233 Journal of Vegetation Science 17: 233-244, 2006 © IAVS; Opulus Press Uppsala. Abstract Questions: How do forest types differ in their distinctiveness among islands in relation to environmental and anthropogenic disturbance gradients? Are biogeographic factors also involved? Location: Tonga, ca. 170 oceanic islands totalling 700 km 2 spread across 8° of latitude in Western Polynesia. Method: Relative basal area was analysed for 134 species of woody plants in 187 plots. We used clustering, indirect gradi- ent analysis, and indicator species analysis to identify continu- ous and discontinuous variation in species composition across geographical, environmental and disturbance gradients. Par- tial DCA related environmental to compositional gradients for each major forest type after accounting for locality. CCA and partial CCA partitioned observed compositional variation into components explained by environment/disturbance, locality and covariation between them. Results: Differences among forest types are related to envi- ronment and degree of anthropogenic disturbance. After ac- counting for inter-island differences, compositional variation (1) in coastal forest types is related to substrate, steepness and proximity to coast; (2) in early-successional, lowland rain forest to proximity to the coast, steepness and cultivation disturbance; (3) in late-successional, lowland forest types to elevation. For coastal/littoral forests, most of the composi- tional variation (71%) is explained by disturbance and envi- ronmental variables that do not covary with island while for both early and late-successional forests there is a higher de- gree of compositional variation reflecting covariation between disturbance/environment and island. Conclusions: There are regional similarities, across islands, among littoral/coastal forest types dominated by widespread seawater-dispersed species. The early-successional species that dominate secondary forests are distributed broadly across islands and environmental gradients, consistent with the gra- dient-in-time model of succession. Among-island differences in early-successional forest may reflect differences in land-use practices rather than environmental differences or biogeo- graphical history. In late-successional forests, variation in composition among islands can be partly explained by differ- ences among islands and hypothesized tight links between species and environment. Disentangling the effects of anthro- pogenic disturbance history versus biogeographic history on late-successional forest in this region awaits further study. Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia Franklin, Janet 1* ; Wiser, Susan K. 2 ; Drake, Donald R. 3 ; Burrows, Larry E. 2,4 & Sykes, William R. 2,5 1 Department of Geography, San Diego State University, San Diego, CA 92182-4614, USA; 2 Landcare Research, Christchurch, New Zealand; E-mail [email protected]; 3 Botany Department, University of Hawai`i at Manoa, Honolulu, HI 96822; E-mail [email protected]; 4 E-mail [email protected]; 5 E-mail [email protected]; * Corresponding author; Fax +1 6195945676; E-mail [email protected] Introduction Island studies have been fundamental to understand- ing influences of geographic and ecological processes on community composition and structure (Carlquist 1974; Vitousek et al. 1995; Whittaker 1998; Mueller- Dombois 2002; Drake et al. 2002). This requires inte- grating (1) biogeography, which creates distinctive pat- terns through isolation and evolutionary divergence and extinction; (2) natural and anthropogenic disturbance; (3) gradients in distance from the sea, elevation and edaphic controls (Mueller-Dombois & Fosberg 1998). As model systems, tropical islands can also increase our understanding of processes that influence rain forest composition worldwide. Studies of continental tropical rain forests have emphasized the importance of environ- mental gradients such as topography, soils, and climate (e.g. Gentry 1988; Pyke et al. 2001; Tuomisto et al. 2003), natural disturbance (Whitmore 1989; Burslem et al. 2000), anthropogenic disturbance (Horn & Kennedy 2001; van Gemerden et al. 2003), and interactions among these factors (e.g. Svenning et al. 2004). Tropical island sys- tems allow these influences to be examined across larger geographic scales, and because the forest communities tend to be simpler, the entire tree community can be studied (cf. Pyke et al. 2001; Svenning et al. 2004). Keywords: Coastal forest; Human impact; Limestone; Litto- ral forest; Lowland rain forest; Pacific; Succession; Tropical forest; Volcanic; Zonation. Abbreviations: GA = Group averaging; MRPP = Multi- response Permutation Procedure; NMS = Non-metric Multidi- mensional Scaling; pCCA = Partial CCA. Nomenclature: Smith (1979, 1981, 1985, 1988, 1991); for species not treated by Smith: Yuncker (1959), Whistler (1991), Wagner et al. (1999).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

- Environment, disturbance history and rain forest composition across the islands of Tonga - 233
Journal of Vegetation Science 17: 233-244, 2006© IAVS; Opulus Press Uppsala.
AbstractQuestions: How do forest types differ in their distinctivenessamong islands in relation to environmental and anthropogenicdisturbance gradients? Are biogeographic factors also involved?Location: Tonga, ca. 170 oceanic islands totalling 700 km2
spread across 8° of latitude in Western Polynesia.Method: Relative basal area was analysed for 134 species ofwoody plants in 187 plots. We used clustering, indirect gradi-ent analysis, and indicator species analysis to identify continu-ous and discontinuous variation in species composition acrossgeographical, environmental and disturbance gradients. Par-tial DCA related environmental to compositional gradients foreach major forest type after accounting for locality. CCA andpartial CCA partitioned observed compositional variation intocomponents explained by environment/disturbance, localityand covariation between them.Results: Differences among forest types are related to envi-ronment and degree of anthropogenic disturbance. After ac-counting for inter-island differences, compositional variation(1) in coastal forest types is related to substrate, steepness andproximity to coast; (2) in early-successional, lowland rainforest to proximity to the coast, steepness and cultivationdisturbance; (3) in late-successional, lowland forest types toelevation. For coastal/littoral forests, most of the composi-tional variation (71%) is explained by disturbance and envi-ronmental variables that do not covary with island while forboth early and late-successional forests there is a higher de-gree of compositional variation reflecting covariation betweendisturbance/environment and island.Conclusions: There are regional similarities, across islands,among littoral/coastal forest types dominated by widespreadseawater-dispersed species. The early-successional speciesthat dominate secondary forests are distributed broadly acrossislands and environmental gradients, consistent with the gra-dient-in-time model of succession. Among-island differencesin early-successional forest may reflect differences in land-usepractices rather than environmental differences or biogeo-graphical history. In late-successional forests, variation incomposition among islands can be partly explained by differ-ences among islands and hypothesized tight links betweenspecies and environment. Disentangling the effects of anthro-pogenic disturbance history versus biogeographic history onlate-successional forest in this region awaits further study.
Environment, disturbance history and rain forest compositionacross the islands of Tonga, Western Polynesia
Franklin, Janet1*; Wiser, Susan K.2; Drake, Donald R.3;Burrows, Larry E.2,4 & Sykes, William R.2,5
1Department of Geography, San Diego State University, San Diego, CA 92182-4614, USA; 2Landcare Research, Christchurch, New Zealand; E-mail [email protected]; 3Botany Department, University of Hawai`i at
Manoa, Honolulu, HI 96822; E-mail [email protected]; 4E-mail [email protected];5E-mail [email protected]; *Corresponding author; Fax +1 6195945676; E-mail [email protected]
Introduction
Island studies have been fundamental to understand-ing influences of geographic and ecological processeson community composition and structure (Carlquist1974; Vitousek et al. 1995; Whittaker 1998; Mueller-Dombois 2002; Drake et al. 2002). This requires inte-grating (1) biogeography, which creates distinctive pat-terns through isolation and evolutionary divergence andextinction; (2) natural and anthropogenic disturbance;(3) gradients in distance from the sea, elevation andedaphic controls (Mueller-Dombois & Fosberg 1998).As model systems, tropical islands can also increase ourunderstanding of processes that influence rain forestcomposition worldwide. Studies of continental tropicalrain forests have emphasized the importance of environ-mental gradients such as topography, soils, and climate(e.g. Gentry 1988; Pyke et al. 2001; Tuomisto et al. 2003),natural disturbance (Whitmore 1989; Burslem et al. 2000),anthropogenic disturbance (Horn & Kennedy 2001; vanGemerden et al. 2003), and interactions among thesefactors (e.g. Svenning et al. 2004). Tropical island sys-tems allow these influences to be examined across largergeographic scales, and because the forest communitiestend to be simpler, the entire tree community can bestudied (cf. Pyke et al. 2001; Svenning et al. 2004).
Keywords: Coastal forest; Human impact; Limestone; Litto-ral forest; Lowland rain forest; Pacific; Succession; Tropicalforest; Volcanic; Zonation.
Abbreviations: GA = Group averaging; MRPP = Multi-response Permutation Procedure; NMS = Non-metric Multidi-mensional Scaling; pCCA = Partial CCA.
Nomenclature: Smith (1979, 1981, 1985, 1988, 1991); forspecies not treated by Smith: Yuncker (1959), Whistler (1991),Wagner et al. (1999).

234 Franklin, J. et al.
Western Polynesian rain forests occur on islandswith a long history of human impacts (Hughes et al.1979; Kirch & Ellison 1994). They are influenced bycyclones (e.g. Woodroffe 1984; Elmqvist et al. 1994;Franklin et al. 2004) and environmental gradients ofcoastal influence, elevation and substrate (Franklin et al.1999; Wiser et al. 2002). They are ideal systems to explorehow interactions among environment, disturbance his-tory and biogeography influence rain forest composi-tion, yet quantitative studies of these forests have thusfar been limited to individual islands or island groups(e.g. Garnock-Jones 1978; Whistler 1980; Kirkpatrick& Hassall 1985; Webb & Fa’aumu 1999; Keppel et al.2005). We assembled data from four previous forestsurveys conducted in Tonga to address three questions:
1. How do forest types differ in distinctiveness amongislands? For example, littoral and coastal forests world-wide harbour wide-ranging, sea-dispersed species(Guppy 1906; Ridley 1930; Whitmore 1985), so weexpect coastal/littoral forests on different island groupsto be less distinctive from one another than inland for-ests. We also expect early-successional forest to be domi-nated by a few widespread species with broad ecologicaltolerances and therefore to be more similar on differentislands than late-successional forest.
2. Do environmental gradients of coastal influenceand elevation and gradients of anthropogenic disturbancehave consistent impacts on forests across island groups?
