2006;66:192-197. Cancer Res Chengsen Xue, Jeffrey Wyckoff, Fubo Liang, et al. Enhanced Intravasation and Metastasis Coordinately with In vivo in Increased Tumor Cell Motility Epidermal Growth Factor Receptor Overexpression Results Updated version http://cancerres.aacrjournals.org/content/66/1/192 Access the most recent version of this article at: Material Supplementary http://cancerres.aacrjournals.org/content/suppl/2006/02/07/66.1.192.DC2.html http://cancerres.aacrjournals.org/content/suppl/2006/02/07/66.1.192.DC1.html Access the most recent supplemental material at: Cited Articles http://cancerres.aacrjournals.org/content/66/1/192.full.html#ref-list-1 This article cites by 49 articles, 24 of which you can access for free at: Citing articles http://cancerres.aacrjournals.org/content/66/1/192.full.html#related-urls This article has been cited by 33 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on November 27, 2013. © 2006 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on November 27, 2013. © 2006 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on November 27, 2013. © 2006 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2006;66:192-197. Cancer Res Chengsen Xue, Jeffrey Wyckoff, Fubo Liang, et al. Enhanced Intravasation and Metastasis
Coordinately withIn vivoin Increased Tumor Cell Motility Epidermal Growth Factor Receptor Overexpression Results
Updated version
http://cancerres.aacrjournals.org/content/66/1/192
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2006/02/07/66.1.192.DC2.html
http://cancerres.aacrjournals.org/content/suppl/2006/02/07/66.1.192.DC1.htmlAccess the most recent supplemental material at:
Cited Articles
http://cancerres.aacrjournals.org/content/66/1/192.full.html#ref-list-1
This article cites by 49 articles, 24 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/66/1/192.full.html#related-urls
This article has been cited by 33 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

Epidermal Growth Factor Receptor Overexpression Results in
Increased Tumor Cell Motility In vivo Coordinately with
Enhanced Intravasation and Metastasis
Chengsen Xue,1Jeffrey Wyckoff,
1,3Fubo Liang,
2Mazen Sidani,
1Stefania Violini,
1Kun-Lin Tsai,
1
Zhong-Yin Zhang,2Erik Sahai,
1John Condeelis,
1,3and Jeffrey E. Segall
1
Departments of 1Anatomy and Structural Biology and 2Molecular Pharmacology and 3Analytical Imaging Facility,Albert Einstein College of Medicine, Bronx, New York
Abstract
Although overexpression of the epidermal growth factorreceptor (EGFR; ErbB1) has been correlated with poorprognosis in breast and other cancers, clinical trials of ErbB1inhibitors have shown limited efficacy in inhibiting tumorproliferation. To evaluate other possible roles of ErbB1 intumor malignancy besides proliferation, we have developed aseries of tools for analysis of intravasation. Overexpression ofErbB1 in MTLn3 mammary adenocarcinoma cells results inincreased intravasation and lung metastasis from tumorsformed by injection of cells in the mammary fat pad. However,increased ErbB1 expression has no effect on primary tumorgrowth and lung seeding efficiency of cells injected i.v.Chemotactic responses to low concentrations of EGF in vitroand cell motility in vivo in the primary tumor measured usingintravital imaging are significantly increased by ErbB1 over-expression. The increased cell motility is restricted to ErbB1-overexpressing cells in tumors containing mixtures of cellsexpressing different ErbB1 levels, arguing for a cell-autonomouseffect of increased ErbB1 expression rather than alterationof the tumor microenvironment. In summary, we proposethat ErbB1 overexpression makes more significant contribu-tions to intravasation than growth in some tumors andpresent a novel model for studying ErbB1 contributions totumor metastasis via chemotaxis and intravasation. (Cancer Res2006; 66(1): 192-7)
Introduction
Metastasis involves multiple steps, including growth of aprimary tumor, intravasation, and arrest and growth in secondarysites. The steps of growth of the primary tumor, as well as arrestand growth in secondary sites have been extensively studied, withgenes affecting these steps being identified and studied (1–5).Studies of intravasation have lagged due to limitations intechniques for analyzing this step. The requirements for studyof intravasation include quantitation of the cell number in thecirculation, the ability to observe cells in the primary tumor, andidentification of cell lines in which intravasation is selectivelyaltered (6).
The epidermal growth factor receptor (EGFR; also referred to asErbB1) is often overexpressed in breast and other cancers (7–9).It has been correlated with poor prognosis (10, 11), and theseobservations have stimulated the development of ErbB1 inhibitors(12). Both antibody inhibitors, such as cetuximab, and smallmolecule inhibitors, such as gefitinib and Tarceva, have beendeveloped to inhibit ErbB1 (13–15). Clinical trials of these ErbB1inhibitors have shown some but much more limited efficacy inblocking tumor growth than would be expected based on theproportions of tumors with overexpressed ErbB1 (16, 17). Theseresults raise the possibility that ErbB1 might contribute to themalignant potential of tumors through affecting other character-istics besides just proliferation. ErbB1 can increase in vitro tumorcell motility and invasion (18–23). However, the in vivo contribu-tion of the increased motility and invasion, which is critical tounderstanding metastasis of tumors, is still not clear.We have developed methods for evaluating cell properties at the
primary tumor, including multiphoton imaging and quantitation oftumor cell number in the blood (6, 24–28). Multiphoton imagingof green fluorescent protein (GFP)–labeled primary tumors enablesdirect visualization of tumor cell behavior in vivo . Quantitation oftumor cell number in the blood is a direct evaluation of theefficiency of intravasation. The combination of these two measure-ments allows identification of changes in cell movement that cancontribute to intravasation efficiency. Using MTLn3 and MTC ratmammary adenocarcinoma cell lines, we showed that there can besignificant differences in intravasation between metastatic andnonmetastatic cells. More specifically, the metastatic MTLn3cells showed greater orientation toward blood vessels within theprimary tumor, whereas nonmetastatic MTC cells fragmentedwhen interacting with vessels (6, 24). The findings illustrate thevalue of a direct visualization of cell properties in vivo fordissection of the metastatic process.In this article, we provide direct in vivo evidence for contributions
of ErbB1 to metastasis independent of effects on proliferation.Overexpression of ErbB1 in MTLn3 rat mammary tumor cellsresults in significantly increased intravasation and metastasis fromthe primary tumor, without any changes in primary tumor growth.Chemotactic efficiency in vitro and tumor cell motility in vivo werealso significantly increased. Increased ErbB1 expression did notaffect lung seeding efficiency of cells injected i.v. Our studies areconsistent with in vitro and in vivo studies showing that ErbB1expression can enhance invasiveness (18–23).
