System. App!. Microhio!. 21, 33-39 (1998) SYSTEM4TIC AND _©_G_us_ta_v _Fi_sc_he_r V_e_rla_ g _________________ APPLIED MICROBIOLOGY Evaluation of Amplified Ribosomal DNA Restriction Analysis for Identification of Acinetobacter Genomic Species LENIE DI]KSHOORN i , BARBARA VAN HARSSELAAR 1 , INGELA TJERNBERG 2 , PHILIPPE]. M. BOUVET 3 and MARIO VANEECHOUTTE 4 I Department of Medical Microbiology, Leiden University Medical Center, Leiden, The Netherlands 2 Department of Medical Microbiology, Malmoe University Hospital, Malmoe, Sweden ] Unite des Enterobacteries, Institut Pasteur, Paris, France 4 Department of Clinical Chemistry, Microhiology and Immunology, University Hospital Ghent, Ghent, Belgium Received August 28,1997 Summary Further to a previous study, the usefulness of amplified ribosomal DNA restriction analysis (ARDRA) for identification of Acinetobacter genomic species (DNA groups) was tested. A set of 202 Acinetobacter strains of 18 described genomic species and 17 unclassified strains were used. Restriction patterns ob- tained with a standard panel of restriction enzymes CfoI, AluI, MboI, RsaI and MspI allowed for separa- tion of 11 DNA groups. With the additional use of restriction enzymes BfaI and BsmAI, five other (ge- nomic) species could be differentiated, leaving only A. haemolyticus and DNA group 13BJI14TU unsep- arated. With the standard panel of enzymes, ten new ARDRA profiles were noted in 14 unclassified strains. Two other unclassified strains had a profile in common with DNA group 15BJ, but were differ- entiated from this DNA group by restriction with BfaI. One remaining unclassified strain could not be differentiated from DNA group 17 by the standard panel of enzymes or hy the enzymes BfaI and BsmAI. Results demonstrate the utility of ARDRA for identification of most genomic species of Acinetobacter. Furthermore, new ARDRA profiles that were shared by several unclassified strains may indicate so far undescribed genomic species in the genus. Key words: Acinetobacter - Taxonomy - Identification - 16S rDNA - Amplification - Restriction analysis Introduction The genus Acinetohacter comprises at least 18 DNA- DNA homology groups (genomic species) and a number of yet ungrouped strains (BOUVET and GRIMONT, 1986; NISHIMURA et al., 1988; BOUVET and JEANJEAN, 1989; TJERNBERG and URSING, 1989; GERNER-SMIDT and TJERN- BERG, 1993). Elucidation of the ecology of the different groups is a challenging task and for this purpose practi- cal identification methods are required. Initial studies with identification schemes based on phenotypic charac- ters were promising (BOUVET and GRIMONT, 1986, 1987), but later studies have shown that some groups are difficult to distinguish phenotypically (GERNER-SMIDT et al., 1991; KAMPFER et al., 1993; KNIGHT et al., 1993; SODDELL et al., 1993; BERNARDS et al., 1995, 1996). In particular, differentiation between the closely related ge- nomic species A. calcoaceticus (DNA group 1), A. hau- mannii (DNA group 2), and the unnamed groups 3 and 13 sensu Tjernberg and Ursing (TU) appeared to be problematic. Recently, restnctlOn analysis of rDNA amplification products (ARDRA) was explored for its usefulness in identification of Acinetohacter genomic species (VANEE- CHOUTTE et al., 1995). For the 53 strains tested, the ARDRA profiles obtained allowed for identification of most genomic species. In the present study, we studied the ARDRA method for identification of Acinetohacter (genomic) species with an important number of strains of all described genomic species and a set of yet unclassified strains. Results indicate that ARDRA can be used for identification of most but not all genomic species. Fur- thermore, new profile groups were found in the unclassi- fied strains which may indicate the existence of addition- al DNA homology groups. Materials and Methods Bacterial strains: A total number of 219 Acinetobacter strains were used, including type and reference strains from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

System. App!. Microhio!. 21, 33-39 (1998) SYSTEM4TIC AND _©_G_us_ta_v _Fi_sc_he_r V_e_rla_g _________________ APPLIED MICROBIOLOGY
Evaluation of Amplified Ribosomal DNA Restriction Analysis for Identification of Acinetobacter Genomic Species
LENIE DI]KSHOORN i , BARBARA VAN HARSSELAAR1, INGELA TJERNBERG2, PHILIPPE]. M. BOUVET3 and MARIO VANEECHOUTTE4
I Department of Medical Microbiology, Leiden University Medical Center, Leiden, The Netherlands 2 Department of Medical Microbiology, Malmoe University Hospital, Malmoe, Sweden ] Unite des Enterobacteries, Institut Pasteur, Paris, France 4 Department of Clinical Chemistry, Microhiology and Immunology, University Hospital Ghent, Ghent, Belgium
Received August 28,1997
Summary
Further to a previous study, the usefulness of amplified ribosomal DNA restriction analysis (ARDRA) for identification of Acinetobacter genomic species (DNA groups) was tested. A set of 202 Acinetobacter strains of 18 described genomic species and 17 unclassified strains were used. Restriction patterns obtained with a standard panel of restriction enzymes CfoI, AluI, MboI, RsaI and MspI allowed for separation of 11 DNA groups. With the additional use of restriction enzymes BfaI and BsmAI, five other (genomic) species could be differentiated, leaving only A. haemolyticus and DNA group 13BJI14TU unseparated. With the standard panel of enzymes, ten new ARDRA profiles were noted in 14 unclassified strains. Two other unclassified strains had a profile in common with DNA group 15BJ, but were differentiated from this DNA group by restriction with BfaI. One remaining unclassified strain could not be differentiated from DNA group 17 by the standard panel of enzymes or hy the enzymes BfaI and BsmAI. Results demonstrate the utility of ARDRA for identification of most genomic species of Acinetobacter. Furthermore, new ARDRA profiles that were shared by several unclassified strains may indicate so far undescribed genomic species in the genus.
