Eukaryotic Gene Regulation • Last lecture we covered elements of the eukaryotic gene and described how each contributed to the creation of a functional mRNA, capable of producing a protein • Basal level transcription = negligible physiological effect • Induced level transcription = caused by transcription factors • The elements of the gene covered last lecture are part of the

Eukaryotic Gene Regulation Last lecture we covered elements of the eukaryotic gene and described how each contributed to the creation of a functional mRNA,
Jan 01, 2016
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eukaryotic Gene Regulation
• Last lecture we covered elements of the eukaryotic gene and described how each contributed to the creation of a functional mRNA, capable of producing a protein
• Basal level transcription = negligible physiological effect
• Induced level transcription = caused by transcription factors
• The elements of the gene covered last lecture are part of the network of gene expression control points

Review: Central Dogma Revised
DNA
1 GENE
RNA PROTEIN
1 ENZYME
• This is an oversimplification
• Multiple genes can arise from the same DNA
• Some genes encode subunits of large enzyme complexes(ex: RNA polymerase)
• Genes can have multiple functions (sometimes tissue-based)
• Catalytic RNAs, tRNAs, rRNAs (not proteins)
• Genes can have multiple reading frames (six possible)

Genes Within Genes
• Three reading frames per strand
• Possible to have promoters at either end of a gene (ie, both strands are template strands)
• Ex: Mitochondrial ATPase genes
Molecular Biology of the Cell

DNA
RNA
mRNA
Protein
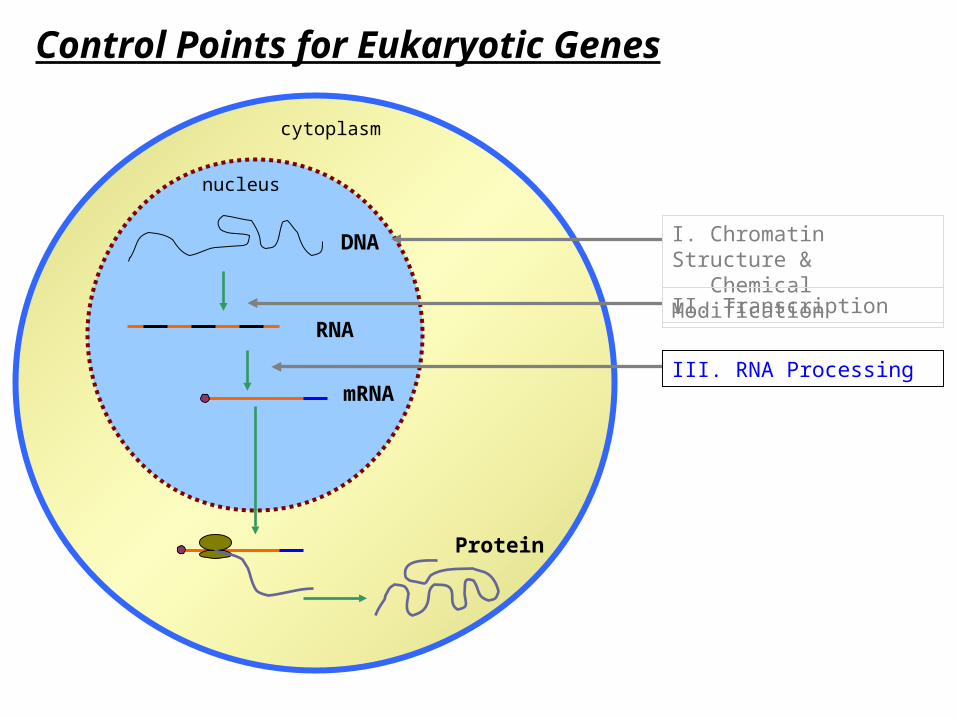
I. Chromatin Structure & Chemical ModificationII. Transcription
III. RNA Processing
IV. Nuclear Export
VI. Protein Modification & Trafficking
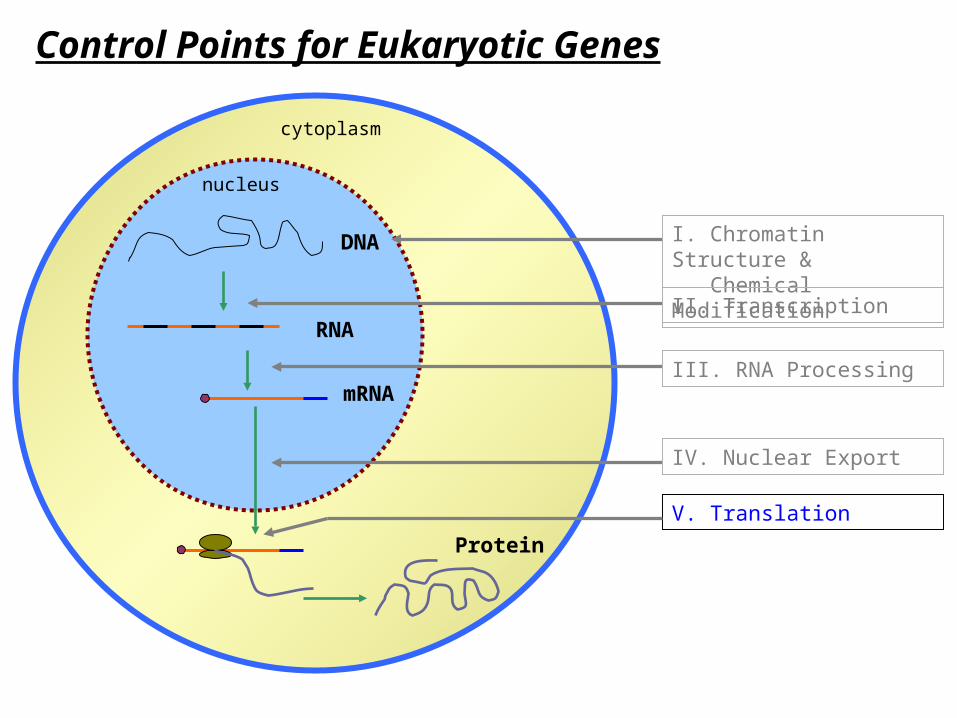
Control Points for Eukaryotic Genes
V. Translation
cytoplasm
nucleus

DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical Modification
Control Points for Eukaryotic Genes
cytoplasm
nucleus

Review: Euchromatin & Heterochromatin
• The structure of chromatin gives an opportunity to control gene expression
• Heterochromatin = highly condensed, unable to transcribe
• Euchromatin = relaxed, areas of transcription
What causes these areas to be condensed?

Histone Modification & Gene Expression
• Remember: Histones bind to DNA because the many (+) charged amino acids (Lys & Arg) bind to (-) PO4 groups of DNA
• Transferring an acetyl group (COCH3) to these amino acids nullifies their (+) charge
• Without this electrostatic interaction, DNA “falls” off the nucleosome
• The N-terminal tails of H3 and H4 are the most common sites of acetylation
• Heterochromatin is less acetylated than euchromatin, which causes its more condensed conformation

Histone Modification & Gene Expression
• Histone acetyl transferases (HATs) are the enzymes that acetylate histone proteins
• Many transcription factors have HAT activity; this is how they unwind the nucleosome structures of promoters
• Transcriptional repressors can recruit histone deacetylases (HDACs) to promoter sites to condense the DNA

DNA Methylation
• Methylation is another way of controlling gene expression
• Histone proteins can be methylated as well as acetylated
• DNA can be methylated directly, mainly at C residues in CG dinucleotide pairs
• Methylated DNA is transcriptionally inactive
• Factors like Methyl-CpG binding protein will bind to methylated sites and have HDAC activity
• Heterochromatin is highly methylated

DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical ModificationII. Transcription
Control Points for Eukaryotic Genes
cytoplasm
nucleus

Transcription Initiation Control
• Primary site of gene expression control
• We know that basal level transcription is achieved by the basal initiation complex of general TFs
• Gene-specific TFs accelerate transcription to significant levels
• These TFs can bind to promoter elements upstream of the RNA polymerase binding site (TATA box) or to enhancers

