EVIDENCE FOR A SPECIALIZED LOCALIZATION OF THE CHLOROPLAST ATP-SYNTHASE SUBUNITS a, b, AND g IN THE EYESPOT APPARATUS OF CHLAMYDOMONAS REINHARDTII (CHLOROPHYCEAE) 1 Melanie Schmidt, Matthias Luff, Anne Mollwo Institute of Biology, Friedrich-Alexander-University, D-91058 Erlangen, Germany Marc Kaminski, Maria Mittag Institute of General Botany and Plant Physiology, Friedrich-Schiller-University Jena, D-07743 Jena, Germany and Georg Kreimer 2 Institute of Biology, Friedrich-Alexander-University, D-91058 Erlangen, Germany The eyespot apparatus (EA) of Chlamydomonas reinhardtii P. A. Dang. consists of two layersof car- otenoid-rich lipid globules subtended by thyla- koids. The outermost globule layer is additionally associated with the chloroplast envelope mem- branes and the plasma membrane. In a recent pro- teomic approach, we identified 202 proteins from isolated EAs of C. reinhardtii via at least two peptides, including, for example, structural com- ponents, signalling-related proteins, and photosyn- thetic-related membrane proteins. Here, we have analyzed the proteins of the EA with regard to their topological distribution using thermolysin to find out whether the arrangement of globules and mem- branes provides protection mechanisms for some of them. From about 230 protein spots separated on two-dimensional gels, the majority were degraded by thermolysin. Five major protein spots were pro- tected against the action of this protease. These pro- teins and some that were degradable were identified by mass spectrometry. Surprisingly, the thermolysin- resistant proteins represented the a and b subunits of the soluble CF 1 complex of the chloroplast ATP synthase. Degradable proteins included typical membrane proteins like LHCs, demonstrating that thermolysin is not in general sterically prevented by the EA structure from reaching membrane-associ- ated proteins. A control experiment showed that the CF 1 complex of thylakoids is efficiently degraded by thermolysin. Blue native PAGE of thermolysin- treated EAs followed by SDS-PAGE revealed that the a and b subunits are present in conjunction with the c subunit in a thermolysin-resistant complex. These results provide strong evidence that a significant proportion of these ATP-synthase subunits have a specialized localization and function within the EA of C. reinhardtii. Key index words: ATP synthase; blue native PAGE; CF 1 complex; Chlamydomonas reinhardtii; eyespot; proteomics; thermolysin Abbreviations: Bis-Tris, bis(2-hydroxyethyl)imino- tris(hydroxymethyl)methane; BN-PAGE, blue na- tive polyacyrlamide gel electrophoresis; EA, eyespot apparatus; GAP, globuli-associated protein; Hepes, N-(2-hydroxyethyl)piperazine-N 0 -2- ethanesulfonic acid; LC–ESI–MS, liquid chroma- tography–electrospray ionization–mass spectrome- try; pI, isoelectric point; Vhr, volt hours; X corr , cross-correlation factor The eyespot apparatus (EA) is almost invariably present in flagellate green algal cells that exhibit pho- totaxis. The ultrastructure of the functional green algal EA is complex and involves local specializations of membranes from different compartments (reviewed in Melkonian and Robenek 1984, Kreimer 2001). Briefly, it consists of one to several highly ordered lay- ers of carotenoid-rich lipid globules inside the chloro- plast. The globules exhibit a remarkably constant diameter of 80–130 nm and usually exhibit a close hexagonal packing. Each layer is subtended by a thyl- akoid membrane; the outermost globule layer is addi- tionally attached to the two chloroplast envelope membranes and the adjacent plasma membrane. In this region, the plasma membrane and the outer chlo- roplast envelope membrane are specialized, as re- vealed by freeze fracturing. Both membranes have an unusually high intramembrane particle density (Naka- mura et al. 1973, Melkonian and Robenek 1984). As a cascade of rapid electrical responses originating from the EA is involved in phototaxis and photoshock (Sineshchekov and Govorunova 1999), it is generally believed that these particles represent proteins in- 1 Received 27 June 2006. Accepted 5 January 2007. 2 Author for correspondence: e-mail GKreimer@biologie. uni-erlangen.de. 284 J. Phycol. 43, 284–294 (2007) r 2007 by the Phycological Society of America DOI: 10.1111/j.1529-8817.2007.00331.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EVIDENCE FOR A SPECIALIZED LOCALIZATION OF THE CHLOROPLASTATP-SYNTHASE SUBUNITS a, b, AND g IN THE EYESPOT APPARATUS OF
CHLAMYDOMONAS REINHARDTII (CHLOROPHYCEAE)1
Melanie Schmidt, Matthias Luff, Anne Mollwo
Institute of Biology, Friedrich-Alexander-University, D-91058 Erlangen, Germany
Marc Kaminski, Maria Mittag
Institute of General Botany and Plant Physiology, Friedrich-Schiller-University Jena, D-07743 Jena, Germany
and
Georg Kreimer2
Institute of Biology, Friedrich-Alexander-University, D-91058 Erlangen, Germany
The eyespot apparatus (EA) of Chlamydomonasreinhardtii P. A. Dang. consists of two layers of car-otenoid-rich lipid globules subtended by thyla-koids. The outermost globule layer is additionallyassociated with the chloroplast envelope mem-branes and the plasma membrane. In a recent pro-teomic approach, we identified 202 proteins fromisolated EAs of C. reinhardtii via at least twopeptides, including, for example, structural com-ponents, signalling-related proteins, and photosyn-thetic-related membrane proteins. Here, we haveanalyzed the proteins of the EA with regard to theirtopological distribution using thermolysin to findout whether the arrangement of globules and mem-branes provides protection mechanisms for some ofthem. From about 230 protein spots separated ontwo-dimensional gels, the majority were degradedby thermolysin. Five major protein spots were pro-tected against the action of this protease. These pro-teins and some that were degradable were identifiedby mass spectrometry. Surprisingly, the thermolysin-resistant proteins represented the a and b subunitsof the soluble CF1 complex of the chloroplast ATPsynthase. Degradable proteins included typicalmembrane proteins like LHCs, demonstrating thatthermolysin is not in general sterically prevented bythe EA structure from reaching membrane-associ-ated proteins. A control experiment showed that theCF1 complex of thylakoids is efficiently degradedby thermolysin. Blue native PAGE of thermolysin-treated EAs followed by SDS-PAGE revealed that thea and b subunits are present in conjunction with thec subunit in a thermolysin-resistant complex. Theseresults provide strong evidence that a significantproportion of these ATP-synthase subunits have a
specialized localization and function within the EAof C. reinhardtii.
Key index words: ATP synthase; blue native PAGE;CF1 complex; Chlamydomonas reinhardtii; eyespot;proteomics; thermolysin
Abbreviations: Bis-Tris, bis(2-hydroxyethyl)imino-tris(hydroxymethyl)methane; BN-PAGE, blue na-tive polyacyrlamide gel electrophoresis; EA,eyespot apparatus; GAP, globuli-associatedprotein; Hepes, N-(2-hydroxyethyl)piperazine-N0-2-ethanesulfonic acid; LC–ESI–MS, liquid chroma-tography–electrospray ionization–mass spectrome-try; pI, isoelectric point; Vhr, volt hours; Xcorr,cross-correlation factor
The eyespot apparatus (EA) is almost invariablypresent in flagellate green algal cells that exhibit pho-totaxis. The ultrastructure of the functional green algalEA is complex and involves local specializations ofmembranes from different compartments (reviewedin Melkonian and Robenek 1984, Kreimer 2001).Briefly, it consists of one to several highly ordered lay-ers of carotenoid-rich lipid globules inside the chloro-plast. The globules exhibit a remarkably constantdiameter of 80–130 nm and usually exhibit a closehexagonal packing. Each layer is subtended by a thyl-akoid membrane; the outermost globule layer is addi-tionally attached to the two chloroplast envelopemembranes and the adjacent plasma membrane. Inthis region, the plasma membrane and the outer chlo-roplast envelope membrane are specialized, as re-vealed by freeze fracturing. Both membranes have anunusually high intramembrane particle density (Naka-mura et al. 1973, Melkonian and Robenek 1984). As acascade of rapid electrical responses originating fromthe EA is involved in phototaxis and photoshock(Sineshchekov and Govorunova 1999), it is generallybelieved that these particles represent proteins in-
1Received 27 June 2006. Accepted 5 January 2007.2Author for correspondence: e-mail GKreimer@biologie.
uni-erlangen.de.
284
J. Phycol. 43, 284–294 (2007)r 2007 by the Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2007.00331.x

volved in signal perception and transduction. Bothelectron microscopic and biochemical analyses alsosupport the presence of proteins at the surface of thecarotenoid-rich eyespot globules (Walne and Arnott1967, Renninger et al. 2001, 2006). Some of theseproteins appear to be involved in globule/globule aswell as globule/membrane interactions and preventingcoalescence of the globules in the hexagonally densepacked layers (Renninger et al. 2001, 2006).
