Enhancing biomass production and yield by maintaining enhanced capacity for CO 2 uptake in response to elevated CO 2 Keshav Dahal 1 , Sarathi M. Weraduwage 2 , Khalil Kane 3 , Shezad A. Rauf 2 , Evangelos D. Leonardos 2 , Winona Gadapati 1 , Leonid Savitch 4 , Jas Singh 4 , Elizabeth-France Marillia 5 , David C. Taylor 5 , Malgre C. Micallef 2 , Vicki Knowles 6 , William Plaxton 6 , John Barron 7 , Fathey Sarhan 3 , Norman Hu ¨ner 1 , Bernard Grodzinski 2,8 , and Barry J. Micallef 2 1 Department of Biology and the Biotron Centre for Experimental Climate Change Research, University of Western Ontario, London, Ontario, Canada N6A 5B7; 2 Department of Plant Agriculture, University of Guelph, Guelph, Ontario, Canada N1G 2W1; 3 Department of Biological Sciences, Universite ´ du Que ´ bec a ` Montre ´ al, Montreal, Quebec, Canada H3C 3P8; 4 Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, Ottawa, Ontario, Canada K1A OC6; 5 National Research Council of Canada, 110, Gymnasium Place, Saskatoon, Saskatchewan, Canada S7N 0W9; 6 Department of Biology, Queen’s University, Kingston, Ontario, Canada K7L 3N6; and 7 Department of Computer Science, University of Western Ontario, London, Ontario, Canada N6A 5B7. Received 9 May 2013, accepted 11 March 2014. Published on the web 10 April 2014. Dahal, K., Weraduwage, S. M., Kane, K., Rauf, S. A., Leonardos, E. D., Gadapati, W., Savitch, L., Singh, J., Marillia, E.-F., Taylor, D. C., Micallef, M. C., Knowles, V., Plaxton, W., Barron, J., Sarhan, F., Hu¨ner, N., Grodzinski, B. and Micallef, B. J. 2014. Enhancing biomass production and yield by maintaining enhanced capacity for CO 2 uptake in response to elevated CO 2 . Can. J. Plant Sci. 94: 10751083. Using four model plants, two members of the Gramineae, rye and wheat, and two Brassicaceae, Brassica napus and Arabidopsis thaliana, two fundamental approaches were exploited to determine how regulating source-sink development would alter photosynthesis, productivity and yield during long-term acclimation to elevated CO 2. In one approach we exploited the cold acclimation response of winter wheat, rye and B. napus. In the other approach we modified the dark respiration in A. thaliana to alter availability of respiratory substrates required for anabolic processes, such as fatty acid metabolism, thus reducing sink limitations on canopy photosynthesis at elevated CO 2 . Taken together, the data show the importance of maintaining strong demand from active sinks when the above- ground canopy is being exposed to elevated levels of the primary substrate of photosynthesis, CO 2 . Key words: Carbon dioxide enrichment, productivity, photosynthesis, respiration, cereals, Brassicaceae Dahal, K., Weraduwage, S. M., Kane, K., Rauf, S. A., Leonardos, E. D., Gadapati, W., Savitch, L., Singh, J., Marillia, E.-F., Taylor, D. C., Micallef, M. C., Knowles, V., Plaxton, W., Barron, J., Sarhan, F., Hu¨ner, N., Grodzinski, B. et Micallef, B. J. 2014. Accroissement de la production et du rendement de la biomasse par le maintien d’une meilleure capacite´ d’absorption du CO 2 dans un milieu plus riche en CO 2 . Can. J. Plant Sci. 94: 10751083. Recourant a` quatre mode`les botaniques, deux gramine´es (le seigle et le ble´) et deux brassicace´es (Brassica napus et Arabidopsis thaliana), les auteurs ont applique´ deux approches fondamentales en vue d’e´tablir comment on pourrait modifier la photosynthe`se, la productivite´ et le rendement des plantes en re´gulant le de´veloppement de sources et de puits de CO 2 , advenant une acclimatation a` long terme a` une concentration e´ leve´ e de ce gaz. Dans la premie` re approche, les auteurs ont exploite´ la re´ action du ble´ d’hiver, du seigle et de B. napus a` l’acclimatation au froid; avec la seconde, ils ont modifie´ la respiration mitochondriale chez A. thaliana afin d’alte´rer la disponibilite´ des substrats respiratoires ne´cessaires a` des proce´de´s anaboliques comme le me´tabolisme des acides gras, ce qui a eu pour effet d’atte´nuer les restrictions de captage lors de la photosynthe`se par la ve´ge´tation, dans un milieu riche en CO 2 . Collectivement, ces donne´ es re´ ve` lent qu’il est faut maintenir la forte demande des puits actifs quand les organes ae´riens des plantes sont expose´s a` une concentration e´leve´e de CO 2 , principal substrat de la photosynthe`se. Mots cle ´s: Hausse du dioxyde de carbone, productivite´, photosynthe`se, respiration, ce´re´ales, Brassicaceae 8 Corresponding author (e-mail: [email protected]). Abbreviations: CA, cold acclimated; mtPDH, mitochondrial pyruvate dehydrogenase; mtPDHK, mitochondrial pyruvate dehydrogenase kinase; dehydrogenase; NA, non-acclimated; NPQ, nonphotochemical dissipation; plPDH, chloroplastic isozyme of PDH; TCA, tricarboxylic acid Can. J. Plant Sci. (2014) 94: 10751083 doi:10.4141/CJPS2013-151 1075 Can. J. Plant Sci. Downloaded from pubs.aic.ca by Agriculture and Agri-food Canada on 07/09/14 For personal use only.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Enhancing biomass production and yield by maintainingenhanced capacity for CO2 uptake in response to
elevated CO2
Keshav Dahal1, Sarathi M. Weraduwage2, Khalil Kane3, Shezad A. Rauf2,Evangelos D. Leonardos2, Winona Gadapati1, Leonid Savitch4, Jas Singh4,
Elizabeth-France Marillia5, David C. Taylor5, Malgre C. Micallef2, Vicki Knowles6,William Plaxton6, John Barron7, Fathey Sarhan3, Norman Huner1,
Bernard Grodzinski2,8, and Barry J. Micallef2
1Department of Biology and the Biotron Centre for Experimental Climate Change Research, University ofWestern Ontario, London, Ontario, Canada N6A 5B7; 2Department of Plant Agriculture, University of Guelph,Guelph, Ontario, Canada N1G 2W1; 3Department of Biological Sciences, Universite du Quebec a Montreal,
Montreal, Quebec, Canada H3C 3P8; 4Eastern Cereal and Oilseed Research Centre, Agriculture andAgri-Food Canada, Ottawa, Ontario, Canada K1A OC6; 5National Research Council of Canada, 110,
Gymnasium Place, Saskatoon, Saskatchewan, Canada S7N 0W9; 6Department of Biology,Queen’s University, Kingston, Ontario, Canada K7L 3N6; and 7Department of Computer Science,
University of Western Ontario, London, Ontario, Canada N6A 5B7.Received 9 May 2013, accepted 11 March 2014. Published on the web 10 April 2014.
