Nutrient Cycling in Agroecosystems 56: 177–193, 2000. © 2000 Kluwer Academic Publishers. Printed in the Netherlands. 177 Estimation of N 2 fixation in Desmodium ovalifolium from the relative ureide abundance of stem solutes: Comparison with the 15 N-dilution and an in situ soil core technique Bruno J.R. Alves 1 , Claudia de P. Rezende 2 , Alexander S. Resende 1 , Robert Macedo 1 , Ricardo M. Tarr´ e 1 , Segundo Urquiaga 1 & Robert M. Boddey 1,* 1 Embrapa-Agrobiologia, Km 47, Serop´ edica, 23851-970, Rio de Janeiro, Brazil; 2 Estação Experimental de Zootecnia de Extremo Sul da Bahia (CEPLAC-ESSUL), Km 757, BR 101, Itabela, BA, Brazil ( * Corresponding author; e-mail: [email protected]) Received 3 November 1999; accepted in revised form 10 May 1999 Key words: 15 N dilution technique, Desmodium ovalifolium, in situ N mineralisation, N 2 fixation, ureide technique Abstract Many, but not all, legumes of tropical origin, transport fixed N from the nodules to the shoot tissue in the form of ureides, and the mineral N absorbed from the soil is principally transported in the form of nitrate. The analysis of stem xylem sap, or hot-water extracts of stem tissue, for ureide and nitrate has been used successfully to quantify BNF contributions to several grain legumes and more recently to some shrub and forage legumes. The objective of this study was to investigate the application of this technique to the quantification of the contribution of BNF to the forage legume Desmodium ovalifolium by comparing the relative ureide abundance (RUA) of stem extracts of this plant with simultaneous estimates of BNF obtained using the 15 N isotope dilution technique. The first experiment was performed in pots of soil, taken from a grazing study, amended with 15 N-labelled organic matter at four different application rates. The ureide concentration in the stem extracts reflected the changes in BNF activity during plant growth and the RUA was closely correlated with the proportion of N derived from BNF as determined from the 15 N technique (r 2 = 0.86 and 0.88 for inoculated and non-inoculated plants, respectively). The use of a calibration curve derived from a previous study where the same legume was fed increasing concentrations of 15 N labelled nitrate in sand/vermiculite culture, resulted in an over-estimation of the BNF contribution which may have been due to a significant uptake of ammonium from this acidic soil. The second experiment was performed in field plots and a good agreement was found between the estimates of BNF derived from using the ureide and 15 N dilution techniques at two harvests six months apart. The uptake of soil N by the D. ovalifolium and two forage grasses (Brachiaria humidicola and Panicum maximum) was estimated using an in situ soil core technique, and, while the uptake of N by the grasses was successfully estimated, this technique underestimated the N derived from the soil by the legume as determined by the ureide and 15 N dilution techniques. Introduction The use of the ratio of ureide-N to total-N in the bleed- ing sap extracted from soybean stems to estimate the contribution of biological nitrogen fixation (BNF) to the nutrition of this legume, was first investigated by McClure et al. (1980) and tested and applied in both greenhouse and field experiments by Herridge and his colleagues (Herridge, 1982, 1984; Herridge and Peoples, 1990; Herridge et al., 1988; 1990). This tech- nique has also been successfully applied to quantify BNF to other ureide producing grain legumes (Rer- kasem et al., 1988; Peoples et al., 1988; Peoples et al., 1989, Hansen et al., 1993) as well as for some woody legumes (Herridge et al., 1996). In an attempt to quantify the contribution of BNF to the ureide-producing forage legumes Desmodium ovalifolium and a Centrosema hybrid, Alves et al. (1999a) found it almost impossible to extract sap from the stems of these legumes even under greenhouse

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nutrient Cycling in Agroecosystems56: 177–193, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
177
Estimation of N2 fixation in Desmodium ovalifoliumfrom the relativeureide abundance of stem solutes: Comparison with the15N-dilution andan in situ soil core technique
Bruno J.R. Alves1, Claudia de P. Rezende2, Alexander S. Resende1, Robert Macedo1, RicardoM. Tarre1, Segundo Urquiaga1 & Robert M. Boddey1,∗1Embrapa-Agrobiologia, Km 47, Serop´edica, 23851-970, Rio de Janeiro, Brazil;2Estação Experimental deZootecnia de Extremo Sul da Bahia (CEPLAC-ESSUL), Km 757, BR 101, Itabela, BA, Brazil(∗Corresponding author; e-mail: [email protected])
Received 3 November 1999; accepted in revised form 10 May 1999
Key words:15N dilution technique,Desmodium ovalifolium, in situN mineralisation, N2 fixation, ureide technique
Abstract
Many, but not all, legumes of tropical origin, transport fixed N from the nodules to the shoot tissue in the form ofureides, and the mineral N absorbed from the soil is principally transported in the form of nitrate. The analysis ofstem xylem sap, or hot-water extracts of stem tissue, for ureide and nitrate has been used successfully to quantifyBNF contributions to several grain legumes and more recently to some shrub and forage legumes. The objectiveof this study was to investigate the application of this technique to the quantification of the contribution of BNFto the forage legumeDesmodium ovalifoliumby comparing the relative ureide abundance (RUA) of stem extractsof this plant with simultaneous estimates of BNF obtained using the15N isotope dilution technique. The firstexperiment was performed in pots of soil, taken from a grazing study, amended with15N-labelled organic matter atfour different application rates. The ureide concentration in the stem extracts reflected the changes in BNF activityduring plant growth and the RUA was closely correlated with the proportion of N derived from BNF as determinedfrom the15N technique (r2 = 0.86 and 0.88 for inoculated and non-inoculated plants, respectively). The use of acalibration curve derived from a previous study where the same legume was fed increasing concentrations of15Nlabelled nitrate in sand/vermiculite culture, resulted in an over-estimation of the BNF contribution which may havebeen due to a significant uptake of ammonium from this acidic soil. The second experiment was performed infield plots and a good agreement was found between the estimates of BNF derived from using the ureide and15Ndilution techniques at two harvests six months apart. The uptake of soil N by theD. ovalifoliumand two foragegrasses (Brachiaria humidicolaandPanicum maximum) was estimated using anin situ soil core technique, and,while the uptake of N by the grasses was successfully estimated, this technique underestimated the N derived fromthe soil by the legume as determined by the ureide and15N dilution techniques.
Introduction
The use of the ratio of ureide-N to total-N in the bleed-ing sap extracted from soybean stems to estimate thecontribution of biological nitrogen fixation (BNF) tothe nutrition of this legume, was first investigated byMcClure et al. (1980) and tested and applied in bothgreenhouse and field experiments by Herridge andhis colleagues (Herridge, 1982, 1984; Herridge andPeoples, 1990; Herridge et al., 1988; 1990). This tech-
nique has also been successfully applied to quantifyBNF to other ureide producing grain legumes (Rer-kasem et al., 1988; Peoples et al., 1988; Peoples et al.,1989, Hansen et al., 1993) as well as for some woodylegumes (Herridge et al., 1996).
In an attempt to quantify the contribution of BNFto the ureide-producing forage legumesDesmodiumovalifolium and a Centrosemahybrid, Alves et al.(1999a) found it almost impossible to extract sap fromthe stems of these legumes even under greenhouse

178
conditions. These authors followed the suggestion ofPeoples et al. (1989), to use hot-water extracts ofstem tissues of these legumes, and found that the ratioof ureide-N:ureide + nitrate-N in these extracts waslinearly correlated (r2 = 0.82 and 0.90 forD. ovalifo-liumand theCentrosemahybrid, respectively) with theestimates of BNF contributions derived from the ap-plication of the15N dilution technique when the plantswere grown in sand/vermiculite culture.
The final aim of the project was to quantify thecontribution of BNF toD. ovalifoliumin pastures un-der grazing at a site in the South of the State of Bahia(NE Brazil - see Rezende et al., 1999). For this reasonin the present study it was decided to compare thiscalibration of the technique with another15N dilutionstudy conducted in pots of soil taken from the pasture,and with the results of a field experiment where BNFwas estimated by the same15N method and frominsitu estimates of mineralisation of soil N using thesoil core technique described by Raison et al. (1987).In the latter technique, the amount of plant-availablesoil mineral N (NH+4 + NO−3 ) is estimated from po-tassium chloride extracts of samples taken from intactsoil cores. The insertion of the soil cores cuts plantsroots and the mineral N which accumulates in thesecores over a period of 7 days, is considered to be theN produced from mineralisation of soil organic matterduring this time. Comparison of the quantity of min-eral N accumulated in these cores compared with thatfound in the soil at the end of this period in neighbour-ing areas where plant uptake had continued, yields anestimate of N uptake by the crop. The estimates de-rived from this procedure were found by Alves et al.(1993) to show a reasonable agreement to the total Nuptake of the grassPanicum maximum.In the case ofa N2-fixing legume the soil N uptake will be less thanthe total N in the crop, and the difference yields anestimate of the N derived from BNF.
Materials and methods
Experimental site
Both experiments were performed at the Animal Hus-bandry Station (16◦39′ S, 39◦30′ W) situated nearItabela in the South of Bahia State belonging to CE-PLAC, the cocoa research organisation. A full descrip-tion of this site, and the grazing experiment referredto above, was given by Rezende et al. (1999). Meanannual rainfall is 1300 mm, with no marked dry sea-son, and temperature ranges from 19 to 29◦C. The soil
is an Ultisol with a sandy surface layer (Typic Paleu-dult, fine loamy, kaolinitic, isohyperthermic) with thefollowing chemical characteristics (0–20 cm): pH 5.5,exchangeable cations (cmol kg−1) Ca, 2.2; Mg, 0.2;K, 0.1; Al 0.1; available P (Mehlich), 7.0 mg kg−1;total C (Walkley Black), 9.0 g kg−1; total N 0.82 gkg−1. The grazing experiment was originally installedin 1987 with 3 pastures (1.Brachiaria humidicolain monoculture, 2.Desmodium ovalifolium/B. hu-midicola, and 3. Pueraria phaseoloides(kudzu)/ B.humidicola) each with three different stocking rates(2, 3 and 4 animals ha−1), with 3 replicates arrangedin a completely randomised design. The pastureswere continuously grazed by three crossbred Brahmansteers (‘novilhos azebuados’) in each paddock of size1.5, 1.0 or 0.75 ha for the three stocking rates, respect-ively. The kudzu did not persist and since 1990 thisthird treatment has been pure grass, fertilised with 4×50 kg N ha−1 yr−1 as urea.
