J. Great Lakes Res. 22(2):264-279 Internat. Assoc. Great Lakes Res., 1996 Evidence for a Reduction of 2,3,7,8-TCDD Toxic Equivalent Concentrations in White Sucker (Catostomus commersoni) Exposed to Bleached Kraft Pulp Mill Emuent, Following Process and Treatment Improvements Michael R. van den Heuvel,l* Mark R. Servos,2 Kelly R. Munkittrick,2 Niels C. Bols,l and D. George Dixon l 1University of Waterloo Department of Biology Waterloo, Ontario N2L 3GI 2Department of Fisheries and Oceans Great Lakes Laboratory for Fisheries and Aquatic Sciences Department of Fisheries and Oceans 867 Lakeshore Rd., P.o. Box 5050 Burlington, Ontario L7R 4A6 ABSTRACT. Over a 4-year period from 1990 to 1993, the H4/lE bioassay was used to measure 2,3,7,8-tetrachlorodibenzo-p-dioxin toxic equivalent concentrations (TECs) in the liver tissue of white sucker exposed to bleached kraft mill effluent. During two of the sampling periods, the H4/lE bioassay- derived TECs (BD-TECs) were compared directly with BD-TECs measured by a similar bioassay using a rainbow trout liver cell line (RTL-W1). From 1989 to 1993, chemistry-derived TECs (CD-TECs) were calculated directly from measured polychlorinated dibenzo-p-dioxin (PCDD) and dibenzofuran (PCDF) concentrations in liver tissue. Chemistry-derived TECs were compared to bioassay-derived TECs using toxic equivalentfactors (TEFs) developed for these systems. During the study period, the pulp mill began secondary treatment of effluent and increased the substitution of chlorine dioxide for molecular chlorine during bleaching. Significant reductions in H4/lE BD-TECs and CD-TECs were observed in liver tissue of both male and female white sucker during summer and fall sampling periods but not during spring sampling periods. Process and treatment improvements at this particular bleached kraft pulp mill appear to have been successful at reducing the levels of PCDDs and PCDFs found in white sucker exposed to the effluent. Despite the decreases in TECs, relative mixed function oxygenase (MFa) induction in the exposed white sucker did not decrease over the study period, suggesting the PCDDs and PCDFs are cur- rently not the dominant MFa-inducing compounds in the effluent of this mill. INDEX WORDS: Kraft paper, white sucker, TCDD, water treatment, bioassay. INTRODUCTION Since 1948, Jackfish Bay, Lake Superior, has re- ceived effluent from a pulp mill located in Terrace Bay, Ontario; this mill has been subject to intense scientific and public scrutiny. The Terrace Bay mill was singled out by Greenpeace (Van Strum and Merrell 1987) to illustrate the potential threat of chlorinated dioxin and furan pollution and to spear- head the continuing lobby against the industrial use •Author to whom correspondence should be addressed 264 of chlorine. In response to environmental concern and effluent toxicity, the mill installed secondary effluent treatment, in the form of an aerated stabi- lization basin (10 d retention) which became opera- tional in October, 1989. In 1990 the mill began to increase the substitution of chlorine dioxide in place of molecular chlorine in its bleaching process, a move designed to reduce the release of organochlorines (Karl 1992). The solutions imple- mented by the mill have greatly improved effluent quality; there has been a 92% reduction in biologi- cal oxygen demand (BOD), a 31 % reduction in lev-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Great Lakes Res. 22(2):264-279Internat. Assoc. Great Lakes Res., 1996
Evidence for a Reduction of 2,3,7,8-TCDD Toxic EquivalentConcentrations in White Sucker (Catostomus commersoni)
Exposed to Bleached Kraft Pulp Mill Emuent,Following Process and Treatment Improvements
Michael R. van den Heuvel,l* Mark R. Servos,2 Kelly R. Munkittrick,2 Niels C. Bols,l and D. George Dixonl
1University ofWaterlooDepartment ofBiology
Waterloo, Ontario N2L 3GI
2Department ofFisheries and OceansGreat Lakes Laboratory for Fisheries and Aquatic Sciences
Department ofFisheries and Oceans867 Lakeshore Rd., P.o. Box 5050
Burlington, Ontario L7R 4A6
ABSTRACT. Over a 4-year period from 1990 to 1993, the H4/lE bioassay was used to measure2,3,7,8-tetrachlorodibenzo-p-dioxin toxic equivalent concentrations (TECs) in the liver tissue of whitesucker exposed to bleached kraft mill effluent. During two of the sampling periods, the H4/lE bioassayderived TECs (BD-TECs) were compared directly with BD-TECs measured by a similar bioassay using arainbow trout liver cell line (RTL-W1). From 1989 to 1993, chemistry-derived TECs (CD-TECs) werecalculated directly from measured polychlorinated dibenzo-p-dioxin (PCDD) and dibenzofuran (PCDF)concentrations in liver tissue. Chemistry-derived TECs were compared to bioassay-derived TECs usingtoxic equivalent factors (TEFs) developed for these systems. During the study period, the pulp mill begansecondary treatment of effluent and increased the substitution of chlorine dioxide for molecular chlorineduring bleaching. Significant reductions in H4/lE BD-TECs and CD-TECs were observed in liver tissueof both male and female white sucker during summer and fall sampling periods but not during springsampling periods. Process and treatment improvements at this particular bleached kraft pulp mill appearto have been successful at reducing the levels ofPCDDs and PCDFs found in white sucker exposed to theeffluent. Despite the decreases in TECs, relative mixed function oxygenase (MFa) induction in theexposed white sucker did not decrease over the study period, suggesting the PCDDs and PCDFs are currently not the dominant MFa-inducing compounds in the effluent ofthis mill.
INDEX WORDS: Kraft paper, white sucker, TCDD, water treatment, bioassay.
INTRODUCTION
Since 1948, Jackfish Bay, Lake Superior, has received effluent from a pulp mill located in TerraceBay, Ontario; this mill has been subject to intensescientific and public scrutiny. The Terrace Bay millwas singled out by Greenpeace (Van Strum andMerrell 1987) to illustrate the potential threat ofchlorinated dioxin and furan pollution and to spearhead the continuing lobby against the industrial use
•Author to whom correspondence should be addressed
264
of chlorine. In response to environmental concernand effluent toxicity, the mill installed secondaryeffluent treatment, in the form of an aerated stabilization basin (10 d retention) which became operational in October, 1989. In 1990 the mill began toincrease the substitution of chlorine dioxide inplace of molecular chlorine in its bleaching process,a move designed to reduce the release oforganochlorines (Karl 1992). The solutions implemented by the mill have greatly improved effluentquality; there has been a 92% reduction in biological oxygen demand (BOD), a 31 % reduction in lev-

2,3,7,8-TCDD TECs After Pulp Mill Improvements 265
els of suspended solids, a 40% reduction in adsorbable organic halide concentration (AOX), andacute toxicity has been eliminated based on 96 hrainbow trout acute lethality bioassays (Karl 1992).
Monitoring of wild fish populations at JackfishBay (McMaster et al. 1991; Munkittrick et al. 1991,1992a, 1992b, 1994) has demonstrated consistentsublethal responses. These studies have repeatedlyshown elevated levels of P4501A, a mixed functionoxygenase (MFO) enzyme, in wild white sucker(Catostomus commersoni), longnose sucker(Catostomus catostomus), and lake whitefish (Coregonus clupeajormis) in Jackfish Bay. Changes inwhole fish reproductive parameters observed inwhite sucker include increased age to maturity, reduced secondary sex characteristics, smaller gonads, and lower fecundity with age. Alteredbiochemical reproductive indicators include lowplasma steroid hormone concentrations (Munkittrick et al. 1991, McMaster et al. 1991), reduced invitro steroid hormone synthesis, lowered plasmagonadotropin concentration, and altered regulationof sex steroid biosynthesis (Van Der Kraak et al.1992) as compared to fish captured at two Lake Superior reference sites.
The induction of the P4501A enzyme is known tobe activated by compounds such as polychlorinateddibenzo-p-dioxins (PCDDs) and polychlorinateddibenzofurans (PCDFs) through a cellular receptor,the aryl hydrocarbon (Ah) receptor (Hahn andStegeman 1992). It is generally held that at concentrations reflecting those found in the environment,the majority, if not all, of the toxicological effectsof PCDDs, PCDFs, and coplanar polychlorinatedbiphenyls (PCBs) are mediated through the bindingof these compounds to the Ah receptor (Goldsteinand Safe 1989, Hahn and Stegeman 1992).P4501Al-induction is thus a sensitive biochemicalindicator of exposure to PCDDs, PCDFs, PCBs,and also polycyclic aromatic hydrocarbons (PAHs).P4501Al-induction may share a common mechanism with other more severe effects, including reproductive dysfunction (Bookstaff et al. 1990a,1990b; Moore et al. 1992; Mably et al. 1992).
Due to human health concerns, a great deal ofemphasis has been placed on reducing levels ofPCDDs and PCDFs released by bleached kraftmills. The release of PCDDs and PCDFs is alsothought to pose a threat to fish and wildlife. Biochemical changes observed in fish exposed to pulpmill effluents have been suggestive of a "dioxinlike" mode of toxic action (Rogers et al. 1989,Smith et ai. 1991, Munkittrick et ai. 1991). Al-
though there is limited evidence that the quantitiesof PCDDs and PCDFs released by bleached kraftmills are detrimental to biota, the highly toxic andpersistent nature of these chemicals has been sufficient to cause alarm. Elevated levels of these compounds have been well documented in biotaresident in Jackfish Bay (Sherman et al. 1990, Servos et al. 1994).
