ORIGINAL ARTICLE Amy K. Sater . Heithem M. El-Hodiri . Mousumi Goswami . Tara B. Alexander . Oday Al-Sheikh . Laurence D. Etkin . J. Akif Uzman Evidence for antagonism of BMP-4 signals by MAP kinase during Xenopus axis determination and neural specification Received July 1, 2003; accepted July 28, 2003 Abstract We have previously shown that mitogen- activated protein (MAP) kinase activity is required for neural specification in Xenopus. In mammalian cells, the BMP-4 effector Smad1 is inhibited by phosphorylation at MAP kinase sites (Kretzschmar et al., 1997). To test the hypothesis that MAP kinase inhibits the BMP-4/ Smad1 pathway during early Xenopus development, we have generated a Smad1 mutant lacking the MAP kinase phosphorylation sites (M4A-Smad1) and com- pared the effects of wild-type (WT)- and M4A-Smad1 on axial pattern and neural specification in Xenopus embryos. Although overexpression of either WT- or M4A-Smad1 produced ventralized embryos, at each mRNA concentration, M4A-Smad1 had a greater ventralizing effect than WT-Smad1. Interestingly, over- expression of either form of Smad1 in ventral blas- tomeres disrupted posterior pattern and morphogenesis; again, more severe defects were produced by expression of M4A-Smad1 than by equal amounts of WT-Smad1. Ectodermal expression of M4A-Smad1 disrupted ex- pression of the anterior neural gene otx2 in vivo and inhibited neural specification in response to endogenous signals in mesoderm–ectoderm recombinates. In con- trast, overexpression of WT-Smad1 at identical levels had little effect on either neural specification or otx2 expression. Comparisons of protein levels following overexpression of either WT- or M4A-Smad1 indicate that WT-Smad1 may be slightly more stable than M4A- Smad1; thus, differences in stability cannot account for the increased effectiveness of M4A-Smad1. Our results demonstrate that mutations disrupting the MAPK phosphorylation sites act collectively as a gain-of- function mutation in Smad1 and that inhibitory phosphorylation of Smad1 may be a significant mechanism for the regulation of BMP-4/Smad1 signals during Xenopus development. Key words Smad1 Á MAP kinase Á BMP-4 Á Xenopus Á neural induction Introduction The establishment of neural fate and anteroposterior neural pattern are governed by a diverse array of positive and negative regulatory signals. Foremost among these are bone morphogenetic protein-4 (BMP- 4) and its antagonists noggin (Zimmerman et al., 1996), chordin (Piccolo et al., 1996), follistatin (Iemura et al., 1998), cerberus (Bouwmeester et al., 1996; Hsu et al., 1998), and gremlin (Hsu et al., 1998), which Amy K. Sater ( . *) . Mousumi Goswami . Tara B. Alexander . Oday Al-Sheikh Division of Molecular and Cell Biology Department of Biology and Biochemistry University of Houston Houston, TX 77204, USA Tel: (713) 743-2688, Fax: (713) 743-2636 E-mail: [email protected] Heithem M. El-Hodiri 1 . Laurence D. Etkin Department of Molecular Genetics University of Texas M.D. Anderson Cancer Center Houston, TX 77030, USA J. Akif Uzman Department of Natural Sciences University of Houston-Downtown Houston, TX 77002, USA 1 Present address: Department of Molecular and Human Genetics, Children’s Research Institute, Columbus, OH 43205, USA U.S. Copyright Clearance Center Code Statement: 0301–4681/2003/7107–434 $ 15.00/0 Differentiation (2003) 71: 434–444 r International Society of Differentiation 2003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Amy K. Sater . Heithem M. El-Hodiri .
Mousumi Goswami . Tara B. Alexander .
Oday Al-Sheikh . Laurence D. Etkin .
J. Akif Uzman
Evidence for antagonism of BMP-4 signals by MAP kinase duringXenopus axis determination and neural specification
Received July 1, 2003; accepted July 28, 2003
Abstract We have previously shown that mitogen-activated protein (MAP) kinase activity is required forneural specification in Xenopus. In mammalian cells, theBMP-4 effector Smad1 is inhibited by phosphorylationat MAP kinase sites (Kretzschmar et al., 1997). To testthe hypothesis that MAP kinase inhibits the BMP-4/Smad1 pathway during early Xenopus development, wehave generated a Smad1 mutant lacking the MAPkinase phosphorylation sites (M4A-Smad1) and com-pared the effects of wild-type (WT)- and M4A-Smad1on axial pattern and neural specification in Xenopusembryos. Although overexpression of either WT- orM4A-Smad1 produced ventralized embryos, at eachmRNA concentration, M4A-Smad1 had a greaterventralizing effect than WT-Smad1. Interestingly, over-
expression of either form of Smad1 in ventral blas-tomeres disrupted posterior pattern and morphogenesis;again, more severe defects were produced by expressionof M4A-Smad1 than by equal amounts of WT-Smad1.Ectodermal expression of M4A-Smad1 disrupted ex-pression of the anterior neural gene otx2 in vivo andinhibited neural specification in response to endogenoussignals in mesoderm–ectoderm recombinates. In con-trast, overexpression of WT-Smad1 at identical levelshad little effect on either neural specification or otx2expression. Comparisons of protein levels followingoverexpression of either WT- or M4A-Smad1 indicatethat WT-Smad1 may be slightly more stable than M4A-Smad1; thus, differences in stability cannot account forthe increased effectiveness of M4A-Smad1. Our resultsdemonstrate that mutations disrupting the MAPKphosphorylation sites act collectively as a gain-of-function mutation in Smad1 and that inhibitoryphosphorylation of Smad1 may be a significantmechanism for the regulation of BMP-4/Smad1 signalsduring Xenopus development.
Key words Smad1 � MAP kinase � BMP-4 � Xenopus �neural induction
Introduction
The establishment of neural fate and anteroposteriorneural pattern are governed by a diverse array ofpositive and negative regulatory signals. Foremostamong these are bone morphogenetic protein-4 (BMP-4) and its antagonists noggin (Zimmerman et al.,1996), chordin (Piccolo et al., 1996), follistatin (Iemuraet al., 1998), cerberus (Bouwmeester et al., 1996;Hsu et al., 1998), and gremlin (Hsu et al., 1998), which
Amy K. Sater ( .*) . Mousumi Goswami . Tara B. Alexander .
Oday Al-SheikhDivision of Molecular and Cell BiologyDepartment of Biology and BiochemistryUniversity of HoustonHouston, TX 77204, USATel: (713) 743-2688, Fax: (713) 743-2636E-mail: [email protected]
Heithem M. El-Hodiri1 . Laurence D. EtkinDepartment of Molecular GeneticsUniversity of TexasM.D. Anderson Cancer CenterHouston, TX 77030, USA
J. Akif UzmanDepartment of Natural SciencesUniversity of Houston-DowntownHouston, TX 77002, USA
1Present address: Department of Molecular and HumanGenetics, Children’s Research Institute, Columbus, OH43205, USA
U.S. Copyright Clearance Center Code Statement: 0301–4681/2003/7107–434 $ 15.00/0
Differentiation (2003) 71: 434–444 r International Society of Differentiation 2003

down-regulate BMP-4 signaling by preventing BMP-4from interacting with its receptor. b-catenin alsocontributes to neural specification by repressing BMP-4 transcription (Baker et al., 1999). BMP-4 inducesepidermis and represses neural fate (Wilson andHemmati-Brivanlou, 1995), and antagonism of BMP-4leads to the initiation of anterior neural development(Harland and Gerhart, 1997; Sasai and De Robertis,1997; Wilson and Hemmati-Brivanlou, 1997; reviewedin Weinstein and Hemmati-Brivanlou, 1999).
Fibroblast growth factors (FGFs) have also beenimplicated in neural induction and posterior regionali-zation in amphibian (Kengaku and Okamoto, 1993,1995; Cox and Hemmati-Brivanlou, 1995; Lamb andHarland, 1995) and chick embryos (Streit et al., 2000;Wilson et al., 2000). FGF signals also participate in theestablishment of the ventral forebrain (Lupo et al.,2002), although their role in earlier anterior neuralspecification is in dispute (cf. Holowacz and Sokol,1999; Hongo et al., 1999). Additional posteriorizingsignals include retinoids (Papalopulu et al., 1991; Ruiz iAltaba and Jessell, 1991) and Xwnt3a, which acts inconjunction with FGF to repress anterior neuraldevelopment and initiate expression of posterior neur-al-specific genes (McGrew et al., 1995, 1997). Otherstudies have also implicated wnt/b-catenin signaling inposterior neural patterning (Domingos et al., 2001).
