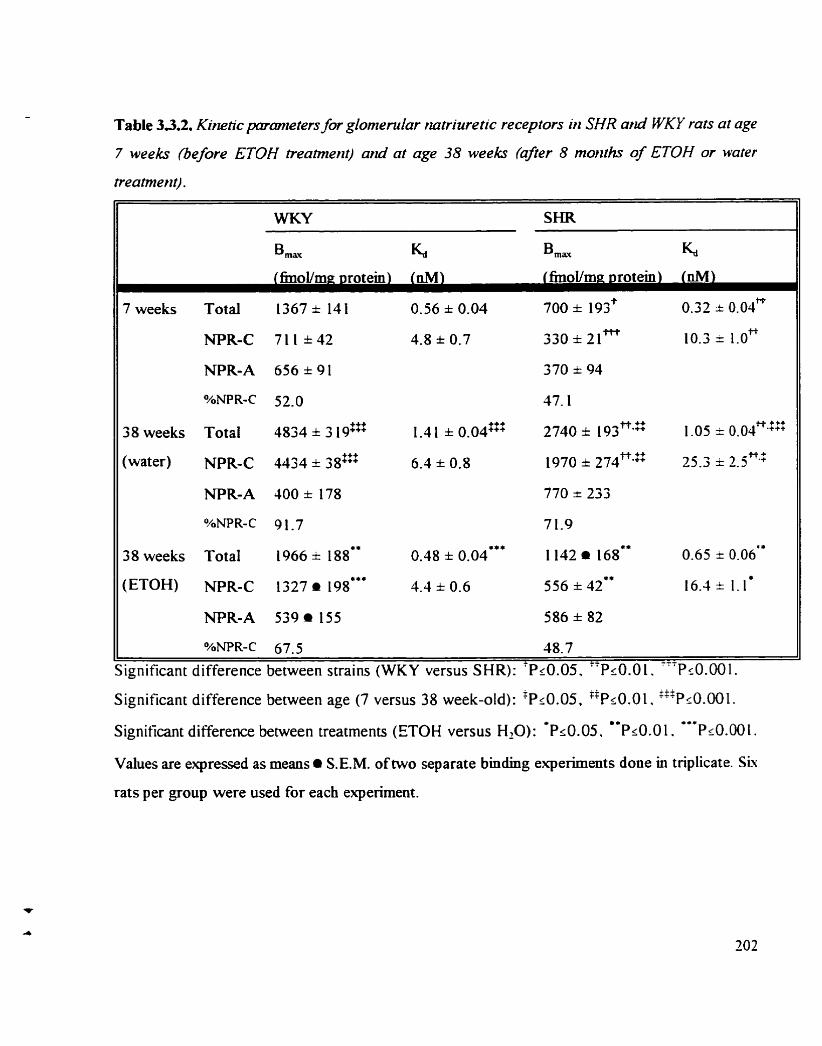
Table 33.2. Kittetic parameters for giornemlor mtriuretic receptors in SHR a d WKY rats ut age
7 weeks @e@e ETOH treatrnent) and ai age 38 w e e b (after 8 month of ETOH or water
treatment) .
WKY SHR
Bm, Bm, Ed
7 weeks Total 1367 * 141 0.56 * 0.04 700 * 193* 0.32 * 0 . 0 4 ~
NPR-C 71 1 * 42 4.8 * 0.7 330 * 21m 10.3 = 1.0"
NPR-A 656*91 370 i 94
%NPR-C 52.0 47.1 +++
38weeks Total 4834*3lgtt: 1.4 1 * 0 . 0 4 ~ ~ ~ 2740 * 193".:: 1 .O5 i 0.04 +++
(water) NPR-C 4434 i 6.4 * 0.8 1970*274w0zz 25.3+2.5"*:
NPR-A 400 * 178 770 s 233
%R-C 91-7 71.9
38weeks Total 1966 * 188" 0.48 * 0.04~" 1142 168" 0.65 r 0.06"
(ETOH) NPR-C 1327 198"' 1.4 * 0.6 556 * 42" 16.3 1 1.1
NPR-A 539 155 586 * 82
%NPR-C 67.5 48.7 ..- ---
ignificant difference between ~trains (WKY versus SHR): 'Pr0.05. "PgO.01. ' "Pc0.001. . . . 7 - Y
Significant difference between age (7 versus 38 week-old): ' ~ i 0 . 0 5 . '*P.-0.0 1. "'PdLOû 1. . iL a+*
Signifiant difference between treatments (ETOH venus H,O): ' ~50 .05 . Pc0.0 1. Ps0.00 1.
Values are expressed as means S.E.M. of two separate bindmg experiments doue in tnplicate. Six
rats per group were used for each experiment.
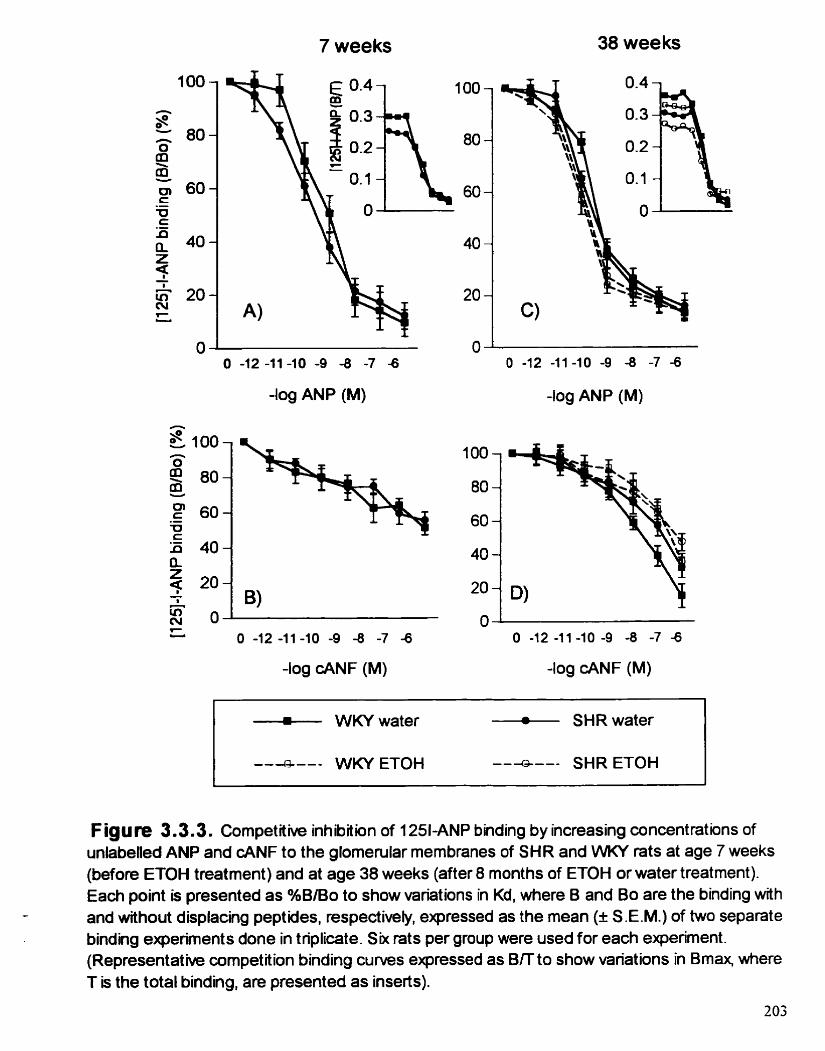
7 weeks 38 wee ks
-log ANP (M) -log ANP (M)
O -12 -11 -10 -9 -8 -7 -6
-log cANF (M) -log cANF (M) - WKY water - SHR water
---I)--- WKY ETOH --a--- SHR ETOH
Figure 3.3.3. Cornpetitive inhibition of 1251-ANP binding by increasing concentrations of unlabelled ANP and cANF to the glomenilar membranes of SHR and W rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (after 8 months of ETOH or water treatment). Each point is presented as %BR30 to show variations in Kd, where B and Bo are the binding with
- and without displacing peptides, respectiveiy, expressed as the mean (I S.E.M.) of two separate binding experiments done in tripkate. Six rats per group were used for each experiment. (Representative competition binding cums expressed as BTT to show variations in Bmax, where T is the total binding, are presented as inserts).
203

The dissociation constant (&) for total glomedar ANP bindmg sites is lower in SHR
compared to WKY rats of the same age. However. the affinity of glornedar NPR-C
receptors is lower m both young and adult SHR compared to age-rnatched WKY rats (Table
3.3.2.).
Age effect:
The total number of gIomedar natriuretic binding sites is increased with age for both
WKY and SHR rats (Table 3.3.2.). This increase is mady due to the augmentation of NPR-C
receptor numbers. The dissociation constant (KJ for the giomedar ANP binding sites is also
increased signifïcantly fiom the 7- to the 38-week-old animals of both strains.
ETOH eEect:
The displacement of ' 3 ~ - ~ by unlabelled ANP produces a significant reduction in the
total glomerular natriuretic bmding sites in chronically ETOH-treated animals of both strains
compared to controls (Figure 3 . 3 . K . ) . Similarly. the displacernent of labelled ANP by cold
cANF produces a sipificant decrease in the glomerular NPR-C receptor binding sites in
ETOH- compared to H20-treated rats (Figure 3.3.3D.). This result mggens a sensitive
discrimination between the two populations of receptors with chroaic ETOH treatment.
Although more specinc agonins of NPR-A are needed. the direct extrapolation of NPR-A
receptor numbers fiom the calculated glomemlar total ANP binding sites minus the NPR-C
receptor density (Kolienda et ai.. 1990: Muhddam-Daher et al., 1995 ) suggests a sipificant
decrease of clearance receptors. d o u t any significant variation in NPR-A receptor numbers
(Table 3.3.2.). Indeed the long-tem ETOH administration decreases the proportion of NPR-
C in glomedar membranes. as calculated by displacement studies with cold ANP and cold
cANF (fiom 71.9 to 48.7% and fiom 9 1.7 to 67.5% oftotal receptors number in SHR and
WKY rats. respectively ).
The dissociation constant (4) for the total glomerular natriuretic receptors and for the
NPR-C receptor bindiig sites are decreased with ETOH in both strains of animals (Table
3.3.2.).

6) Irrrrer meduifury (IM) mttriuretzc receptors:
The hetic parameters obtained fiom the cornpetitive bmdiog curves are shown on Table
3 -3.3. The rend papilla is beiieved to possess mostly NPR-A receptors. Nevertheless. the
possible presence of other natriuretic receptors is investigated in the present experiments by
the displacement of '"I-ANP with cold CANE As expected. mcreashg concentrations of
unlabeued CAM? are not progressively inhibithg the '"1-ANP binding. except at its highest
concentration ( IOn M) where it displaced the labelled peptide by ody 14?h (WKY) and 6%
(SHR). dernonaratkg the absence of NPR-C receptors (figure not show).
Strain effect:
in contraa to the glomedar natriuretic receptors, both young (7 week-old) and adult
(38 week-old) ETOH-naive SHR rats exhibit higher total b e r medullary ANP receptors
compared to age-rnatched ETOH-naive WKY aaimals. as demonstrated by the grearer
disp lacement of "'EANP by unlabelled ANP (Figure 3.3 -4 and Table 3.3.3. ).
The dissociation constant (b) for the total papillary ANP binding sites is not different
between the SHR and WKY rats of the same age (Table 3 - 3 3 . ).
Age effect:
The total inner medullary natriuretic receptors are decreased with age in both strauts of
rats (Table 3.3.3. ). However. the dissociation constant (KJ is unaltered between the 7- and
the 38-week-old SKR and WKY rats.
ETOH effect:
The displacement of "'1-ANP by unlabelled ANP demonstrates a significant
augmentation in the total b e r meddary ANP binding sites in chronkally ETOH-treated
animals of both strains compared to controls (Figure 3 34B. and Table 3 -3.3. ).
The a E t y for the total papillary natriuretic receptors is not altered by the ETOH
treatment in both SHR and WKY rats (Table 3.3.3.) .
3.3.4.4. Autoradio~ra
To furtber illustrate the differential regulation of the glomedar ANP receptor subtypes
by ETOH admhistration in adult SHR and WKY rats, the autoradiography of whole liidney
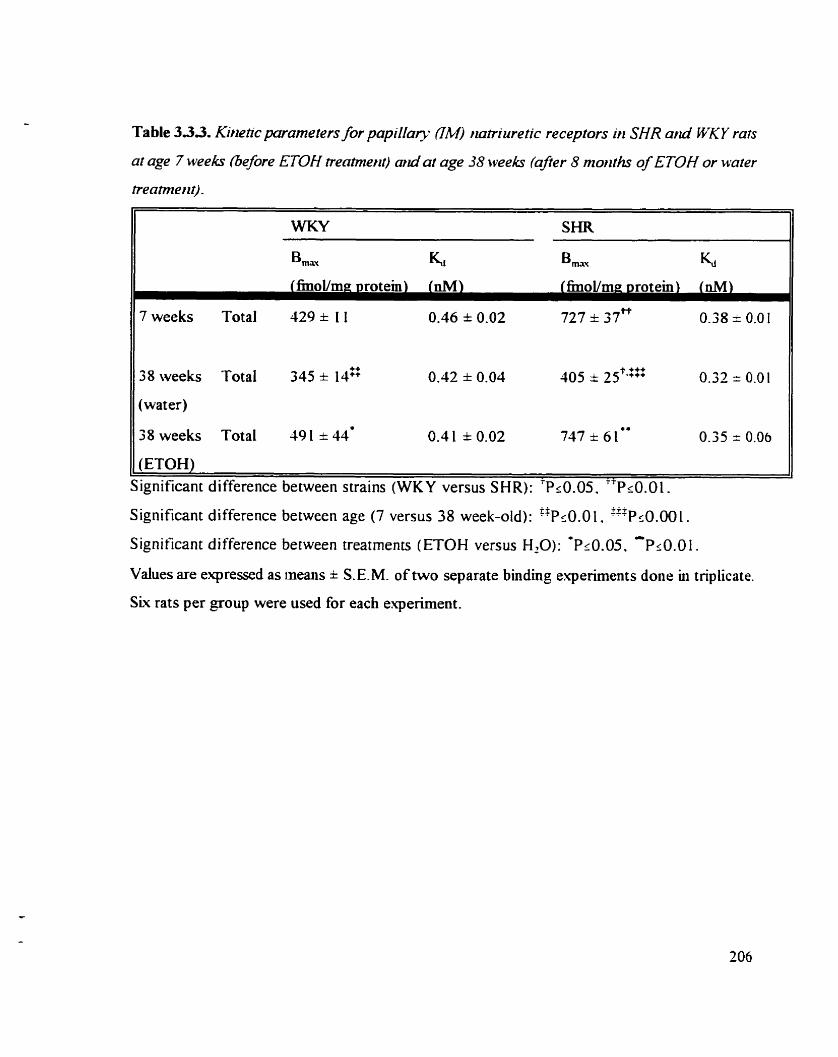
Table 3 3 3 . Kimtic praameters for papillary rzatriuretic receptors irt SHR arid WK Y rats
at age 7 weekr fiefore ETOH trentmertt) midut age 38 iveeh m e r 8 rnor~thr of ETOH or water
II SHR
7 weeks Total 429 * L 1 0.46 * 0.02 727 * 37" 0.38 = 0.0 1
38 weeks Total 345 * 14:: 0.42 * 0.04 105 * 25':tt 0.32 = 0.0 I
(water)
38 weeks Total 49 1 * 44* 0.4 1 * 0.02 717 i. 61" 0.35 - 0.06 11 (ETOH) Significant difference between strains (WKY versus SHR): '~~0.05. "~<0.01.
Significant difference between age (7 versus 38 week-old): ''~~0.01. " i ~ c O . O O 1 . - Significant difference between treatments (ETOH versus H,O): 'Pr0.05. P50.01.
Vahes are expressed as means * S.E.M. of two separate binding experiments done in tnplicate.
Six rats per group were used for each experiment.
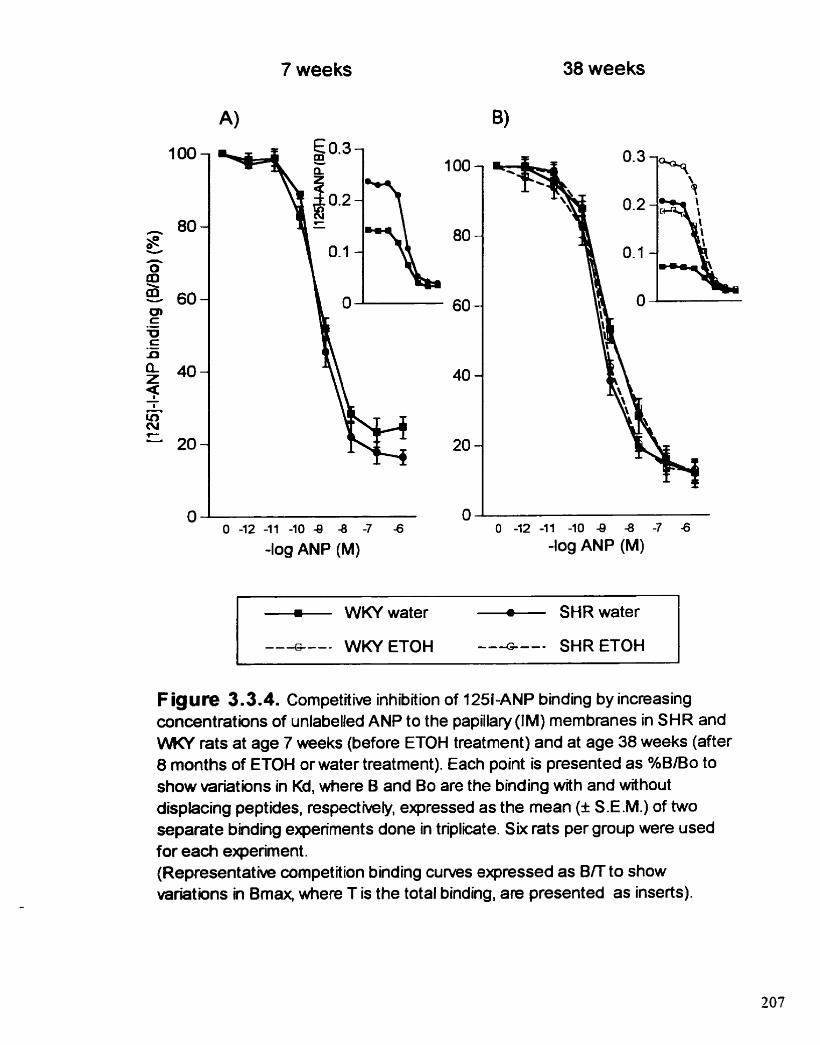
7 weeks
A)
38 weeks
-log ANP (M) -log ANP (M)
- SHR water
--+--- WKY ETOH --a--- SHR ETOH
F ig urê 3 3.4. Cornpetitive inhibition of 1251-ANP binding by increasing concentrations of unlabelled ANP to the papillary (lM) membranes in SHR and W rats at age 7 weeks (before ETOH treatment) and at age 38 weeks (after 8 rnonths of ETOH or water treatment). Each point is presented as %B/Bo to show variations in Kd, where B and Bo are the binding with and wÏthout dispbcing peptides, respectkiy, expressed as the mean (i S.E.M.) of two separate binding experiments done in triplicate. Six rats per group were used for each experiment. (Representative cornpetlion binding cuwes expressed as B K to show variations in Brnax, where T is the total binding, are presented as inserts).

sections ("'1-ANP) displaced by unlabelled cANF is also perfomed.
The displacement of '151-ANP by cold cANF is shown on Figure 3 - 3 5 Because this
synthetic ANP analog binds specifically to NPR-C. it ailows the differentiation between the
"clearance" and guanylyl cyciaç- receptors. 80 to 90% of the total cortical bmding is inhibled
by IO4 M CM, coafirming that the majority of the namuretic receptors in the cortical tissue
is of the NPR-C abtype. In contrast, total inner meddary bindmg is ody partially inhibited
by the highest concentration of unlabelled cANF ( 1on M). suggesting the preponderance of
guanylyl cyclase receptors in the [M (Figure 3.3.5E. and F. ). L
ETOH effect:
Quantitative densitometric analysis shows a lower displacement of total glomerular '"I-
ANP binding by unlabelled cANF ( l ~ - ' and I O - ~ M) m ETOH-treated SHR and WKY rats than
in H@-treated controls (Figure 3.3.5. and 3.3.6A.). This result suggests that in the ETOH-
treated rats. the relative proportion of NPR-C receptors is lower than in the water-treated
controls. supporting the conclusion of the radioreceptor audies.
Although the s e n s i t ~ t y of the autoradiographic technique does not ailow the
visualization of ETOH-induced changes in the total inner medullary (LM) ANP receptors
(Figure 3.3.6C. ). a lower displacement of total outer meduiiary (OM) "'1-ANP binding by
cold cANF is observed for both grains of ETOH- compared to water-treated rats (Figure
3.3.6B.).
3.3.4.5. Urinarv cGMP excretion
in order to link the glomeruiar and papillary ETOKinduced alterations in renal
natriuretic receptors with the possibility of elevated renal effects of the natnuretic peptides.
the urinary excretion of cGMP is examined following 7% months of water or ETOH
treatment.
Chronic ETOH consurnption is associated with a significant increase (in SHR rats) and
with no signifiant change (in WKY rats) in the daily excretion of cGMP when compared to
water-treated controls, despite the lower plasma ANP levels in ETOH- compared to water-
treated animals (Figure 3.3.7.). This result suggests that the ETOH-induced glomerular and
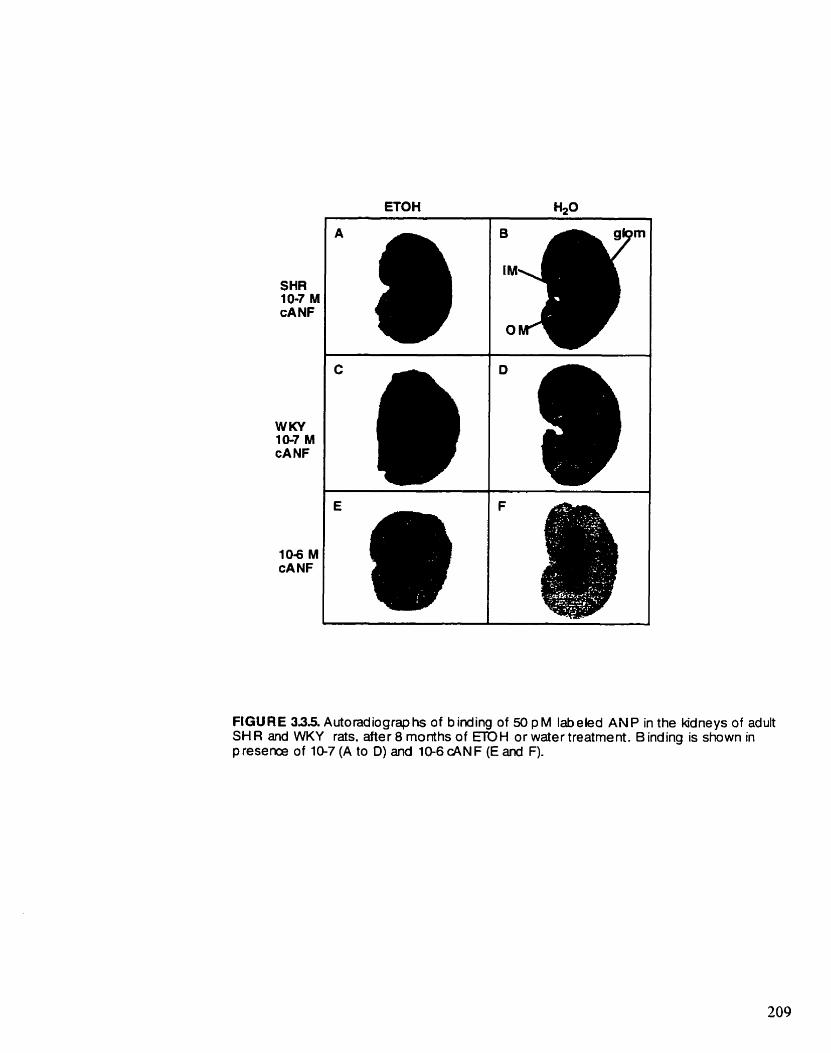
SHR 10-7 M cANF
WKY 10-7 M cANF
106 M cANF
FIGURE 33.5. Autoradiograp hs of b M i n g of 50 p M labeled ANP in the kidneys of adult SHR and WKY rats, after 8 months of €iDH or water treatment. B inding is shown in presence of 10-7 (A to D) and 10-6 cANF (E and F).

1 O water ETOH 1
SHR
F igure 3.3.6. Quantification by densitometry of the displacement of total 1251- ANP binding by 10-7 M unlabelled cANF in the kidneys of adutt SHR and WKY rats, after 8 months of ETOH or water treatment (n=6). Resutts are expressed as: % of total 1251-ANP binding, in (A) glornenili, (B) outer medulla (OM), and (C) inner medulla (IM). ETOH versus H20: p<O.Ol , "p<0.001.
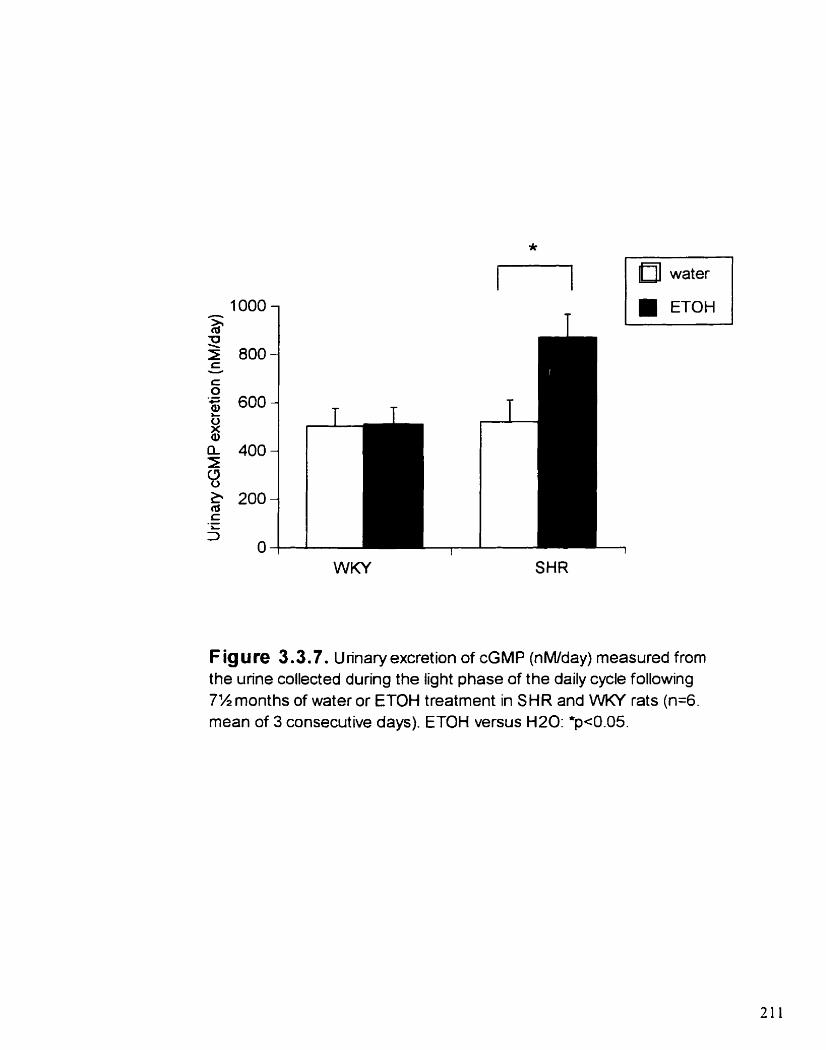
SHR
a water
ETOH
F ig ure 3.3.7. Unnary excretion of cGMP (nMfday) measured from the urine collected during the light phase of the daily cycle following 7% months of water or ETOH treatment in SHR and WKY rats (n=6. mean of 3 consecutive days). ETOH versus H20: *pc0.05.

papillary modifications in renal natriuretic receptors may produce potentially greater cGMP-
mediated renal effects. demonsrrating a fùnctional significance of these specific receptor
alterations.
3.3.5. DISCUSSION
The present experiments have: (a) demonstrated that chronic moderate ETOH
consumption &ers rend ANP receptors. mducing a decrease in total glomenilar ANP binding
sites (mostly NPR-C receptors) and an mcrease m total papillary ANP receptor density in both
SHR and WKY rats; (b) s h o w a lower number of total glomeruiar ANP bindmg sites and a
higher number of total papilIary ANP receptor bmding sites in both young (7 weeks-old) and
aduh (38 weeks-old) SHR compared to WKY animals: and (c) characterized the presence of
an heterogeneous ANP receptor population (NPR-A and NPR-C) in rat renal glomemlar
membranes and of a . homogeneous natriuretic receptor population (NPR-A) in the rat renal
papilla.
Alcoliol has two very difEerent effects (acute and chronic) on the body. Acute
administration of ETOH mcreases the disorder (fluidking effect) of the renal lipid membranes.
and is known to inhibit the a c t ~ t y of the Na'K'ATPase purnp throughout the nephron. thus
affecting the reabsorption of Na' (Rothman et al.. 1992). Water and salt excretion is generally
increased (Wadstein and ohlin. 1979). As tolerance develops. the membranes nSen
(Goldstein, 1987). the activity ofthe Na'K'ATPase enzymes is increased. and Na* and water
retention may occur duruig chroaic alcoholism (Ray et al.. 1992).
The present audies showed that chronic moderate ETOH consumption significantly
prevented the age-dependent increase of the BP in both WKY and SHR rats. This is in
agreement with earlier reports of an hypotensive effect of chronic low and moderate ETOH
administration (Howe er al.. 1989). Dehydration of the ETOH-treated animals is unliliely. as
indicated by the absence of a body weight or fluid intake modïcation fiom water-treated
controls.

a) Glomenr lar natrizïretzc receptors:
The present snidies confbmed earlier reports demonstrating the presence of NPR-A and
NPR-C receptors m renal glomedar membranes. with the "clearance" receptor in excess
amounts (50 to 80% of the total number) (Martin et al.. 1989: Brown and Zuo. 1992). The
two d i h a glomerular ANP receptor populations are mainiy located on the epithelial cells.
with smaller aumbers on the renal microvessels and mesangial cells (Bianchi er ai.. 1986:
Chansel er al., 1990).
Age and strain effects:
Decreased total glornerular ANP binding sites have been demonstrated in various models
of adult hypertensive aniu.uk. such as in the SHR (Ogura er ai.. 1989). the lKlC rat
(Bonhomme and Garcia. 1993). the Lyon hypertensive rat (Gauquelin et al. . 1 993 ) or m the
DOCA-salt treated rat (Nugiozeh et al.. 1990). The present report has extended this result
for young (7 week-old) SHR rats. suggesting the existence of lower total glomerular ANP
receptors during the pre-hypertens~e stage. This strain-induced modification in renal ANP
receptors may be secondary to the elevated circulatmg ANP levels noticed in 7 week-old SHR
compared to 7 week-old WKY rats.
ETOH effect:
In the present experiments. results have s h o w a significant decrease in total glornemlar
natriuretic binding sites in ETOH- compared to water-treated animals of both strains.
calculated fiorn the displacement of '"1-ANP by unlabelled ANP. Furthemore. the
displacement of IabeUed ANP by cold cANF lias suggested that this decrease may be due to
the dom-replation of NPR-C only. A differential regulation of glomedar ANP receptor
subtypes by extemal agents has already been described. indeed. salt load was reponed to
decrease significantly glomerular NPR-C receptors without altering the density of NPR-A
receptors. allowing a greater glomenilar effect of circulating ANP levels in order to remove
the excess salts (Kollenda er al.. 1 990; Fraenkel et ai.. 1994). Similady. the observed
differential regulation ofthe various classes of glomerular ANP receptors by ETOH may have
a funaional significance since a decrease m NPR-C receptors only would mean that a geater
proportion of circulatiag ANP b e l s would bind to the active receptors (NPR-A). Therefore.

the increased proportion of NPR-A receptors in the glomenili of ETOH-treated rats may
increase the glornerular effect of circulating natriuretic peptides. nich as diuresis and
natriuresis. so that it rnay represent one of the mechanisms for the beneficial effect of
moderate ETOH consumption on BP. It is also possîile that this reduction in glomerular
NPR-C binding sites by ETOH is secondary to the decrease in cüculating ANP levels
observed after chronic moderate ETOH consumption in both SHR and WKY rats. increasing
the relative NPR-A a c t ~ t y . To c o h these hypotheses. the exact role of the so-called
"clearance" receptor needs fùrther clarification ( Anand-Srivaçtava and Trachte. 1993). Until
it is h o w n whether the NPR-C receptor has additional fùnctions than ANP buffering. we may
onty speculate on the present dmease of total glomerular ANP bmding sites in ETOH-treated
SKR and WKY animals.
6) lmer medziliary mtrizrretzc receptors:
The present studies confirmed earlier reports dernonarathg the presence of NPR-A
receptors in renal inner medullary tissue (Bianchi er al.. 1987). The IM tissue preparations
used in the present experiments did not allow the distinction between the various ce11 types
in the papilla. Previous studies have dernonstrated that NPR-A alone is present on inner
medda collecting ducts (IMCD) (Bianchi er ai.. 1987) and interstitial cells (Fontoura et al..
1990). Thin loops of Hede and IM capillaries have also been shown to possess natnuretic
peptide receptors (Bianchi et al.. 1987). Furthemore. natnuretic peptide receptors are also
present in the outer medulla. mainly in the vasa recta (Bianchi er ai.. 1987). These papillary
natriuretic peptide receptors have been reported to influence Na* handling and medullary
hypenonicity ( C 3 g and Brenner. 1 992).
Age and strain effects;
Various animal models of hypertension possess higher numbers of natriuretic receptors
m the medulla (Nuglozeh et ai., 1990: Bonhomme and Garcia. 1993). Nugiozeh et al. ( 1990)
reported a significant increase in the density of ANP receptors in the papilla of DOCA-salt
rats, with similar a f i t y . Similarly, Bonhomme and Garcia ( 1993) found a modest
augmentation m the deisity of papillary ANP receptors in l K 1 C rats. Again. the affinity was

mchanged. The present experiments extended this fmding to young (7 week-old) SHR rats.
Several reasons may explain the effect of high BP on papillary ANP receptors. It may be
compensatory to the mcreased BP. The increased number of inner medullary NPR-A
receptors in SHR rats may also be caused by the hypertrophy of the kidneys (Nuglozeh et al. . 1990). However. values calculated as hoVrng protein and not only as hoVpapiUa d l
showed sipifkant elevations in the B,,. Similarly. the assumption that increased plasma
ANP levels must be associated with renal hypertrophy and therefore with mcreased B,, in
the IM is somewhat misleadmg. considering that lower densities of NPR-A were found in the
papilla of rats with congestive heart failure (CHF), a pathophysiological condition where
circulating ANP levels are also signtficantly increased (Yechieli et ai.. 1993 ).
ETOH effect:
The present studies reported increased total papillary ANP bmding sites afier long-tenn
ETOH administration m both WKY and SHR rats. Such an elwatioa depending on the actual
receptor occupancy and renal level of natriuretic peptides. may resdt in enhanced natriuresis
of tlie ETOH-fed rats. Interestingly. the present experiments have shown increased Na-
excretion in ETOH-treated animals. With chronic ETOH consurnption. these alterations in
natriuresis and papillary NPR-A receptors may explain. at least in part. the reduced BP of
ETOH-treated rats. The weak antidiuresis associated with the natnuretic effect of chronic
moderate ETOH connimption is puzzling, although previous experiments have demonstrat ed
the possibility of a dissociation between water and salt excretion followuig both ETOH or
ANP administration (Wadstein and ohlin. 1979: Singer et al., 1987).
Some reports have suggeaed that the hypotensive and natriuretic effects of circulating
ANP levels rnay be independent i?om papuary receptor interactions. because of the necessity
for natriuretic peptides to cross the endopeptidase 24.1 1 "barrier" (main enzyme for ANP
metabolism) of the proximal tubules in order to reach the luminal-situated receptors of the
medda (Goetz. 1991; Gunning and Brenner, 1992). Despite this. tlie discovery of an
intrarenal mernber of the natriuretic peptide family. the NH1-elongated Urodilatin (UD)
(Goetr 199 1 ), synthesized in the diaal tubules, released on the luminal side and binciing to
the same receptors as ANP with equal or greater afnnity (GumÎng and B r e ~ e r . 1992). has

open again the discussion for a role of inner meduilary NPR-A. not only in subtle Na'
variation. but also for BP control. Furthemore. ANP and ANP mRNA are present m the
glomeruli, distal tubules and cortical collecting ducts of rat kidneys (Gunning and Brenner.
1992). Interestingly. CNP and NPR-B mRNAs are also found throughout the nephron
(Terada et al.. 1994). although no expression of these mRNAs have been detected thus far
in the kidney (Mukaddam- Daher et al.. 1995 ). Therefore. the possibility of intrarenal
paracrine effects on papiliary ANP receptors. by UD. by ANP or even by CNP. suggen a
contniution of the IM and its natriuretic receptors for Na' and BP regulation during long-
terni moderate ETOH consumption. Howwer. one would need to determine whether chronic
alcohol treatment rnay increase the synthesis and expression of natriuretic peptides of renal
ongin.
c) U r i i z a ~ cGMP excretiot>
The urinary Ievels of cGMP have been examined in both SHR and WKY rats in order to
link the observed natriuretic receptor modifications of glornenilar and papillary membranes
witb the second messenger syaem of the natriuretic peptides and thus with the possibility of
chronic renal functiooal aherations. nich as mcreased natriuretic a c t ~ t y . that rnay rnodify BP
regdation. It is noteworthy that the cGMP levels are measured from urine s a q l e s collected
d u h g the light phase of the dady cycle. at a time when the animals are not active and are not
consurning ethanol. Accordingly. the blood alcohol levels (BAC) are relatively low and
plasma ANP levels are reduced in ethanol- compared to water-treated animals because of
secondary hemodynarnic effects associated with the lower blood pressure. Nevertheless.
excretory cGMP levels are elevated in ETOH-treated SHR rats whereas they remained
unchanged in ETOH-treated WKY rats. despite their lower plasma ANP levels. This result
directly supports the hypothesis that the natriuretic receptors modifications produced by
ETOH mcrease the rend effects of the natriuretic peptides and may possibly coninbute to the
antihypertensive effect of long-term moderate alcohol consumption.
in addition to the natriuretic receptors. long-term alcohol treatrnent rnay also alter the

activity of other systems implicated in the regdation of water and electrolytes. i.e. AVP or
the renin-angiotensin-aldosterone system. Angiotensin II receptors for example are present
on mesangial membranes of the g l o m d (Sexton et al.. 1992) and possible modifications in
th& deoZty or characteristics by long-term ETOH administration may be responsiile. at least
in part. for the ETOH-mediated modifications in BP.
In summary, the present midies have dernonstrated alterations of renal ANP receptors
with chrooic ETOH treatment and suggested a possible role for the renal natriuretic receptors
m the long-tenn effects of moderate alcohol conaimption on Na' handling and B P regdation.
Specificaliy, increased density of NPR-A in the papilla and decreased quantity of ANP
receptors (mostiy NPR-C) in the glomeruli with chronic moderate ETOH were found by
radioreceptor and autoradiographic studies. for both SHR and WKY rats. Whether these
alterations are responsible for the protective effect of long-term moderate ETOH
consurnption on BP remains to be confirmed.
3.3.6. ACKNOWLEDGEMENTS
The authors wish to e.xpress their gratitude to Mr Ricardo Claudio for his excellent worli
m the handling of the animals. Special thmks are also addressed to Mrs Céline Coderre and
Ms Nathalie Charron for their techoical assistance. P. G. is a recipient of a scholarship from
"Fonds pour la formation de Chercheurs et l'Aide a la Recherche" (FCAR). This work was
supported by gants fiom Canadian medical research council (MT- 10337 and MT- 1 1671).
the Heart and Stroke Foundation. the Canadian Kidney Foundation (J.G.) and the Alcohol
research program at Douglas hospital ( C G . ).

Section 3.4.
ALTERATIONS IN BRAIN LEVELS OF ATRIAL AND C-TYPE
NATRIURETIC PEPTIDES AFTER CHRONIC MODERATE
ETHANOL CONSUMPTION IN SPONTANEOUSLY HYPERTENSIVE
RATS
P. Guillaume, J. Gutkowska and C. Gianoulakis
European Journal of Pharmacology
(Eur Pharmacol, in press, 1997)
Contribution bv CO-authors: Dr. I. Gutkowska and Dr. C. Gianoulakis were my CO-
sup exvisors.
Acknowled~ements: R Claudio was the animal technician. C. Coderre and N. Charron
performed the ANP and CNP iodinations.

3.4. ALTERATIONS lN BRAIN LEVELS OF ATRIAL AND C-TYPE
NATRIURETIC PEPTIDES AFTER CHRONIC MODERATE
ETHANOL CONSUMPTION iN SPONTANEOUSLY
BkTERTENSIVE RATS
3.4.1. ABSTRACT
Atrial (ANP) and C-type (CNP) natriuretic peptides have been found in brain regions
associated with fluid homeostasis and blood pressure. Since chronic moderate ethanol
consumption has been show to prevent the age-dependent increase in blood pressure in
experimental animals the objective of the present studies was to investigate the effect of
ethano1 (20 O/O v h for 8 months) on the total content and concentration of ANP and CNP in
the brain of spontaneously hypertensive (SHR) and Wistar-Kyoto (WKY) rats. Ethanol
mcreased the content and concentration ofboth AM> and CNP m the hypothalamus. pons and
medulla of SHR rats. in contrast. in the WKY rats ethanol had no effect on the levels of ANP
in any of the brain regions studied. but enhanced the concentration of CNP in the
hypothalamus and medulla. Thus. ethanol induced changes in the content of natriuretic
peptides in distinct brain regions associated with control of cardiovascular a c t ~ t y . Such
changes may be partialiy responsible for the effect of chronic moderate ethanol consumption
on blood pressure.
3.4.2. INTRODUCTION
The brain is one of the main targets for a number of ethanol-mediated effects (Nut t
and Peters. 1994. Diamond and Messing. 1994). Indeed. ethanol readily crosses the blood-
brain-barrier to modify electrical and chernical properties of the brain and allers
neuroendocrine, behavioral and cardiovascular responses. Ethanol is also associated with
changes in blood pressure (MacMahon. 1987). In fact, chronic heavy alcohol consumption
has been associated with the developrnent of hypertension (Lian. 19 15. MacMahon. 1987).
whereas low and moderate ethanol consurnption have been found to prevent the age-

dependent increase in blood pressure in both human (MacMahon. 1987) and experimental
animals (Howe et al.. 1989 and Section 3.1 .). This effect is more pronounced in
hypertensive animals. such as in spontaneously hypertensive rats (SHR) (Section 3.1.).
The mechanisms mediating alcohol's effects on blood pressure are still unknown. A
number of brain regions. such as the hypothalamus. pons and medulla have been associated
with the control of cardiovascular activity and blood pressure. It is possible then that
alterations in the activity of some of these brain regions by ethanol may be responsible.
at least in part. for the effects of alcohol on blood pressure.
Atriai natriuretic peptide (ANP). the first rnember of a natriuretic peptide famil y with
potent hypotensive properties. was discovered by De Bold and CO-workers in 198 1. S ince
then, brain natriuretic peptide (BNP) and C-type natriuretic peptide (CNP) have been
isolated frorn the porcine brain (Sudoh et al.. 1988. Sudoh et al.. 1990). These natriuretic
peptides produce hypotension by vasorelaxation. diuresis and nauiures is. and b y inh ib i t ing
the release and activity of several pressor hormones. such as arginine vasopressin ( AVP).
angiotensin II (AH) and aldosterone (Blaine. 1990. Jamison et al.. 1991). In recent years.
the implication of the natriuretic peptides in the central control of blood pressure has been
demonstrated ([mura er al.. 1997). Indeed. immunohistochemical staining has
dernonstrated that ANP is mainly localized in the hypothalamus in the adult rat brain
(Kawata er ai.. 1985. Standaert et al.. 1986. Morii et al.. 1986b). and especially in the
anteroventral third ventricle region (AV3V) ( K u and Zhang. 1994). an area believed to be
important in the regulation of fluid homeostasis and blood pressure (Buggy and Bealer.
1987). ANP is also found throughout the pons. the medulla (Kawata et al.. 1985) and the
olfactory bulbs (OB) (Gutkowska et al.. 1991). Sirnilarly. the presence of CNP has been
demonstrated in the rat hypothalamus, pons. medulla and cerebellum (Kornatsu er al..
199 t . Herman et al., 1993). In contrast, BNP is absent from the rat brain.
Our initial studies have demonstrated that both acute and chronic ethanol exposure
1 alter the activity of the hem ANP and BNP systems (Section
the strong hypotensive effects of natriuretic peptides. it
2.1. and 3.1 .). Considering
was hypothesized that the

natriuretic system (peptides and recepton) may mediate. at least in part. the effects of low
to moderate ethanol consumption on blood pressure. Ethanol may also alter the activity of
the brain natriuretic peptide systems (ANP and CNP). which may play a role in rnediating
ethanol's effects on the cardiovascular system. There is no previous data on the subject.
Thus. the objective of the present studies was to investigate the effects of chronic rnoderate
ethanol consumption on the brai n natriure tic system of spontaneousl y h ypertensive ( S H R)
and Wistar-Kyoto rats (WKY). by estimating the total content and concentration (ng/mg
protein) of ANP and CNP in the hypothalamus. pons and medulla. brain regions important
in the regulation of fiuid homeostasis and cardiovascular responses.
3.4.3. MATERLALS AND METHODS
3.4.3.1. Animal T r e m m
6 week-old male Wistar-Kyoto (WKY) and spontaneously hypertensive rats t SHR)
were used in the presenr studies (Charles River Breeding Laboratories. St-Constant.
Québec. Canada). The animals were randomly separated in 4 different groups: WKY -H,O
( n = 12), WKY-ethano1 ( n = 12). SHR-H20 ( n = 12) and SHR-ethanol ( n = 12). After a week
acclimatization period. alcohol was gradually added to the drinking water of the ethanol
groups. as described previously (Howe et al.. 1989). Brietly . ethanol-treated rats ( WKY -
ethanol and SHR-ethanol) had free access to an ethanol solution of 5 % v/v for 5 days.
10% v/v for 5 days. 15 % v/v for 5 days and 20% v/v for the remaining of the 8 months
experimental period. The corresponding control rats (WKY-H,O and SHR-H,O) had free
access to tap water. Al1 animals had free access to pellet chow (Purina. Richmond. VA) .
Furthermore. from each strain of rats. an additional 10 animals were sacritïced at 7 weeks
of age to estimate the total content and concentration (ng/mg protein) of brain A N P and
CNP prior to the age-dependent increase in blood pressure and ethanol administration.
Blood pressure was measured via the tail-cuff method (Pfeffer et al.. 1971) at the
beginning of the study (7 weeks-old) and at the end of the 8 month experimental period.
In addition, body weight was recorded every 15 days and tluid intake (alcohol solution or
water) was recorded daily.

