JOURNAL OF THE MECHANICAL BEHAVIOR OF BIOMEDICAL MATERIALS 4 (2011) 1196–1205 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/jmbbm Research paper Engineering alginate for intervertebral disc repair Johannes L. Bron a,b,∗ , Lucienne A. Vonk b,c , Theodoor H. Smit a,b , Gijsje H. Koenderink d a Department of Orthopaedic Surgery, VU University Medical Center, Amsterdam, The Netherlands b Skeletal Tissue Engineering Group Amsterdam (STEGA) and Research Institute MOVE, The Netherlands c Department of Oral Cell Biology, Academic Centre for Dentistry Amsterdam (ACTA), University of Amsterdam and VU University Amsterdam, Amsterdam, The Netherlands d Biological Soft Matter group, FOM Institute for Atomic and Molecular Physics (AMOLF), Amsterdam, The Netherlands ARTICLE INFO Article history: Received 14 January 2011 Received in revised form 2 April 2011 Accepted 4 April 2011 Published online 24 April 2011 Keywords: Alginate Intervertebral disc Tissue engineering Gene expression Rheology ABSTRACT Alginate is frequently studied as a scaffold for intervertebral disc (IVD) repair, since it closely mimics mechanical and cell-adhesive properties of the nucleus pulposus (NP) of the IVD. The aim of this study was to assess the relation between alginate concentration and scaffold stiffness and find preparation conditions where the viscoelastic behaviour mimics that of the NP. In addition, we measured the effect of variations in scaffold stiffness on the expression of extracellular matrix molecules specific to the NP (proteoglycans and collagen) by native NP cells. We prepared sample discs of different concentrations of alginate (1%–6%) by two different methods, diffusion and in situ gelation. The stiffness increased with increasing alginate concentration, while the loss tangent (dissipative behaviour) remained constant. The diffusion samples were ten-fold stiffer than samples prepared by in situ gelation. Sample discs prepared from 2% alginate by diffusion closely matched the stiffness and loss tangent of the NP. The stiffness of all samples declined upon prolonged incubation in medium, especially for samples prepared by diffusion. The biosynthetic phenotype of native cells isolated from NPs was preserved in alginate matrices up to 4 weeks of culturing. Gene expression levels of extracellular matrix components were insensitive to alginate concentration and corresponding matrix stiffness, likely due to the poor adhesiveness of the cells to alginate. In conclusion, alginate can mimic the viscoelastic properties of the NP and preserve the biosynthetic phenotype of NP cells but certain limitations like long-term stability still have to be addressed. c ⃝ 2011 Elsevier Ltd. All rights reserved. 1. Introduction Transplantation systems based on scaffolds seeded with stem cells or native cells offer a promising means to repair aged, damaged, or diseased tissues (Hubbell, 2003). Accordingly, there has been much recent effort to design scaffolds that mimic the bioadhesive and physical characteristics ∗ Corresponding address: Department of Orthopaedic Surgery, VU University Medical Center, PO Box 7057, 1007 MB Amsterdam, The Netherlands. Tel.: +31 20 44 45242. E-mail addresses: [email protected], [email protected] (J.L. Bron). of natural extracellular matrices found in tissues and can thus promote tissue-specific cell phenotype (Huebsch and Mooney, 2009; Lutolf et al., 2009). A variety of tissues can already be engineered by this approach, including artery, skin, cartilage, bone, ligament, and tendon. Scaffold stiffness has been recognized as an especially important cue to 1751-6161/$ - see front matter c ⃝ 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.jmbbm.2011.04.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5
available at www.sciencedirect.com
journal homepage: www.elsevier.com/locate/jmbbm
Research paper
Engineering alginate for intervertebral disc repair
Johannes L. Brona,b,∗, Lucienne A. Vonkb,c, Theodoor H. Smita,b, Gijsje H. Koenderinkd
aDepartment of Orthopaedic Surgery, VU University Medical Center, Amsterdam, The Netherlandsb Skeletal Tissue Engineering Group Amsterdam (STEGA) and Research Institute MOVE, The NetherlandscDepartment of Oral Cell Biology, Academic Centre for Dentistry Amsterdam (ACTA), University of Amsterdam and VU UniversityAmsterdam, Amsterdam, The Netherlandsd Biological Soft Matter group, FOM Institute for Atomic and Molecular Physics (AMOLF), Amsterdam, The Netherlands
A R T I C L E I N F O
Article history:
Received 14 January 2011
Received in revised form
2 April 2011
Accepted 4 April 2011
Published online 24 April 2011
Keywords:
Alginate
Intervertebral disc
Tissue engineering
Gene expression
Rheology
A B S T R A C T
Alginate is frequently studied as a scaffold for intervertebral disc (IVD) repair, since it
closely mimics mechanical and cell-adhesive properties of the nucleus pulposus (NP) of the
IVD. The aim of this study was to assess the relation between alginate concentration and
scaffold stiffness and find preparation conditions where the viscoelastic behaviour mimics
that of the NP. In addition, we measured the effect of variations in scaffold stiffness on the
expression of extracellular matrix molecules specific to the NP (proteoglycans and collagen)
by native NP cells. We prepared sample discs of different concentrations of alginate (1%–6%)
by two different methods, diffusion and in situ gelation. The stiffness increased with
increasing alginate concentration, while the loss tangent (dissipative behaviour) remained
constant. The diffusion samples were ten-fold stiffer than samples prepared by in situ
gelation. Sample discs prepared from 2% alginate by diffusion closely matched the stiffness
and loss tangent of the NP. The stiffness of all samples declined upon prolonged incubation
in medium, especially for samples prepared by diffusion. The biosynthetic phenotype of
native cells isolated fromNPs was preserved in alginate matrices up to 4 weeks of culturing.
