Evaluation of adrenal function in serum and feces of Steller sea lions (Eumetopias jubatus): influences of molt, gender, sample storage, and age on glucocorticoid metabolism Kendall L. Mashburn and Shannon Atkinson * University of Alaska, Fairbanks and Alaska SeaLife Center, P.O. Box 1329, 301 Railway Avenue, Seward, AL 99664, USA Received 15 September 2003; revised 26 January 2004; accepted 28 January 2004 Abstract Fecal corticosterone concentrations, measured via radioimmunoassay (RIA), were validated as a method to monitor adrenal function in Steller sea lion physiology. Quantification of adrenal response to an acute stressor and relevance of data produced by developed methodologies was determined through physiological challenge with exogenous administration of adrenocorticotropic hormone (ACTH) to captive adult, reproductively intact, Steller sea lions of both sexes (n ¼ 3, 1 male, 2 female) during seasonal molt. Following ACTH administration, serial blood and fecal samples were collected and analyzed by RIA to determine adrenal response. Storage regimens and weather exposure were examined to establish external impact on fecal corticosterone concentrations. High-pressure liquid chromatography (HPLC) of both serum and feces of Steller sea lions was employed to explore potential gender-based differences extant in either sample media. ACTH challenges produced >3-fold increases in serum cortisol concen- trations which were reflected in >18-fold increases in fecal corticosterone concentrations post-injection at 3.25 and 32 h, respectively, and fecal corticosterone concentrations returned to baseline 52 h post-injection. Neither outdoor exposure to weather nor variation in duration and temperature of freezer storage impacted fecal corticosterone concentrations. HPLC of individual fecal samples produced eluate immunoreactivity profiles that differed consistently with both sex and age class. Techniques developed herein ef- fectively detected physiologically relevant corticosterone data in Steller sea lion feces, unaffected by conditions likely to be en- countered with field collection samples. Additionally, results quantify an acute response to ACTH and provide methodology for examining chronically heightened adrenal activity in Steller sea lions. Ó 2004 Elsevier Inc. All rights reserved. Keywords: Eumetopias jubatus; Steller sea lion; Cortisol; Corticosterone; Fecal glucocorticoids; ACTH; Adrenal glands; Hormones; Stress 1. Introduction Since about 1970, the Western stock of Steller sea lions have declined by an estimated 80% across the geographic range from Cape Suckling, Alaska to the Sea of Okhostk, Japan (Braham et al., 1980; Merrick et al., 1987). To date, there is no single identified cause of the decline in endangered populations of Steller sea lions. The bulk of previous research activities have addressed diet, population trends, and metabolic demands (Mer- rick et al., 1997). Most reproductive studies have been based on observation or gross morphology of animals harvested in the 1970s, 1980s, and 1990s (Ishinazaka and Endo, 1999; Pitcher and Calkins, 1981; Pitcher et al., 1998; Ruam-Suryan et al., 2002). Studies that address the physiology of Steller sea lions are few and limited to basic blood chemistries or related to fasting and lipid metabolism (Bergman and Rea, 2000; Bishop and Morado, 1995; Castellini et al., 1993; Rea et al., 1998; Zenteno-Savin et al., 1997). The endocrine system is the biochemical means of internal homeostatic regu- lation, therefore, lack of knowledge in this area hampers the investigation of factors suspected to impair Steller sea lion physiology. Disruption or alteration in ‘‘nor- mal’’ endocrine patterns can create physiological im- balances that impair the growth, reproduction, and * Corresponding author. Fax: 1-907-224-6320. E-mail address: [email protected] (S. Atkinson). 0016-6480/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2004.01.016 www.elsevier.com/locate/ygcen General and Comparative Endocrinology 136 (2004) 371–381 GENERAL AND COMPARATIVE ENDOCRINOLOGY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GENERAL AND COMPARATIVE
ENDOCRINOLOGY
www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 136 (2004) 371–381
Evaluation of adrenal function in serum and feces of Steller sealions (Eumetopias jubatus): influences of molt, gender, sample
storage, and age on glucocorticoid metabolism
Kendall L. Mashburn and Shannon Atkinson*
University of Alaska, Fairbanks and Alaska SeaLife Center, P.O. Box 1329, 301 Railway Avenue, Seward, AL 99664, USA
Received 15 September 2003; revised 26 January 2004; accepted 28 January 2004
Abstract
Fecal corticosterone concentrations, measured via radioimmunoassay (RIA), were validated as a method to monitor adrenal
function in Steller sea lion physiology. Quantification of adrenal response to an acute stressor and relevance of data produced by
developed methodologies was determined through physiological challenge with exogenous administration of adrenocorticotropic
hormone (ACTH) to captive adult, reproductively intact, Steller sea lions of both sexes (n ¼ 3, 1 male, 2 female) during seasonal
molt. Following ACTH administration, serial blood and fecal samples were collected and analyzed by RIA to determine adrenal
response. Storage regimens and weather exposure were examined to establish external impact on fecal corticosterone concentrations.
High-pressure liquid chromatography (HPLC) of both serum and feces of Steller sea lions was employed to explore potential
gender-based differences extant in either sample media. ACTH challenges produced >3-fold increases in serum cortisol concen-
trations which were reflected in >18-fold increases in fecal corticosterone concentrations post-injection at 3.25 and 32 h, respectively,
and fecal corticosterone concentrations returned to baseline 52 h post-injection. Neither outdoor exposure to weather nor variation
in duration and temperature of freezer storage impacted fecal corticosterone concentrations. HPLC of individual fecal samples
produced eluate immunoreactivity profiles that differed consistently with both sex and age class. Techniques developed herein ef-
fectively detected physiologically relevant corticosterone data in Steller sea lion feces, unaffected by conditions likely to be en-
countered with field collection samples. Additionally, results quantify an acute response to ACTH and provide methodology for
examining chronically heightened adrenal activity in Steller sea lions.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Eumetopias jubatus; Steller sea lion; Cortisol; Corticosterone; Fecal glucocorticoids; ACTH; Adrenal glands; Hormones; Stress
1. Introduction
Since about 1970, the Western stock of Steller sea
lions have declined by an estimated 80% across the
geographic range from Cape Suckling, Alaska to the Seaof Okhostk, Japan (Braham et al., 1980; Merrick et al.,
1987). To date, there is no single identified cause of the
decline in endangered populations of Steller sea lions.
The bulk of previous research activities have addressed
diet, population trends, and metabolic demands (Mer-
rick et al., 1997). Most reproductive studies have been
* Corresponding author. Fax: 1-907-224-6320.
E-mail address: [email protected] (S.Atkinson).