Pot experiment
Experimental design: Soil (0 – 15 cm) was takenfrom the paddocks of the above study, dried, sievedto 2 mm and mixed with dry15N-labelled straw atfour different rates; 3, 6, 9 and 12 g kg soil−1 anddistributed in pots at 4 kg pot−1. The labelled strawwas a mixture of stem and leaf material of soybeanandPanicum maximumobtained from earlier studies(henceforth referred to as labelled straw) and showeda C:N ratio of 24 and a15N enrichment of 0.175 atom% 15N excess.
The experiment consisted of a 4× 4× 5 factorialexperiment, with:a) 4 different levels of labelled straw (3, 6, 9 and 12 g
kg soil−1).b) 4 plant treatments (D. ovalifolium inoculated
with Bradyrhizobium, D. ovalifolium uninocu-lated,Panicum maximumcv. Centenário and rice(Oryza sativacv. Agulhinha).
c) Five harvests at 14-day intervals.The free-draining pots were arranged in complete ran-domised blocks in the open field with 4 replicates, andirrigated when rainfall was insufficient.
The sterile peat-basedBradyrhizobiuminoculantcontained a mixture of the strains BR 2212 and BR2214 (originally isolated fromD. ovalifolium) fromthe CNPAB collection, and contained approximately109 viable cells g−1.
The experiment was planted on 14th February1993 with 9 seeds pot−1 for the D. ovalifolium and

179
the rice, and 15 seeds pot−1 for theP. maximum.Afterone month of growth all plants were thinned to threeper pot and cut to a uniform height of 5 cm.
Harvests and analyses:The 5 harvests were taken at14 day intervals after the uniformization cut. TheD.ovalifolium plants were separated into leaves, stems+ petioles and roots (the latter for the evaluation ofnodulation only), and just the aerial tissue of the riceandP. maximumwas harvested. All plant material wasdried at 65◦C for>72 h, weighed and then ground to< 85µm with a Wiley mill.
For the analysis of nitrogenous solutes in thestem/petiole samples of theD. ovalifolium, sub-samples (500 mg) of the ground material were boiledfor 3 min in 25 ml of distilled water as described byPeoples et al. (1989). When cool the suspension wasfiltered into a 50 ml volumetric flask and the volumecompleted to 50 ml with washings and distilled water.These extracts were analysed for nitrate and ureidecontent using the flow injection technique of Alves etal. (1999b), and the colorimetric procedure of (Youngand Conway, 1942), respectively. All plant sampleswere analysed for total N content (Liao, 1981) andsubsequently for15N enrichment using a VG IsogasVG 903 mass spectrometer as described by Urquiagaet al. (1992).
Calculations: The relative ureide abundance (RUA)in the stem extracts was calculated from the equationof Herridge (1984):
RUA = (4× n mol ureide/
(n mol nitrate + 4× n mol ureide)) (1)
where n mol ureide/nitrate is the concentration ofureide/nitrate in the stem extracts expressed in n mol.
The contribution of biological nitrogen fixation tothe inoculated and non-inoculatedD. ovalifolium ateach harvest was calculated using two methods. Thefirst, conventional calculation was employed using theequation (Chalk, 1985):
%Ndfa =
(1– (atom %15N excess of legume/
atom %15N excess reference crop))× 100 (2)
where % Ndfa is the % of N derived from BNF.The other technique utilised was the ‘plant to
plant simulation’ technique described by Boddey etal. (1996). In this technique, curves of the accumu-lation of total N and excess15N (atom %15N excess
× total N× 100) were simulated from the data fromall 5 harvests using ‘S-shaped’ curves described by theequation (Witty, 1983; Hamilton et al., 1992):
Total N or15N excess = Nmax /
(1 + exp(-k.(DAC – T1/2)) (3)
where Nmax is the maximum N (or excess15N) accu-mulated by the plant, k is the exponential constant ofN (or excess15N) accumulation with time, DAC thenumber of days since the uniformization cut, and T1/2is the time (days) to half maximum N (or excess15N)accumulation. Using this equation it is possible to es-timate the quantity of total and15N-enriched N of thelegume and the rice andP. maximumat every instantthroughout the 70 day growth period. From this data,assuming thatP. maximumand the rice obtained Nonly from the15N-labelled soil, it is possible to estim-ate the15N enrichment of the soil on each day of thestudy [(total15N excess/total N) x 100]. Using this andthe same calculation to estimate the15N enrichmentof the legume on each day, it is possible to make twoseparate estimates of the proportion nitrogen derivedfrom the substrate (PNdfs) by the legume using thetwo different non-N2-fixing reference plants on eachday using the equation:
PNdfs = (recovery of excess15N by the legume/
atom %15N excess of substrate)× 100 (4)
Multiplying the value of PNdfs by the incrementof N accumulated by the legume on each day it ispossible to calculate the quantity of N derived fromthe soil on each day, and hence construct curves of thetotal N and N derived from the soil (and by difference,BNF) over the 70 day period of the study.
Full details of the analytical techniques and calcu-lation procedures used in this study are described inthe accompanying paper of Alves et al. (1999a).
Field experiment
Experimental layout: This experiment was installedin 5 separate blocks (21.6× 7.0 m) situated adja-cent (within 10 m) of the main grazing experimentdescribed above. Each block was divided into threeequal areas (7.2 m wide and 7.5 m in length) randomlyallocated, respectively, for sampling with soil cores(SC) for 15N labelling (EN) and sampling for15Nnatural abundance. Each area was divided into threesub-plots (2.4× 7.5 m) randomly allocated to threeplant species;D. ovalifolium(cv. Itabela),BrachiariahumidicolaandP. maximumcv. KK 16, the latter two
180
species to act as non-N2-fixing reference plants. Theseeds were planted on 23 May 1993 in four 7.5 m rowsspaced 20 cm apart, and after establishment 20 dayslater, the plants were cut to a uniform 5 cm height.Immediately after, in each EN sub-plot,15N-enrichedfertiliser was added to a central micro-plot of 3.0×2.0 m at a rate of 100 mg15N excess m−2 with a solu-tion of 15N-labelled ammonium sulphate (10.1 atom %15N) mixed with sucrose at a C:N ratio of 10:1 to facil-itate the slow release of the isotope and hence a morestable15N enrichment of the soil mineral N (Giller andWitty, 1987).
Harvests and analyses:Shoot tissue was harvestedfrom an area of 2.0× 1.0 m from the centre of the15N-labelled micro-plot, and separately from the remainderof the 3.0× 2.0 micro-plot, on 27 July (after 42 daysof regrowth). A further uniformization cut was madeon 25 November followed by a harvest made in thesame way on 26 January 1994. In the case of theD.ovalifolium the tissue was separated into leaves andstem + petioles.
Sub-samples of the stem + petioles of the legumeswere analysed for nitrate and ureide content, and allplant material was analysed for total N and that fromthe EN treatment for15N enrichment, as describedabove.
Soil core technique: Within each subplot of the SCarea two tubes (35 cm long× 6.5 cm diameter – area3.32× 10−3 m2) made from plastic water pipe, weredriven into the soil to a depth of 30 cm between plantsso as to cut the roots. One of these soil cores (A)was immediately removed and the soil from withinwas dried, mixed and sieved to 2 mm and roots andother debris removed. A 20 g sample was removedfor extraction with 100 ml of 2M KCl and after thesuspension had settled overnight, the clear supernatantwas analysed for nitrate and ammonium content usingthe flow injection techniques of Giné et al. (1980) andAlves et al. (1992), respectively. The second cylinder(B) was left in the soil for 7 days and then removed,and the soil treated as for cylinder A, to estimate thechange in concentration of mineral N in the soil dur-ing this 7-day period. Simultaneous with the removalof soil core B, a further core (C) was inserted andimmediately removed to assess the concentration ofsoil mineral N where active uptake by the roots hadcontinued in the 7-day period (Raison et al., 1987).
The whole procedure was then repeated, the soilcore C acting as the core A for the next 7 day period.
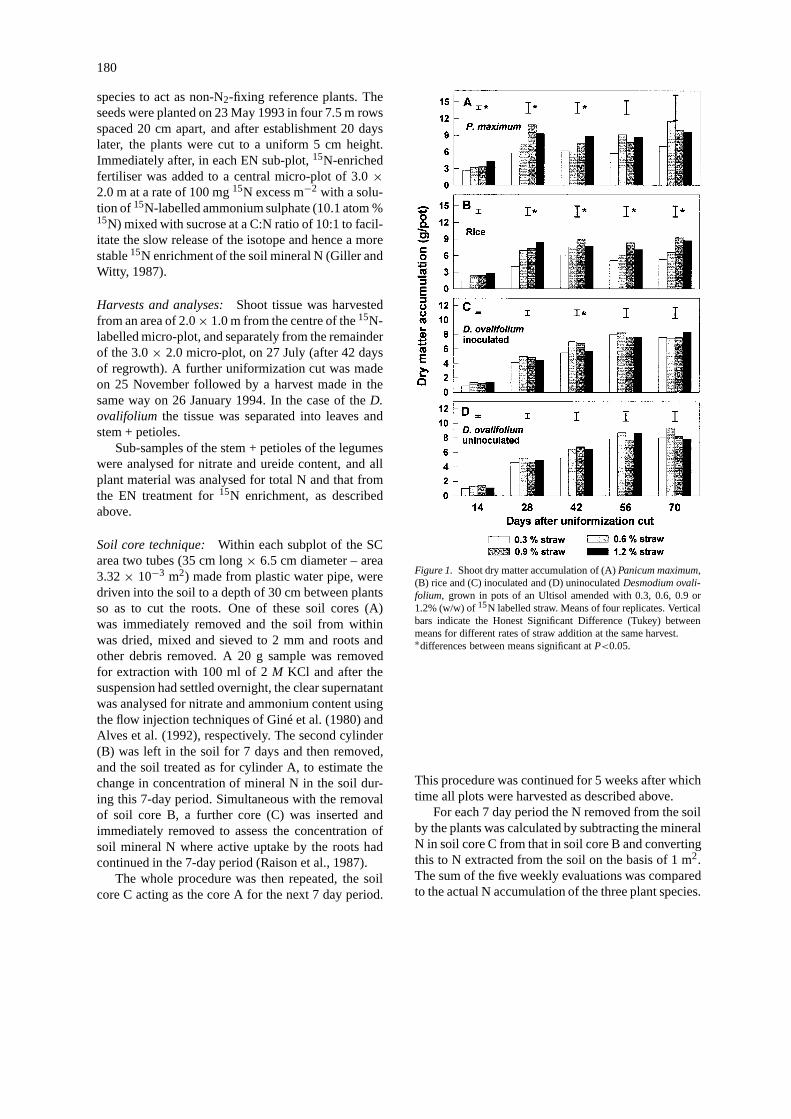
Figure 1. Shoot dry matter accumulation of (A)Panicum maximum,(B) rice and (C) inoculated and (D) uninoculatedDesmodium ovali-folium, grown in pots of an Ultisol amended with 0.3, 0.6, 0.9 or1.2% (w/w) of15N labelled straw. Means of four replicates. Verticalbars indicate the Honest Significant Difference (Tukey) betweenmeans for different rates of straw addition at the same harvest.∗differences between means significant atP<0.05.
