Enhanced secretion of heterologous proteins in Kluyveromyces lactis by overexpression of the GDP-mannose pyrophosphorylase, KlPsa1p Daniela Uccelletti a , Dessislava Staneva b , Silvia Rufini a , Pencho Venkov b , Claudio Palleschi a, * a Department of Developmental and Cell Biology, University of Rome ÔLa SapienzaÕ, Rome, Italy b Department of Molecular Genetics, Institute of Molecular Biology, Bulgarian Academy of Sciences, Sofia 1113, Bulgaria Received 28 July 2004; received in revised form 26 November 2004; accepted 19 January 2005 First published online 2 March 2005 Abstract GDP-mannose is the mannosyl donor for the glycosylation reactions and is synthesized by GDP-mannose pyrophosphorylase from GTP and D-mannose-1-phosphate; in Saccharomyces cerevisiae this enzyme is encoded by the PSA1/VIG9/SRB1 gene. We isolated the Kluyveromyces lactis KlPSA1 gene by complementing the osmotic growth defects of S. cerevisiae srb1/psa1 mutants. KlPsa1p displayed a high degree of similarity with other GDP-mannose pyrophosphorylases and was demonstrated to be the functional homologue of S. cerevisiae Psa1p. Phenotypic analysis of a K. lactis strain overexpressing the KlPSA1 gene revealed changes in the cell wall assembly. Increasing the KlPSA1 copy number restored the defects in O-glycosylation, but not those in N-glycosylation, that occur in K. lactis cells depleted for the hexokinase Rag5p. Overexpression of GDP-mannose pyrophosphor- ylase also enhanced heterologous protein secretion in K. lactis as assayed by using the recombinant human serum albumin and the glucoamylase from Arxula adeninivorans. Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: KlPSA1; Glycosylation; Secretion; Kluyveromyces lactis; GDP-mannose pyrophosphorylase 1. Introduction Protein synthesis, translocation, modification, and targeting are complex and elaborate processes that are highly conserved among eukaryotes. The addition of oli- gosaccharide chains to the protein backbone is the ma- jor posttranslational covalent modification occurring to secretory proteins during their passage through the secretory pathway [1]. In all eukaryotes, mannose is transferred to proteins from its activated form, GDP-mannose (GDP-Man). GDP-Man is directly used as mannosyl donor for the synthesis of cellular carbohydrate moieties on the cyto- plasmic surface of the ER membrane and in all man- nosylation reactions in the Golgi compartments. It can also be used indirectly, via the intermediate dolichyl phosphate mannose (Dol-P-Man). The synthesis of Dol-P-Man from GDP-Man and Dol-P is catalyzed by Dol-P-Man synthase, DPMS. Dol-P-Man is the man- nose donor for all mannosylation reactions carried out in the ER lumen. GDP-Man is not only involved in all kinds of protein glycosylation but it is also required for the synthesis of other mannose-containing glycoconjugates, such as glycolipids, lipopolysaccharides and polysaccharides; 1567-1356/$22.00 Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsyr.2005.01.004 * Corresponding author. E-mail address: [email protected] (C. Palleschi). www.fems-microbiology.org FEMS Yeast Research 5 (2005) 735–746

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.fems-microbiology.org
FEMS Yeast Research 5 (2005) 735–746
Enhanced secretion of heterologous proteins inKluyveromyces lactis by overexpression of theGDP-mannose pyrophosphorylase, KlPsa1p
Daniela Uccelletti a, Dessislava Staneva b, Silvia Rufini a,Pencho Venkov b, Claudio Palleschi a,*
a Department of Developmental and Cell Biology, University of Rome �La Sapienza�, Rome, Italyb Department of Molecular Genetics, Institute of Molecular Biology, Bulgarian Academy of Sciences, Sofia 1113, Bulgaria
Received 28 July 2004; received in revised form 26 November 2004; accepted 19 January 2005
First published online 2 March 2005
Abstract
GDP-mannose is the mannosyl donor for the glycosylation reactions and is synthesized by GDP-mannose pyrophosphorylasefrom GTP and D-mannose-1-phosphate; in Saccharomyces cerevisiae this enzyme is encoded by the PSA1/VIG9/SRB1 gene.
We isolated the Kluyveromyces lactis KlPSA1 gene by complementing the osmotic growth defects of S. cerevisiae srb1/psa1
mutants. KlPsa1p displayed a high degree of similarity with other GDP-mannose pyrophosphorylases and was demonstrated tobe the functional homologue of S. cerevisiae Psa1p. Phenotypic analysis of a K. lactis strain overexpressing the KlPSA1 generevealed changes in the cell wall assembly. Increasing the KlPSA1 copy number restored the defects in O-glycosylation, but not thosein N-glycosylation, that occur in K. lactis cells depleted for the hexokinase Rag5p. Overexpression of GDP-mannose pyrophosphor-ylase also enhanced heterologous protein secretion in K. lactis as assayed by using the recombinant human serum albumin and theglucoamylase from Arxula adeninivorans.� 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: KlPSA1; Glycosylation; Secretion; Kluyveromyces lactis; GDP-mannose pyrophosphorylase
1. Introduction
Protein synthesis, translocation, modification, andtargeting are complex and elaborate processes that arehighly conserved among eukaryotes. The addition of oli-gosaccharide chains to the protein backbone is the ma-jor posttranslational covalent modification occurringto secretory proteins during their passage through thesecretory pathway [1].
In all eukaryotes, mannose is transferred to proteinsfrom its activated form, GDP-mannose (GDP-Man).
1567-1356/$22.00 � 2005 Federation of European Microbiological Societiesdoi:10.1016/j.femsyr.2005.01.004
* Corresponding author.E-mail address: [email protected] (C. Palleschi).
GDP-Man is directly used as mannosyl donor for thesynthesis of cellular carbohydrate moieties on the cyto-plasmic surface of the ER membrane and in all man-nosylation reactions in the Golgi compartments. It canalso be used indirectly, via the intermediate dolichylphosphate mannose (Dol-P-Man). The synthesis ofDol-P-Man from GDP-Man and Dol-P is catalyzed byDol-P-Man synthase, DPMS. Dol-P-Man is the man-nose donor for all mannosylation reactions carried outin the ER lumen.
GDP-Man is not only involved in all kinds of proteinglycosylation but it is also required for the synthesis ofother mannose-containing glycoconjugates, such asglycolipids, lipopolysaccharides and polysaccharides;
. Published by Elsevier B.V. All rights reserved.

736 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
GDP-Man is also a precursor for GDP-fucose synthesisand acts as a substrate for ascorbic acid (vitamin C) bio-synthesis in plants [2].
The synthesis of GDP-Man from mannose-1-phos-phate (Man-1-P) and GTP is catalyzed by the enzymeGDP-mannose pyrophosphorylase (GMPP, EC2.7.7.13). GMPPs from different organisms, includingyeasts, protozoa, plants and mammals, display a highdegree of sequence identity (50–90%) to the Saccharo-
myces cerevisiae GMPP. Complete lack of GMPP activ-ity and the consequent loss of Man-containingglucoconjugates are lethal in eukaryotes [3–5]. Leish-
mania mexicana is the only eukaryote, reported so far,which is still viable in culture upon GMPP gene dele-tion. However, the absence of GMPP activity led tocomplete inability of L. mexicana to infect macrophagesand mice [6].
The S. cerevisiae gene encoding GMPP has beenindependently isolated by several groups and is referredto as PSA1/VIG9/SRB1 [3,7,8]. ScPSA1 gene is a cellcycle-regulated gene with a peak of expression in theG1 phase [3]. Constitutive expression of ScPSA1/SRB1resulted in enhanced cell growth, larger cell volumeand a dominant flocculation phenotype [9]. The Candidaalbicans PSA1/SRB1 homologue might be regulated toachieve differential expression in yeast and hyphal forms[10].