Gene expression levels of extracellular matrix components were insensitive to alginate
concentration and corresponding matrix stiffness, likely due to the poor adhesiveness of
the cells to alginate. In conclusion, alginate can mimic the viscoelastic properties of the NP
and preserve the biosynthetic phenotype of NP cells but certain limitations like long-term
stability still have to be addressed.c⃝ 2011 Elsevier Ltd. All rights reserved.
U
d
1. Introduction
Transplantation systems based on scaffolds seeded with stemcells or native cells offer a promising means to repair aged,damaged, or diseased tissues (Hubbell, 2003). Accordingly,there has been much recent effort to design scaffoldsthat mimic the bioadhesive and physical characteristics
∗ Corresponding address: Department of Orthopaedic Surgery, VUNetherlands. Tel.: +31 20 44 45242.
E-mail addresses: [email protected], [email protected] (J.L. Bro
1751-6161/$ - see front matter c⃝ 2011 Elsevier Ltd. All rights reservedoi:10.1016/j.jmbbm.2011.04.002
niversity Medical Center, PO Box 7057, 1007 MB Amsterdam, The
n).
of natural extracellular matrices found in tissues and canthus promote tissue-specific cell phenotype (Huebsch andMooney, 2009; Lutolf et al., 2009). A variety of tissues canalready be engineered by this approach, including artery,skin, cartilage, bone, ligament, and tendon. Scaffold stiffnesshas been recognized as an especially important cue to
.

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5 1197
guide cell differentiation and extracellular matrix (ECM)production (Breuls et al., 2008; Discher et al., 2005; Engleret al., 2007) and this knowledge is now increasingly beingimplemented in tissue engineering strategies (Discher et al.,2009). The mechanical characteristics of many tissues havebeen documented over the recent years, facilitating thedevelopment of new generations of 3D scaffolds mimickingthese features (Breuls et al., 2008; Iatridis et al., 1996).
Our own research over the past years has focused ontissue engineering strategies to repair damaged intervertebraldiscs (IVDs) (Bron et al., 2009b,a, 2010). The IVD is acartilaginous structure that lies between adjacent vertebrae,where it acts as a shock absorber and allows motionof the otherwise rigid vertebral column (Roberts et al.,2006). The IVD consists of a collagenous outer annulusfibrosus (AF) which surrounds the gelatinous inner nucleuspulposus (NP). Ageing is accompanied by loss of waterand proteoglycans from the gelatinous NP, which becomesmore fibrous, resulting in a more rigid IVD. Although thesechanges are to some extent physiological, they may result insymptomatic degenerative disc disease (Roberts et al., 2006).In some patients, early degeneration of the AF may resultin a posterior tear through which the NP can extrude (discherniation), compromising the neurological structures (spineand nerve roots) that the vertebral column usually protects.The current clinical solution is to evacuate the herniatedNP material (discectomy), thereby relieving the compressednerves (Hegewald et al., 2008). There are, however, seriousadverse effects of disc herniation and subsequent discectomyon spinal biomechanics resulting in discogenic back painthat seriously affects the quality of life in many patients.Much research is therefore directed towards the restorationof the herniated disc either by replacement or regenerativeapproaches. Ideally, current discectomy procedures shouldbe combined with the replacement of the lost NP materialby a scaffold with (native or stem-) cells initiating discregeneration instead of degeneration (Hegewald et al., 2008).