Our previous studies have implicated MAP kinase asa positive regulator of neural fate. First, MAP kinaseactivity is elevated in mid-gastrula neural ectoderm orectoderm overexpressing noggin, relative to uninducedmid-gastrula ectoderm. Second, overexpression of theMAP kinase antagonist MAP kinase phosphatase-1(MKP-1) prevents neural-specific gene expression inresponse to endogenous inductive signals (Uzgare et al.,1998). Although MKP-1 can inhibit activity of theMAP kinase-related proteins Jun N-terminal kinases(JNKs; Liu et al., 1995) and p38 (Mendelson et al.,1996), as well as MAP kinase itself, our results suggestthat MAP kinase participates in neural specification.
MAP kinase can phosphorylate Smad1 at four siteswithin the linker region between the MH1 and MH2domains; once phosphorylated at these sites, Smad1 isunable to enter the nucleus (Kretzschmar et al., 1997).These findings suggest that MAP kinase could promoteneural fate in vivo by phosphorylating and inactivatingSmad1. To evaluate the significance of this inhibitoryphosphorylation on Smad1 function in vivo, wehave generated a mutated form of Smad1 in which allfour consensus MAPK sites have been disrupted. Inthis article, we describe overexpression studies usingthe wild-type and mutated forms of Smad1, whichshow that disruption of the MAPK phosphorylationsites acts as a gain-of-function mutation in Smad1.Our findings indicate that inhibitory phosphorylationplays a significant role in the regulation of Smad1function in vivo.
Methods
Microinjection and microsurgery
Preparation of embryos and microsurgical manipulations wereperformed as described (Uzgare et al., 1998; Uzman et al., 1998).For overexpression studies, synthetic mRNA was prepared by invitro transcription of linearized plasmid DNA, using the AmbionmMessage machine in vitro transcription system. Unless otherwiseindicated, B10 nl of mRNA stock at the appropriate concentrationwas pressure injected near the animal pole into each blastomere atthe two- to four-cell stage. For dorsally or ventrally directedinjections, embryos were injected in the equatorial region of dorsalor ventral blastomeres at the four-cell stage.
Preparation of the M4A-Smad1 construct
The MAP kinase phosphorylation sites (consensus: PXSP) atpositions 187, 195, 205, and 213 were mutated from serine toalanine over two rounds of mutagenesis using the QuickChangeSite Directed Mutagenesis kit (Stratagene, La Jolla, CA; Fig. 1).First, S187 and S195 were mutated by amplification of 5 ng ofpXMad (Thomsen, 1996) according to the manufacturer’s direc-tions. S205 and S213 were mutated in a second set of reactions. Themutagenesis was confirmed by sequencing the mutated construct(University of Texas M.D. Anderson Cancer Center Core Sequen-cing Facility).
Western blots
Immunoblotting was carried out as described in Uzman et al.(1998). For detection of myc-tagged Smad1, isolated animal capswere lysed in kinase buffer (Uzgare et al., 1998) and centrifuged at14,000 rpm at 41C. Supernatants were then mixed with concen-trated SDS sample buffer, boiled, run on 10% SDS-PAGE gels, andtransferred to nitrocellulose. Blots were blocked in 5% milk inTBST (150mM NaCl, 3mM KCl, 20mM Tris [pH 7.4], 0.05%Tween-20) before incubation with anti-myc antibodies (RocheMolecular, Indianapolis, IN), diluted according to the manufac-turer’s instructions. Blots were washed before and after incubationwith secondary antibody; all washes were performed in TBST.Bands were visualized by chemiluminescent detection (ECL).
RT-PCR
RNA isolation, preparation of cDNA, and polymerase chainreaction (PCR) assays were carried out as described (Uzman et al.,1998). PCR primers for Zic3 and N-tubulin were taken from theXenopus Molecular Marker Resource (http://vize222.zo.utexas.edu;accessed June 2003). The RT-PCR products were separated on 6%polyacrylamide gels and visualized by autoradiography.
In situ hybridization and immunohistochemistry
In situ hybridization was carried out as described in Harland (1991),using the modifications suggested by Cox and Hemmati-Brivanlou(1995). Immunohistochemistry was performed as described in Sateret al. (1993), using diaminobenzidine as a substrate.
Results
Antagonism of Smad1 by MAP kinase
Recent work has shown that MAP kinase canphosphorylate Smad1 and prevent it from entering thenucleus (Kretzschmar et al., 1997). Four optimal
435
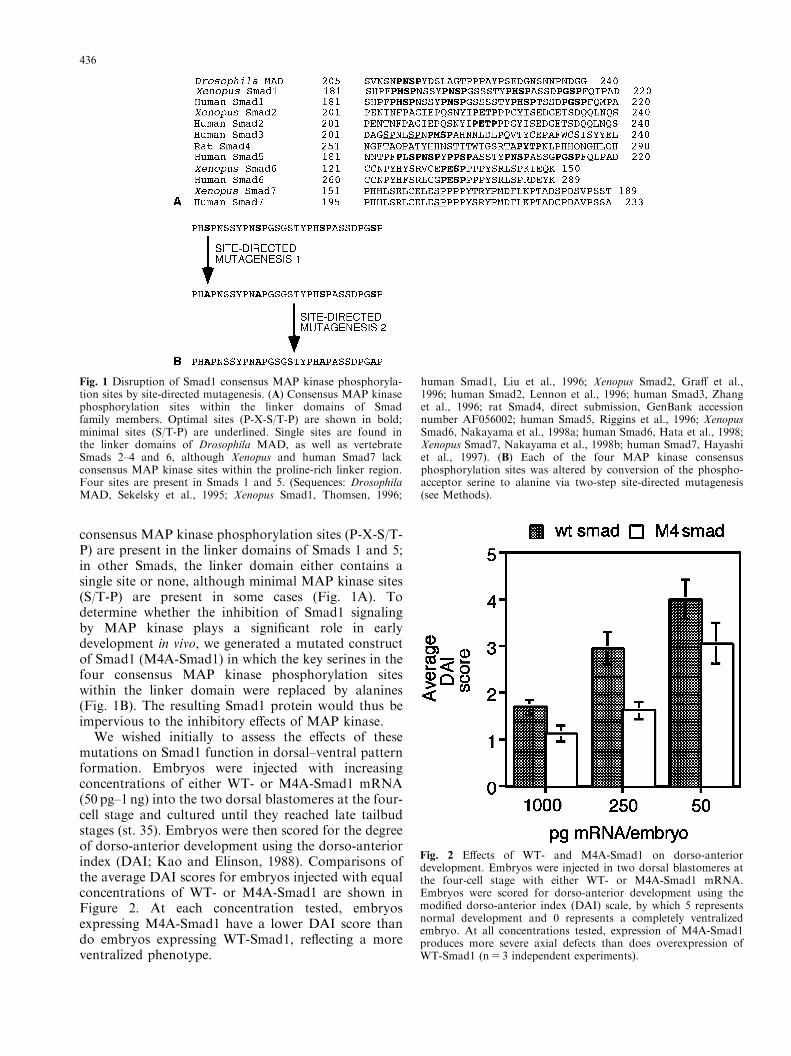
consensus MAP kinase phosphorylation sites (P-X-S/T-P) are present in the linker domains of Smads 1 and 5;in other Smads, the linker domain either contains asingle site or none, although minimal MAP kinase sites(S/T-P) are present in some cases (Fig. 1A). Todetermine whether the inhibition of Smad1 signalingby MAP kinase plays a significant role in earlydevelopment in vivo, we generated a mutated constructof Smad1 (M4A-Smad1) in which the key serines in thefour consensus MAP kinase phosphorylation siteswithin the linker domain were replaced by alanines(Fig. 1B). The resulting Smad1 protein would thus beimpervious to the inhibitory effects of MAP kinase.
We wished initially to assess the effects of thesemutations on Smad1 function in dorsal–ventral patternformation. Embryos were injected with increasingconcentrations of either WT- or M4A-Smad1 mRNA(50 pg–1 ng) into the two dorsal blastomeres at the four-cell stage and cultured until they reached late tailbudstages (st. 35). Embryos were then scored for the degreeof dorso-anterior development using the dorso-anteriorindex (DAI; Kao and Elinson, 1988). Comparisons ofthe average DAI scores for embryos injected with equalconcentrations of WT- or M4A-Smad1 are shown inFigure 2. At each concentration tested, embryosexpressing M4A-Smad1 have a lower DAI score thando embryos expressing WT-Smad1, reflecting a moreventralized phenotype.
