Journal of Alzheimer’s Disease 15 (2008) 11–28 11 IOS Press Environmental Enrichment Provides a Cognitive Reserve to be Spent in the Case of Brain Lesion Laura Mandolesi a,b , Paola De Bartolo a,c , Francesca Foti a,c , Francesca Gelfo a,b , Francesca Federico a,d , Maria Giuseppa Leggio a,c and Laura Petrosini a,c,∗ a IRCCS Santa Lucia Foundation, Rome, Italy b University of Naples “Parthenope”, Naples, Italy c Department of Psychology, University of Rome “La Sapienza”, Rome, Italy d University of Siena, Siena, Italy Communicated by Sigfrido Scarpa Abstract. To experimentally verify the reserve hypothesis, the influence of rearing conditions on the cognitive performances and on dendritic spines following basal forebrain lesions was analyzed. Adult rats reared in enriched or standard conditions were depleted of the cholinergic projection to the neocortex by 192 IgG-saporin injection into Ch4 region of basal forebrain. Their performance in spatial tasks was compared with that of intact animals reared in analogous conditions. Furthermore, number and density of dendritic spines of the layer-III parietal pyramidal neurons were analyzed. Cholinergic depletion of forebrain cortex resulted in impaired performances in most behavioral tasks in animals reared in standard conditions. Conversely, the enriched lesioned animals did not exhibit most deficits evoked by cholinergic lesion, even if some deficits, such as perseverative behaviors, were still present. The pyramidal neurons exhibited an increased spine number and density in the lesioned animals reared in standard conditions. In the enriched lesioned animals, the enhancement of spine number and density elicited by the rearing condition was fully maintained but not further increased in the presence of the lesion. Thus, rearing in an enriched environment results in the development of brain and cognitive reserves that reduce the cognitive impairment following forebrain lesions. Keywords: Basal forebrain, cognitive impairment, 192 IgG-saporin, neuronal morphology, spatial learning INTRODUCTION Since the pioneering work by Mortimer [45] advanc- ing a relationship between psychosocial risk factors and expression of dementia of Alzheimer’s type, it has been repeatedly reported that the cognitive impairment following brain lesions is the end result of the interac- tive effects of pathological tissue damage and the indi- vidual’s capacity to compensate for that damage, i.e., ∗ Corresponding author: Laura Petrosini, Department of Psychol- ogy, University “Sapienza” of Rome, Via dei Marsi 78, 00185 Rome, Italy. Tel./Fax: +39 0649917522; E-mail: laura.petrosini@ uniroma1.it. the so-called “cognitive reserve” [61,70]. The cogni- tive reserve model posits that subjects can develop re- sources that reduce the risk of later cognitive impair- ment, hypothesizing that a subject with greater brain reserve capacity can sustain a greater amount of brain damage before reaching the threshold for clinical ex- pression of cognitive impairment [55]. To build the cognitive reserve, several factors, whether anatomical or functional in nature, interact each other [43]. They include complex experiences, such as high level of ed- ucation, lifestyle choices, and sustained cognitive en- gagement [17,53,62–64]. In addition, there are the components linked to the brain structures whose effi- ciency, enhanced by repeated use, allows effective in- ISSN 1387-2877/08/$17.00 2008 – IOS Press and the authors. All rights reserved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Alzheimer’s Disease 15 (2008) 11–28 11IOS Press
Environmental Enrichment Provides aCognitive Reserve to be Spent in the Case ofBrain Lesion
Laura Mandolesia,b, Paola De Bartoloa,c, Francesca Fotia,c, Francesca Gelfoa,b, Francesca Federicoa,d,Maria Giuseppa Leggioa,c and Laura Petrosinia,c,∗aIRCCS Santa Lucia Foundation, Rome, ItalybUniversity of Naples “Parthenope”, Naples, ItalycDepartment of Psychology, University of Rome “La Sapienza”, Rome, ItalydUniversity of Siena, Siena, Italy
Communicated by Sigfrido Scarpa
Abstract. To experimentally verify the reserve hypothesis, the influence of rearing conditions on the cognitive performances andon dendritic spines following basal forebrain lesions was analyzed. Adult rats reared in enriched or standard conditions weredepleted of the cholinergic projection to the neocortex by 192 IgG-saporin injection into Ch4 region of basal forebrain. Theirperformance in spatial tasks was compared with that of intact animals reared in analogous conditions. Furthermore, number anddensity of dendritic spines of the layer-III parietal pyramidal neurons were analyzed. Cholinergic depletion of forebrain cortexresulted in impaired performances in most behavioral tasks in animals reared in standard conditions. Conversely, the enrichedlesioned animals did not exhibit most deficits evoked by cholinergic lesion, even if some deficits, such as perseverative behaviors,were still present. The pyramidal neurons exhibited an increased spine number and density in the lesioned animals reared instandard conditions. In the enriched lesioned animals, the enhancement of spine number and density elicited by the rearingcondition was fully maintained but not further increased in the presence of the lesion. Thus, rearing in an enriched environmentresults in the development of brain and cognitive reserves that reduce the cognitive impairment following forebrain lesions.
Keywords: Basal forebrain, cognitive impairment, 192 IgG-saporin, neuronal morphology, spatial learning
INTRODUCTION
Since the pioneering work by Mortimer [45] advanc-ing a relationship between psychosocial risk factorsand expression of dementia of Alzheimer’s type, it hasbeen repeatedly reported that the cognitive impairmentfollowing brain lesions is the end result of the interac-tive effects of pathological tissue damage and the indi-vidual’s capacity to compensate for that damage, i.e.,
∗Corresponding author: Laura Petrosini, Department of Psychol-ogy, University “Sapienza” of Rome, Via dei Marsi 78, 00185Rome, Italy. Tel./Fax: +39 0649917522; E-mail: [email protected].
the so-called “cognitive reserve” [61,70]. The cogni-tive reserve model posits that subjects can develop re-sources that reduce the risk of later cognitive impair-ment, hypothesizing that a subject with greater brainreserve capacity can sustain a greater amount of braindamage before reaching the threshold for clinical ex-pression of cognitive impairment [55]. To build thecognitive reserve, several factors, whether anatomicalor functional in nature, interact each other [43]. Theyinclude complex experiences, such as high level of ed-ucation, lifestyle choices, and sustained cognitive en-gagement [17,53,62–64]. In addition, there are thecomponents linked to the brain structures whose effi-ciency, enhanced by repeated use, allows effective in-
ISSN 1387-2877/08/$17.00 2008 – IOS Press and the authors. All rights reserved

12 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
formation processing, enhanced retrieval of memoriesand superior problem solving.
The correlation between cognitive reserve and com-plex experiences is consistent with the model of theenvironmental enrichment (EE). Exposure to complexenvironments strengthens synaptic neurotransmissionand plasticity inducing structural and biochemical mod-ifications of neurons [30,37,54,66,67], increase of sur-vival or decrease of apoptosis of regenerating neu-rons [30,46,66,74], and enhancement of expression ofmolecules involved in neuronal signaling [14,25,40].The EE-induced strengthening of neuronal connectiv-ity provides the means through which the brain mayefficiently utilize already existing neuronal networksand recruit alternative circuits, when required [47]. Asfor behavioral results, cognitive performances are im-proved in normal animals and recovered in brain injuredsubjects [9,23]. Accordingly, the EE-induced plasticchanges might be of special concern to the theory ofbrain and cognitive reserve, providinga basis to explainthe maintenance of normal cognitive function despitethe presence of brain pathology.
The present study was devoted to experimentally ver-ify the brain reserve hypothesis by analyzing the in-fluence of rearing conditions on the cognitive perfor-mances and on neuronal morphology following lesionsof cholinergic basal forebrain system. As paradigmof EE, we used the same protocol demonstrated to en-hance procedural and working memory abilities in spa-tial tasks, as well as to increase dendritic arborisationand spine density in parietal pyramidal neurons [37].
Because the cholinergic system plays a central rolein learning and memory abilities, as a model of brainpathology, we used the immunotoxic lesions of basalforebrain nuclei eliciting cholinergic depletion of neo-cortical areas. Clinical studies indicate that choliner-gic projection from the basal forebrain to cortical areasis the pathway earliest and most severely affected inAlzheimer patients [6]. Attempts to model Alzheimer-like deficits in animals have focused on producingcholinergic hypofunction by manipulating basal fore-brain nuclei [3,5,16,18,38,52,69]. Bilateral injectionsof 192 IgG-saporin into the basal forebrain provide thecholinergicdepletion of the neocortical areas represent-ing an attractive experimental model for analyzing cog-nitive functions and neuronal changes in the absenceof cholinergic projection to the neocortex. This modelof cholinergic depletion of the neocortex allowed us toanalyze whether the cognitive impairment elicited bythe lesion could be beneficially affected by sustainedEE-rearing. This finding would support the occurrence
of a cognitive reserve. In particular, we analyzed theperformances obtained in spatial tasks that are of fun-damental importance to mobile organisms whose spa-tial memories are organized in distinct but coordinatedframes, thus allowing well structured adaptive capaci-ties. Given the heavy involvement of associative pari-etal areas in many spatial aspects, to correlate behav-ioral outcomes and structural features of neocorticalneurons, the synaptic connectivity of the parietal pyra-midal neurons was analyzed. In particular, we analyzedthe layer-III pyramidal cells that integrate both horizon-tal and top-down information [59] and are particularlyresponsive to modified stimulation patterns. Dendriticspines play an important role in neuronal informationprocessing and since their number and density are mod-ified by the activity levels [10,28,33–35,37,54,65,67],they provide the morphological substrate for synapticplasticity either lesion-induced and activity-dependent.On such a basis, differences among groups in behav-ioral outcomes and in neuronal correlates would indi-cate that EE has the potential to provide cognitive andstructural reserves to be spent in the case of brain lesion.The analysis of both types of reserves in the very samesubjects is an attractive approach addressable only inexperimental studies, since the closer a neuronal level(synaptic density, degree of arborization) is focused,the harder is evaluate its features in living humans.
MATERIAL AND METHODS
Animals
Wistar male rats were maintained following theguidelines for ethical conduct developed by the Eu-ropean Communities Council Directive of 24 Novem-ber 1986 (86/609/EEC). The experiments were carriedout on two groups of animals, one reared in an en-riched environment and one reared in standard condi-tions from the postnatal day 21 onward. Followingthree months, both groups were further subdivided intotwo subgroups, one comprising rats that received bi-lateral infusions of 192 IgG-saporin into the region ofnucleus basalis magnocellularis(NBM) and one com-prising rats used as controls. Thus, we obtained an en-riched lesioned group (EL group,n = 13), a standardreared lesioned group (SL group,n = 13), an enrichedcontrol group (EC group,n = 13) and a standard rearedcontrol group (SC group,n = 13).

