6SULQJHU is collaborating with JSTOR to digitize, preserve and extend access to 0LFURELDO (FRORJ\ http://www.jstor.org (QULFKPHQW RI *HREDFWHU 6SHFLHV LQ 5HVSRQVH WR 6WLPXODWLRQ RI )H,,, 5HGXFWLRQ LQ 6DQG\ $TXLIHU 6HGLPHQWV $XWKRUV 2 / 6QRH\HQERV:HVW . 3 1HYLQ 5 7 $QGHUVRQ DQG ' 5 /RYOH\ 6RXUFH 0LFURELDO (FRORJ\ 9RO 1R )HE SS 3XEOLVKHG E\ 6SULQJHU 6WDEOH 85/ KWWSZZZMVWRURUJVWDEOH $FFHVVHG 87& Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/ info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTC All use subject to JSTOR Terms and Conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

is collaborating with JSTOR to digitize, preserve and extend access to
http://www.jstor.org
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/ info/about/policies/terms.jsp
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected].
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

ECOLOGY Microb Ecol (2000) 39:153-167
DOI: 10.1007/s002480000018 ? 2000 Springer-Verlag New York Inc.
Enrichment of Geobacter Species in Response to Stimulation of Fe(III) Reduction in Sandy Aquifer Sediments
O.L. Snoeyenbos-West, K.P. Nevin, R.T. Anderson, D.R. Lovley
Department of Microbiology, University of Massachusetts, Amherst, MA 01003, USA
Received: 19 October 1999; Accepted: 28 December 1999; Online Publication: 25 April 2000
|A B S T R A C T
Engineered stimulation of Fe(III) has been proposed as a strategy to enhance the immobilization of radioactive and toxic metals in metal-contaminated subsurface environments. Therefore, laboratory and field studies were conducted to determine which microbial populations would respond to stimulation of Fe(III) reduction in the sediments of sandy aquifers. In laboratory studies, the addition of either various organic electron donors or electron shuttle compounds stimulated Fe(III) reduction and resulted in Geobacter sequences becoming important constituents of the Bacterial 16S rDNA sequences that could be detected with PCR amplification and denaturing gradient gel elec- trophoresis (DGGE). Quantification of Geobacteraceae sequences with a PCR most-probable- number technique indicated that the extent to which numbers of Geobacter increased was related to the degree of stimulation of Fe(III) reduction. Geothrix species were also enriched in some instances, but were orders of magnitude less numerous than Geobacter species. Shewanella species were not detected, even when organic compounds known to be electron donors for Shewanella species were used to stimulate Fe(III) reduction in the sediments. Geobacter species were also enriched in two field experiments in which Fe(III) reduction was stimulated with the addition of benzoate or aromatic hydrocarbons. The apparent growth of Geobacter species concurrent with increased Fe(III) reduction suggests that Geobacter species were responsible for much of the Fe(1II) reduction in all of the stimulation approaches evaluated in three geographically distinct aquifers. Therefore, strat- egies for subsurface remediation that involve enhancing the activity of indigenous Fe(III)-reducing populations in aquifers should consider the physiological properties of Geobacter species in their treatment design.
Introduction
Engineered stimulation of dissimilatory metal reduction may be a useful method for immobilizing contaminant met-
Correspondence to: D.R. Lovley; Fax: (413) 545-1578; E-mail: dlovley@ microbio.umass.edu
als in subsurface environments [22]. Several metals and met- alloids, including uranium, technetium, chromium, cobalt, and selenate, are more soluble, and thus mobile, in the oxi- dized state and tend to precipitate in the reduced state [22]. For example, many microorganisms capable of dissimilatory metal reduction can reduce soluble U(VI) to insoluble U(IV) [25, 36], and stimulation of microbial U(VI) reduction un-
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

154 O.L. Snoeyenbos-West et al.
der laboratory conditions has been shown to precipitate ura- nium from contaminated groundwaters [34].
Fe(III) is generally the most abundant possible electron acceptor for dissimilatory metal-reducing microorganisms in soils and sediments [23, 25]. Most Fe(JII)-reducing mi- croorganisms can also reduce Mn(IV), which is typically present at concentrations about 10% of those of Fe(III) [23, 25]. Fe(III) and Mn(IV) are abundant in many subsurface soils and are likely to be in higher concentrations than con- taminant metals, even under conditions of severe contami- nation. Therefore, stimulation of dissimilatory metal reduc- tion to immobilize metal contaminants also involves micro- bial reduction of the Fe(III) and Mn(IV) in the soils. The reduction of Fe(III) and Mn(IV) along with contaminant metals may be beneficial because the Fe(II) and Mn(II) pro- duced can serve as a redox buffer to help prevent reoxidation of the reduced contaminant metals. Furthermore, since most of the dissimilatory Fe(III)- and Mn(IV)-reducing microor- ganisms that have been evaluated are capable of reducing contaminant metals [24, 25, 29], stimulation of Fe(III) and Mn(IV) reduction may increase the population size of mi- croorganisms that can also reduce contaminant metals. Such stimulation may thus enhance rates of contaminant metal reduction.
In order to understand how to most effectively accelerate dissimilatory metal reduction in subsurface environments, it is important to know which microbial populations are pro- moted when dissimilatory metal reduction is artificially stimulated. A wide phylogenetic diversity of Bacteria [24] and Archaea [53] are capable of dissimilatory Fe(III) reduc- tion. It is conceivable that the growth and activity of any of these known Fe(III) reducers, or as yet undescribed species, might be enhanced when Fe(III) reduction is stimulated in contaminated subsurface environments. Which dissimila- tory metal reducers become predominant might be affected by such factors as the physiological properties of the Fe(III)- reducing microorganisms, the geochemistry of the environ- ment, the composition of the microbial community prior to stimulation, and the method used to stimulate metal reduc- tion.
Three mechanisms for stimulating Fe(III) reduction in sedimentary environments have been identified. Previous studies have demonstrated that addition of known electron donors for Fe(III) reducers can stimulate Fe(III) reduction in aquatic sediments [33], and this is also a likely strategy for stimulating Fe(III) reduction in organic-poor subsurface en- vironments. The addition of electron shuttling organic com- pounds such as humic acids or other extracellular quinones
[30, 31] can also stimulate Fe(III) reduction in sediments [46]. These compounds can greatly accelerate Fe(III) reduc- tion by alleviating the need for metal reducers to directly contact insoluble metals or metals occluded within pore spaces too tight for metal reducers to access [30, 31, 46]. Fe(I1I) reduction can also be stimulated in subsurface sedi- ments with the addition of Fe(III) chelators that solubilize Fe(III) and make it more readily available for Fe(III) reduc- tion [38, 39, 40]. However, this third option is not an ap- propriate strategy for stimulating dissimilatory metal reduc- tion in metal-contaminated subsurface environments be- cause the chelators that solubilize Fe(III) may also chelate contaminant metals and enhance their mobility.
Highly transmissive sandy aquifers are likely subsurface environments in which contaminant metal mobility may be of concern. Such highly porous sediments are also among the most amenable for in situ treatment strategies that in- volve the addition of soluble stimulants. Therefore, a com- bination of laboratory and field studies was conducted in order to determine which microbial populations are stimu- lated when Fe(III) reduction is accelerated in sandy aquifers. The results from both field and laboratory studies demon- strate that stimulation of dissimilatory metal reduction with a variety of electron donors and/or electron shuttling com- pounds results in the specific enrichment of Geobacter spe- cies.
Materials and Methods Laboratory Incubations
Sediments for laboratory studies were collected from an aquifer located in Bemidji, Minnesota. Portions of the aquifer at this site have been contaminated with crude oil, and the petroleum- contaminated portion of the aquifer contains an extensive zone of Fe(III) reduction [4, 5, 50]. For this study, sediments were collected from a nearby uncontaminated, aerobic portion of the aquifer with a truck-mounted drill rig and sterilized core barrels, as previously described [5]. The cores were sectioned, immediately capped, wrapped in plastic wrap, sealed with duct tape, and placed in plastic bags under an N2 atmosphere prior to shipment in coolers via overnight carrier to the laboratory. In the laboratory, sediments (40 g) were transferred into serum bottles (5 ml) under N2 in a glove bag. The bottles were sealed with thick butyl rubber stoppers and removed from the glove bag, and the headspace was flushed with N2-CO2 (93:7). Acetate (10 mM), lactate (5 mM), formate (5 mM), benzoate (2 mM), or glucose (3.3 mM) were added as electron donors at the stated final concentrations from anaerobic stocks. Triplicate bottles of each treatment were incubated in the dark at 20?C. The sediments were subsampled over time, and HCl- extractable Fe(II) and Fe(III) were determined as previously de-
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

Subsurface Fe(III)-Reducing Community Structure 155
scribed [4]. Subsamples (2 g) of sediment were frozen at -80?C for the molecular analyses described below.
The effect of humics and anthraquinone 2,6-disulfonate (AQDS) on the rate of Fe(III) reduction in the sediments were also determined in a related study [46], where all of the sediments were amended with acetate (5 mM) as the electron donor and various concentrations of humics or AQDS. Subsamples that were frozen at -800C for molecular analysis during that study were analyzed in the study reported here.
Columbus Field Site
A field study was conducted to determine the effect of the addition of petroleum hydrocarbons on the microbiology and geochemistry of a sand and gravel aquifer at the Columbus Air Force Base, Columbus, Mississippi [6, 41]. This is the site of a groundwater research facility for the U.S. Air Force. A synthetic jet fuel mixture was placed within the aquifer in order to simulate the effect of a hydrocarbon spill in an aquifer. The development of the hydrocar- bon plume and its effects on the subsurface microbial ecology were monitored over time. Sediments were cored and samples processed as described above. The predominant terminal electron-accepting process in the sediments was determined with the [2-'4C]acetate technique [26]. Samples from a portion of the Fe(III)-reducing zone that developed in the aquifer as well as a nearby, uncontami- nated site were frozen at -800C for molecular analysis.