This procedure was continued for 5 weeks after whichtime all plots were harvested as described above.
For each 7 day period the N removed from the soilby the plants was calculated by subtracting the mineralN in soil core C from that in soil core B and convertingthis to N extracted from the soil on the basis of 1 m2.The sum of the five weekly evaluations was comparedto the actual N accumulation of the three plant species.
181
Results and discussion
Pot experiment
Dry matter accumulation. The use of 4 differentlevels of addition of15N-labelled straw was intendedto give 4 different levels of N supply to the plants.The dry matter accumulation of theP. maximumandthe rice plants was generally increased (significantly– P<0.05 in some cases) by increased addition of thestraw (Figure 1A and 1B). In contrast, the dry matteraccumulation of theD. ovalifoliumin both inoculatedand uninoculated treatments was not significantly af-fected by the proportion of added straw suggesting thatin treatments where soil N availability was lower, in-puts of BNF compensated for this shortfall (Figure 1Cand 1D).
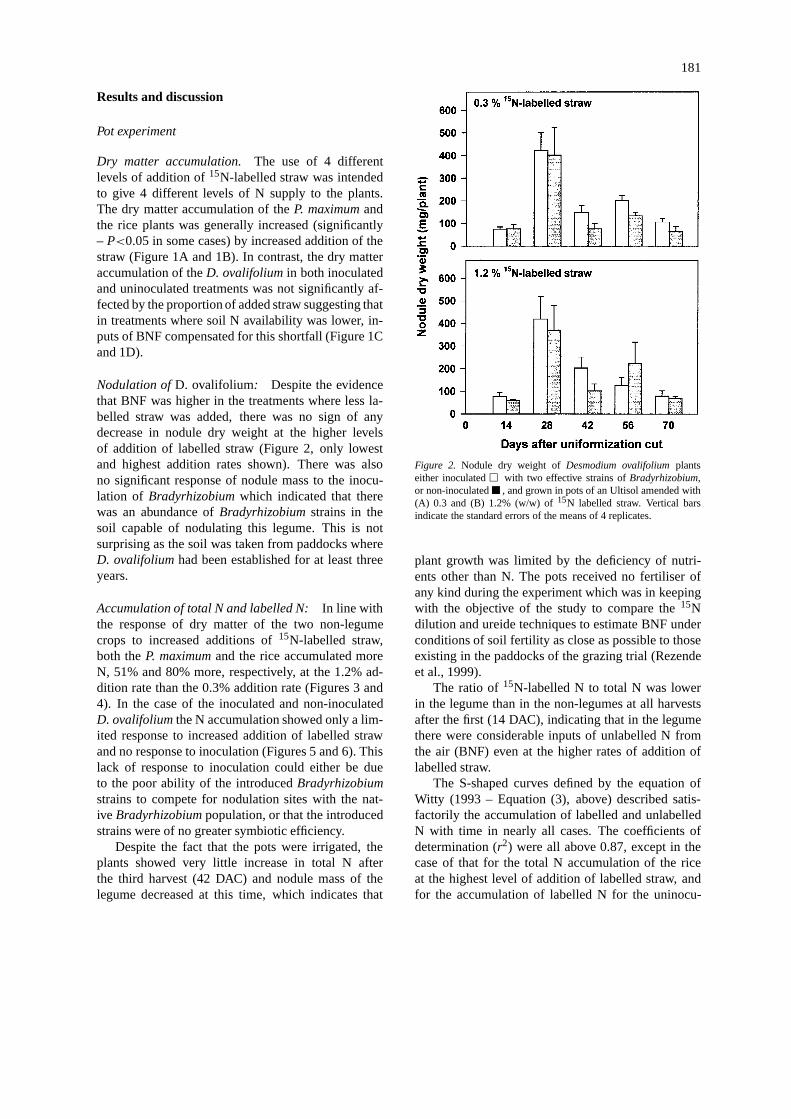
Nodulation ofD. ovalifolium: Despite the evidencethat BNF was higher in the treatments where less la-belled straw was added, there was no sign of anydecrease in nodule dry weight at the higher levelsof addition of labelled straw (Figure 2, only lowestand highest addition rates shown). There was alsono significant response of nodule mass to the inocu-lation of Bradyrhizobiumwhich indicated that therewas an abundance ofBradyrhizobiumstrains in thesoil capable of nodulating this legume. This is notsurprising as the soil was taken from paddocks whereD. ovalifoliumhad been established for at least threeyears.
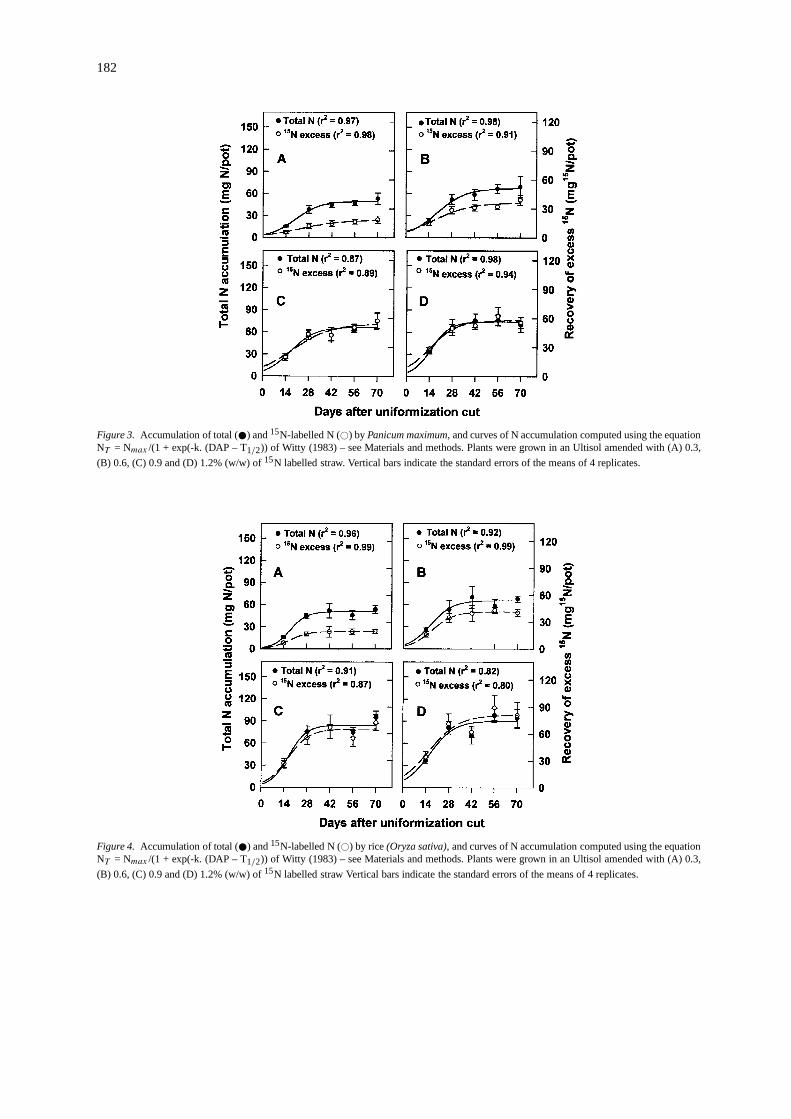
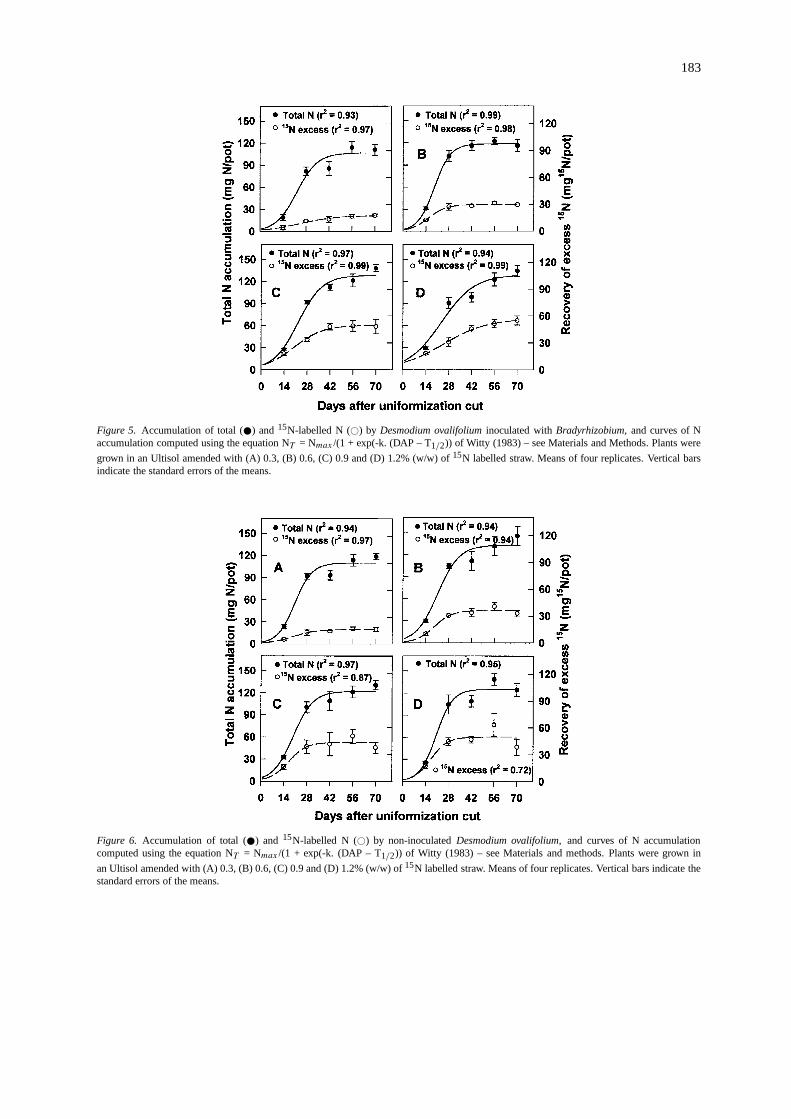
Accumulation of total N and labelled N:In line withthe response of dry matter of the two non-legumecrops to increased additions of15N-labelled straw,both theP. maximumand the rice accumulated moreN, 51% and 80% more, respectively, at the 1.2% ad-dition rate than the 0.3% addition rate (Figures 3 and4). In the case of the inoculated and non-inoculatedD. ovalifoliumthe N accumulation showed only a lim-ited response to increased addition of labelled strawand no response to inoculation (Figures 5 and 6). Thislack of response to inoculation could either be dueto the poor ability of the introducedBradyrhizobiumstrains to compete for nodulation sites with the nat-ive Bradyrhizobiumpopulation, or that the introducedstrains were of no greater symbiotic efficiency.