Several mutant alleles of the ScPSA1 gene have beenidentified so far and each one was shown to cause pleio-tropic defects. For example, vig9-1 and vig9-2 mutantsshowed vanadate resistance, immature protein glycosy-lation and drug hypersensitivity [7]. Mutant cells carry-ing a vig9-3 allele were 2-3 times larger than the parentalcells, temperature sensitive and dependent on osmoticstabilizer for viability, displayed severe N-glycosylationdefects, and excreted cell surface proteins into themedium [11]. The srb1-1 mutation resulted in highlypleiotropic phenotypes, the most significant being sus-ceptibility to hypotonic shock, sorbitol-dependentgrowth [12,13], enhanced transformability [14], andhypersensitivity to a wide range of compounds [15].These mutant phenotypes resulted from compositionand structural alterations in the major cell wall compo-nents, glucan and mannan [16,17]. Similar pleiotropic ef-fects have been observed in two yeast species, S.
cerevisiae and C. albicans, upon methionine-mediatedrepression of pScMET3-ScSRB1 and pCaMET3-CaSRB1. The major phenotypes were cell lysis, impairedbud site selection, failure of cell separation, increasedflocculation, drug sensitivity, and impaired hyphalswitching ability [4]. Recently, a Hansenula polymorpha
mutant has been isolated and shown to carry a mutationin the OPU24 gene, encoding a homologue of GMPP.The mutant exhibited phenotypes similar to those ofsome S. cerevisiae psa1/srb1/vig9 mutants [18]. All thesephenotypes were a result from the GMPP deficiency in
GDP-Man biosynthesis, which in turn causes defectsin glycosylation of secretory and cell wall proteins.
Originally, the ScPSA1 gene was isolated as a multi-copy plasmid suppressor of alg1 [3], and overexpressionof PSA1 was shown to suppress not only the alg1/nes17mutation [19,20] but also that of alg2 and dpm1 [20,21],restoring defects in N-glycosylation, O-mannosylation,GPI anchor synthesis and/or temperature sensitivity ofthe mutants. In fact, overexpression of the GMPP-encoding gene increases the cellular GDP-Man levelabout 2–3-fold [7,19,20,22]. In Trichoderma reesei, over-expression of the mpg1 gene, coding for GMPP, resultedin both N- and O-hypermannosylation of secreted pro-teins as well as in increased DPM1 gene transcriptionand DPMS activity, suggesting that the level of GDP-Man can play a relevant regulatory role in T. reesei
protein glycosylation [22].The lactose-utilizing yeast Kluyveromyces lactis is re-
lated to S. cerevisiae and has a distinctive capacity forextracellular secretion of recombinant proteins. As afurther advantage, K. lactis secretes homologous pro-teins in low abundance, thus greatly facilitating the puri-fication of the secreted product. K. lactis is generallyregarded as safe (GRAS) and is employed for industrialproduction of enzymes, e.g., lactase [23] and chymosin[24], for human use. The degree of genetic manipulationfor this yeast stimulated its use in the study of secretionand glycosylation pathways [25–28]. In the present pa-per, we report on the isolation and characterization ofthe KlPSA1 gene, a functional homologue of theScPSA1/VIG9/SRB1 gene. We found that increasedcopy numbers of KlPSA1 are able to restore the O-gly-cosylation defects that occur in K. lactis cells depleted ofthe hexokinase Rag5p. In addition, overexpression ofGDP-mannose pyrophosphorylase enhances the heter-ologous protein secretion in K. lactis, as indicatedby the extracellular production of glucoamylase fromArxula adeninivorans and recombinant human serumalbumin.
2. Materials and methods
2.1. Strains, media and growth conditions
Yeast strains used in this study are listed in Table 1.Yeast YP-rich medium containing either 2% glucose(YPD) or 2% glucose and 2% ethanol (YPDE), and syn-thetic minimal (SM) media, supplemented with theappropriate nutritional requirements, were preparedaccording to Johnston [29]. G418 was added at a con-centration of 0.1 or 0.3 mg ml�1 as indicated. All typesof media (YPDSorb, SMSorb) for the cultivationof srb1 osmosensitive mutants were supplemented with1-M sorbitol. Yeast strains were incubated at 28 �C, unlessnoted otherwise.

Table 1Yeast strains used in this study
Strain Genotype Source/Reference
S. cerevisiae
7SLU MATa srb1-1 leu2 ura3 [60]4STLU MATa srb1-1 trp1 leu2 ura3 [13]S288C MATa gal2 G. Fink
K. lactis
MW270-7B MATa ura3 leu2 met1-1 Wesolowski-LouvelMW98-8C MATa uraA lys arg rag1 rag2 K+ pKD10 Wesolowski-LouvelJA6 MATa ade1-600 adeT-600 trp1-11 ura3-12 [61]rag5D MATa ade1-600 adeT-600 trp1-11 ura3-12 rag5::URA3 [62]
D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746 737
Escherichia coli strain DH5a(/ 80dlac ZDM15, recA1, end A1, gyr A96, thi-1, hsd R17 ðr�k ;mþ
k Þ, sup E44,rel A1, deo R, D(lacZYA-argF) U169), used to amplifyrecombinant plasmids, was grown in LB broth, supple-mented with Ampicillin (100 lg ml�1) and X-gal(40 lg ml�1) when necessary. For solid media, agarwas added at 2% (w/v).
2.2. Genomic libraries and plasmids
The K. lactis genomic libraries, based on the YCp50centromeric vector or on the YEp352 multicopy vector,were kindly provided by Dr. M. Stark (University ofDundee, Dundee, UK) and Dr. A. Paszewski (Instituteof Biochemistry and Biophysics, Warsaw, Poland),respectively. The plasmids used in this study are de-scribed in Table 2.
For the KlPSA1 overexpression plasmid, pKlPSA1,the gene amplified by PCR was cloned into the PCR 2.1vector (Invitrogen, Carlsbad, CA, USA). The insertioninto such plasmid was sequenced and then digested withXbaI to be cloned into theNheI site of the pYG131/1 vec-tor (kindly provided by Prof. C. Falcone, University ofRome). This final constructwas transformed intoK. lactisstrain JA6, and transformantswere selectedonYPDmed-ium containing 0.1 mg ml�1 of G418.
For the heterologous expression of rHSA the HSA
cDNA was cloned in the HindIII restriction site of thepRS426 plasmid [30], whose SnaBI site was used to
Table 2Plasmids used in this study
Plasmid Relevant details
pRS426+SD11 K. lactis SD11 inserted in SnaBI site of pp426-HSA rHSA cDNA cloned in pRS426+SD11pSRB1-9b ScPSA1 on a ClaI–BamHI fragment in ppYG131/1 pKD1-KanR-shuttle vectorYEp352-KlPSA1 Genomic clone from YEp352-based genoYCp50-KlPSA1 Genomic clone from YCp50-based genopRS316-KlPSA1 XbaI–SacI fragment of YEp352-KlPSA1pKlPSA1 KlPSA1 amplified and cloned in pYG13pBS-KlPSA1 XbaI–SacI fragment of YEp352-KlPSA1pTS32x-GAA a-Glucoamylase gene expression plasmid
add the fragment SD11, the minimal sequence neededfor a plasmid to be maintained in K. lactis. The resultingplasmid was named p426-HSA.
2.3. General methods
Plasmid isolation, restriction enzyme analyses, liga-tions, agarose gel electrophoresis and Southern blotwere performed according to Sambrook et al. [31]. S.cerevisiae transformation was performed as describedpreviously [32], with the modification that all mediaand solutions except for ‘‘the transformation mixture’’contained 1 M sorbitol. K. lactis was transformed byelectroporation with a Biorad Gene-Pulser apparatus,following the supplier�s procedure.
The amplification of the ScPSA1 gene, used as aprobe, was obtained by PCR with S. cerevisiae genomicDNA as a template and the primer pair scpsaF,5 0-AATTTTAGTCGGTGGTTACGG-3 0 and scpsaR,5 0-CCTTTGGAACATTATCGGAGA-3 0. The PCRproduct obtained was purified from 1% agarose geland labelled radioactively with [a-32P]dCTP using acommercial Oligolabelling kit (Pharmacia Biotech).
