TISSUE-SPECIFIC STEM CELLS Epithelial-Mesenchymal Transition-Derived Cells Exhibit Multilineage Differentiation Potential Similar to Mesenchymal Stem Cells VENKATA LOKESH BATTULA, a KURT WILLIAM EVANS, b BRETT GEORGE HOLLIER, b YUEXI SHI, a FRANK C. MARINI, a AYYAKKANNU AYYANAN, c RUI-YU WANG, a CATHRIN BRISKEN, c RUDY GUERRA, d MICHAEL ANDREEFF, a# SENDURAI A. MANI b# a Section of Molecular Hematology and Therapy, Departments of Stem Cell Transplantation and Cellular Therapy and b Molecular Pathology, The University of Texas-M.D. Anderson Cancer Center, Houston, Texas, USA; c ISREC, School of Life Science, Ecole Polytechnique Fe ´de ´rale (EPFL), Lausanne, Switzerland; d Department of Statistics, Rice University, Houston, Texas, USA Key Words. Epithelial-mesenchymal transition • Twist • Snail • MSC • Mesenchymal stem cells • CD140b • PDGFR-b ABSTRACT The epithelial-to-mesenchymal transition (EMT) is an embry- onic process that becomes latent in most normal adult tissues. Recently, we have shown that induction of EMT endows breast epithelial cells with stem cell traits. In this report, we have further characterized the EMT-derived cells and shown that these cells are similar to mesenchymal stem cells (MSCs) with the capacity to differentiate into multiple tissue lineages. For this purpose, we induced EMT by ectopic expression of Twist, Snail, or transforming growth factor-b in immortal- ized human mammary epithelial cells. We found that the EMT-derived cells and MSCs share many properties includ- ing the antigenic profile typical of MSCs, that is, CD44 1 , CD24 2 , and CD45 2 . Conversely, MSCs express EMT-associ- ated genes, such as Twist, Snail, and mesenchyme forkhead 1 (FOXC2). Interestingly, CD140b (platelet-derived growth factor receptor-b), a marker for naive MSCs, is exclusively expressed in EMT-derived cells and not in their epithelial counterparts. Moreover, functional analyses revealed that EMT-derived cells but not the control cells can differentiate into alizarin red S-positive mature osteoblasts, oil red O-posi- tive adipocytes and alcian blue-positive chondrocytes similar to MSCs. We also observed that EMT-derived cells but not the control cells invade and migrate towards MDA-MB-231 breast cancer cells similar to MSCs. In vivo wound homing assays in nude mice revealed that the EMT-derived cells home to wound sites similar to MSCs. In conclusion, we have demonstrated that the EMT-derived cells are similar to MSCs in gene expression, multilineage differentiation, and ability to migrate towards tumor cells and wound sites. STEM CELLS 2010;28:1435–1445 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION Epithelial-to-mesenchymal transition (EMT) is a latent embry- onic process that causes epithelial cells to lose their epithelial traits and acquire properties of mesenchymal cells. During EMT, epithelial cells lose cell polarity by downregulating the expression of cytokeratins and cell–cell adhesion molecules such as E-cadherin [1, 2]. The decrease in epithelial gene expression is accompanied by increased expression of mesen- chymal genes, including vimentin and fibronectin. Following passage through EMT, epithelial cells also acquire a mesen- chymal morphology in adherent culture and increased motility and invasiveness [1, 3, 4]. The EMT program plays an impor- tant role during the morphogenesis of multicellular organisms. For example, during gastrulation, epithelial cells located in the primitive streak undergo EMT followed by ingression and migration to a new location where they form endodermal and mesodermal embryonic tissues. In the absence of the EMT process, gastrulation does not occur and embryonic develop- ment does not progress past the blastula stage [5]. Similarly, during neural crest development, epithelial-looking cells within the neural plate shed their epithelial traits and gain a mesenchymal phenotype via EMT [6]. EMT can be induced by several cytokines and chemo- kines, including transforming growth factor-b (TGF-b), or by the expression of several developmentally important transcrip- tion factors, including Twist and Snail [7, 8]. We recently Author contributions: V.L.B.: conception and design, collection and assembly of data, data analysis and interpretation, manuscript writing; K.E.: collection and assembly of data, data analysis and interpretation, manuscript writing; B.G.H.: collection and assembly of data; Y.S.: collection and assembly of data; F.C.M.: collection and assembly of data, provision of study material; A.A.: collection and assembly of data, provision of study material; R.W.: collection and assembly of data, provision of study material; C.B.: provision of study material; M.A.: conception and design, data analysis and interpretation, financial support, final approval of the manuscript; S.A.M.: conception and design, data analysis and interpretation, financial support, manuscript writing, final approval of the manuscript. # Correspondence: Sendurai A. Mani PhD, (email:[email protected]) or Michael Andreeff M.D., Ph.D., (e-mail: mandreef@ mdanderson.org), 1515 Holcombe Blvd, Houston, TX, 77030; Received October 26, 2009; accepted for publication June 8, 2010; first published online in STEM CELLS EXPRESS June 22, 2010. V C AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.467 STEM CELLS 2010;28:1435–1445 www.StemCells.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

TISSUE-SPECIFIC STEM CELLS
Epithelial-Mesenchymal Transition-Derived Cells Exhibit
Multilineage Differentiation Potential Similar to
Mesenchymal Stem Cells
VENKATA LOKESH BATTULA,a KURT WILLIAM EVANS,b BRETT GEORGE HOLLIER,b YUEXI SHI,a
FRANK C. MARINI,aAYYAKKANNU AYYANAN,
cRUI-YU WANG,
aCATHRIN BRISKEN,
cRUDY GUERRA,
d
MICHAEL ANDREEFF,a# SENDURAI A. MANIb#
aSection of Molecular Hematology and Therapy, Departments of Stem Cell Transplantation and Cellular Therapy
and bMolecular Pathology, The University of Texas-M.D. Anderson Cancer Center, Houston, Texas, USA;cISREC, School of Life Science, Ecole Polytechnique Federale (EPFL), Lausanne, Switzerland; dDepartment of
Statistics, Rice University, Houston, Texas, USA
Key Words. Epithelial-mesenchymal transition • Twist • Snail • MSC • Mesenchymal stem cells • CD140b • PDGFR-b
ABSTRACT
The epithelial-to-mesenchymal transition (EMT) is an embry-
onic process that becomes latent in most normal adult tissues.Recently, we have shown that induction of EMT endowsbreast epithelial cells with stem cell traits. In this report, we
have further characterized the EMT-derived cells and shownthat these cells are similar to mesenchymal stem cells (MSCs)
with the capacity to differentiate into multiple tissue lineages.For this purpose, we induced EMT by ectopic expression ofTwist, Snail, or transforming growth factor-b in immortal-
ized human mammary epithelial cells. We found that theEMT-derived cells and MSCs share many properties includ-ing the antigenic profile typical of MSCs, that is, CD441,
CD242, and CD45
2. Conversely, MSCs express EMT-associ-
ated genes, such as Twist, Snail, and mesenchyme forkhead 1
(FOXC2). Interestingly, CD140b (platelet-derived growth
factor receptor-b), a marker for naive MSCs, is exclusively
expressed in EMT-derived cells and not in their epithelialcounterparts. Moreover, functional analyses revealed thatEMT-derived cells but not the control cells can differentiate
into alizarin red S-positive mature osteoblasts, oil red O-posi-tive adipocytes and alcian blue-positive chondrocytes similar
to MSCs. We also observed that EMT-derived cells but notthe control cells invade and migrate towards MDA-MB-231breast cancer cells similar to MSCs. In vivo wound homing
assays in nude mice revealed that the EMT-derived cellshome to wound sites similar to MSCs. In conclusion, we havedemonstrated that the EMT-derived cells are similar to
MSCs in gene expression, multilineage differentiation, andability to migrate towards tumor cells and wound sites. STEMCELLS 2010;28:1435–1445
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Epithelial-to-mesenchymal transition (EMT) is a latent embry-onic process that causes epithelial cells to lose their epithelialtraits and acquire properties of mesenchymal cells. DuringEMT, epithelial cells lose cell polarity by downregulating theexpression of cytokeratins and cell–cell adhesion moleculessuch as E-cadherin [1, 2]. The decrease in epithelial geneexpression is accompanied by increased expression of mesen-chymal genes, including vimentin and fibronectin. Followingpassage through EMT, epithelial cells also acquire a mesen-chymal morphology in adherent culture and increased motilityand invasiveness [1, 3, 4]. The EMT program plays an impor-
tant role during the morphogenesis of multicellular organisms.For example, during gastrulation, epithelial cells located inthe primitive streak undergo EMT followed by ingression andmigration to a new location where they form endodermal andmesodermal embryonic tissues. In the absence of the EMTprocess, gastrulation does not occur and embryonic develop-ment does not progress past the blastula stage [5]. Similarly,during neural crest development, epithelial-looking cellswithin the neural plate shed their epithelial traits and gain amesenchymal phenotype via EMT [6].