Fig. 1 Disruption of Smad1 consensus MAP kinase phosphoryla-tion sites by site-directed mutagenesis. (A) Consensus MAP kinasephosphorylation sites within the linker domains of Smadfamily members. Optimal sites (P-X-S/T-P) are shown in bold;minimal sites (S/T-P) are underlined. Single sites are found inthe linker domains of Drosophila MAD, as well as vertebrateSmads 2–4 and 6, although Xenopus and human Smad7 lackconsensus MAP kinase sites within the proline-rich linker region.Four sites are present in Smads 1 and 5. (Sequences: DrosophilaMAD, Sekelsky et al., 1995; Xenopus Smad1, Thomsen, 1996;
human Smad1, Liu et al., 1996; Xenopus Smad2, Graff et al.,1996; human Smad2, Lennon et al., 1996; human Smad3, Zhanget al., 1996; rat Smad4, direct submission, GenBank accessionnumber AF056002; human Smad5, Riggins et al., 1996; XenopusSmad6, Nakayama et al., 1998a; human Smad6, Hata et al., 1998;Xenopus Smad7, Nakayama et al., 1998b; human Smad7, Hayashiet al., 1997). (B) Each of the four MAP kinase consensusphosphorylation sites was altered by conversion of the phospho-acceptor serine to alanine via two-step site-directed mutagenesis(see Methods).
Fig. 2 Effects of WT- and M4A-Smad1 on dorso-anteriordevelopment. Embryos were injected in two dorsal blastomeres atthe four-cell stage with either WT- or M4A-Smad1 mRNA.Embryos were scored for dorso-anterior development using themodified dorso-anterior index (DAI) scale, by which 5 representsnormal development and 0 represents a completely ventralizedembryo. At all concentrations tested, expression of M4A-Smad1produces more severe axial defects than does overexpression ofWT-Smad1 (n5 3 independent experiments).
436
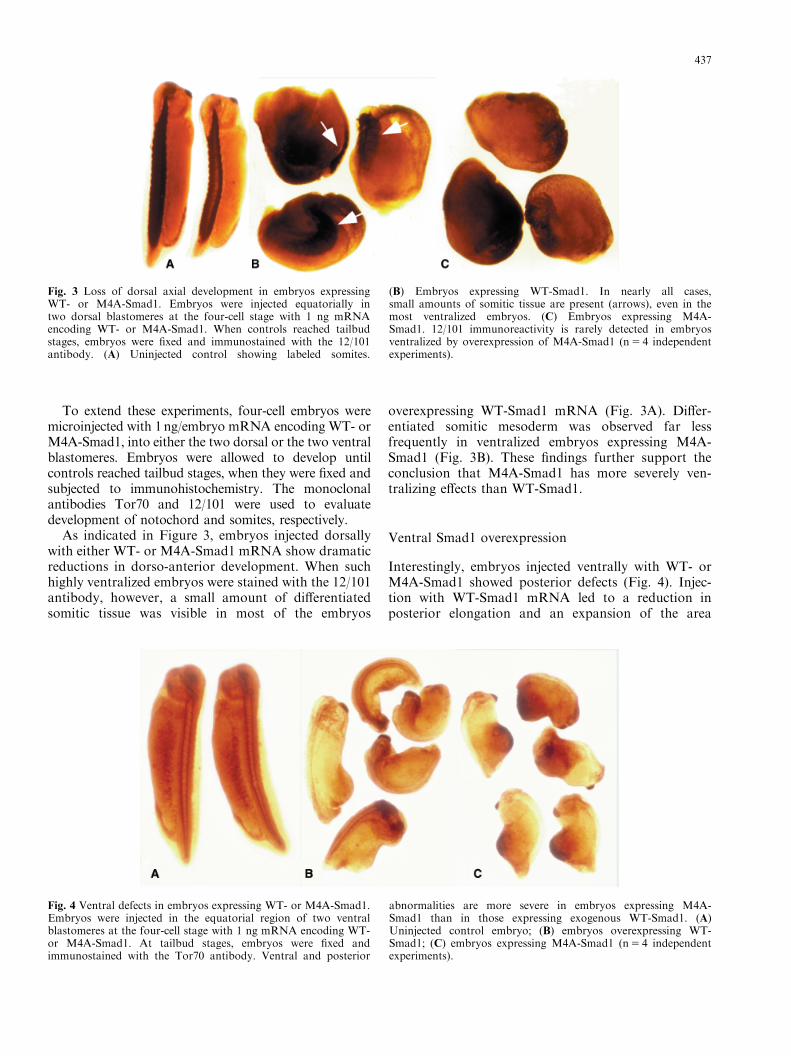
To extend these experiments, four-cell embryos weremicroinjected with 1ng/embryo mRNA encoding WT- orM4A-Smad1, into either the two dorsal or the two ventralblastomeres. Embryos were allowed to develop untilcontrols reached tailbud stages, when they were fixed andsubjected to immunohistochemistry. The monoclonalantibodies Tor70 and 12/101 were used to evaluatedevelopment of notochord and somites, respectively.
As indicated in Figure 3, embryos injected dorsallywith either WT- or M4A-Smad1 mRNA show dramaticreductions in dorso-anterior development. When suchhighly ventralized embryos were stained with the 12/101antibody, however, a small amount of differentiatedsomitic tissue was visible in most of the embryos
overexpressing WT-Smad1 mRNA (Fig. 3A). Differ-entiated somitic mesoderm was observed far lessfrequently in ventralized embryos expressing M4A-Smad1 (Fig. 3B). These findings further support theconclusion that M4A-Smad1 has more severely ven-tralizing effects than WT-Smad1.
Ventral Smad1 overexpression
Interestingly, embryos injected ventrally with WT- orM4A-Smad1 showed posterior defects (Fig. 4). Injec-tion with WT-Smad1 mRNA led to a reduction inposterior elongation and an expansion of the area
Fig. 3 Loss of dorsal axial development in embryos expressingWT- or M4A-Smad1. Embryos were injected equatorially intwo dorsal blastomeres at the four-cell stage with 1 ng mRNAencoding WT- or M4A-Smad1. When controls reached tailbudstages, embryos were fixed and immunostained with the 12/101antibody. (A) Uninjected control showing labeled somites.
(B) Embryos expressing WT-Smad1. In nearly all cases,small amounts of somitic tissue are present (arrows), even in themost ventralized embryos. (C) Embryos expressing M4A-Smad1. 12/101 immunoreactivity is rarely detected in embryosventralized by overexpression of M4A-Smad1 (n5 4 independentexperiments).
Fig. 4 Ventral defects in embryos expressing WT- or M4A-Smad1.Embryos were injected in the equatorial region of two ventralblastomeres at the four-cell stage with 1 ng mRNA encoding WT-or M4A-Smad1. At tailbud stages, embryos were fixed andimmunostained with the Tor70 antibody. Ventral and posterior
abnormalities are more severe in embryos expressing M4A-Smad1 than in those expressing exogenous WT-Smad1. (A)Uninjected control embryo; (B) embryos overexpressing WT-Smad1; (C) embryos expressing M4A-Smad1 (n5 4 independentexperiments).