L. Mandolesi et al. / Enrichment and Cholinergic Depletion 13
Rearing conditions
The enriched rats were housed in groups of 10 in alarge cage (100× 50 × 70 cm), with one extra lev-el constructed of galvanized wire mesh and connectedby ramps of the same material to create two intercon-nected levels. The cage contained wood shavings, arunning wheel, a shelter (a house-shaped toy with aconcave opening in which the rat may enter), plasticcolored toys, and small constructions. Throughout theenrichment period, the shelter and running wheel werekept in the cage, while the toys and constructions werechanged twice a week. Once a week, the feeding boxesand water bottles were moved to different cage pointsto encourage foraging and explorative behaviors. Inthis way, rats were cognitively stimulated by the ob-jects that encouraged exploration or manipulation andby enhanced social interactions. They were also phys-ically trained through exposure to objects that encour-aged aerobic exercise as the running wheel. Further-more, each enriched animal was handled daily for atleast 10 min and allowed to forage in a large space,i.e., the whole lab room for 15 min. The rats reared instandard conditions were pair housed in a standard cage(40× 26 × 18 cm) containing wood shavings but noobjects. Feeding boxes and water bottles were kept inthe same position throughout the animals’ lives. Theseanimals received the usual care by the stabulary staff,even if without particular and prolonged manipulation.This procedure did not result in an impoverished rear-ing and also standard animals were accustomed to the“human” contact. Both groups of animals received thesame type of food. Food and water were providedadlibitum until behavioral testing.
Surgery
At approximately 110 days of age, all rats were anes-thetized with ketamine (90 mg/kg i.p.) and xylazine(15 mg/kg i.p.). In the animals to be lesioned, theimmunotoxin 192 IgG-saporin (Chemicon Internation-al Inc., Harrow, UK) was bilaterally injected througha glass micropipette of 50µL connected to a 10-µLHamilton syringe at the following coordinates [51]:AP: −1.0 (from the bregma); ML:± 2.3 (from themidline); DV: −8 (from the dura). 0.4µg of the im-munotoxin 192 IgG-saporin diluted in 0.4µL of PBSwas injected at a rate of 0.1µL/min for 4 min. Sucha dosage was demonstrated to be the appropriate oneto elicit selective lesions of the cholinergic basal fore-brain [3,38]. At the end of administration, the needle
was left in situ for 5 min. In the remaining rats, onlyPBS solution containing no saporin was injected. Theanimals were put in a standard cage for 24 hours torecover from anesthesia and surgical stress. After thisperiod, they were returned to their respective housingconditions. No significant difference was found in thebody weight of the animals in the four experimentalgroups (mean values recorded the last testing day: SC:440 g± 27; SL: 440 g± 55; EC: 467 g± 68; EL:458 g± 42).
Behavioral testing
From one month after surgery onwards, learning andmemory capabilities were evaluated in three tests ad-ministered in the following order: Morris Water Maze(MWM), Radial Arm Maze (RAM), and Open Field(OF).
Morris water mazeThe rats were placed in a circular white pool (di-
ameter 140 cm), filled with water (24◦C) 60 cm deep,made opaque by the addition of 2 L of milk. An escapeplatform (diameter 10 cm) placed 2 cm below or abovethe water level was sited in the middle of one cardinalquadrant, 30 cm from the side walls. The rat was al-lowed to swim around to find the platform. Startingpoints at the pool wall were pseudo-randomly varied.Each rat was presented with two sessions of four trialsper day with an inter-session interval of 5 hours. In thefirst four sessions (Place I), the platform was hidden inthe northwest pool quadrant; in the next two sessions(Cue phase), the platform was kept visible in the north-east quadrant; in the final four sessions (Place II), theplatform was hidden in the northeast quadrant.
The rats’ trajectories in the pool were monitored bya video camera mounted on the ceiling. The resultingvideo signal was relayed to a monitor and to an imageanalyzer (Ethovision, Noldus).
The MWM parameters taken into account were: la-tencies; heading angles; path length; percentage ofpath length spent in 20-cm wide outer annulus; swim-ming velocity. Following the procedure described else-where [21], exploration behavior was classified in fourmain categories: circling; extended searching; restrict-ed searching; direct finding.
Radial arm mazeIt consisted of a round central platform (diameter
30 cm) from which eight arms (12.5× 60 cm) radiatedlike the spokes of a wheel. A food well (5 cm) waslocated at the end of each arm. Prior to the testingphase, rats were food-restricted to decrease their weightby 20%.

14 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
Full-baited maze procedureAll maze arms were baited with a small piece of
Purina chow at each session. The animal made an errorwhen it completely entered an already visited arm inthe same session. The animals underwent two sessionsa day for 5 consecutive days. The inter-session intervalwas 5 hours.
The parameters considered were: total time to com-plete the task; total errors, number of re-visited arms;spatial span (the longest sequence of correctly visitedarms); percentage of 45◦, 90◦, 135◦, 180◦ or 360◦ an-gles the rats made in each session divided by the num-ber of angles made; perseverations (the consecutive re-entries in the same arm through a 360◦ angle or in afixed sequence of arms).
Forced-choice procedureForty-eight hours after the end of the preceding pro-
tocol, all animals were submitted to the forced-choiceparadigm. In the training phase, four arms (for exam-ple, arms 1,3,4,7) were baited and the remaining armswere closed by a little door at their proximal end. Therat was allowed to explore all the open arms. Then,it spent 60 sec in its cage. In the testing phase, freeaccess to all eight arms was allowed but only the fourpreviously closed arms were baited. This task was re-peated for 5 consecutive days with a different configu-ration of arms closed each day. The working memoryerrors (entries into un-baited arms) were considered.This parameter was further broken down into two er-ror subtypes: across-phase errors (entries into an armentered during the training phase) and within-phase er-rors (re-entries into an arm visited earlier in the testingphase). The perseverations (consecutive re-entries inthe same arm or in a fixed sequence of arms) were alsoconsidered [41].
Open fieldThe apparatus consisted of a circular box (diameter
140 cm) delimited by a wall 30 cm high. During ses-sion 1 (S1) each rat was allowed to move in the emptyopen field, and its baseline level of activity was mea-sured. During S2-S4 (habituation phase), five differentobjects were simultaneously placed in the arena. Fourobjects were placed in a square arrangement in the mid-dle annulus of the arena and the fifth one was placedin the central area. Before S5 (spatial change), thespatial configuration was changed by moving two ob-jects so that the initial square arrangement was changedto a polygon-shaped configuration, without any central
object. Session duration was 6 min, and intersessionintervals were 3 min.
The behavior was assessed by using: Emotional pa-rameters: number of defecation boluses; Motor param-eters: total distance (in m) travelled in the arena; per-centage of the total distance engaged in exploring pe-ripheral sectors; Spatial parameters: total time spent incontacting objects.
Analysis of dendritic spines
We used thein vivoGolgi-like filling of the neuronsdescribed by Jiang et al. [27] that allows high efficiencyof labeling and a complete visualization of dendriticarborizations. At the end of behavioral testing, twentyrandomly selected animals belonging to the four ex-perimental groups were chosen for the dendritic analy-ses. Anesthetized (ketamine: 90 mg/kg i.p.; xylazine:15 mg/kg i.p.) animals received parietal cortical injec-tions according to the stereotaxic co-ordinates derivedfrom the atlas of Paxinos and Watson [51] (ML−5,AP −1.5, DV−1) of 10% biotinylated dextran amineBDA (Molecular Probes, Eugene, OR) and 20mM NM-DA (Sigma, St Louis, MO) in 0.01 M phosphate bufferPB (total injected volumes ranging from 0.1 to 0.2µl).After a survival time of 96 hours, the animals wereanesthetized and transcardially perfused with saline fol-lowed by 4% paraformaldehyde and 0.1% glutaralde-hyde in phosphate buffer saline PBS (4◦C, pH 7.5).After removal, the brains were cryoprotected in 30%buffered sucrose and cut on a freezing microtome in-to coronal sections of 50 or 40µm according to thestaining method. Three 1:4 series of adjacent sectionswere collected for BDA labeling (to visualize neuronalmorphology), Nissl staining (to visualize the cytoar-chitectonic features of the cortex) and for ChAT im-munohistochemistry (to visualize the cholinergic activ-ity). This later technique is described in the “neuro-chemical analysis” section. The BDA labeled neuronswere evidenced using the avidin-biotin complex. Theinjection area centered in primary somato-sensory cor-tex showed evidence of necrosis, probably due to theexcitotoxic action of NMDA. Most labeled cells werefound in the cortex surrounding the experimental le-sion where no signs of excitotoxicity were noted. Thelabeled parietal neurons chosen for the morphologicalanalyses were located in layer III of parietal cortexfrom AP−2.8 to−3.8, areas corresponding to the sec-ondary somato-sensory and posterior parietal cortices.Pyramidal neurons were defined by the presence of abasilar dendritic tree, a distinct single apical dendrite

L. Mandolesi et al. / Enrichment and Cholinergic Depletion 15
and dendritic spines. A researcher unaware of spec-imen identity made the morphological analysis withthe aid of the Neurolucida software (Microbrightfield,Colchester, VT). This system allows computer-assistedreconstruction of the dendritic arbors. Neurons wereselected when their labeling was uniform and extend-ing into the most distal branches of apical and basaldendrites, where spines were clearly marked. All pro-trusions of the dendritic membrane were considered,regardless of their shape or actual function. Further-more, the predominant plan of the dendritic arbors ofthese neurons should be parallel to the plan of the sec-tion. According to these criteria, in layer III of parietalcortex, 10 neurons were selected from the animals be-longing to each of the four experimental group, so 40neurons were totally collected. Because our previousresearch demonstrated that the rearing in an enrichedenvironment deeply modified number and density of thedendritic spines in both apical and basal dendrites [37],in the present study the morphological analysis of theneurons was carried out by analyzing the apical as wellas basal dendrites and by analyzing the whole visual-izable extension of dendritic arborisation, without re-sorting to any kind of sampling. This time-consumingprocedure provided a very accurate investigation of theeffects of EE and cholinergic depletion on both types ofdendrites. A Sholl analysis estimated the amount anddistribution of dendritic material by overlaying a seriesof concentric rings at 10µm intervals centered on thesoma to each pyramidal neuron.
The following spine parameters were analyzed inthe apical and basal trees: total number of dendriticspines, calculated by summing all the spines from theinitial protrusion of apical or basal dendrites (the initial10 µm from the soma were not considered becauseusually no spines were present); mean spine density,calculated as the total number of spines divided bythe number of concentric spheres at 10µm intervals;spine distribution, calculated as spine density (numberof spines per 10-µm shell) along the entire apical andbasal dendrites.
Neurochemical analysis
When the behavioral testing was finished, all theanimals not selected for the neuronal analyses weredeeply anaesthetized and perfused with saline followedby 4% paraformaldehyde and 0.1% glutaraldehyde inPBS (4◦C, pH 7.5). Brains were post-fixed for about 4 hand transferred into a 0.1 M phosphate buffered 25%sucrose solution in which they remained for 36–48 h.