EMT can be induced by several cytokines and chemo-kines, including transforming growth factor-b (TGF-b), or bythe expression of several developmentally important transcrip-tion factors, including Twist and Snail [7, 8]. We recently
Author contributions: V.L.B.: conception and design, collection and assembly of data, data analysis and interpretation, manuscriptwriting; K.E.: collection and assembly of data, data analysis and interpretation, manuscript writing; B.G.H.: collection and assembly ofdata; Y.S.: collection and assembly of data; F.C.M.: collection and assembly of data, provision of study material; A.A.: collection andassembly of data, provision of study material; R.W.: collection and assembly of data, provision of study material; C.B.: provision ofstudy material; M.A.: conception and design, data analysis and interpretation, financial support, final approval of the manuscript;S.A.M.: conception and design, data analysis and interpretation, financial support, manuscript writing, final approval of the manuscript.
#Correspondence: Sendurai A. Mani PhD, (email:[email protected]) or Michael Andreeff M.D., Ph.D., (e-mail: [email protected]), 1515 Holcombe Blvd, Houston, TX, 77030; Received October 26, 2009; accepted for publication June 8, 2010; firstpublished online in STEM CELLS EXPRESS June 22, 2010. VC AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.467
STEM CELLS 2010;28:1435–1445 www.StemCells.com

demonstrated that induction of EMT by TGF-b1, Snail, orTwist in immortalized human mammary epithelial cells(HMECs) results in the acquisition of stem cell characteristics[9]. These characteristics include the ability to form spheresin nonadherent culture [10] and a shift from the more differ-entiated mammary epithelial cell surface marker profile(CD44lowCD24high) to the antigenic phenotype associated withmammary stem cells (CD44highCD24low) [9, 11]. However,these studies only began to delineate the molecular similar-ities between EMT-derived cells and stem cell populationsand did not address the full lineage differentiation potentialacquired by EMT-derived cells.
Mesenchymal stem cells (MSCs) are a small populationof cells within the mesenchymal stromal cell compartmentthat have the capacity to self-renew and differentiate intomultiple cell lineages including three major mesodermal line-ages: osteoblasts, adipocytes, and chondrocytes [12–14].MSCs have been identified and propagated from various adultand fetal tissues including the bone marrow, adipose tissue,umbilical cord, human term placenta, and endometrium [15–19]. MSCs express a panel of cell surface antigens, includingCD105 (Endoglin), CD73 (Ecto-50-nucleotidase), CD44 (Hyal-uronic acid receptor), CD140b (platelet-derived growth factorreceptor-b [PDGFR-b]), CD90 (Thy-1), and are negative formarkers of the hematopoietic lineage, for example, CD45[20–23]. In addition, MSCs have been shown to engraft intowounds and damaged tissues [24, 25].
Herein, we report that EMT-derived cells have a func-tional resemblance to MSCs derived from human bonemarrow, including a similar antigenic phenotype, the abilityto differentiate into multiple cell lineages, and the potential tohome to tumor cells in vitro and wounds in vivo.
METHODS
Isolation and Culture of Primary MSCs
MSCs were isolated from the bone marrow of healthy donorswho were undergoing bone marrow harvest for use in allo-geneic bone marrow transplantation. All bone marrow donors
provided written informed consent, and this study wasconducted according to institutional guidelines under anapproved protocol. Bone marrow was subjected to centrifuga-tion (700g for 15 minutes at 4�C) over a Ficoll–Hypaque gra-dient (Sigma, St. Louis, MO) to separate mononuclear cells.After centrifugation, the buffy coat layer was carefullyextracted and resuspended in a-minimal essential mediumcontaining 20% fetal bovine serum (Gibco BRL, Rockville,MD), L-glutamine, and penicillin–streptomycin (Flow Labora-tories, Rockville, MD) and plated at an initial density of 1 �106 cells/cm2. After 3 days, the cultures were washed withphosphate-buffered saline (PBS), and the remaining adherentcells were cultured until �80% confluence. The cells werethen subcultured at densities of 5,000–6,000 cells/cm2. Thethird or fourth passage was used for the experiments.
HMECs Culture and Generation ofEMT-Derived Cells
The HMECs were transduced and maintained as previouslydescribed [9, 26]. In brief, HMECs obtained from Cloneticswere immortalized with the catalytic subunit of human telo-merase and SV40 Large T antigen. These cells were thentransduced with either pBabe-puro retroviral vector or pBabe-puro vectors expressing Twist, Snail, or TGF-b1. Bright-fieldphotographs were taken with a Nikon Coolpix 950 cameraattached to a Nikon TMS light microscope (Nikon Instru-ments Inc., Melville, NY). Isolation, culturing, and infectionof primary HMECs are described in [27].
Reverse Transcription Polymerase ChainReaction Analysis
Total RNA extraction and real-time reverse transcriptionpolymerase chain reaction (RT-PCR) were performed as pre-viously reported [9] using ABI7900 real-time PCR machine.The primer sets used for detection of EMT-associated geneswere previously described [9]. The primers used in the differ-entiation studies are listed in Supporting Information Table 1.
Flow Cytometry
HMECs stably expressing Twist, Snail, TGF-b1 or empty vec-tor or MSCs were trypsinized, washed once with PBS, once
Table 1. Cell surface marker expression of EMT-derived cells and MSCs using flow cytometry
Gene name Alternate names Distribution Twist Snail TGF-b MSC
CD10 Neprilysin B, T precursors þ þ þ þCD11C Integrin, alpha X Neutrophils, dendritic cells � � � �CD14 LPS-R Monocytes, macrophages � � � �CD24 BA-1, HSA Thymocytes, erythrocytes �/low �/low �/low �CD44 H-CAM Fibroblasts þ þ þ þCD45 PTPRC Hematopoietic cells � � � �CD73 NT5E Epithelial cells, smooth muscle cells þ þ þ þCD90 Thy-1 Fibroblasts, neurons þ þ þ þCD105 Endoglin Endothelial cells þ þ þ þCD106 VCAM-1 Activated endothelial cells þ þ þ þCD140b PDGFRb Fibroblasts, smooth muscle cells þ þ þ þCD166 ALCAM Neurons, fibroblasts þ þ þ þ5 � 105 HMECs stably expressing Twist, Snail, or TGFb1 and MSCs were incubated with the following antibody conjugates: CD44-APC,CD90-PE, CD105-PE, and CD10-PE, CD11c-PE, CD14-PE, CD-24-FITC, CD45-FITC, CD73-PE, CD106-PE, CD140b-PE, CD166-PE.After staining, the cells were analyzed on a LSR-II Flow cytometer. Ten thousand events were acquired for each sample. The flow-cytometric data analysis was performed using FCS Express software. Alternate names and marker distribution is included in a separatecolumn for each marker.Abbreviations: ALCAM, activated leukocyte cell adhesion molecule; EMT, epithelial-to-mesenchymal transition; H-CAM, homing-cellularadhesion molecule; HMEC, human mammary epithelial cell; HSA, heat stable antigen; LPS-R, lipopolysaccharide receptor; MSC,mesenchymal stem cell, NT5E, 50-nucleotidase-ecto; PDGFRb, platelet-derived growth factor receptor-b; PTPRC, protein-tyrosinephosphatase receptor-type C; TGFb1, transforming growth factor-b1; VCAM-1, vascular cell adhesion molecule-1.
1436 EMT Generates MSC-Like Cells

with PBS containing 4% fetal bovine serum (FBS) (FACSbuffer), and then incubated in fluorescence activated cell sort-ing (FACS) buffer for 15 minutes on ice. Live cells (5 � 105)were then incubated with 1 lg of fluorochrome-conjugatedantibodies in 100 ll reaction volume for 15 minutes. The fol-lowing antibody conjugates were used: CD44-APC, CD90-PE,CD105-PE (eBiosciences, San Diego, CA) and CD10-PE,CD11c-PE, CD14-PE, CD-24-FITC, CD45-FITC, CD73-PE,CD106-PE, CD140b-PE, CD166-PE (BD Bioscience, SanJose, CA). After incubation, the cells were washed withFACS buffer containing 0.5 lg/ml 40,6-diamidino-2-phenylin-dole (DAPI) and analyzed on a LSR-II Flow Cytometer (BDBiosciences). A total of 10,000 events were acquired for eachsample. The flow-cytometric data analysis was performedusing FCS Express software (De novo software, Los Angeles,CA).