3. Are there patterns of species composition related tolocality (island group) that may suggest the importanceof biogeographic factors instead? At this extensive geo-graphic scale we expect space (island locality) to repre-sent biogeographic factors (large-scale dispersal limita-tions affecting the regional species pool, local extirpa-tion, speciation), rather than recruitment limitation(Harms et al. 2000) from local species pools (Zobel1997), although it could represent unmeasured environ-mental or disturbance factors that are spatially corre-lated with island locality.
Material and Methods
Study area and the data sets
Tonga is the easternmost island group on the Indo-Australian plate, and comprises about 170 islands total-ling 700 km2 of land spread across 600 000 km2 of ocean(Fig. 1). They are mainly low limestone islands fromless than 1 to 5 - 10 million years old, with activevolcanic islands to the north and west (Tappin 1993;Dickinson et al. 1999). Eruptions of the volcanic outliershave covered the limestone islands with andesite tephrathat has weathered to rich soils.
Tonga is within the western Polynesian biogeographicregion (Stoddart 1992) (Fig. 1), where rain forests arespecies-poor compared to those of Australasia fromwhich their floras are derived by long-distance dispersal(Raven & Axelrod 1972; Carlquist 1996; Mueller-Dombois & Fosberg 1998; Turner et al. 2001). Tonga,for example, has about 450 native flowering plant species(Park & Whistler 2001), Samoa 550 (Whistler 1992)and Fiji 1769 (Ash 1992). Non-native species approxi-mately double the sizes of the floras. Endemism is lowwithin Tonga (3% of the native flora), but fairly highwithin Western Polynesia as a whole (van Balgooy et al.1996).
Previous studies surveyed rain forest composition inthree major island groups in Tonga (App. 1). The exten-sive, relatively undisturbed forests on ’Eua (Drake et al.1996), Kao and Tofua (Park & Whistler 2001) (Fig. 1)were surveyed for conservation planning. On Tongatapu,the largest and most populous island, where forest frag-ments and very early successional forest cover about11% of the island, plots were allocated to capture mappedvariation in soil type and coastal influence and evaluateconsequent composition, regeneration, and alien speciesoccurrence (Wiser et al. 2002). Data from the small
Fig. 1. Western Polynesia Biogeographical Re-gion, showing the islands and island groups ofTonga mentioned in the text (redrawn fromMueller-Dombois & Fosberg 1998).

- Environment, disturbance history and rain forest composition across the islands of Tonga - 235
nearby island of ’Euaiki, collected during the Tongatapusurvey, are analysed here for the first time. In Vava’u,survey objectives included characterizing a small forest(1 km2) for conservation (Bolick 1995), contrastingpaired early- and late-successional forest fragmentsthroughout the island group (Franklin et al. 1999), andassessing the composition and status of littoral/coastalforest (Steadman et al. 1999). We follow Whistler (1992)in using ‘littoral’ to refer to forests adjacent to the coast,i.e. within the spray zone, and ‘coastal’ for foreststransitional between littoral and lowland. Data from thelow islands of eastern Ha’apai (JF, unpubl. data) are alsoanalysed here for the first time. In Vava’u, ’Eua andTongatapu air photos and soil maps were used to stratifysampling; in the other surveys plots were sited subjec-tively to capture variation in forest composition ob-served during reconnaissance.
The data analysed in this study comprise the relativebasal area (RBA) of woody plants ≥ 5 cm DBH of 134non-cultivated species (App. 2) in 187 plots. Plots were600 m2 except for those on Kao and Tofua (1000 m2).Environmental variables on record for all plots reflectindirect topographic gradients correlated with maritimeinfluence and exposure, substrate characteristics, anddisturbance history. These included elevation and dis-tance to the coast (determined from topographic maps),and slope and aspect (measured in the field). We derivedan index of windwardness by calculating how similarplot-aspect is to the windward (SE) aspect, withcos(aspect –135°) equals 1 for SE aspects, –1 for NW(leeward) aspects, and 0 for NE or SW aspects (modi-fied from Beers et al. 1966). Substrate was recorded assand, limestone, volcanic, swamp, coral or soil. Aver-age percent exposed rock was converted to an ordinalscale to minimize observer differences between studies(0 = 0, 1-10 = 1, 11-40 = 2, 41-70 = 3, 71-100 = 4).Finally, because Tonga lacks detailed land-use recordsand synoptic historical air photos, we calculated ananthropogenic disturbance or ‘cultivation’ index as thesum of relative basal areas of six cultivated species(Cocos nucifera, Artocarpus altilis, three Citrus spp.,and Mangifera indica) that were then excluded as de-pendent variables (cf. Wiser et al. 2002). For Tofua andKao, elevation, aspect and substrate were determinedfrom plot descriptions and slope was estimated frommaps. Percent rock was not recorded so the ‘group’average was used.
Data analysis
Classification and ordination were used to detectdiscontinuous and continuous variation in forest com-position and relate this to variation in environmental andgeographical factors and human disturbance history.
Clustering (Greig-Smith 1983), based on Bray-Curtisdistance calculated from relative abundances, and usinggroup averaging (GA) as the linkage method, was car-ried out for the species by plot matrix. For comparison,we used TWINSPAN, Two-way Indicator SpeciesAnalysis (Hill 1979), and examined the two sets ofresults for congruence. Classification was followed by amulti-response permutation procedure (MRPP; Mielke1984) to test the differences among groups of plotsdefined by the classification, and by indicator speciesanalysis (Dufrêne & Legendre 1997) to determine thesignificance (by simulation) of indicator values (theaverage of relative abundance and relative frequency)for species associated with groups. The difference amonggroups in the disturbance-related cultivation index wastested.
Indirect gradient analysis was carried out usingDetrended Correspondence Analysis (DCA; ter Braak1995). Non-metric Multidimensional Scaling (NMS;Clarke 1993) was run in parallel with the DCA and theresults examined for congruence (Økland 1996; e.g.Pyke et al. 2001; Rydgren et al. 2003). In the DCA, rarespecies were downweighted and axes rescaled. NMSwas carried out using default parameters with PC-Ordsoftware (McCune & Mefford 1999). Patterns of groupsidentified by the classification were examined graphi-cally with respect to the ordination axes. Spearman’srank correlations between environmental variables andthe ordination axes derived from the indirect gradientanalysis were calculated.
To describe consistent relationships between veg-etation composition and both major environmental gra-dients and anthropogenic disturbance we used partialindirect ordination (pDCA) (ter Braak & Prentice 1988)with island designated as a covariable. We performedthese analyses for each of three major forest typesidentified by clustering. We then used canonical corre-spondence analysis (CCA) and partial CCA (pCCA) topartition observed compositional variation into compo-nents explained by environment and disturbance alone,island locality (island group, representing biogeographi-cal factors) alone, and covariation between the two (cf.Borcard et al. 1992; Økland 2003). Plots were groupedas: Tongatapu, ’Eua, ’Euaiki, Vava’u, and Kao plusTofua (the two Ha’apai plots were excluded). To avoidover-fitting, we used forward selection to determine theset of environmental variables to include in the analysis.Again, we performed these analyses for each foresttype, applying Monte Carlo tests to determine if rela-tionships between supplied explanatory variables andcomposition were statistically significant (ter Braak &Smilauer 1988; Crowley 1992).
Our interpretation followed Økland (1999), focus-ing on the relative amounts of variation explained by
‹

236 Franklin, J. et al.
each analysis, rather than the ratio between variationexplained and total inertia (e.g. Borcard et al. 1992;Økland & Eilertsen 1994). For each analysis, the sum ofthe canonical eigenvalues represents the variation ex-plained by the constraining variables (after removingvariation explained by any covariables). The percentageaccounted for by each analysis was obtained by dividingthe sum of canonical eigenvalues for that analysis by thetotal variation explained. This was then partitioned asthat accounted for: (1) solely by environment and distur-bance; (2) solely by island group; and (3) by covariationbetween the two. To aid our interpretation of the varia-tion partitioning results, we determined how islandsdiffered by applying MANOVA to continuously distrib-uted variables describing environment and disturbanceand comparing percentage frequency of occurrence ofplots in each substrate category on different islands. Todetermine whether the variables used in these modelswere indeed adequate to explain the observed com-positional variation, we compared eigenvalues and standscore positions between a DCA and DCCA (constrainedby ‘island’ and selected environmental/disturbance vari-ables) conducted for each forest type data set.
Clustering was carried out using PC-Ord (McCune& Mefford 1999), ordination using CANOCO (ter Braak& Smilauer 1998), and other statistical analyses usingS-Plus 6.1 (Anon. 1999) and R (Anon. 2004).
Table 1. Groups of plots based on classification using Group Averaging (see text). Indicator species are shown (p < 0.01). Forestgroup names are based on species dominance; BA = basal area; Number of plots by location (island[s]) is given as: Tof = Kao andTofua; Eu = ’Eua; Va = Vava’u; Ton = Tongatapu; Ik = ’Euaiki near Tongatapu; Ha = eastern Ha’apai.
Forest type/ Group Elevation BA Density Tof Eu Va Ton Ik Ha Indicator(m) (m2 ha–1) (trees ha–1) species
LittoralHernandia/ Barringtonia 0.5-10 68 ± 29 960 ± 366 4 13 6 H. nymphaeifolia, B. asiaticaCasuarina 2-40 32 ± 10 892 ± 540 1 1 1 C. equisetifolia
CoastalPisonia 7-30 38 ± 20 1037 ± 405 1 2 1 P. grandisExcoecaria 0-25 32 ± 13 1313 ± 469 1 11 15 1 E. agallocha
Early-successional lowland rain forestRhus 5-250 29 ± 10 1215 ± 310 5 17 14 2 R. taitensis, Cocos nucifera, Grewia
crenata, Alphitonia zizyphoides,Pometia pinnata
Late-successional lowland rain forestMyristica 8-180 47 ± 18 1024 ± 299 12 5 M. hypargyraeaManiltoa 5-180 40 ± 11 1373 ± 410 2 39 1 M. grandiflora, Cryptocarya
turbinata, Zanthoxylum pinnatum,Pleiogynium timoriense,Garuga floribunda
Calophyllum neo-ebudicum 100-400 55 ± 18 1224 ± 344 5 16 1 C. neo-ebudicum, Garciniamyrtifolia, Neonauclea forsteri,Dysoxylum tongense, Citronellasamoensis, Podocarpus pallidus
Inocarpus 1-35 60 ± 13 992 ± 366 1 8 I. fagiferErythrina 1 75 999 1 E. fusca
Results
Classification
Clustering using GA as the linkage method andretaining ten groups of plots (< 15% similarity) yieldedgroups similar to TWINSPAN’s groupings (ca. 70%correspondence), so only the GA results are shown.Four littoral/coastal groups and six lowland groups wereidentified. One inland group represents early-succes-sional stands, and five represent differences in speciesdominance among late-successional stands (Table 1).These groups are distinct based on MRPP (p < 0.0001),and within-group homogeneity (A = 0.23) is typical ofcommunity data.
