ARTHRITIS & RHEUMATISM Vol. 62, No. 5, May 2010, pp 1349–1360 DOI 10.1002/art.27397 © 2010, American College of Rheumatology ERK-1/2 and p38 in the Regulation of Hypertrophic Changes of Normal Articular Cartilage Chondrocytes Induced by Osteoarthritic Subchondral Osteoblasts Indira Prasadam, 1 Stijn van Gennip, 2 Thor Friis, 1 Wei Shi, 1 Ross Crawford, 3 and Yin Xiao 1 Objective. Previous studies have shown the influ- ence of subchondral bone osteoblasts (SBOs) on pheno- typical changes of articular cartilage chondrocytes (ACCs) during the development of osteoarthritis (OA). The molecular mechanisms involved during this process remain elusive, in particular, the signal transduction pathways. The aim of this study was to investigate the in vitro effects of OA SBOs on the phenotypical changes in normal ACCs and to unveil the potential involvement of MAPK signaling pathways during this process. Methods. Normal and arthritic cartilage and bone samples were collected for isolation of ACCs and SBOs. Direct and indirect coculture models were applied to study chondrocyte hypertrophy under the influence of OA SBOs. MAPKs in the regulation of the cell–cell interactions were monitored by phosphorylated anti- bodies and relevant inhibitors. Results. OA SBOs led to increased hypertrophic gene expression and matrix calcification in ACCs by means of both direct and indirect cell–cell interactions. In this study, we demonstrated for the first time that OA SBOs suppressed p38 phosphorylation and induced ERK-1/2 signal phosphorylation in cocultured ACCs. The ERK-1/2 pathway inhibitor PD98059 significantly attenuated the hypertrophic changes induced by condi- tioned medium from OA SBOs, and the p38 inhibitor SB203580 resulted in the up-regulation of hypertrophic genes in ACCs. Conclusion. The findings of this study suggest that the pathologic interaction of OA SBOs and ACCs is mediated via the activation of ERK-1/2 phosphorylation and deactivation of p38 phosphorylation, resulting in hypertrophic differentiation of ACCs. Explanations concerning the cause of osteoarthri- tis (OA) have long focused on the destruction of artic- ular cartilage, the activating factors of which were thought to be triggered as a result of repetitive loading (1). Pathologic changes of cartilage in OA are associated with changes in the cellular phenotype of articular cartilage chondrocytes (ACCs) to a state of terminal differentiation (2,3). However, the long-term molecular events that are responsible for this transition are not well understood. Recent studies suggest that the subchondral bone plays a major role in OA cartilage changes, an indication of active communication between the subchondral bone and the cartilage in the progression of OA (4,5). Bone anabolic factors, such as osteocalcin, osteopontin, and alkaline phosphatase (ALP) are all up-regulated in OA subchondral bone osteoblasts (SBOs) as compared with normal SBOs, supporting the notion of a dysfunction of osteoblast behavior (6–8). It has been shown in animal models of OA that a thickening of subchondral bone precedes cartilage changes (9,10), and it has further been demonstrated that in vivo factors produced by OA SBOs increase glycosaminoglycan release from the car- tilage (11) and can influence cartilage-specific gene expression (12). It was demonstrated by the application of a coculture model of bovine explant subchondral bone and cartilage that excision of subchondral bone from articular cartilage resulted in increased chondrocyte death, thus demonstrating the important role of sub- Supported by the Prince Charles Hospital Research Founda- tion. 1 Indira Prasadam, MSc, Thor Friis, BSc (Hons), Wei Shi, Dipl, Yin Xiao, PhD: Queensland University of Technology, Brisbane, Queensland, Australia; 2 Stijn van Gennip, BSc: Queensland University of Technology, Brisbane, Queensland, Australia, and University Med- ical Center St. Radboud, Nijmegen, The Netherlands; 3 Ross Crawford, DPhil (Oxon): Queensland University of Technology and Prince Charles Hospital, Brisbane, Queensland, Australia. Address correspondence and reprint requests to Yin Xiao, PhD, Institute of Health and Biomedical Innovation, Queensland University of Technology, Kelvin Grove Campus, 60 Musk Avenue, Brisbane, Queensland 4059, Australia. E-mail: [email protected]. Submitted for publication August 16, 2009; accepted in revised form February 4, 2010. 1349

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTHRITIS & RHEUMATISMVol. 62, No. 5, May 2010, pp 1349–1360DOI 10.1002/art.27397© 2010, American College of Rheumatology
ERK-1/2 and p38 in the Regulation of Hypertrophic Changesof Normal Articular Cartilage Chondrocytes Induced by
Osteoarthritic Subchondral Osteoblasts
Indira Prasadam,1 Stijn van Gennip,2 Thor Friis,1 Wei Shi,1 Ross Crawford,3 and Yin Xiao1
Objective. Previous studies have shown the influ-ence of subchondral bone osteoblasts (SBOs) on pheno-typical changes of articular cartilage chondrocytes(ACCs) during the development of osteoarthritis (OA).The molecular mechanisms involved during this processremain elusive, in particular, the signal transductionpathways. The aim of this study was to investigate the invitro effects of OA SBOs on the phenotypical changes innormal ACCs and to unveil the potential involvement ofMAPK signaling pathways during this process.
Methods. Normal and arthritic cartilage and bonesamples were collected for isolation of ACCs and SBOs.Direct and indirect coculture models were applied tostudy chondrocyte hypertrophy under the influence ofOA SBOs. MAPKs in the regulation of the cell–cellinteractions were monitored by phosphorylated anti-bodies and relevant inhibitors.
Results. OA SBOs led to increased hypertrophicgene expression and matrix calcification in ACCs bymeans of both direct and indirect cell–cell interactions.In this study, we demonstrated for the first time that OASBOs suppressed p38 phosphorylation and inducedERK-1/2 signal phosphorylation in cocultured ACCs.The ERK-1/2 pathway inhibitor PD98059 significantlyattenuated the hypertrophic changes induced by condi-
tioned medium from OA SBOs, and the p38 inhibitorSB203580 resulted in the up-regulation of hypertrophicgenes in ACCs.
Conclusion. The findings of this study suggestthat the pathologic interaction of OA SBOs and ACCs ismediated via the activation of ERK-1/2 phosphorylationand deactivation of p38 phosphorylation, resulting inhypertrophic differentiation of ACCs.
Explanations concerning the cause of osteoarthri-tis (OA) have long focused on the destruction of artic-ular cartilage, the activating factors of which werethought to be triggered as a result of repetitive loading(1). Pathologic changes of cartilage in OA are associatedwith changes in the cellular phenotype of articularcartilage chondrocytes (ACCs) to a state of terminaldifferentiation (2,3). However, the long-term molecularevents that are responsible for this transition are not wellunderstood.
Recent studies suggest that the subchondral boneplays a major role in OA cartilage changes, an indicationof active communication between the subchondral boneand the cartilage in the progression of OA (4,5). Boneanabolic factors, such as osteocalcin, osteopontin, andalkaline phosphatase (ALP) are all up-regulated in OAsubchondral bone osteoblasts (SBOs) as compared withnormal SBOs, supporting the notion of a dysfunction ofosteoblast behavior (6–8). It has been shown in animalmodels of OA that a thickening of subchondral boneprecedes cartilage changes (9,10), and it has furtherbeen demonstrated that in vivo factors produced by OASBOs increase glycosaminoglycan release from the car-tilage (11) and can influence cartilage-specific geneexpression (12). It was demonstrated by the applicationof a coculture model of bovine explant subchondral boneand cartilage that excision of subchondral bone fromarticular cartilage resulted in increased chondrocytedeath, thus demonstrating the important role of sub-
Supported by the Prince Charles Hospital Research Founda-tion.
1Indira Prasadam, MSc, Thor Friis, BSc (Hons), Wei Shi,Dipl, Yin Xiao, PhD: Queensland University of Technology, Brisbane,Queensland, Australia; 2Stijn van Gennip, BSc: Queensland Universityof Technology, Brisbane, Queensland, Australia, and University Med-ical Center St. Radboud, Nijmegen, The Netherlands; 3Ross Crawford,DPhil (Oxon): Queensland University of Technology and PrinceCharles Hospital, Brisbane, Queensland, Australia.
Address correspondence and reprint requests to Yin Xiao,PhD, Institute of Health and Biomedical Innovation, QueenslandUniversity of Technology, Kelvin Grove Campus, 60 Musk Avenue,Brisbane, Queensland 4059, Australia. E-mail: [email protected].
Submitted for publication August 16, 2009; accepted inrevised form February 4, 2010.
1349

chondral bone in maintaining joint homeostasis (13).However, the molecular mechanisms, and, in particular,the signaling pathways, by which normal and OA SBOsregulate the articular cartilage phenotype remain un-known.
