........................................................................................................................... Estrogen receptor b: the guardian of the endometrium D.K. Hapangama 1, * , A.M. Kamal 1,2 , and J.N. Bulmer 3 1 Department of Women’s and Children’s Health, Institute of Translational Medicine, University of Liverpool, Liverpool Women’s NHS Foundation Trust, Liverpool L8 7SS, UK 2 The National Center for Early Detection of Cancer, Oncology Teaching Hospital, Baghdad Medical City, Baghdad, Iraq 3 Reproductive and Vascular Biology Group, Institute of Cellular Medicine, Newcastle University, Newcastle upon Tyne NE2 4HH, UK *Correspondence address. E-mail: [email protected] Submitted on June 30, 2014; resubmitted on August 20, 2014; accepted on August 27, 2014 table of contents † Introduction Human endometrial anatomy Human endometrium as an estrogen-receptive organ † Methods † Estrogen receptors ERb structure ERb receptor activation † ERb expression in normal endometrium Evidence from transgenic mice: ERbKO mice Primate studies ERb in the endometrial vasculature ERb in endometrial immune cells ERb in the human uterus Role of ERb in endometrial regeneration ERb expression in the healthy human post-menopausal endometrium † ERb in endometrial pathology Endometriosis and adenomyosis Polycystic ovary syndrome Endometrial polyps Endometrial hyperplasia Endometrial cancer † Pharmacological regulation of endometrial ERb expression Evidence for regulation of endometrial ERb expression by exogenous hormone treatment Evidence for SERMs (ERb selective pharmacological agents) in endometrium † Summary/conclusion background: The endometrium is the primary target organ for the ‘female’ sex steroid hormone estrogen, which exerts effects in the endo- metrium via two main classical estrogen receptor (ER) isoforms, ERa and ERb. The main function of the endometrium, embryo implantation, appears unperturbed in ERb knockout mice, which has led researchers to disregard other potentially important functional roles that ERb may have in endometrium. This review focuses on ERb in the human endometrium and its protective role from the undesired effects of ERa. methods: We conducted a systematic search using PubMed and Ovid for publications between January 1996 and February 2014. All studies that examined ERb expression or function in non-pregnant endometrium or cells derived from the endometrium were considered, including human and animal studies. & The Author 2014. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved. For Permissions, please email: [email protected] Human Reproduction Update, Vol.21, No.2 pp. 174 – 193, 2015 Advanced Access publication on October 10, 2014 doi:10.1093/humupd/dmu053 at Universitas Diponegoro on March 10, 2015 http://humupd.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

...........................................................................................................................
Estrogen receptor b: the guardianof the endometriumD.K. Hapangama1,*, A.M. Kamal1,2, and J.N. Bulmer3
1Department of Women’s and Children’s Health, Institute of Translational Medicine, University of Liverpool, Liverpool Women’s NHS FoundationTrust, Liverpool L8 7SS, UK 2The National Center for Early Detection of Cancer, Oncology Teaching Hospital, Baghdad Medical City, Baghdad, Iraq3Reproductive and Vascular Biology Group, Institute of Cellular Medicine, Newcastle University, Newcastle upon Tyne NE2 4HH, UK
*Correspondence address. E-mail: [email protected]
Submitted on June 30, 2014; resubmitted on August 20, 2014; accepted on August 27, 2014
table of contents
† IntroductionHuman endometrial anatomyHuman endometrium as an estrogen-receptive organ
† Methods† Estrogen receptors
ERb structureERb receptor activation
† ERb expression in normal endometriumEvidence from transgenic mice: ERbKO micePrimate studiesERb in the endometrial vasculatureERb in endometrial immune cellsERb in the human uterusRole of ERb in endometrial regenerationERb expression in the healthy human post-menopausal endometrium
† ERb in endometrial pathologyEndometriosis and adenomyosisPolycystic ovary syndromeEndometrial polypsEndometrial hyperplasiaEndometrial cancer
† Pharmacological regulation of endometrial ERb expressionEvidence for regulation of endometrial ERb expression by exogenous hormone treatmentEvidence for SERMs (ERb selective pharmacological agents) in endometrium
† Summary/conclusion
background: The endometrium is the primary target organ for the ‘female’ sex steroid hormone estrogen, which exerts effects in the endo-metrium via two main classical estrogen receptor (ER) isoforms, ERa and ERb. The main function of the endometrium, embryo implantation,appears unperturbed in ERb knockout mice, which has led researchers to disregard other potentially important functional roles that ERb mayhave in endometrium. This review focuses on ERb in the human endometrium and its protective role from the undesired effects of ERa.
methods: We conducted a systematic search using PubMed and Ovid for publications between January 1996 and February 2014. All studiesthat examined ERb expression or function in non-pregnant endometrium or cells derived from the endometrium were considered, includinghuman and animal studies.
& The Author 2014. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved.For Permissions, please email: [email protected]
Human Reproduction Update, Vol.21, No.2 pp. 174–193, 2015
Advanced Access publication on October 10, 2014 doi:10.1093/humupd/dmu053
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

results: Studies of the basic function of ERb isoforms in restraining ERa-mediated cell-specific trophic/mitotic responses to estrogen in othertissues has allowed appreciation of the important potential role of ERb in the regulation of cell fate in the human endometrium. Our current under-standing of ERb expression and function in endometrium is, however, incomplete. ERb is dynamically expressed in healthy premenstrual endo-metrium, persists in post-menopausal atrophic endometrium and may play an important role in endometrial disease. All endometrial cell typesexpress ERb and aberrations in ERb expression have been reported in almost all benign and malignant endometrial proliferative disease.
conclusions: The collective evidence suggests that ERb has an important role in normal endometrial function and also in most, if not all,benign and malignant endometrial diseases. However, the conduct of studies of endometrial ERbexpression needs to be standardized: agreementis needed regarding the most appropriate control tissue for endometrial cancer studies as well as development of standardized methods for thequantification of ERb immunohistochemical data, similar to those scoring systems employed for other hormonally regulated tissues such as breastcancer, since these data may have direct clinical implications in guiding therapy.
Key words: endometrium / estrogen receptor b / hormone receptors / endometrial proliferation / endometrial cancer
IntroductionThe endometrium is the primary target organ for the ‘female’ sex steroidhormone estrogen (Katzenellenbogen, 1984; Greaves et al., 2013). Theeffects of estrogens are exerted in the endometrium via two main classic-al estrogen receptor (ER) isoforms, ERa and ERb, and perhaps via therecently described G-protein-coupled estrogen receptor (GPER; for-merly GPR30) (Critchley and Saunders, 2009; Holm and Nilsson,2013). The main function of the endometrium, embryo implantation,appears to be unperturbed in ERb knockout (KO) mice, which has ledresearchers to disregard other potentially important functional rolesthat ERb may have in endometrium (Burns and Korach, 2012). Studiesfrom other organs have confirmed the opposing actions of ERb onERa function (Gustafsson, 2003; Bottner et al., 2014). Evidence for theinvolvement of ERb has been reported in almost all gynaecological path-ologies including menorrhagia, endometriosis, infertility and endometrialcancer (EC), as well as in normal and abnormal pregnancy-related con-ditions (Fernandez et al., 2012; Haring et al., 2012a; Hu et al., 2012).The classical trophic effects of ERa are reviewed elsewhere (Arnalet al., 2013) and this review focuses on the less well-described receptor,ERb, and its role in protecting the human endometrium from theundesired effects of ERa.
Human endometrial anatomyThe endometrium is the mucosal lining of the uterus and is derived fromthe inner layer of the embryonic paramesonephric ductal mesenchyme(McCluggage, 2011). Endometrial development and function in menstru-ating upper order primates (including humans) is complex comparedwith most other mammals (Slayden and Brenner, 2004; Jabbour et al.,2006). The human endometrium is stratified into two functional layers:the transient superficial stratum functionalis and the permanentdeeper stratum basalis adjacent to the myometrium (Ferenczy and Ber-geron, 1991). The superficial stratum functionalis is lined by luminal epi-thelium, contains superficial glandular epithelium and stroma and iscompletely shed and regenerated during the monthly menstrual cycleand after childbirth (Ferenczy and Guralnicik, 1983; Gargett et al.,2008). It can be divided into the deeper zona spongiosa with a looselyorganized stromal zone and a superficial zona compacta with a morecompact stroma (Ferenczy, 1980; Wynn, 1989). The stratum basaliscontains the terminal part of the endometrial glands and densely orga-nized stroma and is not shed during menses or at parturition; it
remains after cessation of ovarian cyclicity as an atrophic, inactive post-menopausal endometrium (Chhieng and Hui, 2011). The germinallayer of the endometrium where the stem cells reside is, therefore, pos-tulated to be the stratum basalis (Padykula et al., 1989; Gargett andMasuda, 2010; Valentijn et al., 2013). The other components of theendometrium, that is the blood vessels and immune cells, exist in bothlayers (Bulmer et al., 1991a; Spencer et al., 2011).
Access to the full thickness of the endometrium containing bothstratum basalis and a stratum functionalis usually requires hysterectomyand, therefore, particularly when studying endometrium either fromhealthy women or those with benign endometrial disease, researchershave mainly considered the stratum functionalis which is easily obtainedwith an outpatient endometrial biopsy (Hapangama et al., 2008a).However, the hormonal responsiveness, for example, has beenpostulated to differ between the stratum functionalis and basalis(Prianishnikov, 1978; Padykula et al., 1989). Many studies of endometrialhormone receptors have not only overlooked the structural and func-tional differences between the stratum basalis and functionalis, but alsohave discounted the exceptionally dynamic nature of the stratum functio-nalis during the normal menstrual cycle (Argenta et al., 2014). Conse-quently, compared with other estrogen-sensitive organs such as thebreast, the exact detailed mechanism of estrogen action in the endomet-rium remains unclear, with seemingly contradictory results.
Human endometrium as anestrogen-receptive organEstrogens, progesterone and androgens are the main three classicalovarian steroid hormones that exert their effects on the endometrialcells mainly via their cognate receptors (Hapangama, 2003; Slaydenand Brenner, 2004). 17b-estradiol (E2) and estrones are the two mainestrogens available for the non-pregnant endometrium, and theseexert their cellular functions through nuclear receptors, ERa and ERb,which are hormone-inducible transcription factors (Vani et al., 2008;Blair, 2010; Crandall and Barrett-Connor, 2013).
Surprisingly, compared with the large number of studies on breasttissue, for example, the number of studies investigating ERb in endomet-rium (despite it being a primary target organ for E2) has been modest.There are many reviews of ERb which discuss most other organs andcompletely disregard endometrium as an organ in which ERb has a func-tional role (Bottner et al., 2014). Furthermore, there are no recent
Estrogen receptor b in the endometrium 175
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

