Evidence for Endocytosis of ROMK Potassium Channel via Clathrin-Coated Vesicles Wei-Zhong Zeng 1 , Victor Babich 1 , Bernardo Ortega 3 , Raymond Quigley 2 , Stanley J. White 3 , Paul A. Welling 4 , and Chou-Long Huang 1 Departments of 1 Medicine and 2 Pediatrics, University of Texas Southwestern Medical Center, Dallas, TX 75390; 3 Department of Biomedical Science, University of Sheffield, Sheffield, S102TN, UK; 4 Department of Physiology, University of Maryland, School of Medicine, Baltimore, MD 21201 Running Title: Endocytosis of ROMK via CCVs. Corresponding Address: Chou-Long Huang, MD PhD Department of Medicine UT Southwestern Medical Center 5323 Harry Hines Blvd. Dallas, TX 75390-8856 Tel: 214-648-8627 Fax: 214-648-2071 E-mail: [email protected] Copyright 2002 by the American Physiological Society. AJP-Renal Articles in PresS. Published on April 16, 2002 as DOI 10.1152/ajprenal.00378.2001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Evidence for Endocytosis of ROMK Potassium Channel
via Clathrin-Coated Vesicles
Wei-Zhong Zeng1, Victor Babich1, Bernardo Ortega3, Raymond Quigley2,
Stanley J. White3, Paul A. Welling4, and Chou-Long Huang1
Departments of 1Medicine and 2Pediatrics, University of Texas Southwestern Medical Center,
Dallas, TX 75390; 3Department of Biomedical Science, University of Sheffield, Sheffield,
S102TN, UK; 4Department of Physiology, University of Maryland, School of Medicine,
Baltimore, MD 21201
Running Title: Endocytosis of ROMK via CCVs.
Corresponding Address:
Chou-Long Huang, MD PhDDepartment of MedicineUT Southwestern Medical Center5323 Harry Hines Blvd.Dallas, TX 75390-8856Tel: 214-648-8627Fax: 214-648-2071E-mail: [email protected]
Copyright 2002 by the American Physiological Society.
AJP-Renal Articles in PresS. Published on April 16, 2002 as DOI 10.1152/ajprenal.00378.2001

ABSTRACT
ROMK channels are present in the cortical collecting ducts (CCDs) of kidney and
are responsible for K+ secretion in this nephron segment. Recent studies suggest that
endocytosis of ROMK channels is important for regulation of K+ secretion in CCDs. We
investigated the molecular mechanisms for endocytosis of ROMK channels expressed in
Xenopus oocytes and cultured Madin-Darby canine kidney (MDCK) cells. When plasma
membrane insertion of newly synthesized channel proteins was blocked by incubation with
brefeldin A, ROMK currents decreased with a half-time of ~6 hrs. Coexpression with the
Lys-446 Ala (K44A) dominant-negative mutant dynamin, but not wild type dynamin,
reduced the rate of reduction of ROMK in the presence of brefeldin A. Mutation of Asn-
371 to Ile in the putative NPXY internalization motif of ROMK1 abolished the effect of
K44A dynamin mutant on endocytosis of the channel. Co-immunoprecipitation study and
confocal fluorescent imaging revealed that ROMK channels associated with clathrin coat
proteins in MDCK cells. These results provide compelling evidence for endocytosis of
ROMK channels via clathrin-coated vesicles.

Keywords: dominant-negative dynamin; MDCK cells; brefeldin A; Xenopus oocytes; tyrosine-
based consensus motif.
.

1
INTRODUCTION
The physiological role of the low-conductance K+ channels in apical membranes of
principal cells of the cortical collecting ducts (CCDs) in regulation of K+ secretion is well
established (43). Secretion of K+ in this nephron segment is mediated by active transport of K+
into the cell through basolateral Na+-K+-ATPase and followed by passive movement of K+ into
the tubular fluid through the apical K+ channels. cDNAs for ROMK1 and its isoforms ROMK-2
and -3 have been isolated (4, 15, 44). Based on the distribution of mRNA and proteins, and
biophysical characterization, it is known that the cDNAs for ROMK encode the low-
conductance K+ channels in the apical membranes of thick ascending limb of Henle’s loop and
CCD (11, 12). As the final common pathway for K+ secretion in CCDs, ROMK channels
are regulated by dietary K+ intake (43). To maintain K+ homeostasis, the ability of the kidney to
secrete K+ increases when dietary intake increases. This response, called K+ adaptation, is
associated with an increase in the number of active channels in rat CCDs. Studies using cell-
attached patch-clamp recording have found that the number of active K+ channels in the apical
membranes of CCDs increases by a factor of 3-4 when animals are fed a high K+ diet for 1-2
weeks (29). The increase in the density of active channels by high K+ intake is not associated
with an increase in the mRNA for ROMK in isolated rat CCDs (10). These results raise the
possibility that ROMK channels in the apical membranes of CCD undergo active trafficking and
the increase in channel density in K+-adapted animals may be due to alterations in these
processes.

2
Recently, Wang et al reported that Src kinase activity in the rat kidney cortex is inversely
correlated with dietary K+ intake in these animals (40). Low dietary K+ intake increases the
activity of Src kinase and whereas high dietary K+ intake decreases the activity. Application of
tyrosine kinase inhibitors and tyrosine phosphatase inhibitors increases and decreases,
respectively, the number of channels in cell-attached recording of CCDs isolated from these
animals as well as in oocytes co-expressing ROMK1 and recombinant Src kinase (24, 41). The
effects of these tyrosine kinase and phosphatase inhibitors on channel activity are prevented by
pretreating the cells with inhibitors of endocytosis (24, 41). These results suggest that alteration
of endocytosis of K+ channels is important for physiological regulation of the K+ channels in
CCDs by dietary K+ intake.
There are multiple pathways for endocytosis in mammalian cells, including phagocytosis,
internalization via caveolae, and clathrin-dependent endocytosis (17, 23, 25). Phagocytosis (or
“cell-eating”) occurs only in specialized cells. Caveolae are small microdomains of plasma
membranes that are enriched in cholesterol and glycosphingolipids. Some
glycophosphatidylinositol (GPI)-anchored proteins and receptors of the seven transmembrane
domain family are endocytosed via caveolae. Endocytosis via clathrin-coated vesicles is a well-
characterized form of pinocytosis and is a common mechanism for retrieval of plasma membrane
proteins (17, 23, 25).
The clathrin coat contains two oligomeric proteins, clathrin and clathrin adaptor protein
(AP) complexes (37). Clathrin is composed of three light (~33 kDa) and three heavy (~180 kDa)
chains that form a three-legged structure called triskelion. The AP complexes are
heterotetrameric. Thus far, 3 adaptor complexes, AP-1, AP-2, and AP-3, have been found (18).

3
The localization and function of the three AP complexes are different. AP-2 complexes are
localized to the coated pits and are involved in plasma membrane endocytosis. Assembly of
clathrin is not sufficient to drive vesicle budding. Budding of CCVs from plasma membranes
requires members of the ~100 kDa GTPase protein family, dynamins (34). The role of dynamins
in clathrin-dependent endocytosis from the plasma membrane is well established. Mutations of
dynamin that interfere with GTP binding and hydrolysis (such as lys-44 to ala) severely inhibit
CCV-mediated endocytosis from plasma membranes (8). In the present study, we sought to
investigate the role of CCVs in endocytosis of ROMK.

4
MATERIALS AND METHODS
Materials. All chemicals are from Sigma-Aldrich (St. Louis, MO) unless stated
otherwise. Restriction endonucleases are from either New England Biolabs, Gibco-BRL or
Roche-Mannheim. Brefeldin A stocks (in ethanol; 15 mM) were stored at -20O C before use.
Molecular biology. Wild type ROMK1 cDNA was in the pSPORT plasmid (15). Site-
directed mutagenesis of ROMK1 was performed using a commercial mutagenesis kit
(Quickchange from Stratagene, La Jolla, CA), and confirmed by nucleotide sequencing as
previously described (16, 19, 20). mCAP cRNAs of the wild type ROMK1 channels were
transcribed in vitro using T7 RNA polymerase (mMESSAGE kit from Ambion, Austin, TX) (16,
19, 20) after linearization of plasmid DNA with Not1 restriction endonuclease. cDNAs for
syntaxin-3 and syntaxin-5 were in pGEM vector which contain Xenopus β-globin 5'- and 3'-
untranslated regions (3). mCAP cRNAs for syntaxins were transcribed in vitro using T7 RNA
polymerase after linearization of plasmid DNA with Nhe1 restriction endonuclease (3). cDNAs
encoding c-myc-tagged wild-type and K44A mutant rat dynamin II were originally constructed
in pCMV-5 vector (2). EcoR1-Xba1 fragment was excised from the original pCMV-dynamin
plasmid and subcloned into pBluescrpit SK vector doubly digested with EcoR1 and Xba1.
mCAP cRNAs for wild-type and K44A dynamins were transcribed in vitro using T7 RNA
polymerase after linearization of pBluescript-dynamin plasmid DNAs with Xba1 restriction
endonuclease. Construction of the N-terminal enhanced green fluorescent protein (EGFP)-tagged
ROMK2 in pEGFP-C2 vector (Clontech), pEGFP-ROMK2, has been described (27).