Materials and Methods
Cell culture and establishing EGFR transfectants in MTLn3 ratmammary adenocarcinoma cells. Rat mammary adenocarcinoma cell
line MTLn3 was obtained from Dr. Garth Nicolson (Institute for Molecular
Note: E. Sahai is currently at the Tumour Cell Biology Laboratory, Cancer ResearchUnited Kingdom, London Research Institute, 44 Lincoln’s Inn Fields, London WC2A3PX, United Kingdom.
Requests for reprints: Jeffrey E. Segall, Department of Anatomy and StructuralBiology, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY10461. Phone: 718-430-4237; Fax: 718-430-8996; E-mail: [email protected].
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-05-1242
Cancer Res 2006; 66: (1). January 1, 2006 192 www.aacrjournals.org
Research Article
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

Medicine, Huntington Beach, CA; ref. 29) and was maintained in aMEM(Life Technologies, Inc., Gaithersburg, MD) supplemented with 5% fetal
bovine serum and penicillin-streptomycin (Life Technologies). For trans-
fection assays, MTLn3 cells were seeded in six-well plates and grown until
70% confluent. These cells were subjected to retroviral transduction ofconstructs containing the retroviral expression vector pLXSN alone or
pLXSN driving expression of ErbB1 (ref. 30; provided by Dr. David Stern,
Yale University). The vectors were packaged in the Phoenix cell line
following standard protocols (31), and geneticin-resistant clones wereisolated. Cell growth rate and morphology of transductants was determined
to be the same as parental cells, and the expression of ErbB1 was confirmed
by flow cytometry and Western blot.
Microchemotaxis chamber assay. A 48-well microchemotaxis chamber(Neuroprobe, Cabin John, MD) was used as previously described (32), except
that L15 containing 0.35% bovine serum albumin was used instead of
aMEMH. For measurement of migration in response to EGF (LifeTechnologies Bethesda Research Laboratories, Gaithersburg, MD), filters
were coated with 27 Ag/mL rat tail collagen (BD Biosciences, Palo Alto, CA).
After inserting the filters in the chamber, 20,000 cells detached by trypsin/
EDTA were plated into the wells of the upper chamber. The chambers wereincubated for 3 hours at 37jC before counting the number of cells crossing
the filter.
To evaluate the in vitro cell-autonomous chemotactic efficiency ofMTLn3-
B1 cells compared with MTLn3-PL cells, mixtures of equal numbers offluorescent MTLn3-B1-GFP and nonfluorescent MTLn3-PL cells were used in
the 48-well chemotaxis assay as described above. After fixation, the filters
were stained with.5 mg/mL 4V,6-diamidino-2-phenylindole (DAPI) for 2minutes, and then images of DAPI and GFP fluorescence were taken using a�10 objective. In each experiment, images of unscrapedwells were also taken
to determine the actual relative percentage of cells plated per experiment,
and this information was used in calculating the relative chemotacticefficiency. ImageJ (developed by Wayne Rasband at the NIH, Bethesda, MD;
http://rsb.info.nih.gov/ij/, 1997-2005) macros were written to identify all cells
using the DAPI image and MTLn3-B1 cells by GFP fluorescence. For each
condition, the relative proportion of MTLn3-B1 cells was determined andused to calculate the relative chemotactic efficiency of MTLn3-B1 cells
compared with MTLn3-PL cells. Iressa (AstraZeneca, Manchester, England)
and AG879 (SigmaT2067) were used to inhibit ErbB1 and ErbB2, respectively.Immunoblotting. Cells were grown to 70% confluency in a 10-cm cell-
culturing dish and then incubated with serum-free medium for 4 hours. The
medium was changed to fresh serum-free medium with or without 5 nmol/L
EGF, and cells were incubated for 30 seconds. Cells were then washed with
cold PBS and lysed in 0.5 mL lysis buffer [50 mmol/L Tris-HCl (pH 7.5),
1% Triton X-100, 5 mmol/L EGTA, 1 mmol/L EDTA, 150 mmol/L NaCl,
10% glycerol, 1 mmol/L phenylmethylsulfonyl fluoride, 50 mmol/L sodium
fluoride, Complete Protease Inhibitor Cocktail (Roche Diagnostics Corp.,
Indianapolis, IN)]. The plates were scraped with a rubber policeman and
incubated on ice for 20 minutes. The lysate was precleared by centrifugation
at 15,000 rpm for 15 minutes. Lysate protein concentration was measured
using bicinchoninic acid protein assay reagent (Pierce, Rockford, IL).
Protein (20 Ag) of each sample was loaded and separated by SDS-PAGE and
transferred electrophoretically to nitrocellulose membranes, which were
immunoblotted by appropriate antibodies followed by incubation with
horseradish peroxidase–conjugated secondary antibodies. The following
antibodies were used: anti-h-actin (Sigma, St. Louis, MO), anti-phosphotyr-
osine (PY20, BD Biosciences, San Diego, CA), anti-phospho-ErbB1 (Tyr1173,
Cell Signaling Technology, Beverly, MA), anti-phospho-ErbB2 (Tyr877, Cell
Signaling Technology), and anti-ErbB2 monoclonal (NeoMarkers, Fremont,
CA). The blots were developed by using SuperSignal West Pico
Chemiluminescent Substrate (Pierce).
Spontaneous and experimental metastasis assays. All animal studies
described here were done according to protocols approved by theInstitutional Animal Care and Use Committee of Albert Einstein College
of Medicine. To measure spontaneous metastasis, tumor cells were grown
to 70% to 85% confluence before being harvested for cell counting. Cells(5 � 105) were injected into the right thoracic mammary fat pads of 5- to 7-
week-old female BALB/c severe combined immunodeficient (SCID) mice
(National Cancer Institute, Bethesda, MD). The cells were injected in avolume of 100 AL of PBS with calcium and magnesium through a 25-gauge
needle. Tumor growth rate was monitored at weekly intervals after inoc-
ulation of tumor cells. The tumors were measured in two dimensions, and
tumor volumes were calculated using the formula: (length � width2) / 2.Five weeks after inoculation, the animals were anesthetized, blood burden
was determined as described below followed by sacrifice of the animals and
removal of primary tumors and lungs for histologic analysis as described
below.For the experimental lung metastasis assay, 2.5 � 105 cells were injected
into the lateral tail veins of 5- to 7-week-old female BALB/c SCID mice
(National Cancer Institute). Two weeks after injection, the mice were
euthanized, and the lungs were removed and subjected to histologicexamination as described below.