reviews that specifically discuss the current evidence regarding thepotential roles of ERb in the endometrium.
Although estrogens are responsible for many physiological functions inboth females and males (Couse and Korach, 1999; Harris, 2006), the evi-dence from ERa, ERb double KO mice confirms that life is possiblewithout E2 action, although ERb is almost universally expressed in allhuman organs (Lubahn et al., 1993; Dupont et al., 2000; Weihua et al.,2000). Furthermore, E2 is essential for reproductive function infemales, yet only ERa seems to be essential in preserving fertility(Lubahn et al., 1993). The main function of ERb is therefore thought tobe particularly in preventing undesired ERa-mediated actions of E2(Hall and McDonnell, 2005; Pettersen, 2011). In this review, we focuson the expression and function of ERb in non-pregnant endometrium.We discuss the available evidence regarding the involvement of ERb inpre- and post-menopausal healthy endometrium, in benign premeno-pausal endometrial pathologies and in EC, with reference to animaldata where appropriate. ERb expression and its possible action on thestratum functionalis and the germinal stratum basalis will also be exam-ined in detail. The effects and expression of ERa on the endometriumare reviewed elsewhere (Brosens et al., 2004; Jabbour et al., 2006;Critchley and Saunders, 2009).
MethodsPubMed (Medline) and Ovid searches using the key words ERb, endomet-rium, endometrial cancer, endometriosis, polycystic ovarian syndrome, in-fertility, menorrhagia and uterus were carried out systematically forpublications from January 1996 until February 2014. All studies examiningERb expression or function in non-pregnant endometrium or primary cellsor tissue explants in culture derived from the endometrium, includinghuman and animal studies and endometrial cell lines, were considered.
Estrogen receptorsThe first known ER subtype, ERa, was identified in the rat uterus in 1966(Toft and Gorski, 1966), followed by the cloning of human ERa cDNA(gene ESR1) in 1985 (Walter et al., 1985). This was followed by the dis-covery of a second ER subtype, ERb, in the prostate and ovaries of rats in1996 (Kuiper et al., 1996). The human ERb gene ESR2 was cloned first in1996 from the testis (Mosselman et al., 1996) and subsequent work hasincreased understanding of the physiological and pathological action ofE2 in human cells and tissues. E2 may also bind to transmembraneGPER, which mediates rapid signalling events traditionally associatedwith G-protein-coupled receptors; this receptor is reviewed elsewhere(Prossnitz and Barton, 2014), and it is therefore outside the remit of thisreview. Interestingly, the pivotal accepted role of ERs in reproduction is alate evolutionary development, as there is evidence suggesting that inearly order invertebrates, the reproductive role of E2 is not mediatedby ER and may take place through ancient, ER-independent pathways(Thornton et al., 2003; Keay et al., 2006). Cloning, genome mappingand phylogenetic analysis studies have indicated that ER isoforms arelikely to have been generated by duplication of the Esr gene early in thevertebrate lineage (Thornton 2001; Wu et al., 2003). It is proposedthat the functions of an ancestral gene are partitioned among duplicategenes by complementary loss of tissue-specific expression (Thornton,2001; Wu et al., 2003). The tissue-specific expression of two ER isoforms
and their splice variants, therefore,provides the potential for very flexibleregulation of target tissues by E2.
ERb can have opposing actions to ERa on the same gene promoter inresponse to E2 (Smith et al., 2004; Thomas and Gustafsson, 2011). Theseinhibitory effects of ERb on ERa activity may be exerted through a com-bination of altered recruitment of key transcription factors and increasedERa degradation (Matthews et al., 2006). In contrast, ERb expression isinduced by E2 acting via ERa and may be suppressed by hypermethyla-tion of the ERb promoter (Rody et al., 2005). The pro-proliferative func-tion of ERa is essential for reproduction, yet is associated with obviousE2-associated health risks in the endometrium (and in other organs)(Koos, 2011; May, 2014). Therefore, the ERa opposing activity of ERbhas been of particular interest with the emergence of new receptorisotype-specific pharmacological modulators.
ERb structureERb is a member of the Class I nuclear hormone receptor superfamily ofligand-inducible transcription factors and shares the common, evolution-arily conserved structural and functionally distinct domains of othersuperfamily members (Matthews et al., 2006) (Fig. 1A ). This includes acentral, highly conserved DNA binding domain (DBD), which binds tothe same estrogen responsive element (ERE) as ERa in the target genepromoters; a multifunctional ligand-binding domain (LBD) at theC-terminal; the ligand-dependent activation function 2 (AF2) at theC-terminal; and the constitutively active AF1 at the N-terminal andflexible-hinge D-domain between the LDB and the DBD (Fig. 1A)(Nilsson et al., 2001; Harnish, 2006). Transcriptional activation of ERbis facilitated by two acidic activation domains, AF1 and AF2, whichrecruit a range of specific co-regulatory protein complexes to the DNA-bound receptor (Fig. 1B) (Benecke et al., 2001). Although there is closehomology in the DBD (97%) and LBD (60%) between the two ER sub-types (Fig. 1A), significant divergence exists between the N-terminalregions, where only 20% of amino acid identity is shared. ER subtype-specific (Mosselman et al., 1996), promoter-specific and cell-specificE2 actions on target genes are therefore thought to be due to thishighly variable N-terminal domain and ligand-independent AF1 (Katze-nellenbogen et al., 2001; Hawse et al., 2008; Kumar et al., 2012).Despite their close homology, the ERb (ESR2) gene is located onchromosome 14, whereas ERa protein is coded by a different gene(ESR1) located on chromosome 6 (Menasce et al., 1993; Enmark et al.,1997). The human ERb (ESR2) gene is highly conserved with that ofother higher order primates such as chimpanzee, rhesus monkey andorang-utan (Lewandowski et al., 2002).
Five alternatively spliced transcript variants of the ERb (ESR2) genehave been described to date as ERb1–5 (Moore et al., 1998) (Fig. 1A,and published primer sequences for splice variants included in Supple-mentary data. Table SI), although the characterization of the functionalisoform pattern in human endometrium is not complete. The530-amino acid human ERb isoform is currently regarded as the wild-type ERb1 (Leygue et al., 1998). Unlike ERa, in addition to ERb1 atleast two other splice variants, ERb2 and ERb5, are transcribed withall three proteins being identical except for the C-terminus; all areexpressed as proteins and have been described in the endometrium(Collins et al., 2009). ERb2 does not bind to the ligand or make homo-dimers and the C-terminus truncations of both ERb2 and ERb5 proteinsmay affect their ligand binding capacities, although they can form
176 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from
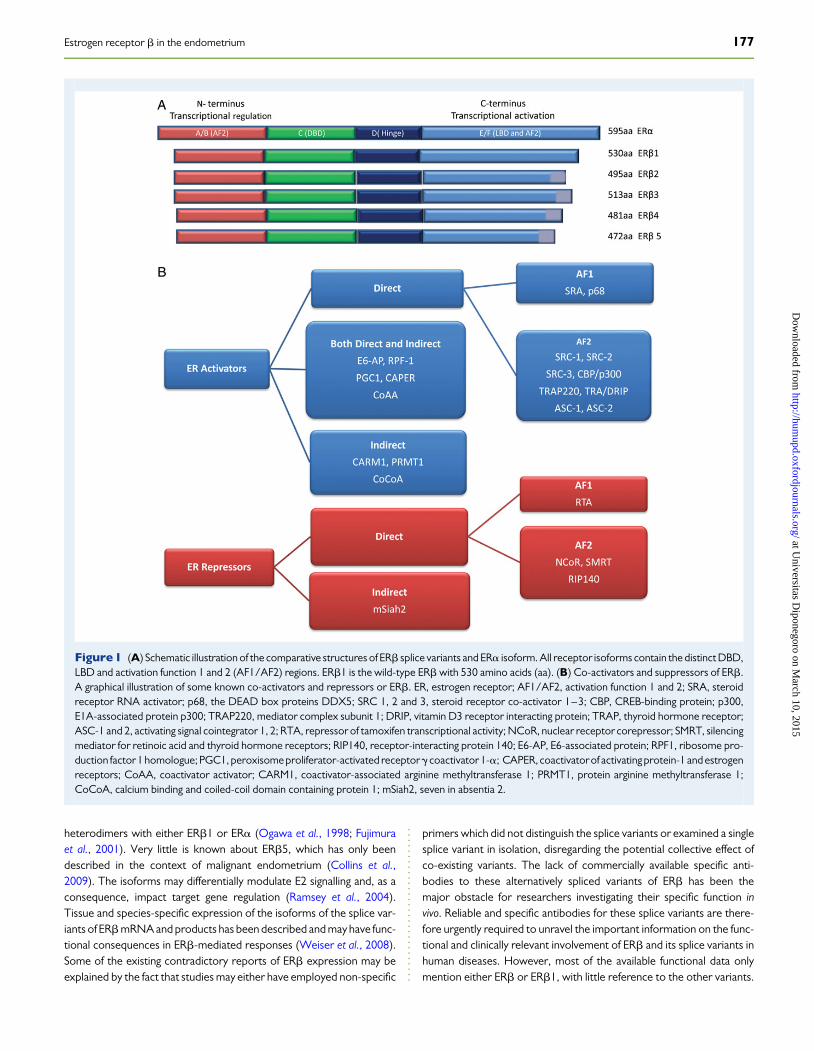
heterodimers with either ERb1 or ERa (Ogawa et al., 1998; Fujimuraet al., 2001). Very little is known about ERb5, which has only beendescribed in the context of malignant endometrium (Collins et al.,2009). The isoforms may differentially modulate E2 signalling and, as aconsequence, impact target gene regulation (Ramsey et al., 2004).Tissue and species-specific expression of the isoforms of the splice var-iants of ERbmRNA and products has been described and may have func-tional consequences in ERb-mediated responses (Weiser et al., 2008).Some of the existing contradictory reports of ERb expression may beexplained by the fact that studies may either have employed non-specific
primers which did not distinguish the splice variants or examined a singlesplice variant in isolation, disregarding the potential collective effect ofco-existing variants. The lack of commercially available specific anti-bodies to these alternatively spliced variants of ERb has been themajor obstacle for researchers investigating their specific function invivo. Reliable and specific antibodies for these splice variants are there-fore urgently required to unravel the important information on the func-tional and clinically relevant involvement of ERb and its splice variants inhuman diseases. However, most of the available functional data onlymention either ERb or ERb1, with little reference to the other variants.
Figure1 (A) Schematic illustration of the comparative structures of ERb splice variants and ERa isoform. All receptor isoforms contain the distinct DBD,LBD and activation function 1 and 2 (AF1/AF2) regions. ERb1 is the wild-type ERb with 530 amino acids (aa). (B) Co-activators and suppressors of ERb.A graphical illustration of some known co-activators and repressors or ERb. ER, estrogen receptor; AF1/AF2, activation function 1 and 2; SRA, steroidreceptor RNA activator; p68, the DEAD box proteins DDX5; SRC 1, 2 and 3, steroid receptor co-activator 1–3; CBP, CREB-binding protein; p300,E1A-associated protein p300; TRAP220, mediator complex subunit 1; DRIP, vitamin D3 receptor interacting protein; TRAP, thyroid hormone receptor;ASC-1 and 2, activating signal cointegrator 1, 2; RTA, repressor of tamoxifen transcriptional activity; NCoR, nuclear receptor corepressor; SMRT, silencingmediator for retinoic acid and thyroid hormone receptors; RIP140, receptor-interacting protein 140; E6-AP, E6-associated protein; RPF1, ribosome pro-duction factor1 homologue; PGC1, peroxisome proliferator-activated receptorg coactivator 1-a; CAPER, coactivatorof activating protein-1 and estrogenreceptors; CoAA, coactivator activator; CARM1, coactivator-associated arginine methyltransferase 1; PRMT1, protein arginine methyltransferase 1;CoCoA, calcium binding and coiled-coil domain containing protein 1; mSiah2, seven in absentia 2.
Estrogen receptor b in the endometrium 177
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

This review of the functional role of ERb therefore mainly focuses onERb1, unless otherwise stated.
ERb receptor activationERb binds E2 with similar affinity to that of ERa and upon binding to theligand, the activated receptor may exert effects involving the classicalhormone signalling pathway (Pastore et al., 2012). This involves ERb di-merizing, translocating to the nucleus, binding to the ERE located in therespective gene promoters to initiate recruitment of co-activators,co-repressors and chromatin-remodelling factors to either activate orrepress transcription of target genes (McDonnell and Norris, 2002;Saxon and Turner, 2005; Zhang and Trudeau, 2006; Zhao et al., 2008;Fig. 2). DNA binding of ERb, and hence its nuclear localization, isreported by some to be rapidly lost at body temperature when theligand, E2, is absent (Pace et al., 1997; Tan et al., 1999). Althoughthese findings have been described in the context of the classical hormo-nal pathway, recent literature examining a new class of ERb-selectivecompounds has demonstrated activation of multiple endogenousgenes through ERb by selectively recruiting ERb and co-activators totarget genes without binding to ERb (Vivar et al., 2010). This novelpathway of ligand-independent ERb activation may play an importantrole and needs to be explored further in the future to understand the col-lective and the full impact of both classical and non-classical pathways inERb function. ERb1 can form homodimers or heterodimers with ERaand ERb splice variants (e.g. ERb2), and therefore, the expression and
availability of each ER subtype in a cell will influence the cell-specific re-sponse to E2 (Kuiper and Gustafsson, 1997; Tremblay et al., 1999). Ata subcellular level, ERb is localized in the nucleus, cytoplasm and mito-chondria and this is regulated by both the availability of the ligand andby the co-expression of ERa/ERb2 (Chen et al., 2007; Milanesi et al.,2009). Furthermore, ligand-bound ERb can interact directly with othertranscription factor complexes and influence the transcription of genesthat do not possess the ERE in their promoter (Kushner et al., 2000).In the presence of E2, ERb is able to oppose the effects of tissue-specificER modulators such as tamoxifen via these indirect pathways (Paechet al., 1997). Therefore, the availability and type of the estrogenicagonist, the cellular expression of steroid receptors (including ERs andtheir respective isoforms, as well as other hormone receptors) and theexpression of co-regulators, all influence cellular expression of ERband ERb-mediated gene expression in response to estrogen in anygiven tissue type.
ERb expression in normalendometrium
Evidence from transgenic mice: ERbKO miceE2 action is essential for normal development of female sexual character-istics;ERa is the main receptor responsible for theseactions, as evidencedby the phenotype of the ERaKO mouse, which demonstrated a non-fertile and infantile phenotype (Lubahn et al., 1993). In contrast the
Figure 2 Schematic diagram of ER activation. ER binds to ligand, dimerizes, translocates to the nucleus and binds to the ERE located in the respective genepromoters to initiate recruitment of co-activators (COA), co-repressors (COR) and chromatin-remodelling factors to either activate or repress transcrip-tion of target genes (Classical pathway). Ligand-bound ER can also interact directly with other transcription factor (TF) complexes and influence the tran-scription of genes that do not possess the ERE in their promoter (non-classical pathway).
178 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

ERbKO mouse was fertile; this observation may have led to a relative lackof interest from reproductive biologists to investigate ERb expression andfunction in the endometrium (Weihua et al., 2000). Although fertile,ERbKO mice did display some reproductive deficiencies with a subfertilephenotype and an exaggeration of the endometrial epithelial proliferativeresponse to E2, suggesting a suppressive roleof ERbon the actions of ERa(Wada-Hiraike et al., 2006). However, there is evidence that ERb couldpartially compensate for the loss of ERa in the genital tract since theuterine phenotype of ERab double KO mice has been described to besimilar to that of an aggravated ERaKO uterine phenotype, whereasthe ERbKO genital tract appeared to be normal (Dupont et al., 2000).The decrease in ovarian production of E2 was thought to be responsiblefor the reduced fertility and smaller litter size seen in ERbKO animals andtherefore, these animals initially were not thought to have a defect in im-plantation or placentation (Weihua et al., 2000).
Normal endometrial epithelial cell maturation was associated with theattainment of apical–basal polarity, which relied on formation of intercel-lular adherent junctions that provides a structural foundation for normalepithelial architecture (Valentijn et al., 2013). E-cadherin, localized tothe lateral membrane of differentiated epithelia, is essential for the main-tenance of functional junctions and was deficient in ERbKO mouse endo-metrium (Wada-Hiraike et al., 2006), resulting in deformed glandulargenesis and loss of glandular differentiation. Recent work has highlightedthe importance of endometrial glands in implantation (Filant and Spencer,2013), suggesting that, despite earlier beliefs, ERbmay indeed have an im-portant role to play in the reproductive function of the endometrium.Treatment with E2 increased both stromal and epithelial ERa and ERbex-pression in wild-type, ovariectomized mice, whilst combined E2 and pro-gesterone treatment decreased expression of ERb in endometrialepithelium (Wada-Hiraike et al., 2006). Conversely, ERbKO miceshowed up-regulation of progesterone receptor (PR) in response to E2compared with the wild-type mice, suggesting that ERb represses epithe-lial PR expression (Wada-Hiraike et al., 2006). There are no reports ofdetailed examination of the proliferative effect of endometrial epithelialcells in ERa, ERb double KO mice in response to E2 and the publishedstudies only describe the uterine phenotype in comparison with wild-typemice (Dupont et al., 2000). ER subtype-specific ligand studies have alsoindicated that ERb can modulate ERa activity in a response specificmanner (Frasor et al., 2003). When ERa was selectively deleted in themouse uterine epithelium, although the E2-induced initial epithelial mito-genic response remained intact, prolonged E2 treatment induced an in-crease in epithelial apoptosis, indicating that the protective effect of E2against uterine epithelial apoptosis is mediated via ERb (Winuthayanonet al., 2010). These observations further suggest that the direct actionof E2 on endometrial epithelial cells via ERb is to induce apoptosis.
Ovariectomized ERbKO mice showed an aberrant hyper-proliferativeresponse in the absence of E2 (Weihua et al., 2000). These mice wouldhave low E2 and absent progesterone, but relatively high androgen levelsof adrenal origin. In breast epithelium, androgen up-regulated ERb via an-drogen receptor (AR) and inhibited proliferation (Rizza et al., 2014); if asimilar mechanism exists in the endometrium, the action of androgens inthe absence of ERb may be to stimulate proliferation.
Primate studiesPrimates menstruate and have an endometrial cycle and structure similarto that of humans. In primates such as macaques, ERb expression does
not change across the menstrual cycle in either the stratum functionalis orthe stratum basalis. Only ERb has been reported to be expressed in theendometrial endothelial cells throughout the menstrual cycle and ERbhas been proposed to regulate the angiogenic and vascular changesthat occur in embryo implantation, early placentation and the mainten-ance of pregnancy (Slayden and Brenner, 2004). In the marmosetmonkey, ERb was highly expressed in endometrial epithelial cellsthroughout the menstrual cycle and in pregnancy. Increased stromalERb expression was observed in the late proliferative phase with thestaining index decreasing by half as the secretory phase progressed andremaining low in pregnancy (Silvestri and Fraser, 2007). Treatmentwith GnRH agonists or ovariectomy caused significant reductions in PRand ERb expression, but not in ERa when compared with the late pro-liferative phase of the normal menstrual cycle. In rhesus macaques,ERb expression was increased with E2 treatment in simulated cyclesand the levels decreased in the epithelial cells in the stratum functionaliswith the subsequent combined treatment with E2 and progesterone(Critchley et al., 2001). These authors also reported static expressionof both ER isoforms in the stratum basalis across the menstrual cycle(Critchley et al., 2001). When compared with other animals, primateendometrium is the closest to that of humans with obvious similarities.However, there are subtle yet striking differences in endocrinology; forexample, E2 levels rise in the mid-secretory phase of the cycle inhumans, but this is not observed in non-human primates (Narkar et al.,2006). Therefore, caution should be exercised when extrapolatingprimate data to human endometrial physiology, including ERb expres-sion and function.
ERb in the endometrial vasculatureBenign angiogenesis is a unique property of endometrium which is essen-tial for normal regeneration of the stratum functionalis after menstrualshedding and is also a fundamental feature of the E2-dominant prolifera-tive endometrium (Nayak and Brenner, 2002). Most described trophiceffects of E2 on this benign endometrial angiogenic process areexerted via ERa either directly or indirectly, acting on endometrial epi-thelial and stromal cells to secrete angiogenic growth factors (Rees andBicknell, 1998). The evidence that E2 has a direct action on endometrialvessels is suggested by reports describing the presence of ER isoforms inthe endometrial vasculature. Nevertheless, there is significant contro-versy regarding cyclical variation in expression of ER subtypes, includingERb, in the endometrial vascular cells. Although all studies report expres-sion of ERb by endometrial vessels (Critchley et al., 2001; Lecce et al.,2001; Greaves et al., 2013), some have suggested that endometrial endo-thelial ERb expression may be dynamically regulated during the men-strual cycle (Lecce et al., 2001), while others reported non-cyclicalconstitutive expression (Critchley et al., 2001). Lecce et al. (2001)reported expression of both ERa and ERb in endometrial endothelialand vascular smooth muscle cells at different phases of the menstrualcycle, although in the menstrual phase, when ovarian hormone levelsare at a nadir, both ER isoforms were reported to be absent from the vas-cular compartment (Lecce et al., 2001). The ERb expression by the vas-cular smooth muscle cells was highest in the late secretory phase,whereas those who reported detection of ERa in the endotheliumnoted the highest endothelial ERa expression to be in E2-dominant mid-cycle endometrium (Lecce et al., 2001). There are also conflictingreports of PR expression by vascular endothelium with some reporting
Estrogen receptor b in the endometrium 179
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