437
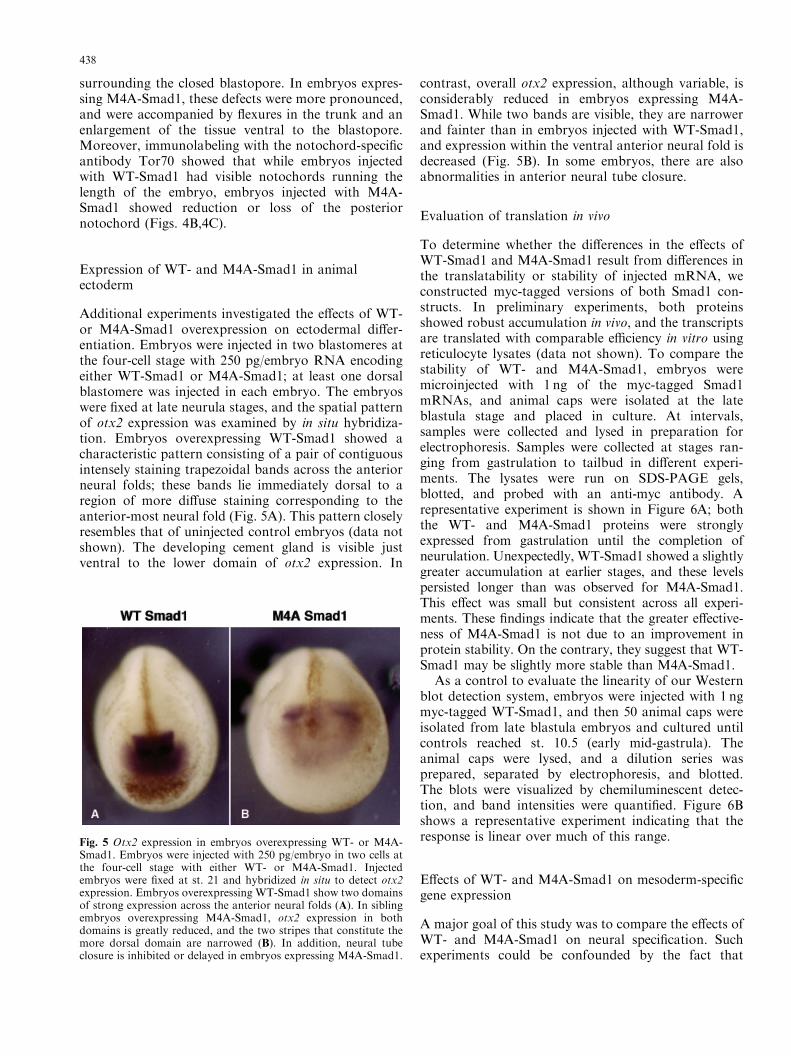
surrounding the closed blastopore. In embryos expres-sing M4A-Smad1, these defects were more pronounced,and were accompanied by flexures in the trunk and anenlargement of the tissue ventral to the blastopore.Moreover, immunolabeling with the notochord-specificantibody Tor70 showed that while embryos injectedwith WT-Smad1 had visible notochords running thelength of the embryo, embryos injected with M4A-Smad1 showed reduction or loss of the posteriornotochord (Figs. 4B,4C).
Expression of WT- and M4A-Smad1 in animalectoderm
Additional experiments investigated the effects of WT-or M4A-Smad1 overexpression on ectodermal differ-entiation. Embryos were injected in two blastomeres atthe four-cell stage with 250 pg/embryo RNA encodingeither WT-Smad1 or M4A-Smad1; at least one dorsalblastomere was injected in each embryo. The embryoswere fixed at late neurula stages, and the spatial patternof otx2 expression was examined by in situ hybridiza-tion. Embryos overexpressing WT-Smad1 showed acharacteristic pattern consisting of a pair of contiguousintensely staining trapezoidal bands across the anteriorneural folds; these bands lie immediately dorsal to aregion of more diffuse staining corresponding to theanterior-most neural fold (Fig. 5A). This pattern closelyresembles that of uninjected control embryos (data notshown). The developing cement gland is visible justventral to the lower domain of otx2 expression. In
contrast, overall otx2 expression, although variable, isconsiderably reduced in embryos expressing M4A-Smad1. While two bands are visible, they are narrowerand fainter than in embryos injected with WT-Smad1,and expression within the ventral anterior neural fold isdecreased (Fig. 5B). In some embryos, there are alsoabnormalities in anterior neural tube closure.
Evaluation of translation in vivo
To determine whether the differences in the effects ofWT-Smad1 and M4A-Smad1 result from differences inthe translatability or stability of injected mRNA, weconstructed myc-tagged versions of both Smad1 con-structs. In preliminary experiments, both proteinsshowed robust accumulation in vivo, and the transcriptsare translated with comparable efficiency in vitro usingreticulocyte lysates (data not shown). To compare thestability of WT- and M4A-Smad1, embryos weremicroinjected with 1 ng of the myc-tagged Smad1mRNAs, and animal caps were isolated at the lateblastula stage and placed in culture. At intervals,samples were collected and lysed in preparation forelectrophoresis. Samples were collected at stages ran-ging from gastrulation to tailbud in different experi-ments. The lysates were run on SDS-PAGE gels,blotted, and probed with an anti-myc antibody. Arepresentative experiment is shown in Figure 6A; boththe WT- and M4A-Smad1 proteins were stronglyexpressed from gastrulation until the completion ofneurulation. Unexpectedly, WT-Smad1 showed a slightlygreater accumulation at earlier stages, and these levelspersisted longer than was observed for M4A-Smad1.This effect was small but consistent across all experi-ments. These findings indicate that the greater effective-ness of M4A-Smad1 is not due to an improvement inprotein stability. On the contrary, they suggest that WT-Smad1 may be slightly more stable than M4A-Smad1.
As a control to evaluate the linearity of our Westernblot detection system, embryos were injected with 1 ngmyc-tagged WT-Smad1, and then 50 animal caps wereisolated from late blastula embryos and cultured untilcontrols reached st. 10.5 (early mid-gastrula). Theanimal caps were lysed, and a dilution series wasprepared, separated by electrophoresis, and blotted.The blots were visualized by chemiluminescent detec-tion, and band intensities were quantified. Figure 6Bshows a representative experiment indicating that theresponse is linear over much of this range.
Effects of WT- and M4A-Smad1 on mesoderm-specificgene expression
A major goal of this study was to compare the effects ofWT- and M4A-Smad1 on neural specification. Suchexperiments could be confounded by the fact that
Fig. 5 Otx2 expression in embryos overexpressing WT- or M4A-Smad1. Embryos were injected with 250 pg/embryo in two cells atthe four-cell stage with either WT- or M4A-Smad1. Injectedembryos were fixed at st. 21 and hybridized in situ to detect otx2expression. Embryos overexpressing WT-Smad1 show two domainsof strong expression across the anterior neural folds (A). In siblingembryos overexpressing M4A-Smad1, otx2 expression in bothdomains is greatly reduced, and the two stripes that constitute themore dorsal domain are narrowed (B). In addition, neural tubeclosure is inhibited or delayed in embryos expressing M4A-Smad1.
438
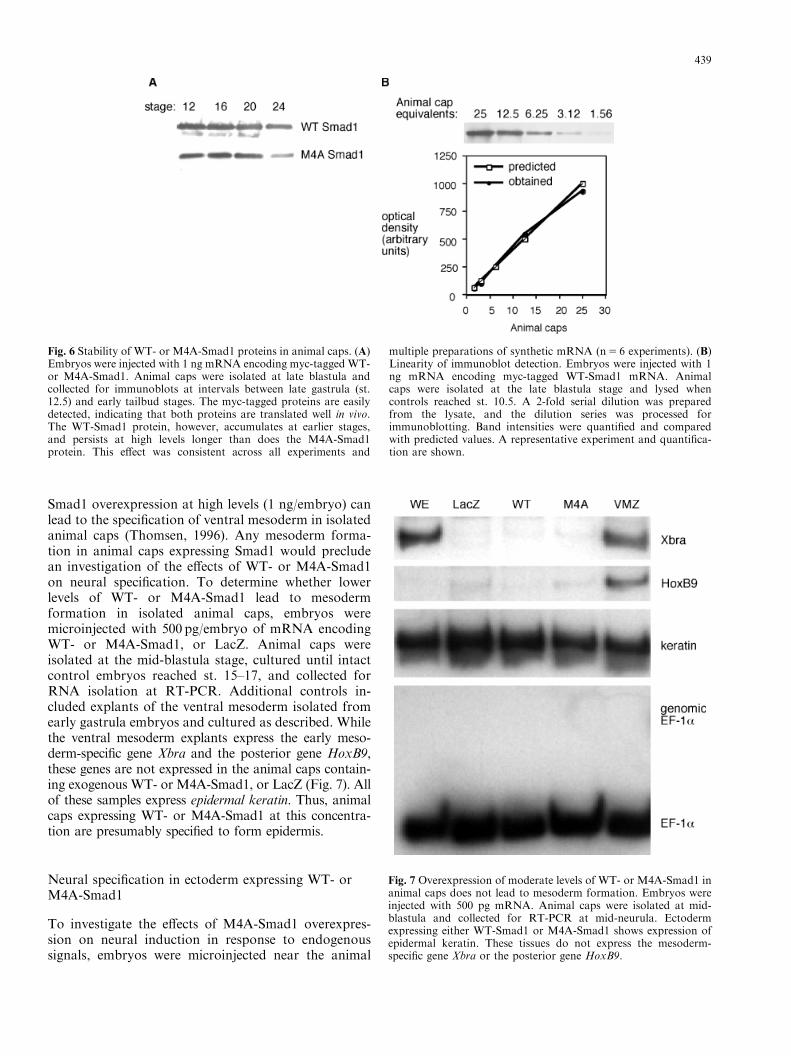
Smad1 overexpression at high levels (1 ng/embryo) canlead to the specification of ventral mesoderm in isolatedanimal caps (Thomsen, 1996). Any mesoderm forma-tion in animal caps expressing Smad1 would precludean investigation of the effects of WT- or M4A-Smad1on neural specification. To determine whether lowerlevels of WT- or M4A-Smad1 lead to mesodermformation in isolated animal caps, embryos weremicroinjected with 500 pg/embryo of mRNA encodingWT- or M4A-Smad1, or LacZ. Animal caps wereisolated at the mid-blastula stage, cultured until intactcontrol embryos reached st. 15–17, and collected forRNA isolation at RT-PCR. Additional controls in-cluded explants of the ventral mesoderm isolated fromearly gastrula embryos and cultured as described. Whilethe ventral mesoderm explants express the early meso-derm-specific gene Xbra and the posterior gene HoxB9,these genes are not expressed in the animal caps contain-ing exogenous WT- or M4A-Smad1, or LacZ (Fig. 7). Allof these samples express epidermal keratin. Thus, animalcaps expressing WT- or M4A-Smad1 at this concentra-tion are presumably specified to form epidermis.
