Ependymoma stem cells are highly sensitive to temozolomide in vitro and in orthotopic models Daniela Meco † , Tiziana Servidei † , Giuseppe Lamorte, Elena Binda, Vincenzo Arena, and Riccardo Riccardi Department of Pediatric Oncology, Catholic University, Rome, Italy (D.M., T.S., R.R.); Istituto CSS - Mendel Laboratory, Rome, Italy (G.L.); Department of Biotechnology and Biosciences, Building U3, University of Milan Bicocca, Milan, Italy (E.B.); Institute of Pathology, Catholic University, Rome, Italy (V.A.) Corresponding Author: Tiziana Servidei, Department of Pediatric Oncology, Catholic University, Largo A. Gemelli, 8, 00168 Rome, Italy ([email protected]). † These authors are co-first authors. Background. Ependymoma management remains challenging because of the inherent chemoresistance of this tumor. To determine whether ependymoma stem cells (SCs) might contribute to therapy resistance, we investigated the sensitivity of ependymoma SCs to temozolomide and etoposide. Methods. The efficacies of the two DNA damaging agents were explored in two ependymoma SC lines in vitro and in vivo models. Results. Ependymoma SC lines were highly sensitive to temozolomide and etoposide in vitro, but only temozolomide impaired tumor- initiation properties. Consistently, temozolomide but not etoposide showed significant antitumoral activity on ependymoma SC-driven subcutaneous and orthotopic xenografts by reducing the mitotic fraction. In vitro temozolomide at the EC 50 (10 mM) induced accu- mulation of cells in the G 2 /M phase that was unexpectedly accompanied by downregulation of p27 and p21 without modulation of full-length p53 (FLp53). Differentiation-committed ependymoma SCs acquired resistance to temozolomide. Inhibition of proliferation was partly due to apoptosis, that occurred earlier in differentiated cells as compared to neurospheres. The activation of apoptosis correlated with an increase in p53b/g isoforms without modulation of FLp53 under both serum-free and differentiation-promoting media. Incubation of cells in both conditions with temozolomide resulted in increased glioneuronal differentiation exhibiting elevated glial fibrillary acidic protein, galactosylceramidase, and bIII-tubulin expression compared to untreated controls. O 6 -methylguanine DNA methyltransferase (MGMT) transcript levels were very low in SCs, and were increased by treatment and, epigenetically, bydiffer- entiation through MGMT promoter unmethylation. Conclusion. Ependymoma growth might be impaired by temozolomide through preferential depletion of a less differentiated, more tumorigenic, MGMT-negative cell population with stem-like properties. Keywords: differentiation, ependymoma stem cells, MGMT, orthotopic models, temozolomide. Ependymomas are chemorefractory tumors of glial origin that occur in all age groups. 1 To date, ependymoma treatment is based on surgery and adjuvant radiotherapy, with a 5-year sur- vival rates as high as 70.8% in adults. However, prognosis is sub- stantially worse in children, because gross total resection is rarely achieved and the use of radiotherapy in very young patients is limited because of adverse side-effects. 2,3 Chemotherapy has been widely used in relapsed patients and in the pediatric popu- lation as adjuvant therapy or to delay, or even replace, radiother- apy. 4 – 7 To date, clinical trials in ependymoma have failed to demonstrate a clear overall survival benefit in the diverse subpopulations of patients. 8 – 11 Progress in this field has been hin- dered by the relatively low incidence of the disease and the pau- city of suitable in vitro and in vivo models. Cellular heterogeneity of many cancers has been explained as functional differences between stem-like, tumorigenic cells and more differentiated non-tumorigenic cells. Cancer stem cells (CSCs) are held to be responsible for tumor initiation, progression and treatment failure due to their intrinsic chemoresistance. Although it is still debated whether and to which tumors this model applies, it might lend the biological support to screen ther- apies that specifically target CSCs. 12,13 Cells that fulfill stemness Received 14 January 2014; accepted 14 January 2014 # The Author(s) 2014. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: [email protected]. Neuro-Oncology Neuro-Oncology 2014; 0, 1 – 11, doi:10.1093/neuonc/nou008 1 of 11 Neuro-Oncology Advance Access published February 12, 2014 by guest on February 14, 2014 http://neuro-oncology.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ependymoma stem cells are highly sensitive to temozolomidein vitro and in orthotopic models
Daniela Meco†, Tiziana Servidei†, Giuseppe Lamorte, Elena Binda, Vincenzo Arena, and Riccardo Riccardi
Department of Pediatric Oncology, Catholic University, Rome, Italy (D.M., T.S., R.R.); Istituto CSS - Mendel Laboratory, Rome, Italy (G.L.);Department of Biotechnology and Biosciences, Building U3, University of Milan Bicocca, Milan, Italy (E.B.); Institute of Pathology, CatholicUniversity, Rome, Italy (V.A.)
Corresponding Author: Tiziana Servidei, Department of Pediatric Oncology, Catholic University, Largo A. Gemelli, 8, 00168 Rome,Italy ([email protected]).†These authors are co-first authors.
Background. Ependymoma management remains challenging because of the inherent chemoresistance of this tumor. To determinewhether ependymoma stem cells (SCs) might contribute to therapy resistance, we investigated the sensitivity of ependymoma SCs totemozolomide and etoposide.
Methods. The efficacies of the two DNA damaging agents were explored in two ependymoma SC lines in vitro and in vivo models.
Results. Ependymoma SC lines were highly sensitive to temozolomide and etoposide in vitro, but only temozolomide impaired tumor-initiation properties. Consistently, temozolomide but not etoposide showed significant antitumoral activity on ependymoma SC-drivensubcutaneous and orthotopic xenografts by reducing the mitotic fraction. In vitro temozolomide at the EC50 (10 mM) induced accu-mulation of cells in the G2/M phase that was unexpectedly accompanied by downregulation of p27 and p21 without modulation offull-length p53 (FLp53). Differentiation-committed ependymoma SCs acquired resistance to temozolomide. Inhibition of proliferationwas partly due to apoptosis, that occurred earlier in differentiated cells as compared to neurospheres. The activation of apoptosiscorrelated with an increase in p53b/g isoforms without modulation of FLp53 under both serum-free and differentiation-promotingmedia. Incubation of cells in both conditions with temozolomide resulted in increased glioneuronal differentiation exhibiting elevatedglial fibrillary acidic protein, galactosylceramidase, and bIII-tubulin expression compared to untreated controls. O6-methylguanineDNA methyltransferase (MGMT) transcript levels were very low in SCs, and were increased by treatment and, epigenetically, by differ-entiation through MGMT promoter unmethylation.
Conclusion. Ependymoma growth might be impaired by temozolomide through preferential depletion of a less differentiated, moretumorigenic, MGMT-negative cell population with stem-like properties.
Keywords: differentiation, ependymoma stem cells, MGMT, orthotopic models, temozolomide.
