Evaluation of d-Amphetamine Effects on the Binding of Dopamine D-2 Receptor Radioligand, 18 F-Fallypride in Nonhuman Primates Using Positron Emission Tomography JOGESHWAR MUKHERJEE, 1 * ZHI-YING YANG, 1 ROBERT LEW, 2 TERRY BROWN, 1 SHARA KRONMAL, 1 MALCOLM D. COOPER, 1 AND LEWIS S. SEIDEN 2 1 Franklin McLean Institute, Department of Radiology, University of Chicago, Chicago, Illinois 60637 2 Department of Pharmacological and Physiological Sciences, University of Chicago, Chicago, Illinois 60637 KEY WORDS 18 F-fallypride; dopamine D 2 receptors; amphetamine; monkey PET ABSTRACT We have investigated the ability of dopamine to compete with the binding of the high affinity dopamine D 2 receptor positron emission tomography (PET) radioligand, 18 F-fallypride. In vitro dissociation of 18 F-fallypride with dopamine in rat striatal homogenates exhibited a dissociation rate, k off , of 1.76 3 10 22 min 21 while the association rate constant, k on , was found to be 5.30 3 10 8 M 21 min 21 . This resulted in a dissociation constant, K D of 33 pM for 18 F-fallypride. For in vivo studies, we investigated the effects of reserpine and d-amphetamine treatment on 18 F-fallypride in an attempt to study competition of endogenous dopamine with the radioligand at the receptor sites in rats and monkeys. PET experiments with 18 F-fallypride in two male rhesus monkeys were carried out in a PETT VI scanner. In control experiments, rapid specific uptake of 18 F-fallypride in the striata was observed (0.05–0.06% injected dose (ID)/g) while nonspecifically bound tracer cleared from other parts of the brain. Striata/cerebellum ratios for 18 F-fallypride were approximately 8 at 80 min postinjection, respectively. The monkeys received various doses (0.25 to 1.50 mg/kg) of d-amphetamine (AMPH) pre- and postinjection of the radioligand. There was a decrease of specifically bound 18 F-fallypride as well as evidence of an enhanced clearance of specifically bound 18 F-fallypride after administering AMPH in the two monkeys. The dissociation rates, k off , of 18 F-fallypride without AMPH was ,10 24 min 21 but after 25 min preadministration of AMPH (1 mg/kg), it was 4.1 3 10 23 min 21 and after 17, 45 and 90 min postadministration of AMPH (1 mg/kg) it was 3.6 3 10 23 to 4.0 3 10 23 min 21 . Lower doses of AMPH (0.25 mg/kg) had a reduced effect on the binding of 18 F-fallypride. No effect was seen until about 30 minutes after the injection of AMPH. Studies with various doses indicated that 18 F-fallypride has a maximum response at doses of 0.75–1.50 mg/kg, with an approximately 16%/hour reduction in binding. These results indicate that AMPH stimulated release of endogenous dopamine reduces the specific binding of 18 F-fallypride. Synapse 27:1–13, 1997. r 1997 Wiley-Liss, Inc. INTRODUCTION Endogenous dopamine present at the synapse has been shown to compete with the binding of several postsynaptic dopamine D 2 receptor radioligands (DeJe- sus et al., 1986; Dewey et al., 1991; Friedman et al., 1984; Innis et al., 1992; Seeman et al., 1989). This ability of dopamine to compete with the radioligands for the receptors has been used to study alterations in the synaptic levels of dopamine by investigating the changes in the binding of the radioligand both in vitro and in vivo (for a recent review see Volkow et al., 1996). At the same time, in studies that involve evaluation of the distribution of postsynaptic D 2 receptors, there is a concern that excessive competition by endogenous dopa- mine with the radioligands could lead to an underesti- Presented in part at the 41st Annual Meeting Society of Nuclear Medicine, Orlando, Florida June 5–8, 1994, and the 42nd Annual Meeting Society of Nuclear Medicine, Minneapolis, Minnesota June 11–16, 1995. Contract grant sponsor: U.S. Department of Energy, Office of Health and Environmental Research; Contract grant number: FG02-94ER61840; Contract grant sponsor: Scottish Rite Schizophrenia Foundation. *Correspondence to: Jogeshwar Mukherjee, Ph.D., Franklin McLean Institute, Department of Radiology, MC 2026, 5841 S. Maryland Avenue, University of Chicago, Chicago, IL 60637. E-mail: [email protected] Received 2 September 1996; Accepted 10 November 1996 SYNAPSE 27:1–13 (1997) r 1997 WILEY-LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Evaluation of d-Amphetamine Effectson the Binding of Dopamine D-2 ReceptorRadioligand, 18F-Fallypride in Nonhuman
Primates Using Positron EmissionTomography
JOGESHWAR MUKHERJEE,1* ZHI-YING YANG,1 ROBERT LEW,2 TERRY BROWN,1SHARA KRONMAL,1 MALCOLM D. COOPER,1 AND LEWIS S. SEIDEN 2
1Franklin McLean Institute, Department of Radiology, University of Chicago, Chicago, Illinois 606372Department of Pharmacological and Physiological Sciences, University of Chicago, Chicago, Illinois 60637
KEY WORDS 18F-fallypride; dopamine D2 receptors; amphetamine; monkey PET
ABSTRACT We have investigated the ability of dopamine to compete with thebinding of the high affinity dopamine D2 receptor positron emission tomography (PET)radioligand, 18F-fallypride. In vitro dissociation of 18F-fallypride with dopamine in ratstriatal homogenates exhibited a dissociation rate, koff, of 1.76 3 1022 min21 while theassociation rate constant, kon, was found to be 5.30 3 108 M21 min21. This resulted in adissociation constant, KD of 33 pM for 18F-fallypride. For in vivo studies, we investigated theeffects of reserpine and d-amphetamine treatment on 18F-fallypride in an attempt to studycompetition of endogenous dopamine with the radioligand at the receptor sites in rats andmonkeys. PET experiments with 18F-fallypride in two male rhesus monkeys were carried outin a PETT VI scanner. In control experiments, rapid specific uptake of 18F-fallypride in thestriata was observed (0.05–0.06% injected dose (ID)/g) while nonspecifically bound tracercleared from other parts of the brain. Striata/cerebellum ratios for 18F-fallypride wereapproximately 8 at 80 min postinjection, respectively. The monkeys received various doses(0.25 to1.50mg/kg) ofd-amphetamine (AMPH)pre- andpostinjectionof the radioligand.Therewas adecrease of specifically bound 18F-fallypride aswell as evidence of an enhanced clearanceof specifically bound 18F-fallypride after administering AMPH in the two monkeys. Thedissociation rates, koff, of 18F-fallypride without AMPH was ,1024 min21 but after 25 minpreadministration of AMPH (1 mg/kg), it was 4.1 3 1023 min21 and after 17, 45 and 90 minpostadministration of AMPH (1 mg/kg) it was 3.6 3 1023 to 4.0 3 1023 min21. Lowerdoses of AMPH (0.25 mg/kg) had a reduced effect on the binding of 18F-fallypride. Noeffect was seen until about 30 minutes after the injection of AMPH. Studies with variousdoses indicated that 18F-fallypride has a maximum response at doses of 0.75–1.50 mg/kg,with an approximately 16%/hour reduction in binding. These results indicate thatAMPHstimulated release of endogenous dopamine reduces the specific binding of 18F-fallypride.Synapse 27:1–13, 1997. r 1997 Wiley-Liss, Inc.
INTRODUCTION
Endogenous dopamine present at the synapse hasbeen shown to compete with the binding of severalpostsynaptic dopamine D2 receptor radioligands (DeJe-sus et al., 1986; Dewey et al., 1991; Friedman et al.,1984; Innis et al., 1992; Seeman et al., 1989). Thisability of dopamine to compete with the radioligands forthe receptors has been used to study alterations in thesynaptic levels of dopamine by investigating the changesin the binding of the radioligand both in vitro and invivo (for a recent review see Volkow et al., 1996). At thesame time, in studies that involve evaluation of the
distribution of postsynaptic D2 receptors, there is aconcern that excessive competition by endogenous dopa-mine with the radioligands could lead to an underesti-
Presented in part at the 41st Annual Meeting Society of Nuclear Medicine,Orlando, Florida June 5–8, 1994, and the 42nd Annual Meeting Society ofNuclear Medicine, Minneapolis, Minnesota June 11–16, 1995.
Contract grant sponsor: U.S. Department of Energy, Office of Health andEnvironmental Research; Contract grant number: FG02-94ER61840; Contractgrant sponsor: Scottish Rite Schizophrenia Foundation.
*Correspondence to: Jogeshwar Mukherjee, Ph.D., Franklin McLean Institute,Department of Radiology, MC 2026, 5841 S. Maryland Avenue, University ofChicago, Chicago, IL 60637. E-mail: [email protected]
Received 2 September 1996; Accepted 10 November 1996
SYNAPSE 27:1–13 (1997)
r 1997 WILEY-LISS, INC.