Dahal, K., Weraduwage, S. M., Kane, K., Rauf, S. A., Leonardos, E. D., Gadapati, W., Savitch, L., Singh, J., Marillia,E.-F., Taylor, D. C., Micallef, M. C., Knowles, V., Plaxton, W., Barron, J., Sarhan, F., Huner, N., Grodzinski, B. andMicallef, B. J. 2014. Enhancing biomass production and yield by maintaining enhanced capacity for CO2 uptake in response to
elevated CO2. Can. J. Plant Sci. 94: 1075�1083. Using four model plants, two members of the Gramineae, rye and wheat,and two Brassicaceae, Brassica napus and Arabidopsis thaliana, two fundamental approaches were exploited to determinehow regulating source-sink development would alter photosynthesis, productivity and yield during long-term acclimationto elevated CO2. In one approach we exploited the cold acclimation response of winter wheat, rye and B. napus. In theother approach we modified the dark respiration in A. thaliana to alter availability of respiratory substrates required foranabolic processes, such as fatty acid metabolism, thus reducing sink limitations on canopy photosynthesis at elevatedCO2. Taken together, the data show the importance of maintaining strong demand from active sinks when the above-ground canopy is being exposed to elevated levels of the primary substrate of photosynthesis, CO2.
Key words: Carbon dioxide enrichment, productivity, photosynthesis, respiration, cereals, Brassicaceae
Dahal, K., Weraduwage, S. M., Kane, K., Rauf, S. A., Leonardos, E. D., Gadapati, W., Savitch, L., Singh, J., Marillia,E.-F., Taylor, D. C., Micallef, M. C., Knowles, V., Plaxton, W., Barron, J., Sarhan, F., Huner, N., Grodzinski, B. etMicallef, B. J. 2014. Accroissement de la production et du rendement de la biomasse par le maintien d’une meilleure capacited’absorption du CO2 dans un milieu plus riche en CO2. Can. J. Plant Sci. 94: 1075�1083. Recourant a quatre modelesbotaniques, deux graminees (le seigle et le ble) et deux brassicacees (Brassica napus et Arabidopsis thaliana), les auteurs ontapplique deux approches fondamentales en vue d’etablir comment on pourrait modifier la photosynthese, la productivite etle rendement des plantes en regulant le developpement de sources et de puits de CO2, advenant une acclimatation a longterme a une concentration elevee de ce gaz. Dans la premiere approche, les auteurs ont exploite la reaction du ble d’hiver, duseigle et de B. napus a l’acclimatation au froid; avec la seconde, ils ont modifie la respiration mitochondriale chez A. thalianaafin d’alterer la disponibilite des substrats respiratoires necessaires a des procedes anaboliques comme le metabolisme desacides gras, ce qui a eu pour effet d’attenuer les restrictions de captage lors de la photosynthese par la vegetation, dans unmilieu riche en CO2. Collectivement, ces donnees revelent qu’il est faut maintenir la forte demande des puits actifs quand lesorganes aeriens des plantes sont exposes a une concentration elevee de CO2, principal substrat de la photosynthese.
Mots cles: Hausse du dioxyde de carbone, productivite, photosynthese, respiration, cereales, Brassicaceae
8Corresponding author (e-mail: [email protected]).
Abbreviations: CA, cold acclimated; mtPDH, mitochondrialpyruvate dehydrogenase; mtPDHK, mitochondrial pyruvatedehydrogenase kinase; dehydrogenase; NA, non-acclimated;NPQ, nonphotochemical dissipation; plPDH, chloroplasticisozyme of PDH; TCA, tricarboxylic acid
Can. J. Plant Sci. (2014) 94: 1075�1083 doi:10.4141/CJPS2013-151 1075
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

The primary goal of our group within the Green CropNetwork is described in the overview article by Smith andZhou (2014; this issue). The overall goal of Theme 3 wasto provide a genetic andmetabolic blueprint for enhancedplant carbon storage and biomass production through anenhanced capacity for CO2 uptake in response to elevatedCO2 that is associated with global warming. To addressthis goal, two fundamentally different experimentalapproaches with different plant material were exploitedto assess how source-sink interactions during CO2 enrich-ment may lead to enhanced plant productivity. The firstapproach exploited the known enhancement inCO2 assim-ilation and biomass production induced by cold acclima-tion of winter cereals, such as wheat and rye as well asBrassica napus under ambient CO2 conditions (Hurryand Huner 1991; Boese and Huner 1992; Oquist et al.2003; Hurry et al. 1994, 1995; Huner et al. 1998; Savitchet al. 2002; Rapacz et al. 2008). The second approachexploited the enhancement of seed oil yield previouslyfound at ambient CO2 in genetic variants of Arabidopsisthaliana altered in respiratory metabolism (Zou et al.1999; Marilla et al. 2003). We hypothesized that thisgenetic modification would reduce sink limitations onphotosynthesis and growth at elevated CO2 throughgreater availability of respiratory substrates requiredfor anabolic processes such as fatty acid synthesis inseeds.
THE EFFECTS OF COLD ACCLIMATION ONPHOTOSYNTHESIS AND PRODUCTIVITY AT
HIGH CO2
During cold acclimation of cold-tolerant cultivars andspecies, increased photosynthetic capacity is, in part,the result of the enhanced activities of key regulatoryenzymes of primary carbon metabolism such as Rubisco,cFBPase, and SPS (Hurry et al. 1995, 2000; Stitt andHurry 2002) and the concomitant export to sinks duringcold acclimation (Leonardos et al. 2003). Although springcereals can grow and develop at low temperatures, theyexhibit decreased photosynthetic capacity estimated aslight-saturated rates of carbon assimilation, Asat, in re-sponse to low temperatures in contrast to winter cultivars(Hurry and Huner 1991). Because spring cultivars exhibita limited sink demand coupled with a retardation ofcarbon export from photosynthetically active sourcetissue to plant sink tissue (Hurry et al. 1995), this resultsin inhibition of cytosolic sucrose biosynthesis, whichleads to Pi limitations within the chloroplast which, inturn, decreases the utilization of stromal phosphorylatedintermediates of the Calvin�Benson cycle. This results inan inhibition of CO2 assimilation due to feedback-limitedphotosynthesis (Hurry et. al. 1995; Savitch et al. 2002).Therefore, spring cultivars exhibit decreased plasticity tolow growth temperatures and exhibit reduced photosyn-thetic capacity during cold acclimation (Hurry et al.1995; Savitch et al. 2002). Winter cultivars overcome the
potential for low-temperature-induced feedback inhibi-tion of photosynthesis by the maintenance of high sinkcapacity, which maintains a high flux of carbon exportfrom the source.
