Evaluation of predation risk by a caterpillar using substrate-borne vibrations IGNACIO CASTELLANOS * & PEDRO BARBOSA † *Centro de Investigaciones Biolo ´ gicas, Universidad Auto ´ noma del Estado de Hidalgo yDepartment of Entomology, University of Maryland, College Park (Received 5 July 2004; initial acceptance 17 September 2005; final acceptance 8 February 2006; published online 7 July 2006; MS. number: A9929) Assessment of predation risk and appropriate defensive responses are critical for most organisms. Prey in- dividuals must distinguish between the cues produced by abiotic factors, competitors and predators, and respond appropriately. We determined whether larvae of Semiothisa aemulataria (Geometridae), which de- fend themselves from invertebrate predators by hanging on a silk thread, can detect predator threats and distinguish between different predators, other herbivores and abiotic factors. Furthermore, we determined the mechanism used by the caterpillars to detect predator threats and the degree to which predation risk was specified. The hanging behaviour of S. aemulataria caterpillars occurred in response to invertebrate predators (wasps and stink bugs) but not in response to birds, other herbivores or abiotic factors. Caterpil- lars distinguish these predators by perceiving their substrate-borne vibrations. These vibrations differed both qualitatively and quantitatively. Caterpillars did not hang in response to mechanically reproduced vibration stimuli representing herbivores, but they did show this defensive behaviour in response to vibra- tion stimuli that duplicated invertebrate predator signals. Caterpillars responded differently to predatory wasps and stink bugs. The length of the silk thread produced by caterpillars in response to foraging wasps was significantly greater than that produced in response to stink bugs. These predator-specific responses led to increased survival. Our results show that invertebrate prey are able to distinguish among the cues produced by abiotic factors, competitors and predators and respond specifically, in accordance with the potential risk posed by a predator. Ó 2006 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. Communication (sensu Markl 1983) between predator and prey individuals determines the outcome of predator- eprey interactions. Most research has focused on what predators perceive and how their responses to prey signals influence predatoreprey interactions. Yet the nature and consequences of communication between predator and prey depend not only on what information is conveyed to the predator, but what, if any, information is perceived and processed by prey. Only recently have studies begun to examine in detail the signals that mediate predatore prey communication and, in particular, the characteristics of the signal that provide information useful in predation risk assessment (Dicke & Grostal 2001; Djemai et al. 2001). To distinguish between abiotic and biotic relevant cues, prey must attend to specific characteristics of signals (Stephens & Krebs 1986). Furthermore, since organisms in natural environments usually encounter a number of different types of predators, and all predators are not equally effective at prey perception and capture (Eisner & Dean 1976; Bauer et al. 1977; Berenbaum et al. 1992), prey individuals must attend to characteristics of signals that give information on the type of risk posed by the sig- naller. The specificity of the information in the signals used by invertebrate prey in a multiple-predator environ- ment is poorly known for most predatoreprey interactions (but see Viitasalo et al. 1998; McIntosh & Peckarsky 1999), and experimental evidence that prey benefit (in terms of survival) by adopting different responses to different pred- ators appears to be lacking. To assess predation risk, invertebrate prey may use chemical (Tollrian & Von Elert 1994; Von Elert & Pohnert 2000), visual (Nalbach 1990; Land & Layne 1995) or me- chanical (i.e. air and substrate-borne vibrations; Markl & Tautz 1975; Plummer & Camhi 1981; Gnatzy & Ka ¨mper Correspondence: I. Castellanos, Centro de Investigaciones Biolo ´gicas, Uni- versidad Auto ´noma del Estado de Hidalgo, Apartado Postal 69-1, Pachuca, Hidalgo, 42001, Me ´xico (email: [email protected]). P. Barbosa is at the Department of Entomology, University of Maryland, College Park, MD 20742, U.S.A. 461 0003e3472/06/$30.00/0 Ó 2006 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. ANIMAL BEHAVIOUR, 2006, 72, 461e469 doi:10.1016/j.anbehav.2006.02.005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ANIMAL BEHAVIOUR, 2006, 72, 461e469doi:10.1016/j.anbehav.2006.02.005
Evaluation of predation risk by a caterpillar using
substrate-borne vibrations
IGNACIO CASTELLANOS* & PEDRO BARBOSA†
*Centro de Investigaciones Biologicas, Universidad Autonoma del Estado de Hidalgo
yDepartment of Entomology, University of Maryland, College Park
(Received 5 July 2004; initial acceptance 17 September 2005;
final acceptance 8 February 2006; published online 7 July 2006; MS. number: A9929)
Assessment of predation risk and appropriate defensive responses are critical for most organisms. Prey in-dividuals must distinguish between the cues produced by abiotic factors, competitors and predators, andrespond appropriately. We determined whether larvae of Semiothisa aemulataria (Geometridae), which de-fend themselves from invertebrate predators by hanging on a silk thread, can detect predator threats anddistinguish between different predators, other herbivores and abiotic factors. Furthermore, we determinedthe mechanism used by the caterpillars to detect predator threats and the degree to which predation riskwas specified. The hanging behaviour of S. aemulataria caterpillars occurred in response to invertebratepredators (wasps and stink bugs) but not in response to birds, other herbivores or abiotic factors. Caterpil-lars distinguish these predators by perceiving their substrate-borne vibrations. These vibrations differedboth qualitatively and quantitatively. Caterpillars did not hang in response to mechanically reproducedvibration stimuli representing herbivores, but they did show this defensive behaviour in response to vibra-tion stimuli that duplicated invertebrate predator signals. Caterpillars responded differently to predatorywasps and stink bugs. The length of the silk thread produced by caterpillars in response to foraging waspswas significantly greater than that produced in response to stink bugs. These predator-specific responsesled to increased survival. Our results show that invertebrate prey are able to distinguish among the cuesproduced by abiotic factors, competitors and predators and respond specifically, in accordance with thepotential risk posed by a predator.
� 2006 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Communication (sensu Markl 1983) between predatorand prey individuals determines the outcome of predator-eprey interactions. Most research has focused on whatpredators perceive and how their responses to prey signalsinfluence predatoreprey interactions. Yet the nature andconsequences of communication between predator andprey depend not only on what information is conveyedto the predator, but what, if any, information is perceivedand processed by prey. Only recently have studies begunto examine in detail the signals that mediate predatoreprey communication and, in particular, the characteristicsof the signal that provide information useful in predationrisk assessment (Dicke & Grostal 2001; Djemai et al. 2001).
Correspondence: I. Castellanos, Centro de Investigaciones Biologicas, Uni-versidadAutonoma del Estado de Hidalgo,Apartado Postal 69-1,Pachuca,Hidalgo, 42001, Mexico (email: [email protected]). P. Barbosais at the Department of Entomology, University of Maryland, CollegePark, MD 20742, U.S.A.