Neural specification in ectoderm expressing WT- orM4A-Smad1
To investigate the effects of M4A-Smad1 overexpres-sion on neural induction in response to endogenoussignals, embryos were microinjected near the animal
Fig. 6 Stability of WT- or M4A-Smad1 proteins in animal caps. (A)Embryos were injected with 1 ng mRNA encoding myc-tagged WT-or M4A-Smad1. Animal caps were isolated at late blastula andcollected for immunoblots at intervals between late gastrula (st.12.5) and early tailbud stages. The myc-tagged proteins are easilydetected, indicating that both proteins are translated well in vivo.The WT-Smad1 protein, however, accumulates at earlier stages,and persists at high levels longer than does the M4A-Smad1protein. This effect was consistent across all experiments and
multiple preparations of synthetic mRNA (n5 6 experiments). (B)Linearity of immunoblot detection. Embryos were injected with 1ng mRNA encoding myc-tagged WT-Smad1 mRNA. Animalcaps were isolated at the late blastula stage and lysed whencontrols reached st. 10.5. A 2-fold serial dilution was preparedfrom the lysate, and the dilution series was processed forimmunoblotting. Band intensities were quantified and comparedwith predicted values. A representative experiment and quantifica-tion are shown.
Fig. 7 Overexpression of moderate levels of WT- or M4A-Smad1 inanimal caps does not lead to mesoderm formation. Embryos wereinjected with 500 pg mRNA. Animal caps were isolated at mid-blastula and collected for RT-PCR at mid-neurula. Ectodermexpressing either WT-Smad1 or M4A-Smad1 shows expression ofepidermal keratin. These tissues do not express the mesoderm-specific gene Xbra or the posterior gene HoxB9.
439
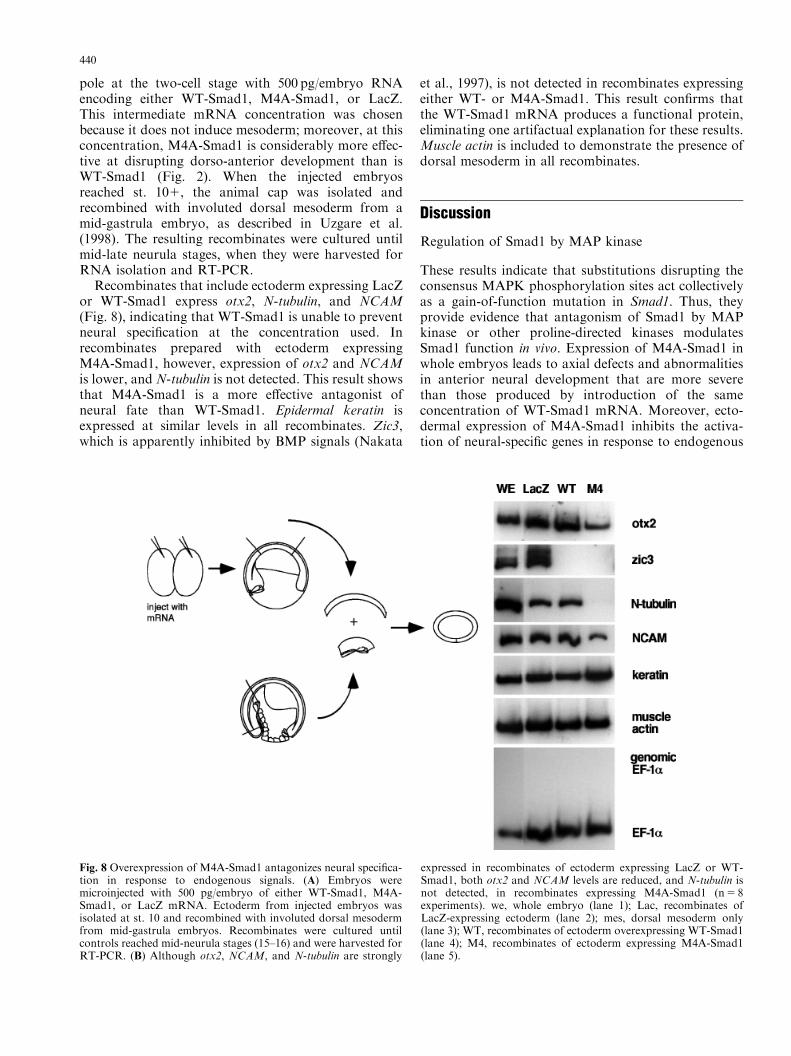
pole at the two-cell stage with 500 pg/embryo RNAencoding either WT-Smad1, M4A-Smad1, or LacZ.This intermediate mRNA concentration was chosenbecause it does not induce mesoderm; moreover, at thisconcentration, M4A-Smad1 is considerably more effec-tive at disrupting dorso-anterior development than isWT-Smad1 (Fig. 2). When the injected embryosreached st. 101, the animal cap was isolated andrecombined with involuted dorsal mesoderm from amid-gastrula embryo, as described in Uzgare et al.(1998). The resulting recombinates were cultured untilmid-late neurula stages, when they were harvested forRNA isolation and RT-PCR.
Recombinates that include ectoderm expressing LacZor WT-Smad1 express otx2, N-tubulin, and NCAM(Fig. 8), indicating that WT-Smad1 is unable to preventneural specification at the concentration used. Inrecombinates prepared with ectoderm expressingM4A-Smad1, however, expression of otx2 and NCAMis lower, and N-tubulin is not detected. This result showsthat M4A-Smad1 is a more effective antagonist ofneural fate than WT-Smad1. Epidermal keratin isexpressed at similar levels in all recombinates. Zic3,which is apparently inhibited by BMP signals (Nakata
et al., 1997), is not detected in recombinates expressingeither WT- or M4A-Smad1. This result confirms thatthe WT-Smad1 mRNA produces a functional protein,eliminating one artifactual explanation for these results.Muscle actin is included to demonstrate the presence ofdorsal mesoderm in all recombinates.