Key words: Acinetobacter - Taxonomy - Identification - 16S rDNA - Amplification - Restriction analysis
Introduction
The genus Acinetohacter comprises at least 18 DNADNA homology groups (genomic species) and a number of yet ungrouped strains (BOUVET and GRIMONT, 1986; NISHIMURA et al., 1988; BOUVET and JEANJEAN, 1989; TJERNBERG and URSING, 1989; GERNER-SMIDT and TJERNBERG, 1993). Elucidation of the ecology of the different groups is a challenging task and for this purpose practical identification methods are required. Initial studies with identification schemes based on phenotypic characters were promising (BOUVET and GRIMONT, 1986, 1987), but later studies have shown that some groups are difficult to distinguish phenotypically (GERNER-SMIDT et al., 1991; KAMPFER et al., 1993; KNIGHT et al., 1993; SODDELL et al., 1993; BERNARDS et al., 1995, 1996). In particular, differentiation between the closely related genomic species A. calcoaceticus (DNA group 1), A. haumannii (DNA group 2), and the unnamed groups 3 and 13 sensu Tjernberg and Ursing (TU) appeared to be problematic.
Recently, restnctlOn analysis of rDNA amplification products (ARDRA) was explored for its usefulness in identification of Acinetohacter genomic species (VANEECHOUTTE et al., 1995). For the 53 strains tested, the ARDRA profiles obtained allowed for identification of most genomic species. In the present study, we studied the ARDRA method for identification of Acinetohacter (genomic) species with an important number of strains of all described genomic species and a set of yet unclassified strains. Results indicate that ARDRA can be used for identification of most but not all genomic species. Furthermore, new profile groups were found in the unclassified strains which may indicate the existence of additional DNA homology groups.
Materials and Methods
Bacterial strains: A total number of 219 Acinetobacter strains were used, including type and reference strains from
34 L. DljKSHOORN et a!.
public culture collections and field strains, mostly from clinical sources in France, Sweden and The Netherlands. Two hundred and two strains had been assigned to existing DNA groups by DNA-DNA hybridization, and many of these had been investigated for other features as well (BOUVET and GRIMONT, 1986; BOUVET and ]EANJEAN, 1989; TJERNBERG and URSING, 1989; DUKSHOORN et a!., 1990; GERNER-SMIDT et a!., 1991; KAMPFER et ai., 1993). Seventeen strains were as yet not identifiable to any described DNA group by DNA-DNA hybridization. Throughout the report these strains are called unclassified strains. A list with detailed information of the strains is available on request.
ARDRA: The ARDRA method was performed as described previously (VANEECHOUTTE et a!., 1995) with minor modifications. Strains were grown on Iso-Sensitest agar (CM 471, Oxoid, Hampshire, England) at 30°C. A 10-pl loopful of colony growth was suspended in 300 pi milliQ water (Millipore B.V., Etten-Leur, The Netherlands), heated at 100°C for 10 min, and briefly centrifuged. With filter-protected tips, 3 pi supernatant was added to 47 pi PCR mix. The mix contained 200 pM of each deoxyribonucleoside triphosphate (Pharmacia, Biotech, Uppsala, Sweden), 0.3 pM of each primer (Eurogentec, Maastricht, The Netherlands), and 0.2 U of SuperTth polymerase (Sphaeroq, Leiden, The Netherlands) in reaction buffer (1.5 mM MgCI2, and 50 mM KCI in 10 mM Tris-HCI (pH 8.3), or 0.3 U of GoldStar* DNA polymerase (Eurogentec, Maastricht, The Netherlands) in reaction buffer supplied by the manufacturer. Reaction mixtures were overlaid with mineral oil and PCR was carried out with an initial denaturation of 6 min at 94°C, followed by 34 cycles of 45 sec at 94 °C, 45 sec at 60°C, and 4 min at 72 °C, and a final extension of 16 min at 72 0C. Oligonucleotide primers derived from conserved regions at the edges of the 16S rDNA were obtained from Eurogentec. Presence and yield of PCR products were verified by agarose electrophoresis using Ultra Pure™ agarose (Gibco BRL, Breda, The Netherlands). Restriction of amplified products was carried out in 20-pl volumes of commercial incubation buffer containing 20 U of restriction enzyme AluI, or 5 U of efaI, MbaI, RsaI or MspI (Gibco), and 3 to 10 pi PCR product depending on its
efo) Alul M M
600
100
I p ttern 1 2 3 4 1
yield. For selected strains, BfaI and BsmAI (New England Biolabs, Beverly, MA) were used in quantities of 5U per 20 pi according to the instructions of the manufacturer. For electrophoresis of restriction fragments, Ultra Pure™ agarose was used.