5
Immuno-staining and laser scanning confocal imaging. MDCK cell lines stably
transfected with pEGFP-C2 empty vector or pEGFP-ROMK2 were grown in DMEM/F12 media
(1:1), 10% fetal calf serum, and selected with geneticin at 0.3 mg/ml as previously described
(27). Cells were sub-cultured at 1:10 dilution twice a week. For immunostaining by monoclonal
antibody against clathrin heavy chain, cells were grown in cover glass to ~50% confluence at 37o
C and further incubated at 25o C for 24-48 hrs. Surface expression of ROMK in MDCK cells is
low at 37o C (6, 27). Growing cells at 25o C increases the stability and surface expression of the
channel (6, 27). MDCK cells (in cover glass) were fixed in 4% formalin in PBS and
permeabilized by 0.1% Triton X-100. After blocking for the non-specific staining by 5% BSA,
cells were incubated sequentially with mouse monoclonal anti-clathrin heavy chain antibody
(ICN Biochemicals; 1:200 dilution), rabbit anti-mouse IgG (1:200) and rhodamine-conjugated
goat anti-rabbit IgG (Jackson Immuno-Research Laboratories). Fluorescent images were
visualized through a Zeiss 100 X objective lens using a Zeiss LSM410 microscope. To detect the
fluorescence of the green fluorescent protein (GFP), samples were excited with a 643 Kr/Ar ion
laser at 488 nm. Fluorescent emissions were passed through a 510/540 band pass filter. To detect
fluorescence given risen by rhodamine, samples were excited at 568 nm and emissions were
passed through a 590 nm long pass filter.
Co-immunoprecipitation of ROMK2 with clathrin coat proteins. MDCK cells stably
transfected with ROMK2 (cultured as above) were lysed in a RIPA buffer containing 150 mM
NaCl, 10 mM Hepes (pH 7.5), 1 mM EGTA, 1 mM DTT, 1% (v/v) NP-40, 1% (w/v) sodium

6
deoxycholate, 0.1% (w/v) SDS, 1 mM pheylmethylsulfonyl fluoride, 10 µM leupeptin, and 1 µM
pepstatin at 4O C for 1 hr. Lysates were centrifuged in a microfuge at 15,000 rpm at 4O C for 30
mins. Supernatants were incubated with either monoclonal antibody against α-adaptin (1:30
dilution; Transduction Lab) or control mouse IgG for 2 hrs and then precipitated by protein G
sepharose. The immunoprecipitates were separated by 10% SDS-PAGE and detected in
immunoblot analysis using monoclonal antibody against α-adaptin, against clathrin heavy chain,
and polyclonal antibody against ROMK1.
Two-electrode voltage-clamp recording. Xenopus laevis oocytes were prepared as
previously described (16, 19, 20). Oocytes were injected with cRNA for ROMK1, cRNA for
wild type or dominant-negative dynamin and/or cRNA for syntaxin as indicated. Current-voltage
(I-V) relationships (-100 mV to +100 mV, in 25 mV steps) were measured in oocytes at ~23O C
by two-electrode voltage-clamp (TEVC) using an OC-725C oocyte clamp amplifier (Warner
Instrument), pCLAMP6 software, and Digidata 1200A digitizer (Axon Instrument). The
resistance of current and voltage microelectrodes (filled with 3M KCl solution) was 1-2 MΩ.
The bath solution contained (in mM) 96 KCl, 1 MgCl2, 1 CaCl2, 5 Hepes (pH 7.5 by KOH).
Whole-cell patch-clamp and single-channel patch-clamp recording. For patch-clamp
recording, MDCK cells were grown to ~50% confluence at 37o C and further incubated at 25o C
for 24-48 hrs. Cells were dissociated by limited trypsin treatment and transferred to a recording
chamber. For ruptured whole-cell recording of MDCK cells, patch-clamp pipettes (pulled from
borosilicate glass, Warner Inst. Co., Hamden, CT) were filled with solutions containing (in mM):

7
140 KCl, 1 MgCl2, 2 Na2ATP, 1 EGTA, and 5 Hepes (pH 7.4 titrated with KOH). Bath solutions
contain (in mM): 140 KCl, 1 MgCl2, 1 CaCl2, 5 Hepes (pH 7.4 with KOH), and 5 glucose. For
cell-attached single-channel recording (in MDCK cells and in oocytes), pipette solutions were
(in mM): 100 KCl, 1 MgCl2, 2 CaCl2, and 5 Hepes (pH 7.4 titrated with KOH). Pipette tip
resistance ranged from 3 to 5 MΩ. Currents (whole-cell or single channel) were recorded with an
Axopatch 200B patch-clamp amplifier (Axon Instruments, Foster City, CA). Single-channel
currents were low-pass filtered at 1 kHz using an 8-pole Bessel filter, sampled every 0.1 ms (10
kHz) with Digidata-1200A interface and stored directly onto computer hard disk (100 GB) using
pCLAMP7 software (19). Data were transferred to CD for long-term storage. For analysis, event
list files were generated using the Fetchan program and analyzed for open probability (Po) and
amplitude histogram using pCLAMP7 pSTAT (version 6.0.5, Axon Instruments, Foster City,
CA). Po was analyzed on segments of continuous recording from patches that contained only one
active channel during the life-time of the recording. Po for opening was determined using the
criteria of threshold crossing of currents (50%) (19).

8
RESULTS
Endocytosis of ROMK1 in oocytes. Xenopus oocytes possess the machinery for
endocytosis via CCVs and have been used for studying CCV-mediated endocytosis of many ion
channels, including the epithelial Na+ channels, ENaC, and the CFTR Cl- channels (5, 38).
Current-voltage (I-V) relationships were recorded in oocytes (~5 oocytes for each time point)
every 24 hrs post-injection of cRNA to monitor plasma membrane expression of ROMK1
channels. After reaching the maximal steady-state expression of ROMK1 (~72 to 96 hrs),
brefeldin A (BFA) was added. BFA is a fungal metabolite that inhibits transport of the newly
synthesized channel proteins to plasma membranes by blocking anterograde trafficking of
vesicles from the endoplasmic reticulum to the Golgi complex (38, 42). BFA has been shown to
be active in Xenopus oocytes and inhibits protein trafficking in these cells (26). BFA does not
affect CCV-mediated endocytosis in plasma membranes (42). I-V relationships were again
recorded from oocytes at 3, 6, 12, 24, and 48 hr after addition of BFA (labeled as “+BFA”, Fig.
1A). After 3 or 24 hrs incubation with BFA, some of the oocytes were transferred to BFA-free
solutions for further incubation (“washout”). Another group of oocytes received vehicle
(ethanol) instead of BFA from the beginning and served as time-controls (“- BFA”).
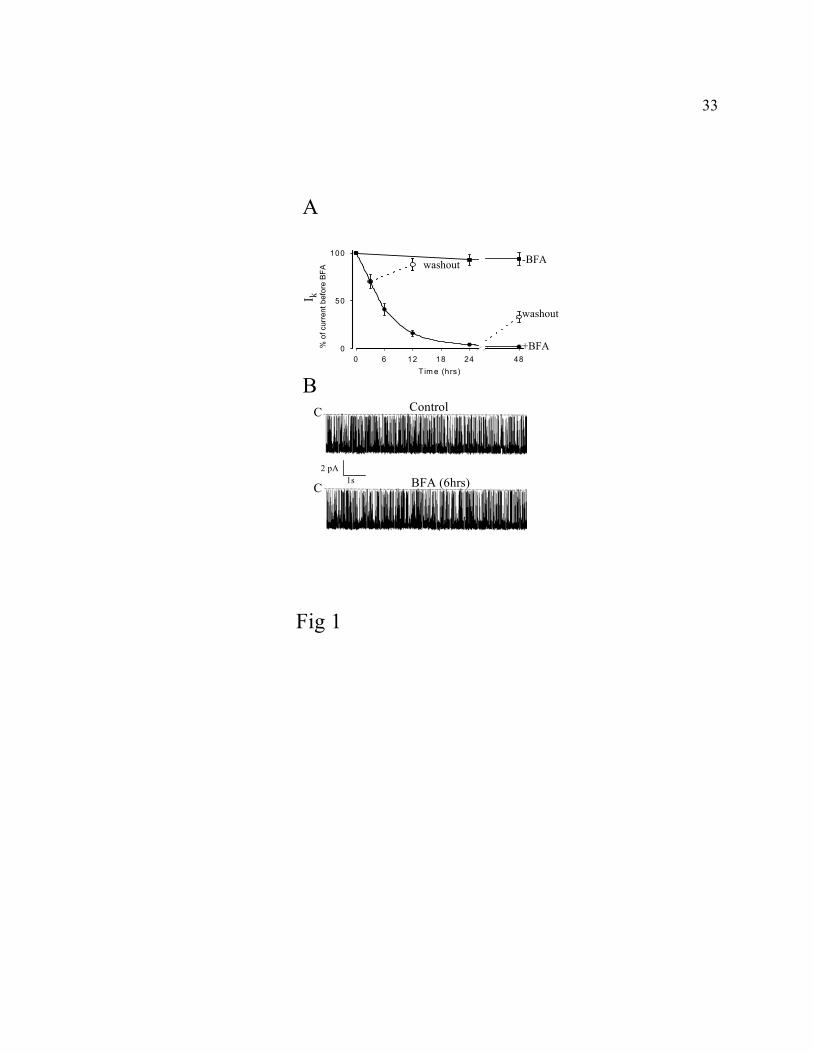
As shown in Fig. 1A, whole-cell inward K+ currents (Ik, measured at -100 mV, shown as
“% of current before BFA”) decreased to the level of background currents during 24 hrs of
incubation with BFA. Currents measured showed the characteristic weak inward rectification in
I-V relations and were sensitive to Ba2+ (not shown). ROMK1 currents recovered if oocytes were
transferred to BFA-free solutions after incubation in BFA, suggesting that the effect of BFA was