Ependymomas are chemorefractory tumors of glial origin thatoccur in all age groups.1 To date, ependymoma treatment isbased on surgery and adjuvant radiotherapy, with a 5-year sur-vival rates as high as 70.8% in adults. However, prognosis is sub-stantially worse in children, because gross total resection is rarelyachieved and the use of radiotherapy in very young patients islimited because of adverse side-effects.2,3 Chemotherapy hasbeen widely used in relapsed patients and in the pediatric popu-lation as adjuvant therapy or to delay, or even replace, radiother-apy.4 – 7 To date, clinical trials in ependymoma have failed todemonstrate a clear overall survival benefit in the diverse
subpopulations of patients.8 – 11 Progress in this field has been hin-dered by the relatively low incidence of the disease and the pau-city of suitable in vitro and in vivo models.
Cellular heterogeneity of many cancers has been explained asfunctional differences between stem-like, tumorigenic cells andmore differentiated non-tumorigenic cells. Cancer stem cells(CSCs) are held to be responsible for tumor initiation, progressionand treatment failure due to their intrinsic chemoresistance.Although it is still debated whether and to which tumors thismodel applies, it might lend the biological support to screen ther-apies that specifically target CSCs.12,13 Cells that fulfill stemness
Received 14 January 2014; accepted 14 January 2014# The Author(s) 2014. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved.For permissions, please e-mail: [email protected].
Neuro-OncologyNeuro-Oncology 2014; 0, 1–11, doi:10.1093/neuonc/nou008
1 of 11
Neuro-Oncology Advance Access published February 12, 2014 by guest on February 14, 2014
http://neuro-oncology.oxfordjournals.org/D
ownloaded from

defining criteria have been isolated from ependymoma and iden-tified in a restricted subpopulation with a molecular signature re-sembling that of radial glial cells.14,15
Temozolomide (TMZ) and etoposide (VP16) are two DNA dam-aging agents that are used in the treatment of ependy-moma.11,16,17 Clinical trials are ongoing to optimize schedulesand dosing of these drugs (http://www.clinicaltrials.gov/).
The effects of TMZ and VP16 on ependymoma SCs have beenaddressed only in vitro studies, that found resistance or minorsensitivity of neurosphere-derived cells to both agents.18 – 20 Wehave previously established orthotopic ependymoma xenograftsdriven by cells with stem-like properties.21 This resource affordedus to explore for the first time the effects of TMZ and VP16 ontumorigenicity of ependymoma stem cells (SCs) and the antitu-moral activity of both agents on subcutaneous and intracranialependymoma SC-driven xenografts. Because only TMZ provedto be effective in vivo, we further investigated the cellular andmolecular mechanisms underlying the response to TMZ in epen-dymoma SCs.
Materials and methods
Cell cultures
We used the ependymoma lines EPP and EPV that we establishedfrom two recurrent infratentorial pediatric ependymoma by usingneural SC permissive conditions.21 Cells were grown in NeuroCultmedium (Stem Cell Technologies, Vancouver, BC, Canada) supple-mented with epidermal growth factor (EGF; 20 ng/mL; Sigma-Aldrich-Aldrich, Dorset, UK) and basic fibroblast growth factor(bFGF; 10 ng/mL; Peprotech, Rocky-Hill, NJ).
To induce differentiation, cells were seeded onto vesselscoated with poly-L-ornithine and laminin (Invitrogen, Carlsbad,CA) in Neurobasal (Invitrogen) supplemented with EGF andbFGF.15,21 After 24 h, medium was changed to 10% fetal bovineserum (FBS, Invitrogen)/Neurobasal without mitogens and cellswere allowed to differentiate for up to 14 days.
Cell proliferation assay
The antiproliferative effects of TMZ and VP16 were determined inundifferentiated and differentiated cells. TMZ was purchasedfrom Sigma-Aldrich and diluted in dimethyl sulfoxide to a stocksolution of 100 mM, whereas VP16 was that for clinical use(Teva Pharma B. V., Utrecht, ND). Vehicle or serial concentrationsof each drug were added to the medium and cells were culturedfor up to 7 days, with medium changed on day 3 and day 5. Cellnumber and viability were assessed by an automated cell counter(NucleoCounter 100TM, ChemoMetec, Allerød, Denmark), andexpressed as a percentage of the control. The half maximal ef-fective concentration (EC50) was calculated using the GraphPadPrism software package version 6.0 (GraphPad Software Inc.,San Diego, CA, USA).
Flow cytometry
For CD133 analysis, cells were exposed to serial concentrationsof TMZ and VP16 for 3 and 5 days. Cells were then collectedand labeled with monoclonal CD133/1-phycoerythrin-conjugatedantibody or isotype control antibody (Miltenyi Biotec, Bergisch
Gladbach, Germany) for 30 min at 48C. After washing, cellswere resuspended in 0.5 mL culture medium and analyzed byflow cytometry (CyAn Flow Cytometer, Beckman Coulter, OrangeCounty, CA).
For cell cycle analysis, ependymoma cells were incubated for24, 48, and 72 h in the presence of TMZ (10 mM and 100 mM) orvehicle and then fixed in 70% ethanol. Cell pellets were treatedwith 0.5 mg/mL RNAse (Sigma-Aldrich) in phosphate buffer saline(PBS) plus 0.1% saponin (Sigma-Aldrich), incubated at 378C for30 min before staining with 20 mg/mL propidium iodide(Sigma-Aldrich) for 30 min at 48C. Cell were then analyzed forDNA content by means of FACSCalibur flow cytometer (BectonDickinson, San Jose, CA).
Subcutaneous and intracranial transplantations
All experimental animal investigations complied with the guide-lines of the ‘‘Istituto Superiore di Sanita‘’’ (National Institute ofHealth, Rome, Italy) and the Ethical Committee of Catholic Uni-versity and all animal care was in accordance with local institu-tional guidelines.
For subcutaneous transplantations, 3×106 cells/100 mL 0.9%sodium chloride solution were inoculated together with an equalvolume of Matrigel (Becton Dickinson, Franklin Lakes, NJ) in bothflanks of five-week-old male nude CD1 nu/nu mice (Charles River,Calco, Italy). Tumor volume (TV) and body weight were monitoredtwice per week throughout the study. TVs were calculated by theformula: TV¼ d2×D/2, where d and D are the shortest and long-est diameters, respectively.
For the orthotopic model, 3×105 cells/10 mL PBS wereimplanted into the third ventricle of nude mice using a stereotaxicinjection frame (Kopf Instrument, Better Hospital EquipmentCorp, Miami Lakes, FL), after administration of general anesthesia(80 mg/kg ketamine + 10 mg/kg xylazine, Sigma-Aldrich). The in-jection coordinates were: lateral to right, 1 mm from bregma;dorsoventral, 23.5 mm. The animals were monitored daily untilsigns of neurologic deficit developed, at which time they wereeuthanized and their brains removed for histopathologic analysis.
Five days after subcutaneous or intracranial transplantations,mice were randomized in separate groups (8–10 mice/group),each group receiving one of the following treatments: only vehicle(control), TMZ (35 mg/kg for five times per week for 10 weeks, oralgavage), VP16 (15 mg/kg in one daily injection on days 1–3 oftreatment, intraperitoneal22) and combination of the two drugsat the indicated doses and schedules. VP16 and TMZ were extem-poraneously prepared as previously described.22,23 The dose ofTMZ that mimics a metronomic administration in mice was extra-polated from literature data.23 – 25
All statistical analyses were performed using the GraphPadPrism software package version 6.0 (GraphPad Software Inc).The survival days of animals were determined using the Kaplan-Meier plots and compared by the log-rank test. P values , .05were considered to be significant.