The large number of compounds that bind the Ahreceptor make toxicological risk assessment of environmental mixtures difficult. For example, thereare 75 PCDD and 135 PCDF congeners all with different toxic potencies. Toxicologists have sought tosimplify this problem by deriving the toxic potencyrelative to the most toxic congener, 2,3,7,8-tetrachlorodibenzo-p-dioxin (2,3,7,8-TCDD). This potency ratio is called a toxic equivalent factor (TEF)and has been determined using a variety of bioassays, including those with mammalian and fish celllines (Clemons et al. 1994) as well as whole organism testing (Parrott et ai. 1995) with P4501A induction as an endpoint. Assuming that compounds thatbind the Ah receptor act in an additive manner, thisallows derivation of a single dose for risk assessment, which is referred to as a 2,3,7,8-TCDD toxicequivalent concentration (TEC).
There are two 'current approaches for derivingTECs: chemistry-derived TECs (CD-TEC) andbioassay-derived TECs (BD-TEC). A CD-TEC iscalculated by multiplying the measured environmental concentration of each congener by its TEFand summing the individual contributions of eachcongener. In this study, the BD-TEC is measured bycomparing P4501A enzyme induction elicited by anenvironmental mixture to the induction potency of2,3,7,8-TCDD. A mammalian-based cell-culturebioassay, the H4IIE rat hepatoma bioassay, has frequently been used to measure the in vitro P4501Ainducing ability of extracts prepared from fish andwildlife tissue (Tillitt et ai. 1990, 1991; van denHeuvel et al. 1994, 1995). The BD-TEC has the advantage of integrating the biological potency of allof the Ah-receptor-binding compounds in a mixture, including those compounds that are not routinely measured by analytical chemistry methods.
For assessing fish exposure and risk, use of BDTECs measured with mammalian systems, and CDTECs that are dependent on mammalian-derivedTEFs, is a clear target for criticism in light of thedifferences observed between responses of fish andmammals to particular PCDDs, PCDFs, and PCBs.Both in vivo and in vitro studies have found significant differences between fish- and mammalian-

266 van den Heuvel et al.
based TEFs (Clemons et al. 1994, Parrott et al.1995, Walker and Peterson 1991). There is clearly aneed to integrate the biological potency of fish extracts using fish-based bioassays and TEFs in orderto validate the use of mammalian-based cell culturebioassays.
The goal of this work was to monitor the 2,3,7,8TCDD BD-TECs and also TECs calculated from directly measured PCDDs and PCDFs in the livertissue of white sucker captured in Jackfish Bay overseveral years. The time frame of this study encompassed the installation of secondary treatment andincreases in the level of chlorine dioxide substitution. Bioassay-derived TECs were measured alongwith TECs calculated (chemistry-derived TECs)from a combination of fish- and mammalian-basedTEFs and measured PCDD and PCDF concentrations. BD-TECs and CD-TECs were compared toliver P450lAl-related enzyme activity in exposedwhite sucker over the same collection period. TECsderived with a rainbow trout liver cell line, RTLWI, were measured in order to compare fish andmammalian (H4IIE) BD-TECs in the livers of whitesucker that were captured in the BKME receivingarea, and also in the livers of sucker that were artificially exposed to receiving water through cagingexperiments.
MATERIALS AND METHODS
Fish Sampling
During spring sampling periods from 1989through 1993, white sucker were caught inovernight hoopnet sets as they entered the spawningstream. Mature prespawning white sucker were captured in Sawmill Creek, the spawning stream adjacent to Jackfish Bay (Fig. 1). The only suitablespawning location adjacent to the bay, SawmillCreek, is not exposed to effluent. White suckerwere also captured from a reference location, theLittle Gravel River which flows into Mountain Bay,a small embayment on Nipigon Bay, Lake Superior.
During summer and fall sampling periods from1989 through 1993, fish were captured at a locationapproximately 250 m from where effluent fromBlackbird Creek enters Moberly Bay, the northernsection of Jackfish Bay. Fish were captured inovernight sets with 10.2 cm gill nets.
Captured fish were rendered unconscious by concussion prior to necropsy. Weight, fork length,gonad weight, and liver weight were recorded foreach fish, and an operculum was removed for
aging. Livers were excised and divided into twosubsamples. One gram of liver for MFO analysiswas placed in a 2 mL cryovial and immediatelyfrozen in liquid nitrogen. The remaining liver tissuewas wrapped in tin foil (previously fired at 400°C),stored at -20°C and retained for chemical or bioassay analysis.
Caging Experiments
Fish caging experiments were previously described (van den Heuvel et al. 1995). Briefly, inspring 1993, white sucker were transported fromthe capture site at Mountain Bay to a holding location in Sawmill Creek. Sucker from both SawmillCreek (BKME exposed) and Mountain Bay weretransported by boat to the effluent plume and suspended in the water column in wire mesh cages approximately 25 m from the mouth of BlackbirdCreek. Ten fish from each site remained caged inSawmill Creek as caging controls. For data presented in this study, caging control fish were sampled after 4 d and plume-exposed fish after 8 d.
Liver Enzyme Analysis
Over the years of study, hepatic P4501Al activitywas quantified using a variety of catalytic techniques, including benzo[a]pyrene hydroxylase(AHH), diphenyloxazole hydroxylase (PPO), and as7-ethoxyresorufin-O-deethylase (EROD). AHH wasperformed according to Nebert and Gelboin (1968).PPO was quantified using the techniques of Luxonet al. (1987). EROD analysis used a modification ofthe method of Pohl and Fouts (1980). Some of theMFO data utilized in this study have been previously published (McMaster et al. 1991; Munkittricket al. 1991, 1992b; Smith et al. 1991).
Chlorinated DioxinlFuran Analysis
Chlorinated dioxin and furan measurements usedin this manuscript were part of a larger long-termstudy of the white sucker population in JackfishBay and will be presented in detail elsewhere (M.R.Servos, Dept. of Fisheries and Oceans, Burlington,Ontario, Canada, unpublished data). Liver samplesfor congener-specific dioxin and furan analyseswere analyzed using the techniques of Huestis et al.(1995). Liver was ground in sodium sulphate,placed in glass columns, spiked with a mixture ofl3C dioxin and furan congeners, and extracted withmethylene chloride. The resulting extract wascleaned up by automated GPC followed by alumina

2,3,7,8-TCDD TECs After Pulp Mill Improvements 267
N
Lake Superior
FIG. 1. White sucker sampling locations at Jackfish Bay and Mountain Bay, Lake Superior.
chromatography. A final cleanup using carbon fibrechromatography isolated the planar compounds.Samples were analyzed using gas chromatographywith high resolution mass spectrometry. Concentrations were corrected for the recovery of the 13C labelled congeners.
Tissue Extraction and Sample Preparationfor CeU Culture Bioassays
Tissue extracts for analysis in the H4IIE andRTL-WI bioassay systems were extracted as previously described (van den Heuvel et ai. 1994). Liversamples were dried with sodium sulphate and soxhlet extracted with methylene chloride. Samplecleanup involved GPC to separate lipids followed bybasic alumina chromatography to remove highly
polar materials. Lipid content was estimated gravimetrically. The final extract was concentrated into adimethylsulfoxide (DMSO) carrier and serial dilutions were made from the final extract to yield atotal of seven doses. This cleanup regime was chosen to be highly non-selective for the compoundsthat are retained in the final extract. Previous validation (van den Heuvel et al. 1995) has demonstratedthat a wide range of PCDDs, PCBs, and PAHsdemonstrate excellent recovery using this technique.
H4IIE and RTL-Wl Bioassay Procedure
Rat hepatoma (H4IIE) cells were purchased fromthe American Type Culture Collection (ATCC #CRL 1548) and cultured using the medium described by Tillitt et ai. (1990). This medium con-

268 van den Heuvel et al.
sisted of Dulbecco's modified Eagle's medium(MEM) supplemented with L-glutamine (IX),MEM essential amino acids (1.5X), MEM non-essential amino acids (2X), MEM vitamins (1.5X),15% fetal bovine serum (FBS), and 0.05 mg mL-Igentamycin sulphate. Cells were maintained in 75cm2 tissue culture flasks in a humidified 5% CO2atmosphere at 37 ± 0.1 0C. The rainbow trout RTLWI cell line (Lee et ai. 1993) was maintained inLeibovitz's L-15 medium supplemented with 10%FBS, 100 units penicillin and 100 f.lg mL-I streptomycin. Cells were grown at 22 ± 1°C in 75 cm2 tissue culture flasks.
For both the cell bioassays, four 2,3,7,8-TCDD(Cambridge Isotope Laboratories) dose-effectcurves were run concurrently with every bioassay.The concentration of the 2,3,7,8-TCDD stock solution was confirmed using GC-MS. All values givenin this paper are based on measured concentrationsof the 2,3,7,8-TCDD standard. Each standard curveexposed cells to seven concentrations ranging between 0.122 and 31.2 pg well- I of 2,3,7,8-TCDD.The seven dilutions of the liver extracts tested hadconcentrations of 0.78, 1.56, 3.13, 6.25, 12.5, 25.0,50.0, and 100% of the original extract.