Activation of the 3 major classes of MAPKs(ERK-1/2, JNK, and p38 MAPK) has been detected inchondrocytes (14). MAPKs are known to be responsiblefor the conversion of a vast number of extracellularstimuli into specific cellular responses, including chon-drocyte proliferation and differentiation (15,16). Therequirement of MAPK signaling pathways, in particular,p38 and ERK-1/2, during various phases of endochon-dral ossification has also been demonstrated in severalstudies (17,18). MAPK signaling pathways have beenshown to play a distinct role in aspects of cartilagebiology, such as cartilage matrix synthesis and homeosta-sis (19,20). The role of MAPK signaling in skeletaldevelopment and in the biology of cartilage pointstoward a possible association of altered MAPK signalingand OA. Indeed, alterations in these signaling pathwaysare reported to play a prominent role in chondrocytedysfunction as a part of OA pathogenesis and diseaseprogression (21).
Since the OA SBOs are reported to alter thecartilage phenotype, it is possible that these alterationsin ACCs may occur via MAPK regulation. However, nostudies to date have explored the role of MAPK signal-ing factors in the cell–cell interactions of SBOs andACCs. The present study was designed to investigateMAPK signaling pathways in the hypertrophic changesof normal ACCs induced by OA SBOs using both directand indirect coculture systems.
PATIENTS AND METHODS
Articular cartilage sample collection and determina-tion of phenotype. Ethical approval for this project was grantedby the Queensland University of Technology and the PrinceCharles Hospital Ethics Committees, and informed consentwas given by all subjects involved. ACCs from OA patients(n � 5) were sourced from the main defective area of themedial compartment cartilage in which there were degenera-tive changes. The mean � SD age of the OA patients in thisstudy was 65.20 � 5.94 years. ACCs from normal subjects (n �3) were obtained from trauma patients, where knee tissue wasavailable. Normal subjects were healthy adults with a mean �SD age of 53.56 � 10.76 years, no clinical signs or symptoms ofjoint, metabolic, or hormonal diseases (osteoporosis), and nohistory of medications that might affect cartilage or bonemetabolism. To eliminate the possibility that these sampleshad early OA, those showing any evidence of cartilage changeswere excluded. These changes included softening of the hya-line articular cartilage, thinning and fibrous dislocation, ulcer-
ations of the cartilage, and light sclerosis of the subchondralbone.
Cartilage features were classified according to theMankin scale (22), based on histologic assessment of SafraninO–and hematoxylin and eosin–stained sections. Chondrocytesfrom the cartilage tissues were isolated according to a previ-ously described method (23). Only early-passage ACCs (pas-sages 0–2) showing strong expression of type II collagen(COL2) and aggrecan (AGG) were used for subsequent exper-iments. To determine phenotype changes in ACCs, chondro-genic (COL2, AGG) and hypertrophic (ALP, COL10, core-binding factor �1 [CBFA1]) marker genes were measuredaccording to their messenger RNA (mRNA) expression.
Subchondral bone sample collection and determina-tion of phenotype. Bone specimens were taken within 5 mm ofthe subchondral bone plate. SBOs from OA patients (n � 5)were sourced from weight-bearing sites, where the cartilagewas degraded and showed prominent subchondral bone ero-sion and density. These samples were obtained from patientswith advanced OA who were undergoing primary total kneereplacement surgery. The mean � SD age of the OA patientsin this study was 65.20 � 5.94 years. SBOs from normalsubjects (n � 3) were collected from patients undergoingsurgery for fracture repair who had no evidence of boneerosion or cartilage degeneration, as judged according tocriteria established by the American College of Rheumatology(24). Normal subjects were healthy adults with a mean � SDage of 53.56 � 10.76 years, no clinical signs or symptoms ofjoint, metabolic, or hormonal diseases (osteoporosis), and nohistory of medications that might affect cartilage or bonemetabolism.
After removing the overlying cartilage, SBOs wereisolated according to the method described by Beresford et al(25,26). Isolated normal and OA SBOs were differentiated inosteogenic medium supplemented with 10% fetal bovine se-rum (In Vitro Technologies), 50 units/ml of penicillin, 50�g/ml of streptomycin, 10 nM dexamethasone, 10 mM�-glycerophosphate, and 50 �g/ml of ascorbic acid. The cellswere then used for characterization of bone cell phenotype, asdetermined by the expression of the bone markers alkalinephosphatase and osteocalcin, as well as by staining with 1%alizarin red solution after 2 weeks of osteogenic induction.
Chondrocyte pellet culture. Cell culture systems knownto preserve the chondrocyte phenotype were used in thecoculture studies. ACCs (2 � 105 cells) were resuspended inserum-free chondrogenic medium (serum-free high-glucoseDulbecco’s modified Eagle’s medium [DMEM; Invitrogen])supplemented with 10 ng/ml of transforming growth factor �3(TGF�3; Bio Scientific), 10 nM dexamethasone, 50 mg/ml ofascorbic acid, 10 mg/ml of sodium pyruvate, 10 mg/ml ofproline, and an insulin–transferrin–selenium supplement (finalconcentration of the supplement 10 �g/ml of insulin, 5.5 �g/mlof transferrin, 5 ng/ml of sodium selenite, 0.5 mg/ml of bovineserum albumin, and 4.7 �g/ml of linoleic acid) and thencentrifuged at 600g for 20 minutes to form a pellet. Pellets wereallowed to differentiate for 2 weeks under 3-dimensionalconditions in 15-ml Falcon tubes containing chondrogenicmedium, which was replenished every 2–3 days. After 2 weeksof chondrogenesis, ACC pellets were cocultured with normalor OA SBOs as described below.
1350 PRASADAM ET AL
High-density micromass culture. High-density micro-mass droplets were prepared as described previously (17).Briefly, ACCs were trypsinized, resuspended in growth me-dium at a final cell density of 2.5 � 107 cells/ml, spotted asdroplets of 10 �l/well in 24-well culture plates, and incubatedat 37°C for 2 hours to allow cell attachment to the plate.Micromasses were cultured for 1 week in chondrogenic me-dium. After 1 week, micromasses containing ACCs werecultured with the conditioned medium generated from normalor OA SBOs as described below.
Direct coculture. ACC pellets were prepared as de-scribed above, placed directly upon the monolayer of normal
or OA SBOs (75,000 cells/well) in 24-well plates, and cocul-tured for another 2 weeks in high-glucose DMEM supple-mented with 1% fetal calf serum (FCS), 0.5% L-glutamine, 50units/ml of penicillin, 50 �g/ml of streptomycin, 50 �M ascor-bic acid, 10 nM dexamethasone, and 10 mM �-glycero-phosphate. After 14 days of coculture, the ACC pellets werewashed 3 times in phosphate buffered saline, fixed in 4%paraformaldehyde for 10 minutes, and stained with 1% alizarinred or 0.5% Alcian blue to assess the effect of normal or OASBOs on ACC matrix deposition. RNA and protein were alsoextracted from some of the pellets. The culture system selectedfor this coculture study was modified from the previously
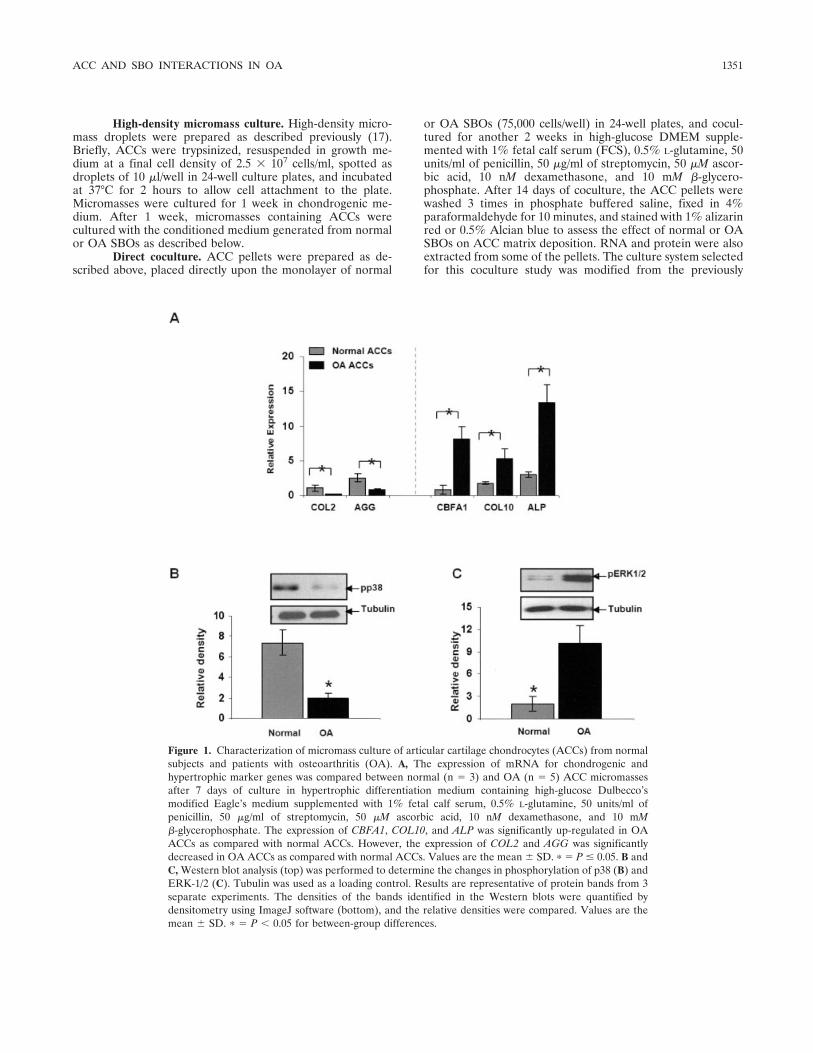
Figure 1. Characterization of micromass culture of articular cartilage chondrocytes (ACCs) from normalsubjects and patients with osteoarthritis (OA). A, The expression of mRNA for chondrogenic andhypertrophic marker genes was compared between normal (n � 3) and OA (n � 5) ACC micromassesafter 7 days of culture in hypertrophic differentiation medium containing high-glucose Dulbecco’smodified Eagle’s medium supplemented with 1% fetal calf serum, 0.5% L-glutamine, 50 units/ml ofpenicillin, 50 �g/ml of streptomycin, 50 �M ascorbic acid, 10 nM dexamethasone, and 10 mM�-glycerophosphate. The expression of CBFA1, COL10, and ALP was significantly up-regulated in OAACCs as compared with normal ACCs. However, the expression of COL2 and AGG was significantlydecreased in OA ACCs as compared with normal ACCs. Values are the mean � SD. � � P � 0.05. B andC, Western blot analysis (top) was performed to determine the changes in phosphorylation of p38 (B) andERK-1/2 (C). Tubulin was used as a loading control. Results are representative of protein bands from 3separate experiments. The densities of the bands identified in the Western blots were quantified bydensitometry using ImageJ software (bottom), and the relative densities were compared. Values are themean � SD. � � P � 0.05 for between-group differences.