2.4. Plasmid segregation
Plasmid segregation was performed as described pre-viously [10], with a minor modification: the media con-tained 1 M sorbitol and aliquots for single colonies
Reference
RS426 Farina F.This study
RS316 [8][38]
mic library containing 2.8 kb insert This studymic library containing about 10 kb insert This studyin pRS316, contains KlPSA1 This study1/1 This studyin pBluescript II KS, contains KlPSA1 This study
[53]

738 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
were spread on SMSorb plates supplemented withuracil.
2.5. Sequence analysis
The nucleotide sequence of the pBS-KlPSA1 insert(XbaI–SacI fragment of YEp352-KlPSA1 ligated toXbaI–SacI digested pBluescript II KS vector) was deter-mined automatically (MWG-Biotech, Ebersberg, Ger-many). Comparison against SGD and PIR-NREFdatabases was done with a BLAST algorithm [33]. Mul-tiple alignments of protein sequences were obtainedusing the DNAMAN program, Version 4.13 (LynnonCorp. Quebec, Canada).
2.6. Analysis of invertase and chitinase
Chitinase and invertase were assayed as previously re-ported [34].
2.7. Porosity assay
The cell wall porosity assay was performed accordingto De Nobel et al. [35].
2.8. Glucoamylase detection
The plasmid pTS32x-GAA (gently provided by Prof.K. Breunig) was used in this study for the expression ofthe A. adeninivorans a-glucoamylase gene in K. lactis.The strains JA6 transformed with the control vectorpYG131/1 and pKlPSA1 were co-transformed withpTS32x-GAA and grown in SM medium supplementedwith 2% EtOH and 0.3 mg ml�1 of G418. The secretionof glucoamylase was determined by measurement ofhydrolytic activity on starch, assayed as described in[36]. No a-glucoamylase activity was observed in sam-ples from the strains transformed with empty vector.
2.9. Secretion and analysis of rHSA
We analyzed the secretion of human serum albuminin the culture media of strains JA6 transformed withthe control vector pYG131/1 and pKlPSA1 co-trans-formed with the plasmid p426-HSA, constructed forthe heterologous expression of rHSA, and grown for72 h at 30 �C in SM supplemented with 2% EtOH and0.3 mg ml�1 of G418. The extracellular medium corre-sponding to 5 · 106 cells was loaded on 10% SDS–PAGE and electroblotted onto a PVDF membrane(Biorad). The primary polyclonal antibodies were usedat 1:10000 (Sigma). The secondary antibodies wereanti-rabbit IgG conjugated with peroxidase (Biorad).The detection procedure was carried out with ECL Wes-tern blotting detection kit (Amersham) according to themanufacturer�s instructions. The densitometric analysis
was done with an image analyzer (Phoretix 1D; NonLinear Dynamics Ltd., Newcastle upon Tyne, UK).
2.10. Northern-blot analysis
Total RNA was extracted, following the hot phenolmethod [37], from cultures grown 24 or 72 h as de-scribed. The probes were generated by random primedlabelling of a 1.8-kb HindIII DNA fragment frompYG132 [38] for rHSA and of a 1-kb PCR-amplifiedDNA fragment from pKlPSA1 for KlPSA1. Hybridiza-tion and washing were done at high stringency accord-ing to Sambrook et al. [31].
2.11. Nucleotide sequence accession number
The K. lactis PSA1 DNA sequence reported in thisarticle is available in the EMBL Data Library underAccession No. AJ551274.
3. Results
3.1. Identification and functional complementation of the
KlPSA1 gene
Total genomic DNA was isolated from two K. lactis
(MW98-8C and MW270-7B) and one S. cerevisiae
(S288C) strains, digested with several enzymes (HindIII,XbaI, XhoI) and subjected to Southern-blot analysis,using the S. cerevisiae PSA1 gene as a probe. In eachof K. lactis genomic DNA digests, as in those of S. cere-visiae, only one hybridizing band was identified. Thehybridization patterns of the two K. lactis genomicDNAs were identical to each other and differed fromthe S. cerevisiae one (data not shown). The results sug-gest that a PSA1-related sequence does exist in theK. lactis genome and that, as in S. cerevisiae, the puta-tive K. lactis PSA1 gene seems to also be unique. Thesedata, together with the ability of VIG9/PSA1 homo-logues of C. albicans (CaVIG9) and C. glabrata
(CgVIG9) to complement S. cerevisiae psa1/srb1-1/vig9-
1 protein glycosylation mutant phenotypes [10,39], stim-ulated us to attempt the isolation of a K. lactis PSA1
homologue by functional complementation.To clone the wild-type KlPSA1 gene, we transformed
7SLUand4STLUmutants ofS. cerevisiaewith twodiffer-entK. lactis genomic libraries, constructed in amulticopy(YEp352) and in a centromeric (YCp50) vector, respec-tively. Sorbitol dependence for growth of the S. cerevisiaesrb 1 mutants was used as selective condition in the clon-ing procedure. Transformants were first selected onSMSorb medium lacking uracil and then screened onSM medium (without uracil and sorbitol). One 7SLUtransformant, carrying a genomic clone of a YEp352-based genomic library, and one 4STLU transformant,

Fig. 1. Complementation ability of different constructs harbouring theputativeKlPSA1 gene. The srb1-1mutant strain 7SLUwas transformedwith control vectors (YCp50, YEp352 or pRS316) or plasmids contain-ing theputativeKlPSA1 (YCp50-KlPSA1,YEp352-KlPSA1orpRS316-KlPSA1). For comparison the mutant 7SLU and a transformantcarrying the S. cereviviae PSA1/SRB1 gene (pRS316-ScPSA1, plasmidpSRB1-9b) were also streaked on the plates containing appropriateselective medium. The plates were incubated for 5 days at 28 �C.
D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746 739
containing an YCp50-based clone, were able to grow onmedia without the osmotic stabilizer. Plasmid DNAs res-cued from these two transformants were designatedYEp352-KlPSA1 and YCp50-KlPSA1, respectively.
Back-transformation and plasmid segregation werecarried out to confirm the plasmid-linked complementa-tion. All transformants from 7SLU and 4STLU, selectedon SMSorb media, grew on SM media as well and testsfor plasmid segregation were performed on two trans-formants of each type. The obtained results demon-strated that, in all cases, the segregants that had lostthe plasmid-born selective marker URA3 were not ableto grow on sorbitol-depleted media (data not shown).Restriction analysis of the complementing plasmidsshowed that YEp352-KlPSA1 contained an insert ofabout 2.8 kb, while YCp50-KlPSA1 carried a K. lactis
genomic fragment of about 10 kb; both plasmids hybrid-ized to the ScPSA1 gene probe, as demonstrated bySouthern analyses (data not shown).
In order to identify the minimal region containing theKlPSA1 gene the 2.8-kb genomic insert from theYEp352-KlPSA1 plasmid was transferred in the centro-meric vector pRS316 and tested for its ability to comple-ment srb1 mutant phenotype when present as a singlecopy in the cell. The resulting plasmid pRS316-KlPSA1was transformed in the 7SLU strain. All the construc-tions containing the KlPSA1 putative gene or ScPSA1
gene were able to complement the sorbitol requirementof the S. cerevisiae 7SLU srb1-1 mutant (Fig. 1). Wetherefore concluded that both YEp352-KlPSA1 andYCp50-KlPSA1 plasmids contained a functional homo-logue of the S. cerevisiae PSA1 gene.
Sequencing of the 2.8-kb complementing genomicfragment revealed the presence of one open readingframe (ORF), 1083 bp in length, which displayed 84%identity to the nucleotide sequence of ScPSA1 ORF.On the basis of sequence similarity with ScPSA1 andfunctional complementation data the isolated K. lactisgene was named KlPSA1.
The KlPSA1 ORF encodes for a putative protein of361 amino-acid residues that, compared with the PIR-NREF protein sequence database, showed significantsimilarities to GMPP proteins of yeasts and other unre-lated species (Fig. 2). The KlPsa1p was more closely re-lated to the Psa1p of S. cerevisiae (91% identity) than toGMPP homologues of C. glabrata, C. albicans and Sch.pombe (88%, 84% and 71% identity, respectively). Theoverall identity among the five yeast proteins was 64%.As shown in Fig. 2, significant similarity betweenKlPsa1p and the GMPPs from higher eukaryotes wasalso found, ranging from 56% (human GMPPB) to60% (Arabidopsis thaliana GMPP).