9
not toxic. The half-time for reduction in currents (t1/2, see legends to Fig. 1A) in the presence of
BFA was 5.7 ± 2.2 hrs (n = 6). For comparison, ENaC currents decay with a half-time of 3.6 hrs
in oocytes in the presence of BFA (38). Cell-attached single-channel recording was performed in
oocytes before addition of BFA (labeled “control”) and in oocytes after incubating in BFA for 6
hrs (Fig. 1B). As shown, neither single-channel conductance nor open probability (Po) of the
channels were affected by BFA (Po: 0.91± 0.08 at -100 mV and conductance 36 ± 2.3 pS from 0
to -100 mV, n = 5, for control vs Po: 0.90 ± 0.14 and conductance 35 ± 1.8 pS, n = 8, for BFA-
treated cells; Fig. 1B). These results suggest that the progressive reduction of ROMK currents in
BFA is not due to an effect of BFA on the opening of the channels, but rather is a result of a
decrease in the number of functional channels in the plasma membrane. This decrease in the
number of functional ROMK channels in the plasma membrane may be due to endocytosis or
may be due to degradation and/or sequestration of channels by other membrane proteins. The
role of endocytosis will be examined in this report.
Inhibition of ROMK1 currents by syntaxin-5. As an independent confirmation for the
results from oocytes incubated with BFA, we used syntaxin-5 over-expression as an additional
tool to inhibit transport of the newly synthesized ROMK1 to plasma membranes. Syntaxins are a
family of membrane proteins that are involved in the trafficking of membrane vesicles (39).
Syntaxin-5 is expressed in the Golgi complex and plays an important role in regulating vesicle
transport between endoplasmic reticulum (ER) and the Golgi (9). Over-expression of syntaxin-5
disrupts the stoichiometric interactions among the endogenous transport-associated regulatory
proteins and thus inhibits ER to Golgi transport (33, 35).

10
ROMK1 currents in plasma membranes of oocytes were monitored using two-electrode
voltage-clamp recording (Fig. 2). After reaching the maximal steady-state expression of ROMK1
currents, one group of oocytes were injected with cRNA for syntaxin-5 (3 ng) while the other
group of oocytes were injected with water to serve as controls (Fig. 2A). As shown, ROMK1
currents decreased by 76 ± 12 % over 24 hrs in oocytes injected with cRNA for syntaxin-5 (open
circles), but not in water-injected oocytes (closed circles). The relatively slower time course of
reduction in ROMK currents by over-expression of syntaxin-5 (compared to by incubation with
BFA) may be due to the delay from expression of syntaxin in oocytes.
Single-channel conductance and Po for ROMK1 channels recorded after 24 hr injection of
syntaxin-5 were not different from those in water-injected controls (not shown), suggesting the
effect of syntaxin-5 was due to a reduction in the number of active channels. Fig. 2B shows the
dose-dependent effect of various concentrations of cRNA for syntaxin-5 injected after maximal
expression of ROMK1 currents in plasma membranes (closed circle and solid line). The amount
of cRNA required for inhibition of ROMK1 currents (1-3 ng) was comparable to the
concentrations of syntaxin cRNA found to regulate other channels in oocytes (3, 28, 31, 36). In
contrast to syntaxin-5, syntaxin-3 is expressed in plasma membranes and regulates vesicle
trafficking at this site (21, 22, 32). As expected from its non-Golgi location, syntaxin-3 (injected
after maximal expression of ROMK1) did not cause a reduction in ROMK1 currents (Fig. 2B,
open circle and dotted line). Expression of syntaxin-3 in plasma membranes of oocytes has been
demonstrated in many studies (28, 31, 36).
Inhibition of endocytosis of ROMK in oocytes by coexpression with a dominant-negative

11
dynamin. The lysine-44 to alanine mutation of dynamin impairs GTP binding and hydrolysis.
Over-expression of this K44A dominant-negative mutant dynamin inhibits budding of CCVs and
endocytosis from plasma membranes, but does not inhibit budding of vesicles from the trans-
Golgi network (1, 8).
The role of CCVs in endocytosis of ROMK1 was examined using K44A mutant rat
dynamin II (14). Oocytes were injected with cRNA for ROMK1 combined with cRNA for either
wild type or K44A mutant dynamin. Seventy-two hrs after injection of cRNAs, I-V relationships
were recorded in oocytes by TEVC. Afterwards, oocytes received either BFA (5 µM) or vehicle
(ethanol) and I-V relationships were recorded every 6 hrs for an additional 24 hrs. As shown in
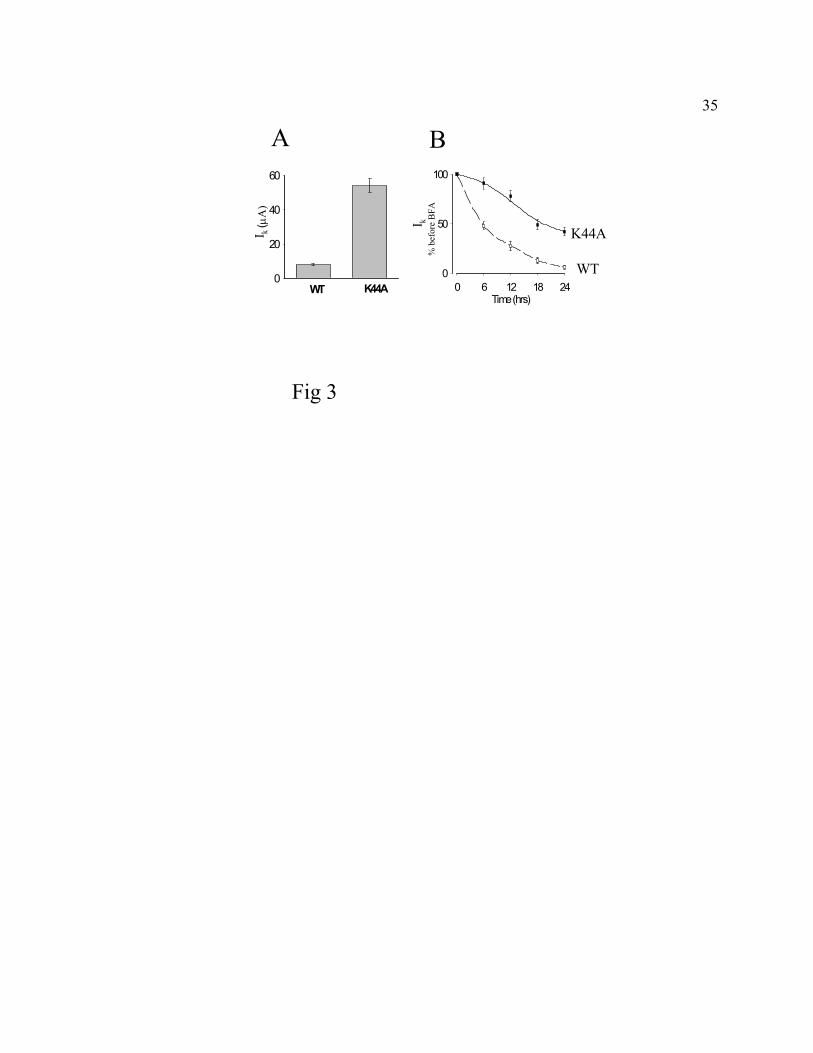
Fig. 3A, oocytes co-expressing ROMK1 and K44A mutant dynamin (“K44A”) for 72 hrs after
injection of cRNA had higher currents (Ik, 54 ± 8 µA, at -100 mV) as compared to those co-
expressing ROMK1 and wild type dynamin (“WT”, 8 ± 1 µA). Furthermore, the reduction in
ROMK1 currents over 24 hrs incubation of BFA was markedly inhibited in oocytes co-
expressing ROMK1 and K44A mutant dynamin (Fig. 3B). The half-time for reduction in
currents in oocytes treated with BFA was 5.5 ± 3.2 hrs (n = 4) and 18.3 ± 3.7 hrs (n = 4)
(p<0.05, unpaired t-test) for oocytes co-expressing wild type and dominant-negative dynamin,
respectively. The relatively smaller reduction in ROMK currents observed in oocytes co-
expressing K44A dynamin mutant may be caused by dynamin-independent internalization and/or
degradation pathways (17, 23, 25). As was reported for ENaC in oocytes (38), baseline
endocytosis of ROMK1 was not affected by over-expression of wild type dynamin (compare the
reduction in currents in BFA for ROMK1 + WT dynamin in Fig. 3B with that for ROMK1 alone
in Fig. 1A). These results suggest that endocytosis of ROMK1 in oocytes is, at least partly,