PR mRNA to be present (Krikun et al., 2005), whilst all existing reportsassert that PR protein is not expressed by these cells (Critchley et al.,2001).
In summary, the human data suggest that ERb is the main ER subtype inendometrial endothelial cells and that expression may be hormonallyregulated, although precise receptor expression remains to be fully clari-fied (Critchley et al., 2001; Lecce et al., 2001; Kayisli et al., 2004; Krikunet al., 2005). Subsequently, in an elegant set of rodent experiments,Masuda et al. (2007) demonstrated that bone marrow-derived endo-thelial stem/progenitor cells preferentially express ERa, and that physio-logical post-natal vascular regeneration is E2 regulated via ERa (Masudaet al., 2007). In the endometrium of rhesus macaques (Macaca mulatta),ERb is the only steroid receptor to be expressed by endothelial cells andperivascular smooth muscle cells, but the perivascular stromal cellsexpressed all types of steroid receptors (Slayden and Brenner, 2004).The exact ER isoform regulating the vascular remodelling and neo-vasculogenesis that occurs in regular human endometrial regenerationduring the menstrual cycle and after parturition is therefore not yet con-clusively established, although there is no doubt that ERb exists in thesecells, with evidence that it is likely to play a pivotal role in that process.
ERb in endometrial immune cellsImmune cells are a major component (.20% of the stromal cells in thelate secretory phase) of the endometrium; the cell numbers change in thestratum functionalis according to the ovarian hormone cycle with higherlevels seen in the secretory phase, particularly in late secretory phaseendometrium (Bulmer et al., 1991b; Berbic and Fraser, 2013). The ma-jority of endometrial leucocytes in the stratum functionalis consist ofthree cell types: T cells, macrophages and uterine natural killer (uNK)cells, with very few neutrophils except in menstrual endometrium, andrare B cells (Vassiliadou and Bulmer, 1996), although B lymphocytesare seen in the stratum basalis (Bulmer et al., 1988; Marshall and Jones,1988). The observation that the stromal leucocytes increase innumber during the window of implantation is a compelling reason for im-plicating endometrial leucocytes with a key role in the implantationprocess, and the immunological maintenance of pregnancy (Blois et al.,2011; Evans et al., 2011). Because of their frequency in the late secretoryphase and early pregnancy, as well as the dramatic increase in numbersaround the time of expected implantation in a fertilized cycle, manystudies have focused on the uNK cells. Recent work has focused ontheir potential role in spiral artery remodelling in early pregnancy(Robson et al., 2012), although other roles suggested relate to controlof trophoblast invasion, immunosuppression and cytokine secretion,amongst others (reviewed in Lash et al., 2010). The available evidenceregarding endometrial stromal immune cell numbers and compositionin the endometrial stratum basalis and in post-menopausal endometriumis particularly scarce and, for the reasons previously alluded to, evidenceon any aberrations of these cell numbers or their function in endometrialpathologies is generally limited to the stratum functionalis and at best isconfusing, due to the lack of established and agreed methods of assess-ment. In humans, uNK cells increase in number dramatically in the mid-secretory phase of the menstrual cycle, although the explanation for thisincrease remains uncertain. This has led to investigation of expression ofsteroid hormone receptors by uNK cells and in particular the expressionof PR since uNK cells are also prominent in progesterone-treated endo-metrium. Mouse uNK cells are devoid of both ER receptor subtypes
(Borzychowski et al., 2003), but human uNK cells purified from earlypregnancy decidua contained mRNA for both ERb1 and ERb2, whilstnot expressing ERa or PR transcripts; using immunohistochemistryonly ERb1 protein was expressed in uNK cells of non-pregnant endo-metrium, with no ERb2 protein (Henderson et al., 2003). Expressionlevels for ERb splice variants in mRNA from purified uNK cells from non-pregnant endometrium are not yet known. This highlights the species-specific differences in regulation and function of the various endometrialcell types. Furthermore, ERbexpression and specific functions have beendescribed in T-cells (Rider et al., 2006) and macrophages (Kramer andWray, 2002), yet the ERb expression of endometrial T-cells and macro-phages has not been specifically examined. Studies in KO mice and inother murine models have suggested that in T-cells, E2 modulation ofthe immune response may depend on the origin of the T-cells, andmay be tissue-specific (Maret et al., 2003; Wu et al., 2013). Hence,there is a need for further studies to characterize steroid receptorexpression by human endometrial immune cells.
ERb in the human uterusExpression of ERbmRNA and proteins has been documented in humanendometrium across the menstrual cycle and in post-menopausal endo-metrium. The method employed in most studies of ERb expression inendometrium has been polymerase chain reaction (PCR), and the levelof mRNA detected for ERb at any time in the menstrual cycle hasbeen reported by most researchers to be significantly lower than thatfor ERa (Matsuzaki et al., 2000; O’Neill et al., 2004). However, manystudies either using immunohistochemistry to localize ERb protein atthe cellular level with analysis by a variety of semi-quantitative methodsor using western blotting reported that the levels of ERb protein inhuman endometrium are either comparable with or in excess of thoseof ERa in the endometrium (Villavicencio et al., 2006; Wu et al.,2012). ERb is expressed in all endometrial cell types, including glandularepithelium and stromal cells. ERb is reported by some groups to be thesole ER expressed in many specific cell types within the endometrium,including the endometrial endothelium (Taylor and Al-Azzawi, 2000;Critchley et al., 2001) and uNK cells (Henderson et al., 2003), althoughas stated before (in the ERb in endometrial immune cells section), othershave reported conflicting results (Lecce et al., 2001).
ERb expression in the developing uterusStudies in ERb KO mice suggest that ERb may play an important role inmaintaining endometrial quiescence, particularly in the immature uterus(Weihua et al., 2000). Similarly in the pre-pubertal human uterus, ERaand ERb are expressed in both endometrial epithelium and stromaduring early development before maturity, whereas menarche(maturation)-associated proliferation coincides with an exclusive in-crease in ERa expression (Spencer et al., 2011).
ERb expression in healthy adult human endometrium across themenstrual cycleThe dynamic expression pattern of ovarian steroid receptor proteins andmRNA in the endometrium according to menstrual cycle phase has beenwell established. Ligand-activated ERa induces expression of both ERsubtypes, PR and AR in the endometrium (Brosens et al., 2004). Proges-terone, working through PR, counteracts these effects of E2 onsteroid receptor expression, and also prevents proliferation of endo-metrial epithelial cells, promoting epithelial differentiation and stromal
180 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

decidualization in preparation for embryo implantation (Critchley andSaunders, 2009).
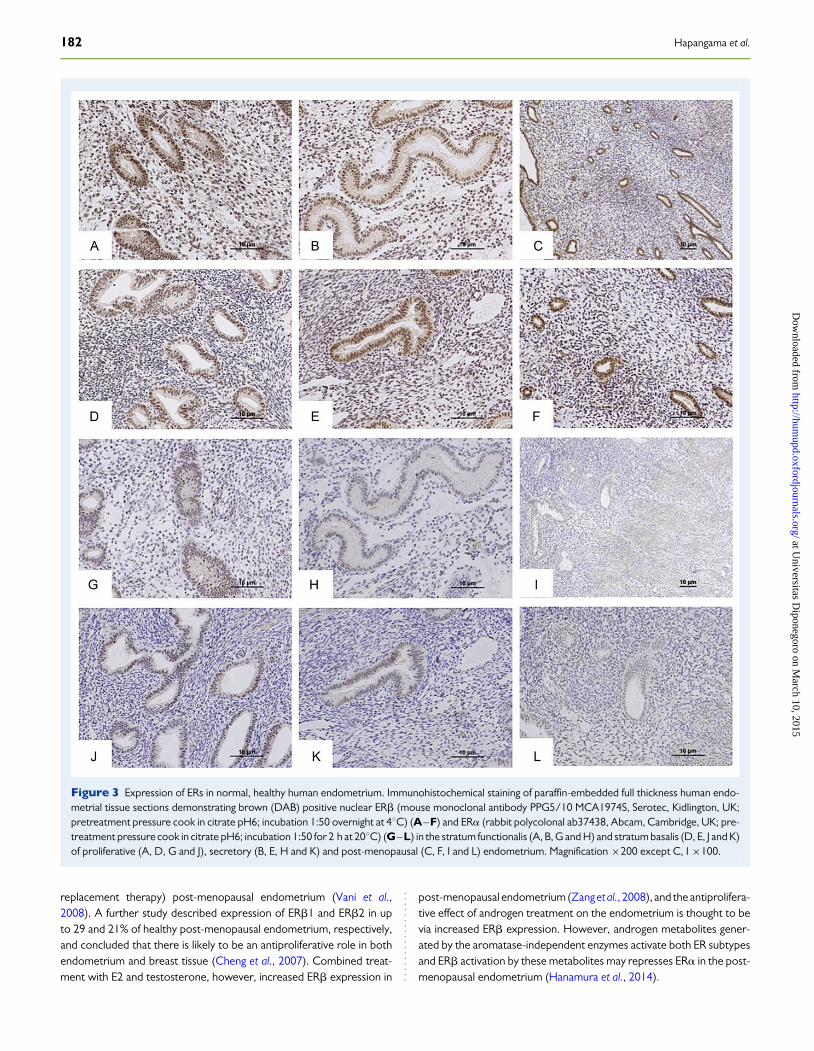
ERbmRNA expression across the menstrual cycle is much lower thanthat of ERa, despite the high levels of ERbprotein expression throughoutthe cycle (Matsuzaki et al., 1999). ERb mRNA is present in epithelial,stromal and endothelial cells, with the highest levels seen in the epithelialcells (Matsuzaki et al., 1999; Critchley et al., 2002). In the estrogen-dominant proliferative phase, nuclear ERa protein levels are high in allendometrial cell types with moderate levels of nuclear ERb protein.Expression of both ER subtypes increases in the late proliferative andearly secretory phases and subsequently decreases in the mid-late secre-tory phase, yet ERb is the predominant ER subtype in the late secretoryphase endometrial stroma (Critchley et al., 2001; Lecce et al., 2001).Whilst stromal ERb expression is increased or maintained in the late se-cretory phase, epithelial expression of ERb protein decreases incommon with ERa (Critchley et al., 2001, 2002; Lecce et al., 2001).Therefore, in the mid-late secretory phase in particular, whereendomet-rial ERa expression diminishes, ERb remains as the predominant ERisoform in the stratum functionalis (Lecce et al., 2001). Interestingly,ERb2 is also expressed in both stromal and epithelial cell compartments,similar to ERb1, but there is a significant decrease in ERb2 in the glandularepithelium of the stratum functionalis in the mid-secretory phase, whilstERb1 persists (Critchley et al., 2002). This may suggest that ERb1 is ableto largely form homo-dimers and become dominant in response to E2 inthe mid-secretory phase endometrial stratum functionalis when circulat-ing E2 levels are still high. Intriguingly, the mid-secretory phase of thecycle in humans is also known to be associated with increased E2levels and a reduction in endometrial glandular proliferative activity(Critchley et al., 2006; Narkar et al., 2006; Cooke et al., 2013). ERb ex-pression in the epithelial compartment of the stratum functionalis corre-lates with the time when highest E2 levels are seen in the menstrual cycle(late proliferative, early secretory phases) (Lecce et al., 2001), whereasstromal and vascular ERb levels peak in the late secretory phase, withthe plateaued second peak of circulating E2 (Lecce et al., 2001)(Fig. 3). Since both E2 via ERa and progesterone via PR have beenshown to increase ERb transcripts, we postulate that ERb is the mainsafety mechanism whereby the potent mitogenic action of E2 is restrictedin the healthy endometrium.
It has been postulated by many authors that the stratum basalis of theendometrium may be less responsive to ovarian hormone regulationthan the stratum functionalis (Prianishnikov, 1978; Padykula et al.,1984). Expression of ERb in the stratum basalis has been reported toremain static across the menstrual cycle (Critchley et al., 2001): this par-ticular study included a modest 32 full-thickness endometrial samples atdifferent time points in the cycle, utilized an immune scoring method thatexamined only the intensity of the staining and also did not examineco-expression of other hormone receptors such as PR and AR. ERb2 ex-pression was very low in the stratum basalis compared with ERb1, sug-gesting that ERb1 is the dominant splice variant in the stratum basalis(Critchley et al., 2002).
The reason why the stratum functionalis of the endometrium but notthe stratum basalis responds to ovarian hormonal signals, despite bothlayers expressing all classical ovarian hormone receptors, is not fullyunderstood. ERa and PR are present, but their levels in the stratumbasalis and the cyclical changes in their expression levels arecontroversialwith reports ranging from alterations in the secretory phase to no changeacross the menstrual cycle (Critchley et al., 2001, 2002; Leyendecker
et al., 2008; Fig. 3). There are no current studies that examine simultan-eously the expression (and therefore interplay) of all ovarian steroidreceptors (ERa, ERb, PR and AR) in these two functionally very distinctendometrial layers of healthy endometrium (Fig. 3). Therefore, data onspatial and temporal differences in the expression of all steroidhormone receptor types in human endometrium remain inconclusive.
Role of ERb in endometrial regenerationThe hypothesis that endometrium regenerates from the hormonally re-sistant stratum basalis came initially from Prianishnikov (1978). Otherstudies suggested that although the initial part of post-menstrual endo-metrial regeneration is independent of estrogen, subsequent post-menstrual repair growth is estrogen-dependent (Ferenczy, 1976). Thestratum basalis is widely accepted as the germinal compartment of theendometrium where stem progenitor cells reside (Padykula et al.,1989; Valentijn et al., 2013) and ERb1 and ERb2 are expressed by allcell types in the stratum basalis (Critchleyet al., 2002). Many investigatorsof endometrial stem progenitor cells have reported a reduction or lack ofERa and PR in the more primitive cells with progenitor activity (Chan andGargett, 2006; Valentijn et al., 2013). In mouse endometrium, the endo-metrial epithelial progenitor cell pool expanded dramatically despite thelack of expression of both ERa and PR (Janzen et al., 2013). However,none of the descriptive papers of endometrial stromal/epithelialstem/progenitor populations in either animal or human studies exam-ined or commented on ERb expression (Chan and Gargett, 2006;Cervello et al., 2010; Masuda et al., 2010; Janzen et al., 2013), and post-partum vascular regeneration (thought to involve stem cells), althoughE2-dependent, has been reported to be mediated via ERa rather thanERb (Masuda et al., 2007). Furthermore, studies on mouse endometrialregeneration provide evidence that the primitive label-retaining cells areestrogen responsive (Gargett et al., 2012). Therefore, further work isneeded to examine expression of ERb and other hormone receptorsin the primitive epithelial and stromal stem/progenitor population,since the observed (possible direct) stimulatory effect of E2 on the pro-genitor population may be mediated via ERb.
ERb expression in the healthy humanpost-menopausal endometriumNormal post-menopausal endometrium is the remaining stratum basalisafter the cessation of the ovarian hormonal cycle. Similarities betweenthe premenopausal stratum basalis and post-menopausal endometriumhave been described, both in the gene expression profile (Nguyen et al.,2012) and in expression of epithelial markers (Valentijn et al., 2013).However, the hormonal milieu of pre- and post-menopausal endomet-rium is clearly different (Labrie, 2014). Post-menopausal endometrium isexposed to relatively low E2 levels, absent progesterone and relativelyunchanged androgen levels of adrenal origin (Yasui et al., 2012). ERbexpression in post-menopausal endometrium was reported to beweaker than ERa expression in both stromal and epithelial compart-ments (Zang et al., 2008). Another small immunohistochemical studywhich included only 11 post-menopausal patients reported down-regulation of both ERs in post-menopausal endometrium comparedwith the late proliferative phase (Mylonas et al., 2007). Others havereported moderate to strong ERb immunostaining in all cell types innormal post-menopausal endometrium which did not change inhormone-treated (continuous combined E2 and progestagen hormone
Estrogen receptor b in the endometrium 181
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from
replacement therapy) post-menopausal endometrium (Vani et al.,2008). A further study described expression of ERb1 and ERb2 in upto 29 and 21% of healthy post-menopausal endometrium, respectively,and concluded that there is likely to be an antiproliferative role in bothendometrium and breast tissue (Cheng et al., 2007). Combined treat-ment with E2 and testosterone, however, increased ERb expression in
post-menopausal endometrium (Zang et al., 2008), and the antiprolifera-tive effect of androgen treatment on the endometrium is thought to bevia increased ERb expression. However, androgen metabolites gener-ated by the aromatase-independent enzymes activate both ER subtypesand ERb activation by these metabolites may represses ERa in the post-menopausal endometrium (Hanamura et al., 2014).
Figure 3 Expression of ERs in normal, healthy human endometrium. Immunohistochemical staining of paraffin-embedded full thickness human endo-metrial tissue sections demonstrating brown (DAB) positive nuclear ERb (mouse monoclonal antibody PPG5/10 MCA1974S, Serotec, Kidlington, UK;pretreatment pressure cook in citrate pH6; incubation 1:50 overnight at 48C) (A–F) and ERa (rabbit polycolonal ab37438, Abcam, Cambridge, UK; pre-treatment pressure cook in citrate pH6; incubation 1:50 for 2 h at 208C) (G–L) in the stratum functionalis (A, B, G and H) and stratum basalis (D, E, J and K)of proliferative (A, D, G and J), secretory (B, E, H and K) and post-menopausal (C, F, I and L) endometrium. Magnification ×200 except C, I ×100.
182 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