Measurement of tumor cell blood burden. At the end point of the
spontaneous metastasis assay, mice were anesthetized with Aerrane(Isoflurane, Baxter Pharmaceutical Products, Inc., Deerfield, IL). The right
chest was exposed by a simple skin flap surgery. Blood was taken from the
right atrium via heart puncture with a 25-gauge needle and 1-mL syringe
coated with heparin. Blood (0.2-1.05 mL) was harvested from each animal.The blood was immediately plated into 150-mm-diameter dishes filled with
5% fetal bovine serum/aMEM growth medium. The next day, the plates
were rinsed with fresh medium containing 0.8 mg/mL geneticin to
selectively grow the tumor cells. After 3 to 7 days, all tumor cell clonesin the dish were counted. Tumor blood burden was calculated as total
colonies in the dish divided by the volume of blood taken. To test tumor
cell viability in the blood colony assay process, cultured MTLn3-PL cellswere removed from dishes by 0.25% trypsin/EDTA, and 1,000 tumor cells
were added to fresh mouse blood. The mixtures were then processed as
described above.
Tumor histology and quantitative assessment of the efficiency ofmetastasis. The primary tumors and lungs from each mouse were used for
histologic analysis. Samples were fixed in formalin and embedded in
paraffin, and 5-Am sections were stained with H&E. For each lung sample,
all micrometastases were counted under a light microscope at �10
magnification, and the total lung area was measured using a UMAX
PowerLook III color scanner (UMAX Technologies, Inc., Dallas, TX) and
Adobe Photoshop 5.5 software. Briefly, after scanning lung sections, the
cross-sectional area in pixels was measured using PhotoShop and converted
to mm2 using a calibration factor of 1 pixel = 0.00179 mm2. The efficiency of
lung metastasis then was calculated as metastases per mm2 of lung area.
Intravital imaging by multiphoton microscopy. Tumor imaging was
done as described previously (6, 24, 27, 33). Cells (5 � 105) were injected into
the mammary fat pads of SCID mice as described above and allowed to grow
for 4 weeks. The animal was placed under isoflurane anesthesia, and the
tumor was exposed using a simple skin flap surgery, with as little disruption
of the surrounding vasculature as possible. The animal was then placed in a
30jC temperature chamber on an inverted Olympus IX70 microscope and
imaged using a �20 objective. Briefly, a 10-W Millennium Xs laser (Spectra
Physics, Mountain View, CA) was used to run a Radiance 2000 multiphoton
system (Bio-Rad, Hercules, CA) at 880 nm using a 450/480-nm filter to image
matrix and CFP and a 515/530-nm filter to image CFP and GFP. Time lapse
images of GFP-labeled, MTLn3-PL- and MTLn3-B1-generated tumors were
taken at 60-second intervals for 30 minutes. The images were collected using
Bio-Rad’s Lasersharp 2000 software. During each 1-minute interval, a z series
of 9 to 12 images was taken at a spacing of 10 Am between images, beginning
at the periphery of the tumor and moving into the tumor. For each tumor,
this image acquisition process was repeated for 30 minutes, resulting in a
time lapse three-dimensional z series for analysis of tumor cell motility.
Sites of moving tumor cells in each plane of the z series were identified
by playing the time lapse movie of that plane in ImageJ and marking sites of
movement using a specially written plugin. The total number of movementsites for each plane over the entire 30 minutes was then calculated. The top
four planes (corresponding to the outermost 40 Am of the tumor)
reproducibly provided good enough images for determination of movementsites and were used to calculate the average number of motility sites for
each tumor type.
EGFR Enhances Motility and Intravasation In vivo
www.aacrjournals.org 193 Cancer Res 2006; 66: (1). January 1, 2006
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
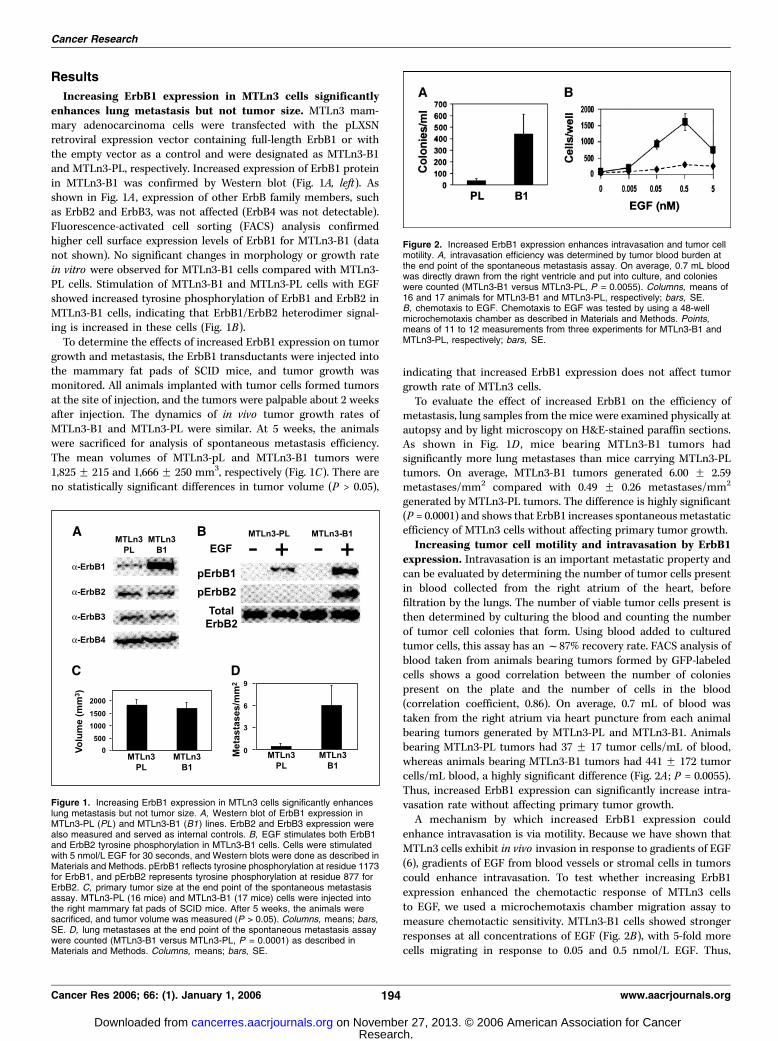
Results
Increasing ErbB1 expression in MTLn3 cells significantlyenhances lung metastasis but not tumor size. MTLn3 mam-mary adenocarcinoma cells were transfected with the pLXSNretroviral expression vector containing full-length ErbB1 or withthe empty vector as a control and were designated as MTLn3-B1and MTLn3-PL, respectively. Increased expression of ErbB1 proteinin MTLn3-B1 was confirmed by Western blot (Fig. 1A, left). Asshown in Fig. 1A , expression of other ErbB family members, suchas ErbB2 and ErbB3, was not affected (ErbB4 was not detectable).Fluorescence-activated cell sorting (FACS) analysis confirmedhigher cell surface expression levels of ErbB1 for MTLn3-B1 (datanot shown). No significant changes in morphology or growth ratein vitro were observed for MTLn3-B1 cells compared with MTLn3-PL cells. Stimulation of MTLn3-B1 and MTLn3-PL cells with EGFshowed increased tyrosine phosphorylation of ErbB1 and ErbB2 inMTLn3-B1 cells, indicating that ErbB1/ErbB2 heterodimer signal-ing is increased in these cells (Fig. 1B).To determine the effects of increased ErbB1 expression on tumor