Thehigher sequence conservation is concentrated in theNH2-terminal half of the polypeptide, as already observednot only amongst eukaryotic GMPPs but also in bacterialproteins that catalyze the formation of NDP sugars [7].
The sequence of the complementing clone YEp352-KlPSA1 revealed a 430-bp sequence upstream of theKlPSA1 ORF, which was enough to promote the geneexpression in S. cerevisiae. This promoter region,characterized by a high A/T content (70%), was in silicoanalyzed for presence of regulatory elements. FourTATA-box-like sequences are present, at nucleotides�303 to�294 (TATATATAA,),�255 to�250 (TATAA),�183 to �177 (TATATAA) and �7 to �1 (TATA-TAA), upstream of the ATG translation start codon.Furthermore, several consensus sequences for bindingof transcription factors in S. cerevisiae were identifiedin the KlPSA1 promoter region after searching the Sac-charomyces cerevisiae Promoter Database (http://cgsigma.cshl.org/jian/HTML/searchputative.html). GCN4 andPHO4 putative binding sites are present in the promoterregion of both KlPSA1 and ScPSA1 promoters. In addi-tion to these, several potential control elements werefound only in the KlPSA1 gene, e.g., SCB element(cell-cycle box) and Bas2p-binding sites.
3.2. Increase dosage of KlPSA1 is able to suppress the
O-glycosylation defect of rag5D
In K. lactis RAG5 encodes the main hexokinase andcells depleted of this activity show defects in theN- and O-glycosylation [40,41]. We then asked whethersuch glycosylation defects, probably originating from

Fig. 2. Alignment of KlPsa1p with other Psa1p homologues from different organisms. Identical residues are shaded in black and the consensus isgiven beneath the protein sequences. Accession numbers are as follows: O44970 for Caenorhabditis elegansGMPP homolog (Cele Psa1p); O22287 forArabidopsis thaliana GMPP (Atha Psa1p); Q9VN61 for Drosophilia melanogaster GMPP (Dmel Psa1p) and Q9Y5P6 for Homo sapiens GMPPB(Hsap GMPPB).
740 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
the reduced availability of hexose-phosphates as inter-mediates in the biosynthetic pathway of GDP-mannose,could be suppressed by increased dosage of the KlPSA1gene. To this aim the rag5D strain was transformed withthe pKlPSA1 plasmid and the glycosylation phenotypeswere analyzed. The determination of KlPSA1 transcrip-tion level by Northern-blot analysis revealed an in-
creased mRNA level of KlPSA1 in the rag5D straintransformed with the pKlPSA1 plasmid (Fig. 3, panelA). The O-linked glycosylation was then analyzed,employing chitinase as a reporter. In S. cerevisiae, thisprotein is secreted from yeast cells into the growth med-ium and is exclusively O-mannosylated [42]. The electro-phoretic profile of chitinase secreted by the rag5D strain

D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746 741
showed a higher mobility due to the lack of the O-man-nan chain extension (Fig. 3, panel B, lane 2). Surpris-ingly, the chitinase produced by the rag5D strainoverexpressing the KlPSA1 gene was indistinguishablefrom the chitinase of wild-type cells (Fig. 3, panel B,lanes 3 and 1, respectively).
In order to investigate whether the N-glycosylationdefects of this mutant were also suppressed, we usedinvertase as a reporter protein. The invertase extractedfrom the K. lactis wild-type strain appeared, on nativegel electrophoresis, as a diffuse band (in the upper
Fig. 3. KlPSA1 overexpression restores O-glycosylation but not N-glycosylation in Klrag5D strains. Panel A, Northern analysis ofKlPSA1 expression in wild-type strain JA6 transformed with the emptyvector pYG131/1 (lane 1), Klrag5D cells transformed with the controlvector pYG131/1 (lane 2) and Klrag5D transformed with pKlPSA1(lane 3). The same amount of RNA (30 lg) was loaded on each lane.The ethidium bromide-stained gel of the autoradiogram is shown inthe lower part of the panel. Densitometric quantification of mRNAwas performed by the computer program Phoretix 1D, and mRNAloading was normalized using the rRNAs bands. Panel B, Immunoblotanalysis of chitinase. Chitinase was isolated from the culture mediaand subjected to SDS–PAGE separation and then immunoreactedwith chitinase antibody. In lane 1, wild-type strain JA6 transformedwith the empty vector pYG131/1, in lane 2, Klrag5D cells transformedwith the control vector pYG131/1 and Klrag5D transformed withpKlPSA1 in lane 3. Molecular-mass marker is on the right, plainarrow. Panel C, Effect of KlPSA1 overexpression on native invertaseglycosylation of rag5D strain. Cells were grown in YPD and total cellproteins corresponding to 1 · 108 cells were subjected to native gelelectrophoresis (3.5% polyacrylamide) and stained for invertaseactivity. Fully glycosylated invertase is present in lane 1 (wild-typestrain JA6), under-glycosylated invertase is produced by rag5D cellsoverexpressing or not the KlPSA1 gene (lanes 2 and 3, respectively).
part of the gel) due to the heterogeneous N-linked gly-cosylation of the protein (Fig. 3, panel C, lane 1).When the GDP-mannose pyrophosphorylase wasoverexpressed in the rag5D strain, the invertase re-sulted as a diffuse band, with a higher mobility ascompared to the wild-type counterpart, and identicalto the one produced by the rag5D cells (Fig. 3, panelC, lanes 2 and 3, respectively). These data indicatethat the defect of N-linked glycosylation, in contrastto the O-glycosylation defect, was not suppressed bythe increased dosage of KlPSA1 in the cells lackingRag5p activity. This conclusion was also supportedby the inability of the KlPSA1 overexpression to sup-press the growth defects of rag5D cells on a mediumcontaining hygromycin B (data not shown); such sen-sitivity is regarded as diagnostic for alterations in theN-glycosylation process [43].
3.3. Overexpression of KlPSA1 causes cell wall
alterations
Yeast mutants altered in Golgi-specific glycoproteinprocessing are characterized by resistance to sodiumorthovanadate and altered sensitivity to the amino-glycoside antibiotic hygromycin-B and to SDS [43–45].We analyzed these phenotypes in the JA6 wild-typestrain transformed with the empty vector and with thepKlPSA1 construct. Serial dilution droplets from over-night cultures of each of the strains were plated onYPDE containing various amounts of hygromycin Bor SDS (Fig. 4, panel A). The increased dosage of thepyrophosphorylase conferred resistance to hygromycinB since the strain overexpressing KlPSA1 was still ableto grow on plates containing 30 lg ml�1 of the antibi-otic, whereas the growth of the JA6 strain, carryingthe empty vector, was almost completely inhibited.These data are in agreement with the observation thatcells carrying mutations affecting the functionality ofthis kind of enzyme show hygromycin-B sensitivity.The KlPSA1 overexpressing strain was sensitive toSDS at a concentration of 0.02% when compared tothe parental strain (Fig. 4, panel A).
The growth defect on SDS, together with altered sen-sitivity to substances that interfere with the synthesis orassembly of cell wall components, such as Calcofluorwhite, can be used as a diagnostic tool for changes inthe yeast cell wall [46–48]. The overexpression ofKlPSA1 indeed conferred to the cells the ability to growin presence of 50 lg ml�1 of Calcofluor white, whereasthe same strain, transformed with the empty plasmid,showed a completely abolished growth at this drug con-centration (Fig. 4, panel A). In addition, the increaseddosage of GDP-mannose pyrophosphorylase did not al-low the cells to grow in presence of 5 mM caffeine, aninhibitor of kinases and especially of those involved inthe cell wall integrity pathway [49,50].
