Kidney International, Vol. 62 (2002), pp. 1160–1170 Enhanced MCP-1 expression during ischemia/reperfusion injury is mediated by oxidative stress and NF-B FION L. SUNG,TONG Y. ZHU,KATHY K.W. AU-YEUNG,YAW L. SIOW, and KARMIN O Department of Pharmacology, Faculty of Medicine, University of Hong Kong, Hong Kong, China Enhanced MCP-1 expression during ischemia/reperfusion in- ciated with high mortality and morbidity [1, 2]. The in- jury is mediated by oxidative stress and NF-B. sults causing renal injury include severe hypotension, Background. Renal ischemia/reperfusion injury is a major interruption of renal blood flow while operating on the cause of acute renal failure in both native kidneys and renal kidney in situ, renal artery stenosis, supra-renal aortic allografts. One important feature of such injury is monocyte/ macrophage infiltration into the renal tissue. The infiltration of cross-clamping and renal pedicular damage [1–3]. In ad- monocytes/macrophages can be induced by chemotactic factors dition, ischemia/reperfusion injury is the most common produced by renal cells. Monocyte chemoattractant protein-1 cause of delayed function of renal allografts and is associ- (MCP-1) is a potent chemoattractant protein for monocyte ated with poor long-term function and survival of the graft recruitment. The objective of the present study was to investi- [4, 5]. It has been suggested that besides genetic dissimi- gate mechanisms of elevated MCP-1 expression in rat kidney during ischemia/reperfusion injury. larity between the donor and recipient, non-immunologi- Methods. The left kidney was subjected to one hour of isch- cal factors are important in the survival of the graft [6, 7]. emia followed by reperfusion for various time periods. The Ischemia/reperfusion injury is a non-immunological fac- expression of MCP-1 mRNA was determined by nuclease pro- tor and is unavoidable in cadaveric renal transplantation. tection assay and MCP-1 protein was identified by immunohis- tochemistry. Activation of a nuclear factor-kappa B (NF-B) Evidence suggests that infiltrated leukocytes including was determined by electrophoretic mobility shift assay and the macrophages may play an important role in the develop- level of lipid peroxides in the kidney was measured. ment of ischemia/reperfusion-mediated injury in the kid- Results. There was a significant increase in MCP-1 expres- ney [8–10]. These infiltrated macrophages can in turn sion in the ischemia/reperfusion kidney 2 hours after reperfu- sion (210% of the control). This increase was accompanied by potentiate the inflammatory response by producing vaso- activation of NF-B, suggesting that this transcription factor constrictors, cytokines and toxic mediators such as free might be involved in the event. The number of monocytes radicals. Increased numbers of monocytes and macro- was significantly elevated in the kidney 3 days after ischemia/ phages were found in transplanted kidneys of patients reperfusion. Pretreatment of rats with NF-B inhibitors not only prevented NF-B activation induced by ischemia/reperfu- suffering from acute tubular necrosis or graft rejection sion, but also inhibited MCP-1 mRNA expression. Further [10] as well as in rat kidneys subjected to ischemia/reper- analysis revealed that oxidative stress and increased IB- fusion injury [11]. Ischemia/reperfusion also makes the phosphorylation might be an underlying mechanism for NF-B graft more prone to rejection by increasing the major activation and subsequent MCP-1 mRNA expression in the ischemia/reperfusion kidney. histocompatibility complex antigen expression [12] and Conclusion. The present study clearly demonstrates that en- adhesion molecule expression [13]. hanced MCP-1 expression in rat kidney during ischemia/reper- The infiltration of monocytes/macrophages is facili- fusion injury is mediated by NF-B activation and oxidative tated by chemotactic factors and/or adhesion molecules. stress. Elevated MCP-1 expression might be responsible for increased monocyte infiltration in the injured kidney. Monocyte chemoattractant protein-1 (MCP-1) is a po- tent chemokine that stimulates monocyte migration into the intima of arterial walls and organs [14, 15]. Little Ischemia/reperfusion injury, a result of decreased MCP-1 is detectable in normal human kidneys. However, blood flow in the kidney followed by reperfusion, is asso- MCP-1 gene expression is greatly increased in kidneys of patients with acute renal transplant rejection [16–18]. Key words: kidney, inflammatory response, monocyte chemoattractant Although the mechanisms of this elevation are not fully protein-1, nuclear factor-B, organ storage, cadaveric renal trans- plantation. understood, ischemia/reperfusion injury-stimulated MCP-1 gene expression may be responsible for enhanced mono- Received for publication November 12, 2001 cyte infiltration and subsequent graft damage. We recently and in revised form April 4, 2002 Accepted for publication May 10, 2002 reported that very low-density lipoprotein (VLDL)- or lipoprotein-X–stimulated MCP-1 expression in mesan- 2002 by the International Society of Nephrology 1160

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Kidney International, Vol. 62 (2002), pp. 1160–1170
Enhanced MCP-1 expression during ischemia/reperfusioninjury is mediated by oxidative stress and NF-�B
FION L. SUNG, TONG Y. ZHU, KATHY K.W. AU-YEUNG, YAW L. SIOW, and KARMIN O
Department of Pharmacology, Faculty of Medicine, University of Hong Kong, Hong Kong, China
Enhanced MCP-1 expression during ischemia/reperfusion in- ciated with high mortality and morbidity [1, 2]. The in-jury is mediated by oxidative stress and NF-�B. sults causing renal injury include severe hypotension,
Background. Renal ischemia/reperfusion injury is a major interruption of renal blood flow while operating on thecause of acute renal failure in both native kidneys and renalkidney in situ, renal artery stenosis, supra-renal aorticallografts. One important feature of such injury is monocyte/
macrophage infiltration into the renal tissue. The infiltration of cross-clamping and renal pedicular damage [1–3]. In ad-monocytes/macrophages can be induced by chemotactic factors dition, ischemia/reperfusion injury is the most commonproduced by renal cells. Monocyte chemoattractant protein-1 cause of delayed function of renal allografts and is associ-(MCP-1) is a potent chemoattractant protein for monocyte
ated with poor long-term function and survival of the graftrecruitment. The objective of the present study was to investi-[4, 5]. It has been suggested that besides genetic dissimi-gate mechanisms of elevated MCP-1 expression in rat kidney
during ischemia/reperfusion injury. larity between the donor and recipient, non-immunologi-Methods. The left kidney was subjected to one hour of isch- cal factors are important in the survival of the graft [6, 7].
emia followed by reperfusion for various time periods. TheIschemia/reperfusion injury is a non-immunological fac-expression of MCP-1 mRNA was determined by nuclease pro-tor and is unavoidable in cadaveric renal transplantation.tection assay and MCP-1 protein was identified by immunohis-
tochemistry. Activation of a nuclear factor-kappa B (NF-�B) Evidence suggests that infiltrated leukocytes includingwas determined by electrophoretic mobility shift assay and the macrophages may play an important role in the develop-level of lipid peroxides in the kidney was measured.
ment of ischemia/reperfusion-mediated injury in the kid-Results. There was a significant increase in MCP-1 expres-ney [8–10]. These infiltrated macrophages can in turnsion in the ischemia/reperfusion kidney 2 hours after reperfu-
sion (210% of the control). This increase was accompanied by potentiate the inflammatory response by producing vaso-activation of NF-�B, suggesting that this transcription factor constrictors, cytokines and toxic mediators such as freemight be involved in the event. The number of monocytes
radicals. Increased numbers of monocytes and macro-was significantly elevated in the kidney 3 days after ischemia/phages were found in transplanted kidneys of patientsreperfusion. Pretreatment of rats with NF-�B inhibitors not
only prevented NF-�B activation induced by ischemia/reperfu- suffering from acute tubular necrosis or graft rejectionsion, but also inhibited MCP-1 mRNA expression. Further [10] as well as in rat kidneys subjected to ischemia/reper-analysis revealed that oxidative stress and increased I�B-�
fusion injury [11]. Ischemia/reperfusion also makes thephosphorylation might be an underlying mechanism for NF-�Bgraft more prone to rejection by increasing the majoractivation and subsequent MCP-1 mRNA expression in the
ischemia/reperfusion kidney. histocompatibility complex antigen expression [12] andConclusion. The present study clearly demonstrates that en- adhesion molecule expression [13].
hanced MCP-1 expression in rat kidney during ischemia/reper-The infiltration of monocytes/macrophages is facili-fusion injury is mediated by NF-�B activation and oxidative
tated by chemotactic factors and/or adhesion molecules.stress. Elevated MCP-1 expression might be responsible forincreased monocyte infiltration in the injured kidney. Monocyte chemoattractant protein-1 (MCP-1) is a po-
tent chemokine that stimulates monocyte migration intothe intima of arterial walls and organs [14, 15]. LittleIschemia/reperfusion injury, a result of decreasedMCP-1 is detectable in normal human kidneys. However,blood flow in the kidney followed by reperfusion, is asso-MCP-1 gene expression is greatly increased in kidneysof patients with acute renal transplant rejection [16–18].