460003e3472/06/$30.00/0 � 2006 The Association for the S
To distinguish between abiotic and biotic relevant cues,prey must attend to specific characteristics of signals(Stephens & Krebs 1986). Furthermore, since organismsin natural environments usually encounter a number ofdifferent types of predators, and all predators are notequally effective at prey perception and capture (Eisner &Dean 1976; Bauer et al. 1977; Berenbaum et al. 1992),prey individuals must attend to characteristics of signalsthat give information on the type of risk posed by the sig-naller. The specificity of the information in the signalsused by invertebrate prey in a multiple-predator environ-ment is poorly known for most predatoreprey interactions(but see Viitasalo et al. 1998; McIntosh & Peckarsky 1999),and experimental evidence that prey benefit (in terms ofsurvival) by adopting different responses to different pred-ators appears to be lacking.
To assess predation risk, invertebrate prey may usechemical (Tollrian & Von Elert 1994; Von Elert & Pohnert2000), visual (Nalbach 1990; Land & Layne 1995) or me-chanical (i.e. air and substrate-borne vibrations; Markl &Tautz 1975; Plummer & Camhi 1981; Gnatzy & Kamper
1tudy of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.

ANIMAL BEHAVIOUR, 72, 2462
1990) information. In terrestrial arthropods, substrate-borne vibrations may mediate courtship behaviour (Marti-nez-Wells & Henry 1992), coordinate and maintain socialstructures (Roces et al. 1993; Kirchner et al. 1994, Cocroft1996), maintain symbiotic relationships (DeVries 1990),mediate competitive interactions (Yack et al. 2001), aswell as facilitate finding food (Pfannenstiel et al. 1995)and mates (Cokl et al. 1999), and detecting predatorsand parasitoids (Markl & Tautz 1975; Djemai et al.2001). More important with regard to risk assessment,substrate-borne vibrations may allow prey to differentiatepredators from nonpredators (Hergenroder & Barth 1983).
Larval Semiothisa aemulataria Walker (Geometridae)hang from silk threads to avoid invertebrate predatorswalking on a leaf (I. Castellanos, personal observation).Therefore, we investigated the potential importance ofsubstrate-borne vibrations produced by predators andthe specificity of prey response to these vibrations. Wealso examined whether the reduced sensory systems typi-cal of insect holometabolous larvae (Scoble 1992) allowfor complex signal discrimination. Specifically, we deter-mined whether S. aemulataria responds differentially tosignals produced by abiotic factors, invertebrate herbi-vores, invertebrate predators and a vertebrate predator.Furthermore, we determined whether signals producedby each of these factors are distinct and thus provide thebasis for predation risk assessment.
To test these hypotheses, we (1) quantified the hangingbehaviour of larvae of S. aemulataria in response to differ-ent predators, herbivores and abiotic factors, (2) character-ized the substrate-borne vibrations produced by thesefactors and experimentally determined the signals towhich S. aemulataria responded (in the absence of theagent that originally produced the signal) and (3) manip-ulated the behavioural responses of S. aemulataria to deter-mine whether differential responses of caterpillars, againstdifferent predators, enhanced survival.
METHODS
Study Taxa
Herbivore speciesSemiothisa aemulataria is a specialist on Acer species in
Central Maryland and has about three generations peryear (P. Barbosa, unpublished data). All instars of S. aemu-lataria are leaf vein and petiole mimics. Hanging behav-iour seems to occur after leaf-mimicry has failed (i.e.once a predator is quite close). Females collected fromthe field were allowed to oviposit in the laboratory and lar-vae were reared individually in 237-ml plastic containerson Acer negundo L. (Aceraceae) (box elder) foliage.
The other herbivores used in the experiments were Or-gyia leucostigma J. E. Smith (Lymantriidae), Oncometopiaorbona Fabricius (Cicadellidae) and Acanalonia conica Say(Acanaloniidae). Orgyia leucostigma is a polyphagous, ex-ternal solitary feeder and has two generations per year(Payne 1917; Barbosa et al. 2003). Oncometopia orbonahas one generation per year and it feeds on a wide vari-ety of herbaceous and woody plant species. Acanalonia
conica feeds on a wide variety of herbaceous plant anddeciduous tree species and is univoltine (Wilson &McPherson 1981).
Predator speciesThe predators included the tufted titmouse, Parus bicolor
L. (Paridae), a visually hunting predator of caterpillars andone of the most abundant avian predators in our study site(Beal et al. 1941; Bent 1946; Medina 1999). This species isan omnivore with a diet made up mainly of caterpillars,but including other arthropods (Beal et al. 1941; Bent1946). Tufted titmice typically attack prey items usinga gleaning manoeuvre that consists of flights betweenbranches and jumps along branches, taking stationaryprey items after landing on a branch (Robinson & Holmes1982; Grubb & Pravosudov 1994). Parus bicolor individualsused in this study were captured in a wooded area inGlenn Dale, Maryland, following the protocol in IACUCPermit No. R-02-38.
Two invertebrate caterpillar predators were used, thepaper wasp Polistes fuscatus Fabricius (Vespidae) and thestink bug Podisus maculiventris Say (Pentatomidae) (Rabb1960; McPherson 1982; Gould & Jeanne 1984; Stamp1992). These two species actively search for prey and oftendetect prey items at very close distances or even afterphysically contacting prey (Evans 1982; Stamp 1992).
Adults of P. fuscatus feed on floral nectars and othersweet exudates, but are major predators of caterpillars,which they capture and process to feed their larvae (Mich-ener & Michener 1951; Rabb 1960; Gould & Jeanne 1984;Raveret Richter 2000). While foraging, wasps activelysearch for prey items by hovering around and walkingon plants (Stamp & Wilkens 1993). To date, P. fuscatushas been assumed to detect prey items visually or chemi-cally. After detection, often at very close distances whilewalking on plants, wasps pounce upon prey (Stamp 1992).
Wasps used in this research were obtained from nestscollected from the Patuxent Wildlife Refuge ResearchCenter, Patuxent, Maryland, in early spring. Each colonywas provided daily with fresh water, honey, and approx-imately five to 20 live lymantriid, noctuid, arctiid, geo-metrid and tenebrionid larvae, as the colony grew.
The stink bug P. maculiventris is a generalist predatorthat feeds primarily on larval Lepidoptera and Coleoptera(McPherson 1982). It actively searches for prey while walk-ing on the foliage of plants, responding to prey within dis-tances of a few millimetres or after physically contactingthe prey (Evans 1982). Podisus maculiventris used in the ex-periments originated from adults collected in early springand were provided with live lymantriid, noctuid, arctiid,geometrid and tenebrionid larvae, water and green beans(as in Mallampalli et al. 2002).