3.4.3.2. m u e E-
After decapitation. the hypothalami. pons and medulla were removed from the brain.
Each brain region was placed in separate tubes containing ice-cold 0.1 N HCI and protease
inhibitors at final concentrations of 1 mg/ml ethylenediamine-tetra-acetate (EDTA). 10
mM phenylrnethylsuifonylfluoride (PMSF) (Sigma Chemical Co.. P-7626) and 5 rnM
pepstatin A (Sigma Chemical Co.. P-4265). The tissues were rapidl y boiled for 5 min and
cooled on ice. The brain regions were then homogenized with a microultrasonic ce11
disrupter (Kondes. Vineland. NJ). Proteins were rneasured by a modification of the
Bradford rnethod (Bradford. 1976). The remaining homogenates were centrifuged for 15
min at 4°C and the supernatants stored frozen at -75°C until assayed for ANP and CNP.
3.4.3.3. m - a v s (B1Bsl .
Hypothalamic. pontine and medullary extracts were appropriatel y diluted and the
content of ANP and CNP was directly quantified by using second-antibody RIA
procedures for both ANP and CNP. The ANP RIA has been described elsewhere
(Gutkowska et al.. 1987a). The antibody (produced by the immunization procedure of
Gutkowska et al.. (1984). final dilution 1:30000) showed a 100% cross-reactivity with the
126 aminoacid prohorrnoiie and the circulating ANP. but less than 5 % cross-reactivity with
oxidized ANP. It also showed less than 0.001 % cross-reactivity with CNP. Iodination of
ANP was done by the lactoperoxidase method. The lowest quantity of ANP rneasurable
with this assay was 0.75 pgltube. The intra- and inter-assay coefficients of variation were
4.3 and 8.3% respectively.
The CNP RIA was performed by a RIA similar to that for ANP which we developed
in Our laboratory. A typical standard curve generated with "'I-CNP and cold CNP is
presented at Fig. 3.4.1. 12SI-CNP displacement by serial dilutions of rat hypothalamic
homogenates paralleled the dose-response standard curve of rat CNP. indicating that the
peptide rneasured by the RIA was immunologically identical with synthetic CNP (Fig.
3.4.1.). Similar parallelisms to the RIA standard curve are demonstrated for pontine and
medullary extracts. The antibody (Peninsula Laboratories. final dilution 1 : 8000) showed
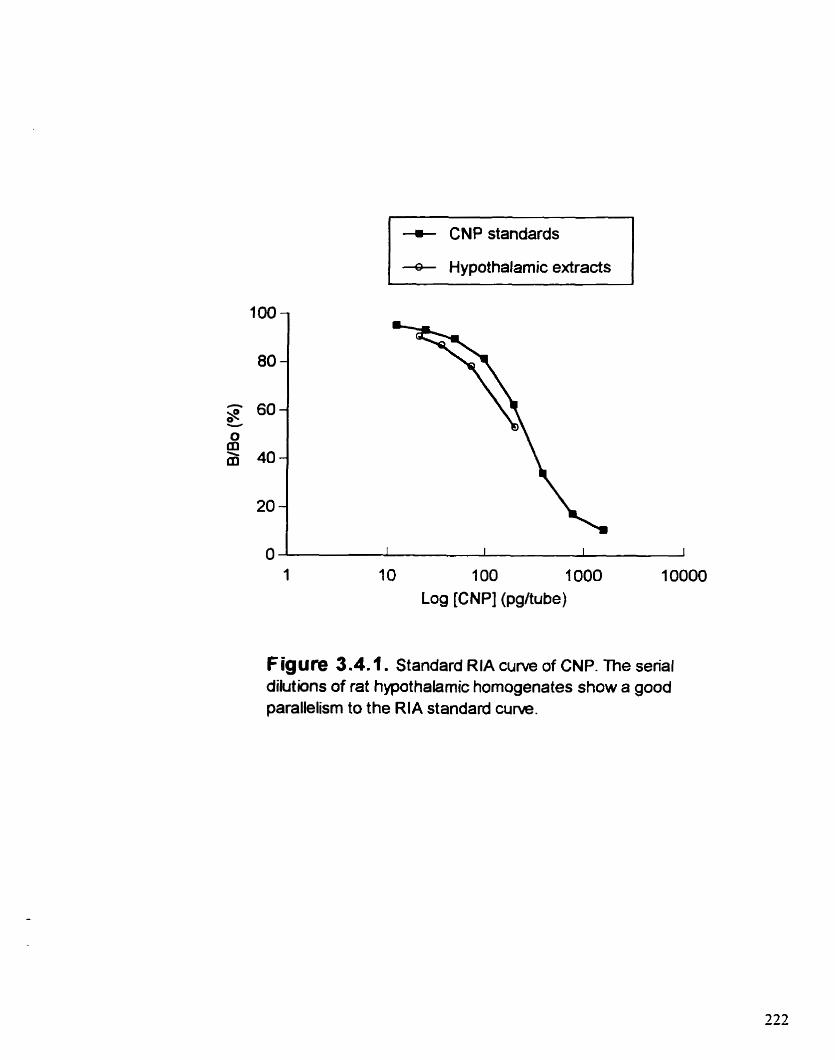
- CNP standards
* Hypothalamic extracts
01 I I 1 I
1 10 100 1000 1 O000 Log [CNP] (pgltube)
Figure 3.4.1. Standard RIA cuwe of CNP. The serial dilutions of rat hypothaiarnic homogenates show a good parallelism to the RIA standard curve.

a 100% cross-reactivity with CNP. but only a 0.015% cross-reactivity with ANP. The
iodination of CNP was also done by the lactoperoxidase method. The minimal quantity of
CNP detectable with this RIA was 12 pgltube. The intra- and inter-assay coefficients of
variation were 4.8 and 9.7 % respective1 y.
. . 3.4.3.4. -1 A m
Results are presented as mean I S.E.M.. Statistical analysis was done by a two-way
analysis of variance (ANOVA). with the treatment or the age as the first independent
variable and the strain as the second independent variable. This analysis was followed by
the Neuman-Keuls multiple cornparison test. A p value of 0.05 or lower was considered
signiticant.
3.4.4. RESULTS
3.4.4.1. J3ffect of a s
There is a significant decrease with age ( p ~ 0 . 0 0 1 ) in hypothalamic. pontine and
medullary ANP levels of both SHR and WKY rats (Fig. 3 - 4 2 . ) .
Total CNP content in the hypothalamus is augmented with age (pc0.001). whereas
its concentration is decreased (pé0.001) (Fig. 3.4.3.). Pontine and medullary CNP content
and concentration are decreased with age in both strains of rats (~50.00 1).
3.4.4.2. Effect of strah
Both the content (pc0.05) and the concentration (ps0.05) of hypothalamic A N P are
higher in young SHRs cornpared to age-matched WKY rats (Fig. 3.4.2.). In adult anirnals.
the total hypothalamic ANP content is higher in the SHR compared to WKY rats (pc0.05).
but the hypothalarnic ANP concentration is sirnilar between the two strains of rats. Pontine
ANP content (pc0.001) and concentration ( p ~ 0 . 0 1 ) are lower in young SHRs compared
to young W K Y rats. However. adult SHRs have significantly higher pontine ANP content
(pc0.01) and A N P concentration ( p ~ 0 . 0 5 ) than age-matched WKY animals. There is no
difference between the medullary ANP levels of 7 week-old SHR and WKY rats. In
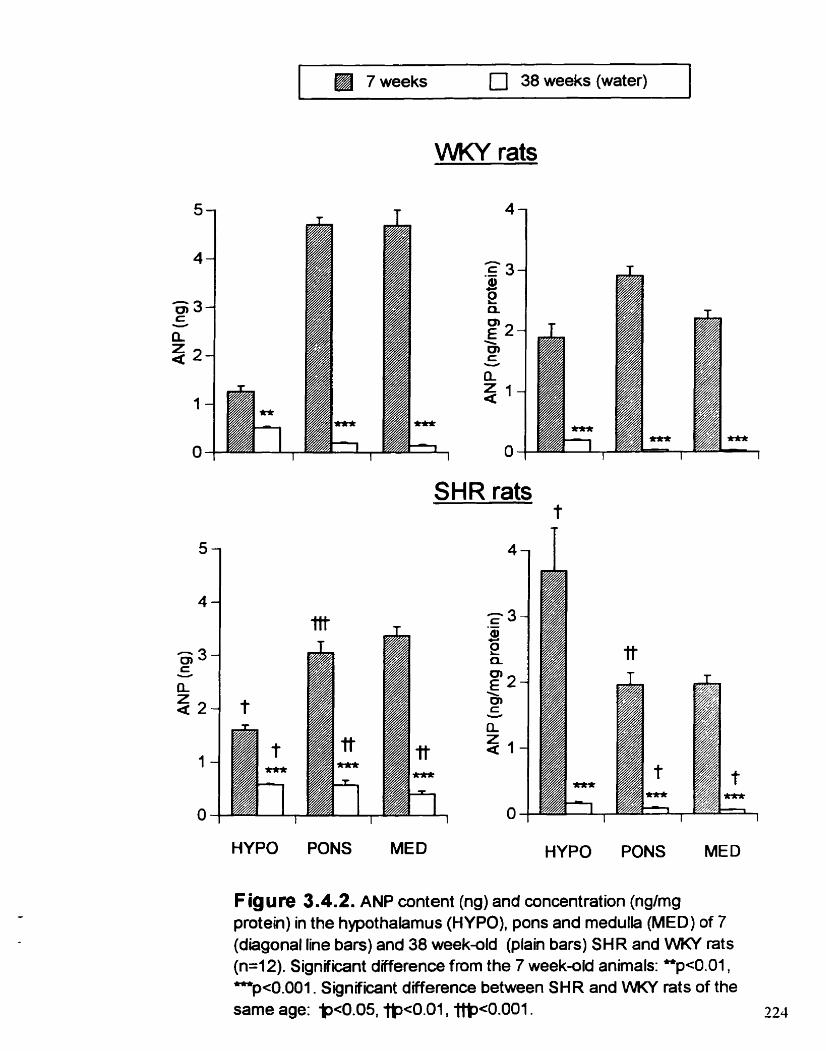
1 7weeks 0 38 weekç (water) 1
WKY rats
SHR rats
HYPO PONS MED
I I I
HYPO PONS MED
Figure 3.4.2. ANP content (ng) and concentration (nglmg proten) in the hypothahmus (HYPO), pons and medulia (MED) of 7 (diagonal line bars) and 38 week-old (plain bars) SHR and WKY rats (n=12). Signlicant difference from the 7 week-old animais: *p<0.01, "p~0.001. Significant difference between SHR and W rats of the same age: Wc0.05, m<O.O1, ttpc0.001.
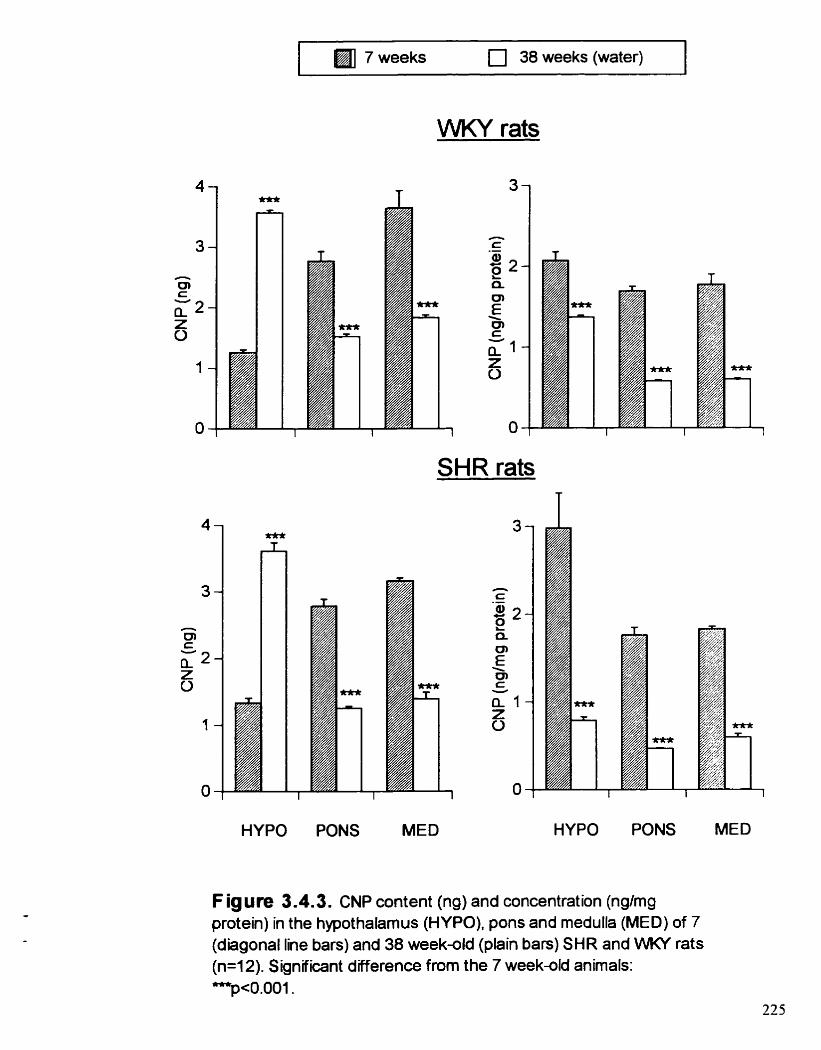
I 7 weeks 0 38 weeks (water) 1
WKY rats
SHR rats
HYPO PONS MED HYPO PONS MED
Figure 3.4.3. CNP content (ng) and concentration (nglmg protein) in the hypothalamus (HYPO), pons and medulla (MED) of 7 (diagonal line bars) and 38 weekold (plain bars) SHR and WKY rats (n=12). Significant difference from the 7 weekold anirnals: -p<0.001.

contrast. both the total content (pi0.01) and concentration (pr0.05) of medullary ANP
are higher in the adult SHRs compared to adult WKY rats.
There is no significant difference in the hypothalamic. pontine and medullary CNP
levels (content and concentration) between young or adult SHR versus young or adult
WKY rats. respectively (Fig. 3.4.3 .).
3.4.4.3. Effect of e t h a d
After 8 monrhs of moderate ethanol consumption. both SHR and WKY rats had
significantly lower blood pressure than water-treated animals of the same strain (table
3.4.1. ). The body weight and the daily fluid consumption are not different between the
ethanol- and water-mted rats (shown for the last week of treatment only) (table 3.4.1. ).
There is a signitïcant increase in the hypothalamic ANP content of SHR. but not
WKY rats. after chronic ethanol consumption (Fig. 3 A.4.). Indeed. total hypothalamic
ANP content is raised from 0.58 f 0.02 ng in SHR-H,O to 1.16 f 0.06 ng (p~O.00 1 ) in
the SHRethanol group. Likewise. a two fold increase in hypothalamic ANP concentration
is observed following the chronic ethanol treatment (from 0.17 t 0.02 ng/mg protein in
SHR-H,O to 0.51 & 0.05 ng/mg protein. p-0.001 in SHR-ethanol). Chronic moderate
ethanol consumption induced an increase in the pontine ANP concentration in SHR. but
not in WKY rats (~50.05). Medullary ANP content (pc0.05) and concentration (pd.00 1 )
were also higher after long-term ethanol consumption in SHR. but not W K Y rats.
Hypothalamic CNP content (~50.01) and concentration ( ~ ~ 0 . 0 5 ) are eievated after
chronic ethanol treatment in both SHR and WKY rats (Fig. 3.4.5.). Pontine CNP
concentration. but not CNP content. is also signitïcantl y increased (ps0.0 1 ) by the erhanol
treatment in the SHR. but not WKY rats. Medullary CNP content and concentration are
increased by the ethanol treatrnent in the SHRs. However. in the rnedulla of WKY rats.
chronic ethanol treatment increased the concentration but not the total content of CNP.
3.4.5. DISCUSSION
The present experiments have demonstrated age, strain and ethanol-induced changes
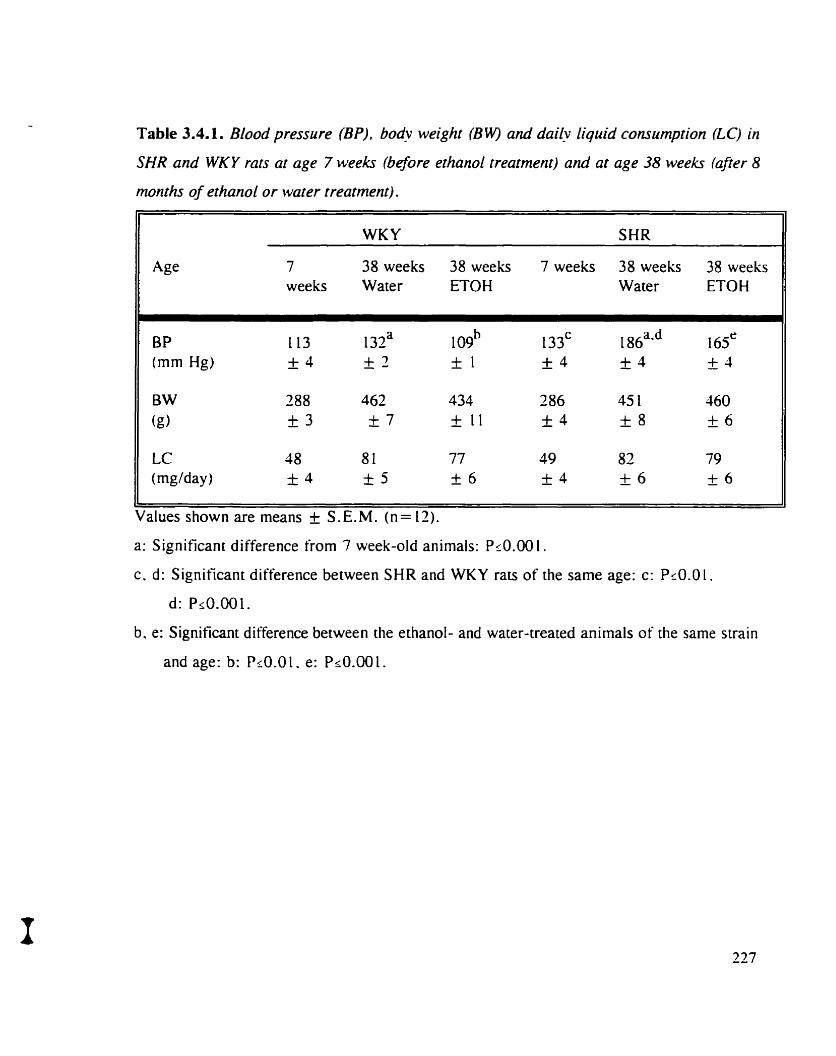
Table 3.4.1. Blood pressure (BP), bodv weight (BW) and dai& Iiquid conrumption (LC) in
SHR and WKY rats at age 7 weeks (befiore ethanol treatrnent) and at age 38 weekr fafier 8
months of ethanol or water treatment) .
WKY S H R II 7 38 weeks 38 weeks 7 weeks 38 weeks 38 weeks weeks Water ETOH Water ETOH
BP 113 1 3~~ 1 0 9 ~ 133' 186"** 165' (mm Hg) - + 4 * 2 I 1 + 4 t 4 & 4
IL Values shown are means f S.E.M. ( n = t2).
a: Significant difference frorn 7 week-old animals: P4L00 1 . c. d: Signitïcant difference between SHR and WKY rats of the same age: c: Pc0.0 1 .
b. e: Signiticant difference between the ethanol- and water-treated animals of the same strain
and age: b: P ~ 0 . 0 1, e: P~0.00 1.
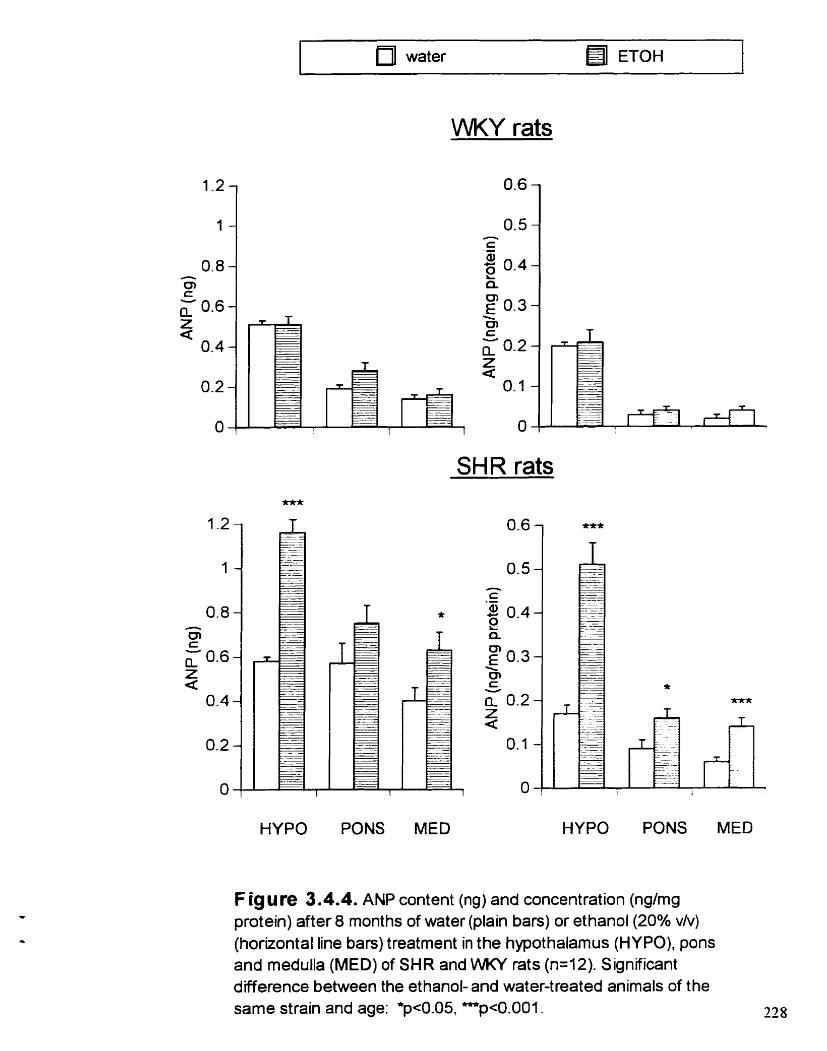
water
WKY rats
HYPO PONS
SHR rats
*
MED HYPO PONS MED
F igure 3.4.4. ANP content (ng) and concentration (ngimg protein) after 8 months of water (plain bars) or ethanol(20% vlv) (horizontal line bars) treatment in the hypothalamus (HYPO), pons and medulla (MED) of SHR and WKY rats (n=12). Significant difference behveen the ethanol- and water-treated animals of the same strain and age: '~~0.05, "p<0.001.
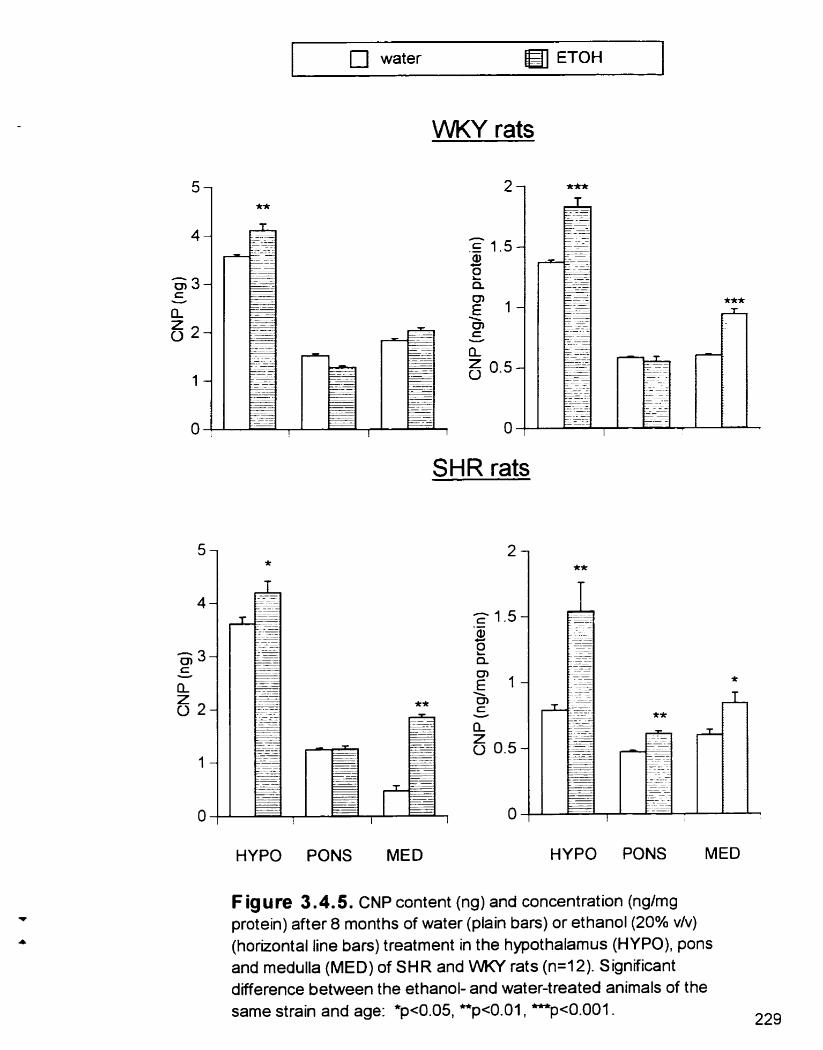
I 0 water ETOH I
WKY rats
SHR rats
HYPO PONS MED HYPO PONS
F ig u re 3.4.5. CNP content (ng) and concentration (nglmg
*
f MED
protein) after 8 months of water (plain bars) or ethanol(20% vlv) (horizontal line bars) treatment in the hypothalamus (HYPO), pons and medulla (MED) of SHR and WKY rats (n=12). Significant difference between the ethanol- and water-treated animais of the same strain and age: *p<0.05, wp<O.O1, "p<0.001.

in natriuretic peptide levels in brain regions involved in cardiovascular homeostasis. ( 1 )
There is a signifiant decrease with age in both ANP and CNP levels in the hypothalamus.
pons and medulla of SHR and WKY rats. (2) There is significantly higher brain ANP, but
not CNP, levels in SHR compared to WKY rats. (3) There is significantly higher brain
ANP levels following chronic moderate ethanol consumption in SHR but not WKY rats.
In contrast, brain CNP levels are higher in both SHR and WKY rats following the chronic
ethanol ueatment.
The importance of the brain in mediating a number of alcohol effects is indicated by
the nurnerous ethanol-induced behavioral effects (Nutt and Peters. 1994). 1 t has been
hypotheskd that some of the ethanol-induced effects on biood pressure may be mediated
through specific modifications in the activity of relevant neurohormonal and
neurommitter systerns of the brain, such as the natriuretic or renin-ang iotensin systerns
(Imura et al.. 1992, Steckelings. 1992). or through secondary modifications in the activity
of peripheral pressor and depressor agents by ethanol-induced changes in hypothalamic and
pituitary hormones (Cicero. 198 1). Ethanol. an amphiphilic dmg, rapidly crosses the
blood-brain-barrier to physically dissolve into brain ce11 membranes (Diamond and
Messing , 1994). This property of alcohol modifies membrane fluidity (Nutt and Peters.
1994) and inhibits brain Na'K'ATP ase activity (Sun, 1976, Nhamburo et al.. 1986.
Swann, 1990). leading to various alterations in the chernical and electrical properties of
brain cells. Interestingly , centrally administered ANP significantly enhanced the release
of Na'K'ATP ase inhibitors (Crabos et al., 1988). It is possible then that elevated brain
ANP levels in ethanol-treated rats serve as a mediator for the ethanol-induced inhibition
of N a X ' ATP ase activity. Further work is needed to elucidate this observation. Acute
ethanol also decreases the sensitivity of N-methyl-D- Aspartate (NMDA) receptors for
excitatory aminoacids like glutamate and potentiates y-aminobutyric acid (GABA)-A
recepton, explaining in part its sedative actions (Lovinger, L989, Wafford et al., 1991).
Ethanol also affects the activity of several neurohormonal and neurotransrnitter s ystems
such as AVP or angiotensin II, some of which are of critical importance in the long-term

control of blood pressure (Cicero. 1 98 1 ) . The eiTect of ethanol on blood pressure is biphasic. Indeed. although high ethanol
consumption has long been shown to produce hypertension (Lian. 1915). low and/or
moderate ethanol consumption has been s hown to prevent the age-dependent increase in
blood pressure ( MacMahon. 1987). Thus. a nurnber of epidemiological studies have
reported lower systolic blood pressure in light and moderate human drinkers than in both
heavy and non-drinkers (Harburg et al.. 1980. Gillman et al.. 1995). Similarly. animal
experiments have demonstrated the prevention of the age-dependent increase in blood
pressure with moderate ethanol consumption. especial l y in the hypertensive strains of rats
(Howe et al.. 1989 and Section 3.1.). The mechanism(s) by which ethanol induces this
antihypertensive effect remain unclear. Some studies have indicated that this ethanol-
mediated protection from this age-dependent increase in blood pressure could be due. at
least in part. to lipoproteins which are differently regulated by low and high ethanol
consumption (Hojnacki et al., 1988). Recently. the natriuretic peptide famil y was added
to the list of systems which centrally regulate blood pressure and fluid homeostasis ( Imura
et al.. 1992). Therefore. alterations in the activity of the brain A N P system may be
partially responsible for ethanol's effect on blood pressure.
In the adult brain. the highest A N P concentration is found in the hypothalamus
(Kawata et al.. 1985. Standaert et al.. 1986). Within this tissue. reg ions involved in the
regulation of fluid homeostasis and blood pressure. such as the AV3V area. m i n strongly
for ANP immunoreactivity (Kawata et ab. 1985). Pro-ANP mRNA is also present in high
quantities within the hypothalamic areas involved in cardiovascular regulation (Gundlach
and Knobe. 1992). Likewise. CNP (Komatsu et al., 1991) and CNP rnRNA (Herman et
al.. 1993) are found in high quantities in the hypothalamus. In addition to the
hypothalamus. natriuretic peptides are also present in the pons. mainly in the dorsal
tegmental area. and in the medulla. in close association with regions involved in
cardiovascular control (Kawata et al.. 1985).
I t has been reported that S H R rats exhibit higher levels of hypothalamic A N P

compared to WKY rats (Morii et al.. 1986b. Debinski et al.. 1989. Jin et al., 199 1 . Chen
et al.. 1997. Komatsu et al.. 1992b). The present studies confirmed this finding.
Furthermore. in contrast to the finding of lmada and collegues ( 1985). who reported no
significant change in hypothalamic A N P levels with age. the present stud ies demonstrated
a reduction in hypothalamic ANP levels with age in both strains of rats. This discrepancy
could be explained by the different ages of the animais used in the two studies (38 weeks-
old in the present study versus 15 weeks-old in that of Imada). Significantly higher content
of pontine and medullary A N P in adult compared to young SHRs was observed in the
present study. confirming previous reports (Imada et al.. 1985. Komatsu et al., 1992b).
Chronic ethanol consumption increases ANP levels in the hypothalamus. pons and
rnedulla of hypertensive. but not normotensive rats. The presence of elevated hypothalamic
ANP levels in SHR rats following chronic ethanol treatment may enhance the contribution
of central natriuretic peptides to cardiovascular homeostasis. I ndeed.
intracerebroventricular (i.c.v. ) injection of ANP (third and lateral ventricles) produces
dose-dependent inhibition of water intake and angiotensin-induced drinking in overniglit-
dehydrated rats (Nakamura et al.. 1985. Antunes-Rodrigues et al.. 1985. Katsuura er al..
1986). S imilarl y. centrally administered ANP (lateral ventricles) suppresses sait appetite
in salt-depleted rats and in strains known to have increased sa1 t preference. such as S H R
(Fitts et al., 1985. ltoh et al.. 1986b). Furthermore. i.c.v. injection of ANP (lateral
ventricles) is associated with diuresis (Israel and Barbella. 1986. Shoji et al., 1987. Lee
et al., 1989). Since i.c.v. injection of A N P also signitkantly blunts the A V P increase
induced by i.c.v. injection of angiotensin I I or by osmotic stimulation. parts of the diuretic
effect of central A N P administration rnay be explained by its effect on the A V P system
(Yamada et al., 1986. litake et al., 1986. Inoue et al.. 1990). Interestingly. the diuretic
effecr of ethanol is also anributed in part to the inhibition of AVP release (Eisenhofer and
Johnson. 1982). Therefore. the elevated brain A N P levels following chronic ethanol
consumption may produce a greater inhibit ion of vasopressinergic neurons and thus
elevated diuresis in these animals compared to water-treated rats. Final ly . central l y

administered ANP inhibits the pressor effects of i.c.v. injection of angiotensin I I in
conscious rats (Itoh et al., 1986a. Shimizu et al.. 1986. Castro et al., 1987). The exact
site of action of ANP injected within the hypothalamus is not clear. but recent evidence
suggests that ir rnay be the AV3V area (Ku and Zhang. 1994). The subfornical organ rnay
also be important since microinjections of ANP into this region blocks angiotensin II-
induced neuronal stimulation (Hanori et al.. 1988. Ehrlish and Fitts. 1990). I nterestingl y.
it was previously demonstrated that hypothalamic ANP concentration was signitïcantly
reduced in S H R rats fol low ing antihypertensive treatment. suggesting that the increased
hypothalamic ANP levels represented a homeostatic response to the elevated blood
pressure in order to control the increase in arter ial pressure (Rus koaho and Leppaluoto.
1988). In contrast. the fact that the hypotensive effect of chronic moderate ethanol
consumption in SHR rats is associated with significant increases in brain ANP leveis
suggests a specific effect of alcohol in central ANP regulation rather than a secondary
response to lower blood pressure. Thus. alcohol either directly or indirectly . through its
effects on other systems known to regulate the activity of hypothalamic ANP. such as the
cholinergie and norepinephrine systems (Nissen et al.. 1989. Baldissera et al.. 1989). rnay
modify the synthesis and release of ANP by the hypothalamus. Therefore. the increased
hypothalamic ANP concentrations associated with the possibil ity of chronic enhancement
of centrally-mediated ANP hypotensive effects rnay represent one mechanism by which
the prevention of the age-dependent increase in blood pressure is explained.
The elevated ANP levels found in the medulla of SHR rats following chronic ethanol
treatment rnay also rnodify ANP-med iated cardiovascular homeostasis. The ANP-rich
region of the nucleus tractus solitarius is the sire of termination of baroreceptor afferent
fibres. Therefore. ANP rnay be involved in lowering blood pressure by interfering with
the baroreceptor reflex. Indeed. microinjections of ANP into the nucleus tractus solitarius
or into adjacent areas such as the ventrolateral medulla caused significant decreases in
b l d pressure (McKitrick and Calaresu. 1988. Bergagl io and Calaresu. 1990). S im ilarl y.
the administration of A N P into the fourth ventricle produced a profound decrease in
arterial blood pressure (Levin et al.. 1989). Therefore. it is possible that elevated

medullary ANP levels in ethanol-neated SHR rats are associated with greater modifications
in the transmission of the baroreceptor information within the nucleus tractus solitarus and
medullary regions. leading to a lowering in blood pressure that may explain in part the
prevention of the age-dependent increase in blood pressure observed following chronic
moderate ethanol consumption.
There was no strain-specific difference in brain CNP levels between WKY and SHR
rats. In contrast. hypothalarnic CNP content was significantly increased with age in both
suains of rats. whereas pontine and medullary CNP levels were decreased during the same
period. To our knowledge. this is the first report of strain (WKY versus SHR) or age (7
versus 38 week-old animals) modifications in brain CNP levels.
C hronic ethanol consumption increases CNP levels in the hypothalamus. pons and
medulla of hypertensive rats. and hypothalamic and medullary CNP levels in WKY rats.
In contrast to ANP. the exact role of CNP in brain tissues and its possible contribution to
cardiovascular regulation is still unclear (Imada et al.. 1992). It is therefore difficult to
decide what the significance of the modifications in the activity of the brain CNP system
by chronic ethanol consumption is and what are its effects on blood pressure.
The reason for the different sensitivity of the central natriuretic system toward ethanol
between SHR and W K Y rats is unclear but may retlect genetic differences between the two
strains of rats. Indeed. grafting embryonic hypothalamic SHR cells into the hypothalamus
of adult WKY rats resulted in rapid elevations of the systolic blood pressure and heart rate.
suggesting profound differences in the brain characteristics between these two strains of
rats (Eilam et al., 199 I ). Other endocrine and neuroendocrine deviations from
normotensive rats have been observed in SHR rats. such as differences in renin, arginine
vasopressin (AVP) or vasoactive intestinal polypeptide (VI P) mRNAs (Lang et al.. 198 1 .
Avidor et al., 1989, Kreutz et al., 1993). Furthermore, alterations in the ANP and B N P
mRNAs of the heart ventricles by ethanol have also been demonstrated in SHR. but not
WKY rats (Sections 3.1. and 3.2.). However, other parts of the natriuretic peptide system
have been similarly modified by chronic moderate ethanol administration in both strains
of rats. such as atrial ANP and BNP contents (Sections 3.1 and 3.2.) . Considering the

genetic differences between the two strains and the more profound ant ih ypertensive e ffects
of moderate ethanol consumption in SHR compared to WKY rats (tabie 3 -4.1. ). a different
activation of the major components of the natriuretic peptide system in the brain. the hem
and the kidney may exist between the two strains of rats. producing faster and greater
effects on the blood pressure of ethanoi-treated SHR than ethanol-treated WKY rats.
In summary. during pathophysiological conditions such as hypertension. the elevated
ANP levels seen in the hypothalamus. pons and medulla are considered compensatory to
the higher blood pressure. On the other hand. the increase in brain ANP and CNP levels
of SHRs observed during chronic moderate ethanol administration. in spite of the
considerably lower blood pressure measured in these animais compared to water-treated
SHRs. suggests a direct or indirect stimulatory effect of ethanol on the activity of the A N P
system which is independent of the hemodynamic control. Such stimulation in the activity
of the brain A N P systern may play an important role in the antihypertensive effect of
chronic moderate ethanol consumption.
3.4.6. ACKNOWLEDGEMENTS
The authors wish to thank Ricardo Claudio for his excellent care of the animals.
Special thanks are also given ro Céline Coderre and Nathalie Charron for their technical
assis tance.
P.G. is the recipient of a doctoral scholarship from the "Fonds pour la formation de
Chercheurs et I 'Aide à la Recherche" (FCAR). This work was supported by gram from
the Medical Council of Canada (MRC) (MT- 10337 and MT- I 1674) (J.G.) and by the
alcohol research program at Douglas hospital (C.G.).

Section 3.5.
CIRCUMNENTRICUIAR ORGAN NATRIURETIC PEPTIDE
RECEPTORS FOLLOWING CHRONIC MODERATE ETHANOL
CONSUMPTION IN SHR AND WKY RATS
P. Guillaume, C. Gianoulakis and J. Gutkowska
Manuscript in preparcrüon
Contribution bv CO-authors: Dr. C. Gianoulakis and Dr. J. Gutkowska were my CO-
s u p e ~ s o r s .
Acknowled~ements: R Claudio was the animal technician. C. Coderre and N. Charron
performed the ANP iodinations.