Tissue processing and immunohistochemistry
Xenograft specimens were fixed with 4% paraformaldehyde,paraffin-embedded and cut into 3-mm sections. Immunohisto-chemical analysis of monoclonal mouse anti-human Ki67
Meco et al.: TMZ antitumor effects in ependymoma SC models
2 of 11 Neuro-Oncology
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

(Novocastra Laboratories, Newcastle, UK) was carried out accord-ing to a standard protocol previously described.21
Western blot analysis
Cells were exposed to 10 mM or 100 mM TMZ for time periods ran-ging from 3 h up to 7 days. At the end of incubation, cells wereimmediately processed in lysis buffer.21 Total lysate was sub-jected to sodium dodecyl sulfate polyacrylamide gel electrophor-esis, transferred to Hybond nitrocellulose membrane (AmershamPharmacia, Buckinghamshire, UK), and probed with antibodiesto p53, p27, p21, caspase 3, Bcl2, anti-poly(ADP-ribose) polymer-ase (PARP), and actin, all from Santa Cruz Biotechnology (SantaCruz, CA). The membranes were incubated with horseradishperoxidase-conjugated secondary antibodies (Vector, Burlin-ghame, CA) and the immunoblots were visualized using the ECLdetection system (Amersham Pharmacia).
Real-Time Quantitative Reverse Transcriptase (RT) PCR
DNA and total RNA were extracted from cells using the AllPrepDNA/RNA Kit (Qiagen GmbH, Hilden, D). RNA was reverse-transcribed with High-Capacity cDNA Reverse Transcription kit(Applied Biosystems, Foster City, CA) according to the manufac-turer’s instructions. TaqMan gene expression assays for CD133,nestin, Olig2, Sox2, glial fibrillary acidic protein (GFAP), bIII-tubulin, O6-methylguanine-DNA methyltransferase (MGMT),galactosylceramidase (GalC), as well as for the reference normal-ization gene (hypoxanthine guanine phosphoribosyltransferase,HPRT) were obtained from Applied Biosystems. Each amplificationreaction was performed in triplicate on a 7500 Real-Time PCR Sys-tem (Applied Biosystems). The amount of each target mRNA wasnormalized to that of HPRT as previously reported.21
Methylation-specific PCR (MSP)
The methylation status of MGMT promoter was determined by atwo-stage methylation-specific PCR (MSP) approach with minormodifications.26 – 28 Genomic DNA (2 mg) was treated with sodiumbisulphite using the Epitech Kit (Qiagen) according to the manu-facturer’s instructions. Stage-1 PCR was performed to amplify a289-bp fragment of the MGMT gene by using primers that recog-nize the bisulfite-modified template but do not discriminate be-tween methylated and unmethylated alleles.27,28 In the stage-2PCR, the primer combinations allowed for the amplification of a122-bp fragment from methylated DNA or a 129-bp fragmentfrom unmethylated DNA.26 Each PCR product was separated on2% agarose gels.
Results
TMZ, but not VP16, impairs tumor-initiation propertiesof ependymoma SCs
We have previously established and characterized two ependy-moma cell lines enriched with stem-like cells, referred to as EPPand EPV.21 Both lines displayed similar levels of the defining mar-kers of ependymoma SCs, but showed a markedly different ex-pression of CD133. In order to investigate the effects of TMZand VP16 on ependymoma SCs, we treated the two lines with
increasing concentrations of each drug. Dose response experi-ments after a 3-day exposure showed a similar reduction in pro-liferation of both lines regardless of the CD133 phenotype(Fig. 1A). TMZ EC50s were 10.0+4.4 mM and 9.1+3.9 mM in EPPand EPV, respectively; these values are in the same range asthat of the glioblastoma cell line A172, previously reported asTMZ sensitive.29 VP16 also was highly effective in inhibiting cellproliferation, with EC50s of 0.5+0.1 mM and 0.2+0.1 mM, re-spectively in EPP and EPV lines.
Cytofluorimetric analysis showed that exposure to equitoxicdoses of TMZ or VP16 (5 mM TMZ and 0.2 mM VP16; 10 mM TMZand 0.5 mM VP16), did not reduce the expression of CD133 evenafter 5 days of treatment at the higher concentration of eachdrug that caused a 75% reduction in survival (Fig. 1B and datanot shown).
We previously found that ex vivo – rather than clonogenic –assays measure the residual tumorigenic potential of cells afterdrug challenging more reliably.21 Therefore, to address whethertreatment with either drug could translate into diminishedtumor initiation properties of ependymoma SCs, we stereotactic-ally injected an equal number of viable EPP cells after treatmentfor 5 days with TMZ or VP16 at a dose that reduces cell prolifer-ation by 50% (5 mM and 0.2 mM, respectively). TMZ treatmentsignificantly increased the latency of tumor development ascompared to vehicle-pretreated controls, in spite of substantiallyunchanged CD133 expression levels in the two experimental con-ditions (log-rank, P¼ .027) (Fig. 1C). By contrast, ex vivo treatmentwith VP16 did not substantially affect tumorigenicity (log-rank,P¼ .055 with respect to vehicle-treated cells). Altogether, thesedata indicate that TMZ selects a less tumorigenic subpopulationof ependymoma cells and reinforce our previously publishedfinding that CD133 expression does not segregate for stemnessproperties in ependymoma.21
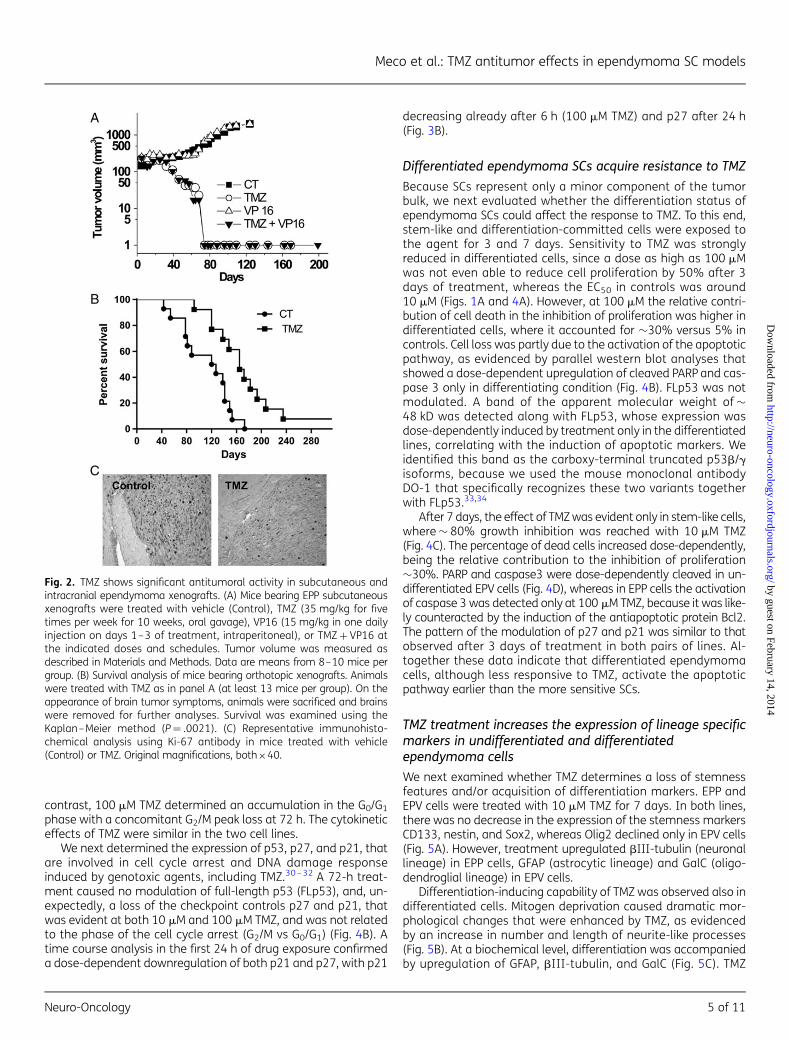
TMZ, but not VP16, shows significant antitumoral activity insubcutaneous and intracranial ependymoma xenografts
We next investigated the antitumoral effects of the two drugsadministered either as single agents or in combination in sub-cutaneous tumor models (Fig. 2A). TMZ alone induced completeresorption of tumors and, importantly, no regrowth was observedafter interruption of treatment up to the end of the experiment inall of the animals (8/8 animals, �120 d from the last administra-tion). VP16 administered alone was ineffective. The combinationof the two drugs did not exert any superior antitumoral efficacy ascompared to TMZ alone. The dose and schedule of VP16 that weused had previously been shown to prolong the growth delay ofhuman small cell lung carcinoma xenografts.22
In parallel experiments in orthotopic models, TMZ caused asignificant prolongation of survival (log-rank, P¼ .0021), whereasVP16 and TMZ + VP16 did not (log-rank, P¼ .88, and P¼ .343,respectively) (Fig. 2B, and data not shown). One mouse out of13 in the TMZ-group was potentially cured. TMZ strongly reducedthe proliferating fraction in orthotopic xenografts, as evidenced bystaining of samples for the expression of Ki-67 (Fig. 2C).