ACC AND SBO INTERACTIONS IN OA 1351
described protocols for the formation of a chondro-osseousrudiment in micromass cultures (27).
Indirect coculture and preparation of SBO condi-tioned medium. Passage 2 SBOs from normal and OA sub-chondral bone (2.5 � 105 cells) were cultured in high-glucoseDMEM supplemented with 1% FCS, 0.5% L-glutamine, 25units/ml of penicillin, 25 �g/ml of streptomycin, 50 �M ascor-bic acid, 10 nM dexamethasone, and 10 mM �-glycer-ophosphate in 25-cm2 flasks for 2 days. The media from theseflasks was collected and centrifuged at 1,000g for 15 minutes.The supernatants were transferred to fresh tubes and mixedwith an equal volume of fresh (preincubated in 37°C in theincubator) medium with the same supplements to form condi-tioned medium. During coculture experiments, ACC micro-masses prepared as described above were grown for 1 week inconditioned medium from either normal or OA SBOs. ControlACCs were cultured in the same medium composition de-scribed above, but were not incubated with SBOs. Medium wasreplenished every 2 days. At the end of the coculture period,protein and total RNA were harvested from the ACCs, andsome cells were fixed with 4% paraformaldehyde and stained
with alizarin red and Alcian blue to assess extracellular matrixdeposition.
Detection of MAPK activation. MAPK-mediated cel-lular interactions were evaluated with the use of MAPKinhibitors SB203580 and PD98059 (both from Novabiochem),which inhibit the p38 and ERK-1/2 pathways, respectively.ACC micromasses were incubated with or without the MAPKinhibitors in conditioned medium prepared from normal orOA SBOs. Stock solutions of each inhibitor were dissolved inDMSO (final concentration of DMSO not exceeding 0.1%[volume/volume]). An equal amount of DMSO vehicle wasadded to control ACCs. Medium was replenished every 2 days.Optimal concentrations for inhibition in ACCs were found tobe 10 �M for ERK-1/2 and 5 �M for p38. At these concentra-tions, there was no evidence of cytotoxicity, nor was cellproliferation influenced by the addition of the inhibitors. Allexperiments were performed in triplicate.
RNA extraction and quantitative reverse transcrip-tion–polymerase chain reaction (RT-PCR). Total RNA wasisolated with TRIzol reagent (Invitrogen), treated with
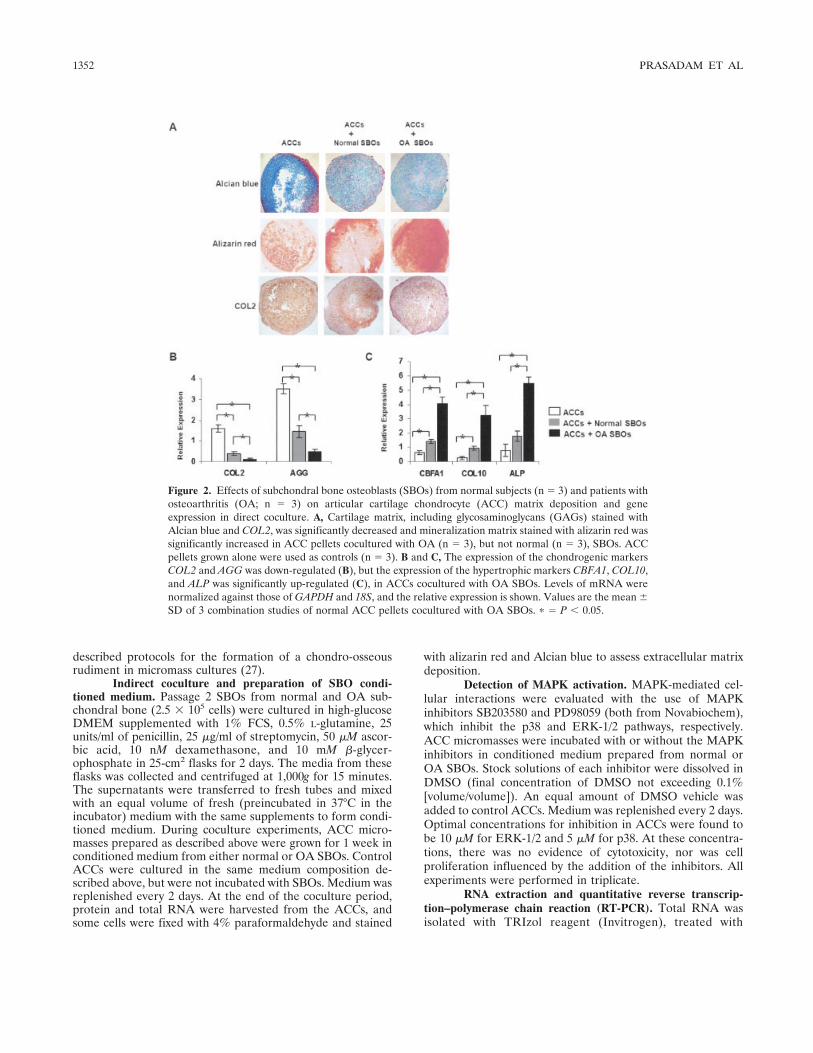
Figure 2. Effects of subchondral bone osteoblasts (SBOs) from normal subjects (n � 3) and patients withosteoarthritis (OA; n � 3) on articular cartilage chondrocyte (ACC) matrix deposition and geneexpression in direct coculture. A, Cartilage matrix, including glycosaminoglycans (GAGs) stained withAlcian blue and COL2, was significantly decreased and mineralization matrix stained with alizarin red wassignificantly increased in ACC pellets cocultured with OA (n � 3), but not normal (n � 3), SBOs. ACCpellets grown alone were used as controls (n � 3). B and C, The expression of the chondrogenic markersCOL2 and AGG was down-regulated (B), but the expression of the hypertrophic markers CBFA1, COL10,and ALP was significantly up-regulated (C), in ACCs cocultured with OA SBOs. Levels of mRNA werenormalized against those of GAPDH and 18S, and the relative expression is shown. Values are the mean �SD of 3 combination studies of normal ACC pellets cocultured with OA SBOs. � � P � 0.05.