Borden Field Site
Sediments from a field study in which benzoate was injected into a sandy aquifer in order to promote anaerobic respiration were kindly provided by B.J. Butler, University of Waterloo. Portions of the Borden aquifer were divided with sheet piling to fashion three treatment gates parallel to groundwater flow. A contaminant solu- tion containing perchloroethylene (6 MtM), carbon tetrachloride (6 riM), and toluene (12 1iM) was added to the groundwater in the gates. Gate 2 received this contaminant solution only and no other additions. However, gate 3, in addition to the contaminants, also received during the treatment period a solution of benzoate (2.1 mM), which was injected into the groundwater at gate 3 approxi- mately every 14 days. This was done in order to promote anaerobic respiration in the gate. In this study, sediments of gate 2 and gate 3 were analyzed, as well as sediments from a nearby site that did not receive any additions. Sediment cores were collected from gates 2 and 3 and the background site over an interval of 1.5 to 3 m below ground surface on 4 July and 4 November 1997, after several pe- riodic injections of benzoate into the gate 3 site. Sediment cores were collected in aluminum core barrels, capped and sealed with duct tape, then returned to the lab where each 1.5-m core sample was cut into 0.3-m sections. Each section was then placed into an N2-filled anaerobic bag. About 1 cm of material from the ends of each section was discarded and the center portion of each section was sampled, homogenized in sterilized aluminum pans, and allo- cated into sterile Whirlpak bags inside sterile canning jars contain-
ing an argon headspace. Samples for molecular analysis were frozen at -80?C.
Concentrations of dissolved hydrogen at the gate 2 and gate 3 sampling sites were determined in October 1997 using the bubble strip method as previously described [28]. HCl-extractable Fe(III) and Fe(II) on the same sediments used for molecular analysis were determined as described above.
PCR-DGGE Analysis of Sediment Microbial Communities
The phylogenetic diversity of the microbial community in the sedi- ments was evaluated with PCR amplification of 16S rDNA genes and denaturing gradient gel electrophoresis (DGGE) [44, 45]. PCR- DGGE methods employed were as previously described [50], but included the following modifications: PCR was performed using bacterial primers 338F-GC (the complement of EUB338) [2] and 907R [20] with touchdown primer annealing from 65 to 55?C (decreasing 0.5?C per cycle), followed by 10 cycles at 55?C. This prevents amplification of nonspecific PCR products. In addition, archaeal diversity was accessed in the sampled sediments with Ar- chaea-specific primers ARC344 (5'-ACGGGGAGCAGCAGGC- GCGA-3') [48] and 0915Ra Reverse (5'-GTGCTCCCCCGC- CAATTCCT-3') [1]. 16S rDNA amplicons were resolved by DGGE with a 40 to 80% denaturing gradient (where 100% is equivalent to 7 M urea and 40% formamide) [45] on a 7% acrylamide gel in lx TAE (40mM Tris-acetate (pH 7.4), 20 mM sodium acetate, 1 mM EDTA). Standards of reference DNA, extracted from various pure cultures as previously described [50], were run along with the en- vironmental samples for ease of comparison among the different gels. DGGE gels were run for 15 h at 65 V and 60?C. PCR products from excised DGGE bands were purified using the QlAquick PCR Purification kit (Qiagen Inc., Valencia, CA), and sequenced using Dye Deoxy Terminator Cycle Sequencing (Perkin-Elmer Cetus) and an ABI 377 automated sequencer (Applied Biosystems, Foster City, CA) at the University of Massachusetts Sequencing Facility.
Phylogenetic Analysis of Sequence Data
The Ribosomal Database Project (RDP) [42] CHECK_CHIMERA program and secondary structure determination were used to check the partial 16S rRNA gene sequences for potential chimeric artifacts [18]. Sequences were analyzed using BLAST (National Center for Biotechnology Information) and SIMILARITY_RANK (RDP) in order to find the most similar available database sequences. Se- quences were then manually aligned with closely related 16S rRNA sequences from RDP and GenBank, using the graphical user inter- face SeqLab (Wisconsin Package version 10; Genetics Computer Group (GCG), Madison, WI). Only those sequence regions that could be aligned with confidence were included in the analyses, and gaps were treated as missing nucleotides. Phylogenetic trees were inferred from unambiguously aligned sequence data, using the dis- tance, maximum-likelihood, and maximum-parsimony tools of PAUP* [51].
Distance and maximum likelihood analyses were performed us- ing heuristic tree searching via simple stepwise addition with tree
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
156 O.L. Snoeyenbos-West et al.
bisection reconnection rearrangement. Unweighted parsimony analysis used the branch-and-bound algorithm. In addition, the distance tools (neighbor joining of Kimura distances) of TREECON for Windows 1.3b [52] were employed. Three different methods of phylogenetic tree construction were used with the same data set in order to test the robustness of the generated tree topo- logy. One thousand bootstrap replicates were performed on the data set. Phylogenetic trees inferred using the two different software packages described above showed the same topologies.
MPN-PCR Analysis of Sediments
The relative numbers of Geobacteraceae, Geothrix, and Shewanella species were determined in sediment samples using an MPN-PCR technique [5, 47]. DNA was extracted from triplicate sediment samples as described above. Serial 10-fold dilutions of the sediment DNA were made in sterile water, and 3-[dl aliquots of the diluted DNA were used as template in the PCR. Partial Geobacteraceae 16S rDNA sequences were amplified with bacterial forward primer 8F (5' -AGAGTTTGATCCTGGCTCAG-3') and Geobacteraceae- specific reverse primer Geo825R (5'-TACCCGCRACACCTAGT- 3') in the first round of a semi-nested PCR protocol, followed by 338F [2] and Geo825R in the second round. The PCR conditions were as previously described [5] except that 1 [l of the first-round reaction product served as template in second-round reactions. Geothrix species were also enumerated with a semi-nested MPN- PCR protocol. The primers for the first round of PCR were 8F and the Geothrix-specific reverse primer Gx.472R (5' -AGGT- ACCGTCAAGTAACASS-3'). The Geothrix-specific forward primer Gx. 1 82F (5' -AGACCTTCGGCTGGGATGCT-3') and Gx.472R were the primers for the second round. Both the Geobacteraceae and Geothrix-specific primers were designed and tested for speci- ficity in our lab prior to use in lab and field experiments (manu- script in preparation). In addition, partial Shewanella 16S rDNA sequences in sediment from the stimulation experiments were enu- merated using the Shewanella-specific primer set Sw.783-F (5'- AAAGACTGACGCTCAKGCA-3') and Sw. 1 245-R (5' -TTYG- CAACCCTCTGTACT-3') (manuscript in preparation).
Results Effect of Electron Donor Amendments to Bemidji Aquifer Sediments
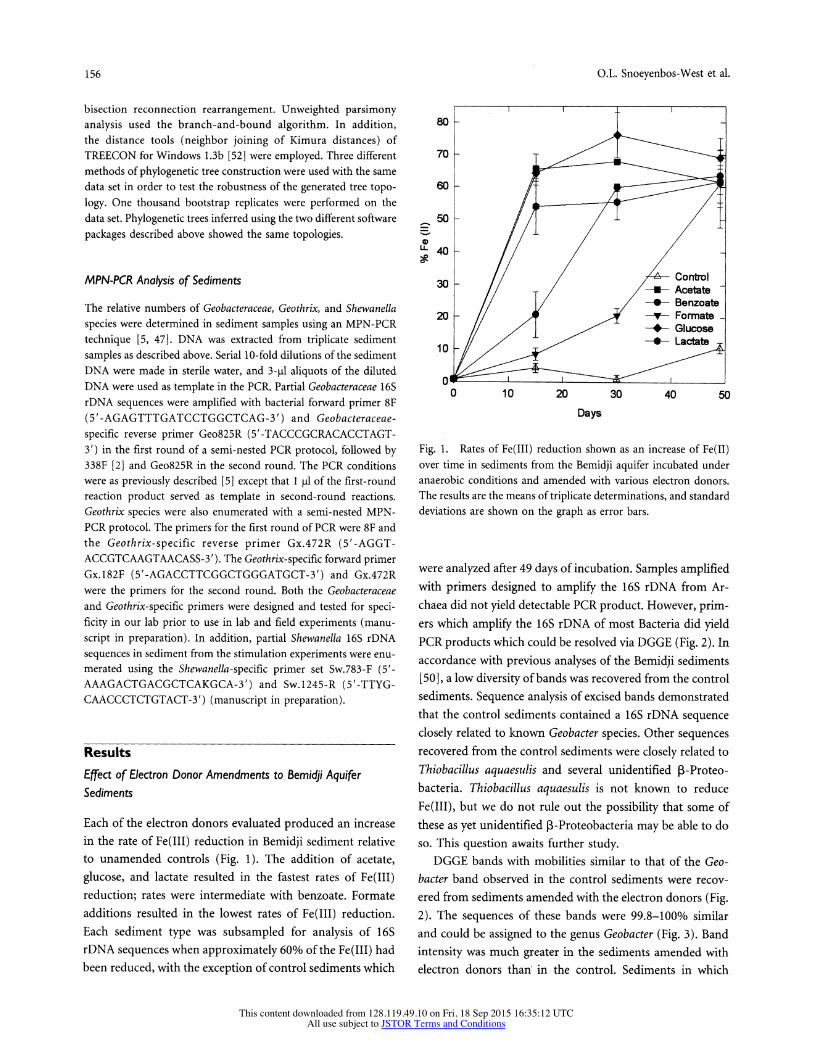
Each of the electron donors evaluated produced an increase in the rate of Fe(III) reduction in Bemidji sediment relative to unamended controls (Fig. 1). The addition of acetate, glucose, and lactate resulted in the fastest rates of Fe(III) reduction; rates were intermediate with benzoate. Formate additions resulted in the lowest rates of Fe(III) reduction. Each sediment type was subsampled for analysis of 16S rDNA sequences when approximately 60% of the Fe(LII) had been reduced, with the exception of control sediments which
80
70
60
50
uL 40
3 0 -0 Cont-ol 30 -U-- ~~~~~~~~Acetate
// Benzoate 20 -v- Formate
-4- Glucose 10 ---~~~~~~~~~ Lactate
10
0 10 20 30 40 50 Days
Fig. 1. Rates of Fe(IIL) reduction shown as an increase of Fe(II) over time in sediments from the Bemidji aquifer incubated under anaerobic conditions and amended with various electron donors. The results are the means of triplicate determinations, and standard deviations are shown on the graph as error bars.
