ETİL ALKOL İLE OLUŞTURULAN KARACİĞER TOKSİKASYONUNDA PUNİCALAGİN’İN OKSİDATİF STRES VE İNFLAMASYON ÜZERİNE ETKİSİ YÜKSEK LİSANS TEZİ Esma YANPAR MERSİN ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ ECZACILIK BİYOKİMYA ANABİLİM DALI MERSİN AĞUSTOS-2019

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ETİL ALKOL İLE OLUŞTURULAN KARACİĞER TOKSİKASYONUNDA PUNİCALAGİN’İN OKSİDATİF STRES VE
İNFLAMASYON ÜZERİNE ETKİSİ
YÜKSEK LİSANS TEZİ
Esma YANPAR
MERSİN ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
ECZACILIK BİYOKİMYA
ANABİLİM DALI
MERSİN
AĞUSTOS-2019

ii
ETİL ALKOL İLE OLUŞTURULAN KARACİĞER TOKSİKASYONUNDA PUNİCALAGİN’İN OKSİDATİF STRES VE
İNFLAMASYON ÜZERİNE ETKİSİ
YÜKSEK LİSANS TEZİ
Esma YANPAR
MERSİN ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
ECZACILIK BİYOKİMYA
ANABİLİM DALI
Danışman
Prof. Dr. A. Erdinç YALIN
MERSİN
AĞUSTOS-2019

iii

iv

v
ÖZET
Etil Alkol İle Oluşturulan Karaciğer Toksikasyonunda Punicalagin’in Oksidatif Stres ve İnflamasyon Üzerine Etkisinin Araştırılması
Punicalagin, punica granatum (nar) bitkisi kabuğunda bulunur ve birçok biyoaktiviteye sahiptir.
Punicalaginin antioksidan, antiinflamatuvar, antikanser ve antiaterosklerotik özellikte olduğu
kanıtlanmıştır. Çalışmamızda deneysel olarak etil alkol toksisitesi oluşturulmuş sıçanlarda
punicalagin etken maddesinin oksidatif stres ve inflamasyon üzerine etkisi incelenmiştir.
Çalışmamızda 24 adet sıçan 4 farklı gruba ayrılmıştır. Grup 1 kontrol grubudur, Grup 2 etil alkol
grubudur, grup 3 punicalagin grubudur, grup 4 punicalagin ve etil alkol grubudur. Deneysel
çalışmamız 21 gün devam etmiştir. Gruplara ayrıdığımız sıçanlara punicalagin, etil alkol ve etil
alkol + punicalagin gavaj yolu ile verilmiştir. Deney protokolü sonucunda deney hayvanları
sakrifiye edilerek karaciğer dokuları izole edilmiştir. İzole edilen bu dokular homojenize edilerek
süperoksit dismutaz (SOD), katalaz (KAT), malondialdehit (MDA), glutatyon (GSH), interlökin-6
(IL-6) ve tümör nekrozis faktör-α (TNF-α) düzeyleri incelenmiştir. Karaciğer dokusunda, etil
alkol ile toksisite oluşturulduktan sonra MDA seviyesinde artma, KAT ve SOD enzim aktivitesinde
azalma gözlenmiştir. Punicalagin uygulaması MDA düzeyini istatistiksel olarak anlamlı bir şekilde
düşürmüştür, KAT ve SOD enzim aktivitesi ise istatistiksel olarak anlamlı bir şekilde artmıştır.
Etil alkol ile toksisite oluşturulmuş KC dokusunda TNF-α ve IL-6 düzeyi istatistiksel anlamlı
olarak artmıştır. Punicalagin uygulaması TNF-α ve IL-6 seviyesini düşürmüştür. TNF-α’nın
düşüşü istatistiksel olarak anlam ifade etmiştir. Sonuç olarak etil alkolün oksidatif stresi
arttırdığı, antioksidan enzim seviyelerinde düşüşe sebep olduğu ve inflamasyon belirteçlerini
arttırdığı gözlenmiştir. Punicalaginin ise oksidatif stresi azalttığı ve antioksidan enzim
seviyelerini arttırdığı inflamasyon belirteçlerini azalttığı saptanmıştır. Bundan dolayı etil alkol
kullanan bireylerde oluşabilecek toksikasyona karşı punicalaginin kullanılabileceği
düşünülmektedir.
Anahtar Kelimeler: Oksidatif stres, Antioksidan, Antiinflamatuvar, Punicalagin
Danışman: Prof. Dr. A. Erdinç YALIN, Mersin Üniversitesi, Eczacılık Fakültesi Biyokimya
Anabilim Dalı, Mersin.

vi
ABSTRACT
Investigation of the Effect of Punicalagin on Oxidative Stress and Inflammation in Ethyl
Alcohol-Induced Liver Toxication
Punicalagin is found in the bark of punica granatum (pomegranate) and has many bioactivities.
Punicalagine has been shown to be antioxidant, antiinflammatory, anticancer and
antiatherosclerotic. In this study, the effect of punicalagin on oxidative stress and inflammation
was investigated experimentally in rats with ethyl alcohol toxicity. In our study, 24 rats were
divided into 4 different groups. In our study, 24 rats were divided into 4 different groups. Group
1 is the control group, Group 2 is the ethyl alcohol group, group 3 is the punicalagin group, group
4 is the punicalagin and ethyl alcohol group. Our experimental study continued for 21 days.
Punicalagin, ethyl alcohol and ethyl alcohol + punicalagin were given by gavage to rats. As a result
of the experimental protocol, the animals were sacrificed and the liver tissues were isolated.
These isolated tissues were homogenized and superoxide dismutase (SOD), catalase (KAT),
malondialdehyde (MDA), glutathione (GSH), interleukin-6 (IL-6) and tumor necrosis factor-α
(TNF-α) levels were examined. Liver toxicity was increased by MDA level and decreased CAT and
SOD enzyme activity after toxicity with ethyl alcohol. Punicalagin administration significantly
decreased MDA levels, whereas CAT and SOD enzyme activity increased significantly. TNF-α and
IL-6 levels were significantly increased in ethyl alcohol-induced toxicity in liver tissue.
Punicalagin administration decreased TNF-α and IL-6 levels. The decrease in TNF-α was
statistically significant. As a result, it was observed that ethyl alcohol increased oxidative stress,
decreased antioxidant enzyme levels and increased inflammation markers. Punicalagine
decreased oxidative stress and increased antioxidant enzyme levels and decreased inflammation
markers. Therefore, it is thought that punicalagine can be used against toxicity in individuals who
use ethyl alcohol.
Keywords: Oxidative stress, Antioxidant, Antiinflammatory, Punicalagin
Advisor: Prof. Dr. A. Erdinç YALIN, Mersin University, Faculty of Pharmacy, Department of
Biochemistry, Mersin.

vii
TEŞEKKÜR
Biyokimya yüksek lisans eğitimim süresince bilgi birikimi, tecrübesi, anlayışı ve sabrı ile daima
desteğini hissettiğim değerli hocam Prof. Dr. Ali Erdinç YALIN’a,
Bilgi paylaşımları ve desteklerinden dolayı değerli bölüm hocalarım Prof. Dr. Serap YALIN’a, Prof.
Dr. Necmiye CANACANKATAN’a ve Dr. Öğr. Üyesi Nil Zeynep ÜNAL’a,
Tez çalışmalarım esnasında ve sonrasında desteklerini benden esirgemeyen Metin YILDIRIM,
Merih AKKAPULU ve Özge ERYİĞİT’e,
Sıçanların beslenmesi, bakımı ve gavaj uygulanması esnasında desteklerini esirgemeyen deney
hayvanları laboratuvarı çalışanlarına,
Ve son olarak da bugünlere gelmemde büyük payı olan başta annem Fatma YANPAR ve babam
Ahmet YANPAR olmak üzere aileme teşekkürü bir borç bilirim.
Bu tez, Mersin Üniversitesi Bilimsel Araştırma Projeleri Birimi tarafından 2018-1-TP2-2804 nolu
proje olarak desteklenmiştir.

viii
İÇİNDEKİLER
Sayfa İÇ KAPAK ii ONAY iii ETİK BEYAN iv ÖZET v ABSTRACT vi TEŞEKKÜR vii İÇİNDEKİLER viii TABLOLAR DİZİNİ xi ŞEKİLLER DİZİNİ xii KISALTMALAR ve SİMGELER xiii 1. GİRİŞ 1 2. KAYNAK ARAŞTIRMALARI 3 2.1. Serbest Radikaller 3 2.1.1. Reaktif Oksijen Türleri 4 2.1.1.1. Süperoksit Radikali 4 2.1.1.2. Singlet Oksijeni 4 2.1.1.3. Hidroksil Radikali 5 2.1.1.4. Hidrojen Peroksit Radikali 5 2.1.2. Reaktif Nitrojen Türleri 6 2.1.2.1. Nitrikoksit Radikali 6 2.1.2.2. Peroksinitrit Radikali 6 2.2. Serbest Radikal Kaynakları 7 2.2.1. Endojen Kaynaklar 7 2.2.2. Eksojen Kaynaklar 7 2.3. Serbest Radikallerin Etkileri 7 2.3.1. Hücre Membran Lipitleri Üzerine Etkisi 7 2.3.2. Proteinler Üzerine Etkileri 9 2.3.3. Karbonhidratlar Üzerine Etkileri 9 2.3.4. Nükleik Asitler Üzerine Etkileri 9 2.4. Oksidatif Hasar 10 2.5. Antioksidan Savunma Sistemleri 11 2.5.1.Enzimatik Antioksidanlar 11 2.5.1.1. Süperoksit Dismutaz 11 2.5.1.2. Katalaz 12 2.5.1.3. Glutatyon Peroksidaz 12 2.5.1.4. Glutatyon Redüktaz 13 2.5.1.5. Glutatyon-S-Transferaz 13 2.5.2 Nonenzimatik Antioksidanlar 14 2.5.2.1. Glutatyon 14 2.5.2.2. Vitamin E 14 2.5.2.3. Vitamin C 15 2.5.2.4. Bilirubin 15 2.5.2.5 β-Karoten 15 2.5.2.6. Koenzim Q10 15 2.5.3.7. α-Lipoik Asit 16 2.5.3.8. Selenyum 16 2.5.4.9. Folik Asit 16 2.5.5.10. Flavanoidler 16 2.6. Punicalagin 17
2.7. Sitokinler 18
2.7.1. Sitokinlerin Genel özellikleri 19

ix
Sayfa 2.7.2. Sitokinlerin Organizmadaki Etkileri 19 2.7.3. Sitokinlerin İşlevlerine Göre Sınıflandırılması 19 2.7.3.1. Doğal İmmüniteye Aracılık Eden Sitokinler 19 2.7.3.1.1. Tip I İnterferonlar (IFN) 20 2.7.3.1.2. Tümör Nekrozis Faktör (TNF ) 20 2.7.3.1.2.1. TNF-α 22 2.7.3.1.2.2. TNF-β 23 2.7.3.1.3. İnterlökin-1 (IL-1) 23 2.7.3.1.4. İnterlökin-6 (IL-6) 24 2.7.3.1.5. Kemokinler 25 2.7.3.2. Lenfosit aktivasyonu, Büyüme ve Farklılaşmasın Düzenleyen Sitokinler 25 2.7.3.2.1. İnterlökin-2 (IL-2) 25 2.7.3.2.2. İnterlökin-4 (IL-4) 26 2.7.3.2.3. Transforming Büyüme Faktörü-β (TGF-β) 27 2.7.3.3. Bağışıklık Aracılığı İle İnflamasyonu Düzenleyen Sitokinler 28 2.7.3.3.1. İnterferon-γ 28 2.7.3.3.2. Lenfotoksin (LT) 28 2.7.3.3.3. İnterlökin-10 29 2.7.3.3.4. İnterlökin-5 29 2.4.3.3.5. İnterlökin-12 29 2.7.3.3.6. Migrasyon İnhibisyon Faktör (MIF) 30 2.7.3.4. İmmatör Lökositlerin Büyüme ve Farklılaşmasına Neden Olan Sitokinler 30 2.7.3.4.1. C-Kit Ligand 30 2.7.3.4.2. İnterlökin-3 (IL-3) 31 2.7.3.4.3. Granülosit-Makrofaj Koloni Uyaran Faktör (GM-CSF) 31 2.7.3.4.4. Monosit-Makrofaj Koloni Uyaran Faktör (M-CSF) 31 2.7.3.4.5. Granülosit-Koloni Uyaran Faktör (G-CSF) 31 2.7.3.4.6. İnterlökin-7 (IL-7) 32 2.7.3.4.7. Diğer Koloni Uyaran Sitokinler 32 2.8. Alkol Metabolizması 32 2.8.1. Alkolün Karaciğerde Oksidatif Hasar Üzerine Etkisi 35 2.8.2. Alkolün Karaciğerde Sitokin Üretimi ve İnflamasyon Üzerine Etkisi 35 3. MATERYAL ve YÖNTEM 37 3.1. Materyal 37 3.1.1. Kullanılan Alet ve Malzemeler 37 3.1.2. Kullanılan Kimyasal Malzemeler 37 3.1.3. Hayvan Materyali 38 3.2. Yöntem 39 3.2.1. Çalışma Örneklerinin Hazırlanması 39 3.2.1.1. Doku Örnekleri 39 3.2.1.2. Homojenizasyon 39 3.2.2. Analiz Yöntemleri 39 3.2.2.1. Glutatyon Tayini 39 3.2.2.2. Katalaz Enzim Aktivitesi Tayini 40 3.2.2.3. Süperoksit Dismutaz Enzim Aktivitesi Tayini 41 3.2.2.4. Malondialdehit Ölçümü 42 3.2.2.5. Sitokin Düzeyleri Ölçümü 43 3.2.2.5.1. TNF-α Düzeyi 43 3.2.2.5.2. IL-6 Düzeyi 44 3.2.3. İstatiksel Analiz 45 4. BULGULAR 46 5. TARTIŞMA ve SONUÇLAR 52 KAYNAKLAR 58

x
Sayfa ÖZGEÇMİŞ 71

xi
TABLOLAR DİZİNİ
Sayfa Tablo 2.1. Reaktif Oksijen Türleri 3 Tablo 2.2. Reaktif Nitrojen Türleri 3

xii
ŞEKİLLER DİZİNİ
Sayfa Şekil 2.1. MDA Oluşum Mekanizması 8 Şekil 2.2. Punicalagin’in Kimyasal Yapısı 18 Şekil 2.3. Punicalagin’in Hidroliz Reaksiyonu 18 Şekil 2.4. Biyolojik Sistemlerde Alkol Metabolizmasından Sorumlu Yolaklar 33 Şekil 2.5. Alkolün Hepatositlerdeki Metabolik Yıkım Yolları 33 Şekil 2.6. Alkol Dehidrogenaz Enzim Yolağı 34 Şekil 4.1. Karaciğer Dokusunda SOD Aktivitesi 46 Şekil 4.2. Karaciğer Dokusunda KAT Aktivitesi 47 Şekil 4.3. Karaciğer Dokusunda MDA Düzeyleri 48 Şekil 4.4. Karaciğer Dokusunda Glutatyon Düzeyleri 49 Şekil 4.5. Karaciğer Dokusunda TNF-α Düzeyleri 50 Şekil 4.6. Karaciğer Dokusunda IL-6 Düzeyleri 51

xiii
KISALTMALAR VE SİMGELER
Kısaltma/Simge Tanım °C Santigrat Derece µL Mikrolitre ADH Alkol Dehidrogenaz ALDH Asetaldehit Dehidrogenaz Cu Bakır DHLA Dihidrolipoik Asit DNA Deoksiribonükleik Asit EA Ellagik Asit EC SOD Ekstrasellüler Süperoksit Dismutaz FAD Flavoadenin Dinükleotit Fe Demir g Gram GM-CSF Granülosit-Makrofaj Koloni Uyaran Faktör GPx Glutatyon Peroksidaz GR Glutatyon Redüktaz GSH Glutatyon H₂O Su H₂O₂ Hidrojen Peroksit HHDP Heksahidroksidifenik Asit IFN İnterferon IL-1 İnterlökin-1 IL-10 İnterlökin-10 IL-4 İnterlökin-4 IL-6 İnterlökin-6 i.p. İntraperitonel KAT Katalaz kg Kilogram LA Lipoik Asit LF Lenfotoksin LOO. Peroksi Radikali MDA Malondialdehit MEOS Mikrozomal Etanol Oksidasyon Sistemi mg Miligram M-GSF Monosit-Makrofaj Koloni Uyaran Faktör MIF Migrasyon İnhibisyon Faktörü mL Mililitre Na⁺ Sodyum NADPH Nikotinamit Adenin Dinükleotitfosfat NO Nitrik Oksit NOS Nitrik Oksit Sentaz O₂ Oksijen OH˙ Hidroksil Radikali PG Punicalagin PUFA Çoklu Doymamış Yağ Asitleri RNA Ribonükleik Asit RNS Reaktif Nitrojen Türleri ROS Reaktif Oksijen Türleri SOD Süperoksit Dismutaz TGF- β Transforming Büyüme Faktörü- β TNF Tümör Nekrozis Faktör UV Ultraviyole

xiv
α Alfa β Beta

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
1
1. GİRİŞ
Kronik alkol tüketimi ile karaciğer hastalıkları arasında pozitif korelasyon olduğu
gözlenmiştir [1]. Günlük ortalama 80 gr ve üstü alkol tüktimi insan sağlığı açısından risk olarak
değerlendirilmektedir [2]. Kronik alkol tüketiminin kötüye kullanımıyla sonuçlanan karaciğer
hasarı, çeşitli genetik ve çevresel faktörleri içeren multifaktöriyel bir durumdur. Bu faktörler
arasında, serbest radikal aracılı oksidatif stresin rolü önemlidir. Etanol toksisitesinde oksidatif
hasarın oluşumu ilk olarak 1963 yılında Di Luzio tarafından önerilmiş ve sonrasında etanolün
çeşitli serbest radikallerin (oksijen türevli radikaller) oluşumunu teşvik ettiğini gösteren
deneysel çalışmalarla kanıtlanmıştır. Reaktif oksijen türleri (serbest radikaller) kolayca
reaksiyon verebilen moleküller olup özellikle mitokondride olmak üzere diğer hücre
organellerinde meydana gelen metabolizmanın doğal bir neticesi olarak veya inflamasyon,
iskemi, radyasyon, yaşlanma, oksijen basıncında artış ve kimyasal ajanlarla temas etme gibi
sebeplerle oluşurlar [3-5].
Kronik etil alkol muamelesi karaciğerde kuppfer hücrelerinden TNF-α, IL-1, IL-6 gibi
proinflamatuar sitokinlerin üretimini aktive eder. Bu sitokinler alkole bağlı gelişen karaciğer
toksisitesinde ve alkole bağlı gelişen karaciğer hastalıklarında rol alır [6].
Dış yörünge orbitallerinde ortaklanmamış bir elektron ihtiva eden elektrik yükü içeren
veya elektrik yükü içermeyen atom veya moleküllere serbest radikal denir. Bu moleküller
biyolojik sistemlerde normal metabolik yolakların aktivitesinin bir sonucu olarak oluştuğu gibi,
çeşitli dış faktörlerle de oluşabilmektedir. Oksijenli solunum yapan memelilerde serbest
radikaller birincil olarak oksijenden türemektedir [7]. Serbest radikallern organizmada yaptığı
zararlı etkileri yok etmek antioksidanlar ile mümkündür. Antioksidanlar serbest radikalleri
temizleyebilen ve hücrede meydana gelen hasarı önleyebilen moleküllerdir. Savunma sisteminin
bir parçası olan antioksidanlar, insan vücudunda doğal olarak üretilirler veya diyetle alınırlar.
Endojen ve eksojen kaynaklı olabilen antioksidanlar canlı organizmada serbest radikal süpürücü
olarak görev yaparlar. Serbest radikal süpürücü etkisiyle vücudun savunma sistemini
kuvvetlendirip, hastalık riskini azaltırlar [8].
Nar, punica granatum, punicaceae ailesine ait bir türdür. Nar antionsidan, antimikrobiyal,
antikarsinojen, antiinflamatuar terapötik etkilere sahip olduğu için hastalıklara karşı tedavide
kullanılmaktadır [9]. Ellagitaninler ve antosiyaninler nar meyvesinin antioksidan aktivitesinden
sorumlu olan polifenollerdir [10]. Özellikle meyvenin kabuk kısmında bol miktarda bulunan
ellagitaninler antioksidan etkinin % 92’sinden sorumludur. Meyve kabuğunda bulunan en temel
ellagitanin punicalagindir ve in vivo olarak ellagik asit ile küçük polifenollere parçalanırlar [11].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
2
Bizim çalışmamızdaki amaç, gavaj yoluyla etil alkol verilmiş ve akabinde etil alkol
toksisitesi oluşturulmuş sıçanlarda yine gavaj yoluyla nar kabuğu ekstraktından elde edilen
punicalagin verilerek antioksidan özellik gösterip göstermediğini ve inflamasyon üzerine nasıl
bir etki gösterdiğini incelemektir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
3
2. KAYNAK ARAŞTIRMALARI
2.1 Serbest Radikaller
Serbest radikaller, dış yörünge orbitallerinde bir ya da birden çok eşlenmemiş elektron
ihtiva ederler. Yüksek enerjiye sahip atom veya moleküllerdir [12-14]. Serbest radikaller sahip
oldukları eşlenmemiş elektron sebebiyle diğer maddelerle oldukça kolay reaksiyon verme
özelliğine sahiptirler. Yapısında eşlenik elektron bulunduran atomlar veya moleküller ise diğer
moleküllerle reaksiyon verme eğilimleri serbest radikallerle kıyaslanamayacak kadar azdır.
Çünkü bu atom veya moleküller kararlı yapıya sahiptirler. Bu nedenle kimyasal olarak
eşlenmemiş elektron bulundurmayıp kararlı yapıya sahip olan ve farklı moleküller ile serbest
radikallere kıyasla çok daha az reaksiyon verme eğilimi olan bu atom veya moleküller
nonradikaller olarak adlandırılırlar. Serbest radikaller oksijen kaynaklı ve nitrojen kaynaklı
olmak üzere iki gruba ayrılırlar. Oksijen kaynaklı olan serbest radikaller reaktif oksijen türleri
(ROS) ve nitrojen kaynaklı olan serbest radikaller ise reaktif nitrojen türleri (RNS) olarak
isimlendirilirler.
Süperoksit, hidroksil, peroksil, lipit peroksil ve alkoksil radikalleri reaktif oksijen türleri
(ROS) olarak adlandırılırlar ve tablo 2.1. ile gösterilmiştir. Nitrik oksit ve nitrojen dioksit ise
reaktif nitrojen türleri (RNS) olarak adlandırılırlar ve tablo 2.2. İle gösterilmiştir [15, 16]. Oksidan
madde olarak değerlendirilen, hidrojen peroksit, ozon, singlet oksijen, hipokloröz asit, nitrik asit,
peroksinitrit, dinitrojen trioksit ve lipit peroksit ise serbest radikal olarak adlandırılmazlar. Fakat
bu oksidan atom ve moleküller patolojik ve fizyolojik olayların doğal bir ürünü olarak canlı
organizmalarca üretilirler. Buna ek olarak canlı organizmada kolayca serbest radikal
reaksiyonlarının oluşmasına neden olurlar [17, 18].
Tablo 2.1. Reaktif Oksijen Türleri (ROS) [15] Radikaller Non-Radikaller Süperoksit, .O2- Hidrojen peroksit, H2O2 Hidroksil, OH. Hipoklorik asit, HOCL Peroksil, LO2. Ozon,O3 Alkoksil, LO. Singlet oksijen Hidroperoksil, HO2. Lipit peroksitleri
Tablo 2.2. Reaktif Nitrojen Türleri (RNS) [16] Radikaller Non-Radikaller Nikrikoksit, NO. Nitröz asit, HNO2 Nitrojen dioksit, NO2. Dinitrojen trioksit, N2O3
Dinitrojen tetraoksit, N2O4 Peroksinitrit, ONOO-

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
4
2.1.1. Reaktif Oksijen Türleri
Oksijen, canlı organizmalar için önemli moleküllerin başında gelir. Hidrojen, karbon,
nitrojen ve kükürt gibi atomlarla beraber, organik moleküllerin ana bileşenlerini meydana getirir
[17]. Oksijenin büyük bir çoğunluğu oksidatif fosforilasyon ile kullanılır [18]. Kullanılan bu
oksijenin bir kısmı hücre içerisinde özellikle mitokondrilerde reaktif oksijen türlerine
dönüştürülür [19].
2.1.1.1. Süperoksit Radikali
Moleküler oksijenin (O₂) elektronlarından birinin transferi neticesinde indirgenme
reaksiyonu ile nonstabil yapıdaki süperoksit radikali meydana gelir [20]. Major olarak ve ksantin
oksidaz gibi flavoenzimlerin katalizlediği reaksiyonların bir sonucu olarak, elektron rezervi
zengin bir ortam olan mitokondrinin iç zarında oluşur.
HO2 → H+ + O2⋅
İndirgenmiş geçiş metallerinin otooksidasyon reaksiyonları sonucunda da süperoksit
radikali oluşabilir.
Fe2+ + O2 → Fe3+ +O2⋅ [21].
2.1.1.2. Singlet Oksijeni
Oksijen molekülünün sahip olduğu elektronlarından bir tanesinin enerji alması ile
elektronun rutin dönüş yönünün tersi yönünde olan farklı bir yörüngeye geçiş yapması sonucu
oluşmasının yanında; süperoksit radikali ile nitrik oksit arasında gerçekleşen reaksiyon sonucu
ve hidrojen peroksit ile hipoklorit arasında gerçekleşen reaksiyon sonucunda da oluşabilir [22].
Yapısında ortaklanmamış elektron bulundurmayan singlet oksijeni (¹O2), serbest radikal
reaksiyonlarının başlangıç ürünlerinden biri olması açısından önemlidir. Bu nedenle
ortaklanmamış elektrona sahip olmadığı halde singlet oksijeni serbest radikal olarak
değerlendirilir. Singlet oksijen çoğunlukla laboratuvar ortamında fotosensitizasyon reaksiyonu
ile üretilir [23].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
5
2.1.1.3. Hidroksil Radikali
Hidroksil radikali, moleküler oksijene üç elektron transfer edilmesi ile meydana gelir
[24]. Hidroksil radikali, biyomoleküller ile oldukça kuvvetli reaksiyon vermesinden kaynaklı
canlı organizmaya diğer reaktif oksijen türleri ile kıyaslandığında çok daha fazla zarar verebilir.
Fe⁺² ve Cu⁺ veya diğer geçiş elementleri (Zn, Mn, Cr, Co, Ni, Mo) varlığında süperoksit radikali
indirgenerek hidroksile dönüştürülür ve bu reaksiyon “fenton reaksiyonu” olarak adlandırılır.
Süperoksit radikali fenton reaksiyonu ile bağlantı kurarak oluşan metal iyonlarının tekrardan
tüketilmesinde önemli bir rol oynar. Geçiş metalleri böylelikle OH˙ Oluşmasında önemli bir rol
oynarlar. Hidrojen peroksit ile süperoksit radikalinin reaksiyonu sonucunda da hidroksil
radikali meydana gelir ve bu reaksiyona ‘Haber-Weiss Reaksiyonu’ denir [25-28].
Fe+3 + O2•- → O2 + Fe+2
Fe+2 + H2O2 → Fe+3 + OH˙
Fenton Reaksiyonu
H2O2 + O2.- OH- + O2 + OH
Haber-Weiss Reaksiyonu
2.1.1.4. Hidrojen Peroksit Radikali
Serbest radikal oluşumuna yol açan önemli bir moleküldür. Bu nedenle serbest radikal
olmadığı halde ROS olarak değerlendirilir. Hücresel kompartımanlarda bulunan ürat oksidaz,
glikoz oksidaz ve D-amino asit oksidaz gibi enzimler sahip oldukları iki elektronu oksijene
transfer ederek hidrojen peroksit radikalini oluştururlar. Fenton reaksiyonu ve Haber-Weiss
reaksiyonu ile oldukça kuvvetli bir radikal olan hidroksil radikalini (OH˙) meydana getirir [29,
30].
Fe+3 + O2•- → O2 + Fe+2
Fe+2 + H2O2 → Fe+3 + OH- + OH•
Fenton reaksiyonu