Results
Comparison with previous results
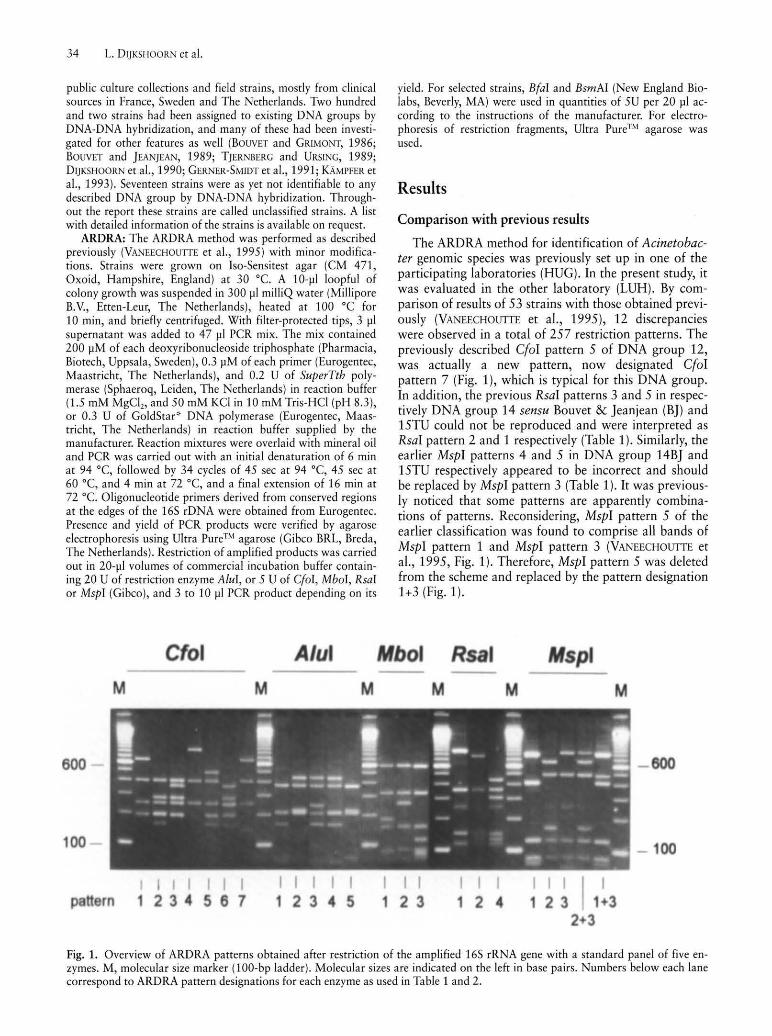
The ARDRA method for identification of Acinetabacter genomic species was previously set up in one of the participating laboratories (HUG). In the present study, it was evaluated in the other laboratory (LUH). By comparison of results of 53 strains with those obtained previously (VANEECHOUTIE et aI., 1995), 12 discrepancies were observed in a total of 257 restriction patterns. The previously described CfaI pattern 5 of DNA group 12, was actually a new pattern, now designated CfaI pattern 7 (Fig. 1), which is typical for this DNA group. In addition, the previous RsaI patterns 3 and 5 in respectively DNA group 14 sensu Bouvet & Jeanjean (BJ) and 15TU could not be reproduced and were interpreted as RsaI pattern 2 and 1 respectively (Table 1). Similarly, the earlier MspI patterns 4 and 5 in DNA group 14BJ and 15TU respectively appeared to be incorrect and should be replaced by MspI pattern 3 (Table 1). It was previously noticed that some patterns are apparently combinations of patterns. Reconsidering, MspI pattern 5 of the earlier classification was found to comprise all bands of MspI pattern 1 and MspI pattern 3 (VANEECHOUTIE et aI., 1995, Fig. 1). Therefore, MspI pattern 5 was deleted from the scheme and replaced by the pattern designation 1+3 (Fig. 1).
Mbol R I
M
pi
1 2 3 1 +3 2+3
- 600
100
Fig. 1. Overview of ARDRA patterns obtained after restriction of the amplified 165 rRNA gene with a standard panel of five enzymes. M, molecular size marker (100-bp ladder). Molecular sizes are indicated on the left in base pairs. Numbers below each lane correspond to ARDRA pattern designations for each enzyme as used in Table 1 and 2.

Interlaboratory reproducibility
Eleven strains, representing the new patterns and the discrepancies with previous results, were investigated blind in both laboratories. The patterns obtained in the laboratories were completely identical for these strains (data not shown). Occasionally, patterns obtained in LUH showed an extra band of approximately 250 bp which was not observed in the UHG laboratory. This band was visible after amplification as a minor band, and was sometimes more clear after digestion, but only in RsaI and MspI patterns (Fig. 1). The presence of the band had no influence on the interpretation of the patterns, but its appearance was reduced by replacement of SuperTth polymerase by GoldStar'~DNA polymerase and by including a hot start in the PCR procedure, by per-
Identification of Acinetobacter genomic species by ARDRA 35
forming annealing during the first five cycles at 65°C, 64°, 63°, 62° and 61°C respectively, followed by 29 cycles with an annealing temperature of 60 0c.