Despite the fact that the pots were irrigated, theplants showed very little increase in total N afterthe third harvest (42 DAC) and nodule mass of thelegume decreased at this time, which indicates that
Figure 2. Nodule dry weight ofDesmodium ovalifoliumplantseither inoculatedE with two effective strains ofBradyrhizobium,or non-inoculatedB , and grown in pots of an Ultisol amended with(A) 0.3 and (B) 1.2% (w/w) of15N labelled straw. Vertical barsindicate the standard errors of the means of 4 replicates.
plant growth was limited by the deficiency of nutri-ents other than N. The pots received no fertiliser ofany kind during the experiment which was in keepingwith the objective of the study to compare the15Ndilution and ureide techniques to estimate BNF underconditions of soil fertility as close as possible to thoseexisting in the paddocks of the grazing trial (Rezendeet al., 1999).
The ratio of15N-labelled N to total N was lowerin the legume than in the non-legumes at all harvestsafter the first (14 DAC), indicating that in the legumethere were considerable inputs of unlabelled N fromthe air (BNF) even at the higher rates of addition oflabelled straw.
The S-shaped curves defined by the equation ofWitty (1993 – Equation (3), above) described satis-factorily the accumulation of labelled and unlabelledN with time in nearly all cases. The coefficients ofdetermination (r2) were all above 0.87, except in thecase of that for the total N accumulation of the riceat the highest level of addition of labelled straw, andfor the accumulation of labelled N for the uninocu-
182
Figure 3. Accumulation of total ( ) and15N-labelled N (#) by Panicum maximum,and curves of N accumulation computed using the equationNT = Nmax /(1 + exp(-k. (DAP – T1/2)) of Witty (1983) – see Materials and methods. Plants were grown in an Ultisol amended with (A) 0.3,
(B) 0.6, (C) 0.9 and (D) 1.2% (w/w) of15N labelled straw. Vertical bars indicate the standard errors of the means of 4 replicates.
Figure 4. Accumulation of total ( ) and15N-labelled N (#) by rice(Oryza sativa),and curves of N accumulation computed using the equationNT = Nmax /(1 + exp(-k. (DAP – T1/2)) of Witty (1983) – see Materials and methods. Plants were grown in an Ultisol amended with (A) 0.3,
(B) 0.6, (C) 0.9 and (D) 1.2% (w/w) of15N labelled straw Vertical bars indicate the standard errors of the means of 4 replicates.
183
Figure 5. Accumulation of total ( ) and15N-labelled N (#) by Desmodium ovalifoliuminoculated withBradyrhizobium,and curves of Naccumulation computed using the equation NT = Nmax /(1 + exp(-k. (DAP – T1/2)) of Witty (1983) – see Materials and Methods. Plants were
grown in an Ultisol amended with (A) 0.3, (B) 0.6, (C) 0.9 and (D) 1.2% (w/w) of15N labelled straw. Means of four replicates. Vertical barsindicate the standard errors of the means.
Figure 6. Accumulation of total ( ) and 15N-labelled N (#) by non-inoculatedDesmodium ovalifolium,and curves of N accumulationcomputed using the equation NT = Nmax /(1 + exp(-k. (DAP – T1/2)) of Witty (1983) – see Materials and methods. Plants were grown in
an Ultisol amended with (A) 0.3, (B) 0.6, (C) 0.9 and (D) 1.2% (w/w) of15N labelled straw. Means of four replicates. Vertical bars indicate thestandard errors of the means.

184
latedD. ovalifoliumand the rice in this same treatment(Figures 3 to 6). These values are within the range,or higher than, those reported in other studies whichutilised this technique (e.g. Witty, 1983; Hamilton etal., 1992; Smith et al., 1992, Boddey et al., 1996),suggesting that this type of equation can generallybe used successfully to describe N accumulation inboth ‘N2-fixing’ and reference in both pot and fieldexperiments.
‘Plant to plant simulation’ of N accumulation:Totest the model of plant to plant simulation (see Ma-terials and methods), the total N accumulated by theP.maximumwas estimated from the data for the dailyaccumulation of total N and15N-labelled N by therice and the15N enrichment of theP. maximum, andvice-versa. When the daily accumulation of labelledand total N of theP. maximumwere utilised to es-timate the15N enrichment of the soil mineral N oneach day, the estimate of the N accumulated from thesoil by the rice was shown to be very close to theactual N accumulation of this crop (data not shown).The estimates of the N accumulated from the soil bythe P. maximum, from the simulated daily accumu-lation of labelled and unlabelled N of the rice, theactual N accumulation of this grass was underestim-ated by between 20 (0.9% added labelled straw) and33% (0.6% added straw) (Figure 7). When this sim-ulation technique was applied in the same manner byBoddey et al. (1996) for non-nodulating soybean andwheat in a field experiment, there were much greaterdiscrepancies between the simulated and actual N ac-cumulation of these two non-N2-fixing plants. Theseauthors suggested that this may have been due to thewheat obtaining more N from lower layers in the soilthan the non-nod soybean, where the surface-appliedlabelled N was in lower concentration. In this presentstudy this could not have been the reason as the15N-labelled straw was mixed uniformly with all soil in thepots. Unless there was a significant ‘associative’ BNFinput to the rice under dryland conditions which, incontrast to wetland conditions (Boddey et al., 1995a;Shrestha and Ladha, 1996), has never been reported,the difference between the simulated estimate and theactual N uptake for theP. maximummay have been dueto differential stimulation of N mineralisation by thetwo species (Huntjens, 1971), or perhaps just due tovariability in the sampling and analyses of the variousparameters measured.
When the uptake of N from the soil by the inocu-latedD. ovalifoliumwas calculated from the simulated
daily accumulation of labelled and total N of theP.maximumand rice (Figure 8A and 8C, respectively, forthe addition of 0.3% labelled straw, and Figure 9A and9C for the addition of 1.2%), there was a very largeunderestimate of total N accumulation by the legume.This is exactly what would be expected if the legumewas obtaining a significant contribution of BNF, andfrom these data the estimates of the daily BNF con-tributions were made (Figures 8B, 8D, 9B and 9D).The same simulations of the accumulation of soil Nand N from BNF for the uninoculated plants (data notshown) were consistent with the data on dry matter andN accumulation and nodule dry weight, in that therewas no marked increase in the BNF input when theplants were inoculated withBradyrhizobium.
The estimates of the overall contribution of BNF atthe final harvest (70 DAC) derived from the use of thesimulation technique with the two different referenceplants were generally in good agreement (Table 1). Atthe final 3 harvests (42, 56 and 70 DAC) the estim-ates of % Ndfa derived from the use of the ‘plant toplant simulation’ technique were almost constant, orincreased slightly with time, which is consistent withthe observation that the N accumulation of the legumedid not increase significantly after 42 DAC (Figures5 and 6) and nodule mass decreased after 28 DAC(Figure 2). In contrast, the estimates derived from thetraditional application of the isotope dilution equation(Equation (2)) at each individual harvest suggestedboth increases and decreases in %Ndfa from 42 to 70DAC. If legume total N did not decrease during thisperiod, then decreases in %Ndfa would suggest thatplants somehow preferentially lost fixed N, for whichthere appears to be no logical explanation. The appar-ently more satisfactory estimates of BNF derived fromthe use of the simulation technique is best explainedby the fact that they are derived from total N and15Nenrichment data from all harvests together, and arebased on a model of plant N accumulation which wasshown to be a good approximation to reality in boththis experiment (Figures 3 to 6) and several earlierstudies (Witty, 1983; Hamilton et al., 1992; Smith etal., 1992, Boddey et al., 1996, Alves et al., 1999a).The use of the simulation technique also clearly re-vealed a peak in N2-fixing activity at 28 DAC (Figures8 and 9) which coincided precisely with the peak innodule mass at this same date (Figure 2).
The total recovery of labelled N at the final harvestby theP. maximumwas similar to that of the inoculatedand non-inoculatedD. ovalifolium, and considerablylower than that of the rice especially at the two highest
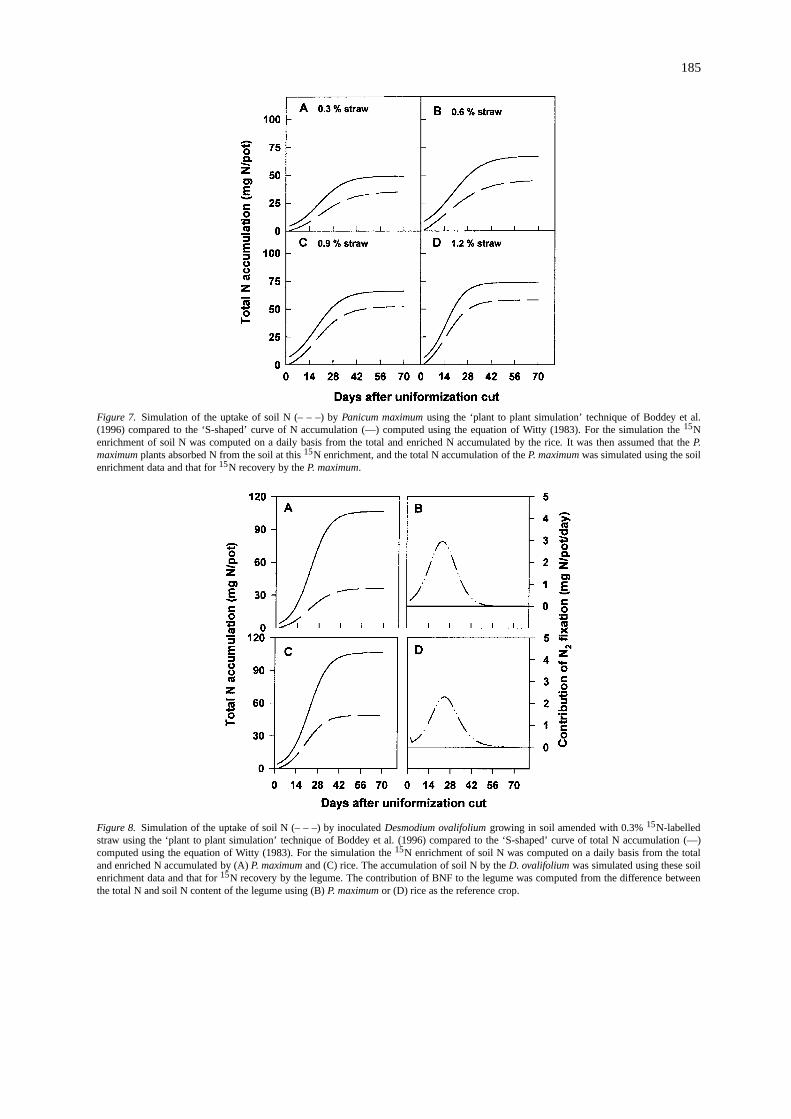
185
Figure 7. Simulation of the uptake of soil N (– – –) byPanicum maximumusing the ‘plant to plant simulation’ technique of Boddey et al.(1996) compared to the ‘S-shaped’ curve of N accumulation (—) computed using the equation of Witty (1983). For the simulation the15Nenrichment of soil N was computed on a daily basis from the total and enriched N accumulated by the rice. It was then assumed that theP.maximumplants absorbed N from the soil at this15N enrichment, and the total N accumulation of theP. maximumwas simulated using the soilenrichment data and that for15N recovery by theP. maximum.