Key words: kidney, inflammatory response, monocyte chemoattractantAlthough the mechanisms of this elevation are not fullyprotein-1, nuclear factor-�B, organ storage, cadaveric renal trans-
plantation. understood, ischemia/reperfusion injury-stimulated MCP-1gene expression may be responsible for enhanced mono-Received for publication November 12, 2001cyte infiltration and subsequent graft damage. We recentlyand in revised form April 4, 2002
Accepted for publication May 10, 2002 reported that very low-density lipoprotein (VLDL)- orlipoprotein-X–stimulated MCP-1 expression in mesan- 2002 by the International Society of Nephrology
1160

Sung et al: MCP-1 expression in I/R injury 1161
gial cells might be responsible for increased macrophage Analysis of MCP-1 mRNAinfiltration and foam cell formation in the kidney of Total RNA was isolated from kidneys with TRIzolpatients with dyslipidemia [19, 20]. Reagent (Invitrogen, Groningen, The Netherlands) and
The expression of MCP-1 and other inflammatory fac- used for the nuclease protection assay (Ambion, Inc.,tors can be up-regulated by a transcription factor, nuclear Austin, TX, USA). In brief, the isolated RNA (10 �g)factor-�B (NF-�B) [21]. The promoter region of the was hybridized with 32P-end labeled MCP-1 oligonucleo-MCP-1 gene consists of several putative binding sites tide probe (Clontech Laboratory Inc., Palo Alto, CA,for transcription factors including two binding sites for USA) overnight at 30�C followed by nuclease digestionNF-�B. This transcription factor is normally present in according to the manufacturer’s instruction. A positivethe cytosol in an inactive form associated with an inhibi- control, 28S rRNA oligonucleotide probe (Ambion Inc.)
was used as an internal control. After digestion, the pro-tory protein, named I�B. NF-�B can be rapidly activatedtected fragments were resolved on a denaturing poly-by many pathogenic stimuli, such as tumor necrosis fac-acrylamide gel (12%) containing 8 mol/L urea followedtor-� (TNF-�), interleukin-1 (IL-1), oxidized low-densityby exposure to X-ray film [20]. The bands correspondinglipoprotein (ox-LDL) and homocysteine [22–25]. Uponto MCP-1 mRNA or 28S rRNA in the resulting autora-stimulation, I�B is phosphorylated and then degradeddiograph were analyzed using a gel documentation sys-rapidly, releasing the free NF-�B that translocates intotem (Bio-Rad Gel Doc1000 and Multi-Analyst� versionthe nucleus and binds to the �B binding site on the1.1; Bio-Rad, Richmond, CA, USA). Values were ex-promoter regions of genes, leading to up-regulation ofpressed as relative expression of MCP-1 mRNA normal-these corresponding genes. Although several inhibitorized to 28S rRNA levels.proteins have been identified, I�B-� is the best-charac-
As a comparison, semiquantitative reverse transcrip-terized form of I�B. It was reported that inhibition oftion (RT) also was performed. The primers for MCP-1NF-�B activation by NF-�B decoy oligodeoxynucleo-mRNA used in the polymerase chain reactions (PCR)tides reduces monocyte infiltration in renal allografts [26].were (1) 5�-ATC ACC AGC AGC AGG TGT CCCAlthough MCP-1 may play a crucial role in the recruit-AAA GAA GCT-3�; (2) 5�-AGA AGT GCT TGA GGTment of monocytes/macrophages into the kidney afterGGT TGT GGA AAA GAG-3� [19], and those forischemia/reperfusion, the underlying mechanisms for el-glyceraldehydes-3-phosphate dehydrogenase (GAPDH)evated MCP-1 expression are not well understood. Thewere (1) 5�-AAT GCA TCC TGC ACC ACC AA-3�aim of the present study was to investigate mechanismsand (2) 5�-GTA GCC ATA TTC CAT TGT CAT A-3�of elevated MCP-1 expression in the rat kidney during(Invitrogen). The reaction mixture for PCR containedischemia/reperfusion injury.1.5 mmol/L MgCl2, 0.2 mmol/L dNTP, 0.4 �mol/L5�primer, 0.4 �mol/L 3�primer, 2 units Taq DNA poly-merase and 1 �g cDNA product from reverse transcrip-METHODStion reaction. After an initial denaturation for five min-Renal ischemia/reperfusionutes at 95�C, 35 cycles of PCR amplification (95�C for
Male Sprague-Dawley rats (200 to 250 g) were anes- 45 seconds, 55�C for 45 seconds and 72�C for 1.5 min)thetized with sodium pentobarbital (50 mg/kg) intraperi- was carried out followed by an additional seven-minutetoneally. Renal ischemia was induced by clamping the extension at 72�C. The cDNA amplification was linearleft renal artery for 60 minutes with a non-traumatic up to 35 cycles of the PCR reaction. The products ofvascular clamp through a midline abdominal incision. PCR were separated by 1.8% agarose gel (containingAfter the clamp was removed, reperfusion was confirmed ethidium bromide) electrophoresis and visualized undervisually. During reperfusion, rats were allowed free ac- UV using gel documentation system (Bio-Rad Gelcess to water and a standard chow diet. At various time Doc1000). The GAPDH was used as an internal standardpoints after reperfusion, the left kidney was harvested to verify equal PCR product loading for each experi-under anesthesia. A sham-operated group of rats were ment. After the RT-PCR reaction, the MCP-1 cDNAsubjected to the same surgical procedure without induc- signal was normalized by comparing it to a GAPDHing ischemia/reperfusion and were sacrificed at corre- cDNA signal from the same sample [27]. The values aresponding time points. Results obtained from sham-oper- expressed as a ratio of MCP-1 to GAPDH. In negativeated kidneys were used as controls. The experimental controls, reverse transcriptase was omitted during re-procedure was approved by the Committee for the Use verse transcription reaction.of Live Animal in Teaching and Research of the Univer-
Immunohistochemistrysity of Hong Kong. For comparison, right kidneys (with-out ischemia/reperfusion) also were harvested for deter- After ischemia/reperfusion, kidney slices were immer-
sion-fixed in 10% neutral-buffered formalin overnightmination of MCP-1 expression.

Sung et al: MCP-1 expression in I/R injury1162
and then embedded in paraffin. Sequential 5-�m paraf- The products of PCR were separated by 1.8% agarosegel (containing ethidium bromide) electrophoresis andfin-embedded sections were immunostained using rabbit
polyclonal antibodies (1:100) against rat MCP-1 (Pepro- visualized under UV using a gel documentation system(Bio-Rad Gel Doc 1000). GAPDH was used as an inter-Tech EC Ltd., London, UK). Endogenous peroxidase
was blocked with 0.3% H2O2 for 20 minutes. The second- nal standard to verify equal PCR product loading foreach experiment. The I�B-� signal was normalized byary antibodies for immunostaining were biotin-conju-
gated anti-rabbit immunoglobulins (1:200; Dako, Glos- comparison to the GAPDH signal from the same sample.Values were expressed as a ratio of I�B-� to GAPDH.trup, Denmark). For detection of monocytes/macrophages
in the kidney, mouse monoclonal antibody against ED-1Western immunoblotting analysis of I�B-� protein(1:100) was used (Serotec, Oxford, UK). The secondary
antibodies for immunostaining were biotin-conjugated The renal I�B-� protein levels were determined byWestern immunoblotting analysis as previous describedanti-mouse immunoglobulins (1:200; Dako). After incuba-
tion with streptavidin-horse radish peroxidase conjugate [25]. Briefly, tissue proteins were separated by sodiumdodecyl sulfate-polyacrylamide gel (12.5%) electropho-(Zymed Laboratories, Inc., San Francisco, CA, USA),
visualization of peroxidase localization was performed us- resis (SDS-PAGE) followed by electrophoretic transferof proteins from the gel to a nitrocellulose membrane.ing diaminobenzidine-hydrogen peroxide (DAB-H2O2)
substrate to give a brown color. After immunostaining, The membrane was probed with rabbit anti-I�B-� poly-clonal antibodies or anti-phospho-I�B-� (Ser32) poly-the slides were counterstained with Mayer’s hematoxy-
lin. For a negative control, normal rabbit IgG was used clonal antibodies (New England Biolabs Inc., Beverly,MA, USA). Bands corresponding to I�B-� or serineas primary antibody.phosphorylated I�B-� proteins were visualized by using
Electrophoretic mobility shift assay anti-rabbit immunoglobulins and enhanced chemilumi-nescence reagents (Amersham, Buckinghamshire, UK).Nuclear proteins were isolated by a previously de-
scribed method [28]. Electrophoretic mobility shift assay The intensity of the bands was analyzed with a gel docu-mentation system (Bio-Rad Gel Doc 1000 and Multi-(EMSA) was performed to determine the NF-�B/DNA
binding activity [25]. In brief, nuclear proteins (10 �g) Analyst� version 1.1). For the detection of phospho-I�B-�(Tyr42), rabbit anti-I�B-� polyclonal antibodies werewere incubated with the reaction buffer for 15 minutes at
room temperature followed by incubation with 32P-end- first pre-absorbed to agarose-immobilize protein-A aga-rose beads (Oncogene Research Products, Boston, MA,labeled oligonucleotide containing the consensus se-
quence for the NF-�B/DNA binding site (5�GGGGAC USA) and an aliquot of kidney homogenate was added toprecipitate all I�B-� proteins. The immunoprecipitatedTTTCC-3�; Invitrogen). The reaction mixture was sepa-
rated in a non-denaturing polyacrylamide gel (6%) that proteins were subjected to electrophoresis on an SDS-polyacrylamide gel (12.5%) followed by an electro-was later exposed to the x-ray film at �70�C. The binding
of labeled oligonucleotide to nuclear proteins was phoretic transfer onto a nitrocellulose membrane. Themembrane was then probed with phospho-tyrosine mono-blocked by adding unlabeled specific competitor oligo-
nucleotide to the reaction mixture. This was to confirm clonal antibody (P-Tyr-100; Biotinylated; New EnglandBiolabs Inc.). Bands corresponding to tyrosine phos-that the binding of 32P-end-labeled oligonucleotide to
NF-�B was sequence-specific. phorylated I�B-� protein were visualized by using horse-radish peroxidase (HRP)-conjugated anti-biotin antibody
Analysis of I�B-� mRNA expression and enhanced chemiluminescence reagents (Amersham).The immunoprecipitated tyrosine phosphorylated I�B-�The expression of I�B-� mRNA in the kidney was
examined by semiquantitative RT-PCR analysis [29]. protein migrated during electrophoresis at the same posi-tion as the 39 kD protein.The primers for I�B-� used in the PCR reactions (1)
5�-CAT GAA GAG AAG ACA CTG ACC ATGDetermination of lipid peroxidation by measuringGAA-3�; (2) 5�-TGG ATA GAG GCT AAG TGTthiobarbituric acid reactive substances (TBARS)AGA CAC GT-3� were synthesized by Clontech Labora-
tory Inc. [30]. RT-PCR was performed. The total RNA Malondialdehyde (MDA) level in the ischemic/reper-fused kidney was measured as described previously(2 �g) was used in the reverse transcription reaction.