Is the enhanced photosynthetic capacity exhibited bycold-acclimated winter cultivars under ambient CO2
conditions related to an enhanced efficiency for lightenergy conversion into biomass and seed production?A major focus of photosynthesis research continues tobe the enhancement of crop yield coupled with minimalincreases in energy inputs. Classical breeding has beenused in attempts to maximize photosynthetic efficiencythrough alteration of canopy structure to minimize lightsaturation effects on individual leaves (Zhu et al. 2010).Under light saturated conditions, photosynthetic effi-ciency is minimal, and plants protect the photosyntheticapparatus from the potential damaging effects of excesslight by dissipating the excess absorbed energy as heatthrough induction of the xanthophyll cycle (Ort 2001).The evolution of the xanthophyll cycle to regulatenonphotochemical dissipation (NPQ) of excess absorbedlight is considered essential for the survival of sun plantsto fluctuations in irradiance (Demmig-Adams andAdams 1992). Enhanced rates of photosynthesis shoulddecrease the propensity for energetically wasteful dis-sipation of absorbed light has heat. Dahal et al. (2012c,2013) reported that cold acclimation of winter wheatand rye increased the quantum requirement to closePSII reaction centers with a concomitant increase in thequantum requirement to induce NPQ relative to non-acclimated plants. This means that it is more difficult toclose PSII reaction centers in cold acclimated versus non-acclimated plants. This is consistent with the increasedflux of electrons through photosynthetic electron trans-port, the enhanced rates of CO2 assimilation and thedecreased light sensitivity of NPQ in cold acclimatedversus nonacclimated plants (Dahal et al. 2012c, 2013).This is not only coupled to an increase in the content ofmajor photosynthetic components (e.g., Rubisco, psaB,psaA, cFBPase) per unit chlorophyll, but also a decreasein the low temperature sensitivities of CO2 assimilation aswell as photosynthetic electron transport (Dahal et al.2012a,b, 2013). Thus, cold acclimation increases energyconversion efficiency at the expense of NPQ such thatmore absorbed energy is trapped as fixed carbon andsubsequently converted into biomass. Interestingly, coldacclimation of spring wheat and spring rye decreasesenergy conversion efficiency, which is coupled to in-creased dissipation of absorbed light as NPQ (Dahalet al. 2012a, 2013). Furthermore, this differential capa-city to modulate energy conversion efficiency is trans-lated into a 40% increase in grain yield (grams of seeds/plant) in winter wheat than spring wheat (Dahal et al.2013).
1076 CANADIAN JOURNAL OF PLANT SCIENCE
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

Can cold acclimated plants maintain this enhancedcapacity for CO2 assimilation and biomass productionduring long-term growth under elevated CO2
conditions?Although an immediate increase in the rates of netCO2 assimilation has been observed following a short-term shift of C3 plants from ambient to elevated CO2 dueto enhanced carboxylation velocity (Cheng et al. 1998;Long et al. 2004; Ainsworth and Rogers 2007), long-termgrowth and development of C3 plants at high CO2 concen-tration may lead to an end product inhibition of photo-synthetic capacity due to accumulation of non-structuralcarbohydrates in the cytosol (Stitt and Quick 1989; Foyer1990). At elevated CO2, photosynthesis is usually limitedeither by the capacity of photosynthetic electron trans-port to supply ATP and NADPH to regenerate RuBP,or by the capacity of starch and sucrose synthesis toutilize triose phosphates and consequently regenerate Pi.
The Pi regeneration-limited photosynthesis is governedby the balance between the source leaves to assimilatecarbon and the sink strength to utilize photoassimilates(Arp 1991; Drake et al. 1997). It has been suggested thatthe increased carbon uptake resulting from initial stim-ulation of photosynthesis alters the balance betweensupply and demand due to limited sink capacity to utilizecarbohydrates and concomitant retardation of carbonexport to the sinks (Kramer 1981; Arp 1991; Drake et al.1997). This results in the accumulation of sucrose in thesource leaves followed by inhibition of sucrose synthesisand a short-term decrease in utilization of phosphorylatedintermediates and depletion in stromal Pi. Low avail-ability of stromal Pi triggers inhibition of ATP synthesisand thereby a decrease in the rate at which PGA is con-verted to triose phosphate, which results in feedbackinhibition of CO2 assimilation (Stitt and Quick 1989;Sharkey and Vaderveer 1989). In the long-term, thefeedback inhibition of photosynthesis may lead to thedownregulation of the expression of key regulatoryenzymes of photosynthetic carbon metabolism (Drakeet al. 1997; Moore et al. 1999).
We hypothesized that the cold acclimation-inducedincrease in photosynthetic performance of winter wheatand winter rye at ambient CO2 is maintained under long-term growth and development at elevated CO2. Consis-tent with our hypothesis, the low-temperature-inducedincrease in photosynthetic capacity of winter cultivars,Norstar andMusketeer, at ambient CO2, was maintainedunder growth and development of plants at elevated CO2
(Dahal et al. 2012). In fact, the cold acclimated (CA)winter cultivars exhibited about 1.4-fold higher Asat thannon-acclimated (NA) controls when grown at elevatedCO2. This 1.4-fold enhancement of photosynthetic capa-city in CA versus NA winter cultivars at elevated CO2
was associated with increased SLW, and, as a conse-quence, an increase in total leaf protein content andsubsequent levels of major photosynthetic enzymes such
as Rubisco (rbcL), cFBPase and components of photo-synthetic electron transport, Lhcb1, PsbA and PsaA on aleaf area basis upon cold acclimation (Dahal et al., inpreparation). These were consistent with increased quan-tum requirements to close PSII reaction centers as wellas to induce energy dissipation by NPQ coupled with alower Ci requirement to open PSII reaction centers anda lower propensity to dissipate absorbed energy throughNPQ under CO2 saturated conditions in CA versus NAwinter cultivars irrespective of growth CO2. This indi-cates that compared with NAwinter cultivars, CA wintercultivars grown at elevated CO2 maintain an enhancedefficiency to utilize absorbed light energy and convert itto biomass with a concomitant decrease in dissipationof absorbed energy through NPQ. Furthermore, theenhanced energy conversion efficiency is translated into agreater increase in grain yield (grams of seeds/plant) inwinter wheat than spring wheat, even during long-termgrowth at elevated CO2 (Dahal et al., in preparation).
Can the target genes that govern the cold acclimation-induced enhancement in photosynthetic capacity andbiomass production be identified?We hypothesized, first, that the cold acclimation-inducedincrease in photosynthetic performance of B. napus atambient CO2 is maintained under long-term growth anddevelopment at elevated CO2 similar to that observed forCA winter cereals (Dahal et al. 2012a,b). Second, basedon our previous results (Savitch et al. 2005), we hypothe-sized that the BnCBF17 over-expressor grown at 208Cshould respond similarly to growth and developmentunder elevated CO2 as does cold acclimatedWTB. napus.The recent results of Dahal et al. (2012c) indicate thatthe over-expression of BnCBF17 mimics the effects ofcold acclimation of B. napus with respect to photosyn-thetic performance, the efficiency of energy conversionand WUE, which is still maintained even after long-termgrowth and development under elevated CO2 conditions.Neither cold acclimated Brassica nor the BNCBF17over-expressor exhibited feedback inhibition of photo-synthesis during long-term growth and development atelevated CO2. Thus, we suggest that the transcriptionfactor, BnCBF17, may be a central component whichgoverns the regulation of photosynthetic capacity andenergy conversion efficiency of crop plants at ambient aswell as at elevated CO2 conditions.