Figure 8. Simulation of the uptake of soil N (– – –) by inoculatedDesmodium ovalifoliumgrowing in soil amended with 0.3%15N-labelledstraw using the ‘plant to plant simulation’ technique of Boddey et al. (1996) compared to the ‘S-shaped’ curve of total N accumulation (—)computed using the equation of Witty (1983). For the simulation the15N enrichment of soil N was computed on a daily basis from the totaland enriched N accumulated by (A)P. maximumand (C) rice. The accumulation of soil N by theD. ovalifoliumwas simulated using these soilenrichment data and that for15N recovery by the legume. The contribution of BNF to the legume was computed from the difference betweenthe total N and soil N content of the legume using (B)P. maximumor (D) rice as the reference crop.
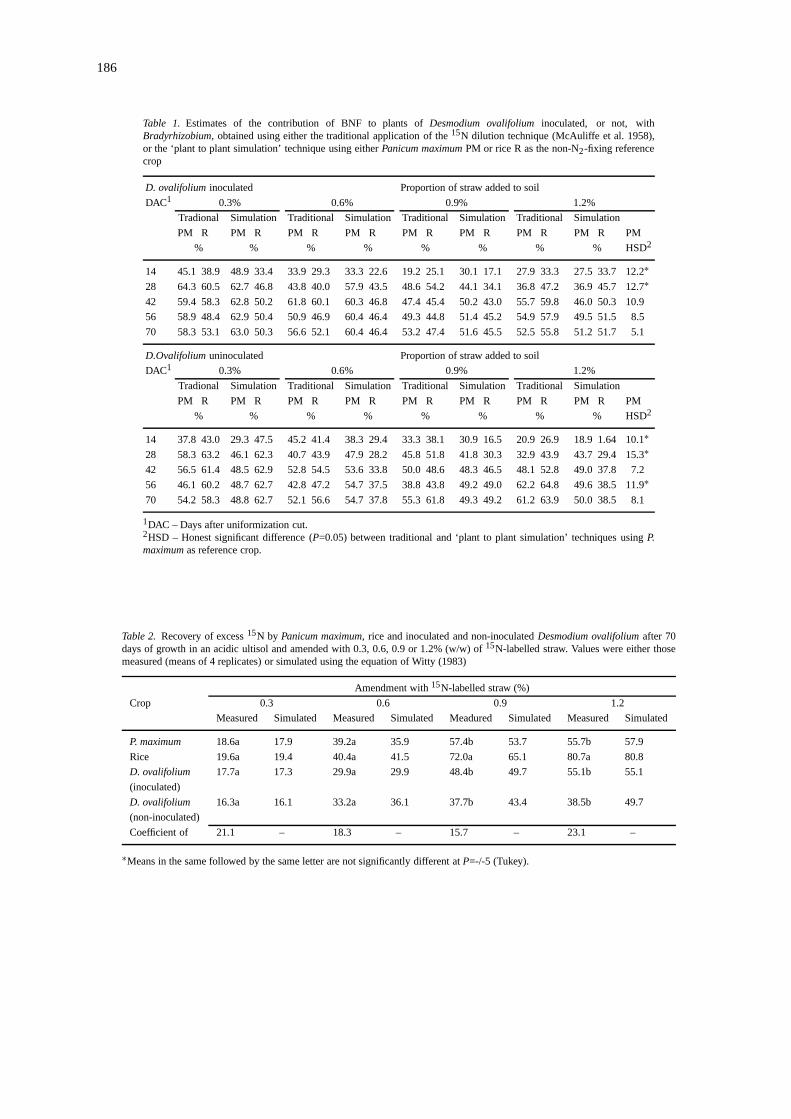
186
Table 1. Estimates of the contribution of BNF to plants ofDesmodium ovalifoliuminoculated, or not, withBradyrhizobium, obtained using either the traditional application of the15N dilution technique (McAuliffe et al. 1958),or the ‘plant to plant simulation’ technique using eitherPanicum maximumPM or rice R as the non-N2-fixing referencecrop
D. ovalifolium inoculated Proportion of straw added to soil
DAC1 0.3% 0.6% 0.9% 1.2%
Tradional Simulation Traditional Simulation Traditional Simulation Traditional Simulation
PM R PM R PM R PM R PM R PM R PM R PM R PM
% % % % % % % % HSD2
14 45.1 38.9 48.9 33.4 33.9 29.3 33.3 22.6 19.2 25.1 30.1 17.1 27.9 33.3 27.5 33.7 12.2∗28 64.3 60.5 62.7 46.8 43.8 40.0 57.9 43.5 48.6 54.2 44.1 34.1 36.8 47.2 36.9 45.7 12.7∗42 59.4 58.3 62.8 50.2 61.8 60.1 60.3 46.8 47.4 45.4 50.2 43.0 55.7 59.8 46.0 50.3 10.9
56 58.9 48.4 62.9 50.4 50.9 46.9 60.4 46.4 49.3 44.8 51.4 45.2 54.9 57.9 49.5 51.5 8.5
70 58.3 53.1 63.0 50.3 56.6 52.1 60.4 46.4 53.2 47.4 51.6 45.5 52.5 55.8 51.2 51.7 5.1
D.Ovalifoliumuninoculated Proportion of straw added to soil
DAC1 0.3% 0.6% 0.9% 1.2%
Tradional Simulation Traditional Simulation Traditional Simulation Traditional Simulation
PM R PM R PM R PM R PM R PM R PM R PM R PM
% % % % % % % % HSD2
14 37.8 43.0 29.3 47.5 45.2 41.4 38.3 29.4 33.3 38.1 30.9 16.5 20.9 26.9 18.9 1.64 10.1∗28 58.3 63.2 46.1 62.3 40.7 43.9 47.9 28.2 45.8 51.8 41.8 30.3 32.9 43.9 43.7 29.4 15.3∗42 56.5 61.4 48.5 62.9 52.8 54.5 53.6 33.8 50.0 48.6 48.3 46.5 48.1 52.8 49.0 37.8 7.2
56 46.1 60.2 48.7 62.7 42.8 47.2 54.7 37.5 38.8 43.8 49.2 49.0 62.2 64.8 49.6 38.5 11.9∗70 54.2 58.3 48.8 62.7 52.1 56.6 54.7 37.8 55.3 61.8 49.3 49.2 61.2 63.9 50.0 38.5 8.1
1DAC – Days after uniformization cut.2HSD – Honest significant difference (P=0.05) between traditional and ‘plant to plant simulation’ techniques usingP.maximumas reference crop.
Table 2. Recovery of excess15N by Panicum maximum, rice and inoculated and non-inoculatedDesmodium ovalifoliumafter 70days of growth in an acidic ultisol and amended with 0.3, 0.6, 0.9 or 1.2% (w/w) of15N-labelled straw. Values were either thosemeasured (means of 4 replicates) or simulated using the equation of Witty (1983)
Amendment with15N-labelled straw (%)
Crop 0.3 0.6 0.9 1.2
Measured Simulated Measured Simulated Meadured Simulated Measured Simulated
P. maximum 18.6a 17.9 39.2a 35.9 57.4b 53.7 55.7b 57.9
Rice 19.6a 19.4 40.4a 41.5 72.0a 65.1 80.7a 80.8
D. ovalifolium 17.7a 17.3 29.9a 29.9 48.4b 49.7 55.1b 55.1
(inoculated)
D. ovalifolium 16.3a 16.1 33.2a 36.1 37.7b 43.4 38.5b 49.7
(non-inoculated)
Coefficient of 21.1 – 18.3 – 15.7 – 23.1 –
∗Means in the same followed by the same letter are not significantly different atP=-/-5 (Tukey).
187
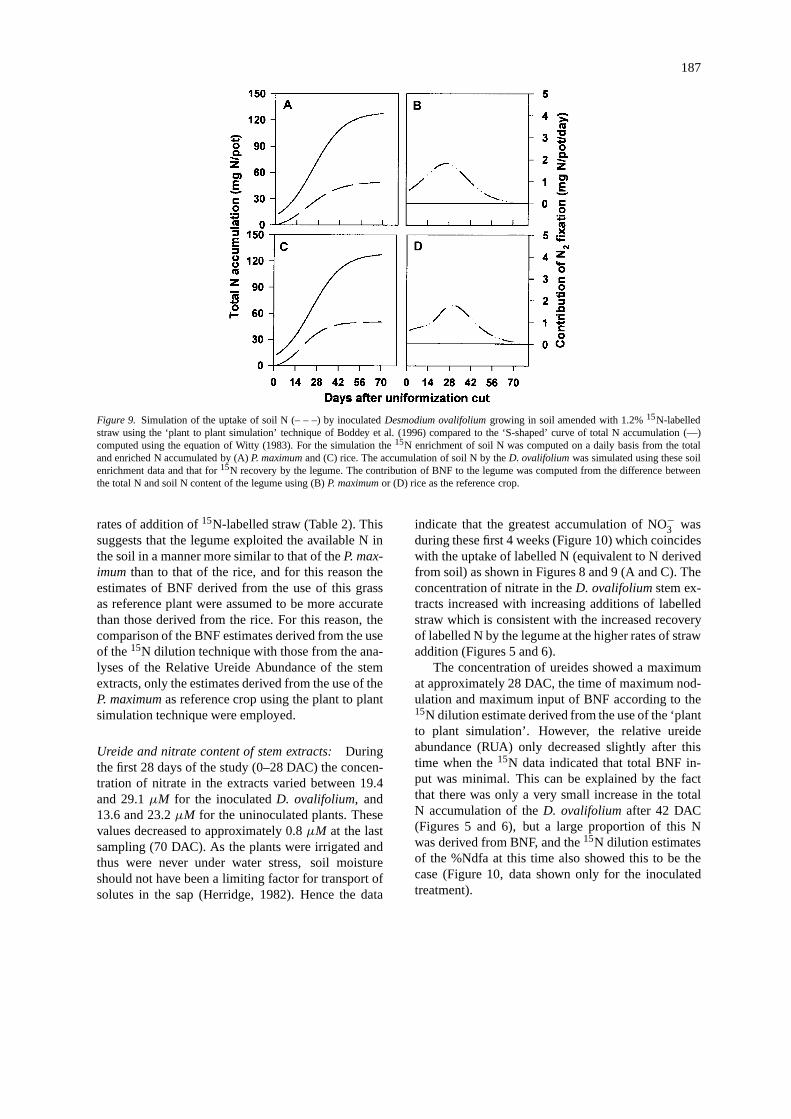
Figure 9. Simulation of the uptake of soil N (– – –) by inoculatedDesmodium ovalifoliumgrowing in soil amended with 1.2%15N-labelledstraw using the ‘plant to plant simulation’ technique of Boddey et al. (1996) compared to the ‘S-shaped’ curve of total N accumulation (—)computed using the equation of Witty (1983). For the simulation the15N enrichment of soil N was computed on a daily basis from the totaland enriched N accumulated by (A)P. maximumand (C) rice. The accumulation of soil N by theD. ovalifoliumwas simulated using these soilenrichment data and that for15N recovery by the legume. The contribution of BNF to the legume was computed from the difference betweenthe total N and soil N content of the legume using (B)P. maximumor (D) rice as the reference crop.