Transcription Factor Structure
• Transcription factors usually have two major domains:
DNA-Binding Domain binds TF to consensus sequence of the promoter
Activation Domain binds TF to consensus sequence of the promoter
• These two domains are modular; ie they can be switched and still function
• Called “domain swapping”
• More on this when we talk about yeast
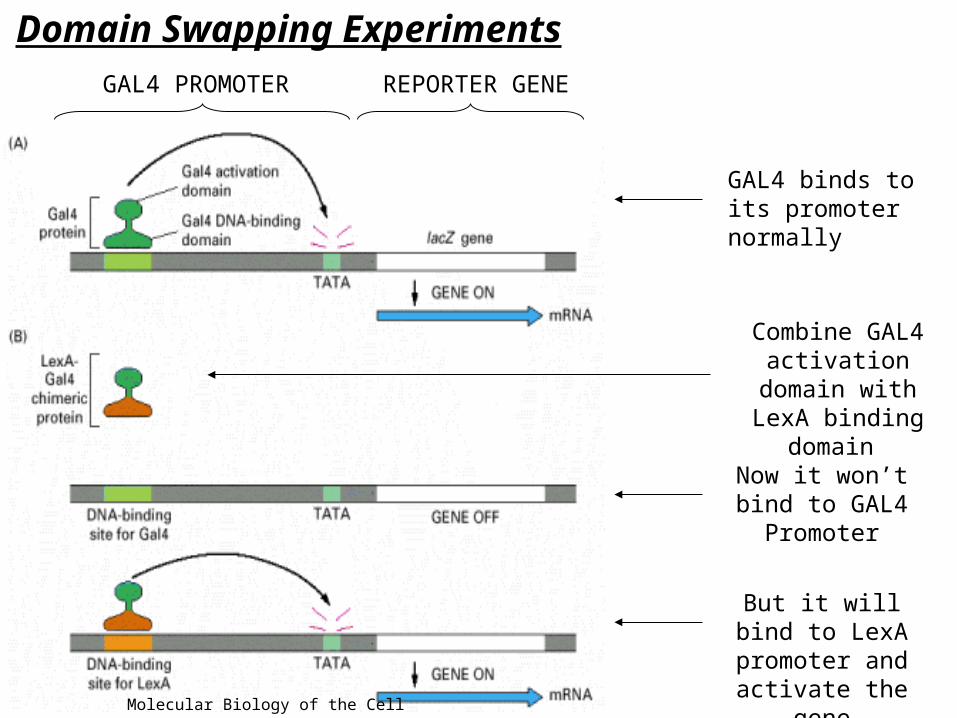
Domain Swapping Experiments
GAL4 binds to its promoter normally
Now it won’t bind to GAL4 Promoter
But it will bind to LexA promoter
and activate the gene
GAL4 PROMOTER REPORTER GENE
Molecular Biology of the Cell
Combine GAL4 activation domain with LexA binding
domain

Transcription Factor Binding
• DNA-binding domains of TFs attach to consensus sequences by hydrogen bonds
• This binding usually occurs on bases exposed in the major groove; minor groove is usually too tight for domains to fit
Leninger Principles of Biochemistry

DNA-Binding Domains: Helix-Turn-Helix
• HTH domain is also called homeodomain, because it is found in homoeobox (hox) genes
• Conserved amino acids of recognition helix bind to major groove bases
• Example: lac repressor
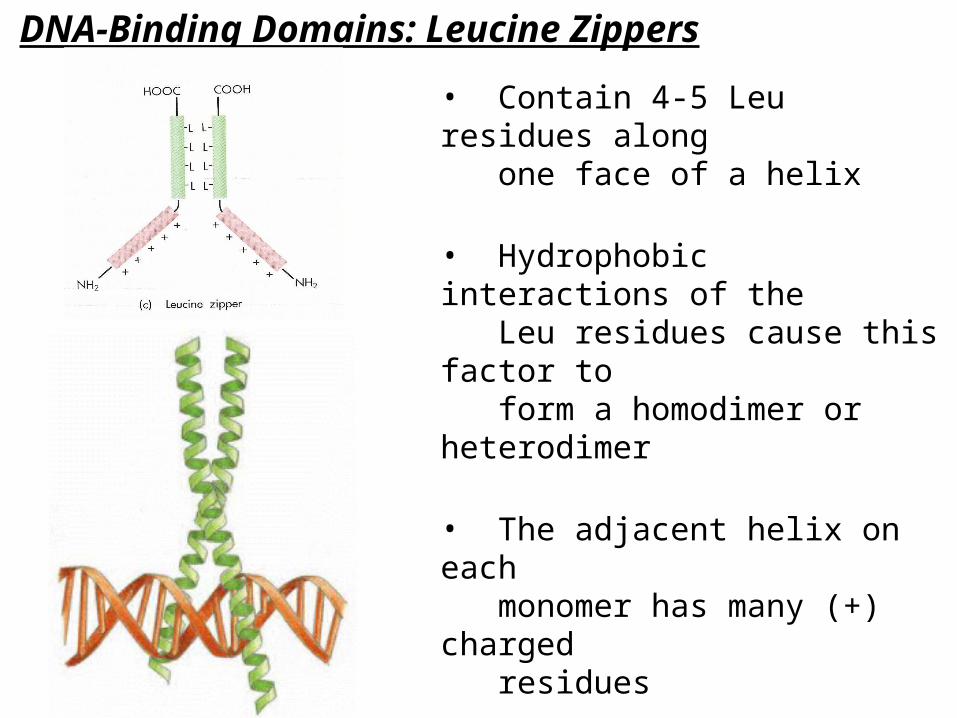
DNA-Binding Domains: Leucine Zippers
• Contain 4-5 Leu residues along one face of a helix
• Hydrophobic interactions of the Leu residues cause this factor to form a homodimer or heterodimer
• The adjacent helix on each monomer has many (+) charged residues
• These helices bind DNA in the major groove in a “scissor grip”
• Ex: c-fos & c-jun (cell cycle TFs, form heterodimers)