ERb in endometrial pathology
Endometriosis and adenomyosisEndometriosis is a common benign gynaecological condition defined bythe presence of endometrium-like tissue outside of the endometrialcavity (Bernardi and Pavone, 2013; Sourial et al., 2014). Adenomyosisis defined as the presence of endometrium within the myometrium(Hapangama and Bulmer, 2014). Aberrant cell proliferation andaltered cell fate have been described in eutopic and ectopically grownendometrium and in adenomyosis; estrogen dependence may play arole in the pathophysiology of both conditions (Yang et al., 2007;Hapangama et al., 2009). Although this may immediately suggest atrophic estrogenic effect via ERa, ectopic endometrial lesions and ade-nomyosis have been reported to express high levels of ERb (.100×)compared with the eutopic endometrium (Bulun et al., 2012). The avail-able evidence regarding levels of ERa and ERb expression in ectopicendometriotic lesions is, however, contradictory (Shao et al., 2014).Factors that are likely to contribute to the variable results include the het-erogeneity of the lesions studied, as well as the fact that when surgicallyexcised ectopic endometriotic lesions are examined using techniquessuch as PCR and western blotting which homogenize the whole tissuethe contribution from endometrial-gland and stroma-like tissue to thelevels reported can vary widely.
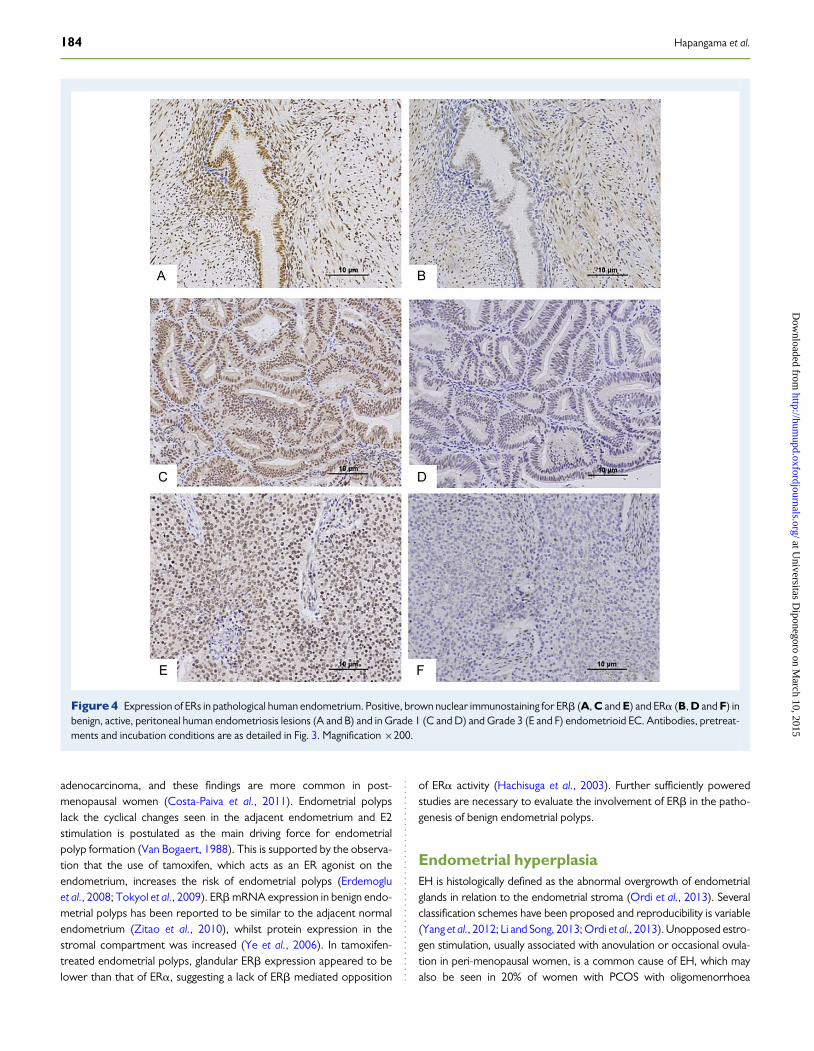
Relative over-expression of ERb mRNA (therefore decreasedERa:ERb mRNA ratio) in ovarian endometriomas compared witheither peritoneal ectopic lesions (Bukulmez et al., 2007) or eutopic endo-metrium (Brandenberger et al., 1999; Fujimoto et al., 1999; Bukulmezet al., 2007) has been described suggesting a unique E2-dependentgrowth of ovarian endometriomas. Deficient methylation of the ERbpromoter resulting in pathological overexpression of ERb in endome-triotic stromal cells has been suggested by some authors who alsoreport relatively low ERa expression in these cells (Bulun et al., 2012).A high level of ERb has been proposed as the reason for low ERa expres-sion, resulting in low PR levels contributing to progesterone resistanceand inflammation. ERb knockdown has been reported to significantly in-crease ERa mRNA and protein levels in endometriotic stromal cells(Trukhacheva et al., 2009; Bulun et al., 2012). This theory is reviewed ex-tensively (Bulun et al., 2012). Conversely, ERb overexpression in endo-metrial stromal cells was reported to decrease ERa mRNA and proteinlevels and ERb knockdown significantly decreased proliferation of endo-metriotic stromal cells (Trukhacheva et al., 2009). In human endometrio-tic lesions (Fujimoto et al., 1999; Bukulmez et al., 2007) and in inducedlesions in endometriosis models in baboons (Fazleabas et al., 2003),and in rodents (Han et al., 2012), there was decreased ERa, while ERbwas maintained. Compared with healthy women, women with endo-metriosis and adenomyosis showed a decrease in the ERa/ERb ratioin proliferative phase eutopic endometrium, suggesting ERb dominance(Juhasz-Bass et al., 2011; Mehasseb et al., 2011), although there is no con-clusive evidence suggesting differential proliferative activity in the prolif-erative phase endometrium of women with endometriosis. Mostcellular aberrations described in the eutopic endometrium have beenobserved in the stratum functionalis in the secretory phase where a per-sistence in proliferative activity is detected (Hapangama et al., 2009,2012) and ERbexpression in women with endometriosis in the secretoryphase has been reported to be unchanged (Hudelist et al., 2005) ordecreased (Hapangama et al., 2008b). In the window of implantation,
endometrial stratum functionalis of women with endometriosis mayalso have a reduction in ERb expression compared with healthy controls(Hapangama et al., 2008b). Further studies have shown higher ERamRNA levels in endometriotic lesions compared with the eutopic endo-metrium (Matsuzaki et al., 2000, 2001; Fig. 4). Furthermore, ERb pro-moter methylation has been proposed as the primary defect resultingin differential ERb expression between ectopic and eutopic endomet-rium (Xue et al., 2007).
It is of interest that the pattern of ERb expression reported in bothadenomyosis and ectopic endometriotic lesions was similar to that ofthe endometrial stratum basalis (Mehasseb et al., 2011), which in turnis proposed to be unresponsive to hormones. There are suggestionsfrom primate (Donnez et al., 2012; Sourial et al., 2014) and humanstudies (Leyendecker et al., 2002; Valentijn et al., 2013) that endometrialstratum basalis plays an essential role in the pathogenesis of endometri-osis. This may suggest that the regulation of cell fate in these pathologicallesions is distinct from that of the differentiated stratum functionalis in theeutopic endometrium but rather is similar to the stratum basalis that con-tains the germinal capacity. Further exploration of their similarities to thegerminal stratum basalis may improve our understanding of endometrialcellular growth and regeneration.
Polycystic ovary syndromeThe clinical phenotype of polycystic ovary syndrome (PCOS) includes re-productive and hormonal aberrations. The endometrial consequencesare those of a high and unopposed effect of estrogen, and possibly andro-gens (Hapangama and Bulmer, 2014). The aetiology of PCOS is not fullyunderstood and using genotyping a +1730 G/A polymorphism in theERb gene has been proposed to be associated with susceptibility toPCOS (Kim et al., 2010). Homozygous ERbKO mice have defective ovu-lation reminiscent of PCOS in humans (Imamov et al., 2005). It has beenpostulated that women with PCOS exhibit a lower pregnancy rate sec-ondary to decreased endometrial expression of both ERa and ERbduring the window of implantation, potentially decreasing endometrialreceptivity, reducing conception and hence lowering fertility (Wanget al., 2011). However, investigation of the differential ER subtype ex-pression in women with PCOS compared with healthy women has notrevealed a definitive pattern (Maliqueo et al., 2003). AnovulatoryPCOS is associated with a 3- to 4-fold increased risk of developing ECcompared with unaffected women and the effect is postulated to bedue to the progesterone unopposed action of E2 (Fearnley et al.,2010; Hapangama and Bulmer 2014), presumed to be via ERa.A gradual increase in ERb levels from anovulatory endometrium to endo-metrial hyperplasia (EH) was seen in women with PCOS, suggestingeither direct involvement of ERb in endometrial proliferation or an indir-ect effect due to the action of ERawhich can cause increased expressionof all steroid receptors including ERb (Villavicencio et al., 2006). Furtherstudies are needed to evaluate the involvement of ER subtypes and theendometrial proliferative aberration that occurs in PCOS.
Endometrial polypsEndometrial polyps are commonly occurring outgrowths of the endo-metrium consisting of a monoclonal overgrowth of endometrialstromal cells with inclusion of a non-neoplastic glandular component(Indraccolo et al., 2013). Endometrial polyps are usually benign butare occasionally associated with focal atypical hyperplasia or even
Estrogen receptor b in the endometrium 183
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from
adenocarcinoma, and these findings are more common in post-menopausal women (Costa-Paiva et al., 2011). Endometrial polypslack the cyclical changes seen in the adjacent endometrium and E2stimulation is postulated as the main driving force for endometrialpolyp formation (Van Bogaert, 1988). This is supported by the observa-tion that the use of tamoxifen, which acts as an ER agonist on theendometrium, increases the risk of endometrial polyps (Erdemogluet al., 2008; Tokyol et al., 2009). ERbmRNA expression in benign endo-metrial polyps has been reported to be similar to the adjacent normalendometrium (Zitao et al., 2010), whilst protein expression in thestromal compartment was increased (Ye et al., 2006). In tamoxifen-treated endometrial polyps, glandular ERb expression appeared to belower than that of ERa, suggesting a lack of ERb mediated opposition
of ERa activity (Hachisuga et al., 2003). Further sufficiently poweredstudies are necessary to evaluate the involvement of ERb in the patho-genesis of benign endometrial polyps.
Endometrial hyperplasiaEH is histologically defined as the abnormal overgrowth of endometrialglands in relation to the endometrial stroma (Ordi et al., 2013). Severalclassification schemes have been proposed and reproducibility is variable(Yang et al., 2012; Li and Song, 2013; Ordi et al., 2013). Unopposed estro-gen stimulation, usually associated with anovulation or occasional ovula-tion in peri-menopausal women, is a common cause of EH, which mayalso be seen in 20% of women with PCOS with oligomenorrhoea
Figure 4 Expression of ERs in pathological human endometrium. Positive, brown nuclear immunostaining for ERb (A, C and E) and ERa (B, D and F) inbenign, active, peritoneal human endometriosis lesions (A and B) and in Grade 1 (C and D) and Grade 3 (E and F) endometrioid EC. Antibodies, pretreat-ments and incubation conditions are as detailed in Fig. 3. Magnification ×200.
184 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