rates of addition of15N-labelled straw (Table 2). Thissuggests that the legume exploited the available N inthe soil in a manner more similar to that of theP. max-imum than to that of the rice, and for this reason theestimates of BNF derived from the use of this grassas reference plant were assumed to be more accuratethan those derived from the rice. For this reason, thecomparison of the BNF estimates derived from the useof the15N dilution technique with those from the ana-lyses of the Relative Ureide Abundance of the stemextracts, only the estimates derived from the use of theP. maximumas reference crop using the plant to plantsimulation technique were employed.
Ureide and nitrate content of stem extracts:Duringthe first 28 days of the study (0–28 DAC) the concen-tration of nitrate in the extracts varied between 19.4and 29.1µM for the inoculatedD. ovalifolium, and13.6 and 23.2µM for the uninoculated plants. Thesevalues decreased to approximately 0.8µM at the lastsampling (70 DAC). As the plants were irrigated andthus were never under water stress, soil moistureshould not have been a limiting factor for transport ofsolutes in the sap (Herridge, 1982). Hence the data
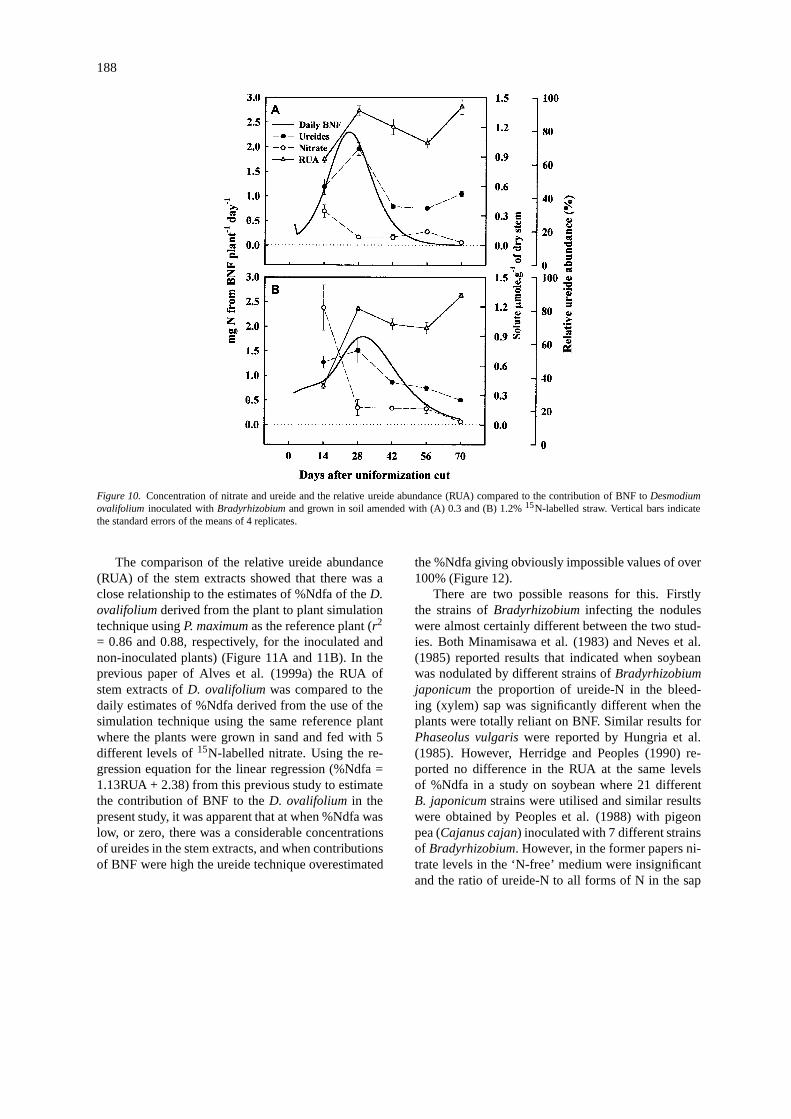
indicate that the greatest accumulation of NO−3 wasduring these first 4 weeks (Figure 10) which coincideswith the uptake of labelled N (equivalent to N derivedfrom soil) as shown in Figures 8 and 9 (A and C). Theconcentration of nitrate in theD. ovalifoliumstem ex-tracts increased with increasing additions of labelledstraw which is consistent with the increased recoveryof labelled N by the legume at the higher rates of strawaddition (Figures 5 and 6).
The concentration of ureides showed a maximumat approximately 28 DAC, the time of maximum nod-ulation and maximum input of BNF according to the15N dilution estimate derived from the use of the ‘plantto plant simulation’. However, the relative ureideabundance (RUA) only decreased slightly after thistime when the15N data indicated that total BNF in-put was minimal. This can be explained by the factthat there was only a very small increase in the totalN accumulation of theD. ovalifolium after 42 DAC(Figures 5 and 6), but a large proportion of this Nwas derived from BNF, and the15N dilution estimatesof the %Ndfa at this time also showed this to be thecase (Figure 10, data shown only for the inoculatedtreatment).
188
Figure 10. Concentration of nitrate and ureide and the relative ureide abundance (RUA) compared to the contribution of BNF toDesmodiumovalifolium inoculated withBradyrhizobiumand grown in soil amended with (A) 0.3 and (B) 1.2%15N-labelled straw. Vertical bars indicatethe standard errors of the means of 4 replicates.
The comparison of the relative ureide abundance(RUA) of the stem extracts showed that there was aclose relationship to the estimates of %Ndfa of theD.ovalifoliumderived from the plant to plant simulationtechnique usingP. maximumas the reference plant (r2
= 0.86 and 0.88, respectively, for the inoculated andnon-inoculated plants) (Figure 11A and 11B). In theprevious paper of Alves et al. (1999a) the RUA ofstem extracts ofD. ovalifoliumwas compared to thedaily estimates of %Ndfa derived from the use of thesimulation technique using the same reference plantwhere the plants were grown in sand and fed with 5different levels of15N-labelled nitrate. Using the re-gression equation for the linear regression (%Ndfa =1.13RUA + 2.38) from this previous study to estimatethe contribution of BNF to theD. ovalifolium in thepresent study, it was apparent that at when %Ndfa waslow, or zero, there was a considerable concentrationsof ureides in the stem extracts, and when contributionsof BNF were high the ureide technique overestimated
the %Ndfa giving obviously impossible values of over100% (Figure 12).
There are two possible reasons for this. Firstlythe strains ofBradyrhizobiuminfecting the noduleswere almost certainly different between the two stud-ies. Both Minamisawa et al. (1983) and Neves et al.(1985) reported results that indicated when soybeanwas nodulated by different strains ofBradyrhizobiumjaponicumthe proportion of ureide-N in the bleed-ing (xylem) sap was significantly different when theplants were totally reliant on BNF. Similar results forPhaseolus vulgariswere reported by Hungria et al.(1985). However, Herridge and Peoples (1990) re-ported no difference in the RUA at the same levelsof %Ndfa in a study on soybean where 21 differentB. japonicumstrains were utilised and similar resultswere obtained by Peoples et al. (1988) with pigeonpea (Cajanus cajan) inoculated with 7 different strainsof Bradyrhizobium. However, in the former papers ni-trate levels in the ‘N-free’ medium were insignificantand the ratio of ureide-N to all forms of N in the sap
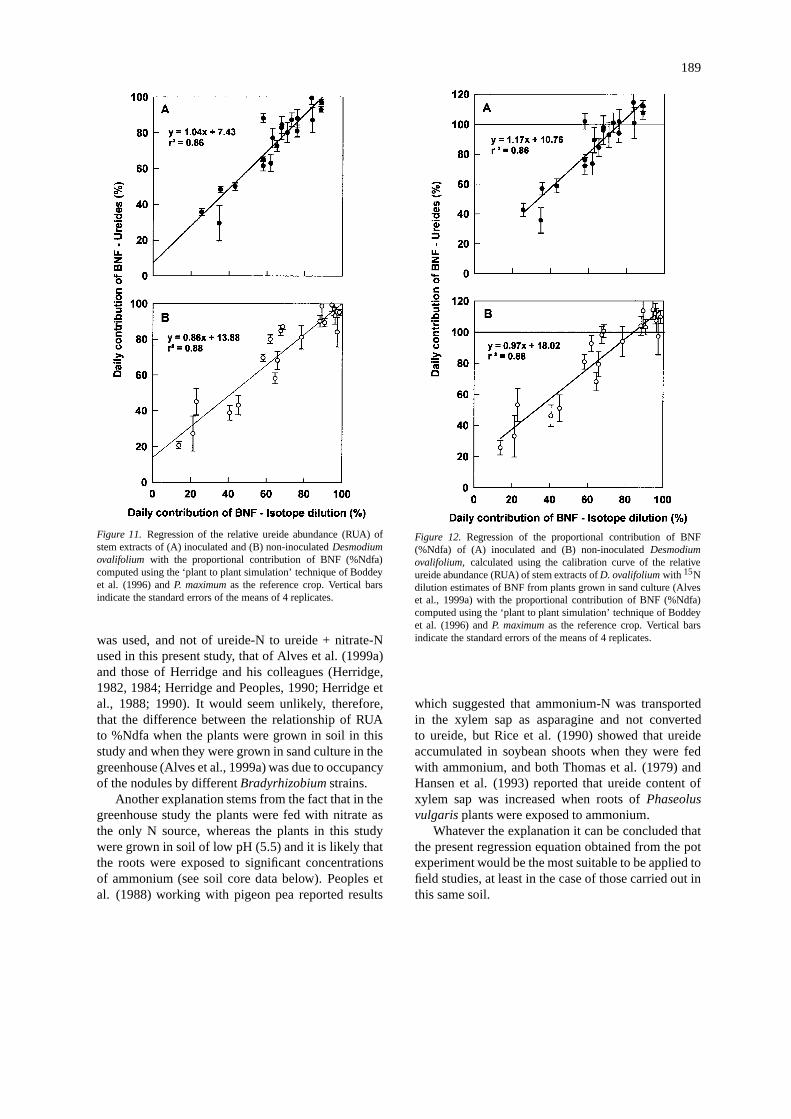
189
Figure 11. Regression of the relative ureide abundance (RUA) ofstem extracts of (A) inoculated and (B) non-inoculatedDesmodiumovalifolium with the proportional contribution of BNF (%Ndfa)computed using the ‘plant to plant simulation’ technique of Boddeyet al. (1996) andP. maximumas the reference crop. Vertical barsindicate the standard errors of the means of 4 replicates.