The H4IIE and RTL-WI bioassay were performed using modifications of the method ofKennedy et al. (1993). For H4IIE bioassays, cellswere plated in 48-well plates (Costar) at a densityof 50,000 cells per well in 0.5 mL of media. Wellswere dosed in triplicate 24 h after plating with either 2.5 f.lL of DMSO containing the extracts, or,2.5 f.lL of DMSO containing 2,3,7,8-TCDD standards. After a 48h incubation, the media was aspirated from the wells, cells were rinsed once withphosphate-buffered saline and EROD activity wasdetermined kinetically at 37°C. Reagent concentrations in the final reaction mixture were: 0.1 MHEPES pH 7.8; 0.5 mM NADPH; 5 mM MgC12;
16 f.lM dicoumarol, and 1.5 f.lM 7-ethoxyresorufin.At time 0 and 10 min. resorufin was measured fluorometrically using a 520 nm excitation filter and a590 nm emission filter. Protein was determined by amodified fluorescamine method against a BSAstandard (Lorenzen and Kennedy 1993). EROD incubation media was aspirated from the plate and thecells were allowed to swell in 250 f.lL of distilledwater. Plates were then frozen to lyse the cells.After thawing, 500f.lL of 0.5 M potassium phosphate buffer, pH 8.0 was added to the wells followed by 250 f.lL of fluorescamine (O.3mg mL-I inacetone). Plates were read with a 340 nm excitation
filter and 420 nm emission filter 5 min after the addition of fluorescamine.
The RTL-W1 bioassay was performed only forextracts from the October 1991 sampling periodand the May 1993 caging experiment. Modifications of the bioassay procedures were made to compensate for the slower growth of RTL-WI cells.The bioassays were performed in a similar mannerexcept that cells were plated using one confluent75 cm2 flask per three 48-well tissue culture plates.RTL-WI cells were allowed to grow to confluency(approximately 7 d) in the 48 well plates prior todosing. Cells were incubated for 72 h prior toEROD analysis at 22°C. The RTL-WI bioassay reaction mixture was identical to that used for H4IIEexcept that it did not contain dicoumarol, and the 7ethoxyresorufin concentration was 0.25 f.lM.
Median effective dose (ED50) and a slope parameter (y) for both the extract dilution curves anddioxin standard curves were determined by fitting alogistic function to the data (equation 1):
ERODMAX - BACKGROUNDEROD = BACKGROUND + 1+ Eyo(LOG(ED50)-LOG(DOSE)) (1)
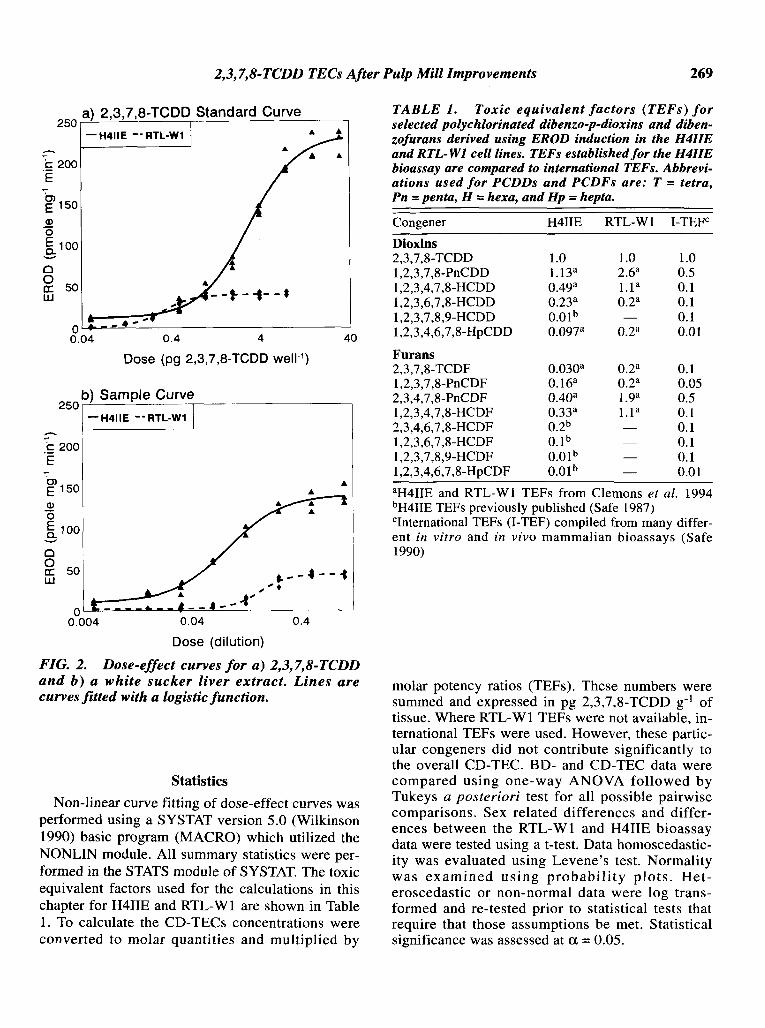
where EROD is the measured activity of each replicate in pmo1-mg protein-I min-I, BACKGROUNDis the activity of the DMSO blanks, and ERODMAX is the plateau of activity reached by the cells.DOSE is the dose given to the cells in units of pgwell- I for dioxin standards or dilution fraction(given that the most concentrated extract has a dilution of 1) for the liver extracts. A comparison of2,3,7,8-TCDD and liver extract curves for theH4IIE and RTL-W1 bioassay are shown in Figure2. Toxic equivalent concentration (TEC) of 2,3,7,8TCDD in pg g-I was derived from the ratio of2,3,7,8-TCDD standard curve ED50s to sample extract ED50s similar to Tillitt et ai. (1990).
Although lipid values were reported for BDTECs, the TECs were not corrected for lipid content in order to facilitate comparison with theCD-TECs for which lipid content dataset was incomplete. In addition, because lipid values did notshow any significant change over time, the lipidcorrected values showed the same trends as the uncorrected values that are presented. Since fish forwhich the bioassay-derived and the CD-TECs weremeasured did not show any significant changes inmean age over time; fish age was eliminated fromfurther consideration as a factor in the accumulationofTECs.
2,3,7,8-TCDD TECs After Pulp Mill Improvements 269
TABLE 1. Toxic equivalent factors (TEFs) forselected polychlorinated dibenzo-p-dioxins and dibenzofurans derived using EROD induction in the H411Eand RTL-WI cell lines, TEFs established for the H411Ebioassay are compared to international TEFs. Abbreviations used for PCDDs and PCDFs are,' T = tetra,Pn =penta, H =hexa, and Hp =hepta.
Congener H4IIE RTL-W1 I-TEF
Dioxins2,3,7,8-TCDD 1.0 1.0 1.0l,2,3,7,8-PnCDD 1.13a 2.6a 0.51,2,3,4,7,8-HCDD OA9a 1.1a 0.11,2,3,6,7,8-HCDD 0.23a 0.2a 0.11,2,3,7,8,9-HCDD O.Ol b 0.1
40 1,2,3,4,6,7,8-HpCDD 0.097a 0.2a 0.01
~a'L}~2~,3:..!.-,7~,~8---.:-T~C=-=D=-=D=,.=S~ta~n_d_a_rd_C_u_rv_e __~1250~
- H411E -- RTL-W1
04-.---",d..-~----------_---.J
0.04 0.4 4
-I:: 200
E
~150Q)
oE 100So~ 50w
Dose (pg 2,3,7,8-TCDD well-I)
-~£ 200E
~150Q)
oE 100So~ 50W
;::b!--}-=S:..=a::...:m-=..!p::..:.l-=-e-=C:....:u::..:.r~ve~ _250
- H411E _. RTL-W1
.. --"--i,,'.__ ..f"oLJ,~.........~'-"'-~--"'--"~=--~------
0.004 0.04 0.4
Dose (dilution)
FIG. 2. Dose-effect curves for a) 2,3,7,8-TCDDand b) a white sucker liver extract. Lines arecurves fitted with a logistic function.
Statistics
Non-linear curve fitting of dose-effect curves wasperformed using a SYSTAT version 5.0 (Wilkinson1990) basic program (MACRO) which utilized theNONLIN module. All summary statistics were performed in the STATS module of SYSTAT. The toxicequivalent factors used for the calculations in thischapter for H4IIE and RTL-Wl are shown in Table1. To calculate the CD-TECs concentrations wereconverted to molar quantities and multiplied by
Furans2,3,7,8-TCDF 0.030a 0.2a 0.11,2,3,7,8-PnCDF 0.16a 0.2a 0.052,3,4,7,8-PnCDF OAoa 1.9a 0.51,2,3,4,7,8-HCDF 0.33a 1.1a 0.12,3,4,6,7,8-HCDF 0.2b 0.11,2,3,6,7,8-HCDF O.lb 0.11,2,3,7,8,9-HCDF O.Ol b 0.11,2,3,4,6,7,8-HpCDF O.Ol b 0.01
aH4IIE and RTL-W1 TEFs from Clemons et al. 1994bH4IIE TEFs previously published (Safe 1987)CIntemational TEFs (I-TEF) compiled from many different in vitro and in vivo mammalian bioassays (Safe1990)
molar potency ratios (TEFs). These numbers weresummed and expressed in pg 2,3,7,8-TCDD g-l oftissue. Where RTL-WI TEFs were not available, international TEFs were used. However, these particular congeners did not contribute significantly tothe overall CD-TEC. BD- and CD-TEC data werecompared using one-way ANOVA followed byTukeys a posteriori test for all possible pairwisecomparisons. Sex related differences and differences between the RTL-Wl and H4IIE bioassaydata were tested using a t-test. Data homoscedasticity was evaluated using Levene's test. Normalitywas examined using probability plots. Heteroscedastic or non-normal data were log transformed and re-tested prior to statistical tests thatrequire that those assumptions be met. Statisticalsignificance was assessed at a =0.05.