In a recent study, we showed that the mechanicalproperties of the NP can be mimicked using dense scaffoldsof collagen I, which is a natural extracellular matrix protein(Bron et al., 2009b). The scaffold stiffness approached thatof the NP, but the viscous modulus was lower. Aside fromthe difference in viscous behaviour, type I collagen is notan optimal replacement of the NP, which is predominantlycomposed of type II collagen and proteoglycans. Other 3Dscaffold materials, such as alginate, agarose and chitosan,have also been studied for NP regeneration, and thesemight allow a closer match both from a mechanical anda biochemical point of view (Gruber et al., 1997; Leoneet al., 2008; Saad and Spector, 2004). Alginate is mostoften studied since it is inexpensive and does not evokeadverse tissue reactions (Leone et al., 2008; Li et al., 2008;Nunamaker et al., 2007). Alginate is a naturally occurring,water soluble polysaccharide block copolymer composed ofβ-L-mannuronic acid (M) and α-L-guluronic acid (G) that canbe ionically crosslinked by divalent ions, such as calcium(Larsen and Haug, 1971). The resulting matrix has a stiffnesswhich is determined by the alginate concentration and bythe ratio between G and M blocks (Nunamaker et al., 2007).Other conditions such as gelation temperature and type of
crosslinker also influence the final network structure andensuing mechanical properties (Augst et al., 2006).
The aim of this study was to design alginate scaffoldswith viscoelastic properties that mimic those of the NPand to assess the biosynthetic response of native NPcells. We therefore investigated the effects of variationsin alginate concentration on the viscoelastic (rheological)characteristics of scaffolds. In addition, we compared twodifferent methods of inducing alginate gelation, by diffusionand by ‘in situ’ gelation. In diffusion-induced gelation,calcium ions are allowed to diffuse into the alginate gelvia a porous membrane, leading to crosslinking (Nunamakeret al., 2007). “In situ” gelation is performed by mixinginsoluble calcium with the alginate solution and thenreleasing calcium ions within the solution by enzymaticallydecreasing the pH level (Kuo and Ma, 2001). Since it hasbeen documented that alginate scaffolds rapidly loose theirstiffness in vivo (Nunamaker et al., 2007), we monitored thetime-dependent stiffness during prolonged incubation in cellculture medium. Finally, to determine whether variationsin alginate concentration affect cell behaviour, we culturednative cells isolated from goat NP and annulus fibrosus (AF)in alginate beads of different alginate concentrations (2%,4% and 6%). We monitored the gene expression levels ofthe main natural components of the ECM of the NP (typesI and II collagen and aggrecan) up to 4 weeks. The geneexpression levels were compared to gene expression levelsfound in chondrocytes from articular cartilage (AC), for whichextensive studies have been performed of the preservation ofphenotype in alginate (De et al., 2004; Hauselmann et al., 1994;Lin et al., 2009; Masuda et al., 2003).
2. Materials and methods
2.1. Preparation of alginate sample discs by calciumdiffusion
Freeze dried alginate (LVCR sodium alginate, Monsanto, SanDiego, CA) was dissolved in water containing 0.9 wt% sodiumchloride. Alginate solutions at four different concentrations(1, 2, 4, and 6 wt%) were sterilized by autoclaving(121 ◦C,15 min). Sample discs were prepared by pouring2 ml of alginate solution into tissue culture inserts (25 mm,pore size 0.4 µm; Nunc, Roskilde, Denmark). The insertswere placed in Petri dishes containing an aqueous solutionof 500 mM calcium chloride, and a polycarbonate filtermembrane (thickness 8 mm) was placed on top, which wasirrigated with 2 ml of the calcium solution. After two hoursat room temperature, alginate gelation was finished and thesample discs were removed from the culture inserts. Thesamples intended for analysis after prolonged storage inmedium were transferred to 6-well plates containing 5 mlDulbecco’s Modified Eagles Medium (DMEM, Gibco, Paisley,UK) supplemented with 1% streptomycin, penicillin andamphotericin B (all from Gibco). The medium was refreshedevery three days. Samples were assayed at three time points(0, 1 and 10 days), using five separate samples for each timepoint and each alginate concentration.