1352 PRASADAM ET AL
DNase, and column purified using an RNeasy Mini kit (Qia-gen). Complementary DNA (cDNA) was synthesized from 1�g of total RNA using Superscript III (Invitrogen) accordingto the manufacturer’s instructions. PCR primers were designedbased on cDNA sequences from the National Center forBiotechnology Information Sequence database using PrimerExpress software; primer specificity was confirmed byBLASTN searches. Quantitative RT-PCR was performed withan ABI Prism 7000 Thermal Cycler (Applied Biosystems)using SYBR Green detection reagent. Briefly, 2 �l of cDNA,20 pmoles of gene-specific primers, and 10 �l of 1� MasterMix were used in a 20-�l reaction volume; each sample wasdetermined in duplicate. Thermocycling conditions were asfollows: 1 cycle of 10 minutes at 95°C for activation of thepolymerase, 40 cycles of 10 seconds at 95°C, and 1 minute at60°C for amplification. Dissociation curve analysis was per-formed to verify the absence of primer dimers and/or nonspe-cific PCR products. The relative expression of the genes ofinterest was normalized against housekeeping genes GAPDHand 18S RNA.
Western blotting. Total protein lysates were harvestedby lysing the cells with a lysis buffer containing 1M Tris HCl(pH 8), 5M NaCl, 20% Triton X-140, 0.5M EDTA, and aprotease inhibitor cocktail (Roche). The cell lysate was clari-fied by centrifugation, and the protein concentration wasdetermined with a bicinchoninic acid protein assay (Sigma).Ten micrograms of protein was separated by electrophoresison a 12% sodium dodecyl sulfate–polyacrylamide gel, trans-ferred to a nitrocellulose membrane, and blocked with aTris–Tween buffer containing 5% nonfat milk. The mem-branes were incubated overnight at 4°C with primary antibod-ies against phospho-p38 (1:1,000 dilution; Genesearch),phospho–ERK-1/2 (1:2,000 dilution; Quantum Scientific), andtubulin (1:5,000 dilution; Quantum Scientific). The mem-branes were washed 3 times in Tris buffered saline–Tween andincubated for 1 hour with anti-rabbit secondary antibody at adilution of 1:2,000. The protein bands were visualized usingECL Plus Western Blotting Detection Reagents (AmershamBiosciences) and exposed on x-ray film (Fujifilm). Immuno-blots were analyzed by densitometry using ImageJ software
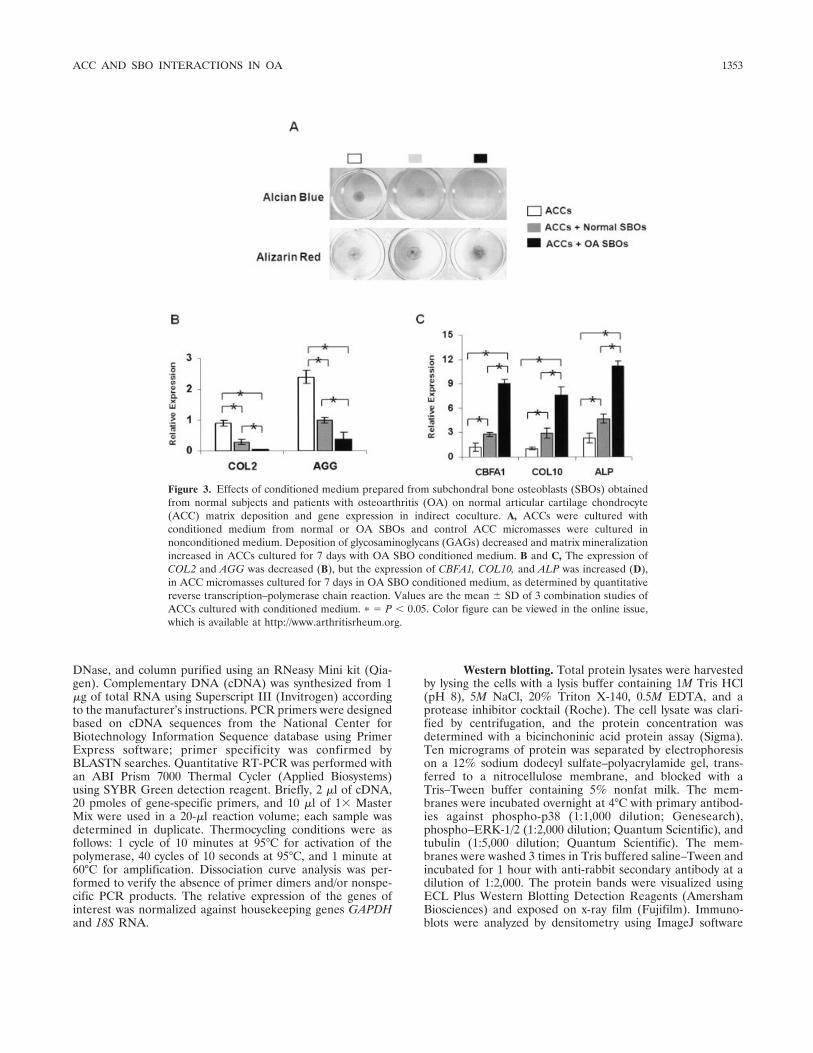
Figure 3. Effects of conditioned medium prepared from subchondral bone osteoblasts (SBOs) obtainedfrom normal subjects and patients with osteoarthritis (OA) on normal articular cartilage chondrocyte(ACC) matrix deposition and gene expression in indirect coculture. A, ACCs were cultured withconditioned medium from normal or OA SBOs and control ACC micromasses were cultured innonconditioned medium. Deposition of glycosaminoglycans (GAGs) decreased and matrix mineralizationincreased in ACCs cultured for 7 days with OA SBO conditioned medium. B and C, The expression ofCOL2 and AGG was decreased (B), but the expression of CBFA1, COL10, and ALP was increased (D),in ACC micromasses cultured for 7 days in OA SBO conditioned medium, as determined by quantitativereverse transcription–polymerase chain reaction. Values are the mean � SD of 3 combination studies ofACCs cultured with conditioned medium. � � P � 0.05. Color figure can be viewed in the online issue,which is available at http://www.arthritisrheum.org.
ACC AND SBO INTERACTIONS IN OA 1353
(NIH Image, National Institutes of Health; online at: http://rsbweb.nih.gov/ij/).
Statistical analysis. Each normal sample of ACCs wascocultured with either normal (n � 3) or OA (n � 3) SBOs,and the study was repeated in 3 normal ACC samples. Resultsare presented as the mean � SD. The relative expressionrepresents the mean of 3 combinations of chondrocytes andosteoblasts in the coculture studies. Repeated-measures ana-lysis of variance with post hoc tests was used to assess statisticalsignificance. P values less than or equal to 0.05 were consid-ered significant.
RESULTS
Expression of chondrogenic and hypertrophicgenes in normal and OA ACCs. The cell proliferationstudy indicated that there was no difference in cellnumbers between normal and OA ACC pellet cultures(data not shown). The expression of messenger RNA(mRNA) for chondrogenic and hypertrophic markergenes was compared between normal and OA ACCs.The expression of CBFA1, COL10, and ALP was signif-icantly up-regulated in OA ACCs as compared with
normal ACCs (P � 0.05 for each comparison), whereasthe expression of COL2 and AGG was significantlydown-regulated in OA ACCs as compared with normalACCs (P � 0.05 for each comparison) (Figure 1A).These results indicated that OA ACCs had greaterpotential to undergo hypertrophic differentiation ascompared with normal ACCs. With regard to MAPKphosphorylation, we observed that phosphorylation ofp38 was down-regulated in OA ACCs as compared withnormal ACCs (Figure 1B). On the other hand, phos-phorylation of ERK-1/2 was significantly up-regulated inOA ACCs as compared with normal ACCs (Figure 1C).