12
mediated by dynamin-dependent CCVs.
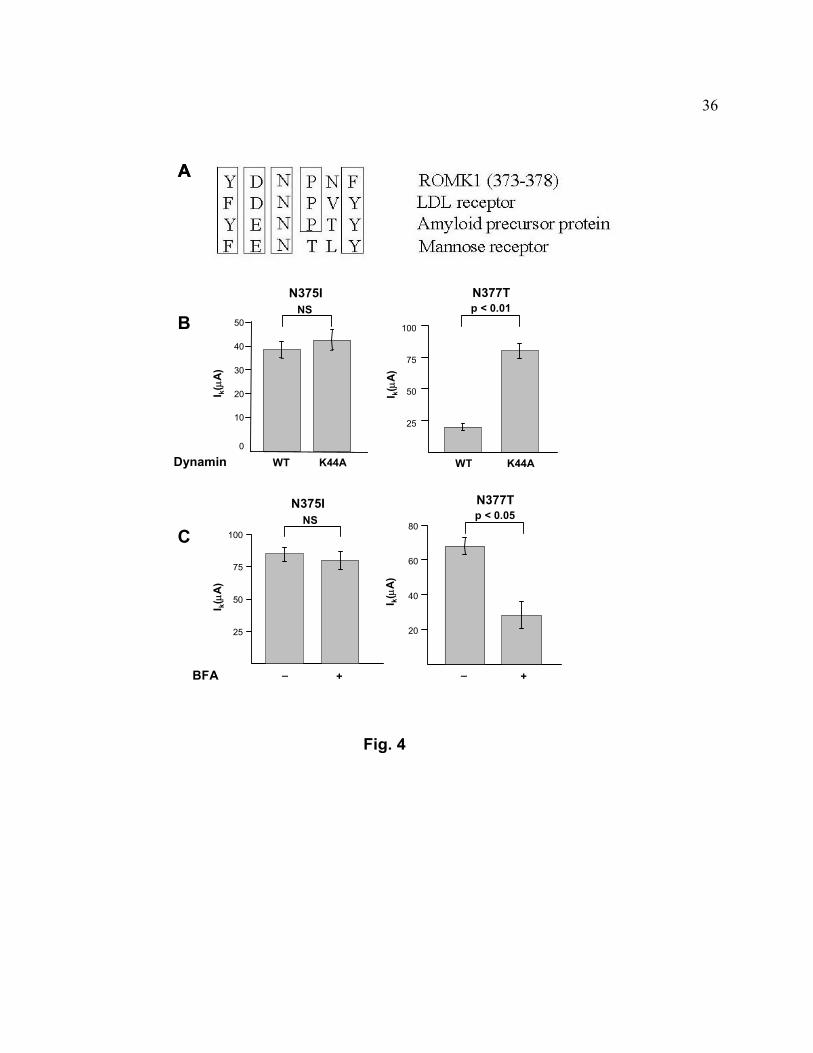
Internalization signal for endocytosis via CCVs. The C-terminal tail of ROMK1 (amino
acids 373-378) contains the sequence YDNPNF which is highly homologous to
(Y/F)(D/E)NPXY internalization motif of many membrane proteins that are internalized via
CCVs (23), such as LDL receptor, β-amyloid precursor protein, and the mannose receptor (Fig.
4A). To test the importance of this region for endocytosis of ROMK1, asparagine-375 was
mutated to isoleucine and the effect on ROMK1 currents in oocytes was examined. We found
that mutation of asn-375, but not of asn-377, abolished the ability of the K44A dynamin mutant
to enhance ROMK currents (Fig. 4B, also compare with Fig. 3A). Moreover, mutation of asn-
375, but not asn-377, abolished the decay of ROMK1 in the presence of BFA for 6 hrs (Fig. 4C,
also compare with Fig. 1A).
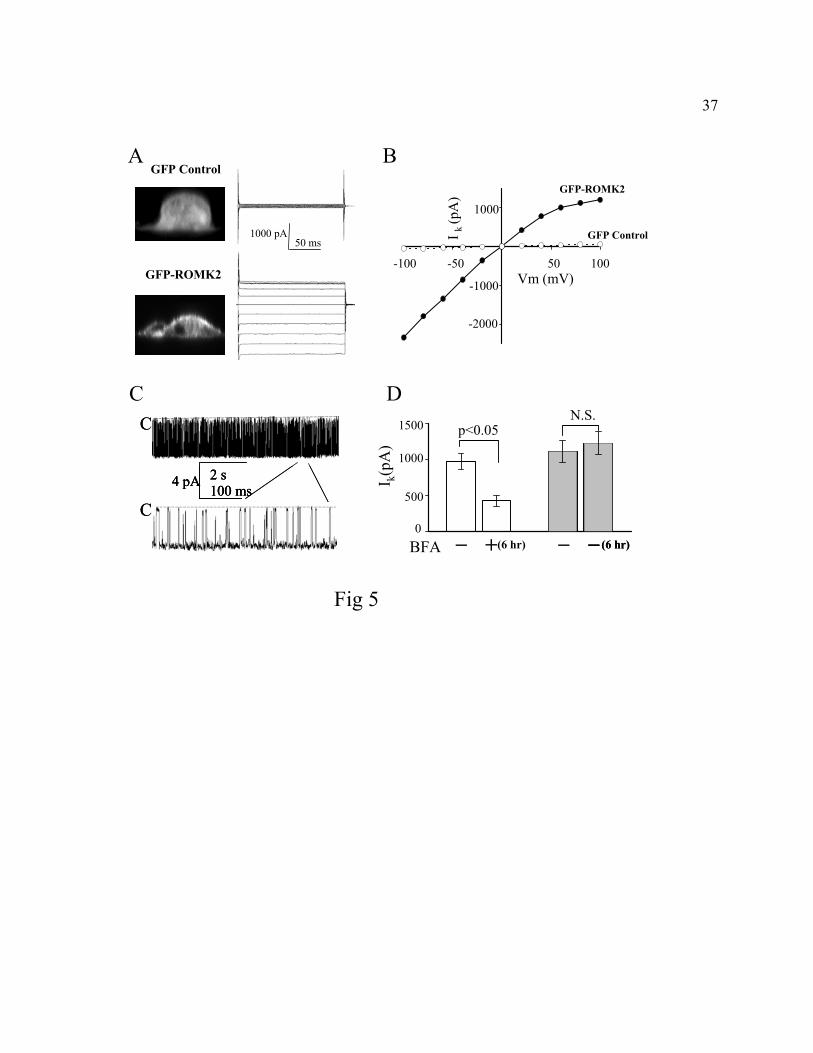
Endocytosis of ROMK2 in MDCK cells. To examine endocytosis of ROMK in a
mammalian cell system, we used a MDCK cell line stably transfected with a green fluorescent
protein-tagged ROMK2. Fusion of GFP N-terminal to the ROMK peptide does not affect
trafficking and function of the channel (27). Stable transfection avoids potential artefacts of
overexpression of proteins commonly observed with transient transfection. As reported
previously (27), GFP was distributed throughout the cytoplasm and nuclei (Fig. 5A, left upper
panel) and GFP-tagged ROMK2 was targeted to the plasma membrane of MDCK cells (lower
panel). Functional channel activity on the cell surface was confirmed by patch-clamp recording.
Whole-cell inwardly rectifying K+ currents were recorded from cells expressing GFP-ROMK2