1198 J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5
2.2. Preparation of alginate sample discs by in situgelation
Insoluble calcium carbonate powder was mixed at aconcentration of 100 mM with alginate solutions in 0.9%NaCl (2%, 4% or 6% alginate) and stirred. The mixture wasacidified by adding the enzyme Glucono Delta-Lactone (GDL,Sigma Chemical Co. (St. Louis, MO)) to a final concentrationof 80 mM. A volume of 2 ml of the acidified mixture wasinjected into the wells of 12-well (well diameter 22mm) platesusing a syringe. After 2 h at room temperature, the gelledsample discs were removed from the wells and transferredto the rheometer for analysis. The samples intended foranalysis after prolonged storage in medium were transferredto 6-well plates containing 5 ml DMEM supplemented with1% antibiotics. The medium was refreshed every threedays. Samples were assayed at two time points (0 and10 days), using five separate samples for each time pointand each alginate concentration. The samples after 10 daysof incubation showed irregular edges and were thereforereduced to a size of 20.0 mm with a cork borer.
2.3. Rheometry
The viscoelastic properties of the alginate discs weremeasured using a stress-controlled rheometer (Paar PhysicaMCR501, Anton Paar, Graz, Austria) equipped with atemperature-controlled steel bottom plate and 20 or 40 mm-diameter steel top plates. The alginate discs showed somevariability in diameter after incubation in culture medium,due to variable degrees of shrinkage. Since variations insample size complicate the interpretation of rheologicaldata, we equalized the sample diameters using cork borers.Samples prepared by diffusion were reduced to a diameter of20 mm at t = 0 and 15.4 mm for t = 1 and 10 days. In situgelled samples were perfectly circular directly after gelation,with a diameter of 22mm; they weremeasured using a 40mmtop plate. After 10 days incubation, the samples showed someedge irregularities. To exclude any edge effects, the incubatedsamples were reduced to a diameter of 20 mm, matchingthe diameter of the 20 mm top plate. For samples with adiameter smaller than the diameter of the rheometer topplate, the absolute values of the shear moduli were correctedas described earlier (Bron et al., 2009b).
To prevent sample slippage, self-adhesive sandpaper(CP918C P180, VSM Abrasives, O’Fallon, Missouri, USA) wasattached to both plates. The discs were loaded between theplates, and the top plate was lowered until the sample wasin good contact with both plates. The tests were performedat a temperature of 37 ◦C in a humidified chamber. Toexclude any time-dependent relaxation during the tests, thesamples were first equilibrated for 10 min. During this time,the normal force decreased to values below 0.25 N in allsamples. Subsequently, we probed the frequency-dependentshear moduli by performing frequency sweep measurementsover an angular frequency range of 0.2–200 rad s−1 at a strainamplitude of 1%, well within the linear regime. Finally, totest the strength of the alginate discs, we subjected them tosinusoidally oscillating shear at a fixed frequency of 0.5 Hzand gradually increasing strain amplitude, until a maximum
of 1000% strain or until sample failure occurred. The shearmodulus G∗(ω) follows from the ratio between stress (σ) andstrain amplitude (γ). G∗ is a complex quantity with an elastic(or storage) modulus (G′) and viscous (or loss) modulus (G′′).The absolute magnitude of the shear modulus, |G∗
|, wascalculated using |G∗
| = ((G′)2 + (G′′)2)0.5. The ratio G′′/G′ isreferred to as the loss tangent, since it equals the tangent ofthe phase angle difference between stress and strain (tan δ).Data reported represent the mean ± S.E. from 5 samples percondition.
2.4. Isolation of native cells and cell culture in alginate
Cartilaginous tissues were obtained from skeletally maturefemale Dutch milk goats (n = 8) that were sacrificedfor other studies. All thoracic and lumbar intervertebraldiscs (IVDs, T1-L2/L6/S1) and articular cartilage (AC) fromthe glenohumeral joint were collected. The IVDs weredissected to separate the nucleus pulposus (NP) from theannulus fibrosus (AF). To assure an adequate cell number,tissues from two goats were mixed for every measurement.Experiments were performed in quadruplicate. The tissueswere dissected and minced, and the cells were releasedby subjected the tissues to sequential treatments first withDMEM supplemented with 1% foetal bovine serum (FBS,HyClone, Logan, UT, USA), 100 U/ml penicillin, 100 µg/mlstreptomycin, 2.5 µg/ml amphotericin B and 2.5% (w/v)Pronase E (Sigma, St. Louis, MO) for 1 h, then with DMEMsupplemented with 25% FBS, 100 U/ml penicillin, 100 µg/mlstreptomycin, 2.5 µg/ml amphotericin B and 0.125% (w/v)collagenase (CLS-2, Worthington, Lakewood, NJ) for 16 h at37 ◦C. After filtering the cell suspension through a 70 µm poresize cell strainer (BD Biosciences, San Diego, CA), isolated cellswere resuspended in an alginate solution (2, 4 and 6 (w/v)in 0.9% NaCl (0.2 µm sterile filtered), creating a suspensionof 4 × 106 cells/ml. The suspension was homogenized byslow pipetting and transferred to a sterile syringe. Alginatebeads were formed by the diffusion method, dripping ∼10 µLdrops of the solution from the syringe needle (26 gauge)into a calcium chloride solution (102 mM). The beads wereallowed to gel by inward diffusion of Ca2+ for 10 min atambient temperature. After washing twice in 0.9% NaCl andtwice in DMEM, the alginate beads were transferred to 24-welltissue culture dishes with 10 beads per well (Greiner Bio-one,Kremsmuenster, Austria). The cells were cultured in 500 µlof DMEM per well, supplemented with 10% FBS, 100 U/mlpenicillin, 100 µg/ml streptomycin, 2.5 µg/ml amphotericinB, and 50 µg/ml ascorbate-2-phosphate (Sigma). We note thatour purpose is to develop a clinical procedure where freshlyharvested cells are immediately transplanted back into thepatient in an alginate scaffold. For this reason, we did not firstdo expansion in 2D culture, but characterized gene expressionfor freshly isolated cells cultured in 3D.