Hypertrophic differentiation of normal ACCs incoculture with OA SBOs. Direct coculture. No differencein cell proliferation rates was observed in SBOs after 14days of culture (data not shown). On day 14 of coculture,glycosaminoglycan matrix deposition was lower in thecoculture groups as compared with the noncoculturedACC pellets. There was slightly lower staining intensityin the coculture with OA SBOs as compared with that inthe coculture with normal SBOs (Figure 2A, top). On
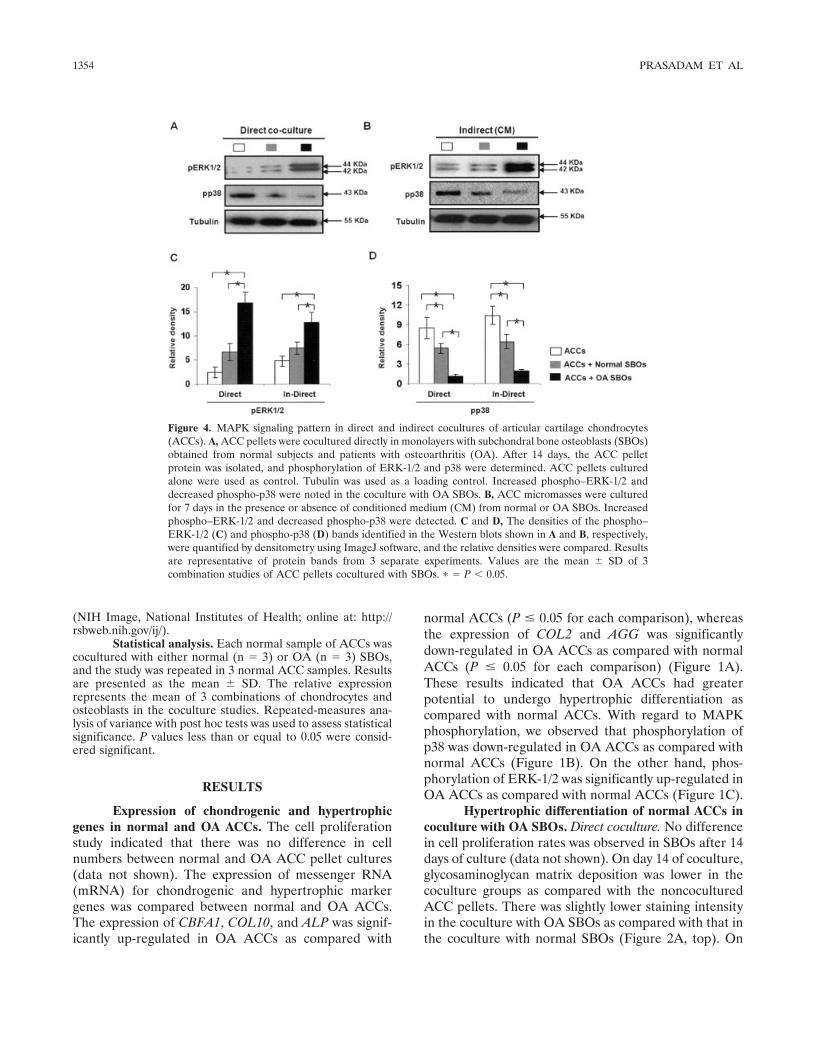
Figure 4. MAPK signaling pattern in direct and indirect cocultures of articular cartilage chondrocytes(ACCs). A, ACC pellets were cocultured directly in monolayers with subchondral bone osteoblasts (SBOs)obtained from normal subjects and patients with osteoarthritis (OA). After 14 days, the ACC pelletprotein was isolated, and phosphorylation of ERK-1/2 and p38 were determined. ACC pellets culturedalone were used as control. Tubulin was used as a loading control. Increased phospho–ERK-1/2 anddecreased phospho-p38 were noted in the coculture with OA SBOs. B, ACC micromasses were culturedfor 7 days in the presence or absence of conditioned medium (CM) from normal or OA SBOs. Increasedphospho–ERK-1/2 and decreased phospho-p38 were detected. C and D, The densities of the phospho–ERK-1/2 (C) and phospho-p38 (D) bands identified in the Western blots shown in A and B, respectively,were quantified by densitometry using ImageJ software, and the relative densities were compared. Resultsare representative of protein bands from 3 separate experiments. Values are the mean � SD of 3combination studies of ACC pellets cocultured with SBOs. � � P � 0.05.
1354 PRASADAM ET AL
the other hand, mineralization in the ACC pellets wassignificantly enhanced in ACCs that had been cocul-tured with OA SBOs as compared with the noncocul-tured ACCs and the ACCs cocultured with normal SBOs(Figure 2A, middle). The expression of COL2 immuno-staining was decreased in the coculture groups as com-pared with the noncocultured ACC pellets. Further-more, the expression of COL2 in ACCs was significantlydecreased in the presence of OA SBOs as comparedwith the normal SBOs (Figure 2A, bottom).
At the gene expression level, cartilage-specificgenes, such as COL2 and AGG, were significantly lowerin ACCs cocultured with OA SBOs than in ACCscocultured with normal SBOs or in ACCs alone (Figure2B). These observations were further validated by themRNA expression of hypertrophy and mineralization
marker genes in ACC pellets. The results of quantitativeRT-PCR analysis indicated that OA SBOs induced asignificant up-regulation of mineralization and hypertro-phic markers, such as COL10, ALP, and CBFA1, ascompared with the coculture group containing normalSBOs and with ACC pellets alone (Figure 2C).
Indirect coculture. When ACC micromasses werecultured with normal or OA SBO conditioned medium,Alcian blue staining revealed that cartilage matrix dep-osition was attenuated by both normal and OA SBOconditioned medium; however, cartilage matrix loss wasmore prominent in OA SBO conditioned medium (Fig-ure 3A, top). Conversely, matrix mineralization wasgreater in ACC micromasses grown for 7 days in OASBO conditioned medium, as demonstrated by alizarinred staining (Figure 3A, bottom). Induction of the
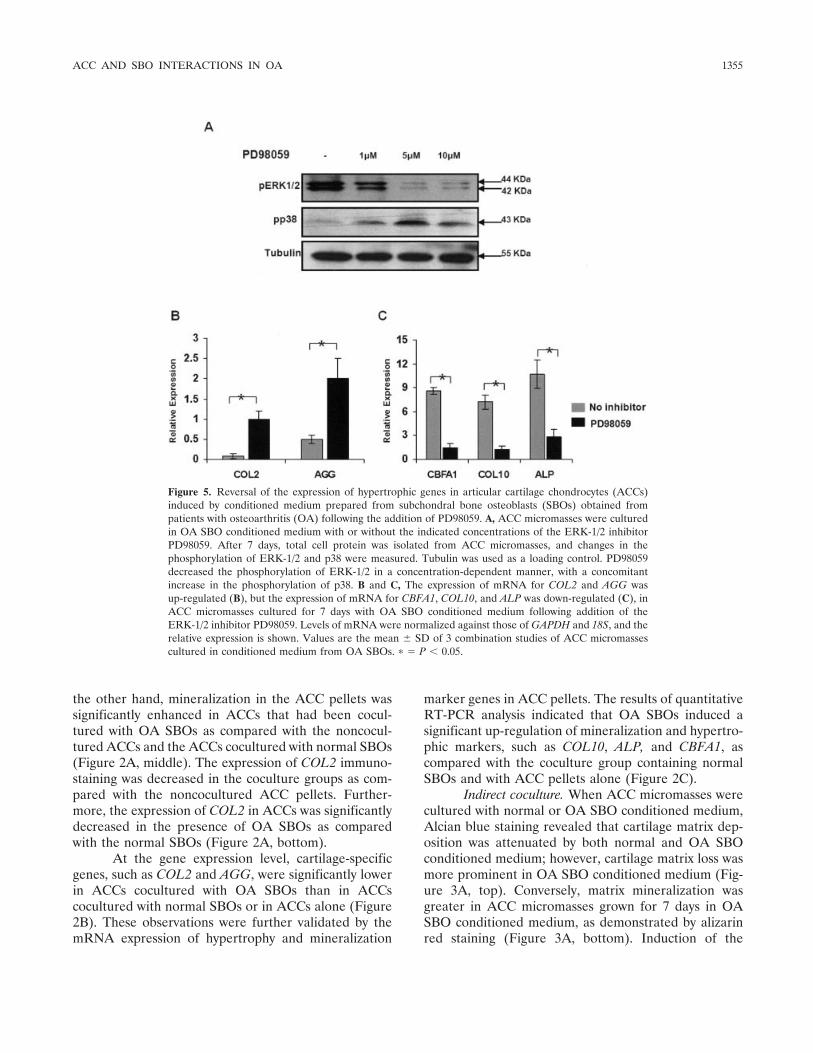
Figure 5. Reversal of the expression of hypertrophic genes in articular cartilage chondrocytes (ACCs)induced by conditioned medium prepared from subchondral bone osteoblasts (SBOs) obtained frompatients with osteoarthritis (OA) following the addition of PD98059. A, ACC micromasses were culturedin OA SBO conditioned medium with or without the indicated concentrations of the ERK-1/2 inhibitorPD98059. After 7 days, total cell protein was isolated from ACC micromasses, and changes in thephosphorylation of ERK-1/2 and p38 were measured. Tubulin was used as a loading control. PD98059decreased the phosphorylation of ERK-1/2 in a concentration-dependent manner, with a concomitantincrease in the phosphorylation of p38. B and C, The expression of mRNA for COL2 and AGG wasup-regulated (B), but the expression of mRNA for CBFA1, COL10, and ALP was down-regulated (C), inACC micromasses cultured for 7 days with OA SBO conditioned medium following addition of theERK-1/2 inhibitor PD98059. Levels of mRNA were normalized against those of GAPDH and 18S, and therelative expression is shown. Values are the mean � SD of 3 combination studies of ACC micromassescultured in conditioned medium from OA SBOs. � � P � 0.05.
ACC AND SBO INTERACTIONS IN OA 1355
cartilage-specific genes COL2 and AGG was signifi-cantly down-regulated in the presence of both normaland OA SBO conditioned medium as compared withACCs cultured alone, although this decrease was moreprominent in ACCs grown in the presence of OA SBOconditioned medium (Figure 3B). In contrast, the ex-pression of cartilage hypertrophy markers CBFA1,COL10, and ALP, were significantly up-regulated in thepresence of OA SBO conditioned medium (Figure 3C)as compared with both normal SBO conditioned me-dium and with the control groups.