Weight loss was �12% in all of the treated groups and micecompletely recovered after drug administration was stopped(data not shown).
By comparing TMZ antitumoral activity in the two models, theefficacy was lower in orthotopic than in subcutaneous xenografts
Meco et al.: TMZ antitumor effects in ependymoma SC models
Neuro-Oncology 3 of 11
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

and this underlies the importance of models that more faithfullymirror the environment where human tumor develops for preclin-ical drug testing.
TMZ induces dose-dependent perturbations of cell cyclein ependymoma SCs
Because we observed that only TMZ inhibited the growth of epen-dymoma SCs-driven xenografts, we focused on determining the
cellular mechanisms underlying the response to this agent in cell-based assays.
To address whether decrease in cell number was due toimpaired proliferation, we analyzed the perturbations of cellcycle after exposure to TMZ (10 mM and 100 mM) for 24, 48 and72 h. Fluorescence-activated cell sorting analysis showed that10 mM TMZ induced an increase in the G2/M phase at 72 h, thatwas preceded by a time-dependent accumulation of cells in theS phase at the expense of the G0/G1 population (Fig. 3A). By
Fig. 1. TMZ and VP16 inhibit the proliferation of ependymoma SCs, but only TMZ reduces tumorigenicity. (A) Dose-dependent inhibition of theproliferation of ependymoma SC lines by TMZ and VP16. Cells were treated with either drug for 72 h. Viable and nonviable cells were counted byNucleoCounter. Each point represents the average value of two independent experiments, each in triplicate. (B) CD133+ fraction of EPP line treatedwith TMZ or VP16 for 3 and 5 days was assessed by flow cytometric analysis. The dual EGF receptor/HER2 inhibitor AEE788 at an equitoxic dosewas used as an example of drug that downregulates the CD133+ fraction.21 (C) Survival of mice (five animals per group) after orthotopic injectionof an equal number of viable EPP cells treated for 5 days with vehicle, or equitoxic doses of TMZ or VP16 (5 mM and 0.2 mM, respectively). Mice weresacrificed when brain tumor symptoms developed. Survival was examined using the Kaplan–Meier method (P¼ .027 for TMZ, and P¼ .055 for VP16).
Meco et al.: TMZ antitumor effects in ependymoma SC models
4 of 11 Neuro-Oncology
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from
contrast, 100 mM TMZ determined an accumulation in the G0/G1
phase with a concomitant G2/M peak loss at 72 h. The cytokineticeffects of TMZ were similar in the two cell lines.
We next determined the expression of p53, p27, and p21, thatare involved in cell cycle arrest and DNA damage responseinduced by genotoxic agents, including TMZ.30 – 32 A 72-h treat-ment caused no modulation of full-length p53 (FLp53), and, un-expectedly, a loss of the checkpoint controls p27 and p21, thatwas evident at both 10 mM and 100 mM TMZ, and was not relatedto the phase of the cell cycle arrest (G2/M vs G0/G1) (Fig. 4B). Atime course analysis in the first 24 h of drug exposure confirmeda dose-dependent downregulation of both p21 and p27, with p21
decreasing already after 6 h (100 mM TMZ) and p27 after 24 h(Fig. 3B).
Differentiated ependymoma SCs acquire resistance to TMZ
Because SCs represent only a minor component of the tumorbulk, we next evaluated whether the differentiation status ofependymoma SCs could affect the response to TMZ. To this end,stem-like and differentiation-committed cells were exposed tothe agent for 3 and 7 days. Sensitivity to TMZ was stronglyreduced in differentiated cells, since a dose as high as 100 mMwas not even able to reduce cell proliferation by 50% after 3days of treatment, whereas the EC50 in controls was around10 mM (Figs. 1A and 4A). However, at 100 mM the relative contri-bution of cell death in the inhibition of proliferation was higher indifferentiated cells, where it accounted for �30% versus 5% incontrols. Cell loss was partly due to the activation of the apoptoticpathway, as evidenced by parallel western blot analyses thatshowed a dose-dependent upregulation of cleaved PARP and cas-pase 3 only in differentiating condition (Fig. 4B). FLp53 was notmodulated. A band of the apparent molecular weight of �48 kD was detected along with FLp53, whose expression wasdose-dependently induced by treatment only in the differentiatedlines, correlating with the induction of apoptotic markers. Weidentified this band as the carboxy-terminal truncated p53b/gisoforms, because we used the mouse monoclonal antibodyDO-1 that specifically recognizes these two variants togetherwith FLp53.33,34
After 7 days, the effect of TMZ was evident only in stem-like cells,where � 80% growth inhibition was reached with 10 mM TMZ(Fig. 4C). The percentage of dead cells increased dose-dependently,being the relative contribution to the inhibition of proliferation�30%. PARP and caspase3 were dose-dependently cleaved in un-differentiated EPV cells (Fig. 4D), whereas in EPP cells the activationof caspase 3 was detected only at 100 mM TMZ, because it was like-ly counteracted by the induction of the antiapoptotic protein Bcl2.The pattern of the modulation of p27 and p21 was similar to thatobserved after 3 days of treatment in both pairs of lines. Al-together these data indicate that differentiated ependymomacells, although less responsive to TMZ, activate the apoptoticpathway earlier than the more sensitive SCs.