Multilineage Differentiation
Osteoblast Differentiation. Ten thousand HMECs express-ing the empty vector, Twist, or Snail as well as MSCs werecultured in NH OsteoDiff (Miltenyi Biotec, Auburn, CA)media for 21 days. The medium was replaced every 3 days.After 21 days, the cells were washed twice with PBS andfixed with 4% paraformaldehyde (PFA). To determine alka-line phosphatase (AP) activity, cells were incubated withFAST BCIP/NBT substrate (Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) for 20 minutes at room temperature.Calcium deposition was analyzed by staining with 1% alizarinred S (Spectrum, Gardena, CA) for 5 minutes at room temper-ature. Mineral deposition was determined by Von-Kossa stain-ing, which was performed by incubating fixed cells with 1%silver nitrate (Sigma-Aldrich) under bright light at room tem-perature for 30 minutes. For the gene expression analysis dur-ing osteoblast differentiation, total mRNA was collected fromcells which were grown in osteoblast differentiation mediumfor either 0, 5, or 10 days during the course of differentiation.Real-time RT-PCR was performed on these samples asdescribed earlier for AP and osteocalcin (OC) using theprimers described in Supporting Information Table 1.
Adipocyte Differentiation. To test differentiation into adipo-cytes, the respective cells types were cultured in NH Adipo-Diff medium (Miltenyi Biotec). In brief, 2 � 104 of HMECsexpressing the empty vector, Twist, or Snail as well as MSCswere cultured in NH AdipoDiff
VRmedium in a 12-well cell
culture dish. The medium was replaced every 3 days. After28 days of culture, the formation of adipocytes was evaluatedfixing cells with 4% PFA and staining with oil red O dye(Sigma-Aldrich) for 15 minutes at RT. Alternatively, oil drop-lets were also stained using LipidTox, a fluorescent lipid dyefrom Invitrogen. Photographs were taken either by using aNikon Coolpix 950 camera attached to Nikon TMS lightmicroscope (Nikon Instruments Inc.) or by Hamamatsu-C4742-95 camera (Hamamtsu, Bridgewater, NJ) attached toOlympus IX-51 inverted fluorescent microscope (OlympusAmerica Inc, Center Valley, PA). For the gene expressionanalysis during adipocyte differentiation, total mRNA wascollected from cells which were grown in adipocyte differen-tiation medium for either 0, 5, or 10 days during the courseof differentiation. Real-time RT-PCR was performed on thesesamples as described above for peroxisome proliferator-acti-vated receptor-gamma (PPAR-c) and lipoprotein lipase (LPL)using the primers described in Supporting InformationTable 1.
Chondrogenic Differentiation. To induce differentiationinto the chondrocyte lineage, the cells were incubated inChondroDiff medium (Miltenyi Biotec). Briefly, 4 � 105 ofHMECs expressing the empty vector, Twist, or Snail as wellas MSCs were washed once with PBS and the cell pelletswere cultured in 1 ml of ChodroDiff medium in 15 ml falcontubes for 21 days at 37�C. The medium was replaced every3 days. After incubation, the resulting cell pellets were fixedwith 3.7% formalin, embedded in paraffin, and cut into 5-lm-thick sections. Following deparaffinization and hydratation,the sections were incubated with Alcian Blue 8GX solution(Sigma-Aldrich) for 30 minutes at room temperature. Theslides were then washed in 3% acetic acid and then indistilled water. After washing, the slides were counterstained with nuclear fast red (Sigma) for 5 minutes and thenwashed with distilled water. Photographs were taken usingHamamatsu-C4742-95 camera (Hamamatsu) attached to Olym-pus BX41 microscope (Olympus America Inc).
Alternatively, the sections were immune-stained withcollagen-I or collagen-II antibody (Abcam, Cambridge, MA)as described earlier using paraffin-embedded tissue sections[28]. Briefly, after blocking with specific blocking buffer(Dako, Carpinteria, CA), the sections were then incubatedwith rabbit anti-human collagen-I or collagen II polyclonalantibody (Abcam) overnight at 4�C. The sections werewashed three times with phosphate buffer saline with tween(PBST) and then incubated with biotinylated secondary anti-body and horseradish peroxidase (HRP)-conjugated streptavi-din complex, as described in the manufacturer’s instructions(Dako). After washing the sections with wash buffer, thestaining was visualized using 3,30-diaminobenzidine. Thepictures were taken with Olympus DP-70 camera attached toOlympus BX-41 microscope (Olympus America Inc.)
Wound Homing Assay
Luciferase Labeling Using Adenoviral Vector Transduction. Arecombinant adenoviral (Ad) vector expressing firefly luci-ferase (ffLuc) and possessing an RGD-modified fiber (AdLuc-F/RGD) was prepared, purified, and tittered as previouslydescribed [29]. HMECs expressing the empty vector, Twist,or Snail as well as MSCs were incubated for 4 hours, in se-rum-free medium with 2,000 AdLuc-F/RGD viral particlesper cell. The transduced cells were assessed for luciferaseexpression by plating 5 � 104 cells into 24-well plates andadding 40 lg of D-Luciferin (Caliper Life Sciences, Hopkin-ton, MA) into 2 ml culture medium. After 30 seconds, thecells were placed into the imager for detection. Using thismultiplicity of infection protocol, we routinely detected >500copies of Ad-delivered ffLuc transcript/cell by quantitativeRT-PCR, and bioluminescence could be detected for up to30 days (data not shown).
In vivo Wound Homing Model in Nude Mice. Nude mice(NU/J, The Jackson laboratory, Bar Harbor, ME, http://www.jax.org) were housed according to institutional standardsand treated with approved protocols. To generate thewounding model, mice were first anesthetized and then 4-mmdiameter punch biopsies were made on the dorsal side of theanimals using Biopsy Punch (4.0 mm, HealthLink, http://www.healthlinkinc.net/). To analyze the wound homingability, 3 � 105 luciferase-labeled cells were transplantedthrough the tail vain on the day of wounding. The mice wereimaged after 3 days as described below. All mouse work wasperformed following institutional approved protocols.
Battula, Evans, Hollier et al. 1437
www.StemCells.com

Bioluminescent In Vivo Imaging. In vivo optical imagingwas performed with a Xenogen IVIS bioluminescence/fluores-cence optical imaging system (Caliper Life Sciences [Xeno-gen], Hopkinton, MA). Five minutes prior to imaging, eachmouse was given an intraperitoneal injection of D-luciferin (ata dose of 125 mg/kg or a 100 Rl injection of 40 mg/ml coe-lenterazine) as described previously. General anesthesia wasgiven (5% isoflurane [IsoSol, Medeva Pharmaceutical PA,Inc.]), and the mice were placed in the light-tight-heatedchamber. Anterior and posterior luminescent images wereacquired with 1- to 3-minute exposure times. Optical imageswere displayed and analyzed with IVIS Living Image (CaliperLife Sciences [Xenogen], Hopkinton, MA) software packages.
In Vitro Matrigel Invasion Assay
An in vitro Matrigel invasion assay was performed using 24-well Biocoat Matrigel Invasion Chambers containing BD Fal-con Cell Culture Inserts with 8-lm-diameter pore size PETmembrane that has been treated with Matirgel Matrix (BDBiosciences). The MSCs were serum-starved for 24 hoursbefore the assay to avoid any receptor blocking factors fromserum. There was no such treatment for HMEC cells, as theyare cultured in serum free medium. In addition, 5 � 104
MDA-MB-231 cells were plated into 24-well cell culturedishes. To perform the assay, at first the inserts containingMatrigel were hydrated using 500 ll of warm culture mediumwithout serum at 37�C for 2 hours. After hydration, themedium was removed from the chambers and the inserts wereplaced on top of each well of the MDA-MB-231 cell contain-ing culture dish. In case of platelet-derived growth factor-bb(PDGF-bb)-mediated invasion assays, medium in the bottomchamber was replaced with fresh medium with or without10 ng/ml PDGF-bb ligand (Peprotech, Rocky Hill, NJ, http://www.peprotech.com). Cell suspensions of vector, Twist, orSnail overexpressing HMEC or MSCs were adjusted to aconcentration of 15 � 104 cells/ml and, and 200 ll of theadjusted cell suspension (3 � 104 cells/insert) was immedi-ately placed in the Matrigel-coated upper chamber. Afterincubation at 37�C for 36 hours in a 5% CO2 incubator, theresidual cells on the upper surface of the filter werecompletely removed with cotton swabs. The membranes werethen stained with HEMA-3 hematoxylin solution (FisherScientific Company L.L.C., Kalamazoo, MI), and the invasivepotential of the cells was determined by counting the numberof cells that had invaded to the lower surface of the filter in10 different areas under a inverted light microscope (OlympusBX41). Each assay was performed in triplicate in threeseparate experiments.