Phospho-p38 and phospho–ERK-1/2 kinase sig-naling patterns in cocultured ACCs. The effect ofnormal and OA SBOs on the ACCs was assessed withrespect to alterations in the MAPK signaling cascade inboth direct and indirect coculture systems. The resultsshowed that the phosphorylation of ERK-1/2 was signif-
icantly augmented when ACCs were cocultured with OASBOs, in both the direct and the indirect coculturemodels, as compared with noncocultured ACCs andACCs cocultured with normal SBOs. Phosphorylation ofp38, on the other hand, was considerably down-regulated in the ACCs cocultured with normal SBOs ascompared with ACCs alone. Nonetheless, the cocultureof ACCs with OA SBOs led to a complete attenuation ofp38 phosphorylation. These results suggest that up-regulation of ERK-1/2 phosphorylation and down-regulation of p38 phosphorylation are involved in theinteraction between ACCs and SBOs, which in turn,leads to hypertrophic changes in ACCs (Figures 4A–D).
Reversal of phenotype changes in ACCs inducedby OA SBO conditioned medium following addition ofthe ERK-1/2 inhibitor PD98059. Incubation of ACCscultured with OA SBO conditioned medium with
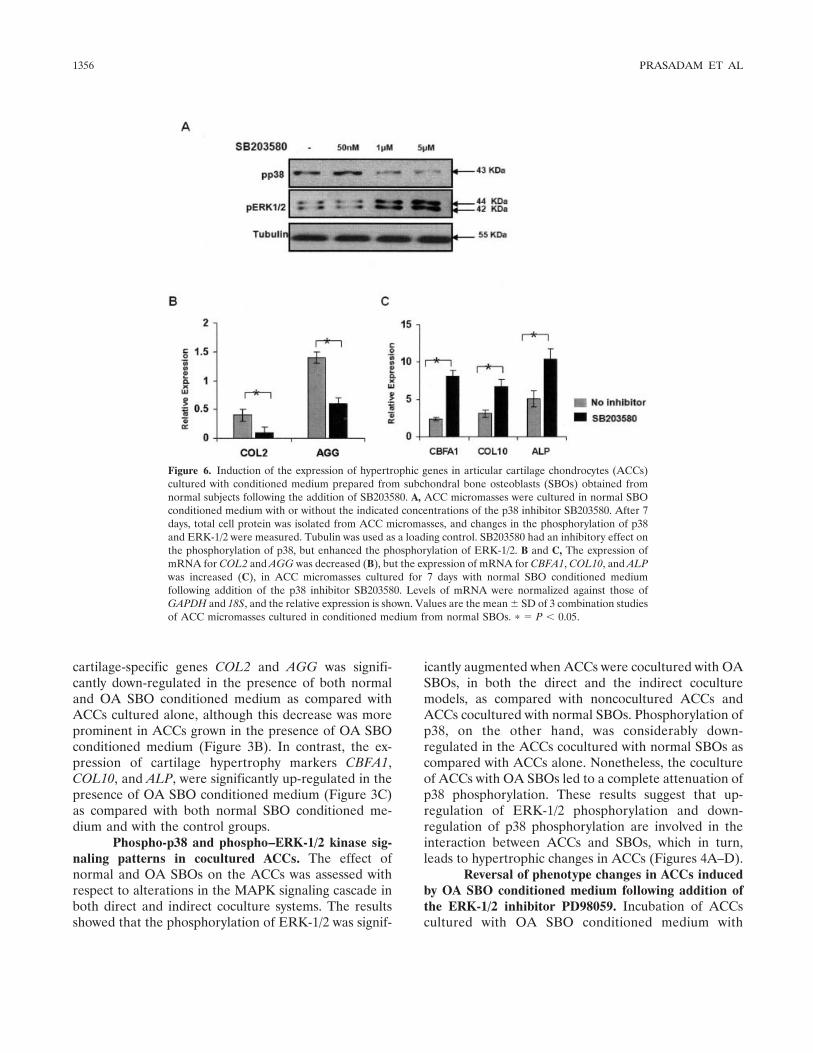
Figure 6. Induction of the expression of hypertrophic genes in articular cartilage chondrocytes (ACCs)cultured with conditioned medium prepared from subchondral bone osteoblasts (SBOs) obtained fromnormal subjects following the addition of SB203580. A, ACC micromasses were cultured in normal SBOconditioned medium with or without the indicated concentrations of the p38 inhibitor SB203580. After 7days, total cell protein was isolated from ACC micromasses, and changes in the phosphorylation of p38and ERK-1/2 were measured. Tubulin was used as a loading control. SB203580 had an inhibitory effect onthe phosphorylation of p38, but enhanced the phosphorylation of ERK-1/2. B and C, The expression ofmRNA for COL2 and AGG was decreased (B), but the expression of mRNA for CBFA1, COL10, and ALPwas increased (C), in ACC micromasses cultured for 7 days with normal SBO conditioned mediumfollowing addition of the p38 inhibitor SB203580. Levels of mRNA were normalized against those ofGAPDH and 18S, and the relative expression is shown. Values are the mean � SD of 3 combination studiesof ACC micromasses cultured in conditioned medium from normal SBOs. � � P � 0.05.
1356 PRASADAM ET AL

PD98059 had the effect of decreasing the expression ofphospho–ERK-1/2 and increasing the expression ofphospho-p38 in a concentration-dependent manner(Figure 5A). ACCs alone did not show significantchanges in response to the addition of PD98059, whichindicates that the observed effects were specific to OASBO conditioned medium. Quantitative RT-PCR analy-sis showed that PD98059 reversed the expression ofhypertrophic gene expression of CBFA1, COL10, andALP in ACC micromasses cultured in the presence ofOA SBO conditioned medium (Figure 5C). In contrast,the expression of COL2 and AGG was up-regulated bythe addition of PD98059 (Figure 5B).
Induction of hypertrophic gene expression inACCs cultured in normal SBO conditioned mediumfollowing addition of the p38 inhibitor SB203580. Inhi-bition of the phosphorylation of p38 by SB203580 led toa reduction of p38 phosphorylation and activated thephosphorylation of ERK-1/2 in ACCs cultured in nor-mal SBO conditioned medium (Figure 6A). In thepresence of SB203580, the hypertrophic markersCBFA1, COL10, and ALP were significantly enhanced,whereas the chondrogenic markers COL2 and AGGwere down-regulated in ACCs in the presence of normalSBO conditioned medium (Figures 6C and D). Theseresults indicate that the use of SB203580 could signifi-cantly shift ACCs toward a more hypertrophic pheno-type. Similar results were obtained in normal ACCscultured alone (data not shown).
DISCUSSION
In this study, we demonstrated the importance ofMAPK signaling pathways as the means by which OAsubchondral bone osteoblasts induce altered phenotypechanges in articular cartilage chondrocytes. Our experi-ments also provide some insight into the cross-talktaking place between the p38 and ERK-1/2 signalingpathways during this pathologic interaction process.
It was observed that ACCs isolated from OApatients produced significantly greater levels of mRNAfor CBFA1, COL10, and ALP genes as compared withACCs isolated from healthy patients. This finding indi-cates that the OA ACCs possessed greater potential toundergo hypertrophic differentiation; these results cor-roborate those of previous studies comparing normaland OA ACCs (28,29).
Applying an in vitro indirect coculture model,Sanchez et al (12) demonstrated that sclerotic OA SBOsdecreased cartilage-specific gene expression, such asSOX9 and COL2. They also showed that inhibitors of
hypertrophic differentiation, such as parathyroidhormone–related protein and parathyroid hormone re-ceptor were significantly down-regulated in ACCs cocul-tured with OA SBOs (12). These findings are evidencethat OA SBOs can decrease the inhibitors of hypertro-phic differentiation, leading to a subsequent mineralizedmatrix deposition in cartilage. In the present study, usingboth direct and indirect coculture methods, we showedthat OA SBOs increased both hypertrophic gene expres-sion and matrix mineralization.
Interestingly, hypertrophic changes are followedby a simultaneous decrease in chondrocyte-specificphenotype. A characteristic change of OA is anup-regulation of hypertrophy-related markers andmineralization-related markers (3) and a down-regulation of chondrocyte-specific markers (COL2 andAGG) in articular cartilage (30). The observations in ourstudy suggest that the interaction of OA SBOs may leadto these typical hypertrophic changes in ACCs. It hasbeen reported that the transition of ACCs to hypertro-phic changes contributes to the activation of matrixmetalloproteinases (MMPs), which precedes cartilagedegeneration (31,32), indicating that the phenotypicconversion of ACCs to hypertrophy is pathologic for thehealth and integrity of articular cartilage, leading to itsdegeneration.