270 van den Heuvel et at.
RESULTS
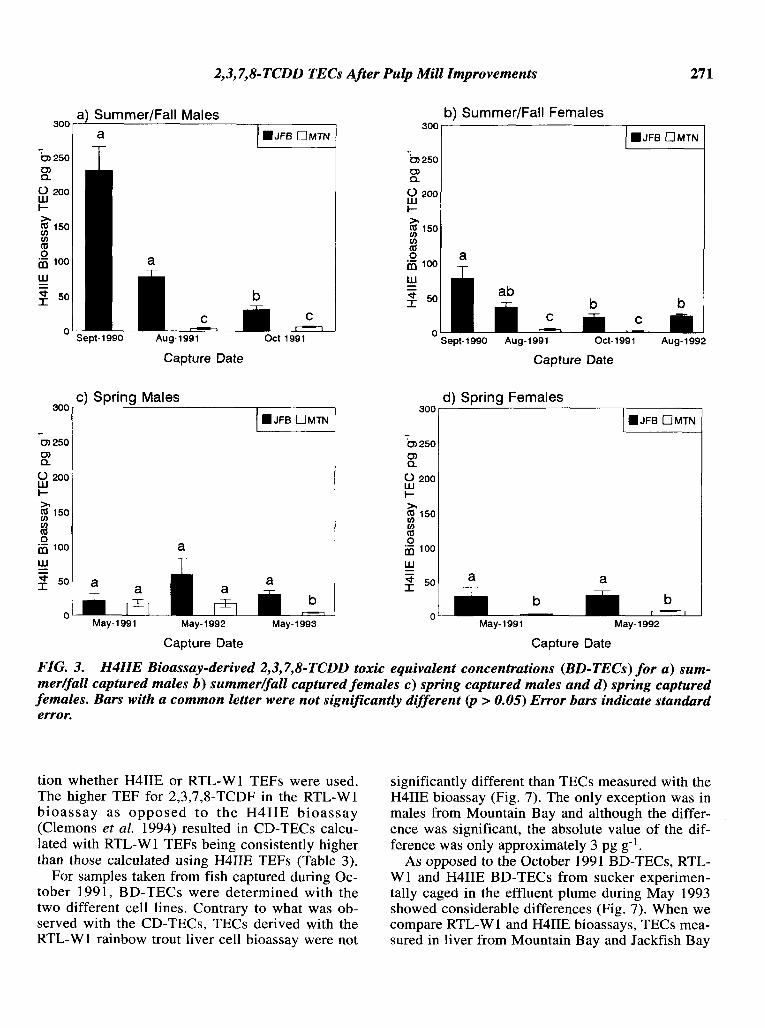
BD-TECs from summer/fall sampling periods(Table 2, Fig. 3) showed significant decreases beginning in 1991 for both male and female fish.Sucker captured in Sawmill Creek in the spring(Table 2, Fig. 3) did not show any pattern of increase or decrease. The TECs for the reference location were significantly lower than all comparableJackfish Bay samples with the exception of springcaptured males in 1991 and 1992; Mountain Baysamples were unusually high during these periods.
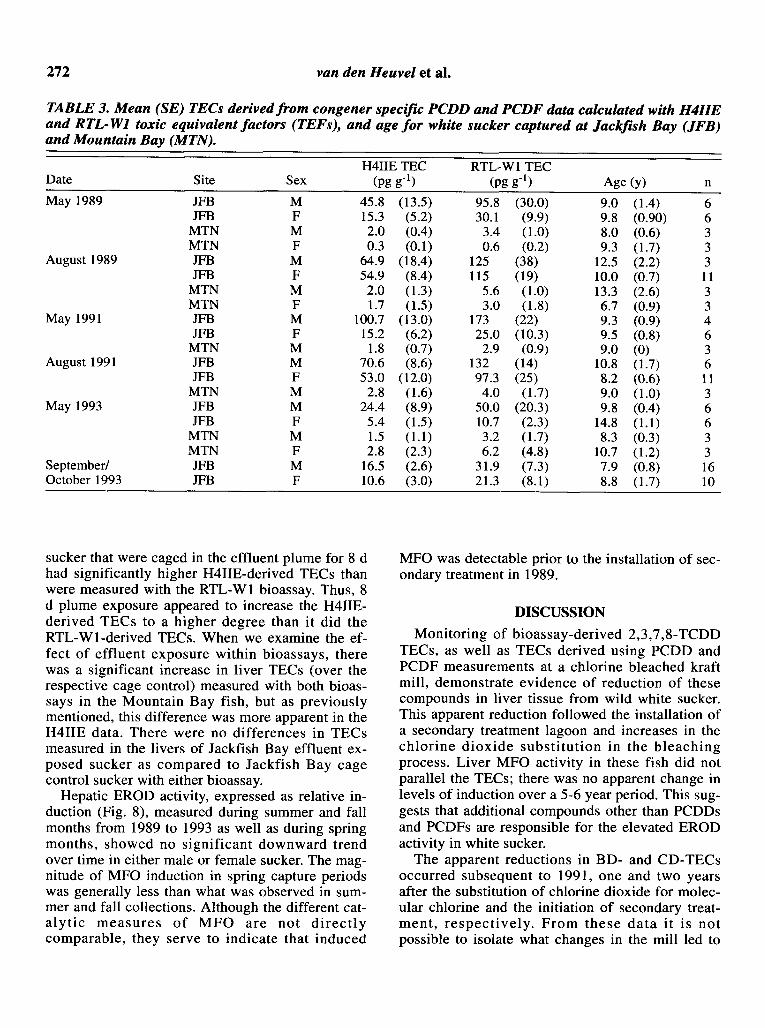
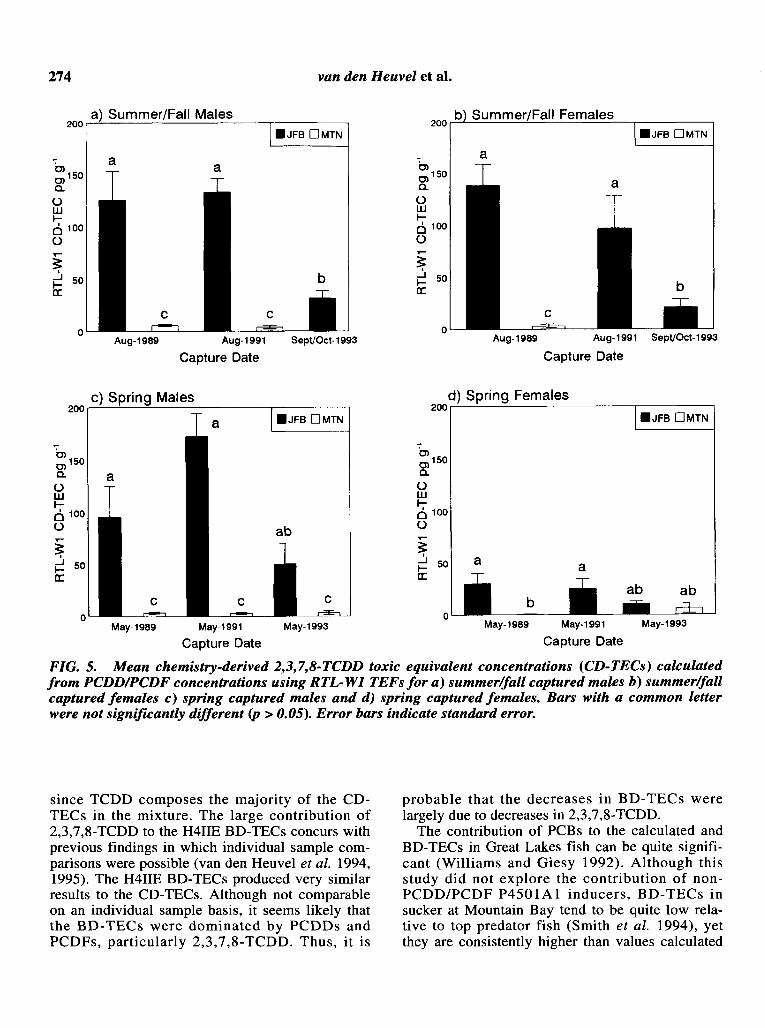
CD-TECs, using both H4IIE and RTL-Wl TEFs,decreased significantly in summer/fall capturedmales and females after 1991 (Table 3, Figs. 4 and5). CD-TECs as calculated with H4IIE and RTLWI TEFs in spring-captured male and femalesucker showed no significant differences, althoughthere is a downward trend for both sexes. TECs calculated from PCDDs and PCDFs in Mountain Baytissues were significantly lower than all JackfishBay TECs from the same group, with the exceptionof the spring captured females, where relatively lowlevels of CD-TECs were evident.
Direct comparison on an individual sample basisof BD-TECs with CD-TECs was not possible; there
were few sampling periods in common, and samples were usually taken from different fish duringthese periods. It was observed that BD- and CDTEC values for sucker livers from Jackfish Bayshowed a similar range for both techniques, 20-230pg g-l for BD-TECs and 18-173 pg g-l for CDTECs. However, the H4IIE bioassay generally revealed higher TEC values at the reference site thanwas indicated by CD-TECs.