mation of the distribution of receptor concentrations(Seeman et al., 1990).Two different classes of radioligands which bind
primarily to the D2 receptor have been investigated formeasuring in vivo alterations of endogenous dopamine.The substituted benzamides, such as 11C-racloprideand 123I-IBZM, are selective, moderate affinity, revers-ible radioligands and the butyrophenones, such as,11C-N-methylspiperone (11C-NMSP) and 18F-N-methyl-spiperone (18F-NMSP), have high affinity but are lessselective for the D2 receptors. It has been shown thatdensity of 3H-raclopride sites was 20% to 45% lowerthan the density of 3H-methylspiperone (3H-NMSP)sites in the presence of 100 nM dopamine added tohuman striata in vitro (Seeman et al., 1989). Similarly,in vivo experiments with and without reserpine pre-treatment of rats and mice indicated a more than 50%increase in in vivo striatal binding of 3H-raclopride inboth rats and mice (Ross and Jackson, 1989; Seeman etal., 1989; Young et al., 1991). No effect of reserpinepretreatment was observed on the binding of 3H-NMSP(Young et al., 1991). This difference between 3H-raclopride and 3H-NMSP has been attributed to thehigher affinity of 3H-NMSP (Seeman et al., 1989).Dopamine release at the synapse in vivo is a known
mechanism of action of d-amphetamine (AMPH, Seidenet al., 1993). Therefore, AMPH is frequently used as ameans of inducing the release of large amounts ofendogenous dopamine at the synapse in order to evalu-ate the effect on receptor-bound radioligands in vivo.Administration ofAMPH (10mg/kg) prior to the radioli-gand injection showed a 41.5% reduction in 3H-raclopride binding (Young et al., 1991). In kinetic experi-ments carried out in rats with 3H-NMSP, there was nosignificant difference in the volume of distribution of 3H-NMSP with and without AMPH (5 mg/kg) pretreatment(Young et al., 1991). This has been attributed to the higheraffinity of 3H-NMSP for the D2 receptor and its slowerkinetics in vivo compared to that of 3H-raclopride(Seeman et al., 1989; Young et al., 1991).A number of imaging studies, positron emission
tomography (PET) and single photon emission com-puted tomography (SPECT), have been carried outusing AMPH challenges in order to evaluate effects onthe binding of the radioligands at the D2 receptor sites.Using the SPECT radioligand, 123I-IBZM, AMPH chal-lenge studies in nonhuman primates show an enhancedwashout rate of 34%/h (Innis et al., 1992). In humans,using 123I-IBZM and a smaller dose of AMPH (0.3mg/kg), a decrease of 15% in the binding of the radioli-gand at the D2 receptor was observed (Laruelle et al.,1995). In PET experiments using 11C-raclopride, an oraldose of 30 mg ofAMPH induced a decrease in binding ofapproximately 10% in humans (Farde et al., 1992). Asimilar AMPH-induced reduction in binding of 11C-raclopride in baboons has also been reported (Dewey etal., 1993). Pretreatment of baboons with AMPH caused
a reduction in the binding of N-methyl-18F-spiperone aswell (Logan et al., 1991).Measurement of alterations in endogenous dopamine
release by methods other than AMPH-induced releasehave also been reported. Dewey et al. (1992) haveshown an increase of 11C-raclopride binding subsequentto gamma aminobutyric acid (GABA)ergic inhibition ofsynaptic dopamine release in baboons. Treatment withvinyl-GABA, a suicide inhibitor of GABAtransaminase,increased the distribution volume of 11C-raclopride inthe striata by approximately 25%. Treatment withlorazepam, a benzodiazepine agonist, also increasedthe binding of 11C-raclopride significantly (rangingbetween 8% and 30%).Attempts have also beenmade tostudy the competition of endogenous dopamine with thebinding of 11C-raclopride in subjects treated with meth-ylphenidate (Volkow et al., 1994). More recently, in vitrostudies using electrical stimulation of rat striatal slicesdemonstrated competition of released dopamine with 3H-raclopride and 123I-epidepride (Gifford et al., 1996).We have developed the substituted benzamide, 18F-
fallypride ((S)-N-[(1-allyl-2-pyrrolidinyl)methyl]-5-(3-18F -fluoropropyl)-2,3-dimethoxybenzamide, Fig. 1) as arelatively selective, high affinity, fluorinated dopamineD2 receptor radioligand (Mukherjee et al., 1995a). Innonhuman primate PET studies, 18F-fallypride bindingis very selective to dopaminergic sites providing highspecific to nonspecific binding ratios. Binding of 18F-fallypride in vivo was found to be rapidly reversible inthe presence of dopamine D2 receptor antagonists, suchas haloperidol and raclopride (Mukherjee et al.,1995a,b). In order to study the ability of endogenousdopamine to compete with specifically bound 18F-fallypride in the striatum, we have carried out a seriesof experiments in rats and PET experiments in rhesusmonkeys which involved competition studies of 18F-fallypridewithAMPH-induced release of endogenousdopa-mine. This paper describes in vivo studies in rats andrhesusmonkeys in order to study the effect ofAMPHon thebinding of 18F-fallypride at the dopamine D2 receptor sites.
MATERIALS AND METHODSRadiopharmaceutical
The radioligand used in the study was the substi-tuted benzamide, 18F-fallypride (Mukherjee et al.,1995a). 18F-Fluoride was prepared by 18O(p,n)18F reac-
Fig. 1. Chemical structure of 18F-fallypride.
2 J. MUKHERJEE ET AL.

tion in a CS-15 cyclotron and the solubilized 18F-fluoride was then reacted with the tosylate precursor toprovide 18F-fallypride as previously described (Mukher-jee et al., 1995a). For purification of the radiolabeledproduct, high performance liquid chromatography(HPLC) separation was carried out on a Gilson Gradi-ent System consisting of two Gilson pumps and one UVdetector with wavelength fixed at 280 nm and a radia-tion flow detector with a NaI(Tl) crystal. Solvent A ofthe gradient was 0.01 M phosphoric acid, pH 4.7, andsolvent B was acetonitrile. Semiprep (250 3 10 mm)C18 columns from Alltech Associates Inc. were used forreverse-phase HPLC. Separation was carried out byusing gradient elution with 0.01 M phosphoric acid andacetonitrile (from 0% acetonitrile and 100% 0.01 Mphosphoric acid at 0 minute, the gradient compositionwent to 60% acetonitrile and 40% 0.01 M phosphoricacid in 10 minutes, and was maintained at this ratio(60:40) up to 40 minutes, the flow rate was 3 ml/min).The retention time of 18F-fallypride was 12.7 minutes(Mukherjee et al., 1995a). The radioligand was pro-duced in approximately 10–20 mCi batches; specificactivities ranged between 800 and 1,000 Ci/mmole. Theradioactivity was also quantified with a Capintec radio-isotope calibrator (CRC-30). The final product (approxi-mately 3.0 mCi) of the radioligand was taken up insterile saline and filtered through a millipore filter(0.22 µm) for intravenous administration to the mon-key. PET experiments on monkeys were carried out in a3-ring PETT VI tomograph (Ter-Pogossian et al., 1982).The animal care and use procedures for all the experi-ments in rats and monkeys were approved by theInstitutional Animal Care and Use Committee of theUniversity of Chicago.
In vitro kinetic studies in rat brain tissue
Male Sprague-Dawley rats (150–175 g) were killed;the brain was removed and dissected, and the striatumwas stored at 280°C if not used on the day of death. Onthe day of assay, the striatum was homogenized with aTekmar Tissumizer (15 sec at half-maximum speed) ina 100-fold (w:v) dilution of a 50-mM Tris HCl buffer, pH7.4, containing 120 mM NaCl, 5 mM KCl, 2 mM CaCl2,1 mM MgCl2, 1 mM NaEDTA, and 0.1 mM Na ascor-bate. The homogenate was centrifuged at 12,0003 g for15 min at 4°C. The pellet was resuspended in the samevolume of buffer, centrifuged a second time, and resus-pended in fresh buffer at a concentration of 10 mg oftissue/ml. For association kinetics, the incubation mix-ture consisted of 0.10 ml of 18F-fallypride (0.5 nM stock,specific activity of 500 Ci/mmol), 0.80 ml of buffer and0.10 ml of tissue (stock of 10 mg/ml) to provide a totalincubation volume of 1.0 ml. The mixtures were incu-bated for 1 hour at 25°C. Binding was initiated byadding tissue homogenate to the incubation mixture atvarious times (range between 1 to 120 minutes) fol-lowed by filtration using Brandel filtration apparatus of
the incubation mixtures followed by 3 3 5 ml washeswith cold buffer. Nonspecific binding was measuredusing 100 µM of sulpiride. Similarly, dissociation kinet-ics were carried out under similar concentrations oftissue and 18F-fallypride. The mixtures were incubatedfor 1 hour at 25°C or 37°C. The dissociation of 18F-fallypride was initiated by adding an excess of thedopamine (10 µM or 100 µM final concentration) andhaloperidol (10 µM final concentration) followed byfiltration of the incubation mixtures at various timepoints (range of 1 to 60 minutes). The filters werecounted in a well-counter connected to an Ortec Mae-stro MCA Emulator for fluorine-18 radioactivity, decaycorrected and evaluated as counts per minute.