3.5. CIRCUMVENTRICULAR ORGAN NATRWRETIC PEITIDE
RECEPTORS FOLLOWING CHRONIC MODERArlTE ETHANOL
CONSUMPTION IN SHR AND WKY RATS
3.5.1. ABSTRACT
Chronic moderate ethanol (ETOH) consumption is associated with the prevention of
the agedependent increase in the blood pressure of experimental animais. The rnechanisms
mediating this antihypertensive effect of ETOH are unclear, but the natriuretic peptide
system has been hypothetized to contribute to this effect. In the present experiments.
ETOH- induced modifications in the characteristics (B, and Ka) of the natriuretic
receptors are investigated in the subfornical organ (SFO), choroid plexus (CP) and area
postrema (AP) of spontaneously hypertensive (SHR) and Wistar-Kyoto (WKY) rats. Both
strains received either water or a 20% v/v ETOH solution for 8 months. A significantly
lower number of total natriuretic recepton is observed in the water-treated SHR compared
to the water-treated WKY rats in the SFO and CP, but not in the AP. This lower B, in
the SFO and CP of hypertensive compared to normotensive animals is associated with
elevated proportions of NPR-C receptors. Furthermore, it was observed that chronic
moderate ETOH consumption significantly decreased the affinity of the NPR-C receptors
in the SFO and CP. In contrast, ETOH does not alter the characteristics (Bu and KJ of
the NPR-A receptors in the SFO, CP or AP of either SHR or WKY rats.
Thus, development of age-dependent hypertension seems to be associated w ith
changes in the total density as well as in the relative proportion of the natriuretic recepror
subtypes in the SFO and CP. Furthermore. the ETOH treatment is associated with
alterations in the circumventricular organ ANP receptors K, that may mediate. at least in
part. the antih ypertens ive effect of moderate alcohol consumption.
INTRODUCTION
Three distinct receptors have been described for the natriuretic peptides fam ily

( Anand-Srivastava and Trachte. 1 993). The natriuretic peptide receptor (NPR)-A and
NPR-B are both bel ieved to be active receptors, consisting of a single transmembrane
domain, with cGMP as their second messenger. Atrial natriuretic peptide (ANP) appears
to bind preferably to NPR-A while C-type natriuretic peptide (CNP) binds preferentially
to NPR-B (Suga et al., 1992a). The NPR-C. on the other hand. is less selective. binding
various ANP analogs, and seerns to be part of a buffer system which major function is the
clearance of excess amounts of natriuretic peptides frorn the circulation (Maack et al.,
1987). In the brain, the natriuretic receptors are found rnostly in the circurnventricular
organs such as the subfornical organ (SFO). the choroid plexus (CP) and the area postrema
(AP) (Quirion et al., 1986, Brown et Czamecki, 1990a), on brain microvessel endothelial
cells (Whitson et al., 1991) and on the blood side of the BBB (Ermish, 1992).
Long-term ethanol (ETOH) drinking has been associated with some antihypertensive
effects in animal experimenü, particularly in hypertensive strains of rats (Howe et al.,
1989 and Section 3.1 .). Likewise. lower systolic blood pressure (BP) has been found in
light and moderate human ETOH drinkers than in both heavy and non-drinkers. suggesting
the existence of some depressor effects of low and moderate alcohol consurnption
(MacMahon. 1987). It has been hypothetized that the diuretic and antihypertensive effects
of low and moderate ETOH consumption may be mediated, at least in part. by the
natriuretic peptide family (Colantonio et al.. 1991 and Section 2.1.). Modifications in
natriuretic receptors have alread y been noticed in the kidneys fol low ing chronic mode rate
ETOH drinking, implicating this system in the antihypertensive effects of alcohol (Section
3 -3 .). Considering the importance of the communication and feedback mechanisms
between the brain and the circulation in the control of long-term BP. it was interesting to
investigare also the possibility of ETOH-induced alterations in :he binding characteristics
of the natriuretic receptors in the circumventricular organs. Structures such as the SFO.
CP and AP lack a blood-brain-barrier (BBB), a unique situation that enables a direct link
between hormonal inputs from the periphery and neural syscems in the brain. For example.
the binding of circulating ANP to natriuretic receptors in the SFO has been shown to
inhibit AII-sensitive neurons projecting to the AV3V area (Hattori et al., 1988, Shibata

er al.. 1992) and therefore to block stimuli for AVP release and tluid intake. supporting
an inhibitory action of ANP on the hypothalamus and AVP system. Modifications in
c ircumventricular natriuretic recep tor characteristics have already been demonstrated in
various conditions. such as hypertension. and have been suggested to be responsibie to
some extent for the development of hypertension (Saavedra. 1986. McCarty et al.. 1986).
Li kewise, ETOH-induced al terations of the circumventricular natriuretic recep tors may
help to explain the prevention of the age-dependent hypertension in experimental animals
such as SHR and WKY by chronic Iow to moderate alcohol consumption.
Thus. in the present stud ies. the natriuretic receptor characteristics (B,, et K,) were
investigated in the circumventricular organs (SFO. CP and AP) following the chronic
administration of 20 % v/v ETOH in hypertensive (SHR) and normotensive (WKY) rats.
3.5.3. MATERIALS AND METHODS
3.5.3.1. Animal Treatmem
6 weeks-old male Wistar-Kyoto (WKY) and spontaneously hypenensive rats (SHR)
were used in the present studies (purchased fiom Charles River Breeding Laboratories. St-
Constant. Québec. Canada). The animais were randomly separated in 4 different groups:
WKY-H,O (n=6). WKY-ETOH (n=6). SHR-H,O (n=6) and SHR-ETOH (n=6). Afrer
a week acclimatization period. alcohol was gradually added to the drinking water of the
ETOH groups. as described previously by us (Section 3.1. ) and by Howe ef al.. ( 1989).
Brietly, rats received 5 % v/v for 5 days. 10% v/v for 5 days. 15 % v/v for 5 days and
20 % v/v for the remaining of the 8 months experimental period. During that time. rats had
free access to their respective water or ETOH solutions and to pellet chow (Purina.
Richmond. VA).
3.5.3.2. Autoradiomhic Studie
The brains were frozen intact in isopentane and kept at -75 OC until used. Frozen. 20-
pm-thick coronal brain sections were cut in a cryostat at - 15 OC through the SFO-CP and
through the AP regions. These tissue sections were thaw-mounted ont0 poly-L-lysine-

treated siides and placed overnight into a desiccator under vacuum at 4°C.
The brain sections were washed for 15 minutes in a preincubation buffer consisting
of O. 1 % PEI in 50 m M Tris-HCI. pH 7.4. and subsequently incubated with 50 pM 131-
ANP (specific activity. Zûûû Ci/mmol) for 1 hour at room temperature. The incubation
buffer consisted of 50 mM HCI, pH 7.4. 150 m M NaCI. 5 rnM MnCl, (Sigma Chemicai
Co.. M-3634). 40 g/ml bacitracin (Sigma Chernical Co.. B-OL25) and 0.5 % bovine serum
albumin (BSA). The competitive inhibition of "'1-ANP binding to the membranes by
unlabeled peptides was examined for each animal by coincubating the membranes with
various concentrations of cold ANP (10"' to l ( r M) and des-(Glnllb. Seri''. GlyllX. Leu1''.
Gly'31ANF,,,.,,l (or cANF) (IO-"'. IO-". [O-' M). cANF is a ring deleted ANP analog
which binds specifically to the clearance receptors (NPR-C) (Maack et al.. 1987).
The highest concentration of cold peptides ( 10'" M) was considered to exhibit non-
specific binding (NSB). After incubation. the sl ides were washed 4 times in 50 mM Tris-
HCl. pH 7.4 (4°C). rinced quickly in distilled water (4°C) and left to dry overnight. The
tissue sections were then exposed on Phosporlmager cassettes for 5 days. The intensity of
the signals were subsequently quantitated by densitometry using a Phosporlmager
( Molecular Dynamics. Sunnyvale. CA). Relative quantities between experiments were
correlated by known quantities of I3I-ANP simultaneously exposed ont0 Phosphorlmager
cassettes. The average of at least 4 values per concentration per rat were calculated for
each individual brain SFO. CP and AP and their optical densities converted to radioactive
concentratioiis (cpm). based on the cornparison with the "'1-ANP standards.
3.5.3.3. Statistical t .
The maximum binding capacity (B-J and the apparent dissociation constant (K,)
were calculated when possible by the LIGAND iterative model-tïtting computer program
(Munson et al.. 1980).
3.5.4. RESULTS
Natriuretic receptors characteristics (B,, and Kd) are calculated by displacing "'1-

ANP with increasing concentrations of unlabeled A N P ( 10-l0 to IO-" M) and cANF ( IO?
IO-". 10-" M) in SFO. CP and AP of water- and ETOH-treated S HR and WKY rats. The
LIGAND -calculated B, and K, values for the three circumventricular organs are
presented on Table 3.5.1.
3.5.4.1. S u b f o ~ c a l orgao (SFO) and choroid plexus (CF')
The average (n=6) displacernent of labeled ANP by cold ANP is presented on Figure
3.5.1 A for SFO and Figure 3.5.2A for CP. As calculated from these values. signitkantly
lower (n =6. p~0.05) total ANP receptor densities are found in hypertensive compared to
normotensive anirnals (Table 3.5.1 .). In contrat. a significant decrease in K, is reported
in the CP only (n=6. ~ ~ 0 . 0 5 ) . When I3I-ANP is incubated with increasing concentrations
of unlabeled cANF (Figures 3.5. I B. and 3.5.2B.). another difference is visible between
SHR and WKY rats. cANF has no (in SFO) or little ( in CP) capacity to displace labeled
ANP in normotensive animals. However. a significant portion of '"1-ANP is displaced by
cANF in hypertensive rats. 1 ndeed. the B,, values for "'1- ANPlcANF displacement
represent 64.8 and 36.4% of maximal binding density calculated with 1251-ANPIANP in
the SFO of SHR-water and SHR-ETOH rats. respectively (Figure 3.5.1B). Likewise. the
B,,, values for 1251-ANP/cANF displacement represent 78.0 and 63.1 % of the ma.. imal
binding density for l'SI-ANP/ANP in the CP of SHR-water and SHR-ETOH animals
respectively. whereas they are calculated as only 28.0 and 35.1 % in WKY-water and
WKY-ETOH rats (Figure 3.5 -18). The availability of complete and specific antagonists
toward the various natriuretic receptors are needed to confirm this result. but these
calculations suggest that the lower numbers of total natriuretic receptors in hypertensive
anirnals are also of a different nature than those of normotensive controls. consisting of an
elevated proportion of NPR-C (or clearance) receptors.
No significant variation is seen between the total ANP receptor characteristics (B,,
and KJ of water- versus ETOH-treated anirnals (Table 3.5.1 .). Similady. the quantity of
NPR-C receptors. evaluated by the displacement of labeled ANP by cold cANF. is not
different between treatments. However, there is a significant increase in the K, measured
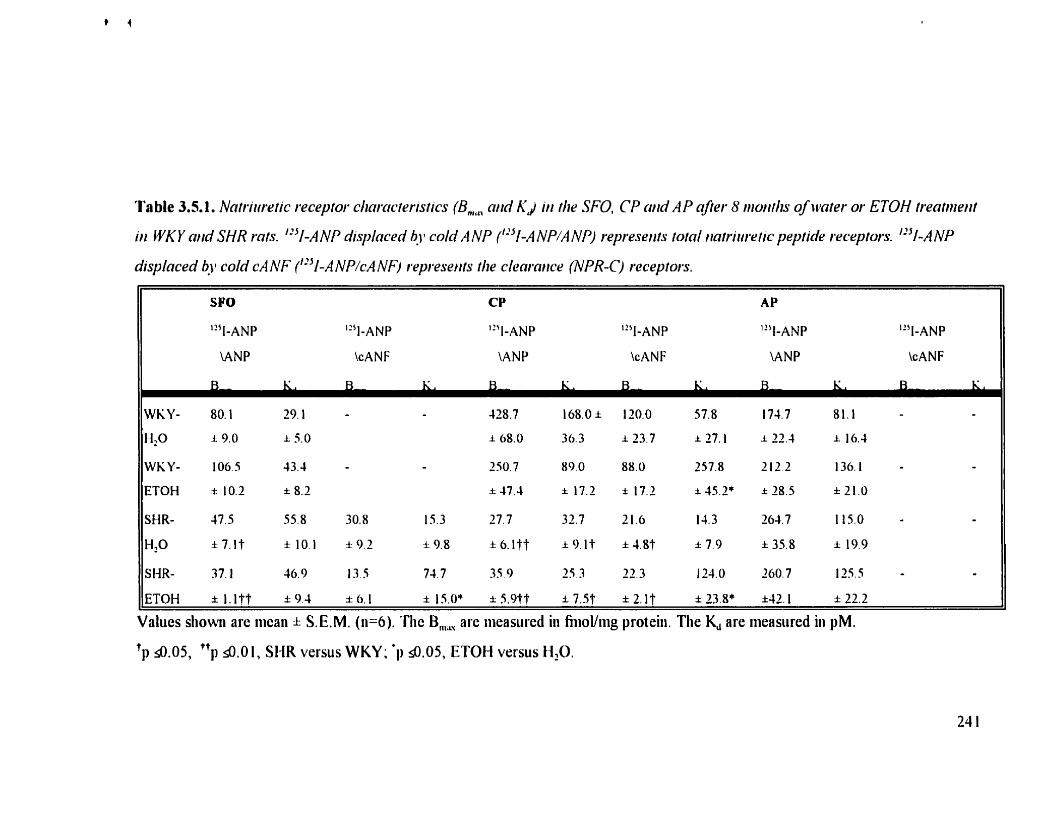
Table 3.5.1. Nott*iitrrtic tecepot* cliot*cicterrstics (B , , , o t d KA in the SFO, CP ritid A P nfret* 8 tirotrtlu. ojiinter or ETOH ttaenttnetit
itr WK Y n id SHR rats. "'1-A NP displc~erl by cold A NP ('"1-ANP~A NP) repesetrts totnl trotriirtatic peptide ivceptors. "%A NP
displnced bjt cold CA NF ('"1-A NP/cA NF) rel>t*esetrts the clenrntice (NPR-C) receptotrr.
WKY- 80.1 29.1 128.7 168.0 ~t 120.0 57.8 174.7 81.1
4 0 k 9.0 5.0 * 68.0 36.3 * 23.7 * 27.1 L 22.4 16.4
WKY- 106.5 43.4 250.7 89.0 88. O 257.8 212.2 136.1
ETOH I 10.2 * 8.2 * 47.4 k17.2 k l 7 . 2 i 45.2* k 28.5 21.0
SHR- 47.5 55.8 30.8 15.3 27.7 32.7 21.6 14.3 264.7 1 15.0
H,O * 7.1t 1 0 1 *9.2 i 9.8 k6.17t k9.l-t i4.81. 7.9 * 35.8 ;t 19.9
SHR- 37.1 46.9 13.5 74.7 35.9 25.3 22.3 124.0 160.7 125.5
ETOH i : l . l t t k9.4 i: 6.1 i: 15.0* * 5.977 i 7 3 * 2.1t *23,8* k4Z.l k 22.2
Values sliowii are inean * S.E.M. (ii=6). Tlie B,,,, are iiieasured iii fiiioViiig proteiii. The Y, are iiieesiired in pM.
'p 4.05, "p 4 . 0 1, SI-IR versus WKY; 'p 4.05, ETOH versus HIO.
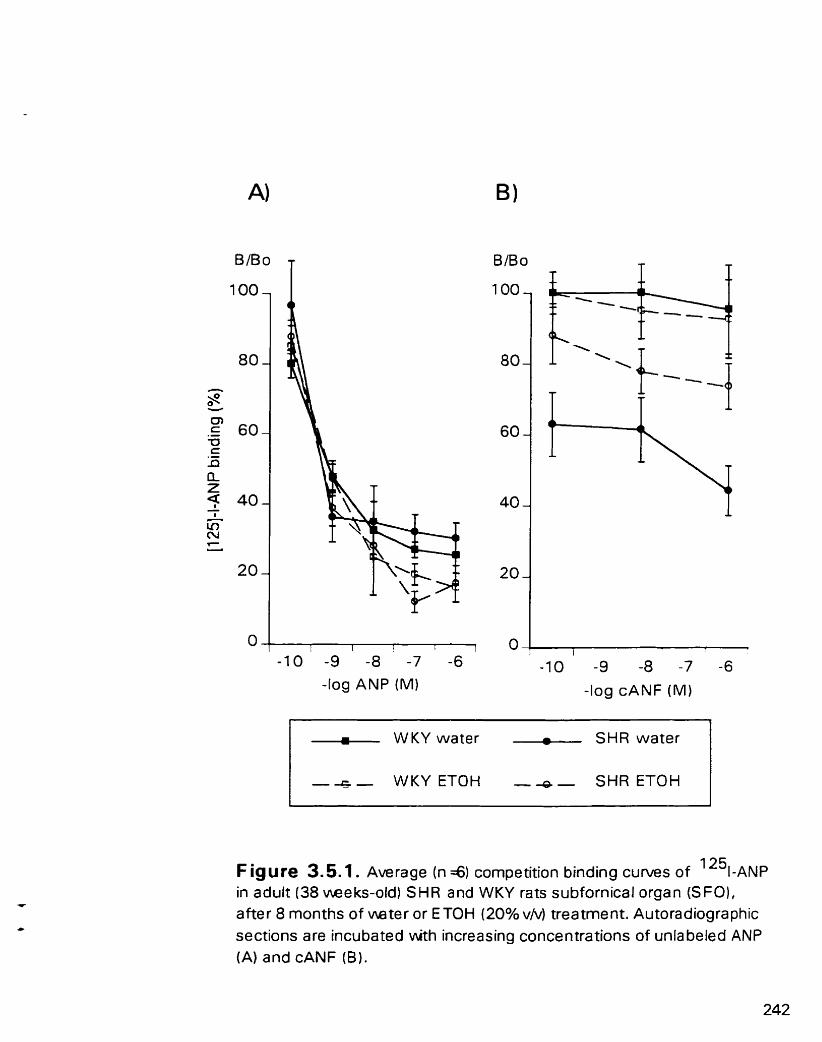
l I I 1 -10 -9 -8 -7 -6
-log ANP (M)
T ?
-10 -9 -8 -7 -6
-log cANF (M)
I WKY water - SHR water
- -C - WKY ETOH - 4 - SHR ETOH
F iguie 3.5.1. Average (n 4) cornpetition binding curves of ' L 3 1 - ~ ~ ~ in adult (38 weks-old) SHR and WKY rats subfornical organ (S FOI, after 8 months of w t e r or ETOH (20% v/v) treatment. Autoradiographic sections are incubated \hith increasing concentrations of unlabeled ANP (A) and cANF (B).

CL 7 e - 40
1
in' CV T L.
20
O I T l I 1 O
-IO -9 -8 -7 -6
-log ANP (M)
1 I 1 I
-10 -9 -8 -7 -6 -log cANF (M)
-- WKY water - SHR water
-4- WKY ETOH 4 SHR ETOH
F igure 3.5.2. Average (n 4) cornpetition binding curves of ' 2 5 1 - ~ ~ ~ in adult (38 m e ks-old) S HR and WKY rats choroid plexus (CP), after 8 months of m t e r or E TOH (20% v/v) treatment. Autoradiographic sections are incu bated mith increasing concentrations of unlabeled ANP (A) and cANF (6).

for the NPR-C receptors of the ETOH-treated animals in the SFO (for SHR rats) and CP
(for both WKY and SHR rats) (Table 3.5.1. ). This result suggests a lower affinity of the
clearance receptors for the circulating natriuretic peptides and the possibility of a greater
proportion of the total plasma natriuretic peptides binding to the active NPR-A. B
receptors .
3.5.4.2.Area (AP)
Mean (n=6) displacement of "51-ANP by cold A N P and cANF (Figure 3.5.3.)
confirms the absence of B, or K, variation between strains (Table 3 S. 1. ). cANF has no
effect in displacing E51-ANP. suggesting the absence of NPR-C receptors in this tissue.
No significant variation is seen between the total ANP receptor characteristics (B,,
and K,) of water- versus ETOH-treated animals (Table 3.5.1 .).
3.5.5. DISCUSSION
The present experiments 1 ) demonstrated significantl y lower natriuretic peptide
receptor binding sites in SFO and CP of S H R cornpared to WKY rats. Furthermore.
rnoditications in the relative proportion of the different ANP receptor subtypes were noted
beween hypenensive and normotensive animals in the two circumventricular organs: and
1) showed a significant reduction by ETOH treatrnent in the affinity of the NPR-C
receptors located in the SFO and CP.
In contrast to the natriuretic peptides found mainly within the brain. most of the
natriuretic receptors in the central nervous system (CNS) are located in circurnventricular
organs. particularly in SFO. CP and AP (Quirion et ai.. 1986. Brown and Czarnecki.
1990a.b). These structures Iack a BBB. a unique situation in the CNS that enables them
to transduce directly hormonal signals from the circulation (for SFO and AP) and the
cerebrospinal tluid (CSF) (for CP) to the neural circuits within the brain. Thus. they
display high concentrations of receptors for the hormones involved in the regulation of
cardiovascular system, such as ANP and angiotensin 11 (AH) (Saavedra et al.. 1987).
S imilarl y. numerous nerve fibers are found connecting these structures to hypothalamic
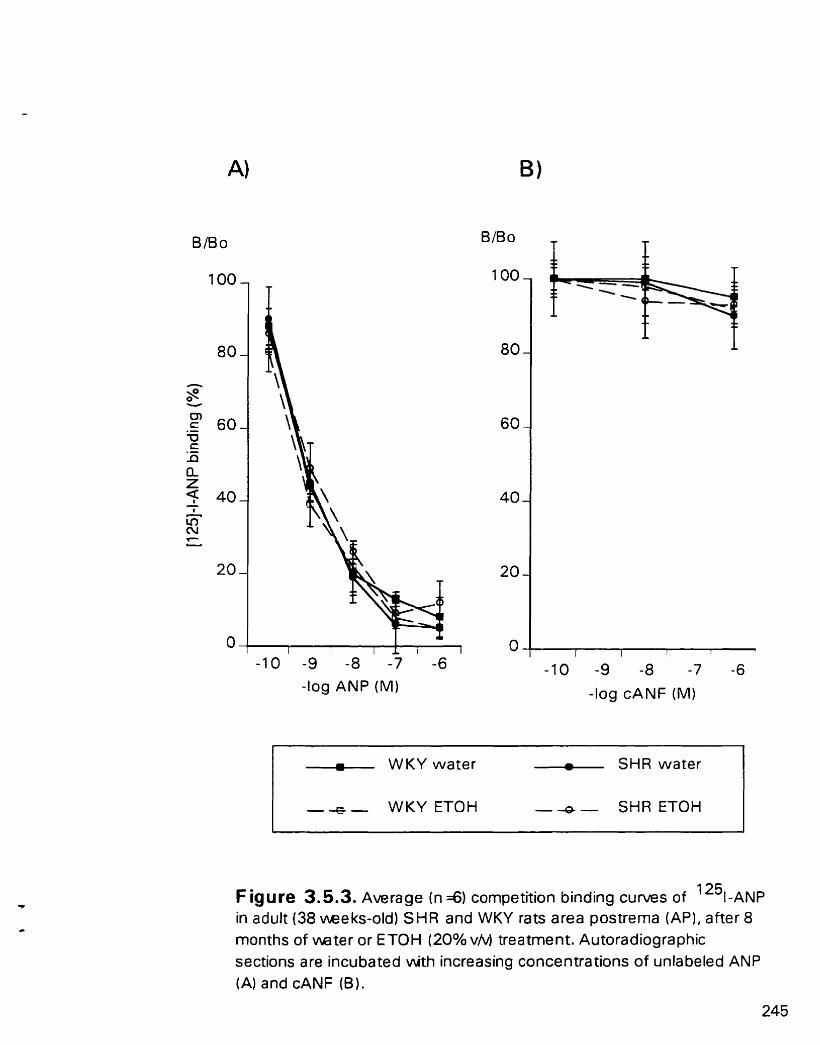
I I I 1 i 1 I
-10 -9 -8 -7 -6
-log ANP (M)
I I 1
-10 -9 -8 -7 -6
-log cANF (M)
I WKY water
- 4- WKY ETOH
.L SHR water
- -8- SHR ETOH
F igure 3.5.3. Average (n 4) cornpetition binding curves of ' 2 5 1 - ~ ~ ~ in adult (38 w e ks-old) S HR and WKY rats area postrema (AP), after 8 months of w t e r or ETOH (20% vM treatment. Autoradiographic sections are incubated increasing concentrations of unlabeled ANP (A) and cANF (BI.
245

or rnedullary cardiovascular centres. such as the AV3V a r a or the NTS-baroretlex region.
For example. Paikovits et al. ( 1992) reported ANP-sensitive nerve fibers and terminals in
the SFO arising from deep into the hypothalamus. Likewise. AH-sens itive neurons have
been shown to connect SFO with the AV3V area (Buggy and Bealer. 1987).
The current hypothesis for the contribution of the brain natriuretic peptides farnily to
BP regulation involves two aspects: the natriuretic receptors present in circumventricuiar
organs and therefore m f e r i n g information from the periphery to the brain. and the ANP
and CNP stores which are located within the cardiovascular centres of the CNS. Brietly,
from the circulation. ANP may bind to specific receptors (in the SFO for example) and
inhibit AH-sensitive cells fibres to the AV3V region (Hattori et al.. 1988). This inhibition
would result in lower stimuli for tluid intake or for AVP secretion from the supra-optic
nucleus (SON). promoting BP lowering (Mangiapane. 1987). Likewise. A N P and CNP
from the hypothalamus rnay directly mediate a depressor action of the AV3V area on BP
(Ku et al.. 1994) and suppress in parts A V P release from SON (Standaert et al., 1987).
Al terations in natriuretic receptor numbers and affinity have been described in
pathophysiological conditions such as hypertension. SHR rats have been reported to exhibit
lower B, for the total natriuretic peptide receptors cornpared to WK Y anirnals in SFO
and CP (Saavedra. 1986. McCarty et ai.. 1986. Brown and Czarnecki. 1990a). but not in
AP (Saavedra et al.. 1987). These results are confirrned by the present studies. Recently.
different natriuretic receptor populations have been suggested to exist in the
circumventricular organs. Konrad et al. ( l992a) described an heterogeneous population of
active nauiuretic receptors (NPR-A and NPR-B). devoid of clearance receptors. in the AP.
This finding is confirmed by the present studies. The SFO has been reported to produce
an hornogeneous population of NPR-A (Himeno er al.. 1992. Brown and Zuo. 1993.
Zorad et ai.. 1993) or NPR-B (Konrad et al.. 1992b). In contrat, the presence of NPR-C
in association with active natriuretic receptors (NPR-A or NPR-B) in various proportions
has been described in the CP (Himeno et al., 1992, Brown and Zuo. 1993. Zorad et al.,
1993). These studies were all perforrned on normotensive anirnals and are in agreement

with the result of the present experiments for WKY rats. However. there are no published
snidies investigating the quantity of the various natriuretic receptor subtypes in conditions
such as hypertension or ETOH intoxication. The results of the present experiments suggest
the existence of a subpopulation of NPR-C receptors in the SFO and an elevated
proportion of NPR-C in the CP of SHR compared to WKY rats. I t is possible that with
high BP and high circulating ANP levels (Gutkowska et al., 1986a). secondary decreases
in natriuretic receptor numbers are seen, but also augmentations in NPR-C receptors to
remove excess ANP from the circulation. It could also be a genetic difference between
SHR and WKY rats, leading to a smaller depressor effect of peripheral ANP to central BP
regulation, and therefore to elevated BP in SHR anirnals.
Long-term moderate ETOH consumption had no effect on the total natriuretic
receptor characteristics (B, and KJ of the three circumventricular organ investigated.
However. the afinity for the NPR-C recepton. evaluated by the displacement of '"1-ANP
by cold cANF. was signiticantly lower in the SFO and CP of ETOH- compared to water-
treated rats. Therefore. it appears that a greater proportion of peripheral ANP rnay bind
to the active NPR-A,B receptors following chronic moderate alcohol consumption. leading
possibly to enhanced signal transduction of plasma (and/or cerebrosp inal fluid) ANP to the
brain cardiovascular centres. Such an effect rnay stimulate ANP-sensitive tïbers and inhibit
AH-sensitive neurons comecting the SFO (andlor the CP) to the AV3V area and produce
some depressor. diuretic, natriuretic and antidipsogenic effects that rnay mediate parts of
the antihypertensive effect of moderate ETOH treacment.
In summary . the significant decrease in natr iuretic recep tor dens ities in hypertens ive
animals rnay be considered secondary to the increased circulating ANP levels. However.
since this reduction in the B, is already present in Young, pre-hypertensive rats
(Saavedra. 86. McCarty et ai., 1986, Saavedra er ai., 1987), it rnay also represent part of
the reason for the elevated BP in SHR rats. The augmented proportion of clearance
receptors in the SFO and CP of hypertensive compared to normotensive rats reported in
the present studies would further diminish any effect of peripheral ANP on the CNS.
Conversely, this rnay also represent a compensatory mechanism to remove from the

circulation the increased plasma ANP levels in SHR rats.
Funhermore. the signifiant decrease in the afi-nity of the clearance receptors located
in the SFO and CP following long-term ETOH administration may contribute. at least in
part. to the prevention of the age-dependent increase in the BP following chronic alcohol
treatment.
3.5.6. ACKNOWLEDGEMENTS
The authors wish to thank Ricardo Claudio for his friendship and excellent care of
the animals. Special thanks are also given to Céline Coderre and Nathalie Charron for their
technical assistance.
P.G. is the recipient of a doctoral scholarship from the " Fonds pour la formation de
Chercheurs et l'Aide à la Recherche" (FCAR). This work was supponed by grants from
the Medical Council of Canada (MRC) (MT- 10337 and MT- 1 1674) (J.G.) and by the
alcohol research program at Douglas hospital ( C . G . ) .

GENERAL DISCUSSION

The present work descnbes the effects of acute and chronic moderate ETOH
consumption on the major components of the natriuretic peptide system (ANP. BNP. CNP.
NPR-A, NPR-B and NPR-C) in the heart, the kidney and the bram. It also coaârms the
existence of a lowering effkt on the BP of long-term moderate alcohol consumption in both
hypertensive and normotensive stragis of rats. Furthemore, the present studies explore a
specific mode1 for the ETOH-mduced preveution of the age-dependent mcrease in BP
observed m both rat and human populations. It thus provides msights into the mteractions of
ETOH with the natriuretic peptide syaem, which mteractions may contniute to the
antihypertensive effect produced by chronic moderate ETOH drinking.
4.1. EFFECT OF ETOH ON THE BLOOD PRESSURE
There is a progressive mcrease with age m BP. particularly in strams of rats with genetic
predisposition to hypertension nich as SHR rats (Howe et al.. 1989). Even though acute
eqosure to moderate concentrations of ETOH did not produce tigdïcant changes in the BP
of either rats (Section 2.1.) or humans (Section 2 - 2 4 the administration of moderate
quantities of ETOH (20% v/v) for 8 months resulted in the partial suppression of the age-
dependent increase in BP in both SHR and WKY rats (Section 3.1. ).
The absence of significant Merences either m body weight mcrease or m liquid
consumption between the water- and ETOH-treated groups indicated that the effect of
chronic moderate ETOH consumption on the BP was not due to secondary d d dehydration.
Thus, the ETOH-induced alterations m BP observed in the present studies agreed with the
majority of the human and animal midies in which an antihypertensive effect of chronic
moderate ETOH consumption has been reported (Section 1.2.2.). Currently, the mechanisms
mediahg this antihypertensive effect of ETOH are not weli understood. The present audies
suggest that the natriuretic peptide f d y may mediate some of the effects of ETOH on the
cardiovascular system.

4.2. EFFECT OF ETOH ON PLASMA A N D HEART NATRIURETIC
PEPTIDES
4.2.1. PLASMA
The administration of a single moderate dose of ETOH ( 1 and 2 g ETOWkg B.W.) by
[.p. mjection resuhed in a rapid but short-lasting mcrease in plasma ANP levels ( Section 2.1 . ).
Circulating ANP lwels were also sigmficantly elevated foilowing the ingestion of a 0.25 and
a 0.50 g/kg B. W. ETOH drink in human volunteers (Section 2.2.).
Ln contrast. the administration of a moderate 20% v/v ETOH solution for 8 months in
WKY and SHR rats was associated with significantly lower iirculating ANP (Section 3.1. )
and BNP (Section 3.2.) Ievels compared to water-treated controls.
Based on experirnental evidence dernonstrating (a ) that acute ETOH exposure induces
a transient increase m the release of natriuretic peptides fiom the heart and (B) that following
mcreases or decreases m BP there are hemodynamic modifications which would respectively
increase or decrease the release of cardiac natnuretic peptides. two hypotheses may br
proposed to explain the low content of natnuretic peptides in the plasma of rats treated
chronically with moderate amounts of alcohol:
( 1 ) According to the fïrst hypothesis. the low levels of circulating natriuretic peptides in rats
treated chronicaNy with ETOH are a direct consequence of the significant decrease in systolic
B P and the associated bemodynarnic modifications reflecting lower requirements for
circulating ANP and BNP levels and thus !ower rates of cardiac release.
(2) According to the second hypothesis. the low levels of circulating natriuretic peptides are
due to either the absence or low levels of ETOH in the circulation during the light phase of
the dady cycle d e n the estimation of plasma naaiuretic peptides were perforrned. During the
Light phase of the daily cycle. rats are not eating and drinking actively. thus the BAC is low
and there is no direct stimulation of the cardiac tissue by ETOH to increase the release of the
natnuretic peptides. in the absence of a direct stimulatory effect of ETOH on the cardiac
tissue. the hemodynamic modifications produced by the significantly lower BP predominate
and. as a consequence, there is a lower rate of ANP and BNP release and lower plasma ANP

and BNP Iwels. It is possible that during the dark phase of the daily cycle when the animals
are drinkmg and eating actively and when the BAC is hi&. the stimulatory effect of ETOH
on the cardiac tissue may predonmiate leadmg to enhanced release of natriuretic peptides and
higher contents of ANP and BNP m the circulation. The absence of down-regulation of the
atrial and ventncular ANP and BNP systems. as a logical consequence of the prolonged
decrease in BP. is in support of this hypothesis.
4.2.2. HE ART ATRL4
A schematic diagram of the heart ANP and BNP systems is presented on figure 4.2.1.
The translation of ANP and BNP mRNAs in the atnal tissue produces ANP and BNP
prohomones which are then stored in specific granules. The ANP contained in atrial gantdes
is the main contributor to cuculating ANP levels (Flynn et al.. 1983). Unlike ANP. BNP is
mostly produced by the heart ventncles (Ogawa et al.. 199 1). Nevertheless. BNP is also
colocalized with ANP m certam aûial granules (Thibault et al.. 1992). With the development
of high BP. such as in SHR rats. a t d ANP mRNA and total BNP content are increased.
resulting in higher plasma levels of natriuretic peptides (Sections 3.1. and 3.2.).
The administration of a single moderate dose of ETOH ( 1 and 2 g ETOWkg B.W.) by
i -p . injection resulted in a rapid decrease in lefi and right atrial content (Section 2.1.). In
contrast. following 8 months of continuous ETOH (20°h v/v) administration. there was a
sign5cant increase in both lefi and rigbt atnal ANP content and concentration in WKY and
SHR rats (Section 3.1 .). Similarly, ETOH also increased atnal BNP levels in WKY rats and
lefi atrial BNP content and concentration m SHR rats (Section 3.2.). There was no signifïcant
daerence in atrial ANP or BNP mRNA levels between the water- and the ETOH-treated
animals of both strains.
Based on our nudies demonstrating that acute. i j i vivo. exposure to ETOH induces a
transient mcrease m plasma ANP levels (retuming to basal levels within 2 hours post-ETOH
admbhration) as a consequence of a short-lasting increase in ANP release. and considering
the eating and drinking pattern of rats during the daily cycle (the eating and drinking occur
during the dark phase of the daiiy cycle), the a c t ~ t y of the atrial natriuretic peptide system
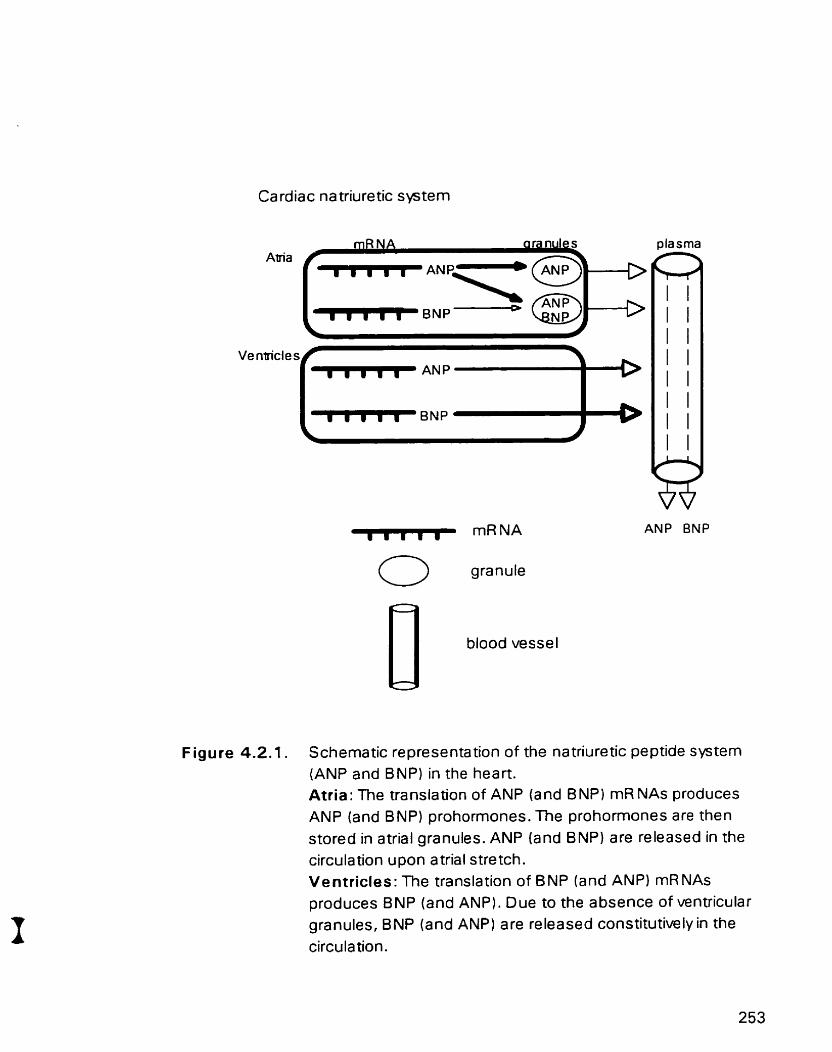
Ca rdiac natriuretic system
Ve nvicles ANP
4- mRNA ANP BNP
0 granule
F igu re 4.2.1 . Schematic representa tion of the natriuretic peptide system (ANP and BNP) in the heart. Atria: The translation of ANP (and BNPi mR NAs produces ANP (and BNP) prohormones. The prohormones are then stored in atrial granules. ANP (and BNP) are released in the circulation upon a trial stretch. Ventricles: The translation of BNP (and ANP) mRNAs produces BNP (and ANP). Due to the absence of ventricular granules, B NP (and ANP) are released constitutively in the circulation.

may be descnïed as outlined in Figure 4.2.2.:
( 1 ) During the dark phase of the daily cycle. the animals are active and thus eating and
conaiming sigaificant amounts of ETOH. Accordingly, BAC levels are hi&. The release of
ANP and BNP rnay be stirnulated by alcohol. leacüng to transient increases in plasma
natriuretic peptide levels and stimulating peptide and mRNA çynthesis in the atria. The only
other published study investigatmg the effect of chronic (6 weeks) moderate ETOH
consumption on the hem ANP system has reported a high BAC ( 172.0 * 13.9 mg/dl) (Wigle
et al.. 1993b). Interestingly. this hi& BAC level was associated with a si@cant increase
in atrial ANP mRNA levels.
(2) hiring the light phase of the daily cycle. the animals are inactive and thus are not eating
or consuming sigdicant amounts of ETOH. Accordmgly. BAC levels are low (28.92 c 12.25
and 39.23 * 18.97 mg/dl for the WKY and SHR rats of the present studies). Therefore. there
is no direct stimulation of ANP and BNP release fiom the atrial tissue by ETOH. so that the
release of the two natriuretic peptides is controlled mainly by the hemodynamic mechanisms.
Since the BP is lower in the ETOH- compared to the water-treated animals. the release of
ANP and BNP is lower. resulting in lower plasma levels and the increase in the accumulation
of ANP and BNP peptides in the avial granules. Iadeed. the higher accumulation of ANP and
BNP in the atria of the ETOH-treated animals compared to the water-treated controls. and
the finding that the contents of atrial ANP and BNP rnRNA are similar in the atria of the
ETOK and water-treated rats indicates that despite the low BP the atnal ANP and BNP
systems are not down-regulated by the chronic moderate ETOH e.xposure. It may then be
proposed that in the ETOH-treated animals when the BAC is high (usually during the dark
phase of the daily cycle), alcohol either directly or indirectly through its effects on vanous
hormonal and neurotransmitter systems. stimulates the release and synthesis of atrial ANP and
BNP (as seen by Wigle et al.. 1993b) and thus maintams the content of ANP and BNP mRNA
in the atria to the same levels as those in the atria of the water-treated animals which evhibit
a significantly higher BP. It rnay be suggeaed that in the water-treated rats. ANP and BNP
synthesis and release are higher probably because of the high BP. whereas in the ETOH-
treated rats. the atrial ANP and BNP systems are stirnulated to a higher level of a c t ~ t y by
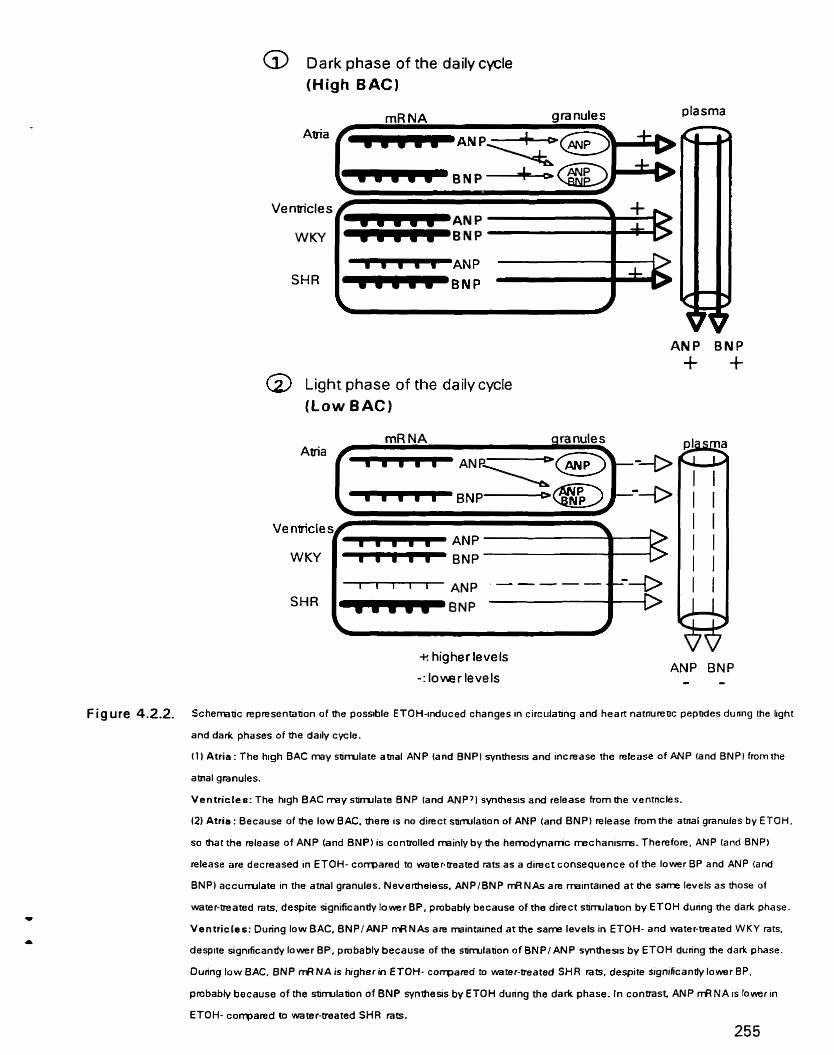
@ Dark phase of the daily cycle (High BAC)
ANP B N P + + a Light phase of the daily cycle (Low BAC)
-
Ve nmcle s I- ANP WKY - BNP
p --- --.-=-O f-b BNP
+ hig he r leve Is ANP BNP
-: lover levels - -
Fig ure 4.2.2. Schemuc representation of the possible ETOH-induced changes in circulating and heart natnurenc pepbdes dunng the Iight
and dark phases of the daily cycle.
(1 ) Atria: The high BAC m y sumlate amal ANP (and BNP) synthesis and increase the release of ANP (and BNPI fromlhe
atnal granules.
Vent r ic les : The high BAC may st imlate BNP (and ANP?] synthesis and release from the ventncles.
12) Atria : Because of the low BAC. there is no direct stimulation of ANP (and BNPi release from the atnal granules by ETOH,
so that the release of ANP (and BNP) is controlled mainly by the hemdynamc mechanism. Therefore. ANP (and BNPI
release are decreased in ETOH- conpared to water-îreated rats as a direct consequence of the lower BP and ANP (and
BNPi accumilate in the amal granules. Neverttieless, ANPlBNP mRNAs are maintained a t the same levels as those of
water-mated rats, despite significandy lower BP, probabty because of the direct stimlaaon by ETOH dunng the dark phase.
Vent r ic les : During low BAC, BNPlANP M N A s are rmintained at the s a m levels in ETOH- and water-treated WKY rats,
despite significandy lower BP, probably because of the sumlation of BNPIANP synthesis by ETOH during the dark phase.
Oudng low BAC, BNP mR N A is higher in ETOH- conpared to water-ireated SHR rats, despite signrficandy lower BP.
probably because of the stin-ulation of BNP synthesis by ETOH during the dark phase. In conûast ANP rrClNA 1s lower in
ETOH- conpared to water-treated SHR rats.
255

a different mechanism than hemodynarnic changes, probably by ETOH. Thus, it may be
hypothesized that this enhanced potential actinty of both atrial ANP and BNP systems is
produced by the repetitive stimdatory effects of the daily ETOH consumption.
4.2.3. aEART VENTRICLES
The schernatic diagram of the ventricdar ANP and BNP systems is presented on figure
4.2.1. Due to the absence of grandes, the translation of ventncular ANP and BNP &As
produces mature natriuretic peptides that are released constitutively in the circulation. in
contrast to atrial ANP and BNP release (Lang et al.. 1992). The m m site of ANP production
in the body is the heart atria and 95% of the circulatmg ANP is produced by the atnal tissue
uuder normal conditions (Ogawa et al.. 199 1 ). However, with the development of hi& BP
and venaicular hypertmphy m the SHR rats. this contribution decreases to 60%. In contrast.
the ventrkular tissue contniutes to more than 60% of total circulatmg BNP levels under
normal conditions (Ogawa et al.. 199 1 ). This contnbution is fùrther increased m certain
pathophysiological conditions, such as hypertension (Yokota et al., 1993).
The admini_ctratiotl of a single moderate dose of ETOH ( 1 and 2 g ETOHkg B. W.) by
i.p. mjection resulted m a dekyed increase m ventficular ANP levels (Section 2.1 .). Following
chronic ETOH treatment, total ventncular ANP content and ventricular ANP mRNA were
sigaificantly lower in SHR, but not WKY rats (Section 3.1.). in contrast, ventricular BNP
conceniration and BNP mRNA levels were significantly increased in S m but not in WKY
rats (Section 3.2.).
Based on our studies demonstratmg that acute ETOH consumption induces a transient
hcrease m plasna ANP levels and a delayed mcrease in ventncular ANP content, and taking
into consideration the eating and drslkmg pattern of rats during the daily cycle, the mode1
proposed for the effect of ETOH on the atrial natriuretic peptide systems may be expanded
to include the ventncular natriuretic peptide systems (figure 4.2.2. ):
(A) WKY rat : The chronic ETOH treatment produced no aherations in ANP and BNP
contents or in ANP and BNP mRNA lwels, even though the BP was significantly lower in
the ETOH-compareci to the water-treated animals for at least a month pnor to sacrifice. The

fàct that a decrease m the act-ivity of the ventricular BNP and ANP qstems was not observed
in the ETOH-treated WKY rats mdicates that chronic ETOH treatment maintained the
potential actMty of the ventricuiar natriuretic peptide systems to the same levels as those of
water-treated WKY animals exhibithg higher BP. Thus, t may be hypothesized that during
the dark phase of the daily cycle, each ETOH drmking episode produces an mcrease m
ventricular ANP and BNP release, leadhg to an mcrease m ventricular ANP and BNP mKNA
b e l s and ANP and BNP biosynthesis. During the light phase of the cycle, the hemodynamic
fàctors predominate and the ventricular natriuretic peptides are not stimulated. In agreement
with this model, the study of Wigle and colleagues ( 1993b), performed in conditions of high
BAC, demonstrated Sgnificant elevations in both ventricular ANP mRNA and ANP content
following the administration of moderate ETOH b e l s for 6 weeks.
(B) SHR rats: The chronic ETOH consumption prevented the development of ventricular
hypertrophy in SHR rats. Indeed, the total ventricular protein content is sigiilficantly lower
in ETOH-compared to water-treated SHR rats, mdicatmg the absence of ventricular
hypertrophy. Thus, the significant increase in ventricular %NP &A levels suggests a
specific effect of ETOH mice the Iower BP of ETOH-treated SHR rats prevents the
amibution of this effect to compensatory mechanisms due to elevated BP. Therefore, it may
be hypothesized that during the dark phase of the daily cycle, the ETOH consumption
Stimulates venhicular BNP activity and thus BNP mRNA levels, producing transient increases
in circulating BNP levels. Interestingly, elevated plasma BNP levels have already been
observed following short-term chronic ETOH drinking, when the animals were sacrifïced
during high BAC conditions (Wigle et al, 1993~). Chrooic stimulation of the ventricular BNP
system by ETOH may then maintain higher basal levels of ventricular BNP mRNA in ETOH-
treated SHR rats as suggeaed by the higher BNP mRNAs, compared to water-treated SHR
rats which have higher BP. In contrast to BNP, the contniution of ventricular ANP content
to the plasma ANP levels must be minimal in ETOH-treated SHR rats. The sigdcant
decrease in ventricular ANP content and ANP mRNA levels shouid reflect the lower
ventncular mass due to ETOH. Therefore, any possible stimulatory effect of ETOH on the
ventricular ANP system of SHR rats may be masked by the sigdicant decrease in ventricular