TMZ treatment increases the expression of lineage specificmarkers in undifferentiated and differentiatedependymoma cells
We next examined whether TMZ determines a loss of stemnessfeatures and/or acquisition of differentiation markers. EPP andEPV cells were treated with 10 mM TMZ for 7 days. In both lines,there was no decrease in the expression of the stemness markersCD133, nestin, and Sox2, whereas Olig2 declined only in EPV cells(Fig. 5A). However, treatment upregulated bIII-tubulin (neuronallineage) in EPP cells, GFAP (astrocytic lineage) and GalC (oligo-dendroglial lineage) in EPV cells.
Differentiation-inducing capability of TMZ was observed also indifferentiated cells. Mitogen deprivation caused dramatic mor-phological changes that were enhanced by TMZ, as evidencedby an increase in number and length of neurite-like processes(Fig. 5B). At a biochemical level, differentiation was accompaniedby upregulation of GFAP, bIII-tubulin, and GalC (Fig. 5C). TMZ
Fig. 2. TMZ shows significant antitumoral activity in subcutaneous andintracranial ependymoma xenografts. (A) Mice bearing EPP subcutaneousxenografts were treated with vehicle (Control), TMZ (35 mg/kg for fivetimes per week for 10 weeks, oral gavage), VP16 (15 mg/kg in one dailyinjection on days 1–3 of treatment, intraperitoneal), or TMZ + VP16 atthe indicated doses and schedules. Tumor volume was measured asdescribed in Materials and Methods. Data are means from 8–10 mice pergroup. (B) Survival analysis of mice bearing orthotopic xenografts. Animalswere treated with TMZ as in panel A (at least 13 mice per group). On theappearance of brain tumor symptoms, animals were sacrificed and brainswere removed for further analyses. Survival was examined using theKaplan–Meier method (P¼ .0021). (C) Representative immunohisto-chemical analysis using Ki-67 antibody in mice treated with vehicle(Control) or TMZ. Original magnifications, both×40.
Meco et al.: TMZ antitumor effects in ependymoma SC models
Neuro-Oncology 5 of 11
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

increased further the expression of these markers, mostly in EPP,where there was a 4-fold increase in GFAP, and approximately a2-fold increase in GalC and bIII-tubulin.
To address whether treatment could modulate differentiationmarkers in vivo, we stained sections for GFAP expression, whichshows the highest expression in vitro. However, no obvious differ-ence was observed between control and treated animals (datanot shown).
MGMT is induced by TMZ treatment and differentiation
The most cytotoxic effects of alkylating agents such as TMZ aredue to methylation of guanine at position O6. The DNA repair en-zyme MGMT removes the methyl group from O6-methylguanine,thus protecting cells against DNA damage and cell death.35
MGMT expression is the best characterized mechanism of TMZ re-sistance.27,35,36 Loss of MGMT expression and diminishedDNA-repair activity have been related to epigenetic silencing ofthe MGMT gene mediated by promoter methylation.
Ependymoma SCs are expected to not express MGMT becauseof their high responsiveness to TMZ. Indeed, levels of MGMT mRNAwere very low in both lines, and dose-dependently induced byTMZ treatment (�5-fold after 100 mM TMZ) (Fig. 6A). Interesting-ly, differentiation highly increased MGMT expression in both lines.
We next assessed MGMT promoter methylation by MSP instem-like conditions and after differentiation. We used the T98Gline (which expresses high level of the protein) and peripheralblood lymphocytes as a control for unmethylated MGMT37 andthe U87 line as a control for methylated MGMT (Fig. 6B).29 EPPand EPV SCs showed hypermethylation of the MGMT promoter,whereas unmethylated MGMT was present in EPP cells and totallyabsent in EPV cells (Fig. 6C). Unmethylated MGMT increased withthe differentiation, consistently with the increase in mRNA ex-pression. TMZ treatment did not determine any change in thelevel of MGMT promoter methylation (data not shown).
DiscussionIn this study we found that ependymoma SCs are highly sensitiveto TMZ at clinically achievable concentrations both in vitro and invivo, whereas differentiated cells acquire resistance to the agent.TMZ preferentially depletes cells with stemness characteristics,because treatment results in a decrease in tumor-initiation prop-erties and induction of differentiation markers. Ependymoma SCsare specifically sensitive to TMZ, because VP16 (present work) andthe dual EGF receptor/HER2 inhibitor AEE78821 do not determinea statistical significant survival difference of mice bearing
Fig. 3. TMZ blocks ependymoma SCs in G0/G1 or G2/M phases of the cell cycle, while downregulating the checkpoint controls p27 and p21.(A) Time-course analysis of the effects of TMZ on cell cycle distribution in ependymoma SCs. Cells were treated with TMZ at 10m and 100 mM forthe indicated periods of time. The percentages of the total cell population in the different phases of the cell cycle were assessed by flow cytometry.(B) Western blot analysis of the regulatory proteins p27 and p21 in the two ependymoma SC lines treated with the indicated concentrations of TMZ fordifferent time intervals.
Meco et al.: TMZ antitumor effects in ependymoma SC models
6 of 11 Neuro-Oncology
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

ependymoma SC-driven orthotopic xenografts in ex vivo and invivo models.
TMZ increased the expression of the lineage-specific markersbIII-tubulin, GalC and GFAP in ependymoma cells in both stem-like and differentiating conditions. This might be a consequenceof the selective depletion of cells with a more immature pheno-type or/and of the inherent differentiation-inducing capability ofTMZ. Eritroid differentiation of leukemia cells after TMZ has beenascribed to either treatment-induced hypomethylation ofdifferentiation-related genes or cell cycle arrest consequent toDNA damage.38 In glioblastoma SCs, TMZ upregulates the expres-sion of bIII-tubulin, microtubule-associated protein 2, and GFAP,and potentiates BMP2-induced differentiation, that is accompan-ied by a parallel decrease in MGMT expression and sensitization ofcells to TMZ through a dramatic increase in apoptosis.39 In our
ependymoma cells, some induction of apoptosis was observedwith a different onset in the two experimental conditions, occur-ring earlier in the slow-dividing differentiated cells, and at a latertime point in the fast-dividing SCs. However, the inhibition of pro-liferation was mainly due to a p53-independent arrest in G2/M orG0/G1 phase after exposure to a low or high concentration of TMZ,respectively. TMZ arrests cells in both G0/G1 and G2/M dependingon the specific cellular context and tumor type, p53 status, andDNA mismatch repair system.32,40 – 42 Multiple effects usuallyoccur in the cell cycle after treatment with a drug, as the cellsflow through G1, S and G2/M checkpoints (blocking activity,death rate, delay and recycling), each one with its own kineticsand concentration thresholds.43 The activation of proteinsinvolved in cell cycle checkpoints after exposure to TMZ is dose-and time-dependent.41 It is likely that ependymoma cells
Fig. 4. Differentiated ependymoma cells, although less sensitive to TMZ than undifferentiated cells, show an earlier onset of apoptosis induction.Ependymoma cells were cultured in defined neurosphere medium (Undiff) or in medium containing 10% FBS (Diff). Cells were treated for 3 days (A)and 7 days (C) with vehicle (Ct), 10m or 100 mM TMZ. Live and dead cells were counted by NucleoCounter. Inhibition of proliferation was calculated by theformula: (1- live drug-treated cells/live vehicle-treated cells)×100. Cell death was the difference between the percentage of dead cells in drug-treatedand vehicle-treated samples. (B), (D). Modulation of the pro-apoptotic (cleaved PARP and caspase 3) and anti-apoptotic (Bcl2) proteins and of FLp53,p53b/g, p27 and p21. Cells were treated with vehicle (Ct) or TMZ at the indicated concentrations (mM) for 3 days (B) or 7 days (D). Cell lysates wereimmunoblotted for the aforementioned proteins.