All herbivores and predators were collected from A. ne-gundo trees at the Patuxent Wildlife Refuge Research Cen-ter. Invertebrate predators were found in a communitystudy in which the canopy of A. negundo trees in the Pa-tuxent Wildlife Refuge Research Center was fogged (P. Bar-bosa, I. Castellanos & A. E. Segarra, unpublished data). Allspecies were maintained in the laboratory at room temper-ature (25 � 2 �C).

CASTELLANOS & BARBOSA: CATERPILLAR RISK ASSESSMENT 463
Experimental Protocol
Quantification of response to predators, herbivoresand abiotic factors
To quantify the hanging response of S. aemulataria inthe presence of invertebrate predators, herbivores and abi-otic factors, we exposed individual fourth instars to a sin-gle predator, herbivore or abiotic factor. For each trial,a fourth instar was placed on the leaf of a 15-cm A. ne-gundo branch positioned 1.5 m above the ground andallowed to acclimate for half an hour. The branch wasplaced inside a vial with water and fixed with a clamp toa vibration-isolated table (Newport LW3048B-OPT). Dif-ferent prey, predator and herbivore individuals wereused for each behavioural trial. Polistes fuscatus individualsflew from their nest to the experimental leaf, whereas P.maculiventris and the herbivores O. leucostigma, O. orbonaand A. conica were individually placed on or near the ex-perimental leaf. We recorded whether caterpillars (thatsurvived predation from wasps and stink bugs) hung in re-sponse to the presence of predators and measured thelength of the silk thread produced by each larva. Semio-thisa aemulataria larvae were recorded as survivors aftera predator (wasp or stink bug) left the experimental leaf.
To determine whether S. aemulataria larvae exposed toother herbivores responded by hanging, we introduceda larva of O. leucostigma, which have a similar weight toP. fuscatus (X� SE fresh weight for O. leucostigma andP. fuscatus, 0.128 � 0.0065 g and 0.125 � 0.0057 g, respec-tively; N ¼ 21). Similarly, the average weight of adultP. maculiventris was approximately the same as that ofO. orbona adults (0.055 � 0.0026 g and 0.052 � 0.0020 g,respectively; N ¼ 21). The third herbivore, A. conica,weighed less than the others (0.027 � 0.0016 g; N ¼ 21),and provided a contrast to herbivores weighing aboutthe same as the predators. The behavioural trials endedwhen the herbivores left the experimental leaf.
We also determined whether caterpillars responded byhanging when exposed to three simulated environmentalabiotic factors, a falling twig, rain drops and wind. Onesingle twig or water drop with weights of 0.505 � 0.0218 g(N ¼ 5) and 0.054 � 0.0016 g (N ¼ 15) respectively, was re-leased from a distance of 10 cm above the leaf. Wind wasgenerated at 0.2 m/s during a 30-s period (the averagewind velocity measured on three different days in Patux-ent Wildlife Refuge Research Center was 0.07 m/s (range0e0.2 m/s; N ¼ 10).
To quantify the hanging response of S. aemulataria tothe tufted titmouse, we exposed a fourth-instar larva toa single bird inside a cage. For each trial, the caterpillarwas placed on a 1e1.15-m high box elder tree insidea cage (1.25 � 1.25 � 0.65 m) framed with fish-netting.Before a bird was allowed to forage on the experimentaltree, the caterpillar was allowed to acclimate for half anhour; this was sufficient for a larva to assume a mimeticposture on a leaf. A single bird was placed inside thecage and allowed to acclimate for 1 h. After the acclima-tion period, the bird was allowed to forage for 1 h onthe experimental tree, because in preliminary observa-tions, 15 min was sufficient for P. bicolor to find and eatan Orgyia leucostigma caterpillar on a box elder tree inside
a cage. Birds were viewed through a cloth opening(15 � 2 cm) on one side of the cage to prevent the birdfrom seeing the observer. For surviving larvae we recordedwhether caterpillars hung in response to birds.
We used 30 replicates for each treatment. For the tuftedtitmouse, we conducted 36 replicates, where a replicateconsisted of a fourth-instar S. aemulataria. Three caterpil-lars were each exposed to 12 birds. All trials were con-ducted in the laboratory at a room temperature of about25 � 2 �C.
Characterization of vibrationsThe substrate-borne vibrations produced by inverte-
brate predators, herbivores and abiotic factors weredetected with a piezoelectric transducer (CB Sciences/iWorx PT-100) attached to an A. negundo leaf with an8.00 � 2.70 � 0.01-cm nylon flexible lever, glued toa 2.5 � 0.008-cm-diameter steel needle. The needle waswaxed to the underside of a leaf, 1e5 mm away froma S. aemulataria caterpillar. Because of the larger size ofP. bicolor, the lever was directly waxed to the undersideof a leaf, 1e5 mm away from a S. aemulataria caterpillar.All vibrations were recorded using a MacLab/400 unit(ADInstruments Colorado Springs, Colorado, U.S.A.)and analysed using Chart (ADInstruments). The vibra-tions produced by invertebrate predators, herbivoresand abiotic factors were recorded on an air-buffered, vi-bration-isolated table (Newport LW3048B-OPT), whereasthose produced by P. bicolor were recorded on a laboratorybench.
We analysed the spectral component of the vibrationsby calculating that fast Fourier transformation of theacceleration. Each frequency spectrum included a 1-speriod before S. aemulataria hung or before the point ofphysical contact between the biotic factors and the cater-pillar. We used 21 replicates to obtain an average spectrumfor each factor.
Experimental confirmation of the role of vibrationsTo determine whether S. aemulataria’s hanging behav-
iour occurred in response to the substrate-borne vibrationsproduced by the invertebrate predators, we exposed cater-pillars to band-limited noise vibration stimuli with a centralfrequency of 200 or 30 Hz. The 200-Hz stimulus repre-sented the frequency of maximum amplitude producedby the predators between 100 and 250 Hz, and the 30-Hzstimulus represented the frequency of maximum ampli-tude produced by the herbivores. White noise was pro-duced with a waveform generator (Agilent Technology/Hewlett Packard 33120A) and filtered with a parametricequalizer (Ashly Audio PQX-571 with a modified band toproduce a frequency range from 2 Hz to 2 kHz and 30 dBof boost). The filtered band-limited noise was amplified(Labworks PA-119) and the stimuli were transmitted tothe leaf using an electrodynamic shaker (Labworks ET-132-2). The 200-Hz stimulus was adjusted to be equal inpeak amplitude to wasp natural signals, and the 30-Hzstimulus was adjusted to be equal in amplitude to the200-Hz stimulus at a distance of approximately 0.5 cmfrom the larvae. The stop-band attenuation was set to

ANIMAL BEHAVIOUR, 72, 2464
30 dB, and the parametric bandwidth filter was set to ap-proximately 0.05 octave for the 200-Hz stimuli and to ap-proximately 0.2 octave for the 30-Hz stimulus. Weexposed 24 fourth-instar S. aemulataria to each of the twotypes of stimuli representing predators or herbivores.