In vivo reserpine studies in rats
A group of eight male Sprague-Dawley rats (250–275g) was administered either reserpine (5 mg/kg in 0.25ml) or saline intraperitoneally, 24 hours before adminis-tration of 18F-fallypride, following reported protocols(DeJesus et al., 1986; Ross and Jackson, 1989; Seemanet al., 1989; Young et al., 1991). The rats were allowedfree access to food and water. The rats were adminis-tered with 18F-fallypride (100 µCi in 0.1 ml) via the tailvein. Sixty minutes after the administration of 18F-fallypride, the rats were killed and the striatum, frontalcortex, cerebellum and blood were isolated into taredvials and counted for fluorine-18 radioactivity. Using astandard of 18F-fallypride, the measured activities inthe tissue and blood samples were converted to percentinjected dose per gram of wet tissue.
In vivo amphetamine studies in rats
A group of eight male Sprague-Dawley rats (150–175g) received, approximately 20 min prior to the adminis-tration of 18F-fallypride, either saline or AMPH (d-amphetamine hydrogen sulfate salt, 5 mg/kg) subcuta-neously in a volume of 0.1 mL. Twenty minutes afterthe saline/AMPH treatment, the rats were adminis-tered with 18F-fallypride (100 µCi in 0.1 mL) via the tailvein. Two hours after the administration of 18F-fallypride, the rats were killed. The brain regionsstriatum, frontal cortex and cerebellum as well as bloodwere isolated into tared vials and counted for fluo-rine-18 radioactivity. Using a standard of 18F-fal-lypride, the measured activities in the tissue and bloodsamples were converted to percent injected dose pergram of wet tissue.
PET studies in monkeys
Two healthy male rhesus monkeys (Macaca mulatta,10.5–13 kg) were used for the PET studies. Each animalwas studied once every 2 weeks. The animal wassedated with ketamine (10 mg/kg) and subsequentlyanesthetized with 0.9–1.5% halothane following anovernight fast. The head of the animal was placed in the
3AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE

gantry of a 3-ring PETT VI tomograph (Ter-Pogossianet al., 1982) and positioned on a ‘‘tailored’’ styrofoamsupport and held in place with the use of adhesive tape.After initial positioning the animal was not moved forthe duration of the scan. Five transaxial image slices(interslice spatial resolution of 8 mm at FWHM and aninterslice distance of 14 mm) parallel to the canthome-atal plane were acquired. One of the slices passedthrough the striata. A transmission scan using a Ge-68/Ga-68 ring source was acquired prior to administrationof the radioligand. The radioligand was administeredintravenously (i.v.) using an indwelling catheter placedin one forearm. Serial venous blood samples of 0.5 mlwere drawn during the course of the PET scan throughan indwelling catheter placed in the other forearm.Dynamic scans of 2.62 min were acquired for <58minutes immediately after administering the radioli-gand followed by one static scan of 7 minutes followedby another set of dynamic scans (2.62-min frames) of<58minutes. Total scanning times ranged between 120to 150 minutes.
Amphetamine studies in monkeys
Pharmacological interventions with AMPH (d-am-phetamine hydrogen sulfate salt, administered intrave-nously over the course of approximately 1 minute) werecarried out at various doses ranging between 0.25mg/kg and 1.5 mg/kg and the administration eitherpreceded the administration of the radioligand or wasadministered following the administration of the radio-ligand. All vital signs, such as heart rate, respiratoryrate and in some instances body temperature wereclosely monitored. A heated recirculating water-filledblanket maintained at 37°C was used to keep themonkeys warm during the course of the PET experi-ment. The peripheral anticholinergic agent, glycopyrro-late (0.01 mg/kg, i.m.) was administered in approxi-mately 1-hour intervals in order tominimize respiratorysecretions in all the PET studies.In order to study the effect of various doses of AMPH
on the binding of 18F-fallypride, a set of experimentswas carried out in which 18F-fallypride was first admin-istered to the monkeys. Subsequently, a dose of AMPHat 45 min postinjection of 18F-fallypride was adminis-tered. The extent of displacement of the specificallybound 18F-fallypride in the striata was evaluated overthe course of next 60 minutes.
PET data analysis
PET scan data were collected using the University ofChicago PETT VI tomograph. For each of the five slicesin each frame, coincidence data were corrected forrandoms and compensated for photon attenuation us-ing measured transmission data. Each slice was thenreconstructed into a 100 3 100 image matrix (2.7 mmpixel size) via filtered backprojection. The counts in
each pixel in the image were then converted to units ofpercent-injected-dose-per-unit-volume through the fol-lowing steps: (1) decay-correction to the time of injec-tion; (2) division by the time duration of each frame toget counts/pixel/sec; (3) conversion to units of concentra-tion (mCi/ml) using calibration factors obtained sepa-rately from a scan of a cylinder phantom (20 cmdiameter by 13 cm length) injected with a knownamount of activity; and finally, (4) division by the dose(in mCi) injected into the monkey. The reduced data infinal form were expressed in units of percent-injecteddose/c.c. An area showing maximal radioligand bindingwithin the striata was delineated in the images. Asimilar area was used for measurement of nonspecifi-cally bound radioligand in the region posterior to thestriatum of the same slice as well as the cerebellumwhich was observed in the lower slice.Time activity curves for the distribution and kinetics
of 18F-fallypride were developed from the total bindingobserved in the striata and nonspecific binding ob-served in the cerebellum. Difference between the bind-ing in the striata and cerebellum was referred to as thespecific binding of 18F-fallypride. The dissociation rateswere measured by plotting time versus ln X/Xo, whereXo is the specific binding of 18F-fallypride at time 0 andX is the specific binding of 18F-fallypride at varioustimes past Xo. The dissociation rate was the slope of thisplot and half-times (T1/2) were computed by using theequation, T1/2 5 0.693/koff. In the case of measuring theeffect of various doses of AMPH, percent change inspecific binding was calculated by the difference inspecific binding at time 0 and 30 min and 30 and 60 minpost-AMPH challenge. A normalized plot (similar tothat reported for other radioligands, Mach et al., 1995)of the specific binding of 18F-fallypride was computed inorder to compare the results amongst several PETexperiments. For purposes of normalization, a conver-sion factor for each PET experiment was obtained fromthe reciprocal of the maximum specific binding (striataminus cerebellum) that was observed in each separatePET experiment.
RESULTSIn vitro studies with 18F-fallypride
In vitro association and dissociation experiments inrat brain tissue with 18F-fallypride were carried out andthe data analyzed using kinetic analysis (Munson andRodbard, 1980). All in vitro binding experiments werecarried out at 25°C. The observed association rateconstant measured from the data shown in Figure 2awas found to be 0.1309 min21. Dissociation of specifi-cally bound 18F-fallypride was initiated at equilibrium(after 60 minutes of incubation of 18F-fallypride withtissue homogenate) by 100 µM dopamine as shown inFigure 2b. The dissociation of 18F-fallypride was rela-tively rapid and at 60 minutes approximately 30% of18F-fallypride still remained specifically bound. The
4 J. MUKHERJEE ET AL.

dissociation rate constant was measured to be 0.0176min21 and is comparable to what has been observed for3H-raclopride (Kohler et al., 1985). The association rateconstant was computed to be 5.30 3 108 M21min21 fromK1 5 Kobs 2 K21/[M], where K1 is the association rateconstant, Kobs is the observed rate constant, K21 is thedissociation rate constant and [M] is the concentrationof the radioligand. The equilibrium dissociation con-stant, KD, obtained from K21/K1 (0.0176/5.30 3 108)was found to be 33 pM which is in close correlation tothe measured Ki of 30 pM for fallypride using 125I-epidepride from competition experiments in rat braintissue (Mukherjee et al., 1995a).Dissociation of 18F-fallypride was also investigated at
37°C using dopamine (10 µM) as well as haloperidol (10µM) in order to compare the in vitro findings with thosefound in vivo using PET. A 10-µM concentration ofdopamine was used in order to maintain the similarityto the extracellular dopamine concentrations found invivo by microdialysis experiments upon AMPH treat-ment (Kuczenski and Segal, 1989; Moghaddam et al.,1993). The results are shown in Figure 3. Dissociationof 18F-fallypride with haloperidol was rapid and almostcomplete (only 2.9% of 18F-fallypride remained undis-placed at 60 minutes) with a dissociation rate constantof 5.24 3 1022 min21 while that with dopamine wassignificantly slower (63.6% of 18F-fallypride remainedundisplaced at 60 minutes) with a dissociation rateconstant of 8.60 3 1023 min21.