Meco et al.: TMZ antitumor effects in ependymoma SC models
Neuro-Oncology 7 of 11
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

exposed to a low concentration are still able to progress slowlythrough S phase to eventually block in G2/M. After a high dose,cells might “freeze” in G0/G1 – where the majority of cells reside –because of high levels of DNA damage, that lead to an irreversibleand immediate block.
Unexpectedly, cell cycle perturbations were accompanied by adecrease in the expression of p21 and p27, that, so far, remainselusive. In glioblastoma, TMZ triggers a canonical p53-dependentpathway with upregulation of p21 leading to cell cycle arrest andapoptosis, but similar effects have been described also in cellscarrying a mutated p53.32,40,42 p21 and p27 function as inhibitorsof cyclin dependent kinases and regulate cell cycle progressionand differentiation.30 However, some evidence suggests addition-al functions for these proteins, including pro-survival properties.44
If this hypothesis holds true, their downregulation might facilitatethe apoptotic process. Indeed, we found that the levels of p21and p27 were inversely related to the levels of apoptotic markers.
The presence of the alternative spliced variants p53b/g is anovel finding in ependymoma. Several isoforms of p53 havebeen found to be differentially expressed in cancer, althoughtheir functional role is not fully understood. Both p53b andp53g are suggested to influence carcinogenesis and drug-sensitivity in a tumor-dependent manner by modulating cellcycle progression, senescence, and apoptosis.34,45 In ependy-moma cells the p53b/g isoforms seem to have functional signifi-cance in that they were modulated by TMZ unlike FLp53, and thisparalleled upregulation of apoptotic markers. Of note, the expres-sion levels of p53b/g isoforms were higher in ependymoma neu-rospheres than in their differentiated counterparts, similarly towhat was previously found in melanoma cells.33 Therefore,p53b/g isoforms might play a role in modulating stemness/differ-entiation pathways.
TMZ is the first-line therapy in glioblastoma and the persist-ence of chemoresistant glioblastoma SCs may account for fre-quent treatment failure. In vitro, TMZ enriches the fraction ofglioblastoma cells with stem-like properties and elevated MGMTexpression, suggesting that MGMT is a key determinant of intrinsicor acquired resistance of glioblastoma SCs to the agent.46 – 48 Inother works TMZ preferentially depletes the glioblastoma SC com-partment, being the sensitivity inversely related to the expressionof the MGMT protein.26,49
Because of the lack of ependymoma models, no study has sofar investigated MGMT expression and promoter methylation sta-tus in SCs from this tumor. In ependymal tumors positive MGMTmethylation status is not frequent, this data correlating with thepoor response to TMZ treatment in ependymoma clinical trials.50
We found that the two ependymoma SC lines displayed highmethylation status of MGMT promoter and lack of MGMT expres-sion, in keeping with their high susceptibility to TMZ. Of note, pro-longed treatment increased MGMT expression. On the whole, ourdata suggest that in ependymoma MGMT might be a mechanism
Fig. 5. Differentiation-inducing capability of TMZ on ependymoma cells.(A) Real time quantitative RT-PCR analysis of expression of CD133, Olig2,Sox2, nestin, GFAP, GalC and bIII-tubulin. Ependymoma SCs were treatedwith 10 mM TMZ for 7 days. mRNA level of each target gene wasnormalized to the level of the reference gene HPRT in each sample.Means+SD relative to untreated controls, which were used ascalibrators (1¼ no change). (B) Phase-bright photomicrographs ofdifferentiated EPP cells treated with vehicle (Control) or 10 mM TMZ for 7days. Original magnification, both×20. (C) Real-time quantitative RT-PCR
of expression of GFAP, GalC and bIII– tubulin in ependymomaneurospheres (Undiff) and differentiated cells (Diff) treated with vehicle(Control) or 10 mM TMZ for 7 days. Level of each target gene wasnormalized to the level of the reference gene HPRT in each sample.Average values from two independent amplifications each in triplicateare shown.
Meco et al.: TMZ antitumor effects in ependymoma SC models
8 of 11 Neuro-Oncology
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

of acquired resistance to TMZ through drug-mediated selection ofan inherently resistant SC subpopulation that expresses MGMT orepigenetic induction of MGMT by promoter hypomethylation.36
Alternatively, the increase in MGMT expression might be relatedto the pro-differentiation properties of TMZ, in agreement withthe increase in unmethylated MGMT promoter and MGMT ex-pression that we found in differentiated ependymoma lines.
Epigenetic changes mediated by DNA methylation ap-pear to be critical in the pathogenesis of tumors, including
ependymoma.51,52 A dynamic transition between active and re-pressive state of gene transcription could provide survival advan-tages to the CSC subpopulation.53 Because MGMT is a mechanismof defense against mutagenic alkylation lesions,36 MGMT-silencedcancer cells, such as our ependymoma SCs, might acquire a mu-tator phenotype that predisposes them to subsequent geneticand epigenetic changes essential in tumorigenesis.54 In thisview, the acquisition of MGMT might be a differentiated functionto protect cells against genotoxic insults. Interestingly, MGMTappears to be developmentally regulated in human brain, in-creasing with gestation age and later in life.36
No clinical nor preclinical studies have so far addressed the ef-ficacy of TMZ given at low doses on continuous (metronomic) ad-ministration in ependymoma. We found that TMZ on ametronomic schedule significantly reduces tumor growth rate inboth subcutaneous and orthotopic models. Our in vitro experi-ments indicate that prolonged exposure to a low dose of TMZis required to induce apoptosis in ependymoma SCs. On theother hand, exposure to TMZ increases the expression of dif-ferentiation markers and of MGMT in both stem-like anddifferentiation-committed cells, both mechanisms being asso-ciated with resistance to TMZ, which suggests that even dailyadministration could spare differentiated cells. However, a metro-nomic schedule could yet increase the therapeutic efficacy ofTMZ, because protracted administration inactivates MGMT moreeffectively than short treatment,55 thus counteracting the emer-gence of MGMT-expressing clones. Indeed, metronomic treat-ment is effective in glioblastoma with unmethylated MGMTstatus.56,57 Extended dosing schedule offers other potentialadvantages, such as enhanced pro-apoptotic activity, antiangio-genic effects, sensitization of tumor cells to radiotherapy, and,because of higher tolerability, association with other chemother-apeutics.57 – 59 Finally, drug-induced differentiation in itself couldcontribute to the therapeutic activity of TMZ, by reducing the“stemness” features of cells, hence tumorigenicity.60 We previ-ously found that ex vivo differentiation of ependymoma SCs sig-nificantly prolonged the survival of xenografted mice.21
In conclusion, ependymoma SCs appear to be resistant toVP16 and specifically sensitive to TMZ, that exerts antiproli-ferative, pro-apoptotic, and pro-differentiation effects on anMGMT-negative cell population with stem-like properties. Opti-mized TMZ dosing schemes and schedules might increase thetherapeutic potential of this agent in the treatment of patientswith ependymoma.