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
6
H2O2 + O2.- OH- + O2 + OH
Haber-Weiss reaksiyon
2.1.2. Reaktif Nitrojen Türleri
Reaktif nitrojen türlerinin en önemlileri nitrikoksit radikali ve peroksinitrit
radikalleridir.
2.1.2.1. Nitrikoksit Radikali
Bir azot atomu ile bir oksijen atomunun çiftleşmemiş elektron vererek reaksiyona
girmesi ile oluşmuştur. Kimyasal yapısı nedeniyle radikal olarak değerlendirilir. NO’nun
yarılanma ömrü 10-20 saniyedir. NO metabolizasyonu, moleküler oksijen ile rekasiyon vererek
azot dioksit (NO2) oluşturması reaksiyonuna dayanır [31].
2NO + O2 2NO2
NO, nötrofiller, nöronlar, endotel hücreleri, makrofajlar, plateletler gibi çok çeşitli
hücrelerde üretilmektedir [32].
NO, biyolojik sistemlerdeki etkilerini hedef hücredeki guanilat siklaz aktivasyonu
yapıp, intrasellüler cGMP artışına sebep olarak yapmaktadır [33]. cGMP ise protein kinaz G
aktivasyonu ile fosforilasyonu uyarmaktadır. Ayrıca NO, O2· ile birleşerek sitotoksik etkilere
sahip bir radikal olan ONOO· ve OH˙ radikallerini meydana getirebilmektedir. Bu özellikleri,
NO’nun sitotoksik ve dejeneratif etkilerini kanıtlayan bir özelliğidir [34].
2.1.2.2. Peroksinitrit Radikali
Beyinde bulunan nöronal ve endoteliyal nitrik oksit sentaz (NOS) enziminin katalizlediği
reaksiyon neticesinde süsperoksit radikali ile gaz radikal olan nitrik oksit peroksinitrit oluşur
[35, 36, 37]. Fizyolojik pH da ONOO· hızla OH˙ ve azot dioksite parçalanır. Çok güçlü bir
prooksidan olan ONOO-, SOD ile reaksiyon vererek yine oldukça güçlü bir nitratlayıcı ajan
meydana getirir. Sonuçta gerçekleşen bu nitratlanma reaksiyonu hücresel disfonksiyon ve hücre
ölümüne neden olabilir [38, 39].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
7
2.2. Serbest Radikal Kaynakları
Hücrede reaktif oksijen ve nitrojen türevlerinin kaynağı endojen ve eksojen kaynaklı
olarak iki kısımda incelenir.
2.2.1. Endojen Kaynaklar
Peroksizomlarda bulunan enzimler, nükleus membranında, mitokondride ve
endoplazmik retikulumda bulunan elektron transport sistemi, makrofajlar ve fagositler, iskemi,
intoksikasyon, katekolaminler, tetrahidroproteinler, hidrokinonlar, tioller gibi küçük moleküllerin
otooksidasyonu, enzimler ve proteinler serbest radikal oluşturan endojen kaynaklardır [40, 41].
2.2.2. Eksojen Kaynaklar
Sigara dumanı, solventler, ilaçlar, antineoplastik ajanlar, metalik katyonlar, iyonize
radyasyon, UV ışınlar, pestisitler, ozon ve kirli hava serbest radikal oluşturan eksojen kaynaklardır
[42, 43].
2.3. Serbest Radikallerin Etkileri
Reaktif oksijen türlerinin biyolojik sistemlerde üretiminin artışı veya antioksidan
maddelerin azalmasına bağlı olarak oksidatif denge bozulur. Bozulan oksidatif dengeye bağlı
olarak artış gösteren serbest radikaller hücrelerdeki DNA, lipit, karbonhidrat, protein ve enzim gibi
önemli bileşenlere etki ederler [44].
2.3.1. Hücre Membran Lipitleri Üzerine Etkisi
Reaktif oksijen türleri hücre membranı üzerinde bulunan poliansatüre yağ asitlerinde
(PUFA) oksidasyon reaksiyonlarına neden olarak lipit peroksidasyon reaksiyonunu başlatırlar
[45].
Rekatif oksijen türleri içerisinde en güçlü ve en zararlı etkiye sahip olan serbest radikal
hidroksil radikalidir. Yüksek reaktiviteye sahip radikallerin hücre membran yapısında bulunan
poliansature yağ asitlerinden bir hidrojen atomunu metilen gruptan (-CH₂) koparması ile zincir
reaksiyonlar başlar. Hidrojen atomu sadece bir elektron içerir. Bu nedenle metilen gruptan bir
elektron kopması neticesinde karbon üzerinde eşleşmemiş bir elektron kalır (-•CH ). Yağ asitinin
kimyasal yapısında bulunan çift bağ, kendisine komşu olan karbon ile hidrojen arasındaki bağı
zayıflatır. Zayıflayan bağ hidrojenin kopması reaksiyonunu kolaylaştırır [46]. Hücresel membran
lipitlerinin poliansatüre yağ asiti zincirleri kimyasal yapısı nedeniyle peroksidasyon

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
8
reaksidasyonuna oldukça duyarlıdır. Hidrojen kaybeden yağ asitinin moleküler yapısında yeni
düzenlenmeler olur ve konjuge dien yapıyı alırlar. Meydana gelen konjuge dien yapı oksijenle
reaksiyon vererek lipit peroksil radikallerine (LOO˙) dönüşür. Meydana gelen bu peroksil
radikalleri ortamdaki yağ asitlerinden hidrojen kopmasına neden olur. Böylece zincirleme
peroksidasyon reaksiyonlarını başlatmış olurlar [47]. Lipit peroksidasyon reaksiyonları
neticesinde meydana gelen lipit peroksitleri son ürün olarak Malondialdehit (MDA), hegzenal ve 4-
Hidroksinonenal (HNE) isimli aldehitleri meydana getirir [48, 49]. Hücre membran lipitlerinde
meydana gelen peroksidasyon reaksiyonları ortama reaksiyon zincirini kıracak bir antioksidan
molekül girene kadar zincirleme olarak devam eder. Ortamda antioksidan yoksa peroksil
radikalleri birbirleri ile çapraz kovalent bağ oluşturarak membranın kimyasal yapısını bozar ve
membranı hasarlanmaya uygun bir duruma getirir. Biyolojik membranlarda meydana gelen
peroksidayon membran akışkanlığında bozulmaya, mebranların H⁺ ve diğer iyonlara karşı
geçirgenliğinde artışa, membran potansiyelinde azalmaya neden olarak membranlarda yırtıkların
oluşmasına ve organel içeriğinin sitoplazmaya sızmasına sebep olur. Bu zincir rekasiyonların
neticesinde hücre hasarı ve akabinde hücre ölümü meydana gelir [50].
Şekil 2.1. MDA Oluşum Mekanizması [47]

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
9
2.3.2. Proteinler Üzerine Etkileri
Biyoljik sistemlerde oluşan reaktif oksijen türleri hücre içindeki proteinlerde meydana
getirdikleri oksidatif değişikliklere bağlı olarak oksidatif hasara sebep olur. Bu oksidatif değişikler
geri dönüşümlü veya geri dönüşümsüz olabilir [51, 52]. Hücre içindeki protein yapılar oksidasyon
reaksiyonu verdiklerinde, yan zincirlerinde (Prolin, Arginin, lizin ve treonin) karbonil gruplar
meydana gelir. Protein karbonil yapılar α-amidasyon yolağı ile ve glutamil yan zincirlerin
oksidasyonu ile proteinlerin kimyasal bütünlüğünün bozulması sonucunda oluşular [53]. Serbest
radikallerin protein yapılarda oluşturdukları oksidatif değişiklikler, hücrenin temel yapısını
oluşturan enzimler ile proteinlerde, yapısal ve fonksiyonel farklılaşmalara sebep olur. Protein
karbonilasyonu ve tirozin nitrasyonu geri dönüşümsüz oksidatif değişikler olarak, sistein üzerinde
meydana gelen değişikler ise geri dönüşümlü oksidatif değişiklikler olarak değerlendirilirler [51,
52].
2.3.3. Karbonhidratlar Üzerine Etkileri
Monosakkaridlerin otooksidasyon reaksiyonu neticesinde hidrojen peroksit, diğer
peroksitler ve okzoaldehidler oluşur. Okzoaldehitler DNA, RNA ve proteinlere bağlanabilmeleri
bakımında mitoz bölünmeye karşı negatif etki gösterirler [54]. Hyalüronik asit bağ dokusunun
önemli bir mukopolisakkariti olmakla birlikte, sinovyal sıvıda bol bulunur. Eklem hastalıklarının
bazılarında sinovyal sıvıya çok sayıda hücre göçü meydana gelir ve ekstraselüler sıvıya H₂O₂ ve O₂˙
salgılarlar [55].
2.3.4. Nükleik Asitler Üzerine Etkileri
Serbest radikallerin sebep olduğu DNA hasarı başta kanser olmak üzere pek çok rahatsızlığın
patogenezinde önemli bir rol oynamaktadır. Yüksek reaktiviteye sahip olan hidroksil radikalleri
protein ve lipitlerde olduğu gibi DNA bazlarında bulunan çift bağlara H atomu katarak veya timin
yapısındaki metil gruplarından ve 2-deoksiribozun C-H bağlarından H atomu kopararak DNA
molekülü ile reaksiyon verir [56]. Sonuçta oluşan timin peroksil radikalleri indirgenir ve
hidroksihidroperoksit, timin glikol, 5-hidroksimetilurasil, 5-formilurasil ve 5-hidroksi 5-
metilhidantoin gibi oksidasyon ürünleri bu reaksiyon sonucunda oluşur. 8-OHdG, DNA baz
mutasyonları arasında en yaygın bilinenidir. Hidroksil radikalleri (OH•), guanin molekülünde 8.
pozisyonundan etkileşerek oksidasyon reaksiyonu verir. Yapısal olarak farklılaşmanın gerçekleştği
DNA’da meydana gelen oksidatif hasarın neticesinde 8-OHdG (8-hidroksi-2′-deoksiguanozin)

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
10
oluşur. Bunun yanında Cu⁺² iyonları özellikle guanin bazları olmak üzere DNA’ya yüksek afinite ile
bağlanır ve H2O2 ile reaksiyon vererek DNA hasarına yol açarlar. Oksidatif olarak değişikliğe
uğrayarak 8-OHdG formuna gelen DNA, DNA hasarının belirlenmesinde önemli bir ölçüttür [57].
Oksidatif stres DNA yapısı üzerinde bir başka etki olarak, baz radikalleri proteinlerin aromatik
aminoasitleri ile beraber çapraz “DNA-protein” bağları oluşturur [58].
Buna ek olarak, hidroksil radikalleri DNA üzerinde bulunan şeker kalıntılarından H atomu
kopararak şeker üzerinde modifikasyonlara ve zincir kırılma reaksiyonlarına da yol açar. Sonuç
olarak hücrelerin oksidan özellikteki serbest radikallere maruz kalması replikasyon ve
transkripsiyon üzerine etkili olup aynı zamanda DNA tamir mekanizmalarını baskılayarak DNA
hasarına neden olur [59]. DNA üzerine oksidatif hasarın göstergesi yükselmiş 8-OH-dG (8-
hidroksiguanin) düzeyleri olarak kabul edilmiştir. 8-OH-dG mutajenik özelliğe sahip olup idrar,
doku ve plazmada biyokimyasal belirteç olarak kullanılabilinmektedir [60].
2.4. Oksidatif Hasar
Oksidan ve antioksidanlar arasındaki dengenin oksidanlar lehine bozulması ile
organizmada hücresel hasarın oluşmasına oksidatif hasar denir. Organizmada oksidatif stres
karşısında antioksidan sistem yetersiz kalırsa yaşlanma, kanser, sepsis, dejeneratif nörolojik
hastalıklar, kardiyovasküler hastalıklar gibi durumların ortaya çıkması söz konusudur [61, 62].
Reaktif oksijen ve reakstif nitrojen türleri normal hücre metabolizması esnasında oluşur.
Canlı organizmada düşük konsatrasyonda yararlı etkileri vardır. Serbest radikaller biyolojik
sistemlerde faydalı oldukları miktarı aşınca bulundukları ortamda lipit, protein ve nükleik asitler
gibi moleküllerle etkileşirler. Bu etikileşim sonucu hücrede yapısal ve işlevsel bozukluklara sebep
olurlar [63]. Serbest radikaller enzim aktivite bozukluklarına, geri dönüşümsüz DNA hasarlarına,
proteinlerin zarar görmesine, lipit peroksidasyonuna neden olarak zar yapısında bozukluklara
neden olmaktadır [64, 65].
Reaktif oksijen türleri, hücre membran yapısında bulunan yağ asit zincirinden bir
hidrojen atomu kopararak membran lipit yapısını değiştiren, bir dizi reaksiyonu içerisinde
barındıran lipit peroksidasyon reaksiyonunu başlatır [64, 54]. Bu reaksiyon engellenmezse hücre
membran harabiyeti ve akabinde hücre ölümü gerçekleşir [64, 65].
Biyolojik sistemlerde serbest radikal üretiminin artması, antioksidankarın miktarsal ya
da işlevsel olarak yetersiz olması oksidan ve antioksidan arasıdaki dengenin bozulmasına ve
oksidatif stres oluşumuna neden olur. Organizmada oksidan ve antioksidan ararasındaki dengenin

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
11
bozulması serbest radikallerin doku ve hücrelerde birikmesine ve işlevsel/yapısal bozukluklara
neden olur. Bu zararlı etkilerden organizmayı koruyabilmek için serbest radikal oluşumunu
engelleyen, oluşan bu serbest radikallerin zararlarını azaltan bazı antioksidan savunma sistemleri
geliştirilmiştir [66]. Oksidan maddelerle karşılaşan antioksidanlar hedef molekülün okside
olmasına engel olurlar ya da okside olmasını geciktirirler [65]. Antioksidan enzimler bu görevi
serbest radikalleri indirgeyerek yapar. Antioksidanlar, serbest radikallerin aktivitelerini azaltarak,
serbest radikallerin sebep oldukları reaksiyonları kırarak ve onarım mekanizmasını kullanarak
oksidatif stres oluşumuna engel olurlar [67].
2.5. Antioksidan Savunma Sistemleri
Biyolojik sistemlerde reaktif oksijen türleri ile bu maddelerin sebep olduğu hasarı
önlemek ve detoksifikasyon ile ortamdan uzaklaştırılmalarını sağlamak üzere görev yapan
sistemlere antioksidan savunma sistemleri denir. Antioksidan savunma sistemlerinde görev alan
maddelere antioksidanlar denir [68]. Antioksidan maddeler serbest radikallerle hızlı bir şekilde
rekasiyon vererek otooksidasyon ve peroksidasyona engel olur. Antioksidanlar serbest
radikallerin zararlı etkilerinden hücreleri koruyarak hastalıkları önlemede aktif rol alır [69, 70].
2.5.1.Enzimatik Antioksidanlar
2.5.1.1. Süperoksit Dismutaz
Biyolojik sistemlerde reaktif oksijen türlerine karşı ilk savunma hattı süperoksit dismutaz
enzimi aracılığı ile olur [71-73]. Süperoksit dismutaz enzimi, süperoksit radikalinin moleküler
oksijen ve hidrojen peroksite dönüşmesi reaksiyonunu katalizasyonunda görev alan bir
antioksidandır. Oluşan hidrojen peroksit radikali katalaz vaya glutatyon peroksidaz enzimleri ile
detoksifiye edilir [74].
İnsanlarda SOD enzimi üç faklı formda bulunur. Sitozolde bakır (Cu) ve çinko (Zn) içeren
süperoksit dismutaz (Cu/Zn SOD), mitokondride manganez (Mn) içeren süperoksit dismutaz (Mn
SOD), hücre dışı sıvılarda ekstrasellüler süperoksit dismutaz (EC SOD) enzimi bulunur [75, 76].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
12
Sitozolik Cu/Zn SOD enzimi 32 kD ağırlığındadır ve birbirine eş iki alt üniteden meydana
gelir. Bu alt ünitelerin biri Cu, diğeri Zn içerir. Biyolojik sistemde en çok bulunan süperoksit
dismutaz formudur [77, 78].
Mitokondride bulunan Mn SOD formu dört eşit alt üniteden oluşan, 80 kD ağırlığında bir
enzimdir. Enzimin aktif bölgesi Mn⁺³ içerir. Sitozolik Cu/Zn SOD enziminden farlı özelliklere de
sahip olduğu halde aynı reaksiyonları katalizlemektedir [79, 80].
Ekstrasellüler süperoksit dismutaz (EC SOD) canlı organizmada çoğunlukla
homotetramer formda bulunan 135.000 kD ağırlığında bir enzimdir. EC SOD enzimi her bir alt
ünitesinde Cu ve Zn atomlarını bulundurur. Bu iki atom enzim aktivitesi için zaruridir. EC SOD, glia
hücreleri, fibroblastlar, endotel hücreler tarafında salgılanıp salınmakla birlikte biyolojik
sistemlerde en fazla hücre yüzeylerinde ve ekstrasellüler matrikste bulunur. Özellikle akciğer
dokusunun tip II epitel hücreleri ile solunum yoları ve kan damarlarını saran düz kas hücrelerinde
yüksek konsatrasyonda bulunur. Süperoksit radikalini ekstrasellüler düzeyde etkisizleştirebilen
tek antioksidan olması bakımından ciddi akciğer rahatsızlıklarından korunmada önemli bir role
sahiptir [81].
2.5.1.2. Katalaz
Katalaz enzimi, her alt birimi bir hem, bir NADPH molekülü içeren dört alt birimden
meydana gelen bir enzimdir [75, 82]. NADPH, molekülü katalaz enizminde genelde yüzeye yakın
ve sıkı bir bağile bağlı olarak bulunur [83]. Hücrede çoğunlukla peroksizomlarda bulunur. Katalaz
enzimi hidrojen peroksitin su ve moleküler oksijene dönüşmesini katalizyelen enzimdir [82].
2.5.1.3. Glutatyon Peroksidaz
Hidrojen peroksitin sebep olabileceği oksidatif hasara karşı koruyucudur. Dolaylı yoldan
hidrojen peroksitten çok tehlikeli bir radikal olan hidroksil radikalin oluşumuna engel olmuş olur.
Dört alt birimden oluşur. Her bir alt birimi bir selenyum içerir [74].
Elektron kaynağı glutatyondur. Hidrojen peroksiti ve organik hidroperoksitleri
detoksifiye eden reksiyonları katalizler. Glutatyon peroksidaz enzimi, biri selenyum içeren
selenyuma bağımlı glutatyon peroksidaz ve selenyum içermeyen yani selenyuma bağımlı olmayan

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
13
glutatyon peroksidaz olmak üzere iki tipten oluşur. Selenyum bağlı glutatyon peroksidaz (Se-GPx)
hidrojen peroksit ve organik hidroperoksitlere karşı etikilidir. Selenyum bağımsız gulutatyon
peroksidaz ise organik hidroperoksitler üzerinde etkilidir.
Bu detoksifikasyon reaksiyonları sırasında GSH’ın elektron verici olarak görev alması
hidrojen peroksit ve organik hidroperoksitleri indirgerken kendisinin yükseltgenmesine neden
olur [84] Glutatyonun (GSH) oksitlenmesi ile glutatyon disülfit oluşur (GSSH). Okside glutatyonun
indirgenmesi için glutatyon redüktaz enzimi gereklidir. Bu indirgenme reaksiyonu sırasında
elektron vericisi olma görevini NADPH üstlenir [74, 83].
2.5.1.4. Glutatyon Redüktaz
Yapısında flavo adenin dinükleotit (FAD) bulunduran flavoprotein yapıda bir enzimdir.
Glutatyon redüktaz, NADPH’ı kullanarak okside glutatyonu disülfit bağlarına elektron transferi
yapar ve glutatyonu tekrardan redükte forma getirir. Bu redüksiyon reaksiyonu için NADPH çok
önemlidir. Serbest radikal hasarını önlemek için ortamda bulunması şarttır. En önemli sentez yolu
da pentoz fosfat yolağıdır [84, 85].
2GSSG + NADPH + H⁺ GR 2GSH + NADP⁺
2.5.1.5. Glutatyon-S-Transferaz
Biyolojik sistemlere dışarıdan girmiş ksenobiyotiklerin biyotransformasyonunda görev
alan bir enzimdir. Lipit peroksitlerin çok daha az toksik olan yağ asitlerine dönüştürülmesi
reaksiyonlarını katalizler [86].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
14
2.5.2 Nonenzimatik Antioksidanlar
2.5.2.1. Glutatyon
Glutatyon, bütün ökaryötik hücrelerde sentezlendiği için biyolojik sistemlerde yüksek
konsatrasyonda bulunan bir moleküldür. Hücrelerin redoks durumunu muhafaza etmede,
detoksifikasyon reaksiyonlarının yürümesinde, apoptoziste, gen ekspresyonunda görev alarak
antioksidan rolü üstlenir [87]. GSH, yüksek oranda stoplazmada bulunmasına rağmen bazen sentez
edildikten sonra mitokondri, peroksizomlar ve çekirdeğe geçebilir [88].
Glutatyon, aşamalı olarak ilerleyen iki reaksiyon sonucunda sentez edilir. İlk reaksiyonda,
glutamil ve sistein, glutamil-sistein ligaz enzimi ile bağlanarak ɣ-glutamilsistein oluşturulur. İkinci
reaksiyonda, ɣ-glutamilsisteine glisin molekülü, glutatyon sentetaz (GSS) enzimi kullanılarak
bağlanır ve GSH sentez edilir. Glutamil-sistein ligaz enzimi, katalitik (GCLC) ve düzenleyici (GCLM)
olmak üzere iki alt birimden meydana gelir. Katalitik alt birim glutamil ve sisteinin
bağlanmasından sorumludur. Düzenleyici alt birim ise katalitik alt birimin etkisini arttırır [89, 90].
Glutamil + Sistein GCL ɣ-glutamilsistein
ɣ-glutamilsistein +Glisin GSS Glutatyon
Glutatyon, lipit peroksitlerin ve hidrojen peroksitin detoksifikasyonunu sağlar ve
hidroksil ile singlet oksijen radikalini süpürücü etkiye sahiptir. Bunun yanında C ve E vitamininin
antioksidan aktivitelerini düzenler [84].
2.5.2.2. Vitamin E
Vitamin E, α, β, ɣ, δ tokotrienol ve α, β, ɣ, δ tokoferol olmak üzere sekiz farklı stereoizomeri
olan, yağda çözülen ve yüksek antioksidan aktivite gösteren bir antioksidandır. İnsandaki aktif
formu α tokoferoldür. Lipit peroksidasyonuna karşı hücre membranını koruyucu olarak görev
yapar [70]. Vitamin E, reaktif oksijen türlerinin zincir reaksiyonlarını kırması ve onları yok
etmesinin yanında endojen savunma sistemlerini kuvvetlendirip bozulan yapıları onararak
antioksidan aktivitesini gerçekleştiren güçlü bir vitamindir. Glutatyon peroksidaz enzimi ve α
tokoferol antioksidan aktivite bakımından bribirlerini tamamlayıcı ve destekleyici niteliktedirler.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
15
α tokoferol peroksit oluşumunu engelleyici non enzimatik bir antioksidan iken, glutatyon
peroksidaz oluşan peroksitleri yok eden enzimatik bir antioksidandır [91, 92].
2.5.2.3. Vitamin C
Vitamin C, kollojen, karnitin ve nörotransmitter sentezinde görev alan, bir diğer ismi de
askorbik asit olan, suda çözünebilen bir vitamindir [93]. Biyolojik sistemlerde meydana gelen
serbest radikalleri süpürücü özelliği yüksek olan bir antioksidandır, hücre membran lipitlerinde
meydana gelen serbest radikallerin yok edilmesi reaksiyonları ile oluşan α tokoferoksil
radikalinden α tokoferol oluşturma yetisine sahip bir antioksidandır. Bu özelliği vitamin C’yi
koantioksidan bir vitamin yapar [94].
2.5.2.4. Bilirubin
Bilirubin, eritrositlerin yapısında bulunan hem molekülünün yıkılması ile oluşur.
Vücüttaki dolaşımı karaciğer tarafından alınıp metabolize edilmesi ile son bulur. Safra veya idrar
yolu atılır. Bilirubin, peroksil radikallerinde zincir kırıcı reaksiyonlara sebep olarak antioksidan
alktivite gösterir [95, 96].
2.5.2.5 β-Karoten
Aktif A vitaminine dönüşebilen bir provitamin olan β-karoten, karotenoidlerin yağda
çözünen bir formudur. Retinada, karanlıkta görmek için gerekli olan retinole dönüşür. Güçlü bir
siglet oksijen temizleyicisi olması bakımından önemli bir antioksidandır [70].
2.5.2.6. Koenzim Q10
Koenzim Q10, oksijenli solunum ile enerji üretiminde oldukça öenmli olan, insan
vücudunda doğal olarak sentezlenen vitamin benzeri bir benzokinondur. İnsanlarda, tirozinden
koenzim Q10 sentez edilir. Yağda çözünürlüğü yüksek olduğu için hücre membran yapısıda
bulunur. Hücre mitokondrisinde bulunan bazı enzimler için kofaktör olarak görev yapar ve
oksidatif fosfosrilasyonda rol alır.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
16
Koenzim Q10, biyolojik sistemlerdeki serbest radikalleri temizler, lipit ve proteinlerin
peroksidasyon reaksiyonlarına engel olur. Koenzim Q10’in indirgenmesi ile lipofilik yapıdaki
ubikinol oluşur. Bu lipofilik antioksidan elektron taşıma sisteminde elektron ve proton
taşınmasından sorumludur. Ubikinol antioksidan aktivitesini oksidanlara elektron vererek
gerçekleştirir. Oldukça güçlü bir antioksidan olma özelliğine sahiptir. En az vitamin E kadar lipit
peroksidasyonu önleyicisidir. Vitamin C’ye benzer bir mekanizma ile antioksidan aktivitesini
gösterir [97].
2.5.3.7. α-Lipoik Asit
Hidroksil, hipokloröz asit, singlet oksijeni ve peroksinitrit anyonunu yok ederek
antioksidan aktivite gösterir. α-Lipoik asit’in indirgenmiş formu olan dihidro lipoik asitte (DHLA)
antioksidan aktivite gösterir. Dihidrolipoik asit süperoksit ve peroksit radikalleri üzerinde de etkili
bir antioksidandır [98].
2.5.3.8. Selenyum
Selenyum, aminoasit sentezi için kullanılan, selenoprotein fonksiyonu için önemli bir
elementtir. Antioksidan ve bağışılık sistemlerinde düzenleyici bir rol üstlenir. GPx enzim
aktivtesini arttırır. Böylece reaktif oksijen türlerinin oluşumunu önler [99].
2.5.4.9. Folik asit
Folik asit, DNA sentezi ve kırmızı kan hücresi üretiminde görev alan, suda çözünebilir
yapıda bir B vitaminidir. Fertilitede, hamilelelik ve çocukluk dönemlerinde hücrelerin büyüme ve
çoğalmasında gerekli olan önemli bir moleküldür. Reaktif oksijen radikallerini süpürücü özellikte
olması bakımında oldukça güçlü bir antioksidandır [100, 101].
2.5.5.10. Flavonoidler
Flavanoidler, polifenolik yaıda ve fenolik yapıda olmaları bakımında yüksek antioksidan
aktivite gösteren moleküllerdir. İnsanlar diyet yolu ile organizmaya alırken bitki metabolitlerinde
yüksek oranda flavanoitlere rastlamak mümkündür. Serbest radikal süpürücü özelliğe sahiptir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
17
Lipit peroksidasyonunu katalizleyen metal iyonlar ile şelat oluşturarak lipit peroksidasyonuna
engel olur [102].
2.6. Punicalagin
Antioksidanlar insan diyetinin temel bileşenleridir ve son zamanlarda vitaminler ile bitki
polifenollerinden oluşan antioksidan içerik bakımından zengin doğal kaynakların kullanımı
yaygınlaşmıştır. Doğal antioksidanların serbest radikallere karşı koruma ve kronik hastalıkları
önlemedeki gücünün sentetik antioksidanlardan çok daha fazla olduğu kanıtlanmıştır [103].
İnsan sağlığı durumunun bitki polifenolleri ile kontrol edilebildiği gözlenmiştir [104] Nar
meyvesi flavoidler, antasiyoninler ve hidrolize edilebilir tanenlerden olan punicalagince oldukça
zengin bir fenolik içeriğe sahiptir. Narın antioksidan içeriğinin %92’si hidrolize edilebilir
tanenlerden gelir [105]. Bu hidroliz ebilebilir tanenler arasında punicalagin en önemli antioksidan
maddedir [106]. Punicalagin (PG), nar kabuğunun asıl bileşenidir [107-109]. Punicalaginin
antioksidan, antiinflamatuar, antikanserojen ve antiaterosklerotik özellikte olduğu kanıtlanmıştır.
Punicalagin ve diğer nar polifenolleri, reaktif oksijen türlerinin vücut üzerindeki zararlı etkilerini
kısıtlayabilmektedir [108, 109]. Ayrıca lipit peroksidasyonu üzerinde ciddi antioksidan aktivite
gösterdiği bilinmektedir [110].
Narda bulunan polifenolik bileşikler TNF-α, IL-8, IL-6, IL-2, IL-1 gibi sitokinlerin
transkripsiyonunu düzenler [111, 112]. Proinflamatuvar sitokinlerden olan IL-6 ve IL-8’in
üretimini azaltır [113].
Antiinflamatuvar, antioksidan ve hemostaz ajanı olarak kullanılan nar kabuğu fenolik
bileşiklerce zengindir [114]. Nar kabukları içinde en bol bulunan polifenoller, güçlü asitlere veya
mikroorganizmalara maruz kaldıklarında ellagik asite (EA) hidrolize edilebilen poliollere bağlı
heksahidroksidifenik asit (HHDP) esterleri olan ellagitanninlerdir. Punicalagin, nar kabuğundaki
başlıca ellagitanindir ve içeriği, nar kabuğundaki toplam polifenollerin %65,75'ine ulaşabilir [115].
Bu polifenollerin en fazla konsantrasyonda bulunanı punicalagindir [116]. Punicalagin, gallik asit
ve ellagik asite bağlı bir glukoz molekülünü ihtiva eden ellagitanin yapıda bir moleküldür ve şekil
2.6.1. ile kimyasal yapısı gösterilmiştir. Punicalagin, narın yüksek antioksidan kapasitesinden asıl
sorumlu olan moleküldür.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
18
Şekil 2.2. Punicalagin’in Kimyasal Yapısı [117].
Lipitleri, aminoasitleri ve guanozini oksidatif hasara karşı koruyucu özelliktedir.
Punicalaginin güçlü biyoaktivitesi, in vivo ve in vitro olarak mitokondrial zar boyunca ellagik asite
hidrolize olması ile ilgilidir [116]. Punicalagin, bir HHDP bırakarak başka bir ellagitanin olan
punikaline (PL) hidrolize edilebilir ve hem punicalagin hem de punicallin, belirli koşullar altında
ellagik asite hidrolize edilebilir [118]. Ellagik asit yüksek antioksidan, antikanserojen ve
antiaterosklerotik kapasiteye sahip bir moleküldür. İnsan plazmasındaki serbest ellagik asit,
punicalaginler gibi ellagitanenlerin hidrolize olup salınmasından kaynak alır.
Şekil 2.3. Punicalagin’in Hidroliz Reaksiyonu [119].
2.7. Sitokinler
Sitokinler ortalama 20–30 kD ağırlığına sahiptirler. Glikoprotein veya peptit yapıya sahip,
immun veya inflamatuvar yanıta katılan hücrelerin etkinliklerini arttırmak için; uyarılmış
lenfositler, makrofajlar ve monositler ile diğer bazı somatik hücrelerce sentezlenen maddelerdir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
19
Düşük molar konsantrasyonlarda bile aktif oldukları gibi çözünebilir formda etkilerini gösterirler.
Sitokinler çok çeşitli hücrelerce üretilebilen polipeptit yapısında maddelerdir. Hücre büyümesi,
enflamasyon, iyileşmesi ve yaralanmaya karşı canlı organizmanın cevabını kapsayan inflamasyon
ve bağışıklık olaylarının düzenlenmesinde rol alırlar. Sitokinler, hormona benzemektedir fakat tam
olarak hormon olarak adlandırılamazlar [120].
2.7.1. Sitokinlerin Genel Özellikleri
Sitokinler bağışıklık, inflamasyon ve hematopoeze aracılık eden ve bu mekanizmaları
düzenleyen eser miktarda salınan protein yapıda maddelerdir. Hücrelerin uyarılması ile kısa
sürede salgılanırlar. Depo edilmezler. Uyarana tepki olarak de novo olarak üretilirler. Genelde kısa
mesafelerde etkilidirler. Kısa zaman aralıklarında ve düşük yoğunlukta hareket ederler. Etkilerini
göstermek için ikinci habercileri kullanırlar. Bu amaçla çoğunlukla tirozin kinazlar kullanılır.
Böylece spesifik membran reseptörlerine bağlanarak hareket ederler. Sitokinlerin salındıkları
ortamda tepkiler, zar proteinlerinin (sitokin reseptörleri dahil) artması veya azalmasına bağlı
olarak efektör moleküllerin salınmasını içerir. Çok çeşitli hücrelerce üretilebilirler. Buna bağlı
olarak çok çeşitli hücrelerde etki gösterebilme yeteneğine sahiptirler. Benzer etkiler ve aralarında
agonist-antagonist etki gösterebilirler. Stokinler polipeptid hormonlardakine benzer şekilde hedef
hücrelerdeki özel reseptörlere bağlanarak etkilerini gösterirler [121].
2.7.2. Sitokinlerin Organizmadaki Etkileri
Biyolojik sistemlerde endotelyal etkileri, fibroblastik etkileri ve sistemik akut faz
reaksiyonlarını tetikleyici etkileri vardır [122, 123].
2.7.3. Sitokinlerin İşlevlerine Göre Sınıflandırılması
2.7.3.1. Doğal İmmüniteye Aracılık Eden Sitokinler
Bu grup sitokinler enflamatuvar reaksiyonları başlatan sitokinlerdir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
20
2.7.3.1.1. Tip I İnterferonlar (IFN)
İki farklı grup proteinleri içerisinde barındırır. Birinci grup; her biri ayrı gen ile kodlanmış
olan, birbiriyle ilişkili 20 aa grubundan oluşan 18 kD’luk IFN-α olarak adlandırılır. IFN-α grup
içerindeki aa sıralamasının farklılığından dolayı IFN-α1 ve IFN-α2 olarak da kendi içerinde ikiye
ayrılır [120, 124]. Lökosit interferon olarak da bilinen IFN-α’ nın asıl üretim yeri mononükleer
fagositlerdir [125]. İkinci grup 20 kD’luk bir protein olan ve tek gen ürününden oluşan IFN-β’dır.
Fibroblast interferon olarak da adlandırılan IFN-β’ nın üretim yeri fibroblastlardır. Antijenlere
karşı bağışık yanıtın verilmesinde birçok hücre tarafında üretilen IFN-α ve IFN-β görev alırlar. Her
ikisi de aynı hücre yüzey reseptörlerine bağlanıp birbirine benzeyen hücre yanıtlarını oluştururlar.
Tip I İnterferonların canlı sisteminde bilinen dört temel etkisi vardır:
1. Viral RNA veya DNA replikasyonunu engelleyen 2-5 oligoadenilat sentetaz gibi enzimlerin
sentezini uyararak viral replikasyonu inhibe ederek antiviral etkisini gösterir.
2. Viral replikasyonu inbibe eden enzimlere bağlı olarak ve özellikle triptofan gibi esansiyel aa’
lerin sentezini önleyerek hücre proliferasyonunu önler.
3. Virüsle enfekte olan hücreleri öldürmekle görevli olan naturel killer hücrelerin (NK) litik
potansiyelini arttırır.
4. Class I MHC molekül ekspresyonunu arttırırken, Class II MHC moleküllerinin ekspresyonunu
azaltır.
Tip I interferonlar bu etkilerinin hepsini virüsleri yok etmek için yapar [126].
2.7.3.1.2. Tümör Nekrozis Faktör (TNF)
Gram negatif bakteriler ve diğer infeksiyöz organizmalara karşı yanıtta görev alır.
Lipopolisakkaritlerce aktive olan mononükleer fagositler tarafından üretilirler. Aktive NK
hücreleri, T lenfosit hücreleri, aktive mast hücreleri bu proteinin üretiminde rol alır. İnsan TNF’ si
17 kD ağırlığında olup nonglikolize bir transmembran proteinidir. Sitotoksik aktiviteyi, TNF
reseptörleri ve fibroblast proliferasyonunu arttıran TNFR Tip I ve T lenfosit proliferasyonuna
neden olan TNFR-Tip II olarak kendi içinde ikiye ayrılırlar [120, 126-128].
TNF’ nin düşük yoğunluktaki (10⁻⁹ M) biyolojik etkileri şunlardır:

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
21
1. Lökositlere karşı damar endotel hücrelerinin yeni yüzey reseptörlerini ekspresse etmelerine
neden olur. Bunu da endotel hücre yüzeyini adhezyon molekülleri (ICAM-I, NCAM-I, ELAM-I gibi)
aracılığı ile daha yapışkan hale getirerek yapar.
2. TNF, inflamatuar lökositleri mikroplar öldürecek ekilde aktive eder. Özellikle nötrofilleri aktive
ederek mikropları yok eder.
3. TNF, mononükler fagositleri ve diğer hücre tiplerini uyararak IL-1, IL-6, kemokinler ve TNF’nin
kendisini üretimini sağlar.
4. TNF, interferon benzeri koruyucu etkisi bakımından viruslara karşı koruyucu özellik gösterir.
TNF’nin düşük yoğunluktaki bu özelliklerinin tamamı mikroplara karşı verilen
inflamatuvar cevapta oldukça önemlidir.
TNFnin infeksiyonlara karşı temel sistemik etkileri şunlardır:
1. TNF, IL-1 ile birlikte endojen pirojen olarak etki ederek ateşi yükseltir. Ateşin yükselmesini
hipotalamusun uyarılıp prostaglandin E2 sentezine öncülük etmesi ile sağlar. 2. TNF, IL-1 ve IL-
6’nın dolaşıma salınmasını sağlar. Bu etkiyi mononükleer fagositler ve vasküler endotel hücreleri
uyararak yapar.
3. TNF, hepatositlere etki ederek serum amyloid A ve P proteini, kompleman faktör 3-
haptoglobulin, creaktif protein, a1-asid glikoprotein, Faktör B gibi bazı akut faz proteinlerinin
sentezini uyarır [127, 129].
4. TNF, pıhtlaşma sistemini aktive edilmesinde görev alır. Bu olayı damar endotel hücrelerinde
antikoagülan ve prokoagülan dengeyi değiştirerek yapar.
5. TNF, kemik iliğinde hücre bölünmesini baskılar. Bu sebeple uzun vadade immün yetmezliğe ve
lenfopeniye sebep olur.
6. TNF’nin uzun süre verilmesi kaşektik metabolik bozulmalara sebep olur. Bu kaşkesi durumu TNF
ile uyarılan organizmada iştah azalması ile birlikte gelişir.
Biyolojik sistemlerde TNF, gram negatif bakterilerin neden olduğu sepsiste oldukça
yüksek konsatrasyonda üretilir. Serumda artan TNF yoğunluğu dolaşımda dissemine intravasküler
koagulasyon (DIC) ile kollapsa sebep olur. Bu özelliği bakımından TNF endotoksik ve septik şokun
önemli bir mediatörüdür.
TNF’nin yüksek konsantrasyonda ki etkileri şunlardır:

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
22
1. TNF kardiyak myositteki nitrik oksit sentaz (NOS) enziminin sentezini uyararak sitrülin, arjinin,
ve NO’ya dönüşümünü sağlar. Oluşan NO myokardın kasılabilme özelliğini azaltır. Bu etkisi ile TNF
doku perfüzyonunu azaltmış olur.
2. TNF, damar endotel hücreleri tarafından salgılanan NO ve prostasiklin gibi damar genişleticileri
uyararak ve düz kas hücrelerine etki ederek kan basıncı ve doku perfüzyonunu azaltır.
3. TNF, doku perfüzyonunu azaltır. Bunu intravasküler koagülasyona sebep olarak yapar.
4. TNF canlı organizmada ağır metabolik bozukluklara sebep olabilir. Yaşamı sonlandıracak
boyutta düşük kan glukoz düzeyinin, glukozun kas tarafından aşırı kullanımı ve karaciğer
tarafından glukozun yerine koyulamamasına bağlı olarak gelişmesi bu duruma örnek olarak
verilebilir.
IFN-ϒ, TNF reseptör sayısını artırır. Bu durum neticesinde TNF’nin birçok fizyolojik etkisi
IFN- ϒ tarafından arttırılmış olur [120, 124, 126, 129].
Aktif makrofajlardan salnan TNF-α (kaşektin) ile aktif T hücrelerinden salınan TNF-β
(Lenfotoksin) olmak üzere iki çeşit TNF vardır.
2.7.3.1.2.1. TNF-α
TNF-α, pro-enflamatuvar etkinlik, proliferasyon, apoptoz, morfogenetik değişiklikler ve
farklılaşma gibi işlevlerde aktif rol alır [130]. TNF-α biyolojik sistemde iki formda bulunur. Bunlar,
membrana yerleşik protein (transmembran) ve çözünür sitokindir. TNF-α, TNFR-I ve TNFR-II
olmak üzere iki adet TNF reseptörüne sahiptir. Vücutta en fazla TNFR-I reseptörü bulunur, TNFR-
II reseptörü ise sinir sitemi ve bağışıklık sistemi hücrelerinde bulunur [131, 132]. Memebrana
yerleşik halde bulunan TNF-α hedef hücre reseptörleri aracılığı ile hücre tipine özel olarak hücresel
temas yöntem ile bölgesel enflamasyonun düzenlenmesinde rol alır. Membrana yerleşik TNF-α’nın
farklı hücre ekspresyonları patolojik ve fziyolojik süreçlerde aktif görev alır.
HIV, leishmania, mycobacterium tuberculosis ve listeria monocytogenes gibi çeşitli
enfeksiyonlara karşı canlı organizmanın savunmasında görev alır. Bu özelliği bağışık sistemin
verdiği yanıtta aktif rol aldığının bir örneğidir [132, 133]. Özellikle tüberkülozda (TB) akut
enfeksiyona karşı etkililik gösterir. Bunu T-hücre aracılığı ile makrofaj göçü ve granülom
oluşumunu aktive ederek yapar [134]. Transmembran formdan TNF-α dönüştürücü enzimin
katalizlediği reaksiyonlar sonucu çözünür formdaki TNF-α proteolitik olarak ayrılır. Akabinde pro-
enflamatuvar hücre proliferasyonu ile apoptoz gibi etkiler yapar. TNF-α’nın bu formu,

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
23
lipopolisakkaritlere, IL-1’e ve diğer bakteriyel ürünlere cevap olarak üretilir [135]. TNF-α’nın
çözünür formu özellikle otoimmün hastalıklar, nörolojik hastalıklar, kardiyovasküler hastalıklar,
kemik hastalıkları, metabolik hastalıklar, kanser gibi hastalıklara karşı patolojik cevabın
oluşmasına görev alır [131]. TNF-α, otoimmün hastalıkların patogenezinde oldukça kritik ve
önemli bir görev alır. Crohn hastalığı, artrit, psöriazis, multipl skleroz, üveit ve sistemik lupus gibi
otoimmün hastalıkların patogenezinde başta TNF-α olmak üzere, TNF süper-ailesi/reseptörleri alt
tipleri aktif olarak görev alır [131, 136-138]. TNF aktivitesinin seviyesi, patolojik süreçte oluşum
zamanı ve aktif görev yaptığı süre bağışıklık fizyolojisi için oldukça önemlidir. Bağışıklık fizyolojisi
için önemli olan bu etkenlerin aralarındaki dengenin bozulması TNF-reseptörü aracılı aktivitelerin
pro-enflamatuvar, bağışıklık baskılayıcı veya bağışıklık aktive edici olmasına bağlı olarak biyolojik
sistemlerde farklı patolojilerin meydana gelmesine sebep olmaktadır [138].
2.7.3.1.2.2. TNF-β
Aynı TNF-α gibi protein yapıdadır. TNF-β glikoprotein yapısındadır. Ortalam 26 kD
ağırlığında olan bu glıkoprotein bir prohormon şeklinde hücre membranına entegre
olabilmektedir. TNF-β’nın 34 aa’lik sinyal dizileri karboksi uçlara dışa gelecek şekilde
ekstrasellüler alanda uygun konumu alır [139, 140]. TNF-α ve TNF-β’nın aminoasit benzerliği %35
dolaylarında olmasına rağmen her iki formun da etki mekanizmaları, reseptörleri ve etki
yelpazeleri benzerdir [141, 142].
2.7.3.1.2. İnterlökin-1 (IL-1)
İnterlökin-1 aktive mononükleer fagositlerden türeyen bir polipeptiddir. TNF’ye benzer
şekilde IL-1 de hormon gibi gram negatif bakterilerin neden olduğu sepsis sonrasında canlı
organizmanın dolaşımında görülür. IL-1 iki farlı genin ürünü olan IL-1α ve IL-1β olarak ikiye ayrılır.
Bu iki formda aynı hücre yüzey reseptörüne bağlanarak etki eder. Biyolojik etki bakımından da
aynı oldukları söylenebilir. IL-1’in bir üçündü üyesi IL-1reseptör antagonistidir. IL-1’in yapısal
olarak fibroblast büyüme faktörü ile de benzerliği vardır. Canlı organizmada IL-1 aktivitesinin
çoğunu IL-1β üstlenir. IL-1 için immun globulin üst ailesine ait olan iki farklı reseptör belirlenmiştir
[120, 127, 129, 143]. IL-1 biyolojik etki bakımında TNF’ye benzer özelliktedir. Düşük
konsatrasyonda bölgesel inflamasyon olaylarına aracılık eder. IL-1 endotel hücrelere ve
mononükleer fagositler etki ederek sentezini etkiler ve arttırır. TNF’ ye ait birçok inflamatuvar
özelliğe de sahiptir. Endotel hücrelere etkisiyle pıhtılaşmayı arttırması buna örnek olarak

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
24
verileblir. IL-1 nötrofiller gibi inflamatuvar fagositleri etileyemezler, bunu mononükleer fagositler
ve endotel hücrelere etki ederek lökositleri aktive eden kemokin sentezini uyararak yapar.
IL-1, yüksek konsantrasyonda salgılanıp kan dolaşımına girdiğinde endokrin etkiler
gösterir. Dolaşıma girmiş IL-1, TNF ile birlikte ateşin oluşumuna aracılık eder [127, 129].
IL-1, hipotalamusa etki ederek kortikotropin serbestleştirici faktörün salgılanmasını
sağlar. Salgılanan bu CRF’de adrenal kortekse etki ederek steroid salınımını uyarır. Oluşan
kortikosteroitler de geri beslenim mekanizmasına uygun olarak TNF ve IL-1 oluşumunu inhibe
eder.
IL-1, kollojen dokuya etki ederek osteklastik aktiviteyi artırır ve böylece kemik
döngüsünün artmasını sağlar. Osteoblastlarda ise alkalen fosfataz aktivitesinde artışa neden olur.
Sinovial hücrelerin ve fibroblastların çoğalmasını sağlar. Kemik iliğinde homopoetik hücrelere etki
ederek, TNF ile beraber nötrofiliye sebep olur.
IL-1, doğal olarak var olan bir inhibitördür. Biyolojik olarak inaktiftirler fakat kompetetif
olarak IL-1’i engelleyici özellikleri vardır. Bu etkileri sebebiyle IL-1 reseptör anatagonisti olarak
adlandırılırlar. Sitokin inhibitörleri, sitokinlerin aşırı ve düzensiz üretildiği durumlarda (septik
şok) biyolojik yanıt düzenleyici olrak görev yaparlar.
TNF ve IL-1 mazenşiyal ve glomerüler endotel hücreleri tarafından üretilir. Oluşan TNF ve
IL-1 otokrin ve parakrin etkiyle glomerüler ve mezenşial hücrelerde Tip IV kollojenaz ve
prostaglandin E2 üretimini sağlar. TNF ve IL-1 mezenşial hücrelerde IL-8, IL-6 ve kollojen
oluşumunu uyarır [120, 144].
2.7.3.1.3. İnterlökin-6 (IL-6)
IL-6, 26 kD ağırlığında bir sitokindir. Damar entotel hücreleri, mononükleer fagositler,
epitel hücreler, fibroblastlar ve bazı aktive T hücreleri IL-6 sentezinden sorumludur. (61,69) IL-
6’nın reseptörü bağlayıcı protein ve sinyal ileten olmak üzere iki alt birimden meydana gelir. IL-
6’nın hepatositler ve B lenfositleri üzerine etkili olduğu bilinmektedir. IL-6 akut veya kronik
ınflamatuvar olay sonucu hemopeksin, fibrinojen, sistein proteinaz inhibitörü gibi akut faz yanıtına
katkıda bulunan birçok plazma proteinin hepatositlerce sentezlenmesine aracılık eder [120, 145].
IL-6, B lenfositlerinin immunoglobulin salınımı için kofaktör olrak görev yapar. B
lenfositlerine ayrışım sıralaması yapıldığında geç döneminde B lenfositler için büyüme faktörü

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
25
görevi yapar. Aynı etkiyi malign plazma hücreleri (plazmasitoma, myelom) için de yapar. Büyüyen
plazamasitom hücreleri otokrin büyüme faktörü olarak IL-6 salgılanmasını uyarır [146, 147].
Diğer sitokinlerle beraber kemik iliği hemopoetik ana hücrelerde erken fazda büyüme
faktörü olarak rol alır [146].
2.7.3.1.4. Kemokinler
Kemokinler yaklaşık olrak 8-10 kD ağırlığında olan ve yapısal olarak biribine benzeyen
geniş bir sitokin ailesidir. Kemokinler kemokinezi (lökosit hareketi), kemotaksisi (yönlendirilmiş
hareketi) uyarırlar. Monositler ve fibrablastlar için kimyasal madde çekici (kemoatraktan) özelliğe
sahiptir. Bu özellikleri sebebiyle kemokinler ismi verilmiştir. İki adet internal disülfit bağı içermesi
sebebiyle α ve β olrak iki alt gruba ayrılmıştır.
α alt grubuna ait kemokinler doku hücreleri (endotel, fibroblast), mononükleer fagositler
ve megakaryositlerce üretilirler. Akut inflamasyonda aracı olarak nötrofiller üzerine etki ederler.
α alt grubun en iyi biline ögesi interlökin-8 ‘dir. Aktif T hücreleri beta alt grubunun oluşumundan
sorumludur. Mononükleer inflamatuvar hücrelerin alt gruplarında etkilidir [129, 147].
Kemokin reseptörleri yedi transmembran α-heliks reseptör ailesine aittir. Reseptöre
bağlanma sonrasında; inozitoltrifosfat oluşumu, hücre içi kalsiyum artışı ve protein kinaz C
aktivasyonu şeklinde bir dizi olay akabinde hücrenin aktivasyonu sağlanır [148]. Ligand
bağlanması sonucunda bu reseptörlerin GDP’nin GTP’ye dönüşümünü katalizlediği
düşünülmektedir [129].
2.7.3.2. Lenfosit aktivasyonu, Büyüme ve Farklılaşmasın Düzenleyen Sitokinler
2.7.3.2.1. İnterlökin-2 (IL-2)
Bir diğer ismi T hücresi büyüme faktörüdür. T lenfositlerin hücre siklusunda G1 fazında S
fazına geçmesinde rol alır. IL-2, az miktarda CD8⁺T hücrelerinden üretilir, büyük çoğunlukla
üretiminden CD4⁺T hücreleri sorumludur. IL-2 otokrin büyüme faktörü özelliğine sahiptir. Bu
etkiyi kendisini üreten hücrelere etki edip üretilmesini katalize ederek yapar [127, 129, 149].
IL-2, 14-17 kD ağırlığında olan bir glikoproteindir. İnsanların 4. Geninde bulunan ve tek
genle kodlanan bir sitokindir.
Temelde etkileri lenfotsitler üzerinedir. T lenfositleri için otokrin büyüme faktörü görevi
yapar. IFN-γ ve lenfotoksin T hücresinde köken alan sitokinlerdir, bu sitokinlerin sentez

