Essentials of Glycobiology Lecture 7 April 12, 2002 Ajit Varki Structure, biosynthesis and general biology of Glycosphingolipids

Essentials of Glycobiology Lecture 7 April 12, 2002 Ajit Varki
Dec 31, 2015
Essentials of Glycobiology Lecture 7 April 12, 2002 Ajit Varki. Structure, biosynthesis and general biology of Glycosphingolipids. CHONDROITIN SULFATE. HYALURONAN. GLYCOSAMINO- GLYCANS. HEPARAN SULFATE. N-LINKED CHAINS. O-LINKED CHAIN. GLYCOPHOSPHO- LIPID ANCHOR. - PowerPoint PPT Presentation
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Essentials of Glycobiology
Lecture 7
April 12, 2002
Ajit Varki
Structure, biosynthesis and general biology of Glycosphingolipids
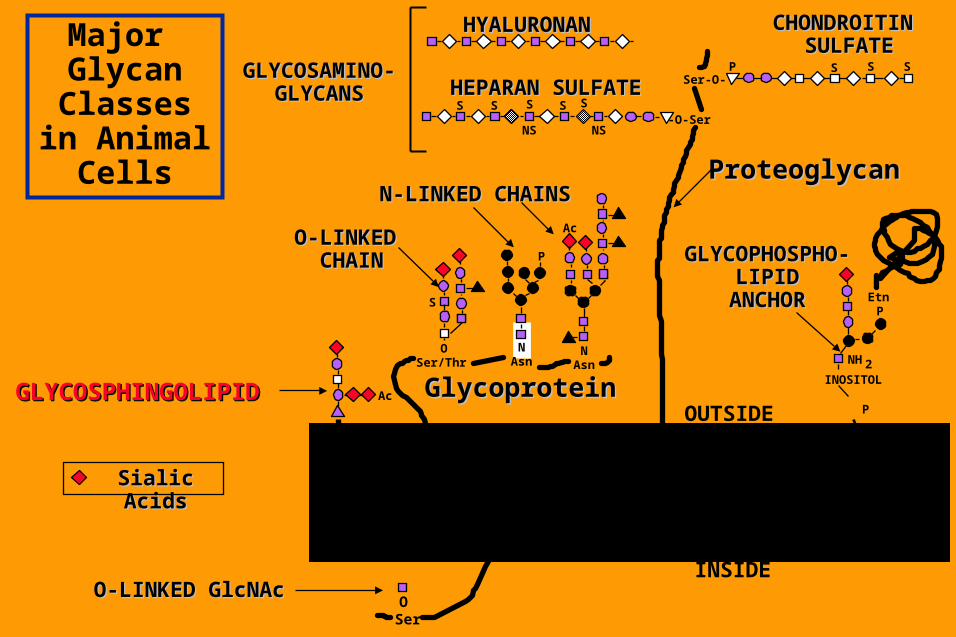
Major Glycan
Classes in Animal Cells
OSer
OSer/Thr
NAsn
Ser-O-
OUTSIDE
INSIDE
NAsn
S S S
-O-SerS SSS S
EtnP
INOSITOL
P
NH
Ac
P
NS NS
Ac
S
2
P
GlycoproteinGlycoprotein
ProteoglycanProteoglycan
GLYCOPHOSPHO-GLYCOPHOSPHO-LIPIDLIPID
ANCHORANCHOR
N-LINKED CHAINSN-LINKED CHAINS
O-LINKED O-LINKED CHAINCHAIN
HYALURONANHYALURONAN
GLYCOSAMINO-GLYCOSAMINO-GLYCANSGLYCANS HEPARAN SULFATEHEPARAN SULFATE
CHONDROITINCHONDROITIN SULFATESULFATE
Sialic AcidsSialic Acids
GLYCOSPHINGOLIPIDGLYCOSPHINGOLIPID
O-LINKED GlcNAcO-LINKED GlcNAc

Lecture Overview
Historical BackgroundDefining Structures and Major ClassesOther Nomenclature IssuesBiosynthesisOccurrence & Structural VariationsIsolation and purificationTrafficking, Turnover and Degradation