4.3. EFFECT OF ETOH ON RENAL NATRKTRETIC RECEPTORS
The natriuretic and diuretic effects of circulating (ANP and BNP) and intrarenal (UD)
natriuretic peptides are mediated by receptors located in the glomenili (NPR-A and NPR-C)
and the coilecting duct of the b e r medda (NPR-A) (Figure 4.3.1. ):
(A) in the glomenili NPR-A receptors are considered effective mediators of the physiological
effects of natriuretic peptides through the activation of the secondary rnessenger. cGMP
(Kolier and Goeddel, 1992). Natriuretic peptide binding to tbese so-called active glomemlar
aatriuretic receptors (NPR-A) has been açsociated with the relaxation of giomemlar mesangial
ceils and with the expansion of the capillary surface area for filtration. producing a modest
increase in GFR (Singhal et al.. 1989). The relaxation of glomedar affereat anerioles and
the constriction of giomerular efferent anerioles produced by the binding of natriuretic
peptides to active natriuretic receptors also increase GFR Ma enhanced glomemlar capillary
hydraulic pressure (Marïn-Grey et ai.. 1986: Kumira et al.. 1990). in contraa. the exact role
of the NPR-C receptors is unclear. Any fuoction beside the weil-defined "clearance" role
remaius highly speculative (Maack et ai.. 1993).
( B ) An homogeneous NPR-A receptor population exias in the CM (Martin et al.. 1989). The
binding of natriuretic peptides to this receptor has been associated with the suppression of
sodium reuptake via amiloride-sensitive sodium channels (Sonnenberg et al.. 1986: Zeidel et
al., 1988) and with the inhibition of the antidiuretic effect of AVP (Zeidel et al.. 1987).
Interestingly, significant increases in IM natriuretic peptide binding sites have been obsewed
in hypertensive animais. suggesting increased NPR-A a c t ~ t y to compensate for the higher
BP (Nuglozeh et al.. 1990; Bonhomme and Garcia. 1993).
Chronic moderate ETOH drinking is associated with a significant reduction in total
glomedar natriuretic peptide bmding sites (Section 3.3. ). Furthemore. quantification of the
various receptor subtypes suggeaed a significant decrease in glomerular NPR-C (or
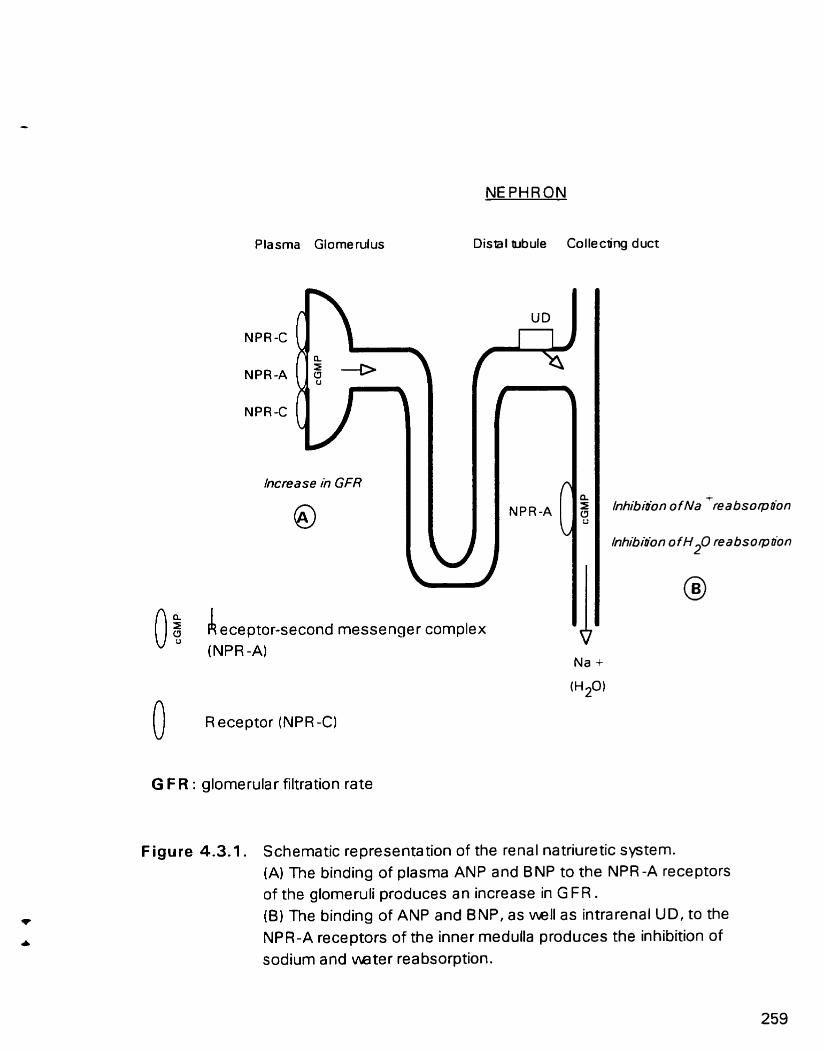
Plasma Glomerulus
NPR-C k NPR-C
""-A F lncrease in GFR
NEPHRON
Distal tubule Collecb'ng duct
O $ k eceptor-second messenger cornpiex (NPR -A)
NPR-A
O R eceptor (NPR-C)
G F R : glomerular filtration rate
Inhibi~on ofNa \eabsorption
Inhibibon o f H 2 0 reabsoipbon
Figure 4.3.1. Schematic representation of the renal natriuretic system. (A) The binding of plasma ANP and BNP to the NPR-A receptors of the glomeruli produces an increase in G FR. (5) The binding of ANP and BNP, as vudl as intrarenal UD, to the NPR -A receptors of the inner medulla produces the inhibition of sodium and w t e r reabsorption.

clearance) receptors, but not m glomenilar NPR-A receptors. Chronic ETOH consumption
is also associated with a significant mcrease m b e r medullary (IM) natriuretic peptide
binding sites (NPR-A) ( Section 3.3. ).
Based on our snidies demonstrating ETOH-induced changes in renal natriuretic peptide
receptors (NPR-A and NPR-C), a mode1 illustrating a possible role of the renal natriuretic
system m the antihypertaisive ef5ect of moderate alcohol connunption is proposed on Figure
43.2:
( 1 ) in the glomeruk the specific dom-regdation of NPR-C receptors suggests that a greater
proportion of circulating ANP and BNP levels may bind to the active receptors (NPR-A).
aUowing a greater glomerular effect at any given concentration of circulating natriuretic
peptides. It has been lmown that various stimuli such as salt Ioad or hypertension specifically
dom-regulate NPR-C receptors in glomenilar tissue in order to reduce the plasma ANP and
BNP butTering by NPR-C and to allow an enhanced renal diuretic and natriuretic effect
through the NPR-A receptors (Ogura et al.. 1989: Kollenda et al.. 1990). However. the
demonstration of a selective dom-replation of glomerular NPR-C receptors by ETOH.
despite the lower BP m these animals compared to water-treated rats. niggests that this effect
is not due to secondary hemodynarnic characteristics. Thus, the physiological consequence
of the effect of ETOH on the glomerular tissue may be enhanced GFR and as consequence
some antihypertensive effects.
(2) The demonstration of higher IM natriuretic peptide binding sites in ETOH-treated rats.
despite the sipificantly lower BP. suggests a specific effect of ETOH in renal tissue. Thus.
circulating ANP and BNP reaching the IM (as weU as intrarenal C I D ) have the possibility to
produce enhanced inhibition of sodium reabsorption by the collecting ducts in ETOH-
compared to water-treated rats.
The ETOH-induced modifications in the kidney. such as the increased NPR-A (LM) and
decreased NPR-C (glomedi), suggest enhanced diuretic and natriuretic effects. which may
lead to a long-lasting decrease of the BP. However, to assig. any functional significance on
the ETOH-induced changes in the renal natriuretic peptide receptors, the increase in the
relative proportion (glomeruli) and number (IM) of NPR-A must be associated with increased
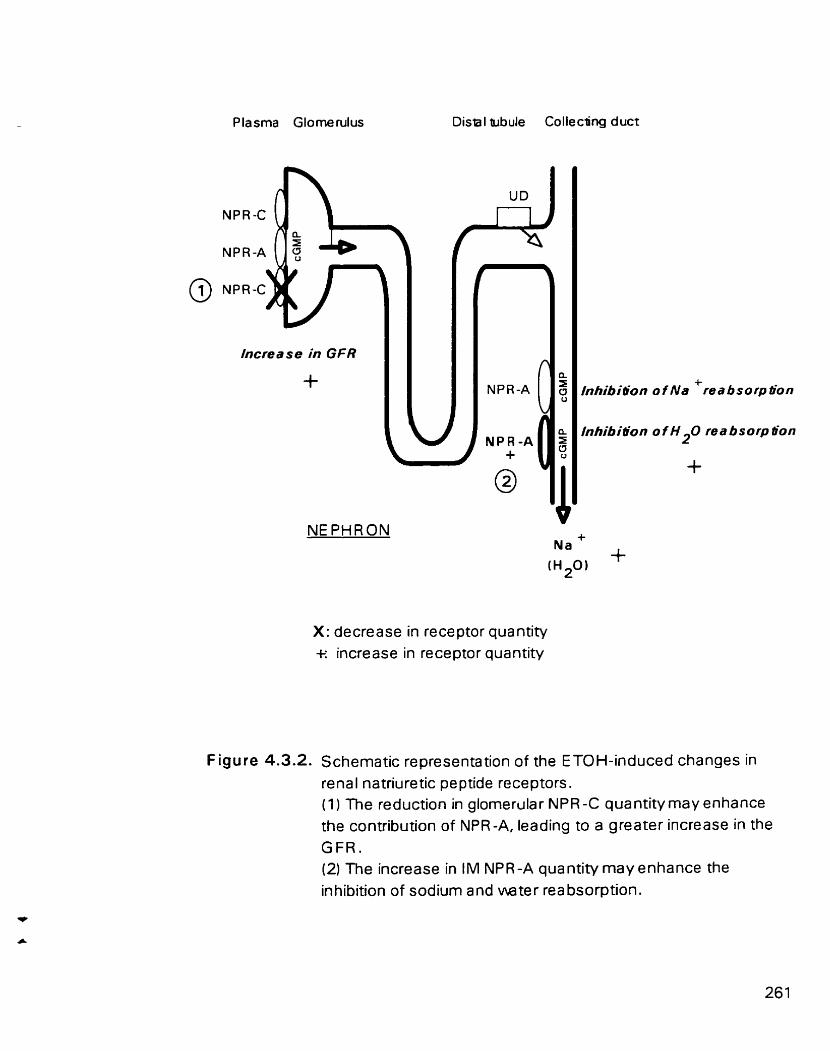
Plasma Glomenilus Disal tubule Cokcbng duct
Increase in GFR
+
NEPHRON
7
NPR-A
NPR-A +
( Inhibi~foon o f H20 rea bsorp don
+
X: decrease in receptor quantity +: increase in receptor quantity
Figure 4.3.2. Schematic representation of the ETOH-induced changes in
renal natriuretic peptide receptors. (1) The reduction in glomerular NPR -C quantity ma y enhance the contribution of NPR-A, leading to a greater increase in the GFR. (2) The increase in IM NPR-A quantity may enhance the inhibition of sodium and vlater rea bsorption.

urinary excretion of cGMP in the ETOH-treated mimals. hterestmgly. there is either no
change (WKY) or an mcrease (SHR) in the h a r y excretion of cGMP in ETOH- compared
to water-treated rats. despite the reduced plasma levels of ANP and BNP observed during the
light phase of the daily cycle as a consequence to the secondary bernodynamic effects
associated with the lower BP (Section 4.2.). This suggests that a sirnilar level of circulating
natriuretic peptide would have a greater effect in ethanol- compared to water-treated SHR
rats at the level of the kidney. because of the ethanol-mediateci modifications of the natnuretic
peptide receptors in the glomenili and the huer medulla. In SHR rats. these receptor
alterations are sdicient to produce higher levels of urinary cGMP during the light cycle. at
a time when the plasma ANP levels are lower. In WKY rats. despite the Iower circulating
ANP and BNP levels, the urinary cGMP levels of the ETOH-treated rats are similar to those
of the water-treated WKY rats which exhibit higher circulating ANP and BNP levels.
indicatmg a functional significance of the ETOH-induced changes of the renal NPR-A
receptors. These r e d s support the theory that the ETOKinduced modifications in the renal
natriuretic receptors may contribute to the antihypertensive effect of long-term moderate
alcohol consumption. The potential signficance ofthe ETOH-induced changes in the renal
natriuretic peptide receptors rnay be even more important during the dark phase of the daily
cycle. when as a consequence of the higher BAC the plasma ANP and BNP levels rnay be
elevated. inducing a further increase in cGMP production and natriuresis. and a tùrther
decrease in BP.
4.4. EFFECT OF ETOH ON BRADJ NATMURETIC PEPTIDES AND
RECEPTORS
Under normal conditions, the basal a c t ~ t y of the brain natriuretic peptides syaem is
detennined by the BP. Two dinerent pathways have been postulated to influence central
natriuretic peptide levels and may serve as sensors of the BP (Figure 4.4.1.):
(A) Natriuretic peptide receptors located in the subfomical organ (SFO) and area postrerna
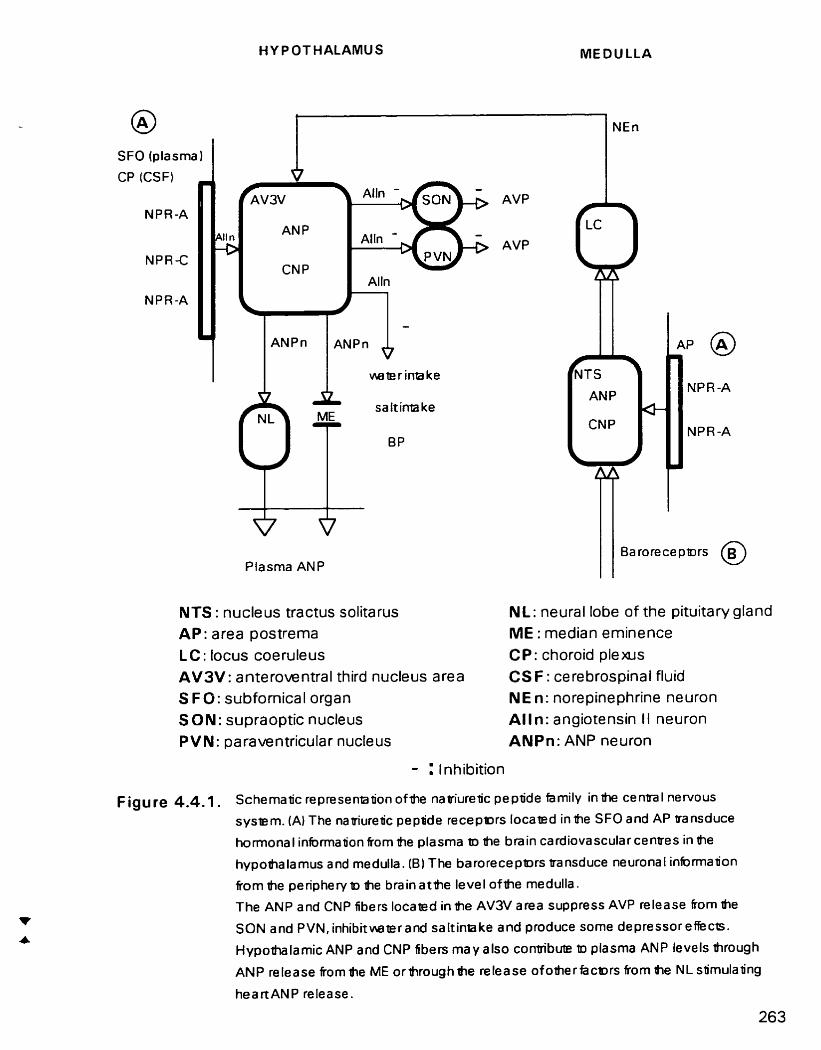
HYPOTHALAMUS MEDULLA
CP (CSFI
NPR-A
NPR-C
NPR-A
Plasma ANP
AVP
AVP
I l Ba roreceptors 0
NTS : nucleus tractus solita rus NL: neural lobe of the pituitary gland AP: area postrema ME : median eminence LC: locus coeruleus CP: choroid plexus AV3V: anteroventral third nucleus area CS F : cerebrospinal fluid S FO: subfornical organ NE n: norepinephrine neuron S ON: supraoptic nucleus All n: angiotensin II neuron PVN: paraventricular nucleus ANPn: ANP neuron
- I Inhibition
Figure 4.4.1. Schemaîic represeniation ofthe naüiurebc peptide farnily in the central neivous
sysem. (A) The naüiuretic peptide recepcrs located in the SFO and AP aansduce
hormonal inbrmation fiom bie plasma a, the brain cardiovascular centres in the
hypothalamus and medulla. (B) The barorecepmrs tansduce neuronal information
fiom the peripheryto the brain atihe level ofthe medulla.
The ANP and CNP fibers located inthe AV3V area suppress AVP release from the
SON and PVN,inhibitwterand saltintake and produce some depressoreffects.
Hypoihalamic ANP and CNP fibers mayalso conûibute î~ plasma ANP levels through
ANP release from lhe ME ordwough the release ofother facmrs fiom the NL siimulabng
heanANP release.
263

( AP), two circumventncular organs lackmg a blood-brain-bamer ( BBB ). transduce diuect ly
hormonal information fiom the circulation to the cardiovascular centres in the hypothalamic
AV3V area and meduliary nucleus tractus solitarü (NTS). respect~ely.
(B) Neuronal Somation ftom the baroreceptors in the nght atria. carotid and aortic suiuses
and in the kidneys are also reachhg the brah through the medulla and the NTS. The impulses
produced by the NTS fiom baroreceptor- and AP-mediated informations activate the locus
c o d e u s (LC) which m tm sends fibers of noradrenergic neurons (NEn) to the AV3V area
of the hypothalamus.
Interestin&. the NTS area is nch m ANP neurons (Kawata et al., 1985). Micromjections
of ANP into that region or into adjacent areas such as the ventrolateral medulIa have also
been associated with important decreases in BP ( McKitrick and Calaresu. 1 988: Bergaglio
and Calaresu, 1990). It seems then that medullary ANP may be involved in BP lowering by
interfering with the noradrenergic neurons and baroreceptor reflex since afferent
baroreceptor fibres terminate in the NTS.
Both the neuronal inputs fiom the medulla and the hormonal information fiom the SFO
(and the choroid plexus) terminate in the A V W area of the hypothalamus. This region is
central to the control of fluid intake and fluid excretion (Buggy and Bealer. 1987).
interestingly. this area bas also the highest concentration of ANP fibres in the brain (Kawata
et ai.. 1985). Intracerebroventncular (i.c.v.) injections of ANP have produced various
antidrpsogenic. dituetic and depressor effects which are believed to be mediated by the A V W
area. as suggeaed by a number of studies such as: (a) Centrally administered ANP
significantly inhibited the water intake d u ~ g water deprivation and NI-induced drinking
(Nakamura et al.. 1985: Antunes-Rodngues e t d . 1985; Katsuura et al.. 1986). ( b ) The i.c.v.
injection of ANP also reduced sigrilficantly the salt appetite during salt deprivation or in
mains usually know to have elevated salt appetite. such as SHR rats (Fitts et ni.. 1986: Itoh
el al.. 1986b). (c) Microinjections of ANP around the hypothalamus produced significant
increases in water excretion. possibly through the inhibitory effect of ANP on AVP release
f?om the supra-optic (SON) and paraventricular (PVN) nuclei (Israel and Barbella. 1986: Lee
et aL, 1989; houe et al., 1990). (d) 1.c.v. mjected ANP also signiticantly inhibited the pressor

action of ic.v. mjected An m conscious. unrestrained rats (Itoh et ai.. 1986a: Shimini er al..
1986; Castro et al., 1987).
Severai hypothalamic ANP neurons ( A m ) terminate in the median emhence (ME) or
the neural lobe of the pituitary gland (NL). probably leadmg to the release of natriuretic
peptides into the vasculature drainhg these two organs. The contribution of hypothalamic
ANP to circulating ANP levels is certaidy minimal. considering the low brain ANP levels
compared to those found in the heart atria. However. it is possible that the activation of
hypothalamic efferent ANP neurons to ME and NL produces the release of other factors fiom
the pmiitary gland. such as oxytocin (Haanwiuckel et al.. 1995), a d o r the activation of other
neuronal pathways to the heart. leading to the synthesis and release of ANP and BNP fiom
the heart atria and ventricles.
Following chronic moderate ETOH connimption. there is a significant increase in
hypothalamic. pontine and medullary ANP levels in SHR. but not WKY rats (Section 3.4. ).
Brain CNP levels are also elevated in SHR-ETOH rats and in the hypothalamus and medulla
of WKY-ETOH animals. Furthemore. chronic ETOH treatment significantly increased the
& of NPR-C receptors in the SFO (for the SHR rats) and CP (for both WKY and SHR rats).
but not in the AP (Section 3.5. ).
( 1) It rnay be speculated that elevated meduilary ANP levels produced by chronic moderate
ETOH consumption in SHR rats d o w a greater and prolonged inhibition of the baroreceptor
reflex. so that the reaing BP is calibrated to a lower level (Figure 4.1.2. ). it is also possible
that the increased concentration of medullary ANP levels in ETOH-treated rats enhances the
impulses of noradrenergic neurons to the hypothalamic ANP areas of the AV3V area.
promoting a greater antidipsogenic. diuretic and depressor effect.
(2) The significantly higher ANP content and concentration in the liypothalamus of SHR rats
following chronic moderate ETOH consumption may contribute to the ETOH-mediated
prevention of the age-dependent hypertension by enhancing the hhibitory effect on water and
sait mtake and the stimulatory effect on water and salt excretion by the ANP released in the
A V W area. Higher brah ANP levels following ETOH may also enhance the impulses of
hypothalamic efferent ANP neurons to the neurohypophysis and the median eminence.
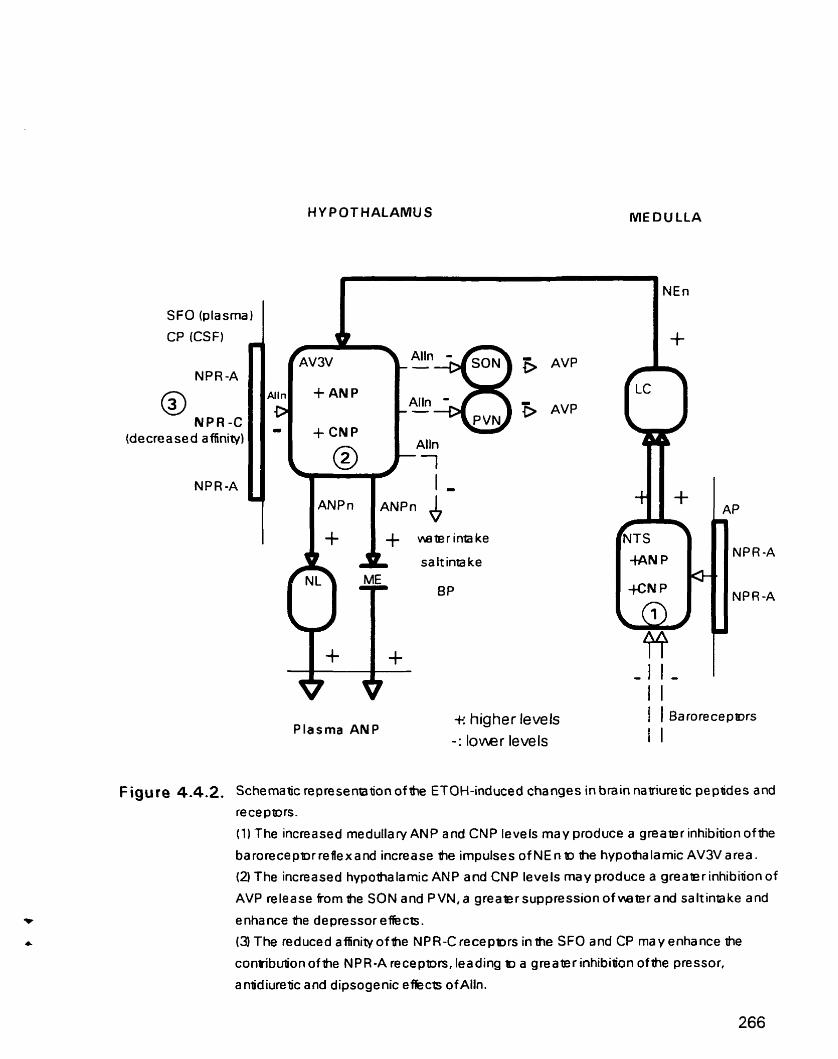
HYPOTHALAMUS MEDULLA
SFO (plasmi
CP (CSFI
NPR-A
O N P R - C
NPR-A
AVP
AVP
+: higher levels Plasma ANP
-: i o w r levels
NPR-A
NPR-A
- I I - I I I 1 1 Barorecepmrs
I I
Figure 4.4.2. Schematic represenation o f h ETOH-induced changes in brain natriureic pepldes and
recepmrs.
(1) The increased medullary ANP and CNP IeveIs ma y produce a greaaer inhibibon ofthe
barorecepmrreflexand increase itie impulses ofNEn lo the hypothalamic AV3V area.
(2 The increased hypothalamic ANP and CNP levels may produce a g r e a ~ r inhibition of
AVP release fiom the SON and PVN,a greaorsuppressionofwoerand saltinmke and
enhance the depressor efkcts.
(3) The reduced affmityofihe NPR-Crecepmrs inthe SFO and CP may enhance the
conûibuiion ofttie NPR-A recepmrs, leading m a greaber inhibition ofthe pressor,
antidiuretic and dipsogenic e f k c ~ ofAlIn.

promoting higher basal levels of heart ANP and BNP synthesis and mRNA levels. despite the
lower BP (Figure 4.2.2.).
(3) The significant decrease m the affinrty of the NPR-C receptors m the SFO and CP
foUowing long-term alcohol consumption may slow a greater proportion of the natriuretic
peptides fiom the circulation (and/or the cerebrospinal fluid) to bind to the "active" NPR-AB
receptors. leading possibly to enhanced inhiibitioo of the pressor. antidiuretic and dipsogenic
effects mediated by the AI1 fibers (Figure 42.2.).
The ETOH-induced changes in brain CNP levels may also mediate some of the
antihypertensive effects of rnoderate ETOH consumption. Interestingly. CNP and CNP
mRNA have been isolated from the hypothalamus, particularly fiom the AV3V area ( H e m
er al.. 1993). However. hi& quantities of CNP have also been found in areas unrelated to
cardiovascular controL such as the cerebelium (Komatsu et al.. 199 1 ). Consequently. the
siflcance of the CNP changes in the mediation of the antihypertensive effect of ETOH are
difficult to assess until it is known to which extent central CNP may contribute to the
cardiovascular regdation.
4.5. GENERAL S U M M M Y AND FUTURE STUDIES
Based on our results for the ETOH-induced alterations in circulating, cardiac. brain and
renal natriuretic peptides and receptors. a tentative general pathway for the lowering of the
BP by alcohol may be outlined (Figure 4.5.1 .). The direction of most of the modifications in
the natriuretic peptide system foster the hypothesis that this hormonal family may mediate.
at least in part. the antihypertensive effect of chronic moderate ETOH consurnption.
( 1 ) Acute moderate ETOH consumption may increase circulating ANP and BNP levels.
During long-term alcohol treatment. this condition may occur during the dark phase of the
daily cycle, associated with high BAC.
(2) The elevated plasma natriuretic peptide levels results probably £?om the release of ANP
and BNP fiom the cardiac tissue. The greater ANP content accumulated in the atnal granules
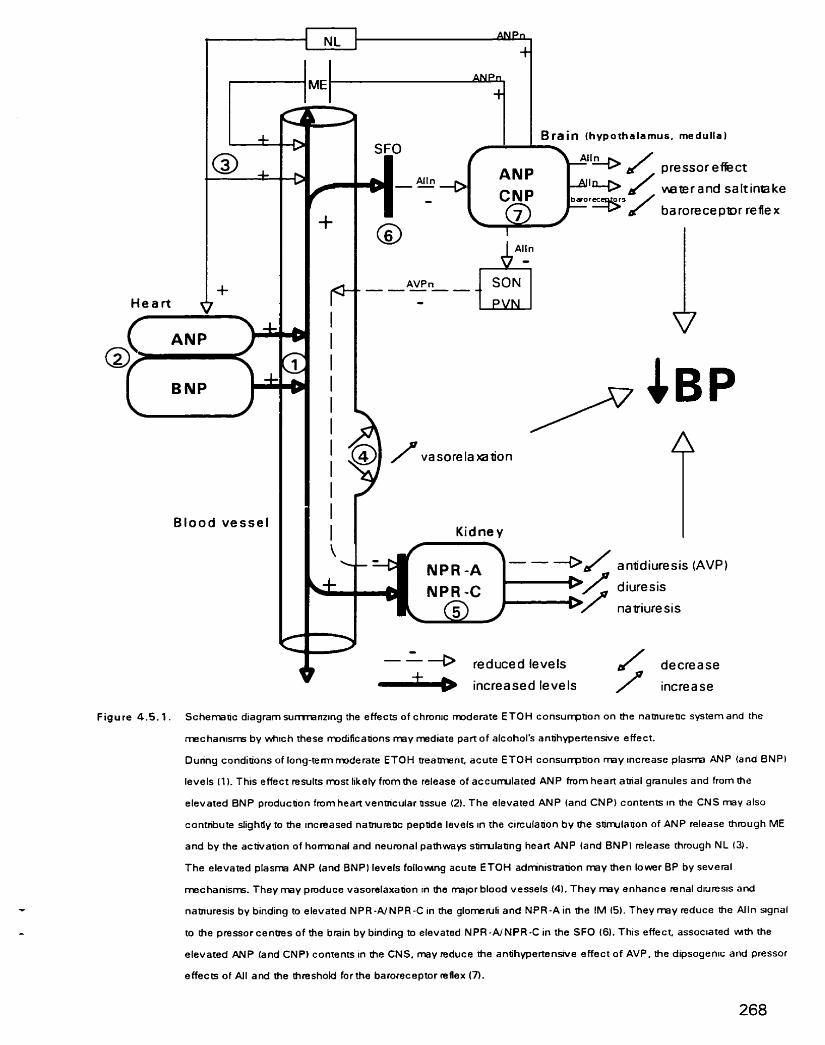
Heart
B lood vesse1
1 1 B ra in (hvpothalamus. medu l la l
Kidne y
pressor efkct
vieterand saltinmke
barorecepbr reflex
a ntidiuresis
d iure s is
naaiuresis
3' -- O 4 reducedlevels / decrease
increased levels / increase
Figure 4.5.1. Schemaoc d iag ramsumnz ing the effects of chronic moderate ETOH consunpnon on the namureoc syçtemand the
m c h a n i s n s by which these modificauons rnay rnediate part of alcohol's antihypertensive effect.
During conditions of long-term moderate ETOH treaûmnt, acute E T O H consunption m y increase plasma ANP (and BNP)
levels (1). This effect results m s r likely from the release of accumilated ANP fmm heart aûial granules and from che
elevated BNP producaon from hean venmcular nssue (2). The elevated ANP (and CNP) contents in the CNS may also
contribute slighdy to the increased namureoc pepude levels in the circulation by ttre stinulauon of ANP release through ME
and by the activanon of horrmnal and neuronal pathways sunxilating heart ANP iand BNP) release through NL (3).
The elevated plasma ANP (and BNP) levels f o t l o ~ n g acute ETOH admnismuon may then lower BP by several
mechanism. They may produce vasorelaxaiion in the major blood vessels (4). They rnay enhance renal diuresis and
namuresis by binding to elevated NPR-AiNPR-C in the glomeruli and NPR-A in the IM (5 ) . They r m y reduce the Alln çignal
to rhe pressorcentres of the brain by binding to elevated NPR-AINPR-C in the SFO (6) . This effect associated w th the
elevated ANP iand CNP) contents in the CNS, m y reduce the antihypertensive effect of AVP, the dipsogenic and pressor
effects of All and che threshold forthe baroreceptor reflex (7).

of ETOH-treated animals during low BAC may represent the major source of elevated plasma
ANP levels following each acute alcohol exposure. Likewise. the enhanced ventricular BNP
mRNA and BNP content suggest elevated BNP production fkom the ventricular tissue and
qeater BNP release mto the circulation.
(3) Ahhough the major source of plasma ANP and BNP variations is the heart. the effect of
ETOH on the central ANP and CNP systems may also contniute slightly to the elevated
natriuretic peptide leveis. indeed the ETOH-mduced mcreases in hypothalamic and medullary
ANP and CNP contents may produce a greater activation of the ANP neurons (ANPn)
terminahg m the median eninence (ME) and neural lobe of the pituitary gland (NL). leading
to the release of natriuretic peptides mto the vasculature draining the ME or to the release of
other hormonal or neuronal pathways invohed in the stimulation of ANP and/or BNP eom
the heart.
The elevated plasma naaiuretic peptide levels foUowing each acute ETOH consumption
(or during hi& BAC) m chronicaUy ETOH-treated anirnals may then lower the BP by several
mechanisms.
(4) nie higher circulating ANP and BNP levels may produce some vasorelaxation in the
major blood vessels.
(5) The higher circulating ANP and BNP levels may cause elevated diuresis and natriuresis.
This effect is enhanced by the higher proportion of active NPR-A (dom-regdation of NPR-
C) in the renal glomeruli and by the increased quantity of NPR-A in the renal inner medulla
(IM) of ETOH-treated rats.
(6) The higher circulating ANP and BNP levels may reduce the pressor sipals to the brain
cardiovascular centres mediated by AI1 neurons ( A h ) . This effect is enhanced by the lower
a££ïnity of the clearance NPR-C in the SFO (and CP) of ETOH-treated rats.
(7) The effect of higher circulating ANP and BNP levels to the CNS is further enhanced by
elevated ANP and CNP contents in the hypothalamus and meduila of ETOH-treated rats.
These natriuretic peptide levels may reduce the antidiuretic effect of AVP. the pressor and
salt/water mtake produced by An and recalibrate the baroreceptor reflex to a lower BP value.

The objective of the present work was to mvestigate the implication of the natriuretic
peptide system in the prwention of the agedependent increase in BP by chronic moderate
ETOH consumption. In general terms, both the major sites of production for this hormonal
family, such as the heart, and the main effector areas, such as the kidneys and the bram.
presented ETOH-mduced modifications that may explain, at least in part, the effect of
moderate ETOH connimption on the BP. Therefore, both components of this endocrine
system, the peptides (ANP. BNP and CM) and the receptors (NPR-A NPR-B and NPR-C)
have undergone specific ETOH-mediated alterations, suggeamg widespread modifications
in the general characteristics of the syaem and ailowhg the possibility of a role in ETOHs
antihypertensive effect.
Smce the present studies are among the few experiments investigatmg the mteractions
of ETOH with the natriuretic peptide system, they are of an exploratory nature. Now that a
significant interaction between ETOH and the various cornponents of the natriuretic syaern
have been demonstrated, W e r studies should be perfonned focused on distinct components
ofthe natriuretic £àmily to evaluate the exact mechanisms mvolved and better defhe the role
ofthe natriuretic system on the lowering effect of chronic moderate ETOH consumption on
B P.
4.5.1. HEART A N D VASCULATURE
(a) The possiiility of an mdirect effect of ETOH on the cardiac natriuretic peptide system
is niggested by the observation that heart myocytes exposed to alcohol iti vitro present
no changes in ANP release (Wigle er al.. 1993b). interestingly, there is also a arong
positive correlation between the changes of plasma ANP and circulating O-endorphin
levels following acute ETOH administration (Section 2.1 .). Since ETOH consumption
rapidly increases plasma Il-endorphin levels (Gianoulakis et al., 1989) and p opioid
receptor agonists, such as fentanyl have been associated with the release of ANP fkom
the heart (Vollmar et al., 1987; Gutkowska et al., 1993), it may be hypothesized that the
ETOH-induced increase on circdating levels of û-endorphin peptides may mediate the
ETOH-induced mcrease in plasma ANP levels. Therefore, the effect of naloxone (an

opioid receptor antagooist) on the cirnilating ANP (and BNP) levels during acute ETOH
adminktmtion should illustrate whether the endogenous opioid system is mediahg the
ETORmduced mcrease in the plasma ANP levels. The use of opioid receptor
antagonists that do not cross the BBB nich as methyhaltrexone may M e r illustrate
whether the e f f i of alcohol on tûe nahiuretic peptides is produced in the heart or CNS.
(b) From the data obtained following acute and chronic ETOH exposure. we have
proposed that ETOH may increase the potential activity of the healt ANP and BNP
systems (section 4.2.). However, lower plasma ANP and BNP levels were observed m
animals chronicaily treated with alcohol, because of the hemodynamic modifications
produced by the lower BP. Yet. it was proposed that the ETOH-specific alterations in
cardiac ANP and BNP levels may enhance the ability of the heart natriuretic peptide
systems to respond to various physiological andlor pharmacological conditions (aich as
high BAC) by a more pronounced expression and release of the natnuretic peptides.
Thus. the effect of an acute ETOH administration in animals chronically treated with
ETOH should be mvestigated. The demonstration of an increased release of ANP andlor
BNP foilowhg an acute ETOH challenge would support the hypothesis of the tight and
dark phases of the daily cycle on ETOH consumption and its effect on the release of the
natriuretic peptides (section 4.2.).
(c) The wahration of the circulating and cardiac ANP and BNP levels at various intervals
during the 8 months of the experimental period should determine the progressive changes
in natriuretic peptide levels and allow their association with the development of the
antihypertensive effect of alcohol. The respective contribution of ETOH and of the
secondary hemodynamic changes to the actual b e l s of both ANP and BNP activity may
then become more clear.
(d) Puise and pulse-chase midies could determine the rate of biosynthesis of ANP and
BNP in the various components of the cardiac tissue and allow a direct correlation

between the effect of ETOH on the mRNA levels and the peptide biosynthesis. Such
experirnents are especiaily important for the BNP system seice the BNP gene contains
an ATlTA motif in the 3'-untranslateci region niggesting mRNA dest abilization ( Ogawa
et al., 1995).
(e) The circulating natriuretic peptide levels may also be modifled by ETOH-induced
alterations in NPR-C receptors either directly at the site of release (Nunez er al.. 1993 )
or m the blood aream nich as in the platelets. Changes in vascular and cardiac NPR-C
receptors should then be detennined following chronic ETOH treatment to see whether
ETOH may alter either their number or their a£Enity for ANP and BNP.
1.5.2. KIDNEY
(a) Chrooic ETOH treatment was associated with the increase in papillary NPR-A
bmding sites (Section 3 2.). However. the effect of circutating ANP and BNP levels on
these receptors is likely to be minimal. considering the important layer of endopeptidase
24.1 1 enrymes located in the proximal tubule of the nephron (Gunning and Brenner.
1992). hstead intrarenal UD. ANP and even CNP may bind to the NPR-A receptors in
the iM, producing paracrine natriuretic and depressor effects (Figueroa et al.. 1 990:
Goetz 199 1 : Terada et al.. 1991). Therefore. experirnents should be performed on the
expression and release of natnuretic peptides of renal ongui to evaluate whether ETOH
modifies their levels and whether nich ETOH-induced modifications parallel the changes
in papillary NPR-A receptors.
(b) The status of the proximal tubule endopeptidase 24.11 enzymes should also be
investigated following chronic moderate ETOH treatrnent to evaluate whether ETOH
alters the acthity of endopeptidase 24.11 and thus aUows a greater proportion of
circulating ANP and BNP levels to reach the natnuretic receptors in the IM and affect
directly the natriuresis of ETOH-treated rats.

(c) The modifications produced by ETOH on both the gIomedar and papillary
natriuretic peptide receptors suggest the possibility of an enhanced diuretic and
nahret ic e f f i ofnatriuretic peptides (Section 3.3.). It may be useful to inject specifïc
concentrations of ANP (and BNP) i v. mto ETOH- and water-treat ed animals t O evaluat e
whether natriuresis and diuresis are greater in the ETOH- than in water-treated rats.
Animais should also be placed in highly accurate metabolic cages to observe the exact
modifications in cGMP production. natriuresis and diuresis during both light and dark
phases of the daily cycle.
4s.3, BRAIN
(a) Chronic ETOH treatment produced significant mcreases in hypothalamic and
medullary ANP and CNP levels in SHR rats (Section 3.4.). ANP immunoreact~ty is
mamly l o c k d m the cardiovascular centres of the brain. nich as the AVW area or the
NTS (Kawata et al.. 1985). in contrast. CNP has been found in additional brah regions
which are unrelated to fluid or BP homeostasis (Komatsu et al.. 199 1). Future
experiments may inveaigate. through Ni sitic hybridization or immunohistocherniary.
which s p e d c nuclei present elevated ANP and CNP peptide levels as well as ANP and
CNP mRNA levels. allowing the exact mapping of the ETOH-induced changes in the
cardiovascular centres of the CNS.
(b) ANP (and C M ) may also be injected ;.C.V. into the third (hypothalamus) or fourth
(meduiia) ventricles of the brain to investigate whether the antidipsogenic. loss of sait
appetite. diuretic. natriuretic and depressor effects of ANP are modifîed in the brain of
ETOH-treated animais compared to the brain of water-treated rats.
(c) Chronic ETOH treatment has been associated with some specifk changes in the
uatrIuretic peptide receptor characteristics m the circumventricular organs (Section 3.5. ).
It is possible that natriuretic peptide receptors located inside the brain are also modZed
following ETOH consumption and that these modifications parallel those of the brain

ANP and CNP peptides. Future experiments may then explore NPR mRNA levels in
various hypothalamic and rneduhq nuclei and correlate the ETOH-induced changes on
NPR levels with the ETOH-induced changes on the ANP and CNP contents and their
specific mRNAs.
In conclusion this work dowed the identification of an hormonal system the natriuretic
peptide system that may be implicated m the prevention of the age-dependent increase in BP
by chronic moderate ETOH consurnption. The data obtained indicated that the general
aspects of this endocrine system such as the organs of production and the major bindmg sites.
have presented ETOH-induced modifications that may mediate ETOHs antihypertensive
effect at least in part. Therefore. the exact and specific role of this natriuretic peptide family
in mediatmg alcohoi's effects on the BP clearly ments further investigation. Future
experiments should provide new msights regarding the exact contribution of each component
of this endocrine system on the antihypertensive effect of moderate ETOH consumption
observed in both human and animal studies.


METHODOLOGIES
a) Estimation of the BP andor atrial pressure
In the present studies, the systolic BP was evaiuated by the non-mvasive mdirect tail-cuff
method using a photoelectnc sensor attached to a polygraph (Grass Instruments, Quincey.
MASS) (Pfeffer et al.. 1971). In general, the tail-cdfmethod mvolves the occlusion of the
arterial blood flow to the tail through an idated cuffand the subsequent detection of the
exact pressure, during the cddeflation, at which the fira arterial pulsation reappears. The
vddity of this technique ultimately dep ends on the sensitMty of the sensor used t O record this
initial pulsation. uiitialiy, anesthetized animals and extemai preheating of the tail were
necessas, to produce çome vasorelaxation and ailow the recording of artenal pulsations (Borg
and Viberg, 1980). However, these conditions have bem show to mod* the measured BP.
For example, preheating may by itself'elevate the BP m SHR rats because of their greater
sensitMty to thermal stress than WKY animals (Yen et a!., 1978). Modem taiI-cuffmethods
have bypassed these technicd düEculties by usmg better sensors, such as photoelectnc rather
than ultrasonic sensors, thus dowing repeated and accurate recordmgs of the BP in unheated
non-anesthetized rats (Bunag and Buttefield, 1982; ikeda et al.. 199 1 ; Spanos et al.. 199 1 ).
in fact, syaolic BP estimations using the mdirect method have shown an aimoa perfect
correlation with direct syaolic BP measurements using an indwehg arterial catheter
(r=0.970 m the experiment of Ikeda et al., 199 1 ). Furthemore, the mdirect technique proved
to be a superior means of BP measmement in long-term or chronic midies, because this non-
invasive method prevented the significant appetite suppression and growth impairment
associated with the chronic arterial catheterization (O'Neill and Kaufinan, 1990).
ANP is released pnmady fkom the cardiac atria m response to elevated atrial pressure and
the concomitant stretch of the atrial walis (Nose et al.. 1994; Javeshghani et ah, 1995).
interestingly, lefi atrial pressure has been shown to be twice as hi& m SHR compared to
WKY rats, explainhg in part the elevated levels of circulating ANP in the hypertensive
animals (Noresson et al., 1979). This effect appears to be secondq and conipensatoly to the
ventricular hypertmphy observeci with elevated BP m SHR rats (Ricksten et al., 1980). ui the

present sudies, chronic moderate ETOH coflsumption prevented the development of the age-
dependent hypertension in SHR rats and thus the ventricular hypemophy. Accordingiy, atrial
pressure should also be reduced in these animals compared to the water-treated SHR rats.
Although catheters could have been placed m both atria, it seems likely that these surgical
procedures would have caused stressfùl reactions of the animals during the chronic studies
and modined the relationships between ETOH and BP and between ETOH and ANP.
Furthemore, previous studies have mdicated that ANP release f?om the atria upon ETOH
stimulation is indirect and mdependent of water-loading and increased atrial stretch
(Colantonio et al.. 199 1 ; Wigle et al.. 1993a).
b) Route of ETOE injection in acute studies (rats)
in the present studies, 1 and 2 g of ETOWkg B.W. (as a 40% v/v solution) were injected
irztraperitor~eal& (i-p. ) in Long-Evans rats. Although btîraverzous (i-v.) injection or
irzîragutric (i.g.) administration couid have been used as well, the i.p. injection was chosen
for the following reasons:
I.D. hieetion conpareci to i. v. iniection: (a) Concentrated injections of ETOH in the blood
are IethaL AccordBigly, 1 and 2 g of ETOWkg B. W. çhould be mjected i. v. in rats as a diluted
solution, increasing greatly the final volume of the injection. In contraa, higher concentrations
of ETOH (up to 40% vh) may be çafely mjected i . p . reducing the chance of a water-Ioading
effect on the BP or the natriuretic peptide system (Roine et al., 199 1). (b) The BAC is
maximum immediately after the i. v. injection. However, because of the gradua1 diffusion of
ETOH mto the blood vessels after the i.p. injection, the BAC is maximum 30 minutes post-
injection and thus better mimics the BAC curve following oral administration of alcohol.
[.o. iniectiorz comoared to i.p admDiisiratzo~z: (a) intragastnc mtubation may stress the
animais. Furthemore, this route of administration may produce elevated levels of ETOH in
the stomach, causing gastric imitation and the possibhty of sympathetic activation. Both
effècts may modiS, the interaction between alcohol and the natriuretic peptide system (b) The
use of i-p. rather than i.g. admhktration prevents the possibility of different BAC levels £iom
the same i-g. ETOH administration m different animals because of the presence of different

amounts of food m the stomach and thus dserent Iweis of gashic first-pas metabolism
(Rome et al., 199 1).