DNA-Binding Domains: Zinc Finger
• Conserved pairs of Cys & His residues bind a Zn atom
• A stem-loop structure (the finger) binds to DNA in the major groove
• These “finger” motifs usually occur in groups of three
• Sometimes the two His residues will be Cys (ie, four Cys total)
• Ex: TFIIIA

DNA-Binding Domains: Basic Helix-Loop-Helix
• Similar to Leu zipper
• Bind as homo- or hetero-dimers
• Basic, (+) charged DNA binding domains, bind in “scissor grip”
• Two pairs of helices join through hydrophobic interactions
• Ex: c-myc, a cell cycle gene

DNA Activation Domains
• Much more varied in structure than DNA-binding domains
• Act in a variety of ways to produce one of two specific effects:
I. Alter the physical structure of the promoter to aid General TF Binding:
II. Increase the binding efficiency of, or stabilize General TFs
• Recruit HATs
• Unwind DNA
• Bind to General TFs and induce conformational changes
• Activate General TFs by chemical modification

Control of Transcription Factors• What induces a transcription factor to bind to DNA?
• Signaling Pathways
Human molecular genetics
c-myc, c-fos, c-jun
Cell Division
• A hormone or other factor binds to a receptor on the cell surface
• Signal transduction pathways activate, the ultimate result of which is the recruitment of a transcription factor
• Signaling can initiate expression of a transcription factor or activate (ex: phosphorylate) a factor that has already been transcribed
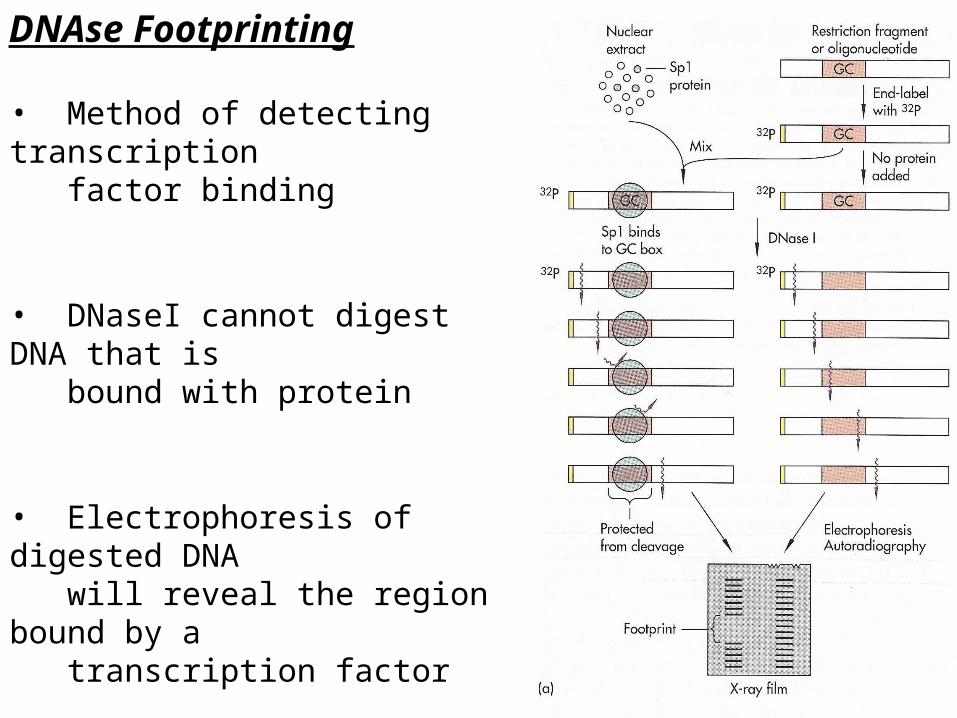
DNAse Footprinting
• Method of detecting transcription factor binding
• DNaseI cannot digest DNA that is bound with protein
• Electrophoresis of digested DNA will reveal the region bound by a transcription factor
DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical ModificationII. Transcription
III. RNA Processing
Control Points for Eukaryotic Genes
cytoplasm
nucleus