Consequences of responses to predatorsOnce we determined differences in the silk lengths of
larvae responses to each predator we determined theconsequences of the differential response to caterpillarsurvival. Fourth-instar S. aemulataria were hung with0.1-mm-diameter transparent nylon thread at 30 cm and10 cm from an A. negundo leaf, or prevented from hanging(thus, 0 cm). Larvae of each type were then exposed toindividual foraging wasps or stink bugs. Larvae exposed toP. fuscatus were tied or hung from an A. negundo leaf oneither of two adjacent 1-m-long branches, whereas caterpil-lars exposed to P. maculiventris were tied or hung from a leafon one 1-m-long branch. A caterpillar was recorded asa survivor when the predator left the branch on which theexperimental caterpillar had been placed. All trials wereconducted in the laboratory at an ambient temperature ofabout 25 � 2 �C. Sample sizes for P. fuscatus were 9 (for30-cm threads), 8 (for 10-cm threads) and 15 (for 0 cm);sample sizes for P. maculiventris were 10 (for 30 cm), 10(for 10 cm) and 14 (for 0 cm).
Statistical Analyses
We analysed the behavioural responses of the caterpillars(hanging versus no hanging) to predators, herbivores,abiotic factors and simulated stimuli representing herbi-vores and predators using chi-square tests for independenceor Fisher’s exact tests when the assumptions of the chi-square test were not met (SAS Institute 1990). The lengths ofthe silk threads produced in response to foraging predatorswere compared with a paired Student’s t test. Comparisonsof survival of larvae hung at different lengths were analysedusing Fisher’s exact tests (SAS Institute 1990).
For spectral analyses, the frequency spectra were com-pared between 0 and 500 Hz in intervals of 50 Hz by ana-lysing the log-transformed areas under the curves withrepeated measures ANOVA (unstructured covariance struc-ture) and planned comparisons (SAS Institute 1990).Planned comparisons were adjusted with a Bonferroni cor-rection (Sokal & Rohlf 1995).
RESULTS
Quantification of Response to Predators,Herbivores and Abiotic Factors
The hanging behaviour of S. aemulataria larvae oc-curred primarily in response to invertebrate predators for-aging on a leaf rather than to herbivores, abiotic factorsor birds (Fisher’s exact tests: P < 0.0001; Fig. 1). No larvaehung in response to birds and only three of the 36 cater-pillars were taken and eaten by this predator (one birdate three caterpillars). Invertebrate predators ate 25 of60 caterpillars (17 caterpillars were eaten by wasps andeight were eaten by stink bugs).
The length of the silk thread produced by S. aemulatariacaterpillars in response to foraging wasps (X� SE ¼ 29.2 �3.62 cm, N ¼ 13) was significantly greater than that pro-duced in response to the stink bugs (9.9 � 1.17 cm,N ¼ 13; Student’s t test: t24 ¼ �5.04, P < 0.0001).
Characterization of Vibrations
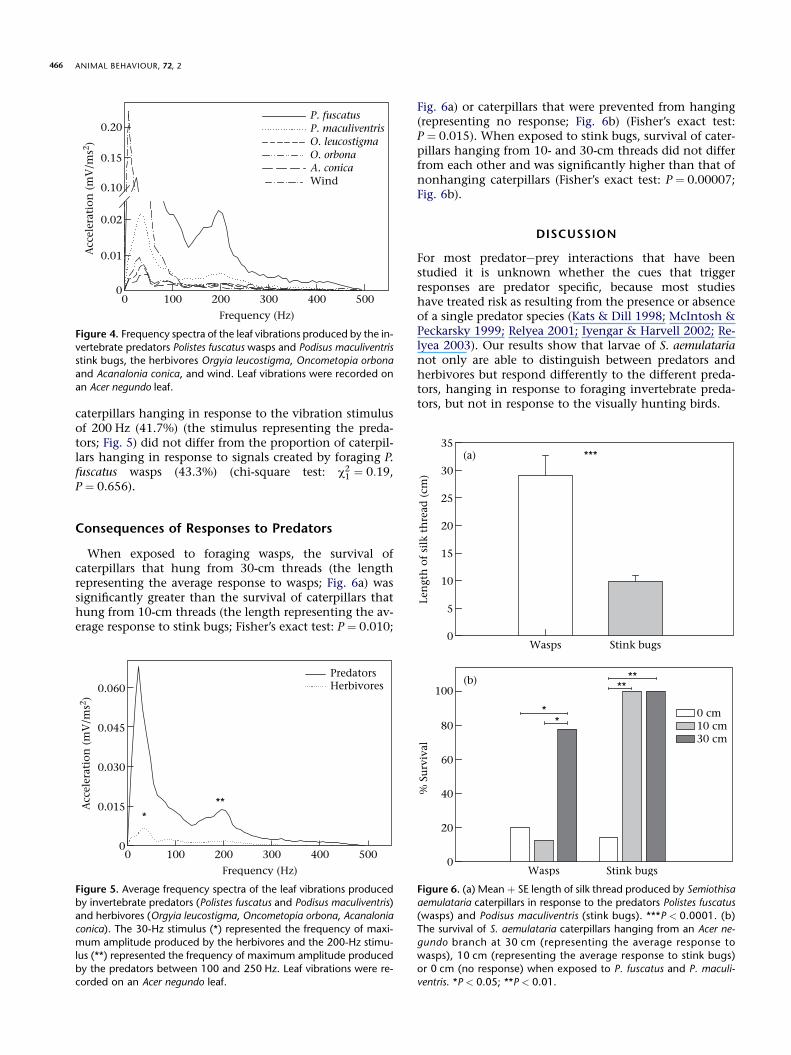
Foraging of invertebrate predators and movement ofherbivores on a leaf were measured as substrate-bornevibrations consisting of relatively continuous waveformsthat lasted a mean � SE of 6.677 � 0.4563 s (N ¼ 105)(Fig. 2a,b). Wind also generated continuous waveforms(Fig. 2c). In contrast, the substrate-borne vibrations pro-duced by P. bicolor consisted of momentary (and dissipat-ing) pulses that originated from random points (Fig. 3a).The pulses produced by P. bicolor lasted 0.9284 �0.1018 s (N ¼ 12) with intervals between them lasting1.8159 � 0.4665 s (N ¼ 12). The amplitude of the pulsesproduced by P. bicolor increased as the substrate distancebetween the point of origin of the pulse and the caterpillardecreased (Fig. 3a, b). Similar substrate-borne vibrationswere produced by falling twigs and drops of water andconsisted of momentary (and dissipating) pulses of1.1026 � 0.0999 s (N ¼ 42) that originated from randompoints (Fig. 3c, d).