13
(Figs. 5A and 5B). Currents recorded from cells expressing GFP alone (Figs. 5A and 5B) were
not significantly different from background currents in the control un-transfected cells (not
shown). The single-channel kinetics and open probability of GFP-ROMK2 in MDCK cells (Fig.
5C and data not shown) were not significantly different from channels expressed in oocytes (19).
When exocytotic insertion of ROMK was blocked by BFA, whole-cell inward-rectifying K+
currents in MDCK cells decreased by 56% over 6 hrs (Fig. 5D, open column). Currents did not
change significantly if cells were incubated with vehicle (ethanol) (Fig. 5D, hatched column).
Incubation with BFA did not affect single-channel conductance or Po of GFP-ROMK2 in MDCK
cells (not shown).
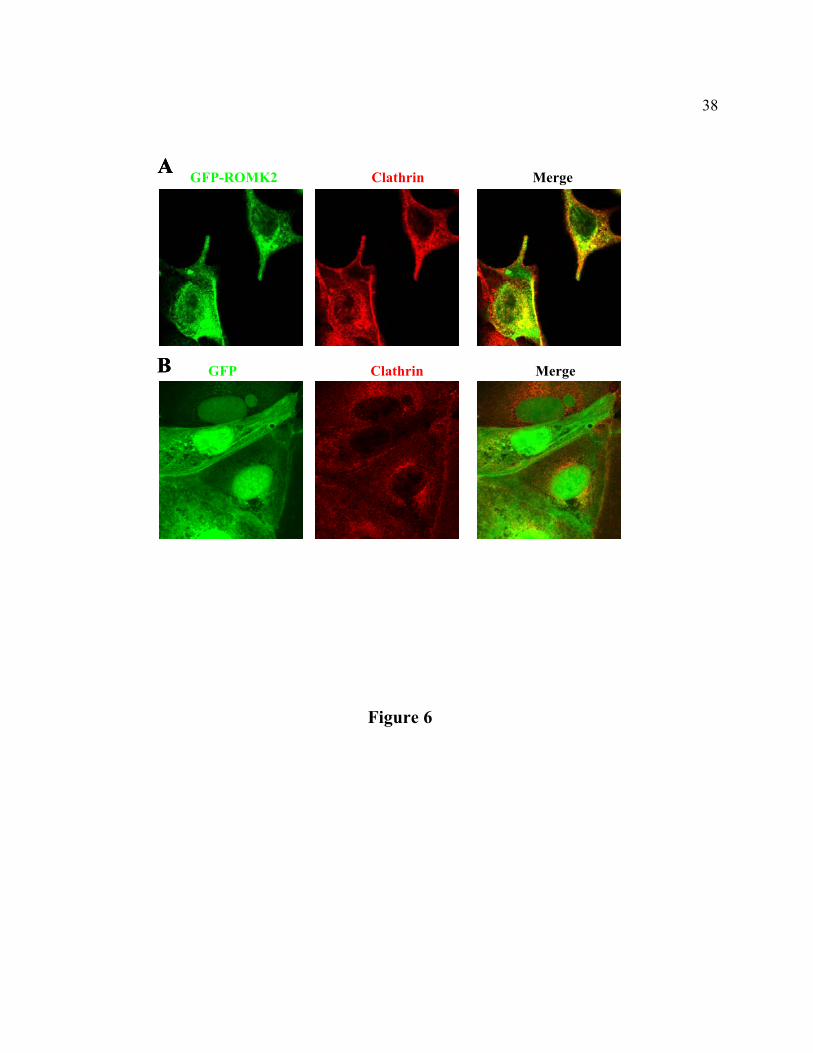
Localization of ROMK in CCVs. To provide direct evidence for endocytosis of ROMK
via CCVs, we examined the subcellular distribution of GFP-ROMK2 in MDCK cells. The
distribution of CCVs was examined by labeling with a monoclonal antibody against clathrin
heavy chain. Due to non-specific interactions of antibodies with the filter membrane, it was not
feasible in these studies to grow cells in transwells for obtaining optimal confocal images along
the z-axis as shown in Fig. 5A. Nevertheless, as shown in the image sectioned at the xy plane
(Fig. 6A, left panel), GFP-ROMK2 in MDCK was evidently distributed to the plasma
membrane. Additionally, GFP-ROMK2 was present intracellularly in a punctate pattern,
suggesting localization to the intracellular organelles. The staining by anti-clathrin heavy chain
antibody was membranous as well as punctate intracellularlly (Fig. 6A, middle panel). This
distribution is consistent with the pattern of distribution of CCVs (1). Partial co-localization of
GFP-ROMK2 with CCVs was evident from the merged image of distribution of GFP-ROMK2

14
and clathrin heavy chain (Fig. 6A, right panel; yellow color indicates co-localization). The
localization of ROMK to the plasma membrane-associated CCVs suggests endocytosis of the
channel at the plasma membrane. Besides endocytosis of plasma membrane proteins, CCVs are
also involved in transport of proteins from trans-Golgi network (TGN) to lysosomes (37).
ROMK channels expressed in MDCK cells are labile (6, 27). A significant fraction of channel
protein is degraded before being targeted to the surface membrane (6, 27). The co-localization of
ROMK with CCVs in the peri-nuclear region likely represents proteins in the synthetic pathway
or the pathway for degradation. As reported previously (27) and also shown in Fig. 5A, GFP was
distributed in cytoplasm and nuclei (Fig. 6B, left panel). For comparison, GFP was not localized
to the plasma membrane-associated CCVs (Fig. 6B, right panel). A small amount of GFP was
found in the peri-nuclear CCVs, probably representing proteins in the synthetic and/or
degradation pathway.
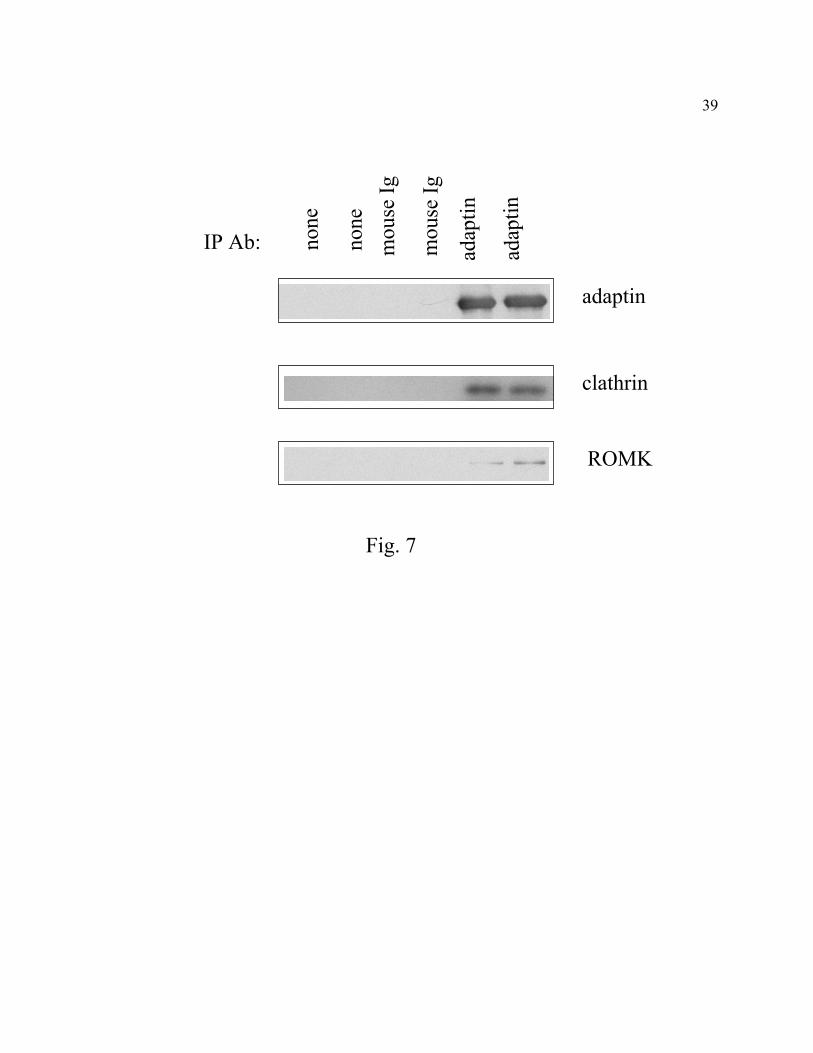
Co-immunoprecipitation of ROMK with clathrin coat proteins. We further investigated
endocytosis of ROMK via CCVs using co-immunoprecipitation. Besides endocytosis of plasma
membrane proteins, CCVs are also involved in transport of proteins from trans-Golgi network
(TGN) to lysosomes (37). CCVs-mediated endocytosis at plasma membrane involves AP2
adaptor proteins (containing α-adaptin subunit) whereas transport of proteins between TGN and
lysosomes involves AP1 adaptor proteins (containing γ-adaptin). To examine association of
ROMK with plasma membrane-associated CCVs, we used antibody against α-adaptin for
immunoprecipitation. As shown in Fig. 7, antibody against α-adaptin but not control IgG co-
immunoprecipitated clathrin heavy chain and ROMK from MDCK cells expressing ROMK2.