0016-6480/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2004.01.016
based on observation or gross morphology of animals
harvested in the 1970s, 1980s, and 1990s (Ishinazaka
and Endo, 1999; Pitcher and Calkins, 1981; Pitcher
et al., 1998; Ruam-Suryan et al., 2002). Studies that
address the physiology of Steller sea lions are few andlimited to basic blood chemistries or related to fasting
and lipid metabolism (Bergman and Rea, 2000; Bishop
and Morado, 1995; Castellini et al., 1993; Rea et al.,
1998; Zenteno-Savin et al., 1997). The endocrine system
is the biochemical means of internal homeostatic regu-
lation, therefore, lack of knowledge in this area hampers
the investigation of factors suspected to impair Steller
sea lion physiology. Disruption or alteration in ‘‘nor-mal’’ endocrine patterns can create physiological im-
balances that impair the growth, reproduction, and

372 K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381
overall health of the sea lions. Detection of these im-balances can more clearly define goals for mitigation or
intervention.
Endocrine signals that indicate deviations from ho-
meostatic balance, or stress, include increased adrenal
output of corticoid hormones (Carrasco and Van de Kar,
2003; Mostl and Palme, 2002; Oki and Atkinson, in
press). Circulatory, salivary, or urinary concentrations
of glucocorticoids, and primarily cortisol or corticoste-rone, are often measured as indicators of stress. The
measurement of elevated glucocorticoids in the blood or
feces indicates that an animal has been exposed to a
stressor, but does not necessarily mean that an animal is
currently stressed. However, chronic elevation of these
hormones above an established baseline is often used as a
diagnostic tool to indicate an individual or population
that is compromised or may be at risk. Sampling forcorticosteroid diagnostics typically occurs in a longitu-
dinal fashion to account for normal circadian rhythms of
corticosteroids and the potential temporal response to
stress caused by collection procedures (Barriga et al.,
2001; Feldman et al., 1978; Walker et al., 2001). Blood,
saliva, or urine collection is not often practical in free-
ranging, non-domestic species due to logistical difficul-
ties and/or stress responses associated with capture orrestraint of animals. Sample collection from marine
mammals is often compounded by the inability to sample
any but the very young, physiologically compromised
(i.e., stranded or entangled), or well trained captive an-
imals. Non-invasive measurement of corticosterone in
feces from free-ranging animals has proven to be a
valuable tool for identification of stress responses in a
variety of species. This method also provides the capa-bility for longitudinal data collection such that stress
responses can be further defined as acute or chronic
(Cavagelli, 1999; Creel et al., 1997; Goymann et al., 1999;
Harper and Austad, 2000; Monfort et al., 1998; Terio
et al., 1999; Turner et al., 2002; Wasser et al., 2000). Once
validated for use in Steller sea lions, measurement of
fecal corticosterone will provide a viable alternative to
blood collection for the determination of adrenal outputand physiological stress in free-ranging animals.
The primary objective of this study was to determine
physiologic relevance of fecal corticosterone measure-
ments as indicators of stress or well-being in Steller sea
lions through detection of adrenal response to exoge-
nously administered adrenocorticotropic hormone
(ACTH). Second, as anesthesia was required for a pe-
riod of 3 h post-ACTH administration, it was necessaryto ensure that restraint and anesthesia did not contrib-
ute to measured adrenal response in either serum or
feces of challenged animals. The geographic distribution
of Steller sea lion rookeries requires that sample col-
lection occur during extended cruises with multiple sites
visited. As a consequence, both storage duration and
temperature vary. Thus, an additional priority for future
field applications was identification of environmental orstorage factors associated with field sample collection as
potential sources for data error as has been seen in other
species (Kahn et al., 2002; Washburn and Millspaugh,
2002). Finally, we endeavored to determine if detectable
gender differences existed in fecal corticosteroid ana-
lytes. Key Steller sea lion sample collection sites (rook-
eries) are typically populated with breeding, territorial
males, breeding females, and breeding females withpups. There is a paucity of data describing reproductive
endocrinology in free-ranging Steller sea lions in the
absence of physical manipulation of animals for sample
collection. Consequently, little information is available
representative of adrenal activity in typical territorial
behavior, reproduction or lactation in this species. In the
absence of these data, sex determination of collected
scats eliminates gender-bias in endocrine analysis of fieldsamples at the population level.
2. Methods
2.1. Animals
Samples for in situ sample exposure, sample storage,anesthesia, and ACTH challenge trials were obtained
from three adult (1 male, 539 kg; 2 females, 200 kg each)
reproductively intact, captive Steller sea lions housed
under ambient conditions at the Alaska SeaLife Center
(ASLC, Seward, Alaska; N 60�120 latitude, W 149�420longitude). Free-range fecal samples for HPLC deter-
mination of sex and age class differences were collected
from Natoa Island (N 59�360 latitude, W 149�360 lon-gitude) and Chiswell Island (N 59�360 latitude, W
149�340 longitude). Age and sex data were obtained
through the use of remote cameras located on each is-
land. Samples were also collected from one female
Steller sea lion pup in the rehabilitation program at the
Alaska SeaLife Center.
2.2. Blood samples
All sera used for this study, whether derived from
whole blood collected via caudal plexus veinipuncture
(Experiment 1, Effects of isoflurane anesthesia) or
through hindflipper vein catheterization (Experiment 5,
ACTH challenge), were collected into serum separator
Vacutainers (Becton–Dickinson, Franklin Lakes, NJ).
All blood samples were refrigerated prior to serumharvest into 1ml aliquots and stored frozen at )80 �Cuntil radioimmunoassay (RIA).
2.3. Fecal sample extraction and preparation
Feces from individual Steller sea lions were fully
mixed, aliquoted (�5 g) and loaded onto a rotary

K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381 373
evaporator (Speed-Vac Plus, SC110A; Savant Instru-ments, Holbrook, NY) and dried without heat. Dried
fecal samples were crushed into powder, after which
0.025–0.03 g was weighed and extracted as previously
described by Monfort et al. (1998). Methanol (MeOH)
extractant (100 ll) was aliquoted into polypropylene
tubes, dried under forced air, reconstituted in 400 llbuffer for a final 1:4 dilution. Sample dilutions were
stored frozen at )20 �C until RIA.
2.4. Cortisol RIA
A solid-phase RIA kit (Diagnostic Products, Los
Angeles, CA) for cortisol was validated for use with
unextracted, undiluted Steller sea lion serum. Radio-
activity of bound portion was determined using a
gamma counter (Gamma C12, Diagnostic Products,Los Angeles, CA). All samples from a single experi-
ment were analyzed in duplicate within the same assay
to reduce variation. The RIAs were performed per
manufacturer instructions with the exception that all
volumes were halved and an additional standard was
added to the curve (i.e., one half the lowest standard)
to increase sensitivity. Manufacturer cross-reactivity
data are as follows: prednisolone (76.0%), 11-deoxy-cortisol (11.4%), prednisone (2.3%), cortisone (0.98%),
corticosterone (0.94%), tetrahydrocortisol (0.34%), 11-
deoxycorticosterone (0.26%), aldosterone (0.03%),
progesterone and pregnenolone (0.02%), flumethasone
(0.017%), and 0.01% or below for all other steroids
tested.