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
26
reaksiyonlarını uyarır. IL-2 reseptörü bünyesinde iki polipeptid zincirini barındırır. Bunlardan
birincisi IL-2Rα’dir, 55 kd ağırlığında ve T hücresi aktivasyonunda görev alan bir reseptördür. IL-
2Rα reseptörü, IL-2’ye Kd:10⁻⁸ M hız sabiti ile bağlanır. Bunlardan ikincisi IL-2Rβ’dır. Bu reseptör
70-75 kD’luk bir polipeptittir. IL-2’nin bu reseptöre bağlanma hızı Kd:10⁻⁹’dur. Yani IL-2’nin IL-
2Rβ reseptörüne bağlanma eğilimi daha yüksektir. IL-2Rβ kendisini 64 kD’luk bir polipeptit olarak
eksprese eder. Buna da IL-2Rβγ denilmektedir. IL-2 yalnızca IL-2Rβγ’ya karşılık gelen hücrelerin
büyümesinde görev alır.
Tirozin kinaz IL-2 aracılığı ile T hücresi büyümesini aktive eder.
IL-2, NK hücrelerinin büyümesini uyararak onların sitolitik özelliklerini arttırır. Sitolitik
özelliğini arttırma işelmini lenfokinle aktive edilmiş hücreleri (LAK) üreterek yapar. IL-2 diğer
sitokin hücreleri ile sinerjik etkileşim içine girerek NK hücreleri aracılığı ile IFN-γ üretimini arttırır.
IL-2 insan B lenfositlerine etki ederek antikor sentezini uyarır. Buna ek olarak büyüme
faktörü olarak görev yapar.
2.7.3.2.2. İnterlökin-4 (IL-4)
IL-4, 20 kd glikoprotein yapsındadır. Alerjik olayların düzenlenmesinde görev alır.
Antijenle stimüle olmuş CD4+T lenfositlerinden TH₂2’den kaynak alarak oluşur. IL-4, B ve T
lenfositlerinin büyüme, aktivite ve farklılaşmasında rol alır. IL-4’ün reseptörü 130 kD ağırlığında
reseptörü vardır. Bazofil hücreler, aktif mast hücreleri ve bazı CD8+T hücreleri tarafından sentez
edilir [127, 129, 150]. IL-4, IgE üretilmesinde görev alır. IgE hızlı gelişen alerjik reaksiyonların aracı
maddesidir ve IL-4 üretiminin artışına sebep olur. IgE, canlı organizmada meydana gelen
antikorları helmintik enfeksiyonlara karşı savunmada görev alır.
IL-4, makrofaj aktivasyonu inhibisyonu yapar. Nitrik oksit, prostaglandin üretimindeki
artış ve IFNϒ’nın makrofaj aktivasyonundaki etkilerini bloke eder.
IL-4, T lenfosit hücrelerinin TH₂ alt grubunun büyüme ve ayrışımında görev alır.
IL-4, damar endotel hücreleri üzerine de etikilir. Bu etkisiyle damar hücresi adezyon
molekülü 1’in (VCAM) ekpresyonunu uyarır. IL-4 ile muamele olmuş endotel hücreleri bir kemokin
olan Monosit Kemotaktik Protein-1 (MCP-1)’i ve eosinofiller üzerine etkili bir kemokin olan
eotaksin salgılanmasını uyarır. Yani, IL-4 yüksek lokal konsatrasyonlarda monosit ve eosinofillerce
oluşturulmuş inflamatuvar yanıtın oluşumuna aracılık eder.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
27
Mast hücreleri için büyüme faktörü olarak görev yapan IL-4, IL-3 ile kombine çalışarak
mast hücre proliferasyonunu artmasını sağlar.
IL-4, eosinofil ve IgE aracılı meydana gelen inflamatuvar olaylarda temel bir görev alır.
2.7.3.2.3. Transforming Büyüme Faktörü-β (TGF-β)
TGF-β, birbirleriyle ilişkili ve çeşitli genlerce kodlanmış moleküllerden oluşur. TGF ailesi
bünyesinde beş üye bulunmaktadır. (β₁, β₂, β₃, β₄, β₅). (123,129). Memelilerde TGF-β’nın β₁, β₂ ve
β₃ izoformaları bulunmaktadır. TGF-β₁, t lenfosit hücreleri ve monositlerce sentez edilir. Özellikle
merkezi sinir sistemi yüksek miktarda TGF-β₃ ihtiva eder. Doğal TGF-β yaklaşık 26 Kd ağırlığında
homodimer bir proteindir. TGF-β₁, latent ve etkisiz bir formda sentez edilir. TGF-β₁, proteazlar ile
aktive olur. LPS ile aktive olan mononükleer fagositler ve antijenle aktive olan T lenfosit hücreler
aktif TGF-β sentezinde görev alan hücre gruplarıdır [127, 129, 151]
Üç tip TGF-β reseptörü vardır. Tip 1, Tip 2 ve Tip 3 olarak adlandırılmışlardır. Sırasıyla
ağırlıkları 65 Kd, 85-90 kD ve 280 kD’ dir.
TGF-β çok çeşitli etkilere sahip bir sitokindir. β hücre proliferasyonu, işlevlerini modüle
etme ve başkalaşımı gibi görevleri vardır. TGF-β immün sistemi inhibe edici özellikte bir sitokindir.
B lenfosit hücrelerinin immün globülin sentezini suprese eder.
TGF-β, hücre tipine göre, hücre proliferasyonunda inhibitör veya stimülatör etki gösterir.
TGF-β, epitel hücreleri, enndotel hücreleri, T ve B lenfositleri, fibroblastlar üzerinde hücre
proliferasyonunu inhibe edici özellikte davranır.
TGF-β; yüksek konsantrasyonlarda hücre büyümesini önler, düşük konsatrasyonlarda ise
mitogenetik etki yapar. Bu özelliği mazenşiyal hücrelerde bifazik etki gösterdiğinin kanıtıdır.
Renal hastalıklarda, TGF-β ekstrasellüler matrikste üretimi ve döngüyü düzenleyici olarak
görev yapar. Bütün maktiks proteinlerinin üretimini arttırır. Bu proteinler; fibronektin, decorin,
biglikan ve Tip IV kollajendir ve ekstrasellüler maktrikste birikim yaparlar [120, 151, 152].
TGF-β, proteazları modüle eder ve buma bağlı olarak matriks yıkımı inhibisyonu yapar.
Proteaz salımını azaltan yönde etki eder. Plazminojen aktivatörlerinin etkisini inhibe eden
inhibitörlerin üretimini destekler.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
28
2.7.3.3. Bağışıklık Aracılığı ile İnflamasyonu Düzenleyen Sitokinler
Antijenle aktive olmuş CD4+ ve CD8+ T lenfositlerinden üretilen sitokinlerdir.
2.7.3.3.1. İnterferon-γ
İnterferon-γ, 20-24 kD’luk alt birimlerden ouşmuş bir glikoproteindir. IFN-γ, Th0 ve Th1
CD4⁺ yardımcı T hücreleri ve CD8+ T hücreleri tarafından üretilmektedir. Aynı zamanda öldüürücü
hücreler olan NK tarafından da üretilirler [127, 129, 153].
IFN-γ, Tip 1 interferon ile benzer etkinliklere sahiptir. IFN-γ antiproliperatif etkiiyi aktive
eder. Antiviral etkinliği arttırır. IFN-γ, hücre yüzeyinde tek bir tip hücre reseptörüne bağlanır. Bu
bağlanılan reseptör Tip 1 interferonun etkileştiği reseptörle benzer özellik gösterir fakat aynı
reseptör değildir.
Tip 1 interferondan farklı birçok işlevsel özelliği vardır. IFN-γ mononükleer fagosit
aktivatörüdür. IFN-γ öyle güçlü bir aktivatördür ki en temel Makrofaj Aktive Edici Faktör (MAF)
denilebilir. Bu aktivasyonu T lenfosit hücreleri aracılığı ile yapar. Class 1 major doku uygunluk
kompleksi (MHC) molekül ekspresyonunu arttırır. Direkt olarak T ve B lenfositlere etki eder ve
onların farklılaimasına neden olur. Nötrofil hücrelerini aktive eder. NK hücrelerinin sitolitik
aktivitelerini Tip 1 interferondan daha fazla uyarır. Damar endotel hücre aktivatörüdür. Buna ek
olarak TNF’nin endotel hücreler üzerine olan etkisini kuvvetlendirir.
2.7.3.3.2. Lenfotoksin (LT)
Lenfotoksin 21-24 kD ağırlığında glikoprotein yapısında bir sitokindir. %30 dolaylarında
TNF’ye homolog bir sitokindir [120, 126-128]. TNF ile aynı hücre reseptörüne bağlanmak için yarış
halindedir. LT’nin üretim yeri aktive T lenfosit hücreleridir. İnsan LT’si bir ya da iki adet N bağlı
oligosakkarit içerir.
Yapılan çalışmalar TNF ile LT’nin biyolojik etkinliklerinin benzer olduğunu göstermiştir.
İki sitokin arasındaki en belirgin fark LT’nin aktive T lenfosit hücreleri tarafından, TNF’ nin ise
büyük çoğunlukla mononükleer fagositler tarafından üretilmesidir [128, 141, 154]
T lenfosit hücreleri tarafından sentez adilmiş LT miktarı, LPS ile uyarılmış mononükleer
fagositlerden üretilmiş TNF’den oldukça azdır. Dolaşımda LT’ye rastlanmamıştır. Bu özelliği
nedeniyle lokal etki gösteren parakrin faktör olarak kabul edilir. Sistemil inflamasyon aracısı olma

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
29
özellğinden yoksundur. LT, damar endotel hücrelerinin ve nötrofilllerin aktivatörüdür. Lökosit
adezyonu, lökosit ekstravasyonu ve sitokin üretiminde aktive edici olarak rol alır.
2.7.3.3.3. İnterlökin-10 (IL-10)
IL-5, aktif B lenfosit hücreleri, aktive makrofajlar ve bazı nonlenfositik hücreler tarafından
üretilebilmesine rağmen büyük çoğunlukla CD4⁺ yardımcı hücrelerinin TH2 alt grubu tarafından
sentez edilir. 18 kD’luk bir sitokindir.
Biyolojik sistemde iki önemli görev yapar. İlki, makrofajlar tarafından, IL-1, TNF, IL-12 ve
kemokin gibi sitokinlerin üretimini durdurmasıdır. Diğeri, makrofajların T lenfosit hücresi
aktivasyonundaki görevlerini bloke etmektir. Makrofajların T lenfosit hücre aktivasyonundaki
işlevinin blokasyonunu, bazı ko-stimülatörlerin ve class 2 MHC moleküllerinin ekspresyonunu
azaltarak yapar. Bu iki etkisi sonucunda T lenfosit hücresi aracığı ile gelişen bağışık yanıtı inhibe
etmiş olur. IL-10’un B lenfositleri üzerine ise uyarıcı etkileri vardır [120, 126, 155].
2.7.3.3.4. İnterlökin-5 (IL-5)
IL-5, 40 kD ağırlığında, homodimerik yapıda bir sitokindir. Üretim yeri aktive mast
hücreleri ve CD4+T hücrelerinin TH2 alt grubudur [126, 127, 156].
IL-5’in en önemli görevi eozinofil hücrelerinin büyüme ve farklılaşmasını uyarmak ve
olgun eozinofillerin helmintleri yok edecek şekilde aktivasyonunu sağlamaktır. Diğer sitokinlerle
sinerji halindedir. IL-5, B lenfosit hücrelerine etki ederek özellikle IgA olmak üzere immunglobulin
sentezinde artışa neden olur.
2.7.3.3.5. İnterlökin-12 (IL-12)
IL-12, 70 Kd ağırlığında, kovalent bağlı iki polipeptit zincirinden oluşan heterodimer yapıda bir
sitokindir. Sahip olduğu bu polipeptit zincirlerinden biri 35 kD (p35) diğeri 40 kD (p40) dir. p35
alt grubunu üretim yeri, T ve B lenfositler, NK hücreleri ve monositlerdir. P40 alt grubunun üretim
yeri ise, B lenfosit hücreleri ve aktive monositlerdir. IL-12’nin p40 alt grubu IL-6 reseptörü ile
homologtur. Aktive T lenfosit ve NK hücreleri IL-12’nin reseptörünü eksprese eder.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
30
IL-12, NK ve T lenfosit hücrelerine etkisi sebebiyle hücresel bağışıklık yanıtının önemli
bir düzenleyicisidir. Bilinen en güçlü Nk hücresi uyaranı IL-12’dir. IL-2 ile güçlü bir sinerjik
etkileşim içerisindedir. NK hücrelerini uyararak IFN-γ’nın transkripsiyonunu tetikler. NK
hücrelerinin sitotoksik aktivitesini arttırır. Bu hücreler için büyüme faktörüdür. IL-12,
olgunlaşmamış CD4⁺ T hücrelerinin TH1 alt grubunun farklılaşması reaksiyonlarını uyarır. IL-12,
CD8⁺ T hücrelerinin aktif sitolitik T lenfositlere farklılaşması reaksiyonlarını da uyarır [120, 126].
2.7.3.3.6. Migrasyon İnhibisyon Faktör (MIF)
MIF, ilk olarak 1966 makrofaj migrasyonunu inhibe edici faktör olrak literatüre girmiştir
[157]. MIF, 115 aa içeren 12,5 kD ağırlığında ve peptit yaısında bir sitokindir [158]. T ve B
lenfositleri, eozinofiller, monosit ve makrofajlar, nötrofiller, epitelyal hücreler, mast hücreleri,
karaciğer, böbrek, beyin, pankreas MIF salgılanmasında görev alan hücre gruplarıdır. [159, 160,
161, 162]. Yapılan çalışmalar MIF’in gecikmiş tip hipersensitivitede görev aldığını göstermiştir
[157].
2.7.3.4. İmmatör Lökositlerin Büyüme ve Farklılaşmasına Neden Olan Sitokinler
Bu grup sitokinler hematopoezin oluşumunu aktive eden sitokinler olarak bilinirler.
İmmün yanıt sırasında üretilen sitokinlerin birçoğu kemik iliği progenitör hücrelerde büyüme ve
farklılaşma yapıcı kuvvetli uyarıcı etkileri vardır. Biyolajilk sistemlerde lökosit konsatrasyonunu
azaltan bağışık ve inflamatuar reaksiyonlar ile enflamatuvar hücrelerin yerine geçecek olan
lökositlerin üretminden de sorumlu olan bir dizi döngü içerisinde yer alır. Kemik iliği progenitor
hücrelerin genişlemesi ve farklılaşması reaksiyonlarını uyaran sitokinlere koloni uyaran faktörler
( CSFs ) denir.
2.7.3.4.1. C-Kit Ligand
C-kit ligand, 24 kD ağırlığında, üretim yeri kemik iliği stroma hücreleri olan bir
monomerdir. C-kit ligandın bir diğer ismi de ana hücre faktörüdür [163]. C-kit ligand ana
hücrelerin diğer CSF’lere yanıt oluşturması bakımından gereklidir. Kendisi koloni
oluşturamamaktadır.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
31
2.7.3.4.2. İnterlökin-3 (IL-3)
Bir diğer ismi de çok nesilli koloni uyaran faktördür (Multi CSF). Büyük bir kısmı CD4⁺T
lenfosit hücrelerinden üretilir. Bunun yanısıra miyelomonositik hücrelerce de sentez edilir. 20-26
kD ağırlığında ve dimer yapıdadır. İmmatör kemik iliği hücrelerinin farlılaşıp büyümesi yönünde
etki eder [126].
2.7.3.4.3. Granülosit-Makrofaj Koloni Uyaran Faktör (GM-CSF)
Granülosit-makrofaj koloni uyaran faktör, 26 kD ağırlığında ve glikoprotein yapısındadır. Üretim
yeri aktive T lenfosit hücreleri, damar endotel hücreleri, aktive monositler ve fibroblastlardır.
Biyolojik sistemlerdeki hücrelerin tamamının büyüme ve farlılaşmasında görev alır [164]. GM-CSF,
üretildiği bölgeye lokal olarak etki eder, canlı organizmanın dolaşım sisteminde bulunmaz.
Dolayısıyla immün inflamatuar yanıt bölgelerinde olgun lökosit aktivasyonu yapar. T lenfosit
hücrelerinin, endotel hücrelerin ve kemik iliğindeki stromal fibroblastların ürettiği GM-CSF’ler
homopoetik özellik gösterir. Rekombinant GM-CSF hemotopoez eksikliği olan hastalarda kemik
iliğini uyarmak için ve sitotoksik kemoterapi ya da kemik iliği transplantasyonu sonrasnda kemik
iliğinin iyileşmesini desteklemek için kullanılmaktadır [120, 126].
2.7.3.4.4. Monosit-Makrofaj Koloni Uyaran Faktör (M-CSF)
Monosit-makrofaj koloni uyaran faktör CSF-1 olarak da bilinmektedir. Üretim yeri
makrofajlar, fibroblastlar ve endotel hücrelerdir. M-CSF, 40 kD’luk dimer yapıda bir moleküldür.
M-CSF’nin reseptörü c-Kit ligand ile yapısal olarak birbirlerine benzerler. M-CSF, olgun monositlere
dönüşecek olan monosit progenitorlar üzerinde etkilidir. M-CSF, organizmanın dolaşım sisteminde
bulunmaz. Koloni uyarıcı etkisini ilik boşluğunda üretildiği yerde lokal olarak yapar [120, 126].
2.7.3.4.5. Granülosit-Koloni Uyaran Faktör (G-CSF)
GCSF, aktive T lenfosit hücreleri, damar endotel hücreleri, aktive mononükleer fagositler
ve fibroblastlar tarafından üretilir. G-CSF, 19 kD ağırlığında dimerik yapıda bir moleküldür. Olgun
granülositlere dönüşecek olan progenitorlara etkilidir, canlı organizmanın dolaşım sisteminde
bulunur. Üretildiği bölgeden farklı bölgelerde de etkili olduğu için nötrofil oluşması ve kemik

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
32
iliğinde serbestleşmesi, kemik iliği dışında meydana gelen inflamatuar reaksiyonlarda etkindir
[120].
2.7.3.4.6. İnterlökin-7 ( IL-7 )
IL-7, kemik iliği stroma hücreleri ve fibroblastlar tarafından salgılanan, 25 kD’luk bir
monomerdir. B lenfositlere dönüşecek olan homepoetik progenitorlar üzerinde etikilidir. T ve B
lenfosit progenitorları üzerinde gelişme faktörü olarak etkindir [165].
2.7.3.4.7. Diğer Koloni Uyaran Sitokinler
IL-9, 30-40 kD ağırlığında protein yapıda bir moleküldür. Kemik iliğinden ve T lenfosit
hücrelerinden köken almış mast hücresi progenitorlarının farklılaşmasında ve gelişmesinde rol
alır. Kemik iliği kaynaklı diğer hücrelerin gelişiminde de etkindir.
IL-11, aktive olmuş kemik iliği hücrelerinden üretilen, 20 kD’luk bir sitokindir.
Megakaryopoezi uyarıcı etkisi vardır ve bu özelliği ile trombositopenide tedavi edici tavır sergiler.
[120, 126].
IL-13, TH2 hücreleri tarafından üretilir. IL-4 ile benzer özelliklere sahiptir. B lenfosit
hücrelerine IgE sentezinde destek olur. Monosit adezyonunu destekler yönde hareket eder [166].
IL-14, T lenfosit hücreleri tarafından üretilir. Aktif B lenfosit hücre proliferasyonu
indükleyicisidir. Antijenle stimüle olmuş B lenfosit hücrelerinin Ig yapımının inhibisyonunda görev
alır [166].
2.8. Alkol Metabolizması
Canlı organizmaya alınan alkolün yaklaşık % 95’i karaciğer dokusunda asetik asite kadar
okside olur. Geriye kalan % 5’lik kısmı ise idrar, ter ve feçes yolu ile biyolojik sistemden atılır.
Alkolün organizmada oksidasyonu 3 farklı yolak ile yapılır. Bu yolaklar, hepatosit sitozolünde
bulunan alkol dehidrogenaz enzim yolağı (ADH), hepatosit endoplazmik retikulumunda bulunan
mikrozomal etanol oksidasyon sistemi (MEOS) ve hepatik peroksizomlarda var olan katalaz enzim
yolağıdır.
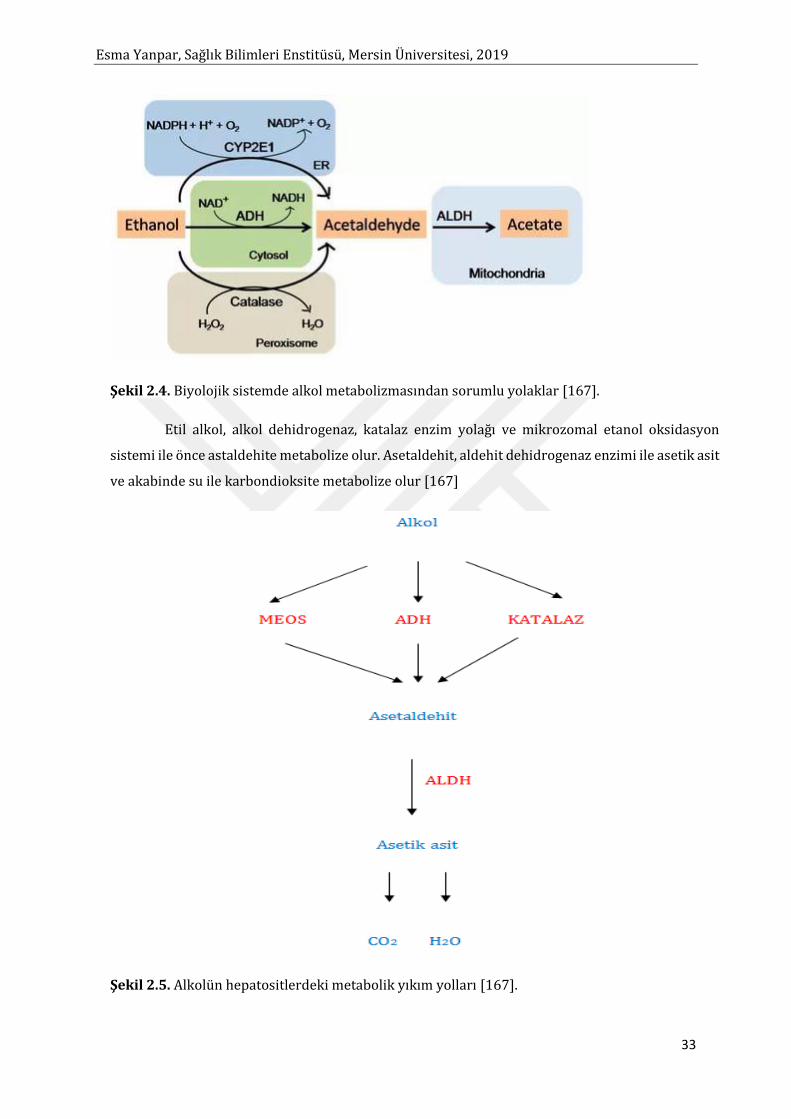
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
33
Şekil 2.4. Biyolojik sistemde alkol metabolizmasından sorumlu yolaklar [167].
Etil alkol, alkol dehidrogenaz, katalaz enzim yolağı ve mikrozomal etanol oksidasyon
sistemi ile önce astaldehite metabolize olur. Asetaldehit, aldehit dehidrogenaz enzimi ile asetik asit
ve akabinde su ile karbondioksite metabolize olur [167]
Şekil 2.5. Alkolün hepatositlerdeki metabolik yıkım yolları [167].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
34
Alkol dehidrogenaz enzim yolağı, hepatositlerde etanol metabolizmasında kullanılan en
önemli yolaktır. NAD⁺, NADH’a indirgenir. Etil alkol, asetaldehite dönüşür. Asetaldehit, aldehit
dehidrogenaz enzimi ile asetik asite dönüştürülür, NAD⁺ yeniden NADH’a indirgenir. [168].
Şekil 2.6. Alkol dehidrogenaz enzim yolağı [168].
ADH’ın, beş farklı geni bulunmaktadır. Bunlardan ADH2 ve ADH3 polimorfizm gösterir.
ADH2 ve ADH3 genine sahip olanlar, bu genlere sahip olmayanlar ile kıyaslandığında çok daha hızlı
bir şekilde etanolü metabolize eder. Doğu Asyalılarda bu genlere daha yaygın rastlanmaktadır
[168]. Etanol metabolizmasında birinci basamak olarak adlandırabileceğimiz ADH enzim aktivitesi
sonucunda alkol, asetaldehite dönüşür. Reaksiyon denkleminin ikinci basamağında asetaldehit
dehidrogenaz (ALDH) enzim aktivitesi ile asetaldehit, toksikolojik olarak etkisiz olan asetik asite
dönüştürülür. ALDH’ın iki önemli izoformu vardır. ALDH2 bu izorformlar arasında en önemlisidir.
ALDH2’de kendi içinde iki farklı gen polimorfizmi içerir. Bu polimorflardan biri aktiftir, fakat diğeri
inaktiftir. Uzak doğu kökenli bireyler ALDH2’nin inaktif genetik polimorfizmine sahiptirler. Bu
nedenle alkol metabolizmaları oldukça yavaş seyreder. Az miktarda alkol aldıklarında bile oldukça
ciddi sağlık sorunları görülmektedir [169].
Mikrozomal etanol oksidasyon yolağı, endoplazmik retikulumda bulunur. Vücuda alınan
alkolün yaklaşık % 20’sinin metabolizmasından sorumlu olan yolaktır. Sitokrom P450’nin CYP2E1
izoformudur. Bu yolakta etanolün metabolizması moleküler oksijenin NADPH’ı okside etmesi ile
gerçekleşir. İki oksijen atomu kullanılır. Bir tanesi NADPH’ı, NADP⁺’ye yükseltgenmesini sağlar.
Diğer oksijen atomu da etanolün asetaldehite okside olmasını sağlar. İndirgenmiş oksijen atomları,
iki mol su oluşumu ile reaksiyonu tamamlar [170].
Katalaz enzim yolağı, peroksizomlarda bulunan bir enzim olan katalaz ile yürüyen bir
yolaktır. Alkolün metabolimasının yaklaşık % 2’lik kısmından sorumludur ve nonspesifiktir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
35
Hidrojen peroksit suya dönüşür. Etil alkol, asetaldehite dönüştürülür. Aldehit dehidrogenaz enzimi
ile asetaldehit asetik asite dönüştürülür [171].
2.8.1. Alkolün Oksidatif Hasar Üzerine Etkisi
Süperoksit ve hidrojen peroksit gibi ROS türlerinin oluşumu birçok hastalıkta oksidatif
hasarın önemli bir nedenini temsil eder. Az miktarda geçiş metali (sıklıkla Fe) varlığında O₂˙ ve
H₂O₂, daha sonrada biyolojik bileşiklerin oksidasyonundan sorumlu OH˙ radikali oluşur. Sitokrom
P450’nin izoformu olan CYP2E1 karaciğerde O₂˙ ve H₂O₂ kaynağıdır [172]. Alkolle muamele olmuş
karaciğer mikrozomlarında CYP2E1 ile lipit peroksidasyonu arasında orantılı bir artış söz
konusudur [173].
Etanolün metabolizmasında kullanılan ADH yolağının kullanılması ile NADH üretiminin
artmasıyla birlikte artan elektron sızıntısı mitokondrilerde solunum zincirini aktive eder ve
mitokondriyal O₂˙ oluşumuna neden olur [174].
Alkol metabolizmasında MEOS sistemi devreye girer. Bu mekanizma ile ortamda serbest
oksijen radikalleri oluşur ve akabinde lipit peroksidasyonu meydana gelir. Lipit peroksidasyonu
reaksiyonu ile hücre içi kalsiyum geçirgenliği artar. Hücre içinde biriken bu kalsiyum iyonları
fibrozis, nekrozis ve mitokondri disfonksiyonuna neden olur.
Alkol metabolizmasının ara ürünü olan asetaldehit oksidatif etki ile serbest radikal
aktiviteside artmaya ve lipit peroksidasyonuna sebep olur. Aynı zamanda toksik lipit aldehitlerinin
ortadan kaldırılmasında ve GSH’ın aktivasyon ve antioksidan fonksiyonunda bozulmalara neden
olur [175, 176].
2.8.2. Alkolün Sitokin Üretimi ve İnflamasyon Üzerine Etkisi
Etanol alımına bağlı gelişen hepatatoksisitenin altında yatan mekanizmalar oldukça
karmaşıktır. TNF-α gibi proinflamatuvar sitokinlerin önemli bir rolü vardır. Karaciğer hasarının
başlangıcında dolaşımda TNF-α’nın arttığı gözlenmiştir [177-179]. Alkol alımına bağlı olarak
kupffer hücreleri TNF-α, IL-6, IL-1, prostaglandin E₂ üretir [177, 178].
Sitokinlerin ana kaynağı olan kuppfer hücre aktivasyonu barsakta bulunan gram negatif
bakterilerin hücre duvarı malzemesinde türeyen endotoksine maruz kalması gerçekleşir. Etanol
uygulaması sonrasında dolaşımda endotoksin varlığı saptanmıştır [180]. Kupffer hücreleri

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
36
tarafından salgılanan kemokinler ve sitokinler karaciğerde savunma yanıtlarını başlatan bir dizi
otokrin ve parakrin etkiye sebep olur. Salınan sitokinler inflamatuvar lökositlerin infiltrasyonunda
ve oksidatif stres patlamalarında aktivasyona neden olur. Bu karmaşık basamaklar ve oksidatif
strese bağlı oluşan hücre savunma mekanizmaları karaciğer hücrelerinde şiddetli akut zorluk
döngüsüne sebep olur.
Normalde TNF-α karaciğer hücresi için toksik değildir. Fakat apaptoz ve nekroz ile hücre
ölümüne yol açan bir dizi reaksiyonu başlatabilir. Hücreler de bu hücre ölüm reaksiyonlarına
karşılık sitoprotektif sinyalleri aktive eder.
Karakteristik olarak TNF-α ve IL-6 karaciğer rejenerasyonunu başlatan temel
bileşenlerdir. TNF-α ve IL-6 hepatik inflamasyon düzeyi ile doğru orantılı olarak yüksektir. Etanol
muamelesi ile sitokinlere olan erken yanıtta bozulmuş rejeneratif yanıt geliştiği gözlenmiştir [181,
182].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
37
3. MATERYAL VE YÖNTEM
3.1. Materyal
3.1.1. Kullanılan Alet ve Malzemeler
Soğutmalı Mikrosantrifuj (Sigma 2-16K)
Derin Dondurucu (Regal, RDD 1145)
Etüv (Binder)
Spektrofotometre (Analytikjena-SPECORD 50)
Hassas Terazi (Mettler Toledo)
Buzdolabı (Regal, RBD 4602 NCF)
Vorteks (REAX)
Mini santrifüj (Eppendorf)
Su Banyosu (GFL-Wasserbad Water Bath)
Distile Su Cihazı (Millipore)
Mikropipet Seti (Gilson- Pipetman- P20-P100-P1000)
pH Metre (Mettler Toledo)
Homojenizatör (IKA)
ELISA Reader (Allsheng AMR-100 Mikroplate)
3.1.2. Kullanılan Kimyasal Malzemeler
1-Butanol (Riedel de Haen)
Amonyum Sülfat (Sigma)
Asetik Asit (Merck)
Bakır Klorür (Sigma)
Disodyum Hidrojen Fosfat (Merck)
Etil Alkol (Etanol) (Merck)
Etilendiamintetraasetikasit (EDTA) (Merck)
Glutatyon Redüktaz (Sigma)
Hidrojen Peroksit (Merck)
Ketamin Pfizer (Ketalar flakon)
Ksalazin Bayer (Rompun flakon)
Ksantin (Sigma)

