ETA Receptor Blockade Induces Tubular Cell Proliferation and Cyst Growth in Rats with Polycystic Kidney Disease BERTHOLD HOCHER,* PHILIPP KALK,* TORSTEN SLOWINSKI,* MICHAEL GODES,* ALEXANDER MACH,* SOPHIA HERZFELD,* DOREEN WIESNER, † PETRA CLARA ARCK, ‡ HANS-H. NEUMAYER,* and BENNO NAFZ † Departments of *Nephrology and † Physiology, and ‡ Biomedical Research Center, University Hospital Charite ´, Humboldt University of Berlin, D-10098 Berlin, Germany. Abstract. Tissue concentrations of ET-1 are markedly elevated in the kidneys of Han:Sprague-Dawley (Han:SPRD) rats, a model of human autosomal dominant polycystic kidney disease (ADPKD). This study analyzed whether disease progression might be attenuated by endothelin receptor antagonists. Het- erozygous Han:SPRD rats received an ETA receptor antagonist (LU 135252), a combined ETA/ETB receptor antagonist (LU 224332), or placebo for 4 mo. Glomerulosclerosis, protein excretion, and GFR remained unchanged, whereas interstitial fibrosis was enhanced by both compounds. BP was not reduced by both compounds in Han:SPRD rats. Renal blood flow (RBF) decreased in ADPKD rats treated with the ETA receptor antagonist. Long-term ETA receptor blockade furthermore in- creased markedly the number of renal cysts (ADPKD rats, 390 119 [cysts/kidney section SD]; LU 135252-treated APKD rats, 1084 314; P 0.001), cyst surface area (ADPKD rats, 7.97 2.04 [% of total section surface SD]; LU 135252- treated ADPKD rats, 33.83 10.03; P 0.001), and cell proliferation of tubular cells (ADPKD rats, 42.2 17.3 [BrdU- positive cells/1000 cells]; LU 135252-treated ADPKD rats, 339.4 286.9; P 0.001). The additional blockade of the ETB receptor attenuated these effects in Han:SPRD rats. Both endothelin receptor antagonists had no effect on BP, protein excretion, GFR, and kidney morphology in Sprague-Dawley rats without renal cysts. It is concluded that ETA receptor blockade enhances tubular cell proliferation, cyst number, and size and reduces RBF in Han:SPRD rats. This is of major clinical impact because endothelin receptor antagonists are upcoming clinically used drugs. Autosomal dominant polycystic kidney disease (ADPKD) is the most common monogenic hereditary kidney disease in humans, which is seen in about 1 in 1000 live births. This disease accounts for up to 10% of all patients requiring renal replacement therapy. Cysts arise from renal tubular segments as focal areas of dilatation. They progressively enlarge with age and might separate from the nephron of origin. Mutations in two genes, PKD1 and PKD2, are associated with this disease in humans (1,2). Progression of ADPKD, resulting in renal failure, varies between affected members of different families, as well as between different members of the same family. In some individuals, kidney cysts are present in early childhood and progress to end-stage renal failure before age of 40 yr, whereas renal function remains unimpaired throughout lifespan in others. Several general factors, such as gender, germ-line mutations, modifier genes, and epigenetic factors, affect dis- ease progression with respect to cyst growth, interstitial in- flammation, and progressive fibrosis (3–5). The renal endothe- lin system seems to be one of these disease-modifying factors. ET-1 transgenic mice develop small cortical kidney cysts (10,31). It was furthermore shown that the renal ET system is markedly activated in patients with autosomal dominant poly- cystic disease (ADPKD) (6 – 8), in polycystic kidneys of cpk mice (9) as well as in Han:SPRD rats (7) (a rat model of ADPKD). The Han:SPRD rat strain develops a form of pro- gressive gender-dependent disease that appears similar in many respects to that seen in the ADPKD in humans (11), even through different genetic defects. The gene responsible for this disease in rats is located on rat chromosome 5 and is not related to the human PKD1 or PKD2 gene (12). ADPKD in humans as well as in Han:SPRD rats is characterized by structural alter- ations of the kidneys, such as thickening of the tubular base- ment membrane, interstitial fibrosis, and formation of cysts leading to end-stage kidney disease (11). Tubular cell prolif- eration seems to be an important step during pathogenesis of kidney cysts in ADPKD, because the main feature of the disease is the development of renal cysts, first occurring in the proximal tubules, and with time dominating all segments of the nephron. It is known that a primarily activated renal endothelin sys- Received May 5, 2002. Accepted September 21, 2002. Correspondence to Priv. Doz. Dr. Berthold Hocher; Universita ¨tsklinikum Charite ´ der Humboldt Universita ¨t zu Berlin, Klinik fu ¨r Nephrologie, Schu- mannstr. 20-21, 10098 Berlin, Germany. Phone: 49-30-450514098; Fax: 49- 30-450514938; E-mail: [email protected] 1046-6673/1401-0367 Journal of the American Society of Nephrology Copyright © 2003 by the American Society of Nephrology DOI: 10.1097/01.ASN.0000042165.63601.65 J Am Soc Nephrol 14: 367–376, 2003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ETA Receptor Blockade Induces Tubular Cell Proliferationand Cyst Growth in Rats with Polycystic Kidney Disease
BERTHOLD HOCHER,* PHILIPP KALK,* TORSTEN SLOWINSKI,*MICHAEL GODES,* ALEXANDER MACH,* SOPHIA HERZFELD,*DOREEN WIESNER,† PETRA CLARA ARCK,‡ HANS-H. NEUMAYER,* andBENNO NAFZ†
Departments of *Nephrology and †Physiology, and ‡Biomedical Research Center, University Hospital Charite,Humboldt University of Berlin, D-10098 Berlin, Germany.
Abstract. Tissue concentrations of ET-1 are markedly elevatedin the kidneys of Han:Sprague-Dawley (Han:SPRD) rats, amodel of human autosomal dominant polycystic kidney disease(ADPKD). This study analyzed whether disease progressionmight be attenuated by endothelin receptor antagonists. Het-erozygous Han:SPRD rats received an ETA receptor antagonist(LU 135252), a combined ETA/ETB receptor antagonist (LU224332), or placebo for 4 mo. Glomerulosclerosis, proteinexcretion, and GFR remained unchanged, whereas interstitialfibrosis was enhanced by both compounds. BP was not reducedby both compounds in Han:SPRD rats. Renal blood flow(RBF) decreased in ADPKD rats treated with the ETA receptorantagonist. Long-term ETA receptor blockade furthermore in-creased markedly the number of renal cysts (ADPKD rats, 390� 119 [cysts/kidney section � SD]; LU 135252-treated APKD
rats, 1084 � 314; P � 0.001), cyst surface area (ADPKD rats,7.97 � 2.04 [% of total section surface � SD]; LU 135252-treated ADPKD rats, 33.83 � 10.03; P � 0.001), and cellproliferation of tubular cells (ADPKD rats, 42.2 � 17.3 [BrdU-positive cells/1000 cells]; LU 135252-treated ADPKD rats,339.4 � 286.9; P � 0.001). The additional blockade of theETB receptor attenuated these effects in Han:SPRD rats. Bothendothelin receptor antagonists had no effect on BP, proteinexcretion, GFR, and kidney morphology in Sprague-Dawleyrats without renal cysts. It is concluded that ETA receptorblockade enhances tubular cell proliferation, cyst number, andsize and reduces RBF in Han:SPRD rats. This is of majorclinical impact because endothelin receptor antagonists areupcoming clinically used drugs.