REFERENCES

Abdel-Rahman A4, Wooles WR: Ethanol-mduced hypertension bvoives impairment of baroreceptors. Hypertension 10; 67-73, 1 987.
AbdeCRahman AA: Acute effects of ethanol on cardiac output and its derivatives m spontaneoudy hyperteosive and normotensive rats. "1 Pharmacol Exp mer 27 1 ; 1 1 50- 1 1 5 8. 1994.
Abe H, Kawano Y, Kojima S, Ashida T, Kuramochi M, Matsuoka H, Omae T: Biphasic effects of repeated alcohol intake on 24-hou blood pressure m hypertensive patients. Circzilatzorz 89; 26626-2633, 1994.
Abe T, Arakawa Y, Rajasekaran AK, Yu T, Wada O: Interaction of atrial natriuretic peptide with its receptors in bovine lung membranes. J Bi01 Chem 270; 7672-7678, 1995.
Aburaya M, Minamino N, Hono J, Kangawa K Matsuo H: Distribution and molecdar forms ofbrain natriuretic peptide m the central nervous syaem, heart and peripheral tissues of rat. Biochem Biophys Res Commun 165; 880-887, 1 989a.
Aburaya M, MmamÏno N, Kangawa K, Tanaka Y Matsuo H: Distnbution and molecdar f o m of braïn naaiuretic peptide m porcine heart and blood. Biochern Biophys Res Cornnititi 165: 872-879. 1989b.
Aburaya M, Hmo .J, Minamho K, Kangawa K, Matsuo H: Isolation and identification of rat brain natriuretic peptides in cardiac atrium. Biochenr Bzophys Res Commtm 1 63 ; 226-23 8. 198%.
Ackerman BK Wyeth RP. Vesely DL, Ngo L, Bisses JK, Wmters CJ, Sallman AL: Warmacokinetic characterization of post-distribution phase of prohormone atrial natriuretic peptides amino acids 1-98,3 1-67, and amal natriuretic fàctor during and followiug rapid right ventncdar pacing in dogs. J Clin Pharmacol 32; 41 5-42 1, 1992.
Adesso VJ, Ritchie SA, Stasiewicz PR: The acute effects of alcohol on the blood pressure of young, normotensive men. J Shrd Alcohol5 1 ; 468-47 1, 1990.
Aguilera G: Merential effects of atrial natriuretic factor on angiotensin II and adrenocorticotropin-stimulated aldosterone secretion. Endocririology 120; 299-304, 1987.
Aiari L, Lewander T, Sjoquist B: The effect of ethanol on the brain catecholarnme system in f e u l e mice, rats, and guinea pigs. Alcohol Clin Exp Res 1 1; 144- 149, 1987.
AUan AM, Harris RA: Acute and chronic ethanol treatments after GABA receptor-operated chloride channels. Phamracol Bzochem Behav 27; 665-670, 1987.

Allen JK, Adena MA: The association between plasma cholesterol, hi&-density iipoprotem choleaerol, triglycerides and uric acid in ethanol consumers. A m Clil1 Biochem 22: 62-66. 1985.
Almeida FA, Suzuki M, Maack T: Aaial natriuretic fàaor mcreases hernatocrit and decreases plasma volume m nephrectomized rats. Llfe Sci 39; 1193- 1 199, 1986.
Altura BM, Altura BT: Microvascular and vascular smooth muscle actions of ethanoL acetaldehyde, and acetate. Federatiorz Proc 4 1 ; 2447-245 1, 1982.
Altura BM, Alma BT: Peripheral vascular actions of ethanol and its interaction with neurohumoral substances. Neztrobehav Toxicol Teratol5: 2 1 1-220, 1983.
Altura BM, Altura BT, Gebrewold A: Alcohol induced spasma of cerebral blood vessels. Science 220; 33 1-333, 1983.
Anand-Srivastava MB, Sakm MR, Cantin M: Ring-deleted analogs of atrial natriuretic factor mhibit adenylate cyclase CAMP system: Possiile couphg of clearance atrial natriuretic factor receptors to adenylate cyclase CAMP signal transduction system. J Biol Chem 265: 8566- 8572, 1990.
Anand-Srivastava MB, Gutkowska J, Cantin M: The presence of atrial natriuretic factor receptors of ANF-R2 subtype in rat platelets. Biochem J 278; 2 1 1-2 17, 199 1.
Anand-Srivastava MB, Trachte GJ: A M natriuretic factor receptors and signal transduction mechanisms. P h t a c o l Rev 45; 455-497. 1993.
Anderson SV, Mülar ND, O'Hare SP, MacKenzie JC, Corral RJ, Bloom SR: Atrial natnuretic peptide: Wysiological release associated with nahiuresh during water immersion in man. Clin Sci 7 1 ; 3 19-322, 1986.
AntunesRodngues J, McCann SM, Rogers LC, Samson WlC Atrial natriuretic factor hhi'bits dehydration- and angiotensin II -mduced water mtake in the conscious, unrestrained rat. Proc Nat1 Acad Sci USA 82; 8720-8723, 1985.
AntmesRodrigues .J, McCann SM, Samson WK: Central administration of atrial natnuretic factor inhi'bits saline preference m the rat. Endocrirzology 1 18; 1726- 1728, 1986.
Arai H, Nakao K, Saito Y, Morü N, Sugawara 4 Yamada T, Itoh H, Shiono S, Mukoyama M., Ohkubo H, Nakanishi S, Imura H: Augmented expression of atrid natriuretic polypeptide gene m ventricles of spontaneoudy hypertensive rats (Sm) and SHR-stroke prone. Circ Res 62; 926-930, 1988.

Argentin S, Drouio J, Nemer M: Thyroid homone stimulates rat pronatriodilatin mRNA levels in primary cardiocyte cultures. Biochem Biophys Res Commun 146; 1 3 36- 1 34 1, 1 987.
Arjamaa O, Vuofteenaho O: Sodium ion stimulates the release of atrial natriuretic polypeptide (ANP) frorn rat auia. Biochem Biophys Res Commun 132: 375-381. 1985.
Arkwright PD, Beilin W, Rouse 1, Armstrong BK, Vandongen R: Effects of alcohol use and other aspects of Mestyle on blood pressure levels and the prevalence of hypertension in a working population. Circulatio~z 66; 60-66, 1982a.
Arkwright PD, Beilin U, Vandongen R, Rouse IA, Lalor C: The pressor effect of moderate alcohol consumption: A search for mechanisms. Circulation 66; 5 15-5 19, l982b.
Arkwright PD, Beilin U, Vandongen R Rouse IL, Masarei IR: Plasma calcium and cortisol as predisposhg fictors to alcobol related blood pressure elevation. J Hypertem 2: 387-392. 1984.
Atlas SA Klemert HD, Camargo MJ, Januszewicz A, Sealey JE, Laragh JK SchiUmg JW. Lewicki IL!, Johnson LK, Maack T: Purification, sequencing and synthesis of natriuretic and vasoactive rat atrial peptide. Nature 309; 7 17-7 19, 1984.
Avidor q Eilam R, Malach R, Gozes 1: VIP-mRNA is mcreased in hypertensive rats. Brait, Res 503; 304-308, 1989.
Babinski K, Roy M, De Léan 4 Ong H: Etude comparative de la régulation de la biosynthèse et de la sécretion d'une troisième famille de peptides natnurétiques dans les cellules c h r o r n a ~ e s en culture. Med Sci 7; 60, 199 1.
Baghura Y Dwyer T: Alcohol consumption and blood pressure m a group of young Australian males. J Htrrnmr Mitr 3 5 ; 257-264, 198 1.
Bahr V, Sander-Baihr C, Ardevol R, Tuchelt H., Beland B, Oelkers W: Effects of atrial natriuretic factor on the renin-aldosterone syaem: hi vivo and il1 vitro studies. J Sterod Biochem Mdec Bzol45; 173478, 1993.
Baldissera S, Menani JW, Sotero dos Santos LF, Favaretto ALV, Gutkowska J , Turrin MQA, McCann SM, Antunes-Rodrigues I: Rote of the hypothalamus in the control of atrial natriuretic peptide release. Proc N d Acad Sci USA 86; 9621-9625, 1989.
Ballantyne FC, Clark RS, Simpson HS, Ballantyne D: High density and low density lipoprotem subfiactions m survivors of myocardial Sarction and in control subjects. Metabolism 3 1; 433-437, 1982.

Baraona E, LR, W, Ma XL, Svegiiati-Baroui G, Lieber CS: Acetaldehyde-collagen adducts m N-nitroçodimethyia-e-mduced liver cirrhosis m rats. Lfe Sei 52; 1249-1255, 1993.
Barboriak JJ, Anderson AJ, Hofbann RG: Interrelationship between coronary artery occhision, hi&-dene lipoprotem cholesterol, and alcohol intake. J Lab C h Med 94: 348- 353, 1979.
Beard ID, b o n DH: Fluid and electrolyte balance during acute withdrawal in chronic alcoholic patients. J Am Med Assoc 204; 13 5- 139, 1968.
Beüm LJ, Puddey IB: Alcobol and hypertension. Clin Exp Hperteris [A] 14; 1 19- 138_ 1992.
Beilin U. Hoffmann P. Nilsson H, Skarphedinsson I, Folkow B: Effect of chronic ethanol consumption upon cardiovascular reactivity , hem rate and blood pressure in spontaneousl y hypertensive and W istar-Kyoto rats. 3 Hypenens 10: 645-650, 1992.
Bergaglio M, Calaresu F: Cardiovascular effects of micromjection of ANP and brain natriuretic peptide mto ventrolaterd medulla. Am J Physiol259; K2-R40, IWO.
Berggren SM, Goldberg L: Absorption of ethyl alcohol fiom gastro-intestinal tract as diffusion process. Acta Physiol Seand 1 ; 246-270, 1940.
Bermudez F, Surks MI, Oppenheimer JH: High incidence of decreased serum trüodotbyronine concentration m patients with nonthyroidal disease. J Clln E~rdocrii~ol Merub 4 1 ; 27-10, 1975.
Bernstein J, Videla L, Israel Y: Hormonal influences m the development of the bypermetabolic Gate ofthe liver by chronic administration of ethanol. J P h t a c o f Exp Ther 192; 583-59 1, 1975.
Bezzegh A, Nyuli L, Kovacs G: a-ahial natriuretic peptide, aldosterone secretion and plasma renin a c t ~ t y during ethanol withdrawal: A correlation with the onset of delirium tremens? Afcohol8; 333-336, 1991.
Bianchi C, Gutkowska J, ïûiiault G, Garcia R, Genest J, Cantin M: Distinct localization of aaial natriuretic factor and angiotensin II bindmg sites in the glomerulus. Am J Physiol25 1 : F594-F602, 1986.
Bianchi C, Gutkowska J, Garcia R thibault G, Genest I, Cantin M: Localization of 1'5~-atrial natriuretic factor (ANFFbinding sites m rat renal medulla: A light and electron microscope autoradiographic study. J Histochern Cytochem 35; 149- 153, 1987.
Bidmon HJ, Gutkowska J, Murakami R Stumpf WE: V i t a e D receptors in heart: Effects

on atrial natriuretic factor. Experientia 47; 958-962, 199 1.
Biollaz J, Callahan LT, Nussberger I, Waeber B, Gomez HJ, Blaine EH, Brunner HR: ebamiacokinetics of synthetic atnal natrRuetic peptides m normal men. Clin Pharmacol Ther 4 1 ; 67 1-677, 1987.
Blaine EW: Atrial natriuretic Bctor plays a significant role m body fluid homeostasis. Hypertension 15; 2-8, 1990.
Blass JP, Gibsm GE: Abnormality of a thiamine-requiring enzyme m patient with Wemicke- Korsakoff syndrome. N Engl J Med 297; 1367- 1370, 1977.
Blaustein MP: Sodium ions, calcium ions, blood pressure reguiation, and hypertension: a reassessement and an hypothesis. Am J Physiol232; C 165-C 173, 1977.
Bloch KD, Scott .l& Zisfem JB, FaUon JT, Margolies MN, Seidman CE? Matsveda GR Homcyn CJ, Graham RM, Seidman JG: Biosynthesis and secretion of proatrial natriuretic factor by cultured rat cardiocytes. Science 23 0; 1 1 68- 1 1 7 1, 1 985.
BIoch KD, Seidman JG, Naftilan JD, FalIan ST, Seidman CE: Neonatal atna and ventricles çecrete atrial natriuretic fàctor via tissue specinc secretory pathways. Cell47; 695-702. 1986.
BoIli P, M d e r FB, Linder L, Raine AE, R e d TJ, Erne P: The vasodilator potency of atrial natriuretic peptide in man. Cirnrlatioti 75; 22 1-228, 1987.
Bonhomme MC, Garcia R Heterogeneous reguiation of rend atriai natriuretic factor receptor subtypes m one-kidney, one-clip hypertensive rats. J Hyperterls 1 1; 389-397, 1993.
Bora PS, Lange LG: Molecular mechanism of ethanol metabolism by human brain to fatty acid ethyl esters. Alcohol Clin Exp Res 17; 28-30, 1993.
Borg E, Viberg A: Role ofheatmg m non-invasive blood pressure measurements in rats. Acta Physiol Scand 108; 73-75, 1980.
Borg S, Kvande H., Rydberg U, Terenius L, Wahlstrom A: Endorphin levels m human cerebrospinal fluid during alcohol intoxication and withdrawal. Psychopharmacol 78: 10 1 - 103, 1982.
Bourassa M, Schmer H, PezaIla PD, Lis M, Chrétien M: Immunoreactive O-endorphin in rat mammotrophic transplatable tumor (MnF4). Cumer Res 38; 1 568- 1 5 7 1, 1978.
Boveris A, Oshino N, Chance B: The cellular production of hydrogen peroxide. Biocheni J 128; 617-630, 1972.

Bovy PR, O'Neal JM, Olins GM, Patton DR: Identification of structural requirements for analogues of atrial natriuretic peptide (ANP) to discriminate between receptor subtypes. J Med Chem 32; 869-874, 1989.
Bowers RV, Burleson WD, Blades IF: Alcohol absorption f?om skh in man. Quart J Stud Alcohol3; 3 1-33, 1942.
Brackett DI, G a d DV, Lemer Holloway F& Wilson MF: Dose- and time-dependent cardiovascular responses mduced by ethanol. J Phmucol Erp Ther 268; 78-84. 1994.
Bradford MM: A rapid and sensitive method for the quantifiication of microgram quantaies of protem utilking the principle of protein-dye bindmg. A n a l Biochem 72; 248-254, 1 976.
Braunwald E, Harrison DC, Chidsey C k The heart as an endocrine organ. Am J Med 36; 1-4. 1964.
Brennan Lq Mahin RL. J o c h KE, Roberts DE: Influence of nght and lefi atrîal receptors on plasma concentrations of ADH and renin. Am J Physiol22 1 ; 273-278. 197 1.
Breuhaus BA, Saneü HH, Brandt C b o s k e y JE: Atriopeptm ïI lowers cardiac output in conscious sheep. Am J Physiol249; R776-R780, 1985.
Brown J, Czarneclci A: Distniution of atriai natriuretic peptide receptor subtypes in rat brain. Am J Physiol258 (Regulatory Iritegmtive Comp Physiol27); R1078-R1083, 1990a.
Brown J , Czamecki A: Autoradiographic localization of atrial and bram natriuretic peptide receptors in rat brain. Am J Physiol258 (Regulatory Integrative Comp Physiol27); R57- R63, 1990b.
Brown J , Zuo 2: Renal receptors for atrial and C-type natriuretic peptides in the rat. Anz J Physiol263 (Rertal Fluid electrotyte Physiol 32); F89-F96, 1992.
Brown J , Zuo 2: C-type natriuretic peptide and atrial natriuretic peptide receptors of rat brain. Am J Physiol264 (Regulatory Iniegrative Comp Physiol3 3 ) ; R5 1 3- R5 23. 1 993.
Buggy J , Bealer SL: Physiological regdation by the AV3V region. In: Circumvetitricuinr orgam and bm&fluidî. Vol. 1, Gross ed., CRC Boca Raton, FL, p. 17 1- 190, 1987.
Bdger RE, Trump BF: Fine structure of the rat rend papilla. Am J AM 1 18; 685-722. 1966.
Bunag RD, Buttedeld J: Tail-cuffblood pressure measurement without extemal preheating in awake rats. Hyperteruiori 4; 89û-903, 1982.

Burke V, Riddey iB, Beilin Li: Monaldy associated with wines, beers, and spirits. Australian data niggest that choice of beverage relates to Mestyle and personality. BMJ 3 1 1; 1 166, 1995.
Burnett JC Jr, Kao PC, Hu DC, Heser DW, Heublem D, Granger JP, Opgenorth TJ. Reeder GS: Aaial natrimetic peptide elevation m congestive heart fdure m the human. Scierice 23 1 : 1 145- 1 147. 1986.
Cahill PA, Redrnond EM, Keenan AK: Vascular atrial natrituetic peptide receptor subtypes are not mdependently regulated by atrial peptides. J Biol Chem 265 ; 2 1 896-2 1906, 1 990.
Caims V, Keil U, Kleinbaurn D, Doering 4 Steiber .J: Alcohol conçumption as a N k factor for high blood pressure: Munich Blood Pressure study. Hypertension 6; 124- 13 1. 1984.
Campbell WB, Currie MG, Needleman P: Inhi'bition of aldosterone biosynthesis by atnopeptins m rat adrenal celis. CN'C Res 57; 1 13- 1 18, 1985.
Canaan-Kuhl S, Jamison RL, Myers BD, Pratt RE: Identification of B receptor for natriuretic peptide in human kidney. Endocric~ology 130; 5 50-552, 1992.
Chansel D. Pham P, Nivez MP, Ardaiilou R: Charactexization of atrial natriuretic factor recepton m human glomerular epithelial and nesangial ceils. Am J Physiol259; F6 19-F627. 1990.
Cantin M, Genest J: The h m as an endocrine gland. Hypertension I O (suppl. 1 ); 1 1 1 8-1 12 1. 1987.
Cantin M, Thibault G, h g JF, Gutkowska J1 Garcia R Jasin G, Hamet P. Genest J: ANF in experimental congestive heart failure. Am J Pathol 130; 5 52-568, 1988.
Capasso JM, Li P, Guideri G, Malhotra 4 Cortese R Anversa P: Myocardial mechanical, biochemicai, and stnictural alterations induced by chronic ethanol ingestion m rats. Circ Res 7 1; 346-356, 1992.
Carmichael FI, Saldivia V, Varghese GA, Israel Y, Orrego H: Ethanol-induced mcrease in ponal blood flow: role of acetate and A,- and A,-adenosine receptors. Am J Physiol255: G4 l7-G423, 1988.
Castefi WP, Gordon T, Hjortland MC, Kagan A, Doyle JY, Harnes CG, Hulley SB, Zukel WJ: Alcohol and blood lipids: The cooperative lipoprotem phenotyping study. Lancer 2; 1 53- 155, 1977.
Castelli WP, Anderson K: A population at Nk: Prevalence of high cholesterol levels in

hypertensive patients m the Framingham midy. Am J Med 80 (suppl. 2A); 23-32, 1986.
Castro Hilbig J, Schroeder G, Stock G: Aûial naaniretic factor inhibits central angiotensin LI pressor responses. Hyperienrion 9; 473-477, 1987.
Chan TCK, Sutter MC: Ethanol consumption and blood pressure. Life Sei 33: 1965- 1973. 1983.
Chan TCK, Wall RA, Sutter MC: Chronic ethanol consumption, stress, and hypertension. Hpertemion 7; 519-524. 1985.
Chang MS, Lowe DG, Lewis M, Hellmiss R, Chen E, Goeddei DV: DifEerential activation by atrial and brain natriuretic peptides of two Mirent guanylate cyclases. Nature 34 1 : 68-72. 1989.
Chapin RE, Breese GR Mueller RD: Possile mechanisms of reduction of plasma IuteiniMg hormone by ethanol. J Phannacol Exp 7ner 2 12; 6- 10, 1980.
Charles CJ; Espiner E 4 Richards AM: Cardiovascular actions of ANP: Contniutions of renal, neurohumoral, and hemodynamic factors m sheep. Am J Physioi 264; R533-R538. 1993.
Chamess ME, Que& Henteleff M: Ethanol Werentially regulates G proteins m neural cells. Bzochem Biophys Res Comrnwi 155; 138- 143, 1988.
Chamess ME: Ethanol and opioid receptor signalling. Experietitia 45; 4 18-428, 1989.
Charness ME: Brain lesions in alcohoiics. Alcohol C h Exp Res 17; 2- 1 1, 1993.
Chartier L, SchiflÎin EL, Thibauh G: Effect of atnal natriuretic factor ( M k r e l a t e d peptides on aldosterone secretion by adrenal glomerulosa cells: Critical role of the intramolecular disulphide bond. Biochem Biophys Res Commun 1 22; 1 7 1 - 174, 1984.
Chen YF, Elton TS, Oparil S: Quantitation of hypothalamic atrial natriuretic peptide messenger RNA in hypertensive rats. Hypertetuion 1 9; 296-302, 1992.
Chien YW, Frohlich ED, Tnppodo NC: Atrial natriuretic peptide mcreases resistance to venous rehun in rats. Am J Physzol252; H894-H899, 1987.
Chin JH, Goldstein DB: Electron paramagnetic resonance studies of ethanol on membrane fluidity. A& Exp MedBiol85A; 1 1 1- 122, 1977.
Chinkers M, Garbers DL: The protein kinase domain of the ANP receptor is required for

signaling. Science 245; 1392- 1394, 1989.
Chinkers M, Garbers DL, Chang MS, Lowe DG, Chin H, Goeddel DV. Schulz S: A membrane fonn of guanylate cyclase is an atriai natriuretic peptide receptor. Nature 3 3 8; 78- 83, 1989.
Chinkers M, Smgh S, Garbers DL: AdenSie nucleotides are required for activation of rat atrial natriuretic peptide receptor: Guanylyl cyclase expressed in a baculoWus system J Bi01 Chem 266; 4088-4093, 199 1.
Chomczynski P, Sacchi N: Single-step method of RNA isolation by acid Guanidiniurn- Thiocyanate-PhenoCChlorofom extraction. A n d Bzochem 162; 1 56- 1 5 9, 1 9 87.
Christensen G: Cardiovascular and rend effects of atrial natriuretic factor. Scarrd J C h Lab Invest 53; 203-209, 1993.
Cicero TJ, Badger TM: Effects of alcohol on the hypothdamic-pihiitary-gonadd axis m the male rat. J Pharmacol Exp Ther 20 1; 427-433, 1977.
Cicero TJ, BeU RD: Effects of ethanol and acetaldehyde on the biosynthesis of testosterone in the rodent testes. Biochern Biophys Res Commuti 94; 8 14-8 19, 1980.
Cicero TJ: Neuroendocrinological effects of alcohol. Amz Rev Med 32; 123- 142, 198 1.
Cicero TJ: Alcohol-mduced deficits in the hypothalamic-pituitary-luteiniting hormone aBs in the male. A kohol Clin Exp Res 6; 207- 2 1 5, 1 982.
Clark VA, Chapman JM, Coulson AH: Effects ofvarious factors on systolic and diastoiic blood pressure in the Los Angeles Heart study. J Chronic Dis 20; 5 7 1 - 5 8 1. 1967.
Clark LT, Friedman HS: Hypertension associated with alcohol withdrawal: Assessment of mechanisms and complications. Alcohol Clirl Exp Res 9; 125- 130. 1985.
Clark M, Camargo MJ: Atrial natriuretic factor in normal subjects and heart failure patients: Plasma levels and renai, hormonal and hemodynamic responses to peptide mfusion. J Clitr hzvest 78; 1362- 1374, 1986.
Clark BA Ehali D, Fi& L, McAiooo-Dyke M, Davis Y Mmake K., Epstein FH: Atnal natriuretic peptide suppresses osmostimulated vasopressin release m young and ederly humans. Am J Physiol (Errdocri,iol Me tub 24) 26 1 ; E25 2-E256, 1 99 1.
Clark BA, Brown RS, Epstein FH: Effect of atrial natriuretic peptide on potassium-aimulated aldosterone secretion: Potential relevance to hypoaldosteronism in man. J Clin Irzvest 75:

Coate D: Moderate drhkhg and coronary heart disease monality: Evidence f?om NHANES 1 and the NHANES I foilow-up. Am JPubl Healrh 83; 888-890, 1993.
Coates RA, Corey PN, Ashley MJ, Steele CA: Alcohol connimption and blood pressure: analysis of data fkom the Canada Health survey. Prev Med 14; 1- 14, 1985.
Cobb CF, Van T'hie1 DH, Ennis MF, Gavder JS, Lester R: Acetaldehyde and ethanol are testicular toxins. Gasiroenterology 75; 958, 1978.
Coca 4 Agdera MT, De la Sierra A, Sinchez M, Picado MJ, Lluch M, Urbano-Marquez A: Chronic alcohol intake mduces reversible disturbances on cellular Na' metabotism in humans: Its reiationshrp with changes m blood pressure. Alcohol C h Eip Res 16; 7 14-720, 1992.
Colantonio D, Casale Desiati P, De Michele G, Mammareiia M, Pasqualeai P: A possible role of atriai natriuretic peptide m ethanol-induced acute diuresis. LIfe Sci 48; 63 5-642. 199 1 .
Colhgridge GL, Lester RA: Excitatoiy amino acid recepton in the vertebrate nervous system. Phannacol Rev 40; 143-2 IO, 1989.
Colsher PL, Wallace RB: 1s modea alcohol consumption better than none at ail? An epidemiological assessment. Atunr Rev Public Health 10; 203-219, 1989.
Conway N: Haemodynamic effects of ethyl alcohol in patients with coronary heart disease. Brit Heart J 30; 638-644, 1968.
Cooke KM, Frost GW, Thomell IR, Stokes GS: Alcohol consumption and blood pressure: w e y of the relation- m a heaith screening chic . Med J Amt 1 ; 65-69, 1982.
Cornian CW. Bridges RJ, Taube JS, Clark AS, Geddes JW, Monaghan DT: The role of the NMDA receptor m central nervous system plasticity and pathology. J NIH Res 1: 65-74. 1989.
Cowley AW, Liard IF: Vasopressin and a r t d pressure regdation. Hypertension 1 1 ; 1-25-E 32, 1988.
Crabb DW: The liver. In: Recent developments irz alcoholisrn. volume i 1, teri years of progress. Gaianter M (ed), New York, Plenum press, p. 207-230, 1993.
Crabos M, Ausiello DA, Haupert GT Jr, Cantiello HF: Atrial natriuretic peptide regulates release of Na+-K+-ATPase gihibitor fiom rat brain. Am J Physiol254; F9 12-F9 17, 1988.

Crandan ME, Gregg CM: In vitro evidence for an inhiiitory effect of atrial natriuretic peptide on vasopressin release. Neuroendmriiiol44; 43 9-445, 1986.
Criqui Wallace RB, Mïshkel M, Barrett-Corner E, Heiss G: Alcohol consumption and blood pressure: the Lipid Research Clinics Prevalence Study. Hypertension 3 ; 5 5 7- 5 6 5 , 1 98 1.
Criqui MH: Alcohol consumption, blood pressure, lipids, and cardiovascular mortality . Alcohol Cliri Exp Res 10; 564- 569, 1986.
Criscione L, Poweli JR Burbet R, Engesser S, Schlager F. Schoepfer A: Alcohol suppresses endothehumdependent relaxation m rat mesenteric vascular beds. Hypertensiori 1 3 : 964-96 7. 1989.
Crozier IG, Nichok MG, Ikrarn H, Espiner Yandle TG, Jans S: Atnd natriuretic peptide m humans: Production and clearance by various tissues. Hypertension 8 [suppl ilJ; II 1 1 -II 15. 1986.
Cden K: Alcohol and the J-curve survivaL Aust NZ JMed 23; 303-305, 1993.
Cuneo RC, Espmer Nicholls MG, Yandle TG, Livesey JH: Effect of physiological levels of atrial natriuretic peptide on hormone secretion: Inhibition of angiotensin-mduced aldosterone secretion and renin release m normal man. J C h Etuiocrinol Metab 65 ; 765- 772, 1987.
Currie MG, Gelier DM, Cole BR, Siegel Fok KF, Adams SP. Eubanks S R Galluppi GR Needleman P: M c a t i o n and sequence analysis of bioactive atnal peptides (atriopeptins). Science 2 10: 67-69. I983.
C h e MG, Oehlenschiager WF, Kuiu DT: Rofound elevation of ventncular and pulmonary atriopeptin m a mode1 of heart faihire. Bimhem Biophys Res Commun 148; 1 1 5 8- 1 1 64. 1 987.
Dagnino L, Drouin J, Nemer M: Merential expression of natriuretic peptide genes in cardiac and extracardiac tissues. Mol E~zdocrirzol 5; 1292- 1300, 199 1.
Dagnino L, Lavigne JP, Nemer M: increased transcripts for B-type natriuretic peptide in spontaneously hypertensive rats. Hypertemiori 20; 690-700, 1992.
Danziger RS, Sakai M, Capogrossi MC, Spurgeon HA, Hansford RG, Lakatta EG: Ethanol acutely and reversibly suppresses excitation-contraction couphg in cardiac myocytes. Cire Res 68; 1660- 1668, 199 1.
Dai Y? Rashba-Step J, Cederbaum AI: Stable expression of human cytochrome P4502E 1 in HepG2 cells: characterization of catalytic activities and production of reactive oxygen

intermediates. Biochernisv 3 2; 6928-693 7, 1993.
Davidson NC, Struthers AD: Brain natriuretic peptide. J Hjperteas 12; 329-336, 1994.
Davis CL, Briggs JP: Effect of atrial natriuretic peptides on rend m e d u h y solute gradients. Am J Physiol253; F679-F684, 1987.
Daynithi G, Antoni FA: Atriopepths are potent inhitors of ACTH secretion by rat anterior pituitary cens in vitro: involvement of the atrial natriuretic factor receptor domam of membrane-bound guanylyl cyclase. J EttdocrznoI 1 25 ; 3 9-44. 1 990.
Debinski W, Gutkowska J, Kuchel O, Racz EC. Buu NT, Cantm M, Genest J: ANP-Like peptide (s) m the peripheral autonomie nervous syaem Biochern Biophys Res Commun 134: 279-284, 1986.
Debinski WK, Kuchel OG, Thibadt G: Iocreased neural natriuretic factor generation in spontaneously hypertensive rats. Hyperterrsion 13; 804-8 12, 1989.
De Bold Al: Heart atria granulanty effects of changes E water-electrolyte balance. Proc Sm Exp Biol Med 161; 508-5 11, 1979.
De Bold AJ, Borenstem HB, Veress AT, Sonnenberg H: A rapid and potent natriuretic response to mtravenous mjection of atrial myocardial extract m rats. Lzfe Sci 28; 89-94. 198 1.
DeFrank RS, Jenkins CD, Rose RM: A longmiciinal investigation of the relationship among dcohol consump tion, p sychosocial fictors, and blood pressure. Psychosom Med 49; 23 6- 249. 1987.
Deitrich RA, Dunwiddie TV, Harris RA, Erwin VG: Mechanism of action of ethanol: Lnitial central nervous syaem actions. Pharmacol Rev 4 1 ; 49 1-537, 1989.
De Léan A Vinay P, Cantin M: Dianiution of atrial natriuretic factor receptors in dog kidney fractions. FEBS Let1 193; 239-242, 1985.
De Léon H, Gauqueh G, Thibault G, Garcia R: Characterization of receptors for the atnal natriuretic factor in rat renal rnicrovessels. J Hypertem 1 1 ; 499- 5 08, 1993.
De Marchi S, Cecchin E, Basile A, Bertotti A, Nardini R, Baitoli E: Rmal tubular dysfunction in chronic alcohol abuse: effeas of abstmence. N Engl J Med 329; 1927- 1934, 1993.
Deng Y, Kaufinan S: The influence of reproductive hormones on ANP release by rat atna. Life Sci 53; 689-696, 1993.

Deshaies Y, Leblanc J, Willemot J: Studies on protem metabolism during isoprotenerol- mduced cardiac hypertropby. Rec A h , Srud Cardiac Stmc Metub 8; 3 87-392, 1975.
Diamond i, Wrubel B, Estrin E, Gordon AS: Basal and adenosbe-receptor stimulated levels of CAMP are reduced m lymphocytes nom alcoholic patients. P roc Nat1 A cud Sci USA 84; 1413-1416, 1987.
Diamond i, Messing RO: Neurologic effects of alcoholism. West J Med 16 1 : 279-287. 1994.
Di Chiara G, ùiiperato A: h u g s abused by humans preferentiaily mcrease synaptic doparnine concentration m the mesolimbic system of fieely moving rats. Proc Nat1 Acad Sci USA 85: 52745278, 1988.
Diehl AK, Fuller JH, Mattock MB, Saher AM, El-Gohari R Keen H: The relationship of high den* $oprotein subhctions to dcohol consumption, other Westyle factors, and coronary heart disease. A therûscfermis 69; 145- 1 53, 1988.
Diehl AM, Bisgaard HC, Kren BT, Steer CJ: Ethanol interferes with regeneration-associated changes m biotransformhg enzymes: a potential mechanimi underlying ethanol's carcmogenicity? Hepatology 13; 722-727, 1991.
Dietz JR: The effect of angiotensin [I and ADH on the secretion of atnal natriuretic factor. Proc Soc Exp Biol Med 187; 366-369. 1988.
Dietz JR, Nazia. SJ, Vesely DL: Release of ANP, proANF 1-98, and proANF 3 1-67 from isolated rat atna by atnal distension. Am J Physiol260; H 1774-H 1778, 199 1 b.
Di Padova C, Womer TM, J h e n Lieber CS: Effects of f aahg and chronic alcohol consumption on the first p a s metabolimi of ethanol Gastroeriteroiogy 92; 1 169- 1 173. 1987.
Dos Reis AM, Fujio N, Dam TV, Mukaddam-Daher S, Jankowski M, Tremblay J. Gutkowska J: Characterization and distriiution of natriuretic peptide receptors in the rat uterus. Etdocrinology 136; 4247-4353, 1995.
Drewett JG, Ziegler RJ, Trachte GJ: Neuromodulatory effect of atnal natriuretic peptides correlate wiith an inhi'bition of adenylyl cyclase but not an activationof guanylyl cyclase. J Pharmacol Exp Ther 260; 689-696, 1992.
Dnimmer C, Fielder F, Konig A, Gerzer R: Urodilatin, a kidoey-derived natriuretic factor. is excreted with a circadian rhythm and is stimdated by salioe inf'usion m man. J Am Soc Nephrol 1; 1109-1 113, 199 1.
Dussaule JC, Stefanski A, Bea ML, Ronco P, Ardailou R: Characterizatiion of neutral

endopeptidase m vascular miooth muscle ceils of rabbt r e n d cortex Am J Physiol264; F45- F52, 1993.
Dyer AR, S t d e r J, Paul O: AIcohol consump tion. cardiovascuiar risk factors and rnortality in two Chicago epidemiologic studies. Ciradation 56; 1067- 1074, 1977.
Dyer Cutter G& Liu Y Armçtrong Mq Friedman GD, Hughes GH, Dolee JJ, Raczynski I, Burke G, Manolio T: Alcohol intake and blood pressure in young aduhs: the CARDIA study. J C h Epidemiol43 ; 1 - 13, 1990.
Edgarian H, Ahura BM: DifEierential efEects of ethanol on proaaglandm responses of arterial and venous smooth muscle. Erperzentia 32: 6 1 8-6 19, 1976.
Edmondson HA. Reynolds TB, Jacobson HG: Rend papillary necrosis with special reference to chronic alcoholism. A report of 20 cases. Arch int Med 1 18: 255-264. 1966.
Ehng T, Boson WF, Li TK: Alcohol and aldehyde dehydrogenase. Alcohol Alcoholism 25: 105- 1 16, 1990.
Ehrlich EU, Fitts DA: Atrial natriuretic peptide m the subfornical organ reduces drinkmg mduced by angiotensh or m response to water deprivation. Behav Neurasci 104: 365-372, 1990.
Eilam Malach R Bergmann F, Segal M: Hypertension induced by hypothalamic transplantation fiom genetically hypertensive to normotensive rats. J Neurosci 1 1 : JO 1-409. 1991.
EÏpper B q Mains RE: Structure and biosynthesis of proadrenocorticotropin/endorphin and related peptides. Emiocr Rev 1 : 1- 17. 1980.
Eisenhofer G, Johnson RH: Effect of ethanol mgestion on plasma vasopressin and water balance in humans. Ani J PhysioI242; 522-527, 1982.
Eisenhofer G. Lambie DG, Johnson RH: Effects of ethanol on plasma catecholarnines and norepinephrine clearance. Clin Pharmacol nter 34; 143- 147, 1983.
Eisenhofer G, Lambie DG, Johnson RH: EEects of ethanol ingestion on a-adrenoceptor- mediated circulatory responses m man. Br J Clitz Pharmacol 18; 58 1-586, 1984.
Eisenhofer G, Lambie DG, Whiteside E 4 Johnson RH: Vaçopressin concentrations during alcohol withdrawal. Br J Addict 80; 195- 199, 1985.
Eisenhofer G, Lambie DG, Johnson RH: Effects of ethanol ingestion on blood pressure

reactMty. Clin Scz 72; 25 1-254, 1987.
Ekman 4 Vakkuri O, Vuoheenaho O, Leppâluoto J: Ethanol decreases nocturnal plasma levels of atrial natriuretic peptide (ANP 99- 126) but not the N-terminal f?agment of pro-atnal natriuretic peptide (ANP 1-98) m man. Clin Sci 86; 285-290, 1994.
EIIiott ME, GoocUkia~d TL: Inhiiition of aldosterone synthesis by atrial natriuretic factor. Fed P m 45; 2376-238 1 , 1986.
Ellis FW: Effect of ethanol on plasna o~rticoçterone Iwels. J Pharmacol Exp 7Xer 153: 12 1- 127, 1966.
Eimér O, Goransson G, Zoucas E: Impairment of primaiy hemostasis and platelet function after alcohol mgestion m man. Haemmtasis 14; 223-228, 1984.
Emmeluth C, Dnunmer C, Gener R Bie P: Roles of cephalic Na' concentration and urodilatm m control of rend Na' excretion. Am J Physiol 262; F5 13-F5 16, 1992.
Emmehith C, Goetz KL, Dnimmer C, Gerzer R, Forssmann WG, Bie P: Renal denervation does not modulate natriuresis caused by carotid hypematremia m conscious dogs (abstract). . W I d Congr Inr Union Physiol Sci, p. 95, 1993.
Emsley RA, Potgieter A, Taljaard JI< Coetzee D, Joubert G, Gledhill RF: lmpaired water excretion and elevated plasma vasopressio in patients with alcohol-withdrawal symptoms. Q J Med 244; 67 1-678, 1987.
Erdos EG, Skidgel RA: Neutrai endopeptidase 24.1 1 (enkephalinase) and related regdations of peptide hormones. FASEB J 3; 145- 15 1 , 1989.
Ermulo R, Ruggeri P, Cogo CE, Molinari C, Calaresu FR: Neuronal and cardiovascular responses to ANF micromjected into nucleus ambiguous. Am J Physiol260; R1089-R1094, 1991.
Ermish A: Peptide recepton of the blood-bram barrier and substrate transport into the brah. Pr Brairz Res 91; 155-161, 1992.
Espmer EA: Physiolology of natriuretic peptides. J Irzt Med 235; 527-54 1, 1994.
Faison EP, Siegl PK, Morgan G, Winquia RI: Regional vasorelaxant seleaivity of atrial natriuretic factor m isolated rabbi vessels. Lrfe Sci 37; 1073- 1079, 1985.
Fewhgs JD, Hama MJD, Walsh JA, Whelan RF: The effects of ethyl alcohol on the blood vessels of the hand and forearm in man. Br J Pharmacol 27; 93- 106, 1966.

Figueroa CD, Lewis HM, MacIver AG, Mackenzie JC, Bhoola KD: Cellular localization of atrial natriuretic fàctor in human kidney. Nephroi Dial Trmplanr 5 ; 25-3 1, 1990.
F i G, Dow RC, Caslq. D, Johnson CI., Lim AT, Copolov DL, Bennie J, Carroll S. Dick H: Atrial natriuretic peptide is a physiological inhibitor of ACTH release: Evidence fiom immunoneutralVation in vivo. J EndocrznoI 13 1; R9-RIZ, 1991.
Fins DA, Thunhorst RL, Simpson JB: Diuresis and reduction of sait appetite by lateral ventricular infusions of atriopeptm II. Brairi Res 348; 1 18- 124, 1985.
Flynn TG, De Bold ML, De Bold AI: The amino acid sequence of an atnal peptide with potent diuretic and natriuretic properties. Biochern Biophys Res Commun 1 1 7 ; 859-865. 1983.
Fontoura BMA, Nussenzveig DR, Pelton KM, Maack T: Atrial naburetic factor receptors in cuitured renomeduhy mterstitial cells. Am J Physiol25 8 (Cell Physioi 27); C692-C699. 1990.
Forsander OA: Muence of the m e t a b o h of ethanol on the lactatdpyruvate ratio of rat-liver slices. Biochem J 98; 244-247, 1966.
Forssmann WG: Urodilatin (Ulantide, MN): A renal natriuretic peptide. Nephroti 69; 2 1 1- 222. 1995.
Forstermaun U, Feuerstain TJ: Decreased systemic function of prostaglandin E and prostacyclin, and unchanged thromboxane function, in alcoholics during withdrawal as estimated from metabolites in mice. Clirz Sci 73; 277-283, 1987.
Fortmaun SP, Haskell WL, Vranizan K Brown BW. Farquhar JW: The association of biood pressure and dietary alcohol: ciifferences by age, sex and enrogen use. Ani J Epideniiol 1 18: 497-507, 1983.
Fraenkel MB, Aldred GP, McDougaii JG: Sodium status aôects GC-B natnuretic peptide receptor mRNA Ievels, but not GC-A or C receptor mRNA levels, m the sheep kidney. C h SCZ 86; 5 17-522, 1994.
Frajria R Angeii A: Alcobol-induced pseudo-Cushing's syndrome. Laricet 1 : 1050- 105 1, 1977.
Franci CR, Anselmo-Franci 5 4 McCann SM: Opposite eEects of central immunoneutralization of angiotensin II or atrial natriuretic peptie on luteinking hormone release in ovariectomized rats. Neuroetidocrznol5 1 ; 683-6 87, 1990.

Franci CR, Anselmo-Franci JA, McCann SM: The role of endogenous atxial natriuretic peptide in restmg and stressinduced release of corticotropin, prolacth, growth hormone. and thyroid-stimulatmg hormone. froc Natl Acad Sci USA 89; 1 13 9 1 - 1 1395, 1992. Frankel EN, Kanner J, German JB, Parks E, Unselfa E: Inhi'bition of oxidation ofhiiman low-den* iipoprotein by phenolic substances in red wine. Lartcet 34 1; 454-457. 1993.
Fuiler F, Porter JG, m e n AE, Miner J, ShiIüng JW, Scarborough RM. Lewicki JA, Schenk DB, Atrial natriuretic clearance receptor: Complete sequence and hnctional expression of cDNA clones. J Biol Chem 263; 9395-940 1 , 1988.
Gagelmaun M, Hock D, Forssmann WG: Urodilatm (CDDI ANP,,-,,,) is not biologically mactivated by a peptidase fia dog kidney cortex membranes in contrast to atnal natriuretic peptide1 cardiolatm ( a - W / CDD,,,,). FEBS Lett 233; 249-254, 1988.
Ganda DP, Savin CT, Iber F, Glennon IA, Mitchell ML: Transient suppression of growth hormone secretion d e r chronic ethanol mtake. Alcohol Ch2 Erp Res 2; 297-299, 1978.
Gangdy 4 Chiou S. West L& Davis JS: Ahial natriuretic factor inhiiits angiotensgl-mduced aldoaerone secretion: Not through cGMP or interference with phospholipase C. Biochenz Biophys Res Cornmm 159; 148- 154, 1989.
Gangdy A: Atrial natriuretic peptide-mduced inhibition of aldosterone secretion: A quest for mediator(s). Am J Physiol 263; E 18 1-E 194, 1992.
Garcia R Tbi'bauit G, Nutt RF: Comparative vasoactive effects of native and synthetic atrial natriuretic factor (ANF). Biochem Biophys Res Comrntot 1 19; 685-688, 1 985.
Garcia R, Cantin M, Gutkowska J, Thibault G: Atrial natriuretic factor during development and reversal of one-kidney, one-clip hypertension. Hypertensiott 9; 144- 149, 1987.
Garcia R, Gauquelin G, Thiibault G, Cantin M, Schiffrm EL: Glomerular atrial natnuretic factor receptors in spontaneously hypertensive rats. Hypertemion 13; 567-574, 1 989.
Gardner DG, Hane S, Trachewsky D, Schenk D, Baxter ID: Atrial natriuretic peptide mRNA is regulated by glucocorticoids irt vivo. Bzochem Biophys Res Cornmwi 139: 1047- 1054. 1986a.
Gardner DG, Deschepper CF, Ganong WF, Hane S, Fiddles J, Baxter ID, Lewicki I: Extra- atrial expression of the gene for atrial natriuretic factor. Proc Natl Acad Sci USA 83 ; 6697- 6701, 1986b.
Gardner DG, Deschepper CF, Baxter JD: The gene for the atrial natriuretic factor is

expressed in the aomc arch. Hypertertîioti 9; 103- 106, 1987.
Gardner DG, Wu P, iapomte MC, Hedges BK, Deschepper CF: Expression of the gene for the a t r d n a h e t i c peptide m cardiac myocytes in vitro. Curdiov h g s & m e r 2: 479-486. 1988.
Gauqueh G, Schifnin EL, Cantin M, Garcia R: Spe&c bmding of atrial natriuretic factor to rend glomenili m DûCA- and DOCA-sdt-treated rats. Correlation with atnd and plasma levels. Biiochem Biophys Res Commun 145 ; 522-5 3 1. 1987.
Gauqueh G, Gutkowska J, Vmcent M, Sassard J, Ghani C: Atrial natriuretic factor: Plasma concentration and specific bmding to rend glornenili during the development of genetic hypertension m rats of the Lyon suiab. Comp Biochem Physiol 1 OSA; 49-5 5. 1993.
Gaziano JM, Buring JE, Bredow .IL, Goldhaber SZ Rosner B, VanDenburgh M, Wiuett W. Hennekens CH: Moderate alcobol mtake, hcreased levels of hi&-density iipoprotein and its subfiactions, and decreased risk of myocardial infaction. N Etlgl J Med 329; 1829- 1834. 1993.
Genazzani AR, Nappi G, Facchinetti F, Mauella GL, Parrini D, Sidoriani E, Petraglia F. Savoldi F: Central deficiency of &endorphin in alcohol addicts. J Clin Endocrirzol Metab 5 5 : 583-586, 1982.
Gerbes AL, Vollmar AM: Degradation and clearance of atnal natriuretic factor (ANF). Lrfe Sci 47; 1 173- 1 180, 1990.
Gianoulakis C, Barcomb A: Effect of acute ethanol in vivo and 112 vitro on the O-endorphin syaem m the rat. Life Sei 40; 19-28, 1987.
Gianoulakis C: Effect of prenatal exposure to ethanol on body growth and the pituitary 0- endorphin. Alcohol Clhl Exp Res 11; 567-573, 1987.
Gianodakis C: The effect of ethanol on the biosynthesis and regulation of opioid peptides. Experzetztzu 4 5 ; 42 8-43 5, 1 989.
Gianoulakis C: Endogenous opioids and excessive aicohol connunption. J Psychian Neiirmci 18; 148-156, 1993.
Giauoulakis C, Krishanan B, Thavumdayd J: Ehanced sensitivity to pituitary 8-endorphin to ethanol in subjects at high risk of alcoholism. Arch Gen Psyfh 53: 250-257, 1996.
Giles TD, Cook JR, Sachitano RA, Iteld BJ: infiuence of alcohol on the cardiovascular response to isometric exercice in normal subjects. Angiology 33; 332-33 8, 1982.