Calculations of the relative contribution ofPCDD and PCDF congeners to the CD-TECs weremade by averaging the contribution of particularcongeners in individual samples (Fig. 6). For CDTECs calculated with both RTL-Wl and H4IIETEFs, there were no significant differences ortrends in the mean relative contribution of the congeners between collection dates. From 1989 to1993, 2,3,7,8-TCDD dominated the mixture in CDTECs calculated with H4IIE TEFs; 62.8% of theTEC was due to 2,3,7,8-TCDD with the majority ofthe remainder being split between 2,3,7,8-TCDFand 2,3,4,7,8-PnCDD. However, for CD-TECs calculated with RTL-Wl TEFs, 2,3,7,8-TCDF was thegreatest contributing congener in the mixture followed closely by 2,3,7,8-TCDD. The remainder ofthe congeners showed a similar pattern of contribu-
TABLE 2. Mean (SE) H4IIE bioassay-derived TEes, liver lipid content, and age for white sucker cap-tured at Jackfish Bay (JFB) and Mountain Bay (MTN).
Date Site Sex TEe (pg g-l) Lipid (%) Age (y) n
September 1990 JFB M 231.6 (45.8) 15.7 (4.4) 10.5 (0.5) 2JFB F 78.1 (17.4) 6.6 (1.1) 11.2 (0.6) 10
May 1991 JFB M 21.1 (9.9) 7.3 (1.9) 9.3 (0.3) 6JFB F 28.1 (14.2) 4.1 (0.4) 11.7 (1.0) 6
MTN M 17.8 (5.3) 11.7 (2.4) 9.2 (0.7) 5MTN F 2.1 (0.2) 4.2 (0.3) 8.6 (0.4) 5
August 1991 JFB M 77.6 (9.8) 14.7 (2.1) 12.2 (1.1) 4JFB F 36.4 (7.3) 10.3 (1.5) 9.6 (0.8) 12
MTN M 4.1 (1.5) 11.7 (1.8) 8.4 (0.5) 5MTN F 4.0 (1.8) 6.3 (1.2) 9.4 (0.9) 5
October 1991 JFB M 31.0 (5.8) 6.4 (0.7) 9.0 (0.8) 8JFB F 21.3 (5.4) 6.3 (1.4) 11.6 (1.4) 8
MTN M 6.5 (1.0) 8.2 (0.9) 11.1 (2.7) 8MTN F 2.5 (0.4) 5.1 (0.3) 8.1 (0.1) 8
May 1992 JFB M 59.6 (24.4) 13.2 (2.3) 12.5 (0.9) 4JFB F 30.5 (2.8) 7.0 (1.4) 11.8 (0.9) 5
MTN M 17.9 (5.7) 17.7 (3.9) 10.7 (2.80) 4MTN F 7.3 (1.1) 6.8 (0.2) 13.0 (0.9) 5
August 1992 JFB F 24.3 (2.6) 9.3 (0.9) 10.2 (0.8) 10May 1993 JFB M 31.0 (5.7) 9.1 (1.1) 10.3 (0.4) 10
MTN M 4.6 (1.1) 7.6 (1.0) 9.7 (0.3) 10
2,3,7,8-TCDD TECs After Pulp Mill Improvements 271
.JFB DMTN
a) Summer/Fall Males300 r-'-------'---------r--------,
a
b) Summer/Fall Females300,------------,--------,
.JFB DMTN
Capture Date
0>250Ola.0200WI->-<1l 150(J)(J)
<1loiii 100
W
~ 50I
oSept-1990 Aug-1991
Capture Date
Oct-1991
c
0>250Ola.0200wI->-m150
~o aiii 100
W
~ 50I
oSept-1990 Aug-1991 Oct-1991 Aug-1992
c) Spring Males300r-'--=--------=---------r--------,
.JFB DMTN
d) Spring Females300 ,--:_c-----" ---, -,
.JFB DMTN
0>250Ola.0200WI->-m150
~oiii 100
W
~ 50I
oMay-1991
a
May-1992
Capture Date
b
May-1993
0>250Ola.0200WI->-m150l/)<1loiii 100
W
~ 50I
o
a
May-1991
a
b
Capture Date
b
May-1992
FIG. 3. H4IIE Bioassay-derived 2,3,7,8-TCDD toxic equivalent concentrations (BD-TECs) for a) summer/fall captured males b) summer/fall captured females c) spring captured males and d) spring capturedfemales. Bars with a common letter were not significantly different (p > 0.05) Error bars indicate standarderror.
tion whether H4IIE or RTL-W1 TEFs were used.The higher TEF for 2,3,7,8-TCDF in the RTL-W1bioassay as opposed to the H4IIE bioassay(Clemons et ai. 1994) resulted in CD-TECs calculated with RTL-WI TEFs being consistently higherthan those calculated using H4IIE TEFs (Table 3).
For samples taken from fish captured during October 1991, BD-TECs were determined with thetwo different cell lines. Contrary to what was observed with the CD-TECs, TECs derived with theRTL-W1 rainbow trout liver cell bioassay were not
significantly different than TECs measured with theH4IIE bioassay (Fig. 7). The only exception was inmales from Mountain Bay and although the difference was significant, the absolute value of the difference was only approximately 3 pg g-l.
As opposed to the October 1991 BD-TECs, RTLWI and H4IIE BD-TECs from sucker experimentally caged in the effluent plume during May 1993showed considerable differences (Fig. 7). When wecompare RTL-WI and H4IIE bioassays, TECs measured in liver from Mountain Bay and Jackfish Bay
272 van den Heuvel et aI.
TABLE 3. Mean (SE) TECs derived from congener specific PCDD and PCDF data calculated with H4IIEand RTL-WI toxic equivalent factors (TEFs), and age for white sucker captured at Jackfish Bay (JFB)and Mountain Bay (MTN).
H4IIE TEC RTL-W1 TECDate Site Sex (pg g-l) (pg g-l) Age (y) n
May 1989 JFB M 45.8 (13.5) 95.8 (30.0) 9.0 (1.4) 6JFB F 15.3 (5.2) 30.1 (9.9) 9.8 (0.90) 6
MTN M 2.0 (0.4) 3.4 (1.0) 8.0 (0.6) 3MTN F 0.3 (0.1) 0.6 (0.2) 9.3 (1.7) 3
August 1989 JFB M 64.9 (18.4) 125 (38) 12.5 (2.2) 3JFB F 54.9 (8.4) 115 (19) 10.0 (0.7) 11
MTN M 2.0 (1.3) 5.6 (1.0) 13.3 (2.6) 3MTN F 1.7 (1.5) 3.0 (1.8) 6.7 (0.9) 3
May 1991 JFB M 100.7 (13.0) 173 (22) 9.3 (0.9) 4JFB F 15.2 (6.2) 25.0 (10.3) 9.5 (0.8) 6
MTN M 1.8 (0.7) 2.9 (0.9) 9.0 (0) 3August 1991 JFB M 70.6 (8.6) 132 (14) 10.8 (1.7) 6
JFB F 53.0 (12.0) 97.3 (25) 8.2 (0.6) 11MTN M 2.8 (1.6) 4.0 (1.7) 9.0 (1.0) 3
May 1993 JFB M 24.4 (8.9) 50.0 (20.3) 9.8 (0.4) 6JFB F 5.4 (1.5) 10.7 (2.3) 14.8 (Ll) 6
MTN M 1.5 (Ll) 3.2 (1.7) 8.3 (0.3) 3MTN F 2.8 (2.3) 6.2 (4.8) 10.7 (1.2) 3
September/ JFB M 16.5 (2.6) 31.9 (7.3) 7.9 (0.8) 16October 1993 JFB F 10.6 (3.0) 21.3 (8.1) 8.8 (1.7) 10
sucker that were caged in the effluent plume for 8 dhad significantly higher H4IIE-derived TECs thanwere measured with the RTL-WI bioassay. Thus, 8d plume exposure appeared to increase the H4IIEderived TECs to a higher degree than it did theRTL-W1-derived TECs. When we examine the effect of effluent exposure within bioassays, therewas a significant increase in liver TECs (over therespective cage control) measured with both bioassays in the Mountain Bay fish, but as previouslymentioned, this difference was more apparent in theH4IIE data. There were no differences in TECsmeasured in the livers of Jackfish Bay effluent exposed sucker as compared to Jackfish Bay cagecontrol sucker with either bioassay.
Hepatic EROD activity, expressed as relative induction (Fig. 8), measured during summer and fallmonths from 1989 to 1993 as well as during springmonths, showed no significant downward trendover time in either male or female sucker. The magnitude of MFO induction in spring capture periodswas generally less than what was observed in summer and fall collections. Although the different catalytic measures of MFO are not directlycomparable, they serve to indicate that induced
MFO was detectable prior to the installation of secondary treatment in 1989.
DISCUSSION
Monitoring of bioassay-derived 2,3,7,8-TCDDTECs, as well as TECs derived using PCDD andPCDF measurements at a chlorine bleached kraftmill, demonstrate evidence of reduction of thesecompounds in liver tissue from wild white sucker.This apparent reduction followed the installation ofa secondary treatment lagoon and increases in thechlorine dioxide substitution in the bleachingprocess. Liver MFO activity in these fish did notparallel the TECs; there was no apparent change inlevels of induction over a 5-6 year period. This suggests that additional compounds other than PCDDsand PCDFs are responsible for the elevated ERODactivity in white sucker.