were analyzed after 49 days of incubation. Samples amplified with primers designed to amplify the 16S rDNA from Ar- chaea did not yield detectable PCR product. However, prim- ers which amplify the 16S rDNA of most Bacteria did yield PCR products which could be resolved via DGGE (Fig. 2). In accordance with previous analyses of the Bemidji sediments [50], a low diversity of bands was recovered from the control sediments. Sequence analysis of excised bands demonstrated that the control sediments contained a 16S rDNA sequence closely related to known Geobacter species. Other sequences recovered from the control sediments were closely related to Thiobacillus aquaesulis and several unidentified ,B-Proteo- bacteria. Thiobacillus aquaesulis is not known to reduce Fe(III), but we do not rule out the possibility that some of these as yet unidentified 13-Proteobacteria may be able to do so. This question awaits further study.
DGGE bands with mobilities similar to that of the Geo- bacter band observed in the control sediments were recov- ered from sediments amended with the electron donors (Fig. 2). The sequences of these bands were 99.8-100% similar and could be assigned to the genus Geobacter (Fig. 3). Band intensity was much greater in the sediments amended with electron donors than' in the control. Sediments in which
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
Subsurface Fe(III)-Reducing Community Structure 157
Standard Standard .. Fotn 3azaormate
Clostridium _
Clostridium red eubacterium
f1-Proteobacten
_ _ ~~~~ M 1i#t r .
~~~~~~Formats Goo2 f3-Proteobacteri' Denftrobancterium Thiobaoillus aq
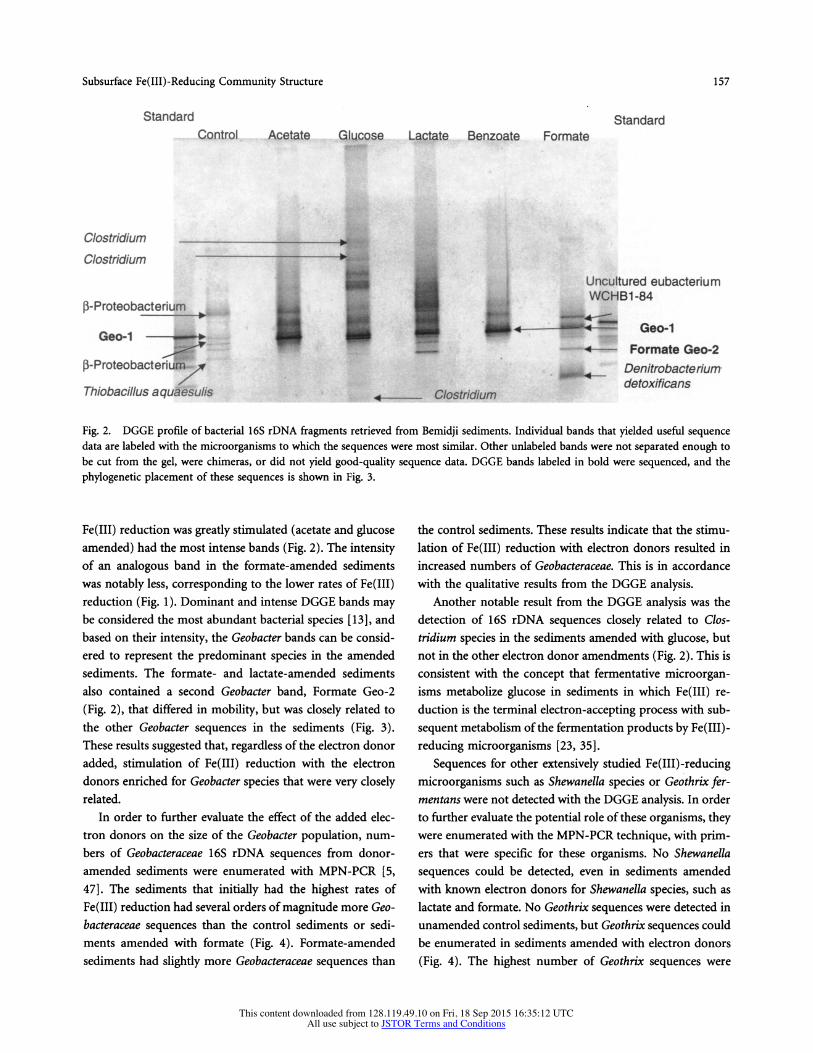
Fig. 2. DGGE profile of bacterial 16S rDNA fragments retrieved from Bemidji sediments. Individual bands that yielded useful sequence data are labeled with the microorganisms to which the sequences were most similar. Other unlabeled bands were not separated enough to be cut from the gel, were chimeras, or did not yield good-quality sequence data. DGGE bands labeled in bold were sequenced, and the phylogenetic placement of these sequences is shown in Fig. 3.
Fe(III) reduction was greatly stimulated (acetate and glucose amended) had the most intense bands (Fig. 2). The intensity of an analogous band in the formate-amended sediments was notably less, corresponding to the lower rates of Fe(III) reduction (Fig. 1). Dominant and intense DGGE bands may be considered the most abundant bacterial species [13], and based on their intensity, the Geobacter bands can be consid- ered to represent the predominant species in the amended sediments. The formate- and lactate-amended sediments also contained a second Geobacter band, Formate Geo-2 (Fig. 2), that differed in mobility, but was closely related to the other Geobacter sequences in the sediments (Fig. 3). These results suggested that, regardless of the electron donor added, stimulation of Fe(III) reduction with the electron donors enriched for Geobacter species that were very closely related.
In order to further evaluate the effect of the added elec- tron donors on the size of the Geobacter population, num- bers of Geobacteraceae 16S rDNA sequences from donor- amended sediments were enumerated with MPN-PCR [5, 47]. The sediments that initially had the highest rates of Fe(III) reduction had several orders of magnitude more Geo- bacteraceae sequences than the control sediments or sedi- ments amended with formate (Fig. 4). Formate-amended sediments had slightly more Geobacteraceae sequences than
the control sediments. These results indicate that the stimu- lation of Fe(III) reduction with electron donors resulted in increased numbers of Geobacteraceae. This is in accordance with the qualitative results from the DGGE analysis.
Another notable result from the DGGE analysis was the detection of 16S rDNA sequences closely related to Clos- tridium species in the sediments amended with glucose, but not in the other electron donor amendments (Fig. 2). This is consistent with the concept that fermentative microorgan- isms metabolize glucose in sediments in which Fe(III) re- duction is the terminal electron-accepting process with sub- sequent metabolism of the fermentation products by Fe(III)- reducing microorganisms [23, 35].
Sequences for other extensively studied Fe(III)-reducing microorganisms such as Shewanella species or Geothrix fer- mentans were not detected with the DGGE analysis. In order to further evaluate the potential role of these organisms, they were enumerated with the MPN-PCR technique, with prim- ers that were specific for these organisms. No Shewanella sequences could be detected, even in sediments amended with known electron donors for Shewanella species, such as lactate and formate. No Geothrix sequences were detected in unamended control sediments, but Geothrix sequences could be enumerated in sediments amended with electron donors (Fig. 4). The highest number of Geothrix sequences were
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