Gill K, France C, Amit 2: Voluntary ethanol consumption: an examination of blood/bram, ethanol leveis and behavior. Alcohol Clin Erp Res 10; 457-462, 1986.
Gïllman MW, Cook N& Evans DA, Rosier B, Heanekens CH: Relationshrp of alcohol intake with blood pressure in young aduhs. Hypertension 25; 1 106- 1 1 10, 1995.
Giridhar J, Peoples RW, Isom GE: Modulation of hypothamalic norepmephrine release by atrial natriuretic peptide: Involvement of cyclic GMP. Eur J Phonnacol 2 13; 3 17-3 2 1 . 1992.
Gleiiemiao L, Iiart,urg E: Alcohol usage and blood pressure: A review. Hum Biol58: 1-3 1. 1986.
Gnadinger MP, Lang RE, Hasler L, Uehhger DE, Shaw S, Weidmann P: Plasma kmetics of synthetic alpha-human a t d naûiuretic peptide m man. Miner EIectroiy~e Metub 1 2 ; 37 1-3 75. 1986.
Goetz KL: Renal natriuretic peptide (urodilatm?) and atriopeptm: Evohing concepts. Am J PhyszoI 26 1 ( R e d Fhid Electrofjte Physioi 30); F92 1-F932, 199 1 .
Goldsmith SR, Dodge D: Response of plasma vasopressin to ethanol in congestive heart failure. Am JCardiol55; 1354-1357, 1985.
Goldstein DB: Ethanol-mduced adaptation in biological membranes. Am NY Acad Sci 192; 103-11 1, 1987.
Goldstein DB: Pharmacokinetics of alcohol. p. 25-47, 1992.
Gordon GG, Southren AL, Altman K, Rubin E, Lieber CS: The effect of alcohol (ethanol) administration on sex hormone metabolisrn in normal men. N Engl J Med 295; 793-797. 1976.
Gordon T, Castefi WP, Hjonland MC, Kannell WB, Dawber TR: Hi& density lipoprotein as a protective hctor agamst coronary heart diseaçe: The Framingham study. Am J Med 62: 707-714, 1977.
Gordon GG, Southren AL, Vittek J, Lieber CS: The effect of alcohol ingestion on hepatic aromatase a c t ~ t y and plasma steroid hormone in the rat. Metabolism 28; 20-24, 1979a.
Gordon GG, Southren AL, Lieber CS: Hypogonadism and feminization in the male: A triple effect of alcohol. Alcohol Clin Exp ïher 3; 2 10-2 12, 1979b.
Gordon GG, Vittek J, Southren AL, Munnangi P, Lieber CS: Effect of chronic alcohol ingestion on the biosynthesk of steroids m rat testicular homogenates in vitro. Eridocrino~ogy

Gordon T, Kame1 WB: hmkmg and its relation to smoking, BP, blood lipids and uric acid. Arch Intem Med 143; 1366-1374, 1983.
Gordon T, Doyle JT: Alcohol consumption and its relationship to smoking, weight, blood pressure, and blood lipids: the Albany study. Arch h e m Med 146; 262-265, 1986.
Gordon AS, Collier K, Diamond 1: Ethanol regulation of adenosine receptor-dependent CAMP levels m a clond neural cell line: An in vitro mode1 of cellular tolerance to ethanol. P m Natl Acad Sei USA 83; 2105-2108, 1986.
Gordon DJ, Probstfield JL, Garrison RJ, Neaton JD, Castefi WP, Knoke JD, Jacobs D R Bangdnivala S, TyroIer HA: High-den* Lip opro tem cholesterol and cardiovascular disease: Four propective Amencan studies. Circularion 79: 8- 1 5, 1989.
Godd L, Zahir M, DeMartino Gomprecht RF: Cardiac effects of a cocktail. JAU4 2 18; 1799-1802, 1971.
Gower WR Jr, Chiou S, Skolnick K, Vesely DL: Molecular fomis of circulating atrial natriuretic peptides in human plasma and their metaboütes. Peptides 15; 86 1-867, 1994.
Grant KA, Valverius P, Hudspith M: Ethanol withdrawal seizures and the NMDA receptor cornplex. Etrr J Pharmacol 176; 289-296, 1990.
Grassi GM, Somers VK, R d WS, Abboud FM, Mark AL: Effects of alcohol intake on blood pressure and sympathetic nerve actMty in nomoteusive humans: a prelimmary report. J Hypertem 7 (suppl6); S20-S2 1, 1989.
Greenberg ST, Xie J, Wang Y, Kolls J, Shellito J, Nelson S, Summer WR: Ethanol relaxes pulmonary artery by release of prostaglandin and nitric oxide. Akohol 10; 2 1-29, 1993.
Greenburg BD, Bencen GH, Seiihamer JL, Lewicki JA, Fiddes JC: Nucieitide sequence of the gene encoding human atrial natriuretic factor precursor. Nature 3 12; 656-658, 1984.
Greenwald JE, Apton M, Hruska Kq Needleman P: Stretch-mduced atriopeptin secretion in the isolated rat myocyte and its negative regdation by calcium J Clin Imest 83; 106 1- 1065, 1989.
Greenwald JE, Ritter D, Tetens E, Rotwein PS: Rend expression of the gene for atrial natriuretic fiidor. Am J Physzoi 263 (Rend Fluid Efectrolyte Physiol32); F974-F978, 1992.
Groban L, Cowley AW, Ebert TJ: Atrial natriuretic peptide augments forearm capillary

filtration m humans. Am J Physiol259; W58-21263, 1990.
Grsnba~k M, Deis A, Snrrensen TI& Becker U, Borch-Johnsen K, Müller C, Schnohr P, Jensen G: Innuence of sex, age, body mass index and smoking on alcohol mtake and mortality. BMJ 308; 302-306, 1994.
Gmchow HW, Sobocmski KA, Barboriak JJ: Alcohoi, nutrient btake, and hypertension in U S . adults. J M 253; 1567- 1570, 1985.
Guaza C, Borrell S: Adrenomedullaryn responses to acute and chronic ethanol administration to rats. Biarhem Pharmacol 32: 3091-3095, 1983.
Gulati 4 Srimal RC, Bhargava HN: Effect of v a w g concentration of ethanol on systemic hemodynamics and regional circulation. Alcohol6; 9- 15, 1989.
Gundlach AL, Knobe KE: Distribution of preproatrial natriuretic peptide mRNA m rat brain detected by m situ hybridization of DNA oligonucleotides: E~chment m hypothalamic and h b i c regions. J Neurochern 59; 755-76 1, 1992.
G h g ME, Brenner BM: Natriuretic peptides and the kidney: Current concepts. Kidriej It~t 42; $127-S133, 1992.
Gunnhg ME, Brady t[R Otuechere G, Brenner BM, Zeidel ML: Atrial natriuretic peptide (3 1-67) inhiiits Na' transport in rabbit ber medullary collecting duct cells: Role of prostaghdm E2. J C h Imest 89; 14 1 1- 14 17, 1992.
Gutkowska J. Thibault G. lanuszewicz P, Cantin M, Genest J: Direct radioimmunoassay of Atrial Natriuretic Factor. Biochem Biophys Res Commun 121: 593-60 1. 1 984.
Gutkowska J, Bourassa M, Roy D. Thibault G. Garcia R. Cantin M. Genest J: lmmunoreactive atrial natriuretic factor (IR-AND in human plasma. Biochem Biophvs Res Commun 128; 1350-1357. 1985.
Gutkowska I, Horky K. Lachance C, Racz K, Garcia R, Thibault G, Kuchel O. Genest J, Cantin M: Atrial Natriuretic Factor in spontaneously hypertensive rats. Hypertension 8 [suppi 1); 1137-1140. 1986a.
Gutkowska J, Racz K, Garcia R, Thibault G, Kuchel O, Genest J, Cantin M: The morphine effect on plasma ANP. Eur J Pharmacol 13 1 ; 9 1-94, 1986b.
Gutkowska I: R a d i o ~ o a s s a y for atrial natriuretic factor. 1nt J Rad (Nucl Med Biol) 14: 323-33 1, 1987.

Gutkowska J, Genest I, Thibautt G, Garcia R, Larochelle P, Cusson JR, Kuchel O, Hamet P. De Léan A, Cantin M: Circulating forms and radioimmunoassay of Atrial Natriuretic Factor. End & Met Clin of N Am 16: 183- 198, 1987a.
Gutkowska J, Racz K, Debmski W, Thibadt G, Garcia R, Kuchel O, CantIn M, Genest J: An atrial natriuretic factor-like activity m rat posterior hypophysis. Peptides 8; 46 1-467, 1987b.
Gutkowska J, Cantin M, Genest J, Sirois P: Release of immunoreactnle atrial natriuretic factor fiom isolated perfùsed rat lungs. FEBS Let? 2 14; 17-20, 1987~.
Gutkowska J, Cantin M: Bioactive atrial natriuretic factor-like peptides in rat antenor pihlltary. Cm JPhysiol Pharmacol 66: 270-275, 1988.
Gutkowska J, Carrier F, St-Louis I, Thiiault G, Cantin M, Genest J: Radioreceptor assay for atrial natrketic factor. Aarzul Biochenl 168; 100- 106, 1988.
Gutkowska J. Nemer M: Structure, expression, and function of atrial natriuretic factor in extraatriai tissues. Erzdocr Rev 10; 5 19-536, 1989.
Gutkowska J, Marcmkiewicz M: Does the olf'actory system mediate water- and minerai- regukting mechanisns? Evidence of immunoreactive atrial natriuretic factor within olfactory mucosa. J Neurochenz 53; 1830- 1837, 1989.
Gutkowska J, Nemer M, Sole MI, Drouin J, Sirois P: Lung is an important source of atnal natriuretic factor in experimental cardiomyopathy. J C h irzvest 839; 1 5 00- 1 504, 1989.
Gutkowska I, Tremblay J, Meyer R Marcinkiewkz M, Nemer M: Evidence for atrial natriuretic peptide (ANP) synthesk and the presence of ANP-transducing receptors in the rat olfactory bulb. J Netirochem 57; 1855- 186 1, 199 1.
Gutkowska J, Tremblay J, Antakly T, Meyer R, Mukaddam-Daher S, Nemer M: The atnal natnuretic peptide syaem m rat ovaries. Erldocrirtology 132; 693-700. 1993.
Guyton AC: Artenal pressure and hypertension. W.B Saunders, Philadelphia, 1980.
Gyntelberg F, Meyer J: Relationship between blood pressure and physical h e s s . smoking and alcohol consumptioo in Copenhagen males aged 40-59. Acta Med Scatzd 195: 375-380. 1974.
Haanwinckel MA, Elias LK, Favaretto AL, Gutkowska J, McCann SM, Antunes-Rodrigues J: Oxytocin mediates atriai natriuretic peptide release and natriuresis after volume expansion in the rat. Proc Natl Acad Sci USA 92; 7902-7906, 1995.

Haass M, Fisher TA, Dietz R: 1s atrial distension the physiological stimulus for release of atrial natriuretic peptide? h c e i 2: 1269- 1270, 1987.
Hafher SM, Applebaum-Bowden D, Wahl PW, Hoover JJ, Warnick GR, Abers JJ, Hazzard W l t Epidemiological correlates of hi& d e n e lipoprotem subfractions, apohprotebs A-1. A-II, and D, and lecahm choleaerol acyltransferase. ArterimcIermis 5 ; 169- 177, 1985.
Harburg E, Erfurt JC, Hauenstein LS, Chape C, S c h d WJ, Schork MA: Socio-ecological stress, suppressed hostility, s h color, and Black-White d e blood pressure. Detroit Psycharom Med 35; 276-296. 1973.
Harburg E, Ozgoren F, Hawthorne VM, Schork MA: Commimity noms of alcohol usage and blood pressure. Am J Public Healih 70; 8 1 3-820, 1 980.
Harburg E, Gleiberman L, Dif?anceisco W, Peele S: Toward a concept of sensible drinking and an illustration of measure. Alcohol A l c o h o h 29; 439-450, 1994.
Harlan WR, Hd AL, Schouder RL, Landis ni Thompson FE, Larkin FA: Blood pressure and nutrition in adults. Am J Epidemiol 120; 17-28. 1984.
Harris PI, Skinner SL: Intra-renal interactions between angiotensin II and atrial natnuretic factor. Kidriey Ir21 38; S87-S9 1, 1990.
Hasegawa K, Fujiwara H, Itoh H, Nakao Y Fujiwara T, Imura H, Kawai C: Light and electron microscopie localization of brain natriuretic peptide in relrtion to atrial natnuretic peptide in porcme atrium Cirnrlutio~z 84; 1 203- 1209, 199 1.
Haskell WL, Camargo C, Williams PT, Vranizan KM, Krauss RM, Lindgren FT. Wood PD: The effect of cessation and resumption of rnoderate alcohol intake on senun high-density- lipoprotem subfiactions: A controlled study. N Er@ J Med 3 10; 805-8 10, 1984.
Hasumura Y, Teschke R Lieber CS: Acetaldehyde oxidation by hepatic mitochondria: its decrease after chronic ethanol consumption. Science 189; 727-729, 1975.
Hatake K, Wakabayashi l7 Kakishita E, Taniguchi T, Ouchi I3, Sakaki N, Hishida S: uihr'bitory effect of ethanol on endothelium-dependent vascuiar responsiveness. E w J Phamtacol 168; 277-283, 1989.
Hatake K, Wakabayashi 1, Hishida S: Mechanism of inhiitory action of ethanol on endothelium-dependent relaxation m rat aorta. Eur J Phamrucol 23 8; 44 1-444, 1993.
Hatake Y Wakabayashi I, Taniguchi T, Hîshida S: increased endothelium-dependent vascular relaxation in ehtanol-fed rats. Alcohol Cliri Exp Res 18; 10 18- 1023, 1994.

Hatton DC, Bukoski RD, Edgar S, McCarron DA: Chronic alcohol consumption lowers blood pressure but enhances v a d a r contractiüty in Wistar rats. J H'ertens 10; 529-537. 1992.
Hattori Y, Kasai M, Uesugi S, Kawata M, Yamashita H: Atrial natriuretic polypeptide depresses angiotensin II induced excitation of neurons m the rat subfomical organ in vitro. Brain Res 443; 355-359, 1988.
Hawkins R, Kalant H, Khama JM: Effects of chronic Mtake of ethanol on rate of ethanol metabolism Can J Physiol Pharmacol 44: 24 1-257. 1966.
Hawley RI, Major LF, Schulman E, Linnoila M: Cerebrospinal fluid 3-methoy-4- hyckoxyphenylglycol and norepinephrine levels m alcohol withdrawal. Arch Gen Psychatr 42; 1056- 1062, 1985.
Herman JP, h g u b MC Jr, Watson RE Jr: Localization of C-type natriuretic peptide mRNA in rat hypothalamus. Endocrinology 133; 1903- 1906, 1993.
Heublein DM, Clavell AL, Stmgo AJ, Lemian A, Wold L, Bumett JC: C-type natriuretic peptide immunoreaaivity in human breast vascular endothelial ceils. Peptides 13 : 10 1 7- 1 O 1 9. 19%.
Higuchi K, Nawata H, Kato K, Ibayashi H, Matsuo H: Alpha-human atnal natriuretic polypeptide mhibits steroidogenesis m cultureci human adrenal cells. J Ch1 Etidocriml Metab 62; 94 1-944, 1986.
Himeno A, Niwa M, Nakao K, Suga S, Yamashita K, Kataoka Y, Nakane Y, h u a H. Taniyama K: C-type natriuretic peptide-22 Werentiates between natriuretic peptide receptors in rat choroid plexus and subfomical organ. Eur J Pharmacol 2 15; 337-340. 1992.
Hira GK, Pang SC, Sarda Watson JD, Flynn TG: Tissue distribution and cellular location of iso-rANP (rat-BNP45) m the rat. FASEB J 5; A1020, 199 1.
Hirata M, Chang CH., Murad F: Stimulatory effects of atrial natriuretic factor on phosphomosinositide hydrolysis in cultured bovine aortic smooth muscle ceiis. Biochim Biophys Acta 10 10; 346-35 1, 1989.
Hirata Y, Fukuda Y, tida T: Endothelin-1 stimulates secretion of atrial and brain natriuretic peptides f?om porcine cardiocytes and mcreases ANP messager mRNA levels in rat cardiocytes. J Hypertem 8; S92, 1990.
Hoemer M, Behrens UJ, Womer T, Lieber CS: Humoral immune response to acetaldehyde adducts m alcoholic patients. Res Comrnuri Chem Pathol Phormacol 54; 3- 12, 1986.

H o b P, Ritmiann R, Walter R, Tabakoff B: Arginme vasopressh maintains ethanol tolerance. Narure 276; 6 14-6 16, 1978.
Elofhm PL, Rabe CS, Moses F: N-methyl-Baspartate receptors and ethanol: Inhi'bition of calcium fhix and cyclic GMP production. J Neurochern 52; 1937- 1940, 1989.
Ho* PL, Tabakoff B: Ethanol and guanine nucleotide bhdmg protems: a selective interaction. FASEB J 4; 26 12-2622, 1990.
Hojnacki JL., CluetteBrown JE, Mdigan JJ, Hagan SM, Mahony KE, Witzgall SK, Osmolski TV, Barboriak JJ: Effect of ethanol dose on low d e n e lipproteins and hi& den* lipoprotein subf?actions. Alcohl C h Exp Res 12; 149- 154, 1988.
Hollister AS, Inagami T: Atrial Natriuretic Factor and hypertension. Am J Hypertens 4: 850-865, 199 1.
Holmes SJ, Espiner EA, Richards AM, Yandle TG, Frampton C: Renai, endocrine, and hemodynamic effects of human brain natriuretic peptide m n o m l man. J Clin Endocrino1 Metab 76; 9 1-96? 1993.
Horio T, Kohno M, Takeda T: E f f i s of arginine vasopressin, angiotensin iI and endothelin- 1 on the release of brain natriuretic peptide in vivo and in vitro. Clin Exp Phormacol Physiol 19; 575-582, 1992.
Horiuchi M., Kohashi N, Nishryama H, Hama J, Takenada T, Kondo H, Katori R: Regdation of aldosterone receptor m rat kidney cytosol by atnal natriuretic factor. Hypertetlsiorl 13: 334-340, 1989.
Horky Y Gutkowska J, Garcia R, Thibault G, Genea .J, Cantin M: Effect of different anesthetics on immunoreactive aaial natriuretic factor concentrations in rat plasma. Biochenz Biophys Res Commun 129; 65 1-657, 1985.
Ho& LD, Atkins SM: Acute effëcts of ethanol on lefi ventricular p erforrnance. Cirnilatiori 49; 124- 128, 1974.
Howe PRC, Rogers PF, Smith RM: Antihypertensive effect of alcohol m spontaneousiy hypertensive rats. Hypertension 13; 607-6 1 1, 1989.
Howes LG, Reid JL: Changes m plasma free 3,4-düiydroxyphenylethylene glycol and noradrenaline levels f i e r acute alcohol administration. Clin Sci 69; 423-428, 1985.
Howes LG, MacGilchna A, Hawksby C, Sumner D, Reid JL: Plasma ['Hlnoradrenaline künetics and blood pressure following regular, moderate ethanol consumption. Br J Clzn

Hsieh S, Sano H, SaÏto K, Kubota Y, Yokoyama M: Magnesium supplementation prevents the development of alcohol-mduced hypertension. Hypertensiorr 19; 175- 1 82, 1992.
Hunt WA: Neuroscience research: How has t contniuted to our understanding of alcobol abuse and alcoholism? A review. Akohol CIin Exp Res 17; 1055- 1065, 1993.
Hutchison WD, Gianoulakk C, Kalant H: Effects of ethanol withdrawal on f3-endorphin levels in rat brah and p i d a r y . Pharmacol Biochem Behav 30: 933-939, 1988.
Hynynen M, Kupari M, Salmenperâ M, Koskien P, Tikkanen 1, Fyhrquist F: Plasma atrial natriuretic factor during ethanol mgestion m volume-loaded subjects. Akohol Aicoholim 27: 277-285, 1992.
Hyytia P, Smclair JD: Responchg for oral ethanol after naloxone treatment by alcobol- prefening AA rats. Aicuhol Clin Erp Res 17; 63 1-636, 1993.
Iibsen H, Christenme NJ, Rasmussen S, Hohagel H, Nielsen MD, Giese J: The influence of chronic high alcohol mtake on blood pressure. piasna noradrenaline concentration and plasma renin concentration. Clin Sci 6 1 ; S3774379, 198 1.
Iida T, Hirata Y, Takemura N, Togashi Y Nakagawa S, Marumo F: Bram natriuretic peptide is cosecreted with atrial natriuretic peptide fiom porcine cardiocytes. FEBS Lett 260; 98- 100. 1990.
litake K, Share L, Crofton JT, Brooks DP, Ouhi Y, Blaine EH: Central atrial natriuretic factor reduces vasopressin secretion in the rat. Etdocrinoiogy 1 19; 438-444, 1986.
Ikeda Y Nara Y, Yamori Y: Indirect systolic and mean blood pressure determination by a new tail-cuffmethod in spontaneously hypertensive rats. Lab Ar2inral.s 25; 26-29. 199 1.
imada T, Takayanagi R, Inagami T: Changes in the content of atrial natriuretic factor with progression of hypertension m spontaneous hypertensive rats. Biochem Biophys Res Conznlw~ 133; 759-765, 1985.
Imura H, Nakao K, Itoh H: The natriuretic peptide system in the brain: implications in the central controt of cardiovascular and neuroendocrine fiinctions. Fr Neuroerrdarirlol 1 3 ; 2 1 7- 249, 1992.
Inagami T, Misono KS, Fukumi FI., Maki M, Tanaka J., Takayanagi R Imada T, Grammer RT. Naruse M, Naruse K, Pandey KN, Parmentier M, Yasujima M, Abe K: Structure and physiological actions of rat atrial natriuretic factor. Hypertensiotz 10 (suppl. 1); Il 13-1 1 17.

Inoue M, Kimura T, Ota K, Iitake K, Shoji M, Sato K, Ota M, Yoshinaga K: Effect of vasopresb m striai natriuretic peptide release and rend functioos m dogs. Am J Physiol255: E449-E455, 1988.
Inoue M, Kimut-a T, Ota K: Effect of atnal natriuretic peptide on the vasopressin release to osmotic and hemorrhagic stimuli m dog S. J Neuroendocrir10l 2; 903-9 1 0 , 1 990.
Iono K R Reinliib L. Tabakoff B: Chronic exposure of cerebellar granule cells to ethanol resuhs m mcreased N-methyl-Daspartate receptor fùnction. M d Phormacol 4 1 ; 1 142- 1 148. 1992.
Ireland MA, Vandongen R Davidson L, Beilin U, Rouse IL: Acute effects of moderate alcohol consumption on blood pressure and plasma catecholamines. C h i Sci 66; 643-648. 1984.
Ishii Y, Watanabe T, Watanabe M, Hasegawa S, Uchiyama Y: Effects of atrial natriuretic peptide on Type II h e o l a r epithelial cens of the rat h g . J A m t 166; 85-95, 1989.
Israel Y, Kalant H: Effect of ethanol on the transport of sodium in frog skm. Nature 200: 476-478, 1963.
Israel MA, Kimura H, Kuriyama K: Changes in actNiry and hormonal sensit~ty of brain adenyly cyclase following chronic ethanol administration. Experieiitia 28; 1322- 1323. 1972.
Israel Y, Waifish PG, Orrego H., Blake S, Kalant H: Thyroid hormones in alcoholic h e r disease. Gustroenrerology 76; 1 16- 122, 1979.
Israel A, Barbella Y: Diuretic and natriuretic action of rat atrial natriuretic peptide (6-33) administered cerebroventricularly in rats. Brairi Res Bull 1 7: 14 1 - 144, 1 986.
Itoh H, Nakao K, Morü N, Yamada T, Shiono S, Sakamoto M, Sugawara A, Saito Y. Katsuura G, Shiomi T, Eigyo M, Matsushita A, [mura H: Central action of atrial natriuretic polypeptide on blood pressure in conscious rats. Brait1 Res Bull 16; 745-749. 1986a.
Itoh H, Nakao G, Katsuura N, M o i N, Shiono S, Sakamoto M, Sugawara 4 Yamada T. Saito Y, Matsushita [mura H: Centrally infùsed atrial natriuretic polypeptide attenuates exaggerated salt appetite in spontaneously hypertensive rats. Circ Res 59; 342-35 1 , 1986b.
Itoh H, Nakao K, Yamada T, Morii N, Shiono S, Sugawara A, Saito Y, Mukoyama M, Arai H, Katsuura G, Imura H: Modulatory role of vasopressin m secretion of atrial natriuretic polypeptide in conscious rats. E~2da:rinology 120; 2 186-2 188, 1987.

Itoh H, Nakao K, Saito Y, Yamada T, Shirakami G, Mukoyama M, Arai H, Hosoda K. Suga S, Mmamho N, Kangawa K, Matsuo H, h u a H: Radioimmunoassay for brain natriuretic peptide (BNP). Detection of BNP m canine brain. Biochem Biophys Res Commun 1 5 8: 120- 135, 1989.
Itoh H, Nakao K, Mukoyama M, Arai H, Yamada T, Sato Y, Shiono S, Hosoda K. Shitakami G, Suga S, Imura H: Pathophysiological role of augmented atrial natnuretic polypeptide gene expression m WCA-sah hypertensive rats. Am J Hyperem 4 ; 39-44, 199 1 .
Itoh H, Sagawa N, Hasegawa M, Nanno H, Kobayashi F, Ihara Y, Mori T, Kornatsu Y. Suga S, Yoshimasa T, Itoh FI, Nakao K. Expression of biologically active receptors for natriuretic peptides m the human uterus during pregnancy. Biochern Biophys Res Commun 602-607. 1994.
Jackson R, Stewart A, Beaglehole R, Scragg R: Alcohol consumption and blood pressure. Am J Epidemiol 122; 1034- 1044, 1985.
James MJ, Walsh JA: Effects of aspirin and alcohol on platelet thromboxane synthesis and vascular prostacyclin synthesis. Thrombosïs Res 39; 587-593, 1985.
Jarnieson JD, Palade GE: Specific granules in attial muscle ceUs. J Ce11 Biol23; 15 1- 172. 1964.
Jamison RL, Canaan-KUhl S, Pratt R: The oatriuretic peptides and their receptors. A m J Kidney Dis 20; 5 19-53 1, 1992.
Jankowski M, Petrone C, Tremblay J, Gutkowska J: Natriuretic peptide syaem m the rat submadary gland. Reg Pepr 62; 53-6 1 , 1996.
Januszewicz P, Thiiault G, Garcia R, Gutkowska J, Genest J. Cantin M: Effect of synthetic atrial natnuretic factor on arginine vasopressh release by the rat hypothalamoneurohypophyd complex m organ culture. Biochem Biophys Res Conrnruri 1 34: 652-658, 1986a.
Januszewicz P, Larose P, Ong H, Gutkowska J, Genest I, C a t h M: Effect of atnal naûiuretic factor on plasna vasopressin and oxytocin m the rats. Peptides 7; 989-993. 1986b.
Jauhonen P, Baraona E, Miyakawa H: Mechanism for the selectivwe perivenular hepatotoxicity of ethanol. Alcohol Cliri Exp Res 6; 350-357, 1982.
Javesbghani D, MukaddamDaher S, Fan L, Guan Z, Gutkowska J, Nuwayhid B, Quillen EW Jr: Control of atrial natriuretic factor by right and left atrial distension m pregnant sheep. Am JPhysiol268; RI41 1-R1417, 1995.

Jenkins JS, Connelly J: Adrenocortical response to ethanol m man. Br Med J 2: 804-805, 1968.
Jennings DB, McKirdy IC, Ohtake PJ, Sarda IR, Flynn TG, Hira GK: infusion of ios- rANP(1-45) or (17-45) mcreases plasma immunoreactive ANP and lowers plasma renin actMty and aldosterone. Cati J Physiol Pharmacol 68; 1292- 1297, 1990.
Jin y Yang R Chen YF, Wyss M, ûparil S: Ahered stores of atrial natriuretic peptide in specific brain nuclei of NaCCsensitive spontaneously hypertençive rats. Am J Hypertetu 4: 349-455, 1991.
Johnson GR Arik L, Pitts Bm Foaer CJ: Rapid receptor mediated catabolimi of '2S~-atrial natriuretiic factor by vascular endothelia1 ceils. Biochem J 268; 77 1-776, 1990.
Jones JV, Raine AEG, Sanderson JE, Carretta R, Graham DI: Adverse effect of chronic alcohol ingestion on cardiac performance in spontaneously hypertensive rats. J Hypertens 6; 4 19-422, 1988.
Jones AW: Excretion of alcohol in urine and diuresis in healthy men in relation to their age, the dose adnhistered and the tirne after drinking. Forens Sei Int 45 : 2 17-224, 1 990.
IuIlamen RJP, Di Padova C, Lieber CS: First pass metabolkm of ethanol: a gaaromtestinal barrier agamst the systemic toxicity of ethanol. Life Sci 37; 567-573. 1985.
Kagan 4 Yano K, Rhoads GG, McGee DL: Alcohol and cardiovascular disease: the Hawaiian experience. Circulatio~r 64 (suppl. 3 ); 27-3 1, 198 1.
Kalant H, LeBlanc AE, Gibbins RJ: Tolerance to, and dependence on, some non-opiate psychotropic drugs. Pharmacol Rev 23: 13 1- 19 1, 197 1.
Kalsmer S: Potentiating effects of ethanol on responses of aortic strips to stimulant drugs. J P h P h a c o l 2 2 ; 877-879, 1970.
Kanazawa, Casley DJ, Sybertz EJ, Haslanger MF, Johnson CI: Locaiization and characterization of neutrd metalloendopeptidase (EC 3.4.24.1 1 ), the degradative enzyme for atnal natriuretic peptide, in rat kidney using a radioiodmated neutral metalloendopeptidase inhibitor. J Pharmacol Exp Ther 26 1; 123 1- 1237, 1992.
Kaneko T, Shirakami G, Nakao K, Nagata 1, Nakagawa O, Hama N, Suga S, Miyamoto S. Kubo H, Hirai O: C-type natriuretic peptide (CNP) is the major natriuretic peptide m human cerebrospinal fluid. Brazn Res 6 12; 104- 109, 1993.
Kangawa K, Matsuo R M c a t i o n and complete amino acid sequence of a-human natriuretic

polypeptide (a-hANP). Biochern Biophys Res Commun 118; 13 1-139, 1984.
Kaplan NM: Bashmg booze: The danger of losing the benefits of moderate alcohol consumption. Am k t J 12 1 ; 18544856, 199 1.
Karanian JW, Stojanov M, Salem N Jr: Effect of ethanol on prostacyclin and thromboxane A2 synthesis m rat aortic rings in vitro. Prostagi Leukot Med 20; 1 75- 1 86, 1985.
Karanian JW, D'Souza NB, Salem N Jr: The effect of chronic alcohol inhalation on blood pressure and the pressor response to noradrenaline and the thromboxane-mimic U466 19. L f e Sei 39; 1245- 1255, 1986.
Karanian JW, Salem N Jr: The effect of alcohol inhalation on the cardiovascular state of the rat. A h Aicohol Subst Abuse 7 ; 22 1-225, 1988.
Kasting NW, Veale WL, Cooper KE: Convulsive and hypothermie effects of vasopressin in the bram of the rat. Cap2 J Physiol Pharmacol 58; 3 16-3 19, 1980.
Katahchi T, Minino T, Hagiwara H, Itakura M, Itoh T, Hirose S: Modulation by NaCl of atrial natriuretic peptide receptor levels and cyclic GMP responsiveness to atnal natriuretic peptide of cultured vascular endothelia1 ceus. J Biol Chem 267; 762497629, 1992.
Katsube N, Schwartz D, Needleman P: Release of atnopeptin in the rat by vasoconstrictors or water immersion correlates with changes in right atrial pressure. Biochem Biophys Res Cornmur2 133; 937-944, 1985.
Katsube N, Schwartz D, Needleman P: Atriopeptin turnover: Quantitative relationship between m vivo changes m plasma Ievels and atrial content. J Pharmacol Exp mer 239: 474- 479, 1986.
Katsuura G, Nakamura M, lnouye K, Kono M, Nakao Y Lmura H: Regulatory role of atrial natriuretic polypeptide m water drinkmg m rats. Eur J Pharmacol 12 1 ; 285-287, 1986.
Kawawo S, Masuda E, Tsuji S, Nagano K, Fusamoto H, Kamada T: Ethanol causes vasoconstriction due to endothelin- 1 release m rabbit gastric vessels. Microvasc Res 4 1 ; 408- 410, 1991.
Kawano Y, Abe H, Kojima S, Ashida T, Yoshida K, Imanishi M, Yoshimi H, Kirnura G, Kuramochi M, Omae T: Acute depressor effect of alcohol in patients with essential hypertension. Hypertensiofi 20; 2 19-226, 1992.
Kawata M, Nakao K, Morii N, Kiso Y, Yamashita H, h a H, Sano Y: Atrial natriuretic polypeptide: Topographical distribution m the rat brah by radioimmunoassay and

Keil U, Chambless L, Remmers A: Alcohol and blood pressure: resuhs nom the Luebeck Blood Fressure study. Prev Med 18; 1-10, 1989.
Keil U, Chambless L., Filipiak B, Hartel U: Alcohol and blood pressure and Rs interaction with smoking and other behavioural variables: results fiom the MONICA Augsburg survey 1984- 1985. J Hpertem 9: 49 1-498, 199 1.
K e i h D, Hartree EF: Properties of catalase: cataiysis of coupled orcidation of aicohols. Bzochem J 39; 293-30 1, 1945.
Kem J: Alcohol and heart disease: The implications of the U-shaped curve. BMJ 307: 1373- 1374, 1993.
Kenny AJ, Boume A. Ingram J: Hydroiysis of human and pig natriuretic peptides, urodilatm, C-type natriuretic peptide and some C-receptor Ligands by endopeptidase-24-11. Biochern J 29 1; 83-88, 1993.
Kettunen R, T i j a M I, Saukko P: The acute dose-related effects of ethanol on nght ventricular h c t i o n in anesthetled dogs. Alcohol9; 149- 153, 1992.
Khalil F. Fie B, Kuriyama S: Increased atrial natriuretüc factor receptor density in cultured vascuiar smooth muscle c e k of the spontaneously hypertensive rats. Clin Exp Hypertem [A] 9; 74 1-749, 1987.
Khetarpal VK. Volicer L: Effects of ethanol on blood pressure of normal and hypenensive rats. J Stud Alcohol40; 732-736, 1979.
Kiberd BA, Larson TS, Robertson CR Iamison RL: Effect of atrial natnuretic peptide on vasa recta blood flow in the rat. Am J Physiol252; F 1 1 12-F 1 1 17, 1987.
Kimura y Hirata Y, Nanba S, Tojo A- Matsuoka FI, Sugimmamoto T: Effects of atnal natriuretic peptide in renal arterioles: Morphometric analysis using microvascular casts. Ani J Physiol259; F936-F944, 1990.
King MS, Baertschi AJ: Physiological concentrations of atrial natnuretic factors with intact N-terminal sequences inhibit corticotropin-releasing factor-stimulated adrenocorticotropin secretion £kom cultured antenor pihiitary cells. E~2docrirzology 124; 286-292, 1989.
King AC, Emco AL, Parsons 04 Lovallo WR Blood pressure dysregulation associated with alcohol withdrawaL Afcohol Clhz Exp Res 15 ; 478-482, 199 1.

Kmnunen P, Vuolteenaho O, Ruskoaho H: Mechanisms of atrial and brah natriuretic peptide release fiom rat ventricular myocardium: Effea of stretchmg. Endocrinology 132; 196 1 - 1966, 1993.
Kisch B: Electron microscopy of the a& of the heart: 1. Guinea pigs. Exp Med Stirg 14; 99- 1 12, 1956.
Klag MJ, He J, Whelton PK, Chen J. Qian M, He G: Alcohol use and blood pressure in an unaccdturated society. Hypertetzsiorz 22; 365-370, 1993.
Klatsky AL, Friedman GD, Siegeland AB, Gerard MJ: Alcohol consumption and blood pressure. N Etigf J Med 296; 1 194- 1200, 1977.
Klatsky AL, Friedman GD, Armstrong MA: The relationshrps between alcoholic beverage use and 0 t h traîts to blood pressure: A new Kaiser Permanente study. Cirnilatto~~ 73 : 628-63 6. 1986.
Knych ET: Endothelium-dependent torelance to ethanol-mduced contraction of rat aorta: effect of inhibition of EDRF action and nitnc oxide synthesis. Alcohol Chi Exp Res 16; 58- 63, 1992.
Kohno M, Yascmai K, Yokokawa K, Murukawa K, Horio T. Takeda T: inhibition by atrial and brain natriuretic peptides of endothelin- 1 secretion after stimulation with angiotensin II and thrombin of cdtured human endothelia1 cells. J C h lrwest 87; 1999-2004, 199 1.
K o h o M, Horio T, Yoçhivama M, Takeda T: Accelerated secretion of brai. natriuretic peptide fiom the hypertrophied ventricles in experimental malignant hypertension. Hypertetsioti 19; 206-2 1 1, 1992a.
Kohno M, Horio T, Toda i, Akioka K, Tahara A Teragaki M: Cosecretion of atrial and brain natriuretic peptides during supraventncular tachyarrhythas. Am Heurt J 123 ; 13 82- 13 84. 1992b.
Kohno M, Horio T, Yokokawa K, Murakawa K, Yasunari K, Akioka K, Tahara A, Toda 1, Takeuchi K, Kurihara N, Takeda T: Brain natriuretic peptide as a cardiac hormone in essential hypertension. Am J Med 92; 29-34, 1992~.
Kohno M, Fukui T, Horio T, Yokohawa K, Yasunari K, Yoshiyama M, Kurihara N' Takeda T: Cardiac hypertrophy and brah natriuretic peptide in experimental hypertension. Am J Physiol266; R45 1-R457, 1994.
Kojena M, MÏnamho N, Kangawa K, Matsuo H: Clonhg and sequence anaiysis of a cDNA encoding a precursor for rat C-type natriuretic peptide (CNP). FEBS Lett 276; 209-2 13.

Kojmia S, Kawano Y. Abe H., Sanai T, Yoshida K, Imanidii M., Ashida T, Kimura G, Yoshimi H , Matsuoka I3, Omae T: Acute effeas of alcohol ingestion on blood pressure and erythrocyte sodium concentration. J Hjpertem 1 1 ; 185- 190, 1993.
Kollenda MC, VoIhnar AM, McEnroe G 4 Gerbes AL: Dehydration mcreases the density of C receptors for ANP on rat glomerular membranes. Am J Physiol25 8 ; RIO84R 1088, IWO.
Koller KI, Lowe DG, Bennett GL, Minamino N, Kangawa K, Matsuo H, Goeddel DV: Selective activation of the B natriuretic peptide receptor by C-type natriuretic peptide (CNP). Science 252; 120- 123, 1991.
KolIer KJ, Goeddel DV: Molecdar biology of the natriuretic peptides and their receptors. Circulatiorz 86; 1081-1088, 1992.
KoUer KJ, De Sauvage FJ, Lowe DG, Goeddel DV: Conservation of the kinase-like regulatory domain is essential for activation of the natriuretic peptide receptor panylyl cyclases. Mol Cell Biol 12; 258 1-2590, 1992.
Kolls J, Deininger P. Cohen JC, Larson I: cDNA equalization for reverse transcription- polymerase chah reaction quantitation. Anal Biochern 208; 264-269, 1993.
Komatsu Y. Nakao K., Suga S. Ogawa Y, Mukoyama M, Arai H, Shirakami G, Hosoda K Nakagawa O, Hama N, Kishimoto 1, [mura H : C-type natriuretic peptide (CNP) in rats and humans. Endocrinology 129; 1 104- 1 106, 199 1 .
Komatsu Y, Nakao Y Itoh H. Suga S, Ogawa Y, h a H: Vascular natriuretic peptide. Lamet 340; 622, 1992a.
Komatsu y Tanaka 1, Funai T, Ichryarna A, Yoshimi T: Increased level of atrial natriuretic peptide messenger RNA in the hypothalamus and brainstem of spontaneously hypertensive rats. J Hypertem 10; 17-23, 1992b.
Kondo K, Ebihara A: Alcohol consumption and blood pressure in a rural community of Japan. in: Nutritional preventiorz ofcurdimmcuIar diseme, Lovenberg W , Yamori Y (eds), Orlando FL, Academic Press, p. 585-592, 1984.
Konrad EM, Thiiault G, Sc- EL: Autoradiographic Msualization of the natriuretic peptide receptor-B m rat tissues. Reg Peptides 39; 177- 189, 1992a.
Konrad EM, Thilault G, Schiffim EL: Atrial natriuretic factor bmding sites in rat area postrema: Autoradiographic study. Am J Physiol263 (Regulatory Inregroiive Comp PhysioI

Kornhuber HH, Lisson G, Suschka-Sauermann L: Alcohol and obegty: a new look at high blood pressure and stroke; an epidemiological midy m prwentive neurology. Eur Arch Psychzaby Neurol Sci 234; 357-362, 1985.
Koskinen P, Virolaben J, Kupari M: Acute alcohol intake decreases short-term heart rate vanabilay in heahhy subjects C h Sci 87: 225-230, 1994.
Kovacs G: a-atrial natriuretic peptide anmuates ethanol withdrawal symptoms. Etir J P h m a c o l 23 8; 4 17-4 19, 1993.
Kozararevic DI, Vojvodic N, Dawber T: Frequency of alcohol consumption and morbidity and mortality: the Yugoslavia Cardiovascular Dwase study. Lamet 1 ; 6 13-6 16, 1980.
Kreutz R, Higuchi M, Ganten D: Molecular genetics of hypertension. CIin Eiper Hypertem [A] 14; 15-34, 1992.
Krey LC, Lu Y Butter W, Hotchkiss J, Piva F, Knobil E: Surgical disco~ections of the media1 basal hypothalamus and pituRary function m the rhesus monkey: GH and cortisol secretion. Endocrinology 96; 1088- 1096, 1975.
Krieter PA, Trapani AI: Metabolism of atrial natriuretic peptide: Extraction by organs in the rat. Drugs Meîab & Disp 17; 14- 19, 1989.
Kromhout D, Bosschieter EB, Coulander CL: Potassium, calcium, alcohol intake and blood pressure: the Zutphen study. Am J Clin Nutr 4 1 ; 1299- 1 304, 1 985.
Kruck F, Krecke HJ: The renal sodium excretion during oral hydration in man. Xephrorr 2: 32 1-333, 1965.
Ku YH, Bang T: Brain atnopeptm mediates AV3V depressor response. Peptides 1 5 : 10% 1056, 1994.
Kuno T, Andresen I, Kamisaki Y, Waldman S, Chang L, Saheki S. Leitman D, Nakane M.
Kumik BRC, Weisberg LS, Askenase AD, Kumik PB: Mannitol atMulates atrial natnuretic peptide release in humans. Am J Kidriey Dis 17; 62-68, 199 1.
Kupari M: Acute cardiovascular e f f i s of ethanol: a controlled non-invasive study. Br Heart J49; 174-182, 1983.
Kurtz 4 Bruna RD, ffeilschifter J, Taugner R, Bauer C: Atnal natriuretic peptide inhibits

renin release fiom juxtaglomenilar cells by a cGMP-mediated process. Proc Nat1 Acad Sci USA 83; 476904773? 1986.
Landolfi q Stemer M: Ethanol raises prostacyclin in vivo and in vitro. Blood 64; 679-682. 1984.
Landry C, Santagata P, Bawab W, F o u r n i d MC, Roques BP, Vmay P: Characterization of neutral endopeptidase 24.1 1 m dog glomenili Biochern J 29 1 ; 773-779, 1993.
Lanese DM, Yuan BH, Falk SA, Conger JD: Effects of atriopeptin III on isolated rat afferent and efferent arterioles. Am J Physiol26 1 ; F 1 102-F 1 109. 199 1 .
Lang RE, Rascher W, Unger T, Ganten D: Reduced content ofvaçopressh m the brab of spontaneously hypertensive as compared to normotensive rats. Neurmci Lett 23; 199-203. 1981.
Lang RE, Tholken H, Gante D: M natriuretic factor: A circulating hormone stimulated by volume loading. Nature 3 14; 265-266, 1985.
Lang T, Degoulet P, Aimé F, Devries C, Jacquhet-Salord M, Fouriaud C: Relationship between alcohol consumption and hypertension prevalence and control in a French population. J Chronic Dis 40; 7 13-720, 1987.
Lang CC, Coutie WJ, Khong TK, Choy AMJ, Struthers AJ3: Dietary sodium loading increases plasma brain natriuretic peptide levels m man. J Hjpertem 9; 779-782, 199 1 .
Lang CC. Choy AJ, Struthers AD: Atrial and bram natriuretic peptides: A dual natriuretic peptide system potentially involved m circulatory homeostasis. C h Sei 83; 5 19-527, 1992.
Lange LG: Mechanisn of htty acid ethyl ester formation and biological significance. Atzrr NY Acad Sci 625; 802-805, 199 1 .
Langer RD, Cnqui MI3, Reed DM: Lipoproteins and blood pressure as biological pathways for effkct of moderate alco ho1 consump tion on coronary heart disease. Circulation 8 5 : 9 1 0- 915, 1992.
Laposata EA, Lange LG: Resence of nonoxidative ethanol metabolisru in human organs commonly damaged by ethanol abuse. Science 23 1 ; 497-499, 1986.
Laposata E q Scherrer DE, Mazow C, Lange LG: Metabolism of ethanol by hurnan brain fatty acid ethyl esters. J Biol Chem 262; 4653-4657, 1987.
Lappe RW, Dinish JL, Bex F, Michalak K, Wendt RL: Effects of atrial natnuretic factor on

drinking responses to central angiotensin LI. Phonnacol Biochem Behav 24; 1 573- 1 5 76. 1986.
Larose L, McNicoll N, Ong H, De Léan A: AUoaenc modulation by ATP of the bovine adrenal nauiuretic factor RI receptor functions. Biochemistry 30; 8990-8995. 199 1.
Lee J, Fong IQ, Marvin RL, Huang BS, Grekin RI: Centrally admHUstered natriuretic factor mcreases rend water extretion. Am J Physiol252, F 10 1 1-F 10 18, 1989.
Leidenheimer NJ, Hams RA: Acute effeas of ethanol on GABA, receptor function: Molecuiar and physiological determinants, In: GABAergic symptzc îran.smissiot~. Biggio G. Concas A, Costa E (eds), New York, Raven Press, p. 269-279, 1992.
Leitman D, Andresen J, Kuno T, Kamisaki Y, Chang J, Murad F: Identification of multiple bmding sites for atrial natriuretic factor by afnnity cross-linking in cuitured endothelia1 cells. J Bi01 Chem 26 1; 1 1650- 1 1655, 1986.
Leianan D, Agnost VL, Tuan JJ, Andresen J, Murad F: Atrial natriuretic factor and sodium nitroprusside mcrease cycEc GMP m cultured rat hmg fibroblasts by activating dflerent f o m of guanylyl cyclase. Bzochem J 244; 69-74, 1987.
Leppaluoto J, Vuolteenaho O, Arjamaa O, Ruskoaho H: Plasma immunoreactRre atrial natriuretic peptide and vasopressin fier ethanol mtake m man. Acta Physiol Scmid 144; 12 1 - 127, 1992.
Lester D, Greenburg LA: inhalation of ethyl alcohol by men. Quart J Slitd Alcohol 1 2: 1 67- 178, 1951.
Lester R, Van Thiel DH: Gonadal function in chronic alcohoüc men. Adv Exp Med Biol85A: 399-414, 1977.
Leslie SW, Barr E, Chandler J, Farrar RP: Inhibition of fast- and slow-phase depolarization- dependent synaptosomal calcium uptake by ethanol. J Pharmacol Exp mer 225 : 57 1-575. 1983.
LeWi ER, Müls S, Weber MA: Central nervous system mediated vasodepressor action of atrial natriuretic factor. Life Sei 44; 16 17- 1624, 1989.
Levm ER: Natiuretic peptide C-receptor: More than a clearance receptor. Am J Physiol264; E4830E489, 1993.
Li GC, Shiu EC, Hahn GM: Shdarities m cellular mactivation by hyperthermia or by ethanol. Rad Res 82; 257-268, 1980.