In vivo studies in rats with 18F-fallypride
In the reserpine pretreatment experiment, binding inthe striata of the two sets of rats (saline-treated andreserpine-pretreated), 60 min postinjection of 18F-fallypride, was approximately 2% injected dose/g (some-what lower due to the larger size of the rats), compa-rable to that previously reported by Mukherjee et al.(1995a). Binding of 18F-fallypride in the reserpinized
rats was marginally elevated compared to controls butwas not statistically significant as seen in Figure 4. Nosignificant difference in the binding of 18F-fallypridewas observed in frontal cortex, cerebellum and blood inthe two sets of rats.Binding in the striata of the two sets of rats (saline-
treated andAMPH-pretreated), 2 hours postinjection of18F-fallypride, was approximately 2.2% to 3.2% injecteddose/g, comparable to that previously reported byMukherjee et al. (1995a). Binding in other parts of thebrain was small relative to that found in the striata,consistent with the distribution of the D2 receptors.Amphetamine pretreatment had a significant effect onthe binding of 18F-fallypride in the striata 2 hourspostadministration of the radioligand. Figure 5 shows
Fig. 3. Dissociation experiments of 18F-fallypride in rat striatalhomogenates at 37°C. Dissociation was initiated with 10 µM dopamineand 10 µM haloperidol. Dissociation rates of 18F-fallypride were 8.6031023 min21 with dopamine and 5.24 3 1022 min21 with haloperidol.
Fig. 2. a:Association experiment of 18F-fallypride in rat striatal homogenates at 25°C. b: Dissociationexperiment of 18F-fallypride in rat striatal homogenates at 25°C. Dissociation was initiated with 100 µMdopamine.
5AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE

an approximately 23% decrease in the percent injecteddose of 18F-fallypride bound in the striata in amphet-amine-treated rats compared to controls that weretreated with saline. No significant difference in thebinding of 18F-fallypride was observed in frontal cortex,cerebellum and blood in the two sets of rats.
PET studies in nonhuman primateswith 18F-fallypride
Uptake of 18F-fallypride in the rhesus monkey brainwas rapid and accumulated in the brain while clearingfrom the blood. Brain uptake varied between levels of0.04% and 0.08% injected dose/cc in the two monkeys.Radioactivity cleared from nonspecific areas, such ascerebellum and cortex quite rapidly, whereas retentionof the tracer was observed in the striata (Mukherjee etal., 1995a,b). Figure 6 shows the time-activity curves inthe striata and cerebellum in three separate experi-ments. The binding of 18F-fallypride in striata did notchange significantly over a period of more than 2 hours(the clearance rate of 18F-fallypride in the uptakeexperiment was negligible (dissociation rates of ,1024
min21), leading to a total decrease in specific bindingover a period of 2 hours of less than 5%). This result washighly reproducible in both the monkeys. Figure 6 alsoshows little effect of the D1 receptor antagonist SCH23390 on the binding of 18F-fallypride while haloperi-dol, a known D2 receptor antagonist, caused a rapiddisplacement of 18F-fallypride from the striata (Mukher-jee et al., 1995a,b).
PET studies of amphetamine effectson 18F-fallypride in primates
In initial studies on the evaluation of endogenousdopamine competition with 18F-fallypride, we investi-gated the effects of AMPH treatment, both prior andafter administration of the radioligand. Several PETexperiments with 18F-fallypride in two male rhesusmonkeys were carried out. The monkeys received inmany cases a dose of 1 mg/kg of AMPH administeredeither before or after injection of 18F-fallypride.
Preinjection of AMPH
Intravenous administration of AMPH (1 mg/kg) 28min prior to the administration of 18F-fallypride showedrapid specific uptake in the striata (0.06–0.07% ID/g)and clearance of nonspecifically bound radioligand fromother parts of the brain as shown in Figure 7. There wasa gradual decrease of specifically bound 18F-fallyprideas well as evidence of an enhanced clearance of specifi-cally bound 18F-fallypride after administeringAMPH inthe two monkeys. The dissociation rates, koff, of 18F-fallypride without AMPH was negligible but after 28-min preadministration of AMPH it was 4.10 3 1023
min21 indicative of the fact that AMPH-stimulatedrelease of endogenous dopamine has a perturbation onthe specific binding of 18F-fallypride.
Postinjection of AMPH
The effect of AMPH on the binding of 18F-fallypridewas also investigated by administering AMPH at vari-
Fig. 4. Comparison of the percent injected dose of 18F-fallypride inthe various brain regions including blood of two groups of maleSprague-Dawley rats. Rats were either pretreated with saline (n 5 4)or reserpine, 5 mg/kg (n 5 4) 24 h prior to the administration of18F-fallypride. All animals were killed 60 min after administration of18F-fallypride.
Fig. 5. Comparison of the percent injected dose of 18F-fallypride inthe various brain regions including blood of two groups of maleSprague-Dawley rats. Rats were either pretreated with saline (n 5 4)or AMPH, 5 mg/kg (n 5 4) 20 min prior to the administration of18F-fallypride. All animals were killed 2 h after administration of18F-fallypride.
6 J. MUKHERJEE ET AL.

ous times after the administration of 18F-fallypride.There was a decrease of specifically bound 18F-fal-lypride as well as evidence of an enhanced clearance of
specifically bound 18F-fallypride after administeringAMPH in the two monkeys as shown in Figure 8.Shown in Figure 8 is the experiment where AMPH was
Fig. 6. PET data of three separate experiments (control study, SCH 23390 challenge study andhaloperidol challenge study) on 18F-fallypride binding in the striata and cerebellum in rhesus monkeys.
Fig. 7. PET data on 18F-fallypride binding in the striata and cerebellum after 28 min preinjection ofAMPH in the rhesus monkey brain. Specific binding was obtained as the difference of striata andcerebellum. Inset shows plot of 1n X/Xo (Xo 5 specific binding of 18F-fallypride at 32.7 min andX 5 specific binding of 18F-fallypride at various times post-Xo) versus time (y 5 0.1353 2 4.103e 2 3x,R2 5 0.984).
7AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE
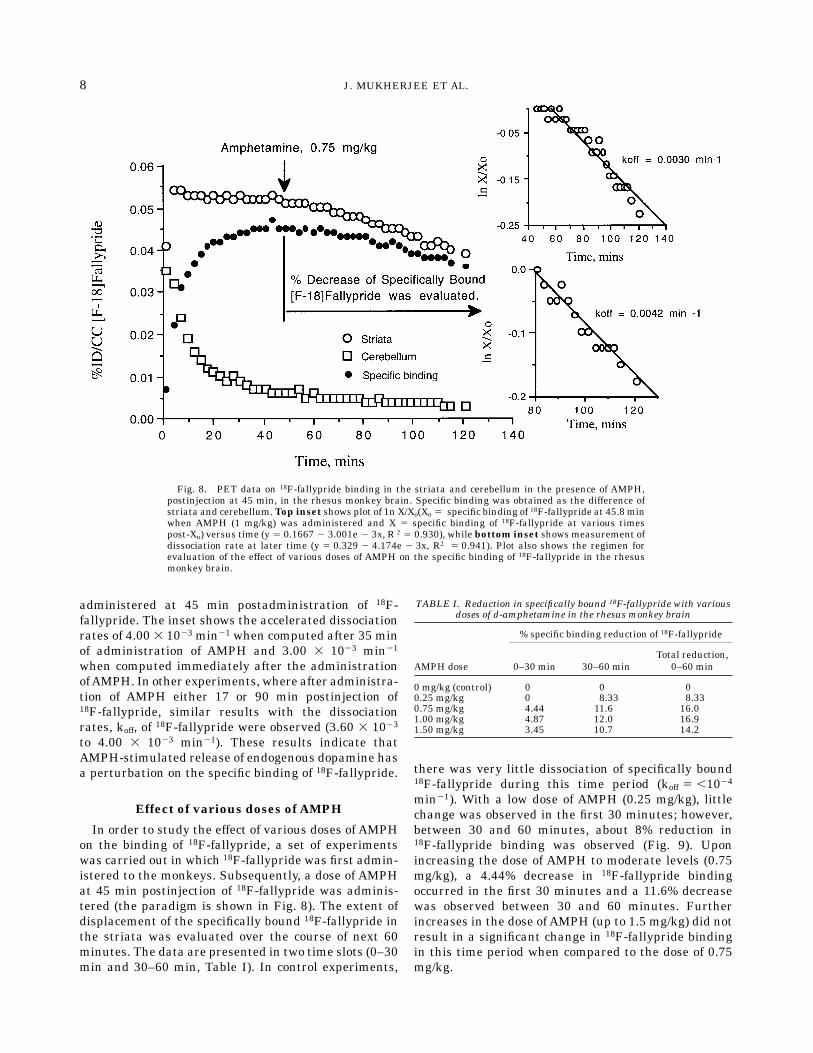
administered at 45 min postadministration of 18F-fallypride. The inset shows the accelerated dissociationrates of 4.00 3 1023 min21 when computed after 35 minof administration of AMPH and 3.00 3 1023 min21
when computed immediately after the administrationofAMPH. In other experiments, where after administra-tion of AMPH either 17 or 90 min postinjection of18F-fallypride, similar results with the dissociationrates, koff, of 18F-fallypride were observed (3.60 3 1023
to 4.00 3 1023 min21). These results indicate thatAMPH-stimulated release of endogenous dopamine hasa perturbation on the specific binding of 18F-fallypride.