The brains were then frozen and cut into 40µm coro-nal sections from the anterior regions of frontal lobe tothe posterior hippocampus. Two 1:4 series of adjacentsections were collected for Nissl staining and for ChATactivity. Sections immunostained for ChAT were pre-incubated in PBS at 4◦C, then in 0.4% Triton X-100 inPBS, finally in 0.1 Triton X-100 plus 1% bovine serumalbumin (Sigma Chemical Company) plus Normal GoatSerum (Vector Laboratories Inc. Burlingame, CA) inPBS. Sections were incubated for 16 h at 4◦C with0.1%Triton X-100 and Normal Goat Serum in PBS withthe primary antibody for ChAT (1:1000). Subsequent-ly, sections were incubated with biotinylated secondaryantibody (Goat Anti-Rabbit IgG-biotin conjugate) and3% Normal Goat Serum (Kit Elite PK 6101, VectorLaboratories Inc.) in PBS for 10 min at room tempera-ture. Staining was visualized with 0.05% diaminoben-zidine and Ammonium Nickel (II) Sulphate (Sigma-Aldrich Chemie, Steinheim, Germany) after incubationwith avidin and biotinylated peroxidase (Kit Elite PK6101, Vector Laboratories Inc.). To exclude artifacts,in each case some random sections were processed aspreviously described, with the only difference of theabsence of the primary antibody.
Statistical analysis
Results of animals of the four experimental groupswere compared by using one-way, two-way (with rear-ing and lesion as between-subjects factors), or three-way (with rearing and lesion as between-subjects fac-tors and sessions as within-subject factor) analysesof variance (ANOVAs), followed by multiple com-parisons using Duncan’s tests. For morphologicalanalyses, the opportunity of applying nested-designANOVAs taking into account the correlation of datawithin the same animal, has been estimated by prelimi-narily evaluating the significance of the interaction be-tween random factor “animal” and fixed factor “group”in two-way ANOVAs (animal x group). No significantinteraction was found in any spine parameter, then nonested-design ANOVA was used.
RESULTS
Histological verification of 192-IgG saporin lesion
The lesion was assessed by counting ChAT-IR neu-rons in the Ch4 region that includes thenucleus basalismagnocellularis(NBM), the substantia innominata,

16 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
Fig. 1. Representative photomicrographs of the Ch4 region in coronal sections located at about−1.40 mm from bregma. ChAT-immunoreactiveneurons in control rats reared in standard (SC) or enriched (EC) conditions are shown on the left side of the figure. On the right side, the sameregion is shown in rats reared in standard (SL) or enriched (EL) conditions subjected to injections of 192 IgG-saporin in Ch4 region. Note thesubstantial absence of ChAT-immunoreactive neurons in lesioned animals. Scale bar= 0.5 mm.
the medial part of the horizontal limb and the lateralpart of the vertical limb of the diagonal band [42]. Inun-lesioned groups in the Ch4 region, the mean numberof cholinergic neurons was 1904± 149.6. The meannumber of cholinergic neurons in the Ch4 region oflesioned animals was 25.25± 17.95.
In the lesioned cases, ChAT-immunoreactive cellsin adjacent nuclei of the basal forebrain were not de-creased in number, apart from the few cholinergic in-terneurons belonging to striatum. No evidence of grossunspecific neuronal damage, such as cavitation, and noor very weak gliosis at the injection sites were found.Representative photomicrographs of the Ch4 region inthe four experimental groups are shown in Fig. 1.
Neurological examination
At the time of testing (one month after the surgery),lesioned rats (EL and SL groups) appeared to have re-covered postural reflexes and locomotor activity to thelevels of both groups of un-lesioned animals (EC andSC groups). When standing, lesioned animals exhibit-ed symmetrical posture with limbs balanced under thetrunk. When stepping, neither ataxic symptoms norstaggering nor dragging a hind-paw were noted. Theoperated animals were able to walk and run withoutany directional bias. When grooming, they were ableto rear on their hind-limbs and maintain an upright pos-
ture. When swimming, all animals exhibited good co-ordination and kept their noses out of the water. Inparticular, they displayed the characteristic swimmingwith forelimb inhibition and alternate hind-limb thrust-ing. Their ability to turn in water was also preserved.
Morris water maze
The MWM allows analyzing the competence onbuilding the spatial cognitive map as well as the explo-rative procedures put into action in searching the plat-form, besides obviously the swimming abilities. Allexperimental groups learned to explore the water mazeand to locate the platform, as demonstrated by progres-sively decreasing escape latencies as the sessions wentby (F9,432 = 71.71;p = 0.00001) (Fig. 2A). No signif-icant effect of lesion (F1,48 = 3.72;p = 0.06) and rear-ing (F1,48 = 3.04;p = 0.087) was found on latencies.The highest mean swimming velocities were displayedby the enriched groups (EC: 27.9; EL: 27.8 cm/sec)and the lowest velocities by the standard groups (SL:20.2; SC: 22.5 cm/sec). A two-way ANOVA (rearingx lesion) revealed a significant rearing effect (F1,48 =105.6;p = 0.00001), while lesion effect (F1,48 = 3.85;p = 0.055) and interaction (F12,48 = 2.84;p = 0.09)were not significant. Heading angles, index linked tothe knowledge of the platform localization, were signif-icantly affected by the presence of the lesion (F1,48 =
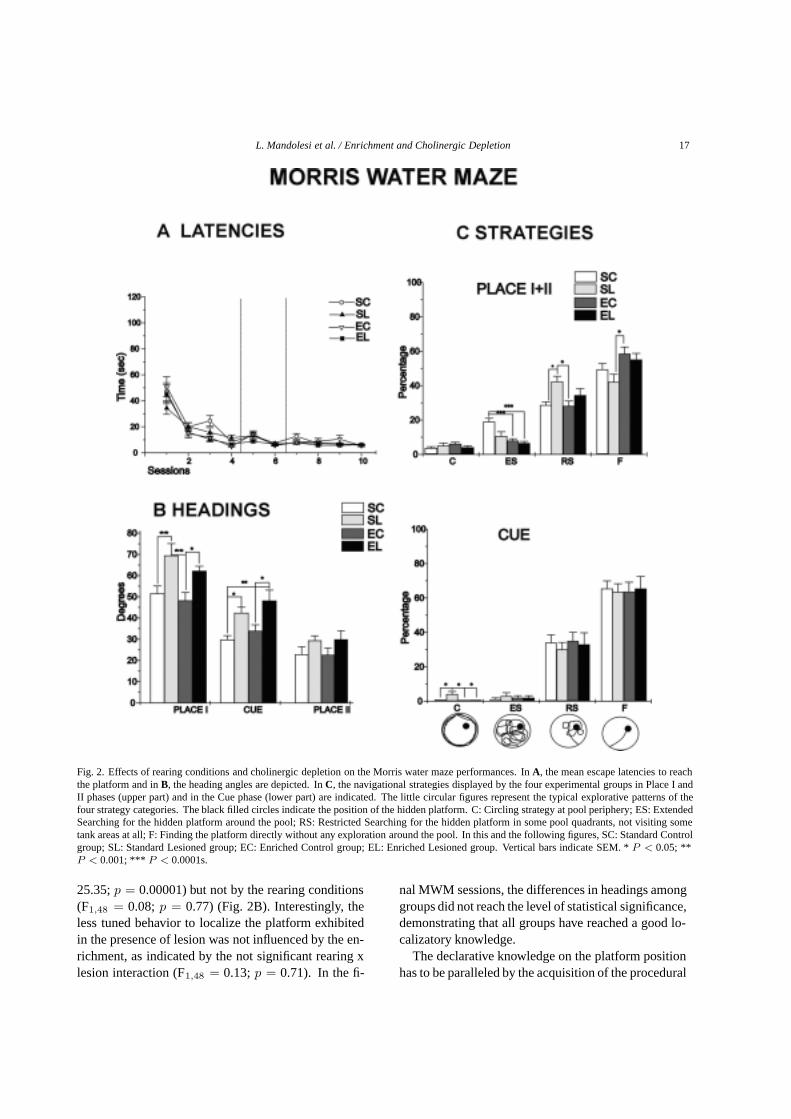
L. Mandolesi et al. / Enrichment and Cholinergic Depletion 17
Fig. 2. Effects of rearing conditions and cholinergic depletion on the Morris water maze performances. InA, the mean escape latencies to reachthe platform and inB, the heading angles are depicted. InC, the navigational strategies displayed by the four experimental groups in Place I andII phases (upper part) and in the Cue phase (lower part) are indicated. The little circular figures represent the typical explorative patterns of thefour strategy categories. The black filled circles indicate the position of the hidden platform. C: Circling strategy at pool periphery; ES: ExtendedSearching for the hidden platform around the pool; RS: Restricted Searching for the hidden platform in some pool quadrants, not visiting sometank areas at all; F: Finding the platform directly without any exploration around the pool. In this and the following figures, SC: Standard Controlgroup; SL: Standard Lesioned group; EC: Enriched Control group; EL: Enriched Lesioned group. Vertical bars indicate SEM. *P < 0.05; **P < 0.001; *** P < 0.0001s.
25.35;p = 0.00001) but not by the rearing conditions(F1,48 = 0.08; p = 0.77) (Fig. 2B). Interestingly, theless tuned behavior to localize the platform exhibitedin the presence of lesion was not influenced by the en-richment, as indicated by the not significant rearing xlesion interaction (F1,48 = 0.13;p = 0.71). In the fi-
nal MWM sessions, the differences in headings amonggroups did not reach the level of statistical significance,demonstrating that all groups have reached a good lo-calizatory knowledge.
The declarative knowledge on the platform positionhas to be paralleled by the acquisition of the procedural
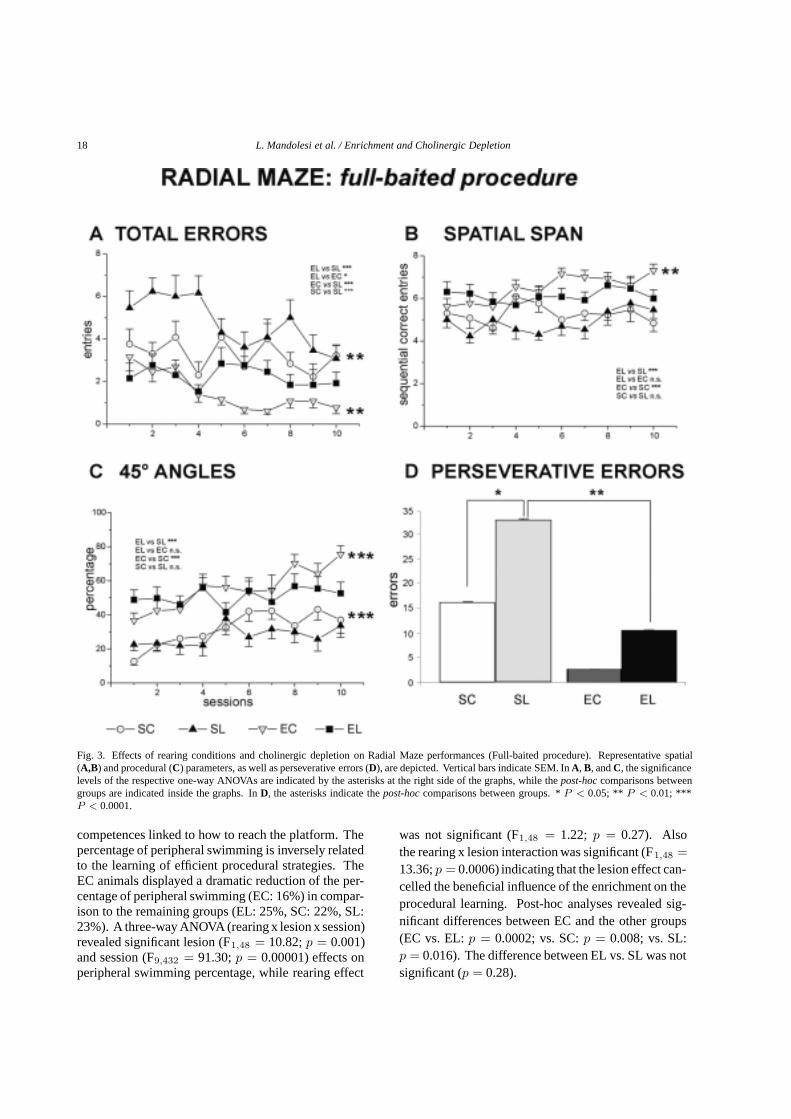
18 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
Fig. 3. Effects of rearing conditions and cholinergic depletion on Radial Maze performances (Full-baited procedure). Representative spatial(A,B) and procedural (C) parameters, as well as perseverative errors (D), are depicted. Vertical bars indicate SEM. InA, B, andC, the significancelevels of the respective one-way ANOVAs are indicated by the asterisks at the right side of the graphs, while thepost-hoccomparisons betweengroups are indicated inside the graphs. InD, the asterisks indicate thepost-hoccomparisons between groups. *P < 0.05; ** P < 0.01; ***P < 0.0001.