2.5. Real-time PCR
Alginate beads were dissolved in alginate dissolving buffer(55 mM Na-citrate, 0.15 M NaCl, 30 mM Na2 EDTA, pH 6.8),total RNA was isolated from the cells with the RNeasy minikit (Qiagen, Gaithersburg, MD), and DNase I treatment was

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5 1199
performed as described by the manufacturer to remove anycontaminating genomic DNA. Total RNA (750 ng) was re-verse transcribed using 250 U/ml Transcriptor Reverse Tran-scriptase (Roche Diagnostics, Mannheim, Germany), 0.08 Urandom primers (Roche diagnostics), and 1 mM of eachdNTP (Invitrogen, Carlsbad, CA) in Transcriptor RT reac-tion buffer at 42 ◦C for 45 min followed by inactivationof the enzyme at 80 ◦C for 5 min. Real-time PCR reac-tions were performed using the SYBRGreen reaction kitaccording to the manufacturer’s instructions (Roche Diag-nostics) in a LightCycler 480 (Roche Diagnostics). The Light-Cycler reactions were prepared in 20 µl total volume with7 µl PCR-H2O,0.5 µl forward primer (0.2 µM), 0.5 µl reverseprimer (0.2 µM), 10 µl LightCycler Mastermix (LightCycler 480SYBR Green I Master; Roche Diagnostics), to which 2 µl of 5times diluted cDNA was added as PCR template. Primers (In-vitrogen) used for real-time PCR are listed in Table 1. Spe-cific primers were designed from sequences available in databanks, based on homology in conserved domains betweenhuman, mouse, rat, dog and cow. The amplified PCR fragmentextended over at least one exon-border (except for 18S). Ty-rosine 3-monooxygenase/tryptophan 5-monooxygenase acti-vation protein, zeta polypeptide (Ywhaz) and hypoxanthine18S (ribosomal RNA) were used as housekeeping genes andthe gene expression levels were normalized using a normal-ization factor calculated with the equation
√(Ywhaz x 18S).
With the LightCycler software (version 4), the crossing pointswere assessed and plotted versus the serial dilution of knownconcentrations of the standards derived from each gene usingthe Fit Points method. PCR efficiency was calculated by Light-Cycler software and the data were used only if the calculatedPCR efficiency was between 1.85 and 2.0.
2.6. Statistical analysis
For the rheological measurements, unpaired Students’T-test was used for statistical analysis. P < 0.05 was con-sidered as significant. For the real-time PCR experiments,Friedman’s non-parametric rank test was used to determinestatistically significant differences within an experiment.When statistically significant differences were detected, as-sessment of differences between individual groups was per-formed using Wilcoxon’s signed-rank test.
3. Results
3.1. Alginate sample discs prepared by different methods
We prepared alginate discs of concentrations between 1and 6 wt% by two different methods, namely by diffusionof Ca2+ ions from outside or by in situ release of Ca2+
from calcium carbonate inside the alginate. To characterizethe viscoelastic properties, we performed small amplitudeoscillatory shear tests on the alginate discs. Both series ofsamples became significantly stiffer with increasing alginateconcentration (square symbols, upper panel Fig. 1). However,the samples prepared by diffusion (black squares) were atleast ten-fold stiffer than the in situ gelated samples (greysquares) at all alginate concentrations (significant with P <
Fig. 1 – Linear viscoelastic behaviour of alginate matrices.Upper panel: dependence of scaffold stiffness (G′, squares),and viscous modulus (G′′, triangles), on alginateconcentration, for samples prepared by diffusion (blacksymbols) and in situ gelation (grey symbols), measured at10 rad/s. Bottom panel: dependence of the loss tangent ofalginate scaffolds on alginate concentration, for samplesprepared by diffusion (black circles) and in situ gelation(grey circles).