Li P, S a DD: Decreased rend activity of vasopressb m spontaneousfy hypertensive rats. J ffyperte~zs 1 1 ; 4 1-48, 1993.
Lian C : L'alcoolisme, cause dliypertension artérielle. Bull Acad Nat! Med (Paris) 74; 525- 528, 1915
Lieber CS, De Cadi LM: Ethanol oxidation by hepatic microsomes: adaptive mcrease &er ethanol feedmg. Science 162; 9 17-9 18, 1968.
Lieber CS, De Carli LM: Hepatic microsornai ethanol oxidizing system: iri vitro characteristics and adaptive properties in vivo. J Biol Chem 245; 2505-25 12, 1970.
Lieber CS: Alcohol and the liver: 1994 update. Gmtroenterology 106; 1085- 1 105. 1994.
Lim RT Jr, Gentry RT, Ito D, Yokoyama H, Baraona E, Lieber CS: First-pass metaboiim of ethanol is predominantly gastnc. Alcohol Clin Exp Res 17; 1337- 1344, 1 993.
Lima-Landma. M m Albuquerque EX: Ethanol potentiates and blocks NMDA-activated singlechanne1 currents in rat hippocampal pyramidal ceus. FEBS Let1 247; 6 1-67. 1989.
Lmdholm J, Fabricius-Bjerre N, Bahsen M, Boiesen P, Bangstrup L, Pedersen ML, Hagen C: Pituitaq-testicular fimction in patients with chronic alcoholism. Eur J C h Imest 8; 269- 272, 1978.
Linkola JI Ylikahri R, Fyhrquist F. Wdenius M: Plasma vasopressin in ethanol intoxication and hangover. Acta Physiol Scarid 104; 180- 187, 1978.
Little HJ: The role of neuronal calcium channels m dependence of ethanol and other sedative1 hypnotics. Phantacol fier 50; 347-365, 199 1.
Littleton JM: Aicohol-induced alterations in clacium handling. Biochem Suc Tram 16; 527- 529, 1988.
Lofton CE, Newman WH, C h e MG: Atrial natnuretic peptide regdation of endothelial perrneability is mediated by cGMP. Biochern Biophys Res Commun 172; 793-799. 1990.
Lovinger DM, White G, Weight FF: Ethanol mhibits NMDA-activated ion current in hrppocampal neurons. Scierice 243; 172 1 - 1 724, 1989.
Lovinger DM: Ethanol potentiation of 5-HT3 receptor-mediated ion current in NCB-20 neuroblastoma cell. Nerrrascz Lett 122; 5 7-60, 199 1.
Lowe DG, Camerato TR, Goeddel DV: cDNA sequence of the human atrial natriuretic

peptide clearance receptor. Nucleic Acidr Res 18; 34 12, 1990.
Luk JICH, Wong EFC, Wong NLM: Absence of C-type natriuretic peptide receptors in hamster glomeruli. Nephron 67; 226-230, 1994.
Lundquist F, Tygstmp N, Winkler K, Mellemgaard K, Munck-Petersen S: Ethanol metabolism and production of fiee acetate m the human liver. J CIin lrrvest 4 1; 955-96 1. 1962.
Luthin GR, Tabakoff B: Activation of adenylate cyclase by alcohols requires the nucleotide- bmdgig protem. J Pharmacol Exp Ther 228; 579-587, 1984.
Maack T, Marion DN, Camargo MJ, Klemert HD, Laragh JH, Vaughan ED Jr: Effects of auriculin ( a h 1 natriuretic h o r ) on blood pressure, rend function, and the renin-aldosterone system m dogs. Am J Med 77; 1069- 1075, 1984.
Maack T, Suzuki M, Almeida FA, Nusseflzveig D. Scarborough RM. McEnroe G A Lewicki JA: Physiological role of silent receptors of atrial natriuretic factor. Scierice 238: 675-678. 1987.
Maack T, Okolicany J. Koh GY, Price DA: Functional properties of atrial natriuretic factor receptors. Sem Nephroi 13; 50-60, 1993.
McBride WJ, Murphy JM, Lumeng L, Li TK: Serotonin, dopamine and GABA involvement in alcohol druik8ig of selectively bred rats. A lcohol7: 199-205, 1990.
McCarty R, Phmkett LM: Forebrain binding sites for atrial natriuretic factor: Alterations iin spontaneously hypertensive (SHR) rats. Neurochern Int 9: 177- 183. 1986.
McCreery MJ, Hunt WA: Physico-chernical correlates of alcohol intoxication. Neurophormacol 17; 45 1-46 1, 1978.
McKenzie JC, Tanaka 1, Misono KS, inagami T: Immunocytochemical localization of atnal natriuretic factor in the kidney, adrenal medulla, pituitary, and atrium of rat. J Histocheni Cytochern 33; 828-834, 1985.
McKitrick DJ, Calaresu FR: Cardiovascular reponses to microinjection of ANF into dorsal medulia of rats. Am J Physiol255; RI 82-R187, 1988.
MacMahon SW, Blacket RB, Macdonald GJ, Hali W: Obesity, alcohol consumption and blood pressure m Australian men and women. J Hypertenr 2; 85-9 1, 1 984.
MacMahon S: Alcohol consumption and hypertension. Hypertension 9; 1 1 1- 12 1, 1987.

McQuenn DV, Celaitano DD: Social factors m the etiology of multiple out cornes: The case of blood pressure and alcohol consumption patterns. Soc Sci Med 16; 397-4 18, 1982.
Maheswaran Gill JS, Davies P, Bewers DG: High blood pressure due to alcohol: A rapidly reversible effect . Hypertemion 1 7; 78% 792, 1 99 1.
Maheswaran R, Beevers M, Beevers DG: Effectiveness of advice to reduce alcohol consumption in hypertensive patients. Hyperfertsion 19; 79-84, 1992.
Maines JE, Aldinger EE: Myocardial depression accompanying chronic connimption of alcohol. Am Heurt J 73; 55-63, 1967.
Mains RE, Eipper BA. Ling N: Common precursor to corticotropins and endorphins. Proc Nain AcadSci USA 73; 3014-3018, 1977.
Malhotra H, Dathur D, Mehta SR, Hkandetwal PD: Pressor effects of alcohol in normotensive and hypertensive subjeas. Larzcet 2; 584-586, 1985.
Mahowska B, Pawlak D, Chabielska E, Buczko W: Cardiovascular effects of ethanol in anaesthetized, conscious and pithed rats. Dmg AIcohol Dep 24; 5 1-56, 1989.
Mangiapane ML: SFO: Cardiovascular regulations. hi: Circumventncular organs and body fluids. vol. 1 . Gross ed., CRC, Boca Raton, FI, p. 95- 108, 1987.
Mann SW, Fuiler GC, Rodil SV, Vidins EI: Hepatic prolyl hydroxylase and collagen synthesis in patients with alcoholic liver k a s e . Gur 20; 825-832, 1979.
Mantyh CR, Kruger L, Brecha NC, Mantyh PW: Localization of specific binding sites for atrial natriuretic fàctor m the central nervous system of rat, guinea pig, cat and human. Brairr Res 4 12; 329-342, 1987.
Marie JP, Guillemot FI, Ha# PY: Le degré de granulation des cardiocytes auriculaires. Path Biol24; 549-554, 1976.
Marin-Grez M., Fleming JM, Steinhausen M: Atrial natriuretic peptide causes preglomerular vasodilation and postglomedar vasoconstriction m rat kidney. Nature 342; 473-476. 1986.
Marmot M, B m e r E: Alcohol and cardiovascular disease: The statu of the U shaped c w e . BMJ 303; 565-568, 199 1.
Marmot MG, Elliott P, Shipley MJ, Dyer A& Ueshima H, Beevers DG, Stamler R, Kesteloot H, Rose G, Stamler J: Alcohol and blood pressure: the INTERSALT study. BMJ 308; 1263- 1267, 1994.

Martin ER, Lewicki JA Scarborough RM, Bail- BJ: Expression and regdation of ANP receptor subtypes m rat renal glomeruli and papillae. Am J Physiol 257 ( R e d Flzrzd Electrolyte Physzol26); F649-F657, 1 989.
Martin DR, Pevahouse JB, Trigg DJ, Vesely DL, Buerkert JE: Three peptides fiom the ANP prohomone Nb-terminus are natriuretic ancilor kaliuretic. Am J Physiol25 8; F 140 1 -F 1408, 1990.
Manuno F, Masuda T. Ando K: Resence of the atriai natriuretic peptide in human cerebrosphd fluid. Biochern Biophys Res Comrnwi 143; 8 13-8 18, 1987.
Matsuoka H, Ishii M, Hirata Y, Ataraçhi K, Sugimoto T, Kangawa K, Matsuo H: Evidence for lack of a role of cGMP in effect of alpha-hANP on aldosterone inhibition. Am J Physiol 252; E643-E64?, 1987,
Made S, Chaudburi Thornaides T, Pavin D, McCleery J, Mathias CJ: Effects of oral alcohol on supenor mesenteric artery blood flow in normal man. horizontal and tilted. Clin Sci 84; 4 19-425, 1993.
Mayes DM, Furuyama S, Kem DC, Nugent CA: Radioimmunoassay of plasma aldosterone. J Clin Ertdocritiol Metab 30; 682-685, 1970.
Meade TW. Chakrabarti R Hames AP, North WRS, Stirling Y: Characteristics affecting fibrinolytic activity and plasma fibrinogen concentrations. BMJ 284; 153- 156, 1979.
Mehta P, Mehta J, Lawson D, Patel S: Ethanol stimulates prostacyclin biosynthesis by human neutrophiles and potentiates anti-platelet aggregatory effects of prostacyclin. irhromb Res 48: 653-66 1 , 1987.
MeIlanby E: Alcohol: its absorption into and dissappearance f?om the blood under different conditions. H M Srat Off 48 (Nat Health Ituur. Med Research Corn. Spec Rep Sem 3 1 ): London, 19 19.
Meloche S, Oog H, Cantin M, De Léan A: A5uty cross-linking of atrid natriuretic factor to its receptor in bovine adrenal zona glomerulosa. JBiol Chem 26 1 ; 1525- 1528, 1986.
Melville DI: Alcohol and hypertension. Laticet 2; 286, 198 1.
Mendelsohn FA, Allen AM, Chai SY, Seaon PM, Figdor R: ûverlapping distniutions of receptors for atrial natriuretic peptide and angiotensin II visualized by itz vitro autoradiography Morphological bask of physiological antagonim. Cati J Physiol Pharmacol 65; 1517-1521, 1987.

Mendelson JH, Stein S: S e m cortisol levels m alcoholics and non-alcoholic abjects during experimentally mduced ethanol intoxication. Psychusm Med 28; 6 16-626, 1966.
Mendelson JH, Ogata M, Mello NK: Adrend function and alcoholism: 1. S e m cortisol. Psychosom Med 33; 145-157, 1971.
Messing RO, Carpenter CL, Greenburg DA: Ethanol regulates calcium channels in clona1 neural cens. Proc Mat/ Acud Sci USA 83; 62 13-62 15, 1986.
Meyer KH: Tram F a r a d ~ Soc 33; 1062- 1068, 1937.
Michaelis E Y Muhraney JJ, Freed WJ: Effects of acute and chronic ethanol mtake on synaptosomal glutamate bmdmg activity. Bzochem Pharmacol 27; 1685- 169 1, 1978.
Michaetk EK, Freed WJ, Galton N: Ghitamate receptor changes in brain synaptic membranes from human alcoholics. Neurochem Res 15; 1055- 1063, 1990.
Michel H, Backer HG Kramer HI: Selective downregulation of rat rend clearance receptors for a t rd natriuretic peptide by chronic hi&-saltb intake: Study on isolated membrane using '31-labeUed c-atnal natnuretic peptide(4-23). C h Sci 83; 139- 142. 1992.
Mikhailidis DP, Jeremy JY, Barrada Dandona P: Effect of ethanol on vascdar prostacyclin (prostaglandin 1,) synthesis, platelet aggregation, and platelet thromboxane reIease. Br Med J 287; 1495- 1498, 1983.
Miller NE, Hammett F, Saltissi S: Relation of angiographically defined coronary artery disease to plasna iipoprotein sub6actions and apolipoprotems. BMJ 282; 174 1- 1744, 198 1.
Miller NE, Bolton C Y Hayes TM, Bainton D, Yamell M G , Baker LA. Sweetnam PM: Associations of alcohol consunq>tion with plasma high density lipoprotein cholesterol and its major subfiactions: The Caerphilly and Speedweli collaborative heart disease studies. J Epidemioi Comm Heaith 42; 220-225, 1988.
Milon H, Froment A, Gaspard P. Guidollet J, Ripole JP: Alcohol consumption and blood pressure in a French epidemiological study. Eur Heart J 3; 59-64, 1982.
Minamino N, Aburaya M, Ueda S, Kangawa K, Matsuo H: The presence of bram natriuretic peptide of 1200 dahons in porcine heart. Biochem Biophys Res Commwz 155; 740-746. 1988.
Minamino N, Makmo Y, Tateyama H, Kangawa K, Matsuo H: Characterization of immunoreactive human C-type natriuretüc peptide m brah and heart. Biochem Biophys Res Commun 179; 535-542, 1991.

Misono K, Fuk- H, Grammer RT, Inagami T: Rat atrial natnuretic factor: Complete amho acid sequence and disufide linkage essential for biological activity. Biochem Bioplys Res Commun 1 19; 524-529, 1984.
Mistra RP: Isolation of glomeniü fkom mammalian kidneys by graded sieving. Am J Clin Pathol58; 135- 139, 1972.
Mitchell PI, Morgan UI, Boadle DJ: The role of alcohol in the aetiology of hypertension. Med J Aust 2 ; 198-200, 1980.
Mizefle HL, Gaillard C A Manning RD, Hall JE: Mechanism of decreased cardiac output d h g ANP intiision in conscious anephnc dogs. Am J Physiol262; R120-R125, 1992.
MinistiHna S, Nara Y, Mano M., Sawamura M, Horie Et, Yamori Y: Alcohol consumption as a risk factor for hi& blood pressure fiom the Cardiovascular Diseases and Alimentary Cornparison mdy. JCardio Pharmacol 16 (suppl8); S35-S37, 1990.
Mogelson S, Lange LG: Nonoxidative ethanol metabolism m rabbit myocardium: purification to homogeneity of fatty acyd ethyl ester synthase. Biochemistry 23; 4075-408 1 . 1984.
Mongo KG, Vassort G: Inhibition by alcohols, halothane and chlorofom of the Ca current in single fiog ventricular cels. J Mol Cell Cardiol22; 939-953, 1990.
Moore RD, Pearson TA: Moderate alcohol conçumption and coronary artery disease. Medicine 65; 242-267, 1986.
Moore RD, Levine DM, Southard J, Entwisle G, Shapiro S: Alcohol connimption and blood pressure in the 1982 Maryland Hypertension w e y . Am J Hypertem 3; 1-7. 1990.
Morü N, Nakao Y Sugawara A, Sakamoto M, Suda M, Shimokura M, Kiso Y, Kihara M. Yamori Y, Irnura H: Occurrence of atrial natriuretic polypeptide m brain. Biochem Biophys Res Commun 127; 413-419, 1985.
Morii N, Nakao K, Kihara M, Sugawara A, Sakamoto M, Yamori Y. Lmura H: Decreased content in lefi atrium and increased plasma concentration of atnal natriuretic polypeptide in spontaneously hypertensive rats (SHR) and SHR stroke-proue. Biochem Biophys Res Commun 135; 74-8 1 , 1986a-
Morü N, Nakao K, Itoh H, Sugawara A, Sakamoto M, Yamada T, Shiono S, Kihara M. Mano M, Kihara M, Yamori Y, Imura H: tncreased tissue level of atrial natriuretic polypeptide in the hypothalamus and septum of spontaneously hypertensive rats. J Hper tem 4 (~uppl. 3), S30943 16, 1986b.

Morrow AL, Suzdak PD, Karanian JW, Paul SM: Chronic ethanol administration alters gamma ~ o b u t y r i c acid, pentobdiai and ethanol-mediated "CI- uptake in cerebral cortical synaptoneurosomes. J Pharrnucol Exp mer 246; 15 8- 164, 1988.
Morrow AL, Montpied P, Paul SM: GABA, receptor fhction and expression following chronic ethanol and barbiturate administration. Anrl NY Acad Sci 625 ; 496-507, 199 1 .
Moshage H, Casini A, Lieber CS: Acetaldehyde stimulates coliagen production m cultured rat iiver fat-storing cells but not m hepatocytes. Hepatology 12; 5 1 1-5 18, 1990.
Mukaddam-Daher S. Gutkowska J. Tremblay J, Dam TV, Quillen EW Jr: Regdation of renal aûiai natriuretic peptide receptors m pregnant sheep. Endocriirulody 136; 4565-457 1 , 1995.
Mukhopadhyay AK, Bohnet HG, Leidenberger FA: Testosterone production by mouse Leydig ceüs is stimulatecl in vitro by atrial natriuretic factor. FEBS Lett 202; 1 1 1 - 1 16. 1986.
Mukoyama M, Nakao K, Hosoda K Suga S, Saito Y, Ogawa Y, Shirakami G, Jougasaki M. Obata K, Yasue H, Kambayashi Y, Inouye K [mura H: Bram natriuretic peptide (BNP) as a novel cardiac hormone m humans. J Clin Invest 87; 1402- 14 12, 199 1 .
Miiller A, Sies H: Role of alcohol dehydrogenase a c t ~ t y and of acetaldehyde in ethanol- induced ethane and pentane production by isolated perfised rat b e r . Biochem J 206; 1 5 3- 156, 1982.
Munson PI, Rodbard D: LIGAND: A versatile computerized approach for characterization of ligand-bindmg synems. A m l Biochenz 107; 220-239, 1980.
Murad F: Co-purification of an atrial natriuretic factor receptor and particdate guanylate cyclase from rat h g . J Biol Chem 26 1 ; 58 17-5823, 1986.
Murphy JM, McBride WJ, Lumeng L, Li TK: Content of monoamines in forebraün regions of alcohol-prefehg (P) and -nonprefening (NP) lines of rats. Pharmacol Biochem Be& 26; 389-392, 1987.
Mmay MM: The diuretic action of alcohol and its relation to pituiûin. J Physiol76; 379-386. 1932.
Murray RD, Itoh S, hagarni T, Misono K, Seto S, Scicii AG, Larretero OA: Effects of synthetic atrial naaniretic Etctor m the isolated perfùsed rat kidney. Am J Physiol249; F603- F609. 1985.
Myrhed M: Blood pressure and alcohol connimption m relation to factors associated with ischaemic heart disease. Acta Med Scand 567 (suppl. 1 ) ; 40, 1974.

Nagaoka & Lovenberg W: Regional changes m the activities of aminergic biosynthetic enzymes in the brain of hypertensive rats. Eur J Pharmacol 43; 297-306, 1977.
Nakamura M, Katsuura G, Nakao K, Imura H: Antidipsogenic action of or-human atnal natriuretic polypeptide administred mtracerebroventridariy in rats. Neurscz Len 58: 1-6. 1985.
Nakamura M, Kawata Y, Yoçhida H, Arakawa N. Koeda T, Ichikawa T: Relationship between plasma atrial and bram natriuretic peptide concentration and haemodynamic parameters during percutaneous transvenous mitral vahlotomy m patients with mitral stenosis. Am Heart J 124; 1283- 1288, 1992.
Nakao K, Katsuura G, Morü N: Inhii'bitory effect of centrab admsiistered atrial natriuretic polypeptide on brain dopaminergic system in rats. Eur J Phormacol 1 3 1 ; 1 7 1 - 1 77. 1 986.
Nakao K, Itoh Y Kambayashi Y: Rat brain natriuretic peptide: Isolation fiom rat heart and tissue distribution. Hypertension 1 5 : 774-778, 1 990.
Nakao K, Ogawa Y, Suga S, h a H: Molecuiar bioIogy and biochemistry of the naniuretic peptide system: il. Natriuretic peptide receptors. J Hyperterrs 10; 1 1 1 1- 1 1 14, 1992.
Nakao K, Itoh H, Suga S, Ogawa Y, Lmura H: The natriuretic peptide family. Czrrr Opiti Nephroi Hypertens 2; 45-50. 1993.
Nanji f i Khwaja S, Khethry U, Sadrzadeh SM: Plasma endothelin levels m chroaic ethanol fed rats: Relationship to pathologie h e r mjury. Life Sci 54; 423-428, 1994.
Napier MA, Vanden RL, Albers-Schonberg G, Nutt RF, Brady S, Lyle T, Winquia R Faison EP, Henei Blame EH: Specific membrane receptors for atnal natriuretic factor in renal and vascdar tissues. Proc Nat1 Acad Sci USA 8 1 ; 5946-5 950, 1984.
Naruse M, Takeyama Y, Tanabe A, Hiroshige J, Nanise K, Yoshimoto T, Tanaka M, Katagin T, Demura H: Atrial and brah natriuretic peptides in cardiovascular diseases. Hypertemioti 23 [suppi 4; 123 1-m4, 1994.
Needleman P, Blaine EH, Greenwald JE, Michener ML, Saper CB, Stockmann PT, Tolunay HE: The biochemical pharrnacology of atnal peptides. Anm Rev Pharmacol ToxicoI 29; 23- 54, 1989.
Nemer M, Chamberland M, Sirosis D, Argentin S, Drouin J, Dixon RAF: Gene structure of human cardiac hormone precursor, proatriodilatin. Nature 3 12; 654-656. 1984.
Nemer M, Lavigne IP, Drouh J, Thiiadt G, Gannon M, Antakly T: Expression of atrial

natriuretic factor gene m heart veutricular tissues. Peptides 7; 1147- 1152, 1986.
Ngo L, Wyeth RP, Bksett JK Prohormone atrial natriuretic peptides 1-30,3 1-67 and 99- 126 hcrease m proportion to right ventricular pacmg rate. Am Heart J 1 17: 385-390, 1989.
Nguyen TT, L a m e C, Babinski K, Chrétien M, ûng H., De Léan A: Aldosterone secretion mhibitory Bctor: A novel neuropeptide m bovine chromaffin cells. Erzd~~rzrroIogy 124: 1 59 1 - 1593, 1989a-
Nguyen Tï, Lapire C, Babmski K, Chrétien M. De Léan A, Ong H: Purification and p r h a q structure of pro-aldosterone secretion inhibitory factor fiorn bovine adrenal chromafEn cells. Mol Etzdocrinol3; 1823- 1829, 1989b.
Nhamburo PT, Salafsky BP, Hoffman PL, Tabakoff B: Effects of short-chah alcohols and norepmephrine on brain (Na+, K')ATPase aaMty. Biochem PhUrrmncol3 5; 1987- 1995, 1986.
Nielsen J: Delirium tremens in Copenhagen. Part of a cross-national mvestigation of delirium tremens m the northern countries. Acta Psychiat Scand 4 1 (suppl. 187); 92- 105, 1965.
Niemela O. Klajner F, Orrego H, Vidins E, Bleudis L, Israel Y: Antibodies against acetaldehyde-mo~ed protein epitopes m human alcoholics HeputoIogy 7; 1 2 1 0- 1 2 1 4, 1 9 87.
Nissen R, Gutkowska J, Renaud LP: Release of atnal natriuretic factor fiom intact and hypophysectomized rat hypothalamic explants. J Netrroendocrinol 1 ; 3 09-3 13, 1989.
Nomura F, Lieber CS: Bhdmg of acetaldehyde to rat h e r microsomes: enhancement afier chronic alcohol connimption. Biochern Biophys Res Commun 1 00; 1 3 1 - 1 3 7, 1 98 1.
Nose H, Takamata 4 Mack GW, Kawabata T, Oda Y, Hashimoto S, Hirose M, Chihara E, Morimoto T: Right atnal pressure and ANP release during prolonged exercise in a hot environment. J Appl Physiol76; 1 882- 1 887, 1994.
Nozuki M, Mouri T, Itoi K, Takahashi Y Tatsune K, Saito T, Yoshinaga K: Plasma concentrations of atrial natriuretic peptide m various diseases. Tohoku J Exp Med 148; 439- 447. 1986.
Nuglozeh E, Gauquelin G, Garcia R Tremblay J, Sc& EL: Atrial natriuretic peptide receptors in renal papilla of DOCA-salt hypertensive rats. Am J Physiol259 (Rerral Fhid ElectroIyte Physiol28); F 130-F 13 7, LWO.
Nunez DJR, Dickson MC, Brown MJ: Natriuretic peptide receptor mRNAs in the rat and human heart. J Clin Invest 90; 1966-1971, 1992.

Nussenzveig DR. Lewicki JA, Maack T: Cellular mechanisms of the clearance fiindon of type-C receptors of atrial natriuretic factor. J Biol Chem 265 ; 20952-2095 8, 1990.
Nutt DJ, Glue P, Molyneux S, Clark E: Alpha-2-adrenoceptor actMty m alcohol withdrawal: A pilot study of the effect s of i. v. clonidine m alcoholics and normals. A kohol Clin Erp Res 12; 14-18, 1988.
Nutt DJ, Peters TJ: Alcohol: The dmg. Br Med Bull 50; 5- 17, 1994.
Obana K, Naruse M, Inagami T, Brown AB, Naruse K, Kurimoto F, Sakurai H, Demura H. S b e K Aaial natriuretic fàctor mhibits vasopressin secretion fiom rat posterior pituitary. Biochern Biophys Res Commun 132; 1088-1094, 1985.
OCallaghan CJ, phinq>s PA, Krum H, Howes G: The etfects of short-terni alcohol intake on c h i c and ambulatory blood pressure in normotensive 'social' drinkers. Am J Hypertem 8; 572-577, 1995.
Ogawa Y, Nakao K, Mukoyama M, Shirakami G, koh H, Hosoda K, Saito Y, Arai H. Suga S, Jougasaki M, Yamada T, Kambayashi Y, Inouye K, Irmira H: Rat brain natriuretic peptide: Tissue distribution and molecular form Etsdocri~~ology 126; 2225-2227, 1990.
Ogawa Y. Nakao Y Mukoyama M, Hosoda K, Shirakami G, Arai H, Saito Y, Suga S. Jougasaki M, Imura H: Natriuretic peptides as cardiac hormones m nonnoteosive and spontaneoudy hypertens~e rats: The ventride is the major site of synthesis and secretion of brain natriuretic peptide. Circ Res 69; 49 1-500, 199 1.
Ogawa Y, Nakao K, Nakagawa O, Komatsu Y. Hosoda K, Suga S. Arai H, Nagata K Yoshida N, Imura H: Human C-type natriuretic peptide: Characterization of the gene and peptide. Hypertertriori 19; 762-765. 1992.
Ogawa Y, Itob E-I, Nakao K: Molecular biology and biochemktry of natriuretic peptide family. CiNl Exp Pharmacol Physiol22; 49-53, 1995.
Ogura T, Yamamoto 1, Ogawa N: Development change of kidney receptor for atrial natriuretic factor m spontaneously hyperteosive rats. Hyperterrîiorz 13: 449-455, 1989.
Ohashi M, Rujio N, Nawata H, Matsuo H, Kato K: Human atrial natriuretic polypeptide binding sites in human adrenal membrane fi-actions. Reg Peptides 21; 271-278, 1988.
Ohnishi K, Lieber CS: Reconstitution of the microsoma1 ethanol oxidkhg system: qualitative and quantitative changes of cytochrome P-450 d e r chronic ethanol consumption. J Bioi Chem 252; 7 124-713 1, 1977.

Oikawa S, Imai M, Ueno A. Tanaka S, Noguchi TI Nakazako H: Clonkg and sequmce analysis of cDNA eicoding a precurror for human atrial natriuretic popypeptide. Nature 309: 724-726, 1984.
O h s GM, Spear KL, Süegel Reinhard EJ, Zurcher-Neely A: Atrial natriuretic peptide mactivation by rabbit-kidney bru&-border membranes. Eur J Biochem 170; 43 1-434. 1987.
Olins GM, Patton DR Bovy PR, Nehta PP: A linear analog of atrd naaiuretic peptide (ANP) es guanylate cyclase-coupled ANP recep tors from non-coup led recep tors. J Biol
Chem 263; 10989- 10993, 1988.
Olson RW, Tobm Al: Molecular biology of GABA, receptors. FASEB J 4; 146% 1480. 1 990.
O'Neill PI, Kaufman LN: Effect of indwelling artenal catheters or physical restramt on food consumption and growth patterns of rats: Advantages of noninvasive blood pressure measurement techniques. Lab Animal Sci 40; 64 1-643, 1 990.
Ong H. L a m e C, Nguyen TT, McNicoll N, Seidah N, Chrétien M, De Léan A: Bovine adrenal chrondfin granules are a site of synthesis of atrial natriuretic factor. Biocheni Bzophys Res Commun 147; 957-963, 1987.
Orlando J, Aronow WS, Cassidy J, Prakash R: Effect of ethanol on angha pectoris. A m h i r
Med 84; 652-655, 1976.
Orth DN: Adrenocorticotrophic hormone (ACTH). hi: Methods of hormone radioirnmunoassay, 2nd ed., vol. 1, Academic Press, New-Y ork, p. 245. 1 979.
Oshino N, Oshino R Chance B: The characteristics of the "p eroxidation" reaction of catalase m ethanol oxidation. Biochem J 13 1 ; 55 5-563, 1973.
Pagala M, Ravindran K., Arnaladevi B, Namba T, Grob D: Effect of ethanol on function of the rat heart and skeletal muscles. Alcohol Clin Exp Res 19; 676-684, 1995.
Pakovits M, Bahner U, Geiger H, Fodor M, Lenkei 2, Heidiand A: Atrial natriuretic factor m the subfornical organ and the organum vasdosum laminae terminalis. In: Progress m brain research, vol 9 1, Ed. by Ermisch A, Landgr& R, W e HJ, Elsevier Science publishers. B. V.. p. 25 1-259, 1992.
Pan L, Gutkowska J: 1s clonidine-induced diuresis mediated by atrial natriuretic factor? Endocrirzology 123; 1259- 1263, 1988.
Pandey KN, Kovacs WJ, Inagami T: The inhiibition of progesterone secretion and the regdation of cyclic nucleotides by atrial natriuretic factor m gonadotropin responsnie murine

Leydig tumor cells. Biochem Biophys Res Commun 133; 800-806, 1985.
Pandey KN, inagami T, Misono KS: Atrial natriuretic factor receptor on culwed Leydig tumor celis: Ligand bmdiag and photoafbty labeling. Biochemisby 25; 8467-8472, 1986.
Pandey K., Orgebin-Crist MC: Atrial natriuretic factor m marumahan testis: Immunological detection m spermatozoa. Biochem Biophys Res Commtin 180; 437-444, 199 1.
Pang SC, Sarda Wigle DA Zhou ZI, Jennings DB, Fiynn TG: Plasma clearance and tissue bmdmg of rANP(99- 126) and iso-rANP( 1-45) m the rat. Reg Peptides 33; 1-9, 199 1 .
Parenti G, Giordana B, Hanozet GM: irr vitro effect of ethanol on sodium and glucose transport m rabbit raial bru& border membrane vesicles. Biochim Biophys Acta 1070; 92-98. 1991.
Parkes DG, Co- JP, McDougall JG, Scoggins BA: Long-term hemodynamiic actions of atrial natriuretic factor (99- 126) m conscious sheep. Am J Physiol254; H8 1 1-H8 15. 1988.
Patel VA, PohorecS, LA: Acute and chronic ethanol treatment on D-endorphin and catecholamine levels. Alcohol6; 59-63, 1 989.
Paulin JM, Simpson FO, Waal-Manning HJ: Alcohol consumption and blood pressure in a New Zealand commUnny study. NZ Med J 98; 425-428, 1985.
Pawlak D, Morelowska-Spierzak D, Azzadin A, Wrobel K, Buczko W: Lack of effect of acetaldehyde on the cardiovascular system in rats. A kohol & A Icoholism 28; 5 29- 533. 1993.
Pell S, D'Alonzo CA: The prevalence of chroaic disease among problem drinkers. Arch Emirorr Health 16; 679-684, 1968.
Periti M, Salvaggüo A, Quaglia G, Di Marzio L: Alcohol consumption and blood pressure: an Italian study. Eur J Epidemiol4; 477-48 1 , 1988.
Perreadt T, Gutkowska J: Role of atrial naniuretic factor in lung physiology and pathology. Am JRespir Crzt Cure Med 15 1 ; 226-242, 1995.
Pesonen 4 Leppaluoto J , Ruskoaho H: Mechanisms of opioid-induced atrial natriuretic peptide relase in conscious rats. J Pharmacol Exp 7ïzer 254; 690-695, 1990.
ffeffer JM, Pfeffer Mq Frohlich ED: Validity of an indirect tail-cuffmethod for determinhg systolic arterial pressure m unanesthetized normoensive and sp ontaneously hyp ertensive rat S. J La& Clirz Med 78; 957-962, 197 1.

Pikaar NA Wedel M, Van der Beek E, Van Dokkum W, Kempen HJM, Mufi C, Ockhuizen T, H e m RJJ: m i s of moderate alcohol consumption on platelet aggregation, fïbrioolysis. and blood lipids. Metubolism 36; 538-547, 1987.
Pikkarainen PY Gordon ER Lebsack ME, Lieber CS: Detemhants of plasma kee acetaldehyde level during the oxidation of ethanol: effects of chronic ethanol feeciing. Bzochem Phumucol30; 799-802, 198 1.
Pohorecky LA, Brick I: Phannacology of ethanol. Phannac Ther 36; 33 5-427, 1988.
Ponticelli C, Montaguino G: Direct renal eEects of alcohol. In: Metabolic eflects ofalcohol. Avogaro P, Sirtori CR Tremoli E (eds), Elsevier/North-HoIland biomedical press, p. 265- 375, 1979.
Porter JG, M e n A, Fuller F, Miller 1 4 Gregory LC, Lewicki JA: Isolation and fundonal eqression of the human atrial natriuretic peptide clearance receptor cDNA Biochem Biophys Res Commun 172; 796-803, 1990.
Potter JF, Beevers DG: Pressor effea of alcohol m hypertension. Lancet 1 ; 1 19- 122, 1 984.
Potter .IF, Watson RDS, Skan W, Beevers DG: The pressor and metabolic effects of alcohol in normotensive abjects. Hypertemioti 8; 625-63 1, 1986.
Potvin W, Varma DR. Bronchodilator actnay of atrial natriuretic peptide m guinea pigs. Cari J Physzol Pharmacol 67; 12 13- 12 18. 1989.
Reedy VR, Richardson PI: Ethanol induced cardiovascular disease. Br Med Bull 50: 152- 163, 1994.
Puddey IB, Beilin U, Vandongen Rouse IL, Rogers P: Evidence for a direct effect of alcohol consumption on blood pressure in normotensive men: a randornized controiled trial. Hypertension 7; 707-7 13, 1985.
Riddey IB, Beilin U, Vandongen R: Effect of regular alcohol use on blood pressure control in treated hypertensive subjects: A controlled study. Clin Exp Pharmacol Physiol 13: 3 15- 3 18, 1986.
Quirion R, Dalpé M, Dam TV: Characterization and distribution of receptors for the atrial natriuretic peptides m mnmmalian brain. P m Nat[ Acad Scz USA 83; 174- 1 78, 1 986.
Rabm R, Molinoff PB: Activation of adelynate cyclase by ethanol in mouse striatal tissue. J Pharrn~cof Exp Ther 2 16; 129- 134, 198 1.

Rachamin G, Macdonald 4 Wahid S, Clapp JJ, Khanna M, Israel Y: Modulation of alcohol dehydrogenase and ethanol metabolism by sex hormones in the spontaneously hypertensive rat. Biochem J 186; 483-490, 1980.
Racz K, Kuchel O, Cantin M, De Léan A: Atrial natriuretic factor inhibits the eariy pathway of steroid biosynthesis in bovine adrenal cortex. FEBS Lett 192; 19-22, 1985.
Ramirez G, Saba SR Dietz JR, Vesely DL: irnmunocytochemical localization of proANF 1- 30, pro- 3 1-67, and atrial natriuretic fàctor (ANF) m the kidney. Kidney h t 4 1 ; 334-34 1. 1992.
Randin D, VoIienweider P, Tappy L, léquier E, Nicod P, Scherrer U: Suppression of alcohol- mduced hypertension by dexamethasone. N Engl J Med 332; 1733- 1737, 1995.
Rascher W, Weidmann E, Gross F: VasopreçSn in the plasma of stroke-prone spontaneously hypertensive rats. Clin Sci 6 1; 295-298, 198 1.
Ray C, Camey SL, Gillies AHB: Effect of ethanol on water and chloride transport in the rat papillary collecting duct. Mirzer Electroiyte Metab 1 8; 370-374, 1992.
Razai G, Heaton KW, Bolton CH, Hughes AO: Alcohol conçumption and its relation to cardiovascular risk factors in bntish women. BMJ 304; 80-83, 1992.
Redei E, Branch BJ, Taylor AN: Direct effect of ethanol on adrenocorticotropm (ACTH) release in vitro. J Pharmacol Exp ther 237: 59-64, 1986.
Redman S, Sanson-Fisher RW, Wilkinson C , Fahey PP, Giibard RW: Agreement between two measures of alcohol consumption. JStudies Alcohol48; 104- 108, 1987.
Redmond GP: Effect of ethanol on spontaneous and stimulated growth hormone secretion. Prog Biochern Pharmacol 18; 58-74, 198 1.
Reed D, McGee D, Katnihiko Y: Biological and social correlates of blood pressure among Japanese men m Hawaii. Hypertension 4; 406-4 14, 1982.
Regan TJ: Alcohol and the cardiovascular system JAMA 264; 377-38 1, 1990.
Renaud S, De Lorgeril M: Wme, alcohoi, platelets and the French paradox for coronary heart disease. Lancet 339; 1523- 1526, 1992.
Richards AM, Nicholls MG, Ikrarn Fi, Webster MW, Yandle TG, Espiner EA: Rend haemodynamic, and hormonal effects of human alpha atrial natriuretic peptide in healthy volunteers. Lancer 1; 545-549, 1985.

Richards AM, Tondo G, Cleland JG, Leckie BI, McIntyre GD, Ingram M, Dargie HJ, Ball SG, Robertson JI: Plasna atrial natriuretic peptide: Responses to modest and severe sodium restriction. J Kperterts 4; S559-S563, 1986.
Richards AM, Crozier IG, Espiner EA, YandIe TG, Nicholk MG: Plasma brain natriuretic peptide and endopeptidase 24.11 iohibition m hypertension. Hjpertensiort 22; 23 1-236, 1993.
Ricksten SE, Yao T, Ljung B, Thoren P: Distensibüity of lefi atrium in nonnotensive and spontaneously hypertensive rats. Acta Physiol Scand 1 1 0; 4 13-4 1 8, 1 980.
Ridker PM, Vaughan DE, Stampfer MJ, Glynn RI, Hennekens CH: Association of moderate alcohol connimption and plasma concentration of endogenous tissue-type plasminogen activator. J M 272; 929-933, 1994.
Riff DP, Jain AC, Doyle JT: Acute hemodynamic effects of ethanol on normal human voiunteers. Am Heart J 78; 592-597, 1969.
Rigter H, Rijk H, Crabbe JC: Tolerance to ethanol and severity of withdrawal m mice are enhanced by a vasopressin fragment. E w J Pharmacol 64; 53-68, 1980.
Ritter D, Chao J, Needleman P, Tetens E, Greenwald JE: Localizatüon, synthetic regulation. and biology of renal atnopeptin-like prohormone. Am J Physzol263; F503-F509. 1992.
Rivier C, Bruhn T, Vale W: Effect of ethanol on the hypothalamic-pituitary-adrenal axis in rat: Role of corticotropin-releasing fàctor (CRF). JPharnwcol Exp mer 229; 127- 13 1. 1984.
Rodrigo R, Vergara L, Oberhauser E: Effect of chronic ethanol consumption on postnatal development of rend (Na+K)-ATPase in the rat. Cell Biochern Furzcci 9; 2 1 5-222. 199 1.
Rome RP, Gentry T, Lim RT Jr, Baraona E, Lieber CS: Effect of concentration of ingested ethanol on blood alcohol levels. A kohol Clin Exp Res 15; 734-73 8, 199 1.
Rosenberg J, Pines M, HuM.itz S: Inhibition of aldosterone secretion by atnal natriuretic peptide in chicken adrenocortical ceils. Biochim Biophys Acta 10 14: 189- 194. 1989.
Rossouw JE, Lai-Tung MT, Jooste PL, Weight MI, Benade AJ: Alcohol intake in relation to lipids lipoproteins and blood pressure. Smth A . Med J 82; 246-250, 1992.
Rothman A, Proverbio T, Femandez E, Proverbio F: Effect of ethanol on the Na' and the Na', KI-ATPase activities of basolateral plasma membranes of kidney proromal tubular cek. Biochem Phumacol43; 2034-2036, 1992.
Roy RN, Ftym TG: Organization of the gene for iso-rANP, a rat B-type natriuretic peptide.

Biochem Biophys Res Commu~z 17 1 ; 4 16-423, 1990.
Roy RN, Pang SC, Fiynn TG: Expression of natriuretic peptide genes m cardiac tissues of hypertensive rats. M d Cell Endmrinol86; RI-R8, 1992.
Rozanski DJ, Delaville FJ, Thomas AP: Depression of contraction and the calcium transient in single cardiomyocytes with acute ethanol exposure. Bzophys J 6 1 ; A 160, 1992.
Rubin E, Lieber CS, Altman K, Gordon GG, Southren AL: Rolonged ethanol connimption mcreases testosterone metabohsm m the liver. Science 19 1 ; 563-564, 1976.
Rubm E: Cardiovascular effects of alcohol. Pharmacol Biochem B e h 13 (suppl. 1 ); 37-40. 1980.
Rubm E, Urbano-Marquez A: Alcoholic cardiomyopathy. Alcohol C h Exp Res 18; 1 1 1- 1 14. 1994.
Rubin R Rand ML: Alcohol and platelet functioo. A kohol C h Exp Res 1 8 ; 1 0 5- 1 1 0, 1 994.
Ruskoaho H, Leppàluoto J: Immunoreactive atrial natriuretic peptide m ventricles, atria. hypothalamus, and plasma of genetically hypertensive rats. Circ Res 62; 3 84-394, 1988.
Ruskoaho H: Atrial natriuretic peptide synthesis, release, and metabolism Pharmacol Rev 44; 479-602, 1992.
Rutherford RA, Wharton DJ, Gordon L, Moscoso G, Yacoub MH, Polak JM: Encocardial localization and characterization of natriuretic peptide bbding sites m human fetal and adult heart. Eur J Pharmacol 2 12; 1-7, 1992.
Saavedra JM: Atrial natriuretic peptide (6-33) bmdsig sites: Decreased number and a&ty m the subfomical organ of spontaneously hypertensive rats. J Hypertem 4; S3 1 3 4 3 16. 1986.
Saavedra IM, Phinkett LM, Correa FM4 Iisrael A, Kurihara M, Shigematsu K: Quantitative autoradiography of angiotensin and atrial natriuretic factor binding sites in brain nuclei of spontaneously hypertensive rats. In: Brain peptides and catecholarnines in cardiovasular regdation, Ed. by Buckley P, Ferrario CM, Raven Press, New-York, p. 245-256, 1987.
Saba SR, Rarnirez G, Vesely DL: Immunocytochemical localization of proANF 1-30. proANF 3 1-67, atrial natriuretic factor (ANF) and urodilatiin m the human kidney. Ani J Nephrd 13; 85-93, 1993.
Sachinidis A, Mengden T, Locher R, Bnuuier C, Vetter W: Novel cellular a c t ~ t i e s for low density lipoprotein m vascular smooth muscle cells. Hypertension 1 5 ; 704-7 1 1, 1 990.