ARDRA profiles in strains of described genomic speCles
In this report, an ARDRA profile is defined as the combination of restriction patterns obtained with the respective enzymes CfoI, AluI, MboI, RsaI and MspI. A summary of the profiles of the classified strains is given in Table 1. Combination patterns for enzymes CfoI, RsaI and MspI, were observed in several DNA groups, with MspI combination patterns being observed most frequently. Intraspecies variation due to the occurrence of two or more restriction patterns and combination pat-
Table 1. ARDRA profiles of 202 Acinetobacter strains previously identified to genomic species by the use of DNA-DNA hybridiza-tiona. Profiles are shown in Figure l.
Genomic species Pattern with enzyme Strains Reference strain
Cfol AZul Mbol RsaI Mspl (N)h
1 (A. calcoaceticus) 2 2 1 1 3 7 ATCC 23055 T
3 2 1 1 3 1 RUH583 2 A. baumannii) 1 1 1 2 3 27 ATCC 17904
1 1 1 2 1 33 ATCC 19606T
1 1 1 2 1+3 4 LMD 82.54 3 2 1 3 1 3 24 ATCC 19004
2+3 1 3 1 3 2 RUH 1163 4 (A. haemolyticus) + 1 4 1 2 2 7 ATCC 17906T
5 (A. junii) e 1 2 1 2 3 6 ATCC 17908 T
6 1 1 2 2 2 2 ATCC 17979 7 (A. johnsonii) + 1 4 1 2 2 8 ATCC 17909T
• 1 4 1 2 2+3 1 RUH2859 8/9 (A. lwoffii) 3 3 2 1 2 19 NCTC 5866T
10 .. 4 2 1 2 3 3 ATCC 17924 11 .. 4 2 1 2 3 7 ATCC 11171 12 (A. radioresistens) 7 3 2 2 1 3 SEIP 12.81
7 3 2 2 3 3 lAM 13186T
7 3 2 2 1+3 2 RUH2862 13 BJI14 TU + 1 4 1 2 2 3 SEIP 5.84
1 4 1 2 3 6 114', RUH 2866
• 1 4 1 2 2+3 6 ATCC 17905 14 BJ 5 5 1 2 3 5 CCUG 14816 15 BJ 1 2 1 2 2 2 79d, RUH 1729 16 1 2 1 4 2 3 ATCC 17988
1 2 1 2+4 2 1 LUH 1734 17 e 1 2 1 2 3 2 SEIP 2/87 13 TU 2 1 1 1 1 2 ATCC 17903
2 1 1 1 3 9 100d
2 1 1 1 1+3 2 RUH2624 15 TU 6 2 1 1 3 2 ISla', LUH 1090
a T, type strain. ATCC, American Type Culture Collection, Rockville, Md, USA; LUH, RUH, collection Leiden University Hospital, Leiden, The Netherlands; LMD, LMD Culture Collection, University of Technology, Delft, The Netherlands; NCTC, National Col-lection of Type Cultures, London, United Kingdom; SEIP, Service des Enterobacteries de l'Institut Pasteur, Paris, France; lAM, Insti-tute of Applied Microbiology, The University of Tokyo, Tokyo, Japan; CCUG, Culture Collection, University of Goteborg, Gote-borg, Sweden. A list with detailed information on all strains is available on request. h (N), Number of strains tested. Markings (+, e, ., .. ) denote profiles occurring in different genomic species. C Strain and designation used by TJERNBERG and URSING, 1989. d Strain and designation used by BOUVET and JEANJEAN, 1989.
36 L. DljKSHOORN et al.
terns was observed in DNA group 1, 2, 3, 7, 12, 13BJI14TU (which are considered to be one DNA group), and in DNA group 16, and BTU. For example, two CfoI patterns, pattern 2 and 3 were observed in A. calcoaceticus (DNA group 1) leading to two ARDRA profiles, i.e. 22113 (in seven strains) and 32113 (in one strain). Remarkably, the type strain of A. calcoaceticus, ATCC 23055T described by BEI]ERINCK as early as in 1911 was shown to have profile 22113 which was indistinguishable from more recent A. calcoaceticus isolates, although it is phenotypically different from these strains (BERNARDS et aI., 1995). The profiles found in DNA group 1, 2, 3, 6, 8/9, 12, 14BJ, 15BJ, 16, BTU, and 15TU were specific for each group in spite of the intraspecies variation within some groups and are thus considered useful for genomic species identification. On the other hand, several genomic species could not be separated, i.e. DNA group 4, 7, 13BJI14TU with the common profile 14122 or 14122+3, DNA group 5 and 17 with profile 12123, and DNA group 10 and 11 with profile 42123.
ARDRA profiles of unclassified strains
Twelve different ARDRA profiles were observed in 17 unclassified strains (Table 2). Profiles 35113, 14143 and 12122 were observed in two, four and two strains, respectively. Profiles 12122 and 12123 were common to those of strains of DNA group 15BJ and 5117 respectively. Ten profiles had not been found before. Four unclassified strains were identified earlier by DNA-DNA hybridization as close to the so-called A. calcoaceticus-A. baumannii (Acb) complex containing A. calcoaceticus,
A. baumannii and the unnamed DNA groups 3 and BTU (GERNER-SMIDT and T]ERNBERG, 1993). Of these, strains 10095 and 10169 were closest to A. calcoaceticus and the unnamed DNA group 3, whereas strains 5804 and 10090 were closest to DNA group BTU. The ARDRA profiles of these pairs of strains were 35113 and 31313/311 +313 respectively, which distinguished them from the Acb complex (Table 1) in spite of the relatively high degree of DNA homology with the complex.