The reaction mixture for PCR contained 1.5 mmol/L [31, 32]. A portion of the kidney was homogenized fol-lowed by centrifugation at 600 � g for 15 minutes. AnMgCl2, 0.2 mmol/L dNTP, 0.4 �mol/L 5�primer, 0.4
�mol/L 3�primer, 2 units Taq DNA polymerase and 1 �g aliquot of supernatant (400 �L) was added to the reac-tion mixture containing 0.2 mL 8.1% SDS, 1.5 mL 20%cDNA product from the reverse transcription reaction.
After 28 cycles of PCR amplification (93�C for 30 sec- acetic acid, 1.5 mL 0.8% thiobarbituric acid and 0.6 mLwater. The reaction mixture was heated up to 95�C foronds, 60�C for 45 seconds and 72�C for 45 seconds), an
additional 10-minute extension was carried out at 72�C. 60 minutes. At the end of the reaction, 1 mL water and

Sung et al: MCP-1 expression in I/R injury 1163
5 mL of n-butanol-pyridine mixture (15:1, vol/vol) were Similar results were obtained for the contralateral kidneyat all reperfusion time points (data not shown).added. The mixture was vigorously shaken and centri-
fuged at 2000 � g for 15 minutes. The absorbance of theDistribution of MCP-1 protein in the kidneysupernatant was read at 532 nm. MDA was used as the
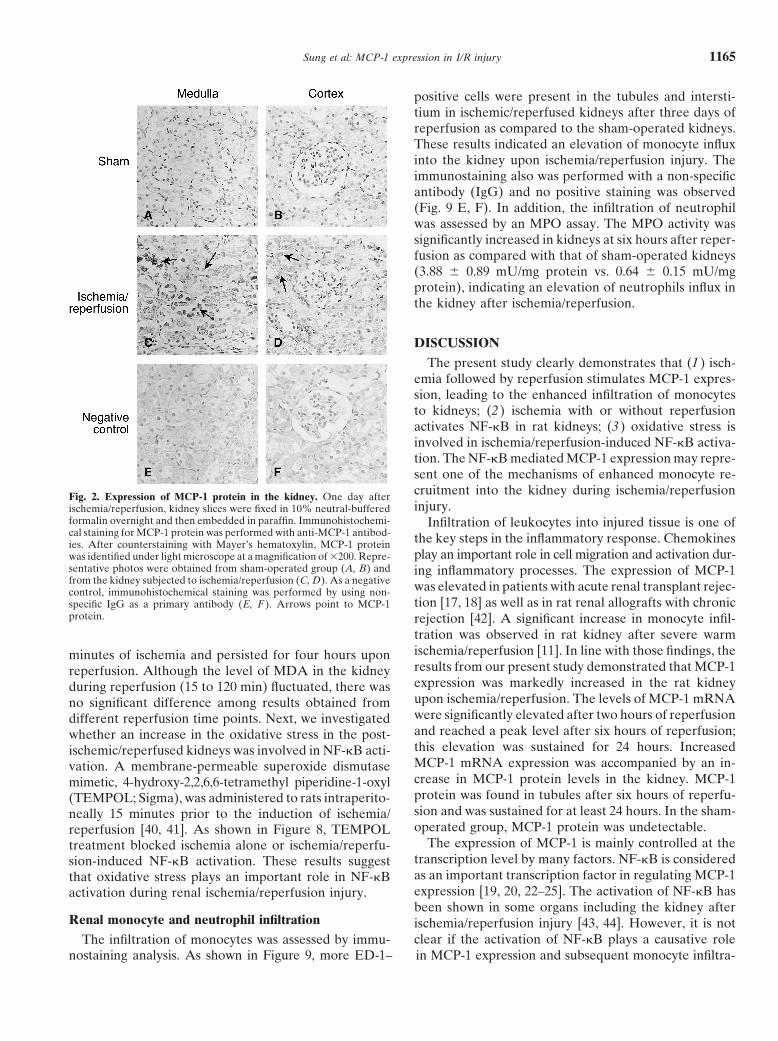
Next, the distribution of MCP-1 protein in the kidneyexternal standard and results were expressed as nmolwas examined by immunohistochemical staining. TheMDA per mg protein.immunostaining for MCP-1 was not evident in the sham-
Myeloperoxidase activity assay operated kidney (Fig. 2 A, B) but was found in tubulesone day after reperfusion (Fig. 2 C, D). A stronger stain-The MPO activity was measured to determine theing for MCP-1 was observed in the medulla. Althoughpresence of neutrophils in the kidney tissue [33, 34].there was mild immunostaining for MCP-1 in tubules ofBriefly, a portion of the kidney was homogenized in 3the cortex, little MCP-1 was detected in glomeruli (Fig.mL 50 mmol/L potassium phosphate buffer (pH 6.0)2D). The immunostaining also was performed with acontaining 0.5% hexadecyltrimethylammoniun bromidenon-specific primary antibody (IgG) and no positive(HTAB). Homogenate was centrifuged at 40,000 � gstaining was observed (Fig. 2 E, F).for 20 minutes and the supernatant was collected. The
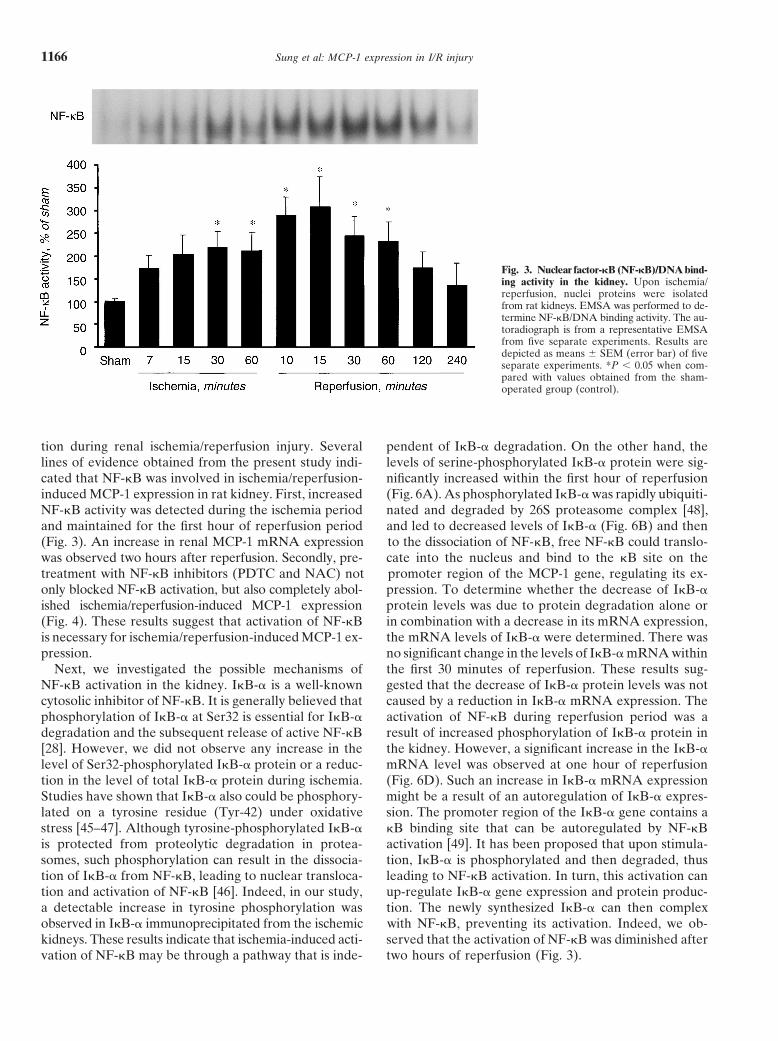
reaction mixture containing 0.6 mmol/L 3,3�,5,5�-tetra-NF-�B activation in the kidneymethylbenzidine (TMB), 0.03 mol/L hydrogen peroxide,
To investigate whether NF-�B was activated in the80 mmol/L sodium phosphate buffer (pH 5.4) and ankidney during ischemia/reperfusion, EMSA was per-aliquot of supernatant was incubated at 25�C for 25 min-formed. As shown in Figure 3, NF-�B/DNA bindingutes. The reaction was stopped by the addition of 2.5activity was significantly elevated in the kidney at 30mL 0.5 mol/L sulfuric acid. The absorbance was read atminutes of ischemia and remained activated through out450 nm. Peroxidase (Sigma Chemical Co., St. Louis, MO,the first hour of reperfusion. The peak activation oc-USA) was used as an external standard. MPO activitycurred at 15 minutes of reperfusion. To further de-was expressed as mU MPO activity per mg protein.termine whether NF-�B activation was necessary forischemia/reperfusion-mediated MCP-1 expression, anStatistical analysisinhibitor of NF-�B activation, pyrrolidine dithiocarba-The results were analyzed using the two-tailed inde-mate (PDTC) (Sigma) was administered to rats intraperi-pendent Student t test. The level of statistical significancetoneally prior to the induction of ischemia/reperfusionwas set at P � 0.05.[35–37]. PDTC treatment significantly blocked the isch-emia/reperfusion-stimulated NF-�B activation (Fig. 4A)