FundingFondazione per l’Oncologia Pediatrica.
AcknowledgmentsThis work was supported by Fondazione per l’Oncologia Pediatrica.
Conflict of interest Statement. The authors disclose any commercialaffiliations or financial interests that may be considered conflicts ofinterest regarding the submitted manuscript.
Fig. 6. MGMT expression is induced by TMZ treatment and differentiation.(A) The transcript expression levels of MGMT gene were measured byreal-time quantitative RT-PCR in undifferentiated (Undiff) anddifferentiated (Diff) ependymoma lines. Undifferentiated cells weretreated with the indicated concentration of TMZ for 7 days. Averagevalues from two independent amplifications each in triplicate areshown. (B) MGMT promoter methylation was examined by a nestedtwo-stage PCR using primers specific for the unmethylated (u) andmethylated (m) promoter in positive controls for unmethylated MGMT(T98G cells and normal peripheral blood lymphocytes - PBL) and formethylated MGMT (U87 cells). Water was used as a negative control forthe PCR. A 100-bp marker ladder was loaded to estimate molecularsize, as shown on the left scale; the sizes of PCR products are indicatedon the right scale. The stage-1 PCR amplified a 289-bp fragment of theMGMT gene. (C) Results of the stage-2 PCR using methylation-specificprimers for MGMT gene in undifferentiated (Undiff) and differentiated(Diff) ependymoma lines.
Meco et al.: TMZ antitumor effects in ependymoma SC models
Neuro-Oncology 9 of 11
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

References1. Witt H, Korshunov A, Pfister SM, et al. Molecular approaches to
ependymoma: the next step(s). Curr Opin Neurol. 2012;25:745–750.
2. Grundy RG, Wilne SA, Weston CL, et al. Children’s Cancer andLeukaemia Group (formerly UKCCSG) Brain Tumour Committee.Primary postoperative chemotherapy without radiotherapy forintracranial ependymoma in children: the UKCCSG/SIOP prospectivestudy. Lancet Oncol. 2007;8:696–705.
3. Koos B, Bender S, Witt H, et al. The transcription factor evi-1 isoverexpressed, promotes proliferation, and is prognosticallyunfavorable in infratentorial ependymomas. Clin Cancer Res. 2011;17:3631–3637.
4. Bouffet E, Perilongo G, Canete A, et al. Intracranial ependymomas inchildren: a critical review of prognostic factors and a plea forcooperation. Med Pediatr Oncol. 1998;30:319–329.
5. de Bont JM, Packer RJ, Michiels EM, et al. Biological backgroundof pediatric medulloblastoma and ependymoma: a review froma translational research perspective. Neuro Oncol. 2008;10:1040–1060.
6. Kilday JP, Rahman R, Dyer S, et al. Pediatric ependymoma: biologicalperspectives. Mol Cancer Res. 2009;7:765–786.
7. Shonka NA. Targets for therapy in ependymoma. Target Oncol. 2011;6:163–169.
8. Atkinson JM, Shelat AA, Carcaboso AM, et al. An integrated in vitroand in vivo high-throughput screen identifies treatment leads forependymoma. Cancer Cell. 2011;20:384–399.
9. Massimino M, Gandola L, Barra S, et al. Infant ependymoma in a10-year AIEOP (Associazione Italiana Ematologia OncologiaPediatrica) experience with omitted or deferred radiotherapy. Int JRadiat Oncol Biol Phys. 2011;80:807–814.
10. Witt H, Mack SC, Ryzhova M, et al. Delineation of two clinically andmolecularly distinct subgroups of posterior fossa ependymoma.Cancer Cell. 2011;20:143–157.
11. Zacharoulis S, Ashley S, Moreno L, et al. Treatment and outcome ofchildren with relapsed ependymoma: a multi-institutionalretrospective analysis. Childs Nerv Syst. 2010;26:905–911.
12. Hadjipanayis CG, Van Meir EG. Brain cancer propagating cells:biology, genetics and targeted therapies. Trends Mol Med. 2009;15:519–530.
13. Magee JA, Piskounova E, Morrison SJ. Cancer stem cells: impact,heterogeneity, and uncertainty. Cancer Cell. 2012;21:283–296.
14. Singh SK, Clarke ID, Terasaki M, et al. Identification of a cancer stemcell in human brain tumors. Cancer Res. 2003;63:5821–5828.
15. Taylor MD, Poppleton H, Fuller C, et al. Radial glia cells are candidatestem cells of ependymoma. Cancer Cell. 2005;8:323–335.
16. Akyuz C, Demir HA, Varan A, et al. Temozolomide in relapsedpediatric brain tumors: 14 cases from a single center. Childs NervSyst. 2012;28:111–116.
17. Teo C, Nakaji P, Symons P, et al. Ependymoma. Childs Nerv Syst. 2003;19:270–285.
18. Guan S, Shen R, Lafortune T, et al. Establishment andcharacterization of clinically relevant models of ependymoma: atrue challenge for targeted therapy. Neuro Oncol. 2011;13:748–758.
19. Hussein D, Punjaruk W, Storer LC, et al. Pediatric brain tumor cancerstem cells: cell cycle dynamics, DNA repair, and etoposide extrusion.Neuro Oncol. 2011;13:70–83.
20. Milde T, Kleber S, Korshunov A, et al. A novel human high-riskependymoma stem cell model reveals the differentiation-inducing
potential of the histone deacetylase inhibitor Vorinostat. ActaNeuropathol. 2011;122:637–650.
21. Servidei T, Meco D, Trivieri N, et al. Effects of epidermal growth factorreceptor blockade on ependymoma stem cells in vitro and inorthotopic mouse models. Int J Cancer. 2012;131:E791–E803.
22. Nemati F, Livartowski A, De Cremoux P, et al. Distinctive potentiatingeffects of cisplatin and/or ifosfamide combined with etoposide inhuman small cell lung carcinoma xenografts. Clin Cancer Res.2000;6:2075–2086.
23. de Vries NA, Bruggeman SW, Hulsman D, et al. Rapid and robusttransgenic high-grade glioma mouse models for therapyintervention studies. Clin Cancer Res. 2010;16:3431–3441.
24. Banissi C, Ghiringhelli F, Chen L, et al. Treg depletion with a low-dosemetronomic temozolomide regimen in a rat glioma model. CancerImmunol Immunother. 2009;58:1627–1634.
25. Clarke JL, Iwamoto FM, Sul J, et al. Randomized phase II trial ofchemoradiotherapy followed by either dose-dense or metronomictemozolomide for newly diagnosed glioblastoma. J Clin Oncol.2009;27:3861–3867.
26. Beier D, Rohrl S, Pillai DR, et al. Temozolomide preferentially depletescancer stem cells in glioblastoma. Cancer Res. 2010;68:5706–5715.