Lecture Overview (Continued)
Relationship to biosynthesis, turnover and signalling functions of other Sphingolipids
Antibodies against GlycosphingolipidsBiological RolesGenetic Disorders in GSL biosynthesisPerspectives and Future Directions

Minimal Defining Structure of a Glycosphingolipid
Glycan-O-Ceramide
Ceramide (Cer)
Sphingosine
Fatty Acyl group
Glycan
*
*Glucose (All animals) Galactose (?Vertebrates only) Mannose (Invertebrates) Inositol-P (fungi)
Find the Missing Double Bond!

Biosynthesis of Ceramide and
Glucosylceramide
Find the Missing Double Bond!

Nomenclature Issues
Glycosphingolipid (GSL) = Glycan + Sphingolipid
(named after the Egyptian Sphinx)
Glycosphingolipids often just referred to as “Glycolipids”.
“Ganglioside": a GSL one or more sialic acid residues
Example of nomenclature:
Neu5Ac3Gal3GalNAc4Gal4Glc1Cer = GM1a
in the Svennerholm nomenclature
OR
II4Neu5Ac-GgOSe4-Cer
in the official IUPAC-IUB designation

Isolation and purification of Glycosphingolipids Most glycosphingolipids obtained in good yield from cells and
tissues by sequential organic extractions of increasing polaritySome separation by polarity achieved by two-phase extractionsSubsequent fractionation away from other lipids in the extract
using:DEAE ion exchange chromatography silica gel thin layer and column separations
HPLC adaptations of these methods are useful
in obtaining complete separations. Principles of structural characterization
of these molecules presented elsewhere
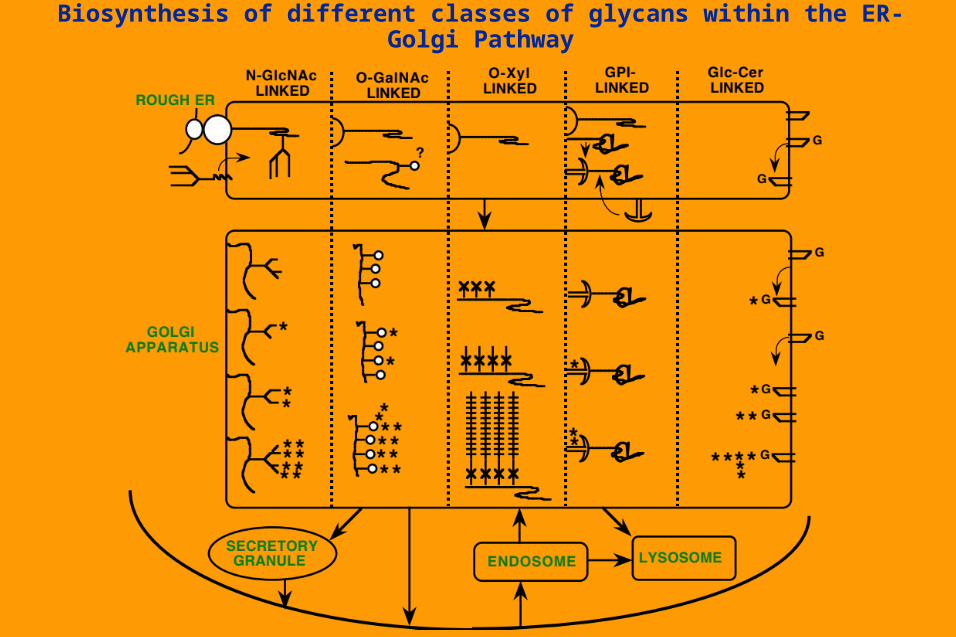
Biosynthesis of different classes of glycans within the ER-Golgi Pathway