Phototaxis requires the cell to determine the direc-tion of incident light. The complex ultrastructure ofthe green algal EA is perfectly suited to give both max-imal directionality and contrast to the photoreceptorsas the cell rotates around its longitudinal axis duringhelical forward swimming. Although the localization ofthe photoreceptors has not yet been analyzed at theultrastructural level, they are generally assumed to belocated in the plasma membrane patch overlying theglobule layer(s). The globule layers modulate the lightintensity reaching the photoreceptors by a dual mech-anism. First, they shield them from light passingthrough the cell body. Second, they act as a quarter-wave interference reflector for light that is fallingdirectly on the EA and is not absorbed by the photo-receptors. Thereby, the whole complex (i.e., the spe-cialized membranes and the eyespot globules) isimportant for precision of the phototactic orientationat low fluence rates (Foster and Smyth 1980, Morel-Laurens and Feinleib 1983, Kreimer and Melkonian1990, Harz et al. 1992, Kreimer et al. 1992, Matsu-naga et al. 2003).
The unicellular green alga Chlamydomonas reinhardtiiis a model for a wide range of biological processes(Harris 2001, Grossman et al. 2003). As its genome se-quences (nuclear, chloroplast, and mitochondria) areavailable, proteomics has also become a powerful toolin recent years for in-depth analysis of cellular sub-fractions. Thus, the subproteomes of chloroplast ribos-omes (Yamaguchi et al. 2002), cilia (Pazour et al.2005), centrioles (Keller et al. 2005), and the EA(Schmidt et al. 2006) have been unraveled. The iden-tified proteins of the EA confirmed and complementedthe earlier genetic studies that have been applied toidentify components of the EA of C. reinhardtii. Severalmutations affecting eyespot assembly and positioningare known (Hartshorne 1953, Morel-Laurens and Fe-inleib 1983, Pazour et al. 1995, Lamb et al. 1999,Nakamura et al. 2001, Roberts et al. 2001). The mu-tant approach led to the identification of two genesinvolved in eyespot assembly. In min1 mutant strains,only miniature eyespots are formed, whereas muta-tions in eye2 induce loss of a visible eyespot (Robertset al. 2001, Dieckmann 2003). Individual eyespot glob-ules are, however, still detectable by electron micros-copy in the mutant eye2 (Lamb et al. 1999). Thus,general formation of the globules is probably not af-fected by this mutation. It is not yet known how thesetwo proteins act at the molecular level to control eye-spot development and size. Additionally, several retinal-binding proteins have been identified, the most
abundant of which is present in two splicing variants.They are not acting as photoreceptors in phototaxisand photoshock, and their function is not yet known(Deininger et al. 1995, Fuhrmann et al. 2001, 2003).In addition, two unique retinal-based photoreceptorsinvolved in the phototactic and photoshock responsehave been identified. Both act as directly light-gatedion channels when expressed in oocytes (Nagel et al.2002, Sineshchekov et al. 2002, Nagel et al. 2003,Suzuki et al. 2003, Govorunova et al. 2004). However,as discussed by Sineshchekov and Spudich (2005),the functional importance of these activities of thephotoreceptors under physiological conditions inC. reinhardtii is not yet clear. The recent large-scaleproteomic approach has led to the identification ofadditional 196 proteins identified via at least two dif-ferent peptides that are present in a fraction enrichedin EAs of C. reinhardtii (Schmidt et al. 2006). This studyhas provided new insights into components and pos-sible signaling elements present in this complex greenalgal cell organelle. A large number of the identifiedproteins have at least a partial hydrophobic character(39.6%) or are membrane proteins (19.3%). In addi-tion, putative structural components have been iden-tified. For example, 4% of the identified proteinspossess a plastid lipid-associated protein (PAP)-fibrillindomain. One function of members of the fibrillin fam-ily in higher plants is stabilization of carotenoid fibrils,plastoglobules, and thylakoids (Deruere et al. 1994,Gillet et al. 1998, Kessler et al. 1999, Rey et al. 2000).Overexpression of fibrillin in higher plants results inorganization of plastoglobules in clusters (Rey et al.2000). Therefore, Schmidt et al. (2006) hypothesizedthat some of the proteins with PAP-fibrillin domainsmight have functions in globule stabilization and mayalso be involved in interactions necessary to form thehighly ordered eyespot globule layers.
Proteolytic methods are often used to analyze thetopology of membrane proteins and membrane-associated proteins. In plastids, the proteases trypsinand thermolysin are commonly applied for this pur-pose (Joyard et al. 1983, Zilber and Malkin 1992,Deruere et al. 1994, Karnauchov et al. 1997, Beckeret al. 2004). Thermolysin was superior to trypsin andalso to the protease mixture pronase when probingsurfaces of intact organelles (Cline et al. 1984). Topo-logical analyses of proteins associated with eyespotglobule preparations of the green alga Spermatozopsissimilis Preisig et Melkonian using thermolysin providedevidence that 19 proteins possess domains hidden inthe globule matrix or that are otherwise not susceptibleto this protease (Renninger et al. 2001, 2006). Mem-bers of this group of proteins were also suggested to beinvolved in globule stabilization. However, none ofthese proteins was identified at the molecular level(e.g., by liquid chromatography–electron ionization–mass spectrometry [LC–ESI–MS] approaches) as themissing genome sequence of this green alga does notallow a straightforward proteome approach. In thepresent study, we applied a similar strategy to the EA
CF1 SUBUNITS IN THE EYESPOT 285

fraction of C. reinhardtii to identify major thermolysin-resistant proteins in this fraction by LC–ESI–MS/MS.Two-dimensional (2-D) gels of proteins from thermol-ysin-treated EA fractions revealed five major proteinspots with unchanged apparent isolectric point (pI)and molecular mass positions in comparison with thereference gels. These protein spots were identifiedwith high peptide coverage as a and b subunits ofthe soluble CF1 complex of the chloroplast ATP syn-thase. In contrast, typical membrane proteins of thephotosynthetic machinery present in the EA fractionwere sensitive toward this protease as well as the ATP-synthase complex from thylakoids. Blue native PAGE(BN-PAGE) analysis of a thermolysin-treated EA frac-tion demonstrated that the a and b subunits are evenpresent as a complex in conjunction with the g-chain inthe EA. Thus, these unexpected results point to a spe-cialized localization and function of a significant frac-tion of these ATP-synthase subunits in the EA fraction.
MATERIALS AND METHODS
Culture conditions and isolation of eyespot apparatuses and thy-lakoids. Chlamydomonas reinhardtii strain cw15 was grown in amodified Gorman–Levine Tris-acetate-phosphate medium(Harris 1989) in 10 L flasks at 151C in a 14:10 light:dark(L:D) cycle. Cultures were illuminated with white fluorescentlamps at 40–60 mmol photons �m�2 � s�1 and bubbled withordinary air. Eyespot apparatuses were isolated from late log-phase cultures (20 L) according to Schmidt et al. (2006)except when the eyespot apparatuses were isolated for 2-Delectrophoresis. In this case, samples were concentrated asfollows: The eyespot-enriched fraction was brought to 45%(w/v) sucrose, and 20 mL of sample was overlaid by 16 mL25% (w/w) sucrose dissolved in 5 mM N-(2-hydroxyethyl)pi-perazine-N 0-2-ethanesulfonic acid (Hepes), 2 mM EDTA, pH7.8, and 2 mL of the same buffer without sucrose and cen-trifuged (100,000 g, 30 min, 41C). The concentrated eyespotfraction was collected from the top of the gradient, extractedeither directly with chloroform:methanol:water (4:8:3) anddissolved in 2� SDS sample buffer (Kreimer et al. 1991) orsupplemented with 1 mM phenylmethylsulfonyl-fluoride andstored for no longer than 2 d at � 801C. Thylakoids wereisolated according to Chua and Bennoun (1975), with themodifications outlined in Schmidt et al. (2006).
Protease treatments. Thermolysin and pronase (Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany) treatmentsof the eyespot fractions (ratio of protease to eyespot pro-tein 5 6:1) were done according to Renninger et al. (2001)with the following modifications: The incubation time wasincreased to 2 h, and after protease treatments, the sampleswere brought to 42% sucrose (w/v) before a flotation centrif-ugation step [8 mL 42% sucrose/sample, 20.5% (w/w) sucrosebuffered with buffer A (10 mM Hepes NaOH, 4 mM MgCl2,pH 7.8), 2 mL buffer A; 100,000 g, 60 min, 41C]. Protease-treated eyespot apparatuses concentrated in buffer A wereremoved and extracted as described above. The ‘‘in gel’’ di-gestion experiments were done as described (Renningeret al. 2001). For protease treatment of isolated thylakoids,the sample was incubated with thermolysin (ratio of proteaseto thylakoid proteins 5 10:1) for 1 h at 41C. The reaction wasstopped by addition of 7.5 mM EDTA, and the thylakoidswere washed three times (5 mM Hepes KOH, pH 7.5;46,000 g, 12 min, 41C). Following extraction with chloro-form:methanol:water (4:8:3), the protein pellet was dissolvedin 2� SDS sample buffer.