Plots characterizing littoral/coastal forest groups werewell distributed among islands (Table 1). Littoral/coastalforests typically occur below 10 m elevation, except onmore exposed windward aspects where they reach 25 m(Fig. 2a). Exceptions were one plot dominated by early-successional Casuarina equisetifolia on young volcanicsubstrate, and two that were transitional between coastaland lowland on small windward islands. The littoralgroups, Hernandia/Barringtonia and Casuarina, occurprimarily within 40 m of the shore, while the coastalgroups, Pisonia and Excoecaria, may occur further in-land (Fig. 2b) and sometimes in association with rainforest species (and also with mangroves for Excoecaria).
‹
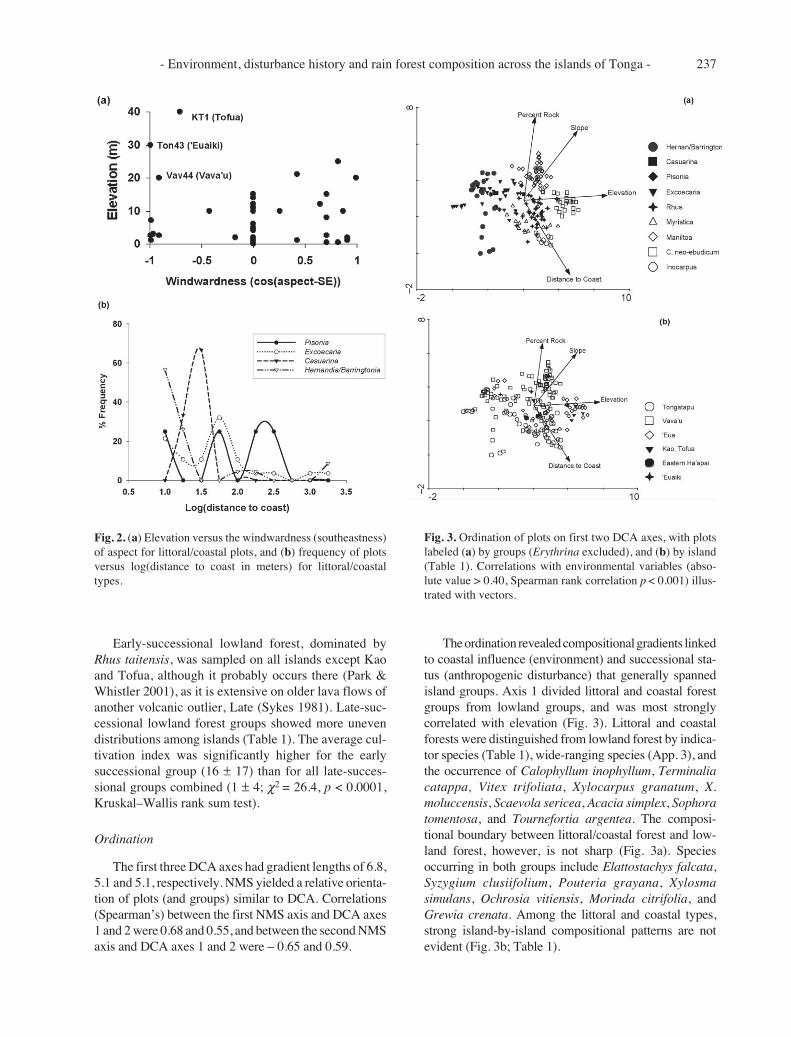
- Environment, disturbance history and rain forest composition across the islands of Tonga - 237
Fig. 2. (a) Elevation versus the windwardness (southeastness)of aspect for littoral/coastal plots, and (b) frequency of plotsversus log(distance to coast in meters) for littoral/coastaltypes.
Fig. 3. Ordination of plots on first two DCA axes, with plotslabeled (a) by groups (Erythrina excluded), and (b) by island(Table 1). Correlations with environmental variables (abso-lute value > 0.40, Spearman rank correlation p < 0.001) illus-trated with vectors.
The ordination revealed compositional gradients linkedto coastal influence (environment) and successional sta-tus (anthropogenic disturbance) that generally spannedisland groups. Axis 1 divided littoral and coastal forestgroups from lowland groups, and was most stronglycorrelated with elevation (Fig. 3). Littoral and coastalforests were distinguished from lowland forest by indica-tor species (Table 1), wide-ranging species (App. 3), andthe occurrence of Calophyllum inophyllum, Terminaliacatappa, Vitex trifoliata, Xylocarpus granatum, X.moluccensis, Scaevola sericea, Acacia simplex, Sophoratomentosa, and Tournefortia argentea. The composi-tional boundary between littoral/coastal forest and low-land forest, however, is not sharp (Fig. 3a). Speciesoccurring in both groups include Elattostachys falcata,Syzygium clusiifolium, Pouteria grayana, Xylosmasimulans, Ochrosia vitiensis, Morinda citrifolia, andGrewia crenata. Among the littoral and coastal types,strong island-by-island compositional patterns are notevident (Fig. 3b; Table 1).
Early-successional lowland forest, dominated byRhus taitensis, was sampled on all islands except Kaoand Tofua, although it probably occurs there (Park &Whistler 2001), as it is extensive on older lava flows ofanother volcanic outlier, Late (Sykes 1981). Late-suc-cessional lowland forest groups showed more unevendistributions among islands (Table 1). The average cul-tivation index was significantly higher for the earlysuccessional group (16 ± 17) than for all late-succes-sional groups combined (1 ± 4; χ2 = 26.4, p < 0.0001,Kruskal–Wallis rank sum test).
Ordination
The first three DCA axes had gradient lengths of 6.8,5.1 and 5.1, respectively. NMS yielded a relative orienta-tion of plots (and groups) similar to DCA. Correlations(Spearman’s) between the first NMS axis and DCA axes1 and 2 were 0.68 and 0.55, and between the second NMSaxis and DCA axes 1 and 2 were – 0.65 and 0.59.
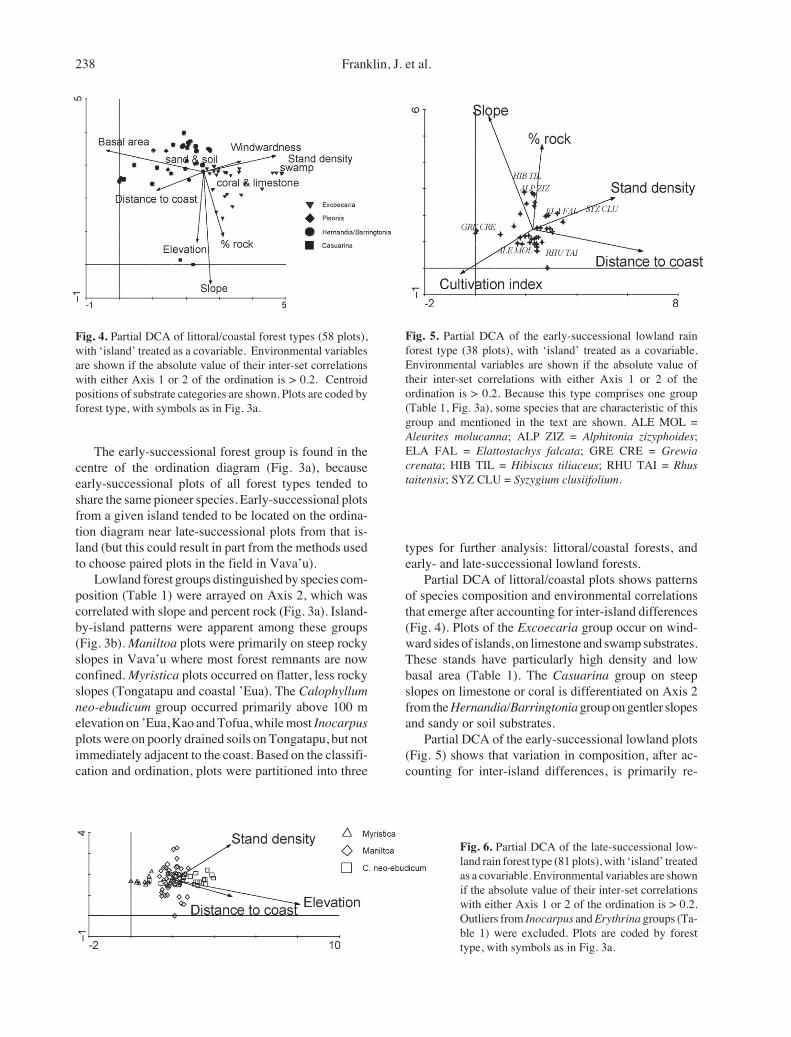
238 Franklin, J. et al.
The early-successional forest group is found in thecentre of the ordination diagram (Fig. 3a), becauseearly-successional plots of all forest types tended toshare the same pioneer species. Early-successional plotsfrom a given island tended to be located on the ordina-tion diagram near late-successional plots from that is-land (but this could result in part from the methods usedto choose paired plots in the field in Vava’u).