The apparent reductions in BD- and CD-TECsoccurred subsequent to 1991, one and two yearsafter the substitution of chlorine dioxide for molecular chlorine and the initiation of secondary treatment, respectively. From these data it is notpossible to isolate what changes in the mill led to

2,3,7,8-TCDD TECs After Pulp Mill Improvements 273
a--.!..)_S_u_m_m---.::.e-.:..r/~F..:..:a..:..:II_M..:..:a..:..:le-=-s=----_,- ---,120.-
.JFB DMTN
b Summer/Fall Females120 ,...=-.<'-=-=..:..:.-'-'-'---'--'--C--C--"--'-"--'-C.C--"--'_-,--- --,
.JFB DMTN
100 100
Cl a~ 80
()WI- 60I
0()
W 40
~I
Aug-1989 Aug-1991
Capture Date
Sept/Oct-1993
en~ 80 a() awI-;' 600()
W 40
~I
c
Aug-1989 Aug-1991 Sept/Oct-1993
Capture Date
May-1993
a
May-1991
Capture Date
bMay-1989
a
.JFB OMTN
d) Spring Females120r..-----'---=---------,------,
100
o
20
en~ 80
()WI- 606()
W 40
~I
c
May-1993
.JFB OMTN
May-1991
Capture Date
May-1989
c) Spring Males
o
20
120 a
100
Cl
~ 80
()UJI-;' 60o()
W 40
~I
FIG. 4. Mean chemistry-derived 2,3,7,8-TCDD toxic equivalent concentrations (CD-TECs) calculatedfrom PCDDIPCDF data using H4IIE TEFs for a) summer/fall captured males b) summer/fall capturedfemales c) spring captured males and d) spring captured females. Bars with a common letter were not significantly different (p > 0.05). Error bars indicate standard error.
the decreases in TECs. However, decreases in theuse of molecular chlorine are believed to reduce theformation of PCDDs and PCDFs (Berry et at. 1989,Luthe et ai. 1994). In the chlorine bleaching ofpulp, PCDOs and PCDFs are believed to be formedfrom the parent molecules dibenzo-p-dioxin anddibenzofuran (Berry et ai. 1989) by chlorinationduring bleaching. Removal of the precursors, lowering the ratio of chlorine to pulp (chlorine multiple), and chlorine dioxide substitution, all serve to
reduce, or even eliminate the production of PCDDsand PCDFs. Mill data (Karl 1992) indicate that therelease of PCDDs and PCDFs in effluent havefallen below detection limits although those limitswere not stated. The effects of biological treatment(aerated lagoon) on the degradation of specificorganochlorines is unknown, but is very site- andseason-dependent (Graves and Joyce 1994).
The decreases in H4IIE BD-TECs noted herewere likely linked to decreases in 2,3,7,8-TCDD
274 van den Heuvel et at
.a..!.)_S_u_m_m_e_r/:..-F_a~II---:M~a:.-le_s__,- ----,200,--
b Summer/Fall Females200..---'----'--'--'-------,--------,
.JFB DMTN .JFB DMTN
Aug-1989 Aug-1991 Sept/Oct-1993
Capture Date
a
c
a0>0)150a.Uw~
0 100
U.-3:..,j
50~0::
0Sept/Oct-1993Aug-1989 Aug-1991
Capture Date
a
o
0>1500)a.Uw~
0 100
U
~..,j 50~0::
May-1993
a
b
a
o
.JFB DMTN
d) Spring Females200 ,----'---=----------.-----,
.-
~~ 500::
0>0)150a.Uw~
0 100
U
c
May-1989o
c) Spring Males200 ~-'----=:::'------------,-------,
0>0)150a.Uw~
0 100U.-
~~ 500::
May-1991 May-1993 May-1989 May-1991
Capture Date Capture Date
FIG. 5. Mean chemistry-derived 2,3,7,8-TCDD toxic equivalent concentrations (CD-TECs) calculatedfrom PCDDIPCDF concentrations using RTL-WI TEFs for a) summerIJall captured males b) summerIJallcaptured females c) spring captured males and d) spring captured females. Bars with a common letterwere not significantly different (p > 0.05). Error bars indicate standard error.
since TCDD composes the majority of the CDTECs in the mixture. The large contribution of2,3,7,8-TCDD to the H4IIE BD-TECs concurs withprevious findings in which individual sample comparisons were possible (van den Heuvel et aZ. 1994,1995). The H4IIE BD-TECs produced very similarresults to the CD-TECs. Although not comparableon an individual sample basis, it seems likely thatthe BD-TECs were dominated by PCDDs andPCDFs, particularly 2,3,7,8-TCDD. Thus, it is
probable that the decreases in BD-TECs werelargely due to decreases in 2,3,7,8-TCDD.
The contribution of PCBs to the calculated andBD-TECs in Great Lakes fish can be quite significant (Williams and Giesy 1992). Although thisstudy did not explore the contribution of nonPCDD/PCDF P4501A1 inducers, BD-TECs insucker at Mountain Bay tend to be quite low relative to top predator fish (Smith et aZ. 1994), yetthey are consistently higher than values calculated

2,3,7,8-TCDD TECs After Pulp Mill Improvements 275
Congener{s)
2,3,7,8-TCDD ~ ~-----'/.
MTN Female
*
MTN MaleJFB FemaleoJFB Male
.....bICI.e:30owI-'0~ 20.;:CDC>-:110IIIIIIoiii
a) October 199140,..-:------------------,
806040
H411E
80 60 40 20 0 20
Relative Contribution (%)
Other PCDFs
Other PCDDs
2,3,7,8-TCDF
2,3,4,7,8-PnCDD
b) May 1993 Caging StUdy80.------------------------,
say, differences between trout and sucker TEFscould also exist. Since sucker TEFs are not available, the use of rat and trout bioassays helps to define the range of TECs expected in sucker.
In contrast to the CD-TECs, RTL-WI BD-TECswere generally very similar to H4IIE BD-TECs.The reason for the similarity of bioassay TECs, de-
FIG. 7. Comparison of mean H411E and RTLWI bioassay-derived TECs (BD-TECs) Jor theOctober 1991 sampling period and the May 1993caging experiment. Asterisks indicate significantdifferences between bioassays (p < 0.05), crossesindicate significant differences (p < 0.05) between8 d plume exposed and 4 d cage control groupswithin a bioassay. Bars indicate SE.
......
ID RTL-W1 Bioassay I
*
JFB 8-day MTN control MTN 8-day
I_ H411E Bioassay
oJFB control
::'blCI.e:60owI-'0~ 40'0:CDC>-=20IIIIIIoiii
from measured PCDDs and PCDFs (Servos et af.1994, van den Heuvel et ai. 1994). This does suggest that unknown compounds, possible coplanarPCBs, contributed to the BD-TECs at the referencesite. However, this contribution generally rangedfrom 1 to 3 pg g-l making it relatively insignificantcompared to the TECs measured at Jackfish Bay.The TECs at Jackfish Bay likely share this smallcontribution of PCBs to the potency of the bioassaymixture. Comparing CD-TECs with BD-TECs isalso limited by the assumption of additivity - thisshould be kept in mind when assessing such data.
When the chemically determined concentrationsof PCDDs and PCDFs were used to calculate TECs,the CD-TECs differed, depending on whether H4IIEor RTL-Wl TEFs were used for the calculations.This difference was exclusively due to a higher TEFfor 2,3,7,8-TCDF derived with the RTL-Wl bioassay relative to the H4IIE bioassay (Clemons et af.1994). The higher TEF for 2,3,7,8-TCDF in tum ledto an increased calculated contribution of 2,3,7,8TCDF to the mixture, hence a higher CD-TEe. Although this suggests that the estimates ofAh-receptor binding compounds in sucker tissue isbetter calculated with TEFs for the fish cell bioas-
FIG. 6. Mean relative contribution oj chlorinated dioxin and Juran congeners to the overallchemistry-derived TECs (CD-TECs) in livers ofwhite sucker captured at lackJish Bay (n =90samples). CD-TECs are calculated either withH411E (dotted bars) toxic equivalent factors(TEFs) or with RTL- WI TEFs from thePCDDIPCDF concentrations measured over theperiod oj this study in white sucker liver tissue.
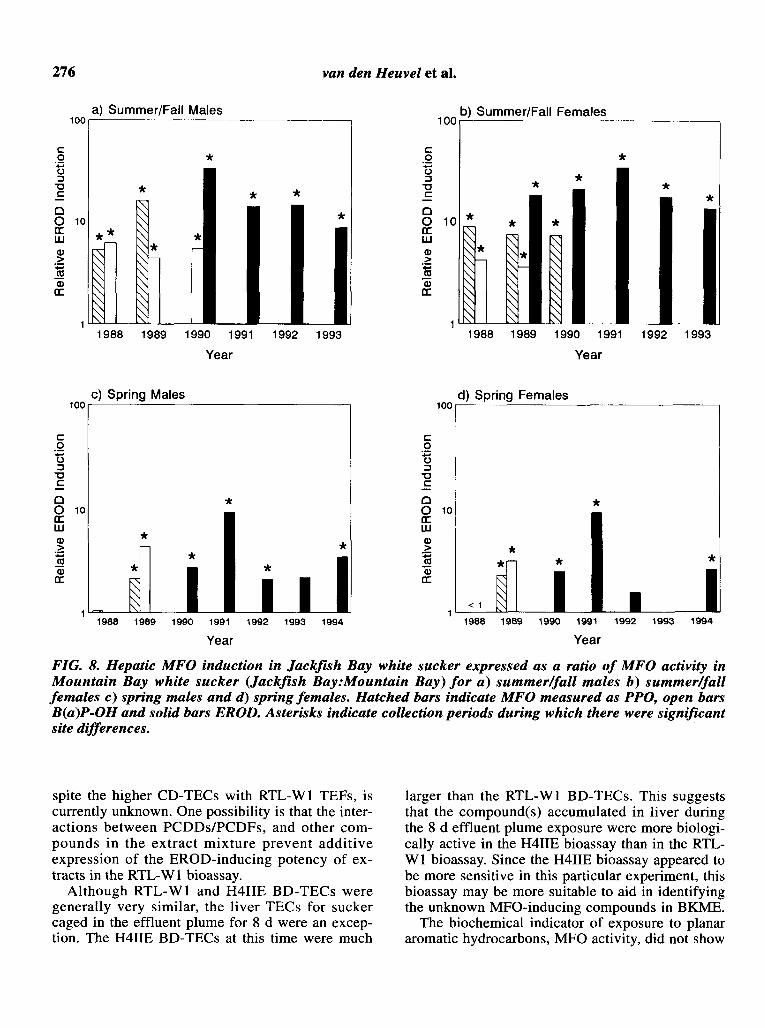
276 van den Heuvel et al.