Discussion
Regulation of Smad1 by MAP kinase
These results indicate that substitutions disrupting theconsensus MAPK phosphorylation sites act collectivelyas a gain-of-function mutation in Smad1. Thus, theyprovide evidence that antagonism of Smad1 by MAPkinase or other proline-directed kinases modulatesSmad1 function in vivo. Expression of M4A-Smad1 inwhole embryos leads to axial defects and abnormalitiesin anterior neural development that are more severethan those produced by introduction of the sameconcentration of WT-Smad1 mRNA. Moreover, ecto-dermal expression of M4A-Smad1 inhibits the activa-tion of neural-specific genes in response to endogenous
Fig. 8 Overexpression of M4A-Smad1 antagonizes neural specifica-tion in response to endogenous signals. (A) Embryos weremicroinjected with 500 pg/embryo of either WT-Smad1, M4A-Smad1, or LacZ mRNA. Ectoderm from injected embryos wasisolated at st. 10 and recombined with involuted dorsal mesodermfrom mid-gastrula embryos. Recombinates were cultured untilcontrols reached mid-neurula stages (15–16) and were harvested forRT-PCR. (B) Although otx2, NCAM, and N-tubulin are strongly
expressed in recombinates of ectoderm expressing LacZ or WT-Smad1, both otx2 and NCAM levels are reduced, and N-tubulin isnot detected, in recombinates expressing M4A-Smad1 (n5 8experiments). we, whole embryo (lane 1); Lac, recombinates ofLacZ-expressing ectoderm (lane 2); mes, dorsal mesoderm only(lane 3); WT, recombinates of ectoderm overexpressing WT-Smad1(lane 4); M4, recombinates of ectoderm expressing M4A-Smad1(lane 5).
440

signals in mesoderm–ectoderm recombinates. In con-trast, neural induction in recombinates containingectoderm expressing similar levels of WT-Smad1 islargely indistinguishable from that of control recombi-nates. While inhibition of Smad function via inhibitoryphosphorylation by MAP kinase has been demon-strated in mammalian cultured cells (Kretzschmar et al.,1997, 1999), our findings demonstrate that this regula-tory mechanism has functional significance duringvertebrate development in vivo.
We have previously shown that MAP kinase activityis considerably higher in mid-gastrula neural ectodermthan in uninduced ectoderm at the same stage, and thatneural specification is blocked by overexpression of theMAP kinase antagonist MKP-1 (Uzgare et al., 1998).Studies in progress indicate that FGF inhibits expres-sion of several BMP-4/Smad1-inducible genes (H. Gohiland A.K.S., unpublished results). In gastrula embryos,the spatial distributions of activated (diphospho-) MAPkinase and activated (phospho-) Smad1 are partiallycomplementary (Schohl and Fagotto, 2002). BecauseMAP kinase is a principal effector of FGF signaling, ourresults suggest that inhibition of Smad1 may be one majorfunction of MAP kinase during neural specification.
While overexpression of M4A-Smad1 producesgreater effects than an equivalent amount of WT-Smad1, the difference is largely quantitative, rather thanqualitative. The absence of a dramatic qualitativephenotype is in keeping with the nature of thesemutations. Instead of producing a constitutively acti-vated protein, they result in a protein that is imperviousto one mechanism of negative regulation. Activationstill requires phosphorylation at C-terminal sites by theBMP-4 receptor complex, and Smad1 activity can stillbe inhibited by dephosphorylation at these sites. Earlierstudies suggest that Smad1 is protected from MAPKphosphorylation by interactions with calmodulin-dependent kinase (CaM kinase), and that phosphoryla-tion at CaM kinase sites promotes Smad1 activity(Scherer and Graff, 2000). However, the quantitativeenhancement of Smad1 activity by mutations at theMAPK sites suggests that phosphorylation by MAPkinase or other proline-directed kinases plays an im-portant role in the modulation of Smad1 activity in vivo.
Our results suggest that these mutations do notincrease apparent Smad1 activity by increasing Smad1stability. Moreover, they suggest that WT-Smad1 maybe slightly more stable than the M4A-Smad1 protein.Any effects of these mutations on Smad1 stability maybe mediated by their effects on Smad1 localization.Previous studies (Zhu et al., 1999; Zhang et al., 2001)have identified two ubiquitin ligases (Smad1 ubiquitina-tion recognition factors1 and 2; Smurf1, Smurf2) thatrecognize a PPXY sequence within the Smad1 linkerdomain near the cluster of consensus MAPK phospho-rylation sites (PXSP). Interactions between Smad1 andSmurf1 or Smurf2 initiate ubiquitin-mediated degrada-
tion and result in a loss of steady-state levels of Smad1in Xenopus embryos. Smurf-mediated ubiquitination ofSmads occurs following translocation to the nucleus(Lo and Massague, 1999). Phosphorylation by MAPKis thought to prevent Smad1 accumulation in thenucleus (Kretzschmar et al., 1997). Thus, if the loss ofsensitivity to MAPK promotes nuclear accumulation ofSmad1, interactions with Smurf1 resulting in ubiquitin-mediated proteolysis might be enhanced as a result.
Inhibition of BMP-4 signals
Modulation of BMP-4 signals can occur at severalpoints along the signaling pathway (reviewed in vonBubnoff and Cho, 2001). As mentioned earlier, theextracellular BMP-4 antagonists can sequester BMP-4within the extracellular space and prevent it frominteracting with the BMP-4 receptor. In Xenopusembryos, the BMP-4 antagonist chordin is inactivatedby Xolloid, an extracellular metalloprotease (Piccoloet al., 1997). Intracellularly, BMP-4 signals are down-regulated by the inhibitory Smads, Smad6 and Smad7.Smad7 competes with Smad1 for binding to the activesite of the BMP-4 type I receptor (Nakao et al., 1997;Casellas and Brivanlou, 1998). Smad6 binds Smad1 andprevents it from interacting with Smad4 (Hata et al.,1998; Nakayama et al., 1998a; Christian and Nakayama,1999).
Studies showing that Smad1 can be inhibited byphosphorylation at MAPK sites within the linkerdomain reveal another mechanism by which BMP-4signals can be down-regulated (Kretzschmar et al.,1997). In contrast to the extracellular BMP-4 antago-nists, regulation of Smad1 activation or functionprovides a way to modulate Smad1 activity withoutdirectly altering the activity of TGF-b activated kinase(TAK1), the second major effector downstream of theBMP-4 receptor (Shibuya et al., 1998).
Antagonism of BMP-4 signaling during neuralspecification in Xenopus may involve each of thesemechanisms. The extracellular antagonists noggin andchordin were first identified as candidate neuralinducers, and their role in the establishment ofamphibian neural fate is undisputed. Moreover, nogginis expressed in the anterior neural ectoderm in mid-gastrula embryos (Vodicka and Gerhart, 1995; Uzmanet al., 1998). Both Smad6 (Nakayama et al., 1998a) andSmad7 (Casellas and Brivanlou, 1998) are expressedmaternally, and thus they should be present in thedorsal ectoderm, where they might modulate BMP-4/Smad1 signals. Our earlier studies have shown thatMAPK activity in mid-gastrula neural ectoderm is 5-fold higher than that of uninduced ectoderm at the samestage. Thus, MAPK could down-regulate Smad1activity via inhibitory phosphorylation in newly in-duced neural ectoderm.
441

Smad1 and the coordination of gastrulation
Ventral expression of either WT- or M4A-Smad1resulted in ventral and posterior abnormalities. Thisresult is surprising, given that BMP-4 is expressed in theventral mesoderm, and thus Smad1 should be active onthe ventral side. These defects presumably arise fromabnormalities in the coordination of gastrulation move-ments. Normally, ventral mesoderm cells begin conver-gence very late in gastrulation, sweeping around theblastopore during neurula stages to form tailbud meso-derm (Keller et al., 1991). Failure of ventral mesodermcells to initiate convergence and extension could lead toa reduction in posterior axial mesoderm and a retentionof mesoderm ventral to the blastopore, as revealed bythe distinctive morphologies of the embryos expressingWT- or M4A-Smad1 on the ventral side. Our resultsinvite the speculation that the BMP-4/Smad1 pathwaymay contribute to the coordination of cell behaviorduring gastrulation by keeping cell movements in checkin the lateral and ventral marginal zone. At the onset ofgastrulation, BMP-4 signals are presumably active inthe marginal zone outside the organizer. The medio-lateral intercalation behavior responsible for conver-gence and extension is initiated within the dorsalmarginal zone and spreads through the lateral marginalzone over the course of gastrulation. Chordin antag-onizes BMP-4 signals within the lateral marginal zonein a dorsal-to-ventral progression, and this sequentialinactivation of BMP-4 signals could play a role in thesequential initiation of cell behavior during gastrulation.