(Prakansamut et al., 2014). When cytological atypia is present, EH is asso-ciated with approximately a 30% risk of developing into or co-existing withEC (Lacey et al., 2007; Park et al., 2011). Transcripts of ERb splice variantshave been described in EH (Witek et al., 2001), and ERb protein expres-sion in EH without cytological atypia in women without PCOS has beenreported to remain unchanged compared with normal proliferative endo-metrium, whilst in pre-cancerous atypical hyperplasia, ERb expressiondecreases (Hu et al., 2005). This suggests that the loss of ERb regulationof ERa-mediated proliferation results in EH.
Endometrial cancerEC is the most common gynaecological malignancy and is usually adisease of the post-menopausal endometrium (Murali et al., 2014). Incontrast with atrophic and inactive post-menopausal endometrium(McCluggage, 2011), the hallmark of EC is dysregulated and inexhaustiblecellular proliferation (Owings and Quick, 2014). At least Type-I endome-trioid EC is thought to be E2-dependent (although recent results indicatethat Types I and II ECs have similar risk factors), and unopposed E2 actionis known to increase the risk of endometrial carcinogenesis (Setiawanet al., 2012). Therefore, conditions such as PCOS and obesity, wherethere is an excessive extra-ovarian conversion of adrenal androgens toestrogens, as well as HRT use, increase the risk of EC (Thanapprapasrand Thanapprapasr, 2013).
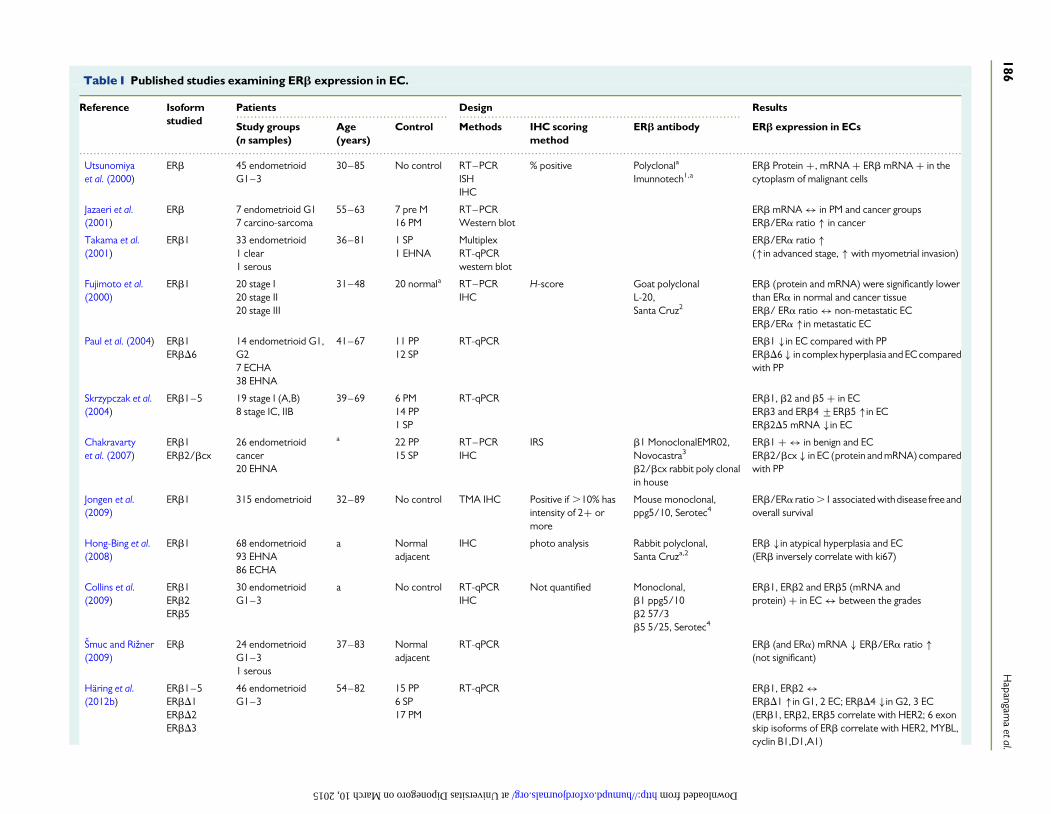
Available data on ERb protein and mRNA expression in EC subtypesare largely confined to endometrioid EC (Table I). The general consensusthat the carcinogenesis process in Type II ECs is E2-independent is likelyto be the reason for the lack of data on ERb expression in Type II ECs inparticular, with virtually no studies examining the ERb expression inserous and clear cell carcinoma. Nevertheless, there are emerging datasuggesting involvement of ERb in carcinogenesis of other tissues, suchas the intestine, that are typically not regarded as E2-responsive(Hogan et al., 2009), and hence future studies examining ERb in type IIECs are warranted.
In addition to the wild-type ERb1, the expression of splice variantisoforms ERb2, b3, b4, b5 and exon deletes (bD1, b2D5, bD4, bD6)has been described in endometrioid EC (Saegusa and Okayasu, 2000;Utsunomiya et al., 2000; Sakaguchi et al., 2002; Paul et al., 2004;Skrzypczak et al., 2004; Mylonas et al., 2005; Chakravarty et al., 2007;Collins et al., 2009; Zannoni et al., 2013). Similar expression patternsto ERa have been reported for ERb variants (ERb1, ERb2 and ERb5)in endometrioid EC samples: ERb1 and ERb2 immuno-expressionwas higher in low-grade ECs, whereas ERb5 expression was consti-tutively intense regardless of the grade (Collins et al., 2009). Intamoxifen-associated ECs, ERb expression was particularly prominentcompared with spontaneous ECs (Negoita and Mihailovici, 2011). In adifferent study that included a semi-quantitative scoring system similarto the quickscore hormone receptor evaluation in breast cancer,ERb1and ERb2 expression did not correlate with tumour grade, FIGO(International Federation of Gynecology and Obstetrics) stage or myo-metrial invasion, but the ERa/ERb2 ratio was reported to be an inde-pendent prognostic factor of overall survival (Zannoni et al., 2013).The ERa/ERb ratio has been used to evaluate the imbalance in the rela-tive expression of ER isoforms in endometrial (Fujimoto et al., 2000;Jazaeri et al., 2001; Takama et al., 2001; Jongen et al., 2009; Smuc andRizner, 2009) and other hormonal-regulated malignancies, such asbreast cancer, and has been suggested as a potential predictor of
disease outcome (Sastre-Serra et al., 2013). Nevertheless, the existingdata on the correlation between clinical outcome and the ERa/ERbratio are conflicting (Jongen et al., 2009; Zannoni et al., 2013). Methodo-logical differences (sample size and scoring methods) are likely to explainthe observed discrepancies in the available literature. There are over 160publications describing ERb involvement in EC, yet the data on ERb ex-pression, correlation with disease stage and grade of histological differen-tiation remain controversial (Table I). Another reason for this is that notall studies included healthy control tissue comparators. Some studieshave only described ERb expression in neoplastic cells and the surround-ing niche (Smuc and Rizner, 2009). When healthy controls were used,they included either pre- or post-menopausal women, although insome studies, the menopausal status of the healthy control womenwas not even mentioned (Jarzabek et al., 2013; Knapp et al., 2013).When premenopausal control tissue was used, attention was not givento the known differences between menstrual cycle phase (Jazaeri et al.,2001), or in full thickness endometrial samples from hysterectomies,the differences between stratum basalis and functionalis, with no infor-mation provided regarding which layer was included in the analysis(Chakravarty et al., 2007; Knapp et al., 2013). Early studies also sufferedfrom deficiencies in antibody and primer specificities, particularly whensplice variants were not appreciated.
Breast and prostatic carcinogenesis has been reported to be asso-ciated with a decrease or loss of ERb expression, and this is the basisfor the hypothesis that ERb plays a tumour suppressor role (Bottneret al., 2014). In breast and ovarian cancer, ERb exerted anti-proliferativeeffects via prevention of ERa transcriptional complexes from activatingc-myc, cyclin-1 and cyclin-A genes and induction of cyclin-dependentkinase inhibitors leading to arrest of the cell cycle in G2 phase (Paruthiyilet al., 2004). Similarly, in the endometrium (as described above), someauthors have reported a decrease in ERb expression in EC suggestingan analogous function. A decrease in both ERb mRNA levels andprotein (immunohistochemistry) has been reported in endometrioidEC compared with either adjacent normal endometrium or normal pro-liferative endometrium from healthy premenopausal control women(Paul et al., 2004; Hong-bing et al., 2008; Smuc and Rizner, 2009).However, a potential oncogenic role has also been proposed by otherreports showing an up-regulation of ERb5 transcript in high-grade endo-metrioid EC, which wasalso associated with HER2 and MYBL2 oncogeneexpression (Skrzypczak et al., 2004; Haring et al., 2012b).
Since ovarian steroid hormone receptor expression is a feature of dif-ferentiated endometrial cells, with primitive or undifferentiated cells notexpressing ER or PR, the diminishing steroid receptor expression levels inEC, including ERb, may merely be a sign of loss of cellular differentiationand/or cellular transformation (Fig. 4). Alternatively, dysregulation ofsteroid receptor homeostasis owing to loss of PR and ERawith possiblepersistant expression and up-regulation of ERb isoforms (ERb5) andsplice variants, particularly in high-grade and advanced stage endome-trioid EC, may reflect a tumour-promoting role for ERb in high-gradeECs. Data from ERabKO mice may suggest ERb could partially compen-sate for the loss of ERa (Dupont et al., 2000) and therefore the reportedincrease in ERb in high-grade ECs may have a tumour-promoting effectsimilar to ERa. However, conclusive data on the functional role of ERbor the splice variants on regulating EC growth or metastasis doesnot exist to date. The reason for the apparent contradictory datafor ERb to be a tumour suppressor and a promotor may be due to atissue-specific dual role of ERb; a tumour suppressor role in healthy
Estrogen receptor b in the endometrium 185
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from
........................................................... ...............................................................................
..........................................................................................................................................................................................................................................................
Table I Published studies examining ERb expression in EC.
Reference Isoformstudied
Patients Design Results
Study groups(n samples)
Age(years)
Control Methods IHC scoringmethod
ERb antibody ERb expression in ECs
Utsunomiyaet al. (2000)
ERb 45 endometrioidG1–3
30–85 No control RT–PCRISHIHC
% positive Polyclonala
Imunnotech1,aERb Protein +, mRNA + ERb mRNA + in thecytoplasm of malignant cells
Jazaeri et al.(2001)
ERb 7 endometrioid G17 carcino-sarcoma
55–63 7 pre M16 PM
RT–PCRWestern blot
ERb mRNA ↔ in PM and cancer groupsERb/ERa ratio � in cancer
Takama et al.(2001)
ERb1 33 endometrioid1 clear1 serous
36–81 1 SP1 EHNA
MultiplexRT-qPCRwestern blot
ERb/ERa ratio �(�in advanced stage, � with myometrial invasion)
Fujimoto et al.(2000)
ERb1 20 stage I20 stage II20 stage III
31–48 20 normala RT–PCRIHC
H-score Goat polyclonalL-20,Santa Cruz2
ERb (protein and mRNA) were significantly lowerthan ERa in normal and cancer tissueERb/ ERa ratio ↔ non-metastatic ECERb/ERa �in metastatic EC
Paul et al. (2004) ERb1ERbD6
14 endometrioid G1,G27 ECHA38 EHNA
41–67 11 PP12 SP
RT-qPCR ERb1 �in EC compared with PPERbD6 � in complex hyperplasia and EC comparedwith PP
Skrzypczak et al.(2004)
ERb1–5 19 stage I (A,B)8 stage IC, IIB
39–69 6 PM14 PP1 SP
RT-qPCR ERb1, b2 and b5 + in ECERb3 and ERb4 +ERb5 �in ECERb2D5 mRNA �in EC
Chakravartyet al. (2007)
ERb1ERb2/bcx
26 endometrioidcancer20 EHNA
a 22 PP15 SP
RT–PCRIHC
IRS b1 MonoclonalEMR02,Novocastra3
b2/bcx rabbit poly clonalin house
ERb1 + ↔ in benign and ECERb2/bcx � in EC (protein and mRNA) comparedwith PP
Jongen et al.(2009)
ERb1 315 endometrioid 32–89 No control TMA IHC Positive if .10% hasintensity of 2+ ormore
Mouse monoclonal,ppg5/10, Serotec4
ERb/ERa ratio .1 associated with disease free andoverall survival
Hong-Bing et al.(2008)
ERb1 68 endometrioid93 EHNA86 ECHA
a Normaladjacent
IHC photo analysis Rabbit polyclonal,Santa Cruza,2
ERb �in atypical hyperplasia and EC(ERb inversely correlate with ki67)
Collins et al.(2009)
ERb1ERb2ERb5
30 endometrioidG1–3
a No control RT-qPCRIHC
Not quantified Monoclonal,b1 ppg5/10b2 57/3b5 5/25, Serotec4
ERb1, ERb2 and ERb5 (mRNA andprotein) + in EC ↔ between the grades
Smuc and Rizner(2009)
ERb 24 endometrioidG1–31 serous
37–83 Normaladjacent
RT-qPCR ERb (and ERa) mRNA � ERb/ERa ratio �(not significant)
Haring et al.(2012b)
ERb1–5ERbD1ERbD2ERbD3
46 endometrioidG1–3
54–82 15 PP6 SP17 PM
RT-qPCR ERb1, ERb2 ↔ERbD1 �in G1, 2 EC; ERbD4 �in G2, 3 EC(ERb1, ERb2, ERb5 correlate with HER2; 6 exonskip isoforms of ERb correlate with HER2, MYBL,cyclin B1,D1,A1)
186H
apangama
etal.
at Universitas Diponegoro on March 10, 2015 http://humupd.oxfordjournals.org/ Downloaded from

endometrium which expresses the full complement of other ovarianhormone receptor types and particularly ERa, and a potential tumourpromoter role in high-grade EC cells that have lost other receptor sub-types. This is an interesting possibility that needs to be explored infuture studies.
In summary, the exact role of ERb in endometrial carcinogenesis andprogression of EC is not yet fully known.Therefore, comprehensivechar-acterization of ERb expression in EC subtypes and further functionalstudies are required to reveal the involvement and contribution of ERbin endometrial carcinogenesis. Since at least some ECs are likely torespond to hormonal chemotherapeutic agents, a detailed examinationof all steroid receptor types and standardization of the methodology forquantification of hormone receptors in EC in a similar approach to thatused for other hormone responsive cancers, such as in breast cancer,is urgently needed.
Pharmacological regulation ofendometrial ERb expression
Evidence for regulation of endometrial ERbexpression by exogenous hormone treatmentThere is evidence from both in vivo and in vitro studies that ERb plays animportant role in determining cellular responses to estrogens and anti-estrogens (Hall and McDonnell, 1999). Progesterone induced ERb tran-scription whilst down-regulating the PR gene in telomerase immortalizedendometrial epithelial cells (Hombach-Klonisch et al., 2005). In contrast,treatment of women with an intrauterine system containing the andro-genic progestagen, Levenorgestrel, decreased epithelial and stromalERb (and also ERa and PR) expression after 3 months (Engemise et al.,2011). In non-human primates, the non-specific progestogen, MPAincreased endometrial stromal ERb expression in ovariectomizedanimals (Wang et al., 2002). The available evidence therefore suggeststhat the endometrial ERb expression in response to exogenous hor-mones depends on the duration and dose as well as the presence ofother steroid hormones and their receptors.
Evidence for SERMs (ERb selectivepharmacological agents) in endometriumStudies in rodents have shown that Puerarin, a selective estrogen recep-tor modulator (SERM) with ERb preferential agonist activity, may affectendometrial expression of implantation-specific proteins and may havea contraceptive effect (Saha et al., 2012). In agreement with these data, aPhase 2 trial investigating the use of a selective ERb agonist for climac-teric symptoms in 164 post-menopausal women reported an increasedincidence of endometrial thickening (18 versus 6% in the placebo group)but with benign histology (Grady et al., 2009). In contrast, others havereported non-uterotrophic activity by ERb selective agonists in ovariec-tomized rats (Hertrampf et al., 2008). Therefore, the expected uterine/endometrial neutral effect of novel ERb selective SERMs has not beenconclusively confirmed in long-term studies and should be exploredprior to administering these agents for their other potential healthbenefits. However, studies of these agents in other tissues have high-lighted the complexities of interplay between ERb and other hormonalnuclear receptor functions. Differential actions of these agents in theendometrial epithelial and stromal compartments also have to be fully
Kna
ppet
al.
(201
3)ER
b35
endo
met
rioid
G1
–3
44–
7629
norm
ala
Wes
tern
blot
ERb
and
ERa�
inEC
ERb
and
ERa��
inG
1
Zan
noni
etal
.(2
013)
ERb
1ER
b2
121
endo
met
rioid
G1
–3
35–
88N
oco
ntro
lIH
CA
llred
Mon
oclo
nal
b1
ppg5
/10,
DA
KO5
b2
57/3
,Se
rote
c4
ERb
/ERa�i
nlo
w-g
rade
ECER
b1
and
ERb
2/E
Ra
ratio
.1
inde
pend
ently
asso
ciat
ew
ithhi
gher
risk
ofde
ath
(ERa
alon
edo
esno
tpre
dict
the
outc
ome)
G1
–3,
Gra
de1
–3;
PP,p
rolif
erat
ive
phas
e;SP
,sec
reto
ryph
ase;
PM,p
ost-
men
opau
sal;
pre
M,p
rem
enop
ausa
l;EH
NA
,end
omet
rialh
yper
plas
iaw
ithno
atyp
ia;E
CH
A,e
ndom
etria
lcom
plex
hype
rpla
sia
with
atyp
ia;I
HC
,im
mun
ohis
toch
emis
try;
TM
A,t
issu
em
icro
arra
y;IS
H,i
nsit
uhy
brid
izat
ion;
IRS,
imm
unor
eact
ive
scor
e;+
,exp
ress
ed;+
,min
imal
expr
essi
onle
vels
;�,i
ncre
ased
;�,d
ecre
ased
;↔,n
otch
ange
d;1 M
arse
ille,
Fran
ce;2
Biot
echn
olog
y,In
c.,S
anta
Cru
z,C
A,U
SA;3 N
ewca
stle
upon
Tyn
e,U
K;4 Se
rote
cLt
d,O
xfor
d,U
K;5 G
lost
rup,
Den
mar
k.a Im
port
anti
nfor
mat
ion
isno
trep
orte
d.
Estrogen receptor b in the endometrium 187
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