Effect of various doses of AMPH
In order to study the effect of various doses of AMPHon the binding of 18F-fallypride, a set of experimentswas carried out in which 18F-fallypride was first admin-istered to the monkeys. Subsequently, a dose of AMPHat 45 min postinjection of 18F-fallypride was adminis-tered (the paradigm is shown in Fig. 8). The extent ofdisplacement of the specifically bound 18F-fallypride inthe striata was evaluated over the course of next 60minutes. The data are presented in two time slots (0–30min and 30–60 min, Table I). In control experiments,
there was very little dissociation of specifically bound18F-fallypride during this time period (koff 5 ,1024
min21). With a low dose of AMPH (0.25 mg/kg), littlechange was observed in the first 30 minutes; however,between 30 and 60 minutes, about 8% reduction in18F-fallypride binding was observed (Fig. 9). Uponincreasing the dose of AMPH to moderate levels (0.75mg/kg), a 4.44% decrease in 18F-fallypride bindingoccurred in the first 30 minutes and a 11.6% decreasewas observed between 30 and 60 minutes. Furtherincreases in the dose of AMPH (up to 1.5 mg/kg) did notresult in a significant change in 18F-fallypride bindingin this time period when compared to the dose of 0.75mg/kg.
Fig. 8. PET data on 18F-fallypride binding in the striata and cerebellum in the presence of AMPH,postinjection at 45 min, in the rhesus monkey brain. Specific binding was obtained as the difference ofstriata and cerebellum.Top inset shows plot of 1n X/Xo(Xo 5 specific binding of 18F-fallypride at 45.8 minwhen AMPH (1 mg/kg) was administered and X 5 specific binding of 18F-fallypride at various timespost-Xo) versus time (y 5 0.1667 2 3.001e 2 3x, R 2 5 0.930), while bottom inset shows measurement ofdissociation rate at later time (y 5 0.329 2 4.174e 2 3x, R2 5 0.941). Plot also shows the regimen forevaluation of the effect of various doses of AMPH on the specific binding of 18F-fallypride in the rhesusmonkey brain.
TABLE I. Reduction in specifically bound 18F-fallypride with variousdoses of d-amphetamine in the rhesus monkey brain
AMPH dose
% specific binding reduction of 18F-fallypride
0–30 min 30–60 minTotal reduction,
0–60 min
0 mg/kg (control) 0 0 00.25 mg/kg 0 8.33 8.330.75 mg/kg 4.44 11.6 16.01.00 mg/kg 4.87 12.0 16.91.50 mg/kg 3.45 10.7 14.2
8 J. MUKHERJEE ET AL.

DISCUSSION
In order to assess the efficacy of dopamine competi-tion with the newly developed high affinity dopamineD2 receptor radioligand 18F-fallypride, we have evalu-ated the effect of dopamine in in vitro experiments andAMPH in in vivo experiments on the binding of 18F-fallypride in the rat and monkey striata. Dopaminecompeted effectively with 18F-fallypride in rat striatalhomogenates. A dissociation rate of 0.0176 min21 wasmeasured in vitro for 18F-fallypride at 25°C with 100µM dopamine. Similar dissociation rates of related highaffinity substituted benzamides like 3H-raclopride (0.023min21 using 1 µM butaclamol, Kohler et al., 1985),125I-NCQ 298 (0.040 min21 at 37°C using 1 µM butacla-mol, Hall et al., 1991) and 125I-epidepride (0.0077 min21
at 25°C using 1 µM haloperidol, Kessler et al., 1993)have been observed at the D2 receptor sites. Theefficient competition of dopamine with 18F-fallypridebinding illustrates fallypride as a competitive inhibitorat the D2 receptor similar to the findings of the competi-tive nature of binding of the related substituted benza-mide, raclopride (Seeman et al., 1989, 1990). The highaffinity of 33 pM for fallypride derived from the rateconstants correlates well with results from equilibriumbinding assays on fallypride (30 pM using 125I-epide-pride, Mukherjee et al., 1995a).The basal extracellular concentration of dopamine in
rats measured by microdialysis has been reported to be23–54 nM (Church et al., 1987; Sharp et al., 1986;Zetterstrom et al., 1983), about 10 nM (Reid et al., 1990)and more recently 14.5 nM (Freeman and Tallarida,
1994). In the in vivo rat studies (saline-pretreated and 5mg/kg reserpine-pretreated, 24 h prior to the adminis-tration of the radioligand), 60 min postinjection of18F-fallypride (a time when 18F-fallypride binding ap-proaches equilibrium levels, Mukherjee et al., 1995a),reserpine had little effect on the extent of binding of18F-fallypride in the striata. Figure 4 shows no signifi-cant difference in reserpine-treated rats compared tocontrols that were treated with saline. These findingsare similar to those reported for 3H-NMSP wherereserpine had little effect, whereas they are differentfrom what has been observed for 3H-raclopride wherereserpine pretreatment increased the binding of 3H-raclopride by about 50% (Ross and Jackson, 1989;Seeman et al., 1989; Young et al., 1991). The elevationin the binding of 3H-raclopride in reserpinized ratsoccurs due to the reduction of dopamine competitionwith 3H-raclopride at the D2 receptors (reserpine, 24 hpostadministration, has been shown to reduce basallevels of dopamine down to undetectable levels bymicrodialysis experiments [Heeringa andAbercrombie,1995] and approximately 3% of control in striatum[DeJesus et al., 1986]). These findings suggest that theconcentration of basal endogenous dopamine (approxi-mately 10 to 50 nM) is sufficient to compete effectivelywith moderate affinity radioligands such as 3H-raclo-pride (Ki 5 1 nM for D2 receptors). However, in the caseof the high affinity radioligands such as 18F-fallypride(Ki 5 30 pM for D2 receptors), competition from the10–50 nM of dopamine is significantly lower.
Fig. 9. PET data on 18F-fallypride binding in the striata and cerebellum in the presence of a low doseAMPH (0.25 mg/kg), postinjection at 45 min, in the rhesus monkey brain. Inset shows plot of 1n X/Xo(Xo 5 specific binding of 18F-fallypride at 45.8 min when AMPH was administered and X 5 specificbinding of 18F-fallypride at various times post-Xo) versus time Displacement of 18F-fallypride occurredafter 35 min with an off-rate of 3.6 3 1023 min21 (y 5 0.2944 2 36422e 2 3x, R2 5 0.905).
9AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE

It has been reported that in anesthetized rats, AMPH(2 mg/kg) resulted in a 20-fold increase in extracellularstriatal dopamine as measured by microdialysis (Humeet al., 1992) and in a freely moving rat, upon subcutane-ous administration of AMPH (5 mg/kg), extracellulardopamine concentration was found to be 1.26 µMwithin 20–40 minutes post-AMPH injection and thedopamine levels remained elevated for more than 3hours (Kuczenski and Segal, 1989). In the in vivo ratAMPH studies with 18F-fallypride (saline-pretreatedand 5 mg/kg AMPH-pretreated 20 minutes prior toradioligand injection), AMPH had a significant effect onthe binding of 18F-fallypride in the striata 2 hourspostinjection of 18F-fallypride. Figure 5 shows an ap-proximately 23% decrease in AMPH-treated rats com-pared to controls that were treated with saline. ThisAMPH-induced decrease in binding of 18F-fallypride isanalogous to that observed for 3H-raclopride althoughsmaller in magnitude (binding of 3H-raclopride de-creased by 41.5% byAMPH at a dose of 10mg/kg, Younget al., 1991). On the other hand, 3H-NMSP, which hasbeen shown to be sensitive to an AMPH dose of 10mg/kg, was not sensitive to AMPH doses of 5 mg/kg(Young et al., 1991).In PET experiments using anAMPH challenge, there
was a decrease of specifically bound 18F-fallypride aswell as evidence of an enhanced clearance of specificallybound 18F-fallypride after administering AMPH com-pared to the control experiments, in the two monkeys.After 25 min preadministration of AMPH, the dissocia-tion rate was 4.10 3 1023 min21. Furthermore, whenAMPHwas administered at either 17 min, 45 min or 90min postadministration, similar dissociation rates inthe range of 3.60 3 1023 to 4.00 3 1023 min21 wereobserved. These results indicate that AMPH-stimu-lated release of endogenous dopamine significantlyalters the specific binding of 18F-fallypride. The dissocia-tion rates of 18F-fallypride in the AMPH challengestudies are about an order of magnitude slower com-pared to displacement of 18F-fallypride with haloperidol(dissociation rate of 3.85 3 1022 min21, Mukherjee etal., 1995a) or raclopride (2.20 3 1022 min21, Mukherjeeet al., 1995b) while they are more than an order ofmagnitude faster than the baseline studies.It is noteworthy to compare the rates of dissociation
of 18F-fallypride using dopamine (an agonist) and halo-peridol (an antagonist). Using the same concentration(10 µM) of the two compounds, the dissociation ratesand the extent of displacement of 18F-fallypride fromrat striatal homogenates in vitro were found to besignificantly different. Dissociation with haloperidol(5.24 3 1022 min21) was found to be more than sixtimes faster than with dopamine (8.60 3 1023 min21).These findings are similar to what has been observed invivo with 18F-fallypride using haloperidol and withAMPH-induced release of dopamine. The in vivo disso-ciation rate with haloperidol was found to be 3.85 3
1022 min21 which compares well with the in vitro rateof 5.24 3 1022 min21. The rates found in vivo withAMPH-induced dopamine release (3.60 3 1023 to4.10 3 1023 min21) were slower and were found to bemore similar to the rates found in vitro with dopamine(8.60 3 1023 min21). It must be noted that the dopa-mine concentration used in the in vitro experimentswas 10 µM, which is somewhat higher than the concen-trations ofAMPH-induced release of extracellular dopa-mine concentrations found in vivo by microdialysis (0.1to 2.0 µM, Kuczenski and Segal, 1989; Moghaddam etal., 1993). This set of experiments provides evidence forthe similarity of the in vitro and in vivo dissociationrates of 18F-fallypride using haloperidol as well asprovide a comparison of dopamine in vitro versusAMPH-induced release of dopamine in vivo. Further-more, they also highlight the difference between effectsof an antagonist (haloperidol) and an agonist (dopa-mine) on the binding of 18F-fallypride. The extent ofdisplacement of 18F-fallypride with 10 µM haloperidolwas complete, whereas 10 µM dopamine was able todisplace less than 60% of specifically bound 18F-fallypride. It is known that dopamine D2 receptors existin two interconvertible affinity states, the high- andlow-affinity states, which have different affinities fordopamine binding (Kd
high 5 7.5 nM and Kdlow 5 4.3 µM,
Seeman et al., 1986). Since 18F-fallypride binds at thetwo sites with a similar affinity (approximately 30 pM),an antagonist such as haloperidol (Kd
high or low 5 1.48nM, Seeman et al., 1986) is able to compete with18F-fallypride equally at the two sites. However, dopa-mine (or agonists) competes with 18F-fallypride effec-tively only at the high-affinity site. At the low-affinitysites, dopamine competes with 18F-fallypride sluggishlydue to its low micromolar affinities and thus leaves asignificant component of specifically bound 18F-fal-lypride undisplaced. Therefore, in comparing displace-ment experiments of radioligands in vitro and in vivo, itis essential to take into account among other factors,the pharmacological differences (such as antagonists oragonists) of the agents that are used to carry out thedissociation.Amphetamine-induced reductions in the specific bind-
ing of various dopamine D2 receptor PET and SPECTradioligands have been reported by various investiga-tors and are summarized in Table II. Based on the invitro data on the sensitivity of raclopride towardsendogenous dopamine, PET experiments in nonhumanprimates with AMPH show a 16% reduction in thestriata to cerebellum distribution volume of 11C-raclopride (Dewey et al., 1993). A similar reduction hasbeen observed in humans when administered with anoral dose of AMPH (Farde et al., 1992). Similar dopa-mine effects have been found for IBZM, where AMPHaccelerated the rates of clearance of the radioligandfrom striata in baboons and humans (Innis et al., 1992;Laruelle et al., 1995). On the other hand, the higher
10 J. MUKHERJEE ET AL.

affinity of 3H-NMSP for the D2 receptor and its slowerkinetics in vivo in comparison to raclopride allows littlecompetitionwith endogenous dopamine in in vivo experi-ments in rats (Seeman et al., 1989; Young et al., 1991).However, experiments in baboons using 18F-NMSPreport some reduction of 18F-NMSPbinding uponAMPH(1 mg/kg) pretreatment (Dewey et al., 1991; Logan etal., 1991). Similar findings have been reported for thehigher affinity substituted benzamide, 123I-epidepride,where little (8–10%) or no effect of AMPH was noted inrhesus monkeys and baboons (Kessler et al., 1993;Al-Tikriti et al., 1994).It has been suggested that a direct relationship
between the affinity of the radioligand for the D2
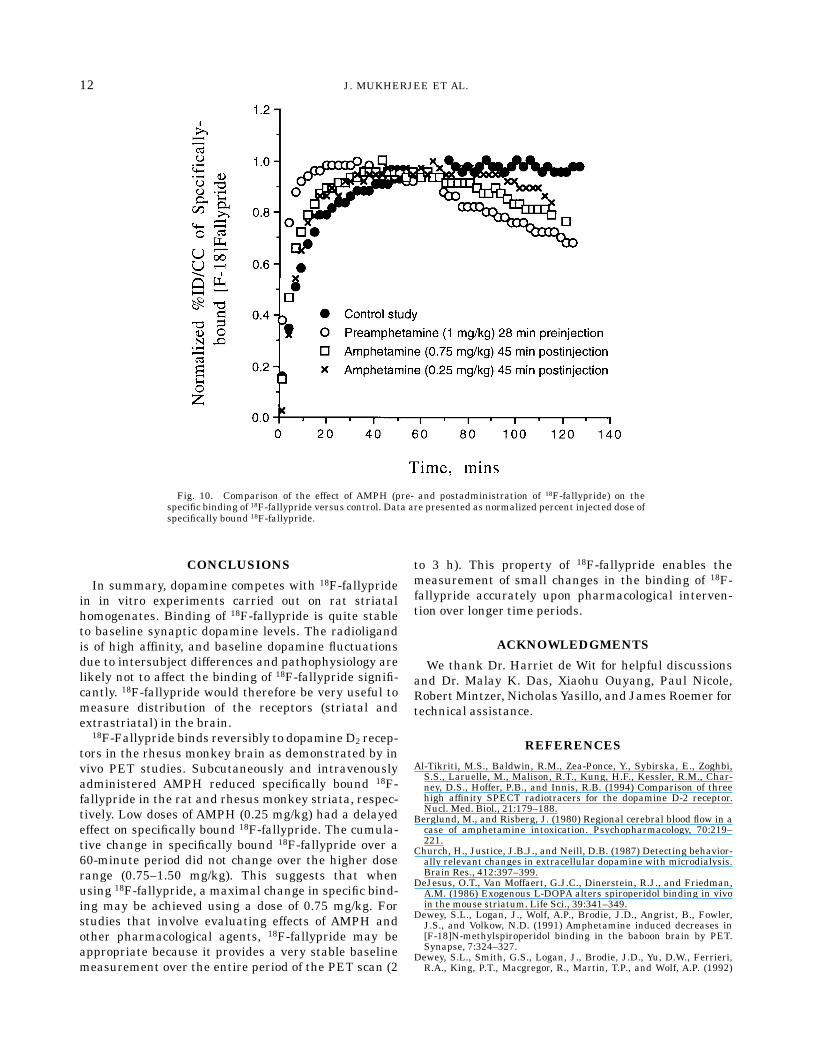
receptors and its propensity of competition with dopa-mine might exist (Seeman et al., 1990). Fallypride(Ki 5 0.03 nM) is a high affinity dopamine D2 receptorantagonist compared to raclopride (Ki 5 1 nM). Al-though the affinity of fallypride is about 30 times betterthan raclopride, AMPH-induced release of endogenousdopamine is able to compete with both raclopride andfallypride. However, raclopride due to its lower affinityis more sensitive to basal dopamine levels. On the otherhand, fallypride due to its higher affinity is more stableto basal dopamine level fluctuations as shown in Figure10 (basal levels measured in anesthetized rhesus mon-key using microdialysis in the caudate-putamen hasbeen reported to be 31 nM, Moghaddam et al., 1993).This stability of the baseline of 18F-fallypride in controlstudies allows evaluation of AMPH effects more pre-cisely, an issue that has been raised previously (Al-Tikriti et al., 1994).