Serial dilutions of serum pools from both male and
female Steller sea lions (neat to 1:16) yielded displace-ment parallel to that of the cortisol standard curve.
Recovery of added cortisol (5–500 ng/ml) was 99.35%
(SD¼ 8.37; CV¼ 8.43%) for males (y ¼ 0:474þ 0:85x,r2 ¼ 0:99) and 92.05% (SD¼ 10.13, CV¼ 11.00%) for
females (y ¼ �0:103þ 0:98x, r2 ¼ 1:00). Interassay co-
efficients of variation for two separate assay controls
were 12.92 and 11.04% (n ¼ 4 for all samples assayed).
Intrassay coefficients of variation were <5% and assaysensitivity was 5.8 ng/ml.
2.5. Corticosterone RIA
A double antibody RIA kit (ICN Biomedicals, Ata-
scadero, CA) for corticosterone was validated for use
with extracted feces. All fecal samples were initially
analyzed at a dilution of 1:4 in diluent buffer providedby the manufacturer. Values from the RIA were cor-
rected for dilution, extraction efficiency, weight of fecal
material extracted, and expressed as ng/g dry weight.
The RIAs were performed according to manufacturer�sinstructions with the exception that all volumes were
halved and an additional standard was added to the
curve (i.e., one-half the lowest standard) to increase
sensitivity. Manufacturer cross-reactivity with othersteroids were: desoxycorticosterone (0.34%), testoster-
one (0.10%), cortisol (0.05%), aldosterone (0.03%),
progesterone (0.02%), and less than 0.01% for all other
steroids tested.
Serial dilutions of fecal extract for both male and
female Steller sea lions (1:1–1:28) yielded displacement
parallel to the standard curve. Recovery of corticoste-
rone added to fecal sample pools (range 12.5–500 ng/ml)was 90.3% (SD¼ 6.1; CV¼ 6.7%) for males (y ¼ 1:95þ0:88x; R2 ¼ 0:99) and 93.3% (SD¼ 15.2; CV¼ 16.4%)
for females (y ¼ 1:85þ 0:80x; R2 ¼ 0:99). Interassay
coefficients of variation for two separate assay controls
were 5.12 and 10.95% (n ¼ 9 for all samples). Intrassay
coefficients of variation were <5% and assay sensitivity
was 13.7 ng/ml.
2.6. High-pressure liquid chromatography
HPLC (Varian ProStar 210/215, Varian, Walnut
Creek, CA) was used to determine immunoreactive
corticosteroid components in Steller sea lion serum and
feces in the validated RIAs. Randomly selected fecal
extracts and unextracted sera were used to create 3ml
pools of each sex and sample type. The 3ml fecal extractpools were dried under forced air, rinsed with MeOH,
and re-dried twice to ensure that the majority of the pool
was washed to the bottom of the tube. Dried fecal pools
were reconstituted in 500 ll phosphate-buffered saline
(PBS) buffer (pH¼ 5.0). Similarly, 500 ll PBS was added
to serum pools and all samples were vortexed for 2min.
All pools were analyzed and fractions collected via
HPLC using a previously described method (Monfortet al., 1998). HPLC for individual animals was identical
to that of the pooled samples, with the exception that
larger portions (0.3–0.4 g) of dried feces from individuals
were extracted to ensure a high concentration of im-
munoreactive components. Resultant immunoreactivity
and radioactivity profiles were then obtained from
analysis of collected eluates and expressed per 1ml
fraction collected.
2.7. Experiment 1. Effects of isoflurane anesthesia
Animals were required to undergo anesthesia the first
3–4 h of an ACTH trial (Experiment 5), therefore, it was
necessary to determine the effects of isoflurane (Isoflu-
rane, USP; Halocarbon Industries, River Edge, NJ)
anesthesia (delivered via Vet Tech Model LAVC-2000,JD Medical Distributing Company, Phoenix, AZ). Ad-
ditional sera collected from both male (n ¼ 1) and fe-
male (n ¼ 2) captive Steller sea lions during routine
veterinary care procedures was analyzed. Samples cho-
sen for the experiment were paired when possible, i.e.,
voluntary or restraint bleeds followed by anesthesia and
additional sampling blood sampling under anesthesia

374 K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381
usually occurred at varying times post-anesthesia so aneffort was made to use those samples obtained between
30 and 40min post-anesthesia. Voluntary samples or
restraint samples were collected between 20 and 30min
after positioning animals in trained ‘‘stationing’’ pos-
tures. Resultant serum concentrations were divided into
three categories, voluntary (n ¼ 8), restraint (n ¼ 7), or
anesthetized (n ¼ 19) and compared statistically be-
tween sexes and treatments.
2.8. Experiment 2. Change in fecal corticosterone con-
centration due to freezing regimen
To assess the impact of storage variation, freshly
voided scat from a male and two pooled fresh scats from
two female Steller sea lions were fully homogenized and
subsampled. Three subsamples, for each sex were im-mediately dried (within 30min post-defecation¼Time
0) on the rotary evaporator and placed in a )20 �Cfreezer until processing and RIA. Thirty subsamples for
each sex (5–6 g wet weight) were placed into tubes, split
into two groups, and stored at either )20 or )80 �C(n ¼ 15 each temperature). Triplicate subsamples for
each sex were then removed weekly from Time 0 for 5
weeks from each freezer, dried without heat, crushed,and stored frozen at )20 �C until RIA. At the end of 5
weeks, all samples (total samples ¼ 66) were processed
then assayed in duplicate to eliminate handling or in-
terassay variation. Resultant fecal corticosterone con-
centrations and freezing regimen were statistically
compared to Time 0 for each sex.
2.9. Experiment 3. Effects of in situ exposure on fecal
corticosterone concentrations
To assess the effects of prolonged weather exposure
on fecal corticosterone, a freshly voided scat from a
male and two pooled fresh scats from two female Steller
sea lions were fully homogenized and subsampled.
(n ¼ 3, Time 0). The remainder was left outdoors, un-
covered, from between June 15 to July 05, 2001, whichcorresponds to typical field collection dates. Exposed
scat was subsampled in triplicate at 1, 2, 3, 4, 5, 6, 8, 10,
12, 18, 24, 36, 48, 60, 72, 96, and 108 h (at which time
scat was completely desiccated) post-defecation. All
subsamples were frozen at )20 �C until concurrent
analysis. Resultant fecal corticosterone concentrations
for each sex and freezing regimen were then statistically
compared to Time 0 for each sex.
2.10. Experiment 4. Differences in captive vs. free-
ranging, sex, and age class fecal immunoreactivity via
RIA of HPLC fractions
To assess similarity of captive and free-range ani-
mals and differences between sex and age class with
regards to fecal corticosterone concentrations, immu-noreactive components of free-range Steller sea lion
scat extracts were separated using HPLC and identified
for comparison with captives via the corticosterone
RIA. Feces of free-range, known sex and age animals
from Chiswell and Natoa Islands, verified through use
of remotely controlled video cameras located on each
island, were collected, processed, then analyzed via
HPLC. Resultant fractions for individuals were ana-lyzed via RIA and immunoreactive corticosterone peak
profiles of free-ranging animals were then compared to
those of captive animals housed at the Alaska SeaLife
Center.