Further discrimination between genomic species by additional restriction enzymes
Various enzymes including BfaI, BsmAI, HaeIII, HinflI, MvaI, Neil, ScrFI, StyI, TaqI, and Tru91 were tested on selected strains for their utility in discriminating genomic species which have ARDRA profiles in common. BfaI and BsmAI showed encouraging results (Table 3), whereas the discriminatory value of the other enzymes was limited (data not shown). All strains of A. haemolyticus, A. johnsonii, and DNA group 13 BJI14TU were tested with enzyme BfaI, all strains of DNA group 10 and 11 were tested with BsmAI, and every strain of DNA group 5 and 17 was investigated with both enzymes. In other cases single strains representative for a given ARDRA profile were tested. With BfaI, ten patterns and a combination pattern were found (Table 3, Fig. 2). Patterns 1 and 2 showed a minor but reproducible difference in migration in a band of an approximate size of 220 bp, while pattern 1+2 was a combination of both. Pattern 2 was only found in DNA group 7, pattern 1 was found in DNA group 4 and in DNA group 13BJI14TU, whereas patterns 1+2,5 and 6
Table 2. ARDRA profiles in strains not yet classifiable by DNA-DNA hybridizationa .
Strain with designation as Specimen Restriction pattern with enzyme ARDRA pattern as received/other designation DNA group
CfoI AluI MboI RsaI MspI
631 b, CCUG 14818 Ear • 1 2 1 2 2 15BJ RUH203 Liquor • 1 2 1 2 2 15BJ 640b, SEIP 1/87 Wound 1 2 1 2 3 5,17 80b, SEIP 14.83 Blood ~ 1 4 1 4 3 930b, SEIP 87.302 Trachea ~ 1 4 1 4 3 944b, SEIP Ac87.316 Wound ~ 1 4 1 4 3 1240h, RUH 65 Clinical ~ 1 4 1 4 3 1271 h, RUH 422 Urine 1 5 1 2 3 MGH 99613, 10095c Abscess e 3 5 1 1 3 MGH 99614,10169' Sputum e 3 5 1 1 3 MGH 99896, 5804c Blood 3 1 3 1 3 MGH 99685, 10090c Ulcer 3 1 1+3 1 3 Isolate patient Kd Blood 1 4 3 4 3 ATCC 13809 Unknown 2 5 1 1 3 RUH 175 Blood 1 2 1 4 3 RUH 581 Soil 3 3 1 1 2 RUH 1139 Pharynx 3 1 1 3
a Markings (., ~, e) denote profiles found in multiple unclassified strains. b BOUVET and JEANJEAN, 1989. c GERNER-SMIDT and TJERNBERG, 1993. d HORREVORTS et aI., 1995.
Identification of Acinetohacter genomic species by ARDRA 37
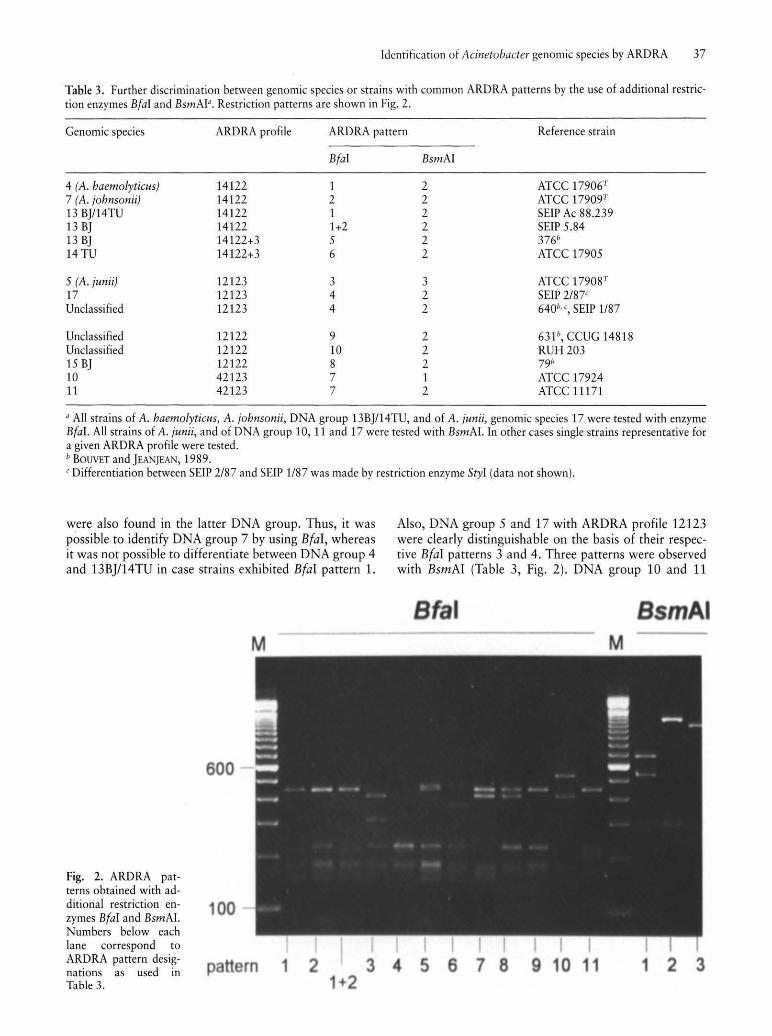
Table 3. Further discrimination between genomic species or strains with common ARDRA patterns by the use of additional restriction enzymes BfaI and BsmAI". Restriction patterns are shown in Fig. 2.