RESULTS as well as abolished the enhanced MCP-1 mRNA expres-sion (Fig. 4B). A similar experiment was performed withMCP-1 expression during ischemia/reperfusionanother known NF-�B inhibitor, N-acetylcysteine (NAC)
The effect of renal ischemia/reperfusion on MCP-1[38, 39]. NAC treatment not only effectively blocked
mRNA expression was examined by a nuclease protec- ischemia/reperfusion-stimulated NF-�B activation (Fig.tion assay. As shown in Figure 1A, there was no signifi- 4A), but also significantly alleviated MCP-1 expressioncant change in MCP-1 mRNA expression in kidneys (Fig. 4B). Both PDTC and NAC have been used assubjected to one or two hours of ischemia. The expres- inhibitors to block NF-�B activation in cultured cellssion of MCP-1 mRNA was significantly elevated (210% and in animal models [35–39].of the control) at two hours after reperfusion. The peakMCP-1 mRNA level was found at six hours after reperfu- Renal I�B-� expression during ischemia/reperfusionsion (270% of the control) and this elevation persisted The activation of NF-�B might be caused by enhancedfor 24 hours (Fig. 1B). There was no significant elevation phosphorylation of the inhibitor protein, I�B-�. There-of MCP-1 mRNA in the kidneys of sham-operated rats fore, the phosphorylation status of I�B-� was examined(used as controls). Similar results were obtained when by Western immunoblotting analysis. During the isch-the levels of MCP-1 mRNA were measured by semi- emia period, there was no change in the levels of phos-quantitative RT-PCR analysis: the peak MCP-1 mRNA pho-I�B-� (Ser32) protein when a specific antibodyexpression was detected at six hours after reperfusion against serine phosphorylated I�B-� was used (Fig. 5A).and the elevation persisted for 24 hours. In addition, However, a time-dependent elevation in the levels ofMCP-1 mRNA also was determined in the contralateral phospho-I�B-� (Try42) protein was observed (15 to 60kidney (right kidney without ischemia/reperfusion). min) in kidneys when the phosphotyrosine-specific anti-There was no elevation of MCP-1 mRNA levels in the body was used (Fig. 5B), indicating that phosphorylationcontralateral kidney six hours after ischemia/reperfusion at the tyrosine residue of I�B-� was increased during
ischemia. The levels of total I�B-� remained unchangedas compared to the sham-operated kidneys (Fig. 1B).
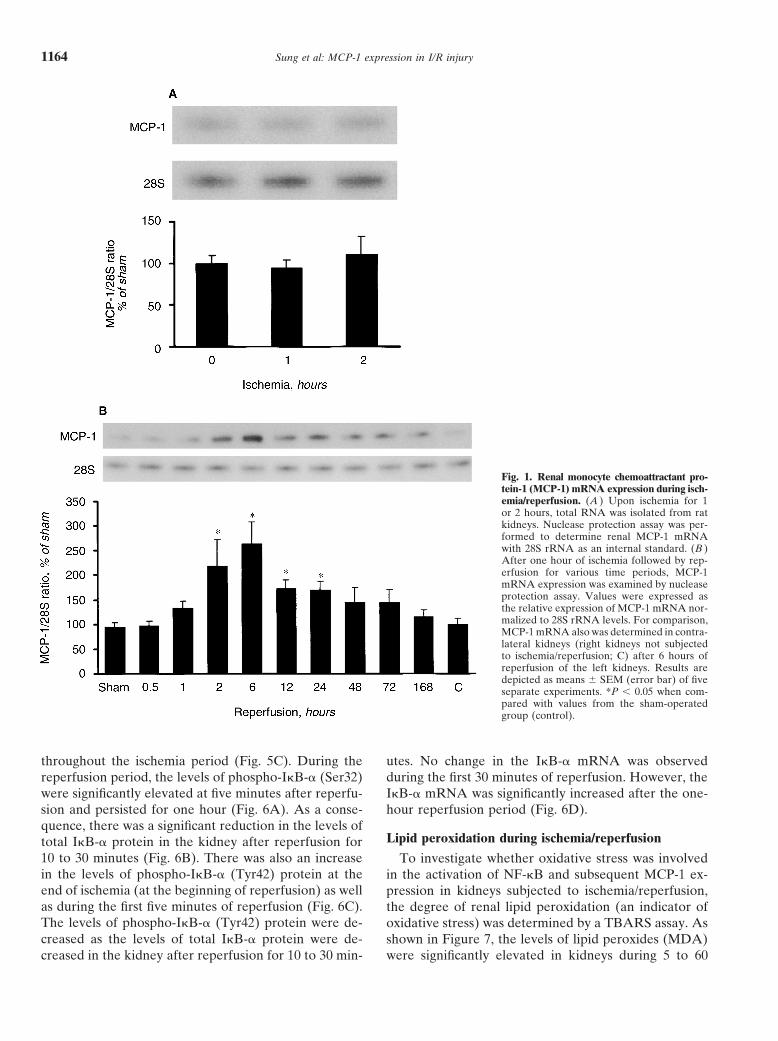
Sung et al: MCP-1 expression in I/R injury1164
Fig. 1. Renal monocyte chemoattractant pro-tein-1 (MCP-1) mRNA expression during isch-emia/reperfusion. (A ) Upon ischemia for 1or 2 hours, total RNA was isolated from ratkidneys. Nuclease protection assay was per-formed to determine renal MCP-1 mRNAwith 28S rRNA as an internal standard. (B )After one hour of ischemia followed by rep-erfusion for various time periods, MCP-1mRNA expression was examined by nucleaseprotection assay. Values were expressed asthe relative expression of MCP-1 mRNA nor-malized to 28S rRNA levels. For comparison,MCP-1 mRNA also was determined in contra-lateral kidneys (right kidneys not subjectedto ischemia/reperfusion; C) after 6 hours ofreperfusion of the left kidneys. Results aredepicted as means SEM (error bar) of fiveseparate experiments. *P � 0.05 when com-pared with values from the sham-operatedgroup (control).