2.11. Experiment 5. ACTH challenge
Adult, reproductively intact Steller sea lions (1 male,2 female) undergoing seasonal molt were treated with a
single injection (2 IU/kg, I.M.) of ACTH in a long-act-
ing gel preparation (synthesized by Hadfield�s Phar-
macy, Edmonds WA). Feces voided from each female,
obtained immediately prior to injection, was considered
Time 0. Time 0 fecal material from the male was col-
lected via the housing for rectal temperature monitor. A
blood sample was drawn from the caudal plexus im-mediately prior to ACTH injection (Time 0) which al-
lowed animals to serve as their own controls. Female sea
lions, due to their smaller size and relative ease of
management were held in an open squeeze cage for 1 h
post-injection followed by 3 h of isoflurane anesthesia.
Blood samples (20ml) were drawn every 15min via an
indwelling catheter (18 gauge, 3 in.) placed in a hind-
flipper vein. The catheter was flushed with approxi-mately 3ml sterile heparinized saline (concentration:
25 IU/ml) after each blood draw. At each new blood
draw, the initial saline portion was discarded. The male
was anesthetized immediately post-injection for 3 h,
during which blood samples were collected as described
above.
After 3 h, animals were removed from isoflourane.
Once fully alert, females were transferred to a largerstainless steel cage (45 in. W� 50 in. H� 10L) with a
slightly sloped floor fitted with gutters on one side to
prevent admixing of urine and feces. The cage was
placed in a sheltered outdoor area and fitted with an
observation camera. Females were then each monitored
constantly for 91.5 h post-ACTH administration for a
total of 96 h. Behaviors were recorded throughout this
time and each defecation was homogenized and sub-sampled. Due to the male�s large size and intractability
in close confinement, a video-monitored dry-run was
utilized during the day until defecation; after defecation
he was in a monitored outdoor pool enclosure. Defe-
cations in the outdoor pool enclosure accumulated until
approximately 20:00 h, at which time they were col-
lected. The pool was subsequently drained each morning
K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381 375
with defecations produced overnight mixed together andsubsampled. All fecal samples were stored at )20 �Cuntil processing and assay.
2.12. Statistical analysis
Data are presented as means� SEM unless otherwise
noted. Comparisons of pre- and post-treatment data
were analyzed using repeated measures analysis withSigmaPlot 2002, v.8.0 (SPSS, Chicago, IL).
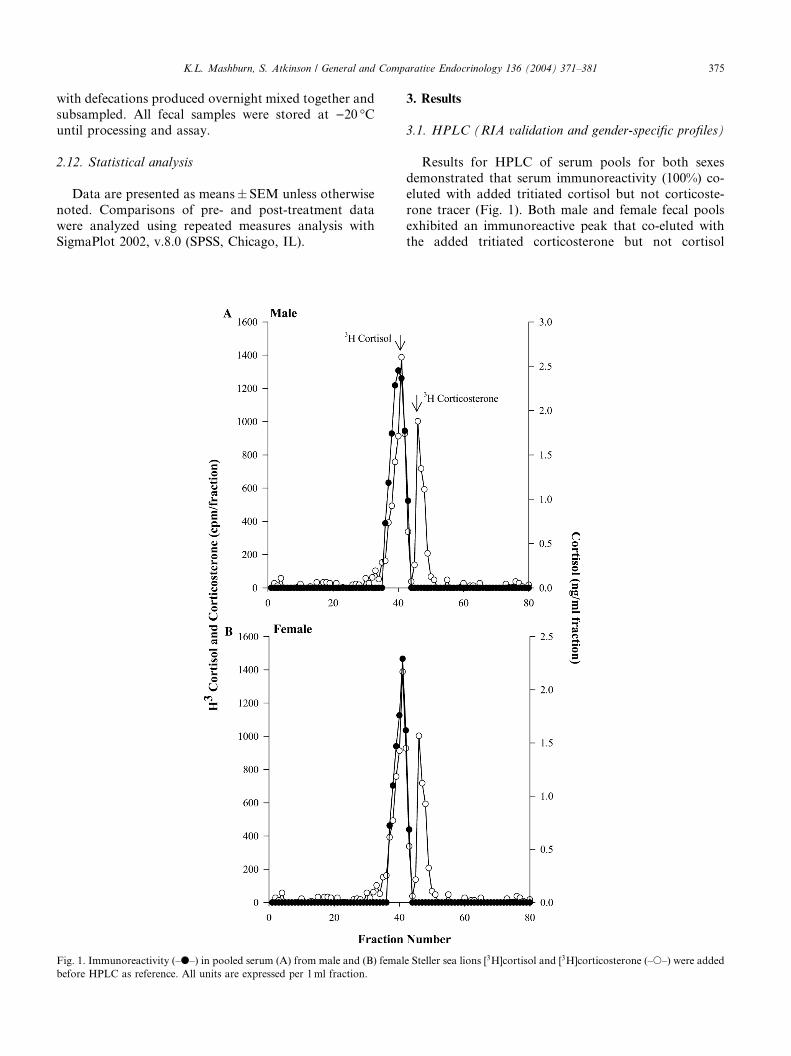
Fig. 1. Immunoreactivity (–d–) in pooled serum (A) from male and (B) femal
before HPLC as reference. All units are expressed per 1ml fraction.