Can this information be used to genetically modifyplants to induce enhanced photosynthetic capacity andbiomass production without the requirement for coldacclimation?We suggest that our results for over-expression ofBNCBF17 inB. napus (Dahal et al. 2012c) provide impor-tant new insights into potential molecular and geneticapproaches focussed on the maintenance or even the en-hancement of plant productivity under suboptimal growth
DAHAL ET AL. * BIOMASS PRODUCTION AND ELEVATED CO2 1077
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

conditions associated with climate change independent ofthe requirement for cold acclimation. Thus, we suggestthat CBFs/DREBs are critical transcription factors thatgovern plant phenotypic plasticity associated with coldacclimation from the level of gene expression and freezingtolerance to whole plant architecture, WUE as well asphotosynthetic energy conversion efficiency into biomassand, ultimately, seed production. Based on our data,over-expression of CBFs circumvents the need for coldacclimation to induce enhanced CO2 assimilation rates,energy conversion efficiency and increased biomass pro-duction in important crop plants such as wheat, rye andB. napus.
THE EFFECT OF RESPIRATORY METABOLISMON SOURCE-SINK BALANCE AT ELEVATED CO2:LESSONS LEARNED FROM GENETIC VARIANTS
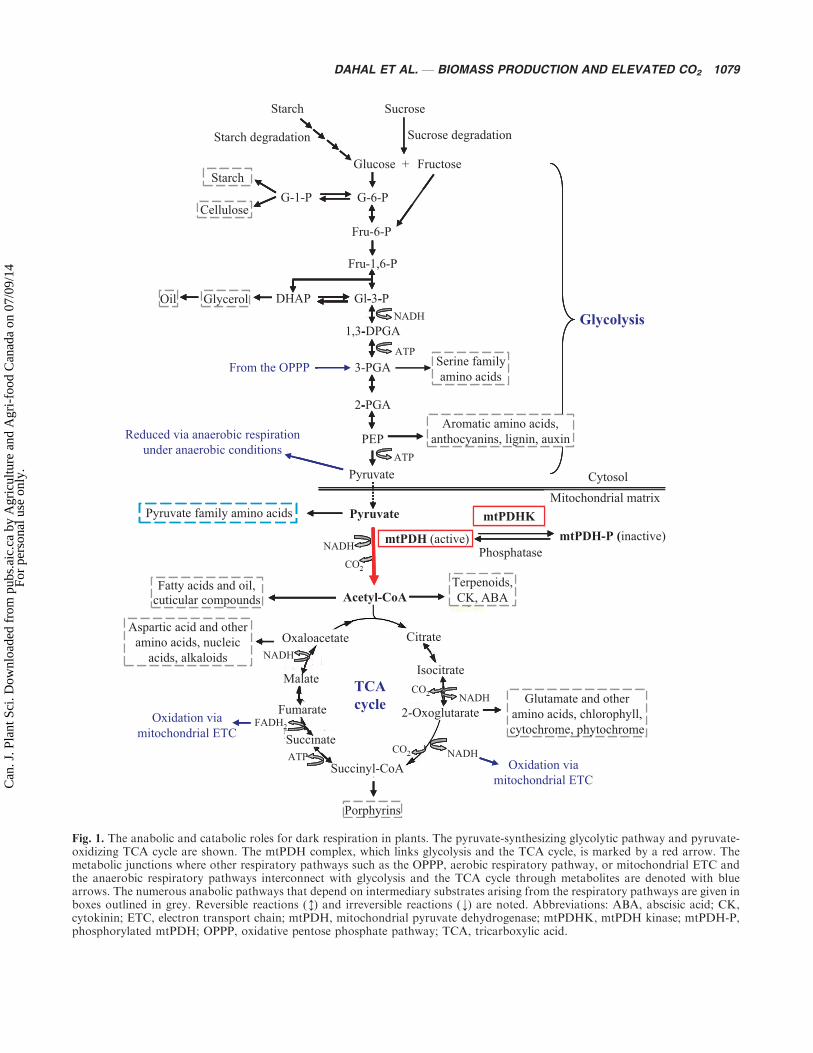
OF ARABIDOPSIS THALIANA ALTERED INMITOCHONDRIAL PYRUVATE DEHYDROGENASEAt elevated atmospheric CO2 as predicted for this century,plant productivity is likely to be limited by the devel-opment and metabolism of sink-tissues (sink-strength),causing feedback effects on photosynthesis (source-strength) (Grodzinski 1992; Morgan et al. 2005; Ainsworthand Rogers 2007). Studies on plants with large sinks, andexperimental manipulations of sinks, have implicatedsink strength as an important factor in maintaining highphotosynthetic rates in plants under CO2-enrichment(Ainsworth et al. 2004). Thus far, studies examiningthe influence of metabolism on sink strength at elevatedCO2 have focused on anabolic processes such as starchsynthesis and nitrogen assimilation (Smidansky et al.2002; Stitt and Krapp 1999). Dark respiration has rarelybeen considered a promising target for increasing sinkstrength and productivity (Long et al. 2006), in part sincedark respiration is associated with catabolism (Fig. 1).Studies at ambient CO2 have demonstrated correlationsbetween decreased respiratory CO2 release and enhancedproductivity (Winzeler et al. 1989).
However, an up-regulation of mitochondrial numbersandmitochondrial genes and proteins can occur in sourcetissues under high CO2 (Robertson et al. 1995; Leakeyet al. 2009).We believe that this respiratory adaptation toelevated CO2 reflects the crucial role of dark respirationin anabolic processes, such as fatty acid and protein syn-thesis (Fig. 1) (Weraduwage et al. 2011). Stated anotherway, we believe that an increased demand for intermedi-ary substrates derived from dark respiration like acetyl-CoA occurs at elevated CO2 that support enhanced plantgrowth (sink strength) and that need to be supplied toprevent feedback effects on photosynthesis (sourcestrength).
To examine the relationship between dark respirationand source-sink balance in plants at elevated CO2, weutilized genetic variants of Arabidopsis thaliana affectedinmitochondrial pyruvate dehydrogenase (mtPDH) activ-ity (Zou et al. 1999; Marilla et al. 2003). Mitochondrialpyruvate dehydrogenase catalyzes the oxidative decar-
boxylation of pyruvate to acetyl-CoA, thus linking gly-colysis with the tricarboxylic acid (TCA) cycle (Fig. 1)(Budde and Randall 1990; Marillia et al. 2003). Acetyl-CoA is required for the synthesis of fatty acids, a numberof plant growth regulators, and secondary metabolitesincluding terpenoids (Fig. 1) (Gemel and Randall 1992;Luethy et al. 1994;Marillia et al. 2003;Weraduwage et al.2011). Up-regulation of mtPDH protein does occur inwheat at elevated CO2 (Robertson et al. 1995).
To modify mtPDH activity in Arabidopsis thaliana weused antisense technology to reduce the activity of mito-chondrial pyruvate dehydrogenase kinase (mtPDHK)(Zou et al. 1999; Marillia et al. 2003). The protein phos-phorylation activity of mtPDHK is believed to deactivatemtPDH (Yeaman et al. 1978; Thelen et al. 2000; Tovar-Mendez et al. 2003). A chloroplastic isozyme of PDH(plPDH) also exists that is not regulated by phosphor-ylation (Patel and Korotchkina 2003). Arabidopsisthaliana transgenic lines showing antisense suppressionof mtPDHK do exhibit increased mtPDH activities inleaves, and an increased 100-seed oil content and 100-seed specific weight (i.e., enhanced sink activity in theseed), particularly in lines showing seed-specific expres-sion of the antisense construct, when grown at 22�258Cand ambient CO2 (Zou et al. 1999; Marillia et al. 2003).However, under these growing conditions overall plantbiomass and yield in the transgenic lines is either notenhanced or it decreases (Zou et al. 1999; Marillia et al.2003). We believe this occurs because source activity atambient CO2 cannot adapt sufficiently to the enhancedrespiratory activity in the mtPDHK transgenics. Expres-sion of mtPDHK in these transgenic lines has not beenexamined at elevated CO2.