Autosomal dominant polycystic kidney disease (ADPKD) isthe most common monogenic hereditary kidney disease inhumans, which is seen in about 1 in 1000 live births. Thisdisease accounts for up to 10% of all patients requiring renalreplacement therapy. Cysts arise from renal tubular segmentsas focal areas of dilatation. They progressively enlarge withage and might separate from the nephron of origin. Mutationsin two genes, PKD1 and PKD2, are associated with this diseasein humans (1,2). Progression of ADPKD, resulting in renalfailure, varies between affected members of different families,as well as between different members of the same family. Insome individuals, kidney cysts are present in early childhoodand progress to end-stage renal failure before age of 40 yr,whereas renal function remains unimpaired throughout lifespanin others. Several general factors, such as gender, germ-line
mutations, modifier genes, and epigenetic factors, affect dis-ease progression with respect to cyst growth, interstitial in-flammation, and progressive fibrosis (3–5). The renal endothe-lin system seems to be one of these disease-modifying factors.ET-1 transgenic mice develop small cortical kidney cysts(10,31). It was furthermore shown that the renal ET system ismarkedly activated in patients with autosomal dominant poly-cystic disease (ADPKD) (6–8), in polycystic kidneys of cpkmice (9) as well as in Han:SPRD rats (7) (a rat model ofADPKD). The Han:SPRD rat strain develops a form of pro-gressive gender-dependent disease that appears similar in manyrespects to that seen in the ADPKD in humans (11), eventhrough different genetic defects. The gene responsible for thisdisease in rats is located on rat chromosome 5 and is not relatedto the human PKD1 or PKD2 gene (12). ADPKD in humans aswell as in Han:SPRD rats is characterized by structural alter-ations of the kidneys, such as thickening of the tubular base-ment membrane, interstitial fibrosis, and formation of cystsleading to end-stage kidney disease (11). Tubular cell prolif-eration seems to be an important step during pathogenesis ofkidney cysts in ADPKD, because the main feature of thedisease is the development of renal cysts, first occurring in theproximal tubules, and with time dominating all segments of thenephron.
It is known that a primarily activated renal endothelin sys-
Received May 5, 2002. Accepted September 21, 2002.Correspondence to Priv. Doz. Dr. Berthold Hocher; UniversitatsklinikumCharite der Humboldt Universitat zu Berlin, Klinik fur Nephrologie, Schu-mannstr. 20-21, 10098 Berlin, Germany. Phone: 49-30-450514098; Fax: 49-30-450514938; E-mail: [email protected]
1046-6673/1401-0367Journal of the American Society of NephrologyCopyright © 2003 by the American Society of Nephrology
DOI: 10.1097/01.ASN.0000042165.63601.65
J Am Soc Nephrol 14: 367–376, 2003

tem causes renal scarring/fibrosis (10); we therefore hypothe-sized that the activated endothelin system in ADPKD contrib-utes to the progression of kidney fibrosis seen in these rats.Thus, a long-term pharmacologic blockade of the endothelinsystem might be a new approach to reduce disease progressionin ADPKD. We tested this hypothesis in Han:SPRD rats usingendothelin receptor antagonists that are currently clinically
tested in large clinical trails with regard to heart failure andpulmonary hypertension.
Materials and MethodsAnimals and Study Design
Male heterozygous (cy/�) Han:SPRD rats (5) were analyzed. Theanimals were maintained as an inbred colony in the Department of
Table 1. Body weight, BP, kidney weight, GFR, renal morphometric data, and clinical chemistry of Sprague-Dawley ratsafter 4 mo of treatment with ETA or combined ETA/ETB receptor antagonistsa
Sprague-Dawley Rats Sprague-Dawley Rats� LU 135252
Sprague-Dawley Rats� LU 224332
Body weight (g) 509.9 � 43.0 518.0 � 29.8 516.9 � 50.7Kidney weight (% of body weight) 0.63 � 0.09 0.64 � 0.11 0.62 � 0.09Mean arterial BP 93.4 � 9.1 92.3 � 9.5 94.2 � 10.9GFR (ml/24 h per 100 g BW) 2198 � 348 2221 � 432 2087 � 378Glomerulosclerosis score 1.70 � 0.31 1.67 � 0.23 1.60 � 0.32Interstitial fibrosis (% fibrotic area) 2.84 � 1.94 2.65 � 1.01 2.97 � 0.87Plasma protein (g/dl) 5.7 � 0.5 5.5 � 0.4 5.4 � 0.2Plasma albumin (g/dl) 2.5 � 0.2 2.4 � 0.3 2.4 � 0.2C-reactive protein (mg/dl) 1.3 � 1.0 1.4 � 0.9 1.4 � 0.7Serum urea (mg/dl) 28.4 � 9.8 29.1 � 10.2 31.2 � 12.0Serum alanine aminotransferase (U/L) 14.6 � 4.0 14.2 � 4.1 14.5 � 3.9Urinary protein excretion (g/24 h) 0.03 � 0.03 0.03 � 0.05 0.04 � 0.03Urinary albumin excretion (g/24 h) 0.01 � 0.01 0.01 � 0.02 0.01 � 0.01
a Sprague-Dawley rats (8-wk-old) were treated for 4 months with placebo, LU 135252 (ETA receptor antagonist), or LU 224332(combined ETA/ETB receptor antagonist), respectively. Urine (24-h urine in a metabolic cage) and blood were taken in the last treatmentweek. Data are given as means � SD. None of the parameters tested were significantly influenced by LU 135252 or LU 224332treatment.