It is known that the primary pharmacological effectof AMPH arises from the release of stored dopamine inthe neurons. However, since AMPH has been shown toincrease blood flow (Berglund andRisberg, 1980;McCul-loch et al., 1977), measurement of changes in thespecific binding of radioligands may be confounded andmay not necessarily be able to exclude alterations in thedegree of binding due to changes in blood flow. Studieson blood flow changes in baboons using AMPH indicatemaximal flows of about 30% within 15 min and returnto baseline values by 45 min (McCulloch et al., 1977).Using microdialysis, striatal dopamine has been shownto rise more than 20-fold with intravenous AMPHtreatment (1 mg/kg) in the monkey in approximately30–40 min (Moghaddam et al., 1993). In our preinjec-tion experiments, AMPH was administered approxi-mately 28 min prior to the injection of 18F-fallypride,thus minimizing effects of flow. Dissociation of 18F-fallypride in this experiment was similar to the ones inwhichAMPHwas administered at later points post-18F-fallypride administration. Also, in our small dose (0.25mg/kg) of AMPH study, reduction in the binding of18F-fallypride was observed approximately 30 min post-AMPH administration, a time when alterations inblood flow have subsided and levels of synaptic dopa-mine have peaked. Together, these results suggest thatthe reduction in the binding of 18F-fallypride in thestriata is most likely due to dopamine competition atthe D2 receptor sites.In AMPH studies with various doses, it was observed
that the degree of reduction was smaller during thefirst 30 min in all the doses studied (Table I) andcorresponds to the slower dissociation rates during thistime period (Fig. 8, top inset). The reduction in specificbinding was more than twice as fast during the secondtime period of 30 to 60 min in all the doses studied (doseresponse studies of AMPH in rats report a similar timecourse for release of dopamine, Kuczenski et al., 1991).Interestingly, this coincides with microdialysis datawhich show maximal increases in extracellular dopa-mine in the caudate-putamen in about 30–40 min(Moghaddam et al., 1993). This indicates the require-ment of a relatively high dopamine concentration in thesynapse in order to effectively compete with 18F-fallypride. Lower doses of AMPH (0.25 mg/kg) did notshow any immediate effect on the binding of 18F-fallypride. A measurable effect occurred during thesecond time period, approximately 30 minutes postad-ministration of AMPH. The extent of reduction in thebinding 18F-fallypride plateued at doses of 0.75 to 1.50mg/kg (Table I) with the total reduction being approxi-mately 16% per hour. The observed half-time of 18F-fallypride clearance in our in vivo AMPH experimentswas 165 min (from the dissociation rate of 4.20 3 1023
min21) which is about two times slower than thehalf-time of 80.6 min for the in vitro 10 µM dopamineinitiated dissociation in rat striata.
TABLE II. Competition of various PET/SPECT radioligands withamphetamine-induced endogenously released dopamine
Radioligand AMPH dose Species%
reduction Reference
11C-Raclopride 1 mg/kg, i.v. Baboons 16.2 130 mg, p.o. Humans 10 6 5 2
18F-Desmethoxy-fallypride 1 mg/kg, i.v. Rhesus 36 3
18F-Fallypride 1 mg/kg, i.v. Rhesus 16/hr 4123I-IBZM 1 mg/kg, i.v. Baboons 22/hr 5
0.3 mg/kg, i.v. Humans 15/hr 6123I-Epidepride 1 mg/kg, i.v. Rhesus none 7
1–2 mg/kg, i.v. Rhesus 8–10 81Female baboons (pappio anubis, 13–18 kg), preinjection of AMPH (5 min beforeradioligand). Reported change is in the striata to cerebellum distribution volume(Dewey et al., 1993).2Human study with AMPH taken orally, 90 min preinjection of the radioligand(Farde et al., 1992).3Male rhesus monkeys (Macaca mulatta, 10–13 kg), preinjection of AMPH, 28min before radioligand. Rate of washout in controls was 20% and in AMPHpretreated it was 56% (Mukherjee et al., 1996).4Male rhesus monkeys (Macaca mulatta, 10–13 kg), postinjection of AMPH (45min after radioligand; Mukherjee et al., this work).5Female baboons (Pappio anubis, 8–13 kg), postinjection of AMPH, 99 6 6 minafter radioligand. Rate of washout in controls was 11%/h andAMPH-treated was33%/h (Innis et al., 1992).6Human study was carried out under steady-state conditions (Laruelle et al.,1995).7Female baboons (Pappio anubis, 8–12 kg), postinjection of AMPH, 90 min afterradioligand (Al-Tikriti et al., 1994).8Male rhesus monkeys (8–10 kg), postinjection of AMPH, 250 min after radioli-gand (Kessler et al., 1993).
11AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE
CONCLUSIONS
In summary, dopamine competes with 18F-fallypridein in vitro experiments carried out on rat striatalhomogenates. Binding of 18F-fallypride is quite stableto baseline synaptic dopamine levels. The radioligandis of high affinity, and baseline dopamine fluctuationsdue to intersubject differences and pathophysiology arelikely not to affect the binding of 18F-fallypride signifi-cantly. 18F-fallypride would therefore be very useful tomeasure distribution of the receptors (striatal andextrastriatal) in the brain.
18F-Fallypride binds reversibly to dopamine D2 recep-tors in the rhesus monkey brain as demonstrated by invivo PET studies. Subcutaneously and intravenouslyadministered AMPH reduced specifically bound 18F-fallypride in the rat and rhesus monkey striata, respec-tively. Low doses of AMPH (0.25 mg/kg) had a delayedeffect on specifically bound 18F-fallypride. The cumula-tive change in specifically bound 18F-fallypride over a60-minute period did not change over the higher doserange (0.75–1.50 mg/kg). This suggests that whenusing 18F-fallypride, a maximal change in specific bind-ing may be achieved using a dose of 0.75 mg/kg. Forstudies that involve evaluating effects of AMPH andother pharmacological agents, 18F-fallypride may beappropriate because it provides a very stable baselinemeasurement over the entire period of the PET scan (2
to 3 h). This property of 18F-fallypride enables themeasurement of small changes in the binding of 18F-fallypride accurately upon pharmacological interven-tion over longer time periods.
ACKNOWLEDGMENTS
We thank Dr. Harriet de Wit for helpful discussionsand Dr. Malay K. Das, Xiaohu Ouyang, Paul Nicole,RobertMintzer, NicholasYasillo, and James Roemer fortechnical assistance.
REFERENCES
Al-Tikriti, M.S., Baldwin, R.M., Zea-Ponce, Y., Sybirska, E., Zoghbi,S.S., Laruelle, M., Malison, R.T., Kung, H.F., Kessler, R.M., Char-ney, D.S., Hoffer, P.B., and Innis, R.B. (1994) Comparison of threehigh affinity SPECT radiotracers for the dopamine D-2 receptor.Nucl. Med. Biol., 21:179–188.
Berglund, M., and Risberg, J. (1980) Regional cerebral blood flow in acase of amphetamine intoxication. Psychopharmacology, 70:219–221.
Church, H., Justice, J.B.J., and Neill, D.B. (1987) Detecting behavior-ally relevant changes in extracellular dopamine with microdialysis.Brain Res., 412:397–399.
DeJesus, O.T., Van Moffaert, G.J.C., Dinerstein, R.J., and Friedman,A.M. (1986) Exogenous L-DOPA alters spiroperidol binding in vivoin the mouse striatum. Life Sci., 39:341–349.
Dewey, S.L., Logan, J., Wolf, A.P., Brodie, J.D., Angrist, B., Fowler,J.S., and Volkow, N.D. (1991) Amphetamine induced decreases in[F-18]N-methylspiroperidol binding in the baboon brain by PET.Synapse, 7:324–327.
Dewey, S.L., Smith, G.S., Logan, J., Brodie, J.D., Yu, D.W., Ferrieri,R.A., King, P.T., Macgregor, R., Martin, T.P., and Wolf, A.P. (1992)
Fig. 10. Comparison of the effect of AMPH (pre- and postadministration of 18F-fallypride) on thespecific binding of 18F-fallypride versus control. Data are presented as normalized percent injected dose ofspecifically bound 18F-fallypride.
12 J. MUKHERJEE ET AL.

GABAergic inhibition of endogenous dopamine release measured invivo with C-11 raclopride and PET. J. Neurosci., 12:3773–3780.
Dewey, S.L., Smith, G.S., Logan, J., Simkowitz, P., Brodie, J.D.,Fowler, J.S., Volkow, N.D., and Wolf, A.P. (1993) Striatal binding ofthe PET ligand, [C-11]raclopride is altered by drugs that modifysynaptic dopamine levels. Synapse, 13:350–356.
Farde, L., Nordstrom, A.L., Wiesel, F.A., Pauli, S., Halldin, C., andSedvall, G. (1992) Positron emission tomography analysis of centralD1 and D2 dopamine receptor occupancy in patients treated withclassical neuroleptics and clozapine. Arch. Gen. Psychiatry, 49:538–544.
Freeman, K.A., and Tallarida, R.J. (1994) A quantitative study ofdopamine control in rat striatum. J. Pharmacol. Exp. Ther., 268:629–638.