The frequency spectra of waveforms produced by in-vertebrate predators, herbivores and wind differed signif-icantly (ANOVA: F45, 120 ¼ 20.18, P < 0.0001; Fig. 4). Theamplitudes of the substrate-borne vibration frequenciesproduced by invertebrate predators were significantlysmaller between 0 and 50 Hz and significantly largerbetween 100 and 450 Hz than the amplitudes of the fre-quencies produced by the wind (planned comparisons(Student’s t tests) with Bonferroni correction: t120 ¼ 3.33,P ¼ 0.0011), and did not differ significantly between 50and 100 Hz (Fig. 4). The amplitudes of the substrate-bornevibration frequencies produced by foraging P. maculiventris
% C
ater
pil
lars
han
gin
g
0
20
40
60
80 ***
Birds Herbivores Abioticfactors
Invertebratepredators
Figure 1. The responses of Semiothisa aemulataria caterpillars to in-vertebrate predators (Polistes fuscatus and Podisus maculiventris),
birds (Parus bicolor), herbivores (Orgyia leucostigma, Oncometopia or-
bona, Acanalonia conica), and abiotic factors (wind, a falling waterdrop and a falling twig). ***P < 0.0001.
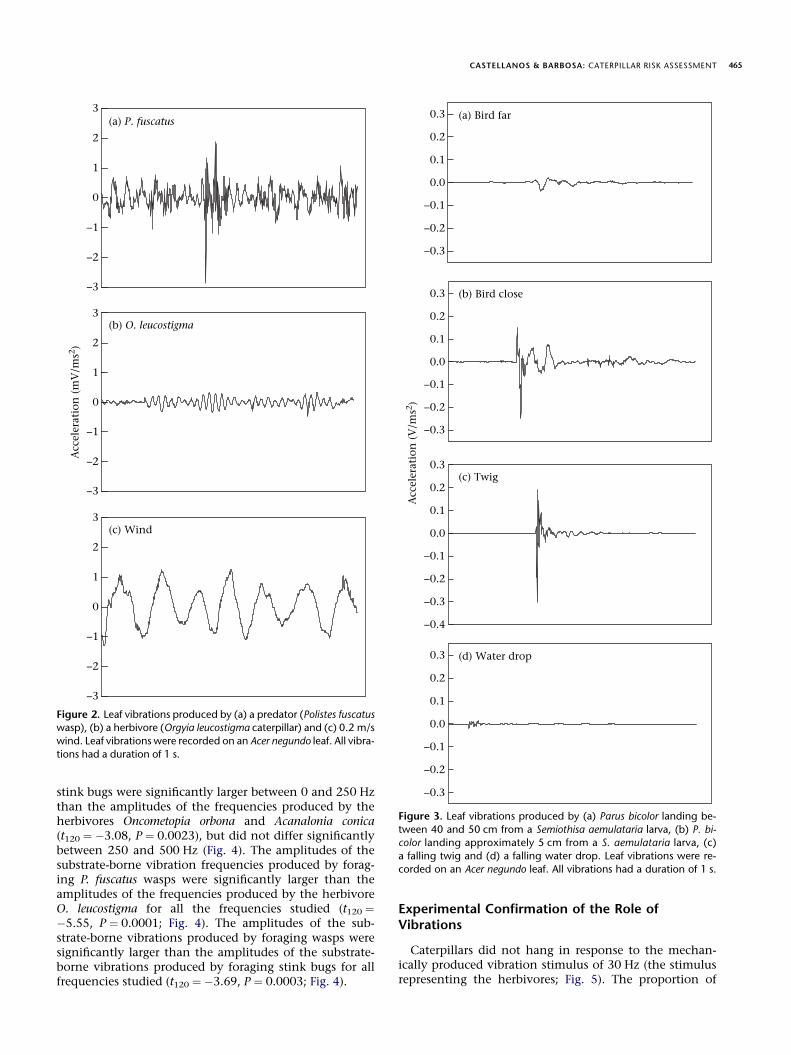
CASTELLANOS & BARBOSA: CATERPILLAR RISK ASSESSMENT 465
stink bugs were significantly larger between 0 and 250 Hzthan the amplitudes of the frequencies produced by theherbivores Oncometopia orbona and Acanalonia conica(t120 ¼ �3.08, P ¼ 0.0023), but did not differ significantlybetween 250 and 500 Hz (Fig. 4). The amplitudes of thesubstrate-borne vibration frequencies produced by forag-ing P. fuscatus wasps were significantly larger than theamplitudes of the frequencies produced by the herbivoreO. leucostigma for all the frequencies studied (t120 ¼�5.55, P ¼ 0.0001; Fig. 4). The amplitudes of the sub-strate-borne vibrations produced by foraging wasps weresignificantly larger than the amplitudes of the substrate-borne vibrations produced by foraging stink bugs for allfrequencies studied (t120 ¼ �3.69, P ¼ 0.0003; Fig. 4).
−3
−2
−1
0
1
2
3(a) P. fuscatus
Acc
eler
atio
n (
mV
/ms2 )
−3
−2
−1
0
1
2
3(b) O. leucostigma
−3
−2
−1
0
1
2
3(c) Wind
Figure 2. Leaf vibrations produced by (a) a predator (Polistes fuscatuswasp), (b) a herbivore (Orgyia leucostigma caterpillar) and (c) 0.2 m/s
wind. Leaf vibrations were recorded on an Acer negundo leaf. All vibra-
tions had a duration of 1 s.
Experimental Confirmation of the Role ofVibrations
Caterpillars did not hang in response to the mechan-ically produced vibration stimulus of 30 Hz (the stimulusrepresenting the herbivores; Fig. 5). The proportion of
−0.3
−0.2
−0.1
0.0
0.1
0.2
0.3 (a) Bird far
−0.3
−0.2
−0.1
0.0
0.1
0.2
0.3 (b) Bird close
Acc
eler
atio
n (
V/m
s2 )
−0.4
−0.3
−0.2
−0.1
0.0
0.1
0.2
0.3(c) Twig
−0.3
−0.2
−0.1
0.0
0.1
0.2
0.3 (d) Water drop
Figure 3. Leaf vibrations produced by (a) Parus bicolor landing be-
tween 40 and 50 cm from a Semiothisa aemulataria larva, (b) P. bi-color landing approximately 5 cm from a S. aemulataria larva, (c)
a falling twig and (d) a falling water drop. Leaf vibrations were re-
corded on an Acer negundo leaf. All vibrations had a duration of 1 s.