Stepwise elongation of
Glucosylceramidegenerates unique Core structures

Major Classes of Glycosphingolipids
Series Designation Core Structure
Lacto (LcOSe4) Gal3GlcNAc3GalGal44GlcGlc1Ceramide1Ceramide
Lactoneo (LcnOSe4) Gal4GlcNAc3GalGal44GlcGlc1Ceramide1Ceramide
Globo (GbOSe4) GalNAc3Gal4GalGal44GlcGlc1Ceramide1Ceramide
Isoglobo (GbiOSe4) GalNAc3Gal3GalGal44GlcGlc1Ceramide1Ceramide
Ganglio (GgOSe4) Gal3GalNAc4GalGal44GlcGlc1Ceramide1Ceramide
Muco (MucOSe4) GalGal3GalGal44GlcGlc1Ceramide1Ceramide
Gala (GalOSe2) Gal4GalGal1Ceramide1Ceramide
Sulfatides 3-0-Sulfo-GalGal1Ceramide1Ceramide
Different Core structures generate unique shapes and are expressed
in a cell-type specific manner
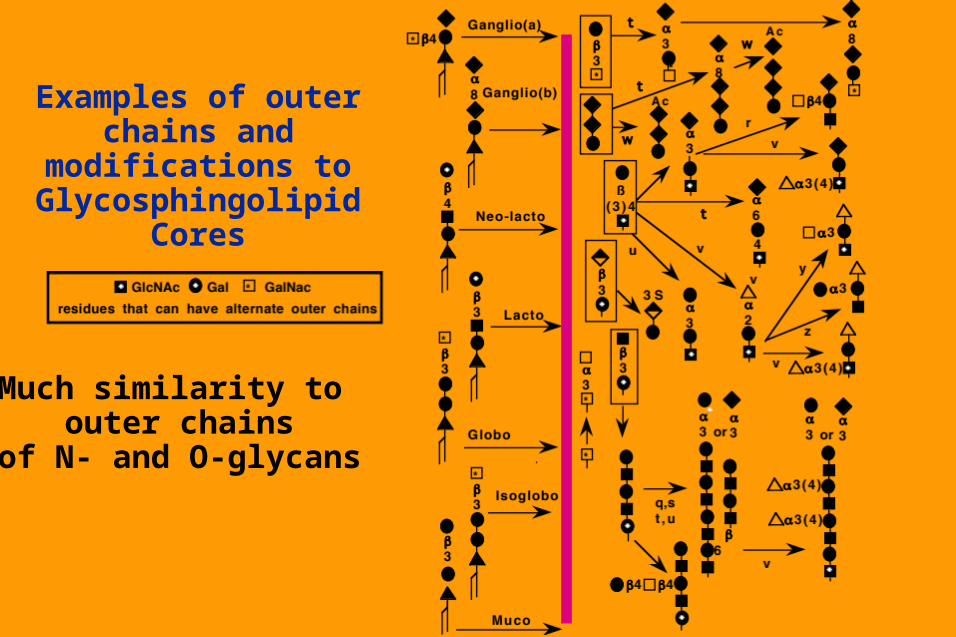
Examples of outer chains and modifications to
Glycosphingolipid Cores
Much similarity to outer chains
of N- and O-glycans
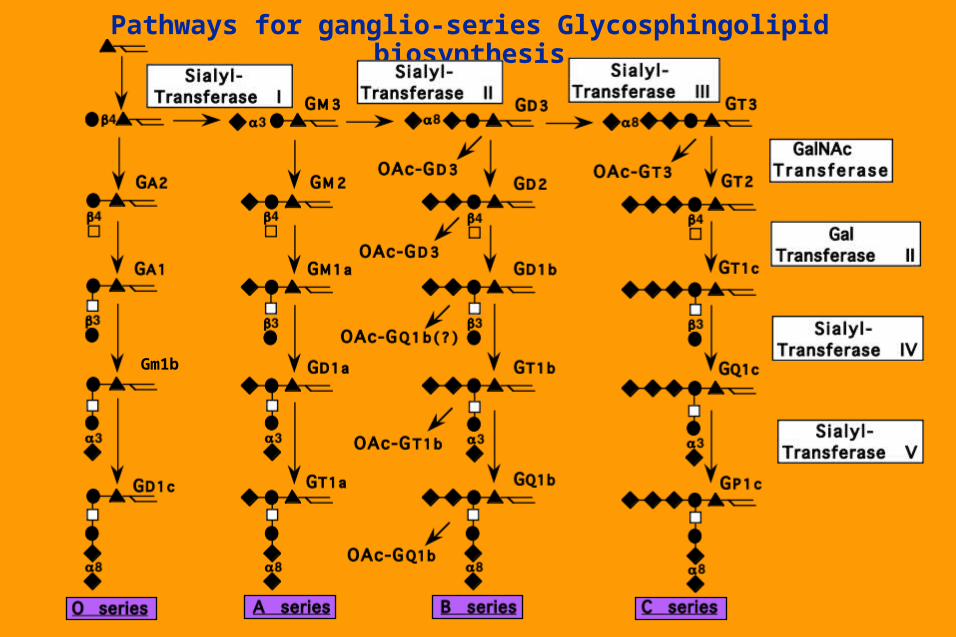
Pathways for ganglio-series Glycosphingolipid biosynthesis
Gm1b
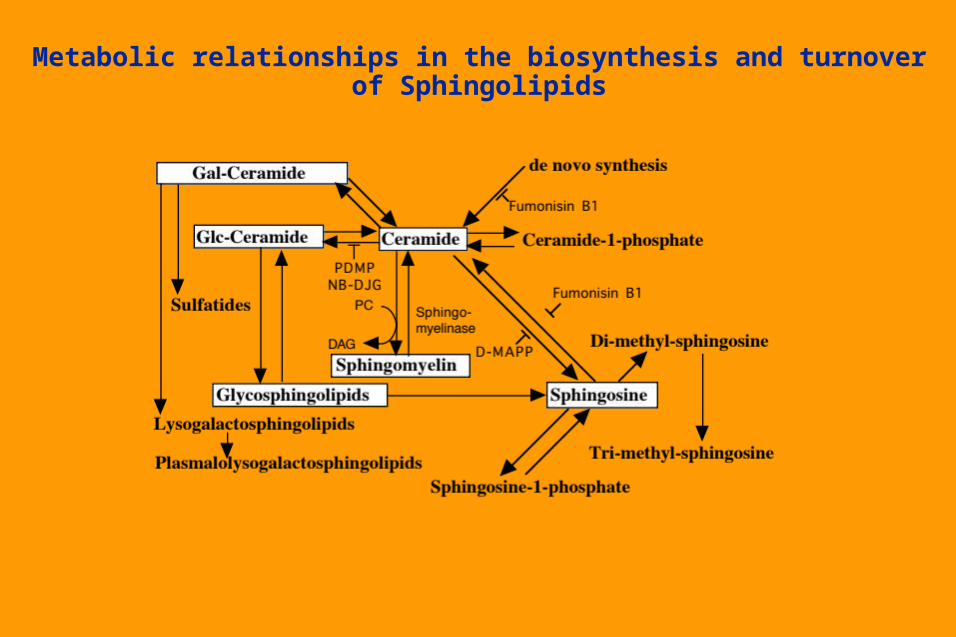
Metabolic relationships in the biosynthesis and turnover of Sphingolipids