158 O.L. Snoeyenbos-West et al.
Borden Geo-1 _ Borden Geo-2
Borden Geo-3 70 Borden Geo-4
Borden Geo-5 Borden Geo-6 Borden Geo-7
62 Borden Geo-8 Borden Geo-9 Borden Geo-1 0
LBorden Geo-1I Humics Geo-82 Humics Geo-24 ControlGeo-1 AQDS Geo-i Humics Geo-1 Humics Geo-2 m a
94. Humics Geo-3 Z L
Formate Geo-2 c o Glucose Geo-1 Acetate Geo-1 Benzoate Geo-1 Formats Geo-1 0 Lactate Geo-1
Bemidji Geo-60 Bemidji Geo-83 O Borden Geo-101 0 0 Bemidji benzene enrichment (Benz-76) @
Columbus Geo-1 0 Columbus Geo-2
"Geobacterbemidjiensis" 82 Geobacter arculus o
Geobacter sulfureducens Q Geobacter metallireducens 0
Geobacter hydrogenophilus L Pelobacterpropionicus
Geobacter chapellei GeobacterALA6 Geobacter humireducens GeobacterALA5
Geobacter akaganeitreducens 84 100 Desulfuromrusa bakii
100 LDesulfuromusa kysingii Desulfuromnusa succinoxidans
Pelobacter carbinolicus r Pelobacter acetylenicus
Desulfuromonas acetoxidans Desulfuromonas acetexigens Pelobacter venetanus 100 Humics Dsf-1
100 FColumbus Dsfi1 Bemidji Geo-1 44 0
62 1 Borden Dsf-1 0 o LI Bemidji Geo-47 uE Bemidji Geo-52 0
100 Bemidji Geo-48 61 r 1Bemidji Geo-58 Ua
00 Bemidji Geo-43 c
q Bemidji Geo-3
Bemidji Geo-2 Desulfovibrio vulgaris
Escherichia coli 0.1 substitutions/site
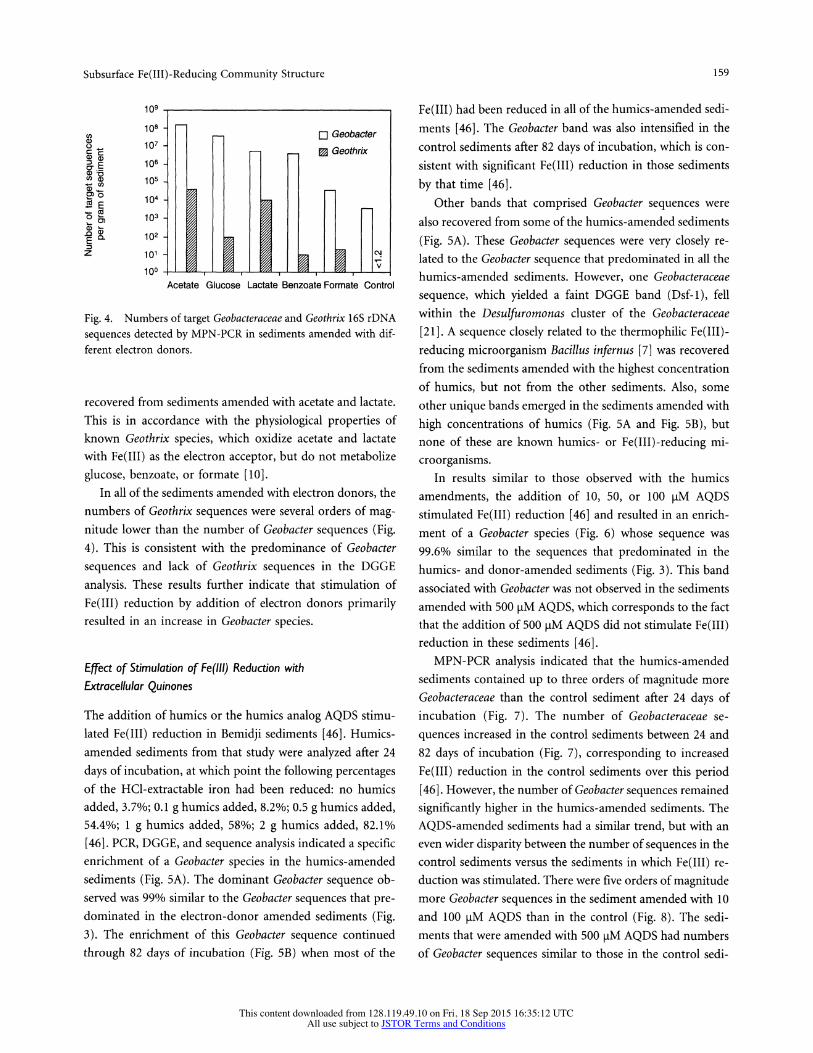
Fig. 3. Phylogenetic tree inferred from 16S rRNA sequences showing the phylogenetic placement of Geo- bacter sequences amplified from sediments in three different aqui- fers, and from environmental iso- lates. Phylogenetic relationships shown here were inferred by using neighbor joining and Kimura two- parameter genetic distances in TREECON [52]. A total of 400 base positions were considered in the analysis. Bootstrap values above 60 are shown adjacent to branch nodes and were calculated from 1,000 re- sampled data sets using neighbor joining. The scale bar shows ex- pected nucleotide substitutions per site per unit of branch length. Op- erational Taxonomic Units (OTUs) shown on the tree in bold are the same sequences obtained using DGGE from amended sediments and fields sites in this study. Those shown in plain text are from a pre- vious study of the Bemidji aquifer [50], and from GenBank and the RDP databases. A similar tree to- pology was generated for trees con- structed using maximum-likeli- hood and maximum-parsimony methods (data not shown).
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
Subsurface Fe(III)-Reducing Community Structure 159
109
1 At8 -
lE Geobacter Fig. 4 Geothrix
106- fee)n dn10 r
1- 104 E z 0 " 100 V
recovered from sediments amended with acetate and lactate. This is in accordance with the physiological properties of known Geothrix species, which oxidize acetate and lactate with Fe(III) as the electron acceptor, but do not metabolize glucose, benzoate, or formate [10].
In all of the sediments amended with electron donors, the numbers of Geothrix sequences were several orders of mag- nitude lower than the number of Geobacter sequences (Fig. 4). This is consistent with the predominance of Geobacter sequences and lack of Geothrix sequences in the DGGE analysis. These results further indicate that stimulation of Fe(III) reduction by addition of electron donors primarily resulted in an increase in Geobacter species.
Effect of Stimulation of Fe(lll) Reduction with Extracellular Quinones
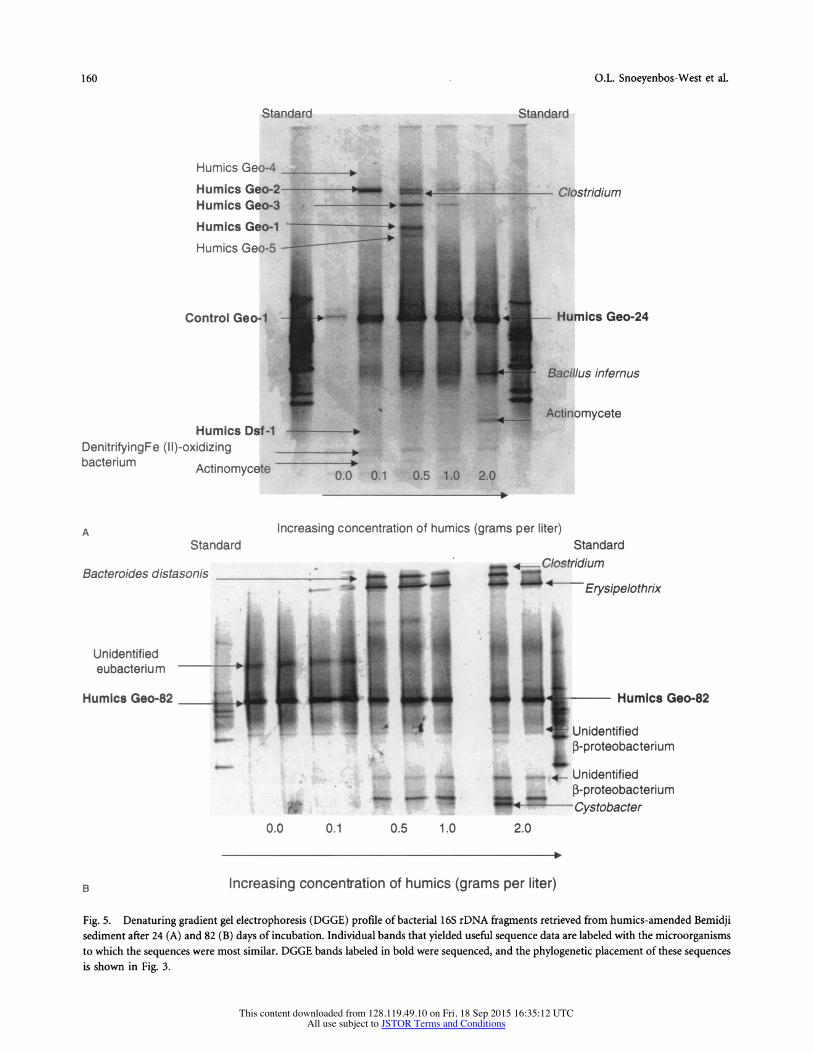
The addition of humics or the humics analog AQDS stimu- lated Fe(III) reductionrin Bemidji sediments [46]. Humics- amended sediments from that study were analyzed after 24 days of incubation, at which point the following percentages of the HCl-extractable iron had been reduced: no humics added, 3.7%; 0.1 g humics added, 8.2%; 0.5 g humics added, 54.4%; 1 g humics added, 58%; 2 g humics added, 82.1% [46]. PCR, DGGE, and sequence analysis indicated a specific enrichment of a Geobacter species in the humics-amended sediments (Fig. 5A). The dominant Geobacter sequence ob- served was 99% similar to the Geobacter sequences that pre- dominated in the electron-donor amended sediments (Fig. 3). The enrichment of this Geobacter sequence continued through 82 days of incubation (Fig. SB) when most of the
Fe(III) had been reduced in all of the humics-amended sedi- ments [46]. The Geobacter band was also intensified in the control sediments after 82 days of incubation, which is con- sistent with significant Fe(III) reduction in those sediments by that time [46].
Other bands that comprised Geobacter sequences were also recovered from some of the humics-amended sediments (Fig. 5A). These Geobacter sequences were very closely re- lated to the Geobacter sequence that predominated in all the humics-amended sediments. However, one Geobacteraceae sequence, which yielded a faint DGGE band (Dsf-1), fell within the Desulfuromonas cluster of the Geobacteraceae [21]. A sequence closely related to the thermophilic Fe(III)- reducing microorganism Bacillus infernus [7] was recovered from the sediments amended with the highest concentration of humics, but not from the other sediments. Also, some other unique bands emerged in the sediments amended with high concentrations of humics (Fig. 5A and Fig. 5B), but none of these are known humics- or Fe(III)-reducing mi- croorganisms.
In results similar to those observed with the humics amendments, the addition of 10, 50, or 100 iM AQDS stimulated Fe(IIJ) reduction [46] and resulted in an enrich- ment of a Geobacter species (Fig. 6) whose sequence was 99.6% similar to the sequences that predominated in the humics- and donor-amended sediments (Fig. 3). This band associated with Geobacter was not observed in the sediments amended with 500 [tM AQDS, which corresponds to the fact that the addition of 500 [M AQDS did not stimulate Fe(III) reduction in these sediments [46].