15
DISCUSSION
Several studies have suggested that ROMK channels undergo endocytosis in the plasma
membrane of Xenopus oocytes and the rat CCDs (24, 40, 41). The purpose of this study was to
examine the kinetics and the role of CCVs in endocytosis of ROMK channels. The abundance of
ROMK proteins in plasma membranes in the steady-state represents the balance between
exocytotic insertion of the newly synthesized proteins and retrieval of the proteins. Using BFA
(or over-expression of syntaxin-5) to block the anterograde transport of the newly synthesized
proteins to plasma membranes, we found that ROMK1 channels undergo endocytosis in plasma
membranes of oocytes with a half-time of ~6 hrs by electrophysiological recording.
Electrophysiological recording was chosen for these studies because it is highly quantitative and
selective for active channels that are present in the plasma membranes only. Additionally, it
allows us to compare our results with the published literature on regulation of K+ channel by
changes in dietary K+ intake (which mainly examine density of active channels using
electrophysiological recordings [10, 24, 29, 30, 40, 41]) (see discussion below).
ROMK channels are expressed in renal tubular epithelial cells (12). To know whether the
endocytosis we observed also occurs in the renal epithelial cells, we further examined
endocytosis of the channel using a MDCK cell line stably expressing ROMK2. We found that
endocytosis of ROMK2 also occurs in these cells with a time course similar to the endocytosis of
ROMK1 in oocytes. ROMK2 lacks the first 19 amino acids of ROMK1 and is otherwise
identical to ROMK1 (12). The similar rate of endocytosis of ROMK1 and 2 is also consistent
with the finding of a NPXY internalization motif in the C-terminus of both ROMK1 and 2.

16
There are multiple pathways for endocytosis (17). Endocytosis of ENaC and CFTR both
involve clathrin-coated pits (5, 38). We therefore examined whether CCVs are involved in
endocytosis of the ROMK channels. Dynamins are a ~100 kDa GTPase protein family which
play an important role in endocytosis mediated via CCVs (1, 8). The role of dynamin-dependent
CCVs in endocytosis of ROMK in oocytes is demonstrated by inhibition of endocytosis by co-
expression of the channels with the dominant-negative dynamin mutants. Dynamin has also been
reported to be involved in internalization of caveolae (13). Direct demonstration of ROMK in
CCVs will provide further confirmation of endocytosis of ROMK via CCVs. We attempted to
examine the localization of ROMK in CCVs in oocytes using immunofluorescent staining. But,
specific staining of CCVs in Xenopus oocytes was not feasible using available commercial
antibodies. Localization of ROMK with plasma membrane-associated CCVs, nevertheless, is
evident in MDCK cells using immunofluorescent co-localization and co-immunoprecipitation.
The cytoplasmic domains of many membrane proteins that are internalized via CCV-
mediated endocytosis contain specific sequence information that facilitates localization to coated
pits (23). Several types of internalization signals have been found (23). One of these is the
“NPXY motif”, which contains consensus amino acid sequence, asparagine, proline, any amino
acids, and tyrosine, respectively (7, 23). In this study, we also reported that C-terminus of
ROMK1 contains a NPXY internalization motif. These results, together with the results of
dominant-negative dynamin and the results of association of ROMK with CCVs, provide strong
evidence that ROMK channels are regulated by CCV-mediated endocytosis. The possibility that
ROMK may also be regulated by other internalization pathways, however, can not be excluded.

17
Endocytosis via CCVs in plasma membranes has multiple roles. These roles include
maintenance of cellular homeostasis by recovering protein components that are inserted into the
plasma membrane by ongoing secretory activity, uptake of molecules from the extracellular
space, and regulation of the number of membrane proteins in physiological and
pathophysiological conditions (17). Several lines of evidence suggest that regulation of the
apical membrane channel density by dietary K+ intake may occur via alterations of endocytosis
of the channels (24, 40, 41). High K+ intake may decrease CCV-mediated endocytosis of the
channels in the apical membrane and thus leads to the increase in the density of the channels. A
recent study reported that the increase in the density of active K+ channel in rat CCDs occurs
within 6 hrs (essentially one meal) of a high K+ diet (30). We found that the half-time for
reduction in ROMK currents in oocytes in the presence of BFA is ~6 hrs. This value may
represent an underestimation of the rate of endocytosis of ROMK in plasma membranes if
significant amounts of the endocytosed channels are recycled back to the plasma membranes. On
the other hand, it may be an overestimation if significant pools of inactive channels not readily
available for endocytosis are present in the plasma membranes. Nevertheless, the time course of
endocytosis of ROMK measured by electrophysiological recording in our studies is in
agreement with the time course of changes in the density of active K+ channels in CCDs
associated with alterations of dietary K+ intake. The similar time course supports the hypothesis
that variations of dietary K+ intake regulate K+ channel density by altering CCV-mediated
endocytosis of the channels.
What might be the signaling mechanism(s) that link the changes in dietary K+ intake to
the control of channel density via alterations of endocytosis of the channels? Studies by Wang et

18
al (24, 40, 41) suggest that changes in the dietary K+ intake affect the activity of the Src tyrosine
kinase and the related tyrosine phosphatases, which in turn alters the rate of endocytosis and/or
exocytosis of the channels. Having found that the mechanism of the endocytosis of ROMK is via
CCVs and determined the kinetics of endocytosis, one can examine in future studies how
tyrosine kinases and/or phosphatases regulate the CCV-mediated endocytosis of the channels.

19
ACKNOWLEDGMENTS
We thank Drs. Barbara Barylko and Joe Albanesi for wild type and mutant dynamin cDNA, Dr.
I. Bezprozvanny for syntaxin-3 and -5 cDNA, Dr. Y. M. Leung for participation in the work in
the early phase, Dr. Xin-Ji Li and Mr. Wei Ding for technical assistance, and Dr. Charles Pak for
support and encouragement. This work was supported in part by NIH grants (DK-54368 to CLH;
DK-54231 to PAW; P01-DK-20543 to Dr. Pak) and grant-in-aid from AHA national center
(0150179N to CLH).

20
REFERENCES
1. Altschuler, Y., S. M. Barbas, L. J. Terlecky, K. Tang, S. Hardy, K. E. Mostov, and S.
L. Schmid. Redundant and distinct functions for dynamin-1 and dynamin-2 isoforms. J.
Cell Biol. 143: 1871-1881, 1998.
2. Barylko, B., D. Binns, D., K.-M. Lin, M. A. Atkinson, D. M. Jameson, H. L. Yin, and J.
P. Albanesi. Synergistic activation of dynamin GTPase by Grb2 and phosphoinositides. J.
Biol. Chem. 273: 3791-3798, 1998.
3. Bezprozvanny, I., R. H. Scheller, and R. W. Tsien. Functional impact of syntaxin on
gating of N-type and Q-type calcium channels. Nature 378: 623-626, 1995.
4. Boim, M. A., K. Ho, M. E. Shuck, M. J. Bienkowski, J. H. Block, J. L. Slightom, Y.
Yang, B. M. Brenner, and S. C. Hebert. ROMK inwardly rectifying ATP-sensitive K+
channel. II. Cloning and distribution of alternative forms. Am. J. Physiol. 268, F1132-
F1140, 1995.
5. Bradbury, N. A., J. A. Clark, S. C. Watkins, C. C. Widnell, H. K. Smith, and R. J.
Bridges. Characterization of the internalization pathways for the cystic fibrosis
transmembrane conductance regulator. Am. J. Physiol. 276: L659-L668, 1999.
6. Brejon, M, S. Le Maout, P. A. Welling, and J. Merot Processing and transport of
ROMK1 channel is temperature-sensitive. Biochem. Biophys. Res. Commun. 261:364-371,
1999.
7. Chen, W. J., J. L. Goldstein, and M. S. Brown. NPXY, a sequence often found in
cytoplasmic tails, is required for coated pits mediated internalization of the low density