Gene Chip Data Analysis and Statistical Methods
Gene chip analysis was performed in triplicate to compare thegene expression changes among the HMECs expressing thecontrol vector, or Snail, with that of MSCs. Prior to statisticalanalysis of gene expression, probe sets were matched betweenthe two generations of Affymetrix chips, HT_HG_U133A andHG_U133_Plus_2. The former chip is a subset of the latterchip and, therefore, the analysis of differential expression wasrestricted to the probe sets on the HT_HG_U133A chip. Probesets between the HT_HG_U133A chip and the HG_U133_Plus_2 chip were matched using the function read.affybatch.hybrid from the CustomCDF R package [30]. A total of22,268 probesets representing 13,294 genes were matched andanalyzed.
The statistical analysis of the Affymetrix CEL data wasperformed using the bioconductor [31] statistical softwareenvironment. To assess differential expression, the intensityvalues in the CEL files were transformed to robust multichip
average (RMA) expression measures [32]. The RMA measureis given in a log base two scale. Using the Limma R package,we fit linear models with the lmFit function to simultaneouslyassess pair wise differential expression among the three RNAsources.
RESULTS
Comparison of EMT-Derived Cells with MSCs
Given that MSCs have mesenchymal features, including mor-phology (Fig. 1A), and exhibit stem cell characteristics [14],we sought to determine whether epithelial cells that haveundergone EMT share molecular and functional propertieswith MSCs. We first determined whether HMECs stablyexpressing Twist, Snail, or TGF-b1 exhibited properties ofMSCs by analyzing the expression of several cell surfacemarkers associated with MSCs in EMT-derived HMECs usingflow cytometry. We found that EMT-derived cells expressedseveral MSCs-associated cell surface markers including,CD10, CD44, CD73, CD90, CD105, CD106, and CD166(Table 1). Interestingly, CD140b (PDGFR-b), a prospectivemarker for isolation of MSCs from human bone-marrow [21],was highly expressed on the surface of EMT-derived HMECsbut not on the vector control HMECs (Fig. 1D). Importantly,both the EMT-derived HMECs and MSCs were negative forCD45, CD11c, and CD14, which are hematopoietic lineagemarkers (Fig. 1D, Table 1). These findings suggested thatmesenchymal-like cells generated from HMECs by EMT andMSCs express a similar set of cell surface antigens.
As EMT-derived HMECs exhibited a similar antigenicprofile to MSCs, we determined whether MSCs obtained fromhuman bone marrow express the mRNAs encoding markersassociated with the mesenchymal-like state generated byEMT. During EMT, epithelial cells lose expression of E-cad-herin and gain expression of mesenchymal-associated genes,such as N-cadherin, fibronectin, and vimentin [33]. Mostimportantly, we found that the MSCs also expressed lowerlevels of E-cadherin and higher levels of vimentin, fibronec-tin, and N-cadherin compared with control epithelial cells(Fig. 1B) [9]. Strikingly, the MSCs expressed high levels ofembryonic transcription factors known to regulate EMT (i.e.,Snail, Twist, Mesenchyme Forkhead 1 [FOXC2], Zeb1) (Fig.1C). Compared with HMECs, the MSCs also expressed highlevels of CD44 and low levels of CD24 at the cell surface,which was similar to the EMT-derived stem-like cells (Fig.1D).
Global gene expression analysis, using Affymetirx micro-arrays, revealed that the sets of genes differentially expressedbetween vector–control HMECs and either MSCs or Snail-induced EMT-derived HMECs were �70% similar. On theother hand, 15% of the genes analyzed were differentiallyexpressed between MSCs and Snail-induced EMT-derivedHMECs. For example, several cytokeratins are more highlyexpressed in EMT-derived cells compared with MSCs (Sup-porting Information Fig. 1B, Supporting Information 1 and 2),which indicated that MSCs and EMT-derived cells havehighly similar gene expression compared to the epithelial con-trol cells but that the Snail-induced HMECs still maintainsome characteristics of their epithelial origin.
Differentiation of EMT-Derived Cells intoMesodermal Lineages
As the EMT-derived HMECs and MSCs had similar cell surfacemarker profiles, morphologies, and gene expression profiles, we
1438 EMT Generates MSC-Like Cells

next investigated whether EMT-derived HMECs have MSCs-likemultipotency including the ability to differentiate into osteo-blasts, adipocytes, and chondrocytes. To test this, we first sub-jected the EMT-derived HMECs (Twist-/Snail-induced), alongwith vector-infected control epithelial cells, to an osteoblast dif-ferentiation assay. After 21 days, the cells were stained withFASTO BCIP/NBT substrate to analyze AP activity. Strikingly,the epithelial cells that had undergone EMT stained positive forAP, a marker of osteoblasts (Fig. 2A, left panel), similar toMSCs isolated from human bone marrow (Fig. 2A, right panel).Furthermore, both Snail and Twist-induced, EMT-derived cellsdisplayed a gradual increase AP activity, in a time-dependentmanner, from week 1–3 of osteoblast differentiation culture(Supporting Information Fig. 2).
To further test if the EMT-derived cells could differentiateinto functional osteoblasts capable of producing mineral depos-
its, cells were grown in osteoblast differentiation media andthen stained with either 1% alizarin red S or 2% silver nitrate(von Kossa staining), which stains calcium deposits. The EMT-derived cells stained positive for alizarin red S and von Kossareagent in a fashion comparable to bone marrow-derived MSCs(Fig. 2B, 2C). These findings showed that the EMT-derivedcells produce calcium in response to differentiation conditionssimilar to MSCs. However, both the vector–control cellsthat were subjected to differentiation condition (Fig. 2B, 2C)and the EMT-derived cells that were not subjected to osteo-genic differentiation were not capable of producing calcium(Supporting Information Fig. 3A). In addition, subjecting EMT-derived cells to an osteoblast differentiation assay increased thelevels of mRNAs encoding osteoblast related genes, OC (FoldChange: Snail, 3.69; Twist, 11.11) and AP (Fold Change: Snail,9.39; Twist, 1.03) after 5 days in the differentiation condition
Figure 1. Morphology and expression pattern of genes in epithelial-to-mesenchymal transition (EMT)-derived HMECs and bone marrowMSCs. (A): Light microscopic images (�10) of HMECs ectopically expressing Twist, Snail, TGF-b1, or the empty vector as well as bonemarrow-derived MSCs. (B, C): Quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis of EMT-associated genes includ-ing E-cadherin, vimentin, fibronectin, and N-cadherin (B) and EMT-regulating transcription factors, including FOXC2, Twist, Snail, and Zeb-1 inMSCs (C). (D): The expression of cell surface markers associated with MSCs in EMT-derived HMECs expressing Twist, Snail, TGF-b, or thecontrol vector. Quantitative RT-PCR was performed in triplicate (mean 6 SD). Abbreviations: TGF-b, transforming growth factor-b; FOXC2,mesenchyme forkhead 1; FSC, forward scatter; HMEC, human mammary epithelial cell; MSC, mesenchymal stem cell; Zeb1, zinc finger E-box-binding homeobox 1.