Table 2. Body weight clinical chemistry of HanSPRD rats after 4 mo of treatment with ETA or combined ETA/ETBreceptor antagonistsa
HanSPRD Rats HanSPRD Rats� LU 135252
HanSPRD Rats� LU 224332
Body weight (g) 402.9 � 24.0 350.7 � 37.4b 433.2 � 33.1d
Kidney weight (% of body weight) 1.22 � 0.04 3.10 � 0.46c 1.43 � 0.07Mean arterial BP (mmHg) 92.9 � 8.5 101.9 � 15.4 106.5 � 12.0b
GFR (ml/24 h per 100 g BW) 475 � 143 429 � 132 356 � 82Glomerulosclerosis score 2.22 � 0.30 2.49 � 0.22 2.09 � 0.20Interstitial fibrosis (% fibrotic area) 15.6 � 3.8 21.1 � 2.13b 23.3 � 3.30b
Plasma protein (g/dl) 5.0 � 0.5 5.5 � 0.4b 4.7 � 0.2d
Plasma albumin (g/dl) 2.4 � 0.2 2.2 � 0.2 2.1 � 0.2b
C-reactive protein (mg/dl) 2.7 � 1.1 1.8 � 0.1b 2.4 � 0.2d
Plasma urea (mg/dl) 78.4 � 17.4 105.0 � 43.2 101.2 � 13.3d
Plasma alanine aminotransferase (U/L) 12.6 � 3.0 15.0 � 3.4 14.8 � 3.7Urinary protein excretion (g/24 h) 0.05 � 0.03 0.08 � 0.04 0.05 � 0.02Urinary albumin excretion (g/24 h) 0.02 � 0.01 0.04 � 0.02 0.02 � 0.01
a HanSPRD rats were treated for 4 mo with placebo, LU 135252 (ETA receptor antagonist), or LU 224332 (combined ETA/ETBreceptor antagonist), respectively. Urine (24-h urine in a metabolic cage) and blood were taken in the last treatment week. Data are givenas means � SD.
b P � 0.05 versus HanSPRD rats.c P � 0.01 versus HanSPRD rats.d P � 0.05 versus LU 135252-treated HanSPRD rats.
368 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 367–376, 2003

Physiology, University Hospital Charite, Humboldt University of Berlin,Germany. Breeding of Han:SPRD rats was performed by mating het-erozygous (cy/�) Han:SPRD rats. Discrimination between heterozygous
(cy/�) Han:SPRD rats and healthy (�/�) Sprague Dawley rats was doneby analyzing serum urea and creatinine concentrations. These tests al-lowed a reliable discrimination between heterozygous (cy/�) Han:SPRD
Figure 1. Typical kidney sections from Han:Sprague-Dawley (Han:SPRD) rats after 4 mo of treatment with the ETA receptor antagonist or thecombined ETA/ETB receptor antagonist: (A) nontreated Han:SPRD rat; (B) LU 135252 (ETA receptor antagonist)–treated Han:SPRD rat; (C)LU 224332 (combined ETA/ETB receptor antagonist). Magnification is the same in all panels; the bar in panel marks 1 mm. Bar graphs shownumber of renal cysts (D) and cyst surface area (E) in Han:SPRD rats after 4 mo of treatment with the ETA receptor antagonist LU 135252or the combined ETA/ETB receptor antagonist LU 224332. Data are given as mean � SD. ## P � 0.01 compared with nontreated HanSPRDrats; ### P � 0.001 compared with nontreated Han:SPRD rats.
J Am Soc Nephrol 14: 367–376, 2003 ADPKD and Endothelin System 369
rats and healthy (�/�) Sprague Dawley rats if the rats were older than 7wk. The Han:SPRD rats were fed a commercial diet (AltrominR, Al-tromin GmbH, Germany) and given water ad libitum. We establishedthree groups:
• A placebo-treated group with heterozygous (cy/�) Han:SPRD rats,n � 12
• A group of heterozygous (cy/�) Han:SPRD rats treated with theETA receptor antagonist LU 135252 (50 mg/kg per d), n � 10
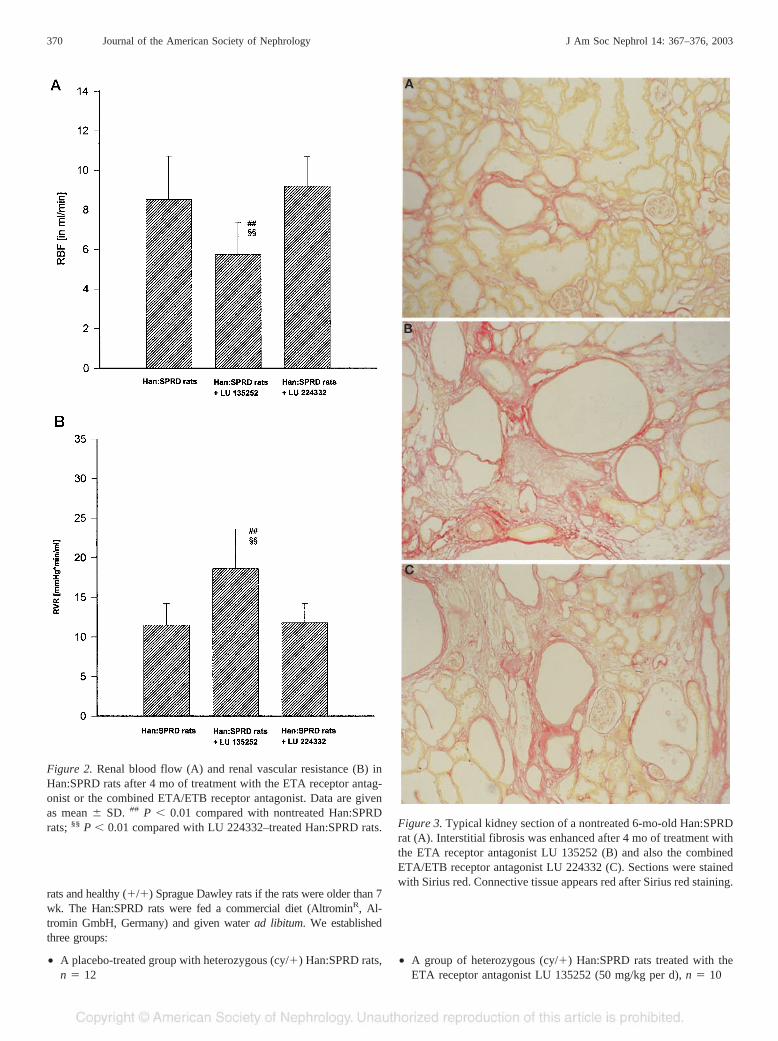
Figure 2. Renal blood flow (A) and renal vascular resistance (B) inHan:SPRD rats after 4 mo of treatment with the ETA receptor antag-onist or the combined ETA/ETB receptor antagonist. Data are givenas mean � SD. ## P � 0.01 compared with nontreated Han:SPRDrats; §§ P � 0.01 compared with LU 224332–treated Han:SPRD rats. Figure 3. Typical kidney section of a nontreated 6-mo-old Han:SPRD
rat (A). Interstitial fibrosis was enhanced after 4 mo of treatment withthe ETA receptor antagonist LU 135252 (B) and also the combinedETA/ETB receptor antagonist LU 224332 (C). Sections were stainedwith Sirius red. Connective tissue appears red after Sirius red staining.