Electrophoretic methods. The SDS-PAGE analyses were con-ducted in a modified high Tris system according to Calenberget al. (1998). For isoelectric focusing, chloroform:metha-nol:water (4:8:3) extracted samples were additionally washedonce with cold 90% acetone before resuspension in 2-D-solubilization buffer. Isoelectric focusing was done with slightmodifications according to Hippler et al. (2001) using an IPG-phor system (Amersham Biosciences, Freiburg, Germany)with Immobiline DryStrip precast gel (IPG) strips pH 3–10(11 and 18 cm; Amersham Biosciences). Focusing was per-formed at 161C with the following program: 500 V (500 Vhr),1000 V (1000 Vhr), 8000 V (11 cm strips: 16,000 Vhr; 18 cmstrips: 32,000 Vhr). For BN-PAGE, concentrated EA fractions(Vivaspin 2, HY-membrane, 10,000 Da molecular mass cut off;VivaScience, Hannover, Germany) were treated for 20 min(41C) with solubilization buffer (2% b-dodecylmaltoside,500 mM e-Amino-n-caproic acid, 50 mM Bis-Tris, pH 7.0) be-fore the first dimension. After centrifugation (15 min,21,900 g, 41C), the upper, orange fraction was withdrawnand used for BN-PAGE according to Schagger et al. (1994).Gels were silver-stained according to Rabilloud et al. (1988)or, when used for MS, according to Wagner et al. (2004).
MS analysis. Bands and spots of interest were cut out ofsilver-stained gels, destained, and in-gel digested with tryp-sin. Peptides were separated by HPLC and analyzed by LC–ESI–MS/MS as described (Schmidt et al. 2006). Data weresearched against the following C. reinhardtii data bases: finalmodel database from the Joint Genome Institute (Version 2;genome.jgi-psf.org/chlre2/chlre2.home.html) and the chlo-roplast database (www.chlamy.org/chloro/default.html). Dataanalysis was done with the Bioworks software (version 3.2;Thermo Electron Corp., San Jose, CA, USA) including theSEQUEST algorithm (Link et al. 1999). Searches were donefor tryptic peptides allowing two missed cleavages. The soft-ware parameters were set to detect a modification of 16 Da onMet, representing its oxidized form. Scores for the cross-cor-relation factor Xcorr (Eng et al. 1994) were set to the followinglimits: Xcorr41.5 if the charge of the peptide was 1; Xcorr42if the charge of the peptide was 2; Xcorr42.5 if the charge ofthe peptide was 3. Only peptides that fulfilled the Xcorr limitsand that had a probability value (P)�0.01 and a deltaCN�0.081 were included in further analysis. The proteinwas considered to be identified only when at least two dif-ferent peptides were present. The peptide sequences of thegene models were compared with the NCBI protein databaseusing basic local alignment search tool (BLAST; Altschulet al. 1997).
Other methods. Western blot analyses with the antiGAP56serum (Renninger et al. 2006) were carried out accordingto Calenberg et al. (1998). Protein content was measuredaccording to Neuhoff et al. (1979) with BSA as standard.Images of gels were taken with a Coolpix 990 (Nikon, Tokyo,Japan) and processed with Photoshop (Adobe Systems, SanJose, CA, USA).
RESULTS AND DISCUSSION
In order to identify specific thermolysin-resistantproteins in the EA fraction of C. reinhardtii, untreatedand treated samples were separated by 2-D-electro-phoresis (Fig. 1). In silver-stained 2-D gels of untreatedsamples, at least 230 spots were resolved, includingvariants of some proteins that are probably posttrans-lationally modified (Fig. 1A). This number correlateswell with the 202 different proteins identified with atleast two different peptides in a recent large-scale pro-teomic approach of the EA fraction (Schmidt et al.
MELANIE SCHMIDT ET AL.286

2006). It also demonstrates that significant protein lossin the 2-D pattern, a problem often occurring with hy-drophobic proteins (Adessi et al. 1997, Santoni et al.2000), was not evident for the EA fraction. All majorbands resolved in the 1-D reference lane correspondedto intense spots in the 2-D map. Most proteins hadapparent pI’s between ~pH 3.4 and ~pH 7.0. About20% of the proteins focused between ~pH 7.3 and~pH 9.9. The vast majority of the proteins were sen-sitive to thermolysin treatments of the isolated eyespots(Fig. 1B). In 2-D gels of thermolysin-treated fractions,most spots completely disappeared or shifted in theirapparent molecular masses and pI’s. Only five domin-ant protein spots were unchanged in their apparentmolecular mass and pI positions in comparison withreference gels. These spots originated from the proteinbands marked in the 1-D reference lane of Fig. 1A at56 (spots 1, 2) and 52 kDa (spots 3–5). Their relativeamounts were increased, thereby further confirmingtheir resistance to thermolysin.
To identify the major thermolysin-resistant pro-teins, the five spots marked in Fig. 1B were cut outof the 2-D gel and digested with trypsin. In addition,the 52 and 56 kDa bands of an untreated EA fractionwere tryptic digested. The resulting peptides weresubjected to nano-LC–ESI–MS analyses using a linearion trap mass spectrometer. For each 2-D spot, be-tween 16 and 25 peptides fulfilling the criteria forXcorr, probability score, and delta CN value were iden-tified (see ‘‘Materials and Methods’’, and supplemen-tary materials, Table S1). Data base analysis revealed
that spots 1 and 2 represent the a chain of the solublechloroplast ATP-synthase CF1 complex, and spots 3 to5 the b chain of this enzyme. Sequence coverage, whichwas relatively high, ranged between 35.6% and 71.3%,respectively (Fig. 2). In addition, in the control 1-Dbands and corresponding spots of untreated samples,both subunits were unequivocally identified (e.g.,56 kDa band, a-chain, 43.3% sequence coverage;52 kDa band, b-chain, 60.7% sequence coverage; datanot shown). Therefore, the possibility that the ATP-synthase subunit spots in the 2-D gel derived artificiallyfrom bands of higher apparent molecular masses bythermolysin treatment can be ruled out. This conclu-sion is further corroborated by the good match of thetheoretical molecular masses and pI’s with those de-termined experimentally for the proteins spots. Theseare 54.75 kDa (pI 5.44) versus 56 kDa (pI ~5.1–5.4)for the spots of the a subunit and 52.04 kDa (pI 5.20)versus 52 kDa (pI ~4.7–5.0) for the spots of the b sub-unit. Interestingly, the a and b subunits resolved in twoand three spots, respectively (Fig. 1B). The intensity ofspots 1–5 varied slightly in independent 2-D experi-ments. Such a row is often indicative of protein mod-ification (e.g., phosphorylation). The observed shifts inpI are, for example, predicted by the program Scansite(http://scansite.mit.edu/calc_mw_pi.html) by assumingfour phosphorylation sites for the a-subunit (pI5.44! 5.12) and three to 10 phosphorylation sites inthe b subunit (pI 5.20 ! 5.02/4.70). Phosphorylationof the ATP-synthase b subunit has been reported forchloroplasts (Kanekatsu et al. 1998). In addition, inter-
FIG. 1. The 2-D electrophoretic separation of proteins from the EA fraction of Chlamydomonas reinhardtii (A) and proteins from afraction treated for 2 h with thermolysin (B). Proteins (30mg) were separated on 18 cm IPG strips in the first dimension and an 11% SDS-PAGE in the second dimension. In addition, 6mg proteins from an EA fraction were separated on a 1-D gel (11% SDS-PAGE). The gelswere silver stained. Positions of molecular mass markers are indicated on the left in kilodaltons. Letters a–f mark thermolysin degradableproteins, which have been analyzed by LC–ESI–MS/MS (see Table 1). Major thermolysin-resistant protein spots have been numbered(1–5). Spots 1 and 2 originate from the 56 kDa protein band marked in the 1-D control lane in (A) by a gray arrow, whereas spots 3–5originate from the 52 kDa protein band (gray arrow head). The black arrow in the 2-D gel of (A) indicates the position of protein spots 1and 2, whereas the black arrowhead points to the position of protein spots 3–5 in the untreated samples.