competences linked to how to reach the platform. Thepercentage of peripheral swimming is inversely relatedto the learning of efficient procedural strategies. TheEC animals displayed a dramatic reduction of the per-centage of peripheral swimming (EC: 16%) in compar-ison to the remaining groups (EL: 25%, SC: 22%, SL:23%). A three-way ANOVA (rearing x lesion x session)revealed significant lesion (F1,48 = 10.82;p = 0.001)and session (F9,432 = 91.30;p = 0.00001) effects onperipheral swimming percentage, while rearing effect
was not significant (F1,48 = 1.22; p = 0.27). Alsothe rearing x lesion interaction was significant (F1,48 =13.36;p = 0.0006) indicating that the lesion effect can-celled the beneficial influence of the enrichment on theprocedural learning. Post-hoc analyses revealed sig-nificant differences between EC and the other groups(EC vs. EL:p = 0.0002; vs. SC:p = 0.008; vs. SL:p = 0.016). The difference between EL vs. SL was notsignificant (p = 0.28).
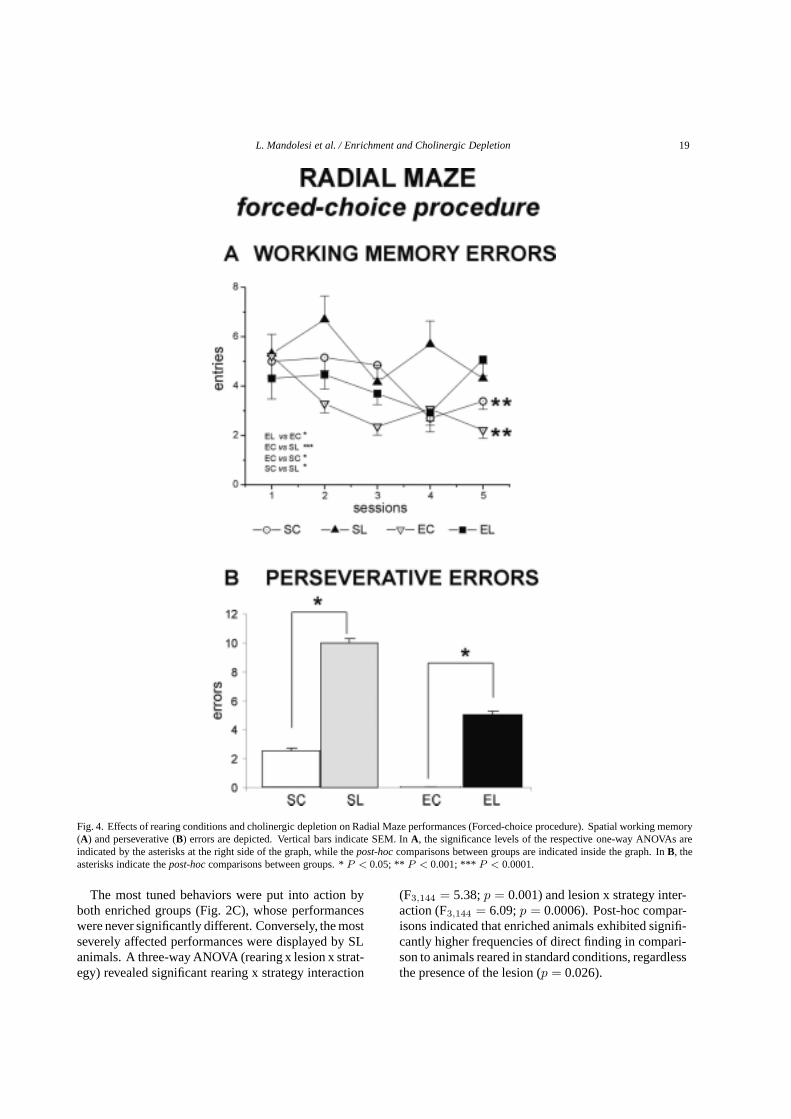
L. Mandolesi et al. / Enrichment and Cholinergic Depletion 19
Fig. 4. Effects of rearing conditions and cholinergic depletion on Radial Maze performances (Forced-choice procedure). Spatial working memory(A) and perseverative (B) errors are depicted. Vertical bars indicate SEM. InA, the significance levels of the respective one-way ANOVAs areindicated by the asterisks at the right side of the graph, while thepost-hoccomparisons between groups are indicated inside the graph. InB, theasterisks indicate thepost-hoccomparisons between groups. *P < 0.05; ** P < 0.001; *** P < 0.0001.
The most tuned behaviors were put into action byboth enriched groups (Fig. 2C), whose performanceswere never significantly different. Conversely, the mostseverely affected performances were displayed by SLanimals. A three-way ANOVA (rearing x lesion x strat-egy) revealed significant rearing x strategy interaction
(F3,144 = 5.38;p = 0.001) and lesion x strategy inter-action (F3,144 = 6.09;p = 0.0006). Post-hoc compar-isons indicated that enriched animals exhibited signifi-cantly higher frequencies of direct finding in compari-son to animals reared in standard conditions, regardlessthe presence of the lesion (p = 0.026).

20 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
RADIAL MAZE
Full-baited procedure
No difference in the time spent to visit the eight armswas found among groups that displayed a significantreduction in time as the sessions went by (F9,432 =6.27; p = 0.00001). Rearing (F1,48 = 0.6; p = 0.4)and lesion (F1,48 = 0.0004;p = 0.98) effects were notsignificant, indicating that even lesioned animals werenot affected by severe motor symptoms. Conversely,groups significantly differed on the parameters mostlinked to the computation of spatial factors. Regardlessof the lesion, the enriched groups performed better thanboth standard reared groups (Fig. 3). In particular, bothenriched groups made a significantly lower number oftotal errors (Fig. 3A). This parameter was significantlyaffected by the rearing conditions (F1,48 = 79.1;p =0.00001), lesion (F1,48 = 21.75; p = 0.00001) andsession (F9,432 = 3.89;p = 0.00001). The significantsecond order interaction (F9,432 = 2.89; p = 0.002)indicated the positive EE-effect even in the presence ofthe lesion. The beneficial influence of enriched rearing(F1,48 = 70.1; p = 0.00001) was evident when thecorrect visits span was analyzed. In fact, the span ofthe enriched groups was longer (6–7 correct visits) thanthat of groups reared in standard conditions (4–6 correctvisits). However, a negative effect of the cholinergiclesion (F1,48 = 6.38; p = 0.015) was also evident,given that in both rearing conditions the cholinergicdepletion reduced the span length (Fig. 3B). The secondorder interaction was not significant (F9,432 = 1.71;p = 0.08).
The 45◦ angles distribution revealed that the best pro-cedural performances were displayed by the enrichedanimals. In fact, it was significantly affected by therearing conditions (F1,48 = 38.3; p = 0.00001), butunaffected by the presence of the cholinergic lesion(F1,48 = 1.38;p = 0.24). The second order interactionwas not significant (F9,432 = 1.53;p = 0.13). Note thatonly the un-lesioned animals were able to increase theirnumber of 45◦ angles as the sessions went by (Fig. 3C),indicating that the cholinergic depletion prevented theimprovement of the procedural competences.
The performances of both lesioned groups were char-acterized by a marked presence of perseverative er-rors. This behavior, mainly present in SC animals, wassignificantly increased by the presence of the lesion(F1,48 = 5.1; p = 0.02), even if significantly reducedby the environmental enrichment (F1,48 = 9.86; p =0.003) (Fig. 3D). The second order interaction was notsignificant (F9,432 = 0.95;p = 0.47).
Forced-choice procedure
To differentiate procedural from working memo-ry factors, the forced-choice protocol has been used.SC and EC groups progressively decreased workingmemory errors, while neither lesioned group displayedany improvement as the sessions went by (Fig. 4A).Working memory errors were significantly decreasedby the environmental enrichment (F1,48= 11.01;p = 0.0017), and increased by the cholinergic deple-tion regardless of rearing conditions (F1,48= 8.64;p = 0.005). The significant second order interaction(F4,192= 3.40;p = 0.01) indicated the beneficial ef-fect of enriched rearing, in spite of the presence of thelesion, as the sessions went by.
A further analysis on the working memory errors re-vealed that enriched animals made fewer across-phaseerrors than standard reared animals (F1,48 = 7.1; p =0.01) and that lesioned animals made a significantlyincreased number of across-phase errors (F1,48 = 8.85;p = 0.004). The second order interaction was signif-icant (F4,192 = 4.12;p = 0.003). Rearing conditionssignificantly affected also within-phase errors (F1,48 =7.71; p = 0.007), while lesion (F1,48 = 2.93; p =0.09) effect and the second order interaction were notsignificant (F4,192 = 1.13;p = 0.33).
Interestingly, the number of perseverative errors wassignificantly influenced by the lesion (F1,48 = 8.64;p = 0.005) and by the rearing conditions (F1,48 =11.01;p = 0.001). Also the interaction was signifi-cant (F4,192 = 3.4; p = 0.01), demonstrating that theperseverative tendencies elicited by cholinergic deple-tion were significantly influenced by the environmentalenrichment (Fig. 4B).