0.05). The sample-to-sample variability was higher for thediffusion series than for the in situ series, as shown by thelarger error bars. This indicates that the diffusion sampleswere less homogeneous than the in situ gelled samples,consistent with prior observations (Nunamaker et al., 2007).The viscous modulus of the samples prepared by diffusion(black triangles) was also significantly larger than that ofthe in situ polymerized samples (grey triangles). The losstangent (G′′/G′) was independent of alginate concentrationfor the diffusion gelated samples (P > 0.05), as shown in thebottom panel of Fig. 1 (black circles). For the in situ gelledsamples (grey circles), the 4% and 6% alginate samples had asignificantly lower loss tangent than the 2% alginate samples(P < 0.05). The loss tangent of the samples prepared bydiffusion (black circles) was significantly higher than that ofsamples that were gelled in situ (grey circles). These findingsimplicate that the diffusion samples are stiffer but also havea higher viscosity.
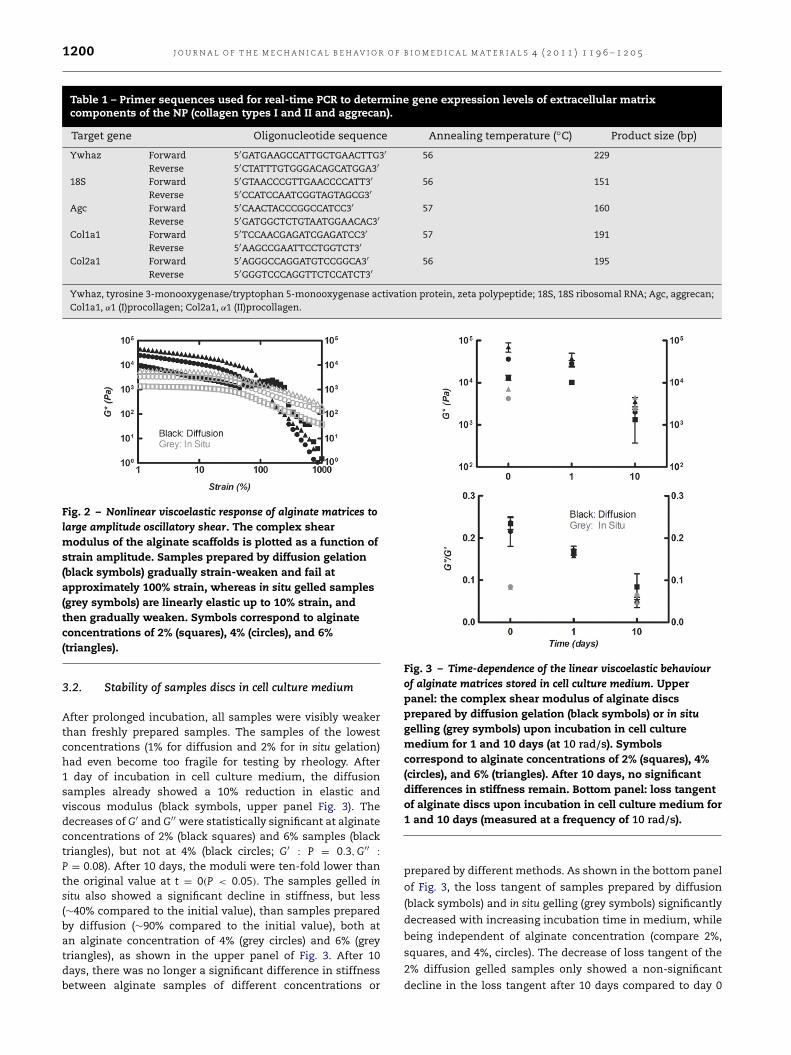
To characterize the nonlinear viscoelastic behaviour, wesubjected the alginate discs to large amplitude oscillatoryshear. The alginate samples prepared by diffusion showedno appreciable linear elastic regime: their shear modulusimmediately started to decrease as the strain amplitude wasraised, and they failed already at strains of about 100% (blacksymbols in Fig. 2). In contrast, the samples prepared by in situgelation were linearly elastic up to strains of about 10%, andthereafter gradually strain-weakened (grey symbols in Fig. 2).