Battula, Evans, Hollier et al. 1439
www.StemCells.com
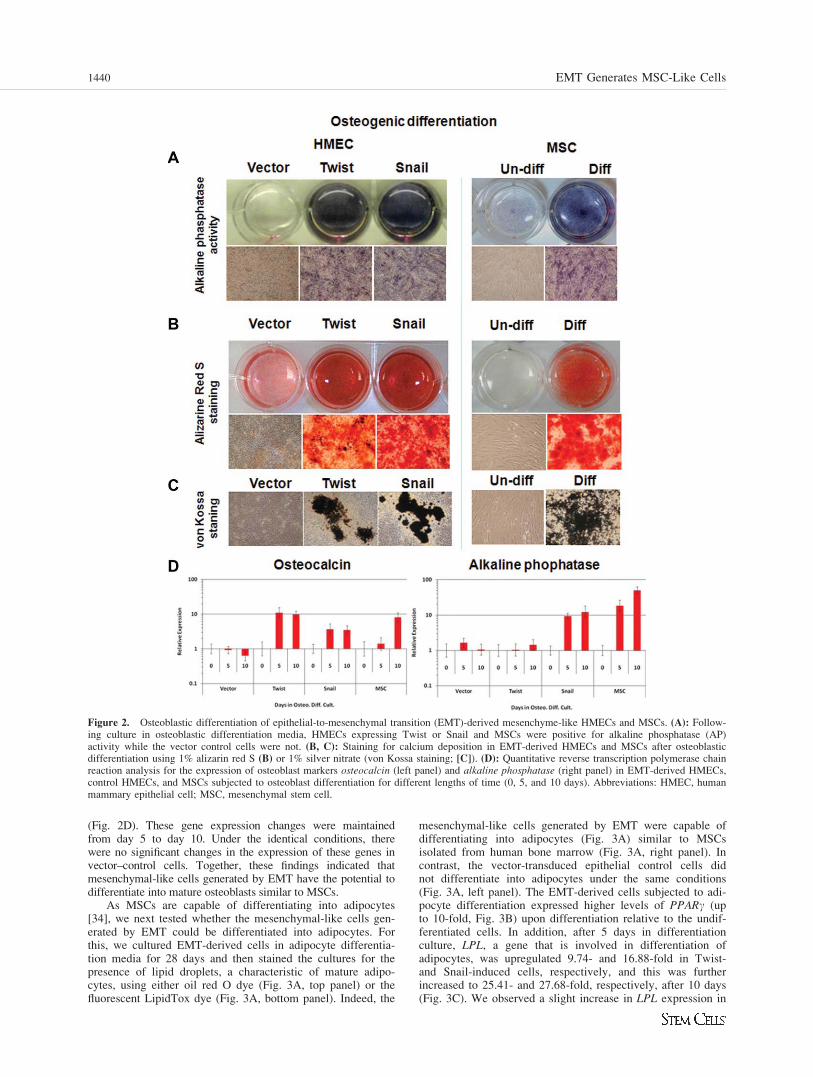
(Fig. 2D). These gene expression changes were maintainedfrom day 5 to day 10. Under the identical conditions, therewere no significant changes in the expression of these genes invector–control cells. Together, these findings indicated thatmesenchymal-like cells generated by EMT have the potential todifferentiate into mature osteoblasts similar to MSCs.
As MSCs are capable of differentiating into adipocytes[34], we next tested whether the mesenchymal-like cells gen-erated by EMT could be differentiated into adipocytes. Forthis, we cultured EMT-derived cells in adipocyte differentia-tion media for 28 days and then stained the cultures for thepresence of lipid droplets, a characteristic of mature adipo-cytes, using either oil red O dye (Fig. 3A, top panel) or thefluorescent LipidTox dye (Fig. 3A, bottom panel). Indeed, the
mesenchymal-like cells generated by EMT were capable ofdifferentiating into adipocytes (Fig. 3A) similar to MSCsisolated from human bone marrow (Fig. 3A, right panel). Incontrast, the vector-transduced epithelial control cells didnot differentiate into adipocytes under the same conditions(Fig. 3A, left panel). The EMT-derived cells subjected to adi-pocyte differentiation expressed higher levels of PPARc (upto 10-fold, Fig. 3B) upon differentiation relative to the undif-ferentiated cells. In addition, after 5 days in differentiationculture, LPL, a gene that is involved in differentiation ofadipocytes, was upregulated 9.74- and 16.88-fold in Twist-and Snail-induced cells, respectively, and this was furtherincreased to 25.41- and 27.68-fold, respectively, after 10 days(Fig. 3C). We observed a slight increase in LPL expression in
Figure 2. Osteoblastic differentiation of epithelial-to-mesenchymal transition (EMT)-derived mesenchyme-like HMECs and MSCs. (A): Follow-ing culture in osteoblastic differentiation media, HMECs expressing Twist or Snail and MSCs were positive for alkaline phosphatase (AP)activity while the vector control cells were not. (B, C): Staining for calcium deposition in EMT-derived HMECs and MSCs after osteoblasticdifferentiation using 1% alizarin red S (B) or 1% silver nitrate (von Kossa staining; [C]). (D): Quantitative reverse transcription polymerase chainreaction analysis for the expression of osteoblast markers osteocalcin (left panel) and alkaline phosphatase (right panel) in EMT-derived HMECs,control HMECs, and MSCs subjected to osteoblast differentiation for different lengths of time (0, 5, and 10 days). Abbreviations: HMEC, humanmammary epithelial cell; MSC, mesenchymal stem cell.
1440 EMT Generates MSC-Like Cells

the vector control cells at day 10 but to a lesser extent thanEMT-derived cells and MSCs. Importantly, the parentalEMT-derived cells did not stain positive for adipocytes differ-entiation markers prior to adipocyte differentiation culture(Supporting Information Fig. 3B). This evidence suggested
that EMT-derived mesenchymal-like cells have the potentialto differentiate into adipocytes.
Another well-known property of MSCs is the ability todifferentiate into chondrocytes, which are the specialized cellsin cartilage tissues that produce and maintain the collagen and
Figure 3. Adipogenic and chondrogenic differentiation of epithelial-to-mesenchymal transition (EMT)-derived HMECs and MSCs. (A): Followingadipogenic differentiation, the EMT-derived HMECs and MSCs stained positive with oil red O dye (top) and fluorescent LipidTox, which stains oildroplets (bottom). Conversely, vector control HMECs did not stain using similar treatment (right panel). (B): Quantitative reverse transcription poly-merase chain reaction (RT-PCR) analysis for the expression of the adipocyte marker PPARc after 21 days in adipocyte differentiation condition.(C): Quantitative RT-PCR analysis for the expression of LPL at different time points as indicated in the figure in EMT-derived HMECs, controlHMECs and MSCs subjected to adipocyte differentiation. Quantitative RT-PCR was performed in triplicate (mean 6 SD). (D): Chondrocytic nod-ules formed by EMT-derived HMECs or MSCs stained positive with alcian blue 8GX (top panel). These sections were counter stained with nuclearfast red solution. Immunohistochemistry was performed on chondrocyte sections using collagen-I antibody (middle panel) and collagen-II (bottompanel). The vector-infected HMECs did not form any chondrocytes nodules under identical conditions. Abbreviations: HMEC, human mammary epi-thelial cell; LPL, lipoprotein lipase; MSC, mesenchymal stem cell; PPARc, peroxisome proliferator-activated receptor-gamma.
Battula, Evans, Hollier et al. 1441
www.StemCells.com
proteoglycan [35]. To test whether EMT-derived cells candifferentiate into chondrocytes, we cultured the cells that hadundergone EMT due to ectopic expression of Twist or Snailin chondrocyte differentiation medium. After 21 days in sus-pension culture, the EMT-derived HMECs formedchondrocyte nodules similar to MSCs. In contrast, the vector–control cells did not form any chondrocyte nodules underidentical culture conditions. To further confirm chondrocytedifferentiation, we fixed, sectioned, and stained the chondro-cyte nodules with alcian blue reagent to detect acid mucoussubstances and acetic mucins. We found that these nodulesstained positive for alcian blue (Fig. 3D, top panel), whichrevealed that the EMT-derived HMECs can differentiate intochondrocytes. Furthermore, the chondrocytes derived fromEMT-derived cells and MSCs were positive for collagen I(Fig. 3D, middle panel), and weakly positive for collagen II(Fig. 3D, bottom panel). Together, these findings indicate thatthe mesenchymal-like cells generated by EMT are multipotentand have the potential to differentiate into the three majormesodermal lineages.