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
38
Ksantin Oksidaz (Sigma)
Nitroblue Tetrazolium Klorür (NBT) (Sigma)
Piridin (Riedel de Haen)
Potasyum Dihidrojen Fosfat (Merck)
Redükte L-Glutatyon (Sigma)
Redükte β-NADPH (Sigma)
Sodyum Azid (Sigma)
Sodyum Dodesil Sülfat (Sigma)
Sodyum Hidroksit (Merck)
Sodyum Karbonat (Merck)
Sodyum Klorür (Merck)
Tetrametoksipropan (Aldrich)
Tiobarbitürik Asit (TBA) (Merck)
Trikloroasetik Asit (TCA) (Merck)
Punicalagin (Sigma)
3.1.3. Hayvan Materyali
Deneylerde ortalama 250-300 gram ve 12 saat karanlıkta 12 saat aydınlıkta senkronize
edilen 24 adet erişkin Wistar albino cinsi dişi sıçan kullanılmıştır. Sıçanlar rastgele seçilerek 4
gruba ayrılmıştır. Grupların kendi içinde homojen olmasına ve gruplardaki sıçan ağırlıklarının
toplamının yaklaşık aynı değerde olmasına dikkat edilerek gruplar oluşturulmuştur. Her grup ayrı
kafesler yer alacak şekilde özel kafesler içinde standart sıçan yemi ile beslenmiştir. Kullanılan
sıçanlar Mersin Üniversitesi Tıp Fakültesi Deneysel Tıp Araştırma Merkezinden sağlanmıştır.
Deney süresince gün aşırı içme suları değiştirlmiş ve kafes temizliği yapılmıştır. Bu çalışmada
sıçanlar üzerine yapılan tüm deneysel işlemler için Mersin Üniversitesi Hayvan Deneyleri Yerel
Etik kurulundan onay alınmıştır (2017/HADYEK/24).
Çalışmamız her grupta 6 hayvan olacak şekilde 4 gruptan oluşmuştur.
Bu gruplar;
Grup 1 (Kontrol Grubu) (n=6): Herhangi bir madde uygulanmadan 1 cc % 0,9’luk NaCl
sıçanlara 21 gün boyunca gavaj yolu ile uygulanmıştır. Punicalagin maddesi verilmemiştir. 21
günün sonunda hayvanlar sakrifiye edilmişir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
39
Grup 2 (Etil Alkol Grubu) (n=6): Bu gruptaki sıçanlara gavaj yolu ile 4 g/kg etil alkol 21
gün boyunca gavaj yolu ile uygulanmıştır. 21 günün sonunda hayvanlar sakrifiye edilmiştir.
Grup 3 (Punicalagin Grubu) (n=6): Bu gruptaki sıçanlara gavaj yolu ile 4 mg/kg
punicalagin gavaj yolu ile 21 gün boyunca uygulanmıştır. 21 günün sonunda hayvanlar sakrifiye
edilmiştir.
Grup 4 (Punicalagin+Etil Alkol Grubu) (n=6): Bu gruptaki sıçanlara 4 mg/kg punicalagin
ile 4 g/kg etil alkol 21 gün boyunca gavaj yolu ile uygulanmıştır. 21 günün sonunda hayvanlar
sakrifiye edilmiştir.
3.2. Yöntem
3.2.1. Çalışma Örneklerinin Hazırlanması
3.2.1.1. Doku Örnekleri
İntraperitoneal olarak uygulanan ketamin (200 mg/kg i.p.) anestezisi altındaki dişi
sıçanların karaciğer dokusu izole edilmiş ve dokular serum fizyolojik solüsyonundan geçirilerek
temizlenmiştir ve çalışma gününe kadar dokular derin dondurucuda saklanmıştır.
3.2.1.2. Homojenizasyon
Yaş ağırlıkları 100 mg olmak üzere dokular tartıldı, üzerine 1/10 (w/v) oranı sağlanacak şekilde
serum fizyolojik çözeltisi eklenerek homojenize edildi. Cam tüplere aktarılan doku örnekleri
numaralandırılarak 13.000 rpm'de, 10 dakika, +4°C’de santrifüj işlemi gerçekleştirildi.
Homojenizatlarda lipid peroksidasyon göstergesi olarak MDA düzeyi, antioksidan olarak katalaz,
SOD enzim aktiviteleri ve GSH düzeyi ölçülmüştür. İnflamasyon belirteçlerinden olan IL-6, TNF-α
düzeylerine bakıldı.
3.2.2. Analiz Yöntemleri
3.2.2.1. Glutatyon Düzeyinin Ölçümü
GSH düzeyleri Sedlak and Lindsay’in bildirdikleri yönteme göre spektrofotometre cihazı ile
belirlendi [183]. GSH tayini için gerekli solüsyonlar; %10 triklorikasetik asit (TCA) solüsyonu Tris

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
40
tamponu (0,4 M pH:8,9): 48,46 gram Tris-hydroxymethil-aminomethan’ın hidroklorik asit (HCl)
ile pH 8,9 olacak şekilde 1 litrede çözülmesi ile hazırlandı.
Deneyin yapılışı: 0,1 mL doku homojenatı 0,4 mL TCA solüsyonu ile karıştırıldı. 20 sn
vorteks kullanılarak karıştırıldı ve 13000 rpm de 5 dk santrifüj edildi, 0,1 mL süpernatant temiz
bir tüp içine alındı. Üzerine 0,9 mL distile su, 2,0 mL Tris tamponu ve 0,1 mL DTNB solüsyonu
eklendi. Oluşan sarı renk distile suya karşı spektrofotometrede 412 nm dalga boyunda okundu.
3.2.2.2. Katalaz Enzim Aktivitesi Tayini
Katalaz enzim aktivite tayini Aebi’nin belirttiği yönteme göre gerçekleştirilmiştir [184].
Yöntemin prensibi, H2O2 substratının katalaz enzimi ile yıkılmasının 240 nm’de izlenmesi esasına
dayanır.
Kullanılan Reaktifler:
Potasyum Dihidrojen Fosfat Çözeltisi (A): 0,681 g KH₂PO₄ bidistile su içerisinde
çözülerek 100 mL hacime tamamlandı.
Disodyum Hidrojen Fosfat Çözeltisi (B): 2,77 g Na₂HPO₄.12H2O bidistile su içerisinde
çözülerek 155 mL hacime tamamlandı.
Fosfat Tamponu (50 mM pH 7,0): pH 7olacak şekilde, A/B oranı 1/1,55 olacak biçimde
karıştırılarak hazırlandı.
Hidrojen Peroksid Çözeltisi (30 mM): 34 μl %30 luk H₂O₂, fosfat tamponu ile 10 Ml
hacime tamamlanarak hazırlandı.
Deneyin Yapılışı: Kuvarzt spektrofotometre küvetlerinin içerisine, 10 μL 10.000 g
süpernatant fraksiyonu ve bu eklenen fraksiyonların üzerlerine 1990 μL fosfat tamponu eklendi.
Örnek küveti içerisine 1 mL 30 mM H2O2 eklendi ve hızla karıştırılarak örneğin absorbansındaki
azalma köre karşı 1 dakika boyunca 240 nm’de izlendi. Spesifik aktivite ünite/mg protein cinsinden
belirlendi.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
41
3.2.2.3. Süperoksit Dismutaz Enzim Aktivitesi Tayini
Deneyin Prensibi: Endojen ve eksojen kaynaklı olabilen, oksidatif fosforilasyon yolu ile
enerji üretimi sırasında meydana gelen süperoksit radikellerinin su ve moleküler oksijene
dismutasyonunun katalizinden sorumlu SOD enzim aktivitesinin ölçüm esası, ksantin varlığında
ksantin oksidazın oluşturduğu süperoksit radikallerinin nitroblue tetrazolium (NBT) ile 560 nm’de
absorblanan rengin ölçülmesine dayanır (185).
Kullanılan Reaktifler:
Sodyum Hidroksid Çözeltisi (0,1 N): 0,4 g NaOH, bidistile su içerisinde çözülerek 100 mL
hacime tamamlandı.
Stok Ksantin Çözeltisi (3 mM): 4,6 mg ksantin, 1 mL 0,1 N NaOH çözeltisi içerisinde hafif
ısı hafif ısı verilip çözülerek bidistile su ile 10 mL hacime tamamlandı. Çözelti +4 °C’de bir hafta
stabildir.
Ksantin Çözeltisi (0,3 mM): Stok olarak hazırlanan ksantin çözeltisinden 1 mL alınarak
bidistile su ile 10 mL hacime tamamlandı. Deney günü çözelti taze olarak hazırlandı.
Etilendiamintetraasetik Asid Çözeltisi (0,6 mM): 22,32 mg Na2EDTA. 2H2O bidistile su
içerisinde çözülerek 100 mL hacime tamamlandı.
Sodyum Karbonat Çözeltisi (400 mM): 4,24 g Na2CO3, bidistile su içerisinde çözülerek
100 mL hacime tamamlandı.
BSA Çözeltisi (1 g/L): 10 mg BSA bidistile su içerisinde çözülerek 10 mL hacime
tamamlandı.
Nitroblue Tetrazolium Klorür (NBT) Çözeltisi (0,15 mM): 1,226 mg NBT tartılıp,
bidistile su içeriside çözülerek 10 mL hacime tamamlandı. Çözelti deney günü taze olarak
hazırlandı.
SOD Çalışma Reaktifi: 10 mL 0,3 mM ksantin, 3 mL 400 mM Na2CO3, 5 mL 0,15 mM NBT,
5 mL 0,6 mM Na2EDTA, 1,5 mL 1 g/L BSA karıştırılarak deney günü taze olarak hazırlandı.
Amonyum Sülfat Çözeltisi (2 M): 26,428 g (NH4)2SO4, bidistile su içerisinde çözülerek 100
mL hacime tamamlandı.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
42
Ksantin Oksidaz Çözeltisi (167 U/L): 2 mg ksantin oksidaz tartılarak soğutulmuş
(NH4)2SO4 çözeltisi içerisinde çözülerek 1 mL hacime tamamlandı.
Bakır Klorür Çözeltisi (0,8 mM): 13,64 mg CuCl2.2H2O tartılarak, bidistile su içerisinde
çözülerek 100 mL hacime tamamlandı.
Deneyin Yapılışı: 2,85 mL olarak ölçülen SOD reaktifi örnek tüpüne akabinde kör tüpüne
aktarıldı. Örnek tüpüne 100 μL süpernatant eklendi ve her iki tüpe de 50 μL ksantin oksidaz ilave
edildi. Tüpler 25 °C’de 20 dakika inkübasyon işeleminden sonra tüplere 100 μL CuCl2 çözeltisi ilave
edildi. Son işlem olarak da kör tüpüne 100 μL süpernatant eklenip karıştırıldı. Örnek tüpünün ve
kör tüpünün 560 nm’de absorbansları ayrı ayrı ölçülüp SOD aktivite tayini yapıldı. Spesifik aktivite
U/mg protein türünden belirtildi.
3.2.2.4. Malondialdehit Ölçümü
Lipit peroksidasyon ürünlerinden en stabili olan MDA’nın tiobarbütirik asit ile reaksiyon
vermesi sonucu oluşan pembe-kırmızı rengin absorbansının spektrofotometrik olarak ölçülmesi
esasına dayanır [186].
Kullanılan Reaktifler:
Stok tetrametoksipropan çözeltisi: 0,92 gr tetrametoksipropan 1 mL’de çözüldü.
Günlük tetrametoksipropan çözeltisi: 10 mL’lik stok çözelti 100 mL’ye distile su ile
tamamlandı. Çalışma esnasında günlük hazırlanan çözelti yeniden 1/10 oranında seyreltildi.
SDS çözeltisi: 8,1 gr SDS tartıldıktan sonra 100 mL’ye distile su ile tamamlanılıp, çözüldü.
Asetik Asit çözeltisi: 20 mL asetik asit ölçülüp distile su ile 100 mL hacime tamamlandı ve
pH’sı 3,5’e ayarlandı.
TBA çözeltisi: 0,8 gr TBA tartımı yapıldıktan sonra 100 mL hacime distile su ile
tamamlandı ve ısıtılarak çözülme işlemi tamamlandı.
n-bütanol-piridin çözeltisi (15:1): Stok piridinin 1 mL’si 15 mL n-bütanol ile karıştırıldı.
Deneyin Yapılışı:
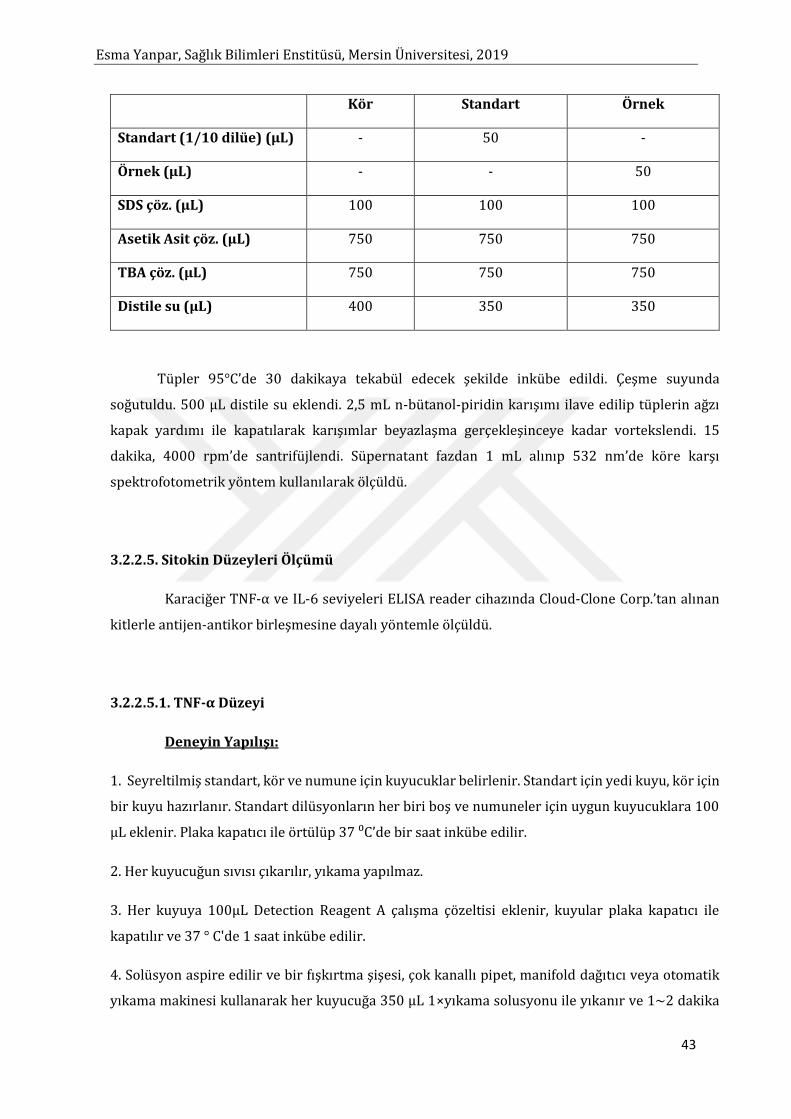
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
43
Kör Standart Örnek
Standart (1/10 dilüe) (µL) - 50 -
Örnek (µL) - - 50
SDS çöz. (µL) 100 100 100
Asetik Asit çöz. (µL) 750 750 750
TBA çöz. (µL) 750 750 750
Distile su (µL) 400 350 350
Tüpler 95°C’de 30 dakikaya tekabül edecek şekilde inkübe edildi. Çeşme suyunda
soğutuldu. 500 µL distile su eklendi. 2,5 mL n-bütanol-piridin karışımı ilave edilip tüplerin ağzı
kapak yardımı ile kapatılarak karışımlar beyazlaşma gerçekleşinceye kadar vortekslendi. 15
dakika, 4000 rpm’de santrifüjlendi. Süpernatant fazdan 1 mL alınıp 532 nm’de köre karşı
spektrofotometrik yöntem kullanılarak ölçüldü.
3.2.2.5. Sitokin Düzeyleri Ölçümü
Karaciğer TNF-α ve IL-6 seviyeleri ELISA reader cihazında Cloud-Clone Corp.’tan alınan
kitlerle antijen-antikor birleşmesine dayalı yöntemle ölçüldü.
3.2.2.5.1. TNF-α Düzeyi
Deneyin Yapılışı:
1. Seyreltilmiş standart, kör ve numune için kuyucuklar belirlenir. Standart için yedi kuyu, kör için
bir kuyu hazırlanır. Standart dilüsyonların her biri boş ve numuneler için uygun kuyucuklara 100
µL eklenir. Plaka kapatıcı ile örtülüp 37 ⁰C’de bir saat inkübe edilir.
2. Her kuyucuğun sıvısı çıkarılır, yıkama yapılmaz.
3. Her kuyuya 100μL Detection Reagent A çalışma çözeltisi eklenir, kuyular plaka kapatıcı ile
kapatılır ve 37 ° C'de 1 saat inkübe edilir.
4. Solüsyon aspire edilir ve bir fışkırtma şişesi, çok kanallı pipet, manifold dağıtıcı veya otomatik
yıkama makinesi kullanarak her kuyucuğa 350 μL 1×yıkama solusyonu ile yıkanır ve 1~2 dakika