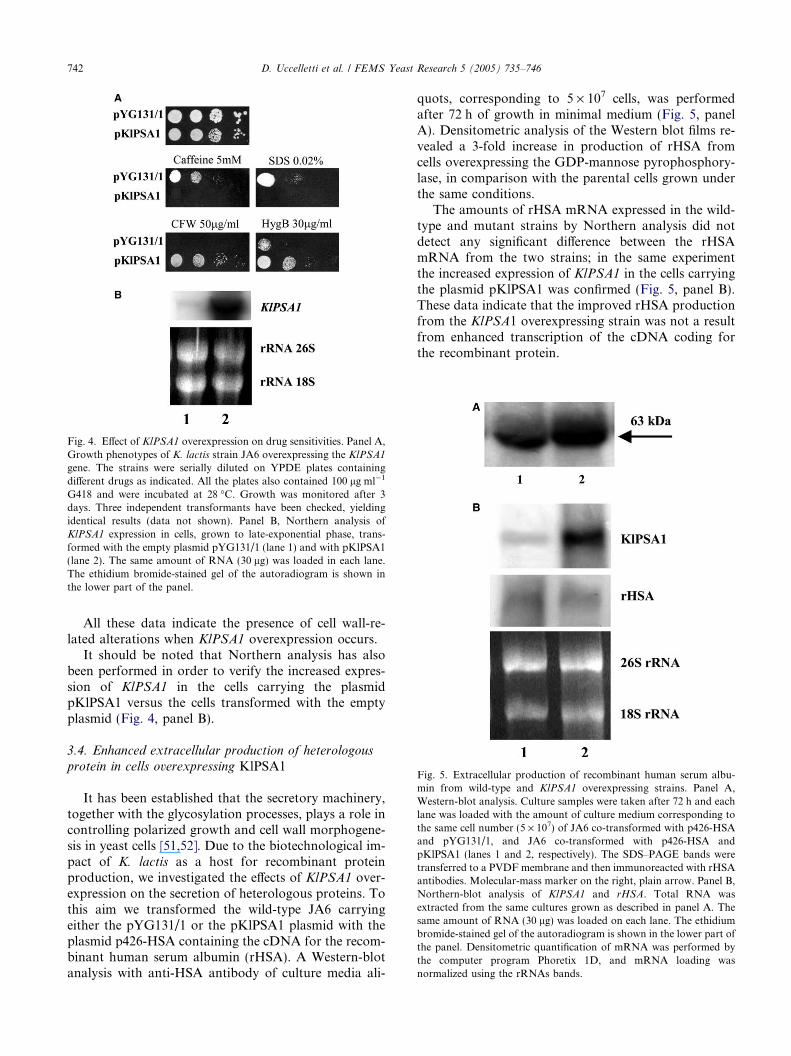
Fig. 5. Extracellular production of recombinant human serum albu-min from wild-type and KlPSA1 overexpressing strains. Panel A,Western-blot analysis. Culture samples were taken after 72 h and eachlane was loaded with the amount of culture medium corresponding tothe same cell number (5 · 107) of JA6 co-transformed with p426-HSAand pYG131/1, and JA6 co-transformed with p426-HSA andpKlPSA1 (lanes 1 and 2, respectively). The SDS–PAGE bands weretransferred to a PVDF membrane and then immunoreacted with rHSAantibodies. Molecular-mass marker on the right, plain arrow. Panel B,Northern-blot analysis of KlPSA1 and rHSA. Total RNA wasextracted from the same cultures grown as described in panel A. Thesame amount of RNA (30 lg) was loaded on each lane. The ethidiumbromide-stained gel of the autoradiogram is shown in the lower part ofthe panel. Densitometric quantification of mRNA was performed bythe computer program Phoretix 1D, and mRNA loading wasnormalized using the rRNAs bands.
Fig. 4. Effect of KlPSA1 overexpression on drug sensitivities. Panel A,Growth phenotypes of K. lactis strain JA6 overexpressing the KlPSA1gene. The strains were serially diluted on YPDE plates containingdifferent drugs as indicated. All the plates also contained 100 lg ml�1
G418 and were incubated at 28 �C. Growth was monitored after 3days. Three independent transformants have been checked, yieldingidentical results (data not shown). Panel B, Northern analysis ofKlPSA1 expression in cells, grown to late-exponential phase, trans-formed with the empty plasmid pYG131/1 (lane 1) and with pKlPSA1(lane 2). The same amount of RNA (30 lg) was loaded in each lane.The ethidium bromide-stained gel of the autoradiogram is shown inthe lower part of the panel.
742 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
All these data indicate the presence of cell wall-re-lated alterations when KlPSA1 overexpression occurs.
It should be noted that Northern analysis has alsobeen performed in order to verify the increased expres-sion of KlPSA1 in the cells carrying the plasmidpKlPSA1 versus the cells transformed with the emptyplasmid (Fig. 4, panel B).
3.4. Enhanced extracellular production of heterologous
protein in cells overexpressing KlPSA1
It has been established that the secretory machinery,together with the glycosylation processes, plays a role incontrolling polarized growth and cell wall morphogene-sis in yeast cells [51,52]. Due to the biotechnological im-pact of K. lactis as a host for recombinant proteinproduction, we investigated the effects of KlPSA1 over-expression on the secretion of heterologous proteins. Tothis aim we transformed the wild-type JA6 carryingeither the pYG131/1 or the pKlPSA1 plasmid with theplasmid p426-HSA containing the cDNA for the recom-binant human serum albumin (rHSA). A Western-blotanalysis with anti-HSA antibody of culture media ali-
quots, corresponding to 5 · 107 cells, was performedafter 72 h of growth in minimal medium (Fig. 5, panelA). Densitometric analysis of the Western blot films re-vealed a 3-fold increase in production of rHSA fromcells overexpressing the GDP-mannose pyrophosphory-lase, in comparison with the parental cells grown underthe same conditions.
The amounts of rHSA mRNA expressed in the wild-type and mutant strains by Northern analysis did notdetect any significant difference between the rHSAmRNA from the two strains; in the same experimentthe increased expression of KlPSA1 in the cells carryingthe plasmid pKlPSA1 was confirmed (Fig. 5, panel B).These data indicate that the improved rHSA productionfrom the KlPSA1 overexpressing strain was not a resultfrom enhanced transcription of the cDNA coding forthe recombinant protein.

D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746 743
We also employed another recombinant protein tostudy the effect of KlPSA1 overexpression on heterolo-gous protein secretion, the glucoamylase (GAA) fromthe salt-tolerant yeast A. adeninivorans. We transformedthe wild-type and the wild-type overexpressing KlPSA1
strains with the plasmid pTS32x-GAA [53] and mea-sured the enzymatic activity in the culture media ofthe transformants. The experiment shown in Fig. 6 is abatch production experiment in which the cultures weremaintained for 4 days. The data, normalized by cellmass, revealed that cells overexpressing KlPSA1 ex-creted about four times more GAA activity than thecontrol cells while the cell mass yields were highly simi-lar for the mutant and wild type.
We also measured the GAA activity present in thecell wall by using intact cells for the essays. Wild-typecells overexpressing KlPSA1 yielded a noticeable GAAactivity (7.8 ± 0.59 U/minGAA/lg d.w.) after 48 h ofgrowth, whereas in the isogenic cells not overexpressingKlPSA1 the GAA activity was undetectable.
Since we used two different reporter proteins withtwo different secretion signals, the enhanced secretorycapabilities of cells overexpressing the GDP-mannosepyrophosphorylase do not seem to be related to a partic-ular recombinant protein but rather originated from thehigh gene dosage of KlPSA1. In order to get some in-sight on the enhanced production of heterologous pro-teins we performed on these cells a porosity assay incomparison with the wild-type counterpart. The assay,based on the polycation-induced leakage of UV-absorb-ing compounds [35], revealed a slight increase in the cellwall porosity of cells overexpressing the KlPSA1 gene,compared to the cells carrying the empty plasmid(46 ± 0.7 versus 34 ± 0.6).