21
lipoprotein receptor. J. Biol. Chem. 265: 3116-3123, 1990.
8. Damke, H., T. Baba, D. E. Warnock, and S. L. Schmid. Induction of mutant dynamin
specifically blocks endocytic coated vesicle formation. J. Cell Biol. 127: 915-934, 1994.
9. Dascher, C., J. Matteson, and W. E. Balch. Syntaxin 5 regulates endoplasmic reticulum to
Golgi transport. J. Biol. Chem. 269: 29363-29366, 1994.
10. Frindt, G., H. Zhou, H., Sackin, and L. G. Palmer. Dissociation of K channel density and
ROMK mRNA in rat cortical collecting tubule during K adaptation. Am. J. Physiol. 274:
F525-F531, 1998.
11. Giebisch, G. Renal potassium channels: An overview. Kid. Int. 48: 1004-1009, 1995
12. Hebert, S. C. An ATP-regulated, inwardly rectifying potassium channel from rat kidney
(ROMK). Kid. Int. 48: 1010-1016, 1995.
13. Henley, J. R, E. W. Krueger, B. J. Oswald, and M. A. McNiven. Dynamin-mediated
internalization of caveolae. J. Cell Biol. 141:85-99, 1998.
14. Herskovitis, J., C. Burgess, R. Obar, and R. Vallee. Effects of mutant rat dynamin on
endocytosis. J. Cell Biol. 122: 565-576, 1993.
15. Ho, K., C. G. Nichols, W. J. Lederer, J. Lytton J, P. M. Vassilev, M. V. Kanazirska,
and S. C. Hebert. Cloning and expression of an inwardly rectifying ATP-regulated
potassium channel. Nature 362: 31-38, 1993.
16. Huang, C. L, S. Feng, and D. W. Hilgemann. Direct activation of inward rectifier
potassium channels and its stabilization by Gβγ. Nature 391:803-806, 1998.
17. Lamaze, C., and S. L. Schmid. The emergence of clathrin-independent pinocytic
pathways. Cur. Opin. Cell Biol. 7: 573-580, 1995.

22
18. Le Borgne, R., and B. Hoflack. Mechanisms of protein sorting and coat assembly: insights
from the clathrin-coated vesicle pathway. Curr. Opin. Cell Biol. 10: 499-503, 1998.
19. Leung, Y. M, W. Z. Zeng, H.-H. Liou, C. R. Solaro, and C.-L. Huang.
Phosphatidylinositol 4,5-bisphosphate and intracellular pH regulate the ROMK1 potassium
channel via separate but interrelated mechanisms. J Biol Chem. 275:10182-10189, 2000
20. Liou, H. H, S. S. Zhou, and C.-L. Huang. Regulation of ROMK1 channel by protein
kinase A via a phosphatidylinositol 4,5-bisphosphate-dependent mechanism. Proc. Natl.
Acad. Sci. USA 96:5820-5825, 1999.
21. Low, S.-H., S. J. Chapin, T. Weimbs, and L. G. Kömüves, M. K. Bennett, and K. E.
Mostov. Differential localization of syntaxin isoforms in polarized Madin-Darby canine
kidney cells. Mol. Biol. Cell 7: 2007-2018, 1996.
22. Mandon, B., S. Nielsen, B. A. Kishore, and M. A. Knepper. Expression of syntaxins in
rat kidney. Am. J. Physiol. 273: F718-F730, 1997.
23. Mellman, I. Endocytosis and molecular sorting. Annu. Rev. Cell Dev. Biol. 12: 575-625,
1996.
24. Moral, Z., K. Dong, Y. Wei, H. Sterling, H., Deng, S. Ali, R. Gu, R., X.-Y. Huang, S. C.
Hebert, G. Giebisch, W.-H. Wang, W.-H. Regulation of ROMK1 channels by
protein-tyrosine kinase and -tyrosine phosphatase. J. Biol. Chem. 276: 7156-7163, 2001.
25. Mukherjee, S. A., R. N. Ghosh, and F. R. Maxfield. Endocytosis. Physiol. Rev. 77: 759-
803, 1997.
26. Mulner-Lorillon, O., R. Belle, P. Cormier, S. Drewing, O. Minella, R. Poulhe, and G.
Schmalzing. Brefeldin A provokes indirect activation of cdc2 kinase (MPF) in Xenopus

23
oocytes, resulting in meiotic cell division. Dev. Biol. 170: 223-229, 1995.
27. Ortega, B., I. D. Millar, A. H. Beesley, L. Robson, and S. J. White. Stable, polarized,
functional expression of Kir1.1b channel protein in Madin-Darby canine kidney cell line. J.
Physiol. 528:5-13, 2000.
28. Naren, A. P., D. J. Nelson, W. Xie, B. Jovov, J. Persner, M. K. Bennett, D. J. Benos,
M. W. Quick, and K. L. Kirk. Regulation of CFTR chloride channels by syntaxin and
munc 18 isoforms. Nature 390: 303-305, 1997.
29. Palmer, L. G., L. Antonian, L., and G. Frindt. Regulation of apical K and Na channels
and Na/K pumps in rat cortical collecting tubule by dietary K. J. Gen. Physiol. 104: 693-
710, 1994.
30. Palmer, L. G., and G. Frindt. Regulation of apical K channels in rat cortical collecting
tubule during changes in dietary K intake. Am. J. Physiol. 277: F805-F812, 1999.
31. Peters, K. W., J. Qi, S. C. Watkins, and R. A. Frizzell. Syntaxin 1A inhibits regulated
CFTR trafficking in Xenopus oocytes. Am. J. Physiol. 277: C174-C180, 1999.
32. Prekeris, R., B. Yang, V. Oorschot, J. Klumperman, and R. H. Scheller. Differential
roles of syntaxin 7 and syntaxin 8 in endosomal trafficking. Mol. Biol. Cell 10: 3891-3908,
1999.
33. Rabouille, C., H. Kondo, R. Newman, N. Hui, P. Freemont, and G. Warren, G.
Syntaxin 5 is a common component of the NSF- and p97-mediated reassembly pathways of
Golgi cisternae from mitotic Golgi fragments in vitro. Cell 92: 603-610, 1998.
34. Robinson, M. S. The role of clathrin, adaptors and dynamin in endocytosis. Curr. Opin.
Cell Biol. 6: 538-544, 1994.

24
35. Rowe, T., C. Dascher, S. Bannykh, H. Plutner, and W. E. Balch. Role of
vesicle-associated syntaxin 5 in the assembly of pre-Golgi intermediates. Science 279: 696-
700, 1998.
36. Saxena, S. S., M. W. Quick, A. Tousson, A., Y. Oh, and D. G. Warnock. Interaction of
syntaxins with the amiloride-sensitive epithelial sodium channel. J. Biol. Chem. 274: 20812-
20817, 1999.
37. Schmid, S. L. Clathrin-coated vesicle formation and protein sorting: An integrated process.
Annu. Rev. Biochem. 66: 511-548, 1997.
38. Shimkets, R. A., R. P. Lifton, R. P., and C. M. Canessa. The activity of the epithelial
sodium channel is regulated by clathrin-mediated endocytosis. J. Biol. Chem. 272: 25537-
25541, 1997.
39. Südhof, T. C. The synaptic vesicle cycle: a cascade of protein-protein interaction. Nature
375: 645-653, 1995.
40. Wang, W. -H., K. M. Lerea, M. Chan, and G. Giebisch. Protein tyrosine kinase regulates
the number of renal secretory channels Am. J. Physiol. 278: F165-F172, 2000.
41. Wei, Y., P. Bloom, R.-M. Gu, and W.-H. Wang. Protein tyrosine phosphatase reduces the
number of apical small-conductance K+ channels in the rat CCD. J. Biol. Chem. 275:
20502-20507, 2000.
42. Wood, S. A., J. E. Park, and W. J. Brown. Brefeldin A causes a microtubule-mediated
fusion of the Trans-Golgi Network and early endosomes. Cell 67: 591-600, 1991.
43. Wright, F. S., and G. Giebisch. Regulation of potassium excretion. The kidney:
Physiology and Pathophysiology, Second Edition, by D. W. Seldin and G. Giebisch.1992;

25
pp 2209-2246.
44. Zhou, H., S. S. Tate, and L. G. Palmer. Primary structure and functional properties of an
epithelial K+ channel. Am. J. Physiol. 266, C809-C824, 1994.