Using four transgenic lines each of Arabidopsis thali-ana that express an antisense construct for mtPDHKin either a constitutive or seed-specific manner (eighttransgenic lines total) and three control lines [an un-transformed control (Wt) and a plasmid-only controlcorresponding to either the constitutive or seed-specificlines], we tested the following hypothesis: antisense sup-pression of mtPDHK in Arabidopsis thaliana will en-hance photosynthesis and overall plant productivity atelevated CO2 due to complementary increases in sourceand sink strength particularly for seed-specific lines.
Objective 1 � What is the effect of atmospheric CO2
levels on mtPDHK expression and the enzymeactivities of mtPDH and mtPDHK in leaves andseeds of Arabidopsis?Leaf mtPDHK transcript levels were lower in bothcontrol and constitutive transgenic lines at elevated CO2
relative to ambient CO2, and mtPDHK transcript levelsin leaves were lower in constitutive transgenic linesrelative to control lines at both ambient and elevatedCO2 (Weraduwage 2013). To quantify mtPDH andmtPDHK enzyme activities in leaves and seeds, twohigh-throughput in vitro assays using crude extracts were
1078 CANADIAN JOURNAL OF PLANT SCIENCE
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.
Mitochondrial matrix
Oxidation via mitochondrial ETC
Oxidation via mitochondrial ETC
CO2
GlycerolOil
mtPDH-P (inactive)Phosphatase
mtPDH (active)
mtPDHK
G-6-P
Fru-6-P
Fru-1,6-P
DHAP Gl-3-P
1,3-DPGA
3-PGA
2-PGA
PEP
Pyruvate
Acetyl-CoA
G-1-PStarch
Cellulose
Citrate
2-Oxoglutarate
Serine family amino acids
Aromatic amino acids, anthocyanins, lignin, auxin
Fatty acids and oil, cuticular compounds
TCA cycle
Glycolysis
Succinyl-CoA
Malate
Oxaloacetate
Porphyrins
Glutamate and other amino acids, chlorophyll, cytochrome, phytochrome
Aspartic acid and other amino acids, nucleic
acids, alkaloids
Fructose
Pyruvate Cytoplasm
NADH
ATP
ATP
NADH
NADH
NADHATP
CO2
Isocitrate
FADH2
NADH
From the OPPP
Reduced via anaerobic respiration under anaerobic conditions
Terpenoids, CK, ABA
Fumarate
Succinate
SucroseStarch
1
3
4, 5
6
12
2
8
9
10
11
13
14
15
16
1718
19
20
21
Glucose +
Starch degradation Sucrose degradation
1
7
CO2
CytosolMitochondrial matrix
Oxidation via mitochondrial ETC
Oxidation via mitochondrial ETC
CO2
GlycerolOil
mtPDH-P (inactive)Phosphatase
mtPDH (active)
mtPDHK
G-6-P
Fru-6-P
Fru-1,6-P
DHAP - -
-
3-PGA
-
Acetyl-CoA
G-1-PStarch
Cellulose
Citrate
2-Oxoglutarate
Serine family amino acids
Aromatic amino acids, anthocyanins, lignin, auxin
Fatty acids and oil, cuticular compounds
TCA cycle
Succinyl-CoA
Malate
Oxaloacetate
Porphyrins
Glutamate and other amino acids, chlorophyll, cytochrome, phytochrome
Aspartic acid and other amino acids, nucleic
acids, alkaloids
Fructose
Cytoplasm
NADH
ATP
NADH
NADH
NADHATP
CO2
Isocitrate
FADH2
NADH
From the OPPP
Reduced via anaerobic respiration under anaerobic conditions
Terpenoids, CK, ABA
Fumarate
Succinate
1
3
4, 5
6
12
2
8
9
10
11
13
14
15
16
1718
19
20
21
Glucose +
1
7
CO2
CytosolMitochondrial matrix
Oxidation via mitochondrial ETC
Oxidation via mitochondrial ETC
CO2
GlycerolOil
mtPDH-P (inactive)Phosphatase
mtPDH (active)
mtPDHK
G-6-P
Fru-6-P
Fru-1,6-P
- -
-
3-PGA
-
Acetyl-CoA
G-1-PStarch
Cellulose
Citrate
2-Oxoglutarate
Serine family amino acids
Aromatic amino acids, anthocyanins, lignin, auxin
Fatty acids and oil, cuticular compounds
TCA cycle
Succinyl-CoA
Malate
Oxaloacetate
Porphyrins
Glutamate and other amino acids, chlorophyll, cytochrome, phytochrome
Aspartic acid and other amino acids, nucleic
acids, alkaloids
Fructose
Cytoplasm
NADH
ATP
NADH
NADH
NADHATP
CO2
Isocitrate
FADH2
NADH
From the OPPP
Reduced via anaerobic respiration under anaerobic conditions
Terpenoids, CK, ABA
Fumarate
Succinate
1
3
4, 5
6
12
2
8
9
10
11
13
14
15
16
1718
19
20
21
Glucose +
1
7
CO2
Cytosol
Pyruvate family amino acids
Fig. 1. The anabolic and catabolic roles for dark respiration in plants. The pyruvate-synthesizing glycolytic pathway and pyruvate-oxidizing TCA cycle are shown. The mtPDH complex, which links glycolysis and the TCA cycle, is marked by a red arrow. Themetabolic junctions where other respiratory pathways such as the OPPP, aerobic respiratory pathway, or mitochondrial ETC andthe anaerobic respiratory pathways interconnect with glycolysis and the TCA cycle through metabolites are denoted with bluearrows. The numerous anabolic pathways that depend on intermediary substrates arising from the respiratory pathways are given inboxes outlined in grey. Reversible reactions (M) and irreversible reactions (¡) are noted. Abbreviations: ABA, abscisic acid; CK,cytokinin; ETC, electron transport chain; mtPDH, mitochondrial pyruvate dehydrogenase; mtPDHK, mtPDH kinase; mtPDH-P,phosphorylated mtPDH; OPPP, oxidative pentose phosphate pathway; TCA, tricarboxylic acid.
DAHAL ET AL. * BIOMASS PRODUCTION AND ELEVATED CO2 1079
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

developed that favor the mtPDH isozyme over plPDHbased both on precipitation of plPDH and optimizationof substrates and co-factors for mtPDH (Weraduwage2013). The mtPDHK activity assay examines the inacti-vation of mtPDH over time in the presence of 5 mMATP, and it allows the determination of contaminatingplPDH activity in crude extracts since plPDH is notphosphorylated like mtPDH (Weraduwage 2013). Theseassays show that mtPDH activities are enhanced andmtPDHK activities are reduced in leaves at elevated CO2
for transgenic lines relative to controls, which is con-sistent with changes in leaf mtPDHK transcript levels(Weraduwage 2013). Seed mtPDH activities are alsoenhanced in constitutive and seed-specific transgenic linescompared with controls at both ambient and elevatedCO2 (Weraduwage 2013). Collectively, the data supportboth an up-regulation of mtPDH activity in leaves andseeds at elevated CO2 relative to ambient CO2, and anenhancement of mtPDH activity at elevated CO2 in bothconstitutive and seed-specific transgenic lines showingantisense repression of mtPDHK relative to control lines.