3. Results
3.1. HPLC (RIA validation and gender-specific profiles)
Results for HPLC of serum pools for both sexes
demonstrated that serum immunoreactivity (100%) co-
eluted with added tritiated cortisol but not corticoste-
rone tracer (Fig. 1). Both male and female fecal pools
exhibited an immunoreactive peak that co-eluted withthe added tritiated corticosterone but not cortisol
e Steller sea lions [3H]cortisol and [3H]corticosterone (–s–) were added
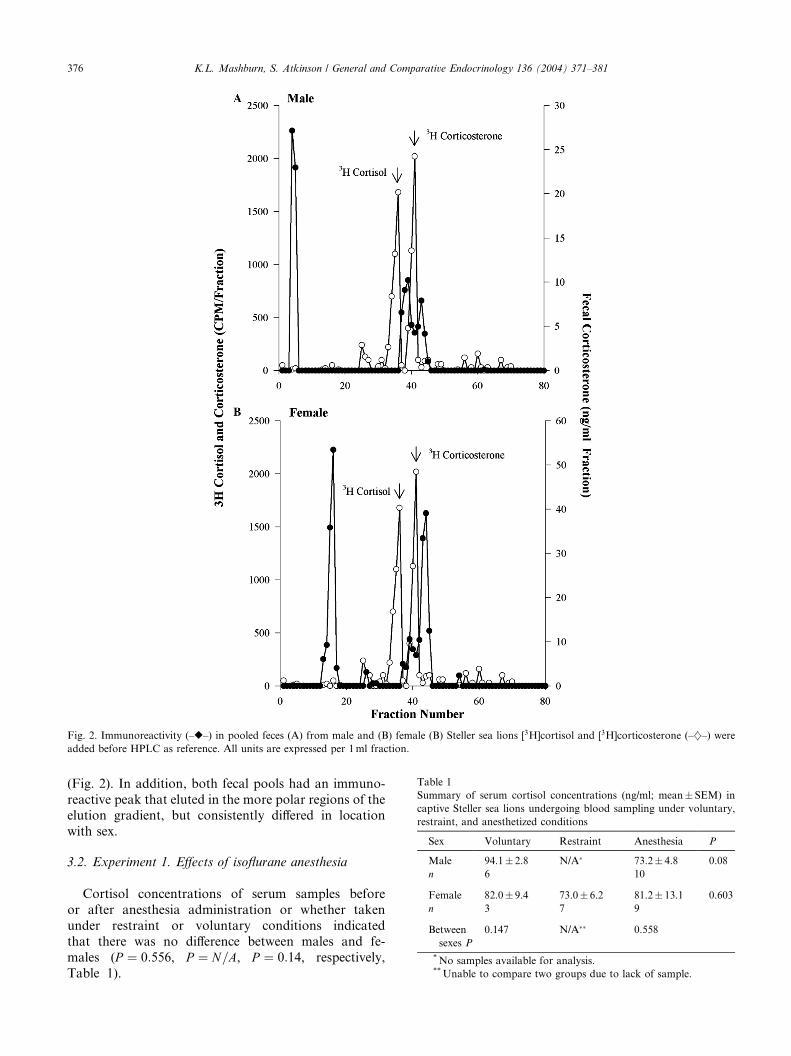
Fig. 2. Immunoreactivity (–r–) in pooled feces (A) from male and (B) female (B) Steller sea lions [3H]cortisol and [3H]corticosterone (–}–) were
added before HPLC as reference. All units are expressed per 1ml fraction.
Table 1
Summary of serum cortisol concentrations (ng/ml; mean� SEM) in
captive Steller sea lions undergoing blood sampling under voluntary,
restraint, and anesthetized conditions
Sex Voluntary Restraint Anesthesia P
Male 94.1� 2.8 N/A� 73.2� 4.8 0.08
n 6 10
Female 82.0� 9.4 73.0� 6.2 81.2� 13.1 0.603
n 3 7 9
Between
sexes P
0.147 N/A�� 0.558
*No samples available for analysis.**Unable to compare two groups due to lack of sample.
376 K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381
(Fig. 2). In addition, both fecal pools had an immuno-
reactive peak that eluted in the more polar regions of theelution gradient, but consistently differed in location
with sex.
3.2. Experiment 1. Effects of isoflurane anesthesia
Cortisol concentrations of serum samples before
or after anesthesia administration or whether taken
under restraint or voluntary conditions indicatedthat there was no difference between males and fe-
males (P ¼ 0:556, P ¼ N=A, P ¼ 0:14, respectively,
Table 1).

K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381 377
3.3. Experiment 2. Change in fecal corticosterone con-
centration due to freezing regimen
Fecal corticosterone concentrations in triplicate
subsamples of the same scat did not differ between Time
0 and those frozen at )20 or )80 �C for up to five weeks
for either male or female samples. Mean concentration
values for male scat were 60.17� 16.81 ng/g dry weight
at Time 0, 46.28� 4.99 at )20 �C (P ¼ 0:522), and50.85� 5.17 at )80 �C (P ¼ 0:687) at the end of five
weeks. Mean concentration values for female scat were
97.06� 6.22 ng/g dry weight at Time 0, 82.37� 6.89 at
)20 �C (P ¼ 0:140), and 92.47� 9.41 at )80 �C(P ¼ 0:517) at the end of five weeks.
3.4. Experiment 3. Effects of in situ exposure on fecal
corticosterone concentrations
Triplicate subsamples of both male and female scat
left exposed to ambient conditions for up to five days
exhibited fecal corticosterone concentrations no differ-
ent than those measured in the same scat at Time 0.
Mean concentration values for male scat were
50.13� 5.72 ng/g dry weight at Time 0 and 45.11� 8.32
(P ¼ 0:311) at the end of five days. Mean concentrationvalues for female scat were 212.78� 17.36 ng/g dry
weight at Time 0 and 196.93� 13.08 (P ¼ 0:795) at theend of five days.
3.5. Experiment 4. Differences in captive vs. free-ranging,
sex, and age/class fecal corticosterone immunoreactivity
via corticosteroid RIA of HPLC fractions
As is indicated by the HPLC profiles, there was no
difference between relative positions of immunoreactive
corticosterone peaks in free-ranging and captive animals
(Fig. 3). Although adult animals exhibited differences in
immunoreactivity in the more polar regions of the elu-
tion gradient, pups of different genders did not. In
contrast, juvenile males presented HPLC profiles with
both immunoreactive peaks in the more polar regions ofthe elution gradient that had previously occurred in
association with a specific gender in adults tested.
3.6. Experiment 5. ACTH challenge
Serum samples were averaged for each collection time
for all animals (Fig. 4). To account for variation in
defecation times, fecal samples were grouped, and av-eraged according to 4-h blocks. Serum cortisol concen-
trations increased 2-fold within 60min with a >3-fold
peak at 195min (65.3–220.0 ng/ml) and were reflected in
peak fecal corticosterone concentrations 18-fold above
baseline (88.5–1605.3 ng/g dry weight) 32 h post-ACTH
injection (beginning at 23.5 h with a return to baseline at
48.5 h). There were no gender-specific differences in
timing, duration or fecal corticosterone concentrationsassociated with the adrenal response to ACTH admin-
istration.
4. Discussion
In this study, the ACTH challenge was used to sim-
ulate an acute stress on Steller sea lions and evaluate theanimal�s adrenal response. The results demonstrate that,
as an acute stressor, ACTH initiates activity at the level
of the adrenal glands by 30min post-ACTH injection.
The adrenal response was clearly reflected in a fecal
corticosterone peak 32 h following the stimulus and was
cleared within 52 h, indicating that the feces integrated
and cleared the signal. Quantification of the response to
a known acute stressor enables a definition for the du-ration and maximal amplitude of adrenal activity in the
feces and provides a benchmark to which responses to
other stimuli can be compared. Such a benchmark al-
lows for the comparison of fecal corticosterone con-
centrations to be used to evaluate physiological function
of Steller sea lions in response to a variety of potential
stressors.