Invasion of EMT-Derived Cells Towards PDGF-bb
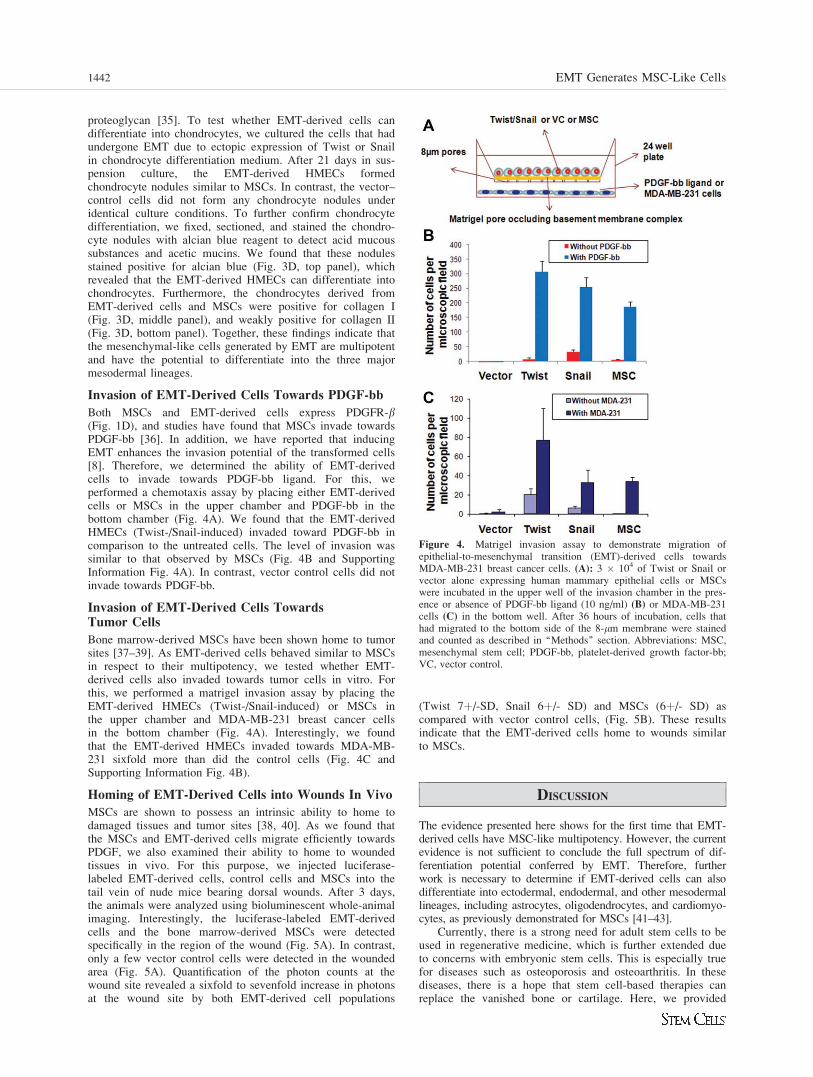
Both MSCs and EMT-derived cells express PDGFR-b(Fig. 1D), and studies have found that MSCs invade towardsPDGF-bb [36]. In addition, we have reported that inducingEMT enhances the invasion potential of the transformed cells[8]. Therefore, we determined the ability of EMT-derivedcells to invade towards PDGF-bb ligand. For this, weperformed a chemotaxis assay by placing either EMT-derivedcells or MSCs in the upper chamber and PDGF-bb in thebottom chamber (Fig. 4A). We found that the EMT-derivedHMECs (Twist-/Snail-induced) invaded toward PDGF-bb incomparison to the untreated cells. The level of invasion wassimilar to that observed by MSCs (Fig. 4B and SupportingInformation Fig. 4A). In contrast, vector control cells did notinvade towards PDGF-bb.
Invasion of EMT-Derived Cells TowardsTumor Cells
Bone marrow-derived MSCs have been shown home to tumorsites [37–39]. As EMT-derived cells behaved similar to MSCsin respect to their multipotency, we tested whether EMT-derived cells also invaded towards tumor cells in vitro. Forthis, we performed a matrigel invasion assay by placing theEMT-derived HMECs (Twist-/Snail-induced) or MSCs inthe upper chamber and MDA-MB-231 breast cancer cellsin the bottom chamber (Fig. 4A). Interestingly, we foundthat the EMT-derived HMECs invaded towards MDA-MB-231 sixfold more than did the control cells (Fig. 4C andSupporting Information Fig. 4B).
Homing of EMT-Derived Cells into Wounds In Vivo
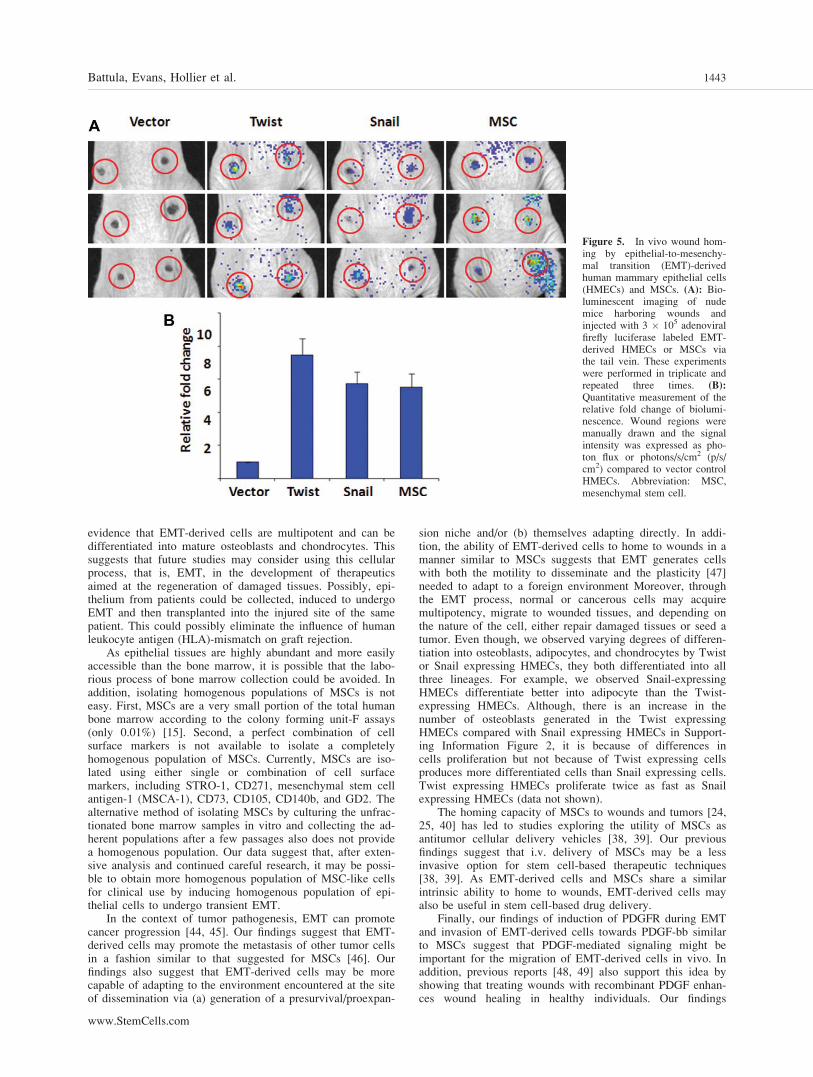
MSCs are shown to possess an intrinsic ability to home todamaged tissues and tumor sites [38, 40]. As we found thatthe MSCs and EMT-derived cells migrate efficiently towardsPDGF, we also examined their ability to home to woundedtissues in vivo. For this purpose, we injected luciferase-labeled EMT-derived cells, control cells and MSCs into thetail vein of nude mice bearing dorsal wounds. After 3 days,the animals were analyzed using bioluminescent whole-animalimaging. Interestingly, the luciferase-labeled EMT-derivedcells and the bone marrow-derived MSCs were detectedspecifically in the region of the wound (Fig. 5A). In contrast,only a few vector control cells were detected in the woundedarea (Fig. 5A). Quantification of the photon counts at thewound site revealed a sixfold to sevenfold increase in photonsat the wound site by both EMT-derived cell populations
(Twist 7þ/-SD, Snail 6þ/- SD) and MSCs (6þ/- SD) ascompared with vector control cells, (Fig. 5B). These resultsindicate that the EMT-derived cells home to wounds similarto MSCs.
DISCUSSION
The evidence presented here shows for the first time that EMT-derived cells have MSC-like multipotency. However, the currentevidence is not sufficient to conclude the full spectrum of dif-ferentiation potential conferred by EMT. Therefore, furtherwork is necessary to determine if EMT-derived cells can alsodifferentiate into ectodermal, endodermal, and other mesodermallineages, including astrocytes, oligodendrocytes, and cardiomyo-cytes, as previously demonstrated for MSCs [41–43].