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
44
bekletilir. Plakayı emici kağıta geçirerek kalan sıvı tüm kuyucuklardan tamamen çıkarılır.
Tamamen 3 kez yıkanır. Son yıkamadan sonra, kalan yıkama tamponu aspire edilerek veya
boşaltılarak çıkarılır. Plaka ters çevirilir ve emici kağıta karşı lekelenir.
5. Her bir oyuğa 100 μL Detection Reagent B çalışma çözeltisi eklenir, kuyucuklar plaka kaplayıcı
ile kapatılır ve 37°C'de 30 dakika inkübe edilir.
6. Aspirasyon/yıkama işlemi 4. adımda yapıldığı gibi toplam 5 kez tekrarlanır.
7. Her kuyucuğa 90 μL substrat solusyonu eklenir. Yeni bir plaka kapatıcı ile örtülür. 37°C'de 10-
20 dakika inkübe edilir (30 dakikayı aşmadan). Işıktan koruyarak işlem gerçekleştirilir. Sıvı,
Substrat Solüsyonu eklenerek maviye döndürülür.
8. Her kuyucuğa 50 μL durdurma solüsyonu eklenir. Durdurma çözeltisi eklendiğinde sıvı sararır.
Plakanın tarafına dokunarak sıvı karıştırılır. Renk değişimi aynı görünmüyorsa, iyice karışmasını
sağlamak için plakaya hafifçe dokunulur.
9. Plakanın altındaki su damlası ile parmak izi kaldırılır ve sıvının yüzeyinde kabarcık olmadığı
doğrulanır. Ardından, mikroplaka okuyucuyu çalıştırılıp ve hemen 450 nm'de ölçüm yapılır.
3.2.2.5.2. IL-6 Düzeyi
Deneyin Yapılışı:
1. Seyreltilmiş standart, boş ve numune için kuyucuklar belirlenir. Standart için yedi kuyu, kör için
bir kuyu hazırlanır. Standart dilüsyonların her biri boş ve numuneler için uygun kuyucuklara 100
µL eklenir. Plaka kapatıcı ile örtülüp 37⁰C’de bir saat inkübe edilir.
2. Her kuyucuğun sıvısı çıkarılır, yıkama yapılmaz.
3. Her kuyuya 100μL Detection Reagent A çalışma çözeltisi eklenir, kuyular plaka kapatıcı ile
kapatılır ve 37 ° C'de 1 saat inkübe edilir.
4. Solüsyon aspire edilir ve bir fışkırtma şişesi, çok kanallı pipet, manifold dağıtıcı veya otomatik
yıkama makinesi kullanarak her kuyucuğa 350 μL 1×yıkama solusyonu ile yıkanır ve 1~2 dakika
bekletilir. Plakayı emici kağıta geçirerek kalan sıvı tüm kuyucuklardan tamamen çıkarılır.
Tamamen 3 kez yıkanır. Son yıkamadan sonra, kalan yıkama tamponu aspire edilerek veya
boşaltılarak çıkarılır. Plaka ters çevirilir ve emici kağıta karşı lekelenir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
45
5. Her bir oyuğa 100 μL Detection Reagent B çalışma çözeltisi eklenir, kuyucuklar plaka kaplayıcı
ile kapatılır ve 37°C'de 30 dakika inkübe edilir.
6. Aspirasyon/yıkama işlemi 4. adımda yapıldığı gibi toplam 5 kez tekrarlanır.
7. Her kuyucuğa 90 μL Substrat Solusyonu eklenir. Yeni bir plaka kapatıcı ile örtülür. 37°C'de 10-
20 dakika inkübe edilir (30 dakikayı aşmadan). Işıktan koruyarak işlem gerçekleştirilir. Sıvı,
Substrat Solüsyonu eklenerek maviye döndürülür.
8. Her kuyucuğa 50 μL durdurma solüsyonu eklenir. Durdurma çözeltisi eklendiğinde sıvı sararır.
Plakanın tarafına dokunarak sıvı karıştırılır. Renk değişimi aynı görünmüyorsa, iyice karışmasını
sağlamak için plakaya hafifçe dokunulur.
9. Plakanın altındaki su damlası ile parmak izi kaldırılır ve sıvının yüzeyinde kabarcık olmadığı
doğrulanır. Ardından, mikroplaka okuyucuyu çalıştırılıp ve hemen 450 nm'de ölçüm yapılır.
3.2.3. İstatiksel Analiz
Gruplarda yer alan sonuçların normal dağılıma sahip olup olmadıklarını belirlemek için
nonparametrik testlerden one-sample Kolmogorov-Smirnov testi kullanılmıştır. Elde edilen veriler
ortalama±standart sapma şeklinde ifade edilmiş ve p değeri 0,05 ve daha küçük olan veriler
istatistiksel olarak anlamlı kabul edilmiştir. Biyokimyasal parametrelerin değerlendirilmesinde
tek yönlü varyans analizi (ANOVA) ve Tukey testi kullanılmıştır.
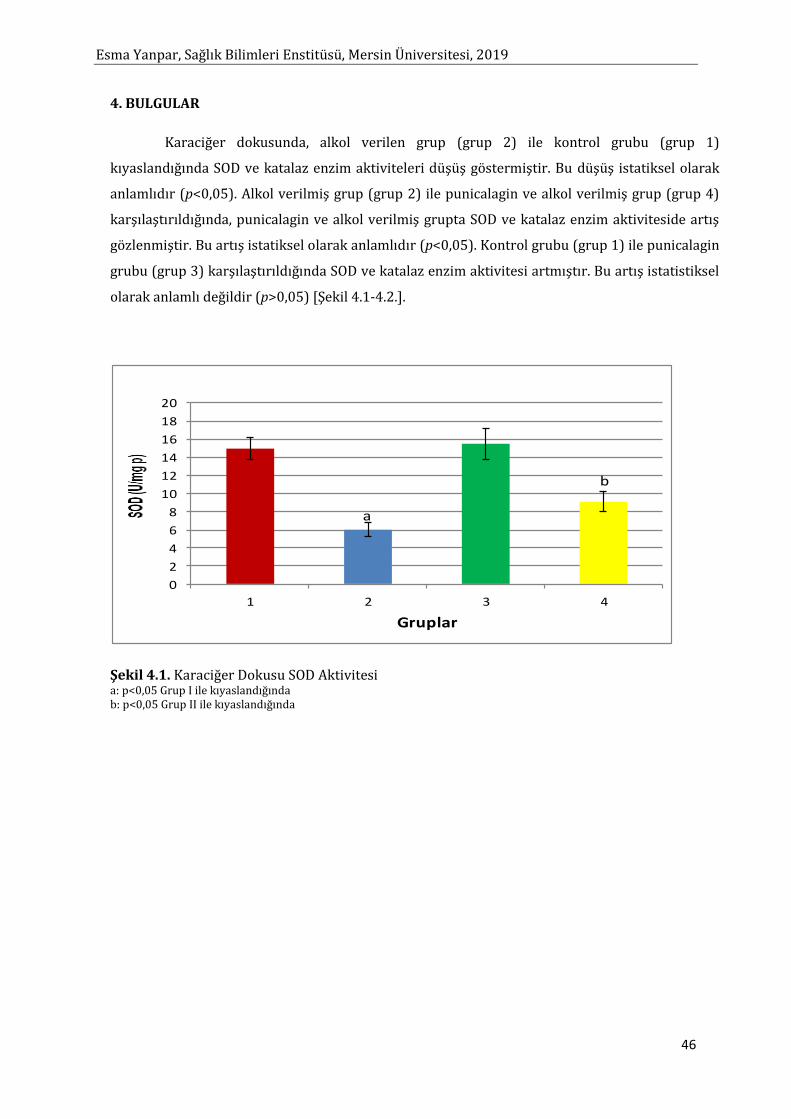
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
46
4. BULGULAR
Karaciğer dokusunda, alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
kıyaslandığında SOD ve katalaz enzim aktiviteleri düşüş göstermiştir. Bu düşüş istatiksel olarak
anlamlıdır (p<0,05). Alkol verilmiş grup (grup 2) ile punicalagin ve alkol verilmiş grup (grup 4)
karşılaştırıldığında, punicalagin ve alkol verilmiş grupta SOD ve katalaz enzim aktiviteside artış
gözlenmiştir. Bu artış istatiksel olarak anlamlıdır (p<0,05). Kontrol grubu (grup 1) ile punicalagin
grubu (grup 3) karşılaştırıldığında SOD ve katalaz enzim aktivitesi artmıştır. Bu artış istatistiksel
olarak anlamlı değildir (p>0,05) [Şekil 4.1-4.2.].
Şekil 4.1. Karaciğer Dokusu SOD Aktivitesi a: p<0,05 Grup I ile kıyaslandığında b: p<0,05 Grup II ile kıyaslandığında
0
2
4
6
8
10
12
14
16
18
20
1 2 3 4
Gruplar
a
b
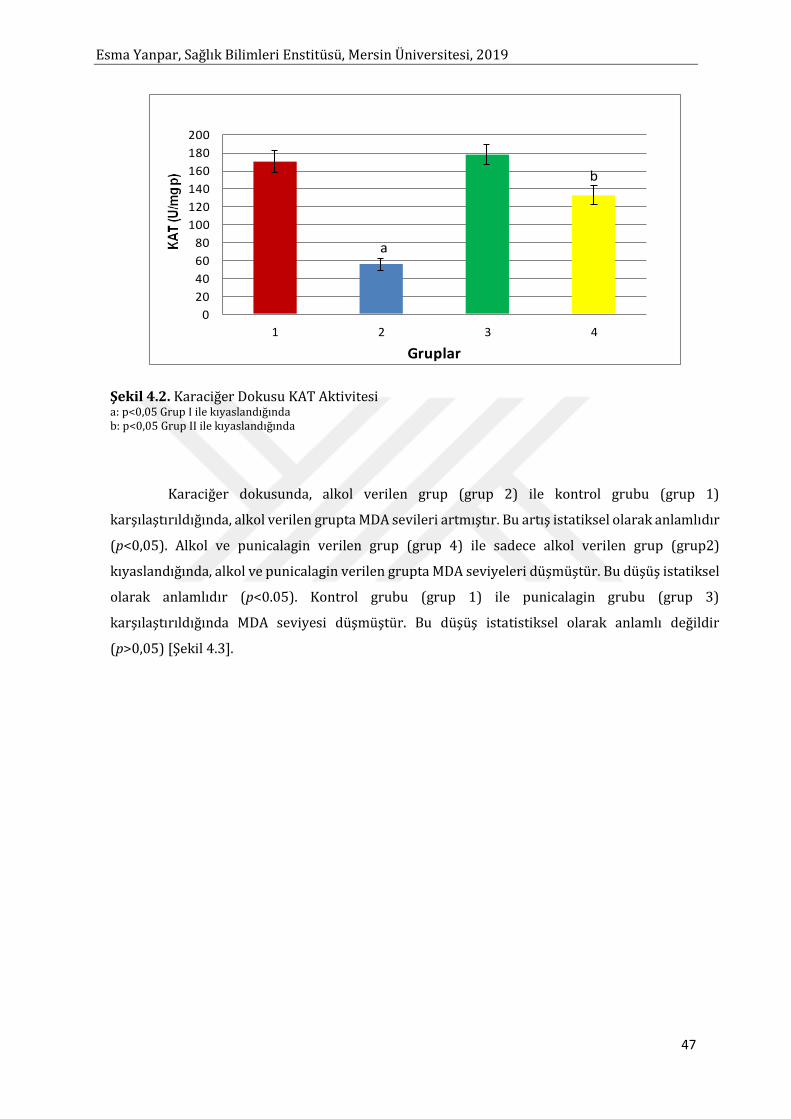
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
47
Şekil 4.2. Karaciğer Dokusu KAT Aktivitesi a: p<0,05 Grup I ile kıyaslandığında b: p<0,05 Grup II ile kıyaslandığında
Karaciğer dokusunda, alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
karşılaştırıldığında, alkol verilen grupta MDA sevileri artmıştır. Bu artış istatiksel olarak anlamlıdır
(p<0,05). Alkol ve punicalagin verilen grup (grup 4) ile sadece alkol verilen grup (grup2)
kıyaslandığında, alkol ve punicalagin verilen grupta MDA seviyeleri düşmüştür. Bu düşüş istatiksel
olarak anlamlıdır (p<0.05). Kontrol grubu (grup 1) ile punicalagin grubu (grup 3)
karşılaştırıldığında MDA seviyesi düşmüştür. Bu düşüş istatistiksel olarak anlamlı değildir
(p>0,05) [Şekil 4.3].
0
20
40
60
80
100
120
140
160
180
200
1 2 3 4
Gruplar
a
b
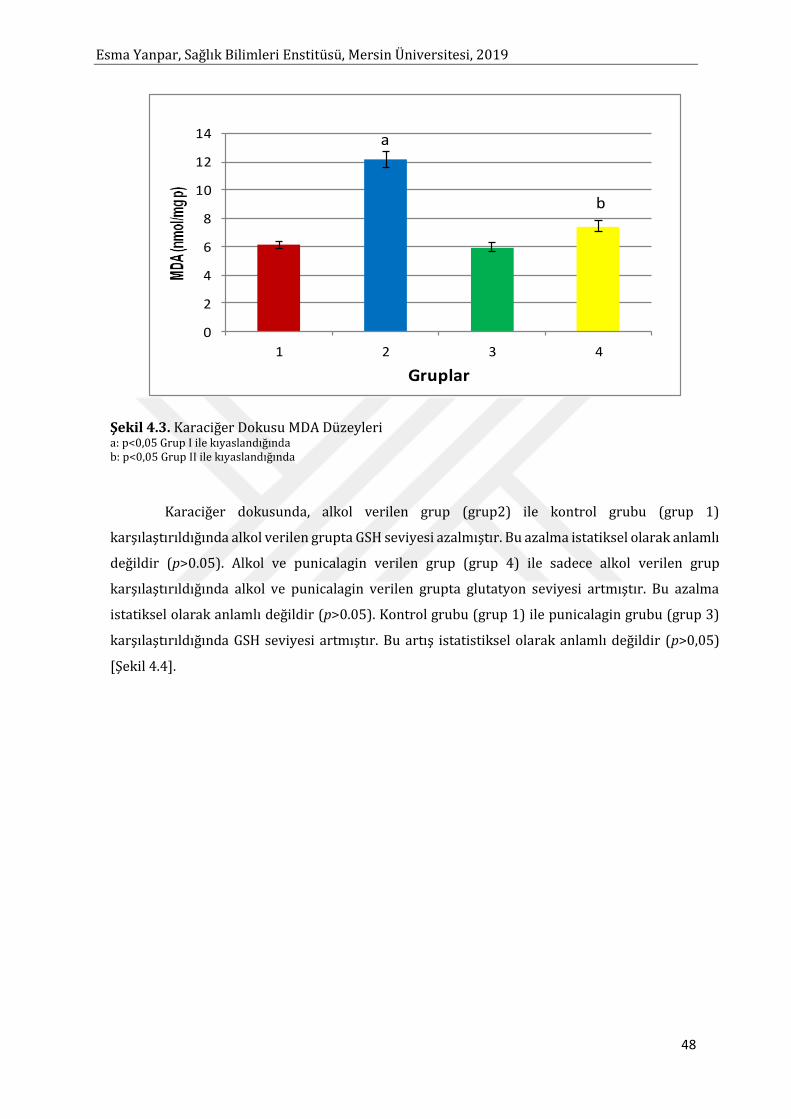
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
48
Şekil 4.3. Karaciğer Dokusu MDA Düzeyleri a: p<0,05 Grup I ile kıyaslandığında b: p<0,05 Grup II ile kıyaslandığında
Karaciğer dokusunda, alkol verilen grup (grup2) ile kontrol grubu (grup 1)
karşılaştırıldığında alkol verilen grupta GSH seviyesi azalmıştır. Bu azalma istatiksel olarak anlamlı
değildir (p>0.05). Alkol ve punicalagin verilen grup (grup 4) ile sadece alkol verilen grup
karşılaştırıldığında alkol ve punicalagin verilen grupta glutatyon seviyesi artmıştır. Bu azalma
istatiksel olarak anlamlı değildir (p>0.05). Kontrol grubu (grup 1) ile punicalagin grubu (grup 3)
karşılaştırıldığında GSH seviyesi artmıştır. Bu artış istatistiksel olarak anlamlı değildir (p>0,05)
[Şekil 4.4].
0
2
4
6
8
10
12
14
1 2 3 4
Gruplar
a
b
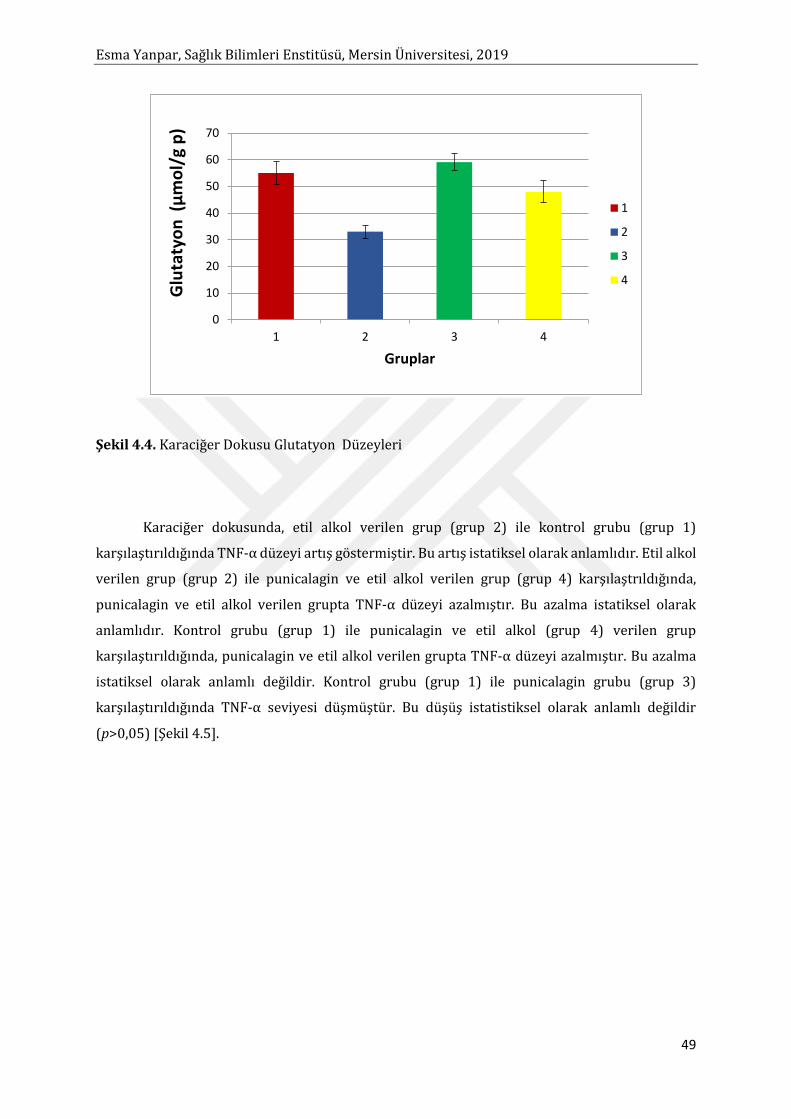
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
49
Şekil 4.4. Karaciğer Dokusu Glutatyon Düzeyleri
Karaciğer dokusunda, etil alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
karşılaştırıldığında TNF-α düzeyi artış göstermiştir. Bu artış istatiksel olarak anlamlıdır. Etil alkol
verilen grup (grup 2) ile punicalagin ve etil alkol verilen grup (grup 4) karşılaştrıldığında,
punicalagin ve etil alkol verilen grupta TNF-α düzeyi azalmıştır. Bu azalma istatiksel olarak
anlamlıdır. Kontrol grubu (grup 1) ile punicalagin ve etil alkol (grup 4) verilen grup
karşılaştırıldığında, punicalagin ve etil alkol verilen grupta TNF-α düzeyi azalmıştır. Bu azalma
istatiksel olarak anlamlı değildir. Kontrol grubu (grup 1) ile punicalagin grubu (grup 3)
karşılaştırıldığında TNF-α seviyesi düşmüştür. Bu düşüş istatistiksel olarak anlamlı değildir
(p>0,05) [Şekil 4.5].
0
10
20
30
40
50
60
70
1 2 3 4
Gruplar
1
2
3
4
Glu
taty
on
(µ
mo
l/g
p)
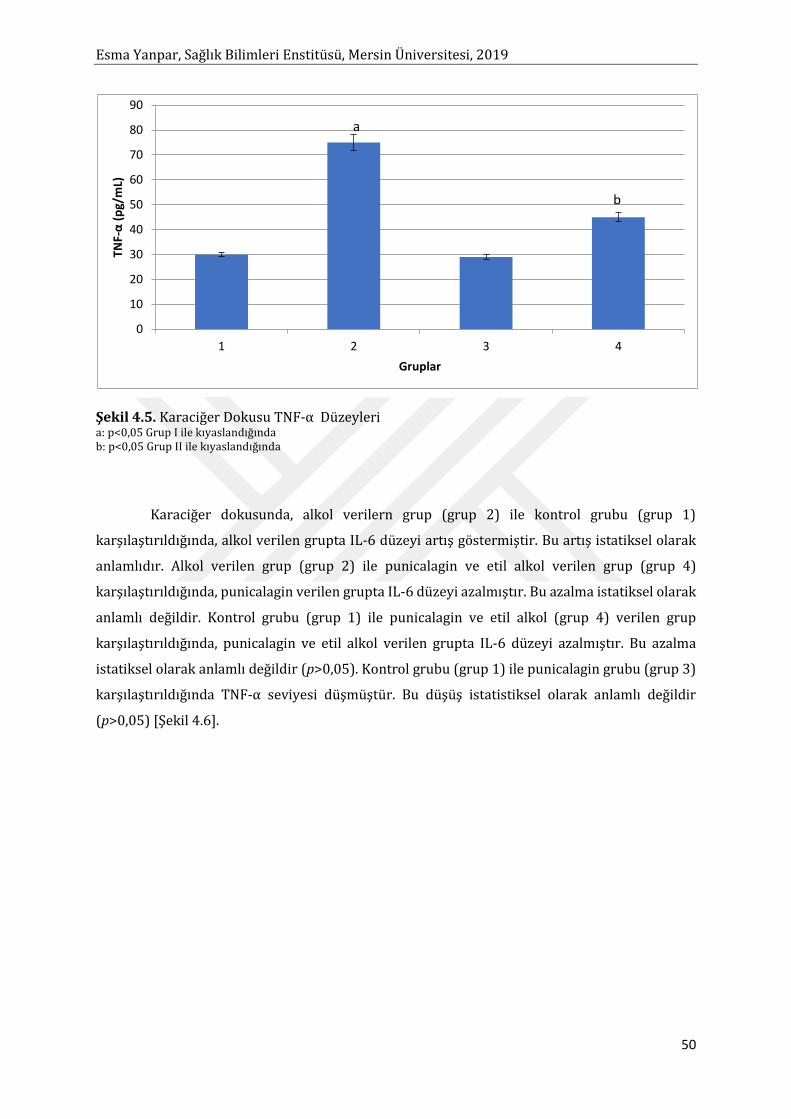
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
50
Şekil 4.5. Karaciğer Dokusu TNF-α Düzeyleri a: p<0,05 Grup I ile kıyaslandığında b: p<0,05 Grup II ile kıyaslandığında
Karaciğer dokusunda, alkol verilern grup (grup 2) ile kontrol grubu (grup 1)
karşılaştırıldığında, alkol verilen grupta IL-6 düzeyi artış göstermiştir. Bu artış istatiksel olarak
anlamlıdır. Alkol verilen grup (grup 2) ile punicalagin ve etil alkol verilen grup (grup 4)
karşılaştırıldığında, punicalagin verilen grupta IL-6 düzeyi azalmıştır. Bu azalma istatiksel olarak
anlamlı değildir. Kontrol grubu (grup 1) ile punicalagin ve etil alkol (grup 4) verilen grup
karşılaştırıldığında, punicalagin ve etil alkol verilen grupta IL-6 düzeyi azalmıştır. Bu azalma
istatiksel olarak anlamlı değildir (p>0,05). Kontrol grubu (grup 1) ile punicalagin grubu (grup 3)
karşılaştırıldığında TNF-α seviyesi düşmüştür. Bu düşüş istatistiksel olarak anlamlı değildir
(p>0,05) [Şekil 4.6].
0
10
20
30
40
50
60
70
80
90
1 2 3 4
TNF-
α(p
g/m
L)
Gruplar
b
a
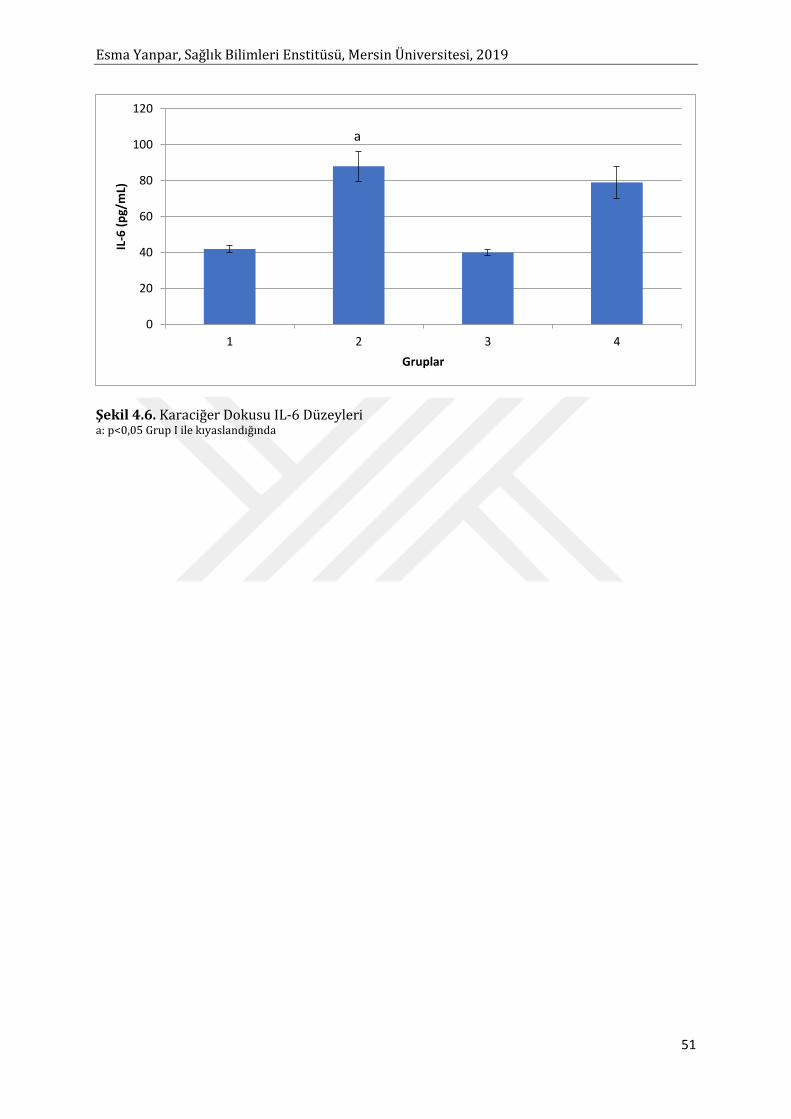
Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
51
Şekil 4.6. Karaciğer Dokusu IL-6 Düzeyleri a: p<0,05 Grup I ile kıyaslandığında
0
20
40
60
80
100
120
1 2 3 4
IL-6
(p
g/m
L)
Gruplar
a

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
52
5. TARTIŞMA VE SONUÇ
Zararlı kimyasal maddelerin çevre kirliliği, endüstri atıkları, güneş ışınları, egzoz gazları,
ağır metaller, sigara, alkol, ozon ve çeşitli zararlıların salınmasıyla serbest radikallerin meydana
gelmesi arasındaki ilişki doğru orantılı olarak artmaktadır. Biyolojik sistemlerde bu faktörlerin
etkisi ile serbest radikaller meydana gelmektedir. Bu durumda antioksidan savunma sisteminin
yetersiz kalabildiği durumlar da olabilmektedir ve bu şartlar altında çeşitli hastalıkların meydana
gelmesi beklenilen bir sonuçtur. Diyabet, kalp hastalıkları, romatoid artrit, kas, deri, göz
hastalıkları, hipertansiyon, psoriasis, kanser gibirhatsızlıklarda serbest radikal ve lipid
peroksidasyon ürünlerinin oluşumunda artma sonucu, antioksidan savunma sistemlerinin
yetersiz kaldığı saptanmıştır. Bu rahatsızlıkların patogenezisinde serbest radikallerin hastalığın
nedeni mi yoksa doğal bir sonucu olarak mı ortaya çıktıkları kesin olarak bilinmemekle birlikte,
hasta dokularda yoğun bir şekilde oksijenin varlığı gözlenmiştir [187-189].
Yapılan deneysel çalışmalarda, serbest radikaller, lipid peroksidasyonu ve peroksidasyon
ürünleri ile kanser gelişimi arasında pozitif korelasyon olduğu ortaya konulmuştur. Birçok
ksenobiyotiğin biyolojik sistemlerdeki oksidatif hasarı arttırıp, kansere neden olduğu
bildirilmektedir. Bu kimyasal maddeler hücrenin antioksidan savunmasında hızlı ve daimi bir
düşüşe neden olur. Serbest radikaller, kanserin her döneminde etkilidir. Fakat bu etki ilerleme
döneminde daha kuvvetlidir. Kanserin diğer dönemlerinde ise kısmen daha az görülmektedir.
Serbest radikaller kromozom ve DNA’da yapısal bozulmalara ve onkogenlerde aktivasyon
artışına sebep olmaktadır [54, 190].
Reaktif oksijen türleri ve diğer serbest radikallerin oluşması biyolojik sistemlerdeki
fizyolojik fonksiyonlar ve hücresel redoks olaylarının bir sonucu olmaktadır. Meydana gelen
serbest radikaller vücuttaki antioksidan savunma sistemleri tarafından yok edilmektedir [191].
Organizmada meydana gelen serbest radikaller ile antioksidan savunma sistemi arasında bir
denge bulunmaktadır. Serbest radikal üretiminin artması veya antioksidan savunma
sistemlerinin yetersiz kalması durumunda bu dengenin serbest radikaller yönünde kaydığı
gözlenmektedir. Bu bozulan denge sonucunda oksidatif stres meydana gelir ve bu da biyolojik
sistemlerde hasarlara neden olur [191, 192].
Oksidatif etki genelde proteinlerin modifikasyonu, mitokondride fonksiyon bozukluğu
oluşması ve lipid peroksidasyonu ile DNA’daki harabiyetinin artması şeklinde kendini gösterir.
Ksenobiyotikler önemli bir serbest radikal kaynağıdır. Önemli bir ksenobiyotik olan pestisitlerin
sebep olduğu toksik etkiler ile serbest radikal oluşumunun arasında pozitif bir korelasyon olduğu
bilinmektedir [193].

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
53
Bu çalışmada oksidatif stresi oluşturmak için etil alkol seçilmiştir. Etil alkol seçilmesinin
sebebi, alkolün oral yoldan canlı bünyesine kolaylıkla girebilmesi ve canlılarda deneysel
karaciğer harabiyeti ve oksidatif stresi oluşturmak amacıyla yaygın olarak kullanılan peroksidant
özellikte bir madde olmasıdır [194-197]. Yüksek miktarda alkol tüketiminin karaciğer dokusunda
harabiyete sebep olduğu yapulan çalışmalarca desteklenmiştir ve bu olumsuzlukların alınan doza
ve alınma süresine, diyete, bireysel farklılıklara ve diğer faktörlere bağlı olarak değişiklik
gösterdiği bildirilmiştir [198].
Ülkemizde çokça tüketilen etil alkolün ve nar (punica granatum) kabuğu ekstraktı olan
punicalagin etken maddesine subkronik uygulamalarına maruz bırakılan sıçanların karaciğer
dokusundan alınan örneklerde; etil alkolün sebep olduğu karaciğer toksikasyonuna karşı
punicalaginin antioksidan rolü ile inflamasyon üzerine etkisi olarak değerlendirilebilecek
biyobelirteçlerden, süperoksit dismutaz, katalaz enzim aktivitesine, malondialdehit ile glutatyon
seviyelerine ve TNF-α ile IL-6 düzeylerine bakılmıştır. Bu parametrelerin seçiliş nedeni ise, etil
alkolün moleküler toksisitesini ve bu fonksiyonel gıdaların gıda biyokimyası açısından sıcakkanlı
canlılar üzerindeki iyileştirici özelliklerini ortaya koymak açısından önem taşıdığı için tercih
edilmiştir.
Fareden sonra araştırmalarda en çok kullanılan omurgalı hayvan olan sıçanlar bu
çalışmada kullanılmıştır. Sıçanlar, temel tıp, ilaç, gıda, davranış ve toksisite çalışmalarında yaygın
olarak kullanılmaktadır [199].
Serbest radikallere karşı antioksidan korumada glutatyonun rolü büyüktür. Glutatyonun
asıl kaynağı kükürt ihtiva eden aminoasitlerden olan metiyonin ve sisteindir [200]. Çalışılan
karaciğer dokusunda düşük GSH düzeyinin, GSH’ın sentezinin yetersiz olmasından kaynaklandığı
düşünülmektedir. Bulgularımız bunu destekler niteliktedir.
Glutatyon, serbest radikal artışının doğal bir sonucu olarak meydana gelen lipid
peroksidasyon ürünleri ile hızlıca reaksiyona girerek biyolojik sistemler için zararlı olan lipid
peroksidasyon ürünlerinin ortamdan uzaklaştırılmasında rol alan önemli ve güçlü bir
antioksidandır. Glutatyon lipid peroksidasyon ürünleri ile reaksiyona girerek okside glutatyona
dönüşür [197, 198]. GSH ve GSH metabolizmasından sorumlu antioksidan enzimler, reaktif
oksijen türevlerinin sebep olduğu hasar karşısında önemli bir savunma sağlar [201, 202]. GSH
antioksidan koruma özelliğinden dolayı hücre ve dokularda tüketildiği bilinmektedir. Reaktif
oksijen türevlerinin sebep olduğu oksidatif stresin azaltılması yönünde kullanıyor olmasından
dolayı GSH seviyesinde azalma olduğu düşünülmektedir. GSH’da görülen bu azalmaya bağlı
olarak oksidatif stres riskinide artma meydana gelmektedir [203-205]. Glutatyon, hücrelerin

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
54
serbest radikal kaynaklı oksidatif hasardan korunmanın yanında yabancı toksik bileşiklerin
metabolizasyonunda görev alan reaksiyonlarda da yer almaktadır [206].
Bizim çalışmamızda, karaciğer dokusunda, alkol verilen grup (grup2) ile kontrol grubu
(grup 1) karşılaştırıldığında alkol verilen grupta GSH seviyesi azalmıştır fakat bu azalma istatiksel
olarak anlamlı değildir. Alkol ve punicalagin verilen grup (grup 4) ile sadece alkol verilen grup
(grup 2) karşılaştırıldığında alkol ve punicalagin verilen grupta glutatyon seviyesi artmıştır. Bu
azalma istatiksel olarak anlamlı bulunmamıştır. Çalışmamızda GSH seviyesindeki artışın kimyasal
strese karşı antioksidan sistemlerin indüklenmesine bağlı gelişen adaptasyonun bir sonucu
olduğu düşünülmektedir.
Membran fosfolipidlerinde bulunan doymamış yağ asitlerinin oksijen ile reaksiyonu
sonucu lipid hidroperoksitleri oluşur, bu reaksiyona lipit peroksidasyonu reaksiyonu denir. Bu
reaksiyon zincirleri neticesinde MDA, konjuge dienler, bazı aldehitler, uçucu hidrokarbonlar gibi
ürünler meydana gelmektedir [54, 207]. Serbest radikaller, antioksidan sistemin koruyucu
etkisini aşacak şekilde çoğalırsa zararlı etkilerini en hassas bileşikler olan lipidler üzerinde
gösterirler. Membranların yapısında yer alan kolesterol ve doymamış fosfolipidler, serbest
radikallerle kolay ve hızlı bir şekilde reaksiyona girerek lipid peroksidasyonunu gerçekleştirir
[208, 209].
Bu çalışmada, karaciğer dokusunda, alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
karşılaştırıldığında, alkol verilen grupta MDA sevileri artmıştır ve bu artış istatiksel olarak
anlamlıdır. Alkol ve punicalagin verilen grup (grup 4) ile sadece alkol verilen grup (grup 2)
kıyaslandığında, alkol ve punicalagin verilen grupta MDA seviyeleri düşmüştür ve bu düşüş
istatiksel olarak anlamlı bulunmuştur. MDA’nın düşüşündeki istatiksel anlam, bize yaptığımız
çalışmada punicalaginin, lipit peroksidasyon reaksiyonlarına karşı koruyucu nitelikte olduğunu
işaret etmektedir.
Süperoksit dismutaz enziminin görevi, oksijen metabolizasyonunda sorumlu hücreleri
oluşan süperoksit radikalinin olumsuz etkilerinden korumaktır. Bu şekilde lipit peroksidasyon
reaksiyonlarını da inhibe etmiş olur. SOD enzim aktivitesi oksijen kullanımının yoğun olduğu
dokularda daha fazladır [210]. Oksidatif stresle doğru orantılı olarak SOD enzim aktivitesinde
artış gözlenmiştir [54]. SOD enzim aktivitesindeki azalmanın nedeninin uzun süreli lipit
peroksidasyon reaksiyonlarına bağlı olarak aşılan antioksidan koruma sistemin neden olduğu
azalmış enzim aktiviteleri olabileceği düşünülmektedir [202, 211].
Bu çalışmada, karaciğer dokusuna alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
kıyaslandığında SOD enzim aktiviteleri düşüş göstermiştir. Bu düşüş istatiksel olarak anlam teşkil