Alternative Splicing
• mRNA is capped, poly-adenylated and spliced
• Splicing can occur in different patterns, thus producing totally different gene products
• Since exons often represent protein domains, excluding an exon by splicing results in a protein with different function (ex: localization signal)
• Trans-splicing can occur; exchanges exons between different mRNAs
Ex: trypanosomes

Drosophila sxl: Sex Determination by Splicing
Stop codon
present in this exon
Stop codon
present in this exon
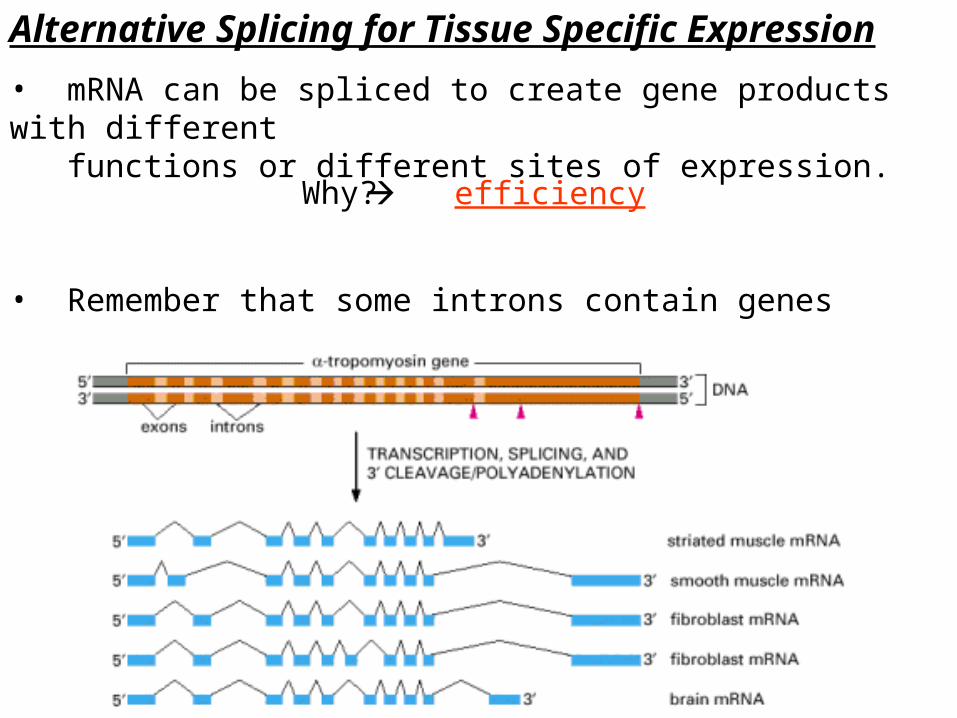
Alternative Splicing for Tissue Specific Expression• mRNA can be spliced to create gene products with different functions or different sites of expression.
• Remember that some introns contain genes
efficiencyWhy?

DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical ModificationII. Transcription
III. RNA Processing
IV. Nuclear Export
Control Points for Eukaryotic Genes
cytoplasm
nucleus

mRNA Export
• mRNA must be translocated from the nucleus to the cytoplasm for translation
• mRNA is inherently unstable; easily degrades (mRNA decay)
• Capping and poly-A tract aids stability and against nucleases
• mRNA is not usually linear; forms secondary structures that help keep it stable
• Secondary folding also helps compact it for travel through the nuclear pore complex

mRNA Secondary Structure
Hairpin
Stem-Loop
3-Stem Junction
SymmetricInternal Loop
AsymmetricLoop
Bulge
DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical ModificationII. Transcription
III. RNA Processing
IV. Nuclear Export
V. Translation
Control Points for Eukaryotic Genes
cytoplasm
nucleus
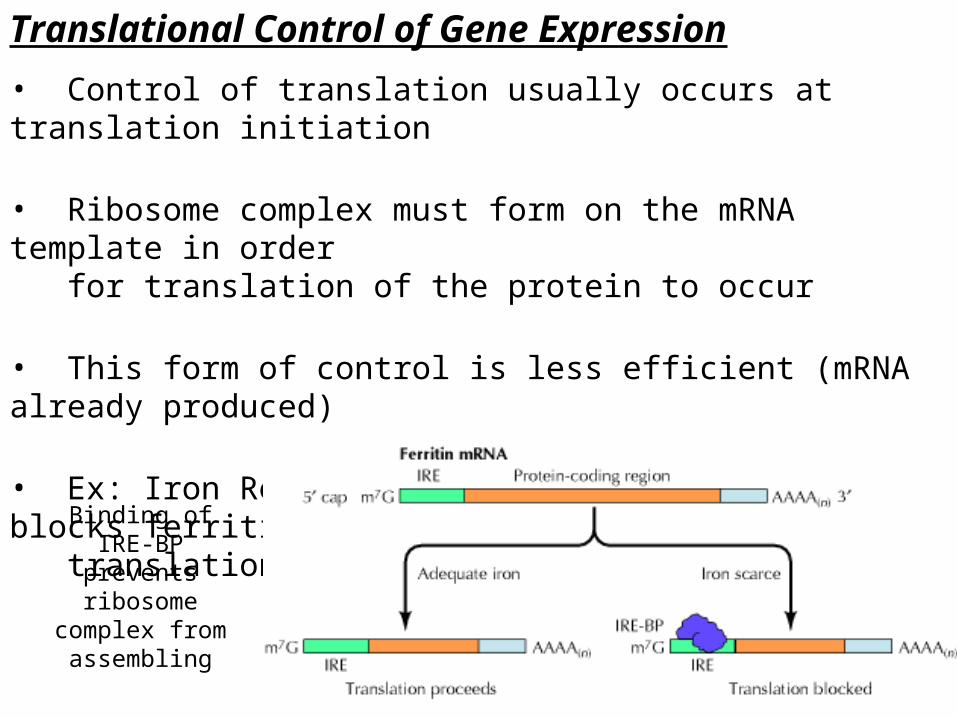
Translational Control of Gene Expression
• Control of translation usually occurs at translation initiation
• Ribosome complex must form on the mRNA template in order for translation of the protein to occur
• This form of control is less efficient (mRNA already produced)
• Ex: Iron Response Element Binding Protein – blocks ferritin translation when Fe is absent
Binding of IRE-BP prevents ribosome
complex from assembling

DNA
RNA
mRNA
Protein
I. Chromatin Structure & Chemical ModificationII. Transcription
III. RNA Processing
IV. Nuclear Export
V. Translation
VI. Protein Modification & Trafficking
Control Points for Eukaryotic Genes
cytoplasm
nucleus

Protein Modification and Trafficking
• After translation, proteins are folded & chemically modified
• One polypeptide can have multiple functions if processed different (again, 1 gene ≠ 1 enzyme)
• Protein modification occurs in the Golgi apparatus
• Gene expression is not complete until the protein reaches the region of the cell where it is useful
• Protein trafficking generally occurs via vesicles that bud off the Golgi apparatus and direct the protein to an organelle
• The destination of a protein is determined by its sequence and modifications and can be controlled to accommodate the cell’s metabolic needs

Post-Translation Modifications
Peptide bond cleavage
MODIFICATION PURPOSE
Expose an active site (ex: zymogens)
Glycosylation Variable (ex: cell-surface antigens)
Addition of Lipids Membrane-surface proteins primarily
Disulfide Bond Formation Enhance Structure & Stability
Hydroxylation Structural (ex: collagen)
Phosphorylation Activate Enzymes or TFs
Related Documents