Interestingly, at ambient CO2 maximal mtPDH activ-ity was exhibited by lines showing intermediate repres-sion of mtPDHK (Weraduwage 2013), which challengesthe notion that mtPDHK is strictly a negative regulatorof mtPDH activity (Yeaman et al. 1978; Thelen et al.2000; Tovar-Mendez et al. 2003). Thus, at ambient CO2
either a high or a low level of phosphorylation appears todecrease mtPDH activity, although this requires con-firmation using phospho-specific antibodies directed tomtPDH (Weraduwage 2013). We hypothesize that thisparabolic relationship between mtPDHK and mtPDHactivities at ambient CO2 reflects a role for mtPDHK inbalancing photosynthetic and respiratory processes inplants based on the energy status (e.g., ATP levels) of theplant, since ATP is a substrate for mtPDHK.
Objective 2 � Will sink activity be enhanced by arepression of mtPDHK?To examine the impact of mtPDHK expression on sinkactivity at ambient and elevated CO2, we quantified anumber of vegetative and reproductive growth parametersincluding total seed and oil production (Weraduwage2013). A number of growth and seed oil parameters wereimproved in transgenic lines, particularly at elevatedCO2;many of these parameters showed a significant linear orquadratic correlationwithmtPDHK transcript levels andmtPDH activity in the leaf and seed (Weraduwage 2013).We consistently found enhanced 100-seed oil con-tent and 100-seed specific weight for all transgenic linescompared with control lines as found in previous studies(Zou et al. 1999; Marillia et al. 2003). The greatest en-hancement in total seed andoil productivitywas found fortwoconstitutive lines at elevatedCO2 (up to2.8 times) thatalso exhibit a significant increase in inflorescence size, thelowest rate of mtPDH complex inactivation by ATP inleaves, and an intermediary enhancement of mtPDH
complex activity in seeds (Weraduwage 2013). Increasesin productivity at elevated CO2 for these two constitutivelines also correlate with altered developmental para-meters, including enhanced seed number per silique, en-hanced harvest index, and a larger inflorescence canopy(Weraduwage 2013). Three-dimensional laser scanningimaging also show that these two constitutive lines havelarger canopy areas, larger stem volumes, and a fastergrowth rate (Zhao 2009; Yang 2010). Interestingly, theenhanced productivity found for these two transgeniclines at elevatedCO2was greatest under lowhumidity andhigh plant density (Weraduwage 2013), suggesting thatmtPDHKmay be involved in stress adaptation in plants.These data show that sink activity at elevated CO2
can be altered through dosage-dependent expression ofmtPDHK.
Objective 3 � Is photosynthesis and respiratoryCO2 release altered at elevated CO2 for transgenicArabidopsis showing reduced expression of mtPDHK?We used both leaf-level and whole-plant gas exchangetechniques to examine the effects of altered mtPDHKexpression on net carbon exchange rates in the light anddark, to assess the impact of mtPDHK on source activityand to establish if respiratory CO2 release is increased ordecreased in Arabidopsis grown at elevated CO2 (Rauf2012; Weraduwage 2013). Gas exchange measurementsof individual rosette leaves demonstrated enhanced pho-tosynthesis for constitutive and seed-specific transgeniclines at both ambient and elevated CO2, providingevidence for reduced feedback effects on photosynthesisearly in plant development before a significant inflores-cence is established (Weraduwage 2013). An altered in-florescence canopy architecture described in Objective 2above for constitutive transgenic lines is significant, sincewe have also demonstrated clearly that the inflorescencein Arabidopsis contributes up to 90% of whole plant Cgain through photosynthesis at later stages of develop-ment (Rauf 2012). Thus, enhanced inflorescence devel-opment later in plant development significantly impactswhole-plant photosynthesis and carbon gain during rep-roductive development.
Measurements of net C exchange for both single leavesand the whole plant consistently showed that both photo-synthesis and respiratory CO2 release are enhanced atelevated CO2 for Arabidopsis (Rauf, 2012). The enhance-ment of respiratory CO2 release at elevated CO2 providesevidence for enhanced sink activity, and these data helpto resolve the controversy on whether plants grown atelevated CO2 show reduced or enhanced respiration(Grodzinski 1992; Drake et al. 1999; Leaky et al. 2009).These data are consistent with recent work by Leakeyet al. (2009) showing an up-regulation of dark respiratorygenes in source tissues under high CO2. The effect ofCO2 was greatest on photosynthesis and (growth andmaintenance) respiration at stages of development whenwe have very active sink development, first during the
1080 CANADIAN JOURNAL OF PLANT SCIENCE
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

leaf expansion phase and then at the silique-to-seed fillingstage (Rauf 2012). Possibly the effects on canopyarchitecture and stress response discussed above occurdue to alterations in synthesis of terpenoid-based plantgrowth regulators in transgenic plants showing alteredmtPDH activity and thus altered capacity for acetyl-CoAsynthesis.
Objective 4 � Is mtPDH is a viable target site forimproving crop productivity at elevated CO2?Our study is the first attempt at increasing dark respira-tion at elevated CO2 to enhance sink activity in plants,thus challenging the belief that decreased respiratory fluxwill enhance crop productivity (Weraduwage et al. 2011).Our definition of respiratory flux is the rate at whichcarbon enters the TCA cycle through mtPDH. Oncecarbon enters the TCA cycle, it can either be oxidized toCO2 or shunted into anabolic pathways (Fig. 1). Inter-estingly, we did not find differences in the rate of res-piratory CO2 release on a leaf area or dry matter basisfor transgenic lines compared with controls, althoughmtPDH activity was enhanced in transgenic lines (Rauf2012; Weraduwage 2013). We have shown previously byfeeding 14C-pyruvate to developing seeds that the fluxthrough mtPDH is increased in transgenic lines showingrepression of mtPDHK as evidenced by enhanced 14C-label in fatty acids (Marilla et al. 2003). It is possible thatPEP carboxylase has been enhanced in our transgeniclines owing to its anapleurotic role in replenishing TCAcycle intermediates (Weraduwage et al. 2011), thusreducing CO2 respiratory release in the transgenic lines.
By using transgenic Arabidopsis having repressedmtPDHK expression, we have shown that dark respira-tory processes play an important role in regulatingsource-sink balance in plants subjected to elevated CO2.Contrary to our original hypothesis, transgenic linesshowing constitutive and not seed-specific expression ofthe mtPDHK antisense construct showed the greatestimprovement inproductivityat elevatedCO2 (Weraduwage2013). This finding is consistent with our observationsthat these yield increases for constitutive lines correlatedwith a number of morphological parameters, includingsize of the inflorescence canopy, and thus a number oftissues did respond to altered mtPDHK expression inthe constitutive lines. We have discovered a method togreatly enhance both plant and oil productivity at ele-vated CO2 in Arabidopsis by directly increasing darkrespiratory flux to harness its anabolic properties in adosage-responsive manner. We believe this approach willinspire future studies to exploit the anabolic aspects ofdark respiration to its fullest potential so that futureagricultural systems can reap maximal benefits of ele-vated CO2 under both field and controlled environments,including the production of plant oils that are importantfor the future bioeconomy.