370 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 367–376, 2003

• A group of heterozygous (cy/�) Han:SPRD rats treated with thecombined ETA/ETB receptor antagonist LU 224332 (50 mg/kg perd), n � 5
To control for potential toxic effects of the endothelin receptorantagonists, we furthermore established the following three groups:
• A placebo-treated group with Sprague-Dawley rats, n � 10• A group of Sprague-Dawley rats treated with the ETA receptor
antagonist LU 135252 (50 mg/kg per d), n � 10• A group of Sprague-Dawley rats treated with the combined ETA/
ETB receptor antagonist LU 224332 (50 mg/kg per d), n � 10
We used Sprague-Dawley rats as controls, because the geneticbackground of the Han:SPRD rats are Sprague-Dawley rats.
Treatment with the endothelin receptor antagonists (13) started in8-wk-old male heterozygous (cy/�) Han:SPRD rats or male Sprague-Dawley rats of the same age. The animals were treated for 4 mo.Drugs were given orally within the food. At the end of the drugtreatment period, all rats were placed into a metabolic cage for 24 h.Urine was collected, and blood was taken at the end of the urine-collection period. All animal experiments were conducted in accor-dance with local institutional guidelines for the care and use oflaboratory animals.
SurgeryAnesthesia was introduced and maintained by 1 g/kg body wt urethane
intraperitoneally (Sigma). The animals were then placed on a thermo-stated table to maintain normal body temperature. After an incision in theleft groin, a polyethylene catheter (ID, 0.54 mm; OD, 0.96 mm withnarrowed tip) was implanted into the left femoral artery in such a way thatthe tip was placed distally to both renal arteries. Another catheter of thesame dimensions was inserted into the femoral vein. The latter line wasused to continuously infuse a warmed (37°C) solution of 0.9% NaCl (10ml/h per kg) throughout the surgery and the following experiment. Theabdominal cavity was then opened, and the left renal artery and vein wereprepared. An ultrasound transit time flow probe (Type 1RB, TransonicSystems) was positioned around the left renal artery.
Measurement of BP, Renal Blood Flow, and RenalVascular Resistance
BP was measured in the abdominal aorta by means of a Stathampressure transducer (Type P23Db) and a Gould pressure processor. Heartrate (HR) was recorded instantaneously with a rate meter (4600 Gouldpressure processor). Whole kidney blood flow (RBF) was measuredcontinuously via the ultrasound transit time flowprobe placed around the
renal artery (ID, 1 mm). For further details, see reference 14. Renalvascular resistance (RVR) was calculated using the following formula:RVR � (BP-20mmHg)/RBF. The offset of 20 mmHg is an adjustmentfor the so-called waterfall phenomenon of the kidney.
GFR and Clinical ChemistrySerum concentrations of alanine aminotransferase, creatinine, pro-
tein, and albumin and urine concentrations of creatinine, protein, andalbumin were determined by using the appropriate kits in an automaticanalyzer in the department of clinical biochemistry and laboratorymedicine of the Charite University hospital. The absolute excretion ofproteins was calculated by the equation: Ux � V24, where Ux is theconcentration of protein in urine, and V24 is the amount of urineexcreted in 24 h. The endogenous creatinine clearance was calculatedusing the formula C � Uc � Uvol/Sc, where C is creatinine clearance,Uc is urinary creatinine concentration, Uvol is urine volume, and Scis serum creatinine concentration.
Histologic EvaluationFor pathohistologic evaluation, all samples were embedded in
paraffin, cut in 3-�m sections, and submitted to hematoxylin-eosin,Sirius red, and periodic acid-Schiff (PAS) staining and analyzed asrecently described (15). The severity of interstitial matrix depositionwas evaluated after Sirius red staining using a computer-aided image-analyzing system. We measured the relationship of red stained inter-stitial area (connective tissue) to total interstitial area of the wholekidney section. Interstitial area was defined as total section area minuscyst area minus glomerular area minus area of blood vessels. Glo-merulosclerosis was defined by the presence of PAS-positive materialwithin the glomeruli. To consider differences in the degree of glo-merulosclerosis, a semiquantitative scoring system was used as re-cently described (15). All tissue samples for scoring were evaluatedindependently by two investigators without prior knowledge of thegroup to which the rats belonged.
In Vivo BrdU IncorporationIn vivo BrdU incorporation was performed exactly as recently
described (16). Briefly, we injected BrdU intraperitoneally to label theDNA in vivo 24 h before scarifying the rats. Paraffin sections weredewaxed and enzymatic digested with trypsin. After washing, thesections were treated with 4 M HCl followed by a neutralizing step.The probes were subsequently incubated with anti-BrdU-AP antibodysolution. Substrate reaction was then started with a sufficient amountof the freshly prepared substrate solution and incubated at room
Table 3. Cell proliferation in kidneys of HanSPRD rats after 4 mo of treatment with ETA or combined ETA/ETB receptorantagonistsa
HanSPRDRats
HanSPRD Rats� LU 135252
HanSPRD Rats� LU 224332
Glomerular cell proliferation (BrdU-positive cells/1000 cells) 5.5 � 2.9e 18.4 � 10.8e 12.9 � 4.5c
Interstitial cell proliferation (BrdU-positive cells/1000 cells) 16.9 � 8.0 26.4 � 13.9 16.5 � 3.5Tubular cell proliferation (BrdU-positive cells/1000 cells) 42.2 � 17.3b 339.4 � 286.9e 123.9 � 19.5d
Epithelial cyst wall cell proliferation (BrdU-positive cells/1000 cells) 9.1 � 6.4 7.2 � 4.0 8.2 � 1.7
a Cell proliferation was analyzed by in vivo BrdU incorporation. Data are given as means � SD.b P � 0.05 compared with glomerular, interstitial, and cyst wall epithelial cells of nontreated Han:SPRD rats.c P � 0.05 compared with the same cell type of nontreated heterozygous (cy/�) Han:SPRD rats.d P � 0.01 compared with the same cell type of nontreated heterozygous (cy/�) Han:SPRD rats.e P � 0.001 compared-with the same cell type of nontreated heterozygous (cy/�) Han:SPRD rats.
J Am Soc Nephrol 14: 367–376, 2003 ADPKD and Endothelin System 371

temperature for 20 min until a clearly visible color developed. Theslides were washed and embedded with glycerine/PBS. We countedBrdU-positive cells and the total cell number to calculate a prolifer-ation index (BrdU-positive cells/total cell number). Cell proliferationwas analyzed separately in the glomeruli, tubular cells, blood vessels,interstitial cells, and epithelial cells of the cystic wall.