Validation of commercial RIA methodology, whenmodified as described herein, detected increased corti-
costeroids released in response to exogenous adminis-
tration of ACTH. Analogous HPLC elution profiles in
free-ranging and captive fecal samples indicate that the
extraction/RIA is an effective tool for the assessment of
adrenal activity in both wild and captive Steller sea li-
ons. The temporal relationship of serum cortisol to fecal
corticosteroids established through the ACTH challengedata enables appropriate protocols such that the timing
of the collection does not interfere with fecal cortico-
sterone concentrations. Further, knowledge of the
‘‘time-lag’’ relationship of serum to fecal corticosterone
allows extrapolation from collection dates to determine
relative contribution of potential stressors (e.g., weather
or entanglement) to corticosterone concentrations in
fecal samples.Outdoor exposure and freezer storage variability did
not significantly alter concentrations of corticosteroids
measured in the RIA utilized. From a field perspective,
this increases the number of samples that can be col-
lected from rookery or haul out sites by not limiting
collections for corticosteroid analysis to only freshly
voided scats. There is also increased confidence in the
comparison of scats from different research cruises withvariable storage temperature and duration.
The ability to determine the sex of a collected scat can
be of significant importance. The current means of scat
collection for adult Steller sea lions is typically on-site
collection without the presence of animals. Research
teams are rarely able to identify scats from individuals
collected from rookery or haul-out sites. Historically,
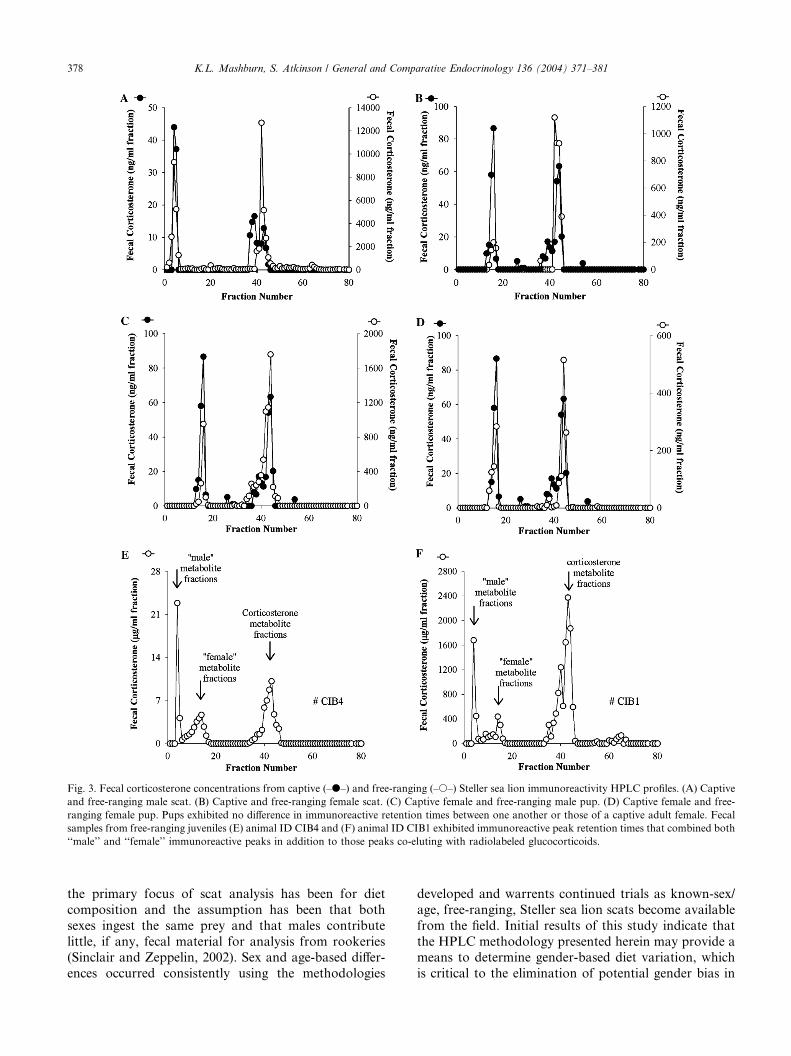
Fig. 3. Fecal corticosterone concentrations from captive (–d–) and free-ranging (–s–) Steller sea lion immunoreactivity HPLC profiles. (A) Captive
and free-ranging male scat. (B) Captive and free-ranging female scat. (C) Captive female and free-ranging male pup. (D) Captive female and free-
ranging female pup. Pups exhibited no difference in immunoreactive retention times between one another or those of a captive adult female. Fecal
samples from free-ranging juveniles (E) animal ID CIB4 and (F) animal ID CIB1 exhibited immunoreactive peak retention times that combined both
‘‘male’’ and ‘‘female’’ immunoreactive peaks in addition to those peaks co-eluting with radiolabeled glucocorticoids.
378 K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381
the primary focus of scat analysis has been for diet
composition and the assumption has been that both
sexes ingest the same prey and that males contribute
little, if any, fecal material for analysis from rookeries(Sinclair and Zeppelin, 2002). Sex and age-based differ-
ences occurred consistently using the methodologies
developed and warrents continued trials as known-sex/
age, free-ranging, Steller sea lion scats become available
from the field. Initial results of this study indicate that
the HPLC methodology presented herein may provide ameans to determine gender-based diet variation, which
is critical to the elimination of potential gender bias in
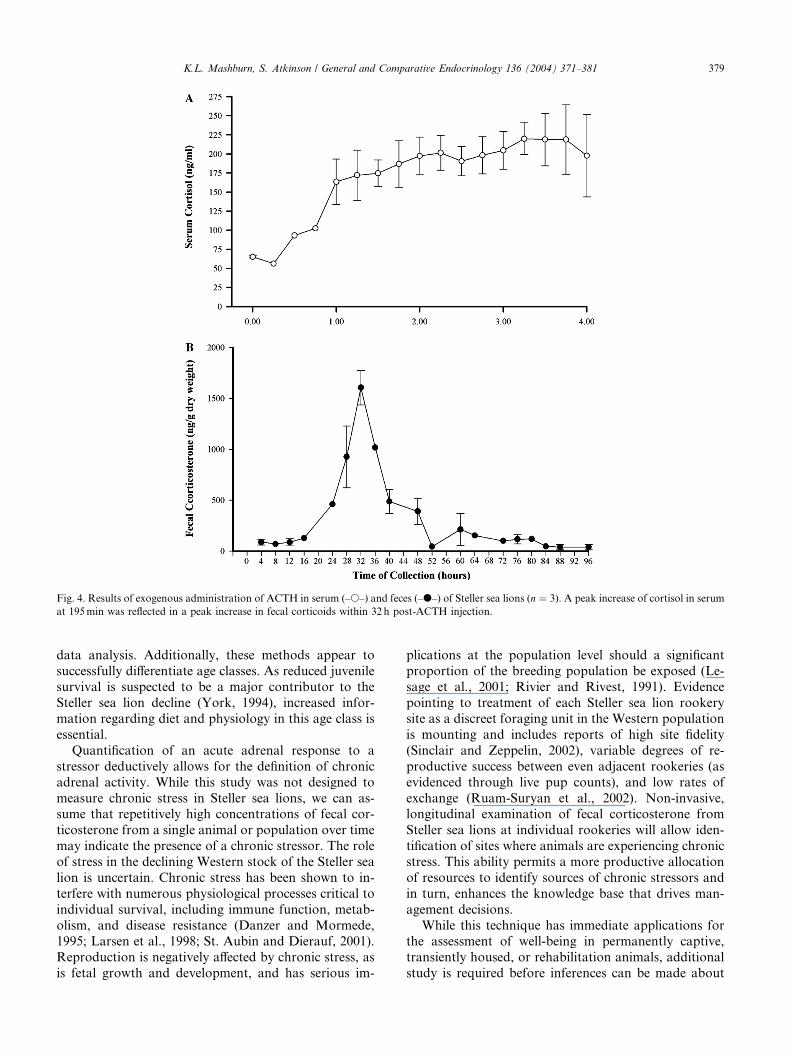
Fig. 4. Results of exogenous administration of ACTH in serum (–s–) and feces (–d–) of Steller sea lions (n ¼ 3). A peak increase of cortisol in serum
at 195min was reflected in a peak increase in fecal corticoids within 32 h post-ACTH injection.