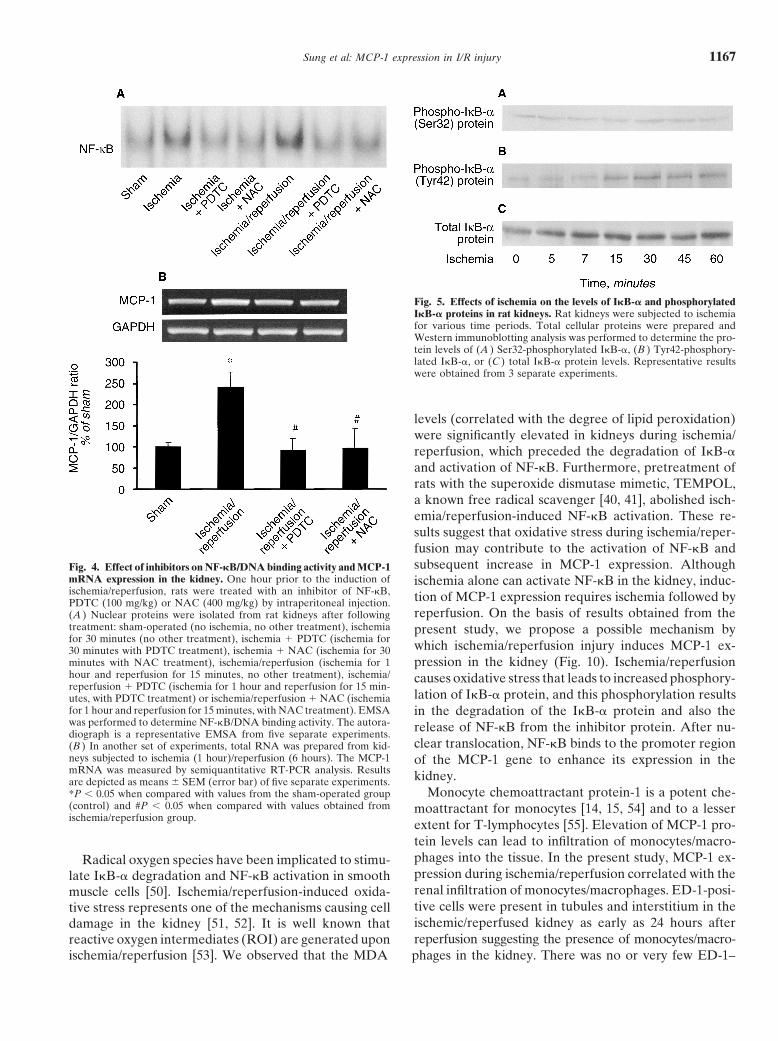
throughout the ischemia period (Fig. 5C). During the utes. No change in the I�B-� mRNA was observedduring the first 30 minutes of reperfusion. However, thereperfusion period, the levels of phospho-I�B-� (Ser32)
were significantly elevated at five minutes after reperfu- I�B-� mRNA was significantly increased after the one-hour reperfusion period (Fig. 6D).sion and persisted for one hour (Fig. 6A). As a conse-
quence, there was a significant reduction in the levels ofLipid peroxidation during ischemia/reperfusiontotal I�B-� protein in the kidney after reperfusion for
10 to 30 minutes (Fig. 6B). There was also an increase To investigate whether oxidative stress was involvedin the levels of phospho-I�B-� (Tyr42) protein at the in the activation of NF-�B and subsequent MCP-1 ex-end of ischemia (at the beginning of reperfusion) as well pression in kidneys subjected to ischemia/reperfusion,as during the first five minutes of reperfusion (Fig. 6C). the degree of renal lipid peroxidation (an indicator ofThe levels of phospho-I�B-� (Tyr42) protein were de- oxidative stress) was determined by a TBARS assay. Ascreased as the levels of total I�B-� protein were de- shown in Figure 7, the levels of lipid peroxides (MDA)
were significantly elevated in kidneys during 5 to 60creased in the kidney after reperfusion for 10 to 30 min-
Sung et al: MCP-1 expression in I/R injury 1165
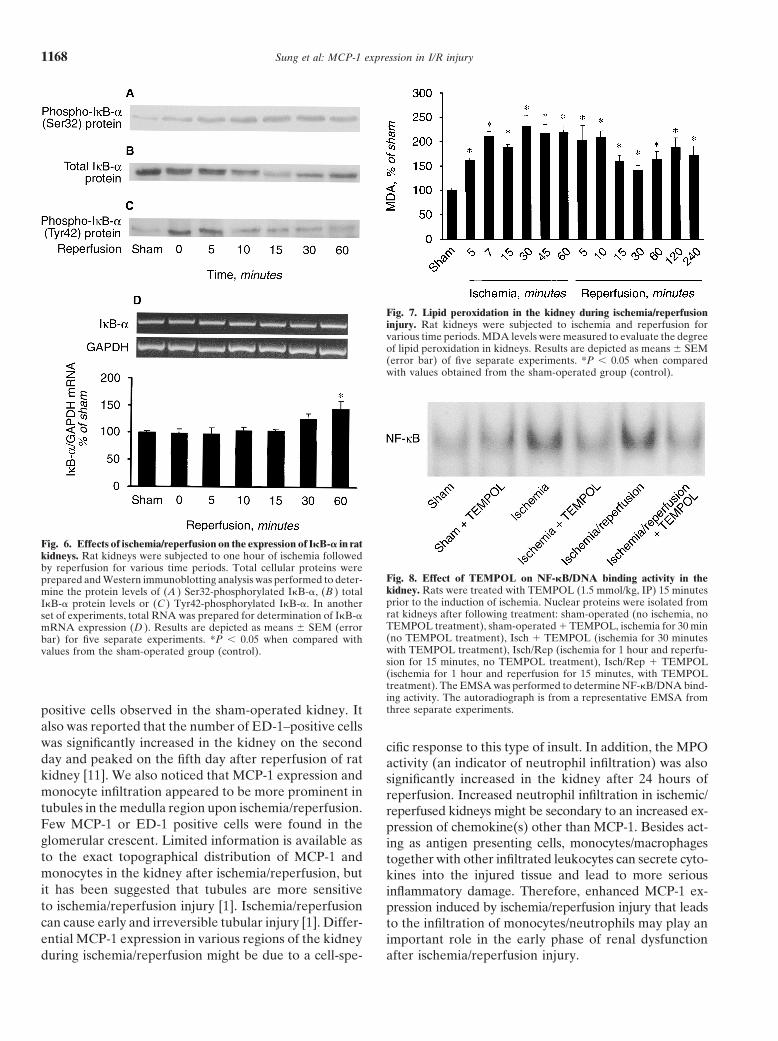
positive cells were present in the tubules and intersti-tium in ischemic/reperfused kidneys after three days ofreperfusion as compared to the sham-operated kidneys.These results indicated an elevation of monocyte influxinto the kidney upon ischemia/reperfusion injury. Theimmunostaining also was performed with a non-specificantibody (IgG) and no positive staining was observed(Fig. 9 E, F). In addition, the infiltration of neutrophilwas assessed by an MPO assay. The MPO activity wassignificantly increased in kidneys at six hours after reper-fusion as compared with that of sham-operated kidneys(3.88 0.89 mU/mg protein vs. 0.64 0.15 mU/mgprotein), indicating an elevation of neutrophils influx inthe kidney after ischemia/reperfusion.
DISCUSSION
The present study clearly demonstrates that (1) isch-emia followed by reperfusion stimulates MCP-1 expres-sion, leading to the enhanced infiltration of monocytesto kidneys; (2) ischemia with or without reperfusionactivates NF-�B in rat kidneys; (3) oxidative stress isinvolved in ischemia/reperfusion-induced NF-�B activa-tion. The NF-�B mediated MCP-1 expression may repre-sent one of the mechanisms of enhanced monocyte re-cruitment into the kidney during ischemia/reperfusionFig. 2. Expression of MCP-1 protein in the kidney. One day afterinjury.ischemia/reperfusion, kidney slices were fixed in 10% neutral-buffered
formalin overnight and then embedded in paraffin. Immunohistochemi- Infiltration of leukocytes into injured tissue is one ofcal staining for MCP-1 protein was performed with anti-MCP-1 antibod- the key steps in the inflammatory response. Chemokinesies. After counterstaining with Mayer’s hematoxylin, MCP-1 protein
play an important role in cell migration and activation dur-was identified under light microscope at a magnification of �200. Repre-sentative photos were obtained from sham-operated group (A, B) and ing inflammatory processes. The expression of MCP-1from the kidney subjected to ischemia/reperfusion (C, D). As a negative was elevated in patients with acute renal transplant rejec-control, immunohistochemical staining was performed by using non-
tion [17, 18] as well as in rat renal allografts with chronicspecific IgG as a primary antibody (E, F ). Arrows point to MCP-1protein. rejection [42]. A significant increase in monocyte infil-
tration was observed in rat kidney after severe warmischemia/reperfusion [11]. In line with those findings, theminutes of ischemia and persisted for four hours uponresults from our present study demonstrated that MCP-1reperfusion. Although the level of MDA in the kidneyexpression was markedly increased in the rat kidneyduring reperfusion (15 to 120 min) fluctuated, there wasupon ischemia/reperfusion. The levels of MCP-1 mRNAno significant difference among results obtained fromwere significantly elevated after two hours of reperfusiondifferent reperfusion time points. Next, we investigatedand reached a peak level after six hours of reperfusion;whether an increase in the oxidative stress in the post-this elevation was sustained for 24 hours. Increasedischemic/reperfused kidneys was involved in NF-�B acti-MCP-1 mRNA expression was accompanied by an in-vation. A membrane-permeable superoxide dismutasecrease in MCP-1 protein levels in the kidney. MCP-1mimetic, 4-hydroxy-2,2,6,6-tetramethyl piperidine-1-oxylprotein was found in tubules after six hours of reperfu-(TEMPOL; Sigma), was administered to rats intraperito-sion and was sustained for at least 24 hours. In the sham-neally 15 minutes prior to the induction of ischemia/operated group, MCP-1 protein was undetectable.reperfusion [40, 41]. As shown in Figure 8, TEMPOL
The expression of MCP-1 is mainly controlled at thetreatment blocked ischemia alone or ischemia/reperfu-transcription level by many factors. NF-�B is consideredsion-induced NF-�B activation. These results suggestas an important transcription factor in regulating MCP-1that oxidative stress plays an important role in NF-�Bexpression [19, 20, 22–25]. The activation of NF-�B hasactivation during renal ischemia/reperfusion injury.been shown in some organs including the kidney after
Renal monocyte and neutrophil infiltration ischemia/reperfusion injury [43, 44]. However, it is notclear if the activation of NF-�B plays a causative roleThe infiltration of monocytes was assessed by immu-
nostaining analysis. As shown in Figure 9, more ED-1– in MCP-1 expression and subsequent monocyte infiltra-
Sung et al: MCP-1 expression in I/R injury1166
Fig. 3. Nuclear factor-�B (NF-�B)/DNA bind-ing activity in the kidney. Upon ischemia/reperfusion, nuclei proteins were isolatedfrom rat kidneys. EMSA was performed to de-termine NF-�B/DNA binding activity. The au-toradiograph is from a representative EMSAfrom five separate experiments. Results aredepicted as means SEM (error bar) of fiveseparate experiments. *P � 0.05 when com-pared with values obtained from the sham-operated group (control).