The reasons why OA SBOs seem to induce thealtered ACC phenotypes remain unclear; however, sev-eral potential pathways may be responsible. Both ourown studies (data not shown) and those by other groupsof investigators have demonstrated that OA SBOs pro-duce abnormal levels of osteogenic markers, growthfactors, and cytokines. Specifically, increased productionof growth factors, such as insulin-like growth factor 1(IGF-1) (33), and TGF� (34), have been reported in OASBOs. Among these factors, IGF-1 has been implicatedin the induction of cartilage hypertrophic changes ingrowth plate chondrocytes (35,36). In addition, it hasbeen reported that OA SBOs produce abnormal levelsof cytokines such as interleukin-1 and interleukin-6 (34),tumor necrosis factor, and MMP-13 (37,38), all of whichhave the ability to activate a diverse array of signalingpathways in cartilage hypertrophy. It is therefore possi-ble that the biomolecules secreted from OA SBOscommunicate either individually or cooperatively withACCs, thereby mediating the induction of phenotypechanges of ACCs. Further studies are required to delin-eate the soluble factors from OA SBOs that are respon-sible for triggering the hypertrophic changes of ACCs inOA.
ACC AND SBO INTERACTIONS IN OA 1357

Among the signaling factors, the MAPK subtypesERK-1/2 and p38 play a key role in the signaling processof chondrocyte cellular differentiation and homeostasis,depending on the nature of extracellular stimuli (14,39).This knowledge prompted us to investigate MAPKsignaling in the context of the influence of normal andOA SBOs on the differentiation of ACCs. This study isthe first of its kind to show that OA SBOs induceERK-1/2 phosphorylation and suppress p38 phosphory-lation in ACCs, indicating that the alterations of thesepathways accompany the pathologic phenotype changesin ACCs. Indeed, we have demonstrated that basal levelsof ERK-1/2 phosphorylation increased and p38 de-creased in OA ACCs as compared with normal ACCs,an indication of the pathologic relevance of these path-ways in the pathogenesis of OA.
When the influence of ERK-1/2 phosphorylationwas blocked by an inhibitor, p38 was activated in ACCsgrown in the presence of OA SBO conditioned medium.The application of the ERK-1/2 inhibitor in OA SBOconditioned medium reversed ACC hypertrophy, andthere was a return to the chondrogenic phenotype ofACCs. This observation implies that OA SBOs inducedaltered phenotype changes in ACCs via a deactivation ofp38 and an activation of ERK-1/2 phosphorylation. Thisnotion was further supported by results showing thatwhen p38 was neutralized by an inhibitor in ACCscocultured with normal SBOs, ERK-1/2 phosphorylationwas augmented, and a weakening of chondrogenic geneexpression and increase of hypertrophic gene expressionwas observed. Taken together, these data indicate thatOA SBOs decrease p38 phosphorylation and increaseERK-1/2 activity, with a resulting reduction in chondro-genic phenotype and an increase in hypertrophic phe-notype.
MAPKs are regulated at several levels, includingkinase–kinase and kinase–substrate interactions and in-hibition of cross-talk/output by the MAPKs themselves(40,41). The activities of p38 are primarily governed byextensive cross-talk with ERK-1/2, a process that in-volves protein phosphatase, resulting in a reciprocalbidirectional equilibrium between ERK-1/2 and p38phosphorylation, where an increase in p38 activity sup-presses the activation of ERK-1/2 and vice versa (42).Such cross-talk appears to play a role in the OASBO–regulated ACC phenotype, the existence of whichhas been shown in chondrocytes. For example, theopposing roles of ERK-1/2 and p38 have been demon-strated in chondrogenesis regulation (43). The findingthat ERK-1/2 activation increased the hypertrophic dif-ferentiation of ACCs is consistent with study findings of
a strong activation of the ERK-1/2 pathway in thehypertrophic zone of the growth plate (44). Further-more, it has been demonstrated that the inhibition ofERK-1/2 delayed hypertrophic differentiation in growthplate chondrocytes during endochondral ossification(45).
It is possible that components of the p38 andERK-1/2 pathways may interact directly in the transcrip-tion complex. The intermediate p38 and ERK-1/2 path-way substrates involved in these interaction are notknown, but it is interesting that PD98059 (anti–ERK-1/2) significantly reduced the expression of the transcrip-tion factor CBFA1, whereas SB203580 (anti-p38) acti-vated this transcription factor. During earlyskeletogenesis, chondrocyte hypertrophy is stimulatedthrough the expression of CBFA1 in prehypertrophicchondrocytes, most likely by up-regulation of COL10expression (46). Continuous expression of CBFA1 inchondrocytes induces hypertrophic differentiation andendochondral ossification, which is suggestive of animportant role of this transcription factor in triggeringhypertrophic changes (47). It is therefore possible thatOA SBO–induced altered phenotype changes are trig-gered in ACCs via the activation of the MAPK/CBFA1pathway.
In conclusion, this study demonstrated that OASBOs could induce the activation of ERK-1/2 and thedeactivation of p38 in articular cartilage chondrocytes,resulting hypertrophic changes of the chondrocytes.These data provide insight into the MAPK signalingpathways involved in the molecular mechanisms of thepathogenesis of OA, which may have significant clinicalimplications.
AUTHOR CONTRIBUTIONS
All authors were involved in drafting the article or revising itcritically for important intellectual content, and all authors approvedthe final version to be published. Dr. Xiao had full access to all of thedata in the study and takes responsibility for the integrity of the dataand the accuracy of the data analysis.Study conception and design. Prasadam, Crawford, Xiao.Acquisition of data. Prasadam, van Gennip, Friis, Shi, Crawford, Xiao.Analysis and interpretation of data. Prasadam, van Gennip, Friis, Shi,Crawford, Xiao.
REFERENCES
1. Loeser RF. Aging and osteoarthritis: the role of chondrocytesenescence and aging changes in the cartilage matrix. Osteoarthri-tis Cartilage 2009;17:971–9.
2. Sandell LJ, Aigner T. Articular cartilage and changes in arthritis,an introduction: cell biology of osteoarthritis [review]. ArthritisRes 2001;3:107–13.
1358 PRASADAM ET AL

3. Pullig O, Weseloh G, Ronneberger D, Kakonen S, Swoboda B.Chondrocyte differentiation in human osteoarthritis: expression ofosteocalcin in normal and osteoarthritic cartilage and bone. CalcifTissue Int 2000;67:230–40.
4. Karsdal MA, Leeming DJ, Dam EB, Henriksen K, AlexandersenP, Pastoureau P, et al. Should subchondral bone turnover betargeted when treating osteoarthritis? Osteoarthritis Cartilage2008;16:638–46.
5. Raynauld JP, Martel-Pelletier J, Berthiaume MJ, Abram F, Cho-quette D, Haraoui B, et al. Correlation between bone lesionchanges and cartilage volume loss in patients with osteoarthritis ofthe knee as assessed by quantitative magnetic resonance imagingover a 24-month period. Ann Rheum Dis 2008;67:683–8.
6. Truong LH, Kuliwaba JS, Tsangari H, Fazzalari NL. Differentialgene expression of bone anabolic factors and trabecular bonearchitectural changes in the proximal femoral shaft of primary hiposteoarthritis patients. Arthritis Res Ther 2006;8:R188.
7. Sanchez C, Deberg MA, Bellahcnee A, Castronovo V, Msika P,Delcour JP, et al. Phenotypic characterization of osteoblasts fromthe sclerotic zones of osteoarthritic subchondral bone. ArthritisRheum 2008;58:442–55.
8. Lajeunesse D, Hilal G, Pelletier JP, Martel-Pelletier J. Subchon-dral bone morphological and biochemical alterations in osteoar-thritis. Osteoarthritis Cartilage 1999;7:321–2.
9. Anderson-MacKenzie JM, Quasnichka HL, Starr RL, Lewis EJ,Billingham ME, Bailey AJ. Fundamental subchondral bonechanges in spontaneous knee osteoarthritis. Int J Biochem CellBiol 2005;37:224–36.
10. Fahlgren A, Messner K, Aspenberg P. Meniscectomy leads to anearly increase in subchondral bone plate thickness in the rabbitknee. Acta Orthop Scand 2003;74:437–41.
11. Westacott CI, Webb GR, Warnock MG, Sims JV, Elson CJ.Alteration of cartilage metabolism by cells from osteoarthriticbone. Arthritis Rheum 1997;40:1282–91.
12. Sanchez C, Deberg MA, Piccardi N, Msika P, Reginster JY,Henrotin YE. Subchondral bone osteoblasts induce phenotypicchanges in human osteoarthritic chondrocytes. Osteoarthritis Car-tilage 2005;13:988–97.