Genomic species ARDRA profile ARDRA pattern Reference strain
4 (A. haemolyticus) 14122 7 (A. johnsonii) 14122 13 BJI14TU 14122 13 BJ 14122 13 BJ 14122+3 14TU 14122+3
5 (A. junii) 12123 17 12123 Unclassified 12123
Unclassified 12122 Unclassified 12122 15 BJ 12122 10 42123 11 42123
BfaI
1 2 1 1+2 5 6
3 4 4
9 10 8 7 7
BsmAI
2 2 2 2 2 2
3 2 2
2 2 2 1 2
ATCC 1790e ATCC 17909T
SEIP Ac 88.239 SEIP 5.84 376h
ATCC 17905
ATCC 17908T
SEIP 2/87' 640h " SEIP 1/87
631h, CCUG 14818 RUH203 791> ATCC 17924 ATCC 11171
a All strains of A. haemolyticus, A. johnsonii, DNA group 13BJI14TU, and of A. junii, genomic species 17 were tested with enzyme BfaI. All strains of A. junii, and of DNA group 10, 11 and 17 were tested with BsmAI. In other cases single strains representative for a given ARDRA profile were tested. h BOUVET and JEANjEAN, 1989. 'Differentiation between SEIP 2/87 and SEIP 1/87 was made by restriction enzyme StyI (data not shown).
were also found in the latter DNA group. Thus, it was possible to identify DNA group 7 by using Bfal, whereas it was not possible to differentiate between DNA group 4 and 13BJ/14TU in case strains exhibited Bfal pattern 1.
Fig. 2. ARDRA patterns obtained with additional restriction enzymes BfaI and BsmAI. Numbers below each lane correspond to ARDRA pattern designations as used in Table 3.
M
600
,00
pattern 1 2 1 2
Also, DNA group 5 and 17 with ARDRA profile 12123 were clearly distinguishable on the basis of their respective Bfal patterns 3 and 4. Three patterns were observed with BsmAl (Table 3, Fig. 2). DNA group 10 and 11
fal mA M
5 7 11 1 2 3

38 L. DUKSHOORN et al.
were discriminated using this enzyme on the basis of their respective patterns 1 and 2. DNA group 5 and 17 also showed differences with BsmAI, although the respective BsmAI patterns 3 and 2 of these groups were less distinct than their BfaI patterns (Fig. 2).
The unclassified strain 5EIP 1187 could not be differentiated from DNA group 17 by the standard panel of enzymes or by restriction with BfaI and BsmAI (Table 3). It was not allocated to DNA group 17 by DNA-DNA hybridization (BOUVET and JEANJEAN, 1989) and in the present study, the strain was distinguished from DNA group 17 by restriction with enzyme StyI (data not shown). Two other unclassified strains, CCUG 14818 and RUH 203, with ARDRA profile 12122 which is specific for DNA group 15BJ (Table 1) were differentiated from strain 79 of DNA group 15BJ by restriction with BfaI.
Discussion
The delineation of the different DNA hybridization groups (genomic species) within the genus Acinetobacter has prompted numerous studies to search for practical methods for identification of the groups. Initial studies were aimed at the exploration of phenotypical methods, but the present trend is to assess the performance of molecular methods. In a previous study (VANEECHOUTTE et a!., 1995), ARDRA was found promising for Acinetobacter genomic species identification. In the present study, it was decided to challenge the method with a large set of strains, and to compare its reproducibility between two participating laboratories. A standard panel of enzymes including CfoI, AZul, MboI, RsaI, and MspI was used for all strains, whereas RsaI was not used for all strains in the first study. Discrepancies with the previous study were checked by both laboratories and the original scheme was revised accordingly. Except for the reassignment of the CfaI pattern of DNA group 12, the revisions did not change the identification scheme nor did they affect the discriminatory power of the method. Apart from the newly recognized CfaI pattern 7, several combination patterns were new. The presence of these combination patterns may be explained by the simultaneous occurrence of multiple copies of 165 ribosomal genes (DOLZANI et a!., 1995) that differ slightly in base sequences.
The profiles obtained with the standard panel of five enzymes were specific for 11 DNA groups (Table 1), whereas the additional use of BfaI and BsmAI allowed for separation of DNA group 10 and 11, DNA group 5 and 17 and discrimination of DNA group 7 from DNA group 4 and 13BJI14TU (Table 3). The value of MspI was limited, since it was only required to separate DNA group 15BJ and 17. A practical approach would be to start with the enzymes CfaI, AZul, MbaI, RsaI, and, depending on the outcome, perform additional restrictions with MspI, BfaI and/or BsmAI. It was important to discover that the genomic groups within the Acb complex which have a relatively high DNA group inter relatedness (65-70%) (TJERNBERG and URSING, 1989) and are phe-
notypically very similar, could be differentiated by ARDRA, even when using CfaI, AZul, and MbaI alone. The (genomic) species A. baumannii, DNA group 3 and 13TU, which are included in the Acb complex, can be highly prevalent in hospitals during outbreaks of infection, and identification of isolates to genomic species is required to establish the sources and mode of transmission in these circumstances.