growth and metastasis, the ErbB1 transductants were injected intothe mammary fat pads of SCID mice, and tumor growth wasmonitored. All animals implanted with tumor cells formed tumorsat the site of injection, and the tumors were palpable about 2 weeksafter injection. The dynamics of in vivo tumor growth rates ofMTLn3-B1 and MTLn3-PL were similar. At 5 weeks, the animalswere sacrificed for analysis of spontaneous metastasis efficiency.The mean volumes of MTLn3-pL and MTLn3-B1 tumors were1,825 F 215 and 1,666 F 250 mm3, respectively (Fig. 1C). There areno statistically significant differences in tumor volume (P > 0.05),
indicating that increased ErbB1 expression does not affect tumorgrowth rate of MTLn3 cells.To evaluate the effect of increased ErbB1 on the efficiency of
metastasis, lung samples from the mice were examined physically atautopsy and by light microscopy on H&E-stained paraffin sections.As shown in Fig. 1D , mice bearing MTLn3-B1 tumors hadsignificantly more lung metastases than mice carrying MTLn3-PLtumors. On average, MTLn3-B1 tumors generated 6.00 F 2.59metastases/mm2 compared with 0.49 F 0.26 metastases/mm2
generated by MTLn3-PL tumors. The difference is highly significant(P = 0.0001) and shows that ErbB1 increases spontaneousmetastaticefficiency of MTLn3 cells without affecting primary tumor growth.Increasing tumor cell motility and intravasation by ErbB1
expression. Intravasation is an important metastatic property andcan be evaluated by determining the number of tumor cells presentin blood collected from the right atrium of the heart, beforefiltration by the lungs. The number of viable tumor cells present isthen determined by culturing the blood and counting the numberof tumor cell colonies that form. Using blood added to culturedtumor cells, this assay has anf87% recovery rate. FACS analysis ofblood taken from animals bearing tumors formed by GFP-labeledcells shows a good correlation between the number of coloniespresent on the plate and the number of cells in the blood(correlation coefficient, 0.86). On average, 0.7 mL of blood wastaken from the right atrium via heart puncture from each animalbearing tumors generated by MTLn3-PL and MTLn3-B1. Animalsbearing MTLn3-PL tumors had 37 F 17 tumor cells/mL of blood,whereas animals bearing MTLn3-B1 tumors had 441 F 172 tumorcells/mL blood, a highly significant difference (Fig. 2A ; P = 0.0055).Thus, increased ErbB1 expression can significantly increase intra-vasation rate without affecting primary tumor growth.A mechanism by which increased ErbB1 expression could
enhance intravasation is via motility. Because we have shown thatMTLn3 cells exhibit in vivo invasion in response to gradients of EGF(6), gradients of EGF from blood vessels or stromal cells in tumorscould enhance intravasation. To test whether increasing ErbB1expression enhanced the chemotactic response of MTLn3 cellsto EGF, we used a microchemotaxis chamber migration assay tomeasure chemotactic sensitivity. MTLn3-B1 cells showed strongerresponses at all concentrations of EGF (Fig. 2B), with 5-fold morecells migrating in response to 0.05 and 0.5 nmol/L EGF. Thus,
Figure 1. Increasing ErbB1 expression in MTLn3 cells significantly enhanceslung metastasis but not tumor size. A, Western blot of ErbB1 expression inMTLn3-PL (PL ) and MTLn3-B1 (B1) lines. ErbB2 and ErbB3 expression werealso measured and served as internal controls. B, EGF stimulates both ErbB1and ErbB2 tyrosine phosphorylation in MTLn3-B1 cells. Cells were stimulatedwith 5 nmol/L EGF for 30 seconds, and Western blots were done as described inMaterials and Methods. pErbB1 reflects tyrosine phosphorylation at residue 1173for ErbB1, and pErbB2 represents tyrosine phosphorylation at residue 877 forErbB2. C, primary tumor size at the end point of the spontaneous metastasisassay. MTLn3-PL (16 mice) and MTLn3-B1 (17 mice) cells were injected intothe right mammary fat pads of SCID mice. After 5 weeks, the animals weresacrificed, and tumor volume was measured (P > 0.05). Columns, means; bars,SE. D, lung metastases at the end point of the spontaneous metastasis assaywere counted (MTLn3-B1 versus MTLn3-PL, P = 0.0001) as described inMaterials and Methods. Columns, means; bars, SE.
Figure 2. Increased ErbB1 expression enhances intravasation and tumor cellmotility. A, intravasation efficiency was determined by tumor blood burden atthe end point of the spontaneous metastasis assay. On average, 0.7 mL bloodwas directly drawn from the right ventricle and put into culture, and colonieswere counted (MTLn3-B1 versus MTLn3-PL, P = 0.0055). Columns, means of16 and 17 animals for MTLn3-B1 and MTLn3-PL, respectively; bars, SE.B, chemotaxis to EGF. Chemotaxis to EGF was tested by using a 48-wellmicrochemotaxis chamber as described in Materials and Methods. Points,means of 11 to 12 measurements from three experiments for MTLn3-B1 andMTLn3-PL, respectively; bars, SE.