*
*
*
**
*~~~~~~~~ ~ L..- L..-
1990 1991 1992 1993
Year
***
1989
**
< 1
1988
1988 1989 1990 1991 1992 1993 1994
Year
d) Spring Females100 r----'---=--------------,
b) Summer/Fall Females100,..e-------------------,
c::oU:J"0c::
oo 10II:UJQl>~Q)a:
-------_._~--
*
* * *[\*~
** ~ *r-
~*~r-
~"-~ ~~ ~~ ~~ t\' '-- '-- -
a) Summer/Fall Males100
1988 1989 1990 1991 1992 1993
Year
c) Spring Males100
c::.Q-u:J"0c::
0 *0 10II:UJQl *> *:;:::;
*.!!!* *Ql
II:
1988 1989 1990 1991 1992 1993 1994
Year
c::o
:;:::;U:J"0c::
oo 10II:UJQl>~QlII:
FIG. 8. Hepatic MFO induction in Jackfish Bay white sucker expressed as a ratio of MFO activity inMountain Bay white sucker (Jackfish Bay:Mountain Bay) for a) summerIJall males b) summerIJallfemales c) spring males and d) spring females. Hatched bars indicate MFO measured as PPO, open barsB(a)P-OH and solid bars EROD. Asterisks indicate collection periods during which there were significantsite differences.
spite the higher CD-TECs with RTL-WI TEFs, iscurrently unknown. One possibility is that the interactions between PCDDs/PCDFs, and other compounds in the extract mixture prevent additiveexpression of the EROD-inducing potency of extracts in the RTL-Wl bioassay.
Although RTL-WI and H4IIE BD-TECs weregenerally very similar, the liver TECs for suckercaged in the effluent plume for 8 d were an exception. The H4IIE BD-TECs at this time were much
larger than the RTL-WI BD-TECs. This suggeststhat the compound(s) accumulated in liver duringthe 8 d effluent plume exposure were more biologically active in the H4IIE bioassay than in the RTLWI bioassay. Since the H4IIE bioassay appeared tobe more sensitive in this particular experiment, thisbioassay may be more suitable to aid in identifyingthe unknown MFO-inducing compounds in BKME.
The biochemical indicator of exposure to planararomatic hydrocarbons, MFO activity, did not show

2,3,7,8-TCDD TECs After Pulp Mill Improvements 277
decreases that paralleled the measured TECs inwhite sucker at Jackfish Bay. In addition, BD- andCD-TECs from the livers of more recently capturedsucker both approach the threshold tissue concentration for MFO induction in rainbow trout of approximately 20 pg g-l (Parrott et at. 1995). Thesedata suggest that although PCDDs and PCDFs inthe liver extracts accounted for most of the ERODinduction in the bioassays, this may not be the casein wild white sucker exposed to BKME. The factthat the PCDDs and PCDFs dominate the bioassaymixture, despite the likely presence of other inducers suggests that the other inducers may be metabolized more rapidly than PCDDs and PCDFs.PAH-type compounds fit this profile. Recent evidence has shown that a large number of compounds, including non-priority PAHs, chlorinatedfluorenes and alkylfluorenes, are produced in theBKME mill processes (Koistenen et ai. 1994a,1994b).
The findings of this study agree with recentlypublished data that indicates that the presence ofnon-PCDD/PCDF MFO inducers in pulp mill effluent. Studies with fish exposed to effluents thoughexperimental caging have indicated that the rate ofEROD induction, and reduction upon removal toclean water, is very rapid (Courtenay et ai. 1993,Munkittrick et ai. 1995). These rapid kinetics arenot suggestive of PCDDs or PCDFs which arefound in effluent only at trace levels (low ppq). Asurvey conducted in Ontario, Canada, of whitesucker exposed to a large selection of pulp millsdemonstrated that male fish captured at mills thatdid not use significant amounts of chlorine had induced EROD despite levels of PCDDs and PCDFsin their liver tissue that were similar to uninducedreference populations (Munkittrick et ai. 1994, Servos et at. 1994). An experimental caging studyusing white sucker at Jackfish Bay measured increases in MFO inducing compounds in the livertissue of white sucker using the H4IIE bioassay;this was not accompanied by a concurrent increasein PCDDs and PCDFs (van den Heuvel et at. 1995).The weight of the experimental evidence in thisstudy and also in previous studies suggests thatMFO-inducing compounds other than PCDDs andPCDFs accumulate to sufficiently high levels to bebiochemically active in fish exposed to pulp mill effluents.
Measures implemented to remediate adverse affects of the pulp and paper mill receiving environment at Jackfish Bay have greatly improved thegross environmental conditions as well as appar-
ently reducing levels of PCDDs and PCDFs in biotaexposed to this receiving area. However, there hasyet to be improvement in the MFO induction observed at the site. These results suggest that currentimprovements may not be sufficient to removecompounds that have the potential to exert sublethaleffects on exposed biota. This requires further scientific work to identify these biologically activecompounds in pulp and paper effluent.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the assistance of Susan Huestis, Andrea Farwell, CameronPortt, Mark McMaster, and Leo and Mae Marchand. This work was supported by grants from theOntario Ministry of the Environment, a Natural Sciences and Engineering Research Council operatinggrant to D.G.D. and by the Canadian Department ofFisheries and Oceans.
REFERENCESBerry, R.M., Fleming, B.I., Voss, R.H., Luthe, C.E., and
Wrist, P.E. 1989. Towards preventing the formationof dioxins during chemical pulp bleaching. PulpPaper Can. 90:48-58.
Bookstaff, R.c., Kamel, F., Moore, R.W., Bjerke, D.L.,and Peterson, R.E. 1990a. Altered regulation of pituitary gonadotropin-releasing hormone (GNRH) receptor number and pituitary responsiveness to GHRH in2,3,7,8,-tetrachlorodibenzo-p-dioxin-treated male rats.Toxicol. Appl. Pharmacol. 105:78-92.
____, Moore, R.W., and Peterson, R.E. 1990b.2,3,7,8-tetrachloro-p-dioxin increases the potency ofandrogens and estrogens as feedback inhibitors ofluteinizing hormone secretion in male rats. Toxicol.Appl. Pharmacol. 104:212-224.
Clemons, J.H, van den Heuvel, M.R., Stegeman, J.J.,Dixon, D.G., and BoIs, N.C. 1994. A comparison oftoxic equivalent factors for selected dioxins andfurans congeners derived using fish and mammaliancell lines. Can. J. Fish. Aquat. Sci. 51:1577-1584.
Courtenay, S., Grunwald, C., Kreamer, G.-L., Alexander,R., and Wirgin, I. 1993. Induction and clearance ofcytochrome P4501A mRNA in Atlantic tomcod cagedin bleached kraft mill effluent in the Miramichi River.Aquat. Toxicol. 27:225-244.
Goldstein, J.A., and Safe, S. 1989. Mechanism of actionand structure-activity relationships for the chlorinateddibenzo-p-dioxins and related compounds. In Halogenated biphenyls, terphenyls, naphthalenes, dibenzodioxins and related products, 2nd Ed., ed. Kimbroughand Jensen, pp. 239-293. Amsterdam: Elsevier Science Publishers.
Graves, J.W., and Joyce, T.W. 1994. A critical review of

278 van den Heuvel et a1.
the ability of biological treatment systems to removechlorinated organics discharged by the paper industry.Water SA 20:155-160.
Hahn, M.E., and Stegeman, J. 1992. Phylogenetic distribution of the Ah receptor in non-mammalian species:implications for dioxin toxicity and Ah receptor evolution. Chemosphere 25:931-937.
Huestis, S.Y., Servos, M.R., Sergeant, D.B., Leggett, M.,and Dixon, D.G. 1995. Methods for determination oforganochlorine pesticides, polychlorinated biphenylcongeners and clorinated dibenzo-p-dioxins andfurans in fish. Can. J. Fish. Aquat. Sci. Tech. Rep.
Karl, W. 1992. The new Terrace Bay. Kimberly-Clarkturns over a new leaf at Ontario Mill. Pulp Paper J.91:43-46
Kennedy, S.W., Lorenzen, A., James, C.A., and Collins,B.T. 1993. Ethoxyresorufin-O- deethylase and porphyrin analysis in chicken embryo hepatocyte cultureswith a fluorescence multiwell plate reader. Anal.Biochem. 211: 102-112.
Koistinen, 1., Paasivirta, 1., Nevalainen, T., andLahtipera, M. 1994a. Chlorinated fluorenes and alkylfluorenes in bleached kraft pulp and pulp mill discharges. Chemosphere 28:2139-2150.
___, Paasivirta, J., Nevalainen, T., and Lahtipera,M. 1994b. Chlorophenanthrenes, alky1chlorophenanthrenes and alkylchloronaphthalenes in kraftpulp mill products and discharges. Chemosphere28: 1261-1277.