tion during renal ischemia/reperfusion injury. Several pendent of I�B-� degradation. On the other hand, thelevels of serine-phosphorylated I�B-� protein were sig-lines of evidence obtained from the present study indi-
cated that NF-�B was involved in ischemia/reperfusion- nificantly increased within the first hour of reperfusion(Fig. 6A). As phosphorylated I�B-� was rapidly ubiquiti-induced MCP-1 expression in rat kidney. First, increased
NF-�B activity was detected during the ischemia period nated and degraded by 26S proteasome complex [48],and led to decreased levels of I�B-� (Fig. 6B) and thenand maintained for the first hour of reperfusion period
(Fig. 3). An increase in renal MCP-1 mRNA expression to the dissociation of NF-�B, free NF-�B could translo-cate into the nucleus and bind to the �B site on thewas observed two hours after reperfusion. Secondly, pre-
treatment with NF-�B inhibitors (PDTC and NAC) not promoter region of the MCP-1 gene, regulating its ex-pression. To determine whether the decrease of I�B-�only blocked NF-�B activation, but also completely abol-
ished ischemia/reperfusion-induced MCP-1 expression protein levels was due to protein degradation alone orin combination with a decrease in its mRNA expression,(Fig. 4). These results suggest that activation of NF-�B
is necessary for ischemia/reperfusion-induced MCP-1 ex- the mRNA levels of I�B-� were determined. There wasno significant change in the levels of I�B-� mRNA withinpression.
Next, we investigated the possible mechanisms of the first 30 minutes of reperfusion. These results sug-gested that the decrease of I�B-� protein levels was notNF-�B activation in the kidney. I�B-� is a well-known
cytosolic inhibitor of NF-�B. It is generally believed that caused by a reduction in I�B-� mRNA expression. Theactivation of NF-�B during reperfusion period was aphosphorylation of I�B-� at Ser32 is essential for I�B-�
degradation and the subsequent release of active NF-�B result of increased phosphorylation of I�B-� protein inthe kidney. However, a significant increase in the I�B-�[28]. However, we did not observe any increase in the
level of Ser32-phosphorylated I�B-� protein or a reduc- mRNA level was observed at one hour of reperfusion(Fig. 6D). Such an increase in I�B-� mRNA expressiontion in the level of total I�B-� protein during ischemia.
Studies have shown that I�B-� also could be phosphory- might be a result of an autoregulation of I�B-� expres-sion. The promoter region of the I�B-� gene contains alated on a tyrosine residue (Tyr-42) under oxidative
stress [45–47]. Although tyrosine-phosphorylated I�B-� �B binding site that can be autoregulated by NF-�Bactivation [49]. It has been proposed that upon stimula-is protected from proteolytic degradation in protea-
somes, such phosphorylation can result in the dissocia- tion, I�B-� is phosphorylated and then degraded, thusleading to NF-�B activation. In turn, this activation cantion of I�B-� from NF-�B, leading to nuclear transloca-
tion and activation of NF-�B [46]. Indeed, in our study, up-regulate I�B-� gene expression and protein produc-tion. The newly synthesized I�B-� can then complexa detectable increase in tyrosine phosphorylation was
observed in I�B-� immunoprecipitated from the ischemic with NF-�B, preventing its activation. Indeed, we ob-served that the activation of NF-�B was diminished afterkidneys. These results indicate that ischemia-induced acti-
vation of NF-�B may be through a pathway that is inde- two hours of reperfusion (Fig. 3).
Sung et al: MCP-1 expression in I/R injury 1167
Fig. 5. Effects of ischemia on the levels of I�B-� and phosphorylatedI�B-� proteins in rat kidneys. Rat kidneys were subjected to ischemiafor various time periods. Total cellular proteins were prepared andWestern immunoblotting analysis was performed to determine the pro-tein levels of (A ) Ser32-phosphorylated I�B-�, (B ) Tyr42-phosphory-lated I�B-�, or (C ) total I�B-� protein levels. Representative resultswere obtained from 3 separate experiments.
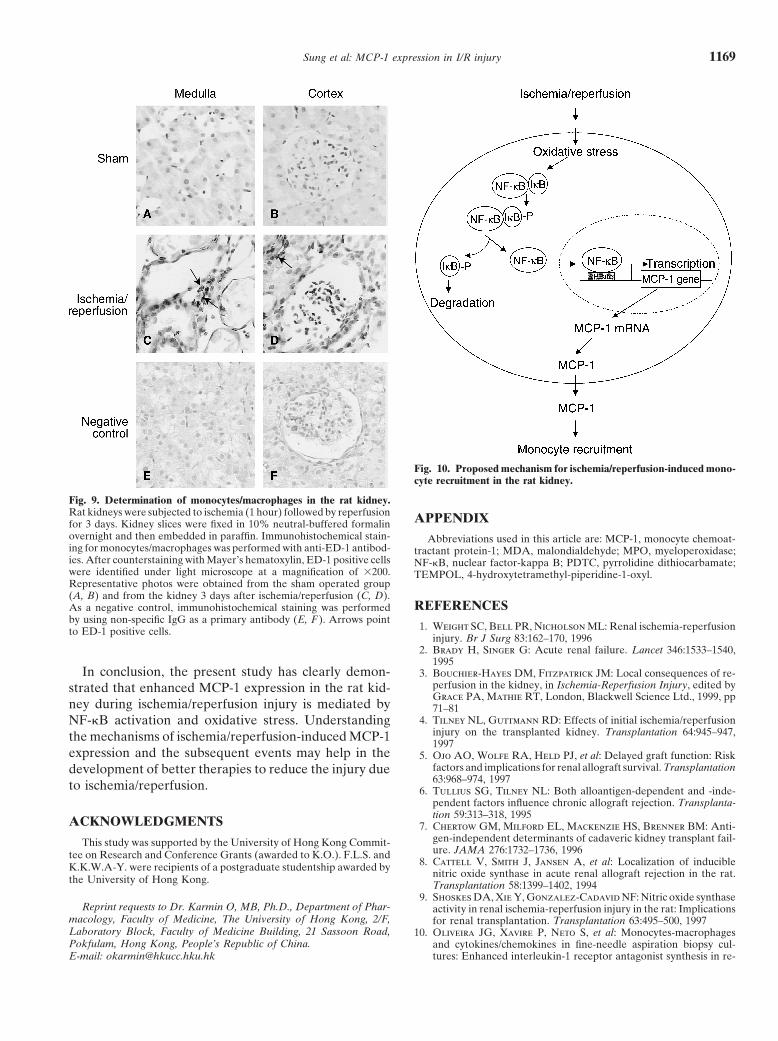
levels (correlated with the degree of lipid peroxidation)were significantly elevated in kidneys during ischemia/reperfusion, which preceded the degradation of I�B-�and activation of NF-�B. Furthermore, pretreatment ofrats with the superoxide dismutase mimetic, TEMPOL,a known free radical scavenger [40, 41], abolished isch-emia/reperfusion-induced NF-�B activation. These re-sults suggest that oxidative stress during ischemia/reper-fusion may contribute to the activation of NF-�B andsubsequent increase in MCP-1 expression. AlthoughFig. 4. Effect of inhibitors on NF-�B/DNA binding activity and MCP-1
mRNA expression in the kidney. One hour prior to the induction of ischemia alone can activate NF-�B in the kidney, induc-ischemia/reperfusion, rats were treated with an inhibitor of NF-�B, tion of MCP-1 expression requires ischemia followed byPDTC (100 mg/kg) or NAC (400 mg/kg) by intraperitoneal injection.
reperfusion. On the basis of results obtained from the(A ) Nuclear proteins were isolated from rat kidneys after followingtreatment: sham-operated (no ischemia, no other treatment), ischemia present study, we propose a possible mechanism byfor 30 minutes (no other treatment), ischemia PDTC (ischemia for which ischemia/reperfusion injury induces MCP-1 ex-30 minutes with PDTC treatment), ischemia NAC (ischemia for 30minutes with NAC treatment), ischemia/reperfusion (ischemia for 1 pression in the kidney (Fig. 10). Ischemia/reperfusionhour and reperfusion for 15 minutes, no other treatment), ischemia/ causes oxidative stress that leads to increased phosphory-reperfusion PDTC (ischemia for 1 hour and reperfusion for 15 min-
lation of I�B-� protein, and this phosphorylation resultsutes, with PDTC treatment) or ischemia/reperfusion NAC (ischemiafor 1 hour and reperfusion for 15 minutes, with NAC treatment). EMSA in the degradation of the I�B-� protein and also thewas performed to determine NF-�B/DNA binding activity. The autora- release of NF-�B from the inhibitor protein. After nu-diograph is a representative EMSA from five separate experiments.
clear translocation, NF-�B binds to the promoter region(B) In another set of experiments, total RNA was prepared from kid-neys subjected to ischemia (1 hour)/reperfusion (6 hours). The MCP-1 of the MCP-1 gene to enhance its expression in themRNA was measured by semiquantitative RT-PCR analysis. Results kidney.are depicted as means SEM (error bar) of five separate experiments.