Cancer Research
Cancer Res 2006; 66: (1). January 1, 2006 194 www.aacrjournals.org
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
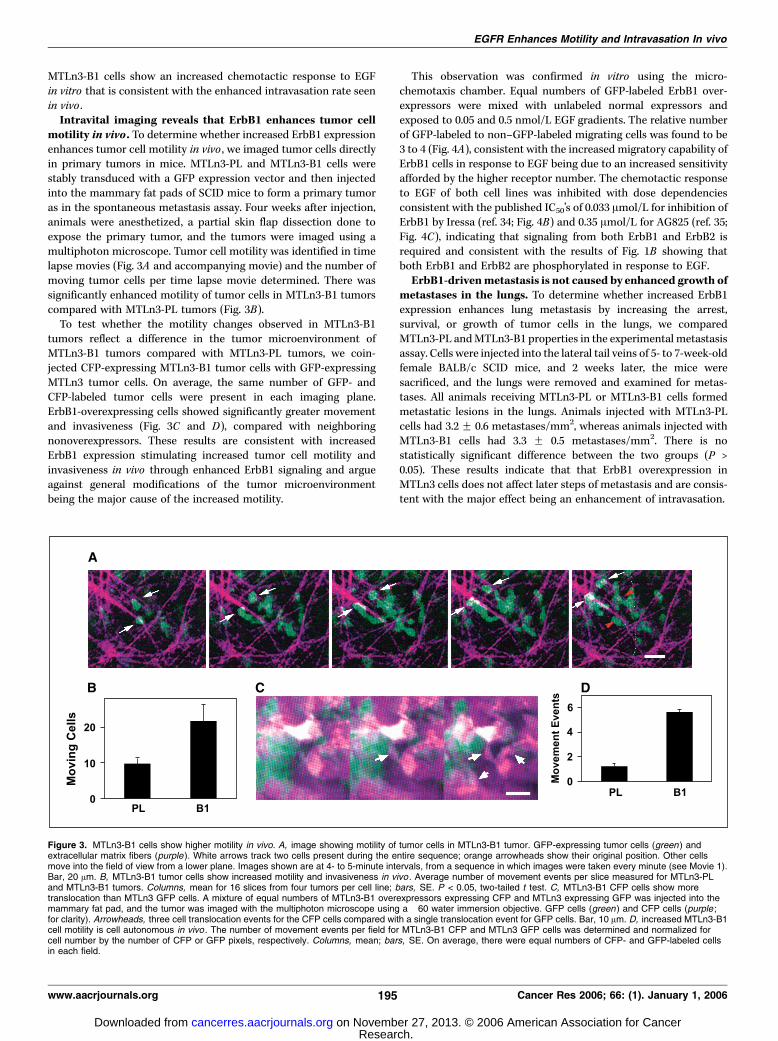
MTLn3-B1 cells show an increased chemotactic response to EGFin vitro that is consistent with the enhanced intravasation rate seenin vivo .Intravital imaging reveals that ErbB1 enhances tumor cell
motility in vivo . To determine whether increased ErbB1 expressionenhances tumor cell motility in vivo , we imaged tumor cells directlyin primary tumors in mice. MTLn3-PL and MTLn3-B1 cells werestably transduced with a GFP expression vector and then injectedinto the mammary fat pads of SCID mice to form a primary tumoras in the spontaneous metastasis assay. Four weeks after injection,animals were anesthetized, a partial skin flap dissection done toexpose the primary tumor, and the tumors were imaged using amultiphoton microscope. Tumor cell motility was identified in timelapse movies (Fig. 3A and accompanying movie) and the number ofmoving tumor cells per time lapse movie determined. There wassignificantly enhanced motility of tumor cells in MTLn3-B1 tumorscompared with MTLn3-PL tumors (Fig. 3B).To test whether the motility changes observed in MTLn3-B1
tumors reflect a difference in the tumor microenvironment ofMTLn3-B1 tumors compared with MTLn3-PL tumors, we coin-jected CFP-expressing MTLn3-B1 tumor cells with GFP-expressingMTLn3 tumor cells. On average, the same number of GFP- andCFP-labeled tumor cells were present in each imaging plane.ErbB1-overexpressing cells showed significantly greater movementand invasiveness (Fig. 3C and D), compared with neighboringnonoverexpressors. These results are consistent with increasedErbB1 expression stimulating increased tumor cell motility andinvasiveness in vivo through enhanced ErbB1 signaling and argueagainst general modifications of the tumor microenvironmentbeing the major cause of the increased motility.
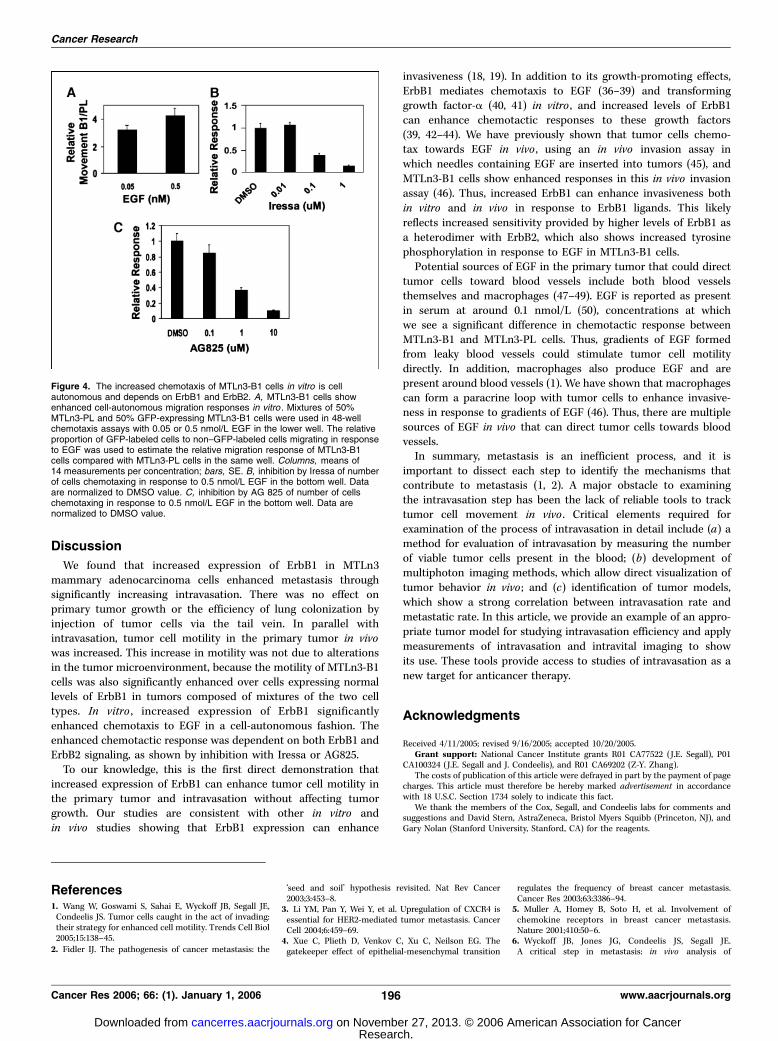
This observation was confirmed in vitro using the micro-chemotaxis chamber. Equal numbers of GFP-labeled ErbB1 over-expressors were mixed with unlabeled normal expressors andexposed to 0.05 and 0.5 nmol/L EGF gradients. The relative numberof GFP-labeled to non–GFP-labeled migrating cells was found to be3 to 4 (Fig. 4A), consistent with the increased migratory capability ofErbB1 cells in response to EGF being due to an increased sensitivityafforded by the higher receptor number. The chemotactic responseto EGF of both cell lines was inhibited with dose dependenciesconsistent with the published IC50’s of 0.033 Amol/L for inhibition ofErbB1 by Iressa (ref. 34; Fig. 4B) and 0.35 Amol/L for AG825 (ref. 35;Fig. 4C), indicating that signaling from both ErbB1 and ErbB2 isrequired and consistent with the results of Fig. 1B showing thatboth ErbB1 and ErbB2 are phosphorylated in response to EGF.ErbB1-drivenmetastasis is not caused by enhanced growth of
metastases in the lungs. To determine whether increased ErbB1expression enhances lung metastasis by increasing the arrest,survival, or growth of tumor cells in the lungs, we comparedMTLn3-PL andMTLn3-B1 properties in the experimental metastasisassay. Cells were injected into the lateral tail veins of 5- to 7-week-oldfemale BALB/c SCID mice, and 2 weeks later, the mice weresacrificed, and the lungs were removed and examined for metas-tases. All animals receiving MTLn3-PL or MTLn3-B1 cells formedmetastatic lesions in the lungs. Animals injected with MTLn3-PLcells had 3.2 F 0.6 metastases/mm2, whereas animals injected withMTLn3-B1 cells had 3.3 F 0.5 metastases/mm2. There is nostatistically significant difference between the two groups (P >0.05). These results indicate that that ErbB1 overexpression inMTLn3 cells does not affect later steps of metastasis and are consis-tent with the major effect being an enhancement of intravasation.