1200 J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5
Table 1 – Primer sequences used for real-time PCR to determine gene expression levels of extracellular matrixcomponents of the NP (collagen types I and II and aggrecan).
Target gene Oligonucleotide sequence Annealing temperature (◦C) Product size (bp)
Ywhaz Forward 5′GATGAAGCCATTGCTGAACTTG3′ 56 229Reverse 5′CTATTTGTGGGACAGCATGGA3′
18S Forward 5′GTAACCCGTTGAACCCCATT3′ 56 151Reverse 5′CCATCCAATCGGTAGTAGCG3′
Agc Forward 5′CAACTACCCGGCCATCC3′ 57 160Reverse 5′GATGGCTCTGTAATGGAACAC3′
Col1a1 Forward 5′TCCAACGAGATCGAGATCC3′ 57 191Reverse 5′AAGCCGAATTCCTGGTCT3′
Col2a1 Forward 5′AGGGCCAGGATGTCCGGCA3′ 56 195Reverse 5′GGGTCCCAGGTTCTCCATCT3′
Ywhaz, tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide; 18S, 18S ribosomal RNA; Agc, aggrecan;Col1a1, α1 (I)procollagen; Col2a1, α1 (II)procollagen.
Fig. 2 – Nonlinear viscoelastic response of alginate matrices tolarge amplitude oscillatory shear. The complex shearmodulus of the alginate scaffolds is plotted as a function ofstrain amplitude. Samples prepared by diffusion gelation(black symbols) gradually strain-weaken and fail atapproximately 100% strain, whereas in situ gelled samples(grey symbols) are linearly elastic up to 10% strain, andthen gradually weaken. Symbols correspond to alginateconcentrations of 2% (squares), 4% (circles), and 6%(triangles).
3.2. Stability of samples discs in cell culture medium
After prolonged incubation, all samples were visibly weakerthan freshly prepared samples. The samples of the lowestconcentrations (1% for diffusion and 2% for in situ gelation)had even become too fragile for testing by rheology. After1 day of incubation in cell culture medium, the diffusionsamples already showed a 10% reduction in elastic andviscous modulus (black symbols, upper panel Fig. 3). Thedecreases of G′ and G′′ were statistically significant at alginateconcentrations of 2% (black squares) and 6% samples (blacktriangles), but not at 4% (black circles; G′
: P = 0.3,G′′:
P = 0.08). After 10 days, the moduli were ten-fold lower thanthe original value at t = 0(P < 0.05). The samples gelled insitu also showed a significant decline in stiffness, but less(∼40% compared to the initial value), than samples preparedby diffusion (∼90% compared to the initial value), both atan alginate concentration of 4% (grey circles) and 6% (greytriangles), as shown in the upper panel of Fig. 3. After 10days, there was no longer a significant difference in stiffnessbetween alginate samples of different concentrations or
Fig. 3 – Time-dependence of the linear viscoelastic behaviourof alginate matrices stored in cell culture medium. Upperpanel: the complex shear modulus of alginate discsprepared by diffusion gelation (black symbols) or in situgelling (grey symbols) upon incubation in cell culturemedium for 1 and 10 days (at 10 rad/s). Symbolscorrespond to alginate concentrations of 2% (squares), 4%(circles), and 6% (triangles). After 10 days, no significantdifferences in stiffness remain. Bottom panel: loss tangentof alginate discs upon incubation in cell culture medium for1 and 10 days (measured at a frequency of 10 rad/s).
prepared by different methods. As shown in the bottom panel
of Fig. 3, the loss tangent of samples prepared by diffusion
(black symbols) and in situ gelling (grey symbols) significantly
decreased with increasing incubation time in medium, while
being independent of alginate concentration (compare 2%,
squares, and 4%, circles). The decrease of loss tangent of the
2% diffusion gelled samples only showed a non-significant
decline in the loss tangent after 10 days compared to day 0

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5 1201
Table 2 – Summary of reported stiffnesses ofcartilaginous tissues (Iatridis et al., 1996).
Tissue G∗ (kPa)
Nucleus pulposus 11Meniscus 100Annulus Fibrosus 540Articular cartilage 600
and 1 (P = 0.07 and P = 0.16). After 10 days, the loss tangentsof all samples were statistically indistinguishable.