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
55
etmektedir. Alkol verilmiş grup (grup 2) ile punicalagin ve alkol verilmiş grup (grup 4)
karşılaştırıldığında, punicalagin ve alkol verilmiş grupta SOD enzim aktivitesinde artış
gözlenmiştir ve bu artış istatiksel olarak anlamlı bulunmuştur. Yapılan deneysel çalışma
sonucunda punicalaginin SOD enzim aktivitesini istatiksel olarak arttırması antioksidan
aktivitesini kanıtlar niteliktedir.
SOD enzim aktivitesi sonucu meydana gelen hidrojen peroksit radikali, katalaz enzim
aktivitesi ile oksijen ve suya dönüşmektedir [212]. Hidrojen peroksitin yüksek
konsantrasyonlarında katalaz enzimi oldukça aktif çalışır [213]. Hidrojen peroksit katalaz ve
glutatyon peroksidaz enzim aktivitesi ile ortamdan uzaklaştırılır. Glutatyon peroksidaz düşük
konsantrasyondaki detoksifikasyondan sorumlu iken, katalaz yüksek konsantrasyondaki
detoksifikasyondan sorumludur [214].
Bu çalışmada, karaciğer dokusunda, alkol verilen grup (grup 2) ile kontrol grubu (grup 1)
kıyaslandığında katalaz enzim aktiviteleri düşüş göstermiştir. Bu düşüş istatiksel olarak anlam
teşkil etmektedir. Alkol verilmiş grup (grup 2) ile punicalagin ve alkol verilmiş grup (grup 4)
karşılaştırıldığında, punicalagin ve alkol verilmiş grupta katalaz enzim aktivitesinde artış
gözlenmiştir ve bu artış istatiksel olarak anlamlı bulunmuştur. Yapılan deneysel çalışma
sonucunda punicalaginin katalaz enzim aktivitesini istatiksel olarak arttırması antioksidan
aktivitesini destekler niteliktedir.
Alkolün sebep olduğu TNF-α üretimi, endotoksinler ve oksidatif stres aracılı olarak
gerçekleşmektedir. Bununla birlite endotoksinler ve oksidatif stres arasındaki arasındaki neden-
sonuç ilişkisi net olarak aydınlatılamamıştır. Yapılan bazı çalışmalarda akut alkol tüketimi
sonrasında plazma endotoksin seviyesinde yükselmeyi, TNF-α üretimi ve hepatik oksidatif stres
arasındaki bağlatıyı belirtmek amacıyla, alkol hepatotoksisitesi oluşturulan sıçanlar kullanılarak
hepatik TNF-α üretimi ile hepatik lipid peroksidasyonu ve plazma endotoksin ile ilişki içinde
olduğunu göstermişlerdir [215].
Bizim çalışmamızda, karaciğer dokusunda, etil alkol verilern grup (grup 2) ile kontrol
grubu (grup 1) karşılaştırıldığında TNF-α düzeyi artış göstermiştir. Bu artış istatiksel olarak
anlamlıdır. Etil alkol verilen grup (grup 2) ile punicalagin verilen grup (grup 3)
karşılaştrıldığında, punicalagin verilen grupta TNF-α düzeyi azalmıştır. Bu azalma istatiksel
olarak anlamlıdır. Bu istatiksel azalma, punicalaginin antiinflamatuvar özellikte olduğunu
göstermektedir. Yani punicalaginin, inflamasyon belirteçlerinden biri olan TNF-α üzerine
etkisinin antiinflamatuvar özelliğini destekler nitelikte olduğunu söyleyebiliriz.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
56
Sitokinlerin alkolik karaciğer hastalığındaki rolünü konu alan birçok çalışma vardır.
Kronik etanol tüketiminden sonra oluşan ROS ve endotoksinler NF-кB aracılığıyla
proinflamatuvar sitokinlerin ekspresyonunu stimüle etmektedir. Alkolik hepatitte aktive olan T-
hücreleri ve makrofajlar etanolün aracılık ettiği karaciğer hasarının sürdürülmesinde önemli bir
role sahip olan TNF-α, IL-6, IL-1β ve IFN-γ’yı içeren sitokinleri salgılarlar. TNF-α fibroblast
proliferasyonunu uyararak sitotoksik lenfosit aktivitesi ile kollejenaz enzim aktivitesini arttırır.
Apoptoz ve nekroz gerçekleştirmektedir. Apaptoz ve nekrozun akabinde hücre ölümü meydana
gelir [216-219].
Bizim çalışmamızda, karaciğer dokusunda, alkol verilen grup (grup 2) ile kontrol grubu
(grup 1) karşılaştırıldığında, alkol verilen grupta IL-6 düzeyi artış göstermiştir. Bu artış istatiksel
olarak anlamlıdır. Alkol verilen grup (grup 2) ile punicalagin ve alkol verilen grup (grup 4)
karşılaştırıldığında, punicalagin ve alkol verilen grupta IL-6 düzeyi azalmıştır. Bu azalma
istatiksel olarak anlamlı değildir. Kontrol grubu (grup 1) ile punicalagin ve etil alkol verilen grup
(grup 4) karşılaştırıldığında, punicalagin ve etil alkol verilen grupta IL-6 düzeyi azalmıştır. Bu
azalma istatiksel olarak anlamlı değildir. Yani punicalaginin, inflamasyon belirteçlerinden biri
olan IL-6 üzerine etkisinin antiinflamatuvar özelliğini destekler nitelikte olmadığını
söyleyebiliriz.
Sonuç olarak, bu çalışmada elde edilen biyokimyasal verilerin doğrultusunda etanolün
neden olduğu serbest radikal üretiminin göstergesi olarak lipid peroksidasyon reaksiyonunun
son ürünü olan malondialdehid karaciğer dokusunda artarken, yine karaciğer dokusunda
antioksidan savunma sistemi biyobelirteçlerinde dalgalanmalara neden olmuştur. Punicalagin ve
etil alkol verilmiş rat karaciğeri ile sadece alkol verilmiş sıçan karaciğeri karşılaştırıldığında MDA
düzeyinde meydana gelen anlamlı düşüş bize punicalaginin lipit peroksidasyonunu
engelleyebildiğini göstermiştir. Etil alkol ve punicalagin verilmiş sıçan karaciğerinde SOD ve KAT
seviyesi ise sadece alkol verilmiş sıçan karaciğerine göre anlamlı olarak artmıştır. Bu artış bize
punicalaginin antioksidan aktivitesinin olduğunu gösterir niteliktedir. Etil alkolün sebep olduğu
oksidatif stres sonucu olarak karaciğer dokusunda artan malondialdehit punicalagin uygulaması
ile iyileştirdiği ve yine karaciğer dokusunda dalgalanan antioksidan savunma sistemi
biyobelirteçlerinde de etil alkol grubuna göre iyileştirme sağladığı tespit edilmiştir. Punicalaginin
inflamasyon belirteçlerinden olan TNF-α ve IL-6 üzerine etkisine bakıldığında, punicalagin ve
alkol verilen sıçan karaciğeri ile sadece alkol verilen şıçan karaciğeri karşılaştırıldığında TNF-
α’nın istatiksel olarak anlamlı olacak şekilde düşmüş olduğu saptanmıştır. Bu durum
punicalaginin antiinflamatuvar özelliğinden ileri gelebilir. Çalışmamızın sonucu olarak,
punicalaginin alkol toksisitesine bağlı oluşabilecek hastalıkların tedavisinde alternatif bir yöntem
olarak kullanılabileceği söylenebilir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
57
Literatürde punicalaginin antioksidan ve antiinflamtuvar özelliğini destekleyen
çalışmalar vardır. Lansky ve arkadaşlarının [220] yapmış olduğu çalışmada, narın kanser ve kalp
hastalıklarının önlenmesine sebep olduğu, AIDS için kullanılan yiyecekler sınıfında yer alan
önemli ve güçlü bir antioksidan olduğu belirtilmiştir. Aviram ve arkadaşları [221] yaptıkları
araştırmada, nar içerisinde bulunan ellagik asitin antioksidan, antiinflamatuvar, antiöstrojenik,
antikanserojenik ve antimutajenik etkilerinin bulunduğu belirtilmiştir. Priyadarsini ve
arkadaşları [222] yapmış oldukları araştırmada, ellagik asitin serbest radikallere bağlanmak
suretiyle etkileşime girerek, oksidatif harabiyete ve serbest radikallerin neden olduğu bazı
rahatsızlıklara karşı organizmayı koruduğu bildirilmektedir. Fischer ve arkadaşlarının [223]
yapmış olduğu çalışmada, narın kurusu, kabuğu ve çekirdeklerinde yüksek miktarda fenolik
içeriğin bulunduğu ve bu fenolik bileşenler arasında punicalaginin antioksidan aktiviteden asıl
sorumlu olan molekül olduğu belirtilmiştir.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
58
KAYNAKLAR
[1]. Gürsoy, F., Etanolün İndüklediği Karaciğer Hasarında Matriks Metalloproteinazların Rolü ve Antioksidanların Etkisi, Yüksek Lisans Tezi, Afyon Kocatepe Üniversitesi Sağlık Bilimleri Enstitüsü , Afyonkarahisar, 2005.
[2]. Adachi, M.; Higuchi, H.; Miura, S.; Azuma, T.; Inokuchi, S.; Saito, H.; Kato, S.; Ishii, H., Bax interacts with voltage-dependent anion channel and mediates ethanol-induced apoptosis in rat hepatocytes. American Journal of Physiology 2004, 287, 695–705.
[3]. Yan, L. J.; Sohal, R. S., Mitochondrial adenine nucleotide translocase is modified oxidatively during aging. Proceedings of the National Academy of Sciences 1998, 95, 896-901.
[4]. Yan L. J., Positive oxidative stress in aging and aging-related disease tolerance. Redox Biology 2014, 9, 165-169.
[5]. Dokuyucu, R.; Karateke, A.; Gökce, H.; Kurt, R. K.; Özcan, O.; Öztürk S, Antioxidant effect of erdosteine and lipoic acid in ovarian ischemia-reperfusion injury. European Journal of Obstetrics & Gynecology and Reproductive Biology 2014, 183, 23-27.
[6]. Yalın, A. E.; Yalın, S.; Yıldız, Ş. M.; Yüzbaşıoğlu, S.; Aksoy, K., Eritrosit membran proteinlerine serbest radikallerin etkisi. Türk Biyokimya Dergisi 2001, 26, 123-129.
[7]. Bagchi, K.; Puri, S., Free radicals and antioxidants in health and disease. Eastern Mediterranean Health Journal 1998, 4, (2), 350-360.
[8]. Duman, A., D.; Ozgen, M.; Dayisoylu, K. S.; Erbil, N.; Durgac, C., Antimicrobial Activity of Six Pomegranate (Punica granatum L.) Varieties and Their Relation to Some of Their Pomological and Phytonutrient Characteristics. Molecules, 2009, 14, (5), 1808-1817.
[9]. Durgac, C.; Özgen, M.; Şimşek, Ö.; Kaçar, Y., A.; Kıyga, Y.; Çelebi, S.; Gündüz, K.; Serçe, S., Molecular and Pomological Diversity Among Pomegranate (Punica granatum L.) Cultivars in Eastern Mediterranean Region of Turkey. African Journal of Biotechnology 2008, 7, (9), 1294-1301.
[10]. Ekşi, A.; Ozhamamci, I., Chemical Composition and Guide Values of Pomegranate Juice. Gıda, 2009, 34, 265-270.
[12]. Bast, A.; Haenen, G.; Goelmen, J. A., Oxidants and antioxidants. State of the articles Am J Medicine 1991, 91, (3), 2-13.
[13]. Halliwell, B.; Gutteridge, J. M. C., The Importance of Free Radicals and Catalytic Metal Ions In Human Diseases. Molecular Aspects of Medicine 1985, 8, (2), 89-193.
[14]. Nawar, W. W., Lipids In “Food Chemistry”. 3rd ed.; Marcel Dekker: New York, 1996; p 225-319.
[15]. Halliwell, B., Antioxidant Defence Mechanisms: From The Beginning To The End (of the beginning). Free Radical Research 1999, 31, (4), 261-272.
[16]. Valko, M.; Leibfritz, D.; Moncola, J., Free radicals and antioxidants in normal physiological functions and human disease. The İnternational Journal Of Biochemistry & Cell Biology 2007, 39, 44-84.
[17]. Pham-Huy, L. A.; He, H.; Pham-Huyc, C., Free Radicals, Antioxidants in Disease and Health. International Journal of Biomedical Science 2008, 4, (2), 89-96.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
59
[18]. Cankurtaran, M., Yaşlılık, yaşlanma mekizmaları, antiaging ve yaşam tarzı değişiklikleri, 7. Ulusal İç Hastalıkları Kongresi, Kongre Kitapçığı. 2005, pp. 1-5.
[19]. Muller, F. L,; Liu, Y.; Van Remmen, H., Complex III releases superoxide to both sides of the inner mitochondrial membrane. Journal of Biological Chemistry 2004, 279, (47), 49064-49073.
[20]. Becker, L. B.; Vanden Hoek, T. L.; Shao, Z. H.; Li, C. Q,.; Schumacker, P. T., Generation of superoxide in cardiomyocytes during ischemia before reperfusion. The American Journal Of Physiology 1999, 277, 2240-2246.
[21]. Valko, M.; Morris, H.; Cronin, M. T., Metals, toxicity and oxidative stress. Current Medicinal Chemistry 2005, 12, 1161-1208.
[22]. Wright, A.; Bubb, W. A.; Hawkins, C. L.; Davies, M. J., Singlet oxygen-mediated protein oxidation: evidence for the formation of reactive side chain peroxides on tyrosine residues. Photochemistry and Photobiology 2002, 76, 35-46.
[23]. Mailloux, R. J., Teaching the fundamentals of electron transfer reactions in mitochondria and the production and detection of reactive oxygen species. Redox Biology 2015, 4, 381-398.
[24]. Lloyd, R. V.; Hanna, P. M.; Mason, R. P., The origin of the hydroxyl radical oxygen in the Fenton reaction. Free Radical Biology and Medicine 1997, 22, 885-888.
[25]. Betteridge, D. J., What is oxidative stress?. Metabolism 2000, 49, (2), 3-8.
[26]. Halliwell, B., Oxidants and human disease: some new concepts. FASEB Journal 1987, 1, (5), 358-364.
[27]. Halliwell, B., Antioxidant defence mechanisms: from the beginning to the end (of the beginning). Free Radical Research 1999, 31, (4), 261-272.
[28]. Lloyd, R. V.; Hanna, P. M.; Mason, R, P., The origin of the hydroxyl radical oxygen in the Fenton reaction. Free Radical Biology and Medicine 1997, 22(5), 885-888.
[29]. Moncada, S.; Palmer, R. M.; Higgs, E. A., Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacological Reviews 1991, 43, (2), 109-142.
[30]. Jomova, K.; Valko, M., Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, (2-3), 65-87.
[31]. Rayner, B. S.; Hua, S.; Sabaretnam, T.; Witting, P. K., Nitric oxide stimulates myoglobin gene and protein expression in vascular smooth muscle. Biochemical Journal 2009, 423, (2), 169-77.
[32]. Dawson, W. l.; Dawson, T. M.; London, E. D.; Bradt, D. S.; Snyder, S. H., Nitric Oxyde Mediates Glutamate Neurotoxicity in Primary Cortical Cultures. Proceedings of the National Academy of Sciences 1991, 88, (14), 6868-6871.
[33]. Andus, T.; Bauer, J.; Gerok, W. , Effect of ctokines on the liver. Hepatology 1991, 13, (2), 364-375.
[34]. Beckman, J. S.; Beckman, T. W.; Chen, J.; Marshall, P. A.; Freeman, B. A., Apparent Hydroxyl Production By Peroxynitrite: Implication for Endothelial Injury From Nitric Oxide and Super Oxide. Proceedings of the National Academy of Sciences 1990, 87, 1620-1624.
[35]. Ischiropoulos, H.; Zhu, L.; Beckman, J. S., Peroxynitrite formation from macrophage-derived nitric oxide. Archives of Biochemistry and Biophysics 1992, 298, (2), 446-51.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
60
[36]. Huıe, R. T.; Padmaja, S., The reaction of NO with superoxide. Free Radical Research Communications 1993, 18, (4), 195-199.
[37]. Chan, P. H., Role of oxidants in ischemic brain damage. Stroke 1996, 27, (6), 1124-1129.
[38]. Beckman, J. S.; Carson, M.; Smıth, C. D.; Koppenol, W. H., ALS, SOD and peroxynitrite. Nature 1993, 18, 195-199.
[39]. Patel, R. K.; Mcadrew, J.; Sellak, H.; Whıte, C. R.; Jo, H.; Freeman, B. A.; Darley-Usmar, V. M., Biological aspects of reactive nitrogen series. Biochimica et Biophysica Acta 1999, 1411, 385-400.
[40]. Gökpınar, S.; Koray, T.; Akçiçek, E.; Göksan, T.; Durmaz, Y., Algal antioksidanlar. Ege Üniversitesi Su Ürünleri Dergisi 2006, 23, (1), 85-89.
[41]. Wenzel, U.; Nickel, A.; Kuntz, S.; Daniel, H., Ascorbic asid suppresses drug induced apoptozis in human colon cancer cells by scavenging mitochondrial süperoxide anions. Carcinogenesis 2004, 25, (5), 703-712.
[42]. Çaylak, E., Hayvan ve bitkilerde oksidatif stres ile antioksidanlar. Tıp Araştırmaları Dergisi 2011, 9, (1), 73-83.
[43]. Trimarchi, J. R.; Liu, L.; Porterfield, D. M.; Smith, P. J.; Keefe, D. L., Oxidative phosphorylation dependent and independent oxygen consumption by individual preimplantation mouse embryos. Biology of Reproduction 2000, 62, (6), 1866-1874.
[44]. Sies, H., Oxidative stress: from basic research to clinical application. The American Journal of Medicine 1991, 91, 31-38.
[45]. Gupta, R. K.; Patel, A. K.; Shah, N., Oxidative stress and antioxidants in disease and cancer: a review. Asian Pacific Journal of Cancer Prevention 2014, 15, (11), 4405-4409.
[46]. Gutteridge, J.M., Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clinical Chemistry 1995, 41, (12), 1819-1828.
[47]. Girotti, A. W., Lipid hydroperoxide generation, turnover, and effector action in biological systems. The Journal of Lipid Research 1998, 39, (8), 1529-1542.
[48]. Kanner, J.; German, J. B.; Kinsella, J. E., Initiation of lipid peroxidation in biological systems. Critical Reviews in Food Science and Nutrition 1987, 25, 317-364.
[49]. Esterbauer, H.; Schaur, R. J.; Zollner, H., Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radical Biology and Medicine 1991, 11, (1), 81-128.
[50]. Catalá, A., An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. International Journal of Biochemistry & Cell Biology 2006, 38, 1482-1495.
[51]. Prokai, L.; Yan, L. J.; Vera-Serrano, J. L., Mass spectrometry-based survey of age-associated protein carbonylation in rat brain mitochondria. Journal of Mass Spectrometry 2007, 42, (12), 1583-1589.
[52]. Rao, R. S.; Moller, I. M., Pattern of occurrence and occupancy of carbonylation sites in proteins. Proteomics 2011, 11, (21), 4166-4173.
[53]. Dalle-Donne, I.; Rossi, R.; Giustarini, D., Protein carbonyl groups as biomarkers of oxidative stress. Clinica Chimica Acta 2003, 329, (1-2), 23-38.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
61
[54]. Akkuş, İ., Serbest Radikaller ve Fizyopatolojik Etkileri. 1. Baskı; Mimoza Yayınları: Konya, 1995; s 34-54.
[55]. Kavas, G. Ö., Serbest Radikaller ve Organizma Üzerine Etkileri. Türkiye Klinikleri 1989, 9, (1), 1-8.
[56]. Breen, A. P.; Murphy, J. A., Reactions of oxyl radicals with DNA. Free Radical Biology and Medicine 1995, 18, (6), 1033-1077.
[57]. Helbock, H. J.; Beckman, K. B.; Ames, B. N., 8-Hydroxydeoxyguanosine and 8-Hydroxy-guanine as biomarkers of oxidative DNA damage. Methods Enzymol 1999, 300, 156-166.
[58]. Cooke, M. S.; Evans, M. D.; Dizdaroglu, M.; Lunec, J., Oxidative DNA damage: mechanisms, mutation, and disease. FASEB Journal 2003, 17, (10), 1195-214.
[59]. Hu, J. J.; Dubin, N.; Kurland, D.; Ma, B. L.; Roush, G. C., The effects of hydrogen peroxide on DNA repair activities. Mutation Research 1995, 336, (2), 193-201.
[60]. Hardie, L. J.; Briggs, J. A.; Davidson, L. A., The effect of hOGG1 and glutathione peroxidase I genotypes and 3p chromosomal loss on 8-hydroxydeoxyguanosine levels in lung cancer. Carcinogenesis 2000, 21, (2), 167-172.
[61]. Ercan, N.; Fidancı, U. R., Piyodermalı köpeklerde idrarda 8-hidroksi-2’-deoksiguanozin (8-OHdG) düzeyleri. Ankara Üniversitesi Veteriner Fakültesi Dergisi 2012, 59, (3), 163-168.
[62]. Gutteridge, J. M. C., Free radicals in disease processes: A compilation of cause and consequence. Free Radical Research 1993, 19, (3), 141–1583.
[63]. Pekcan, Z.; Çınar, M.; Gürkan, M.; Kumandaş, A., Ankara Keçilerinde Propofol ve İzofluran Anestezisinin Oksidatif Stres Üzerine Etkileri. Atatürk Üniversitesi Veteriner Bilimleri Dergisi 2011, 6, (3), 217-222
[64]. Basaga, H. S., Biochemical aspects of free radicals. Biochemistry and Cell Biology 1990, 68, (7-8) 989-998.
[65]. Gutteridge, J. M. C., Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clinical Chemistry 1995, 41, (12), 1819-1828
[66]. Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.; Mazur, M.; Telser, J., Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry & Cell Biology 2007, 39, (1), 44–84.
[67]. Traber, M. G.; Packer, L., Vitamin E. Beyond antioxidant function. The American Journal of Clinical Nutrition 1995, 62, (6), 1501-1509
[68]. Şener, G.; Yeğen, B. C.; Berrak, Ç., İskemi Reperfüzyon Hasarı. Klinik Gelişim Dergisi 2009, 22, 5-13.
[69]. Dündar, Y.; Aslan, R., Hücre Moleküler Statüsünün Anlaşılması ve Fizyolojik Önem Açısından Radikaller, Antioksidanlar. İnsizyon Cerrahi Tıp Bilim Dergisi 1999, 2, (2), 134-142.
[70]. Pham-Huy, L. A.; He, H.; Pham-Huy, C., Free radicals, antioxidants in disease and health. The International Journal of Biomedical Science 2008, 4, (2), 89-96.
[71]. Sen, S.; Chakraborty, R.; Sridhar, C.; Reddy, Y. S. R.; De, B., Free radicals, antioxidants, diseases and phytomedicines: Current status and future prospect. International Journal of Pharmaceutical Sciences and Research 2010, 3, (1), 91-100.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
62
[72]. Aydemir, B.; Karadağ Sarı, E., Antioksidanlar ve büyüme faktörleri ile ilişkisi. Kocatepe Veterinary Journal 2009, 2, (2), 56-60.
[73]. Young, I. S.; Woodside, J. V., Antioxidants in Health and Disease. Journal of Clinical Pathology 2001, 54, (3), 176-186.
[74]. Sen, S.; Chakraborty, R., The Role of Antioxidants in Human Health. Oxidative Stress: Diagnostics, Prevention and Therapy, American Chemical Society, Washington DC, 2011, Chapter 1: pp. 1-37.
[75]. Mruk, D. D.; Silvestrini, B.; Meng-Yun, M. O.; Cheng, C. Y., Antioxidant superoxide dismutase - a review: its function, regulation in the testis, and role in male fertility. Contraception 2002, 65, (4), 305-311.
[76]. Nordberg, J.; Arner, S. J., Reactive oxygen species, antioxidant, and the mammalian thioredoxin system. Free Radical Biology and Medicine 2001, 31, (11), 1287-1312.
[77]. Fridovich, I., Superoxide Radical and Superoxide Dismutase. Annual Review of Biochemistry 1995, 64, (1), 97-112.
[78]. Halliwell, B.; Gutteridge, J.M.C., Free Radicals in Biology and Medicine. 3rd ed.; Oxford University Press: New York, 1999; p 10-121.
[79]. Gao, F.; Kinnula, V. L.; Myllärniemi, M.; Oury, T. D., Extracellular Superoxide Dismutase in Pulmonary Fibrosis. Antioxidants & Redox Signaling 2008, 10, (2), 343-354.
[80]. Kirkman, H. N.; Galiano, S.; Gaetani, G. F., The function of catalase-bound NADPH. Journal of Biological Chemistry 1987, 262, (2), 660–666.
[81]. Zamocky, M.; Koller, F., Understanding the structure and function of catalases: clues from molecular evolution and in vitro mutagenesis. Biophysics & Molecular Biology 1999, 72, (1), 19-66.
[82]. Limon-Pacheco, J.; Gonsebatt, M. E., The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutation Research 2009, 674, (1-2), 137-147.
[83]. Reiter, R. J.; Melchiorri, D.; Sewerynek, E.; Poeggeler, B.; Barlow-Walden, L.; Chuang, J.; Ortiz, G. G.; Acuna-Castroviejo, D. A., Review of the evidence supporting melatonin‘s role as an antioxidant. Journal of Pineal Research 1995, 18, (1), 1-11.
[84]. Sen, S.; Chakraborty, R.; Sridhar, C.; Reddy, Y. S. R.; De, B., Free radicals, antioxidants, diseases and phytomedicines: Current status and future prospect. International Journal of Pharmaceutical Sciences and Research 2010, 3, (1), 91-100.
[85]. Özkan, A.; Fışkın, K., Serbest oksijen radikalleri, karsinogenez ve antioksidant enzimler. Türk Hematoloji Onkoloji Dergisi 2004, 14, 52-60.
[86]. Hayes, J. D.; Pulford, D. J., The glutathione S-transferase supergene family: regulation of GST and the contribution of the isoenzymes to cancer chemoprotection and drug resistance. Critical Reviews in Biochemistry and Molecular Biology 1995, 30, (6), 445-600.
[87]. Townsend, D. M.; Tew, K. D.; Tapiero, H., The importance of glutathione in human disease. Biomedicine & Pharmacotherapy 2003, 57, (3-4), 145-155.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
63
[88]. Green, R. M.; Graham, M.; O’Donovan, M. R.; Chipman, J. K.; Hodges, N. J., Subcellular compartmentalization of glutathione: correlations with parameters of oxidative stress related to genotoxicity. Mutagenesis 2006, 21, (6), 383-390.
[89]. Lagman, M.; Ly, J.; Saing, T.; Kaur Singh, M.; Vera Tudela, E.; Morris, D., Investigating the causes for decreased levels of glutathione in individuals with type II diabetes. PLOS ONE 2015, 10, (3), 1-19.
[90]. Pei, S.; Minhajuddin, M.; Callahan, K. P.; Balys, M.; Ashton, J. M.; Neering, S. J.; Lagadinou, E. D.; Corbett, C.; Ye, H.; Liesveld, J. L.; O’Dwyer, K. M.; Li, Z.; Shi, L.; Greninger, P.; Settleman, J.; Benes, C.; Hagen, F. K.; Munger, J.; Crooks, P. A.; Becker, M. W.; Jordan, C. T., Targeting aberrant glutathione metabolism to eradicate human acute myelogenous leukemia cells. Journal of Biological Chemistry 2013, 288, (47), 33542–33558.
[91]. Dündar, Y.; Aslan, R., Oksidan-antioksidan denge ve korunmasında vitaminlerin rolü. Hayvancılık Araştırma Dergisi 1999, 9, (12), 32-39.
[92]. Aydın, A.; Sayal, A.; Işımer, A., Serbest radikaller ve antioksidan savunma sistemi. Gülhane Askeri Tıp Akademisi Ayın Kitabı No:20; GATA Basımevi: Ankara, 2001; s 1-12.
[93]. Li, Y.; Schellhorn, H. E.; New developments and novel therapeutic perspectives for vitamin C. Journal of Nutrition 2007, 137, (10), 2171-2184.
[94]. Carr, A. C.; Frei, B., Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. The American Journal of Clinical Nutrition 1999, 69, (6), 1086-1107.
[95]. Gutteridge, J. M. C., Lipid Peroxidation and antioxidants as biomarkers of tissue damage. Clinical Chemistry 1995, 41, (12), 1819-1828.
[96]. Burtis, C. A.; Ashwood, E. R., Klinik Kimyada Temel İlkeler. 4th ed.; Palme Yayınları: Ankara, 2005; s 463-493.
[97]. Gürkan, A. S.; Bozdağ-Dündar, O., Coenzyme Q10. Journal of Faculty of Pharmacy of Ankara 2005, 34, (2), 129-154.
[98]. Packer, L.; Kraemer, K.; Rimbach, G., Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition 2001, 17, (10), 888-895.
[99]. Kim, Y.; Kim, D. C.; Cho, E. S.; Ko, S. O.; Kwon, W. Y.; Suh, G. J.; Shin, H. K., Antioxidant and antiinflammatory effects of selenium in oral buccal mucosa and small intestinal mucosa during intestinal ischemia-reperfusion injury. Journal of Inflammation Research 2014, 11, (36), 1-8.
[100]. Hussein, H. K.; Elnaggar, M. H.; Al-Zahrani, N. K., Antioxidant role of folic acid against reproductive toxicity of cyhalothrin in male mice. Global Advanced Research Journal of Environmental Science and Toxicology 2012, 1, (4), 66-71.
[101]. Ebaid, H.; Bashandy, S. A. E.; Alhazza, I. M.; Rady, A.; El-Shehry, S., Folic acid and melatonin ameliorate carbon tetrachlorideinduced hepatic injury, oxidative stress and inflammation in rats. Nutrition & Metabolism 2013, 10, (20), 1-10.
[102]. Heim, K. E.; Tagliaferro, A. R.; Bobilya, D. J., Flavonoid antioxidants: chemistry, metabolism and structure-activity relationships. The Journal of Nutritional Biochemistry 2002, 13, (10), 572-584.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
64
[103]. Finley, J. W.; Kong, A. N.; Hintze, K. J.; Jeffery, E. H.; Ji, L. L.; Lei, X. G., Antioxidants in foods: state of the science important to the food industry. Journal of Agricultural and Food Chemistry 2011, 59, (13), 6837-6846.
[104]. Gulcin, I., Antioxidant activity of food constituents: an overview. Archives of Toxicology 2012, 86, (3), 345-391.
[105]. Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F., The stomach as a site for anthocyanins absorption from food. FEBS Letters 2003, 544, (1-3), 210-213
[106]. Gil, M. I.; Tomas-Barberan, F. A.; Hess-Pierce, B.; Holcroft, D. M.; Kader, A. A., Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. Journal of Agricultural and Food Chemistry 2000, 48, (10), 4581-4589.
[107]. Chen, P. S.; Li, J. H., Chemopreventive effect of punicalagin, a novel tannin component isolated from terminaliacatappa, on H-ras-transformed NIH3T3 cells. Toxicology Letters 2006, 163, (1), 44-53.
[108]. Kulkarni, A. P.; Mahal, H. S.; Kapoor, S.; Aradhya, S. M., In vitro studies on the binding, antioxidant, and cytotoxic actions of punicalagin. Journal of Agricultural and Food Chemistry 2007, 55, (23), 1491-1500.
[109]. Lee, S. I.; Kim, B. S.; Kim, K. S.; Lee, S.; Shin, K. S.; Lim, J. S., Immune-suppressive activity of punicalagin via ınhibition of NFAT activation. Biochemical and Biophysical Research Communications 2008, 371, (4), 799-803.
[110]. Malik, A.; Afaq, F.; Sarfaraz, S.; Adhami, V. M.; Syed, D. N.; Mukhtar, H., Pomegranate fruit juice for chemoprevention and chemotherapy of prostate cancer. Proceedings of the National Academy of Sciences USA 2005, 102, (41), 14813-14818.
[111]. Adams, L. S.; Seeram, N. P.; Aggarwal, B. B.; Takada, Y.; Sand, D.; Heber, D., Pomegranate juice, total pomegranate ellagitannins, and punicalagin suppress inflammatory cell signaling in colon cancer cells. Journal of Agricultural and Food Chemistry 2006, 54, (3), 980–985.
[112]. Lansky, E. P.; Newman, R. A., Punica granatum (pomegranate) and its potential for prevention and treatment of inflammation and cancer. Journal of Agricultural and Food Chemistry 2007, 109, (2), 177–206.
[113]. Conner, E. M.; Grisham, M. B., Inflammation, free radicals, and antioxidants. Nutrition 1996, 12, (4), 274–277.
[114]. Lei, F.; Xing, D. M.; Xiang, L.; Zhao, Y. N.; Wang, W.; Zhang, L. J.; Du, L. J., Pharmacokinetic study of ellagic acid in rat after oral administration of pomegranate leaf extract. Journal of Chromatography B 2003, 796, (1), 189–194.
[115]. Li, J. K.; Li, G. X.; Zhao, Y. H.; Yu, C. Z., Composition of pomegranate peel polyphenols and its antioxidant activities. Scientia Agricultura Sinica 2009, 42, (11), 4035–4041.
[116]. Kulkarni, A. P.; Mahal, H. S.; Kapoor, S.; Aradhya, S. M., In vitro studies on the binding, antioxidant and cytotoxic actions of punicalagin. Journal of Agricultural and Food Chemistry 2007, 55, (4), 1491–1500.
[117]. Yang, C. S.; Landau, J. M.; Huang, M. T.; Newmark, H. L., Inhibition of carcinogenesis by dietary polyphenolic compounds. The Annual Review of Nutrition 2001, 21, 381–406.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
65
[118]. Cerdá, B.; Llorach, R.; Cerón, J. J.; Espín, J. C.; Tomás-Barberán, F. A., Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. European Journal of Nutrition 2003, 42, (1), 18–28.
[119]. Seeram, N. P.; Lee, R.; Heber, D., Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate juice. Clinica Chimica Acta 2004, 348, (1-2), 63–68.
[120]. Nororiha, I. L.; Niemir, Z.; Stein, H.; Waldher, R., Cytokines and growth factors in renal disease. Nephrology Dialysis Transplantation 1995, 10, 775-786.
[121]. Judith, A.; Owen, J.P.; Sharon, A., Kuby Immunology, 7th ed.; Macmillan: Stranford, 2007; 12-44.
[122]. Bienvenu, J., Exploration of cytokines in biological fluids. Comptes Rendus des Seances de la Societe de Biologie et de Ses Filiale 1995, 189, (4), 545-55. [123]. Armstrong, L.; Jordan, N.; Millar, A., Interleukin 10 regulation of TNF-α from human alveolar macrophages and pheripheral blood monocytes. Thorax 1996, 51, (2), 143-149.
[124]. Farrar, M. A.; Schreiber, R. D., The moleculer cell biology of interferon-gamma and its receptor. Annual Reviews 1993, 11, (1), 571-611.
[125]. Sen, G. C.; Lengyel, P., The interferon system. A birds eye view of its biochemistry. Journal of Biological Chemistry 1992, 267, (8), 5017-5020.
[126]. Abbas, A. K.; Lichtman, A. H.; Poper, J. S., Cytokines. Cellular and Molecular Immunology. WB Saunders Company: Philadelphia, 1994; 240-261.
[127]. Stites, D.P.; Terr, A.I., Basic and Clinical Immunology. 8th ed.; Mcgraw-hill/appleton & Lange: USA, 1994; p 105-123.
[128]. Jaatela, M., Biologic activities and mechanism of action of tumor necrosis factor-α. Laboratory Investigation 1991, 64, 724-742.
[129]. Abbas, A.K.; Lichtman, A.H.; Poper, J.S., Cytokines. Cellular and Molecular Immunology Philadelphia. WB Saunders Company: 1994; p 240-261.
[130]. Vasalli, P., The pathophysiology of tumor necrosis factors, The Annual Review of Immunology 1992, 10, 411-452.
[131]. Aggarwal, B.B.; Gupta, S.C.; Kim, J.H., Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 2012, 119, (3), 651-65.
[132]. Horiuchi, T.; Mitoma, H.; Harashima, S.I.; Tsukamoto, H.; Shimoda, T., Transmembrane TNF-α: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, (7), 1215-28.
[133]. Flynn, J.L.; Chan, J., Immunology of tuberculosis. The Annual Review of Immunology 2001, 19, (1), 93-129.
[134]. Saunders, B.M.; Tran, S.; Ruuls, S.; Sedgwick, J.D.; Briscoe, H.; Britton, W.J., Transmembrane TNF is sufficient to initiate cell migration and granuloma formation and provide acute, but not long-term, control of mycobacterium tuberculosis infection. Journal of Immunology 2005, 174, (8), 4852-9.
[135]. Mootoo, A.; Stylianou, E.; Arias, M.A.; Reljic, R., TNF-in tuberculosis: a cytokine