K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381 379
data analysis. Additionally, these methods appear to
successfully differentiate age classes. As reduced juvenilesurvival is suspected to be a major contributor to the
Steller sea lion decline (York, 1994), increased infor-
mation regarding diet and physiology in this age class is
essential.
Quantification of an acute adrenal response to a
stressor deductively allows for the definition of chronic
adrenal activity. While this study was not designed to
measure chronic stress in Steller sea lions, we can as-sume that repetitively high concentrations of fecal cor-
ticosterone from a single animal or population over time
may indicate the presence of a chronic stressor. The role
of stress in the declining Western stock of the Steller sea
lion is uncertain. Chronic stress has been shown to in-
terfere with numerous physiological processes critical to
individual survival, including immune function, metab-
olism, and disease resistance (Danzer and Mormede,1995; Larsen et al., 1998; St. Aubin and Dierauf, 2001).
Reproduction is negatively affected by chronic stress, as
is fetal growth and development, and has serious im-
plications at the population level should a significant
proportion of the breeding population be exposed (Le-sage et al., 2001; Rivier and Rivest, 1991). Evidence
pointing to treatment of each Steller sea lion rookery
site as a discreet foraging unit in the Western population
is mounting and includes reports of high site fidelity
(Sinclair and Zeppelin, 2002), variable degrees of re-
productive success between even adjacent rookeries (as
evidenced through live pup counts), and low rates of
exchange (Ruam-Suryan et al., 2002). Non-invasive,longitudinal examination of fecal corticosterone from
Steller sea lions at individual rookeries will allow iden-
tification of sites where animals are experiencing chronic
stress. This ability permits a more productive allocation
of resources to identify sources of chronic stressors and
in turn, enhances the knowledge base that drives man-
agement decisions.
While this technique has immediate applications forthe assessment of well-being in permanently captive,
transiently housed, or rehabilitation animals, additional
study is required before inferences can be made about

380 K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381
stress in Steller sea lions from unknown samples col-lected in the field. Future investigations to confirm the
presence of gender-specific corticosterone metabolites
and quantify the relative contribution of those identified
to total corticosterone concentrations are needed. Ad-
ditionally, the relationship between normal fecal corti-
costerone concentrations and the onset of parturition
and lactation in females, as well as territorial behavior
and breeding in males must be determined. Fecal sam-ples are typically collected when reproduction and ma-
ternal care are at their height and increased adrenal
activity that influence these events has been well docu-
mented in several mammalian species (Challis et al.,
2000; Whittle et al., 2001). Furthermore, because of the
highly seasonal nature of many physiological processes
in pinnipeds, the relationship of adrenal function and
photoperiod in Steller sea lions needs to be explored.Use of the reported methodology is particularly well-
suited to non-invasive acquisition of longitudinal data
necessary to investigate of the role of adrenal hormones
in Steller sea lion physiology.
Acknowledgments
All procedures were approved by Alaska SeaLife
Center and University of Alaska, Fairbanks Institu-tional Animal Care and Use Committees and were
conducted under NOAA/NMFS permits #782-1532 and
881-1668. We would like to acknowledge Carol Ste-
phens, Danielle O�Neil, Dennis Christen, Don Calkins,
Elizabeth Moundalexis, Howard Ferren, Janelle Schuh,
Jen Dailer, Jo-Ann Mellish, John Maniscalco, Lisa
Hartman, Lisa Petrauskas, Lynn Turcotte, Mary Bozza,
Matt Myers, Millie Gray, Pam Parker, Pam Tuomi,Peter Nilsson, Russ Andrews, and Tim Lebling of the
Alaska SeaLife Center, and Daniel Zatz of SeeMore
Wildlife (Homer, Alaska). Support for this project was
provided by National Oceanic and Atmospheric Ad-
ministration appropriations through the Alaska SeaLife
Center.
References
Barriga, C., Martin, M.I., Tabla, R., Ortega, E., Rodriguez, A.B.,
2001. Circadian rhythm of melatonin, corticosterone and phago-
cytosis: effect of stress. J. Pineal Res. 30, 180–187.
Bergman, M., Rea, L.D., 2000. The effects of food limitation on serum
lipid concentration and content in Steller sea lions. In: Proc. 3rd
Comp. Nutr. Soc. Symp. Pacific Grove, CA, USA, pp. 13–16.
Bishop, D.H., Morado, J.F., 1995. Results on blood cell morphology
and differential blood cell counts from seventeen Steller sea lion
(Eumetopias jubatus) pups. Dis. Aqu. Org. 23, 1–6.
Braham, H.W., Everitt, R.D., Rugh, D.J., 1980. Northern sea lion
decline in the eastern Aleutian Islands. J. Wildlife Manage. 44, 25–
33.
Carrasco, G.A., Van de Kar, L.D., 2003. The neuroendocrine
pharmacology of stress. Eur. J. Pharmacol. 463 (1–3), 235–
272.
Castellini, M.A., Davis, R.W., Loughlin, T.R., Williams, T.M., 1993.
Blood chemistries and body condition of Steller sea lion pups at
Marmot Island, Alaska. Mar. Mammal Sci. 9 (2), 202–204.
Cavagelli, S.A., 1999. Behavioral patterns associated with faecal
cortisol levels in free-ranging female ring-tailed lemurs, (Lemur
catta). Anim. Behav. 57 (4), 935–944.
Challis, J.R.G., Matthews, S.G., Gibb, W., Lye, S.J., 2000. Endocrine
and paracrine regulation of birth at term and preterm. Endo. Rev.
21 (5), 514–550.
Creel, S., Creel, N.M., Monfort, S.L., 1997. Radiocollaring and
stress hormones in African wild dogs. Conserv. Biol. 11 (2), 544–
548.
Danzer, R., Mormede, P., 1995. Psychonueroimmunology of stress. In:
Leonard, B., Miller, K. (Eds.), Stress, the Immune System and
Psychiatry. Wiley, West Sussex, UK, pp. 47–53.