CF1 SUBUNITS IN THE EYESPOT 287

action of the b subunit with phosphoserine/phospho-threonine-binding 14-3-3 proteins in a phosphoryla-tion-dependent manner occurs and regulates theactivity of the ATP synthase in chloroplasts and mito-chondria (Bunney et al. 2001). Specific sites of poten-tial phosphorylation are of interest for future studieson the ATP-synthase subunits localized in the EA butwill require enrichment of phosphopeptides (e.g., viaIMAC) to obtain reliable results.
Because of the special ultrastructure of the EAwhere the globules are subtended by the thylakoidmembrane, the presence of the chloroplast ATP syn-thase in EA fractions is not surprising, and several sub-units (including the a and b subunits and two subunits
of the membrane spanning CF0 complex) of this en-zyme have been identified in a recent proteomic ap-proach with this EA fraction of C. reinhardtii (Schmidtet al. 2006). However, resistance of these subunits thatbelong to the soluble CF1 part, and not to the mem-brane-spanning CF0 part, of the complex against thenonspecific metalloprotease thermolysin was clearlyunexpected. Both subunits contain several cleavagesites for thermolysin, which hydrolyze peptide bondsinvolving the amino group of hydrophobic amino acidswith bulky side chains. In line with this, in-gel digestionof the 52 and 56 kDa bands derived from the EA frac-tion by SDS-PAGE with thermolysin confirmed in vitrosensitivity of both bands toward this protease (data not
CF1 ATP-synthase chain
Spot 1 (coverage: 42.9%) MAMRTPEELSNLIKDLIEQYTPEVKMVDFGIVFQVGDGIARIYGLEKAMSGELLEFEDGTLGIALNLEAN NVGAVLLGDGLKITEGSRVRCTGKIAEIPVGEAYLGRVVDGLARPVDGKGAVQTKDSRAIESPAPGIVARRSVYEPLATGLVAVDAMIPVGRGQRELIIGDRQTGKTAIAVDTILNQKGKGVICVYVAIGQKASSVAQVLNTLKERGALDYTIIVMANANEPATLQYLAPYTGATLAEYFMYTGRPTLTIYDDLSKQAQAYREMSLLLRR PPGREAYPGDVFYLHSRLLERAAKLNNALGEGSMTALPIVETQEGDVSAYIPTNVISITDGQIFLAAGLF NSGLRPAINVGISVSRVGSAAQPKAMKQVAGKLKLELAQFAELEAFSQFASDLDQATQNQLARGARLREI LKQPQSSPLSVEEQVASLYAGTNGYLDKLEVSQVRAYLSGLRSYLANSYPKYGEILRSTLTFTPEAEGLVKQAINEYLEEFKSQAKAA
Spot 2 (coverage: 35.6%) MAMRTPEELSNLIKDLIEQYTPEVKMVDFGIVFQVGDGIARIYGLEKAMSGELLEFEDGTLGIALNLEAN NVGAVLLGDGLKITEGSRVRCTGKIAEIPVGEAYLGRVVDGLARPVDGKGAVQTKDSRAIESPAPGIVARRSVYEPLATGLVAVDAMIPVGRGQRELIIGDRQTGKTAIAVDTILNQKGKGVICVYVAIGQKASSVAQVL NTLKERGALDYTIIVMANANEPATLQYLAPYTGATLAEYFMYTGRPTLTIYDDLSKQAQAYREMSLLLRR PPGREAYPGDVFYLHSRLLERAAKLNNALGEGSMTALPIVETQEGDVSAYIPTNVISITDGQIFLAAGLF NSGLRPAINVGISVSRVGSAAQPKAMKQVAGKLKLELAQFAELEAFSQFASDLDQATQNQLARGARLREI LKQPQSSPLSVEEQVASLYAGTNGYLDKLEVSQVRAYLSGLRSYLANSYPKYGEILRSTLTFTPEAEGLVKQAINEYLEEFKSQAKAA
CF1 ATP-synthase ß chain
Spot 3 (coverage: 61.7%) MSDSIETKNMGRIVQIIGPVLDIVFAKGQVPNIYNALTIRAKNSAGTEMAVTCEVQQLLGDNCVRAVSMNPTEGLMRGMEVVDTGKPLSVPVGKVTLGRIFNVLGEPVDNMGNVKVEETLPIHRTAPAFVDLDTRLSIFETGIKVVDLLAPYRRGGKIGLFGGAGVGKTVLIMELINNIAKAHGGVSVFAGVGERTREGNDLYTEMKESGVIVEKNLSDSKVALVYGQMNEPPGARMRVALTALTMAEYFRDVNKQDVLFFIDNIFRFVQAGAEVSALLGRMPSAVGYQPTLATEMGGLQERITSTKDGSITSIQAVYVPADDLTDPAPATTFAHLDATTVLSRNLAAKGIYPAVDPLESTSTMLQPWILGEKHYDSAQSVKKTLQRYKELQDIIAILGLDELSEEDRLIVARARKIERF LSQPFFVAEVFTGSPGKYVSLAETIEGFGKIFAGELDDLPEQAFYLVGNITEAISKAASLK
Spot 4 (coverage: 71.3%) MSDSIETKNMGRIVQIIGPVLDIVFAKGQVPNIYNALTIRAKNSAGTEMAVTCEVQQLLGDNCVRAVSMNPTEGLMRGMEVVDTGKPLSVPVGKVTLGRIFNVLGEPVDNMGNVKVEETLPIHRTAPAFVDLDTRLSIFETGIKVVDLLAPYRRGGKIGLFGGAGVGKTVLIMELINNIAKAHGGVSVFAGVGERTREGNDLYTEMKESGVIVEKNLSDSKVALVYGQMNEPPGARMRVALTALTMAEYFRDVNKQDVLFFIDNIFRFVQAGAEVSALLGRMPSAVGYQPTLATEMGGLQERITSTKDGSITSIQAVYVPADDLTDPAPATTFAHLDATTVLSRNLAAKGIYPAVDPLESTSTMLQPWILGEKHYDSAQSVKKTLQRYKELQDIIAILGLDELSEEDRLIVARARKIERF LSQPFFVAEVFTGSPGKYVSLAETIEGFGKIFAGELDDLPEQAFYLVGNITEAISKAASLK
Spot 5 (coverage: 49.7%) MSDSIETKNMGRIVQIIGPVLDIVFAKGQVPNIYNALTIRAKNSAGTEMAVTCEVQQLLGDNCVRAVSMNPTEGLMRGMEVVDTGKPLSVPVGKVTLGRIFNVLGEPVDNMGNVKVEETLPIHRTAPAFVDLDTRLSIFETGIKVVDLLAPYRRGGKIGLFGGAGVGKTVLIMELINNIAKAHGGVSVFAGVGERTREGNDLYTEMKESGVIVEKNLSDSKVALVYGQMNEPPGARMRVALTALTMAEYFRDVNKQDVLFFIDNIFRFVQAGAEVSALLGRMPSAVGYQPTLATEMGGLQERITSTKDGSITSIQAVYVPADDLTDPAPATTFAHLDATTVLSRNLAAKG IYPAVDPLESTSTMLQPWILGEKHYDSAQSVKKTLQRYKELQDIIAILGLDELSEEDRLIVARARKIERF LSQPFFVAEVFTGSPGKYVSLAETIEGFGKIFAGELDDLPEQAFYLVGNITEAISKAASLK
FIG. 2. Amino acid sequencesof the chloroplast ATP-synthasesubunits a and b of Chlamydomo-nas reinhardtii. Peptides identi-fied by LC–ESI–MS/MS arehighlighted in bold. Spot num-bering corresponds to that ofFigure 1B. Further details aboutthe peptides identified by LC–ESI–MS/MS are given in theSupplementary materials, TableS1.