MPN-PCR analysis indicated that the humics-amended sediments contained up to three orders of magnitude more Geobacteraceae than the control sediment after 24 days of incubation (Fig. 7). The number of Geobacteraceae se- quences increased in the control sediments between 24 and 82 days of incubation (Fig. 7), corresponding to increased Fe(III) reduction in the control sediments over this period [46]. However, the number of Geobacter sequences remained significantly higher in the humics-amended sediments. The AQDS-amended sediments had a similar trend, but with an even wider disparity between the number of sequences in the control sediments versus the sediments in which Fe(III) re- duction was stimulated. There were five orders of magnitude more Geobacter sequences in the sediment amended with 10 and 100 MM AQDS than in the control (Fig. 8). The sedi- ments that were amended with 500 [MM AQDS had numbers of Geobacter sequences similar to those in the control sedi-
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
160 O.L. Snoeyenbos-West et al.
.e~~~~~~~~~~~~~~~~~1 I
Humics G Humics Humics Humics Humics GA *
A Increasing concentration of humics (grams per l_fer) Stndr Standard2
Control Go le trII
j~~~~~~~~~~ _
| tLFS ~~~~~~~~~~infemus
Humics mct DenitrffyingFe (d)-oxidizing aedum Abateou
A Increasing concentration of humics (Grams per ler) Standard Standard
-post baduum Bac-tq dirostasonisum * -t!t w _ w -. + ~~~~Erysipelothnx
~~~~~~~~~~~~~~~~~~~~~~Unidentified e-e-
Cysprotoobacter u_
B Increasing concentration of humics (gramsperliter)um
.... X_X .......... s -s.s s :s:.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.... ..
Fi. .Deaurn gadet e eetroporsi (G E) prfl of t1 >obacteria6rDAfgmnsetivdroh umic-mne eij
2 .^;.: ;rffl ̂ ^ w --Cyst ystbacter 0.0 0.1 0.5 1.0 2-o
sediment after 24 (A) and 82 (B) days of incubation. Individual bands that yielded useful sequence data are labeled with the microorganisms to which the sequences were most similar. DGGE bands labeled in bold were sequenced, and the phylogenetic placement of these sequences is shown in Fig. 3.
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
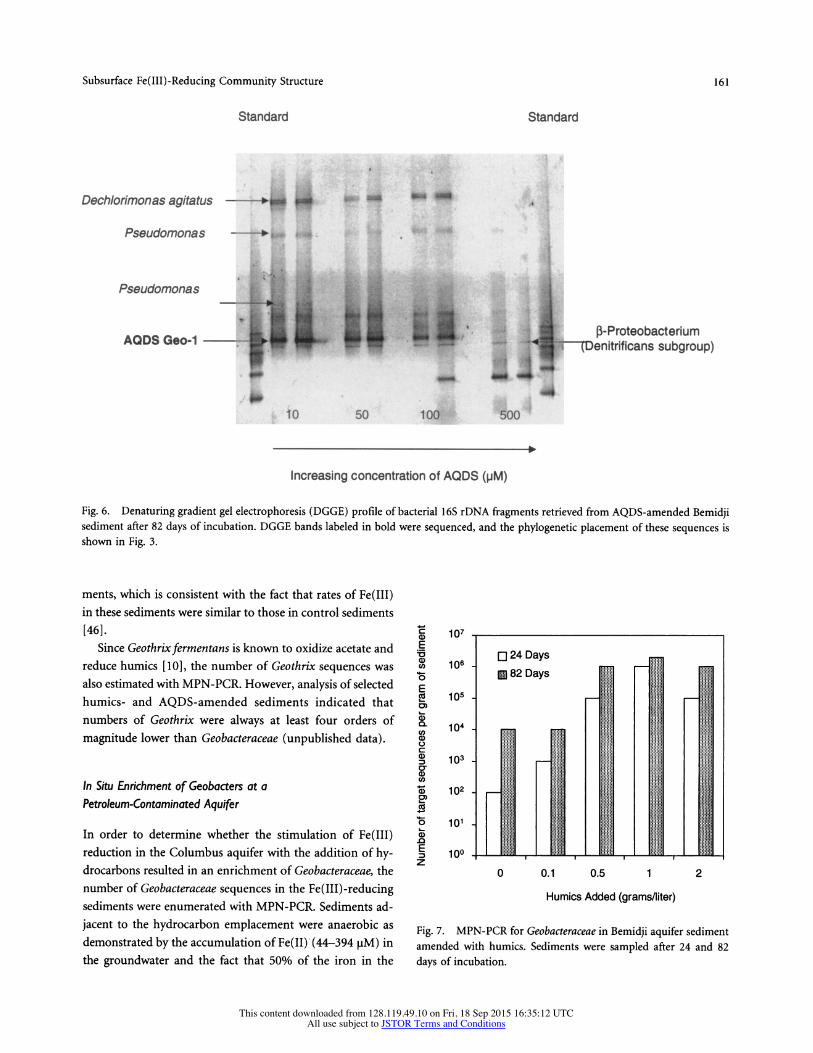
Subsurface Fe(III)-Reducing Community Structure 161
Stadard Standard
Dechiorimonas aiau PDsudnmonas
Pseudomonas
AQDS Go-
10-Protob terIum Itr_MIcans subgroup)
Increasing co raton of AQDS (pM)
Fig. 6. Denaturing gradient gel electrophoresis (DGGE) profile of bacterial 16S rDNA fragments retrieved from AQDS-amended Bemidji sediment after 82 days of incubation. DGGE bands labeled in bold were sequenced, and the phylogenetic placement of these sequences is shown in Fig. 3.
ments, which is consistent with the fact that rates of Fe(III) in these sediments were similar to those in control sediments [46].
Since Geothrixfermentans is known to oxidize acetate and reduce humics [10], the number of Geothrix sequences was also estimated with MPN-PCR. However, analysis of selected humics- and AQDS-amended sediments indicated that numbers of Geothrix were always at least four orders of magnitude lower than Geobacteraceae (unpublished data).
In Situ Enrichment of Geobacters at a Petroleum-Contaminated Aquifer
In order to determine whether the stimulation of Fe(III) reduction in the Columbus aquifer with the addition of hy- drocarbons resulted in an enrichment of Geobacteraceae, the number of Geobacteraceae sequences in the Fe(III)-reducing sediments were enumerated with MPN-PCR. Sediments ad- jacent to the hydrocarbon emplacement were anaerobic as demonstrated by the accumulation of Fe(II) (44-394 iiM) in the groundwater and the fact that 50% of the iron in the
c 107 E .5 fl~~24 Days '106
dyof incuban82.Days E
aL 104 CD)
103 CO) CD
CIS
0; 101 0) 10
z 0 0.1 0.5 1 2
Humics Added (grams/liter)
Fig. 7. MPN-PCR for Geobacteraceae in Bemidji aquifer sediment amended with humics. Sediments were sampled after 24 and 82 days of incubation.
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
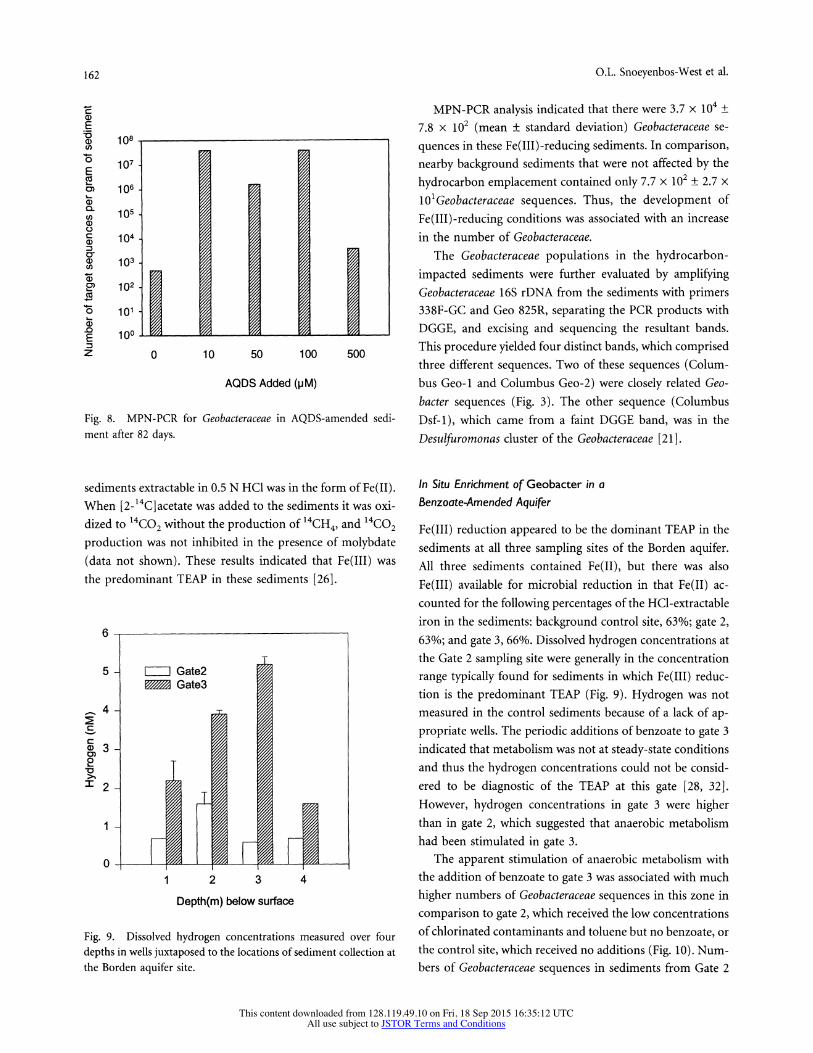
162 O.L. Snoeyenbos-West et al.
CD E a) 108.-
0 107 - CY 106 L)
CL 105 U"
2' 0102
a) 101 aI)
ment after 82 days.
sediments extractable in 0.5 N HCl was in the form of Fe(II). When [2-'4C] acetate was added to the sediments it was oxi- dized to '4C02 without the production of 14CH4, and '4C02 production was not inhibited in the presence of molybdate (data not shown). These results indicated that Fe(III) was the predominant TEAP in these sediments [26].