Figure 3. MTLn3-B1 cells show higher motility in vivo. A, image showing motility of tumor cells in MTLn3-B1 tumor. GFP-expressing tumor cells (green ) andextracellular matrix fibers (purple ). White arrows track two cells present during the entire sequence; orange arrowheads show their original position. Other cellsmove into the field of view from a lower plane. Images shown are at 4- to 5-minute intervals, from a sequence in which images were taken every minute (see Movie 1).Bar, 20 Am. B, MTLn3-B1 tumor cells show increased motility and invasiveness in vivo . Average number of movement events per slice measured for MTLn3-PLand MTLn3-B1 tumors. Columns, mean for 16 slices from four tumors per cell line; bars, SE. P < 0.05, two-tailed t test. C, MTLn3-B1 CFP cells show moretranslocation than MTLn3 GFP cells. A mixture of equal numbers of MTLn3-B1 overexpressors expressing CFP and MTLn3 expressing GFP was injected into themammary fat pad, and the tumor was imaged with the multiphoton microscope using a �60 water immersion objective. GFP cells (green ) and CFP cells (purple ;for clarity). Arrowheads, three cell translocation events for the CFP cells compared with a single translocation event for GFP cells. Bar, 10 Am. D, increased MTLn3-B1cell motility is cell autonomous in vivo . The number of movement events per field for MTLn3-B1 CFP and MTLn3 GFP cells was determined and normalized forcell number by the number of CFP or GFP pixels, respectively. Columns, mean; bars, SE. On average, there were equal numbers of CFP- and GFP-labeled cellsin each field.
EGFR Enhances Motility and Intravasation In vivo
www.aacrjournals.org 195 Cancer Res 2006; 66: (1). January 1, 2006
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Discussion
We found that increased expression of ErbB1 in MTLn3mammary adenocarcinoma cells enhanced metastasis throughsignificantly increasing intravasation. There was no effect onprimary tumor growth or the efficiency of lung colonization byinjection of tumor cells via the tail vein. In parallel withintravasation, tumor cell motility in the primary tumor in vivowas increased. This increase in motility was not due to alterationsin the tumor microenvironment, because the motility of MTLn3-B1cells was also significantly enhanced over cells expressing normallevels of ErbB1 in tumors composed of mixtures of the two celltypes. In vitro , increased expression of ErbB1 significantlyenhanced chemotaxis to EGF in a cell-autonomous fashion. Theenhanced chemotactic response was dependent on both ErbB1 andErbB2 signaling, as shown by inhibition with Iressa or AG825.To our knowledge, this is the first direct demonstration that
increased expression of ErbB1 can enhance tumor cell motility inthe primary tumor and intravasation without affecting tumorgrowth. Our studies are consistent with other in vitro andin vivo studies showing that ErbB1 expression can enhance
invasiveness (18, 19). In addition to its growth-promoting effects,ErbB1 mediates chemotaxis to EGF (36–39) and transforminggrowth factor-a (40, 41) in vitro , and increased levels of ErbB1can enhance chemotactic responses to these growth factors(39, 42–44). We have previously shown that tumor cells chemo-tax towards EGF in vivo , using an in vivo invasion assay inwhich needles containing EGF are inserted into tumors (45), andMTLn3-B1 cells show enhanced responses in this in vivo invasionassay (46). Thus, increased ErbB1 can enhance invasiveness bothin vitro and in vivo in response to ErbB1 ligands. This likelyreflects increased sensitivity provided by higher levels of ErbB1 asa heterodimer with ErbB2, which also shows increased tyrosinephosphorylation in response to EGF in MTLn3-B1 cells.Potential sources of EGF in the primary tumor that could direct
tumor cells toward blood vessels include both blood vesselsthemselves and macrophages (47–49). EGF is reported as presentin serum at around 0.1 nmol/L (50), concentrations at whichwe see a significant difference in chemotactic response betweenMTLn3-B1 and MTLn3-PL cells. Thus, gradients of EGF formedfrom leaky blood vessels could stimulate tumor cell motilitydirectly. In addition, macrophages also produce EGF and arepresent around blood vessels (1). We have shown that macrophagescan form a paracrine loop with tumor cells to enhance invasive-ness in response to gradients of EGF (46). Thus, there are multiplesources of EGF in vivo that can direct tumor cells towards bloodvessels.In summary, metastasis is an inefficient process, and it is
important to dissect each step to identify the mechanisms thatcontribute to metastasis (1, 2). A major obstacle to examiningthe intravasation step has been the lack of reliable tools to tracktumor cell movement in vivo . Critical elements required forexamination of the process of intravasation in detail include (a) amethod for evaluation of intravasation by measuring the numberof viable tumor cells present in the blood; (b) development ofmultiphoton imaging methods, which allow direct visualization oftumor behavior in vivo ; and (c) identification of tumor models,which show a strong correlation between intravasation rate andmetastatic rate. In this article, we provide an example of an appro-priate tumor model for studying intravasation efficiency and applymeasurements of intravasation and intravital imaging to showits use. These tools provide access to studies of intravasation as anew target for anticancer therapy.