Currently, there is a strong need for adult stem cells to beused in regenerative medicine, which is further extended dueto concerns with embryonic stem cells. This is especially truefor diseases such as osteoporosis and osteoarthritis. In thesediseases, there is a hope that stem cell-based therapies canreplace the vanished bone or cartilage. Here, we provided
Figure 4. Matrigel invasion assay to demonstrate migration ofepithelial-to-mesenchymal transition (EMT)-derived cells towardsMDA-MB-231 breast cancer cells. (A): 3 � 104 of Twist or Snail orvector alone expressing human mammary epithelial cells or MSCswere incubated in the upper well of the invasion chamber in the pres-ence or absence of PDGF-bb ligand (10 ng/ml) (B) or MDA-MB-231cells (C) in the bottom well. After 36 hours of incubation, cells thathad migrated to the bottom side of the 8-lm membrane were stainedand counted as described in ‘‘Methods’’ section. Abbreviations: MSC,mesenchymal stem cell; PDGF-bb, platelet-derived growth factor-bb;VC, vector control.
1442 EMT Generates MSC-Like Cells
evidence that EMT-derived cells are multipotent and can bedifferentiated into mature osteoblasts and chondrocytes. Thissuggests that future studies may consider using this cellularprocess, that is, EMT, in the development of therapeuticsaimed at the regeneration of damaged tissues. Possibly, epi-thelium from patients could be collected, induced to undergoEMT and then transplanted into the injured site of the samepatient. This could possibly eliminate the influence of humanleukocyte antigen (HLA)-mismatch on graft rejection.
As epithelial tissues are highly abundant and more easilyaccessible than the bone marrow, it is possible that the labo-rious process of bone marrow collection could be avoided. Inaddition, isolating homogenous populations of MSCs is noteasy. First, MSCs are a very small portion of the total humanbone marrow according to the colony forming unit-F assays(only 0.01%) [15]. Second, a perfect combination of cellsurface markers is not available to isolate a completelyhomogenous population of MSCs. Currently, MSCs are iso-lated using either single or combination of cell surfacemarkers, including STRO-1, CD271, mesenchymal stem cellantigen-1 (MSCA-1), CD73, CD105, CD140b, and GD2. Thealternative method of isolating MSCs by culturing the unfrac-tionated bone marrow samples in vitro and collecting the ad-herent populations after a few passages also does not providea homogenous population. Our data suggest that, after exten-sive analysis and continued careful research, it may be possi-ble to obtain more homogenous population of MSC-like cellsfor clinical use by inducing homogenous population of epi-thelial cells to undergo transient EMT.
In the context of tumor pathogenesis, EMT can promotecancer progression [44, 45]. Our findings suggest that EMT-derived cells may promote the metastasis of other tumor cellsin a fashion similar to that suggested for MSCs [46]. Ourfindings also suggest that EMT-derived cells may be morecapable of adapting to the environment encountered at the siteof dissemination via (a) generation of a presurvival/proexpan-
sion niche and/or (b) themselves adapting directly. In addi-tion, the ability of EMT-derived cells to home to wounds in amanner similar to MSCs suggests that EMT generates cellswith both the motility to disseminate and the plasticity [47]needed to adapt to a foreign environment Moreover, throughthe EMT process, normal or cancerous cells may acquiremultipotency, migrate to wounded tissues, and depending onthe nature of the cell, either repair damaged tissues or seed atumor. Even though, we observed varying degrees of differen-tiation into osteoblasts, adipocytes, and chondrocytes by Twistor Snail expressing HMECs, they both differentiated into allthree lineages. For example, we observed Snail-expressingHMECs differentiate better into adipocyte than the Twist-expressing HMECs. Although, there is an increase in thenumber of osteoblasts generated in the Twist expressingHMECs compared with Snail expressing HMECs in Support-ing Information Figure 2, it is because of differences incells proliferation but not because of Twist expressing cellsproduces more differentiated cells than Snail expressing cells.Twist expressing HMECs proliferate twice as fast as Snailexpressing HMECs (data not shown).
The homing capacity of MSCs to wounds and tumors [24,25, 40] has led to studies exploring the utility of MSCs asantitumor cellular delivery vehicles [38, 39]. Our previousfindings suggest that i.v. delivery of MSCs may be a lessinvasive option for stem cell-based therapeutic techniques[38, 39]. As EMT-derived cells and MSCs share a similarintrinsic ability to home to wounds, EMT-derived cells mayalso be useful in stem cell-based drug delivery.
Finally, our findings of induction of PDGFR during EMTand invasion of EMT-derived cells towards PDGF-bb similarto MSCs suggest that PDGF-mediated signaling might beimportant for the migration of EMT-derived cells in vivo. Inaddition, previous reports [48, 49] also support this idea byshowing that treating wounds with recombinant PDGF enhan-ces wound healing in healthy individuals. Our findings
Figure 5. In vivo wound hom-ing by epithelial-to-mesenchy-mal transition (EMT)-derivedhuman mammary epithelial cells(HMECs) and MSCs. (A): Bio-luminescent imaging of nudemice harboring wounds andinjected with 3 � 105 adenoviralfirefly luciferase labeled EMT-derived HMECs or MSCs viathe tail vein. These experimentswere performed in triplicate andrepeated three times. (B):
Quantitative measurement of therelative fold change of biolumi-nescence. Wound regions weremanually drawn and the signalintensity was expressed as pho-ton flux or photons/s/cm2 (p/s/cm2) compared to vector controlHMECs. Abbreviation: MSC,mesenchymal stem cell.
Battula, Evans, Hollier et al. 1443
www.StemCells.com

suggest this effect may be mediated by the recruitment ofboth MSCs and EMT-derived cells from the near proximityvia PDGF-mediated signaling.
CONCLUSION
Collectively, these findings indicate that EMT-derived cellsnot only express markers that are relevant to MSCs but alsoexhibit functional similarities to MSCs. These EMT-derivedcells are capable of differentiating into mature osteoblasts,adipocytes, and chondrocytes similar to MSCs obtained fromhuman bone marrow. Interestingly, EMT-derived cells migratetowards wounds in vivo and to tumor cells in vitro similar toMSCs. In this report, we show, for the first time, that EMT-derived cells behaved functionally and phenotypically similarto MSCs, a subpopulation of adult stem cells.
ACKNOWLEDGMENTS
We are grateful to Robert Weinberg for his support, reagentsand critical reading of the article. We also thank Nathalie
Sphyris, Joe Taube, and Agata Tinnirello for critical readingof the article and Sreedevi Kumar for the technical help. Weare indebted to anonymous reviewers for their helpful sug-gestions, which improved the article significantly. Thisresearch was supported in part by the M.D. Anderson CancerCenter Research Trust and The V Foundation (to S.A.M.);grants from the National Cancer Institute (RC1CA146381,CA-109451, and CA-116199 [to F.C.M.], CA-55164, CA-16672, and CA-49639 [to M.A.]); and by the Paul and MaryHaas Chair in Genetics (to M.A.).
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
S.A.M., K.W.E., V.L.B., and M.A. are inventors of a patentapplication in part based on findings described in this article.The other authors have no potential conflicts of interest. S.A.M.owns stock, acted as a consultant, and received money withinthe last 2 years from SathGen Biotech. He also served as anofficer or member of the Board within the last 2 years forTheracrine.
REFERENCES
1 Hay ED. An overview of epithelio-mesenchymal transformation. ActaAnat (Basel) 1995;154:8–20.
2 Kalluri R, Neilson EG. Epithelial-mesenchymal transition and itsimplications for fibrosis. J Clin Invest 2003;112:1776–1784.
3 Shook D, Keller R. Mechanisms, mechanics and function of epithe-lial-mesenchymal transitions in early development. Mech Dev 2003;120:1351–1383.
4 Thiery JP. Epithelial-mesenchymal transitions in development andpathologies. Curr Opin Cell Biol 2003;15:740–746.
5 Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mes-enchymal transitions. Nat Rev Mol Cell Biol 2006;7:131–142.
6 Duband JL, Monier F, Delannet M et al. Epithelium-mesenchymetransition during neural crest development. Acta Anat (Basel) 1995;154:63–78.
7 Nawshad A, LaGamba D, Hay ED. Transforming growth factor beta(TGFbeta) signalling in palatal growth, apoptosis and epithelial mes-enchymal transformation (EMT). Arch Oral Biol 2004;49:675–689.
8 Yang J, Mani SA, Donaher JL et al. Twist, a master regulator of mor-phogenesis, plays an essential role in tumor metastasis. Cell 2004;117:927–939.