ANIMAL BEHAVIOUR, 72, 2466
caterpillars hanging in response to the vibration stimulusof 200 Hz (41.7%) (the stimulus representing the preda-tors; Fig. 5) did not differ from the proportion of caterpil-lars hanging in response to signals created by foraging P.fuscatus wasps (43.3%) (chi-square test: c2
1 ¼ 0:19,P ¼ 0.656).
Consequences of Responses to Predators
When exposed to foraging wasps, the survival ofcaterpillars that hung from 30-cm threads (the lengthrepresenting the average response to wasps; Fig. 6a) wassignificantly greater than the survival of caterpillars thathung from 10-cm threads (the length representing the av-erage response to stink bugs; Fisher’s exact test: P ¼ 0.010;
Frequency (Hz)
0 100 200 300 400 500
Acc
eler
atio
n (
mV
/ms2 )
0
0.01
0.02
0.10
0.15
0.20P. fuscatusP. maculiventrisO. leucostigmaO. orbonaA. conicaWind
Figure 4. Frequency spectra of the leaf vibrations produced by the in-
vertebrate predators Polistes fuscatus wasps and Podisus maculiventrisstink bugs, the herbivores Orgyia leucostigma, Oncometopia orbona
and Acanalonia conica, and wind. Leaf vibrations were recorded on
an Acer negundo leaf.
Frequency (Hz)
0 100 200 300 400 500
Acc
eler
atio
n (
mV
/ms2 )
0
0.015
0.030
0.045
0.060PredatorsHerbivores
***
Figure 5. Average frequency spectra of the leaf vibrations produced
by invertebrate predators (Polistes fuscatus and Podisus maculiventris)and herbivores (Orgyia leucostigma, Oncometopia orbona, Acanalonia
conica). The 30-Hz stimulus (*) represented the frequency of maxi-
mum amplitude produced by the herbivores and the 200-Hz stimu-
lus (**) represented the frequency of maximum amplitude producedby the predators between 100 and 250 Hz. Leaf vibrations were re-
corded on an Acer negundo leaf.
Fig. 6a) or caterpillars that were prevented from hanging(representing no response; Fig. 6b) (Fisher’s exact test:P ¼ 0.015). When exposed to stink bugs, survival of cater-pillars hanging from 10- and 30-cm threads did not differfrom each other and was significantly higher than that ofnonhanging caterpillars (Fisher’s exact test: P ¼ 0.00007;Fig. 6b).
DISCUSSION
For most predatoreprey interactions that have beenstudied it is unknown whether the cues that triggerresponses are predator specific, because most studieshave treated risk as resulting from the presence or absenceof a single predator species (Kats & Dill 1998; McIntosh &Peckarsky 1999; Relyea 2001; Iyengar & Harvell 2002; Re-lyea 2003). Our results show that larvae of S. aemulatarianot only are able to distinguish between predators andherbivores but respond differently to the different preda-tors, hanging in response to foraging invertebrate preda-tors, but not in response to the visually hunting birds.
(a)
Wasps Stink bugs
Len
gth
of
silk
th
read
(cm
)
0
5
10
15
20
25
30
35
(b)
Wasps Stink bugs
% S
urv
ival
0
20
40
60
80
100
0 cm10 cm30 cm
***
****
**
Figure 6. (a) Mean þ SE length of silk thread produced by Semiothisa
aemulataria caterpillars in response to the predators Polistes fuscatus(wasps) and Podisus maculiventris (stink bugs). ***P < 0.0001. (b)
The survival of S. aemulataria caterpillars hanging from an Acer ne-
gundo branch at 30 cm (representing the average response to
wasps), 10 cm (representing the average response to stink bugs)or 0 cm (no response) when exposed to P. fuscatus and P. maculi-
ventris. *P < 0.05; **P < 0.01.

CASTELLANOS & BARBOSA: CATERPILLAR RISK ASSESSMENT 467
The higher incidence of hanging by larvae of S. aemula-taria in response to the movement of invertebrate pre-dators compared to that of herbivores was based ondifferences in the amplitude of the high-frequency com-ponents of substrate-borne vibrations. However, althoughthe weight of the invertebrate predators and herbivoreswas controlled, we cannot rule out the importance of phy-logeny. Current research is being conducted to control forthis variable. However, the different herbivores producedessentially the same type of signal, although their weights,taxa and life stage were different. Furthermore, predatorsignals were different and induced a different response.
Since the two invertebrate predators evaluated imposedifferent levels of mortality on S. aemulataria (I. Castella-nos & P. Barbosa, unpublished data), and the differentialhanging response of larvae results in increased survival,one can infer that this species uses substrate-borne vibra-tions to assess levels of risk and respond accordingly.That is, S. aemulataria larvae appear to respond to differ-ences in the amplitude of the high-frequency componentsof the substrate-borne vibrations of predatory stink bugsand wasps by producing silk threads of different lengths(which may be determined in part by the cost of produc-ing silk; Berenbaum et al. 1993; Craig et al. 1999; Stevenset al. 1999), and thus provide differential larval survivalfrom the two invertebrate predators. Podisus maculiventrisforage for prey by walking and cannot attack an out ofreach larva suspended on a thread (Yeargan & Braman1986). Polistes fuscatus on the other hand, can fly, andtherefore can still attack suspended larvae. Flying P. fusca-tus individuals usually cover a 10e15-cm radius aroundtarget leaves, so caterpillars may hang lower on their silkthreads to separate themselves from the searching wasp(I. Castellanos, personal observation).
Although air currents generated by the wingbeats ofa wasp as it approaches a leaf can cause a caterpillar toreact defensively (Markl & Tautz 1975; Tautz & Markl1978), this explanation is unlikely to account for thehanging behaviour of S. aemulataria in the present study,because larvae showed hanging behaviour only aftera wasp had landed and began walking on a leaf.
Three possible explanations may account for the lack ofresponse of S. aemulataria to the presence of P. bicolor.Semiothisa aemulataria caterpillars may not be able to de-tect foraging P. bicolor. It is probable that this species ofcaterpillar has evolved a sensory system matched to theircommon predators, which could be the invertebrate pred-ators (see Jablonski 1998; Djemai et al. 2001). Alterna-tively, larvae may be able to detect avian signals but thevibrations produced by foraging birds may make birdsvery difficult to differentiate from abiotic factors. Theshort substrate-borne pulses produced by the landing ofthe birds are similar in duration and amplitude to thepulses generated by falling twigs and water drops. Further-more, the amplitude of the pulses produced by the birds isdetermined by the distance between the point of origin ofthe pulse and the caterpillar, and pulses originate fromrandom points. Thus, the pulses may not provide reliableor predictable spatial and temporal information.
A third possibility is that larvae may detect avianpredators but do not respond by hanging, because they
rely instead on their mimetic posture, which requiresimmobility to be effective (Stamp & Wilkens 1993; I. Cas-tellanos & P. Barbosa, unpublished data).