Lowland forest groups distinguished by species com-position (Table 1) were arrayed on Axis 2, which wascorrelated with slope and percent rock (Fig. 3a). Island-by-island patterns were apparent among these groups(Fig. 3b). Maniltoa plots were primarily on steep rockyslopes in Vava’u where most forest remnants are nowconfined. Myristica plots occurred on flatter, less rockyslopes (Tongatapu and coastal ’Eua). The Calophyllumneo-ebudicum group occurred primarily above 100 melevation on ’Eua, Kao and Tofua, while most Inocarpusplots were on poorly drained soils on Tongatapu, but notimmediately adjacent to the coast. Based on the classifi-cation and ordination, plots were partitioned into three
types for further analysis: littoral/coastal forests, andearly- and late-successional lowland forests.
Partial DCA of littoral/coastal plots shows patternsof species composition and environmental correlationsthat emerge after accounting for inter-island differences(Fig. 4). Plots of the Excoecaria group occur on wind-ward sides of islands, on limestone and swamp substrates.These stands have particularly high density and lowbasal area (Table 1). The Casuarina group on steepslopes on limestone or coral is differentiated on Axis 2from the Hernandia/Barringtonia group on gentler slopesand sandy or soil substrates.
Partial DCA of the early-successional lowland plots(Fig. 5) shows that variation in composition, after ac-counting for inter-island differences, is primarily re-
Fig. 4. Partial DCA of littoral/coastal forest types (58 plots),with ‘island’ treated as a covariable. Environmental variablesare shown if the absolute value of their inter-set correlationswith either Axis 1 or 2 of the ordination is > 0.2. Centroidpositions of substrate categories are shown. Plots are coded byforest type, with symbols as in Fig. 3a.
Fig. 5. Partial DCA of the early-successional lowland rainforest type (38 plots), with ‘island’ treated as a covariable.Environmental variables are shown if the absolute value oftheir inter-set correlations with either Axis 1 or 2 of theordination is > 0.2. Because this type comprises one group(Table 1, Fig. 3a), some species that are characteristic of thisgroup and mentioned in the text are shown. ALE MOL =Aleurites molucanna; ALP ZIZ = Alphitonia zizyphoides;ELA FAL = Elattostachys falcata; GRE CRE = Grewiacrenata; HIB TIL = Hibiscus tiliaceus; RHU TAI = Rhustaitensis; SYZ CLU = Syzygium clusiifolium.
Fig. 6. Partial DCA of the late-successional low-land rain forest type (81 plots), with ‘island’ treatedas a covariable. Environmental variables are shownif the absolute value of their inter-set correlationswith either Axis 1 or 2 of the ordination is > 0.2.Outliers from Inocarpus and Erythrina groups (Ta-ble 1) were excluded. Plots are coded by foresttype, with symbols as in Fig. 3a.

- Environment, disturbance history and rain forest composition across the islands of Tonga - 239
lated to proximity to the coast, disturbance by formercultivation, and slope steepness, but not substrate type(unlike littoral/coastal forest) or elevation (unlike late-successional forest). Those occurring inland on lessdisturbed sites are characterized by species such asElattostachys falcata and Syzygium clusiifolium. Plotson less disturbed sites but closer to the coast have morecoastal species (e.g. Hibiscus tiliaceus). The most dis-turbed sites also occur near the coast and are character-ized by Grewia crenata and Aleurites moluccana.
Partial DCA of late-successional lowland rain forestplots (Fig. 6) excluded the ten Inocarpus and Erythrinaplots (Table 1) which were extreme outliers in terms of
species composition. Axis 1 is positively related toelevation and distance from the coast and differentiatedthe Myristica (mean elevation = 70 m) and Maniltoa (61m) groups from the C. neo-ebudicum group, foundabove 100 m elevation on ’Eua, Kao and Tofua (Table1). The only Vava’u plot in this group was from one ofthe highest elevations in that island group (180 m). Thesecond axis appears to differentiate rockier Maniltoagroup plots, co-dominated by Garuga floribunda andPleiogynium timoriense, from the remainder of thisgroup, although rockiness was only weakly correlatedwith the second pDCA axis.
Variation partitioning showed that for all three for-est types, differences among islands unrelated to envi-ronment and anthropogenic disturbance account for simi-lar levels of compositional variation (23-28%). Forcoastal forests, most of the remaining compositionalvariation (71%) is explained by disturbance and envi-ronmental variables that do not covary with island. Thiscontrasts with both early and late-successional forestswhere there is a much higher degree of compositionalvariation reflecting covariation between disturbance/environment and island (Fig. 7). And indeed, differ-ences in environment among islands are primarily influ-encing inland, non-coastal areas. Plots from differentisland groups differ significantly in mean elevation (Ta-ble 2), reflecting the increasing maximum elevation ofthe islands from Tongatapu (80 m) to Kao (1046 m;App. 1). Differences in distance from the coast (Table 2)reflect the larger size of Tongatapu (App. 1) and thedistribution of forest remnants on different islands orisland groups (e.g. on small islands and on escarpmentsnear the coast in Vava’u). Islands also differ in slope androckiness (Table 2). This both reflects true differences,i.e. Tongatapu and Ha’apai are largely flat with little
Fig. 7. Relative percentage of compositional variation that canbe explained by island (or island group) alone, covariationbetween island and environment/disturbance and environ-ment/disturbance alone for the three major forest types ofTonga. Littoral/coastal forest: n = 58; Early-successional for-est: n = 38; Late-successional forest: n = 91.
Table 2. Values of environmental variables (described in text) that differ among plots representing the major islands or islandgroups (Table 1). (a) Mean ± SD values demonstrated by Scheffe’s multiple comparison test in a MANOVA to differsignificantly between islands; those not sharing the same letter differ at p < 0.01. (b) Percentage frequency of occurrences of plotsin different substrate classes. Summary statistics are shown for the islands of ’Euaiki and Ha’apai, but they were excluded fromthe MANOVA because of the small numbers of plots sampled on each.
Variable Tofua, Kao ’Eua Vava’u Tongatapu ’Euaiki Ha’apai(n = 6) (n = 40) (n = 84) (n = 52) (n = 3) (n = 2)
(a) Continuous variablesElevation (m) 248 ± 138 a 132 ± 85 b 43 ± 51 c 12 ± 9 c 25 ± 9 7 ± 2Slope (º) 14 ± 6 ab 15 ± 14 a 13 ± 10 a 1 ± 3 b 18 ± 13 1 ± 1Rockiness index — 0.87 ± 1.14 a 1.62 ± 1.05 b 0.25 ± 0.59 a 1.67 ± 0.58 0 ± 0Distance to Coast (m) 1232 ± 878 a 690 ± 836 a 177 ± 318 c 1364 ± 1320 b 162 ± 95 35 ± 21
(b) Substrate classesVolcanic 100 2.5 0 0 0 0Limestone 0 35 83 17 100 0Soil 0 50 0 67 0 0Coral 0 2.5 3 0 0 0Sand 0 10 14 0 0 100Swamp 0 0 0 15 0 0

240 Franklin, J. et al.
exposed rock, and that sampling in other areas focusedon less disturbed forests, which occur mainly on steeper,often rockier, slopes that are unsuitable for cultivation.
Island, including contrasts in environment and an-thropogenic disturbance, explains proportionally the mostvariation in late-successional forests (55%), followed byearly successional forests (51%) and the least in littoraland coastal forests (29%). To further understand thesource of these patterns we compared species that arewide-ranging across islands (≥ 5% frequency in 3-4 is-land groups; App. 3) with those that are indicators forislands (App. 4). A high proportion of littoral/coastal(Guettarda speciosa, Hibiscus tiliaceus, Pandanustectorius) and early-successional (Rhus taitensis, Morindacitrifolia, Grewia crenata) species are wide ranging rela-tive to lowland forest species (the most common beingElaeocarpus tonganus, Ellattostachys falcata, Ficusscabra, Pouteria grayana and Vavaea amicorum). Indi-cator species for Vava’u (Zanthoxylum pinnatum, Crypto-carya turbinata), Kao and Tofua (Psychotria insularum,Canarium vitiense), and ’Eua (Garcinia myrtifolia,Dysoxylum tongense; App. 4) are typically associatedwith late-successional forest (Table 1).
A comparison of eigenvalues of DCA and DCCAshowed that the variables used for variation partitioningaccounted for most of the explainable variation in early-and late-successional forests (ratio of DCA and DCCAaxis 1 and axis 2 eigenvalues ranges from 0.78 to 0.97),but less in coastal forests (ratio of DCA and DCCA axis1 and axis 2 are 0.70 and 0.54 respectively).
Discussion
Our synthesis of forest vegetation data for Tonga inWestern Polynesia both confirms the generality of com-positional patterns and their environmental correlatesnoted in previous studies (Whistler 1992; Drake et al.1996; Franklin et al. 1999; Wiser et al. 2002), andextends their interpretation with quantitative archipelago-wide analysis of the influence of environment, anthro-pogenic disturbance, and biogeography on forest com-position. Littoral and coastal forests had characteristicassemblages of widespread species known to be seawaterdispersed (Guppy 1906), such as Barringtonia asiatica,Calophyllum inophyllum, Excoecaria agallocha, Her-nandia nymphaeifolia, and Terminalia catappa.
Worldwide, disturbance strongly influences tropicalforest composition, typically favouring widely-distrib-uted species (Connell 1978; Whitmore 1985; Brown &Lugo 1990). Our results are consistent with this. Early-successional rain forest composition in Tonga varieswith a cultivation index but is broadly similar acrossenvironmental gradients, and throughout the region
(Straatmans 1964; Schmid 1975; Garnock-Jones 1978;Whistler 1980; Morat & Veillon 1985; Whitmore 1985;Webb & Fa’aumu 1999). Our data are also consistentwith the gradient-in-time model of succession that pre-dicts that early-successional species have broader nichesthan late-successional species (Peet 1992).
Late-successional lowland rain forest compositionvaried along environmental gradients, particularly el-evation. These patterns are again consistent with thegradient-in-time model where niche breadth should de-crease during succession (Peet 1992). Calophyllum neo-ebudicum forest was found at higher elevations thanMyristica and Maniltoa-dominated forests, although el-evations in Tonga do not extend high enough to supportmontane rain forest found elsewhere in Western Poly-nesia (Whistler 1980; Kirkpatrick & Hassall 1985; Ash1992; Keppel 2005; Tuiwawa 2005). The distribution ofC. neo-ebudicum forest seems to be determined byelevation rather than substrate because it was found onboth limestone and volcanic substrates.