13. Amin AK, Huntley JS, Simpson AH, Hall AC. Chondrocytesurvival in articular cartilage: the influence of subchondral bone ina bovine model. J Bone Joint Surg Br 2009;91:691–9.
14. Stanton LA, Underhill TM, Beier F. MAP kinases in chondrocytedifferentiation. Dev Biol 2003;263:165–75.
15. Han YS, Bang OS, Jin EJ, Park JH, Sonn JK, Kang SS. High doseof glucose promotes chondrogenesis via PKC� and MAPK signal-ing pathways in chick mesenchymal cells. Cell Tissue Res 2004;318:571–8.
16. Nakamura K, Shirai T, Morishita S, Uchida S, Saeki-Miura K,Makishima F. p38 mitogen-activated protein kinase functionallycontributes to chondrogenesis induced by growth/differentiationfactor-5 in ATDC5 cells. Exp Cell Res 1999;250:351–63.
17. Stanton LA, Beier F. Inhibition of p38 MAPK signaling inchondrocyte cultures results in enhanced osteogenic differentia-tion of perichondral cells. Exp Cell Res 2007;313:146–55.
18. Hoffman LM, Weston AD, Underhill TM. Molecular mechanismsregulating chondroblast differentiation. J Bone Joint Surg Am2003;85-A Suppl 2:124–32.
19. Zhen X, Wei L, Wu Q, Zhang Y, Chen Q. Mitogen-activatedprotein kinase p38 mediates regulation of chondrocyte differenti-ation by parathyroid hormone. J Biol Chem 2001;276:4879–85.
20. Watanabe H, de Caestecker MP, Yamada Y. Transcriptionalcross-talk between Smad, ERK1/2, and p38 mitogen-activatedprotein kinase pathways regulates transforming growth factor-�-induced aggrecan gene expression in chondrogenic ATDC5 cells.J Biol Chem 2001;276:14466–73.
21. Loeser RF, Erickson EA, Long DL. Mitogen-activated protein
kinases as therapeutic targets in osteoarthritis. Curr Opin Rheu-matol 2008;20:581–6.
22. Mankin HJ, Dorfman H, Lippiello L, Zarins A. Biochemical andmetabolic abnormalities in articular cartilage from osteo-arthritichuman hips. II. Correlation of morphology with biochemical andmetabolic data. J Bone Joint Surg Am 1971;53:523–37.
23. Patti AM, Gabriele A, Della Rocca C. Human chondrocyte celllines from articular cartilage of metatarsal phalangeal joints.Tissue Cell 1999;31:550–4.
24. Altman R, Asch E, Bloch D, Bole G, Borenstein D, Brandt K, etal. Development of criteria for the classification and reporting ofosteoarthritis: classification of osteoarthritis of the knee. ArthritisRheum 1986;29:1039–49.
25. Beresford JN, Gallagher JA, Gowen M, McGuire MK, Poser JW,Russell RG. Human bone cells in culture: a novel system for theinvestigation of bone cell metabolism. Clin Sci (Colch) 1983;64:38–9.
26. Beresford JN, Gallagher JA, Poser JW, Russell RG. Production ofosteocalcin by human bone cells in vitro: effects of 1,25(OH)2D3,24,25(OH)2D3, parathyroid hormone, and glucocorticoids. MetabBone Dis Relat Res 1984;5:229–34.
27. Muraglia A, Corsi A, Riminucci M, Mastrogiacomo M, CanceddaR, Bianco P, et al. Formation of a chondro-osseous rudiment inmicromass cultures of human bone-marrow stromal cells. J CellSci 2003;116:2949–55.
28. Pullig O, Weseloh G, Gauer S, Swoboda B. Osteopontin isexpressed by adult human osteoarthritic chondrocytes: protein andmRNA analysis of normal and osteoarthritic cartilage. Matrix Biol2000;19:245–55.
29. Terkeltaub RA. What does cartilage calcification tell us aboutosteoarthritis? J Rheumatol 2002;29:411–5.
30. Eid K, Thornhill TS, Glowacki J. Chondrocyte gene expression inosteoarthritis: correlation with disease severity. J Orthop Res2006;24:1062–8.
31. D’Angelo M, Yan Z, Nooreyazdan M, Pacifici M, Sarment DS,Billings PC, et al. MMP-13 is induced during chondrocyte hyper-trophy. J Cell Biochem 2000;77:678–93.
32. Johnson K, Terkeltaub R. Upregulated ank expression in osteo-arthritis can promote both chondrocyte MMP-13 expression andcalcification via chondrocyte extracellular PPi excess. Osteoarthri-tis Cartilage 2004;12:321–35.
33. Massicotte F, Fernandes JC, Martel-Pelletier J, Pelletier JP,Lajeunesse D. Modulation of insulin-like growth factor 1 levels inhuman osteoarthritic subchondral bone osteoblasts. Bone 2006;38:333–41.
34. Massicotte F, Lajeunesse D, Benderdour M, Pelletier JP, Hilal G,Duval N, et al. Can altered production of interleukin-1�, interleu-kin-6, transforming growth factor-� and prostaglandin E2 byisolated human subchondral osteoblasts identify two subgroups ofosteoarthritic patients. Osteoarthritis Cartilage 2002;10:491–500.
35. Wang J, Zhou J, Bondy CA. Igf1 promotes longitudinal bonegrowth by insulin-like actions augmenting chondrocyte hypertro-phy. FASEB J 1999;13:1985–90.
36. Mushtaq T, Bijman P, Ahmed SF, Farquharson C. Insulin-likegrowth factor-I augments chondrocyte hypertrophy and reversesglucocorticoid-mediated growth retardation in fetal mice metatar-sal cultures. Endocrinology 2004;145:2478–86.
37. Hulejova H, Baresova V, Klezl Z, Polanska M, Adam M, Senolt L.Increased level of cytokines and matrix metalloproteinases inosteoarthritic subchondral bone. Cytokine 2007;38:151–6.
38. Sakao K, Takahashi KA, Mazda O, Arai Y, Tonomura H, Inoue A,et al. Enhanced expression of interleukin-6, matrix metalloprotein-ase-13, and receptor activator of NF-�B ligand in cells derivedfrom osteoarthritic subchondral bone. J Orthop Sci 2008;13:202–10.
39. Chun JS. Expression, activity, and regulation of MAP kinases incultured chondrocytes. Methods Mol Med 2004;100:291–306.
ACC AND SBO INTERACTIONS IN OA 1359

40. Krishna M, Narang H. The complexity of mitogen-activatedprotein kinases (MAPKs) made simple. Cell Mol Life Sci 2008;65:3525–44.
41. McClean MN, Mody A, Broach JR, Ramanathan S. Cross-talk anddecision making in MAP kinase pathways. Nat Genet 2007;39:409–14.
42. Xiao YQ, Malcolm K, Worthen GS, Gardai S, Schiemann WP,Fadok VA, et al. Cross-talk between ERK and p38 MAPKmediates selective suppression of pro-inflammatory cytokines bytransforming growth factor-�. J Biol Chem 2002;277:14884–93.
43. Oh CD, Chang SH, Yoon YM, Lee SJ, Lee YS, Kang SS, et al.Opposing role of mitogen-activated protein kinase subtypes, Erk-1/2 and p38, in the regulation of chondrogenesis of mesenchymes.J Biol Chem 2000;275:5613–9.
44. Provot S, Nachtrab G, Paruch J, Chen AP, Silva A, Kronenberg
HM. A-raf and B-raf are dispensable for normal endochondralbone development, and parathyroid hormone-related peptide sup-presses extracellular signal-regulated kinase activation in hyper-trophic chondrocytes. Mol Cell Biol 2008;28:344–57.
45. Bobick BE, Kulyk WM. Regulation of cartilage formation andmaturation by mitogen-activated protein kinase signaling. BirthDefects Res C Embryo Today 2008;84:131–54.
46. Ueta C, Iwamoto M, Kanatani N, Yoshida C, Liu Y, Enomoto-Iwamoto M, et al. Skeletal malformations caused by overexpres-sion of Cbfa1 or its dominant negative form in chondrocytes. J CellBiol 2001;153:87–100.
47. Takeda S, Bonnamy JP, Owen MJ, Ducy P, Karsenty G. Continuousexpression of Cbfa1 in nonhypertrophic chondrocytes uncovers itsability to induce hypertrophic chondrocyte differentiation and par-tially rescues Cbfa1-deficient mice. Genes Dev 2001;15:467–81.
1360 PRASADAM ET AL
Related Documents







![The subchondral bone in articular cartilage repair ... · the subchondral plate as the initiating event in osteoarthritis [13]. While the entire osteochondral unit remains the same](https://static.cupdf.com/doc/110x72/60f326de55812e0e3d2df913/the-subchondral-bone-in-articular-cartilage-repair-the-subchondral-plate-as.jpg)