Previous studies have suggested that differential re-sponses of insect prey to predators and competitors mightbe based on differences in the amplitude of the frequencycomponents of the signals (Fullard & Belwood 1988;Gnatzy & Kamper 1990; Fullard 1998). Other studieshave characterized the cues used by invertebrate prey todistinguish between predators and nonpredators (Plum-mer & Camhi 1981). However, the results presented hereshow that caterpillars, which typically have limited visualand chemical perception capabilities (Scoble 1992), cannevertheless determine what is and is not a potentialthreat using substrate-borne signals, and respond accord-ingly, and that the specificity of their response has fitnessconsequences because it enhances survival.
Acknowledgments
We acknowledge the able assistance of Holliday Obrechtof the Patuxent Wildlife Research Center for his invalu-able support of this research. This project was funded, inpart, by the University of Maryland, Agricultural Experi-ment Station Project MD-H-201. We acknowledge theimmensely helpful suggestions provided by A. A. Agrawal,D. G. Bottrel, M. C. Christman, R. F. Denno, D. K.Letourneau, D. R. Papaj, J. C. Schultz, D. S. Thaler, G. S.Wilkinson, D. D. Yager, I. Zuria and three anonymousreferees. Finally, we acknowledge N. Breisch, University ofMaryland, and S. H. McKamey, Systematic EntomologyLaboratory, U.S.D.A. for species determinations and J. R.Aldrich for providing Podisus maculiventris pheromonetraps. I. Castellanos thanks I. Zuria for help in rearingthe insect colonies and the Fulbright program, the Depart-ment of Entomology and the Program in Behavior, Ecol-ogy, Evolution and Systematics, University of Maryland,the Universidad Autonoma del Estado de Hidalgo, Mexico,and the Programa de Mejoramiento del Profesorado(PROMEP), Secretarıa de Educacion Publica, Mexico, fortheir generous support.
References
Barbosa, P., Caldas, A. & Robinson, G. 2003. Host plant associa-tions among species in two macrolepidopteran assemblages. Jour-
nal of Entomological Sciences, 38, 41e47.
Bauer, T., Brauner, U. & Fischerleitner, E. 1977. The relevance of
brightness to visual acuity predation, and activity of visually hunt-
ing ground-beetles (Coleoptera, Carabidae). Oecologia, 30,63e77.
Beal, F. E. L., McAtee, W. L. & Kalmbach, E. R. 1941. Common
birds of southeastern United States in relation to agriculture. U.S.Fish and Wildlife Service Conservation Bulletin, 15.
Bent, A. C. 1946. Life histories of North American jays, crows andtitmice. U.S. National Museum Bulletin, 191.
Berenbaum, M. R., Moreno, B. & Green, E. 1992. Soldier bug pre-dation on swallowtail caterpillars (Lepidoptera: Papilionidae): cir-
cumvention of defensive chemistry. Journal of Insect Behavior, 5,
547e553.

ANIMAL BEHAVIOUR, 72, 2468
Berenbaum, M. R., Green, E. S. & Zangerl, A. R. 1993. Web costs
and web defense in the parsnip webworm (Lepidoptera: Oeco-
phoridae). Environmental Entomology, 22, 791e795.
Cokl, A., Virant-Doberlet, M. & McDowell, A. 1999. Vibrational di-
rectionality in the southern green stink bug, Nezara Viridula (L.), ismediated by female song. Animal Behaviour, 58, 1277e1283.
Cocroft, R. B. 1996. Insect vibrational defence signals. Nature, 382,679e680.
Craig, C. L., Hsu, M., Kaplan, D. & Pierce, N. E. 1999. A compar-ison of the composition of silk proteins produced by spiders and
insects. International Journal of Biological Macromolecules, 24,
109e118.
DeVries, P. J. 1990. Enhancement of symbiosis between butterfly
caterpillars and ants by vibrational communication. Science, 248,
1104e1106.
Dicke, M. & Grostal, P. 2001. Chemical detection of natural ene-
mies by arthropods: an ecological perspective. Annual Review ofEcology and Systematics, 32, 1e23.
Djemai, I., Casas, J. & Magal, C. 2001. Matching host reactions toparasitoid wasp vibrations. Proceedings of the Royal Society of
London, Series B, 268, 2403e2408.
Eisner, T. & Dean, J. 1976. Ploy and counterploy in predatoreprey in-
teractions: orb-weaving spiders versus bombardier beetles. Proceed-
ings of the National Academy of Sciences, U.S.A., 73, 1365e1367.
Evans, E. W. 1982. Feeding specialization in predatory insects: hunt-
ing and attack behavior of two stinkbug species (Hemiptera: Pen-
tatomidae). American Midland Naturalist, 108, 96e104.
Fullard, J. H.1998. The sensory coevolution of moths and bats. In: Com-
parative Hearing: Insects (Ed. by R. R. Hoy, A. N. Popper & R. R. Fay),pp. 279e326. New York: Springer-Verlag.
Fullard, J. H. & Belwood, J. J. 1988. The ecological assemblage:acoustic ensembles in a Neotropical habitat. In: NATO ASI Series,
Series A: Life Sciences. Vol. 156, Animal Sonar (Ed. by P. E.
Natchigall & P. W. B. Moore), pp. 639e643. New York: Plenum.
Gnatzy, W. & Kamper, G. 1990. Digger wasp against crickets. II. An
airborne signal produced by a running predator. Journal of Com-
parative Physiology, A, 167, 551e556.
Gould, W. P. & Jeanne, R. L. 1984. Polistes wasps (Hymenoptera:
Vespidae) as control agents for lepidopterous cabbage pests. Envi-ronmental Entomology, 13, 150e156.
Grubb, T. C. & Pravosudov, V. V. 1994. Tufted titmouse (Parus
bicolor). In: The Birds of North America. No. 86 (Ed. byA. Poole & A. Gill). Philadelphia: The Birds of North America.
Hergenroder, R. & Barth, F. 1983. The release of attack and escapebehavior by vibratory stimuli in a wandering spider (Cupiennieus
salei Keys.). Journal of Comparative Physiology A, 152, 347e358.
Iyengar, E. V. & Harvell, C. D. 2002. Specificity of cues inducing
defensive spines in the bryozoan Membranipora membranacea.
Marine Ecology Progress Series, 225, 205e218.
Jablonski, P. G. 1998. A rare predator exploits prey escape behavior:
the role of tail-fanning and plumage contrast in foraging of
the painted redstart (Myioborus pictus). Behavioral Ecology, 10,7e14.
Kats, L. B. & Dill, L. M. 1998. The scent of death: chemosensory as-sessment of predation risk by prey animals. Ecoscience, 5, 361e394.