Lee, L.E.J., Clemons, J.H., Bechtel, D.G., Caldwell, S.J.,Han, K.-B., Pasitschniak-Arts, M., Mosser, D.D., andBoIs, N.C. 1993. Development and characterization ofa rainbow trout liver cell line expressing cytochromeP450-dependent monooxygenase activity. Cell BioI.Toxico!. 9:279-294.
Lorenzen, A., and Kennedy, S.W. 1993. A fluorescencebased protein assay for use with a microplate reader.Anal. Biochem. 214:346-348.
Luthe, C.E., Berry, R.M., and Voss, RH. 1994. Arechlorinated phenols precursors of bleach plant dioxins. Chemosphere 28:1883-1894.
Luxon, P.L., Hodson, P.V., and Borgmann, U. 1987.Hepatic aryl hydrocarbon hydroxylase activity of laketrout (Salve linus namaycush) as an indicator oforganic pollution. Environ. Toxicol. Chem. 6:649-658.
Mably, T.A., Bjerke, D.L., Moore, R.W., Gendron-Fitzpatrick, A., and Peterson, R.E. 1992. In utero and lactational exposure of male rats to 2,3,7 ,8,-tetrachlorodibenzo-p-dioxin 3. effects on spermatogenesisand reproductive capability. Toxico!. App!. Pharmacol. 114:118-126.
McMaster, M.E., van der Kraak, G.J., Portt, C.B.,Munkittrick, K.R., Sibley, P.K., Smith, I.R., andDixon, D.G. 1991. Changes in hepatic mixed-functionoxygenase (MFO) activity, plasma steroid levels andage at maturity of a white sucker (Catostomus com-
mersoni) population exposed to bleached kraft milleffluent. Aquat. Toxico!. 21:199-218.
Moore, R.W., Bookstaff, R.C., Mably, T.A., and Peterson, RE. 1992. Differential effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on responsiveness of malerats to androgens, 17B-estradiol, luteinizing hormone,gonadotropin-releasing hormone, and progesterone.Chemosphere 25:91-94.
Munkittrick, K.R., Portt, C.B., Van Der Kraak, G.J.,Smith, I.R., and Rokosh, D.A. 1991. Impact ofbleached kraft mill effluent on population characteristics, liver MFO activity, and serum steroid levels of aLake Superior white sucker (Catostomus commersoni)population. Can. J. Fish. Aquat. Sci. 48:1371-1380.
___, McMaster, M.E., Portt, C.B., Van Der Kraak,G.J., Smith, I.R, and Dixon, D.G. 1992a. Changes inmaturity, plasma sex steroid levels, hepatic mixedfunction oxygenase activity, and the presence ofexternal lesions in lake whitefish (Coregonus clupeaformis) exposed to bleached kraft mill effluent.Can. J. Fish. Aquat. Sci. 49:1560-1569.
___, Van Der Kraak, G.J., McMaster, M.E., andPortt, c.B. 1992b. Response of hepatic MFO activityand plasma sex steroids to secondary treatment ofbleached kraft pulp mill effluent and mill shutdown.Environ. Toxico!. Chem. 11:1427-1439.
___, Van Der Kraak, G.J., McMaster, M.E., Portt,C.B., van den Heuvel, M.R, and Servos, M.R. 1994.Survey of receiving water environmental impactsassociated with discharges from pulp mills II. Gonadsize, liver size, hepatic MFO activity and plasmasteroid levels in white sucker. Environ. Toxicol.Chem. 13:1089-1101.
___, Servos, M.R, Gorman, K., Blunt, B., McMaster, M.E., and Van Der Kraak, GJ. 1995. Characteristics of EROD induction associated with exposure topulp mill effluent. In Environmental Toxicology andRisk Assessment: Fourth Volume, T.W. LaPointe, F.T.Price, and E.E. Little [Eds.] ASTM, Philadelphia. INPRESS.
Nebert, D.W., and Gelboin, H.V. 1968. Substrateinducible microsomal aryl hydroxylase in mammaliancell culture I. assay and properties of inducedenzymes. J. Bio!. Chem. 243:6242-6249.
Parrott, J.L., Hodson, P.V., Servos, M.R., Huestis, S.L.,and Dixon, D.G. 1995. Relative potency of polychlorinated dibenzo-p-dioxins and dibenzofurans forinducing MFO activity in rainbow trout. Environ.Toxicol. Chem. 14:1041-1050.
Pohl, R.J., and Fouts, J.R. 1980. A rapid method forassaying the metabolism of 7-ethoxyresorufin bymicrosomal subcellular fractions. Anal. Biochem.107:150-155.
Rogers, I.H., Levings, C.D., Lockhart, W.L., andNorstrom, RJ. 1989. Observations on overwinteringjuvenile chinook salmon (Onchorhynchustshawytscha) expsed to bleached kraft mill effluent in

2,3,7,8-TCDD TECs After Pulp Mill Improvements 279
the Upper Fraser River, British Columbia. Chemosphere 19:1853-1869.
Safe. S. 1987. Determination of 2,3,7,8-TCDD toxicequivalent factors (TEFs): Support for the use of the invitro AHH induction"assay. Chemosphere 16:791-802.
___. 1990. Polychlorinated biphenyls (PCBs),dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs),and related compounds: environmental and mechanistic considerations which support the development oftoxic equivalency factors (TEFs). Crit. Rev. Toxicol.21:51-88.
Servos, M.R., Huestis, S., Whittle, D.M., and Munkittrick, K.R. 1994. Survey of receiving water environmental impacts associated with discharges form pulpmills III. Levels of dioxins and furans in muscle andliver as measured by GC/MS. Environ. Toxicol.Chem.13:1103-1115.
Sherman, R.K., Clement, R.E., and Tashiro, C. 1990.The distribution of polychlorinated dibenzo-p-dioxinsand dibenzofurans in Jackfish Bay, Lake Superior, inrelation to a kraft pulp mill effluent. Chemosphere20: 1641-1648.
Smith, I.R., Portt, c.B., and Rokosh, D.A. 1991. Hepaticmixed function oxidases induced in populations ofwhite sucker, (Catostomus commersoni), from areasof Lake Superior and the St. Marys River. J. GreatLakes Res. 17:382-393.
___, Marchant, B., van den Heuvel, M.R., Clemons,J.H., and Frimeth, J. 1994. Embryonic mortality,bioassay derived 2,3,7,8-tetrachlorodibenzo-p-dioxinequivalents, and organochlorine contaminants inPacific salmon from Lake Ontario. J. Great LakesRes. 20:497-509.
Tillitt, D.E., Giesy, J.P., and Ankley, G.T. 1990. Characterization of the H4IIE rat hepatoma cell bioassay as atool for assessing toxic potency of planar halogenatedhydrocarbons in environmental samples. Environ. Sci.Technol. 25:87-92.
___, Ankley, G.T., Verbrugge, D.A., Giesy, J.P.,Ludwig, J.P., and Kubiak, T.J. 1991. H4IIE rathepatoma cell bioassay-derived 2,3,7,8-tetrachlorodibenzo-p-dioxin equivalents in colonial fisheating waterbird eggs from the Great Lakes. Arch.Environ. Contam. Toxicol. 21 :91-101.
van den Heuvel, M.R., Munkittrick, K.R., Van DerKraak, G.J., McMaster, M.E., Portt, C.B., Servos,M.R., and Dixon, D.G. 1994. Survey of receivingwater environmental impacts associated with discharges from pulp mills. 4. Bioassay-derived 2,3,7,8tetrachlorodi benzo-p-dioxin toxic eq ui valentconcentration in white sucker (Catostomus commersoni) in relation to biochemical indicators. Environ.Toxicol. Chem. 13: 1117-1126.
___, Munkittrick, K.R., Van Der Kraak, G.J., Servos, M.R., and Dixon, D.G. 1995. Hepatic-7ethoxyresorufin-O-deethylase activity, plasma steroidhormone concentration and liver bioassay-derived2,3,7,8-TCDD toxic equivalent concentrations in wildwhite sucker (Catostomus commersoni) caged inbleached kraft pulp mill effluent. Can. J. Fish. Aquat.Sci. 52: 1339-1350.
Van Der Kraak, G.J., Munkittrick, K.R., McMaster,M.E., Portt, c.B., and Chang, J.P. 1992. Exposure tobleached kraft pulp mill effluent disrupts the pituitarygonadal axis of white sucker at multiple sites. Toxicol.Appl. Pharmacol. 115:224-233.
Van Strum, C., and Merrell, P. 1987. No margin ofsafety: A preliminary report on dioxin pollution andthe need for emergency action in the pulp and paperindustry. Greenpeace USA, Inc.
Walker, M.K., and Peterson, R.E. 1991. Potencies ofpolychlorinated dibenzo-p-dioxin, dibenzofuran, andbiphenyl congeners, relative to 2,3,7,8-tetrachlorodibenzo-p-dioxin, for producing early life stagemortality in rainbow trout (Oncorhynchus mykiss).Aquat. Toxicol. 21:219-238.
Wilkinson, L. 1990. SYSTAT: The System for Statistics.Evanston IL: SYSTAT Inc.
Williams, L.L., and Giesy, J.P. 1992. Relationshipsamong concentrations of individual polychlorinatedbiphenyl (PCB) congeners, 2,3,7,8-tetrachlorodibenzo-p-dioxin equivalents (TCDD-EQ), and rearing mortality of chinook salmon (Oncorhynchustshawytscha) eggs from Lake Michigan. J. GreatLakes Res. 18:108-124.
Submitted: 18 August 1995Accepted: 26 January 1996
Related Documents