elucidated in health and in endometrial pathologies prior to consideringthe therapeutic potential of the many available selective pharmacologic-al modulators of ERb for patient benefit.
Summary/conclusionDespite early predictions to the contrary, the collective evidence sug-gests that ERb has an important role in normal endometrial tissuehomeostasis, cell turnover and regeneration and also in most if not allbenign and malignant endometrial diseases. Information gained fromstudies of other tissues on the basic function of ERb isoforms in restrain-ing the ERa-mediated cell-specific trophic/mitotic responses to E2 hasallowed appreciation of the important role of ERb in regulation of cellfate in human endometrium. Our current understanding of ERb expres-sion and function in endometrium is, however, far from complete. ERb isdynamically expressed in healthy premenstrual endometrium and per-sists in post-menopausal atrophic endometrium. The possible dysregu-lation of ERb expression that has been reported in benign endometrialproliferative diseases, such as endometriosis and PCOS, as well as in EChighlights the importance of exploring the potential use of this ERisoform for targeted therapy with estrogenSERMs. However, thereare several voids in our current knowledge; for example, the existenceof ERb splice variants and their influence on ERb1 function and E2 re-sponsiveness in health and in endometrial disease. Research clarifyingthese areas and functional studies may explain conflicting data thatexist in the current literature. Future studies should specificallyexamine the inter-regulatory effect of the ERb splice variants,whereby each of the splice variants regulates the effect of the other var-iants, as well as the co-regulatory effect of ERb with other steroidhormone receptors and their combined effects on endometrial cellularresponses to steroid hormone signals, as these are all pivotal for predict-ing the effects of pharmacological hormone receptor modulators inendometrial function. Our current understanding of the role of ERb inthe endometrium from many animal studies remains to be confirmedby functional studies (both in vitro and in vivo) on human endometrium,or human endometrial epithelial cells in particular. However, there isalso an urgent need to standardize the way that immunohistochemicalstudies of steroid receptor expression in benign and malignant endo-metrium are conducted and assessed, especially in EC; agreement isneeded regarding the most appropriate control tissue (post- or preme-nopausal endometrium), especially in the light of increased levels ofobesity and the associated estrogenic effects on the endometrium, aswell as the development of a robust method for quantification/assess-ment of immunohistochemical data on ERb expression, similar to thescoring systems that are employed in other hormonally regulatedtissues, such as breast cancer, since these data may have obviousdirect clinical implications in guiding therapy.
Supplementary dataSupplementary data are available at http://humupd.oxfordjournals.org/online.
AcknowledgementsThe authors are grateful for Dr Nicola Tempest and Mrs Jo Drury fortheir assistance in manuscript preparation.
Authors’ rolesD.K.H. conceived the manuscript, D.K.H. and A.M.K. prepared the firstdraft and J.N.B. revised the manuscript critically for important intellectualcontent. A.M.K. drafted the figures and tables. All authors revised andread the manuscript and approved the submitted final version.
FundingWe acknowledge the support by Wellbeing of Women project grantRG1073 (D.K.H.), RG1342 (J.N.B.) and RG1487 (D.K.H.), Iraqi Govern-ment (A.M.K.) and BBSRC BB/E016790/1 (J.N.B.).
Conflict of interestNone declared.
ReferencesArgenta P, Svendsen C, Elishaev E, Gloyeske N, Geller MA, Edwards RP, Linkov F.
Hormone receptor expression patterns in the endometrium of asymptomaticmorbidly obese women before and after bariatric surgery. Gynecol Oncol 2014;133:78–82.
Arnal JF, Gourdy P, Lenfant F. In vivo dissection of the estrogen receptor alpha:uncoupling of its physiological effects and medical perspectives. Ann Endocrinol2013;74:82–89.
Benecke A, Chambon P, Gronemeyer H. Synergy between estrogen receptor alphaactivation functions AF1 and AF2 mediated by transcription intermediary factorTIF2. EMBO Rep 2001;1:151–157.
Berbic M, Fraser IS. Immunology of normal and abnormal menstruation. WomensHealth 2013;9:387–395.
Bernardi LA, Pavone ME. Endometriosis: an update on management. Womens Health2013;9:233–250.
Blair IA. Analysis of estrogens in serum and plasma from postmenopausal women: pastpresent, and future. Steroids 2010;75:297–306.
Blois SM, Klapp BF, Barrientos G. Decidualization and angiogenesis in early pregnancy:unravelling the functions of DC and NK cells. J Reprod Immunol 2011;88:86–92.
Borzychowski AM, Chantakru S, Minhas K, Paffaro VA, Yamada AT, He H, Korach KS,Croy BA. Functional analysis of murine uterine natural killer cells genetically devoid ofestrogen receptors. Placenta 2003;24:403–411.
Bottner M, Thelen P, Jarry H. Estrogen receptor beta: tissue distribution and the stilllargely enigmatic physiological function. J Steroid Biochem Mol Biol 2014;139:245–251.
Brandenberger AW, Lebovic DI, Tee MK, Ryan IP, Tseng JF, Jaffe RB, Taylor RN.Estrogen receptor (ER)-alpha and ER-beta isoforms in normal endometrial andendometriosis-derived stromal cells. Mol Hum Reprod 1999;5:651–655.
Brosens JJ, Tullet J, Varshochi R, Lam EWF. Steroid receptor action. Best Pract Res ClinObstet Gynaecol 2004;18:265–283.
Bukulmez O, Hardy DB, Carr BR, Word RA, Mendelson CR. Inflammatory statusinfluences aromatase and steroid receptor expression in endometriosis.Endocrinology 2007;149:1190–1204.
Bulmer JN, Lunny DP, Hagin SV. Immunohistochemical characterization of stromalleucocytes in nonpregnant human endometrium. Am J Reprod Immunol Microbiol1988;17:83–90.
Bulmer JN, Longfellow M, Ritson A. Leukocytes and resident blood cells inendometrium. Ann N Y Acad Sci 1991a;622:57–68.
Bulmer JN, Morrison L, Longfellow M, Ritson A, Pace D. Granulated lymphocytes inhuman endometrium: histochemical and immunohistochemical studies. HumReprod 1991b;6:791–798.
Bulun SE, Monsavais D, Pavone ME, Dyson M, Xue Q, Attar E, Tokunaga H, Su EJ. Roleof estrogen receptor-beta in endometriosis. Semin Reprod Med 2012;30:39–45.
Burns KA, Korach KS. Estrogen receptors and human disease: an update. Arch Toxicol2012;86:1491–1504.
188 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

Cervello I, Gil-Sanchis C, Mas A, Delgado-Rosas F, Martinez-Conejero JA, Galan A,Martinez-Romero A, Martinez S, Navarro I, Ferro J et al. Human endometrial sidepopulation cells exhibit genotypic, phenotypic and functional features of somaticstem cells. PLoS One 2010;5:e10964.
Chakravarty D, Srinivasan R, Ghosh S, Gopalan S, Rajwanshi A, Majumdar S. Estrogenreceptor b1 and the b2/bcx isoforms in nonneoplastic endometrium and inendometrioid carcinoma. Int J Gynecol Cancer 2007;17:905–913.
Chan RW, Gargett CE. Identification of label-retaining cells in mouse endometrium.Stem Cells (Dayton, Ohio) 2006;24:1529–1538.
Chen JQ, Russo PA, Cooke C, Russo IH, Russo J. ERbeta shifts from mitochondria tonucleus during estrogen-induced neoplastic transformation of human breastepithelial cells and is involved in estrogen-induced synthesis of mitochondrialrespiratory chain proteins. Biochim Biophys Acta 2007;1773:1732–1746.
Cheng G, Wilczek B, Warner M, Gustafsson JA, Landgren BM. Isoflavone treatment foracute menopausal symptoms. Menopause 2007;14:468–473.
Chhieng D, Hui P. Normal histology of female genital organs. In: Chhieng D, Hui P (eds).Current Clinical Pathology. NJ, USA: Humana Press, 2011, 1–13.
Collins F, Macpherson S, Brown P, Bombail V, Williams A, Anderson R, Jabbour H,Saunders P. Expression of estrogen receptors, ERalpha, ERbeta, and ERbetavariants, in endometrial cancers and evidence that prostaglandin F may play a rolein regulating expression of ERalpha. BMC Cancer 2009;9:330.
Cooke PS, Spencer TE, Bartol FF, Hayashi K. Uterine glands: development, function andexperimental model systems. Mol Hum Reprod 2013;19:547–558.
Costa-Paiva L, Godoy CE Jr, Antunes A Jr, Caseiro JD, Arthuso M, Pinto-Neto AM.Risk of malignancy in endometrial polyps in premenopausal and postmenopausalwomen according to clinicopathologic characteristics. Menopause 2011;18:1278–1282.
Couse JF, Korach KS. Estrogen receptor null mice: what have we learned and where willthey lead us? Endocr Rev 1999;20:358–417.
Crandall CJ, Barrett-Connor E. Endogenous sex steroid levels and cardiovasculardisease in relation to the menopause: a systematic review. Endocrinol Metab ClinNorth Am 2013;42:227–253.
Critchley HOD, Saunders PTK. Hormone receptor dynamics in a receptive humanendometrium. Reprod Sci 2009;16:191–199.
Critchley HOD, Brenner RM, Henderson TA, Williams K, Nayak NR, Slayden OD,Millar MR, Saunders PTK. Estrogen receptor beta, but not estrogen receptoralpha, is present in the vascular endothelium of the human and nonhuman primateendometrium. J Clin Endocrinol Metab 2001;86:1370–1378.
Critchley HOD, Henderson TA, Kelly RW, Scobie GS, Evans LR, Groome NP,Saunders PTK, Critchley HOD, Henderson TA, Kelly RW et al. Wild-typeestrogen receptor (ERbeta1) and the splice variant (ERbetacx/beta2) are bothexpressed within the human endometrium throughout the normal menstrualcycle. J Clin Endocrinol Metab 2002;87:5265–5273.
Critchley HO, Kelly RW, Baird DT, Brenner RM. Regulation of human endometrialfunction: mechanisms relevant to uterine bleeding. Reprod Biol Endocrinol 2006;4(Suppl. 1):S5.
Donnez O, Van Langendonckt A, Defrere S, Colette S, Van Kerk O, Dehoux JP,Squifflet J, Donnez J. Induction of endometriotic nodules in an experimentalbaboon model mimicking human deep nodular lesions. Fertil Steril 2012;99:783–789.e783.
Dupont S, Krust A, Gansmuller A, Dierich A, Chambon P, Mark M. Effect of single andcompound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) onmouse reproductive phenotypes. Development 2000;127:4277–4291.
Engemise SL, Willets JM, Taylor AH, Emembolu JO, Konje JC. Changes in glandular andstromal estrogen and progesterone receptor isoform expression in eutopic andectopic endometrium following treatment with the levonorgestrel-releasingintrauterine system. Eur J Obstet Gynecol Reprod Biol 2011;157:101–106.
Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, Fried G,Nordenskjold M, Gustafsson JA. Human estrogen receptor beta-gene structure,chromosomal localization, and expression pattern. J Clin Endocrinol Metab 1997;82:4258–4265.
Erdemoglu E, Guney M, Karahan N, Mungan T. Expression of cyclooxygenase-2, matrixmetalloproteinase-2 and matrix metalloproteinase-9 in premenopausal andpostmenopausal endometrial polyps. Maturitas 2008;59:268–274.
Evans T, Sany O, Pearmain P, Ganesan R, Blann A, Sundar S. Differentialtrends in the rising incidence of endometrial cancer by type: data from aUK population-based registry from 1994 to 2006. Br J Cancer 2011;104:1505–1510.
Fazleabas AT, Brudney A, Chai D, Langoi D, Bulun SE. Steroid receptor andaromatase expression in baboon endometriotic lesions. Fertil Steril 2003;80(Suppl. 2):820–827.
Fearnley EJ, Marquart L, Spurdle AB, Weinstein P, Webb PM. Polycystic ovarysyndrome increases the risk of endometrial cancer in women aged less than 50years: an Australian case-control study. Cancer Causes Control 2010;21:2303–2308.
Ferenczy A. Studies on the cytodynamics of human endometrial regeneration. II.Transmission electron microscopy and histochemistry. Am J Obstet Gynecol 1976;124:582–595.
Ferenczy A. Regeneration of the human endometrium. In: Wolff CMFaM (ed). Progressin Surgical Pathology. New York, USA: Masson, 1980, 157–173.
Ferenczy A, Guralnicik M. Endometrial microstructure: structure–functionrelationships throughout the menstrual cycle. Semin Reprod Endocrinol 1983;1:205–219.
Ferenczy A, Bergeron C. Histology of the human endometrium: from birth tosenescence. Ann N Y Acad Sci 1991;622:6–27.
Fernandez JW, Grizzell JA, Wecker L. The role of estrogen receptor beta and nicotiniccholinergic receptors in postpartum depression. Prog Neuropsychopharmacol BiolPsychiatry 2012;40:199–206.
Filant J, Spencer TE. Endometrial glands are essential for blastocyst implantation anddecidualization in the mouse uterus. Biol Reprod 2013;88:93.
Frasor J, Barnett DH, Danes JM, Hess R, Parlow AF, Katzenellenbogen BS.Response-specific and ligand dose-dependent modulation of estrogen receptor(ER) alpha activity by ERbeta in the uterus. Endocrinology 2003;144:3159–3166.
Fujimoto J, Hirose R, Sakaguchi H, Tamaya T. Expression of estrogen receptor-alphaand -beta in ovarian endometriomata. Mol Hum Reprod 1999;5:742–747.
Fujimoto J, Sakaguchi H, Aoki I, Khatun S, Toyoki H, Tamaya T. Review: steroidreceptors and metastatic potential in endometrial cancers. J Steroid Biochem MolBiol 2000;75:209–212.
Fujimura T, Takahashi S, Urano T, Ogawa S, Ouchi Y, Kitamura T, Muramatsu M,Inoue S. Differential expression of estrogen receptor beta (ERbeta) and itsC-terminal truncated splice variant ERbetacx as prognostic predictors in humanprostatic cancer. Biochem Biophys Res Commun 2001;289:692–699.
Gargett CE, Masuda H. Adult stem cells in the endometrium. Mol Hum Reprod 2010;16:818–834.
Gargett CE, Chan RW, Schwab KE. Hormone and growth factor signaling inendometrial renewal: role of stem/progenitor cells. Mol Cell Endocrinol 2008;288:22–29.
Gargett CE, Nguyen HP, Ye L. Endometrial regeneration and endometrial stem/progenitor cells. Rev Endocr Metab Disord 2012;13:235–251.
Grady D, Sawaya GF, Johnson KC, Koltun W, Hess R, Vittinghoff E, Kristof M,Tagliaferri M, Cohen I, Ensrud KE. MF101, a selective estrogen receptor betamodulator for the treatment of menopausal hot flushes: a phase II clinical trial.Menopause (New York, NY) 2009;16:458–465.
Greaves E, Collins F, Critchley HO, Saunders PT. ERbeta-dependent effects on uterineendothelial cells are cell specific and mediated via Sp1. Hum Reprod (Oxford, England)2013;28:2490–2501.
Gustafsson JA. What pharmacologists can learn from recent advances in estrogensignalling. Trends Pharmacol Sci 2003;24:479–485.
Hachisuga T, Miyakawa T, Tsujioka H, Horiuchi S, Emoto M, Kawarabayashi T. K-rasmutation in tamoxifen-related endometrial polyps. Cancer 2003;98:1890–1897.
Hall JM, Mcdonnell DP. The estrogen receptor beta-isoform (ERbeta) of the humanestrogen receptor modulates ERalpha transcriptional activity and is a key regulatorof the cellular response to estrogens and antiestrogens. Endocrinology 1999;140:5566–5578.
Hall JM, Mcdonnell DP. Coregulators in nuclear estrogen receptor action—fromconcept to therapeutic targeting. Mol Interv 2005;5:343–357.
Han SJ, Hawkins SM, Begum K, Jung SY, Kovanci E, Qin J, Lydon JP, Demayo FJ,O’malley BW. A new isoform of steroid receptor coactivator-1 is crucial forpathogenic progression of endometriosis. Nat Med 2012;18:1102–1111.
Hanamura T, Niwa T, Gohno T, Kurosumi M, Takei H, Yamaguchi Y, Ito K-I, Hayashi S-I.Possible roleof the aromatase-independent steroidmetabolismpathways inhormoneresponsive primary breast cancers. Breast Cancer Res Treat 2014;143:69–80.
Hapangama D. Mifepristone: the multi-faceted nti-hormone. J Drug Eval 2003;1:149–175.
Hapangama D, Bulmer J. Contemporary approaches to heavy menstrual bleeding. In:Justin Clark T, Natalie AM (eds). Pathophysiology of HMB. London, UK: FutureMedicine Ltd, 2014.
Estrogen receptor b in the endometrium 189
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