6r
theBodenaqifGaste3
co
0 0 a)3 -0) 10 0
men ate 1 2a3s4
Fig.e9 Dissolvedat hydoge conentrationsedmeatsured over four-
prdepthsion wels jutaosdto ihbthed oaions ofe prsedimen cfollecionate
the Boredemnaqufe sTE.Pi hs eiet 2]
MPN-PCR analysis indicated that there were 3.7 x 1 + 7.8 x 102 (mean ? standard deviation) Geobacteraceae se- quences in these Fe(III) -reducing sediments. In comparison, nearby background sediments that were not affected by the hydrocarbon emplacement contained only 7.7 x 102 ? 2.7 x lI1Geobacteraceae sequences. Thus, the development of Fe(III)-reducing conditions was associated with an increase in the number of Geobacteraceae.
The Geobacteraceae populations in the hydrocarbon- impacted sediments were further evaluated by amplifying Geobacteraceae 16S rDNA from the sediments with primers 338F-GC and Geo 825R, separating the PCR products with DGGE, and excising and sequencing the resultant bands. This procedure yielded four distinct bands, which comprised three different sequences. Two of these sequences (Colum- bus Geo-1 and Columbus Geo-2) were closely related Geo- bacter sequences (Fig. 3). The other sequence (Columbus Dsf- 1), which came from a faint DGGE band, was in the Desulfuromonas cluster of the Geobacteraceae [21].
In Situ Enrichment of Geobacter in a Benzoate-Amended Aquifer
Fe(IIJ) reduction appeared to be the dominant TEAP in the sediments at all three sampling sites of the Borden aquifer. All three sediments contained Fe(JI), but there was also Fe(III) available for microbial reduction in that Fe(II) ac- counted for the following percentages of the HCl-extractable iron in the sediments: background control site, 63%; gate 2, 63%; and gate 3, 66%. Dissolved hydrogen concentrations at the Gate 2 sampling site were generally in the concentration range typically found for sediments in which Fe(II) reduc- tion is the predominant TEAP (Fig. 9). Hydrogen was not measured in the control sediments because of a lack of ap- propriate wells. The periodic additions of benzoate to gate 3 indicated that metabolism was not at steady-state conditions and thus the hydrogen concentrations could not be consid- ered to be diagnostic of the TEAP at this gate [28, 32]. However, hydrogen concentrations in gate 3 were higher than in gate 2, which suggested that anaerobic metabolism had been stimulated in gate 3.
The apparent stimulation of anaerobic metabolism with the addition of benzoate to gate 3 was associated with much higher numbers of Geobacteraceae sequences in this zone in comparison to gate 2, which received the low concentrations of chlorinated contaminants and toluene but no benzoate, or the control site, which received no additions (Fig. 10). Num- bers of Geobacteraceae sequences in sediments from Gate 2
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
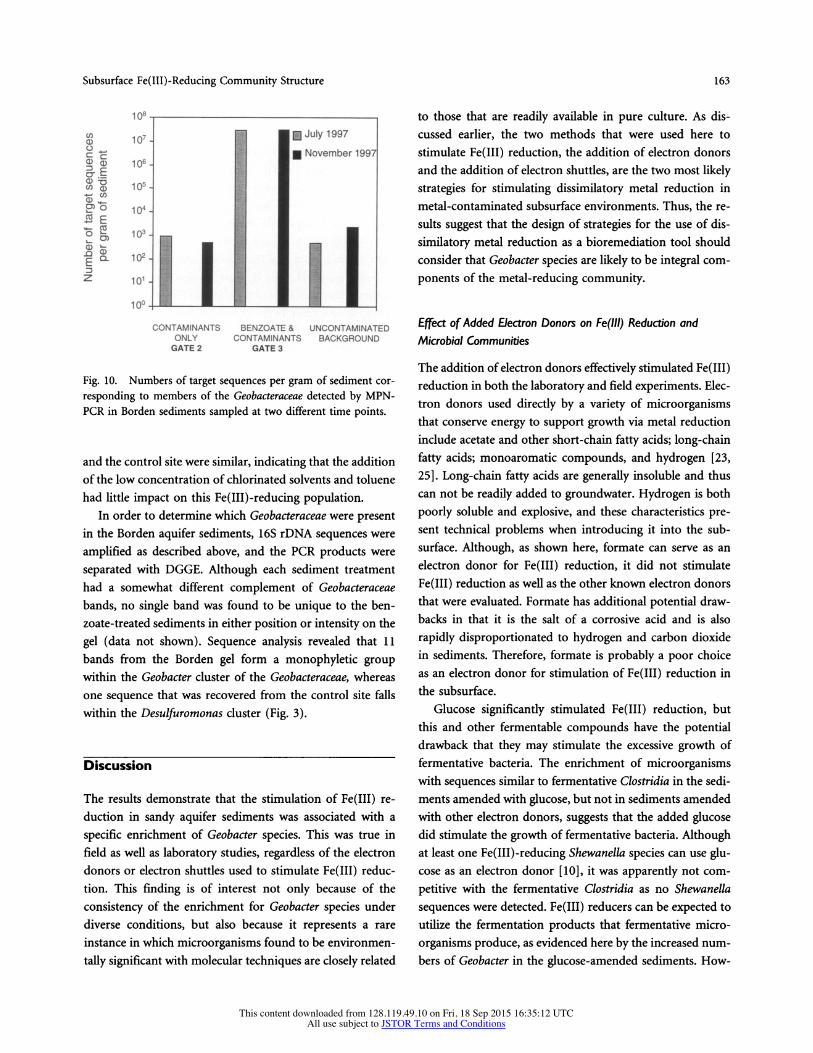
Subsurface Fe(III)-Reducing Community Structure 163
108 I l)
0 1~07 July 1997
li X 10_ 1 November 19
OD 0 101 - CD~ 10?.
10 ICP
102 Z 101
100
CONTAMINANTS BENZOATE & UNCONTAMINATED ONLY CONTAMINANTS BACKGROUND
GATE 2 GATE3
Fig. 10. Numbers of target sequences per gram of sediment cor- responding to members of the Geobacteraceae detected by MPN- PCR in Borden sediments sampled at two different time points.
and the control site were similar, indicating that the addition of the low concentration of chlorinated solvents and toluene had little impact on this Fe(III)-reducing population.
In order to determine which Geobacteraceae were present in the Borden aquifer sediments, 16S rDNA sequences were amplified as described above, and the PCR products were separated with DGGE. Although each sediment treatment had a somewhat different complement of Geobacteraceae bands, no single band was found to be unique to the ben- zoate-treated sediments in either position or intensity on the gel (data not shown). Sequence analysis revealed that 11 bands from the Borden gel form a monophyletic group within the Geobacter cluster of the Geobacteraceae, whereas one sequence that was recovered from the control site falls within the Desulfuromonas cluster (Fig. 3).
Discussion
The results demonstrate that the stimulation of Fe(III) re- duction in sandy aquifer sediments was associated with a specific enrichment of Geobacter species. This was true in field as well as laboratory studies, regardless of the electron donors or electron shuttles used to stimulate Fe(III) reduc- tion. This finding is of interest not only because of the consistency of the enrichment for Geobacter species under diverse conditions, but also because it represents a rare instance in which microorganisms found to be environmen- tally significant with molecular techniques are closely related
to those that are readily available in pure culture. As dis- cussed earlier, the two methods that were used here to stimulate Fe(III) reduction, the addition of electron donors and the addition of electron shuttles, are the two most likely strategies for stimulating dissimilatory metal reduction in metal-contaminated subsurface environments. Thus, the re- sults suggest that the design of strategies for the use of dis- similatory metal reduction as a bioremediation tool should consider that Geobacaer species are likely to be integral com- ponents of the metal-reducing community.
Effect of Added Electron Donors on Fe(lIl) Reduction and Microbial Communities
The addition of electron donors effectively stimulated Fe(III) reduction in both the laboratory and field experiments. Elec- tron donors used directly by a variety of microorganisms that conserve energy to support growth via metal reduction include acetate and other short-chain fatty acids; long-chain fatty acids; monoaromatic compounds, and hydrogen [23, 25]. Long-chain fatty acids are generally insoluble and thus can not be readily added to groundwater. Hydrogen is both poorly soluble and explosive, and these characteristics pre- sent technical problems when introducing it into the sub- surface. Although, as shown here, formate can serve as an electron donor for Fe(III) reduction, it did not stimulate Fe(III) reduction as well as the other known electron donors that were evaluated. Formate has additional potential draw- backs in that it is the salt of a corrosive acid and is also rapidly disproportionated to hydrogen and carbon dioxide in sediments. Therefore, formate is probably a poor choice as an electron donor for stimulation of Fe(III) reduction in the subsurface.
Glucose significantly stimulated Fe(III) reduction, but this and other fermentable compounds have the potential drawback that they may stimulate the excessive growth of fermentative bacteria. The enrichment of microorganisms with sequences similar to fermentative Clostridia in the sedi- ments amended with glucose, but not in sediments amended with other electron donors, suggests that the added glucose did stimulate the growth of fermentative bacteria. Although at least one Fe(III)-reducing Shewanella species can use glu- cose as an electron donor [10], it was apparently not com- petitive with the fermentative Clostridia as no Shewanella sequences were detected. Fe(III) reducers can be expected to utilize the fermentation products that fermentative micro- organisms produce, as evidenced here by the increased num- bers of Geobacter in the glucose-amended sediments. How-
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

164 O.L. Snoeyenbos-West et al.
ever, stimulating the growth of fermentative microorganisms with glucose or other more complex fermentable substrates could increase the possibility of plugging the aquifer with biomass that does not directly contribute to Fe(III) reduc- tion.