Detection of Apoptotic CellsTo evaluate apoptotic cells, we used a combined TUNEL double-
staining method (Roche, Mannheim, Germany) as recently described(17). Briefly, paraffin kidney sections were incubated with digoxige-nin-dUTP in the presence of TdT. TUNEL-positive cells were visu-alized by anti-digoxigenin FITC-conjugated F(ab[prime])2 fragments.Counterstaining was performed using DAPI dye (1 �g/ml methanol)in a subsequent incubating step. Finally, sections were mounted usingVectaShield (Vector Laboratories, Burlingame, VT). Spleen served as
positive control. Negative controls for TUNEL staining were made byomitting TdT, according to the manufacturer’s protocol.
Statistical AnalysisThe Mann-Whitney U test was used to compare groups. The level
of significance was set at P � 0.05. Statistical analysis was performedusing SPSS for Windows, Version 11.0.
ResultsLong-term treatment with LU 135252 and LU 224332 had
no nonspecific or toxic side effects. All Sprague-Dawleyrats (the genetic background of the Han:SPRD rats areSprague-Dawley rats) survived. They grew well and werehealthy (Table 1). Light microscopy of the kidneys revealedcompletely normal kidney morphology with no abnormali-ties within the tubules. Cysts were not detectable in LU135252-treated and LU 224332-treated Sprague-Dawleyrats.
The heterozygous (cy/�) Han:SPRD rats also toleratedtreatment with both endothelin receptor antagonists (LU135252 or LU 224332) without adverse events. No toxic sideeffects (elevated liver enzymes, infectious diseases, clinicalsigns of diseases, or increased mortality) were seen in the LU135252-treated or LU 224332-treated ADPKD rats. We ob-served differences with respect to serum protein and C-reactiveprotein between nontreated and treated Han:SPRD rats; how-ever, all the values were within the normal range for theseparameters for rats in our laboratory. We thus suggest thatthese differences, although statistically significant, are of mi-nor pathophysiologic impact. The body weight of rats treatedwith the ETA receptor antagonist, on the other hand, wassignificantly lower as compared with nontreated ADPKD rats(Table 2). Treatment with the endothelin receptor antagonistsaffected neither urinary total protein excretion nor urinaryalbumin excretion (Table 2).
ETA receptor antagonist–treated ADPKD rats had a mark-edly increased kidney weight, increased number of renal cysts,and increased cyst surface area (Table 2 and Figure 1). Theadditional blockade of the ETB receptor using the combinedETA/ETB receptor antagonist LU 224332 blunted the effectson kidney weight, number of renal cysts, and cyst surface areaas compared with a sole ETA blockade (Table 2 and Figure 1).
GFR determined by the endogenous creatinine clearancewas slightly lower in heterozygous (cy/�) Han:SPRD rats after4 mo of treatment with the ETA receptor antagonist and thecombined ETA/ETB receptor antagonist (Table 2). However,these differences were statistically NS. Mean arterial BP wassignificantly elevated in heterozygous (cy/�) Han:SPRD ratstreated for 4 mo with the combined ETA/ETB receptor antag-onist LU 224332 (Table 2).
Total renal blood flow (RBF), measured at the arteria rena-lis, decreased markedly in LU 135252–treated heterozygous(cy/�) Han:SPRD rats as compared with nontreated and LU224332–treated ADPKD rats (Figure 2A). The ETA antagonistLU 135252 caused a sustained increase of renal vascular re-sistance (RVR) (Figure 2B), because BP did not fall in this
Figure 4. Typical kidney section showing tubular cell proliferation innontreated ADPKD rats (A) and LU 135252 (ETA receptor antago-nist)–treated ADPKD rats (B). Cell proliferation was analyzed by thein vivo BrdU incorporation method. BrdU incorporation is visible byred granular. The already enhanced tubular cell proliferation in AD-PKD rats was increased about eightfold after Lu 135252 treatment.Treatment of ADPKD rats with LU 224332 blunted the effects of asole ETA receptor blockade.
372 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 367–376, 2003

group. The additional blockade of the ETB receptor using LU224332 attenuated this effect.
Glomerulosclerosis was analyzed using a semiquantita-tive score (see Materials and Methods). Long-term treat-ment with both endothelin receptor antagonists (LU 135252and LU 224332) had no effect on glomerulosclerosis inheterozygous (cy/�) Han:SPRD rats (Table 2), whereasinterstitial fibrosis was enhanced in LU 135252-treated aswell as LU 224332-treated Han:SPRD rats (Table 2 andFigure 3).
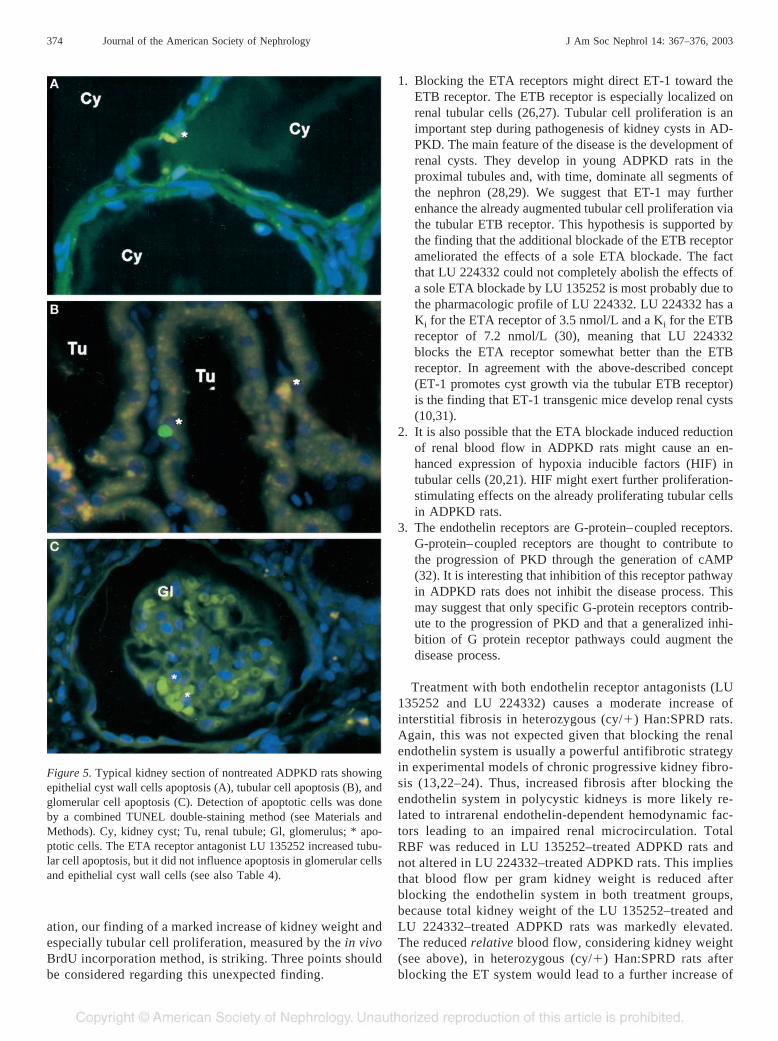
The most remarkable findings were the huge increasedkidney weight, increased number of renal cysts, and in-creased cyst surface area in Han:SPRD rats after long-termblockade of the ETA receptor. We thus analyzed cell pro-liferation in the kidneys of Han:SPRD rats by the in vivoBrdU incorporation method in glomerular cells, interstitialcells, tubular cells, and epithelial cells of the cyst wall.These analyses revealed that cell proliferation is highest intubular cells as compared with all other cell types in non-treated Han:SPRD rats. Long-term treatment with the ETAreceptor antagonist led to a markedly increased (�804.2%;P � 0.001) cell proliferation in tubular cells of Han:SPRDrats (Table 3 and Figure 4). Cell proliferation was alsoelevated (�334.5%; P � 0.001) in glomerular cells of LU135252–treated Han:SPRD rats. The additional blockade ofthe ETB receptor ameliorated the effects of a sole ETAreceptor blockade on tubular and on glomerular cell prolif-eration (Table 3). To analyze the relationship