0
5
10
15
20
25
30
0 20 40 60 80 1000
4
8
12
16
20
Time (h)
U/m
inG
AA /
µg d
.w.
mg
d.w
./l
Fig. 6. Batch culture of glucoamylase from wild-type and KlPSA1
overexpressing strains. Extracellular glucoamylase production inKlPSA1 overexpressing cells (JA6 co-transformed with pTS32x-GAAand pKlPSA1) (open squares) and wild-type strain (JA6 co-trans-formed with pTS32x-GGA and pYG131/1) (open triangles) in batchcultures in SM medium containing 2% EtOH. The biomass productionis reported on the right ordinate for the wild-type strain (JA6 co-transformed with pTS32x-GGA and pYG131/1) (closed triangles) andthe cells overexpressing KlPSA1 (JA6 co-transformed with pTS32x-GAA and pKlPSA1) (closed squares). The values reported are themeans of three independent experiments with less than 10% variation.
4. Discussion
The KlPSA1 gene was cloned by complementation ofthe osmotic phenotype displayed by the S. cerevisiae
srb1-1/psa1 mutant. The ability of the K. lactis counter-part to give functional complementation of SRB1/PSA1, mutated alleles strongly supports the notion thatKlPSA1 indeed encodes GDP-mannose pyrophosphory-lase in K. lactis. The protein appears to be highly con-served as shown by the high level of identity andsimilarity with GDP-mannose pyrophosphorylases fromdifferent organisms.
Sequence analysis showed that a 615-bp portion ofthe 3 0 end of the KlMBP1 ORF was localized on thecomplementary strand of the studied K. lactis genomicfragment. These results indicate synteny for this chro-mosomal region in the two yeast species since, in S. cere-
visiae, the MBP1 and PSA1 genes are located onchromosome IV with the same order and orientationas observed in K. lactis [54]. Further investigations areneeded to determine whether PSA1 is a cell cycle-regu-lated gene in K. lactis, as it is in S. cerevisiae and C. albi-cans [3,10] and whether any of the predicted regulatorysites found in the promoter region is indeed involvedin the transcriptional regulation of KlPSA1.
The GDP-mannose pyrophosphorylase activity has arelevant role in replenishing the pool of GDP-mannosenecessary for the outer chain extension of glycoproteinsin the Golgi apparatus; this enzyme has been recently in-volved also in early glycosylation steps such as assemblyof the dolichol-linked core-oligosaccharide and Dol-P-Man formation in S. cerevisiae [20]. These authors dem-onstrated that the availability of GDP-mannose, limitedunder steady-state conditions, could be increased byoverexpressing the GDP-mannose pyrophosphorylase;this compensates the reduced activity of defective mann-osyltransferases. We found that overexpression of theKlPSA1 gene was able to suppress only the O-glycosyl-ation defects, but not the reduced N-glycosylation, occu-ring in rag5D strains [41]. The rag5D cells have a verylow hexokinase activity, less than 5% of the isogenicwild-type strain [40]. The GDP-Man pool is thenprobably undersized in such cells, due to the reducedsynthesis originating from the low availability of hexose-6-phosphates. The overabundance of GDP-mannosepyrophosphorylase, catalyzing the last step in GDP-Man biosynthesis, could only partially replenish theGDP-Man pool under such conditions. The mann-osyltransferases specific for O-glycosylation could stillbe able to function with a reduced GDP-Man poolwhereas the N-mannosyltransferases, possibly havingdifferent Km for the GDP-Man, could not properly com-plete the glycosylation processes.
Another interpretation is suggested by the report fol-lowing the deletion of the KlGDA1 gene, encoding for aGDP/UDPase localized in the Golgi apparatus of

744 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
K. lactis [55]. When mannose, from GDP-mannose, istransferred to mannans by specific mannosyltransfe-rases, the GDP produced in the reaction is hydrolyzedby GDPase to GMP, which then exits the Golgi lumenin a coupled, equimolar exchange with cytosolic GDP-mannose [56]. The in vivo effect of the KlGDA1 deletion,that perturbs the transport of GDP-Man inside theorganelle, results in cells defective only in the O-glyco-sylation, while the N-glycosylated proteins are correctlyprocessed. The unifying picture for all the above datacould then be the existence of separate sub-compart-ments for the O-glycosylation and for the N-glycosyla-tion reactions, together with their respective GDP-Manpools that could be differentially supplied. A recentreport from Ecker and co-workers [57] has shown that,in the ER of S. cerevisiae, O-mannosylation precedesN-glycosylation for the cell wall glycoprotein Ccw5pthat undergoes both modifications; this argues in favorof the above hypothesis.
Finally, GDP-Man may have also a regulatory func-tion in addition to its role as glycosyl donor. It has beenfound in fact to stimulate the in vitro formation of Dol-PP-GlcNAc1-2, the first intermediate in the N-glycosyla-tion reaction [58]. In addition, GDP-mannose inhibitsglucose transfer from Dol-P-Glc to Dol-PP-GlcNAc-Man9 [59]. Since the absence of glucoses in the N-oligo-saccharide significantly impairs its transfer to the acceptorprotein, this could explain the failure of the overexpres-sion of KlPSA1 to suppress the N-glycosylation defectof the rag5D mutant.
The PSA1 gene has been shown to be involved in cellwall biogenesis both in S. cerevisiae and in H. polymor-
pha with mutations resulting in changes in the major cellwall components such as glucans and mannoproteins[16–18]. InK. lactis we found that, by increasing the copynumber of PSA1, the cells showed defects in cell wallassembly as indicated by altered sensitivity to Calcofluorwhite, SDS, caffeine and by altered cell wall porosity.These defects may explain, at least in part, the enhancedheterologous protein secretion observed with rHSA andglucoamylase as reporters. The growth rate of cells carry-ing the pKlPSA1 plasmid was almost identical to thecells carrying the empty plasmid and the strain did notshow any lysis defects (unpublished results), suggestingthat the cell wall changes we observed do not preventpotential biotechnological applications for this strain.
In H. polymorpha, mutations affecting the OPU24
gene, highly similar to PSA1 gene, result in cell wall de-fects and enhanced secretory capabilities similar to thoseobserved in K. lactis when the GDP-mannose pyrophos-phorylase is overexpressed [18]. Since the opu24 mutantcells show also DTT sensitivity, the authors hypothesizethat the influence of OPU24 on ER-associated reten-tion-degradation events of secretory proteins is a resultfrom defective glycosylation of protein component(s)of the ERAD machinery.
At present, the cellular basis underlying the enhancedsecretion phenotype in cells overexpressing the K. lactis
GDP-mannose pyrophosphorylase remains unexplored.It should be taken into account that, probably, glycosyl-ation differences per se are not responsible for the en-hanced secretion since GAA is a glycosylated proteinwhile rHSA has no N-glycosylation sites. The transcrip-tion analysis of rHSA indicated that no significant dif-ferences in the mRNA level were observed betweenwild-type and KlPSA1 overexpressing strains. Thesedata indicate that post-transcriptional events, e.g.changes in the transport and/or control activities to-wards the exterior of the cells are involved in the‘‘super-secretory’’ phenotype, possibly in cooperationwith the increased porosity of the cell wall.
Acknowledgements
We are indebted to Dr. M. Stark and Dr. A. Paszew-ski for the genomic libraries and Dr. P. Goffrini and Dr.M. Wesolowski-Louvel for K. lactis strains.
We thank G. Miloshev and L. Gigova for their sup-port and helpful advice and Francesco Castelli for skill-ful technical assistance.
This work was partially supported by NationalScience Fund, Bulgarian Ministry of Education andScience, Project Grant No. K-1304 and by a grant fromthe Ministero Universita e Ricerca Scientifica e Tecno-logica – Cofin 2002 (Project code 2002052349); D.S.was recipient of UNESCO-MCBN short-term fellow-ship no. SC/RP 206.720.1/3.
References
[1] Orlean, P. (1997) Biogenesis of yeast wall and surface components(Pringle, J.R., Broach, J.R. and Jones, E.W., Eds.), The Molec-ular and Cellular Biology of the Yeast Saccharomyces, Vol. 3, pp.229–362. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
[2] Conklin, P.L., Norris, S.R., Wheeler, G.L., Williams, E.H.,Smirnoff, N. and Last, R.L. (1999) Genetic evidence forthe role of GDP-mannose in plant ascorbic acid (vitaminC) biosynthesis. Proc. Natl. Acad. Sci. USA 96, 4198–4203.