Acknowledgments
Received 4/11/2005; revised 9/16/2005; accepted 10/20/2005.Grant support: National Cancer Institute grants R01 CA77522 (J.E. Segall), P01
CA100324 (J.E. Segall and J. Condeelis), and R01 CA69202 (Z-Y. Zhang).The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank the members of the Cox, Segall, and Condeelis labs for comments andsuggestions and David Stern, AstraZeneca, Bristol Myers Squibb (Princeton, NJ), andGary Nolan (Stanford University, Stanford, CA) for the reagents.
Figure 4. The increased chemotaxis of MTLn3-B1 cells in vitro is cellautonomous and depends on ErbB1 and ErbB2. A, MTLn3-B1 cells showenhanced cell-autonomous migration responses in vitro . Mixtures of 50%MTLn3-PL and 50% GFP-expressing MTLn3-B1 cells were used in 48-wellchemotaxis assays with 0.05 or 0.5 nmol/L EGF in the lower well. The relativeproportion of GFP-labeled cells to non–GFP-labeled cells migrating in responseto EGF was used to estimate the relative migration response of MTLn3-B1cells compared with MTLn3-PL cells in the same well. Columns, means of14 measurements per concentration; bars, SE. B, inhibition by Iressa of numberof cells chemotaxing in response to 0.5 nmol/L EGF in the bottom well. Dataare normalized to DMSO value. C, inhibition by AG 825 of number of cellschemotaxing in response to 0.5 nmol/L EGF in the bottom well. Data arenormalized to DMSO value.
References1. Wang W, Goswami S, Sahai E, Wyckoff JB, Segall JE,Condeelis JS. Tumor cells caught in the act of invading:their strategy for enhanced cell motility. Trends Cell Biol2005;15:138–45.
2. Fidler IJ. The pathogenesis of cancer metastasis: the
‘seed and soil’ hypothesis revisited. Nat Rev Cancer2003;3:453–8.
3. Li YM, Pan Y, Wei Y, et al. Upregulation of CXCR4 isessential for HER2-mediated tumor metastasis. CancerCell 2004;6:459–69.
4. Xue C, Plieth D, Venkov C, Xu C, Neilson EG. Thegatekeeper effect of epithelial-mesenchymal transition
regulates the frequency of breast cancer metastasis.Cancer Res 2003;63:3386–94.
5. Muller A, Homey B, Soto H, et al. Involvement ofchemokine receptors in breast cancer metastasis.Nature 2001;410:50–6.
6. Wyckoff JB, Jones JG, Condeelis JS, Segall JE.A critical step in metastasis: in vivo analysis of
Cancer Research
Cancer Res 2006; 66: (1). January 1, 2006 196 www.aacrjournals.org
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

intravasation at the primary tumor. Cancer Res 2000;60:2504–11.
7. Sherwood ER, Lee C. Epidermal growth factor-relatedpeptides and the epidermal growth factor receptor innormal and malignant prostate. World J Urol 1995;13:290–6.
8. Muller-Tidow C, Schwable J, Steffen B, et al. High-throughput analysis of genome-wide receptor tyrosinekinase expression in human cancers identifies potentialnovel drug targets. Clin Cancer Res 2004;10:1241–9.
9. Barnes CJ, Kumar R. Epidermal growth factor receptorfamily tyrosine kinases as signal integrators and thera-peutic targets. Cancer Metastasis Rev 2003;22:301–7.
10. Klijn JG, Look MP, Portengen H, Alexieva-Figusch J,van Putten WL, Foekens JA. The prognostic value ofepidermal growth factor receptor (EGF-R) in primarybreast cancer: results of a 10 year follow-up study.Breast Cancer Res Treat 1994;29:73–83.
11. Chrysogelos SA, Dickson RB. EGF receptor expres-sion, regulation, and function in breast cancer. BreastCancer Res Treat 1994;29:29–40.
12. Normanno N, Maiello MR, Mancino M, De Luca A.Small molecule epidermal growth factor receptortyrosine kinase inhibitors: an overview. J Chemother2004;16 Suppl 4:36–40.
13. Elkind NB, Szentpetery Z, Apati A, et al. Multidrugtransporter ABCG2 prevents tumor cell death inducedby the epidermal growth factor receptor inhibitor Iressa(ZD1839, Gefitinib). Cancer Res 2005;65:1770–7.
14. Nakamura Y, Oka M, Soda H, et al. Gefitinib (‘‘Iressa’’,ZD1839), an epidermal growth factor receptor tyrosinekinase inhibitor, reverses breast cancer resistanceprotein/ABCG2-mediated drug resistance. Cancer Res2005;65:1541–6.
15. Brehmer D, Greff Z, Godl K, et al. Cellular targets ofgefitinib. Cancer Res 2005;65:379–82.
16. Twombly R. Failing survival advantage in crucial trial,future of Iressa is in jeopardy. J Natl Cancer Inst 2005;97:249–50.
17. Kobayashi S, Boggon TJ, Dayaram T, et al. EGFRmutation and resistance of non-small-cell lung cancer togefitinib. N Engl J Med 2005;352:786–92.
18. Price JT, Wilson HM, Haites NE. Epidermal growthfactor (EGF) increases the in vitro invasion, motility andadhesion interactions of the primary renal carcinomacell line, A704. Eur J Cancer 1996;32A:1977–82.
19. Turner T, Chen P, Goodly LJ, Wells A. EGF receptorsignaling enhances in vivo invasiveness of DU-145human prostate carcinoma cells. Clin Exp Metastasis1996;14:409–18.
20. Rosen EM, Goldberg ID. Protein factors whichregulate cell motility. In Vitro Cell Dev Biol 1989;25:1079–87.
21. Shibata T, Kawano T, Nagayasu H, et al. Enhancingeffects of epidermal growth factor on human squamouscell carcinoma motility and matrix degradation but notgrowth. Tumour Biol 1996;17:168–75.
22. Wells A. Tumor invasion: role of growth factor-induced cell motility. Adv Cancer Res 2000;78:31–101.
23. Yang Z, Bagheri-Yarmand R, Wang RA, et al. Theepidermal growth factor receptor tyrosine kinaseinhibitor ZD1839 (Iressa) suppresses c-Src and Pak1pathways and invasiveness of human cancer cells. ClinCancer Res 2004;10:658–67.
24. Condeelis JS, Wyckoff J, Segall JE. Imaging of cancerinvasion and metastasis using green fluorescent protein.Eur J Cancer 2000;36:1671–80.