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
66
with a split personality. Inflammation & Allergy - Drug Targets 2009, 8, (1), 53-62.
[136]. Vinay, D.S.; Kwon, B.S., The tumour necrosis factor/TNF receptor superfamily: Therapeutic targets in autoimmune diseases. Clinical and Experimental Immunology 2011, 164, (2), 145-57.
[137]. Pfeffer, K., Biological functions of tumor necrosis factor cytokines and their receptors. Cytokine & Growth Factor Reviews 2003, 14, (3-4), 185-91.
[138]. Kollias, G.; Kontoyiannis, D., Role of TNF/TNFR in autoimmunity: specific TNF receptor blockade may be advantageous to anti-TNF treatments. Cytokine & Growth Factor Reviews 2002, 13, (4-5), 315-21.
[139]. Schreiber, R.D.; Chaplin, D.D.; Frank, M.M., Cytokines, Inflammation, and Innate immunity. 5th ed.; Brown and Company: USA, 1995; p 279-286.
[140]. Tracy, K.J.;Cerami, A., Tumor necrosis factor: A pleiotropic cytokin anad therapeutic target The Annual Review of Medicine 1994, 45, 491-503.
[141]. Vassali, P., The pathophysiology of Tumor Necrosis Factor. The Annual Review of Immunology 1992, 10, 411, 452.
[142]. Smith, C.A.; Davis, T.; Anderson, D., A receptor fot tumor necrosis factor defines an unusual family of celluler and viral proteins. Sience 1990, 248, (4958), 1019-1023.
[143]. Dinarello, C.A., IL -1 and IL -1 antagonism. Blood 1991, 77, 1627.
[144]. Pereina, B.J.G., Implication of cytokine measurements during in vitro and clinical haematodialysis. Nephrology Dialysis Transplantation 1995, 10, (10), 18-26.
[145]. Kishimoto, T., The biology of interleukin – 6. Blood 1989, 74, (1), 1-10.
[146]. Horii, Y.; Muraguchi, A.; Iwano, M., Involvement of IL -1 in mesengial proliferative glomerulonephritis. Journal of Immunology 1989, 143, 3949-3955.
[147]. Stahl, R.A.K., Chemoattractive cytokines (Chemokines) and immune renal injury. Nephrology Dialysis Transplantation 1995, 10, 307-319.
[148]. Laudanna, C.; Campbell, J.J.; Butcher, E.C., Role of Rho in chemoatractant-activated leucocyte adhesion through integrins. Science 1996, 271, (5251), 981-3.
[149]. Smith, K.A., Low-dose IL-2 immunothrapy. Blood 1993, 81, 1402.
[150]. Smith, K.A., Interleukin -2 :Inception,impact and implications. Science 1988, 240, (8), 1169-1176.
[151]. Mackoy, K.; Striker, L.J.; Tauffer, J.W.; Doi, T.; Agodoa, L.Y.; Striker, GE., Transforming Growth Factor - B : Murine glomerular receptors and responses of isolated glomerular cells. Journal of Clinical Investigation 1989, 83, (4), 1160-1167.
[152]. Tikkanen, I.; Uhlenius, N.; Tikkanen, T.; Miettinen, A.; Trnroth, T.; Fyhrauist, F.; Holthfer, H., Increased renal expression of cytokines and growth factors induced by DOCA-Nacl treatment in Heymann nephritis. Nephrology Dialysis Transplantation 1995, 10, (12), 2192-2198.
[153]. Müllüoğlu, E., Sitokinler. İmmnoloji. Saray Tıp Kitabevleri: İzmir, 1993; s 77-97.
[154]. Deng, M.C.; Erren, M.; Kamnerling, L.; Gunther, F.; Kerber, S.; Fahrenkamp, A.; Assman, G.; Brethardt, G.; Scheld, H.H., The relation of interleukin-6, Tumor Necrosis Factor - a, IL - 2 and IL -

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
67
2 receptor levels to cellular rejection, allograft dysfunction and clinical events early after cardiac transplantation. Transplantation 1995, 60, 1118-1124.
[155]. Moore, K. W.; de Waal Malefyt, R.; Coffman, R. L.; O’Garra, A., Interleukin-10 and the interleukin-10 receptor. Annual Review of Immunology 2001, 19, (1), 683–765.
[156]. Sanderson, C.F., Interleukin-5,eosinophils and disease. Blood 1992, 79, (12), 3101-3109.
[157]. Bloom, B.R.; Bennett, B., Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science 1966, 153, (3731), 80-82.
[158]. Mitchell, R.; Bacher, M.; Bernhagen, J., Cloning and characterization of the gene for mouse macrophage migration inhibitory factor (MIF). Journal of Immunology 1995, 154, (8), 3863-3870.
[159]. Calandra, T.; Roger, T., Macrophage migration inhibitory factor: a regulator of innate immunity. Nature Reviews Immunology 2003, 3, 791-800.
[160]. Imamura, K.; Nishihira, J.; Suzuki, M., Identification and immunohistochemical localization of macrophage migration inhibitory factor in human kidney. Biochemistry and Molecular Biology International 1996, 40, (6), 1233-1242.
[161]. Shimizu, T.; Ohkawara, A.; Nishihira, J.; Sakamoto, W., Identification of macrophage migration inhibitory factor (MIF) in human skin and its immmunohistochemical localization. FEBS Letter 1996, 381, (3), 199-202.
162. Bacher, M.; Meinhardt, A.; Lan, H.Y.,. MIF expression in the rat brain: implications for neuronal function. Molecular Medicine 1998, 4, (4), 217-230.
[163]. Kılıçturgay, K., Sitokinler İmmunolojiye Giriş. Yargıçoğlu matbaası: Ankara, 1987; 72-83.
[164]. Heaney, M.L.; Golde, D.W., Soluble Cytokine Receptors. Blood 1996, 87, (3), 847-857.
[165]. Lohnstan, J.A.; Wang, L.M.; Hanson, E.P.; Sun, X.J.; White, M.F., IL-2,4,7 an 15 stimulate tyrozine phosphorylation of insulin receptr substrates. Journal of Biological Chemistry 1995, 270, (48), 2827-30.
[166]. Pollmacher, T.; Korth, C.; Mullington, J.; Schreiber, W.; Sauer, J.; Vedder, H.; Galanos, C.; Holsber, W., Effects of Granulocyte Colony- Stimolating Factor on plasma cytokine and cytokine receptor levels and on the n vivo host response to endotoxin in healthy men. Blood 1996, 87, (3), 900-905.
[167]. Buemann, B.; Astrup, A., How does the body deal with energy from alcohol. Nutrition 2001, 17, (7-8), 638-641.
[168]. Ashmarin, I.P.; Danilova, R.A.; Obakhova, M.F.; Moskvitina, T.A.; Prosorovsky, V.N., Main ethanol metabolizing alcohol dehydrogenases (ADH I and ADH IV): biochemical functions and the physiological manifestation. FEBS Letters 2000, 486, (1), 49-51.
[169]. Suter,P.M.; Je’quier, E.; Schutz, Y., Effect of ethanol on energy expenditure. American Journal of Physiology 1994, 266, (4), 1204-1212.
[170]. Lands, W.E.M., Alcohol and energy intake. American Journal of Clinical Nutrition 1995, 62, 1101-1106.
[171]. Amit, Z.; Aragon, C.M.G.; Smith, B.R., Alcohol metabolizing enzymes as possible markers mediating voluntary consumption. Canadian Journal of Public Health 1986, 1, 15-20.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
68
[172]. Albano, E., Free radicals and alcohol-induced liver injury. In Ethanol and the Liver. Preedy, V.R.; Watson, R.R. ed.; London, 2002; p 153-190.
[173]. Ronis, M.J.J.; Lindros, K.O.; Ingelman-Sundberg, M., The CYP2E family. In Cytochromes P450: Metabolic and Toxicological Aspects. CRC Press: USA, 1996; p 211–239.
[174]. Bailey, S.M.; Cunningham, C.C., Contribution of mitochondria to oxidative stress associated with alcohol liver disease. Free Radical Biology and Medicine 2002, 32, (1), 11–16.
[175]. Steinbauer, J.R.; Cantor, S.B.; Holzer, C.E.; Volk, R.J., Ethnic and sex bias in primary care screening tests for alcohol use disorders. Annals of Internal Medicine 1998, 129, (5), 353-62.
[176]. Bradley, K.A.; Boyd-Wickizer, J.; Powell, S.H.; Burman, M.L., Alcohol screening questionnaires in women: a critical review. JAMA 1998, 280, (2), 166-71.
[177]. McClain, C. J.; Barve, S.; Deaciuc, I.; Kugelmas, M.; Hill, D., Cytokines in alcoholic liver disease. Seminars Liver Disease 1999, 19, (2), 205–219.
[178]. Thurman, R. G.; Bradford, B. U.; Iimuro, Y.; Frankenberg, M. V.; Knecht, K. T.; Connor, H. D.; Adachi, Y.; Wall, C.; Arteel, G. E.; Raleigh, J. A.; Forman, D. T.; Mason, R. P., Mechanisms of alcohol-induced hepatotoxicity: studies in rats. Frontiers in Bioscience 1999, 4, (5), 42–46.
[179]. Tilg, H.; Diehl, A. M., Cytokines in alcoholic and nonalcoholic steatohepatitis. The New England Journal of Medicine 2000, 343, (20), 1467–1476.
[180]. Nanji, A. A.; Khettry, U.; Sadrzadeh, S. M., Lactobacillus feeding reduces endotoxemia and severity of experimental alcoholic liver disease. Experimental Biology and Medicine 1994, 205, (3), 243–247.
[181]. Chen, J.; Bao, H.; Sawyer, S.; Kunos, G.; Gao, B., Effects of short and long term ethanol on the activation of signal transducer and activator transcription factor 3 in normal and regenerating liver. Biochemical and Biophysical Research Communications 1997, 239, (3), 666–669.
[182]. Yang, S. Q.; Lin, H. Z.; Yin, M.; Albrecht, J. H.; Diehl, A. M., Effects of chronic ethanol consumption on cytokine regulation of liver regeneration. American Journal of Physiology 1998, 275, (4), 696–704.
[183]. Sedlak, J.; Lindsay, R. H., Estimation of Total Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Analytical Biochemistry 1968 25, (1), 1192-1205.
[184]. Aebi, H., Catalase In Vitro. Methods in Enzymology 1984, 105, 121-126.
[185]. Sun, Y.; Oberley, L.W.; Li, Y. A, Simple Method for Clinical Assay of Superoxide Dismutase. Clinical Chemistry 1988, 34, (3), 497-500. [186]. Ohkawa, H.; Ohishi, N.,; Yagi, K., Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Analytical Biochemistry 1979, 95, (2), 351-358. [187]. Halliwell, B.; Gutteridge, M.C., Role of free radicals and catalytic metal ions in human disease : An overview. Methods in Enzmology 1990, 186, 1-85.
[188]. Ikeda, Y.; Long, D.M., The molecular basis of brain iınjury and brain edema: The role of oxygen free radicals. Neurosurgery, 1990, 27, (1), 1-11.
[189]. Janssen, Y.M.W.; Houten, B.V.; Borm, P.J.A.; Mossmon, B.T.; Biology of disease, cell and tissue responses to oxidative damage. Laboratory Investigation 1993, 69, (3), 261-274.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
69
[190]. Özdem, S.; Şadan, G., Serbest oksijen radikallerinin oluşum ve klinik açıdan önemi, Ankara Üniversitesi Tıp Fakültesi Dergisi 1994, 11, 1, 63-71.
[191]. Özden, S., Bazı Pestisidlerin Oksidatif Stres Oluşturma Potansiyellerinin ve Antioksidan Sistemler Üzerine Etkilerinin Sıçanlarda Araştırılması, Doktora Tezi, İstanbul Üniversitesi Sağlık Bilimleri Enstitüsü, İstanbul, 2006.
[192]. Mercan, U., Toksikolojide serbest radikallerin önemi. Van Yüzüncü Yıl Üniversitesi Veterinerlik Fakültesi Dergisi 2004, 15, (1-2), 91-96.
[193]. Serafini, M.; Rio, D., Understanding the association between dietray antioxidants, redox status and disease: Is the Total Antioxidant Capacity the Right Tool. Redox Report 2004, 9, (3),145-152.
[194]. Aykaç, G.; Uysal, M.; Süha Yalçin, A.; Koçak-Toker, N.; Sivas, A.; Öz, H., The effect of chronic ethanol ingestion on hepatic lipid peroxide, glutathione, glutathione peroxidase and glutathione transferase in rats. Toxicology, 1985, 36, (1), 71-76.
[195]. Sonde, V.; D’souza, A.; Tarapore, R.; Pereira, L. Khare, M.P.; Sinkar, P.; Krishnan, S.; Rao, C.V., Simultaneous administration of diethylphthalate and ethyl alcohol and its toxicity in male Sprague-Dawley rats. Toxicology, 2000, 147, (1), 23-31.
[196]. Kolankaya, D.; Selmanoğlu, G.; Sorkun, K.; Salih, B., Protective effects of Turkish propolis on alcohol-induced serum lipid changes and liver injury in male rats. Food Chemistry 2002, 78, (2), 213-217.
[197]. Yurt, B., Etil Alkol ile Oksidatif Stres Oluşturulan Sıçanlarda Kayısı ve Kayısı Çekirdeğinin Karaciğer Koruyucu ve Antioksidan Etkilerinin Belirlenmesi, Doktora Tezi, Van Yüzüncü Yıl Üniversitesi Fen Bilimleri Enstitüsü ,Van, 2010.
[198]. Doğan A., Etil Alkol ile Deneysel Oksidatif Stres Oluşturulan Sıçanlarda Üzüm (vitis vinifera l. ) Çekirdeğinin Karaciğer Koruyucu ve Antioksidan Rolünün Belirlenmesi, Yüksek Lisans Tezi, Van Yüzüncü Yıl Üniversitesi Fen Bilimleri Enstitüsü, Van, 2010.
[199]. Murray R. K.; Granner D. K.; Mayes P. A.; Radwell V. W., Harper’s Biochemistry. 21.th ed.; Long Medical Book: London, 1988; p 57-83.
[200]. Van Zutphen, L.F.M.; Baumans, V.; Beynen, A.C., Laboratuvar Hayvanları Biliminin Temel İlkeleri. 1.baskı; Medipres yayıncılık: Malatya, 2003.
[201]. Roediger, W.E.W., New views on the pathogenesis of kwashiorkor: methionine and other amino acids. Journal of Pediatric Gastroenterology and Nutrition 1995, 21, (2), 130-136.
[202]. Meister, A.; Anderson, M.E., Glutathione. Annual Review of Biochemistry 1983, 52, 711-760.
[203]. Yagi, K., Free Radicals in Diagnostic Medicine. Armstrong Donald ed.; Springer Nature: Basingstoke, 1994; s 165-169.
[204]. Avellini, L.; Spaterna, A.; Rebold, G.P.; Gaiti, A., Defense mechanisim against free radical-induced damage in sheep, cattle and dog erythrocytes. Comparative Biochemistry & Physiology 1993, 106, (2), 391-394.
[205]. Kurata, M., Suzuki, M., Agar, N.S., Antioxidant system and erythrocyte life span in mammals. Comparative Biochemistry & Physiology 1999, 106, (3), 477-487

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
70
[206]. Halliwell, B.; Murcia, M.A.; Chirico, S.; Auroma, O.I., Free radicals and antioxidants in food and in vivo: what they do and how they work. Critical Reviews in Food Science and Nutrition 1995, 35, (1), 7-20.
[207]. Slater, T.F., Overview of methods used for detecting lipid peroxidation. Methods in Enzymology 1984, 105, 283-305.
[208]. Sushil, J.K.; Mcvie, R.; Duett, J.; Herbst, J.J., Erythrocyte membrane lipid peroxidation and glycosylated hemoglobin in diabetes. Diabetes 1989, 38, (12) ,1539-1543.
[209]. Porter, N.A., Chemistry of lipid peroxidation. Methods in Enzymology 1984, 105, 273-283.
[210]. Niki, E., Antioxidant in relation to lipid peroxidation. Chemistry and Physics of Lipids 1987, 44, (2-4), 227-253.
[211]. Koner, B.C.; Banerjee, B.D.; Ray, A., Organochlorine pesticide induced oxidative stress and immune suppression in rats. Indian Journal of Experimental Biology 1988, 36, (4), 395-398.
[212]. Duthie, G.G.; Wahle, K.W.J.; James, W.P.T., Oxidants, antioxidants and cardiovascular disease. Nutrition Research Reviews 1989, 2, (1), 51-62.
[213]. Clemens, M.C.; Waller, H.D., Lipid peroxidation in erythrocytes. Chemistry and Physics of Lipids 1987, 45, (2-4), 251-268.
[214]. Miller, D.R.; Baehner, R.L., Blood Diseases of Infancy and Childhood. 6th ed.; The C.V. Mosby Company: St. Louis, 1990; p 295-304.
[215]. Jurczuk, M.; Brzoska, M.M.; Moniuszko-Jakoniuk, J.; GalazynSidorczuk, M.; Kulikowska-Karpinska, E., Antioxidant enzymes activity and lipid peroxidation in liver and kidney of rats exposed to cadmium and ethanol. Food and Chemical Toxicology 2004, 42, (3), 429-438.
[216]. McClain, C.J.; Shedlofsky, S.; Barve, S.; Hill, D.B., Cytokines and alcoholic liver disease. Alcohol Research and Health 1997, 21, (4), 317-320.
[217]. McClain, C.J.; Barve, S.; Deacıuc, I.; Kugelmas, M.; Hill, D., Cytokines in alcoholic liver disease. Seminars in Liver Disease 1999, 19, (2), 205-219.
[218]. Nanji, A.A., Role of kupffer cells in alcoholic hepatitis. Alcohol 2002, 27, (1), 13-15.
[219]. Thurman, R.G., Mechanism of hepatic toxiciti II. Alcoholic liver injury involves activation of kupffer cell by endotoxin. American Journal of Physiology 1998, 275, (38), 605-611.
[220]. Lansky, E.; Shubert, S.; Neeman, I., Pharmacological and therapeutic properties of
pomegranate, Serie A Seminaires Mediterraneennes, Zaragoza, 2000, s 231-235.
[221]. Aviram, M.; Dornfeld, L.; Rosenblat, M.; Volkona N.; Kaplan, M.; Coleman, R.; Hayek, T.; Presser, D.; Fuhrman, B., Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LLD, and platelet aggregation. The American Journal of Clinical Nutrition 2000, 71, (5), 1062-1076.
[222]. Priyadarsini, K. I.; Khopde, S. M.; Kumar, S. S.; Mohan, H., Free radical studies of ellagic acid, a natural phenolic antioxidant. Journal of Agricultural and Food Chemistry 2002, 50, (7), 2200-2206.
[223]. Fischer, U. A.; Carle, R.; Kammerer, D. R., Identification and quantification of phenolic compounds from pomegranate peel. Food Chemistry 2011, 127, 807-821.

Esma Yanpar, Sağlık Bilimleri Enstitüsü, Mersin Üniversitesi, 2019
71
ÖZGEÇMİŞ
Adı ve Soyadı : Esma YANPAR Doğum Tarihi : 01/02/1991 E-mail : [email protected] Öğrenim Durumu :
Derece Bölüm/Program Üniversite Yıl Lisans Eczacılık Fakültesi Ankara Üniversitesi 2009-2014 Yüksek Lisans Eczacılık Biyokimya Mersin Üniversitesi 2016-2019
Related Documents