Turnover and Degradation of Glycosphingolipids Internalized from plasma membrane via endocytosisPass through endosomes (some remodelling possible?)Terminal degradation in lysosomes - stepwise reactions by specific
enzymes. Some final steps involve cleavages close to the cell membrane, and
require facilitation by specific sphingolipid activator proteins (SAPs). Individual components, available for
re-utilization in various pathways. At least some of glucosylceramide may
remain intact and be recycledHuman diseases in which specific enzymes
or SAPS are genetically deficient.

Monoclonal antibodies (Mabs) against Glycosphingolipids
Many “tumor-specific” MAbs directed against glycans Majority react best with glycosphingolipids. Most MAbs are actually detecting “onco-fetal” antigens Some used for diagnostic and prognostic applications in
human diseasesFew being exploited for attempts at monoclonal
antibody therapy of tumors. Many used to demonstrate cell type-specific regulation
of specific GSL structures in a temporal and
spatial manner during development.Precise meaning of findings for cancer
biology and development being explored..

Biological Roles of Glycosphingolipids

Biological Roles of Glycosphingolipids
Thought to be critical components of the epidermal (skin) permeability barrier
Organizing role in cell membrane. Thought to associate with GPI anchors in the trans-Golgi, forming “rafts” which target to apical domains of polarized epithelial cells
May also be in distinct glycosphingolipid enriched domains (“GEMs”) which are associated with cytosolic oncogenes and signalling molecules
Physical protection against hostile environnments Binding sites for the adhesion of symbiont bacteria. Highly specific receptor targets for a variety
of bacteria, toxins and viruses.

Biological Roles of Glycosphingolipids
Specific association of certain glycosphingolipids with certain membrane receptors.
Can mediate low-affinity but high specificity carbohydrate-carbohydrate interactions between different cell types.
Targets for autoimmune antibodies in Guillian-Barre and Miller-Fisher syndromes following Campylobacter infections and in some patients with human myeloma
Shed in large amounts by certain cancers - these are found to have a strong immunosuppressive effects,
via as yet unknown mechanisms

Examples of interactions between
glycosphingolipids and bacterial toxins
or receptors
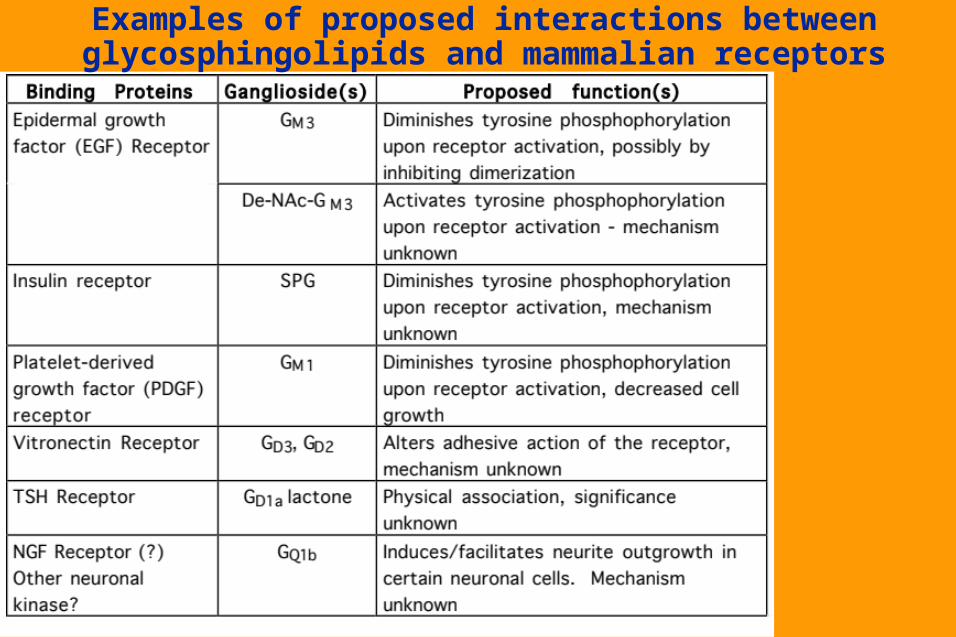
Examples of proposed interactions between glycosphingolipids and mammalian receptors

Natural and induced Genetic Disorders in Glycosphingolipid biosynthesis
Large number of known genetic defects in lysosomal enzymes or SAP proteins result in “storage disorders” characterized by the accumulation of specific intermediates.
Very few naturally-defined genetic defects in the biosynthesis of glycosphingolipids. A cultured cell line is completely deficient in the glucosylceramide synthase - thus,
glycosphingolipids are not essential for growth of single cells in a culture dishTargetted gene disruption of Ceramide Galactosyltransferase loss of Gal Cer and
sulfatides in nervous system myelin. Mice form myelin with GlcCer, which replaces GalCer. Despite myelin of relatively normal appearance mice have generalized tremors and mild ataxia, and electrophysiological evidence for conduction deficits. With increasing age, progressive hindlimb paralysis and severe vacuolation of the ventral region of the spinal cord.
Transgenic overexpression of the GalNAc transferase I (GM2/GD2 synthase) gives higher expression of complex gangliosides. No gross morphological changes observed, but much stronger inflammatory reactions involving neutrophils
Targetted gene disruption of the GalNAc transferase I (GM2/GD2 synthase) no major histological defects in nervous system nor in gross behavior, only a reduction in neural conduction velocity in some nerves. Compensatory increase in GM3 and GD3 in the brain seems sufficient to compensate for lack of complex gangliosides.