Open field
A three-way ANOVA (rearing x lesion x session)indicated the absence of significant differences on thenumber of defecation boluses demonstrating a compa-rable level of anxiety in all animals. Only the sessioneffect was significant (F3,192 = 60.04;p = 0.00001),given the already reduced number of boluses collapsedas the task went by. As regards the distance travelledduring S1, the SL group was the most active one, fol-lowed by the SC animals. The two enriched groupsexhibited significantly lower values (F1,48 = 30.47;p = 0.000001) (Fig. 5A). The lesion effect was alsosignificant (F1,48 = 9.26; p = 0.004). The interac-tion was not significant (F1,48 = 0.09;p = 0.75). In-terestingly, the percentage of the path in the periph-
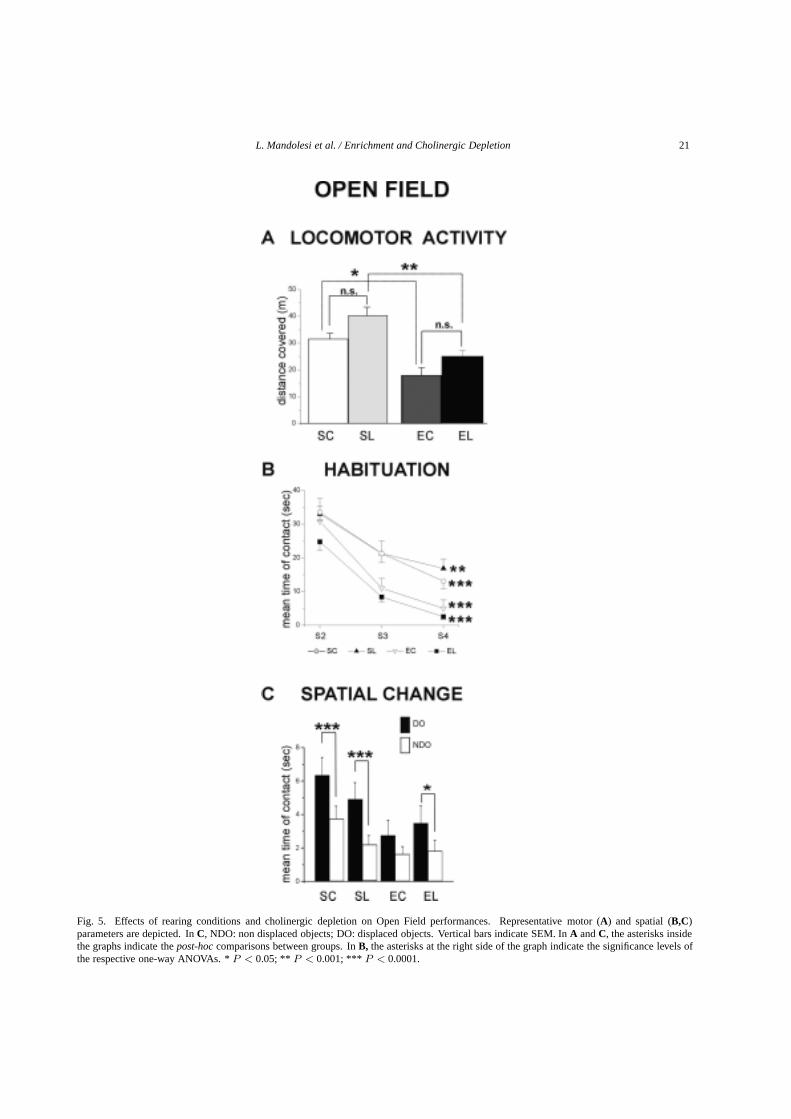
L. Mandolesi et al. / Enrichment and Cholinergic Depletion 21
Fig. 5. Effects of rearing conditions and cholinergic depletion on Open Field performances. Representative motor (A) and spatial (B,C)parameters are depicted. InC, NDO: non displaced objects; DO: displaced objects. Vertical bars indicate SEM. InA andC, the asterisks insidethe graphs indicate thepost-hoccomparisons between groups. InB, the asterisks at the right side of the graph indicate the significance levels ofthe respective one-way ANOVAs. *P < 0.05; ** P < 0.001; *** P < 0.0001.
22 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
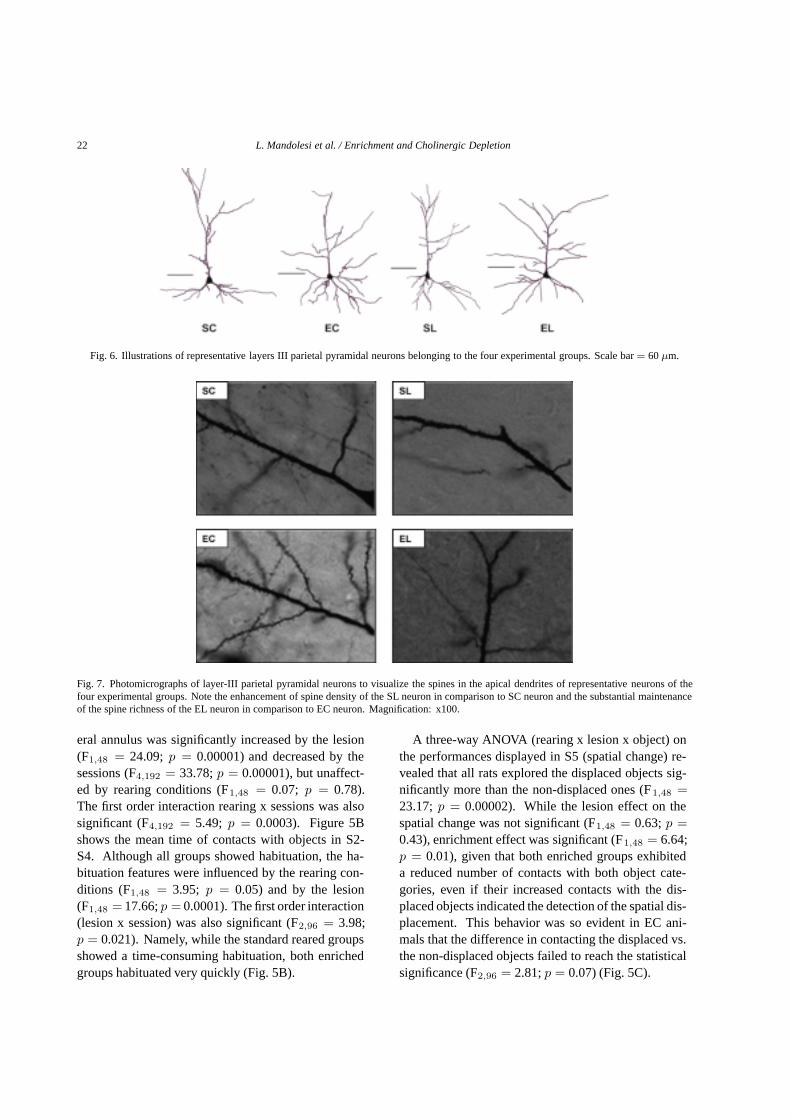
Fig. 6. Illustrations of representative layers III parietal pyramidal neurons belonging to the four experimental groups. Scale bar= 60µm.
Fig. 7. Photomicrographs of layer-III parietal pyramidal neurons to visualize the spines in the apical dendrites of representative neurons of thefour experimental groups. Note the enhancement of spine density of the SL neuron in comparison to SC neuron and the substantial maintenanceof the spine richness of the EL neuron in comparison to EC neuron. Magnification: x100.
eral annulus was significantly increased by the lesion(F1,48 = 24.09; p = 0.00001) and decreased by thesessions (F4,192 = 33.78;p = 0.00001), but unaffect-ed by rearing conditions (F1,48 = 0.07; p = 0.78).The first order interaction rearing x sessions was alsosignificant (F4,192 = 5.49; p = 0.0003). Figure 5Bshows the mean time of contacts with objects in S2-S4. Although all groups showed habituation, the ha-bituation features were influenced by the rearing con-ditions (F1,48 = 3.95; p = 0.05) and by the lesion(F1,48 = 17.66;p = 0.0001). The first order interaction(lesion x session) was also significant (F2,96 = 3.98;p = 0.021). Namely, while the standard reared groupsshowed a time-consuming habituation, both enrichedgroups habituated very quickly (Fig. 5B).
A three-way ANOVA (rearing x lesion x object) onthe performances displayed in S5 (spatial change) re-vealed that all rats explored the displaced objects sig-nificantly more than the non-displaced ones (F1,48 =23.17;p = 0.00002). While the lesion effect on thespatial change was not significant (F1,48 = 0.63;p =0.43), enrichment effect was significant (F1,48 = 6.64;p = 0.01), given that both enriched groups exhibiteda reduced number of contacts with both object cate-gories, even if their increased contacts with the dis-placed objects indicated the detection of the spatial dis-placement. This behavior was so evident in EC ani-mals that the difference in contacting the displaced vs.the non-displaced objects failed to reach the statisticalsignificance (F2,96 = 2.81;p = 0.07) (Fig. 5C).
L. Mandolesi et al. / Enrichment and Cholinergic Depletion 23
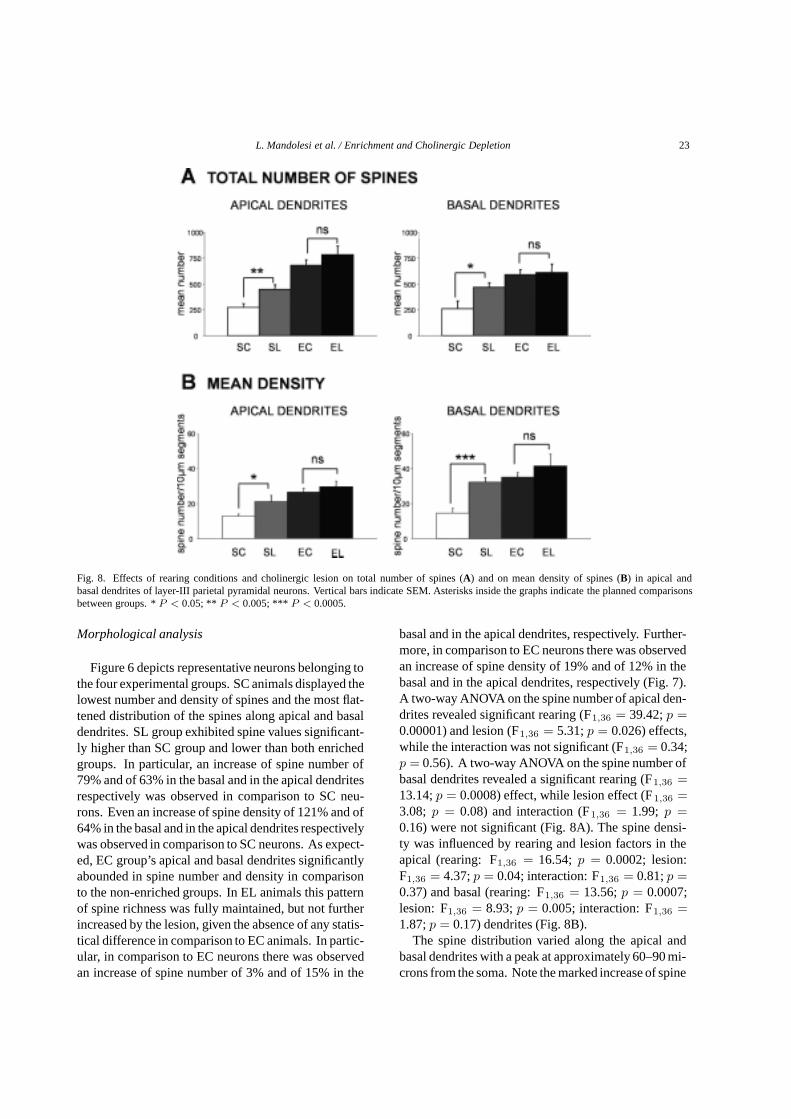
Fig. 8. Effects of rearing conditions and cholinergic lesion on total number of spines (A) and on mean density of spines (B) in apical andbasal dendrites of layer-III parietal pyramidal neurons. Vertical bars indicate SEM. Asterisks inside the graphs indicate the planned comparisonsbetween groups. *P < 0.05; ** P < 0.005; *** P < 0.0005.