was used, and not of ureide-N to ureide + nitrate-Nused in this present study, that of Alves et al. (1999a)and those of Herridge and his colleagues (Herridge,1982, 1984; Herridge and Peoples, 1990; Herridge etal., 1988; 1990). It would seem unlikely, therefore,that the difference between the relationship of RUAto %Ndfa when the plants were grown in soil in thisstudy and when they were grown in sand culture in thegreenhouse (Alves et al., 1999a) was due to occupancyof the nodules by differentBradyrhizobiumstrains.
Another explanation stems from the fact that in thegreenhouse study the plants were fed with nitrate asthe only N source, whereas the plants in this studywere grown in soil of low pH (5.5) and it is likely thatthe roots were exposed to significant concentrationsof ammonium (see soil core data below). Peoples etal. (1988) working with pigeon pea reported results
Figure 12. Regression of the proportional contribution of BNF(%Ndfa) of (A) inoculated and (B) non-inoculatedDesmodiumovalifolium, calculated using the calibration curve of the relativeureide abundance (RUA) of stem extracts ofD. ovalifoliumwith 15Ndilution estimates of BNF from plants grown in sand culture (Alveset al., 1999a) with the proportional contribution of BNF (%Ndfa)computed using the ‘plant to plant simulation’ technique of Boddeyet al. (1996) andP. maximumas the reference crop. Vertical barsindicate the standard errors of the means of 4 replicates.
which suggested that ammonium-N was transportedin the xylem sap as asparagine and not convertedto ureide, but Rice et al. (1990) showed that ureideaccumulated in soybean shoots when they were fedwith ammonium, and both Thomas et al. (1979) andHansen et al. (1993) reported that ureide content ofxylem sap was increased when roots ofPhaseolusvulgarisplants were exposed to ammonium.
Whatever the explanation it can be concluded thatthe present regression equation obtained from the potexperiment would be the most suitable to be applied tofield studies, at least in the case of those carried out inthis same soil.
190
Table 3. Estimates of the proportional contribution of BNF toDesmodium ovalifoliumgrown in field plots estimatedusing the15N dilution and the relative ureide abundance techniques. Means of 5 replicates
15N enrichment Estimates of % Ndfa
Time of (Atom %15N excess)
sampling Reference Desmodium Relative ureide abundance
crop ovalifolium Isotope calibrated against15N dilution in:
dilution Sanda Soilb Soilb
inoculated non-inoculated
July 1993 0.269 (0.030)c 0.177 (0.012) 30.4 (9.9)c 49.2 (12.6) 32.9 (10.7) 32.1 (9.5)
0.307 (0.022)d 40.6 (7.2)d
January 1994 0.125 (0.019)c 0.059 (0.021) 53.0 (9.2)c 71.8 (14.1) 52.1 (12.3) 55.5 (11.3)
0.121 (0.029)d 52.6 (10.7)d
Values in parentheses correspond to the standard error of the means.aCalibration from plants fed15N-labelled nitrate (Alves et al. 1999a).bCalibration from plants grown in soil amended with15N-labelled straw (this study).cB. humidicolareference plant.dP. maximumreference plant.
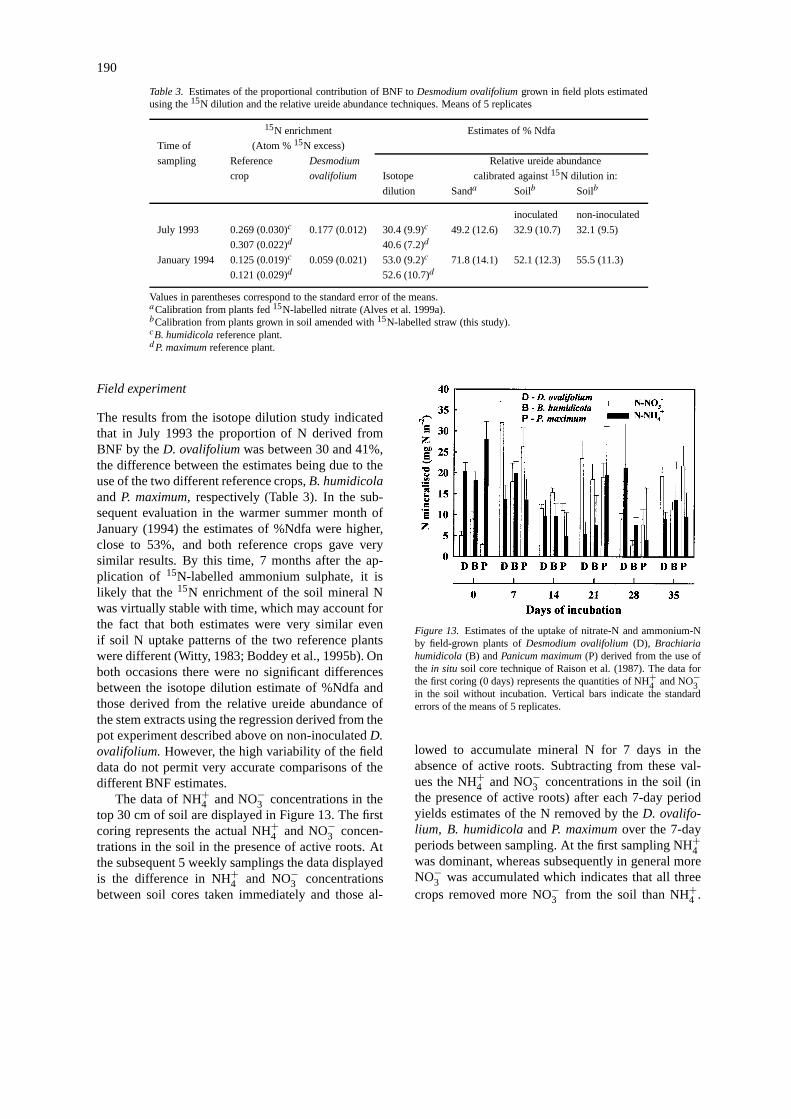
Field experiment
The results from the isotope dilution study indicatedthat in July 1993 the proportion of N derived fromBNF by theD. ovalifoliumwas between 30 and 41%,the difference between the estimates being due to theuse of the two different reference crops,B. humidicolaand P. maximum, respectively (Table 3). In the sub-sequent evaluation in the warmer summer month ofJanuary (1994) the estimates of %Ndfa were higher,close to 53%, and both reference crops gave verysimilar results. By this time, 7 months after the ap-plication of 15N-labelled ammonium sulphate, it islikely that the15N enrichment of the soil mineral Nwas virtually stable with time, which may account forthe fact that both estimates were very similar evenif soil N uptake patterns of the two reference plantswere different (Witty, 1983; Boddey et al., 1995b). Onboth occasions there were no significant differencesbetween the isotope dilution estimate of %Ndfa andthose derived from the relative ureide abundance ofthe stem extracts using the regression derived from thepot experiment described above on non-inoculatedD.ovalifolium.However, the high variability of the fielddata do not permit very accurate comparisons of thedifferent BNF estimates.
The data of NH+4 and NO−3 concentrations in thetop 30 cm of soil are displayed in Figure 13. The firstcoring represents the actual NH+4 and NO−3 concen-trations in the soil in the presence of active roots. Atthe subsequent 5 weekly samplings the data displayedis the difference in NH+4 and NO−3 concentrationsbetween soil cores taken immediately and those al-
Figure 13. Estimates of the uptake of nitrate-N and ammonium-Nby field-grown plants ofDesmodium ovalifolium(D), Brachiariahumidicola(B) andPanicum maximum(P) derived from the use ofthe in situ soil core technique of Raison et al. (1987). The data forthe first coring (0 days) represents the quantities of NH+4 and NO−3in the soil without incubation. Vertical bars indicate the standarderrors of the means of 5 replicates.
lowed to accumulate mineral N for 7 days in theabsence of active roots. Subtracting from these val-ues the NH+4 and NO−3 concentrations in the soil (inthe presence of active roots) after each 7-day periodyields estimates of the N removed by theD. ovalifo-lium, B. humidicolaandP. maximumover the 7-dayperiods between sampling. At the first sampling NH+4was dominant, whereas subsequently in general moreNO−3 was accumulated which indicates that all threecrops removed more NO−3 from the soil than NH+4 .

191
Figure 14. Actual accumulation of total N and estimates of the ac-cumulation of soil N derived from the use of thein situ soil coretechnique of Raison et al. (1987). Vertical bars indicate the standarderrors of the means of 5 replicates.
The individual weekly estimates of plant uptake werehighly variable and no consistent difference betweencrops in uptake of either NH+4 or NO−3 was observed.The data indicate that considerable quantities of NH+
4were absorbed by the plants, and this may explain whythe ureide content of the stem extracts ofD. ovalifo-lium was higher for a given %Ndfa, than for the sameplants fed only with NO−3 in sand/vermiculite culture(Alves et al., 1999a).
The estimates of the total N accumulated by thetwo grasses agreed well with the actual amount ofN accumulated over the 42 day period, but grosslyunderestimated the total N accumulated by theD.ovalifolium (Figure 14). This is to be expected if theextra N was derived from BNF. From these data the%Ndfa of the legume was estimated to be 58%, con-siderably above that derived from the15N dilution orureide techniques.