between cellgrowth and cell death, we also performed a combinedTUNEL double-staining assay to detect apoptotic cells inthe kidneys of Han:SPRD rats. Apoptotic cells in nontreatedHan:SPRD rats were seen in epithelial cyst wall cells.Tubular cell apoptosis was also detectable in Han:SPRDrats, and the ETA receptor antagonist LU 135252 specifi-cally enhances tubular cell apoptosis (Table 4 and Figure 5).Although tubular cell apoptosis increases after LU 135252treatment, it was obvious that LU 135252 enhances theimbalance between cell growth and cell death, because thedifference between the total number of proliferating tubularcells and apoptotic tubular cells increases after LU 135252treatment. In nontreated Han:SPRD rats, we saw 42.2 �17.3 proliferating cells per 1000 tubular cells and 0.06 �0.08 apoptotic cells per 10000 tubular cells; in LU 135252-treated Han:SPRD rats, we detected 339.4 � 286.9 prolif-
erating cells per 1000 tubular cells and 2.23 � 1.87 apo-ptotic cells per 10000 tubular cells (Tables 3 and 4).
DiscussionThe endothelin system is activated in Han:SPRD rats as well
as in humans with ADPKD (7). The reasons for this activationare presently unknown. The known genetic defects in patientswith ADPKD as well as in the rat model of ADPKD analyzedin this study are not related to genes of the endothelin system(1,2,12). However, it was suggested that cyst growth causesfocal ischemia (18); ischemia on its own, via an induction ofhypoxia-inducible factors (19,20), is one of the most potentstimuli of the endothelin system in vivo (21). Given this po-tential pathway of activating the endothelin system in ADPKD,long-term treatment studies with endothelin receptor antago-nists using Han:SPRD rats are most probably of major impactalso for the human disease.
A blockade of especially the ETA receptor induces amarked increase of kidney weight, increased number ofrenal cysts, and increased cyst surface area accompanied byan approximately eightfold increased cell proliferation rateof tubular cells. The (in terms of absolute numbers) negli-gible increase in tubular cell apoptosis has only a minorimpact on the ETA receptor–induced alterations in tubularcell turnover in Han:SPRD rats. To our knowledge, this isthe first study showing that long-term blockade of the ETAreceptor increases cell proliferation in vivo. The additionalblockade of the ETB receptor seems to attenuate the effectsof a sole ETA receptor blockade. Induction of growth/cellproliferation by blocking the ETA receptor is only seen inthe polycystic kidney; other organs of these ADPKD rats arenot affected (data not shown). Neither kidney function norkidney morphology was altered by both endothelin receptorantagonists in healthy Sprague-Dawley rats (the geneticbackground of the Han:SPRD rats).
Blocking the ETA receptor usually decreases cell prolif-eration. The ETA receptor antagonist used in our study (LU135252) decreases cell proliferation and kidney fibrosis invarious models of progressive kidney failure/fibrosis likediabetic nephropathy (13), chronic renal allograft rejection(22,23), and rats with surgical renal mass ablation (24). ETAreceptor antagonism was furthermore shown to inhibit pros-tate cancer cell proliferation (25). Given the strong evidencethat ETA receptor antagonists usually inhibit cell prolifer-
Table 4. Apoptotic cells in kidneys of HanSPRD rats after 4 mo of treatment with ETA or combined ETA/ETB receptorantagonistsa
HanSPRD Rats HanSPRD Rats� LU 135252
HanSPRD Rats� LU 224332
Apoptotic glomerular cells (apoptotic cells/10000 cells) 0.61 � 1.29 0.85 � 1.56 0.61 � 0.84Apoptotic interstitial cells (apoptotic cells/10000 cells) 0.21 � 0.18 0.32 � 0.33 0.21 � 0.13Apoptotic tubular cells (apoptotic cells/10000 cells) 0.06 � 0.08 2.23 � 1.87b 0.06 � 0.13Apoptotic epithelial cyst wall cells (apoptotic cells/10000 cells) 1.05 � 1.81 0.23 � 0.41 0.92 � 0.58
a Apoptosis was analyzed by a combined TUNEL double-staining method. Data are given as means � SD.b P � 0.001 compared with the same cell type of nontreated heterozygous (cy/�) Han:SPRD rats.
J Am Soc Nephrol 14: 367–376, 2003 ADPKD and Endothelin System 373
ation, our finding of a marked increase of kidney weight andespecially tubular cell proliferation, measured by the in vivoBrdU incorporation method, is striking. Three points shouldbe considered regarding this unexpected finding.
1. Blocking the ETA receptors might direct ET-1 toward theETB receptor. The ETB receptor is especially localized onrenal tubular cells (26,27). Tubular cell proliferation is animportant step during pathogenesis of kidney cysts in AD-PKD. The main feature of the disease is the development ofrenal cysts. They develop in young ADPKD rats in theproximal tubules and, with time, dominate all segments ofthe nephron (28,29). We suggest that ET-1 may furtherenhance the already augmented tubular cell proliferation viathe tubular ETB receptor. This hypothesis is supported bythe finding that the additional blockade of the ETB receptorameliorated the effects of a sole ETA blockade. The factthat LU 224332 could not completely abolish the effects ofa sole ETA blockade by LU 135252 is most probably due tothe pharmacologic profile of LU 224332. LU 224332 has aKi for the ETA receptor of 3.5 nmol/L and a Ki for the ETBreceptor of 7.2 nmol/L (30), meaning that LU 224332blocks the ETA receptor somewhat better than the ETBreceptor. In agreement with the above-described concept(ET-1 promotes cyst growth via the tubular ETB receptor)is the finding that ET-1 transgenic mice develop renal cysts(10,31).