Interactions between MAP kinase and BMP-4pathways
Sequence comparisons of Smad family members showthat only Smads 1 and 5, the transducers of BMP-2/4/7signals, have four optimal MAP kinase consensusphosphorylation sites (P-X-S/T-P; Fig 1A). Althougha recent study has shown that Smad2 and Smad3 can beinhibited via phosphorylation at minimal MAP kinasephosphorylation sites (S/T-P; Kretzschmar et al., 1999),Smad1 and Smad5 may be more sensitive to inhibitionat physiological levels of MAP kinase activity. Further-more, a recent study of TGF-b signaling in MDCK cellsfound that activation of MAPK had no significant effecton Smad2/3 function (Lehmann et al., 2000). Thus, wespeculate that among TGF-b family members, antag-onism by MAP kinase is most significant for membersthat signal through Smad1 and Smad5, such as BMP-4.
We have previously shown that overexpression of theBMP-4 antagonist noggin leads to a 5-fold increase inMAP kinase activity in Xenopus ectoderm (Uzgare et al.,1998). This result suggests that signaling through theBMP-4 receptor inhibits MAP kinase activity, which wehave confirmed in subsequent experiments (Goswamiet al., 2001). Thus, the MAP kinase and BMP-4
pathways are mutually antagonistic in Xenopus ecto-derm. FGF is a principal activator of MAP kinase.Therefore, antagonistic interactions between BMP-4pathway members and MAP kinase may play a criticalrole in many developmental processes in which FGFsand BMPs have antagonistic effects. Such processesinclude the control of proliferation in the vertebratelimb (Niswander and Martin, 1993), the regulation ofapoptosis in the embryonic heart (Zhao and Rivkees,2000), and the positioning of feather germs (Jung et al.,1998) and tooth primordia (Neubuser et al., 1997).
Acknowledgments We wish to thank Naoto Ueno, AtasushiSuzuki, Gerry Thomsen, and Ken Cho for cDNA constructs;Richard Harland for Tor70 monoclonal antibody; and JohnWallingford for comments on the manuscript. The 12/101 mono-clonal antibody developed by J.P. Brockes was obtained fromthe Developmental Studies Hybridoma Bank maintained by theDepartment of Biological Sciences, University of Iowa, underthe auspices of the National Institute of Child Health and HumanDevelopment. H.M.E.-H. was supported by an NIH training grantto the Department of Molecular Genetics, M.D. Anderson CancerCenter, University of Texas. The U.T.M.D.A.C.C. Core Sequen-cing Facility was supported by NIH core grant # CA 16672. Thiswork was supported by NSF grant IBN-9723183 to A.K.S.
References
Baker, J.C., Beddington, R.S. and Harland, R.M. (1999) Wntsignaling in Xenopus embryos inhibits bmp4 expression andactivates neural development. Genes Dev 13:3149–3159.
Bouwmeester, T., Kim, S., Sasai, Y., Lu, B. and De Robertis, E.M.(1996) Cerberus is a head-inducing secreted factor expressed inthe anterior endoderm of Spemann’s organizer. Nature 382:595–601.
Casellas, R. and Brivanlou, A.H. (1998) Xenopus Smad7 inhibitsboth the activin and BMP pathways and acts as a neural inducer.Dev Biol 198:1–12.
Christian, J.L. and Nakayama, T. (1999) Can’t get no SMADisfac-tion: Smad proteins as positive and negative regulators of TGF-bfamily signals. Bioessays 21:382–390.
Cox, W.G. and Hemmati-Brivanlou, A. (1995) Caudalization ofneural fate by tissue recombination and bFGF. Development121:4349–4358.
Domingos, P.M., Itasaki, N., Jones, C.M., Mercurio, S., Sargent,M.G., Smith, J.C. and Krumlauf, R. (2001) The Wnt/b-cateninpathway posteriorizes neural tissue in Xenopus by an indirectmechanism requiring FGF signalling. Dev Biol 239:148–160.
Goswami, M., Uzgare, A.R. and Sater, A.K. (2001) Regulation ofMAP kinase by the BMP-4/TAK1 pathway in Xenopusectoderm. Dev Biol 236:259–270.
Graff, J.M., Bansal, A. and Melton, D.A. (1996) Xenopus Madproteins transduce distinct subsets of signals for the TGF-bsuperfamily. Cell 85:479–487.
Harland, R.M. (1991) In situ hybridization: an improved whole-mount method for Xenopus embryos. In: Kay, B. and Peng, B.(eds) Xenopus laevis: practical uses in cell and molecular biology.Methods in cell biology, vol. 36. Academic Press, San Diego, pp685–695.
Harland, R. and Gerhart, J. (1997) Formation and function ofSpemann’s organizer. Annu Rev Cell Dev Biol 11:611–667.
Hata, A., Lagna, G., Massague, J. and Hemmati-Brivanlou, A. (1998)Smad6 inhibits BMP/Smad1 signaling by specifically competingwith the Smad4 tumor suppressor. Genes Dev 12:186–197.
Hayashi, H., Abdollah, S., Qui, Y., Xu, Y.-Y., Grinnell, B.W.,Richardson, M.A., Topper, J.N., Gimbrone, M.A. Jr., Wrana,
442

J.L. and Falb, D. (1997) The MAD-related protein, Smad7,associates with the TGF-b receptor and functions as anantagonist of TGF-b signaling. Cell 89:1165–1173.
Holowacz, T. and Sokol, S. (1999) FGF is required for posteriorneural patterning but not for neural induction. Dev Biol 205:296–308.
Hongo, I., Kengaku, M. and Okamoto, H. (1999) FGF signalingand the anterior neural induction in Xenopus. Dev Biol 216:561–581.
Hsu, D.R., Economides, A.N., Wang, X., Eimon, P.M. andHarland, R.M. (1998) The Xenopus dorsalizing factor Gremlinidentifies a novel family of secreted proteins that antagonizeBMP activities. Mol Cell 1:673–683.
Iemura, S., Yamamoto, T.S., Takagi, C., Uchiyama, H., Natsume,T., Shimasaki, S., Sugino, H. and Ueno, N. (1998) Direct bindingof follistatin to a complex of bone morphogenetic protein and itsreceptor inhibits ventral and epidermal cell fates in the earlyXenopus embryo. Proc Natl Acad Sci USA 95:9337–9342.
Jung, H.S., Francis-West, P.H., Widelitz, R.B., Jiang, T.X.,Ting-Berreth, S., Tickle, C., Wolpert, L. and Chuong, C.M.(1998) Local inhibitory action of BMPs and their relationshipswith activators in feather formation: implications for periodicpatterning. Dev Biol 196:11–23.
Kao, K.R. and Elinson, R.P. (1988) The entire mesodermal mantlebehaves as Spemann’s organizer in dorsoanterior enhancedXenopus laevis embryos. Dev Biol 127:64–77.
Keller, R., Shih, J., Wilson, P. and Sater, A. (1991) Patterns of cellmotility, cell interactions, and mechanism during convergenceand extension in Xenopus. In: Gerhart, J. (ed.) Cell-cellinteractions in early development. 49th Symposium, Society forDevelopmental Biology. Alan R. Liss, Inc., New York.
Kengaku, M. and Okamoto, H. (1993) Basic fibroblast growthfactor induces differentiation of neural tube and neural crestlineages of cultured ectoderm cells from Xenopus gastrula.Development 119:1067–1078.
Kengaku, M. and Okamoto, H. (1995) bFGF as a possiblemorphogen for the antero-posterior axis of the central nervoussystem in Xenopus. Development 121:3121–3130.
Kretzschmar, M., Doody, J. and Massague, J. (1997) OpposingBMP and EGF signaling pathways converge on the TGF-bfamily mediator Smad1. Nature 389:618–622.
Kretzschmar, M., Doody, J., Timokhina, I. and Massague, J.(1999) A mechanism of repression of TGFb/Smad signaling byoncogenic Ras. Genes Dev 13:804–816.
Lamb, T.M. and Harland, R.M. (1995) Fibroblast growth factor isa direct neural inducer, which combined with noggin generatesanterior-posterior neural pattern. Development 121:3627–3636.
Lehmann, K., Janda, E., Pierreux, C.E., Rytomaa, M., Schulze, A.,McMahon, M., Hill, C.S., Beug, H. and Downward, J. (2000)Raf induces TGFb production while blocking its apoptotic butnot invasive responses: a mechanism leading to increasedmalignancy in epithelial cells. Genes Dev 14:2610–2622.
Lennon, G., Auffray, C., Polymeropoulos, M. and Soares, M.B.(1996) The I.M.A.G.E. consortium: an integrated molecularanalysis of genomes and their expression. Genomics 33:151–152.