Ainsworth, E. A., Rogers, A., Nelson, R. and Long, S. P. 2004.
Testing the source-sink hypothesis of down-regulation of pho-tosynthesis in elevated [CO2] in the field with single genesubstitutions in Glycine max. Agric. For. Meteorol. 122: 85�94.Ainsworth, E. A. and Rogers, A. 2007. The response of photo-synthesis and stomatal conductance to rising CO2: mechanismsand environmental interactions. Plant Cell Environ. 30: 258�270.Arp, W. J. 1991. Effects of source�sink relations on photo-synthetic acclimation to elevated CO2. Plant Cell Environ. 14:869�875.Boese, S. R. and Huner, N. P. A. 1992. Developmental historyaffects the susceptibility of spinach leaves to in vivo low tem-perature photoinhibition. Plant Physiol. 99: 1141�1145.Budde, R. J. A. and Randall, D. D. 1990. Pea leaf mitochon-drial pyruvate dehydrogenase complex is inactivated in vivo ina light-dependent manner. Proc. Natl. Acad. Sci. USA 87:673�676.Cheng, S. H., Moore, B. D. and Seemann, J. R. 1998. Effects ofshort and long-term elevated CO2 on the expression ofRibulose-1,5-bisphosphate carboxylase/oxygenase genes andcarbohydrate accumulation in leaves of Arabidopsis thaliana(L.) Heynh. Plant Physiol. 116: 715�723.Dahal, K., Kane, K., Gadapati, W., Webb, E., Savitch, L. V.,
Singh, J., Sharma, P., Sarhan, F., Longstaffe, F. J., Grodzinski,
B. and Huner, N. P. A. 2012a. The effects of phenotypicplasticity on photosynthetic performance in winter Rye, winterwheat and Brassica napus. Physiol. Plant. 144: 169�188.Dahal, K., Kane, K., Sarhan, F., Grodzinski, B. and Huner,
N. P. A. 2012b. Cold acclimation inhibits CO2-dependentstimulation of photosynthesis in spring wheat and spring rye.Botany 90: 433�444.Dahal, K., Gadapati, W., Savitch, L. V. and Singh, J. 2012c.
Cold acclimation and BnCBF17-over-expression enhancephotosynthetic performance and energy conversion efficiencyduring long-term growth of Brassica napus under elevated CO2
conditions. Planta 236: 1639�1652.Dahal, K., Knowles, V. L., Plaxton, W. C. and Huner, N. P. A.
2013. Enhancement of photosynthetic performance, water useefficiency and grain yield during long-term growth underelevated CO2 in wheat and rye is growth temperature andcultivar dependent. Environ. Exp. Bot. (in press) doi: 10.1016/j.envexpbot.2013.11.015.Demmig-Adams, B. and Adams III, W. W. 1992. Photoprotec-tion and other responses of plants to high light stress. Annu.Rev. Plant Physiol. Plant Mol. Biol. 43: 599�626.Drake, B. G., Gonzalez-Meler, M. A. and Long, S. P. 1997.
More efficient plants: a consequence of rising atmosphericCO2? Annu. Rev. Plant Physiol. Plant Mol. Biol. 48: 609�639.Drake, B. G., Azcon-Bieto, J., Berry, J., Bunce, J., Dijkstra, J.,
Farrar, J., Gifford, R. M., Gonzalez-Meler, M. A., Koch, G.
and Lambers, H. et al. 1999. Does elevated atmospheric CO2
concentration inhibit mitochondrial respiration in green plants?Plant Cell Environ. 22: 649�657.Foyer, C. 1990. The effect of sucrose and mannose on cyto-plasmic protein phosphorylation sucrose phosphate synthetaseactivity and photosynthesis in leaf protoplasts from spinach.Plant Physiol. Biochem. 28: 151�160.Gemel, J. and Randall, D. D. 1992. Light regulation of leafmitochondrial pyruvate dehydrogenase complex: role of pho-torespiratory carbon metabolism. Plant Physiol. 100: 908�914.Grodzinski, B. 1992. Plant nutrition and growth regulation byCO2 enrichment. Bioscience 42: 517�525.
DAHAL ET AL. * BIOMASS PRODUCTION AND ELEVATED CO2 1081
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

Huner, N. P. A., .Oquist, G. and Sarhan, F. 1998. Energybalance and acclimation to light and cold. Trend Plant Sci. 3:224�230.Hurry, V. M. and Huner, N. P. A. 1991. Low growthtemperature effects a differential inhibition of photosynthesisin spring and winter wheat. Plant Physiol. 96: 491�497.Hurry, V. M., Malmberg, G., Gardestrom, P. and Oquist, G.
1994. Effects of a short-term shift to low temperature and oflong-term cold hardening on photosynthesis and ribulose-1,5-bisphosphate carboxylase/oxygenase and sucrose phosphatesynthase activity in leaves of winter rye (Secale cereale L.).Plant Physiol. 106: 983�990.Hurry, V., Strand, A., Furbank, R. and Stitt, M. 2000. Therole of inorganic phosphate in the development of freezingtolerance and the acclimatization of photosynthesis to lowtemperature is revealed by the pho mutants of Arabidopsisthaliana. Plant J. 24: 383�396.Hurry, V. M., Strand, A., Tabiaeson, M., Gardestrom, P. and
.Oquist, G. 1995. Cold hardening of spring and winter wheatand rape results in differential effects on growth, carbonmetabolism, and carbohydrate content. Plant Physiol. 109:697�706.Kramer, P. J. 1981. Carbon dioxide concentration, photo-synthesis and dry matter production. Bioscience 31: 29�33.Leakey, A. D. B., Xu, F., Gillespie, K. M., McGrath, J. M. and
Ainsworth, E. A. 2009. Genomic basis for stimulated respira-tion by plants growing under elevated carbon dioxide. Proc.Natl. Acad. Sci. USA 106: 3597�3602.Leonardos, E. D., Savitch, L. V., Huner, N. P. A., .Oquist, G.
and Grodzinski, B. 2003. Daily photosynthetic and C-exportpatterns in winter wheat leaves during cold stress and accli-mation. Physiol. Plant. 117: 521�531.Long, S. P., Ainsworth, E. A., Rogers, A. and Ort, D. R. 2004.
Rising atmospheric carbon dioxide: plants FACE the future.Ann. Rev. Plant. Biol. 55: 591�628.Long, S. P., Zhu, X.-G., Naidu, S. L. and Ort, D. R. 2006. Canimprovement in photosynthesis increase crop yields? Plant CellEnviron. 29: 315�330.Luethy, M. H., Miernyk, J. A. and Randall, D. D. 1994. Thenucleotide and deduced amino acid sequences of a cDNAencoding the E1b-subunit of the Arabidopsis thaliana mito-chondrial pyruvate dehydrogenase complex. Biochim. Biophys.Acta 1187: 95�98.Marillia, E., Micallef, B. J., Micallef, M., Weninger, A.,
Pedersen, K. K., Zou, J. and Taylor, D. C. 2003. Biochemicaland physiological studies of Arabidopsis thaliana transgeniclines with repressed expression of the mitochondrial pyruvatedehydrogenase kinase. J. Exp. Bot. 54: 259�270.Morgan, P. B., Bollero, G. A., Nelson, R. L., Dohleman, F. G.
and Long, S. P. 2005. Smaller than predicted increase inaboveground net primary production and yield of field-grownsoybean under fully open-air [CO2] elevation. Global ChangeBiol. 11: 856�1865.Moore, B. D., Cheng, S. H., Sims, D. and Seemann, J. R. 1999.