[3] Benton, B.K., Plump, S.D., Roos, J., Lennarz, W.J. and Cross,F.R. (1996) Over-expression of S. cerevisiae G1 cyclins restoresthe viability of alg1 N-glycosylation mutants. Curr. Genet. 29,106–113.
[4] Warit, S., Zhang, N., Short, A., Walmsley, R.M., Oliver, S.G. andStateva, L.I. (2000) Glycosylation deficiency phenotypes resultingfrom depletion of GDP-mannose pyrophosphorylase in two yeastspecies. Mol. Microbiol. 36, 1156–1166.
[5] Lukowitz, W., Nickle, T.C., Meinke, D.W., Last, R.L.,Conklin, P.L. and Somerville, C.R. (2001) Arabidopsis cyt1mutants are deficient in a mannose-1-phosphate guanylyl-transferase and point to a requirement of N-linked glycosyl-ation for cellulose biosynthesis. Proc. Natl. Acad. Sci. USA98, 2262–2267.

D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746 745
[6] Garami, A. and Ilg, T. (2001) Disruption of mannose activation inLeishmania mexicana: GDP-mannose pyrophosphorylase isrequired for virulence, but not for viability. EMBO J. 20, 3657–3666.
[7] Hashimoto, H., Sakakibara, A., Yamasaki, M. and Yoda, K.(1997) Saccharomyces cerevisiae VIG9 encodes GDP-mannosepyrophosphorylase, which is essential for protein glycosylation. J.Biol. Chem. 272, 16308–16314.
[8] Tomlin, G.C., Hamilton, G.E., Gardner, D.C., Walmsley, R.M.,Stateva, L.I. and Oliver, S.G. (2000) Suppression of sorbitoldependence in a strain bearing a mutation in the SRB1/PSA1/
VIG9 gene encoding GDP-mannose pyrophosphorylase by PDE2
overexpression suggests a role for the Ras/cAMP signal-trans-duction pathway in the control of yeast cell-wall biogenesis.Microbiology 146, 2133–2146.
[9] Zhang, N., Gardner, D.C., Oliver, S.G. and Stateva, L.I.(1999) Down-regulation of the expression of PKC1 andSRB1/PSA1/VIG9, two genes involved in cell wall integrityin Saccharomyces cerevisiae, causes flocculation. Microbiology145, 309–316.
[10] Warit, S., Walmsley, R.M. and Stateva, L.I. (1998) Cloning andsequencing of the Candida albicans homologue of SRB1/PSA1/
VIG9, the essential gene encoding GDP-mannose pyrophosphor-ylase in Saccharomyces cerevisiae. Microbiology 144, 2417–2426.
[11] Yoda, K., Kawada, T., Kaibara, C., Fujie, A., Abe, M.,Hashimoto, H., Shimizu, J., Tomishige, N., Noda, Y. andYamasaki, M. (2000) Defect in cell wall integrity of the yeastSaccharomyces cerevisiae caused by a mutation of the GDP-mannose pyrophosphorylase gene VIG9. Biosci. Biotechnol.Biochem. 64, 1937–1941.
[12] Venkov, P.V., Hadjiolov, A.A., Battaner, E. and Schlessinger, D.(1974) Saccharomyces cerevisiae: sorbitol-dependent fragilemutants. Biochem. Biophys. Res. Commun. 56, 599–604.
[13] Kozhina, T., Stateva, L. and Venkov, P. (1979) Genetic analysisof an osmotic sensitive Saccharomyces cerevisiae mutant. Mol.Gen. Genet. 170, 351–354.
[14] Heale, S.M., Stateva, L.I. and Oliver, S.G. (1994) Introduction ofYACs into intact yeast cells by a procedure which shows lowlevels of recombinagenicity and co-transformation. Nucleic AcidsRes. 22, 5011–5015.
[15] Venkov, P.V., Milchev, G.I. and Hadjiolov, A.A. (1975) Rifam-picin susceptibility of ribonucleic acid synthesis in a fragileSaccharomyces cerevisiae mutant. Antimicrob. Agents Chemo-ther. 8, 627–632.
[16] Maerkisch, U., Reuter, G., Stateva, L. and Venkov, P. (1983)Mannan structure analysis of the fragile Saccharomyces cerevisiae
mutant VY1160. Int. J. Biochem. 15, 1373–1377.[17] Blagoeva, J., Stoev, G. and Venkov, P. (1991) Glucan structure in
a fragile mutant of Saccharomyces cerevisiae. Yeast. 7, 455–461.[18] Agaphonov, M.O., Packeiser, A.N., Chechenova, M.B., Choi,
E.S. and Ter-Avanesyan, M.D. (2001) Mutation of the homo-logue of GDP-mannose pyrophosphorylase alters cell wall struc-ture, protein glycosylation and secretion in Hansenula
polymorpha. Yeast 18, 391–402.[19] Shimma, Y., Nishikawa, A., bin Kassim, B., Eto, A. and Jigami,
Y. (1997) A defect in GTP synthesis affects mannose outer chainelongation in Saccharomyces cerevisiae. Mol. Gen. Genet. 256,469–480.
[20] Janik, A., Sosnowska, M., Kruszewska, J., Krotkiewski, H.,Lehle, L. and Palamarczyk, G. (2003) Overexpression of GDP-mannose pyrophosphorylase in Saccharomyces cerevisiae correctsdefects in dolichol-linked saccharide formation and proteinglycosylation. Biochim. Biophys. Acta 1621, 22–30.
[21] Cullen, P.J., Schultz, J., Horecka, J., Stevenson, B.J., Jigami, Y.and Sprague Jr., G.F. (2000) Defects in protein glycosylationcause SHO1-dependent activation of a STE12 signaling pathwayin yeast. Genetics 155, 1005–1018.
[22] Zakrzewska, A., Palamarczyk, G., Krotkiewski, H., Zdebska, E.,Saloheimo, M., Penttila, M. and Kruszewska, J.S. (2003) Over-expression of the gene encoding GTP:mannose-1-phosphateguanyltransferase, mpg1, increases cellular GDP-mannose levelsand protein mannosylation in Trichoderma reesei. Appl. Environ.Microbiol. 69, 4383–4389.
[23] Hussein, L., Elasyed, S. and Fod, S. (1989) Reduction of lactosein milk by purified lactase produced by Kluyveromyces lactis. J.Food Prot. 52, 30–34.
[24] Van den Berg, J.A., van der Laken, K.J., van Ooyen, A.J.J.,Renniers, T.C.H.M., Rietveld, K., Schaap, A., Brake, A.J.,Bishop, R.J., Schultz, K., Moyer, D., Richman, M. and Shuster,J.R. (1990) Kluyveromyces as a host for heterologous geneexpression: expression and secretion of prochymosin. Biotechnol-ogy 8, 135–139.
[25] Swinkels, B.W., van Ooyen, A.J. and Bonekamp, F.J. (1993)The yeast Kluyveromyces lactis as an efficient host forheterologous gene expression. Antonie Van Leeuwenhoek 64,187–201.
[26] Gellissen, G. and Hollenberg, C.P. (1997) Application of yeasts ingene expression studies: a comparison of Saccharomyces cerevi-
siae, Hansenula polymorpha and Kluyveromyces lactis. Gene 190,87–97.
[27] Ferminan, E. and Dominguez, A. (1998) Heterologous proteinsecretion directed by a repressible acid phosphatase system ofKluyveromyces lactis: characterization of upstream region-acti-vating sequences in the KlPHO5 gene. Appl. Environ. Microbiol.64, 2403–2408.
[28] Bartkeviciute, D. and Sasnauskas, K. (2003) Studies on yeastKluyveromyces lactis mutations conferring super-secretion ofrecombinant proteins. Yeast 20, 1–11.
[29] Johnston, J.R. (1994) In: Molecular Genetics of Yeast: a PracticalApproach (Rickwood, D. and Hames, B.D., Eds.), The PracticalApproach Series, Vol. 141. Oxford University Press, Oxford, UK.
[30] Christianson, T.W., Sikorski, R.S., Dante, M., Shero, J.H. andHieter, P. (1992) Multifunctional yeast high-copy-number shuttlevectors. Gene 110, 119–122.