MELANIE SCHMIDT ET AL.288
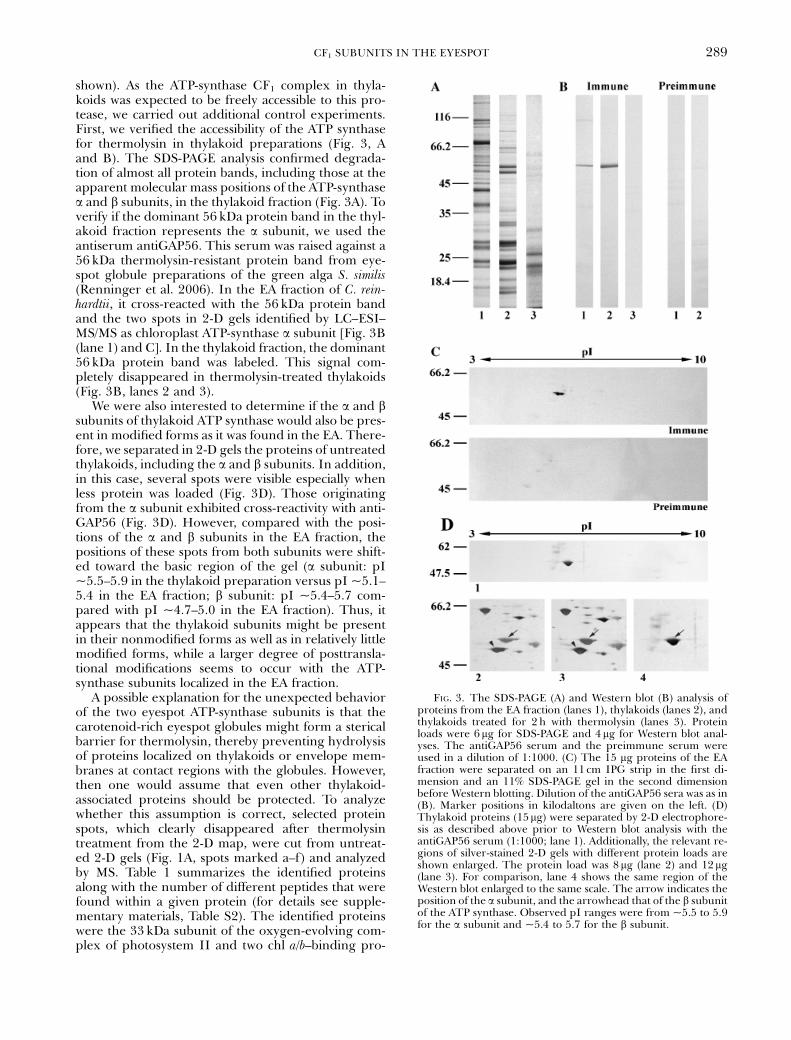
shown). As the ATP-synthase CF1 complex in thyla-koids was expected to be freely accessible to this pro-tease, we carried out additional control experiments.First, we verified the accessibility of the ATP synthasefor thermolysin in thylakoid preparations (Fig. 3, Aand B). The SDS-PAGE analysis confirmed degrada-tion of almost all protein bands, including those at theapparent molecular mass positions of the ATP-synthasea and b subunits, in the thylakoid fraction (Fig. 3A). Toverify if the dominant 56 kDa protein band in the thyl-akoid fraction represents the a subunit, we used theantiserum antiGAP56. This serum was raised against a56 kDa thermolysin-resistant protein band from eye-spot globule preparations of the green alga S. similis(Renninger et al. 2006). In the EA fraction of C. rein-hardtii, it cross-reacted with the 56 kDa protein bandand the two spots in 2-D gels identified by LC–ESI–MS/MS as chloroplast ATP-synthase a subunit [Fig. 3B(lane 1) and C]. In the thylakoid fraction, the dominant56 kDa protein band was labeled. This signal com-pletely disappeared in thermolysin-treated thylakoids(Fig. 3B, lanes 2 and 3).
We were also interested to determine if the a and bsubunits of thylakoid ATP synthase would also be pres-ent in modified forms as it was found in the EA. There-fore, we separated in 2-D gels the proteins of untreatedthylakoids, including the a and b subunits. In addition,in this case, several spots were visible especially whenless protein was loaded (Fig. 3D). Those originatingfrom the a subunit exhibited cross-reactivity with anti-GAP56 (Fig. 3D). However, compared with the posi-tions of the a and b subunits in the EA fraction, thepositions of these spots from both subunits were shift-ed toward the basic region of the gel (a subunit: pI~5.5–5.9 in the thylakoid preparation versus pI ~5.1–5.4 in the EA fraction; b subunit: pI ~5.4–5.7 com-pared with pI ~4.7–5.0 in the EA fraction). Thus, itappears that the thylakoid subunits might be presentin their nonmodified forms as well as in relatively littlemodified forms, while a larger degree of posttransla-tional modifications seems to occur with the ATP-synthase subunits localized in the EA fraction.
A possible explanation for the unexpected behaviorof the two eyespot ATP-synthase subunits is that thecarotenoid-rich eyespot globules might form a stericalbarrier for thermolysin, thereby preventing hydrolysisof proteins localized on thylakoids or envelope mem-branes at contact regions with the globules. However,then one would assume that even other thylakoid-associated proteins should be protected. To analyzewhether this assumption is correct, selected proteinspots, which clearly disappeared after thermolysintreatment from the 2-D map, were cut from untreat-ed 2-D gels (Fig. 1A, spots marked a–f) and analyzedby MS. Table 1 summarizes the identified proteinsalong with the number of different peptides that werefound within a given protein (for details see supple-mentary materials, Table S2). The identified proteinswere the 33 kDa subunit of the oxygen-evolving com-plex of photosystem II and two chl a/b–binding pro-
FIG. 3. The SDS-PAGE (A) and Western blot (B) analysis ofproteins from the EA fraction (lanes 1), thylakoids (lanes 2), andthylakoids treated for 2 h with thermolysin (lanes 3). Proteinloads were 6mg for SDS-PAGE and 4 mg for Western blot anal-yses. The antiGAP56 serum and the preimmune serum wereused in a dilution of 1:1000. (C) The 15 mg proteins of the EAfraction were separated on an 11 cm IPG strip in the first di-mension and an 11% SDS-PAGE gel in the second dimensionbefore Western blotting. Dilution of the antiGAP56 sera was as in(B). Marker positions in kilodaltons are given on the left. (D)Thylakoid proteins (15mg) were separated by 2-D electrophore-sis as described above prior to Western blot analysis with theantiGAP56 serum (1:1000; lane 1). Additionally, the relevant re-gions of silver-stained 2-D gels with different protein loads areshown enlarged. The protein load was 8 mg (lane 2) and 12mg(lane 3). For comparison, lane 4 shows the same region of theWestern blot enlarged to the same scale. The arrow indicates theposition of the a subunit, and the arrowhead that of the b subunitof the ATP synthase. Observed pI ranges were from ~5.5 to 5.9for the a subunit and ~5.4 to 5.7 for the b subunit.
CF1 SUBUNITS IN THE EYESPOT 289

teins, as well as a putative membrane-associated pro-tein of unknown function already previously identifiedin the EA fraction (Schmidt et al. 2006). Therefore, ageneral unspecific steric hindrance of the protease todegrade thylakoid-associated proteins by the eyespotglobules, as well as the presence of a significant amountof inside-out thylakoid vesicles in the EA fraction, canbe excluded as the simplest reason for failure of ther-molysin to degrade the ATP-synthase subunits. A pos-sible localization of the subunits inside the globulematrix also appears to be unlikely, as pronase—a mix-ture of several unspecific endo- and exoproteases gen-erally digesting proteins down to single amino acids—degraded all proteins in the EA fraction (data notshown). Notably, an identical behavior is reported forthe thermolysin-resistant 52 and 56 kDa bands in theeyespot globule preparations of S. similis (Renningeret al. 2001, 2006), which could represent these ATP-synthase subunits according to their apparent molec-ular masses and pI’s. In summary, these unexpectedresults strongly suggest a specialized localization of asignificant portion of the chloroplast ATP-synthase aand b subunits in the region of the EA in flagellategreen algae, possibly in between the eyespot globules.The close packing density of the globules could wellprevent thermolysin action by steric hindrance, where-as the protease mixture pronase is apparently able toweaken such sterical constraints and therefore allowscomplete degradation of all proteins in the EA fraction.
To gain information about the organization of thethermolysin-resistant a- and b-ATP-synthase subunitsin the EA fraction, BN-PAGE was applied. It is capableof resolving protein complexes (Schagger et al. 1994,Arnold et al. 1998) and has also been used successfullyfor the isolation of the chloroplast ATP synthase and itssubcomplexes, CF1 and CF0 (Neff and Dencher 1999,Rexroth et al. 2004). Resolution of a thermolysin-treated EA fraction by BN-PAGE followed by SDS-PAGE in the second dimension demonstrated thatthree dominant protein spots at apparent molecularmass positions of ~58, 54, and 38 kDa migrated as acomplex (Fig. 4). The LC–ESI–MS/MS analysis of thesespots identified them as chloroplast a-, b-, and g-ATP-synthase subunits (for details see Table 2). Other com-plexes usually resolved by BN-PAGE of untreated EA
fractions were absent; therefore, incomplete digestioncan be ruled out. The protein pattern in the lowermolecular mass range presumably represents degra-dation products, as it is also not present in gels of un-treated samples. This result strongly supports the ideathat the thermolysin-resistant a, b, and g subunits ofthe CF1 complex are arranged in a tight complex inthe EA region. Currently, it is not known why thesesubunits are sorted to the EA and become protectedagainst the protease thermolysin. This specializedlocalization might, for example, be induced upon spe-cific posttranslational modifications. The a- and b-ATP-
TABLE 1. Thermolysin-sensitive proteins of the EA fraction identified by LC–ESI–MS/MS analysis.