There are also patterns of species composition re-lated to island that cannot be explained by environmen-tal and disturbance variables measured in this study. Wewere surprised that so much variation in early-succes-sional forest was explained by island and covariationbetween island and disturbance/environment (51%) giventhat many of the wide-ranging species (App. 3) are earlysuccessional. Because early-successional species tendto be widespread owing to dispersal ability and otherlife-history traits (e.g. long seed viability, Denslow 1996;Whitmore 1998; Gitay et al. 1999), we expected localand regional species pools (sensu Zobel 1997) to besimilar for the early-successional forests. However, al-though they were not significantly associated with aparticular island, Aleurites moluccana (a prehistoricintroduction), Bischofia javanica (a preserved tree) andPometia pinnata (commonly planted in villages andplantations) were important in early-successional foreston Tongatapu but infrequent elsewhere. This suggeststhat variation in species composition of secondary forestamong islands could be related to cultivation practicesand land-use history. Other gaps in the distributions ofwidespread, locally abundant early-successional nativespecies (Polyscias multijuga and Dendrocnide harveyifor example) cannot clearly be linked to cultivationhistory, however.
Differences in species composition among islandgroups were clearly seen among late-successional for-ests (e.g. Table 1, App. 4). This may be due tobiogeographic history (dispersal limitations on a re-gional scale), lower dispersal ability of late- (relative toearly-) successional species, and tighter links betweenspecies distributions and environment (where environ-mental gradients could not be characterized by our data)

- Environment, disturbance history and rain forest composition across the islands of Tonga - 241
over the course of succession. For example, Myristica-dominated forests occurred on ’Eua, and Tongatapu,and in Samoa (Whistler 1980) and the Lau Goup, Fiji(Garnock-Jones 1978), but not Vava’u. This patterncannot be explained by latitude or eastern range limits(Fig. 1). Myristica is dispersed by large pigeons (Guppy1906; Meehan et al. 2002), including several species ofwhich have been lost from Tonga since the arrival ofpeople (Steadman 1993, 1995), but also by Pacific pi-geons (Ducula pacifica) (Steadman & Freifeld 1999;McConkey et al. 2004; Meehan et al. 2005) which arefound throughout Tonga, but arrived after human colo-nization (Steadman 1997). Therefore, it is unknown ifthis distribution gap is due to human impacts on nativedispersers or to biogeographical factors (dispersal, colo-nization).
Other late-successional species found throughoutWestern Polynesia are common in one island group butuncommon or absent elsewhere in Tonga, e.g. Crypto-carya turbinata, Chionanthus vitiensis. Their absencefrom Tongatapu may be related to greater impacts ofanthropogenic disturbance there. Zanthoxylum pinnatumshows a low-latitude (Fiji, Samoa, Vava’u, Niuatoputapu,Fig. 1) distribution in Western Polynesia, consistentwith environmental (climatic) limits to its range, whileregional endemics such as Maniltoa grandiflora, Crypto-carya hornei and Pouteria membranacea are only foundin a portion of Western Polynesia, suggesting dispersallimitations.
Island biogeography theory (MacArthur & Wilson1967) suggests that a rare species could be absent froman island in an archipelago, especially a small or remoteone, because it never colonized, or did not maintain aviable population when island areas and proximitiesvaried as sea level fluctuated (Nunn 1994). When wide-spread and locally abundant species show gaps in distri-bution in a region with a long history of forest clearingfor agriculture, however, and when those gaps are unre-lated to island size or isolation, it may have been lost dueto anthropogenic disturbance. This is especially com-pelling when, as in this study, the islands with thehighest potential levels of natural disturbance, but low-est intensities of human land use, the active volcanoesTofua and Kao, support species not found elsewhere inTonga (App. 4), and, with the possible exception ofFagraea, they are not known to be particularly associ-ated with volcanic substrates. Detailed palynologicalstudies to elucidate long-term vegetation patterns wouldbe particularly illuminating (Fall 2005).
Our study furthers the qualitative synthesis of earlierwork on Tongan vegetation provided by Mueller-Dombois & Fosberg (1998). We note however that,while extensive, this data set on Tongan forests is stillfar from comprehensive, and in particular, mangroves,
forest stands dominated by single species, e.g. Erythrinafusca, Pandanus tectorius, Hibiscus tiliaceus, and eco-logically important (Janzen 1979; Terborgh 1986)banyans (Ficus obliqua, F. prolixa), have been ob-served by the authors but are not well represented.Further, the steep southeastern slopes of ’Eua haveforests dominated by the palm Pritchardia pacifica, aspecies we recorded on only a single plot on ’Euaiki.
Analyses of data from multiple surveys (cf. Knapp etal. 2004) spanning island groups allowed us to quantifyregional similarities among littoral/coastal forests (par-ticularly those related to seawater-dispersed species)and among early-successional forests whose speciesdistributions indicates broad environmental tolerances.Some among-island differences in secondary forest com-position involve preserved or planted species and seemto reflect differences in land-use practices, althoughthese differences may also result from covariation be-tween island and environment in our sample. In late-successional forests, the variation in species composi-tion among islands can be partly explained by envi-ronmental differences that exist among islands and hy-pothesized tight links between species and environ-ment, but in some cases may also result from a lack ofrepresentative sampling. Disentangling the effects ofanthropogenic disturbance history versus biogeographichistory on lowland rain forest in this region awaitsfurther study.
Acknowledgements. We thank W. A. Whistler for graciouslyproviding the data from Kao and Tofua. Data collection andanalysis were supported by the East-West Center, NationalGeographic Society, Georgia Southern University, San DiegoState University, University of Hawaii, New Zealand Founda-tion for Research, Science and Technology (ContractCO9X004), and the New Zealand Ministry of Foreign Affairsand Trade. We acknowledge the use of data from Tongatapudrawn from New Zealand’s National Vegetation SurveyDatabank (NVS). We are grateful to the government of Tonga,especially the Office of the Prime Minister, the Ministry ofAgriculture and Forestry, and the Ministry of Land and Sur-vey, and to numerous individuals, especially S. Fotu, N.Prescott, M. Halefihi, T. Hoponoa, D. Steadman, L. Bolick, D.Smith, T. Motley, T. Fine, K. McConkey, F. Tonga, M.Pomelile, C. Imada, Aunofo, Ongo, V. Latu, M. Havea, ’I.Kamoloni, T. Faka’osi, T. Savage and M. Breach. C. Bezar, A.Whistler, R. Ejrnæs and two anonymous reviewers improvedthe manuscript with their comments and corrections.

242 Franklin, J. et al.
References
Anon. 1999. S-Plus 2000 user’s guide. Data Analysis Prod-ucts Division, Mathsoft Inc., Seattle, WA, US.
Anon. (R Development Core Team) 2004. R: A language andenvironment for statistical computing. R Foundation forStatistical Computing, Vienna, Austria. http://www.R-project.org.
Ash, J. 1992. Vegetation ecology of Fiji: past, present andfuture perspectives. Pac. Sci. 46: 111-127.
Beers, T.W., Dress, P.E. & Wensel, L.C. 1966. Aspect trans-formations in site productivity research. J. For. 64: 691-692.
Bolick, L.A. 1995. Conservation and park development atMount Talau, Vava’u, Kingdom of Tonga. M.Sc. Thesis,San Diego State University, San Diego, CA, US.
Borcard, D., Legendre, P. & Drapeau, P. 1992. Partialling outthe spatial component of ecological variation. Ecology 73:1045-1055.
Brown, S. & Lugo, A.E. 1990. Tropical secondary forests. J.Trop. Ecol. 6: 1-32.
Burslem, D.F.R.P., Whitmore, T.C. & Brown, G.C. 2000.Short-term effects of cyclone impact and long-term recov-ery of tropical rain forest on Kolombangara, SolomonIslands. J. Ecol. 88: 1063-1078.
Carlquist, S. 1974. Island biology. Columbia University Press,New York, NY, US.
Carlquist, S. 1996. Plant dispersal and the origin of Pacificisland floras. In: Keast, A. & Miller, S.E. (eds.) The originand evolution of Pacific island biotas, New Guinea toEastern Polynesia: patterns and processes, pp. 153-164.SPB Academic Publishing, Amsterdam, The Netherlands.
Clarke, K.R. 1993. Non-parametric multivariate analyses ofchanges in community structure. J. Ecol. 18: 117-143.
Connell, J.H. 1978. Diversity in tropical rain forest and coralreefs. Science 199: 1302-1309.
Crowley, P.H. 1992. Resampling methods for computation-intensive data analysis in ecology and evolution. Annu.Rev. Ecol. Syst. 23: 405-447.
Denslow, J.S. 1996. Functional group diversity and responseto disturbance. In: Orians, G.H., Dirzo, R. & Cushman,J.H. (eds.) Biodiversity and ecosystem processes in tropi-cal forests, pp. 127-151. Springer-Verlag, Berlin, DE.
Dickinson, W.R., Burley, D.V. & Shutler Jr., R. 1999. Holocenepaleoshoreline record in Tonga: geomorphic features andarchaeological implications. J. Coast. Res. 15: 682-700.
Drake, D.R., Whistler, W.A., Motley, T.J. & Imada, C.T.1996. Rain forest vegetation of ’Eua Island, Kingdom ofTonga. N. Z. J. Bot. 34: 65-77.
Drake, D.R., Mulder, C.P.H., Towns, D.R. & Daugherty, C.H.2002. The biology of insularity: an introduction. J.Biogeogr. 29: 563-569.
Dufrêne, M. & Legendre, P. 1997. Species assemblages andindicator species: the need for a flexible asymmetricalapproach. Ecol. Monogr. 67: 345-366.
Elmqvist, T., Rainey, W.E., Pierson, E.D. & Cox, P.A. 1994.Effects of tropical cyclones Ofa and Val on the structure ofa Samoan lowland rain forest. Biotropica 26: 384-391.