Morphological analysis
Figure 6 depicts representative neurons belonging tothe four experimental groups. SC animals displayed thelowest number and density of spines and the most flat-tened distribution of the spines along apical and basaldendrites. SL group exhibited spine values significant-ly higher than SC group and lower than both enrichedgroups. In particular, an increase of spine number of79% and of 63% in the basal and in the apical dendritesrespectively was observed in comparison to SC neu-rons. Even an increase of spine density of 121% and of64% in the basal and in the apical dendrites respectivelywas observed in comparison to SC neurons. As expect-ed, EC group’s apical and basal dendrites significantlyabounded in spine number and density in comparisonto the non-enriched groups. In EL animals this patternof spine richness was fully maintained, but not furtherincreased by the lesion, given the absence of any statis-tical difference in comparison to EC animals. In partic-ular, in comparison to EC neurons there was observedan increase of spine number of 3% and of 15% in the
basal and in the apical dendrites, respectively. Further-more, in comparison to EC neurons there was observedan increase of spine density of 19% and of 12% in thebasal and in the apical dendrites, respectively (Fig. 7).A two-way ANOVA on the spine number of apical den-drites revealed significant rearing (F1,36 = 39.42;p =0.00001) and lesion (F1,36 = 5.31;p = 0.026) effects,while the interaction was not significant (F1,36 = 0.34;p = 0.56). A two-way ANOVA on the spine number ofbasal dendrites revealed a significant rearing (F1,36 =13.14;p = 0.0008) effect, while lesion effect (F1,36 =3.08; p = 0.08) and interaction (F1,36 = 1.99; p =0.16) were not significant (Fig. 8A). The spine densi-ty was influenced by rearing and lesion factors in theapical (rearing: F1,36 = 16.54; p = 0.0002; lesion:F1,36 = 4.37;p = 0.04; interaction: F1,36 = 0.81;p =0.37) and basal (rearing: F1,36 = 13.56;p = 0.0007;lesion: F1,36 = 8.93;p = 0.005; interaction: F1,36 =1.87;p = 0.17) dendrites (Fig. 8B).
The spine distribution varied along the apical andbasal dendrites with a peak at approximately 60–90 mi-crons from the soma. Note the marked increase of spine
24 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
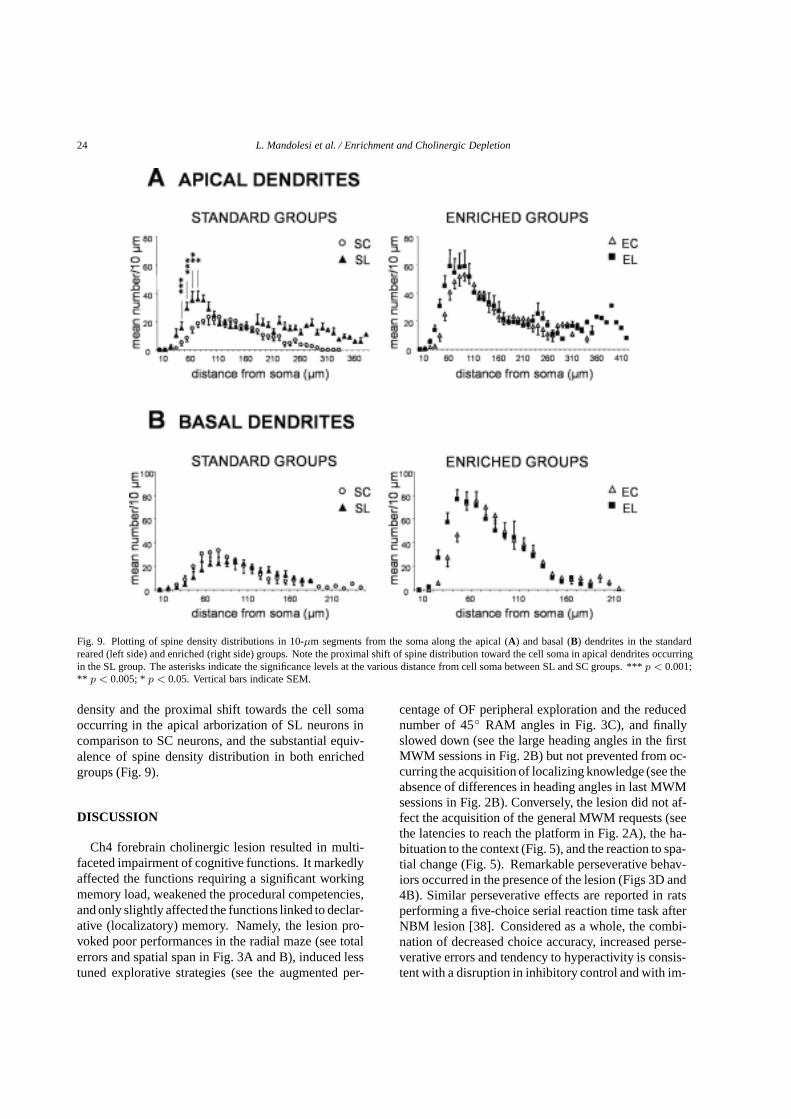
Fig. 9. Plotting of spine density distributions in 10-µm segments from the soma along the apical (A) and basal (B) dendrites in the standardreared (left side) and enriched (right side) groups. Note the proximal shift of spine distribution toward the cell soma in apical dendrites occurringin the SL group. The asterisks indicate the significance levels at the various distance from cell soma between SL and SC groups. ***p < 0.001;** p < 0.005; *p < 0.05. Vertical bars indicate SEM.
density and the proximal shift towards the cell somaoccurring in the apical arborization of SL neurons incomparison to SC neurons, and the substantial equiv-alence of spine density distribution in both enrichedgroups (Fig. 9).
DISCUSSION
Ch4 forebrain cholinergic lesion resulted in multi-faceted impairment of cognitive functions. It markedlyaffected the functions requiring a significant workingmemory load, weakened the procedural competencies,and only slightly affected the functions linked to declar-ative (localizatory) memory. Namely, the lesion pro-voked poor performances in the radial maze (see totalerrors and spatial span in Fig. 3A and B), induced lesstuned explorative strategies (see the augmented per-
centage of OF peripheral exploration and the reducednumber of 45◦ RAM angles in Fig. 3C), and finallyslowed down (see the large heading angles in the firstMWM sessions in Fig. 2B) but not prevented from oc-curring the acquisition of localizing knowledge (see theabsence of differences in heading angles in last MWMsessions in Fig. 2B). Conversely, the lesion did not af-fect the acquisition of the general MWM requests (seethe latencies to reach the platform in Fig. 2A), the ha-bituation to the context (Fig. 5), and the reaction to spa-tial change (Fig. 5). Remarkable perseverative behav-iors occurred in the presence of the lesion (Figs 3D and4B). Similar perseverative effects are reported in ratsperforming a five-choice serial reaction time task afterNBM lesion [38]. Considered as a whole, the combi-nation of decreased choice accuracy, increased perse-verative errors and tendency to hyperactivity is consis-tent with a disruption in inhibitory control and with im-

L. Mandolesi et al. / Enrichment and Cholinergic Depletion 25
paired working memory abilities resulting from aber-rant modulation of the prefrontal cortex [44] caused bythe cholinergic depletion.
In line with the effects provoked in the frontal pyra-midal neurons [22], the cholinergic deafferentation in-duced significant increases of spine number and densi-ty in the parietal pyramidal neurons in the apical andbasal dendrites. This finding supports the hypothe-sis that neocortical neurons upregulate spine numberas a compensatory response to the lesion-induced de-creased innervation [13]. In frontal pyramidal neu-rons, the lesion-induced increment of dendritic spineshas been related to an increased expression of GluR1subunit of the AMPA receptors [19,31], suggesting thespines as the morphological substrate for the increaseof AMPA receptors induced by the cholinergic lesion.Given the similarity of the compensatory reaction inthe parietal pyramidal neurons, it appears reasonableto suggest that the mechanisms of action proposed toact in frontal neurons might be common to all neocor-tical areas. However, it is important to underline thatthe increase in spine number and density observed inthe present research does not necessary imply an in-creased number of functional synapses. In fact, a studyon slice preparations of hippocampal neurons showedthat new spines form when synapses are inactivatedand persist for many hours in the absence of functionalactivity [32]. The presence of marked spatial deficitsobserved in SL animals could suggest that the newlyformed spines of the parietal pyramidal neurons arefunctionally inactive and thus not directly influencingthe spatial behavior. The analysis of behavioral deficitsof lesioned animals in which the spine formation is pre-vented by pharmacological factors [19] would clarifythe possible functional role of compensatory spines.
The environmental enrichment positively affectedcognitive performances even in the presence of thecholinergic depletion of the neocortex. In fact, on mosttasks, EL animals displayed substantial preservation ofmnesic functions (see the absence of significant differ-ences between EL and EC performances in RAM spanin Fig. 3B) and of procedural competencies (see theefficient MWM exploration strategies in Fig. 2C andthe increase of RAM 45◦ angles in Fig. 3C). Even inthe OF parameters, both enriched groups displayed nosignificant between-group differences but significantlydiffered from SC and SL groups (Fig. 5). In brief, en-riched experience preserved most performances evenin the presence of forebrain lesions, providing strongevidence that the increase of social interactions, cog-nitive stimulations and physical activity was able to
mitigate the cognitive deficits evoked by the choliner-gic depletion of the neocortex, in agreement with otherresearches [26,56]. However, some symptoms of theneurodegenerative damage were anyway observable inEL animals. Both lesioned groups exhibited markedperseverative tendencies (Figs 3D and 4B) and someimpairment in spatial working memory functions andprocedural competencies, as demonstrated by their flatcurves in RAM errors and 45◦ angles (Fig. 3A and C).Considered as a whole, these findings indicate on onehand that cholinergic system of the basal forebrain iscritical for optimizing performances in response to spe-cific behavioral challenges [2] and on the other handthat most lesion-induced deficits were beneficially in-fluenced by EE-rearing.