Hapangama DK, Turner MA, Drury JA, Martin-Ruiz C, Von Zglinicki T,Farquharson RG, Quenby S. Endometrial telomerase shows specific expressionpatterns in different types of reproductive failure. Reprod Biomed Online 2008a;17:416–424.
Hapangama DK, Turner MA, Drury JA, Quenby S, Saretzki G, Martin-Ruiz C,Von Zglinicki T. Endometriosis is associated with aberrant endometrialexpression of telomerase and increased telomere length. Hum Reprod 2008b;23:1511–1519.
Hapangama DK, Turner MA, Drury JA, Quenby S, Hart A, Maddick M, Martin-Ruiz C,Von Zglinicki T. Sustained replication in endometrium of women with endometriosisoccurs without evoking a DNA damage response. Hum Reprod 2009;24:687–696.
Hapangama DK, Raju RS, Valentijn AJ, Barraclough D, Hart A, Turner MA,Platt-Higgins A, Barraclough R, Rudland PS. Aberrant expression of metastasis-inducing proteins in ectopic and matched eutopic endometrium of women withendometriosis: implications for the pathogenesis of endometriosis. Hum Reprod2012;27:394–407.
Haring J, Schuler S, Lattrich C, Ortmann O, Treeck O. Role of estrogen receptor b ingynecological cancer. Gynecol Oncol 2012a;127:673–676.
Haring J, Skrzypczak M, Stegerer A, Lattrich C, Weber F, Gorse R, Ortmann O,Treeck O. Estrogen receptor beta transcript variants associate with oncogeneexpression in endometrial cancer. Int J Mol Med 2012b;29:1127–1136.
Harnish DC. Estrogen receptor ligands in the control of pathogenic inflammation. CurrOpin Investig Drugs (London, England: 2000) 2006;7:997–1001.
Harris HA. Estrogen receptor-beta: recent lessons from in vivo studies. Mol Endocrinol2006;21:1–13.
Hawse JR, Subramaniam M, Monroe DG, Hemmingsen AH, Ingle JN, Khosla S,Oursler MJ, Spelsberg TC. Estrogen receptor beta isoform-specific induction oftransforming growth factor beta-inducible early gene-1 in human osteoblast cells:an essential role for the activation function 1 domain. Mol Endocrinol 2008;22:1579–1595.
Henderson TA, Saunders PT, Moffett-King A, Groome NP, Critchley HO. Steroidreceptor expression in uterine natural killer cells. J Clin Endocrinol Metab 2003;88:440–449.
Hertrampf T, Seibel J, Laudenbach U, Fritzemeier KH, Diel P. Analysis of the effects ofestrogen receptor alpha (ERalpha)- and ERbeta-selective ligands given incombination to ovariectomized rats. Br J Pharmacol 2008;153:1432–1437.
Hogan AM, Collins D, Baird AW, Winter DC. Estrogen and gastrointestinal malignancy.Mol Cell Endocrinol 2009;307:19–24.
Holm A, Nilsson BO. Identification and characterization of new mechanisms in vascularestrogen signalling. Basic Clin Pharmacol Toxicol 2013;113:287–293.
Hombach-Klonisch S, Kehlen A, Fowler PA, Huppertz B, Jugert JF, Bischoff G,Schluter E, Buchmann J, Klonisch T. Regulation of functional steroid receptors andligand-induced responses in telomerase-immortalized human endometrialepithelial cells. J Mol Endocrinol 2005;34:517–534.
Hong-Bing C, Yun-Feng Z, Bi-Cheng W, Ling-Ling G. Expression of estrogen receptorsa and b protein in endometrial carcinoma. Cancer Ther 2008;6:907–911.
Hu K, Zhong G, He F. Expression of estrogen receptors ER[alpha] and ER[beta] inendometrial hyperplasia and adenocarcinoma. Int J Gynecol Cancer 2005;15:537–541.
Hu J, Wang J, Xiang H, Li Z, Wang B, Cao Y, Ma X. Association of polymorphisms inthe estrogen receptor beta (ESR2) with unexplained recurrent spontaneousabortion (URSA) in Chinese population. J Matern Fetal Neonatal Med 2012;25:1727–1729.
Hudelist G, Keckstein J, Czerwenka K, Lass H, Walter I, Auer M, Wieser F, Wenzl R,Kubista E, Singer CF. Estrogen receptor beta and matrix metalloproteinase 1 arecoexpressed in uterine endometrium and endometriotic lesions of patients withendometriosis. Fertil Steril 2005;84(Suppl. 2):1249–1256.
Imamov O, Shim GJ, Warner M, Gustafsson JA. Estrogen receptor beta in health anddisease. Biol Reprod 2005;73:866–871.
Indraccolo U, Di Iorio R, Matteo M, Coron G, Greco P, Indraccolo SR. Thepathogenesis of endometrial polyps: a systematic semi-quantitative review. Eur JGynaecol Oncol 2013;34:5–22.
Jabbour HN, Kelly RW, Fraser HM, Critchley HOD. Endocrine regulation ofmenstruation. Endocr Rev 2006;27:17–46.
Janzen DM, Cheng D, Schafenacker AM, Paik DY, Goldstein AS, Witte ON,Jaroszewicz A, Pellegrini M, Memarzadeh S. Estrogen and progesteronetogether expand murine endometrial epithelial progenitor cells. Stem Cells 2013;31:808–822.
Jarzabek K, Koda M, Walentowicz-Sadlecka M, Grabiec M, Laudanski P, Wolczynski S.Altered expression of ERs, aromatase, and COX2 connected to estrogen action intype 1 endometrial cancer biology. Tumour Biol 2013;34:4007–4016.
Jazaeri AA, Nunes KJ, Dalton MS, Xu M, Shupnik MA, Rice LW. Well-differentiatedendometrial adenocarcinomas and poorly differentiated mixed mullerian tumorshave altered ER and PR isoform expression. Oncogene 2001;20:6965–6969.
Jongen V, Briet J, De Jong R, Ten Hoor K, Boezen M, Van Der Zee A, Nijman H,Hollema H. Expression of estrogen receptor-alpha and -beta and progesteronereceptor-A and -B in a large cohort of patients with endometrioid endometrialcancer. Gynecol Oncol 2009;112:537–542.
Juhasz-Bass I, Fischer C, Lattrich C, Skrzypczak M, Malik E, Ortmann O, Treeck O.Endometrial expression of estrogen receptor I2 and its splice variants in patientswith and without endometriosis. Arch Gynecol Obstet 2011;284:885–891.
Katzenellenbogen BS. Biology and receptor interactions of estriol and estriol derivativesin vitro and in vivo. J Steroid Biochem 1984;20:1033–1037.
Katzenellenbogen B, Sun J, Harrington WR, Kraichely DM, Ganessunker D,Katzenellenbogen JA. Structure–function relationships in estrogen receptors andthe characterization of novel selective estrogen receptor modulators with uniquepharmacological profiles. Ann NY Acad Sci 2001;949:6–15.
Kayisli UA, Luk J, Guzeloglu-Kayisli O, Seval Y, Demir R, Arici A. Regulation ofangiogenic activity of human endometrial endothelial cells in culture by ovariansteroids. J Clin EndocrinolMetab 2004;89:5794–5802.
Keay J, Bridgham JT, Thornton JW. The Octopus vulgaris estrogen receptor is aconstitutive transcriptional activator: evolutionary and functional implications.Endocrinology 2006;147:3861–3869.
Kim JJ, Choi YM, Choung SH, Yoon SH, Lee GH, Moon SY. Estrogen receptor beta gene+1730 G/A polymorphism in women with polycystic ovary syndrome. Fertil Steril2010;93:1942–1947.
Knapp P, Chabowski A, Blachnio-Zabielska A, Walentowicz-Sadlecka M, Grabiec M,Knapp PA. Expression of estrogen receptors (alpha, beta), cyclooxygenase-2 andaromatase in normal endometrium and endometrioid cancer of uterus. Adv MedSci 2013;58:96–103.
Koos RD. Minireview: putting physiology back into estrogens’ mechanism of action.Endocrinology 2011;152:4481–4488.
Kramer PR, Wray S. 17-Beta-estradiol regulates expression of genes that function inmacrophage activation and cholesterol homeostasis. J Steroid Biochem Mol Biol2002;81:203–216.
Krikun G, Schatz F, Taylor R, Critchley HO, Rogers PA, Huang J, Lockwood CJ.Endometrial endothelial cell steroid receptor expression and steroid effects ongene expression. J Clin Endocrinol Metab 2005;90:1812–1818.
Kuiper GG, Gustafsson JA. The novel estrogen receptor-beta subtype: potential role inthe cell- and promoter-specific actions of estrogens and anti-estrogens. FEBS Lett1997;410:87–90.
Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novelreceptor expressed in rat prostate and ovary. Proc Natl Acad Sci USA 1996;93:5925–5930.
Kumar R, Zakharov MN, Khan SH, Miki R, Jang H, Toraldo G, Singh R, Bhasin S, Jasuja R.The dynamic structure of the estrogen receptor. J Amino Acids 2012;2011:812540.
Kushner PJ, Agard DA, Greene GL, Scanlan TS, Shiau AK, Uht RM, Webb P. Estrogenreceptor pathways to AP-1. J Steroid Biochem Mol Biol 2000;74:311–317.
Labrie F. All sex steroids are made intracellularly in peripheral tissues by themechanisms of intracrinology after menopause. J Steroid Biochem Mol Biol 2014;2014:06.001.
Lacey JV Jr, Ioffe OB, Ronnett BM, Rush BB, Richesson DA, Chatterjee N, Langholz B,Glass AG, Sherman ME. Endometrial carcinoma risk among women diagnosed withendometrial hyperplasia: the 34-year experience in a large health plan. Br J Cancer2007;98:45–53.
Lash GE, Robson SC, Bulmer JN. Review: functional role of uterine natural killer (uNK)cells in human early pregnancy decidua. Placenta 2010;31(Suppl.):S87–S92.
Lecce G, Meduri G, Ancelin M, Bergeron C, Perrot-Applanat M. Presence of estrogenreceptor beta in the human endometrium through the cycle: expression in glandular,stromal, and vascular cells. J Clin Endocrinol Metab 2001;86:1379–1386.
Lewandowski S, Kalita K, Kaczmarek L. Estrogen receptor beta. Potential functionalsignificance of a variety of mRNA isoforms. FEBS Lett 2002;524:1–5.
Leyendecker G, Herbertz M, Kunz G, Mall G. Endometriosis results from thedislocation of basal endometrium. Hum Reprod 2002;17:2725–2736.
Leyendecker G, Herbertz M, Kunz G, Mall G. Endometriosis results from thedislocation of basal endometrium. Hum Reprod 2008;17:2725–2736.
190 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

Leygue E, Dotzlaw H, Lu B, Glor C, Watson PH, Murphy LC. Estrogen receptor beta:mine is longer than yours? J Clin Endocrinol Metab 1998;83:3754–3755.
Li XC, Song WJ. Endometrial Intraepithelial Neoplasia (EIN) in endometrial biopsyspecimens categorized by the 1994 World Health Organization classification forendometrial hyperplasia. Asian Pac J Cancer Prev 2013;14:5935–5939.
Lubahn DB, Moyer JS, Smithies O, Golding TS, Couse JF, Korach KS. Alteration ofreproductive function but not prenatal sexual development after insertionaldisruption of the mouse estrogen receptor gene. Proc Natl Acad Sci USA 1993;90:11162–11166.
Maliqueo M, Clementi M, Gabler F, Johnson MC, Palomino A, Sir-Petermann T, Vega M.Expression of steroid receptors and proteins related to apoptosis in endometria ofwomen with polycystic ovary syndrome. Fertil Steril 2003;80:812–819.
Maret A, Coudert JD, Garidou L, Foucras G, Gourdy P, Krust A, Dupont S, Chambon P,Druet P, Bayard F et al. Estradiol enhances primary antigen-specific CD4T cellresponses and Th1 development in vivo. Essential role of estrogen receptor alphaexpression in hematopoietic cells. Eur J Immunol 2003;33:512–521.
Marshall RJ, Jones DB. An immunohistochemical study of lymphoid tissue in humanendometrium. Int J Gynecol Pathol 1988;7:225–235.
Masuda H, Kalka C, Takahashi T, Yoshida M, Wada M, Kobori M, Itoh R, Iwaguro H,Eguchi M, Iwami Y et al. Estrogen-mediated endothelial progenitor cell biology andkinetics for physiological postnatal vasculogenesis. Circ Res 2007;101:598–606.
Masuda H, Matsuzaki Y, Hiratsu E, Ono M, Nagashima T, Kajitani T, Arase T, Oda H,Uchida H, Asada H et al. Stem cell-like properties of the endometrial sidepopulation: implication in endometrial regeneration. PLoS One 2010;5:e10387.
Matsuzaki S, Fukaya T, Suzuki T, Murakami T, Sasano H, Yajima A. Estrogen receptoralpha and beta mRNA expression in human endometrium throughout themenstrual cycle. Mol Hum Reprod 1999;5:559–564.
Matsuzaki S, Uehara S, Murakami T, Fujiwara J, Funato T, Okamura K. Quantitativeanalysis of estrogen receptor alpha and beta messenger ribonucleic acid levels innormal endometrium and ovarian endometriotic cysts using a real-time reversetranscription-polymerase chain reaction assay. Fertil Steril 2000;74:753–759.
Matsuzaki S, Murakami T, Uehara S, Canis M, Sasano H, Okamura K. Expression ofestrogen receptor alpha and beta in peritoneal and ovarian endometriosis. FertilSteril 2001;75:1198–1205.
Matthews J, Wihlen B, Tujague M, Wan J, Strom A, Gustafsson JA. Estrogen receptor(ER) beta modulates ERalpha-mediated transcriptional activation by altering therecruitment of c-Fos and c-Jun to estrogen-responsive promoters. Mol Endocrinol2006;20:534–543.
May FE. Novel drugs that target the estrogen-related receptor alpha: their therapeuticpotential in breast cancer. Cancer Manag Res. 2014; 23:225–252.
Mccluggage WG. Benign Diseases of the Endometrium. In: Kurman R, Ellenson L,Ronnett B (eds). Pathology of Female Genetal Tract. Boston, USA: Springer, 2011,305–358.
Mcdonnell DP, Norris JD. Connections and regulation of the human estrogen receptor.Science 2002;296:1642–1644.
Mehasseb MK, Panchal R, Taylor AH, Brown L, Bell SC, Habiba M. Estrogen andprogesterone receptor isoform distribution through the menstrual cycle in uteriwith and without adenomyosis. Fertil Steril 2011;95:2228–2235.
Menasce LP, White GR, Harrison CJ, Boyle JM. Localization of the estrogen receptorlocus (ESR) to chromosome 6q25.1 by FISH and a simple post-FISH bandingtechnique. Genomics 1993;17:263–265.
Milanesi L, Vasconsuelo A, De Boland AR, Boland R. Expression and subcellulardistribution of native estrogen receptor beta in murine C2C12 cells and skeletalmuscle tissue. Steroids 2009;74:489–497.
Moore JT, Mckee DD, Slentz-Kesler K, Moore LB, Jones SA, Horne EL, Su JL,Kliewer SA, Lehmann JM, Willson TM. Cloning and characterization of humanestrogen receptor beta isoforms. Biochem Biophys Res Commun 1998;247:75–78.
Mosselman S, Polman J, Dijkema R. ER beta: identification and characterization of anovel human estrogen receptor. FEBS Lett 1996;392:49–53.
Murali R, Soslow RA, Weigelt B. Classification of endometrial carcinoma: more thantwo types. Lancet Oncol 2014;15:e268–e278.
Mylonas I, Jeschke U, Shabani N, Kuhn C, Kriegel S, Kupka MS, Friese K. Normal andmalignant human endometrium express immunohistochemically estrogenreceptor alpha (ER-alpha), estrogen receptor beta (ER-beta) and progesteronereceptor (PR). Anticancer Res 2005;25:1679–1686.
Mylonas I, Jeschke U, Shabani N, Kuhn C, Kunze S, Dian D, Friedl C, Kupka MS, Friese K.Steroid receptors ERalpha, ERbeta, PR-A and PR-B are differentially expressed innormal and atrophic human endometrium. Histol Histopathol 2007;22:169–176.
Narkar M, Kholkute S, Chitlange S, Nandedkar T. Expression of steroid hormonereceptors, proliferation and apoptotic markers in primate endometrium. Mol CellEndocrinol 2006;246:107–113.
Nayak NR, Brenner RM. Vascular proliferation and vascular endothelial growth factorexpression in the rhesus macaque endometrium. J Clin Endocrinol Metab 2002;87:1845–1855.
Negoita M, Mihailovici MS. Expression of hormonal receptors (alpha-estrogen,beta-estrogen, progesteron), Ki-67 and P53 in endometrium of tamoxifen treatedbreast cancer patients. Rev Med Chir Soc Med Nat Iasi 2011;115:834–838.
NguyenHP, Sprung CN, Gargett CE. Differential expression of Wnt signaling moleculesbetween pre- and postmenopausal endometrial epithelial cells suggests a populationof putative epithelial stem/progenitor cells reside in the basalis layer. Endocrinology2012;153:2870–2883.
Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E,Pettersson K, Warner M, Gustafsson JA. Mechanisms of estrogen action. PhysiolRev 2001;81:535–1565.
Ogawa S, Inoue S, Watanabe T, Orimo A, Hosoi T, Ouchi Y, Muramatsu M. Molecularcloning and characterization of human estrogen receptor betacx: a potentialinhibitor ofestrogen action in human. Nucleic Acids Res 1998;26:3505–3512.
O’neill PA, Davies MPA, Shaaban AM, Innes H, Torevell A, Sibson DR, Foster CS.Wild-type estrogen receptor beta (ERbeta1) mRNA and protein expression intamoxifen-treated post-menopausal breast cancers. Br J Cancer 2004;91:1694–1702.
Ordi J, Bergeron C, Hardisson D, Mccluggage WG, Hollema H, Felix A, Soslow RA,Oliva E, Tavassoli FA, Alvarado-Cabrero I et al. Reproducibility of currentclassifications of endometrial endometrioid glandular proliferations: furtherevidence supporting a simplified classification. Histopathology 2013;64:284–292.
Owings RA, Quick CM. Endometrial intraepithelial neoplasia. ArchPathol LabMed 2014;138:484–491.
Pace P, Taylor J, Suntharalingam S, Coombes RC, Ali S. Human estrogen receptor betabinds DNA in a manner similar to and dimerizes with estrogen receptor alpha. J BiolChem 1997;272:25832–25838.
Padykula HA, Coles LG, Mccracken JA, King NW, Longcope C, Kaiserman-Abramof I.A zonal pattern of cell proliferation and differentiation in the rhesus endometriumduring the estrogen surge. Biol Reprod 1984;31:1103–1118.
Padykula HA, Coles LG, Okulicz WC, Rapaport SI, Mc Cracken JA, King NW Jr,Longcope C, Kaiserman-Abramof I. The basalis of the primate endometrium: abifunctional germinal compartment. Biol Reprod 1989;40:681–690.
Paech K, Webb P, Kuiper GG, Nilsson S, Gustafsson J, Kushner PJ, Scanlan TS.Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1sites. Science 1997;277:1508–1510.
Park JC, Lim SY, Jang TK, Bae JG, Kim JI, Rhee JH. Endometrial histology and predictableclinical factors for endometrial disease in women with polycystic ovary syndrome.Clin Exp Reprod Med 2011;38:42–46.
Paruthiyil S, Parmar H, Kerekatte V, Cunha GR, Firestone GL, Leitman DC. Estrogenreceptor beta inhibits human breast cancer cell proliferation and tumor formationby causing a G2 cell cycle arrest. Cancer Res 2004;64:423–428.
Pastore MB, Jobe SO, Ramadoss J, Magness RR. Estrogen receptor-alpha and estrogenreceptor-beta in the uterine vascular endothelium during pregnancy: functionalimplications for regulating uterine blood flow. Semin Reprod Med 2012;30:46–61.
Paul M, Chotewa K, Mazurek U, Witek A, Wilezok T. Estrogen receptorbD6 (ERbD6)isoform in human endometrial hyperplasia and adenocarcinoma. Cancer Invest 2004;22:211–218.
Pettersen BG. Osteopontin—an important downstream effector of S100A4-mediatedinvasion and metastasis. Int J Cancer 2011;129:780–790.
Prakansamut N, Sirayapiwat P, Triratanachat S. The percentages of endometrialhyperplasia and endometrial cancer among polycystic ovary syndrome (PCOS)patients presenting with abnormal menstrual pattern. J Med Assoc Thai 2014;97:159–164.
Prianishnikov VA. A functional model of the structure of the epithelium of normal,hyperplastic and malignant human endometrium: a review. Gynecol Oncol 1978;6:420–428.
Prossnitz ER, Barton M. Estrogen biology: new insights into GPER function and clinicalopportunities. Mol Cell Endocrinol 2014;389:71–83.
Ramsey TL, Risinger KE, Jernigan SC, Mattingly KA, Klinge CM. Estrogen receptorbeta isoforms exhibit differences in ligand-activated transcriptional activity in anestrogen response element sequence-dependent manner. Endocrinology 2004;145:149–160.
Estrogen receptor b in the endometrium 191
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