Fall, P.L. 2005. Vegetation change in the coastal-lowland rain
forest at Avai’o’vuna Swamp, Vava’u, Kingdom of Tonga.Quat. Res. 64: 451-459.
Franklin, J. 2003. Regeneration and growth of pioneer andshade-tolerant rain forest trees in Tonga. N. Z. J. Bot. 41:669-684.
Franklin, J., Drake, D.R., Bolick, L.A., Smith, D.S. & Motley,T.J. 1999. Rain forest composition and patterns of second-ary succession in the Vava’u island group, Tonga. J. Veg.Sci. 10: 51-64.
Franklin, J., Drake, D.R., McConkey, K.R., Tonga, F. &Smith, L.B. 2004. The effects of Cyclone Waka on thestructure of lowland tropical rain forest in Vava’u, Tonga.J. Trop. Ecol. 20: 409-420.
Garnock-Jones, P.J. 1978. Plant communities on Lakeba andsouthern Vanua Balavu, Lau Group. Fiji. Bull. R. Soc. N.Z. 17: 95-117.
Gentry, A.H. 1988. Changes in plant community diversity andfloristic composition on environmental and geographicalgradients. Ann. Mo. Bot. Gard. 75: 1-34.
Gitay, H., Noble, I.R. & Connell, J.H. 1999. Deriving func-tional types for rain-forest trees. J. Veg. Sci. 10: 641-650.
Greig-Smith, P. 1983. Quantitative plant ecology. BlackwellScientific Publications, Oxford, UK.
Guppy, H.B. 1906. Observations of a naturalist in the Pacificbetween 1896 and 1899. Vol. II. Plant-dispersal.MacMillan, London, UK.
Harms, K.E., Wright, S.J., Caldera, O., Hernandez, A. &Herre, E.A. 2000. Pervasive density-dependent recruit-ment enhances seedling diversity in a tropical forest. Na-ture 404: 493-495.
Hill, M.O. 1979. TWINSPAN – A FORTRAN program forarranging multivariate data in a two-way table by classi-fication of the individuals and attributes. Cornell Univer-sity, Ithaca, NY, US.
Horn, S.P. & Kennedy, L.M. 2001. Pollen evidence of maizecultivation 2700 B.P. at La Selva Biological Station, CostaRica. Biotropica 33: 191-196.
Hughes, P.J., Hope, G., Latham, M. & Brookfield, H.C. 1979.Prehistoric man induced degradation of the Lakeba land-scape: evidence from two inland swamps. In: Brookfield,H.C. (ed.) Lakeba: resources, population and economy,pp. 93-110. ANU, Canberra, Australia.
Janzen, D.H. 1979. How to be a fig. Annu. Rev. Ecol. Syst. 10:13-51.
Keppel, G. 2005. Botanical studies within the PABITRA Wet-Zone Transect, Viti Levu, Fiji. Pac. Sci. 59: 165-174.
Keppel, G., Navuso, J.C., Naikatini, A., Thomas, N.T., Rounds,I.A., Osborne, T.A., Batinamu, N. & Senivasa, E. 2005.Botanical diversity at Savura, a lowland rain forest sitealong the PABITRA Gateway Transect, Viti Levu, Fiji.Pac. Sci. 59: 175-191.
Kirch, P.V. & Ellison, J. 1994. Paleoenvironmental evidencefor human colonization of remote Oceanic islands. Antiq-uity 68: 310-321.
Kirkpatrick, J.B. & Hassall, D.C. 1985. The vegetation andflora along an altitudinal transect through tropical forest atMount Korobaba, Fiji. N. Z. J. Bot. 23: 33-46.
Knapp, A.K., Smith, M.S., Collins, S.L., Zambatis, N., Peel,M., Emery, S., Wojdak, J., Horner-Devine, M.C., Biggs,

- Environment, disturbance history and rain forest composition across the islands of Tonga - 243
H., Kruger, J. & Andelman, S.J. 2004. Generality inecology: testing North American grassland rules in SouthAfrican savannas. Front. Ecol. Environ. 9: 483-491.
MacArthur, R.H. & Wilson, E.O. 1967. The theory of islandbiogeography. Princeton University Press, Princeton, NJ,US.
McConkey, K.R., Meehan, H.J. & Drake, D.R. 2004. Seeddispersal by Pacific Pigeons (Ducula pacifica) in Tonga,Western Polynesia. Emu 104: 369-376.
McCune, B. & Mefford, M.J. 1999. PC-ORD. Multivariateanalysis of ecological data, Version 4. MjM SoftwareDesign, Glendenen Beach, OR, US.
Meehan, H.J., McConkey, K.R. & Drake, D.R. 2002. Potentialdisruptions to seed dispersal mutualisms in Tonga, west-ern Polynesia. J. Biogeogr. 29: 695-712.
Meehan, H.J., McConkey, K.R. & Drake, D.R. 2005. Earlyfate of Myristica hypergyraea seeds dispersed by Duculapacifica in Tonga, Western Polynesia. Austral Ecol. 30:374-382.
Mielke Jr., P.W. 1984. Meteorological applications of permu-tation techniques based on distance functions. In:Krishnaiah, P.R. & Sen, P.K. (eds.) Handbook of statis-tics, pp. 813-830. Elsevier Science Publishing, Amster-dam, NL.
Morat, P. & Veillon, J.-M. 1985. Contribution a la connaissancede la végétation et de la flore de Wallis et Futuna. Adanso-nia 3: 259-329.
Mueller-Dombois, D. 2002. Forest vegetation across the tropi-cal Pacific: a biogeographically complex region with manyanalogous environments. Plant Ecol. 163: 155-176.
Mueller-Dombois, D. & Fosberg, F.R. 1998. Vegetation of thetropical Pacific islands. Springer-Verlag, Berlin, DE.
Nunn, P.D. 1994. Oceanic islands. Blackwell, Oxford, UK.Økland, R.H. 1996. Are ordination and constrained ordination
alternative or complementary strategies in general eco-logical studies? J. Veg. Sci. 7: 289-292.
Økland, R.H. 1999. On the variation explained by ordinationand constrained ordination axes. J. Veg. Sci. 10: 131-136.
Økland, R.H. 2003. Partitioning the variation in a plot-by-species data matrix that is related to n sets of explanatoryvariables. J. Veg. Sci. 14: 693-700.
Økland, R.H. & Eilertsen, O. 1994. Canonical correspondenceanalysis with variation partitioning: some comments andan application. J. Veg. Sci. 5: 117-126.
Park, G. & Whistler, W.A. 2001. The terrestrial ecology andbotany of Tofua and Kao Islands in Ha’apai, Kingdom ofTonga: a survey of biodiversity conservation. Report SouthPacific Regional Environmental Programme, Apia, SM.
Peet, R.K. 1992. Community structure and ecosystem func-tion. In: Glenn-Lewin, D., Peet, R.K. & Veblen, T.T.(eds.) Plant succession: theory and prediction, pp. 103-151. Chapman and Hall, London, UK.
Pyke, C.R., Condit, R., Aguilar, S. & Lao, S. 2001. Floristiccomposition across a climatic gradient in a neotropicallowland forest. J. Veg. Sci. 12: 553-566.
Raven, P.H. & Axelrod, D.I. 1972. Plate tectonics and Aus-tralasian biogeography. Science 176: 1379-1386.
Ridley, H.N. 1930. The dispersal of plants throughout theworld. Reeve, Ashford, Kent, UK.
Rydgren, K., Økland, R.H. & Økland, T. 2003. Species re-sponse curves along environmental gradients. A case studyfrom SE Norwegian swamp forests. J. Veg. Sci. 14: 869-880.
Schmid, P.M. 1975. La flore et la végétation de la partieméridionale de l’Archipel des Nouvelles Hébrides. Phil.Trans. R. Soc. Lond. Ser. B Biol. Sci. 272: 329-342.
Smith, A.C. 1979, 1981, 1985, 1988, 1991. Flora vitiensisnova: a new flora of Fiji. Pacific Tropical Botanical Gar-den, Lawai, HI, US.
Steadman, D.W. 1993. Biogeography of Tongan birds beforeand after human impact. Proc. Natl. Acad. Sci. USA 90:818-822.
Steadman, D.W. 1995. Prehistoric extinctions of Pacific Is-land birds: biodiversity meets zooarchaeology. Science267: 1123-1131.
Steadman, D.W. 1997. The historic biogeography and com-munity ecology of Polynesian pigeons and doves. J.Biogeogr. 24: 737-753.
Steadman, D.W. & Freifeld, H.B. 1999. The food habits ofPolynesian pigeons and doves: a systematic and biogeo-graphic review. Ecotropica 5: 13-33.
Steadman, D.W., Franklin, J., Drake, D.R., Freifeld, H.B.,Bolick, L.A., Smith, D.S. & Motley, T.J. 1999. Conserva-tion status of forests and vertebrate communities in theVava’u Island Group, Tonga. Pac. Conserv. Biol. 5: 191-207.
Stoddart, D.R. 1992. Biogeography of the tropical Pacific.Pac. Sci. 46: 276-293.
Straatmans, W. 1964. Dynamics of some Pacific island forestcommunities in relation to the survival of the endemicflora. Micronesica 1: 113-122.
Svenning, J.-C., Kinner, D.A., Stallard, R.F., Engelbrecht,B.M.J. & Wright, S.J. 2004. Ecological determinism inplant community structure across a tropical forest land-scape. Ecology 85: 2526-2538.
Sykes, W.R. 1981. The vegetation of Late, Tonga. Allertonia2: 323-353.
Tappin, D.R. 1993. The Tongan frontal arc basin. In: Balance,P.F. (ed.) South Pacific sedimentary basins, pp. 157-176.Elsevier, Amsterdam, NL.
ter Braak, C. 1995. Ordination. In: Jongman, R., ter Braak, C.& van Tongeren, O. (eds.) Data analysis in communityand landscape ecology, pp. 91-173. Cambridge Univer-sity Press, Cambridge, UK.
ter Braak, C.J.F. 1987. The analysis of vegetation-environ-ment relationships by canonical correspondence analysis.Vegetatio 69: 69-77.
ter Braak, C.J.F. & Prentice, I.C. 1988. A theory of gradientanalysis. Adv. Ecol. Res. 18: 271-317.
ter Braak, C.J.F. & Smilauer, P. 1998. CANOCO Referencemanual and user’s guide to Canoco for Windows: Soft-ware for canonical community ordination (version 4).Microcomputer Power, Ithaca, NY, US.