Friedman, A.M., DeJesus, O.T., Revenaugh, J., and Dinerstein, R.J.(1984) Measurements in vivo of parameters of the dopamine system.Ann. Neurol., 15 Suppl:S66–76.
Gifford, A.N., Gatley, S.J., and Ashby, C.R. (1996) Endogenouslyreleased dopamine inhibits the binding of dopaminergic PET andSPECT ligands in superfused rat striatal slices. Synapse, 22:232–238.
Hall, H., Hogberg, T., Halldin, C., Kohler, C., Strom, P., Ross, S.B.,Larsson, S.A., and Farde, L. (1991) NCQ 298, a new selectiveiodinated salicylamide ligand for the labeling of dopamine D2receptors. Psychopharmacology, 103:6–18.
Heeringa, M.J., andAbercrombie, E.D. (1995) Biochemistry of somato-dendritic dopamine release in substantia nigra: An in vivo compari-son with striatal dopamine release. J. Neurochem., 65:192–200.
Hume, S.P., Myers, R., Bloomfield, P.M., Opacka-Juffry, J., Cremer,J.E., Ahier, R.G., Luthra, S.K., Brooks, D.J. and Lammertsma, A.A.(1992) Quantitation of carbon-11 labeled raclopride in rat striatumusing positron emission tomography. Synapse, 12:47–54.
Innis, R.B., Malison, R.T., Al-Tikriti, M., Hoffer, P.B., Sybirska, E.H.,Seibyl, J.P., Zoghbi, S.S., Baldwin, R.M., Laruelle, M., Smith, E.O.,Charney, D.S., Heninger, G., Elsworth, J.D., and Roth, R.H. (1992)Amphetamine stimulated dopamine release competes in vivo for123I-IBZM binding to the D-2 receptor in non-human primates.Synapse, 10:177–184.
Kessler, R.M., Votaw, J.R., Schmidt, D.E., Ansari, M., Holdeman, K.P.,dePaulis, T., Clanton, J.A., Pfeffer, R., Manning, R.G., and Ebert,M.H. (1993) High affinity dopamine D-2 receptor radioligands. 3.[123I] and [125I]Epidepride: In vivo studies in rhesus monkey brainand comparison with in vitro pharmacokinetics in rat brain. LifeSci., 53:241–250.
Kohler, C., Hall, H., Ogren, S-O., and Gawell, L. (1985) Specific in vitroand in vivo binding of 3H-raclopride. Biochem. Pharmacol., 34:2251–2259.
Kuczenski, R., and Segal, D. (1989) Concomitant characterization ofbehavioral and striatal neurotransmitter response to amphetamineusing in vivo microdialysis. J. Neurosci., 9:2051–2086.
Kuczenski, R., Segal, D., and Aizenstein, M.L. (1991) Amphetamine,cocaine, and fencamfamine: Relationship between locomotor andstereotypy response profiles and caudate and accumbens dopaminedynamics. J. Neurosci., 11:2703–2712.
Laruelle, M., Abi-Dargham, A., van Dyck, C.H., Rosenblatt, W.,Zea-Ponce, Y., Zoghbi, S.S., Baldwin, R.M., Charney, D.S., Hoffer,P.B., Kung, H.F., and Innis, R.B. (1995) SPECT imaging of striataldopamine release after amphetamine challenge. J. Nucl. Med.,36:1182–1190.
Logan, J., Dewey, S.L., Wolf, A.P., Fowler, J., Brodie, J.D., Angrist, B.,Volkow, N.D., and Gatley, S.J. (1991) Effects of endogenous dopa-mine on measures of [F-18]NMSP binding in the basal ganglia:Comparison of simulations and experimental results from PETstudies in baboons. Synapse, 9:195–207.
Mach, R.H., Ehrenkaufer, R.L.E., Nader, M.A., Line, S.W., Smith,C.R., Hubbard, C., Kung, M-P., Kung, H.F., Luedtke, R.R., and
Morton, T.E. (1995) Validation studies of [18F]fluoroclebopride([18F]FCP): A benzamide analogue that binds reversibly to dopa-mine D2 receptors. J. Label. Compds. Radiopharm., 37:21–23.
McCulloch, J., Deshmukh, V.D., and Harper, A.M. (1977) Indirectsympathomimetics and cerebral blood flow and metabolism. ActaNeurolg. Scand., S64:94–95.
Moghaddam, B., Berridge, C.W., Goldman-Rakic, P.S., Bunney, B.S.,and Roth, R.H. (1993) In vivo assessment of basal and drug-induceddopamine release in cortical and subcortical regions of the anesthe-tized primate. J. Neurosci., 13:215–222.
Mukherjee, J., Yang, Z.-Y., Das, M.K., and Brown, T. (1995a) Fluori-nated benzamide neuroleptics-3. Development of (S)-N-[(1-allyl-2-pyrrolidinyl)methyl]-5-(3-[F-18]fluoropropyl)-2,3-dimethoxy- benza-mide as an improved dopamine D-2 receptor tracer. Nucl. Med. Biol.,22:283–296.
Mukherjee, J., Yang, Z.-Y., Brown, T., Jiang, M., Kapp, O., Chen, C.-T.,and Cooper, M. (1995b) Potential anomalies of dopamine D-2receptor: Challenges for non-invasive imaging studies by positronemission tomography. Med. Chem. Res., 5:174–192.
Mukherjee, J., Yang, Z-Y., Brown, T., Roemer, J., and Cooper, M. (1996)18F-Desmethoxyfallypride: A fluorine-18 labeled radiotracer withproperties similar to carbon-11 raclopride for PET imaging studiesof dopamine D-2 receptors. Life Sci., 59:669–678.
Munson, P.J., and Rodbard, D. (1980) LIGAND: A versatile approachfor the characterisation of ligand binding systems. Anal. Biochem.,107:220–239.
Ross, S.B., and Jackson, D.M. (1989) Kinetic properties of the accumu-lation of 3H-raclopride in the mouse brain in vivo. Archives Pharma-col., 340:6–12.
Seeman, P., Watanabe, M., Grigoriadis, D., Tedesco, J.L., George, S.R.,Svensson, U., Nilsson, J.L.G., and Neumeyer, J.L. (1986) DopamineD-2 receptor binding sites for agonists: A tetrahedral model. Mol.Pharmacol. 28:391–399.
Seeman, P., Guan, H.-C., and Niznik, H.B. (1989) Endogenous dopa-mine lowers the dopamine D-2 receptor density as measured by[H-3]raclopride: Implications for positron emission tomography ofthe human brain. Synapse, 3:96–97.
Seeman, P., Niznik, H.B., and Guan, H.-C. (1990) Evaluation ofdopamine D-2 receptors in schizophrenia is underestimated byradioactive raclopride. Arch. Gen. Psych., 47:1170–1172.
Seiden, L.S., Sabol, K.E., and Ricaurte, G.A. (1993) Amphetamine:Effects on catecholamine systems and behavior. Ann. Rev. Pharma-col. Toxicol., 32:639–677.
Sharp, T., Zetterstorm, T., and Ungerstedt, U. (1986) An in vivo studyof dopamine release and metabolism in rat brain regions usingintracerebral dialysis. J. Neurochem., 47:113–122.
Ter-Pogossian, M.M., Ficke, D.C., Hood, J.T., Yamamoto, M., andMullani, N.A. (1982) PETT VI: A positron emission tomographutilizing cesium fluoride scintillation detectors. J. Comput. Assist.Tomogr., 6:125–133.
Volkow, N.D., Wang, G.-J., Fowler, J.S., Logan, J., Schlyer, D.J.,Hitzemann, R., Lieberman, J., Angrist, B., Pappas, N., MacGregor,R., Burr, G., Cooper, T., and Wolf, A.P. (1994) Imaging endogenousdopamine competition with [11C]raclopride in the human brain.Synapse, 16:255–262.
Volkow, N.D., Fowler, J.S., Gatley, S.J., Logan, J., Wang, G.J., Ding,Y-S., and Dewey, S.L. (1996) PET evaluation of the dopamine systemof the human brain. J. Nucl. Med., 37:1242–1256.
Young, L.T., Wong, D.F., Goldman, S., Minkin, E., Chen, C., Mat-sumura, K., Scheffel, U., and Wagner, H.N., Jr. (1991) Effects ofendogenous dopamine on kinetics of [3H]N-methylspiperone and[3H]raclopride binding in the brain. Synapse, 9:188–194.
Zetterstorm, T., Sharp, T., Marsden, C.A., and Ungerstedt, U. (1983)In vivo measurement of dopamine and its metabolites by intracere-bral dialysis: Changes after d-amphetamine. J. Neurochem., 41:1769–1773.
13AMPHETAMINE EFFECTS ON 18F-FALLYPRIDE
Related Documents