2. It is also possible that the ETA blockade induced reductionof renal blood flow in ADPKD rats might cause an en-hanced expression of hypoxia inducible factors (HIF) intubular cells (20,21). HIF might exert further proliferation-stimulating effects on the already proliferating tubular cellsin ADPKD rats.
3. The endothelin receptors are G-protein–coupled receptors.G-protein–coupled receptors are thought to contribute tothe progression of PKD through the generation of cAMP(32). It is interesting that inhibition of this receptor pathwayin ADPKD rats does not inhibit the disease process. Thismay suggest that only specific G-protein receptors contrib-ute to the progression of PKD and that a generalized inhi-bition of G protein receptor pathways could augment thedisease process.
Treatment with both endothelin receptor antagonists (LU135252 and LU 224332) causes a moderate increase ofinterstitial fibrosis in heterozygous (cy/�) Han:SPRD rats.Again, this was not expected given that blocking the renalendothelin system is usually a powerful antifibrotic strategyin experimental models of chronic progressive kidney fibro-sis (13,22–24). Thus, increased fibrosis after blocking theendothelin system in polycystic kidneys is more likely re-lated to intrarenal endothelin-dependent hemodynamic fac-tors leading to an impaired renal microcirculation. TotalRBF was reduced in LU 135252–treated ADPKD rats andnot altered in LU 224332–treated ADPKD rats. This impliesthat blood flow per gram kidney weight is reduced afterblocking the endothelin system in both treatment groups,because total kidney weight of the LU 135252–treated andLU 224332–treated ADPKD rats was markedly elevated.The reduced relative blood flow, considering kidney weight(see above), in heterozygous (cy/�) Han:SPRD rats afterblocking the ET system would lead to a further increase of
Figure 5. Typical kidney section of nontreated ADPKD rats showingepithelial cyst wall cells apoptosis (A), tubular cell apoptosis (B), andglomerular cell apoptosis (C). Detection of apoptotic cells was doneby a combined TUNEL double-staining method (see Materials andMethods). Cy, kidney cyst; Tu, renal tubule; Gl, glomerulus; * apo-ptotic cells. The ETA receptor antagonist LU 135252 increased tubu-lar cell apoptosis, but it did not influence apoptosis in glomerular cellsand epithelial cyst wall cells (see also Table 4).
374 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 367–376, 2003

the already existing cyst growth–related focal ischemia inADPKD rats (18). Ischemia on its own is a well-knownpro-fibrotic stimulus (for review, see reference 33). A veryrecent study also demonstrates that long-term treatment withan ETA receptor antagonist in rats with two kidney-one clip(2K-1C) renovascular hypertension increases fibrosis in theclipped (ischemic) kidney (15).
The combined ETA/ETB receptor antagonist led to a mildincrease in mean arterial BP as compared with nontreatedADPKD rats. Beside the effects on kidney fibrosis and cystgrowth, the combined ETA/ETB receptor antagonist mighthave additional effects, via the ETB receptor, on renal waterand salt excretion in ADPKD rats. It was recently shown thatthe complete absence of a functional ETB receptor in rats maycause salt-sensitive elevation of BP due to an enhanced tubularsalt reuptake (16,34).
In conclusion, our study indicates that the activated endo-thelin system in heterozygous (cy/�) Han:SPRD rats is sub-stantially involved in the regulation of tubular cell proliferationand cyst growth (cyst size and number). It also plays animportant role in the development of interstitial fibrosis. Thus,the endothelin system is a major disease-modifying system inADPKD. This is of clinical impact, because endothelin recep-tor antagonists will become new clinically used drugs in nearfuture. On the basis of our data, especially sole ETA receptorantagonists but also combined ETA/ETB receptor antagonistsmight be harmful for patients with ADPKD.
AcknowledgmentsThis study was supported by the Else-Kroner Fresenius Stiftung to
B. Hocher and B. Nafz and partially by a grant from the DeutscheForschungsgemeinschaft (Ho 1665/5–1) to B. Hocher.
References1. Qian Q, Harris PC, Torres VE; Treatment prospects for autoso-
mal-dominant polycystic kidney disease. Kidney Int 59: 2005–2022, 2001
2. Arnaout MA: Molecular genetics and pathogenesis of autosomal dom-inant polycystic kidney disease. Annu Rev Med 52: 93–123, 2001
3. Ecder T, Schrier RW: Hypertension in autosomal-dominant poly-cystic kidney disease: Early occurrence and unique aspects. J AmSoc Nephrol 12: 194–200, 2001
4. Peters DJ, Breuning M: Autosomal dominant polycystic kidneydisease: Modification of disease progression. Lancet 358: 1439–1444, 2001
5. Gretz N, Ceccherini I, Kranzlin B, Kloting I, Devoto M, Rohm-eiss P, Hocher B, Waldherr R, Romeo G: Gender-dependentdisease severity in autosomal polycystic kidney disease of rats.Kidney Int. 48: 496–500, 1995
6. Giusti R, Neri M, Angelini D, Carlini A, Fiorini I, Bigongiari P,Antonelli A: Plasma concentration of endothelin and arterial pres-sure in patients with ADPKD. Contrib Nephrol 115: 118–121, 1995
7. Hocher B, Zart R, Schwarz A, Vogt V, Braun C, Thone-ReinekeC, Braun N, Neumayer HH, Koppenhagen K, Bauer C, RohmeissP: Renal endothelin system in polycystic kidney disease. J AmSoc Nephrol 9: 1169–1177, 1998
8. Munemura C, Uemasu J, Kawasaki H: Epidermal growth factorand endothelin in cyst fluid from autosomal dominant polycystic
kidney disease cases: Possible evidence of heterogeneity in cys-togenesis. Am J Kidney Dis 24: 561–568, 1994