The behavioral findings were complemented by theanalyses on dendritic spines. Relatively to the ECgroup, EL rats exhibited spine values almost unmodi-fied by the presence of the lesion. In fact, spine numberand density values as well as distribution pattern weresubstantially similar to those of EC animals in bothapical and basal dendrites. Since in EL animals thismorphological pattern went together with the almosttotal maintenance of efficient spatial performances, itmay be advanced that the increased presence of spinesexhibited by EL rats enhanced the synaptic efficacy andthe excitability of the networks involved in learningprocesses. Afterwards, it is consequent to wonder whythe EL animals did not further increase spine values incomparison to the un-lesioned enriched group, as SLrats did in an attempt of compensating the presence ofthe lesion. This differential rearing effect suggests thatthe lesion-induced up-regulation of dendritic spines isa reaction not necessary in a situation of increased neu-ronal plasticity. As suggested by Young et al. [74],EE optimizes the structural morphology of the brainand the enriched brain already gets the right shapingof neuronal connectivity. This reasoning supports thepreviously advanced suggestion that the compensatorynewly formed spines observed in SL animals could beinactive and thus not functionally useful. The presenceof an augmented number of spinesper sedoes not war-rant improved performances, it seems necessary thatthe spines are not only “many” but also “good”. Futureresearch might be aimed to distinguish functional fromnon-functional spines and thus “useful” compensationphenomena from non-specific generalized reaction tocell damage. However, it has to be noted that cellular ormolecular changes, such as modified levels of synapto-physin, NGF and neprilysin, could be also responsiblefor the establishment of the “cognitive reserve”.

26 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
The maintenance of the functional synaptic patrimo-ny gained through the exposure to enriched environ-ment displayed by EL rats represents the strongest in-dication of the presence of a brain reserve to be spentin the case of lesion. Then, the enrichment-inducedstrengthening of synaptic connectivity (brain reserve)provides a potent mechanism to efficiently utilize theexisting neuronal networks and to engage alternativeneuronal circuits in order to improve behavioral perfor-mances (cognitive reserve).
CONCLUSIONS
A number of researches have indicated that enrichedhousing can dramatically improve cognitive abilitieseither in intact animals [37,72] and in brain-lesionedanimals [7,9,23,57,71]. However, most researchers re-ported beneficial effects of EE used as a therapeuticapproach following brain diseases. Namely, exposureto enriched living conditions results in improved be-havioral outcomes on several sensory-motor tasks afterfocal ischemic injury [11,20,48,50,69,73], followingstatus epilepticus [12,58], or subicular lesions [8]. Thebeneficial behavioral effects of cholinergic cell graftsinto the frontal cortex are amplified by subsequent EErearing [29]. Recently, it has been suggested that EEcould be used as a therapeutic technique after cholin-ergic damage [49] or in transgenic mouse models ofAlzheimer’s disease [1,39] to stabilize or improve cog-nitive functions. Furthermore, EE could mitigate thecognitive decline associated with aging [15]. Con-versely, only few studies have analyzed the effects ofthe exposure to complex environments on the outcomesof subsequent pathological accident. Delayed onsetof symptoms has been reported in transgenic modelsof Huntington’s disease [24,60,66], but these studiesreferred to the appearance of motor symptomatology.Furthermore, the response of animals enriched for 15days to a successive brain injury resulted in markedneurodegeneration and motor dysfunction with a high-er capacity for recovery [36]. Studies more focusedon cognition, although without morphological corre-lates, demonstrated that learning and memory deficitsobserved in transgenic mouse models for AD-like neu-rodegeneration can be ameliorated by EE [4,26].
The present behavioral and morphological findingsdemonstrate that complex experiences (rearing in en-riched environment) induce a more powerful synapticresponse (increase of number and density of dendrit-ic spines) and provide the means by which the brain
adaptively remodels to meet environmental demands(improvement of behavioral performances) even in thepresence of damage or degeneration (lesions of cholin-ergic basal forebrain system). In other words, the neu-roplasticity linked to complex cognitive experience en-hances brain properties of resilience to cope with latedamage, enabling the maintenance of normal cognitivefunction despite the presence of brain pathology [68].Since this is the fundamental assumption of reserve hy-pothesis, rearing in an enriched environment appearsto be a very suitable experimental method to build thebrain and cognitive reserves in analogy with humaneducational attainment and occupational status [43].
As Stern [62,63] has stated, subjects may rely ontwo interrelated kinds of resources, the neural “hard-ware”, such as more synapses and more extended den-dritic branches, and the neural “software”, such as moretuned cognitive skills and strategies. According tothe present results demonstrating impressive improve-ment in cognitive functions accompanied by increasedsynaptic connectivity, both reserves are strengthened inthe experimental model of rearing in complex environ-ments.
ACKNOWLEDGMENTS
This work was supported by MIUR grants to LP.
References
[1] G.W. Arendash, M.F. Garcia, D.A. Costa, J.R. Cracchiolo,I.M. Wefes and H. Potter, Environmental enrichment improvescognition in aged Alzheimer’s transgenic mice despite stablebeta-amyloid deposition,Neuroreport15 (2004), 1751–1754.
[2] M.G. Baxter and A.A. Chiba, Cognitive functions of the basalforebrain,Curr Opin Neurobiol9 (1999), 178–183.
[3] M.G. Baxter, D.J. Bucci, L.K. Gorman, R.G. Wiley and M.Gallagher, Selective immunotoxic lesions of basal forebraincholinergic cells: effects on learning and memory in rats,Behav Neurosci109 (1995), 714–722.
[4] N. Berardi, C. Braschi, S. Capsoni, A. Cattaneo and L. Maffei,Environmental enrichment delays the onset of memory deficitsand reduces neuropathological hallmarks in a mouse modelof Alzheimer-like neurodegeneration,J Alzheimers Dis11(2007), 359–370.
[5] J. Berger-Sweeney, S. Heckers, M.M. Mesulam, R.G. Wi-ley, D.A. Lappi and M. Sharma, Differential effects on spa-tial navigation of immunotoxin-induced cholinergic lesions ofthe medial septal area and nucleus basalis magnocellularis,JNeurosci14 (1994), 4507–4519.
[6] J.T. Coyle, D.L. Price and M.R. DeLong, Alzheimer’s disease:A disorder of cortical cholinergic innervation,Science219(1983), 1184–1190.

L. Mandolesi et al. / Enrichment and Cholinergic Depletion 27
[7] P. Dahlqvist, A. Ronnback, S.A. Bergstrom, I. Soderstromand T. Olsson, Environmental enrichment reverses learningimpairment in the Morris water maze after focal cerebral is-chemia in rats,Eur J Neurosci19 (2004), 2288–2298.
[8] A. Dhanushkodi, B. Bindu, T.R. Raju and B.M. Kutty, Expo-sure to enriched environment improves spatial learning per-formances and enhances cell density but not choline acetyl-transferase activity in the hippocampus of ventral subicular-lesioned rats,Behav Neurosci121 (2007), 491–500.
[9] M.D. Dobrossy and S.B. Dunnett, Environmental enrichmentaffects striatal graft morphology and functional recovery,EurJ Neurosci19 (2004), 159–168.
[10] C.J. Faherty, D. Kerley and R. J. Smeyne, A Golgi-Cox mor-phological analysis of neuronal changes induced by environ-mental enrichment,Brain Res Dev141 (2003), 55–61.
[11] R. Farrell, S. Evans and D. Corbett, Environmental enrichmentenhances recovery of function but exacerbates ischemic celldeath,Neuroscience107 (2001), 585–592.
[12] S.Faverjon, D.C. Silveira, D.D. Fu, B.H. Cha, C. Akman, Y.Hu and G.L. Holmes, Beneficial effects of enriched environ-ment following status epilepticus in immature rats,Neurology59 (2002), 1356–1364.
[13] J.C. Fiala, J. Spacek and K.M. Harris, Dendritic spine pathol-ogy: cause or consequence of neurological disorders?,BrainRes Rev39 (2002), 29–54.
[14] A. Fischer, F. Sananbenesi, X. Wang, M. Dobbin and L.H.Tsai, Recovery of learning and memory is associated withchromatin remodelling,Nature447 (2007), 178–182.
[15] K.M. Frick and S.M. Fernandez, Enrichment enhances spatialmemory and increases synaptophysin levels in aged femalemice,Neurobiol Aging24 (2003), 615–626.
[16] K.M. Frick, J.J. Kim and M.G. Baxter, Effects of completeimmunotoxin lesions of the cholinergic basal forebrain on fearconditioning and spatial learning,Hippocampus14 (2004),244–254.
[17] R.P. Friedland, T. Fritsch, K.A. Smyth, F. Koss, A.J. Lern-er, C.H. Chen, G.J. Petot and S.M. Debanne, Patients withAlzheimer’s disease have reduced activities in midlife com-pared with healthy control-group members,Proc Natl AcadSci USA98 (2001), 3440–3445.
[18] R. Galani, O. Lehmann, T. Bolmont, E. Aloy, F. Bertrand, C.Lazarus, H. Jeltsch and J.C. Cassel, Selective immunolesionsof CH4 cholinergic neurons do not disrupt spatial memory inrats,Physiol Behav76 (2002) ,75–90.
[19] J.E. Garret, I. Kim, R.E. Wilson and C.L. Wellman, Effect ofN-methyl-d-aspartate receptor blockade on plasticity of frontalcortex after cholinergic deafferentation in rat,Neuroscience140 (2006), 57–66.
[20] O.L. Gobbo and S.M. O’Mara, Impact of enriched-environment housing on brain-derived neurotrophic factor andon cognitive performance after a transient global ischemia,Behav Brain Res152 (2004), 231–241.
[21] A. Graziano, L. Petrosini and A. Bartoletti, Automatic recog-nition of explorative strategies in the Morris water maze,JNeurosci Meth130 (2003), 33–44.
[22] K.M. Harmon and C.L. Wellman, Differential effects ofcholinergic lesions on dendritic spines in frontal cortex ofyoung adult and aging rats,Brain Res992 (2003), 60–68.
[23] R.R. Hicks, L. Zhang, A. Atkinson, M. Stevenon, M. Ven-eracion and K.B. Seroogy, Environmental enrichment attenu-ates cognitive deficits but does not alter neurotrophin gene ex-pression in the hippocampus following lateral fluid percussionbrain injury,Neuroscience112 (2002), 631–637.
[24] E. Hockly, P.M. Cordery, B. Woodman, A. Mahal, A. VanDellen, C. Blakemore, C.M. Lewis, A.J. Hannan and G.P.Bates, Environmental enrichment slows disease progressionin R6/2 Huntington’s disease mice,Ann Neurol51 (2002),235–242.
[25] B.R. Ickes, T.M. Pham, L.A. Sanders, D.S. Albeck, A.H.Mohammed and A.C. Granholm, Long-term environmentalenrichment leads to regional increases in neurotrophin levelsin rat brain,Exp Neurol64 (2000), 45–52.
[26] J.L. Jankowsky, T. Melnikova, D.J. Fadale, G.M. Xu, H.H.Slunt, V. Gonzales, L.H. Younkin, S.G. Younkin, D.R.Borchelt and A.V. Savonenko, Environmental enrichment mit-igates cognitive deficits in a mouse model of Alzheimer’s dis-ease,J Neurosci25 (2005), 5217–5224.
[27] X. Jiang, R.R. Johnson and A. Burkhalter, Visualization ofdendritic morphology of cortical projection neurons by retro-grade axonal tracing,J Neurosci Meth50 (1993), 45–60.