In contrast to fermentable sugars, acetate, lactate, and monoaromatic compounds can be directly oxidized to car- bon dioxide by Fe(III)-reducing microorganisms [23, 24]. It was previously speculated [23] that Fe(III) reducers with a metabolism like that of the various Shewanella species might first partially oxidize lactate to acetate and carbon dioxide in Fe(III)-reducing environments. However, it has subse- quently been found that some Fe(III) reducers, including Geobacter species [ 11 ], can completely oxidize lactate to car- bon dioxide. The finding that a Geobacter population be- came dominant in the lactate-amended sediments and that there was no enrichment of Shewanella sequences in lactate- amended sediments suggests that Geobacter species capable of completely oxidizing lactate were most effective at scav- enging the added lactate.
Acetate was very effective in stimulating Fe(III) reduc- tion. Numerous members of the Geobacteraceae family can oxidize acetate with the reduction of Fe(III) [24], as can Geovibrio ferrireducens [9], Ferribacterium limneticum [ 12], and Geothrixfermentans [10]. However, acetate additions led primarily to the enrichment of a Geobacter population. No sequences related to the other known acetate-oxidizing Fe(III)-reducing microorganisms were among the most prominent sequences recovered in the DGGE analysis of the sediments. Enumeration of Geothrix 16S rDNA sequences with specific PCR primers indicated that the growth of this bacterium was stimulated with the addition of acetate and other electron donors. However, Geobacter sequences were typically three to five orders of magnitude greater than those of Geothrix.
Benzoate additions stimulated Fe(III) reduction in the laboratory studies, and benzoate and aromatic hydrocarbons resulted in the development of Fe(III)-reducing conditions in the field studies. This result might have been expected based on previous studies, which have demonstrated that the release of monoaromatic hydrocarbons into groundwater as the result of hydrocarbon contamination greatly stimulates Fe(III) reduction in aquifers [27, 26]. Geobacter species are the only Fe(III)-reducing microorganisms that are known to have the ability to oxidize monoaromatic compounds with the reduction of Fe(III) [24]. In previous studies of the pe- troleum-contaminated portion of the Bemidji aquifer [5,
50], it was found that there was a significant enrichment of Geobacter in the zone where the most rapid oxidation of aromatic hydrocarbons such as benzene [5] and toluene [4] was taking place. DGGE analysis indicated that Geobacter species were major components of the community in those aromatic hydrocarbon-degrading sediments, and MPN-PCR demonstrated that the Geobacteraceae sequences in the most active Fe(III)-reducing sediments were four orders of mag- nitude more numerous than in the uncontaminated back- ground sediments. This increase in Geobacter in the con- taminated sediments was attributed to Geobacter species oxi- dizing the aromatic hydrocarbons at that site. The findings reported here that the growth of Geobacter species was en- hanced with the addition of benzoate to Bemidji sediments, with the addition of benzoate to the Borden aquifer, and in the presence of petroleum hydrocarbons at the Columbus site demonstrate that Geobacter species are likely to be im- portant components of the Fe(JII)-reducing community in a variety of sandy aquifers in which Fe(III) reduction is stimu- lated with monoaromatic compounds.
Stimulation of Fe(lIl) Reduction with Extracellular Quinones
Recent studies have suggested that another strategy for stimulating Fe(III) reduction in subsurface environments is the addition of humics or other extracellular quinones [30, 31, 39, 46]. Fe(III) reducers can reduce the quinones in these organics to the hydroquinone state, and the hydroquinones can then abiotically reduce Fe(III) [30, 31] as well as Mn(IV) and contaminant metals such as uranium and technetium (unpublished data). The Fe(III) reduction step oxidizes the hydroquinone to the quinone, which can then undergo an- other cycle of reduction and oxidation. This alleviates the need for Fe(III)-reducing microorganisms to come into di- rect physical contact with insoluble Fe(III) oxides in order to reduce them. In addition, hydroquinones generated via qui- none reduction by Fe(III) reducers may be able to enter pore spaces that are too small for Fe(III) reducers to enter and permit reduction of Fe(III) and other metals that would otherwise be inaccessible [31]. Although there has not yet been an opportunity to conduct a field trial on the effect of the addition of extracellular quinones on Fe(III) reduction in sandy aquifers, the laboratory results suggest that the addition of humics and other extracellular quinones will stimulate the growth and activity of Geobacter species in aquifer sediments.
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

Subsurface Fe(III)-Reducing Community Structure 165
Implications of the Apparent Ubiquity of Geobacter Species in Fe(lIl)-Reducing Sediments of Sandy Aquifers
The enrichment of Geobacter species in response to the stimulation of Fe(IIL) reduction in the studies reported here is consistent with the known physiological characteristics of Geobacter species that have been studied in pure culture. However, it was not necessarily expected that stimulation of Fe(III) reduction in sandy aquifer sediments would result in the growth of organisms so closely related to those already in pure culture. For example, it is commonly considered diffi- cult to culture the microorganisms that are the most envi- ronmentally significant [3]. Yet, the Geobacter sequences that predominate in the aquifer sediments are closely related to Geobacter species in pure culture, and we have been able to recover organisms in culture from the Bemidji aquifer that are closely related to the sequences that predominate there.
The consistent enrichment of similar Geobacter species, regardless of the method for stimulating Fe(JII) reduction, also could not have been anticipated in advance. For ex- ample, from the known physiological characteristics of the Fe(III)-reducing microorganisms in pure culture it seemed possible that Shewanella species that can oxidize lactate and formate with the reduction of Fe(III) [8, 37] might have been stimulated when these electron donors were added. The finding with molecular techniques that Shewanella spe- cies were either absent from the sediments or present in very low numbers is in accordance with recent culturing studies, which have indicated that even when lactate or formate are used as the electron donor, Geobacter species rather than Shewanella are recovered from soils (Coates, manuscript in preparation).
One of the most interesting results was the finding that the Geobacter species that predominated in the Fe(III)- reducing sediments from three geographically distinct aqui- fers had 16S rDNA sequences that were very similar, desig- nated here as the "Subsurface Geobacter Cluster." The Geo- bacter sequences that predominated in the Fe(III)-reducing sediments from Columbus (Columbus Geo-1 and Geo-2) and the Borden (Borden Geo-1 through Geo-11) aquifer sediments were not only closely related to the Geobacter sequences in the Bemidji sediments from the laboratory studies reported here, but were also very similar to the pre- viously reported [50] Geobacter sequences that were abun- dant in the Fe(III)-reducing zone of the Bemidji aquifer (Geo-60 and Geo-83) and in a benzene-oxidizing, Fe(III)-
reducing enrichment culture (Benz-76) from this site (Fig. 3). An additional Geobacter sequence (Borden Geo-101) that is 98% similar to Geo-83 was recovered from the Borden sediments when the 16S rDNA was amplified with the PCR primer set designed to amplify the 16S rDNA of most Bac- teria (Fig. 3). Although Desulfuromonas sequences were less predominant than Geobacter sequences, three partial 16S rDNA sequences (Humics Dsf- 1, Columbus Dsf- 1, and Bor- den Dsf- 1) that fall within the Desulfuromonas Cluster of the family Geobacteraceae [21] were retrieved from each of the aquifer sites (Fig. 3). They are very similar to one another and to those found in a previous study of the Bemidji aquifer [50] and are designated here as the "Subsurface Desulfu- romonas Cluster" (Fig. 3).
It is becoming increasingly apparent [14, 15] that similar microbial species may be ubiquitous in freshwater [43, 54], marine [16, 17], and aerobic soil environments [19, 49]. The results presented here indicate that such microbial ubiquity may also be extended to the anaerobic zones of sandy aqui- fers.
Conclusions
The results of this study, as well as previous investigations of a petroleum-contaminated aquifer [5, 50], demonstrate that organisms with 16S rDNA sequences closely related to known species of Geobacter species are significantly enriched under a variety of conditions that promote dissimilatory Fe(III) reduction in sandy aquifers. The finding that it may be possible to isolate the most important Fe(III)-reducing microorganisms in pure culture provides a unique oppor- tunity to combine environmental and culture studies to si- multaneously investigate the biochemical and geochemical factors influencing the rate and extent of Fe(III) reduction in the subsurface.
Acknowledgments
The NABIR Program of the Department of Energy (Grant DEFG0297ER62475) supported this work. We thank G. De- lin and W. Larson for Bemidji sediment sample collection, T. Stauffer and R. Stapleton for coordinating sample collection at the Columbus site, and B.J. Butler for sample collection at Borden.