9. Nakamura T, Ebihara I, Fukui M, Osada S, Tomino Y, Masaki T,Goto K, Furuichi Y, Koide H: Increased endothelin and endo-thelin receptor mRNA expression in polycystic kidneys of cpkmice. J Am Soc Nephrol 4: 1064–1072, 1993
10. Hocher B, Thone-Reineke C, Rohmeiss P, Schmager F, Slowin-ski T, Burst V, Siegmund F, Quertermous T, Bauer C, NeumayerHH, Schleuning WD, Theuring F Endothelin-1 transgenic micedevelop glomerulosclerosis, interstitial fibrosis, and renal cystsbut not hypertension. J Clin Invest 99: 1380–1389, 1997
11. Grantham JJ: The etiology, pathogenesis, and treatment of auto-somal dominant polycystic kidney disease: Recent advances.Am J Kidney Dis 28: 788–803, 1996
12. Nagao S, Ushijima T, Kasahara M, Yamaguchi T, Kusaka M,Matsuda J, Nagao M, Takahashi H: Closely linked polymorphicmarkers for determining the autosomal dominant allele (Cy) inrat polycystic kidney disease. Biochem Genet 37: 227–235, 1999
13. Hocher B, Schwarz A, Reinbacher D, Jacobi J, Lun A, Priem F,Bauer C, Neumayer HH, Raschack M: Effects of endothelinreceptor antagonists on the progression of diabetic nephropathy.Nephron 87: 161–169, 2001
14. Nafz B, Berger K, Rosler C, Persson PB: Kinins modulate thesodium-dependent autoregulation of renal medullary blood flow.Cardiovasc Res 40: 573–579, 1998
15. Hocher B, George I, Diekmann F, Zart R, Rebstock J, SchwarzA, Thone-Reineke C, Neumayer HH, Bauer C: ETA receptorblockade induces fibrosis of the clipped kidney in two-kidney-one-clip renovascular hypertensive rats. J Hypertens 18: 1807–1814, 2000
16. Hocher B, Dembowski C, Slowinski T, Friese ST, Schwarz A,Siren AL, Neumayer HH, Thone-Reineke C, Bauer C, Nafz B,Ehrenreich. Impaired sodium excretion, decreased glomerularfiltration rate and elevated blood pressure in endothelin receptortype B deficient rats. J Mol Med 78: 633–641, 2001
17. Arck PC, Handjiski B, Hagen E, Joachim R, Klapp BF, Paus R:Indications for a ’brain-hair follicle axis (BHA)’: Inhibition ofkeratinocyte proliferation and up-regulation of keratinocyte ap-optosis in telogen hair follicles by stress and substance P. FASEBJ 15: 2536–2538, 2001
18. Wang D, Strandgaard S: The pathogenesis of hypertension inautosomal dominant polycystic kidney disease. J Hypertens 15:925–933, 1997
19. Yoshida T, Kuwahara M, Maita K, Harada T Immunohistochem-ical study on hypoxia in spontaneous polycystic liver and kidneydisease in rats. Exp Toxicol Pathol 53: 123–128, 2001
20. Kakinuma Y, Miyauchi T, Yuki K, Murakoshi N, Goto K,Yamaguchi I: Novel molecular mechanism of increased myocar-dial endothelin-1 expression in the failing heart involving thetranscriptional factor hypoxia-inducible factor-1alpha inducedfor impaired myocardial energy metabolism. Circulation 103:2387–2394, 2001
21. Schiffrin EL: Role of endothelin-1 in hypertension and vasculardisease. Am J Hypertens 14: 83S–89S, 2001
22. Braun C, Conzelmann T, Vetter S, Schaub M, Back WE, Yard B,Kirchengast M, Tullius SG, Schnulle P, van der Woude FJ,Rohmeiss. Prevention of chronic renal allograft rejection in ratswith an oral endothelin A receptor antagonist. Transplantation68: 739–746, 1999
23. Orth SR, Odoni G, Amann K, Strzelczyk P, Raschack M, Ritz E:The ET(A) receptor blocker LU 135252 prevents chronic trans-
J Am Soc Nephrol 14: 367–376, 2003 ADPKD and Endothelin System 375

plant nephropathy in the “Fisher to Lewis” model. J Am SocNephrol 10: 387–391, 1999
24. Brochu E, Lacasse S, Moreau C, Lebel M, Kingma I, Grose JH,Lariviere R Endothelin ET(A) receptor blockade prevents theprogression of renal failure and hypertension in uraemic rats.Nephrol Dial Transplant 14: 1881–1888, 1999
25. Nelson JB, Hedican SP, George DJ, Reddi AH, Piantadosi S,Eisenberger MA, Simons JW: Identification of endothelin-1 inthe pathophysiology of metastatic adenocarcinoma of the pros-tate. Nat Med 1: 944–949, 1995
26. Hocher B, Rohmeiss P, Diekmann F, Zart R, Vogt V, Schiller S,Bauer C, Koppenhagen K, Distler A, Gretz N: Distribution ofendothelin receptor subtypes in the rat kidney. Renal and haemo-dynamic effects of the mixed (A/B) endothelin receptor antagonistbosentan. Eur J Clin Chem Clin Biochem 33: 463–472, 1995
27. Dean R, Zhuo J, Alcorn D, Casley D, Mendelsohn FA: Cellularlocalization of endothelin receptor subtypes in the rat kidney followingin vitro labeling. Clin Exp Pharmacol Physiol 23: 524–531, 1996
28. Ramasubbu K, Gretz N, Bachmann S: Increased epithelial cellproliferation and abnormal extracellular matrix in rat polycystickidney disease. J Am Soc Nephrol 9: 937–945, 1998
29. Murcia NS, Sweeney WE Jr, Avner ED: New insights into themolecular pathophysiology of polycystic kidney disease. KidneyInt 55: 1187–1197, 1999
30. Raschack M, Gock S, Unger L, Hahn A, Amberg W, Jansen R,Alken P, Weber A, Hergenroder S: LU 302 872 and its racemate(LU 224 332) show balanced endothelin-A/B receptor affinity,high oral activity, and. inhibit human prostate tissue contractions.J Cardiovasc Pharmacol. 31: S241–S24, 1998
31. Shindo T, Kurihara H, Maemura K, Kurihara Y, Ueda O, SuzukiH, Kuwaki T, Ju KH, Wang Y, Ebihara A, Nishimatsu H,Moriyama N, Fukuda M, Akimoto Y, Hirano H, MoritaH, Kumada M, Yazaki Y, Nagai R, Kimura K: Renal dam-age and salt-dependent hypertension in aged transgenicmice overexpressing endothelin-1. J Mol Med 80: 105–116, 2002
32. Sullivan LP, Wallace DP, Grantham JJ: Epithelial transport inpolycystic kidney disease. Physiol Rev 78: 1165–1191, 1998
33. Meyrier A, Hill GS, Simon P: Ischemic renal diseases: Newinsights into old entities. Kidney Int. 54: 2–13, 1998
34. Gariepy CE, Ohuchi T, Williams SC, Richardson JA, Yanagi-sawa M: Salt-sensitive hypertension in endothelin-B receptor-deficient rats. J Clin Invest 105: 925–933, 2000
376 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 367–376, 2003
Related Documents