Monocyte chemoattractant protein-1 is a potent che-*P � 0.05 when compared with values from the sham-operated group(control) and #P � 0.05 when compared with values obtained from moattractant for monocytes [14, 15, 54] and to a lesserischemia/reperfusion group.
extent for T-lymphocytes [55]. Elevation of MCP-1 pro-tein levels can lead to infiltration of monocytes/macro-phages into the tissue. In the present study, MCP-1 ex-Radical oxygen species have been implicated to stimu-pression during ischemia/reperfusion correlated with thelate I�B-� degradation and NF-�B activation in smoothrenal infiltration of monocytes/macrophages. ED-1-posi-muscle cells [50]. Ischemia/reperfusion-induced oxida-tive cells were present in tubules and interstitium in thetive stress represents one of the mechanisms causing cellischemic/reperfused kidney as early as 24 hours afterdamage in the kidney [51, 52]. It is well known thatreperfusion suggesting the presence of monocytes/macro-reactive oxygen intermediates (ROI) are generated upon
ischemia/reperfusion [53]. We observed that the MDA phages in the kidney. There was no or very few ED-1–
Sung et al: MCP-1 expression in I/R injury1168
Fig. 7. Lipid peroxidation in the kidney during ischemia/reperfusioninjury. Rat kidneys were subjected to ischemia and reperfusion forvarious time periods. MDA levels were measured to evaluate the degreeof lipid peroxidation in kidneys. Results are depicted as means SEM(error bar) of five separate experiments. *P � 0.05 when comparedwith values obtained from the sham-operated group (control).
Fig. 6. Effects of ischemia/reperfusion on the expression of I�B-� in ratkidneys. Rat kidneys were subjected to one hour of ischemia followedby reperfusion for various time periods. Total cellular proteins were
Fig. 8. Effect of TEMPOL on NF-�B/DNA binding activity in theprepared and Western immunoblotting analysis was performed to deter-kidney. Rats were treated with TEMPOL (1.5 mmol/kg, IP) 15 minutesmine the protein levels of (A ) Ser32-phosphorylated I�B-�, (B ) totalprior to the induction of ischemia. Nuclear proteins were isolated fromI�B-� protein levels or (C ) Tyr42-phosphorylated I�B-�. In anotherrat kidneys after following treatment: sham-operated (no ischemia, noset of experiments, total RNA was prepared for determination of I�B-�TEMPOL treatment), sham-operated TEMPOL, ischemia for 30 minmRNA expression (D ). Results are depicted as means SEM (error(no TEMPOL treatment), Isch TEMPOL (ischemia for 30 minutesbar) for five separate experiments. *P � 0.05 when compared withwith TEMPOL treatment), Isch/Rep (ischemia for 1 hour and reperfu-values from the sham-operated group (control).sion for 15 minutes, no TEMPOL treatment), Isch/Rep TEMPOL(ischemia for 1 hour and reperfusion for 15 minutes, with TEMPOLtreatment). The EMSA was performed to determine NF-�B/DNA bind-ing activity. The autoradiograph is from a representative EMSA fromthree separate experiments.positive cells observed in the sham-operated kidney. It
also was reported that the number of ED-1–positive cellswas significantly increased in the kidney on the second cific response to this type of insult. In addition, the MPOday and peaked on the fifth day after reperfusion of rat activity (an indicator of neutrophil infiltration) was alsokidney [11]. We also noticed that MCP-1 expression and significantly increased in the kidney after 24 hours ofmonocyte infiltration appeared to be more prominent in reperfusion. Increased neutrophil infiltration in ischemic/tubules in the medulla region upon ischemia/reperfusion. reperfused kidneys might be secondary to an increased ex-Few MCP-1 or ED-1 positive cells were found in the pression of chemokine(s) other than MCP-1. Besides act-glomerular crescent. Limited information is available as ing as antigen presenting cells, monocytes/macrophagesto the exact topographical distribution of MCP-1 and together with other infiltrated leukocytes can secrete cyto-monocytes in the kidney after ischemia/reperfusion, but kines into the injured tissue and lead to more seriousit has been suggested that tubules are more sensitive inflammatory damage. Therefore, enhanced MCP-1 ex-to ischemia/reperfusion injury [1]. Ischemia/reperfusion pression induced by ischemia/reperfusion injury that leadscan cause early and irreversible tubular injury [1]. Differ- to the infiltration of monocytes/neutrophils may play anential MCP-1 expression in various regions of the kidney important role in the early phase of renal dysfunction
after ischemia/reperfusion injury.during ischemia/reperfusion might be due to a cell-spe-
Sung et al: MCP-1 expression in I/R injury 1169
Fig. 10. Proposed mechanism for ischemia/reperfusion-induced mono-cyte recruitment in the rat kidney.
Fig. 9. Determination of monocytes/macrophages in the rat kidney.Rat kidneys were subjected to ischemia (1 hour) followed by reperfusion APPENDIXfor 3 days. Kidney slices were fixed in 10% neutral-buffered formalinovernight and then embedded in paraffin. Immunohistochemical stain- Abbreviations used in this article are: MCP-1, monocyte chemoat-ing for monocytes/macrophages was performed with anti-ED-1 antibod- tractant protein-1; MDA, malondialdehyde; MPO, myeloperoxidase;ies. After counterstaining with Mayer’s hematoxylin, ED-1 positive cells NF-�B, nuclear factor-kappa B; PDTC, pyrrolidine dithiocarbamate;were identified under light microscope at a magnification of �200. TEMPOL, 4-hydroxytetramethyl-piperidine-1-oxyl.Representative photos were obtained from the sham operated group(A, B) and from the kidney 3 days after ischemia/reperfusion (C, D).
REFERENCESAs a negative control, immunohistochemical staining was performedby using non-specific IgG as a primary antibody (E, F ). Arrows point
1. Weight SC, Bell PR, Nicholson ML: Renal ischemia-reperfusionto ED-1 positive cells.injury. Br J Surg 83:162–170, 1996
2. Brady H, Singer G: Acute renal failure. Lancet 346:1533–1540,1995
In conclusion, the present study has clearly demon- 3. Bouchier-Hayes DM, Fitzpatrick JM: Local consequences of re-perfusion in the kidney, in Ischemia-Reperfusion Injury, edited bystrated that enhanced MCP-1 expression in the rat kid-Grace PA, Mathie RT, London, Blackwell Science Ltd., 1999, pp
ney during ischemia/reperfusion injury is mediated by 71–814. Tilney NL, Guttmann RD: Effects of initial ischemia/reperfusionNF-�B activation and oxidative stress. Understanding
injury on the transplanted kidney. Transplantation 64:945–947,the mechanisms of ischemia/reperfusion-induced MCP-11997
expression and the subsequent events may help in the 5. Ojo AO, Wolfe RA, Held PJ, et al: Delayed graft function: Riskfactors and implications for renal allograft survival. Transplantationdevelopment of better therapies to reduce the injury due63:968–974, 1997to ischemia/reperfusion. 6. Tullius SG, Tilney NL: Both alloantigen-dependent and -inde-pendent factors influence chronic allograft rejection. Transplanta-tion 59:313–318, 1995ACKNOWLEDGMENTS 7. Chertow GM, Milford EL, Mackenzie HS, Brenner BM: Anti-gen-independent determinants of cadaveric kidney transplant fail-This study was supported by the University of Hong Kong Commit-ure. JAMA 276:1732–1736, 1996tee on Research and Conference Grants (awarded to K.O.). F.L.S. and
8. Cattell V, Smith J, Jansen A, et al: Localization of inducibleK.K.W.A-Y. were recipients of a postgraduate studentship awarded bynitric oxide synthase in acute renal allograft rejection in the rat.the University of Hong Kong. Transplantation 58:1399–1402, 1994
9. Shoskes DA, Xie Y, Gonzalez-Cadavid NF: Nitric oxide synthaseReprint requests to Dr. Karmin O, MB, Ph.D., Department of Phar- activity in renal ischemia-reperfusion injury in the rat: Implications
macology, Faculty of Medicine, The University of Hong Kong, 2/F, for renal transplantation. Transplantation 63:495–500, 1997Laboratory Block, Faculty of Medicine Building, 21 Sassoon Road, 10. Oliveira JG, Xavire P, Neto S, et al: Monocytes-macrophagesPokfulam, Hong Kong, People’s Republic of China. and cytokines/chemokines in fine-needle aspiration biopsy cul-
tures: Enhanced interleukin-1 receptor antagonist synthesis in re-E-mail: [email protected]

Sung et al: MCP-1 expression in I/R injury1170
jection-free kidney transplant patients. Transplantation 63:1751– with increased oxygen free radical activity and altered nitric oxide1756, 1997 metabolism in chronic experimental hemosiderosis. Lab Invest
11. Ysebaert DK, De Greef KE, Vercauteren SR, et al: Identification 80:1905–1914, 2000and kinetics of leukocytes after severe ischemia/reperfusion renal 33. Suzuki K, Ota H, Sasagawa S, et al: Assay method for myeloper-injury. Nephrol Dial Transplant 15:1562–1574, 2000 oxidase in human polymorphonuclear leukocytes. Anal Biochem
12. Shoskes DA, Parfrey NA, Halloran PF: Increased major histo- 132:345–352, 1983compatibility complex antigen expression in unilateral ischemic 34. Chatterjee PK, Cuzzocrea S, Brown PAJ, et al: Tempol, a mem-acute necrosis in the mouse. Transplantation 49:201–207, 1990 brane-permeable radical scavenger, reduces oxidant stress-medi-
13. Dargun D, Hoff U, Park JK, et al: Ischemia-reperfusion injury ated renal dysfunction and injury in the rat. Kidney Int 58:658–673,in renal transplantation is independent of the immunologic back- 2000ground. Kidney Int 58:2166–2177, 2000 35. Schreck R, Baeuerle PA: Assessing oxygen radicals as mediators
14. Rollins BJ, Yoshimura T, Leonard EJ, Pober JS: Cytokine- in activation of inducible eukaryotic transcription factor NF-�B.activated human endothelial cells synthesize and secret a monocyte Methods Enzymol 234:151–163, 1994chemoattractant, MCP-1/JE. Am J Pathol 136:1229–1233, 1990 36. Satoh A, Shimosegawa T, Fujita M, et al: Inhibition of nuclear
15. Valente AJ, Rozek MM, Sprague EA, Schwartz CJ: Mecha- factor-�B activation improves the survival of rats with taurocholatenisms in intimal monocyte-macrophage recruitment. A special role pancreatitis. Gut 44:253–258, 1999for monocyte chemotactic protein-1. Circulation 86:III20–III25, 37. Liu SF, Ye X, Malik AB: Inhibition of NF-�B activation by pyrroli-1992 dine dithiocarbamate prevents in vivo expression of proinflamma-
16. Robertson H, Morley AR, Talbot D, et al: Renal allograft rejec- tory genes. Circulation 100:1330–1337, 1999tion: Beta-chemokine involvement in the development of tubulitis. 38. Schreck R, Baeuerle PA: Assessing oxygen radicals as mediatorsTransplantation 69:684–687, 2000 in activation of inducible eukaryotic transcription factor NF-�B.