DNA group 7 was differentiated from DNA group 4 and 13BJI14TU on the basis of a minor difference in BfaI pattern. Therefore, confirmation of the identification of DNA group 7 by a few phenotypic tests (BOUVET and GRIMONT, 1986) is recommended. Of the 17 unclassified strains, three strains were identified incorrectly to DNA groups 5117 or 15BJ by the standard panel of five enzymes (Table 2). Two of these strains were differentiated from the classified strains by the additional use of BfaI and BsmAI (Table 3). Thus, inclusion of these enzymes is recommended for strains with ARDRA pattern 12122 or 12123. In the other unclassified strains, ten new profiles were found which underpins the heterogeneity of the genus. The occurrence of new, identical profiles in different strains may indicate that these strains belong to new, yet undescribed genomic species of the genus Acinetabacter. One possible new group is represented by four strains with profile 14143 (Table 2). Recently, these four strains were also found very similar using AFLpTM, a high resolution genomic fingerprinting method (JANSSEN et a!., 1997). Polyphasic studies including DNA-DNA hybridization, and other genotypic and phenotypic methods are required for further delineation of this group.
Apart from ARDRA, many other molecular methods to identify or differentiate between Acinetabacter DNA groups have been described such as amplification of the 165-235 intergenic spacer region (NOWAK et a!., 1995), restriction analysis of 165-235 rRNA intergenic spacer sequences (DOLZANI et a!', 1995), of the 165 + spacer + complete 23rRNA gene (IBRAHIM et a!., 1996), and of the recA gene (NOWAK and KUR, 1995), tRNA intergenic spacer length polymorphism analysis (WIEDMANN-ALAHMAD et a!., 1994; EHRENSTEIN et a!., 1996), restriction analysis of multiplex PCR products of 165 rDNA and the recA gene (NOWAK and KUR, 1996), sequence analysis of 165 rDNA (RAINEY et a!., 1994; IBRAHIM et a!., 1997) and the gyrB genes (YAMAMOTO and HARAYAMA, 1996) and high resolution genomic fingerprinting by AFLpTM (JANSSEN et a!', 1996). 50me studies were based on a small number of strains or did not include all of the described genomic groups, but most results were promismg.
In conclusion, many methods are of potential use for identification of Acinetabacter genomic species. Among these, ARDRA performs well in the discrimination of most genomic species including those included in the Acb complex, which are difficult to identify phenotypically. Identification by ARDRA can be performed relatively easily and without the need of expensive equipment. Overall, results were consistent and reproducible between the two laboratories. Therefore, with this technique, studies of the ecology of different genomic species

are feasible. However, it is noted that the present classification of acinetobacters is largely based on strains from clinical origin.
Acinetobacters can occupy a wide range of ecological niches, and the genetic diversity of the genus is probably not sufficiently reflected in the present classification. Thus, it is conceivable that strains will be recovered from e.g. environmental sources, which cannot yet be allocated to DNA group by ARDRA or any other by identification system.
Acknowledgement The authors thank Dr. PETER GERNER-SMIDT for providing us
with strains. This work was supported by the Stichting Microbiologie Leiden.
References
BERNARDS, A. T., DIJKSHOORN, L., VAN DER TOORN, J., BOCHNER, B. R., VAN BOVEN, C. P. A.: Phenotypic characterisation of Acinetobacter strains of 13 DNA-DNA hybridisation groups by means of the Biolog system. J. Med. Microbiol. 42, 113-119 (1995).
BERNARDS, A. T., VAN DER TOORN, J., VAN BOVEN, C. P. A., DljKSHOORN, L.: Evaluation of the ability of a commercial system to identify Acinetobacter genomic species. Eur. ]. Clin. Microbiol. Infect. Dis. 15,303-308 (1996).
BEI]ERINCK, M. w.: Uber Pigmentbildung bei Essigbakterien. Zentralbl. Bakteriol. Parasit. 29,169-176 (1911).
BOUVET, P. J. M., GRIMONT, P. A. D.: Taxonomy of the genus Acinetobacter with the recognition of Acinetobaeter baumannii sp. nov., Aeinetobaeter haemolytieus sp. nov., Aeinetobaeter johnsonii sp. nov., and Aeinetobaeter junii sp. nov. and emended descriptions of Aeinetobaeter ealcoacetieus and Acinetobacter lwoffii. Int. ]. System. Bacteriol. 36, 228-240 (1986).
BOUVET, P. J. M., GRIMONT, P. A. D.: Identification and biotyping of clinical isolates of Acinetobacter. Ann. Inst. Pasteur/Microbiol. 138,569-578 (1987).
BOUVET, P. J. M., JEANjEAN, S.: Delineation of new proteolytic genomic species in the genus Acinetobaeter. Res. Microbiol. 140,291-299 (1989).
DIjKSHOORN, L., TjERNBERG, I., POT, B., MICHEL, M. E, URSING, J., KERSTERS, K.: Numerical analysis of cell envelope protein profiles of Acinetobaeter strains classified by DNA-DNA hybridization. System. Appl. Microbiol. 13,338-344 (1990).