Terborgh, J. 1986. Keystone plant resources in the tropicalforest. In: Soulé, M.E. (ed.) Conservation biology: thescience of scarcity and diversity, pp. 330-344. SinauerAssociates, Sunderland, MA, US.
Tuiwawa, M. 2005. Recent changes in the upland watershed
‹

244 Franklin, J. et al.
forest of Monasavu, a cloud forest site along the PABITRAGateway Transect on Viti Levu, Fiji. Pac. Sci. 59: 159-163.
Tuomisto, H., Poulsen, A.D., Ruokolainen, K., Moran, R.C.,Quintana, C., Celi, J. & Ca’as, G. 2003. Linking floristicpatterns with soil heterogeneity and satellite imagery inEcuadorian Amazonia. Ecol. Appl. 13: 352-371.
Turner, H., Hovenkamp, P. & van Welzen, P.C. 2001. Bioge-ography of Southeast Asia and the West Pacific. J.Biogeogr. 28: 217-230.
van Balgooy, M.M.J., Hovenkamp, P.H. & van Welzen, P.C.1996. Phytogeography of the Pacific – floristic and his-torical distribution patterns in plants. In: Keast, A. &Miller, S.E. (eds.) The origin and evolution of Pacificisland biotas, New Guinea to Eastern Polynesia: Patternsand processes, pp. 191-213. SPB Academic Publishing,Amsterdam, NL.
van Gemerden, V.S., Olff, H., Parren, M.P.E. & Bongers, F.2003. The pristine rain forest? Remnants of historicalhuman impacts on current tree species composition anddiversity. J. Biogeogr. 30: 1381-1390.
Vitousek, P.M., Loope, L.L. & Adsersen, H. (eds.) 1995.Islands: biological diversity and ecosystem function.Springer-Verlag, New York, NY, US.
Wagner, W.L., Herbst, D.R. & Sohmer, S.H. 1999. Manual ofthe flowering plants of Hawai’i, revised ed. 2 vols. BerniceP. Bishop Museum and University of Hawaii Press, Hono-lulu, HI, US.
Webb, E.L. & Fa’aumu, S. 1999. Diversity and structure oftropical rain forest of Tutuila, American Samoa: effects ofsite age and substrate. Plant Ecol. 144: 257-274.
Whistler, W.A. 1980. The vegetation of eastern Samoa.Allertonia 2: 45-190.
Whistler, W.A. 1991. The ethnobotany of Tonga: the plants,their Tongan names, and their uses. Bishop Mus. Bull. Bot.2: 1-155.
Whistler, W.A. 1992. Vegetation of Samoa and Tonga. Pac.Sci. 46: 159-178.
Whitmore, T.C. 1985. Tropical rain forests of the Far East.Clarendon Press, Oxford, UK.
Whitmore, T.C. 1989. Changes over twenty-one years in theKolombangara rain forests. J. Ecol. 77: 469-483.
Whitmore, T.C. 1998. An introduction to tropical rain forests.2nd. ed. Oxford University Press, Oxford, UK.
Whittaker, R.J. 1998. Island biogeography: ecology, evolu-tion, and conservation. Oxford University Press, Oxford,UK.
Wiser, S.K., Drake, D.R., Burrows, L.E. & Sykes, W.R. 2002.The potential for long-term persistence of forest frag-ments on Tongatapu, a large island in western Polynesia.J. Biogeogr. 29: 767-787.
Woodroffe, C.D. 1984. The impact of cyclone Isaac on thecoast of Tonga. Pac. Sci. 37: 181-210.
Yuncker, T.G. 1959. Plants of Tonga. Bishop Mus. Bull. 220:1-283.
Zobel, M. 1997. The relative role of species pools in determin-ing plant species richness: an alternative explanation ofspecies coexistence? Trends Ecol. Evol. 12: 266-269.
Received 24 August 2005;Accepted 15 December 2005.
Co-ordinating Editor: R. Ejrnæs.
For App. 1, see JVS/AVS Electronic Archives;www.opuluspress.se/

Ap
p. 1. Internet supplem
ent to: Franklin, J.; W
iser, S.K
.; Drake, D
.R.; B
urrows, L.E
. & S
ykes, W.R
. 2006.E
nvironment, disturbance history and rain forest com
position across the islands of Tonga, W
estern Polynesia.
J. Veg. S
ci. 17: 233-244.
App. 1. D
ata sets used in this study. Group =
Tonga’s m
ain island groups (Fig. 1). Plots = no. of plots in each dataset and Spp. =
total no. of trees and woody species (>
5 cm D
BH
)recorded in that group of plots including cultivated species. D
ate = year of survey. L
at(itude) and Long(itude) are approxim
ate locations of center of island group or island sampled. #Isl
= num
ber of islands sampled; A
rea is of island(s) sampled; E
lev is maxim
um elevation sam
pled (maxim
um elevation of island(s) in parentheses). Sources for island size, elevation and
location: Steadman et al. (1999), D
rake et al. (1996), Park & W
histler (2001), Wiser et al. (2002), topographic m
aps.
Group
Island(s)D
escriptionPlots
Spp.D
ateSource
Lat
Long
# IslA
rea (km2)
Elev (m
)
Tongatapu
Tongatapu
Forest fragments
5273
1997W
iser et al. 200221°10°S
175°10° W1
26130 (80)
Tongatapu
‘Euaiki
Forest fragments &
coastal forest3
391997
Wiser et al. (unpubl.)
21°05°S174°50° W
11
30 (40)T
ongatapu‘E
uaU
ndisturbed forest40
861990
Drake et al. 1996
21°25°S174°55° W
181
280 (312)V
ava’uV
ava‘u groupForest fragm
ents64
771993,1995
Bolick 1995;
18°40°S174°02” W
13119
180 (200)Franklin et al. 1999
Vava’u
Vava‘u group
Coastal forest
2050
1995Steadm
an et al. 1999“
““
“7
Ha’apai
Eastern, low
, limestone
Forest fragments
220
2004Franklin (unpubl.)
20°00°S174°30° W
217
8 (60)H
a’apaiK
ao, volcanicU
ndisturbed forest1
171997
Park & W
histler 200119°40°S
174°00° W1
13250 (1046)
Ha’apai
Tofua, volcanic
Undisturbed forest
535
““
19°45°S174°05° W
147
400 (558)T
otal187
140
App. 2-4. Internet supplement to:Franklin, J.; Wiser, S.K.; Drake, D.R.; Burrows, L.E. & Sykes, W.R. 2006.Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia.J. Veg. Sci. 17: 233-244.
2
App. 2. Species list. A total of 140 taxa were used in the analysis including six cultivated species used in the cultivation index. Somespecies could not be distinguished in the field as the plants lacked flowers or fruits, and were grouped for analysis. Geniostomarupestre and G. insulare were grouped as G. rupestre. Ficus obliqua and F. prolixa were grouped as ‘Banyan’. Two species in twoplots from Vava’u remain unidentified and were included as unknown but separate taxa in the analysis (‘Pouteria species unknown’and ‘Sapotaceae genus unknown’), and one species from Kao and Tofua is tentatively identified (Celtis harperi?). Tong1 = Wiseret al. (2002) and Wiser et al. unpubl. for ’Euaiki; Vav2 = Franklin et al. (1999); Vav3 = Steadman et al. (1999); Eua4 = Drake et al.(1996); KaoTofua5 = Park & Whistler (2001); EHap6 = JF unpubl. * This is really a Syzygium species but the combination in thatgenus has not yet been made from Eugenia, the genus in which most of these Pacific island trees were once treated.
Family Species name Species authority Tong Vav Vav Eua Kao EHap1 2 3 4 Tofua 5 6
Mimosaceae Acacia simplex (Sparr.) Pedley X XMimosaceae Adenanthera pavonina L. X XMeliaceae Aglaia heterotricha A. C. Sm. XRubiaceae Aidia racemosa (Cav.) D. D. Tirveng. X XEuphorbiaceae Aleurites molucanna (L.) Willd. X XRhamnaceae Alphitonia zizyphoides (Spreng.) A. Gray X X X X X XOlacaceae Anacolosa lutea Gillespie XAnnonaceae Annona muricata L. XRubiaceae Antirhea inconspicua (Seem.) Christoph. XMoraceae Artocarpus altilis (Parkinson) Fosberg X XBarringtoniaceae Barringtonia asiatica (L.) Kurz X XEuphorbiaceae Bischofia javanica Bl. X X XSapotaceae Burckella richii (A. Gray) Lam X XClusiaceae Calophyllum inophyllum L. X X XClusiaceae Calophyllum neo-ebudicum Guillaumin X X XAnnonaceae Cananga odorata (Lam.) Hooker f. & Thomps. XBurseraceae Canarium harveyi Seem. X XBurseraceae Canarium vitiense A. Gray XCaricaceae Carica papaya L. X XCasuarinaceae Casuarina equisetifolia L. X X X X XUlmaceae Celtis harperi? Horne XApocynaceae Cerbera odollam Gaertn. X X XOleaceae Chionanthus vitiensis (Seem.) A. C. Sm. X XRutaceae Citrus grandis (L.) Osbeck XRutaceae Citrus maxima (Burm.) Merr. XRutaceae Citrus reticulata L. XIcacinaceae Citronella samoensis (A. Gray) Howard X XArecaceae Cocos nucifera L. X X X X X XRubiaceae Coffea liberica Bull. XBoraginaceae Cordia subcordata Lam. X X XAgavaceae Cordyline fruticosa (L.) A. Chev. X X X XLauraceae Cryptocarya hornei Gillespie X XLauraceae Cryptocarya turbinata Gillespie X XCyatheaceae Cyathea lunulata (G.Forst.) Copel. XRubiaceae Cyclophyllum barbatum (G.Forst.) Hallé & Florence X X X X XCycadaceae Cycas rumphii Miq. X X X XUrticaceae Dendrocnide harveyi (Seem.) Chew XFabaceae Desmodium heterocarpon (L.) DC. XEbenaceae Diospyros elliptica (J. R. & G. Forst.) P. S. Green X X X X XEbenaceae Diospyros major (Forst. f.) Bakh. XEbenaceae Diospyros samoensis A. Gray X X X XEuphorbiaceae Drypetes vitiensis Croizat X XMeliaceae Dysoxylum forsteri (Juss.) C. DC. X X XMeliaceae Dysoxylum tongense A. C. Sm. XElaeocarpaceae Elaeocarpus graeffei Seem. XElaeocarpaceae Elaeocarpus tonganus Burkill X X X XSapindaceae Elattostachys falcata (A. Gray) Radlk. X X X X XApocynaceae Ervatamia orientalis Markgr. X X XFabaceae Erythrina fusca Lour. XFabaceae Erythrina variegata L. XMyrtaceae Eugenia reinwardtiana (Bl.) Bl. XEuphorbiaceae Excoecaria agallocha L. X X X XLoganiaceae Fagraea berteroana A. Gray ex Benth. X X

3
App. 2-4. Internet supplement to:Franklin, J.; Wiser, S.K.; Drake, D.R.; Burrows, L.E. & Sykes, W.R. 2006.Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia.J. Veg. Sci. 17: 233-244.