25. Condeelis JS, Wyckoff JB, Bailly M, et al. Lamellipodiain invasion. Semin Cancer Biol 2001;11:119–28.
26. Condeelis J, Segall JE. Intravital imaging of cellmovement in tumours. Nat Rev Cancer 2003;3:921–30.
27. Wang W, Wyckoff JB, Frohlich VC, et al. Single cellbehavior in metastatic primary mammary tumorscorrelated with gene expression patterns revealed bymolecular profiling. Cancer Res 2002;62:6278–88.
28. Wang W, Goswami S, Lapidus K, et al. Identificationand testing of a gene expression signature of invasivecarcinoma cells within primary mammary tumors.Cancer Res 2004;64:8585–94.
29. Neri A, Welch D, Kawaguchi T, Nicolson GL.Development and biologic properties of malignant cellsublines and clones of a spontaneously metastasizingrat mammary adenocarcinoma. J Natl Cancer Inst1982;68:507–17.
30. Riese DJ, Kim ED, Elenius K, et al. The epidermalgrowth factor receptor couples transforming growthfactor-a, heparin-binding epidermal growth factor-likefactor, and amphiregulin to Neu, ErbB-3, and ErbB-4.J Biol Chem 1996;271:20047–52.
31. Grignani F, Kinsella T, Mencarelli A, et al. High-efficiency gene transfer and selection of humanhematopoietic progenitor cells with a hybrid EBV/retroviral vector expressing the green fluorescenceprotein. Cancer Res 1998;58:14–9.
32. Segall JE, Tyerech S, Boselli L, et al. EGF stimulateslamellipod extension in metastatic mammary adeno-carcinoma cells by an actin-dependent mechanism. ClinExp Metastasis 1996;14:61–72.
33. Johnson SA, Hunter T. Kinomics: methods fordeciphering the kinome. Nat Methods 2005;2:17–25.
34. Wakeling AE, Guy SP, Woodburn JR, et al. ZD1839(Iressa): an orally active inhibitor of epidermal growthfactor signaling with potential for cancer therapy.Cancer Res 2002;62:5749–54.
35. Levitzki A, Gazit A. Tyrosine kinase inhibition: anapproach to drug development. Science 1995;267:1782–8.
36. Chen P, Xie H, Sekar MC, Gupta K, Wells A.Epidermal growth factor receptor-mediated cellmotility: phospholipase C activity is required, butmitogen-activated protein kinase activity is not suffi-cient for induced cell movement. J Cell Biol 1994;127:847–57.
37. Nolte C, Kirchhoff F, Kettenmann H. Epidermalgrowth factor is a motility factor for microglial cells
in vitro : evidence for EGF receptor expression. Eur JNeurosci 1997;9:1690–8.
38. Lamb DJ, Modjtahedi H, Plant NJ, Ferns GA. EGFmediates monocyte chemotaxis and macrophage pro-liferation and EGF receptor is expressed in atheroscle-rotic plaques. Atherosclerosis 2004;176:21–6.
39. Bailly M, Wyckoff J, Bouzahzah B, et al. Epidermalgrowth factor receptor distribution during chemotacticresponses. Mol Biol Cell 2000;11:3873–83.
40. El Obeid A, Bongcam-Rudloff E, Sorby M, Ostman A,Nister M, Westermark B. Cell scattering and migrationinduced by autocrine transforming growth factor a
in human glioma cells in vitro . Cancer Res 1997;57:5598–604.
41. Zhou R, Skalli O. TGF-a differentially regulates GFAP,vimentin, and nestin gene expression in U-373 MGglioblastoma cells: correlation with cell shape andmotility. Exp Cell Res 2000;254:269–78.
42. Chen P, Gupta K, Wells A. Cell movement elicited byepidermal growth factor receptor requires kinase andautophosphorylation but is separable from mitogenesis.J Cell Biol 1994;124:547–55.
43. Caric D, Raphael H, Viti J, Feathers A, Wancio D,Lillien L. EGFRs mediate chemotactic migration inthe developing telencephalon. Development 2001;128:4203–16.
44. Price JT, Tiganis T, Agarwal A, Djakiew D, ThompsonEW. Epidermal growth factor promotes MDA-MB-231breast cancer cell migration through a phosphatidyli-nositol 3V-kinase and phospholipase C-dependentmechanism. Cancer Res 1999;59:5475–8.
45. Wyckoff JB, Segall JE, Condeelis JS. The collection ofthe motile population of cells from a living tumor.Cancer Res 2000;60:5401–4.
46. Wyckoff J, Wang W, Lin EY, et al. A paracrine loopbetween tumor cells and macrophages is required fortumor cell migration in mammary tumors. Cancer Res2004;64:7022–9.
47. Che W, Asahi M, Takahashi M, et al. Selectiveinduction of heparin-binding epidermal growth factor-like growth factor by methylglyoxal and 3-deoxygluco-sone in rat aortic smooth muscle cells. The involvementof reactive oxygen species formation and a possibleimplication for atherogenesis in diabetes. J Biol Chem1997;272:18453–9.
48. Higashiyama S, Abraham JA, Miller J, Fiddes JC,Klagsbrun M. A heparin-binding growth factor secretedby macrophage-like cells that is related to EGF. Science1991;251:936–9.
49. Temelkovski J, Kumar RK, Maronese SE. Enhancedproduction of an EGF-like growth factor by parenchy-mal macrophages following bleomycin-induced pulmo-nary injury. Exp Lung Res 1997;23:377–91.
50. Futamura T, Toyooka K, Iritani S, et al. Abnormalexpression of epidermal growth factor and its receptorin the forebrain and serum of schizophrenic patients.Mol Psychiatry 2002;7:673–82.
EGFR Enhances Motility and Intravasation In vivo
www.aacrjournals.org 197 Cancer Res 2006; 66: (1). January 1, 2006
Research. on November 27, 2013. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

EGFR Enhances Motility and Intravasation In vivo
In the article on how EGFR enhances motility and intravasationin vivo in the January 1, 2006 issue of Cancer Research (1), inFig. 1A , the label reading ‘‘a-ErbB4’’ should have read ‘‘a-actin.’’ Thecorrected figure appears below.
1. Xue C, Wyckoff J, Liang F, Sidani M, Violini S, Tsai K-L, Zhang Z-Y, Sahai E, CondeelisJ, Segall JE. Epidermal growth factor receptor overexpression results in increasedtumor cell motility in vivo coordinately with enhanced intravasation and metastasis.Cancer Res 2006;66:192–7.
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-66-8-COR
www.aacrjournals.org 4547 Cancer Res 2006; 66: (8). April 15, 2006
Correction
Related Documents