Liu, Y., Gorospe, M., Yang, C. and Holbrook, N.J. (1995) Role ofmitogen-activated protein kinase phosphatase during the cellularresponse to genotoxic stress. Inhibition of c-Jun N-terminalkinase activity and AP-1-dependent gene activation. J Biol Chem270:8377–8380.
Liu, F., Hata, A., Baker, J.C., Doody, J., Carcamo, J., Harland,R.M. and Massague, J. (1996) A human Mad protein acting as aBMP-regulated transcriptional activator. Nature 381:620–623.
Lo, R.S. and Massague, J. (1999) Ubiquitin-dependent degradationof TGF-b-activated smad2. Nat Cell Biol 1:472–478.
Lupo, G., Harris, W.A., Barsacchi, G. and Vignali, R. (2002)Induction and patterning of the telencephalon in Xenopus laevis.Development 129:5421–5436.
McGrew, L.L., Lai, C.J. and Moon, R.T. (1995) Specification ofthe anteroposterior neural axis through synergistic interaction of
the Wnt signaling cascade with noggin and follistatin. Dev Biol172:337–342.
McGrew, L.L., Hoppler, S. and Moon, R.T. (1997) Wnt and FGFpathways cooperatively pattern anteroposterior neural ectodermin Xenopus. Mech Dev 69:105–114.
Mendelson, K.G., Contois, L.R., Tevosian, S.G., Davis, R.J. andPaulson, K.E. (1996) Independent regulation of JNK/p38mitogen-activated protein kinases by metabolic oxidative stressin the liver. Proc Natl Acad Sci USA 93:12908–12913.
Nakao, A., Afrakhte, M., Moren, A., Nakayama, T., Christian,J.L., Heuchel, R., Itoh, S., Kawabata, M., Heldin, N.E.,Heldin, C.H. and ten Dijke, P. (1997) Identification of Smad7,a TGFb-inducible antagonist of TGF-b signalling. Nature 389:631–635.
Nakata, K., Nagai, T., Aruga, J. and Mikoshiba, K. (1997)Xenopus Zic3, a primary regulator both in neural and neural crestdevelopment. Proc Natl Acad Sci USA 94:11980–11985.
Nakayama, A., Gardner, H., Berg, L.K. and Christian, J.L. (1998a)Smad6 functions as an intracellular antagonist of some TGF-bfamily members during Xenopus embryogenesis. Genes Cells3:387–394.
Nakayama, A., Snyder, M.A., Grewal, S.S., Tsuneizumi, K.,Tabata, T. and Christian, J.L. (1998b) Xenopus Smad8 actsdownstream of BMP-4 to modulate its activity during vertebrateembryonic patterning. Development 125:857–867.
Neubuser, A., Peters, H., Balling, R. and Martin, G.R. (1997)Antagonistic interactions between FGF and BMP signalingpathways: a mechanism for positioning the sites of toothformation. Cell 90:247–255.
Niswander, L. and Martin, G.R. (1993) FGF-4 and BMP-2 haveopposite effects on limb growth. Nature 361:68–71.
Papalopulu, N., Clarke, J.D., Bradley, L., Wilkinson, D.,Krumlauf, R. and Holder, N. (1991) Retinoic acid causesabnormal development and segmental patterning of the anteriorhindbrain in Xenopus embryos. Development 113:1145–1158.
Piccolo, S., Sasai, Y., Lu, B. and De Robertis, E.M. (1996)Dorsoventral patterning in Xenopus: inhibition of ventral signalsby direct binding of chordin to BMP-4. Cell 86:589–598.
Piccolo, S., Agius, E., Lu, B., Goodman, S., Dale, L. and DeRobertis, E.M. (1997) Cleavage of Chordin by Xolloid metallo-protease suggests a role for proteolytic processing in theregulation of Spemann organizer activity. Cell 91:407–416.
Riggins, G.J., Thiagalingam, S., Rosenblum, E., Weinstein, C.L.,Kern, S.E., Hamilton, S.R., Willson, J.K., Markowitz, S.D.,Kinzler, K.W. and Vogelstein, B.V. (1996) Mad-related genes inthe human. Nat Genet 13:347–348.
Ruiz i Altaba, A. and Jessell, T.M. (1991) Retinoic acid modifiesthe pattern of cell differentiation in the central nervous system ofneurula stage Xenopus embryos. Development 112:945–958.
Sasai, Y. and De Robertis, E.M. (1997) Ectodermal patterning invertebrate embryos. Dev Biol 182:5–20.
Sater, A.K., Steinhardt, R.A. and Keller, R. (1993) Induction ofneuronal differentiation by planar signals in Xenopus embryos.Dev Dyn 197:268–280.
Scherer, A. and Graff, J.M. (2000) Calmodulin differentiallymodulates Smad1 and Smad2 signaling. J Biol Chem275:41430–41438.
Schohl, A. and Fagotto, F. (2002) b-catenin, MAPK and Smad sig-naling during early Xenopus development. Development 129:37–52.
Sekelsky, J.J., Newfeld, S.J., Raftery, L.A., Chartoff, E.H. andGelbart, W.M. (1995) Genetic characterization and cloning ofmothers against dpp, a gene required for decapentaplegicfunction in Drosophila melanogaster. Genetics 139:1347–1358.
Shibuya, H., Iwata, H., Masuyama, N., Gotoh, Y., Yamaguchi, K.,Irie, K., Matsumoto, K., Nishida, E. and Ueno, N. (1998) Roleof TAK1 and TAB1 in BMP signaling in early Xenopusdevelopment. EMBO J 17:1019–1028.
Streit, A., Berliner, A.J., Papanayotou, C., Sirulnik, A. and Stern,C.D. (2000) Initiation of neural induction by FGF signallingbefore gastrulation. Nature 406:74–78.
443

Thomsen, G.H. (1996) Xenopus mothers against decapentaplegic isan embryonic ventralizing agent that acts downstream of theBMP-2/4 receptor. Development 122:2359–2366.
Uzgare, A.R., Uzman, J.A., El-Hodiri, H.M. and Sater, A.K.(1998) MAP kinase and neural specification in Xenopus. ProcNatl Acad Sci USA 95:14833–14838.
Uzman, J.A., Patil, S., Uzgare, A.R. and Sater, A.K. (1998) Therole of intracellular alkalinization in the establishment of anteriorneural fate in Xenopus. Dev Biol 193:10–20.
Vodicka, M.A. and Gerhart, J.C. (1995) Blastomere derivation anddomains of gene expression in the Spemann Organizer ofXenopus laevis. Development 121:3505–3518.
von Bubnoff, A. and Cho, K.W. (2001) Intracellular BMP signalingregulation in vertebrates: pathway or network? Dev Biol 239:1–14.
Weinstein, D.C. and Hemmati-Brivanlou, A. (1999) Neuralinduction. Ann Rev Cell Dev Biol 15:411–433.
Wilson, P.A. and Hemmati-Brivanlou, A. (1995) Induction of epidermisand inhibition of neural fate by Bmp-4. Nature 376:331–333.
Wilson, P.A. and Hemmati-Brivanlou, A. (1997) Vertebrate neuralinduction: inducers, inhibitors, and a new synthesis. Neuron18:699–710.
Wilson, S.I., Graziano, E., Harland, R., Jessell, T.M. and Edlund,T. (2000) An early requirement for FGF signalling in theacquisition of neural cell fate in the chick embryo. Curr Biol10:421–429.
Zhang, Y., Feng, X.-H., Wu, R.-Y. and Derynck, R. (1996)Receptor-associated Mad homologues synergize as effectors ofthe TGF-b response. Nature 383:168–172.
Zhang, Y., Chang, C., Gehling, D.J., Hemmati-Brivanlou, A. andDerynck, R. (2001) Regulation of Smad degradation and activityby Smurf2, an E3 ubiquitin ligase. Proc Natl Acad Sci USA98:974–979.
Zhao, Z. and Rivkees, S.A. (2000) Programmed cell death in thedeveloping heart: regulation by BMP4 and FGF2. Dev Dyn217:388–400.
Zhu, H., Kavsak, P., Abdollah, S., Wrana, J.L. and Thomsen,G.H. (1999) A SMAD ubiquitin ligase targets the BMPpathway and affects embryonic pattern formation. Nature 400:687–693.
Zimmerman, L.B., DeJesus-Escobar, J.M. and Harland,R.M. (1996) The Spemann organizer signal noggin bindsand inactivates bone morphogenetic protein 4. Cell 86:599–606.
444
Related Documents