Moraceae Ficus obliqua G. Forst. X X X XMoraceae Ficus prolixa G. Forst. XMoraceae Ficus scabra G. Forst. X X X X XMoraceae Ficus tinctoria G. Forst. X X X XBurseraceae Garuga floribunda Decne. XClusiaceae Garcinia myrtifolia A. C. Sm. X XLoganiaceae Geniostoma rupestre J. R. & G. Forst. X X X X X XEuphorbiaceae Glochidion ramiflorum G. Forst. X X X X XTiliaceae Grewia crenata (J. R. & G. Forst.) Schinz & Guillaumin X X X X X XRubiaceae Guettarda speciosa L. X X X X XSapindaceae Guioa lentiscifolia Cav. X XGyrocarpaceae Gyrocarpus americanus Jacq. XSapindaceae Harpullia arborea (Blanco) Radlk. XMonimiaceae Hedycarya dorstenioides A. Gray XSterculiaceae Heritiera littoralis Ait. XSterculiaceae Heritiera ornithocephala Kostermans XHernandiaceae Hernandia moerenhoutiana Guill. XHernandiaceae Hernandia nymphaeifolia (Presl) Kubitzki X X XMalvaceae Hibiscus tiliaceus L. X X X X X XFlacourtiaceae Homalium whitmeeanum H. St. John X XFabaceae Inocarpus fagifer (Parkinson) Fosberg X XRubiaceae Ixora calcicola A. C. Sm. X X XVerbenaceae Lantana camara L. XFabaceae Leucaena leucocophala (Lam.) de Wit X XLauraceae Litsea mellifera A. C. Sm. XEuphorbiaceae Macaranga harveyana (Muell. Arg.) Muell. Arg. X X X XAnacardiaceae Mangifera indica L. X XSapotaceae Manilkara dissecta (L. f.) Dubard XCaesalpiniaceae Maniltoa grandiflora (A. Gray) Scheff. X X XRutaceae Melicope retusa A. Gray XMelastomataceae Memecylon vitiense A. Gray X XAraliaceae Meryta macrophylla (Rich ex A. Gray) Harms X X X XRutaceae Micromelum minutum (G. Forst.) Seem. X X X X XRubiaceae Morinda citrifolia L. X X X X X XRubiaceae Mussaenda raiateensis J. W. Moore XMyristicaceae Myristica hypargyraea A. Gray X X XApocynaceae Neisosperma oppositifolium (Lam.) Fosberg & Sachet X X X X XRubiaceae Neonauclea forsteri (Seem. ex Havil.) Merr. X XApocynaceae Ochrosia vitiensis (Markgr.) Pichon X XPandanaceae Pandanus tectorius Parkinson X X X X X XLythraceae Pemphis acidula J. R. & G. Forst. XThymelaeaceae Phaleria disperma (G.Forst.) Baill. X XMyrtaceae Pimenta dioica (L.) Merr. XNyctaginaceae Pisonia grandis R. Br. X X X XNyctaginaceae Pisonia umbellifera (G. Forst.) Seem. XPittosporaceae Pittosporum arborescens Rich ex A. Gray X X X X X XSapotaceae Pouteria garberi (Christoph.) Baehni XSapotaceae Pouteria grayana (H. St. John) Fosberg X X X X X XSapotaceae Pouteria membranacea (Lam) Baehni XSapotaceae Pouteria species unknown XAnacardiaceae Pleiogynium timoriense (DC.) Leenh. X X X XPodocarpaceae Podocarpus pallidus N. E. Gray X XAraliaceae Polyscias multijuga (A. Gray) Harms X XSapindaceae Pometia pinnata J. R. & G. Forst. X XRubiaceae Porterandia crosbyi (Burk.) A. C. Sm. & S. Darwin XVerbenaceae Premna serratifolia L. X XArecaceae Pritchardia pacifica Seem. & H. Wendl. X XMyrtaceae Psidium guajava L. XRubiaceae Psychotria carnea (G.Forst.) A. C. Sm. XRubiaceae Psychotria insularum A. Gray XAnacardiaceae Rhus taitensis Guill. X X X X XSantalaceae Santalum yasi Seem. X X X
Family Species name Species authority Tong Vav Vav Eua Kao EHap1 2 3 4 Tofua 5 6
App. 2, cont.
App. 2-4. Internet supplement to:Franklin, J.; Wiser, S.K.; Drake, D.R.; Burrows, L.E. & Sykes, W.R. 2006.Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia.J. Veg. Sci. 17: 233-244.
4
Sapindaceae Sapindus vitiensis A. Gray X X XGoodeniaceae Scaevola sericea Vahl XMimosaceae Schleinitzia insularum (Guill.) Burkhart X XAnacardiaceae Semecarpus vitiensis (A. Gray) Engl. XMimosaceae Serianthes melanesica Fosberg XSolanaceae Solanum mauritianum Scop. XFabaceae Sophora tomentosa L. XMoraceae Streblus anthropophagorum (Seem). Corner XMyrtaceae Syzygium brackenridgei (A. Gray) C. Muell. X XMyrtaceae Syzygium clusiifolium (A. Gray) C. Muell. X X XMyrtaceae Eugenia crosbyi* Burk. X XMyrtaceae Syzygium dealatum (Burk.) A. C. Sm. XMyrtaceae Syzygium richii (A. Gray) Merr. & Perry X XRubiaceae Tarenna sambucina (G.Forst.) Durand ex Drake X X X X XCombretaceae Terminalia catappa L. X X XCombretaceae Terminalia litoralis Seem. X X X XMalvaceae Thespesia populnea (L.) Sol. ex Correa X X X XBoraginaceae Tournefortia argentea (L. f.) Heine X X XUlmaceae Trema cannabina Lour. X XSapotaceae Genus unknown XMeliaceae Vavaea amicorum Benth. X X X X XVerbenaceae Vitex trifolia L. XOlacaceae Ximenia americana L. XMeliaceae Xylocarpus granatum Koenig XMeliaceae Xylocarpus moluccensis (Lam.) M. Roem. X XFlacourtiaceae Xylosma simulans A.C. Sm. X X X XRutaceae Zanthoxylum pinnatum (J. R. & G. Forst.) W. R.B. Oliver X X
Family Species name Species authority Tong Vav Vav Eua Kao EHap1 2 3 4 Tofua 5 6
App. 2, cont.
5
App. 2-4. Internet supplement to:Franklin, J.; Wiser, S.K.; Drake, D.R.; Burrows, L.E. & Sykes, W.R. 2006.Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia.J. Veg. Sci. 17: 233-244.
App. 3. Common (wide-ranging) species associated with forest types, frequency (% of plots in which species occurred in an islandgroup) shown for island groups with > 5 plots. * Geniostoma rupestre and G. insulare could not be distinguished in the field, and weregrouped as G. rupestre (see App. 1). Ton = Tongatapu; Vav = Vava’u group; Eua = ’Eua; KT = Kao and Tofua (no. of plots inparentheses). Ha’apai and ’Euaiki were excluded because of the small number of plots.
Species name Ton (55) Vav (84) Eua (40) KT (6)
Littoral/Coastal Guettarda speciosa 5 23 13 17Hibiscus tiliaceus 40 35 20 17Neisosperma oppositifolium 29 21 28 0Pandanus tectorius 31 37 10 17Terminalia litoralis 9 11 5 0
Early successional Alphitonia zizyphoides 4 52 20 67Cocos nucifera 35 46 5 17Geniostoma rupestre* 11 32 3 83Glochidion ramiflorum 2 14 8 33Grewia crenata 51 35 18 17Macaranga harveyana 15 5 8 17Morinda citrifolia 51 38 10 17Rhus taitensis 44 39 18 50
Late successional Diospyros elliptica 9 30 0 17Elaeocarpus tonganus 22 25 18 100Elattostachys falcata 31 60 50 100Ficus scabra 20 19 25 50Micromelum minutum 16 44 5 0Pittosporum arborescens 7 24 3 17Pouteria grayana 16 57 25 17Vavaea amicorum 40 55 25 50Xylosma simulans 33 57 38 0
App. 4. Importance value (IV, average of relative abundance and relative frequency) for species that are indicators for island groups.*Found only in plots on the associated island group.
Island group Species name IV p
Vava’u Zanthoxylum pinnatum* 57 0.026Cryptocarya turbinata 45 0.033
’Eua Garcinia myrtifolia 45 0.013Dysoxylum tongense* 40 0.030Citronella samoensis 38 0.036
Kao, Tofua Psychotria insularum* 100 0.001Canarium vitiense* 83 0.001Heritiera ornithocephala* 83 0.001Neonauclea forsteri 49 0.005Fagraea berteroana 40 0.035Melicope retusa* 33 0.027Celtis cf. harperi* 17 0.035
Related Documents