Kirchner, W. H., Broecker, I. & Tautz, J. 1994. Vibrational alarmcommunication in the damp-wood termite Zootermopsis nevaden-
sis. Physiological Entomology, 19, 187e190.
Land, M. & Layne, J. 1995. The visual control of behaviour in fiddler
crabs I. Resolution, thresholds and the role of the horizon. Journal
of Comparative Physiology, A, 177, 81e90.
McIntosh, A. R. & Peckarsky, B. L. 1999. Criteria for determining
behavioural responses to multiple predators by a stream may fly.
Oikos, 85, 554e564.
McPherson, J. E. 1982. The Pentatomoidea (Hemiptera) of Northeast-
ern North America. Carbondale: Southern Illinois University Press.
Mallampalli, N., Castellanos, I. & Barbosa, P. 2002. Evidence for
intraguild predation by Podisus maculiventris on a ladybeetle, Co-
leomegilla maculata: implications for biological control of Colo-rado potato beetle, Leptinotarsa decemlineata. Biocontrol, 47,
387e398.
Markl, H. 1983. Vibrational communication. In: Neuroethology and
Behavioral Physiology (Ed. by F. Huber & H. Markl), pp.
332e353. New York: Springer-Verlag.
Markl, H. & Tautz, J. 1975. The sensitivity of hair receptors in cater-
pillars of Barathra brassicae L. (Lepidoptera, Noctuidae) to particle
movement of sound field. Journal of Comparative Physiology, 99,79e87.
Martinez-Wells, M. & Henry, C. S. 1992. Behavioural responses ofgreen lacewings (Neuroptera: Chrysopidae: Chrysoperla) to syn-
thetic mating songs. Animal Behaviour, 44, 641e652.
Medina, R. F. 1999. Predation of Orgyia leucostigma (J. E. Smith)
(Lepidoptera: Lymantriidae) larvae on Acer negundo (L.) (Sapin-
dales: Aceraceae) by invertebrate and vertebrate predators. M.S.
thesis, University of Maryland, College Park.
Michener, C. D. & Michener, M. H. 1951. American Social Insects.
New York: D. Van Nostrand Rheinhold.
Nalbach, H. O. 1990. Visually elicited escape in crabs. In: Frontiers in
Crustacean Neurobiology (Ed. by K. Wiese, W. D. Krent, J. Tautz, H.Reichert & B. Mulloney), pp. 165e172. Basel: Birkauser Verlag.
Payne, H. G. 1917. The white marked tussock moth. Proceedings ofthe Entomological Society of Nova Scotia, 3, 62e69.
Pfannenstiel, R. S., Hunt, R. E. & Yeargan, K. V. 1995. Orientationof a hemipteran predator to vibrations produced by feeding cater-
pillars. Journal of Insect Behavior, 8, 1e9.
Plummer,M.R.& Camhi, J. M. 1981. Discriminationof sensory signals
from noise in the escape system of the cockroach: the role of wind
acceleration. Journal of Comparative Physiology A, 142, 347e357.
Rabb, R. L. 1960. Biological studies of Polistes in North Carolina
(Hymenoptera: Vespidae). Annals of the Entomological Society of
America, 53, 111e121.
Relyea, R. A. 2001. Morphological and behavioral plasticity of larval
anurans in response to different predators. Ecology, 82, 523e540.
Relyea, R. A. 2003. How prey respond to combined predators: a re-
view and an empirical test. Ecology, 84, 1827e1839.
Raveret Richter, M. 2000. Social wasp (Hymenoptera: Vespidae)
foraging behavior. Annual Review of Entomology, 45, 121e150.
Robinson, S. K. & Holmes, R. T. 1982. Foraging behavior of forest
birds: the relationships among search tactics, diet, and habitat
structure. Ecology, 63, 1918e1931.
Roces, F., Tautz, J. & Holldobler, B. 1993. Stridulation in leaf-cut-
ting ants. Naturwissenschaften, 80, 521e524.
SAS Institute 1990. SAS Procedures Guide, Version 6. 3rd edn. Cary,
North Carolina: SAS Institute.
Scoble, M. J. 1992. The Lepidoptera: Form, Function and Diversity.
Oxford: Oxford University Press.
Sokal, R. R. & Rohlf, F. J. 1995. Biometry. New York: W. H. Freeman.
Stamp, N. E. 1992. Relative susceptibility to predation of two species
of caterpillar on plantain. Oecologia, 92, 124e129.
Stamp, N. E. & Wilkens, R. T. 1993. On the cryptic side of life: being
unapparent to enemies and the consequences for foraging and
growth of caterpillars. In: Caterpillars: Ecological and Evolutionary
Constraints on Foraging (Ed. by N. E. Stamp & T. M. Casey), pp.283e330. New York: Chapman & Hall.
Stephens, D. W. & Krebs, J. R. 1986. Foraging Theory. Princeton,New Jersey: Princeton University Press.
Stevens, D. J., Hansell, M. H., Freel, J. A. & Monaghan, P. 1999.Developmental trade-offs in caddis flies: increased investment in

CASTELLANOS & BARBOSA: CATERPILLAR RISK ASSESSMENT 469
larval defence alters adult resource allocation. Proceedings of the
Royal Society of London, Series B, 266, 1049e1054.
Tautz, J. & Markl, H. 1978. Caterpillars detect flying wasps by hairs
sensitive to airborne vibration. Behavioral Ecology and Sociobiology,
4, 101e110.
Tollrian, R. & Von Elert, E. 1994. Enrichment and purification of
Chaoborus kairomone from water: further steps toward its chem-ical characterization. Limnology and Oceanography, 39,
788e796.
Viitasalo, M., Kiorboe, T., Flinkman, J., Pedersen, L. W. & Visser,A. 1998. Predatory vulnerability of planktonic copepods: conse-
quences of predator foraging strategies and prey sensor abilities.
Marine Ecology Progress Series, 175, 129e142.
Von Elert, E. & Pohnert, G. 2000. Predator specificity of kairomones
in diel vertical migration of Daphnia: a chemical approach. Oikos,
88, 119e128.
Wilson, S. W. & McPherson, J. E. 1981. Life histories of Acanalonia
bivittata and A. conica with descriptions of immature stages. An-nals of the Entomological Society of America, 74, 289e298.
Yack, J. E., Smith, M. L. & Weatherhead, P. J. 2001. Caterpillar talk:acoustically mediated territoriality in larval lepidoptera. Proceed-
ings of the National Academy of Sciences, U.S.A., 98, 11371e11375.
Yeargan, K. V. & Braman, K. 1986. Life history of the parasite Dio-
lcogaster facetosa (Weed) (Hymenoptera: Braconidae) and its be-
havioral adaptation to the defensive response of a lepidopteran
host. Annals of the Entomological Society of America, 79, 1029e1033.
Related Documents