SpotaProtein ID Vs2b/Protein ID Vs3c
No. of identifiedpeptides Function and/or homologies of depicted proteins determined by NCBI BLASTp
a, b 171502/130316 9 PSBO oxygen-evolving enhancer protein 1(33 kDa subunit of oxygen-evolving complex of photosystem II)
c 159358/186064 4 LhcbM3 chl a/b–binding protein of LHCIc 155818/138069 3 LhcbM4 chl a/b–binding protein of LHCIId–f 157114/121991 10, 11, 10 Similar to conserved plant/cyanobacterial proteins of unknown functions;
contains DUF1350 domain(s)
aLabeled in Figure 1A.bJGI, version 2 of the Chlamydomonas reinhardtii genome.cJGI, version 3 of the C. reinhardtii genome.
FIG. 4. The 2-D BN/SDS-PAGE separation of proteins from athermolysin-treated EA fraction of Chlamydomonas reinhardtii. Theb-dodecylmaltoside solubilized proteins were separated on a BN-PAGE gradient gel (5%–15%). Following denaturation of theproteins in the gel strip, a high Tris SDS-PAGE gel (11%) wasused in the second dimension. The gel was silver stained, andspots 1–3 were analyzed after tryptic digestion by LC–ESI–MS/MS (for details see Table 2). The box highlights the vertical areawhere proteins of the CF1 complex are observed. Positions ofmolecular mass markers are given in kilodaltons on the left.
MELANIE SCHMIDT ET AL.290

TABLE 2. Identified thermolysin-resistant proteins after BN-PAGE (protein spots 1, 2, and 3 in Fig. 4).
Spot no. Identified peptides Z Xcorr
Function and protein ID Vs2/protein ID Vs3 or Cp
1 LKLELAQFAELEAFSQFASDLDQATQNQLAR 3 8.86 CF1 ATP-synthase a-chainMVDFGIVFQVGDGIAR 2 6.64 CpSVYEPLATGLVAVDAMIPVGR 2 6.43IAEIPVGEAYLGR 2 5.15QPQSSPLSVEEQVASLYAGTNGYLDKLEVSQVR 3 5.09LELAQFAELEAFSQFASDLDQATQNQLAR 3 5.05EILKQPQSSPLSVEEQVASLYAGTNGYLDK 3 4.82TAIAVDTILNQK 2 4.53DLIEQYTPEVK 2 4.22MoVDFGIVFQVGDGIAR 2 4.09AIESPAPGIVAR 2 4.05SVYEPLATGLVAVDAMoIPVGR 2 4.04STLTFTPEAEGLVK 2 3.80EAYPGDVFYLHSR 2 3.72TPEELSNLIKDLIEQYTPEVK 2 3.49TPEELSNLIK 2 3.45QAINEYLEEFK 2 3.44VVDGLARPVDGK 2 3.40PAINVGISVSR 2 3.25SYLANSYPK 2 3.03ASSVAQVLNTLK 2 2.95PTLTIYDDLSK 2 2.81YGEILR 2 2.40AYLSGLR 2 2.24IYGLEK 1 2.10
2 DVNKQDVLFFIDNIFR 2 6.77 CF1 ATP-synthase b-chainMPSAVGYQPTLATEMGGLQER 2 6.74 CpMoPSAVGYQPTLATEMGGLQER 2 6.58IFAGELDDLPEQAFYLVGNITEAISK 3 5.96VALVYGQMNEPPGAR 2 5.93GMEVVDTGKPLSVPVGK 2 5.69YKELQDIIAILGLDELSEEDRLIVAR 3 5.59MoPSAVGYQPTLATEMoGGLQER 2 5.36VALTALTMoAEYFR 2 5.28YKELQDIIAILGLDELSEEDR 2 5.26IFNVLGEPVDNMoGNVK 2 5.26IFNVLGEPVDNMGNVK 2 5.18VALVYGQMoNEPPGAR 2 5.05MPSAVGYQPTLATEMoGGLQER 2 4.87GIYPAVDPLESTSTMLQPWILGEK 2 4.85TVLIMoELINNIAK 2 4.72VALTALTMAEYFR 2 4.65GMoEVVDTGKPLSVPVGK 2 4.58AHGGVSVFAGVGER 2 4.45TVLIMELINNIAK 2 4.36IVQIIGPVLDIVFAK 2 4.23FVQAGAEVSALLGR 2 4.21TREGNDLYTEMK 2 4.13GIYPAVDPLESTSTMoLQPWILGEK 2 4.12QDVLFFIDNIFR 2 4.03YVSLAETIEGFGK 2 3.76AVSMNPTEGLMR 2 3.74AVSMNPTEGLMoR 2 3.64AVSMoNPTEGLMoR 2 3.46DGSITSIQAVYVPADDLTDPAPATTFAHLDATTVLSR 3 3.40AVSMoNPTEGLMR 2 3.39GQVPNIYNALTIR 2 3.36VEETLPIHR 2 3.23TAPAFVDLDTR 2 2.93TREGNDLYTEMoK 2 2.84LSIFETGIK 2 2.70IGLFGGAGVGK 2 2.67IFNVLGEPVDNMoGNVKVEETLPIHR 3 2.55EGNDLYTEMK 2 2.42ESGVIVEK 1 2.39VVDLLAPYRR 2 2.35NLSDSK 1 2.32EGNDLYTEMoK 2 2.20VVDLLAPYR 2 2.18
CF1 SUBUNITS IN THE EYESPOT 291

synthase subunits localized on the thylakoids eachexhibit several spots in 2-D gel separations. However,these spots focused at slightly more basic pH values incomparison with those in the EA fraction, indicatingthat they might occur in the thylakoids in their non-modified as well as slightly modified forms. The intro-duction of additional posttranslational modificationsmight serve as a signal to localize these subunits to theregion of the EA. In addition, the thermolysin-resistantsubunits may also interact with proteins specific to theEA region. Thereby, for example, a specifically mod-ified subset of the a and b subunits might be localizedto the EA, especially as no indications of the presenceof thermolysin-resistant subunits of the membrane-spanning CF0 complex were observed in either the2-D gels or the BN-PAGE analysis of the EA fraction,albeit the presence of CF0 in the EA fraction was shown(Schmidt et al. 2006). The function of the thermolysin-resistant complex in the EA is also still enigmatic. If aconsiderable portion of this complex is indeed local-ized on the eyespot globule surfaces and in the narrowspace between them, it may well have a stabilizing/structural role. The ATP synthase in both mitochon-dria and chloroplasts is capable of forming dimers(Arnold et al. 1998, Rexroth et al. 2004). Multiplefunctions, even outside the mitochondrion, are addi-tionally reported for the e subunit of the mitochondrialATP synthase. On the one hand, it is essential for max-imal ATP synthesis by mitochondria, but it was alsounexpectedly identified as a ligand for the G-protein-coupled receptor Methuselah in Drosophila (Cvejicet al. 2004, Kidd et al. 2005). In addition, for the mi-tochondrial ATP synthase, evidence for a link betweendimerization and cristae morphology does exist (Pau-mard et al. 2002). In mitochondria, dimerization ismediated by two nonessential subunits associated withthe complex and seems to be driven by the membrane-
intrinsic F0 subcomplex (Paumard et al. 2002, Toma-setig et al. 2002). In contrast, contact sites for the chlo-roplast ATP synthase are probably located on the CF1
subunit and involve a different mechanism. Thephysiological significance of dimerization of the chlo-roplast ATP synthase is not yet known, but roles instabilization and/or membrane curvature have beensuggested (Rexroth et al. 2004). Interactions betweenthe CF1-subunit complexes in the EA could, in prin-ciple, also help in formation of stable, hexagonallyclosed-packed eyespot layers. However, this suggestionis still speculative until detailed localization and furtherfunctional studies have been carried out.
We thank Nicole Eitzinger and Tobias Rei�enweber for helpwith BN-PAGE analysis, and Volker Wagner and Susan Hawatfor initial help with MS analysis. We appreciate the free deliv-ery of information by the USA (DOE) genome project of C.reinhardtii very much. This study was supported by grants ofthe Deutsche Forschungsgemeinschaft to Georg Kreimer andMaria Mittag.
Adessi, C., Miege, C., Albrieux, C. & Rabilloud, T. 1997. Two-di-mensional electrophoresis of membrane proteins: a currentchallenge for immobilized pH gradients. Electrophoresis18:127–35.
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z.,Miller, W. & Lipman, D. J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search pro-grams. Nucleic Acids Res. 25:3389–402.
Arnold, I., Pfeiffer, K., Neupert, W., Stuart, R. A. & Schagger, H.1998. Yeast mitochondrial F1F0-ATP synthase exists as adimer: identification of three dimer-specific subunits. EMBO(Eur. Mol. Biol. Organ.) J. 17:7170–8.