Feldman, E.C., Tyrell, J.B., Bohannon, N.V., 1978. The synthetic
ACTH stimulation test and measurement of endogenous plasma
ACTH levels: useful diagnostic indicators for adrenal disease in
dogs. J. Am. Anim. Hosp. Assoc. 14, 524–531.
Goymann, W., Mostl, E., Van�t Hof, T., East, M.L., Hofer, H., 1999.
Noninvasive fecal monitoring of glucocorticoids in spotted
hyenas (Crocuta crocuta). Gen. Comp. Endocrinol. 114 (3), 340–
348.
Harper, J.M., Austad, S.N., 2000. Fecal glucocorticoids: a non-
invasive method of measuring adrenal activity in wild and captive
rodents. Physiol. Biochem. Zool. 73 (1), 12–22.
Ishinazaka, T., Endo, T., 1999. The reproductive status of Steller sea
lions in the Nemuro Strait, Hokkaido, Japan. Biosphere Conserv. 2
(1), 11–19.
Kahn, M.Z., Altman, J., Isani, S.S., Yu, J., 2002. A matter of time:
evaluating the storage of fecal samples for steroid analysis. Gen.
Comp. Endocrinol. 128 (1), 57–64.
Larsen, P.R., Davies, T.F., Hay, I.D., 1998. The thyroid gland. In:
Wilson, J.D., Foster, D.W., Kronenberg, H.M., Larsen, P.R.
(Eds.), Williams Textbook of Endocrinology. W.B. Saunders,
Philadelphia, pp. 389–516.
Lesage, J., Blondeau, B., Grino, M., Breant, B., Duouy, J.P., 2001.
Maternal undernutrition during late gestation induces fetal over-
exposure to glucocorticoids and intrauterine growth retardation,
and disturbs the hypothalamo–pituitary adrenal axis in the
newborn rat. Endocrinology 142 (5).
Merrick, R.L., Chumbley, M.K., Byrd, G.V., 1997. Diet diversity of
Steller sea lion s and their population decline: a potential
relationship. Can. J. Fish Aquat. Sci. 54, 1342–1348.
Merrick, R.L., Loughlin, T.R., Calkins, D.G., 1987. Decline in
abundance of the northern sea lion (Eumetopias jubatus) in Alaska.
Fish. Bull. U.S..
Monfort, S.L., Mashburn, K.L., Brewer, B.A., Creel, S.R., 1998.
Evaluating adrenal activity in African wild dogs (Lycaon pictus) by
fecal corticosteroid analysis. J. Zoo Wildlife Med. 29 (2), 129–133.
Mostl, E., Palme, R., 2002. Hormones as indicators of stress. Domest.
Anim. Endocrinol. 23 (1–2), 67–74.
Oki, C., Atkinson, S., in press. Cortisol and thyroid hormones
secretory patterns and concentrations in the harbor seal (Phoca
vitulina) in summer and winter seasons. Gen. Comp. Endocrinol.
Pitcher, K., Calkins, D., 1981. Reproductive biology of Steller sea lions
in the Gulf of Alaska. J. Mammal. 62, 599–605.
Pitcher, K.W., Calkins, D.G., Pendleton, G.W., 1998. Reproductive
performance of female Steller sea lions: an energetics-based
reproductive strategy? Can. J. Zool. 76, 2075–2083.
Rea, L.D., Castellini, M.A., Fadely, B.S., Loughlin, T.R., 1998.
Health status of young Steller sea lion pups (Eumetopias jubatus) as
indicated by blood chemistry and hematology. Comp. Biochem.
Physiol. A 120, 617–623.

K.L. Mashburn, S. Atkinson / General and Comparative Endocrinology 136 (2004) 371–381 381
Rivier, C., Rivest, S., 1991. Review: effect of stress on the activity of
the hypothalamic–pituitary–gonadal axis – peripheral and central
mechanisms. Biol. Reprod. 45, 523–532.
Ruam-Suryan, K.L., Pitcher, K.W., Calkins, D.G., Sease, J.L.,
Loughlin, T.R., 2002. Dispersal, rookery fidelity, and metapopu-
lation structure of Steller sea lions (Eumetopias jubatus) in an
increasing and decreasing population in Alaska. Mar. Mammal Sci.
18 (3), 746–764.
Sinclair, E.H., Zeppelin, T.K., 2002. Seasonal and spatial differences in
diet in the Western stock of Steller Sea lions (Eumetopias jubatus).
J. Mammal. 83 (4), 973–990.
St. Aubin, D.J., Dierauf, L.A., 2001. Stress and marine mammals. In:
Dierauf, L.A., Gulland, F.M.D. (Eds.), CRC Handbook of Marine
Mammal Medicine, second ed. CRC Press, New York.
Terio, K.A., Citino, S.B., Brown, J.L., 1999. Fecal cortisol metabolite
analysis for noninvasive monitoring of adrenocortical function
in the cheetah (Acinonyx jubatus). J. Zoo Wildlife Med. 30 (4),
484–491.
Turner Jr, J.W., Tolson, P., Hamad, N., 2002. Remote assessment of
stress in white rhinoceros (Ceratotherium simum) and black
rhinoceros (Diceros bicornis) by measurement of adrenal steroids
in feces. J. Zoo Wildlife Med. 33 (3), 214–221.
Walker, E.F., Walder, D.J., Reynolds, F., 2001. Developmental
changes in cortisol secretion in normal and at risk youth. Dev.
Psychopathol. 13 (3), 721–732.
Washburn, B.E., Millspaugh, J.J., 2002. Effects of simulated
conditions on glucocorticoid metabolite measurements in white-
tailed deer feces. Gen. Comp. Endocrinol. 127 (3), 217–222.
Wasser, S.K., Hunt, K.E., Brown, J.L., Cooper, K., Crockett, C.M.,
Bechert, U., Millspaugh, J.J., Larson, S., Monfort, S.L., 2000. A
generalized fecal glucocorticoid assay for use in a diverse array of
nondomestic mammalian and avian species. Gen. Comp. Endocri-
nol. 120 (3), 260–275.
Whittle, W.L., Patel, F.A., Alfaidy, N., Holloway, A.C., Fraser, M.,
Gyomorey, S., Lye, S.J., Gibb, W., Challis, J.R.G., 2001. Gluco-
corticoid regulation of human and ovine parturition: The relation-
ship between fetal hypothalamic–pituitary–adrenal axis activation
and intrauterine prostaglandin production. Biol. Reprod. 64, 1019–
1032.
York, A., 1994. The population dynamics of Northern sea lions 1975–
1985. Mar. Mammal Sci. 10, 38–51.
Zenteno-Savin, T., Castellini, M.A., Rea, L.D., Fadely, B.S., 1997.
Plasma haptoglobin levels in threatened Alaskan pinniped popu-
lations. J. Wildlife Dis. 33, 64–71.
Related Documents