Sagnella GA, Markandu ND, Shore AC, MacGregor GA: Raised circulahg lwels of atrial natriuretic peptides m essential hypertension. Lancet 1 ; 179- 1 8 1 , 1986.
Saheki T, Minmo T, Iwata T, Saito Y, Nagasawa T, Mizuno K.: structure of the bovine atrial natriuretic peptide receptor (type C ) gene. J Bioi Chent 266; 1 1 122- 1 1 125, 1 99 1 .
Saito H, Inui Y Matsukawa Y, Okano T , Maegawa Y Nakao Y Morii N, h a E3, Makino S, Hori R: Specifïc bmding of atrial natriuretic polypeptide to rend basolateral membranes in sp ontaneously hypertensive rats ( SHR) and stro ke-prone SHR Biiochern Biophys Res Commztrl 1 3 7 1079- 1085. 1986.
Saito Y, Nakao Y Itoh H, Yamada T, Mukoyarna M, Arai A, Shirakami K, Suga S. Minamino N, Kangawa Y Matsuo H, Imura H: Brain natxhetic peptide is a novel cardiac hormone. Biochem Biophys Res Contrnurr 158; 360-368, 1989.
Sakamoto M, Nakao K, Morii N, Sugawara A, Yamada T, Itoh H, Shiono S, Saito Y, h a H: The lung as a possible target organ for atrial natriuretic polypeptide secreted fiom the heart. Biochem Biophys Res Comrnur~ 135; 5 15-520, 1986.
Salaspuro MP, Shaw S, Jayatilleke E: Attenuation of the ethanol induced hepatic redox change d e r chronic alcohol consumption in baboons: metabolic consequences in vivo and iri vitro. Hepatology 1 ; 33-38, 1981.
Salazar FI, Fiksen-Olsen MJ, Opgenorth TJ, Granger JP Jr, Bumet C , Romero JC: Renal effects of ANP without changes m glornerular filtration rate and blood pressure. Am J Physiol 25 1 ; F532-F536, 1986.
S h e n JT, Tuomilehto J, Tanskanen A: Relation ofblood pressure to reported htake of salt. saturated fats, and alcohol in a healthy middle-aged population. J Epidemioi Commzitz@ Health 37; 32-37, 1983.
Samson WK: Dehydration-induced alterations in rat brain vasopressin and atrial natriuretic factor immunoreactivity. Endocrirzology 1 17; 1 279- 128 1 , 1985.
Samson WK, Aguila MC, Martinovic J, AntunesRodrigues J , Norris M: Hypothalamic action of atrial natnuretic factor to bhi'bit vasopressin secretion. Peptides 8; 449-454, 1987.
Samson WK, Bianchi R: Further aidence for a hypothalanric site of action of atrial natriuretic factor: Inhibition of prolactin secretion in the conscious rat. Can J Physiol Pharmmol66; 30 1-305, 1988.
Samson A g d a MC, Bianchi R: Atrial natriuretic factor inhibits luteinieng hormone secretion m the rat: Evidence for a hypothalamic site of action. Endocrinology 122; 1573-

Samson WK, Huang FLS, Fulton RJ: C-type natriuretic peptide mediates the hypothalamic actions of the natriuretic peptide to inhiid h d d g hormone secretion. Endixrinofogy 1 3 2; 504-509, 1993.
Sanderson JE, Jones JV, Graham DI: Effect of alcohol ingestion on the heart and blood pressure of spontaneousiy hypertensive rats. Clin Erp Hyperem [A] 5; 673-689. 1983.
Sargent WQ, Simpson Beard JD: The effect of acute and cironic alcohol administration on rend hemodynamics and monovalent ion excretion. J Pharmacol Erp mer 1 88: 46 1-47 1. 1974.
Saunders JB, Beevers DG, Paton A: Alcohol-induced hypertension. Laticet 2; 65 3-656, 1 98 1.
Savdie E, Grodght GM, Adena MA Relation of alcohol and cigarette conçumption to blood pressure and semm creatinme levels. J Chronic Dis 3 7; 6 17-623, 1984.
Savolainen VT, Lieao Y Mimikk6 A, Penttilà A, Karhunen PJ: Alcohol consumption and alcoholic h e r disease: evidence of a threshold Ievel of effects of ethanol. Alcohol Chi Erp Res 17; 1 1 12-1 117, 1993.
Schenk DB, Phelps MN, Porter JG, Scarborough RM, McEnroe GA, Lewicki JA: Identification of the receptor for atrial natriuretic factor on cultured vascular cells. J Biol Chem 260; 14887- 14890, 1985.
Schiebinger RI, Linden J: Effect of atrial contraction frequency on atnal natriuretic peptide secretion. Am J Physiol25 1 ; H 1095-H 1099, 1986.
Schiebinger RJ, Greenhg KM: Interaction between aretch and hormonaiiy stimulated atrial natriuretic peptide secretion. Am J Physiol262; H78-H83, 1992.
Sc& EL, Chartier L, Thiiauit G, St-Louis J, Cantm M, Genest I: Vascular and adrenal receptors for atrial natriuretic factor in the rat. Cire Res 56; 80 1-807, 1985.
Schifnm EL, Carrier F, Thibault G, Deslongchamps M: Solubilization and molecular characterization of the atrial natriuretic peptide (ANP) receptor m human platelet: Cornparison with ANP receptors m rat tissues. J Ciin Endocrino/ Metab 72; 484-49 1, 199 1.
Schuckit MA, Gold EO, Risch SC: Plasma cortisol levels following ethanol m sons of alcoholic and controls. Arch Gen Psychiairy 44; 942-945, 1987.
Schuckit MA, Risch SC, Goid EO: AIcohol consumption, ACTH level and f d y history of

alcoholism Am J Psychiotry 145; 139 1- 1395, 1988.
Schulz HD, Gardner DG, Deschepper CF, Coleridge HM, Coleridge JC: Vagal C-fiber blockade abolishes sympathetic inhibition by atrial natnuretic factor. Am J Physzol255; R6- R13, 1988.
Schulz S, Smgh S, Benet Smgh G, Tubb DJ, ,Chin H, Garbers DL: The primary structure of plasna membrane guanyhte cyclase demonstrates diversity withm this new receptor famiIy. Cell58; 1 155- 1162, 1989.
Schulz-Knappe P. Forssnann K, Herbst F, Hock D, Pipkom R, Forssnann WG: Isolation and structural analysis of 'urodilatin', a new peptide of the cardiodilatm (ANP) f d y , extracted fiom human urine. Kliri Wocherischr 66; 752-759, 1988.
Seeman P: The membrane actions of anesthetics and tranquilizers. Pharmacoi Rev 24: 583- 655, 1972.
Seeman P: The membrane expansion theory of anesthetics: Direct evidence using ethano1 and a hi&-precision density meters. Experieritia 30; 759-760, 1974.
Sei C 4 Hand GL, Murray SF. Glembotski CC: The cosecretional maturation of atrial natriuretic factor by primary atnal myocytes. Mol Etrdocrinol6; 309-3 19, 1992.
Seihamer JJ, M e n 4 Miller J& Lundquiist P. Scarborough RM, Lewicki JA: Human and canine gene homologs of porcine bram natriuretic peptide. Biochern Bzophys Res Comnzw~ 165; 650-658, 1989.
Selzer ML: The Michigan Alcoholism Screening Test: The quest for a new diagnostic instrument. An2 JPsychiatry 127; 165301658, 1971.
Seppa Y Laippala P, Sillanaukee P: Drinking pattern and blood pressure. Am J Hyperterzs 7; 249-254, 1994.
Seymour AA, Norman J& Asaad MM, Fenneil SA, Abboa-Offei B. Little DK, Kratunis VJ. Delaney NG, Hunt JT, Di Donato G: Possiile regulation of atrial natriuretic factor by neural endopeptidase 24.1 1 and clearance receptors. J Phannacol Exp f i e r 256; 1002- 1009. 199 1.
Sexton PM, Zhuo J, Mendelsohn FAO: Localization and regulation of renal receptors for angiotensin ïiI and atrial natriuretic peptide. Tohoku J E q Med 166; 4 1-56, 1992.
Shaper AG, Wmamethee G, Whincup P: Alcohol and blood pressure in rniddle-age British men. J Hum Hyperterrr 2; 7 1-78, 1988.

Shaper AG: AIcohol and morta&c A review of prospective shidies. Br J Addict 85 ; 83 7-847. 1990.
Sharkey KA, Gall DG, MacNaughton WK: Distribution and fùnction of brain natrüuretic peptide in the stomach and small mtestine of the rat. Regul Pept 34; 6 1-70, 199 1.
Sheehy TW: Alcohol and the heart: how it helps, how it harms. Part Med 9 1 ; 27 1-277. 1992.
Shehon J, HoIlister LE, Gocka ET: The drinküng behavior i n t e ~ e w (an attempt to quante alcoholic impairment). Dis Nerv Syst 30; 164-467, 1969.
Shiiata K, Sakiunjra M., Furukawa T: Antagonism of central pressor response to angiotensin II by a-human atrial natriuretic polypeptide at the preoptic area and posterior hypothalamus in rats. Neuropharmacol32; 175- 184, 1 992.
Shimiai T, Katsuura G, Nakamura M, Nakao K, Morii N, Itoh Y, Shiono S, imura H: Effect of mtracerebroventncular atnal natriuretic polypeptide on blood pressure and urine production in rats. Lrfe Sei 39; 1263- 1279, 1986.
Shiono S, Nakao K, Morii N, Yamada T, Itoh H, Sakamoto M, Sugawara A, Saito Y. Katsuuni G, irnura H: Nature of atrial natriuretic polypeptide in rat brain. Blochem Biophys Res Comrnurz 135; 728-736, 1986.
Shoji M, Kimura T, Matsui K, Ota K, Iitake K, inoueant MM, Yasujima M, Abe K Yoshmaga K: Effects of centrally administered atrial natriuretic peptide on renal fünctions. Acta Endocrir~ol 1 15; 433-440, 1987.
Sirnonnet G, AUard M, Legendre P, Gabrion J, Vincent ID: Characteristics and specific localization of receptors for atrial naûiuretic peptides in non-neuronal cens in cultured mouse spmal cord cells. Neuroscietice 29; 189- 199, 1989.
Singer DU, Shore AC, Markandu ND, Buckley MG, Sagneila GA. MacGregor GA: Dissociation between plasma atrial natriuretic peptide and urhary sodium excretion following intravenous saline mfusion m normal abjects. Chi Sci 72 (suppl. 16); 43P-49P, 1987.
Smgh S, Lowe D, Thorpe D, Rodriguez H, Kuang W, Dmgoa L, Chmkers M, Goeddel D. Garbers D: Membrane guanylate cyclase is a cell d a c e receptor with homology to protein kinases. Nature 334; 708-7 12, 1988.
Smghd PC, DeCandido S, Satriano J 4 Schlondorff D, Hayes RM: Atriai natriuretic peptide and nitroprusside cause relaxation of cultured rat mesangial cells. Am J Physiol257; C86- C93, 1989.

Skowsky MG, Rosenbloom A A Fisher DA: Radioimmunoassay measurement of ar@e vasopreçssi m senun: Development and application. J Clin Endocrino2 Metab 38; 278-287. 1974.
Smah htl, Hopkinson D& Hanis H: Alcohol dehydrogenase isozymes in adult human stomch and liver: Evidence for activity of the ADH 3 locus. A m Hum Genetics 3 5 ; 243-253, 1972.
Stnyth EM, Keenan AK. The vascular ANP-C receptor: Role in atnal peptide signalling. Cefl Sigrzc~I 6; 125- 133, 1994.
Solomon LR, Atherton JC, Bobinski H, Green R: Effect of posture on plasma immunoreactive atrial natriuretic peptide concentrations m man. Clin Sei 7 1 : 299-3 05. 1986.
Solomon L k Evidence for the grneration of transaminase W i t o r ( s ) d u h g ethanol metabolism by rat liver bomogenates: a potential mechanism for alcohol toxicity. azochern MedMetabolBiol 38; 9-18, 1987.
Sorniyo AP, Somlyo AV: Smooth muscle: Excitation-contraction couphg, contractile regdation, and the cross-bridge cycle. Alcohol C'lin Exp Res 18; 13 8- 143, 1 994.
Sonnenberg H, Honrath U, Chong CK, Wilson DR: Atrial natriuretic factor inhibits sodium transpon in meddary coilecting duct. Am J PhvsioI 250; F963-F966, 1986
Spanos HG, DiNicolantonio R, Morgan TO: The use of non-invasive blood pressure measurements to measure pressor responses in rats during air stress. Clin E ~ D Phcrrmacol Physiol 18; 761-765, 199 1.
Spmow MG, Roggendorf H, Vogel WH: Effect of ethaool on heart rate and blood pressure in nonstressed and stressed rats. Lrfe Sci 40; 25 5 1-25 59, 1987.
Springall DR Bhatnagar M, Wharton J, Hamid Q, GuIbenkian S, Hedges M, Meleagros L, Bloom SR, Polak JM: expression of the atnal natriuretic peptide gene in the cardiac muscle of rat extrapulmonary and mtrapulmonary veins. Thorm 43; 44-52, 1988.
Stampfer MJ, Sacks FM, Salvini S. Willett WC. Heunekens CH: A prospective study of cholesterol apoliproteins, and the risk of myocardial mfarction. N Engf J Med 325 ; 373-3 8 1. 1991.
Standaert DG, Needleman P, Saper CB: ûrganization of atriopeptin-like immunoreactive neurons in the central nervous syaem of the rat. J Comp Neurol253; 3 15-34 1, 1986.
Standaert DG, Cechetto DF, Needleman P, Saper CB: [nhiition of the firing of vasopressin neurons by atriopepti.. Nature 329: 15 1- 153, 1987.

Starling EH: The fhiuids ofthe body. In: n e Herter Lectures. WT Keener and Co., Chicago, p. 106, 1909.
Steardo L, Nathanson JA: Brain barrier tissues: End organs for atriopeptins. Scierice 235: 470-473, 1987.
Steckehgs U, Lebrun C, Qadri F, Vehmar 4 Unger T: Role of brain angiotensin m cardiovascular regulation. J Curdiovasc Pharmacol 1 9 ; S72- S79, 1 992.
Steinhelper ME, Cochrane KL, Field U: Hypotension m transgenic mice expressing atnal natriuretic factor fusion genes. Hypertetrrion 16; 30 1-307, 1990.
Steinhelper ME: Structure, expression, and genornic mappmg of the mouse natriuretic peptide type-B gene. Circ Res 72; 984-992, 1993.
Stençtrom S, Enloe L, ff&g M, Richelson E: Acute effects of etbanol and other short- chah alcohols on the guanylate cyclase system of murine neuroblastoma cells (clone N I E- 1 15). J Pharmacol Exp ïher 236; 458-463, 1986.
Stephenson SL, Kenny AT: The hydroiysis of or-human ahid natriuretic peptide by pig kidney microviIlar membranes is initiated by endopepidase-24.1 1. Biochem J 243; 1 83- 1 87. 1987.
Stevens VJ, Fanti WJ. Newman CB, Sims RV, Cerami 4 Peterson CM: Acetaldehyde adduas with hemoglobin. J C h Itzvest 67; 36 1-369, 198 1.
Stmgo AJ, Clavell AL, Heublem DM, Wei CM, Pittekow MR Bumett JC: Presence of C- type natriuretic peptide m cultured human endothelial cells and plasma. Am J Physiol263: Hl3 18-1321, 1992.
Stott DJ, Bal1 SG, Inglis GC, Davies DL, Fraser R, Murray GD, M c h e s GT: Effects of a single moderate dose of alcohol on blood pressure, heart rate and associate metabolic and endocrine changes. Clin Sci 73; 4 1 1-4 16, 1987.
Strickland JA, Wooles WR Blood ethanol level and physiologie meawements during ethanol-induced hypertension. Alcohol6; 109- 1 14, 1989.
Strogatz DS, James SA, Haines PS, Elmer PJ, Gerber AM, Browning SR Ammemian AS, Keenan NL: Alcohol consumption and blood pressure in black adults: the Pitt County midy. Am JEpidemiol 133; 442-450, 1991.
Subramanian MG, Savoy-Moore RT, Bergeski BA, Kruger ML, Abel EL: Acute ethanol mtùSon does not alter plasna gonadotropms or prolacth m ovariectomized rats. Alcohol Clirt

Eup Res 14; 19 1-194, 1990.
Sudoh T, Kangawa K, Minamino N, Matsuo H: A new natriuretic peptide m porche brain, Nature 332; 78-81, 1988.
Sudoh T, Maekawa K, Kojima M, Mmamino N, Kangawa K, Matsuo H: Clonhg and sequence analysis of cDNA encodmg a precursor for human brain natriuretic peptide. Biochem Biophys Res Commun 159; 1427- 1434, 1989.
Sudoh T, f i a m i n o N, Kangawa K, Matsuo H: C-type natriuretic peptide (CNP): A new member of natriuretic peptide fm identified m porcine brain. Biochem Biophys Res Commun 168; 863-870, 1990.
Suga S, Nakao K, Hosoda K, Mukoyama M, Ogawa Y, Shirakami G, Arai H, Saito Y, Kambayashi Y, Inouye Y Lmura H: Recep tor selectivity of natriuretic peptide fa&, atrial natrRiretic peptide, brah natriuretic peptide, and C-type nahuretic peptide. Endocrinology 130; 229-239, 1992a.
Suga S, Nakao K, Itoh H, Komatsu Y, Ogawa Y, Hama N, Imura H: Endothelial production of C-type natriuretic peptide and its marked augmentation by transforming growth factor-i3: Possible existence of "vascular natriuretic peptide system". J Clin lrwest 90; 1 145- 1 149. 1992b.
Suga S, Itoh El, Komatsu Y, Ogawa Y, Hama N, Yoshimasa T, Nakao K: Cytokine-mduced C-type natriuretic peptide (CNP) secretion fiom vascular endothelial ceils: Evidence for CNP as a novel autocrine/paracrine regulator from endothelial cells. Erzdmrinology 133 ; 303 8- 3041, 1993.
Sumners C, Tang W: Atrial natriuretic peptide recqtor subtypes m rat neuronal and astrocyte glial cultures. Am J Physiol262; C 1 134-C 1 143, 1992.
Sun AY: Alcohol-membrane mteraction in the brain. A m NY Acad Sci 273; 295-3 15. 1976.
Sun AY: Biochemical and biophysical approaches in the çhidy of ethanol-membrane interactions. In Biochemistry and pharmacology of ethanol, Majchrowicz E, Noble EP (Eds), New York/London, Plenum Press, p. 8 1-97, 1979.
Suzdak PD, Schwartz RD, Skohiick P, Paul SM: Ethanol stimulates y-aminobutyruc acid receptor-mediated chloride transport m rat brain synaptoneurosomes. Proc N d Acad Sci USA 83; 4071-4075, 1986.
SunilO E, Hirata Y, Hayakawa Y Omata M, Kojima M, Kangawa K, Minamho N, Matsuo H: Evidence for C-type natriuretic peptide production in the rat kidney. Biochem Biophys Res

Swann AC: Ethanol and (Na+, R)-ATPase: Aheration of Na+-K' selectMty. Alcohol Clin Ecp Res 14; 922-930, 1990.
Swithers SE, Stewart RE, McCarty R: Bmding sites for atnal natriuretic factor (ANF) in kidneys and adrend glands of spontaneously hypertensive (SHR) rats. Lzjie Sci 40; 16 73- 168 1. 1987.
Szabo J, Nosztray K, Tabaks 1, Szegi J : Thyroxine-mduced cardiomegaly: assessrnent of nucleic acid, protein content and myosin ATPase of rat hem. Act Phys Acad Sci Hurzg 46: 28 1-288, 1979.
Sze PY. Yanai J , Ginsburg BE: Adrenal glucocorticoids as a required factor in the development of ethanol withdrawal seizures m mice. Brairz Res 80; 155, 1974.
Sze PY: The permissive role of ghicocorticoids m the development of ethanol dependence and tolerance. Drug Alcohol Deperui 2; 3 8 1-396, 1977.
Tabakoff B: Neurotrausmitter function and alcoholism AIcohol Clhl Erp Res 3; 35 1-352. 1979.
Tabakoff B, Yanai I: Cortexolone antagonizes development of alcohol tolerance in rnice. Psychopharntacol64; 123- 124, 1979.
Tabrizchi R, Pang CCY: Innuence of mtravenous infusion of ethanol on regional blood flow in conscious rats. J Pham Pharmacol 45; 15 1- 153, 1992.
Taivainen H, Laitinen K, Tahtela R, Kiianmaa K, Viilimaki MJ: Role of plasma vasopressin m changes of water balance accompanying acute alcohol intoxication. Alcohol Clhi Exp Res 19; 759-762, 1995.
Takahashi T, Lasker JM, Rosman AS, Lieber CS: Induction of P45O2El in human h e r by ethanoi is due to a correspondhg increase in encodmg mRNA. Hepatology 17: 236-245, 1993.
Takayanaki Inagami T, Snajdar RM, h d a T, Tamura M, Misono KS: Two distinct forms of receptors for atrial natriuretic factor in bovine adrenocortical cells. J Bi01 Chem 262; 12104-121 13, 1987.
Takezawa K, Cowley AW Jr, Skehon M, Roman RI: Atriopeptin DI alters rend medullary hemodyaamics and the pressure-diuresis response m rats. Am J Physiol252; F992-F1002. 1987.

Talesnik J, Belo S, Israel Y: Enhancement of noradrenaline mduced metabolic coronary dilation by ethanol. Eur J Phannacol 6 1 : 279-286, 1980.
Tailerico-Mehyk T, Yip CC, Watt VM: Widespread colocalization of messenger RNAs encoding the guanylate cyclase-coupled natriuretic peptide receptors m rat tissues. Biochern Bioplys Res Commun 189; 6 10-6 16, 1992.
Tanaka I, Misono KS, Inagami T: Atrial natmiretic factor m rat hypothalamus, atna and plasma: det ermination by specific radioimmunoassay. Biochem Biophys Res Commun 1 24: 663-668, 1984.
Taraschi TF, Rubm E: Biology of disease: Effects of ethanol on the chemical and structural properties of biological membranes. Lab lnvest 5 2; 120- 13 1, 1985.
Tarjan E, Denton DA, Weisinger RS: Atrial natriuretic peptide &bits water and sodium intake in rabbits. Reg Peptides 23; 63-75, 1988.
Tateyama H, Hmo J, Miinamino N, Kangawa K, Ogihara T, Matsuo H: Characterization of immunoreactive brain natriuretic peptide m human cardiac atrium Biochem Biophys Res Cornmzirz 166; 1080- 1087, 1990.
Tateyama H, Hmo J, Minamino N. Kangawa K, Minamino T. Sakai K: Concentrations and molecuiar f o m of human brain natriuretic peptide m piasna. Biochem Biophys Res C m w l 185; 760-767, 1992.
Tawaragi Y, Fuchimura K, Nakazato H, Tanaka S, Minamino N. Kangawa K, Matsuo H: Gene and precursor structure of porcine C-type natriuretic peptide. Biochem Biophys Res Cornmiin 172; 627-632, 1990.
Tawaragi Y, Fuchimura K, Tanaka S, Minamino N, Kangawa K, Matsuo H: Gene and precursor structures of human C-type natriuretic peptide. Bzochem Biophys Res Contmii~l 175; 645-651, 1991.
Terada Y, Tomita K, Nonoguchi H., Yang T, Manuno F: PCR localization of C-type natriuretic peptide and B-type receptor ruRNAs m rat nephron segments. Am J Physiol267 ( R e d Fluid Electrolyte Physiol3 6 ) ; F2 1 5-F222, 1 994.
Theiss G, John A, Morich F, Neuser D, Schroder W, Stasch JP, Wohlfeid S: Alpha-h-ANP is the only form of circulating ANP in humans. FEBS Lett 2 1 8 : 1 5 9- 1 62, 1 987.
Thibauh G, Garcia R Gutkowska J, Lazure C, Seidah NG, Chrétien M: Identification of the released form of atrial natriuretic factor by the pefised rat heart. Proc Soc Exp Bi01 Med 182; 137-141, 1986.

Thïibault G, Garcia Gutkowska J, Bilodeau J. Lazure C, Seidah NG: The propeptide Am I- Tyr 126 is the storage form of rat atnal natriuretic factor. Bzochem J 24 l ; 265-272, 1987.
Thibauh G, Charbonneau C, Bilodeau J, SchBih EL, Garcia R: Rat brain natriuretic peptide is locabzed m atrial granules and released mto the circulation. Am J Physiol263 (Regdatory Integrutive C m p PhysioZ32); R301-R309, 1992.
Thomas AP, Sass EJ, Tun-Kirchmann TT, Rubm E: Ethanol inhibits electricdy-induced calcium ransients m isolated rat cardiac myocytes. J Mol CelZ Cardiol2 1 ; 555-565, 1989.
Thomas AP, Rozaoski DJ, Renard DC, Rubm E: Effects of ethanol on the contractile b c t i o n of the heart : a review. A kohol Clin Exp Res 1 8; 1 2 1 - 13 1, 1 994.
Thompson KE, Adams MA: DifEierential effect of short-term ethanol on cardiac and vascular growth responses. J Hypertem 1 2; 409-4 1 8, 1 994.
Tikkanen 1, Fyhrquist F, Metsarinne K, Leidenious R: Plasma atrial natriuretic peptide in cardiac disease and during infusion in healthy volunteers. &et 2; 66-69, 1985.
Toda N, Konishi M M j W M, Komura S: The effects of ethanol and acetaldehyde on dog arterial smooth muscle. Q J Shrd AIcohol44; 1- 15, 1983.
Togashi K Kameya T, Kurosawa T. Hasegawa N, Kawkami M: Concentrations and molecula. forms of C-type natriuretic peptide m brain and cerebrospinal fluid. Clin Chenr 38: 2 136-2 139, 1992.
Treistman SN, Bayley H, Lemos IR: EEects of ethanol on calcium channels, potassium channels, and vasopressin release. A m NY Acad Sci 625; 249-263, 199 1.
Tremblay J, Huot C, Willenbrock RC, Bayard F, Gossard F. Fujio N, Koch C. Kuchel O. Debinski W, Hamet P: tncreased cyclic guanosine monophosphate production and overeqression of atrial natriuretic peptide A-receptor mRNA in spontaneously hypenensive rats. J Chil Itrvest 92; 2499-2508. 1993.
Trevisan M, Krogh V, Farinaro E, Panico S, Mancini M: Alcohol consumption, drinking pattern and blood pressure: aoaiysis of data fiom the Italian National Research Council study. bit J Epidemiol 16; 520-527, 1987.
Trippodo NC, Barbee RW: Atrial natriuretic fàctor decreases whole-body capillary absorption in rats. Am J Physiol252; W15-R920. 1987.
Tseng YC, La& S. Sellitti DF, Burman KD, D'Avis JC, Wartofsky L: Characterization by affindy cross-linking of a receptor for atriai natriuretic peptide in cultured human thyroid ceos

associated with reductions in both adenosine 3', 5'-monophosphate production and thyroglobulin secretion. J Clin Erufocrïtzol Metab 70; 528- 5 33, 1990.
Tsunoda K, Hodsman GP, Sumithran E, Johnson CI: Atrial natriuretic peptide in chronic heart fdure in the rat: A correlation with ventricular dysfunction. Cire Res 59; 256-26 1 . 1986.
Tsutsumi K, NRNa M, Kawano T, Ibaragi MA, Ozaki M, Mori K: Atrial natriuretic polypeptides elevate the level of cyclic GMP m the rat choroid plexus. Neurmci 79; 174- 178. 1987.
Tsutsumi M, Lasker JM. Shimizu M. Rosman AS, Lieber CS: The mtralobdar distniution of ethanol-mducible P45OIIE 1 m rat and hunian liver. Hepatology 10; 437-446, 1989.
Tuk R: Alcohol and the brain. MedJ Austr 156; 749-750, 1992.
Tuma DI, Casey C A Sorrell MF: Effects of ethanol on hepatic protem trafficking: Impairment of receptor-mediated endocytosis. A icohol AIcoholisrn 25 ; 1 17- 125, 1990.
Turlapaty PDMV, Altura BT, Amira BM: Ethanol reduces Ca2+ concentrations in arterial and venous smooth muscle. Experientia 3 5 ; 639-640, 1979.
Turner C: How m c b alcohol is a standard drink? An analysis of 125 midies. Br J Addict 84: 1171- 1175, 1990.
Uchida K, MiPlno T, Shimonaka M, Sugiura N, Nara K, Ling N, Hagiwara H Hirose S: Purification and properties of active atrial natriuretic peptide receptor (type C) f?om bovine h g . Biochern J 263; 67 1-678, 1989.
Ueda S, Sudoh T, Fukuda K, Kangawa Y Minamino N, Matsuo H: Indentification of alpha natriuretic peptide (4-28) and (5-28) in porcine brain. Biochern Bzophys Res Cornrnzoi 149: 1055- 1062, 1987.
Ueda S, MinamHio N, Sudoh T, Kangawa Y Matsuo H: Regional distniution of immunoreactive brai. natriuretic peptide in porcine bram and spmal cord. Biochern Biophys Res Cgmrnurt 155; 733-739, 1988.
Ueda S, Minamino N, Aburaya M, Kangawa K, Matsukura S, Matsuo H: Distribution and characterizition of immunoreactive porcine C-type natriuretic peptide. Biochem Biophys Res Commurt 175; 759-767, 1991.
Ueshima l3, Shmiamoto T, Iada M: Alcohol mtake and hypertension among urban and rural Japanese populations. J Chronzc Dis 37; 5 85-592, 1984.

Ueshima H, Mürawa K, Baba S, Sasaki S, Ozawa H, Tsushima M, Kawaguchi 4 ûrnae T. Katayama Y, Kayamori Y, Ito K Effect of reduced alcobol consumption on blood pressure m untreated hypertensive men. Hypertension 21; 248-252, 1993.
Urbano-Marquez 4 Estruch R Navarro-Lopez F, Grau JM, Mont L, Rubin E: 7%e effects of alcoholism on skeletal and cardiac muscle. N Engl J Med 320; 409-4 1 5, 1989.
Valentin JP, Sechi Qiu C, Schambelan M, Humphreys MH: Urodilatm binds to and activates renal receptors for atriai natriuretic peptides. Hypertension 2 1; 432-438, 1993.
Valimaki M ' Harkonen M, Eriksson CJP, Ytikahri RH: Sex hormones and adrenocortical steriods m man acutely mtoxicated with ethanol. A Icohof 1 ; 89-93, 1984.
Valverius P, Ho£Einan PL, Tabakoff B: Hippocampal and cerebellar LI-adrenergic receptors and adenylate cyclase are differentdly ahered by chronic ethanol mgestion. J Neurochem 52; 392-497, 1989.
Van de Wiel JAG, Dutjf CMP, Pertijs JCLM, Peereboom-Stegeman M C , Bos RP: Growth and ber morpholog- der long-term ethanol consumption of rats. La& Anzmals 24; 265-272. 1990.
Van Thiel DH, Gavaler JS, Lester E t Ethanol: A gonadal toxh in the fernale. Dmg Alcohol Depend 2; 373, 1977.
Van Thiel DH: Ethanol: Its adverse effects upon the hypothalamic-pituitary-gonadal a i s . J Lab Clin Med 101; 21033, 1983.
Vasdev S, Gupta IP, Sampson C A Longerich L, Parai S: Ethanol hduced hypertension in rats: Reversibility and role of intracellular cytosolic calcium. Artery 20; 19-43, 1993.
Vatta Ms, Papouchado ML, Locatelli AS, Bianciotti LG, Femandez BE: Effects of atrial natriuretic factor on norepmephrine release m the rat hypothalamus. Reg Peptides 4 1 ; 17 1- 181, 1992.
Veenstra J, te Wierik E, KI& C: Alcohol and fïbrinolysis. Fibririolysis 4 (suppl. 2) ; 64-68. 1990.
Veenstra J: Moderate alcohol use and coronary heart disease: A U-laped curve? W d d Rev Nutr Diet 65; 38-71, 1991.
Veldkamp PJ, Carmhes PK, Inscho EW, Navar LG: Direct evaluation of the microvascular actions of ANP m juxtarnedullary nephrons. Am J Physiol254; F440-F444, 1988.

Vesely DL, Nonis JS, Waiters JM, Jespersen Baeyens DA: Atrid natriuretic prohormone peptides 1- 10, 3 1-67, and 79-98, vasodilate the aorta. Biochem Biophys Res Commun 148; 1540- 1548, 1987.
Vesely DL, Wmters CJ, SaIlman AL: Atrial natriuretic fàctor in the pleural Duid of congestive heart faihire patients. Chest 95; 1 10% 1 109, 1989.
Vesely DL, Cornett LE, McCleod SL, Nash A4 Norris IS: Specific bmding sites for prohormone atriai natriuretic peptides 1-30,3 1-67, and 99- 126. Peptides 1 1; 193- 197, 1990.
Vesely DL, Palmer PA, Giordano AT: Atrial natriuretic factor prohormone is presnet in a vanety of tissues m addition to heart and brah. Peptides 13; 165- 170, 1992.
Vesely DL, Douglas Mq Dietz JR, Giordano AT, McCormick MT, Rodriguez-Paz G, Schocken DD: Tkee peptides f?om the atrial natriuretic factor prohormone animo terminus lower blood pressure and produce a diuresis natriuresis a d o r kaliuresis in humans. Circulation 90; 1 12% 1 140, 1994.
Vesely DL: Atrial natriuretic hormones origmatmg from the N-terminus of the atrial natriuretic factor prohomone. C h Exp Pharmacol Physiol22; 108- 1 14, 1995.
Victor M, A h RA, C o b s GH: The Wemicke-Korsakoff syndrome and related disorders due to alcoholism and malnutrition. Wiladelphia, F.A. Davis, 1989.
Vollmar AM, Arendt RM, Schulz R. The effects of opioids on rat atnal natriuretic peptide. Ezir J Pharmacol 143; 3 15-32 1, 1987.
Vollmar AM, Gerbes AL, Nemer M, Schuiz ik Detection of C-type natriuretic peptide (CNP) transcript m the rat hem and immune organs. Endocrino/ogy 132; 1872- 1874, 1993.
Volpe M, Ode1 G, Kleinert HD, Camargot MJ, Laragh JH, Lewicki JA, Maack T, Vaughan ED Jr, Atlas SA: Antihypertensive and aldosterone-lowering effects of synthetic atrial natriuretic factor in renh-dependent renovascular hypertension. J Hypertem 2; S3 13-S3 15. 1984.
Von Schroeder HP, Ni.chimura E, McIntosh CHS, Buchan AMJ, Wilson N, LeCsorne JR: Autoradiographic localization of bindmg sites for atrial natriuretic factor. Con J Physiol ~ h a c o f 63; 1373-1377, 1985.
Von Wartburg JP, Bethune E, VaIlee BL: Human iiver-alcohol dehydrogenase: kinetic and physicochemical properties. Biochem 3; 1775- 1782, 1964.
Vuolteenaho O, Arjamaa O, Vakkun O, Maksniemi L, Nikkila L, Kangas J, Puurunen J,

Ruskoaho H, Leppahioto J: Atrial natriuretic peptide (ANP) m rat gastromtestmd tract. FEBS Lett 233; 79-82, 1988.
Wadstem J, OhIin H: Changes m water, senun electrolytes and basic acid balance in alcoholism In: Merabolic efects of alcohol. Avogaro P, Sirton CR Tremoli E (eds), ELsaierNorth-Hohd biomedical press, p. 3 5 7-3 64, 1 979.
Wafford KA, Bmett DM, Leidenheimer NJ: Ethaool saiativity of the GABA, receptor expressesi m Xenopus oocytes reqiires eight amino aQds contained in the y z, subunit of the receptor complex Neuron 4; 27-34, 199 1.
Wakabayashi K Nakamura K, Kouo S, Shinchi K, Imanishi K: Alcohol consumption and blood pressure: an extenteci shidy of self-defence officiais m Japan. Int J Epiderniol23 ; 3 07- 3 11, 1994.
Wang Y, Pang CCY: Suppression by ethanol of pressor response caused by the inhiiition of nihic oxide synthesis. Eur J Pharmacoi 233; 275-278, 1993.
Wang YL, Wei IW, Sun AY: Effect of ethanol on braiu monoamine content of spontaneously hypertensive rats (SHR). Neurmhem Res 18; 1293- 1297, 1993.
Wehr H., Rodo M, Lieber CS, Baraona E: Acetaldehyde adduas and autoantibodies against VLDL and LDL m alcoholics. J Lipid Res 34; 1237- 1244, 1993.
Wei C, Heublem DM, Perrella M: Natriuretic peptide system in human heait failure. Circulation 88; 1004- 1009, 1 993.
Weidmann P, Hasler L, Gnadinger MP, Lang RE, Uehlinger DE, Shaw S, Rascher W, Reubi FC: Blood levels and rend effects of atrial natriuretic peptide m normal man. J C h Imest 77: 734-742, 1986.
Weidner G, Connor SL, Chesiey Bums JW, Connor WE, Mataravo ID, Mendell NR: Sex Merences m hi& density lipoprotem cholesterol among low-level alcohol consumers. Cimlation 83; 176- 180, 199 1.
Weisinger RS, Blair-West Denton DA, Tarjan E: Central anministration of a t n d natriuretic peptide suppresses sodium and water mtake of sheep. Brazn Res 579; 1 13- 1 18. 1992.
Weissfeld JL, Johnson EH, Brock BM, Hawthorne VM: Sex and age interactions E the association berneen alcohol and blood pressure. Am J Epidemiol 128; 5 59-569, 1988.
White G, LoWiger DM, Weight FF: Ethanol mhiits NMDA-activated current but does not

alter GABA-activated current m an isolated aduh mammalian neuron. Brazn Res 5 07; 33 2- 336, 1990.
Whitson PA, Huls MH, Sam CF: Characterization of atriai natriuretic peptide receptors in brah microvessel endothelid cells. J Cell Physol 1 46; 43-5 1, 1 99 1.
Widera W, Kokot F, Wiecek A: Do opioid receptors participate m the regulation of atrial natriuretic peptide (ANP) secretion m hypertensive patients? Clin NephroI 38; 209-2 13, 1992.
WigIe DA Pang SC, Sarda iR, Watson ID, Radakovic NN, Roy R, Rynn TG: Acute ethanol mgestion modifies the cirdating plasma levels of avial natriuretic peptide. Alcohol 10; 275- 280, 1993a.
Wigle DA, Pang SC, Radakovic NN, Sarda IR, Watson JD, Roy & Flynn TG: Chronic ethanol ingestion modifies the renin-aldosterone axis mdependent of alterations m the regulation of avial natriuretic peptide. Alcohol C h Exp Res 17; 84 1-846, 1993b.
Wigle DA, Pang SC, Watson ID, Sarda IR, Radakovic NN, Flynn TG: Chronic ethanol treatment mcreases the circulating plasna levels of B-type natriuretic peptide (BNP-45 ) in the rat. Am J Hypertens 6; 7 19-722, 1993c.
Wigle D A Watson JD, Pang SC, Sada IR, Roy RN, F i y ~ TG: Gene expression of A- and B-type natriuretic peptides in response to acute ethanol mgestion. Alcohol C h Exp Res 19; 13 17-1320, 1995.
Wilke N, Sganga M, Barhite S, Mites MF: Effects of alcohol on gene expression in neural cells. in: Tavard a rnolecuIm basis of alcohol me md abuse. Jansson B, J6mvall H, Rydberg U, Terenius L, VaiIee BL (eds), Basel, Switzerland, Birkhêuser Verlag, p. 49-59. 1994.
Williams JW, Tada M, Katz AM, Rubin E: Effect of ethanol and acetaldehyde on the (Na'+K?)-activated adenosine triphosphate actMty of cardiac plasma membranes. Biochem Phannacol 24; 27-32, 1975.
Winquin RJ, Faison EP, Waldman SA, Schwartz K, Murad F1 Rapoport RM: Atnal natriuretic factor elicits an endothelium-independent relaxation and activates particdate guanylate cyclase in vascular smooth muscle. Proc Nat1 A cad Sci USA 8 1 ; 76 6 1 -7664, 1 9 84.
Winters CJ, S a l h AL, Meadows J, Rico DM, Vesely DL: Two new hormones: Rohonnone atrial natriuretic peptides 1-30 and 3 1-67 circulate m man. Biochem Biophys Res Commun 150; 23 1-236, 1988.
Wong M: Depression of cardiac performance by ethanol lmmasked durin autonomie blockade. Am Heart J 86; 508-5 15, 1973.

Wu PH, Naranjo C A Fan T: Chronic ethanol mhiibits rat hippocampal "stirmili-secretion" coupling mechanism for 5-hydroxytqptamine in vitro. Neurochem Res 1 1 ; 80 1-8 12. 1986.
Yarnada T, Nakao K, Morii N: Centrai effect of atrial natriuretic polypeptide on angiotensin II sbmuLated vasopressh secretion in conscious rats. Eur J P h m a c o l 125; 453-458, 1986.
Yamaguchi M, Rutledge LT, Garbers DL: The primary structure of the rat guanylyl cyclase A/ a t r d natriuretic peptide receptor gene. J Biol Chem 265; 204 14-20420, 1990.
Yandle TG, Fisher S, Charles C, Espiner E& Richards AM: The ovine hypothalamus and pmutary have markediy Mirent distn'bution of C-type natriuretic peptide f o m . Peptides 14: 713-716, 1993.
Yandle TG, Richards AM, Gilbert 4 Fisher S, Hoimes S, Espmer EA: Assay of bram natriuretic peptide (BNP) in human plasma: Evidence for high molecular weight BNP as a major plasma component in heart failure. J Clin Endocrinol Metab 76; 832-83 8, 1993.
Yandle TG: Biochemistry of natriuretic peptides. J Int Med 235; 56 1-576, 1994.
Yasujima M, Abe K, Kohjuki M, Tanno M, Kasai Y, Sato M, Omata K, Kudo K, Tsunada K, Takeuchi K: Aaial nahniretic factor mhibits the hypertension mduced by chronic infùsion of norepmephrine in conscious rats. Circ Res 57; 470-474, 1985.
Yasujima M, Abe K, Kohzuki M, Tanno M, Kasai Y, Sato M, Omata K, Kudo K, Tsunoda K, Takeuchi K: Effect of atrial natriuretic Bctor on angiotensin Liinduced hypertension in rats. Hypertemioti 8; 748-753, 1986.
Yechieli H, Kahana L, Haramati 4 H o b A, Winaver J: Regdation of rend glornerular and papilhy ANP receptors m rat with experimental heart failure. Am J Physiol265 ; F 1 1 9- F125, 1993.
Yokota N, Aburaya M, Yamamoto Y, Kato J, Kitamura Y Kida O, Eto T, Minamino N. Kangawa K, Matsuo H, Tanaka K: Increased plasma bram natriuretic peptide levels in DOCA-salt hypertensive rats: Releation to blood pressure and cardiac concentrations. Biochern Biophys Res Commun 173,632-645, 1990.
Yokota N, Yamamoto Y, Kitamura K, Kangawa K, Minamino N, Matsuo H, Eto T: Alterations m circulatmg and cardiac tissue concentrations of brain natriuretic peptide in spontaneously hypertensive rats. Cardlovmc Res 27; 13 12- 13 1 5, 1993.
York JL: Body water content, ethanol pharmacokinetics, and the responsiveness to ethanol m young and old rats. Dev Pharmacol Ther 4; 106- 116, 1982.

Zeidel ML, Brenner BM: Actions of atnal natriuretic peptides on the kidney. Semin Nephrd 7; 9 1-97, 1987.
Zeidel ML, Kikeri D, Siha P, Burrowes M, Brenner BM: Atnal natriuretic peptides inhiiit conductive sodium uptake by rabbit niner medullary coleaing duct ce&. J Clin lnvest 82; 1067-1074, 1988.
Zeidel ML: Hormonal regulatioo of b e r meddiary coUectmg duct sodium transport. Am J Physiol265; F159-F173, 1993.
Zeidel ML: Regdation of collecting duct Na' reabsorptioo by ANP 3 1-67. Clirz Eup Pharmacol Physiol22; 121-124, 1995.
Zgombick IM, Erwin G, CorneII K: Etbanol-mduced adrenomedullary catecholamine secretion in LS/Ibg and SS/Ibg mice. J Pharmacol Exp ï3er 236; 634-640. 1986.
Zorad S, Tsutsumi K, Bhatia AJ, Saavedra JM: Localization and characteristics of atrial natriirretic peptide receptors in prenatal and postnatal rat brah Eur J Pharmacol 24 1 : 195- 200, 1993.
Zsotér TT, Selers EM: Effect of alcohoi on cardiovascular reflexes. J Sttid Alcohol38: 1- 10. 1977.
IMAGE EVALUATION TEST TARGET (QA-3)
APPLIED IMAGE. Inc 1653 East Main Street - -. , Rochester. NY 14609 USA -- -- , - Phone: 716/482-0300 -- -- - - Fax: 7 t 6/288-5989
O 1993. Applied image. inc.. Ali Righls Reserved
Related Documents