Conclusions
As in our previous greenhouse study (Alves et al.,1999a), the results of the pot experiment showed thatthere was a good correlation between the RUA of stemextracts ofD. ovalifolium and the %Ndfa estimatedusing the15N dilution technique. The use of the re-gression equation for these two parameters derivedfrom the greenhouse study where plants were fed15N-labelled nitrate to estimate %Ndfa in plants grown inpots of soil, showed that the contribution of BNF to thelegume was overestimated owing to the considerablecontent of ureide in the extracts when BNF was verylow or zero. Whether this is likely to occur generally
cannot be determined as so few studies to compare thecalibration of the ureide technique with actual con-tributions of BNF under field conditions have beenperformed. Herridge et al. (1990) compared the rel-ative ureide content of vacuum extracted sap and (atearly harvests) stem extracts of soybean grown at 5different field sites with estimates of %Ndfa derivedfrom use of the15N natural abundance technique. Theureide-based estimates of %Ndfa were computed froma previous calibration study performed in the green-house which used the15N dilution technique on plantsfed 15N labelled nitrate (Herridge and Peoples, 1990).There was a very satisfactory agreement of the twodifferent estimates although the ureide technique gavesomewhat lower estimates of %Ndfa when %Ndfa wasabove 40%. However, at the five different sites usedsoil NO−3 ranged from 70 to 280 kg N ha−1 at planting(0–120 cm depth) whereas NH+4 concentrations wereconsiderably lower in all cases (2 to 5 mg N kg soil−1
or approximately 8 to 20 kg N ha−1 in the 0–30 cmlayer).
Again in the field experiment the results confirmedthat the RUA of stem extracts could be used to es-timate the proportional contribution of BNF to thelegume, in that these estimates agreed well with thosederived from the use of the isotope dilution technique.This is the first application of the soil core techniquedeveloped by Raison et al. (1987) to estimate the con-tribution of BNF to a legume, and the results indicatedonly partial success. The estimates of the total N ac-cumulated by the non-N2-fixing grasses were close tothe actual as had been reported in an earlier study atEmbrapa-Agrobiologia (Alves et al., 1993), but thesoil N uptake of the legume was apparently seriouslyunderestimated. The technique involves a great dealof soil coring and analyses for NH+4 and NO−3 , but iffacilities for 15N isotope studies are not available andit is desired to quantify BNF inputs to non-ureide pro-ducing forage legumes (eg.Stylosanthesspp.,Arachispintoi) this method may be the best available.
Acknowledgments
The authors wish to express their gratitude to AltibertoM. Baêta and Roberto G. de Souza, for the tech-nical assistance with the nitrogen and15N analyses.The authors gratefully acknowledge the PhD fellow-ship from the Brazilian National Research Council(CNPq) for the senior author (B.J.R. Alves), and re-search fellowships for R.M. Boddey, R. Macedo, A.S.

192
Resende, R.M. Tarré and S. Urquiaga. This study wasfunded by the Fundação Banco do Brasil (Project No.10/04296-XI).
References
Alves BJR, Boddey RM & Urquiaga S (1992) A rapid and sensitiveflow injection technique for the analysis of ammonium in soilextracts. Commun Soil Sci Plant Anal 24: 277–284
Alves BJR, Urquiaga S, Cadisch G, Souto CM & Boddey RM(1993)In situ estimation of soil nitrogen mineralization. In: Mu-longuy K & Merckx R (eds) Dynamics of Soil Organic Matter inRelation to Sustainability of Tropical Agriculture, pp 173–180.Exeter, UK: Sayce Publishing
Alves BJR, Resende AS, Urquiaga S & Boddey RM (1999a) Bio-logical nitrogen fixation by two tropical forage legumes assessedfrom the relative ureide abundance of stem solutes:15N calibra-tion of the technique in sand culture. Nutr Cycl Agroecosyst 56:165–176 (this issue)
Alves BJR, Resende AS, Polidoro JC, Urquiaga S & Boddey RM(1999b) Rapid and sensitive determination of nitrate in plant tis-sue using flow injection analysis. Commun Soil Sci Plant Anal:in press
Boddey RM, Oliveira OC de, Urquiaga S, Reis VM, Olivares FL de,Baldani VLD & Dobereiner J (1995) Biological nitrogen fixationassociated with sugar cane and rice: Contributions and prospectsfor improvement. Plant Soil 174: 195–209
Boddey RM, Oliveira OC de, Alves BJR & Urquiaga S (1995b)Field application of the15N isotope dilution technique for thereliable quantification of plant-associated biological nitrogenfixation. Fert Res 42: 77–87.
Boddey RM, Müller SH & Alves BJR (1996) Estimation of thecontribution of biological N2 fixation to two Phaseolus vul-garis genotypes using simulation of plant nitrogen uptake from15N-labelled soil. Fert Res 45: 169–185
Chalk PM (1985) Estimation of N2 fixation by isotope dilution:An appraisal of techniques involving15N enrichment and theirapplication. Soil Biol Biochem 17: 389–410
Giller KE & Witty JF (1987) Immobilized15N-fertilizer sourcesimprove the accuracy of field estimates of N2-fixation by isotopedilution. Soil Biol Biochem 19: 459–463
Giné MF, Bergamin Filho H, Zagatto EAG & Reis BF 1980. Sim-ultaneous determination of nitrate and nitrite by flow injectionanalysis. Anal Chim Acta 114: 191–197
Hamilton SD, Smith CJ, Chalk PM & Hopmans P (1992) A modelbased on measurement of soil and plant15N enrichment to es-timate N2 fixation by soybean (Glycine maxL Merrill) grown inpots. Soil Biol Biochem 24: 71–78
Hansen AP, Rerkasem B, Lordkaew S & Martin P (1993) Xylem-solute technique to measure N2 fixation by Phaseolus vulgarisL.: Calibration and sources of error. Plant Soil 150: 223–231
Herridge DF (1982) Relative abundance of ureides and nitrate inplant tissues of soybean as a quantitative assay of nitrogenfixation. Plant Physiol 70: 1–6
Herridge DF (1984) Effects of nitrate and plant development on theabundance of nitrogenous solutes in root bleeding and vacuum-extracted exudates of soybean. Crop Sci 24: 173–179
Herridge DF & Peoples MB (1990) Ureide assay for measuring ni-trogen fixation by nodulated soybean calibrated by15N methods.Plant Physiol 93: 495–503
Herridge DF, O’Connell P & Donnelly K (1988) The xylem ureideassay of nitrogen fixation: Sampling procedures and sources oferror. J Exp Bot 39: 12–22
Herridge DF, Bergersen FJ & Peoples MB (1990) Measurement ofnitrogen fixation by soybean in the field using the ureide andnatural15N abundance methods. Plant Physiol 93: 708–716
Herridge DF, Palmer B, Nurhayati DP & Peoples MB (1996) Eval-uation of the xylem ureide method for measuring N2 fixation insix tree legume species. Soil Biol Biochem 28: 281–289
Hungria M, Neves MCP & Victoria RL (1985) Assimilação do ni-trogênio pelo feijoeiro. II. Absorção e translocação do N minerale do N2 fixado. R Bras Ci Solo 9: 201–209
Huntjens JLM (1971) The influence of living plants on mineraliza-tion and immobilization of nitrogen. Plant Soil 35: 77–94
Liao CFH (1981) Devarda’s alloy method for total nitrogen determ-ination. Soil Sci Soc Am J 45: 852–855
McAuliffe C, Chamblee DS, Uribe Arango H & Woodhouse WW(1958) Influence of inorganic nitrogen on nitrogen fixation bylegumes as revealed by N15. Agron J 50: 334–337
McClure PR, Israel DW & Volk RJ (1980) Evaluation of the relativeureide content of xylem sap as an indicator of N2 fixation insoybeans. Plant Physiol 66: 720–725
Minamisawa K, Arima Y & Kumazawa K (1983) Transport of fixednitrogen from soybean nodules inoculated with H2-uptake pos-itive and negativeRhizobium japonicumstrains. Soil Sci PlantNutr 29: 85–92
Neves MCP, Didonet AD, Duque FF & Döbereiner J (1985)Rhizobiumstrain effects on nitrogen transport and distributionin soybeans. J Exp Bot 36: 1179–1192
Peoples MB, Hebb DM, Gibson AH & Herridge DF (1988) De-velopment of the xylem ureide assay for the measurement ofnitrogen fixation by pigeonpea (Cajanus cajan(L.) Millsp.) JExp Bot 40: 535–542
Peoples MB, Faizah AW, Rerkasem B & Herridge DF (1989) Meth-ods for evaluating nitrogen fixation by nodulated legumes in thefield. ACIAR Monograph No. 11. 76 pp. Canberra, Australia:Australian Centre for International Agricultural Research
Raison RJ, Connell MJ & Khanna PK (1987) Methodology forstudying fluxes of soil mineral-Nin situ. Soil Biol Biochem 19:521–530
Rerkasem B, Rerkasem K, Peoples MB, Herridge DF & Ber-gersen FJ (1988) Measurement of N2 fixation in maize (ZeamaysL.)-ricebean (Vigna umbellata[Thunb.] Ohwi and Ohashi)intercrops. Plant Soil 108: 125–135
Rezende C de P, Cantarutti RB, Braga JM, Gomide JA, PereiraJM, Ferreira E, Tarré RM, Macedo R, Alves BJR, Urquiaga S,Cadisch G, Giller KE & Boddey RM (1999) Litter depositionand disappearance inBrachiaria pastures in the Atlantic forestregion of the South of Bahia, Brazil. Nutr Cycl Agroecosyst 54:99–112
Rice CF, Lukaszewsky KM, Walker S, Blevins DG, Winkler RG& Randall DD (1990) Changes in ureide synthesis, transportand assimilation following ammonium nitrate fertilization ofnodulated soybeans. J. Plant Nutr 13: 1539–1553
Shrestha RK & Ladha JK (1996) Genotypic variation in promotionof rice dinitrogen fixation as determined by nitrogen-15 dilution.Soil Sci Soc Am J 60: 1815–1821
Smith CJ, Chalk PM, Hamilton SD & Hopmans P (1992) Estimat-ing N2 fixation by field-grown lupins (Lupinus angustifoliusL.)using soil and plant15N enrichment. Biol Fert Soils 13: 235–241
Thomas RJ, Feller U & Erismann KH (1979) The effect of differentinorganic nitrogen sources and plant age on the composition ofbleeding sap ofPhaseolus vulgaris. New Phytol 82: 657–670

193
Urquiaga S, Cruz KHS & Boddey RM (1992) Contribution of ni-trogen fixation to sugar cane – Nitrogen-15 and nitrogen-balanceestimates. Soil Sci Soc Am J 56: 105–114
Witty JF (1983) Estimating N2-fixation in the field using15N-
labelled fertilizer: some problems and solutions. Soil Biol Bio-chem 15: 631–639
Young EG & Conway CF (1942) On the estimation of allantoin bythe Rimini-Schryver reaction. J Biol Chem 142: 839–853
Related Documents