9 Mani SA, Guo W, Liao MJ et al. The epithelial-mesenchymal transi-tion generates cells with properties of stem cells. Cell 2008;133:704–715.
10 Dontu G, Al-Hajj M, Abdallah WM et al. Stem cells in normal breastdevelopment and breast cancer. Cell Prolif 2003;36(suppl 1):59–72.
11 Al-Hajj M, Wicha MS, Benito-Hernandez A et al. Prospective identifi-cation of tumorigenic breast cancer cells. Proc Natl Acad Sci USA2003;100:3983–3988.
12 Friedman MS, Long MW, Hankenson KD. Osteogenic differentiationof human mesenchymal stem cells is regulated by bone morphogeneticprotein-6. J Cell Biochem 2006;98:538–554.
13 in ’t Anker PS, Noort WA, Scherjon SA et al. Mesenchymal stemcells in human second-trimester bone marrow, liver, lung, and spleenexhibit a similar immunophenotype but a heterogeneous multilineagedifferentiation potential. Haematologica 2003;88:845–852.
14 Short B, Brouard N, Occhiodoro-Scott T et al. Mesenchymal stemcells. Arch Med Res 2003;34:565–571.
15 Battula VL, Bareiss PM, Treml S et al. Human placenta and bonemarrow derived MSC cultured in serum-free, b-FGF-containingmedium express cell surface frizzled-9 and SSEA-4 and give rise tomultilineage differentiation. Differ Res Biol Divers 2007;75:279–291.
16 Erices A, Conget P, Minguell JJ. Mesenchymal progenitor cells inhuman umbilical cord blood. Br J Haematol 2000;109:235–242.
17 Friedenstein AJ, Gorskaja JF, Kulagina NN. Fibroblast precursors innormal and irradiated mouse hematopoietic organs. Exp Hematol1976;4:267–274.
18 Morizono K, De Ugarte DA, Zhu M et al. Multilineage cells fromadipose tissue as gene delivery vehicles. Hum Gene Ther 2003;14:59–66.
19 Dimitrov R, Timeva T, Kyurkchiev D et al. Characterization of clono-genic stromal cells isolated from human endometrium. Reproduction2008;135:551–558.
20 Battula VL, Treml S, Abele H et al. Prospective isolation and charac-terization of mesenchymal stem cells from human placenta using afrizzled-9-specific monoclonal antibody. Differ Res Biol Divers 2008;76:326–336.
21 Buhring HJ, Battula VL, Treml S et al. Novel markers for the pro-spective isolation of human MSC. Ann N Y Acad Sci 2007;1106:262–271.
22 Caplan AI. Mesenchymal stem cells. J Orthop Res 1991;9:641–650.23 Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and
disease. Nat Rev Immunol 2008;8:726–736.24 Herdrich BJ, Lind RC, Liechty KW. Multipotent adult progenitor
cells: Their role in wound healing and the treatment of dermalwounds. Cytotherapy 2008;10:543–550.
25 Spees JL, Whitney MJ, Sullivan DE et al. Bone marrow progenitorcells contribute to repair and remodeling of the lung and heart in a ratmodel of progressive pulmonary hypertension. FASEB J 2008;22:1226–1236.
26 Elenbaas B, Spirio L, Koerner F et al. Human breast cancer cells gen-erated by oncogenic transformation of primary mammary epithelialcells. Genes Dev 2001;15:50–65.
27 Ayyanan A, Civenni G, Ciarloni L et al. Increased Wnt signaling trig-gers oncogenic conversion of human breast epithelial cells by aNotch-dependent mechanism. Proc Natl Acad Sci USA 2006;103:3799–3804.
28 Zeng Z, Shi YX, Samudio IJ et al. Targeting the leukemia microenvir-onment by CXCR4 inhibition overcomes resistance to kinase inhibi-tors and chemotherapy in AML. Blood 2009;113:6215–6224.
29 Klopp AH, Spaeth EL, Dembinski JL et al. Tumor irradiation increasesthe recruitment of circulating mesenchymal stem cells into the tumormicroenvironment. Cancer Res 2007;67:11687–11695.
30 Dai C, Liu J. Inducing pairwise gene interactions from time-seriesdata by EDA based Bayesian network. Conf Proc IEEE Eng Med BiolSoc 2005;7:7746–7749.
31 Huber W, Gentleman R. Matchprobes: A bioconductor package forthe sequence-matching of microarray probe elements. Bioinformatics2004;20:1651–1652.
32 Irizarry RA, Ooi SL, Wu Z et al. Use of mixture models in a microar-ray-based screening procedure for detecting differentially representedyeast mutants. Stat Appl Genet Mol Biol 2003;2:Article 1.
33 Zeisberg M, Neilson EG. Biomarkers for epithelial-mesenchymal tran-sitions. J Clin Invest 2009;119:1429–1437.
34 Kolf CM, Cho E, Tuan RS. Mesenchymal stromal cells. Biology ofadult mesenchymal stem cells: regulation of niche, self-renewal anddifferentiation. Arthritis Res Ther 2007;9:204.
1444 EMT Generates MSC-Like Cells

35 Mackay AM, Beck SC, Murphy JM et al. Chondrogenic differentia-tion of cultured human mesenchymal stem cells from marrow. TissueEng 1998;4:415–428.
36 Tokunaga A, Oya T, Ishii Y et al. PDGF receptor beta is a potent reg-ulator of mesenchymal stromal cell function. J Bone Miner Res 2008;23:1519–1528.
37 Spaeth E, Klopp A, Dembinski J et al. Inflammation and tumor micro-environments: Defining the migratory itinerary of mesenchymal stemcells. Gene Ther 2008;15:730–738.
38 Studeny M, Marini FC, Champlin RE et al. Bone marrow-derivedmesenchymal stem cells as vehicles for interferon-beta delivery intotumors. Cancer Res 2002;62:3603–3608.
39 Studeny M, Marini FC, Dembinski JL et al. Mesenchymal stemcells: Potential precursors for tumor stroma and targeted-deliveryvehicles for anticancer agents. J Natl Cancer Inst 2004;96:1593–1603.
40 Bittira B, Shum-Tim D, Al-Khaldi A et al. Mobilization and homingof bone marrow stromal cells in myocardial infarction. Eur J Cardio-thorac Surg 2003;24:393–398.
41 Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migratethroughout forebrain and cerebellum, and they differentiate into astro-cytes after injection into neonatal mouse brains. Proc Natl Acad SciUSA 1999;96:10711–10716.
42 Neuhuber B, Gallo G, Howard L et al. Reevaluation of in vitro differ-entiation protocols for bone marrow stromal cells: Disruption of actin
cytoskeleton induces rapid morphological changes and mimics neuro-nal phenotype. J Neurosci Res 2004;77:192–204.
43 Quevedo HC, Hatzistergos KE, Oskouei BN et al. Allogeneic mesen-chymal stem cells restore cardiac function in chronic ischemic cardio-myopathy via trilineage differentiating capacity. Proc Natl Acad SciUSA 2009;106:14022–14027.
44 Janda E, Lehmann K, Killisch I et al. Ras and TGFb cooperativelyregulate epithelial cell plasticity and metastasis: Dissection of Ras sig-naling pathways. J Cell Biol 2002;156:299–313.
45 Kiemer AK, Takeuchi K, Quinlan MP. Identification of genesinvolved in epithelial-mesenchymal transition and tumor progression.Oncogene 2001;20:6679–6688.
46 KarnoubAE, DashAB, VoAP et al.Mesenchymal stem cells within tumourstroma promote breast cancer metastasis. Nature 2007;449:557–563.
47 Savagner P, Kusewitt DF, Carver EA et al. Developmental transcrip-tion factor slug is required for effective re-epithelialization by adultkeratinocytes. J Cell Physiol 2005;202:858–866.
48 Bhattacharyya J, Mondal G, Madhusudana K et al. Single subcutane-ous administration of RGDK-lipopeptide:rhPDGF-B gene complexheals wounds in streptozotocin-induced diabetic rats. Mol Pharm2009;6:918–927.
49 Moore DC, Ehrlich MG, McAllister SC et al. Recombinant humanplatelet-derived growth factor-BB augmentation of new-bone forma-tion in a rat model of distraction osteogenesis. J Bone Joint Surg Am2009;91:1973–1984.
See www.StemCells.com for supporting information available online.
Battula, Evans, Hollier et al. 1445
Related Documents