Natural and induced Genetic Disorders in Glycosphingolipid biosynthesis
Targetted gene disruption of Ceramide Galactosyltransferase loss of Gal Cer and sulfatides in nervous system myelin. Mice form myelin with GlcCer, which replaces GalCer. Despite myelin of relatively normal appearance mice have generalized tremors and mild ataxia, and electrophysiological evidence for conduction deficits. With increasing age, progressive hindlimb paralysis and severe vacuolation of the ventral region of the spinal cord.
Transgenic overexpression of the GalNAc transferase I (GM2/GD2 synthase) gives higher expression of complex gangliosides. No gross morphological changes
observed, but much stronger inflammatory
reactions involving neutrophils

Consequences of GalNAc Transferase I gene disruption

Natural and induced Genetic Disorders in Glycosphingolipid biosynthesis
Targetted gene disruption of the GalNAc transferase I (GM2/GD2 synthase) no major histological defects in nervous system nor in gross behavior, only a reduction in neural conduction velocity in some nerves. Compensatory increase in GM3 and GD3 in the brain seems sufficient to compensate for lack of complex gangliosides.
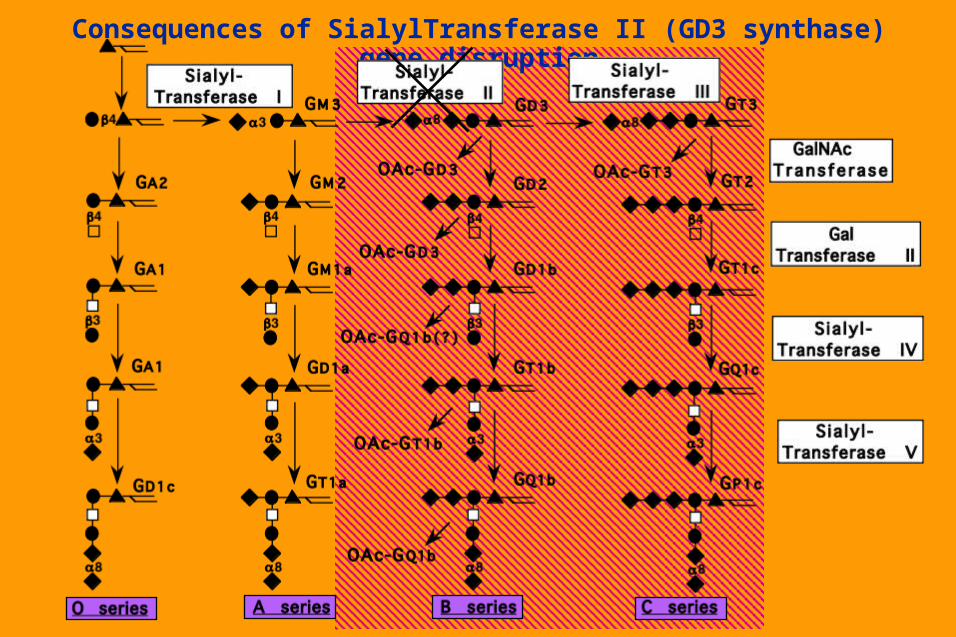
Consequences of SialylTransferase II (GD3 synthase) gene disruption

Consequences of SialylTransferase I (GM3 synthase) gene disruption

Consequences of Lactosylceramide Synthase gene disruption

Consequences of Glycosylceramide Synthase gene disruption
Related Documents