[28] B.B. Johansson and P.V. Belichenko, Neuronal plasticity anddendritic spines: effect of environmental enrichment on intactand postischemic rat brain,J Cereb Blood Flow Metab22(2002), 89–96.
[29] C. Kelche, C. Roeser, H. Jeltsch, J.C. Cassel and B. Will,The effects of intrahippocampal grafts training and postoper-ative housing on behavioral recovery after septohippocampaldamage in the rat,Neurobiol Learn Mem63 (1995), 155–166.
[30] G. Kempermann, H.G. Kuhn and F.H. Gage, More hippocam-pal neurons in adult mice living in an enriched environment,Nature386 (1997), 493–495.
[31] I. Kim, R.E. Wilson and C.L. Wellman, Aging and cholinergicdeafferentation alter GluR1 expression in rat frontal cortex,Neurobiol Aging26 (2005), 1073–1081.
[32] S.A. Kirov and K.M. Harris, Dendrites are more spiny onmature hippocampal neurons when synapses are inactivated,Nat Neurosci2 (1999), 878–883.
[33] B. Kolb, M. Forgie, R. Gibb, G. Gorny and S. Rowntree, Age,experience and the changing brain,Neurosci Biobehav Rev22(1998), 143–159.
[34] B. Kolb, R. Gibb and G. Gorny, Experience-dependentchanges in dendritic arbor and spine density in neocortexvary qualitatively with age and sex,Neurobiol Learn Mem79(2003), 1–10.
[35] B. Kolb, G. Gorny, A.H.V. Soderpalm and T.E. Robinson, En-vironmental complexity has different effects on the structureof neurons in the prefrontal cortex versus the parietalcortex ornucleus accumbens,Synapse48 (2003), 149–153.
[36] D.A. Kozlowski, B.V. Nahed, D.A. Hovda and S.M. Lee, Para-doxical effects of cortical impact injury on environmentallyenriched rats,J Neurotrauma21 (2004), 513–519.
[37] M.G. Leggio, L. Mandolesi, F. Federico, F. Spirito, B. Ricci,F. Gelfo and L. Petrosini, Environmental enrichment promotesimproved spatial abilities and enhanced dendritic growth inthe rat,Behav Brain Res163 (2005), 78–90.
[38] O. Lehmann, A.J. Grottick, J.C. Cassel and G.A. Higgins, Adouble dissociation between serial reaction time and radialmaze performance in rats subjected to 192 IgG-saporin lesionsof the nucleus basalis and/or the septal region,Eur J Neurosci18 (2003), 651–666.
[39] O. Levi, A.L. Jongen-Relo, J. Feldon, A.D. Roses and D.M.Michaelson, ApoE4 impairs hippocampal plasticity isoform-specifically and blocks the environmental stimulation ofsynaptogenesis and memory,Neurobiol Dis13 (2003), 273–282.
[40] T. Malm, M. Ort, L. Tahtivaara, N. Jukarainen, G. Gold-steins, J Puolivali, A. Nurmi, R. Pussinen, T. Ahtoniemi,

28 L. Mandolesi et al. / Enrichment and Cholinergic Depletion
T.K. Miettinen, K. Kanninen, S. Leskinen, N. Vartiainen, J.Yrj anheikki, R. Laatikainen, M.E. Harris-White, M. Koistina-ho, S.A. Frautschy, J. Bures and J. Koistinaho, Beta-amyloidinfusion results in delayed and age-dependent learning deficitswithout role of inflammation or beta-amyloid deposits,ProcNatl Acad Sci USA103 (2006), 8852–8857.
[41] L. Mandolesi, M.G. Leggio, F. Spirito and L. Petrosini, Cere-bellar contribution to spatial event processing: do spatial pro-cedures contribute to formation of spatial declarative knowl-edge?Eur J Neurosci18 (2003), 2618–2626.
[42] M. Mesulam, E.J. Mufson, A.I. Levey and B.H. Wainer, Cen-tral cholinergic pathways in the rat: an overview based onalternative nomenclature (Ch1-Ch6),Neuroscience10 (1983),1185–1201.
[43] N.W. Milgram, C.T. Siwak-Tapp, J .Arahujo and E. Head,Neuroprotective effects of cognitive enrichment,Ageing ResRev5 (2006), 354–369.
[44] L.A. Miner, M. Ostrander and M. Sarter, Effects of ibotenicacid induced loss of neurons in the medial prefrontal cortex ofrats on behavioral vigilance: evidence of executive dysfunc-tion, J Psychopharmachol11 (1997), 169–178.
[45] J.A. Mortimer, Do psychosocial risk factors contribute toAlzheimer’s disease? in:Etiology of dementia of Alzheimer’stype, A.S. Henderson and J.H. Henderson, eds, Wiley, NewYork, 1988, pp. 39–52.
[46] M. Nilsson, E. Perfilieva, U. Johansson, O. Orwar and P.S.Eriksson, Enriched environment increases neurogenesis in theadult rat dentate gyrus and improves spatial memory,J Neu-robiol 39 (1999), 569–578.
[47] J. Nithianantharajah and A.J. Hannan, Enriched environments,experience-dependent plasticity and disorders of the nervoussystem,Nat Rev Neurosci7 (2006), 697–709.
[48] R.J. Nudo, B.M. Wise, F. SiFuentes and G.W. Milliken, Neuralsubstrates for the effects of rehabilitative training on motorrecovery after ischemic infarct,Science272 (1996), 1791–1794.
[49] V. Paban, M. Jaffard, C. Chambon, M. Malafosse and B.Alescio-Lautier, Time course of behavioral changes follow-ing basal forebrain cholinergic damage in rats: Environmentalenrichment as a therapeutic intervention,Neuroscience132(2005), 13–32.
[50] M.J. Passineau, E.J. Green and W.D. Dietrich, Therapeuticeffects of environmental enrichment on cognitive function andtissue integrity following severe traumatic brain injury in rats,Exp Neurol168 (2001), 373–384.
[51] G. Paxinos and C. Watson,The Rat Brain in Stereotaxic Co-ordinates, Academic Press, San Diego, 1998.
[52] T. Perry, H. Hodges and J.A. Gray, Behavioural, histologicaland immunocyto-chemical consequences following 192 IgG-saporin immuno-lesions of the basal forebrain cholinergic sys-tem,Brain Res Bull54 (2001), 29–48.
[53] S.K. Pope, V.M. Shue and C. Beck, Will a healthy lifestylehelp prevent Alzheimer’s disease?Ann Rev Public Health24(2003), 111–132.
[54] C. Rampon, Y.P. Tang, J. Goodhouse, E. Shimizu, M. Kyinand J.Z. Tsien, Enrichment induces structural changes andrecovery from nonspatial memory deficits in CA1 NMDAR1-knockout mice,Nat Neurosci3 (2000), 238–244.
[55] M. Richards, I.J and Deary, A life course approach to cognitivereserve: A model for cognitive aging and development?AnnNeurol58 (2005), 617–622.
[56] A. Risedal, B. Mattsson, P. Dahlqvist, C. Nordborg, T. Olssonand B.B. Johansson, Environmental influences on functional
outcome after a cortical infarct in the rat,Brain Res Bull58(2002), 315–321.
[57] M.R. Rosenzweig and E.L. Bennett, Psychobiology of plastic-ity: effects of training and experience on brain and behaviour,Behav Brain Res78 (1996), 57–65.
[58] A. Rutten, M. van Albada, D.C. Silveira, B.H. Cha, X. Liu,Y.N. Hu, M.R. Cilio and G.L. Holmes, Memory impairmentfollowing status epilepticus in immature rats: time-course andenvironmental effects,Eur J Neurosci16 (2002), 501–513.
[59] D. Schubert, R. Kotter, K. Zilles, H.J. Luhmann and J.F.Staiger, Cell type-specific circuits of cortical layer IV spinyneurons,J Neurosci23 (2003), 2961–2970.
[60] T.L. Spires, H.E. Grote, N.K. Varshney, P.M. Cordery, A. VanDellen, C. Blakemore and A.J. Hannan, Environmental en-richment rescues protein deficits in a mouse model of Hunt-ington’s disease indicating a possible disease mechanism,JNeurosci24 (2004), 2270–2276.
[61] R.T. Staff, A.D. Murray, I.J. Deary and L.J. Whalley, Whatprovides cerebral reserve?Brain 127 (2004), 1191–1199.
[62] Y. Stern, The concept of cognitive reserve: a catalyst forresearch,J Clin Exp Neuropsychol25 (2003), 589–593.
[63] Y. Stern, Cognitive reserve and Alzheimer disease,AlzheimerDis Assoc Disorder20 (2006), 112–117.
[64] Y. Stern, B. Gurland, T.K. Tatemichi, M.X. Tang, D. Wilderand R. Mayeux, Influence of education and occupation on theincidence of Alzheimer’s disease,JAMA 271 (1994), 1004–1010.
[65] C.A. Turner, M.H. Lewis and M.A. King, Environmental en-richment: effect on stereotyped behavior and dendritic mor-phology,Dev Psychobiol43 (2003), 20–27.
[66] A. Van Dellen, C. Blakemore, R. Deacon, D. York and A.J.Hannan, Delaying the onset of Huntington’s in mice,Nature404 (2000), 721–722.
[67] H. Van Praag, G. Kempermann and F.H. Gage, Neural conse-quences of environmental enrichment,Nature Rev Neurosci1(2000), 191–198.
[68] I.M. Van Rijzingen, W.H. Gispen and B.M. Spruijt, Post-operative environmental enrichment attenuates fimbria-fornixlesion-induced impairments in Morris maze performance,Neurobiol Learn Mem67 (1997), 21–28.
[69] A.K. Wagner, A.E. Kline, J. Sokoloski, R.D. Zafonte, E. Ca-pulong and C.E. Dixon, Intervention with environmental en-richment after experimental brain trauma enhances cognitiverecovery in male but not female rats,Neurosci Lett334 (2002),165–168.
[70] L.J. Whalley, I.J. Deary, C.L. Appleton and J.M. Starr, Cogni-tive reserve and the neurobiology of cognitive aging,AgeingRes Rev3 (2004), 369-382.
[71] B. Will, R. Galani, C. Kelche and M.R. Rosenzweig, Re-covery from brain injury in animals: relative efficacy of en-vironmental enrichment physical exercise or formal training(1990–2002),Progr Neurobiol72 (2004), 162–182.
[72] E.A. Woodcock and R. Richardson, Effects of environmentalenrichment on rate of contextual processing and discriminativeability in adult rats,Neurobiol Learn Mem73 (2000), 1–10.
[73] C. Xerri and Y. Zennou-Azogui, Influence of the postlesionenvironment and chronic piracetam treatment on the organiza-tion of the somatotopic map in the rat primary somatosenso-ry cortex after focal cortical injury,Neuroscience118 (2003),161–177.
[74] D. Young, P.A. Lawlor, P. Leone, M. Dragunow and M.J. Dur-ing, Environmental enrichment inhibits spontaneous apoptosisprevents seizures and is neuroprotective,Nature Med5 (1999),448–453.
Related Documents