DOLZANI, L., TONIN, E., LAGATOLLA, c., PRANDIN, L., MONTIBRAGADIN, c.: Identification of Acinetobacter isolates in the A. calcoaceticus-A. baumannii complex by restriction analysis of the 16S-23S rRNA intergenic spacer sequences. J. Clin. Microbiol. 33, 1108-1113 (1995).
EHRENSTEIN, B., BERNARDS, A. T., DI]KSHOORN, L., GERNERSMIDT, P., TOWNER, K. J., BOUVET, P. J. M., DASCHNER, ED., GRUNDMANN, H.: Acinetobaeter species identification by using tRNA spacer fingerprinting. J. Chn. Microbiol. 34, 2414-2420 (1996).
GERNER-SMIDT, P., TjERNBERG, I.: Aeinetobacter in Denmark: II. Molecular studies of the Acinetobacter ealcoacetieus-Aeinetobaeter baumannii complex. APMIS 101, 826-832 (1993).
GERNER-SMIDT, P., TjERNBERG, I., URSING, J.: Reliability of phenotpyic tests for identification of Aeinetobaeter species. J. Clin. Microbiol. 29,277-282 (1991).
Identification of Aeinetobaeter genomic species by ARDRA 39
IBRAHIM, A., GERNER-SMIDT, P., LIESACK, W.: Phylogenetic relationship of the twenty-one DNA groups of the genus Acinetobaeter as revealed by 16S ribosomal DNA sequence analysis.lnt. J. System. Bacteriol. 47, 837-841 (1997).
IBRAHIM, A., GERNER-SMIDT, P., SjOSTEDT, A.: Amplification and restriction endonuclease digestion of a large fragment of genes coding for rRNA as a rapid method for discrimination of closely related pathogenic bacteria. J. Clin. Microbiol. 34, 2894-2896 (1996).
JANSSEN, P., COOPMAN, R., Huys, G., SWINGS, J., BLEEKER, M., Vos, P., ZABEAU, M., KERSTERS, K.: Evaluation of the DNA fingerprinting method AFLP as a new tool in bacterial taxonomy. Microbiology, 142, 1881-1893 (1996).
JANSSEN, P., MAQUELIN, K., COOPMAN, R., TjERNBERG, I., BouVET, P., KERSTERS, K., DIjKSHOORN, L.: Discrimination of Acinetobacter genomic species by AFLP fingerprinting. Int. J. System. Bacteriol. 47 (1997) (accepted for publication).
KAMPFER, P., TjERNBERG, I., URSING, J.: Numerical classification and identification of Aeinetobacter genomic species. J. Appl. Bacteriol. 75,259-268 (1993).
KNIGHT, G. c., McDoNNELL, S. A., SEVIOUR, R. J., SOD DELL, J. A.: Identification of Acinetobacter isolates using the Biolog identification system. Lett. Appl. Microbiol. 16, 261-264 (1993).
NISHIMURA, Y., INO, T., II7UKA, H.: Acinetobacter radioresistens sp. nov. isolated from cotton and soil. Int. J. System. Bacteri-01. 38,209-211 (1988).
NOWAK, A., KUR, J.: Genomic species typing of acinetobacters by polymerase chain reaction amplification of the recA gene. FEMS Microbiol. Lett. 130,327-332 (1995).
NOWAK, A., KUR, J.: Differentiation of seventeen genospecies of Acinetobacter by multiplex polymerase chain reaction and restriction fragment lengthe polymorphism analysis. Mol. Cell. Probes 10,405-411 (1996).
NOWAK, A., BURKIEWICZ, A., KUR, J.: PCR differentiation of seventeen genospecies of Aeinetobacter. FEMS Microbiol. Lett. 126,181-188 (1995).
RAINEY, E A., LANG, E., STACKEBRANDT, E.: The phylogenetic structure of the genus Acinetobacter. FEMS Microbiol. Lett. 124,349-354 (1994).
SODDELI, J. A., BEACHAM, A. M., SEVIOUR, R. J.: Phenotypic identification of non-clinical isolates of Acinetobacter species. J. Appl. Bacteriol. 74,210-214 (1993).
T]ERNBERG, I., URSING, J.: Clinical strains of Acinetobacter classified by DNA-DNA hybridization. APMIS 97, 595-605 (1989).
VANEECHOUTTE, M., DljKSHOORN, L., TjERNBERG, 1., ELAICHOUNl, A., DE Vos, P., CLAEYS, G., VERSCHRAEGEN, G.: Identification of Acinetobacter genomic species by amplified ribosomal DNA restriction analysis. J. Clin. Microbiol. 33, 11-15 (1995).
WIEDMANN-AL-AHMAD, M., TICHY, H.-V., SCHON, G.: Characterization of Acinetobacter type strains and isolates obtained from wastewater treatment plants by PCR fingerprinting. App!. Environ. Microbio!. 60, 4066-4071.
YAMAMOTO, S., HARAYAMA, S.: Phylogenetic analysis of Acinetobaeter strains based on the nucleotide of gyrB genes and on the amino sequences of their products. Int. J. System. Bacteri-01. 46, 506-511 (1996).
Corresponding author: Dr. L. DIjKSHOORN, Department of Medical Microbiology, Leiden University Medical Center L4-P, P.O. Box 9600, 2300 RC Leiden, The Netherlands Phone: +31 71 5263582; Fax: +31 715248148; e-mail: [email protected]
Related Documents