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

166 O.L. Snoeyenbos-West et al.
References
1. Achenbach LM, Woese C (1995) 16S and 23S rRNA-like prim- ers. In: Robb FT, Place AR (eds) Archaea, a laboratory manual, Thermophiles. Cold Spring Harbor Laboratory Press, New York, pp 201-203
2. Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA (1990) Combination of 16S rRNA-targeted oligo- nucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl Environ Microbiol 56:1919-1925
3. Amann RI, Ludwig W, Schleifer K-H (1995) Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev 59:143-169
4. Anderson RT, Lovley DR (1999) Naphthalene and benzene degradation under Fe(III)-reducing conditions in petroleum- contaminated aquifers. Bioremediation J 3:121-135
5. Anderson RT, Rooney-Varga J, Gaw CV, Lovley DR (1998) Anaerobic benzene oxidation in the Fe(III)-reduction zone of petroleum-contaminated aquifers. Environ Sci Technol 32:1222-1229
6. Boggs JF, Young SC, Beard LM (1992) Field study of disper- sion in a heterogeneous aquifer, 1. Overview and site descrip- tion. Wat Resour Res 28:3281-3291
7. Boone DR, Liu Y, Zhao Z-J, Balkwill DL, Drake GT, Stevens TO, Aldrich HC (1995) Bacillus infernus sp nov, an Fe(III)- and Mn(IV)-reducing anaerobe from the deep terrestrial sub- surface. Int J Sys Bacteriol 45:441-448
8. Caccavo Jr F, Blakemore RP, Lovley DR (1992) A hydrogen- oxidizing, Fe(III)-reducing microorganism from the Great Bay Estuary, New Hampshire. Appl Environ Microbiol 58:3211- 3216
9. Caccavo Jr F, Coates JD, Rossello-Mora RA, Ludwig W, Schleifer KH, Lovley DR, McInerney MJ (1996) Geovibrio fer- rireducens, a phylogenetically distinct dissimilatory Fe(III)- reducing bacterium. Arch Microbiol 165:370-376
10. Coates JD, DJ, Ellis DR, Lovley (1999) Geothrixfermentans gen nov sp nov, an acetate-oxidizing Fe(III) reducer capable of growth via fermentation. Internat J Sys Bacteriol 49:1615- 1622
11. Coates JD, Lonergan DJ, Jenter H, Lovley DR (1996) Isolation of Geobacter species from diverse sedimentary environments. Appl Environ Microbiol 62:1531-1536
12. Cummings DE, Caccavo Jr F, Spring S, Rosenzweig RF (1999) Ferribacterium limneticum, gen nov, sp nov, an Fe(III)- reducing microorganism isolated from mining-impacted freshwater lake sediments. Arch Microbiol 17:183-188
13. Fantroussi SA, Verschuere 1, Verstraete W, Top EA (1999) Effect of phenylurea herbicides on soil microbial communities estimated by analysis of 16S rRNA gene fingerprints and com- munity-level physiological profiles. Appl Environ Microbiol 65:982-988
14. Fenchel T, Esteban GF, Finlay BJ (1997) Local versus global diversity of microorganisms: Cryptic diversity of ciliated pro- tozoa. Oikos 80:220-225
15. Finlay BJ, Clarke KL (1999) Ubiquitous dispersal of microbial species. Nature 400:828
16. Fuhrman JA, McCallum K, Davis AA (1993) Phylogenetic di- versity of subsurface marine microbial communities from the Atlantic and Pacific Oceans. Appl Environ Microbiol 59:1294- 1302
17. Giovannoni SJ, Britschgi TB, Moyer C, Field KG (1990) Ge- netic diversity in Sargasso Sea bacterioplankton. Nature 345:60-63
18. Kopczynski ED, Bateson AM, Ward DM (1994) Recognition of chimeric small-subunit ribosomal DNAs composed of genes from uncultivated microorganisms. AppI Environ Mi- crobiol 60:746-748
19. Kuske CR, Barns SM, Busch JD (1997) Diverse uncultivated bacterial groups from soils of the arid southwestern United States that are present in many geographic regions. Appl En- viron Microbiol 63:3614-3621
20. Lane DL, Pace B, Olsen GJ, Stahl D, Sogin ML, Pace NR (1985) Rapid determination of 16S ribosomal RNA sequences for phylogenetic analysis. Proc Nat Acad Sci USA 82:6955- 6959
21. Lonergan DJ, Jenter H, Coates JD, Phillips EJP, Schmidt T, Lovley DR (1996) Phylogenetic analysis of dissimilatory Fe(III)-reducing bacteria. J Bacteriol 178:2402-2408
22. Lovley DR (1995) Bioremediation of organic and metal con- taminants with dissimilatory metal reduction. J Industr Mi- crobiol 14:85-93
23. Lovley DR (1991) Dissimilatory Fe(I1I) and Mn(IV) reduc- tion. Microbiol Rev 55:259-287
24. Lovley DR (2000) Fe(III) and Mn(IV) reducing prokaryotes. In: Dworkin M, Falkow S, Rosenberg E, and Stackebrandt E (eds) The Prokaryotes, Springer-Verlag, New York
25. Lovley DR (1995) Microbial reduction of iron, manganese, and other metals. Adv Agron 54:175-231
26. Lovley DR (1997) Potential for anaerobic bioremediation of BTEX in petroleum-contaminated aquifers. J Industr Micro- biol 18:75-81
27. Lovley DR, Baedecker MJ, Lonergan DJ, Cozzarelli IM, Phil- lips EJP, Siegel DI (1989) Oxidation of aromatic contaminants coupled to microbial iron reduction. Nature 339:297-299
28. Lovley DR, Chapelle FH, Woodward JC (1994) Use of dis- solved H2 concentrations to determine the distribution of mi- crobially catalyzed redox reactions in anoxic ground water. Environ Sci Technol 28:1205-1210
29. Lovley DR, Coates JD (1997) Bioremediation of metal con- tamination. Curr Opin Biotech 8:285-289
30. Lovley DR, Coates JD, Blunt-Harris EL, Phillips EJP, Wood- ward JC (1996) Humic substances as electron acceptors for microbial respiration. Nature 382:445-448
31. Lovley DR, Fraga JL, Blunt-Harris EL, Hayes LA, Phillips EJP, Coates JD (1998) Humic substances as a mediator for micro- bially catalyzed metal reduction. Acta Hydrochim Hydrobiol 26:152-157
32. Lovley DR, Goodwin 5 (1988) Hydrogen concentrations as an indicator of the predominant terminal electron accepting re-
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions

Subsurface Fe(III)-Reducing Community Structure 167
actions in aquatic sediments. Geochim Cosmochim Acta 52:2993-3003
33. Lovley DR, Phillips EJP (1986) Organic matter mineralization with reduction of ferric iron in anaerobic sediments. Appl Environ Microbiol 51:683-689
34. Lovley DR, Phillips EJP (1992) Bioremediation of uranium contamination with enzymatic uranium reduction. Environ Sci Technol 26:2228-2234
35. Lovley DR, Phillips EJP (1989) Requirement for a microbial consortium to completely oxidize glucose in Fe(IIJ)-reducing sediments. Appl Environ Microbiol 55:3234-3236
36. Lovley DR, Phillips EJP, Gorby YA, Landa ER (1991) Micro- bial reduction of uranium. Nature 350:413-416
37. Lovley DR, Phillips EJP, Lonergan DJ (1989) Hydrogen and formate oxidation coupled to dissimilatory reduction of iron or manganese by Alteromonas putrefaciens. Appl Environ Mi- crobiol 55:700-706
38. Lovley DR, Woodward JC (1996) Mechanisms for chelator stimulation of microbial Fe(IlI)-oxide reduction. Chem Geol 132:19-24
39. Lovley DR, Woodward JC, Chapelle FH (1996) Rapid anaero- bic benzene oxidation with a variety of chelated Fe(III) forms. Appl Environ Microbiol 62:288-291
40. Lovley DR, Woodward JC, Chapelle FH (1994) Stimulated anoxic biodegradation of aromatic hydrocarbons using Fe(III) ligands. Nature 370:128-131
41. Maclntyre WG, Boggs M, Antworth CP, Stauffer TB (1993) Degradation kinetics of aromatic organics solutes introduced into a heterogeneous aquifer. Water Resour Res 20:4045-4051
42. Maidak BL, Cole JR, Charles T, Parker J, Garrity GM, Larsen N, Li B, Lilburn TG, McCaughey MJ, Olsen GJ, Overbeek R, Pramanik S, Schmidt TM, Tiedje JM, Woese CR (1999) A new version of the RDP (Ribosomal Database Project). Nucl Acid Res 27:171-173
43. Methe BA, Hiorns WD, Zehr JP (1998) Contrasts between marine and freshwater bacterial community composition: Analyses of communities in Lake George and six other lakes. Limnol Oceanog 43:368-374
44. Muyzer G (1999) DGGE/TGGE, a method for identifying
genes from natural ecosystems. Curr Opin Microbiol 2:317- 322
45. Muyzer G, Wall ECD, Uitterlinden G (1993) Profiling of com- plex microbial populations by denaturing gradient gel electro- phoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695-700
46. Nevin KP, Lovley DR (2000) Potential for nonenzymatic re- duction of Fe(IJJ) during microbial oxidation of organic mat- ter coupled to Fe(III) reduction. Environ. Sci. Technol.
47. Picard C, Ponsonnet C, Paget E, Nesme X, Simonet P (1992) Detection and enumeration of bacteria in soil by direct DNA extraction and polymerase chain reaction. Appl Environ Mi- crobiol 58:2717-2722
48. Raskin L, Stromley JM, Rittmann BE, Stahl DA (1994) Group- specific 16S rRNA hybridization probes to describe natural communities of methanogens. Appl and Environ Microbiol 60:1232-1240
49. Roberts MS, Cohan FM (1995) Recombination and migration rates in natural populations of Bacillus subtilis and Bacillus mojavensis. Evolution 49:1081-1094
50. Rooney-Varga JN, Anderson RT, Fraga JL, Ringelberg D, Lov- ley DR (1999) Microbial communities associated with anaero- bic benzene mineralization in a petroleum-contaminated aquifer. Appl Environ Microbiol 65:3056-3063
51. Swofford DL (1998) PAUP*, Phylogenetic Analysis Using Par- simony (* and other methods) Version 4. Sinauer Associates, Sunderland, MA
52. Van de Peer Y, De Wachter Y (1994) TREECON for Windows: A software package for the construction and drawing of evo- lutionary trees for the Microsoft Windows environment. Comput Applic Biosci 10:569-570
53. Vargas M, Kashefi K, Blunt-Harris EL, Lovley DR (1998) Mi- crobiological evidence for Fe(IIJ) reduction on early Earth. Nature 395:65-67
54. Zwart G, Hiorns WD, Methe BA, Agterveld MP, Huismans R, Nold SC, Zehr JP, Laanbroek HJ (1998) Near-identical 16S rRNA sequences recovered from lakes in North America and Europe indicate the existence of clades of freshwater bacteria with global distribution. Syst Appl Microbiol 21:546-556
This content downloaded from 128.119.49.10 on Fri, 18 Sep 2015 16:35:12 UTCAll use subject to JSTOR Terms and Conditions
Related Documents