Rees MC, Bicknell R. Angiogenesis in the endometrium. Angiogenesis 1998;2:29–35.Rider V, Li X, Peterson G, Dawson J, Kimler BF, Abdou NI. Differential expression of
estrogen receptors in women with systemic lupus erythematosus. J Rheumatol2006;33:1093–1101.
Rizza P, Barone I, Zito D, Giordano F, Lanzino M, De Amicis F, Mauro L, Sisci D,Catalano S, Wright KD et al. Estrogen receptor beta as a novel target of androgenreceptor action in breast cancer cell lines. Breast Cancer Res 2014;16:R21.
Robson A, Harris LK, Innes BA, Lash GE, Aljunaidy MM, Aplin JD, Baker PN,Robson SC, Bulmer JN. Uterine natural killer cells initiate spiral artery remodelingin human pregnancy. FASEB J 2012;26:4876–4885.
Rody A, Holtrich U, Solbach C, Kourtis K, Von Minckwitz G, Engels K, Kissler S,Gatje R, Karn T, Kaufmann M. Methylation of estrogen receptor beta promotercorrelates with loss of ER-beta expression in mammary carcinoma and is anearly indication marker in premalignant lesions. Endocr Relat Cancer 2005;12:903–916.
Saegusa M, Okayasu I. Changes in expression of estrogen receptors alpha and beta inrelation to progesterone receptor and pS2 status in normal and malignantendometrium. Jpn J Cancer Res 2000;91:510–518.
Saha P, Saraswat G, Chakraborty P, Banerjee S, Pal BC, Kabir SN. Puerarin, a selectiveestrogen receptor modulator, disrupts pregnancy in rats at pre-implantation stage.Reproduction 2012;144:633–645.
Sakaguchi H, Fujimoto J, Aoki I, Toyoki H, Khatun S, Tamaya T. Expression of estrogenreceptor alpha and beta in uterine endometrial and ovarian cancers. Eur J Cancer2002;38(Suppl. 6):S74–S75.
Sastre-Serra J, Nadal-Serrano M, Pons DG, Valle A, Garau I, Garcia-Bonafe M, Oliver J,Roca P. The oxidative stress in breast tumors of postmenopausal women is ERalpha/ERbeta ratio dependent. Free Radic Biol Med 2013;61C:11–17.
Saxon LK, Turner CH. Estrogen receptor beta: the antimechanostat? Bone 2005;36:185–192.
Setiawan VW, Pike MC, Karageorgi S, Deming SL, Anderson K, Bernstein L, Brinton LA,Cai H, Cerhan JR, Cozen W et al. Age at Last Birth in Relation to Risk of EndometrialCancer: Pooled Analysis in the Epidemiology of Endometrial Cancer Consortium. USA:Oxford University Press, 2012, 269–278.
Shao R, Wang X, Feng Y, Billig H, Cao S. The elusive and controversial roles of estrogenand progesterone receptors in human endometriosis. Am J Transl Res 2014;6:104–113.
Silvestri A, Fraser HM. Estrogen and progesterone receptors in the marmosetendometrium: changes during the ovulatory cycle, early pregnancy and afterinhibition of vascular endothelial growth factor, GnRH or ovariectomy.Reproduction 2007;134:341–353.
Skrzypczak M, Bieche I, Szymczak S, Tozlu S, Lewandowski S, Girault I, Radwanska K,Szczylik C, Jakowicki JA, Lidereau R et al. Evaluation of mRNA expression of estrogenreceptor beta and its isoforms in human normal and neoplastic endometrium. Int JCancer 2004;110:783–787.
Slayden D, Brenner RM. Hormonal regulation and localization of estrogen, progestinand androgen receptors in the endometrium of nonhuman primates: effects ofprogesterone receptor antagonists. Arch Histol Cytol 2004;67:393–409.
Smith LH, Coats SR, Qin H, Petrie MS, Covington JW, Su M, Eren M, Vaughan DE.Differential and opposing regulation of PAI-1 promoter activity by estrogenreceptor alpha and estrogen receptor beta in endothelial cells. Circ Res 2004;95:269–275.
Smuc T, Rizner TL. Aberrant pre-receptor regulation of estrogen and progesteroneaction in endometrial cancer. Mol Cell Endocrinol 2009;301:74–82.
Sourial S, Tempest N, Hapangama D. Theories on the pathogenesis of endometriosis.Int J Reprod Med 2014;2014:179515.
Spencer TE, Dunlap KA, Filant J. Comparative developmental biology of the uterus:insights into mechanisms and developmental disruption. Mol Cell Endocrinol 2011;354:34–53.
Takama F, Kanuma T, Wang D, Kagami I, Mizunuma H. Estrogen receptorb expressionand depth of myometrial invasion in human endometrial cancer. Br J Cancer 2001;84:545–549.
Tan NS, Frecer V, Lam TJ, Ding JL. Temperature dependence of estrogen binding:importance of a subzone in the ligand binding domain of a novel piscine estrogenreceptor. Biochim Biophys Acta 1999;1452:103–120.
Taylor AH, Al-Azzawi F. Immunolocalisation of estrogen receptor beta in humantissues. J Mol Endocrinol 2000;24:145–155.
Thanapprapasr D, Thanapprapasr K. Molecular therapy as a future strategy inendometrial cancer. Asian Pac J Cancer Prev 2013;14:3419–3423.
Thomas C, Gustafsson J-A. The different roles of ER subtypes in cancer biology andtherapy. Nat Rev Cancer 2011;11:597–608.
Thornton JW. Evolution of vertebrate steroid receptors from an ancestral estrogenreceptor by ligand exploitation and serial genome expansions. Proc Natl Acad SciUSA 2001;98:5671–5676.
Thornton JW, Need E, Crews D. Resurrecting the ancestral steroid receptor: ancientorigin of estrogen signaling. Science 2003;301:1714–1717.
Toft D, Gorski J. A receptor molecule for estrogens: isolation from the rat uterus andpreliminary characterization. Proc Natl Acad Sci USA 1966;55:1574–1581.
Tokyol C, Aktepe F, Dilek FH, Sahin O, Arioz DT. Expression of cyclooxygenase-2 andmatrix metalloproteinase-2 in adenomyosis and endometrial polyps and itscorrelation with angiogenesis. Int J Gynecol Pathol 2009;28:148–156.
Tremblay GB, Tremblay A, Labrie F, Giguere V. Dominant activity of activationfunction 1 (AF-1) and differential stoichiometric requirements for AF-1 and -2 inthe estrogen receptor alpha–beta heterodimeric complex. Mol Cell Biol 1999;19:1919–1927.
Trukhacheva E, Lin Z, Reierstad S, Cheng YH, Milad M, Bulun SE. Estrogen receptor(ER) beta regulates ERalpha expression in stromal cells derived from ovarianendometriosis. J Clin Endocrinol Metab 2009;94:615–622.
Utsunomiya H, Suzuki T, Harada N, Ito K, Matsuzaki S, Konno R, Sato S, Yajima A,Sasano H. Analysis of estrogen receptor alpha and beta in endometrialcarcinomas: correlation with ER beta and clinicopathologic findings in 45 cases. IntJ Gynecol Pathol 2000;19:335–341.
Valentijn AJ, Palial K, Al-Lamee H, Tempest N, Drury J, Hapangama DK, Murray P, VonZglinicki T, Saretzki G, Gargett CE. SSEA-1 isolates human endometrial basalglandular epithelial cells: phenotypic and functional characterization andimplications in the pathogenesis of endometriosis. Hum Reprod 2013;28:2695–2708.
Van Bogaert LJ. Clinicopathologic findings in endometrial polyps. Obstet Gynecol 1988;71:771–773.
Vani S, Critchley HO, Fraser IS, Hickey M. Endometrial expression of steroid receptorsin postmenopausal hormone replacement therapy users: relationship to bleedingpatterns. J Fam Plann Reprod Health Care 2008;34:27–34.
Vassiliadou N, Bulmer JN. Quantitative analysis of T lymphocyte subsets in pregnantand nonpregnant human endometrium. Biol Reprod 1996;55:1017–1022.
Villavicencio A, Bacallao K, Avellaira C, Gabler F, Fuentes A, Vega M. Androgen andestrogen receptors and co-regulators levels in endometria from patients withpolycystic ovarian syndrome with and without endometrial hyperplasia. GynecolOncol 2006;103:307–314.
Vivar OI, Saunier EF, Leitman DC, Firestone GL, Bjeldanes LF. Selective activation ofestrogen receptor-beta target genes by 3,3′-diindolylmethane. Endocrinology2010;151:1662–1667.
Wada-Hiraike O, Hiraike H, Okinaga H, Imamov O, Barros RP, Morani A, Omoto Y,Warner M, Gustafsson JA. Role of estrogen receptor beta in uterine stroma andepithelium: insights from estrogen receptor beta2/2 mice. Proc Natl Acad SciUSA 2006;103:18350–18355.
Walter P, Green S, Greene G, Krust A, Bornert JM, Jeltsch JM, Staub A, Jensen E,Scrace G, Waterfield M et al. Cloning of the human estrogen receptor cDNA.Proc Natl Acad Sci USA 1985;82:7889–7893.
Wang H, Isaksson E, Von Schoultz B, Cline JM, Sahlin L. The effect of long-termtreatment with steroid hormones or tamoxifen on estrogen receptors (alpha andbeta) in the endometrium of ovariectomized cynomolgus macaques. J Endocrinol2002;175:673–681.
Wang A, Ji L, Shang W, Li M, Chen L, White RE, Han G. Expression of GPR30,ERalpha and ERbeta in endometrium during window of implantation inpatients with polycystic ovary syndrome: a pilot study. Gynecol Endocrinol 2011;27:251–255.
Weihua Z, Saji S, Makinen S, Cheng G, Jensen EV, Warner M, Gustafsson J-A. Estrogenreceptor (ER) b, a modulator of ERa in the uterus. Proc Natl Acad Sci USA 2000;97:5936–5941.
Weiser MJ, Foradori CD, Handa RJ. Estrogen receptor beta in the brain: from form tofunction. Brain Res Rev 2008;57:309–320.
Winuthayanon W, Hewitt SC, Orvis GD, Behringer RR, Korach KS. Uterine epithelialestrogen receptor alpha is dispensable for proliferation but essential for completebiological and biochemical responses. Proc Natl Acad Sci USA 2010;107:19272–19277.
Witek A, Mazurek U, Paul M, Bierzynska-Macyszyn G, Wilczok T. Quantitative analysisof estrogen receptor-alpha and -beta and exon 5 splicing variant mRNA in
192 Hapangama et al.
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from

endometrial hyperplasia in perimenopausal women. Folia Histochem Cytobiol 2001;39:119–121.
Wu KH, Tobias ML, Thornton JW, Kelley DB. Estrogen receptors in Xenopus:duplicate genes, splice variants, and tissue-specific expression. Gen CompEndocrinol 2003;133:38–49.
Wu J, Wang C, Li X, Song Y, Wang W, Li C, Hu J, Zhu Z, Li J, Zhang W et al.Identification, characterization and application of a G-quadruplex structured DNAaptamer against cancer biomarker protein anterior gradient homolog 2. PLoS One2012;7:1–9.
Wu WF, Tan XJ, Dai YB, Krishnan V, Warner M, Gustafsson JA. Targeting estrogenreceptor beta in microglia and T cells to treat experimental autoimmuneencephalomyelitis. Proc Natl Acad Sci USA 2013;110:3543–3548.
Wynn RM. Cyclic and gestational changes. In: Wynn, Jollie WP (eds). The HumanEndometrium. New York, USA: Plenum Publishing Corp, 1989, 289–331.
Xue Q, Lin Z, Cheng YH, Huang CC, Marsh E, Yin P, Milad MP, Confino E,Reierstad S, Innes J et al. Promoter methylation regulates estrogenreceptor 2 in human endometrium and endometriosis. Biol Reprod 2007;77:681–687.
Yang JH, Wu MY, Chen CD, Chen MJ, Yang YS, Ho HN. Altered apoptosis andproliferation in endometrial stromal cells of women with adenomyosis. HumReprod(Oxford, England) 2007;22:945–952.
Yang YF, Liao YY, Peng NF, Li LQ, Xie SR, Wang RB. Prediction of coexistentcarcinomas risks by subjective EIN diagnosis and comparison with WHOclassification in endometrial hyperplasias. Pathol Res Pract 2012;208:708–712.
Yasui T, Matsui S, Tani A, Kunimi K, Yamamoto S, Irahara M. Androgen inpostmenopausal women. J Med Investig 2012;59:12–27.
Ye H, Xu XR, Liu YK, Liu ZH, Zhao AZ. Study on the relationship betwwen eastrogenreceptor beta and etiology of human endometrial polyps. Zhonghua Fu Chan Ke ZaZhi 2006;41:814–816.
Zang H, Sahlin L, Masironi B, Hirschberg AL. Effects of testosterone and estrogentreatment on the distribution of sex hormone receptors in the endometrium ofpostmenopausal women. Menopause (New York, NY) 2008;15:233–239.
Zannoni GF, Monterossi G, De Stefano I, Gargini A, Salerno MG, Farulla I, Travaglia D,Vellone VG, Scambia G, Gallo D. The expression ratios of estrogen receptor alpha(ERalpha) to estrogen receptor beta1 (ERbeta1) and ERalpha to ERbeta2 identify poorclinical outcome in endometrioid endometrial cancer. Hum Pathol 2013;44:1047–1054.
Zhang D, Trudeau VL. Integration of membrane and nuclear estrogen receptorsignaling. Comp Biochem Physiol A Mol Integr Physiol 2006;144:306–315.
Zhao C, Dahlman-Wright K, Gustafsson JA. Estrogen receptor beta: an overview andupdate. Nucl Receptor Signal 2008;6:e003.
Zitao L, Kuokkanen S, Pal L. Steroid hormone receptor profile of premenopausalendometrial polyps. Reprod Sci 2010;17:377–383.
Estrogen receptor b in the endometrium 193
at Universitas D
iponegoro on March 10, 2015
http://humupd.oxfordjournals.org/
Dow
nloaded from
Related Documents