26
FIGURE LEGENDS
Fig. 1. Effect of BFA on ROMK1 channels in oocytes. A, Oocytes were injected with cRNA for
ROMK1 (5 ng). I-V relationships (-100 to +100 mV, in 25 mV steps) were measured by two-
electrode voltage-clamp to monitor expression of ROMK1 currents. After reaching maximal
expression, BFA (5 µM) (-, labeled “+BFA”) or vehicle (-, labeled “-BFA”) was added
and I-V relations were recorded again at indicated time points after addition of BFA (BFA was
added at time 0). Dotted lines and open circles indicate “washout” (see text). Background
currents (after 10 mM Ba2+) account for <5% of total currents at all time points and were
subtracted. Currents ( Ik, whole-cell K+ currents measured at -100 mV) are presented as % of
total currents before addition of BFA. Each data point represents mean ± SEM of currents from
at least 5 oocytes. The half-time (t1/2) for reduction in currents was calculated by fitting data to
the equation ln(xo/xt) = t/t1/2, where xo is 100%, xt is % of currents at time t (38). Similar
findings were observed in 5 additional experiments. B, Cell-attached single-channel recordings
were performed in ROMK1-expressing oocytes before (top trace, “Control”) and after 6 hrs of
BFA treatment (bottom trace, “BFA”). Recording of a single channel in a patch at -100 mV was
shown. “C” indicates current level for closed channel.

27
Fig.2. Effect of co-expression of syntaxin-5 on ROMK1 channels in oocytes. A, Oocytes were
injected (at time 0) with cRNA for ROMK1 (5 ng) and I-V relations were recorded by TEVC.
Seventy two hrs later, oocytes were injected with either cRNA for syntaxin-5 (3 ng, open circles)
or water (closed circles). I-V relations were further recorded over 48 hrs. Ba2+-insensitive
currents were subtracted from the total K+ currents (at -100 mV) to give inward currents (Ik, µA)
as shown. B, Experimental paradigm was as in A except that various concentrations of cRNA for
syntaxin-5 or for syntaxin-3 were injected at time 72 hrs. Maximal ROMK1 currents (at -100
mV) before injection of syntaxin cRNA were presented as 100% of control. Currents after 24 hrs
of injection of syntaxin cRNA were compared to the maximal ROMK1 currents and shown as
“% of control”.

28
Fig. 3. Effect of co-expression of K44A dominant-negative mutant dynamin on ROMK1
channels. A, Inward K+ currents (Ik, µA) were measured by TEVC from oocytes co-expressing
ROMK1 (5 ng cRNA) and K44A mutant dynamin (cRNA 15 ng) or co-expressing ROMK1 (5
ng cRNA) and wild type dynamin (15 ng cRNA) for 72 hrs. Currents shown are mean ±SEM, n
= 5; p<0.01, WT vs. K44A. Similar findings were observed in 3 additional experiments. B, The
effects of BFA on oocytes co-expressing ROMK1 and wild type dynamin (“WT) and oocytes co-
expressing ROMK1 and K44A mutant dynamin (“K44A”) were studied as in Fig. 1A.

29
Fig. 4. NPXY internalization signal in the C-terminus of ROMK1. A, Alignment of a.a. 373-378
of ROMK1 with NPXY consensus motif of LDL receptor, amyloid precursor, and mannose
receptor. Conserved amino acids are boxed. B, Inward K+ currents (Ik, µA) were measured by
TEVC from oocytes after co-injection of cRNAs for N375I or N377T ROMK1 mutant with wild
type dynamin or K44A mutant dynamin for 72 hrs. Currents shown are mean ±SEM, n = 6.
Similar findings were observed in 2 additional experiments. C, Oocytes were injected with
cRNA for N375I or N377T ROMK1 mutant. BFA or vehicle was added 72 hrs after injection.
Inward K+ currents (Ik, µA) were measured 6 hrs after BFA. Currents shown are mean ±SEM, n
= 6. Similar findings were observed in 2 additional experiments.

30
Fig. 5. Endocytosis of ROMK in MDCK cells. A, Left panels: Confocal fluorescent images in
MDCK cells stably transfected with GFP (top panel) or with GFP-tagged ROMK2 (bottom
panel). Confocal images (section along xz plane) were from individual sub-confluent cells grown
on permeable support. GFP-ROMK2 was predominantly targeted to the plasma membrane. In
contrast, GFP was diffusely present in cytoplasm and nuclei. Right panels: Whole-cell K+
currents recorded from dissociated MDCK cells (see “METHODS” for preparation of cells)
stably transfected with GFP (top panel) or with GFP-tagged ROMK2 (bottom panel). Inward-
rectifying K+ currents were observed in cells transfected with GFP-ROMK2 but not in cells
transfected with GFP. The currents were inhibited by 5 mM Ba2+ in the bath (not shown). B, I-V
relationships of whole-cell K+ currents as recorded in panel A. C, Cell-attached single-channel
recording of the GFP-ROMK2 channel in MDCK cells. Recording of a single channel in a patch
at -100 mV was shown. A portion of recording was also shown at an expanded time scale. D,
Effect of BFA on whole-cell ROMK currents. Each column is mean ± SEM (n=10) of inward
whole-cell K+ current (Ik, pA) at -100 mV. MDCK cells were recorded before and 6 hrs after
incubation with BFA (open column) or with vehicle (ethanol, hatched column). N.S. indicates
statistically not significant by unpaired t-test.

31
Fig. 6. Confocal fluorescent images of MDCK cells. A, MDCK cells stably transfected with
GFP-ROMK2. Cells were stained by a monoclonal antibody against clathrin heavy chain,
followed by rabbit anti-mouse IgG, and by rhodamine-conjugated goat anti-rabbit IgG. Images
were visualized with a laser scanning fluorescent microscope with the excitation wavelength
selected for green fluorescent protein (left) or for rhodamine (middle). Merged image is shown in
right panel. B, MDCK cells stably transfected with GFP. Cells were stained as described in panel
A.

32
Fig. 7. Co-immunoprecipitation of ROMK2 with clathrin coat proteins. Solubilized membrane
fractions from MDCK cells stably expressing ROMK2 were immunoprecipitated by monoclonal
antibodies against α-adaptin or control mouse IgG. No antibody was added in the left two lanes
labeled “None”. Immunoprecipitates were separated by 10% SDS-PAGE, transferred to
nitrocellulose membrane, and immunoblotted sequentially using antibodies against α-adaptin,
clathrin heavy chain, and ROMK.
33
A
C
C
Control
BFA (6hrs)2 pA
1s
Tim e (hrs)0 6 12 18 24 48
% o
f cur
rent
bef
ore
BFA
0
50
100
B
I k
Fig 1
-BFA
washout
+BFA
washout

34
Syntaxin cRNA (ng)0 5 10 15 20
0
50
100
Syntaxin 5
Time (hrs)0 24 48 72 96 120
0
5
10
15
20
I k (µ
Α)
I k (%
of c
ontro
l)+ Syntaxin 5
A
BSyntaxin 3
Fig 2
35
0
20
40
60
( µΑ
)
WT K44A
A
Time (hrs)0 6 12 18 24
0
50
100
K44A
WT
I k
B
I k
Fig 3%
bef
ore
BFA
36
AA
0
10
20
30
40
50
I k(µA
)
BN375I
NS
25
50
75
100
N377Tp < 0.01
I k(µA
)
Dynamin WT K44A WT K44A
25
50
75
100
I k(µA
)
C
BFA _ +
N375INS
20
40
60
80
_ +
N377Tp < 0.05
I k(µA
)
Fig. 4
37
4 pA100 ms2 s
C
C
4 pA100 ms2 s4 pA100 ms2 s
C
C
50 100-50-100Vm (mV)
1000
-1000
-2000
I k ( p
A) GFP-ROMK2
GFP Control1000 pA50 ms
500
1000
1500
I k(pA
)
BFA_ +(6 hr) _
p<0.05N.S.
A B
C D
GFP Control
GFP-ROMK2
0 _ (6 hr)_ (6 hr)
Fig 5
38
A
B
AA
B
Figure 6
GFP-ROMK2 Clathrin Merge
GFP Clathrin Merge
39
adaptin
clathrin
ROMK
IP Ab: adap
tin
adap
tin
mo u
se Ig
mou
se Ig
none
none
Fig. 7
Related Documents