27. Hegi ME, Diserens AC, Gorlia T, et al. MGMT gene silencing and benefitfrom temozolomide in glioblastoma. N Engl J Med. 2005;352:997–1003.
28. Palmisano WA, Divine KK, Saccomanno G, et al. Predicting lungcancer by detecting aberrant promoter methylation in sputum.Cancer Res. 2000;60:5954–5958.
29. Gaspar N, Marshall L, Perryman L, et al. MGMT-independenttemozolomide resistance in pediatric glioblastoma cells associatedwith a PI3-kinase-mediated HOX/stem cell gene signature. CancerRes. 2010;70:9243–9252.
30. Lapenna S, Giordano A. Cell cycle kinases as therapeutic targets forcancer. Nat Rev Drug Discov. 2009;8:547–566.
31. Mhaidat NM, Zhang XD, Allen J, et al. Temozolomide inducessenescence but not apoptosis in human melanoma cells. Br JCancer. 2007;97:1225–1233.
32. Sato Y, Kurose A, Ogawa A, et al. Diversity of DNA damage responseof astrocytes and glioblastoma cell lines with various p53 status totreatment with etoposide and temozolomide. Cancer Biol Ther.2009;8:452–457.
33. Avery-Kiejda KA, Zhang XD, Adams LJ, et al. Small molecular weightvariants of p53 are expressed in human melanoma cells and areinduced by the DNA-damaging agent cisplatin. Clin Cancer Res.2012;14:1659–1668.
34. Khoury MP, Bourdon JC. The isoforms of the p53 protein. Cold SpringHarb Perspect Biol. 2010;2:a000927.
35. Weller M, Stupp R, Reifenberger G, et al. MGMT promoter methylationin malignant gliomas: ready for personalized medicine?. Nat RevNeurol. 2010;6:39–51.
36. Christmann M, Verbeek B, Roos WP, et al. O(6)-Methylguanine-DNAmethyltransferase (MGMT) in normal tissues and tumors: Enzymeactivity, promoter methylation and immunohistochemistry.Biochim Biophys Acta. 2012;1816:179–190.
37. Sciuscio D, Diserens AC, van Dommelen K, et al. Extent and patternsof MGMT promoter methylation in glioblastoma- and respectiveglioblastoma-derived spheres. Clin Cancer Res. 2011;17:255–266.
38. Zucchetti M, Catapano CV, Filippeschi S, et al. Temozolomide induceddifferentiation of K562 leukemia cells is not mediated by genehypomethylation. Biochem Pharmacol. 1989;38:2069–2075.
Meco et al.: TMZ antitumor effects in ependymoma SC models
10 of 11 Neuro-Oncology
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from

39. Persano L, Pistollato F, Rampazzo E, et al. BMP2 sensitizesglioblastoma stem-like cells to Temozolomide by affecting HIF-1astability and MGMT expression. Cell Death Dis. 2012;3:e412.
40. Bocangel DB, Finkelstein S, Schold SC, et al. Multifaceted resistance ofgliomas to temozolomide. Clin Cancer Res. 2002;8:2725–2734.
41. Caporali S, Falcinelli S, Starace G, et al. DNA damage induced bytemozolomide signals to both ATM and ATR: role of the mismatchrepair system. Mol Pharmacol. 2004;66:478–491.
42. Roos WP, Batista LF, Naumann SC, et al. Apoptosis in malignantglioma cells triggered by the temozolomide-induced DNA lesionO6-methylguanine. Oncogene. 2007;26:186–197.
43. Ubezio P, Lupi M, Branduardi D, et al. Quantitative assessment of thecomplex dynamics of G1, S, and G2-M checkpoint activities. CancerRes. 2009;69:5234–5240.
44. Coqueret O. New roles for p21 and p27 cell-cycle inhibitors: a functionfor each cell compartment?. Trends Cell Biol. 2003;13:65–70.
45. Silden E, Hjelle SM, Wergeland L, et al. Expression of TP53 isoformsp53b or p53g enhances chemosensitivity in TP53(null) cell lines.PLoS One. 2013;8:e56276.
46. Bleau AM, Hambardzumyan D, Ozawa T, et al. PTEN/PI3K/Akt pathwayregulates the side population phenotype and ABCG2 activity in gliomatumor stem-like cells. Cell Stem Cell. 2009;4:226–235.
47. Pistollato F, Abbadi S, Rampazzo E, et al. Intratumoral hypoxicgradient drives stem cells distribution and MGMT expression inglioblastoma. Stem Cells. 2010;28:851–862.
48. Sato A, Sunayama J, Matsuda K, et al. MEK-ERK signaling dictatesDNA-repair gene MGMT expression and temozolomide resistance ofstem-like glioblastoma cells via the MDM2-p53 axis. Stem Cells.2011;29:1942–1951.
49. Blough MD, Westgate MR, Beauchamp D, et al. Sensitivity totemozolomide in brain tumor initiating cells. Neuro Oncol. 2010;12:756–760.
50. Koos B, Peetz-Dienhart S, Riesmeier B, et al. O6-methylguanine-DNAmethyltransferase (MGMT) promoter methylation is significantly lessfrequent in ependymal tumours as compared to malignantastrocytic gliomas. Neuropathol Appl Neurobiol. 2010;36:356–358.
51. Mack SC, Witt H, Wang X, et al. Emerging insights into theependymoma epigenome. Brain Pathol. 2013;23:206–209.
52. Rogers HA, Kilday JP, Mayne C, et al. Supratentorial and spinalpediatric ependymomas display a hypermethylated phenotypewhich includes the loss of tumor suppressor genes involved in thecontrol of cell growth and death. Acta Neuropathol. 2012;123:711–725.
53. Munoz P, Iliou MS, Esteller M. Epigenetic alterations involved incancer stem cell reprogramming. Mol Oncol. 2012;6:620–636.
54. Martinez R, Esteller M. The DNA methylome of glioblastomamultiforme. Neurobiol Dis. 2010;39:40–46.
55. Tolcher AW, Gerson SL, Denis L, et al. Marked inactivation ofO6-alkylguanine-DNA alkyltransferase activity with protractedtemozolomide schedules. Br J Cancer. 2003;88:1004–1011.
56. Kong DS, Lee JI, Kim JH, et al. Phase II trial of low-dose continuous(metronomic) treatment of temozolomide for recurrentglioblastoma. Neuro Oncol. 2010;12:289–296.
57. Neyns B, Tosoni A, Hwu WJ, et al. Dose-dense temozolomideregimens: antitumor activity, toxicity, and immunomodulatoryeffects. Cancer. 2010;116:2868–2877.
58. Chen C, Xu T, Lu Y, et al. The efficacy of temozolomide for recurrentglioblastoma multiforme. Eur J Neurol. 2013;20:223–230.
59. Gasparini G. Metronomic scheduling: the future of chemotherapy?.Lancet Oncol. 2001;2:733–740.
60. Campos B, Wan F, Farhadi M, et al. Differentiation therapy exertsantitumor effects on stem-like glioma cells. Clin Cancer Res. 2010;16:2715–2728.
Meco et al.: TMZ antitumor effects in ependymoma SC models
Neuro-Oncology 11 of 11
by guest on February 14, 2014http://neuro-oncology.oxfordjournals.org/
Dow
nloaded from
Related Documents