The biochemical and molecular basis for photosyntheticacclimation to elevated atmospheric CO2. Plant Cell Environ.22: 567�582..Oquist, G. and Huner, N. P. A. 2003. Photosynthesis ofoverwintering evergreen plants. Ann. Rev. Plant Biol. 54:329�355.Ort, D. R. 2001. When there is too much light. Plant Physiol.125: 29�32.
Patel, M. S. and Korotchkina, L. G. 2003. The biochemistry ofthe pyruvate dehydrogenase complex. Biochem. Mol. Biol.Educ. 31: 5�15.Rapacz, M., Wolanin, B., Hura, K. and Tyrka, M. 2008. Theeffects of cold acclimation on photosynthetic apparatus and theexpression of COR14b in four genotypes of barley (Hordeumvulgare) contrasting in their tolerance to freezing and high-light treatment in cold conditions. Ann. Bot. 101: 689�699.Rauf, S. 2012. The effect of elevated CO2 on whole-plantrespiration, photosynthesis and net carbon gain of Arabidopsisthaliana having altered mitochondrial pyruvate dehydrogenasekinase expressed constitutively. M.Sc. thesis. University ofGuelph, Guelph, ON.Robertson, E. J., Williams, M., Harwood, J. L., Lindsay, J. G.,
Leaves, C. J. and Leech, R. M. 1995. Mitochondria increasethree-fold and mitochondrial proteins and lipid change dra-matically in postmeristematic cells in young wheat leavesgrown in elevated CO2. Plant Physiol. 108: 469�474.Savitch, L. V., Allard, G., Seki, M., Robert, L. S., Tinker,
N. A., Huner, N. P. A., Shinozaki, K. and Singh, J. 2005.
The effect of overexpression of two Brassica CBF/DREB1-liketranscription factors on photosynthetic capacity and freezingtolerance in Brassica napus. Plant Cell Physiol. 46: 1525�1539.Savitch, L. V., Leonardos, E. D., Krol, M., Jansson, S.,
Grodzinski, B., Huner, N. P. A. and .Oquist, G. 2002. Twodifferent strategies for light utilization in photosynthesis inrelation to growth and cold acclimation. Plant Cell Environ.25: 761�771.Sharkey, T. D. and Vanderveer, P. J. 1989. Stromal phosphateconcentration is low during feedback limited photosynthesis.Plant Physiol. 91: 679�684.Smidansky, E. D., Clancy, M., Meyer, F. D., Lanning, S. P.,
Blake, N. K., Talbert, L. E. and Giroux, M. J. 2002. EnhancedADP-glucose pyrophosphorylase activity in wheat endo-sperm increases seed yield. Proc. Natl. Acad. Sci. USA 99:1724�1729.Smith, D. and Zhou, X. 2014. An effective integrated researchapproach to study climate change in Canada: Preface. Can. J.Plant Sci. 94: 995�1008.Stitt, M. and Hurry, V. M. 2002. A plant for all seasons:alterations in photosynthetic carbon metabolism during coldacclimation in Arabidopsis. Curr. Opin. Plant Biol. 5: 199�206.Stitt, M. and Krapp, A. 1999. The interaction between elevatedcarbon dioxide and nitrogen nutrition: the physiological andmolecular background. Plant Cell Environ. 22: 583�621.Stitt, M. and Quick, W. P. 1989. Photosynthetic carbonpartitioning: its regulation and possibilities for manipulation.Physiol. Plantarum. 77: 633�641.Thelen, J. J., Miernyk, J. A. and Randall, D. D. 2000. Pyruvatedehydrogenase kinase from Arabidopsis thaliana: a proteinhistidine kinase that phosphorylates serine residues. Biochem.J. 349: 195�201.Tovar-Me
.ndez, A., Miernyk, J. A. and Randal, D. D. 2003.
Regulation of pyruvate dehydrogenase complex activity inplant cells. Eur. J. Biochem. 270: 1043�1049.Winzeler, M. D., McCullough, E. and Hunt, L. A. 1989. Leafgas exchange and plant growth of winter rye, Triticale, andwheat under contrasting temperature regimes. Crop Sci. 29:1256�1260.Weraduwage, S. M. 2013. Harnessing the anabolic propertiesof dark respiration to enhance sink activity at high CO2
using Arabidopsis thaliana L. with partially suppressed
1082 CANADIAN JOURNAL OF PLANT SCIENCE
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

mitochondrial pyruvate dehydrogenase kinase. Ph.D. thesis.University of Guelph, Guelph, ON.Weraduwage, S. M., Micallef, B. J., Grodzinski, B., Taylor,
D. C. and Marillia, E.-F., 2011. Roles of dark respirationin plant growth and productivity. Pages 191�207 in M. M.Young, ed. Comprehensive biotechnology. 2nd ed. Vol. 4.Elsevier, Oxford, UK.Yang, P. 2010. Quantitative 3D growth of Arabidopsis thalianausing 3D range images. M.Sc. thesis. University of WesternOntario, London, ON.Yeaman, S. J., Hutcheson, E. T., Roche, T. E., Pettit, F. H.,
Brown, J. R., Reed, L. J., Watson, D. C. and Dixon, G. H.
1978. Sites of phosphorylation on pyruvate dehydrogenasefrom bovine kidney and heart. Biochemistry 17: 2364�2370.Zhao, C. 2009. 3D plant growth measurements using theShapeGrabber laser scanner. M.Sc. thesis. University ofWestern Ontario, London, ON.Zhu, X.-G., Long, S. P. and Ort, D. R. 2010. Improvingphotosynthetic efficiency for greater yield. Annu. Rev. PlantBiol. 61: 235�261.Zou, J., Qi, Q., Katavic, V., Marillia, E.-F. and Taylor, D. C.1999. Effects of antisense repression of an Arabidopsis thalianapyruvate dehydrogenase kinase cDNA on plant development.Plant Mol. Biol. 41: 837�849.
DAHAL ET AL. * BIOMASS PRODUCTION AND ELEVATED CO2 1083
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.

This article has been cited by:
1. Evangelos D. Leonardos, Shezad A. Rauf, Sarathi M. Weraduwage, Elizabeth-France Marillia, David C. Taylor, Barry J.Micallef, Bernard Grodzinski. 2014. Photosynthetic capacity of the inflorescence is a major contributor to daily-C-gain andthe responsiveness of growth to elevated CO2 in Arabidopsis thaliana with repressed expression of mitochondrial-pyruvate-dehydrogenase-kinase. Environmental and Experimental Botany 107, 84-97. [CrossRef]
Can
. J. P
lant
Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Agr
icul
ture
and
Agr
i-fo
od C
anad
a on
07/
09/1
4Fo
r pe
rson
al u
se o
nly.
Related Documents