17. Grandaliano G, Gesualdo L, Ranieri E, et al: Monocyte chemo- Method Enzymol 234:151–163, 1994tactic peptide-1 expression and monocyte infiltration in acute renal 39. DiMari J, Megyesi J, Udvarhelyi N, et al: N-acetyl cysteine ame-transplant rejection. Transplantation 63:414–420, 1997 liorates ischemic renal failure. Am J Physiol 272:F292–F298, 1997
18. Prodjosudjadi W, Daha MR, Gerritsma JS, et al: Increased uri- 40. Schnackenberg CG, Welch WJ, Wilcox CS: Normalization ofnary excretion of monocyte chemoattractant protein-1 during acute blood pressure and renal vascular resistance in SHR with a mem-renal allograft rejection. Nephrol Dial Transplant 11:1096–1103, brane-permeable superoxide dismutase mimetic. Role of nitric ox-1996 ide. Hypertens 32:59–64, 1998
19. Lynn EG, Siow YL: O K: Very low-density lipoprotein stimulates 41. Zou A-P, Li N, Cowley AW Jr: Production and actions of superox-the expression of monocyte chemoattractant protein-1 in mesangial ide in the renal medulla. Hypertension 37:547–553, 2001cells. Kidney Int 57:1472–1483, 2000 42. Nadeau KC, Azuma H, Tilney NL: Sequential cytokine dynamics
20. Lynn EG, Siow YL, Frohlich J, et al: Lipoprotein-X Stimulates in chronic rejection of rat renal allografts: Role for cytokinesmonocyte chemoattractant protein-1 expression in mesangial cells RANTES and MCP-1. Proc Natl Acad Sci USA 92:8729–8733,via nuclear factor-�B. Kidney Int 60:520–532, 2001 1995
21. Gawaz M, Neumann FJ, Dickfeld T, et al: Activated platelets 43. Li C, Browder W, Kao RL: Early activation of transcription factorinduce monocyte chemotactic protein-1 secretion and surface ex- NF-�B during ischemia in perfused rat heart. Am J Physiol (Heartpression of intercellular adhesion molecule-1 on endothelial cells. Circ Physiol) 276:H543–H522, 1999Circulation 98:1164–1171, 1998 44. Donnahoo KK, Meldrum DR, Shenkar R, et al: Early renal
22. Tomita N, Morishita R, Tomita S, et al: Inhibition of TNF-alpha, ischemia, with or with without reperfusion, activates NF�B andinduced cytokine and adhesion molecule. Expression in glomerular increases TNF-� bioactivity in the kidney. J Urol 163:1328–1332,cells in vitro and in vivo by transcription factor decoy for NF 2000kappaB. Exp Nephrol 9:181–190, 2001 45. Imbert V, Rupec RA, Livolsi A, et al: Tyrosine phosphorylation23. Stylianou E, Nie M, Ueda A, Zhao L: c-Rel and p65 trans- of I�B� activates NF-�B without proteolytic degradation of I�B�.activate the monocyte chemoattractant protein-1 gene in interleu- Cell 86:787–798, 1996kin-1 stimulated mesangial cells. Kidney Int 56:873–882, 1999 46. Canty TG Jr, Boyle EM Jr, Farr A, et al: Oxidative stress induces24. Li D, Saldeen T, Romeo F, Mehta JL: Oxidized LDL upregulates NF-kappa B nuclear translocation without degradation of Ikappaangiotensin II type 1 receptor expression in cultured human coro- B alpha. Circulation 100(19 Suppl):II361–II364, 1999nary artery endothelial cells: The potential role of transcription 47. Singh S, Darnay BG, Aggarwal BB: Site-specific tyrosine phos-factor NF-kappa B. Circulation 102:1970–1976, 2000
phorylation of I�B� negatively regulates its inducible phosphoryla-25. Wang G, Siow YL: O K: Homocysteine induces monocyte chemo-tion and degradation. J Biol Chem 271:31049–31054, 1996attractant protein-1 expression by activating NF-kappa B in THP-1
48. May MJ, Ghosh S: Signal transduction through NF-�B. Immunolmacrophages. Am J Physiol (Heart Circ Physiol) 280:H2840–Today 19:80–88, 1998H2847, 2001
49. Chiao PJ, Miyamoto S, Verma IM: Autoregulation of I�B� activ-26. Vos IH, Govers R, Grone HJ, et al: NFkappaB decoy oligodeoxy-ity. Proc Natl Acad Sci USA 91:28–32, 1994nucleotides reduce monocyte infiltration in renal allografts. FA-
50. Wang G, Siow YL: O K: Homocysteine stimulates nuclear factorSEB J 14:815–822, 2000�B activity and monocyte chemoattractant protein-1 expression in27. Sung FL, Siow YL, Wang G, et al: Homocysteine stimulates thevascular smooth-muscle cells: a possible role for protein kinase C.expression of monocyte chemoattractant protein-1 in endothelialBiochem J 352:817–826, 2000cells leading to monocyte chemotaxis. Mol Cell Biochem 216:121–
51. Kelly KJ, Bonventre JV: Ischemia/reperfusion injury in trans-128, 2001plantation (chapt 26), in Transplantation Biology: Cellular and28. Simeonidis S, Stauber D, Chen G, et al: Mechanisms by whichMolecular Aspects, edited by Tilney NL, Strom TB, Paul LC,I�B proteins control NF-�B activity. Proc Natl Acad Sci USAPhiladelphia, Lippincott-Raven Publishers, 1996, pp 257–27496:49–54, 1999
52. Torras J, Seron D, Herrero I, et al: Renal protective effect of29. Landry DB, Couper LL, Bryant SR, Linder V: Activation of theliposome superoxide dismutase in an experimental warm ischemiaNF-�B and I�B system in smooth muscle cells after rat arterialmodel. Transplant Int 7(Suppl 1):S472–S475, 1994injury. Induction of vascular cell adhesion molecule-1 and mono-
53. Parra T, de Arriba G, Conejo JR, et al: Cyclosporine increasescyte chemoattractant protein-1. Am J Pathol 151:1085–1095, 1997local glomerular synthesis of reactive oxygen species in rats: Effect30. Galea E, Glickstein SB, Feinstein DL, et al: Stimulation of cere-of vitamin E on cyclosporine nephrotoxicity. Transplantationbellar fastigial nucleus inhibits interleukin-1�-induced cerebrovas-66:1325–1329, 1998cular inflammation. Am J Physiol (Heart Circ Physiol) 275:H2053–
54. Leonard EJ, Yoshimura T: Human monocyte chemoattractantH2063, 1998protein-1 (MCP-1). Immunol Today 11:97–101, 199031. Ohkawa H, Ohishi N, Yagi K: Assay for lipid peroxides in animal
55. Carr MW, Roth SJ, Luther E, et al: Monocyte chemoattractanttissues by thiobarbituric acid reaction. Anal Biochem 95:351–358,protein 1 acts as a T-lymphocyte chemoattractant. Proc Natl Acad1979
32. Zhou XJ, Laszik Z, Wang XQ, et al: Association of renal injury Sci USA 91:3652–3656, 1994
Related Documents