Becker, T., Hritz, J., Vogel, M., Caliebe, A., Bukau, B., Soll, J. &Schleiff, E. 2004. Toc12, a novel subunit of the intermembranespace preprotein translocon of chloroplasts. Mol. Biol. Cell15:5130–44.
Bunney, T. D., van Walraven, H. S. & de Boer, A. H. 2001. 14-3-3protein is a regulator of the mitochondrial and chloroplastATP synthase. Proc. Natl. Acad. Sci. U. S. A. 98:4249–54.
TABLE2 (Continued)
Spot no. Identified peptides Z Xcorr
Function and protein ID Vs2/protein ID Vs3 or Cp
3 SFSLGAAPSTK 2 3.36 CF1 ATP-synthase g-chainSLQEALASELAAR 2 3.25 159538/134235EVSSFDLVNRSQPLTYKVR 2 3.18RAQEAVVNGRPFSENLVK 3 3.16AQEAVVNGRPFSENLVK 3 3.13YRELTAMGVK 2 3.11MoNAMNNASDNAK 2 3.08MNAMNNASDNAK 2 3.04AQEAVVNGRPFSENLVKVLYGVNQR 3 2.99EAQGIADEIFASFIAQESDK 2 2.95VATGLLQGGFSVALK 2 2.69VLYGVNQR 2 2.51KGAQYFAR 2 2.42GGEFAVER 1 2.42IAEIPVGEAYLGR 2 2.41GLTVQYNK 2 2.30PFSENLVK 2 2.20QSGEVRYNGAELDQFNVR 2 2.09
Z, charge; Vs2, JGI version 2 of the Chlamydomonas reinhardtii genome; Vs3, JGI version 3 of the C. reinhardtii genome; Cp,chloroplast genome; Mo, oxidized methionine.
MELANIE SCHMIDT ET AL.292

Calenberg, M., Brohsonn, U., Zedlacher, M. & Kreimer, G. 1998.Light- and Ca2þ -modulated heterotrimeric GTPases in theeyespot apparatus of a flagellate green alga. Plant Cell 10:91–103.
Chua, N.-H. & Bennoun, P. 1975. Thylakoid membrane polypep-tides of Chlamydomonas reinhardtii: wild-type and mutant strainsdeficient in photosystem II reaction center. Proc. Natl. Acad.Sci. U. S. A. 72:2175–9.
Cline, K., Werner-Washburne, M., Andrews, J. & Keegstra, K.1984. Thermolysin is a suitable protease for probing thesurface of intact pea chloroplasts. Plant Physiol. 75:675–8.
Cvejic, S., Zhu, Z., Felice, S. Y., Berman, Y. & Huang, X.-Y. 2004.The endogenous ligand Stunted of the GPCR Methuselaextends lifespan in Drosophila. Nat. Cell Biol. 6:540–6.
Deininger, W., Kroger, P., Hegemann, U., Lottspeich, F. & Hege-mann, P. 1995. Chlamyrhodopsin represents a new type ofsensory photoreceptors. EMBO (Eur. Mol. Biol. Organ.) J.14:5849–58.
Deruere, J., Romer, S., D0Harlingue, A., Backhaus, R. A., Kuntz,M. & Camara, B. 1994. Fibril assembly and carotenoid over-accumulation in chromoplasts: a model for supramolecularlipoprotein structures. Plant Cell 6:119–33.
Dieckmann, C. L. 2003. Eyespot placement and assembly in thegreen alga Chlamydomonas. Bioessays 25:410–6.
Eng, J., McCormack, A. L. & Yates, J. R. 1994. An approach tocorrelate tandem mass spectral data of peptides with aminoacid sequences in a protein database. J. Am. Soc. Mass. Spectrom.5:976–89.
Foster, K. W. & Smyth, R. D. 1980. Light antennas in phototacticalga. Microbiol. Rev. 44:572–630.
Fuhrmann, M., Deininger, W., Kateriya, S. & Hegemann, P. 2003.Rhodopsin-related proteins, cop1, cop2, and chop1, inChlamydomonas reinhardtii. In Batschauer, A. [Ed.] Photoreceptorsand Light Signaling. Royal Society of Chemistry, Cambridge,UK, pp. 124–35.
Fuhrmann, M., Stahlberg, A., Govorunova, E., Rank, S. & Hege-mann, P. 2001. The abundant retinal protein of the Chlamy-domonas eye is not the photoreceptor for phototaxis andphotophobic responses. J. Cell Sci. 114:3857–63.
Gillet, B., Beyly, A., Peltier, G. & Rey, P. 1998. Molecular charac-terization of CDSP 34 a chloroplastic protein induced by waterdeficit in Solanum tuberosum L. plants and regulation of CDSP34 expression by ABA and high illumination. Plant J. 16:257–62.
Govorunova, E. G., Jung, K.-H., Sineshchekov, O. A. & Spudich, J.L. 2004. Chlamydomonas sensory rhodopsins A and B: cellularcontent and role in photophobic responses. Biophys. J.86:2342–9.
Grossman, A. R., Harris, E. E., Hauser, C., Lefebvre, P. A., Mar-tinez, D., Rokhsar, D., Shrager, J., Silflow, C. D., Stern, D.,Vallon, O. & Zhang, Z. 2003. Chlamydomonas reinhardtii at thecrossroads of genomics. Eukaryot. Cell 2:1137–50.
Harris, E. H. 1989. The Chlamydomonas Sourcebook. Academic Press,San Diego, California, 25 pp.
Harris, E. H. 2001. Chlamydomonas as a model organism. Annu. Rev.Plant Physiol. Plant Mol. Biol. 52:363–406.
Hartshorne, J. N. 1953. The function of the eyespot in Chlamy-domonas. New Phytol. 52:292–7.
Harz, H., Nonnengasser, C. & Hegemann, P. 1992. The photore-ceptor current of the green alga Chlamydomonas. Phil. Trans. R.Soc. Lond. B Biol. Sci. 338:39–52.
Hippler, M., Klein, J., Fink, J., Allinger, T. & Hoerth, P. 2001. To-wards functional proteomics of membrane protein complexes:analysis of thylakoid membranes from Chlamydomonas rein-hardtii. Plant J. 28:595–606.
Joyard, J., Billecocq, A., Bartlett, S. G., Block, M. A., Chua, N.-H. &Douce, R. 1983. Localization of polypeptides to the cytosolicside of the outer envelope membrane of spinach chloroplasts.J. Biol. Chem. 258:10000–6.
Kanekatsu, M., Saito, H., Motohashi, K. & Hisabori, T. 1998. Thebeta subunit of chloroplast ATP synthase (CF0CF1-ATPase) isphosphorylated by casein kinase II. Biochem. Mol. Biol. Int.46:99–105.
Karnauchov, I., Herrmann, R. G. & Klosgen, R. B. 1997. Trans-membrane topology of the Rieske Fe/S protein of the cyto-chrome b6/f complex from spinach chloroplasts. FEBS Lett.408:206–10.
Keller, L. C., Romijn, E. P., Zamora, I., Yates III, J. R. & Marshall,W. F. 2005. Proteomic analysis of isolated Chlamydomonas cen-trioles reveals orthologs of ciliary-disease genes. Curr. Biol.15:1090–8.
Kessler, F., Schnell, D. & Blobel, G. (1999). Identification of pro-teins associated with plastoglobules isolated from pea (Pisumsativum L.) chloroplasts. Planta 208:107–13.
Kidd, T., Abu-Shumays, R., Katzen, A., Sission, J. C., Jimenez, G.,Pinchin, S., Sullivan, W. & Ish-Horowicz, D. 2005. The e-sub-unit of mitochondrial ATP synthase is required for normalspindle orientation during the Drosophila embryonic divisions.Genetics 170:697–708.
Kreimer, G. 2001. Light reception and signal modulation duringphotoorientation of flagellate algae. In Lebert, M. & Hader, D.-P. [Eds.] Comprehensive Series in Photosciences. Photomovement.Vol. 1. Amsterdam Elsevier, North Holland, pp. 193–227.
Kreimer, G., Brohsonn, U. & Melkonian, M. 1991. Isolation andpartial characterization of the photoreceptive organelle forphototaxis of a flagellate green alga. Eur. J. Cell Biol. 55:318–27.
Kreimer, G. & Melkonian, M. 1990. Reflection confocal laser scan-ning microscopy of eyespots in flagellate green alga. Eur. J. CellBiol. 53:101–11.
Kreimer, G., Overlander, C., Sineshchekov, O. A., Stolzis, H., Nu-ltsch, W. & Melkonian, M. 1992. Functional analysis of theeyespot in Chlamydomonas reinhardtii mutant ey 627, mt� . Planta188:513–21.
Lamb, M. R., Dutcher, S. K., Worley, C. K. & Dieckmann, C. L.1999. Eyespot assembly mutants in Chlamydomonas reinhardtii.Genetics 153:721–9.