[31] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) MolecularCloning: a Laboratory Manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, New York.
[32] Gietz, R.D., Schiestl, R.H., Willems, A.R. and Woods, R.A.(1995) Studies on the transformation of intact yeast cells by theLiAc/SS-DNA/PEG procedure. Yeast 11, 355–360.
[33] Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang,Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.Nucleic Acids Res. 25, 3389–3402.
[34] Uccelletti, D., Farina, F., Morlupi, A. and Palleschi, C. (1999)Mutants of Kluyveromyces lactis with altered protein glycosyla-tion are affected in cell wall morphogenesis. Res. Microbiol. 150,5–12.
[35] De Nobel, J.G., Klis, F.M., Munnik, T., Priem, J. and van denEnde, H. (1990) An assay of relative cell wall porosity inSaccharomyces cerevisiae, Kluyveromyces lactis and Schizosaccha-
romyces pombe. Yeast 6, 483–490.[36] Morlino, G.B., Tizzani, L., Fleer, R., Frontali, L. and Bianchi,
M.M. (1999) Inducible amplification of gene copy number andheterologous protein production in the yeast Kluyveromyces lactis.Appl. Environ. Microbiol. 65, 4808–4813.
[37] Schmitt, M.E., Brown, T.A. and Trumpower, B.L. (1990) A rapidand simple method for preparation of RNA from S. cerevisiae.Nucleic Acids Res. 18, 3091–3092.
[38] Saliola, M., Mazzoni, C., Solimando, N., Crisa, A., Falcone, C.,Jung, G. and Fleer, R. (1999) Use of the KlADH4 promoter forethanol-dependent production of recombinant human serumalbumin in Kluyveromyces lactis. Appl. Environ. Microbiol. 65,53–60.

746 D. Uccelletti et al. / FEMS Yeast Research 5 (2005) 735–746
[39] Ohta, A., Chibana, H., Arisawa, M. and Sudoh, M. (2000) TheVIG9 gene products from the human pathogenic fungi Candidaalbicans and Candida glabrata encode GDP-mannose pyrophos-phorylase. Biochim. Biophys. Acta 1475, 265–272.
[40] Goffrini, P., Ficarelli, A. and Ferrero, I. (1995) Hexokinaseactivity is affected in mutants of Kluyveromyces lactis resistant toglucose repression. Microbiology 141, 441–447.
[41] Donnini, C., Farina, F., Neglia, B., Compagno, C., Uccelletti,D., Goffrini, P. and Palleschi, C. (2004) Improved productionof heterologous proteins by a glucose repression-defectivemutant of Kluyveromyces lactis. Appl. Environ. Microbiol. 70,2632–2638.
[42] Orlean, P., Kuranda, M.J. and Albright, C.F. (1991) Analysis ofglycoproteins from Saccharomyces cerevisiae. Methods Enzymol.194, 682–697.
[43] Dean, N. (1995) Yeast glycosylation mutants are sensitive toaminoglycosides. Proc. Natl. Acad. Sci. USA 92, 1287–1291.
[44] Ballou, L., Hitzeman, R.A., Lewis, M.S. and Ballou, C.E. (1991)Vanadate-resistant yeast mutants are defective in protein glyco-sylation. Proc. Natl. Acad. Sci. USA 88, 3209–3212.
[45] Kanik-Ennulat, C., Montalvo, E. and Neff, N. (1995) Sodiumorthovanadate-resistant mutants of Saccharomyces cerevisiae
show defects in Golgi-mediated protein glycosylation, sporulationand detergent resistance. Genetics 140, 933–943.
[46] Ram, A.F., Wolters, A., Ten Hoopen, R. and Klis, F.M. (1994) Anew approach for isolating cell wall mutants in Saccharomyces
cerevisiae by screening for hypersensitivity to calcofluor white.Yeast 10, 1019–1030.
[47] Van der Vaart, J.M., Caro, L.H., Chapman, J.W., Klis, F.M. andVerrips, C.T. (1995) Identification of three mannoproteins in thecell wall of Saccharomyces cerevisiae. J. Bacteriol. 177, 3104–3110.
[48] Moukadiri, I., Armero, J., Abad, A., Sentandreu, R. and Zueco,J. (1997) Identification of a mannoprotein present in the innerlayer of the cell wall of Saccharomyces cerevisiae. J. Bacteriol. 179,2154–2162.
[49] Hohmann, S. (2002) Osmotic stress signaling and osmoadaptationin yeasts. Microbiol. Mol. Biol. Rev. 66, 300–372.
[50] Lorberg, A., Schmitz, H.P., Gengenbacher, U. and Heinisch, J.J.(2003) KlROM2 encodes an essential GEF homologue in Kluy-
veromyces lactis. Yeast 20, 611–624.[51] Mondesert, G., Clarke, D.J. and Reed, S.I. (1997) Identifi-
cation of genes controlling growth polarity in the buddingyeast Saccharomyces cerevisiae: a possible role of N-glycosyl-ation and involvement of the exocyst complex. Genetics 147,421–434.
[52] Sheu, Y.J., Barral, Y. and Snyder, M. (2000) Polarized growthcontrols cell shape and bipolar bud site selection in Saccharomy-
ces cerevisiae. Mol. Cell. Biol. 20, 5235–5247.[53] Bui, D.M., Kunze, I., Horstmann, C., Schmidt, T., Breunig, K.D.
and Kunze, G. (1996) Expression of the Arxula adeninivorans
glucoamylase gene in Kluyveromyces lactis. Appl. Microbiol.Biotechnol. 45, 102–106.
[54] Gardner, D.C., Tomlin, G.C., Cele, T., Hamilton, G.A., James,C.M., Stateva, L.I. and Oliver, S.G. (1996) Physical mapping of acentromere-proximal region of chromosome IV-L defines theplacement of genes USO1, MBP1, PSA1 and SLC1. Yeast 12,411–413.
[55] Lopez-Avalos, M.D., Uccelletti, D., Abeijon, C. and Hir-schberg, C.B. (2001) The UDPase activity of the Kluyver-
omyces lactis Golgi GDPase has a role in uridine nucleotidesugar transport into Golgi vesicles. Glycobiology 11, 413–422.
[56] Berninsone, P. and Hirschberg, C.B. (2000) Nucleotide sugartransporters of the Golgi apparatus. Curr. Opin. Struct. Biol. 10,542–547.
[57] Ecker, M., Mrsa, V., Hagen, I., Deutzmann, R., Strahl, S. andTanner, W. (2003) O-mannosylation precedes and potentiallycontrols the N-glycosylation of a yeast cell wall glycoprotein.EMBO Rep. 4, 628–632.
[58] Kean, E.L. (1980) Stimulation by GDP-mannose of the biosyn-thesis of N-acetylglucosaminylpyrophosphoryl polyprenols by theretina. J. Biol. Chem. 255, 1921–1927.
[59] Ronin, C., Caseti, C. and Bouchilloux, S. (1981) Transfer ofglucose in the biosynthesis of thyroid glycoproteins. I. Inhibitionof glucose transfer to oligosaccharide lipids by GDP-mannose.Biochim. Biophys. Acta 674, 48–57.
[60] Stateva, L.I., Oliver, S.G., Trueman, L.J. and Venkov, P.V. (1991)Cloning and characterization of a gene which determines osmoticstability in Saccharomyces cerevisiae. Mol. Cell. Biol. 11, 4235–4243.
[61] Breunig, K.D. and Kuger, P. (1987) Functional homologybetween the yeast regulatory proteins GAL4 and LAC9:LAC9-mediated transcriptional activation in Kluyveromyces
lactis involves protein binding to a regulatory sequencehomologous to the GAL4 protein-binding site. Mol. Cell.Biol. 7, 4400–4406.
[62] Prior, C., Mamessier, P., Fukuhara, H., Chen, X.J. and Weso-lowski-Louvel, M. (1993) The hexokinase gene is required fortranscriptional regulation of the glucose transporter gene RAG1
in Kluyveromyces lactis. Mol. Cell. Biol. 13, 3882–3889.
Related Documents