Link, A. J., Eng, J., Schieltz, D. M., Carmack, E., Mize, G. J., Morris,D. R., Garvik, B. M. & Yates III, J. R. 1999. Direct analysis ofprotein complexes using mass spectrometry. Nat. Biotechnol.17:676–82.
Matsunaga, S., Watanabe, S., Sakushi, S., Miyamura, S. & Hori, T.2003. Screening effect diverts the swimming directions fromdiaphototactic to positive phototactic in a disk-shaped greenflagellate Mesostigma viride. Photochem. Photobiol. 77:322–32.
Melkonian, M. & Robenek, H. 1984. The eyespot apparatus ofgreen algae: a critical review. Prog. Phycol. Res. 3:193–268.
Morel-Laurens, N. & Feinleib, M. E. 1983. Photomovement in an‘‘eyeless’’ mutant of Chlamydomonas. Photochem. Photobiol.37:189–94.
Nagel, G., Ollig, D., Fuhrmann, M., Kateriya, S., Musti, A. M.,Bamberg, E. & Hegemann, P. 2002. Channelrhodopsin-1: alight-gated proton channel in green algae. Science 296:2395–8.
Nagel, G., Szellas, T., Huhn, W., Kateriya, S., Adeishvili, N.,Berthold, P., Ollig, D., Hegemann, P. & Bamberg, E. 2003.Channelrhodopsin-2, a directly light-gated cation-selectivemembrane channel. Proc. Natl. Acad. Sci. U. S. A. 100:13940–5.
Nakamura, K., Bray, D. F., Costerton, J. W. & Wagenaar, E. B.1973. The eyespot of Chlamydomonas eugametos: a freeze-etchstudy. Can. J. Bot. 51:817–9.
Nakamura, S., Ogihara, H., Jinbo, K., Tateishi, M., Takahashi, T.,Yoshimura, K., Kubota, M., Watanabe, M. & Nakamura, S.2001. Chlamydomonas reinhardtii Dangeard (Chlamydomona-dales, Chlorophyceae) mutant with multiple eyespots. Phycol.Res. 49:115–21.
Neff, D. & Dencher, N. A. 1999. Purification of multisubunit mem-brane protein complexes: isolation of chloroplast F0F1-ATPsynthase, CF0 and CF1 by blue native electrophoresis. Biochem.Biophys. Res. Commun. 259:569–75.
Neuhoff, V., Phillip, K., Zimmer, H. G. & Mesecke, S. 1979. A sim-ple versatile sensitive and volume-independent method forquantitative protein determination which is independent ofother external influences. Hoppe-Seyler’s Z. Physiol. Chem.360:1657–70.
Paumard, P., Vaillier, J., Coulary, B., Schaeffer, J., Soubannier, V.,Mueller, D. M., Brethes, D., di Rago, J.-P. & Velours, J. 2002.
CF1 SUBUNITS IN THE EYESPOT 293

The ATP synthase is involved in generating mitochondrial cris-tae morphology. EMBO (Eur. Mol. Biol. Organ.) J. 21:221–30.
Pazour, G., Sineshchekov, O. A. & Witman, G. B. 1995. Mutationalanalysis of the phototransduction pathway of Chlamydomonasreinhardtii. J. Cell Biol. 131:427–40.
Pazour, G. J., Agrin, N., Leszyk, J. & Witman, G. B. 2005. Pro-teomic analysis of a eukaryotic cilium. J. Cell Biol. 170:103–13.
Rabilloud, T., Carpentier, G. & Tarroux, P. 1988. Improvementand simplification of low-background silver staining by usingsodium dithionite. Electrophoresis 9:288–91.
Renninger, S., Backendorf, E. & Kreimer, G. 2001. Subfractiona-tion of eyespot apparatuses from the green alga Spermatozopsissimilis: isolation and characterization of eyespot globules.Planta 213:51–63.
Renninger, S., Dieckmann, C. L. & Kreimer, G. 2006. Towards aprotein map of the green algal eyespot: analysis of eyespotglobule-associated proteins. Phycologia 45:199–212.
Rexroth, S., Meyer zu Tittingdorf, J. M. W., Schwa�mann, H. J.,Krause, F., Seelert, H. & Dencher, N. A. 2004. DimericHþ -ATP synthase in the chloroplast of Chlamydomonasreinhardtii. Biochim. Biophys. Acta 1658:202–11.
Rey, P., Gillet, B., Romer, S., Eymery, F., Massimino, J., Peltier, G. &Kuntz, M. 2000. Over-expression of a pepper plastid-lipid-associated protein in tobacco leads to changes in plastid-ultra-structure and plant development upon stress. Plant J. 21:483–94.
Roberts, D. G. W., Lamb, M. R. & Dieckmann, C. L. 2001. Char-acterization of the eye2 gene required for eyespot assembly inChlamydomonas reinhardtii. Genetics 158:1037–49.
Santoni, V., Molloy, M. & Rabilloud, T. 2000. Membrane proteinsand proteomics: un amour impossible? Electrophoresis 21:1054–70.
Schagger, H., Cramer, W. A. & Jagow, G. V. 1994. Analysis of mo-lecular masses and oligomeric states of protein complexes byblue native electrophoresis and isolation of membrane proteincomplexes by two-dimensional native electrophoresis. Anal.Biochem. 217:220–30.
Schmidt, M., Ge�ner, G., Luff, M., Heiland, I., Wagner, V.,Kaminski, M., Geimer, S., Eitzinger, N., Rei�enweber, T.,Voystekh, O., Fiedler, M., Mittag, M. & Kreimer, G. 2006.Proteomic analysis of the eyespot of Chlamydomonas reinhardtiiprovides novel insights into its components and tactic move-ments. Plant Cell 18:1908–30.
Sineshchekov, O. A. & Govorunova, E. G. 1999. Rhodopsin-medi-ated photosensing in green flagellated algae. Trends Plant Sci.4:58–63.
Sineshchekov, O. A., Jung, K.-H. & Spudich, J. L. 2002. Two rhod-opsins mediate phototaxis to low- and high-intensity lightin Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. U. S. A. 99:8689–94.
Sineshchekov, O. A. & Spudich, J. L. 2005. Sensory rhodopsinsignaling in green flagellate algae. In Spudich, J. L. & Briggs,W. R. [Eds.] Handbook of Photosensory Receptors. Wiley-VCH,Weinheim, Germany, pp. 25–42.
Suzuki, T., Yamasaki, K., Fujita, S., Oda, K., Iseki, M., Yoshida, K.,Watanabe, M., Daiyasu, H., Toh, H., Asamizu, E., Tabata, S.,Miura, K., Fukuzawa, H., Nakamura, S. & Takahashi, T. 2003.Archael-type rhodopsins in Chlamydomonas: model structureand intracellular localization. Biochem. Biophys. Res. Commun.301:711–7.
Tomasetig, L., di Pancratio, F., Harris, D. A., Mavelli, I. & Lippe, G.2002. Dimerization of F0F1 ATP synthase from bovine heart isindependent from the binding of the inhibitor protein IF1.Biochim. Biophys. Acta 1556:133–41.
Wagner, V., Fiedler, M., Markert, C., Hippler, M. & Mittag, M.2004. Functional proteomics of circadian expressed proteinsfrom Chlamydomonas reinhardtii. FEBS Lett. 559:129–35.
Walne, P. L. & Arnott, H. J. 1967. The comparative ultrastructureand possible function of eyespots: Euglena granulata andChlamydomonas eugametos. Planta 77:325–53.
Yamaguchi, K., Prieto, S., Beligni, M. V., Haynes, P. A., McDonald,W. H., Yates III, J. R. & Mayfield, S. P. 2002. Proteomic char-acterization of the small subunit of Chlamydomonas reinhardtiichloroplast ribosome: identification of a novel S1 domain-con-taining protein and unusually large orthologs of bacterial S2,S3, and S5. Plant Cell 14:2957–74.
Zilber, A. L. & Malkin, R. 1992. Organization and topology ofphotosystem I subunits. Plant Physiol. 99:901–11.
Supplementary Material
The following supplementary material is availa-ble for this article:
Table S1. LC-ESI-MS/MS analysis of proteinspots 1–5 (see Fig. 1B).
Table S2. LC-ESI-MS/MS analysis of proteinspots a–f (see Fig. 1A).
This material is available as part of the online art-icle from: http://www.blackwell-synergy.com/doi/abs/10.1111/j.1529-8817.2007.00331.x
(This link will take you to the article abstract).
Please note: Blackwell Publishing is not respon-sible for the content or functionality of any supple-mentary materials supplied by the authors. Anyqueries (other than missing material) should be di-rected to the corresponding author for the article.
MELANIE SCHMIDT ET AL.294
Related Documents