3.3. ECM gene expression
To assess the influence of the alginate matrices on thebiosynthetic phenotype of native tissue cells, we cultured NP,AC, and AF cells isolated from goat IVDs and cartilage insidealginate beads with concentrations of 2%–6% alginate. Wemeasured the expression levels of NP-specific extracellularmatrix components (collagen types I and II and aggrecan)by real-time PCR. Cells freshly isolated from the AF showedthe highest level of type I collagen gene expression andchondrocytes from AC showed the lowest expression level(Fig. 4, T = 0; differences statistically significant with p <
0.05). Upon culturing in alginate beads, there was an increasein type I collagen gene expression by all the cells after 1 week(p < 0.001) which was sustained after 2 and 4 weeks (Fig. 4).However, this increase was not significantly influenced by thealginate concentration over a range of 2%–6% (p > 0.05). Theincrease in gene expression of type I collagen was strongestfor the NP cells. After 4 weeks of culture, the expressionlevel of type I collagen for NP cells was similar to the levelsfound in AF cells (p = 0.08). The levels found in AC cellsconsistently remained the lowest (p < 0.05). The highest typeII collagen gene expression levels directly after cell isolationwere found in AC cells and the lowest in AF cells (Fig. 5T = 0; differences statistically significant with p < 0.05).The gene expression level of type II collagen for all cell typeswas decreased significantly after 1 week of culture in alginate(p < 0.001), but thereafter remained constant (Fig. 5, p > 0.3).Levels of type II collagen expression remained the lowest inAF cells (p < 0.05), while NP and AC cells had similar levelsof expression after 1 or more weeks culturing in alginate(p = 0.067). Levels of aggrecan gene expression were highestfor NP cells after isolation (Fig. 6 T = 0,p < 0.05). Culturein alginate led to a steady decrease of the aggrecan geneexpression levels of all three cell types, which was alreadynoticeable after 1 week (p < 0.001). NP cells had higheraggrecan gene expression levels than AF and AC cells (p <
0.05). No significant differences in the gene expression levelsfor type I and II collagen and aggrecan could be found in anyof the cell populations when cultured in beads with alginateconcentrations of 2% (white bars), 4% (grey bars), or 6% (blackbars) (Figs. 4–6; p > 0.05).
4. Discussion
The principal aim of the current study was to design alginatescaffolds with viscoelastic properties that mimic those of
the NP. We showed that the stiffness of alginate scaffolds,crosslinked by diffusion of calcium into the alginate solution,can be varied over two orders of magnitude (between 1 kPaand almost 100 kPa) by varying the alginate concentration.The loss tangent was not affected by variations in polymerconcentration. The closest matching of the stiffness as wellas loss tangent of a healthy NP, which has a stiffness of11 kPa and loss tangent of about 0.24 (Iatridis et al., 1996), wasfound for 2% alginate scaffolds. Other cartilaginous tissuesare stiffer than even the most concentrated alginate discs(6%) (Table 2). It is difficult to prepare more concentratedalginate discs because the high viscosity of the pre-gelledsolution renders it difficult to process and mould the gel andto mix in cells (Augst et al., 2006). However, this difficultymay be counteracted by stirring the alginate solutions, whichare shear-thinning (Augst et al., 2006). Moreover, the stiffnessof alginate scaffolds may be tuned by other factors, suchas the alginate source, G/M ratio, cross linker type, andtemperature (Augst et al., 2006; Kuo and Ma, 2001; Leoneet al., 2008; Nunamaker et al., 2007). However, these factorsare not always accessible for manipulation in the contextof tissue engineering. The G/M ratio depends on alginatesource and processing and is therefore usually fixed upondelivery (Kuo and Ma, 2001; Larsen and Haug, 1971). Changesin Ca2+ concentration and temperature influence gelationtime and thereby matrix organization and stiffness (Augstet al., 2006), but these variations are not always well toleratedby seeded cells. We also compared samples derived bydiffusion gelation to samples prepared by in situ release ofCa2+ (Nunamaker et al., 2007). The “in situ” method hasseveral practical advantages over the diffusion method forclinical applications in tissue engineering. The liquid solutioncan be injected via a syringe and gelation occurs insidethe tissue of interest. Moreover, in situ gelation results inmore homogeneous scaffolds with less spatial and sample-to-sample variation in biomechanical properties. Scaffoldsprepared by diffusion are notoriously inhomogeneous due tothe diffusion kinetics of calcium ions. Although we preparedthe diffusion scaffolds under standardized circumstances, thestructural inhomogeneity appeared to affect the rheology andmay have affected cell phenotype. The stiffness of alginatesamples prepared by in situ gelation was much lower thanthat of the diffusion samples, consistent with prior findings(Li et al., 2008).
To assess the applicability of alginate discs for IVD repair, itis also crucial to characterize the biological response of nativetissue cells to prolonged culture in an alginate matrix. Wetherefore screened the effects of alginate scaffolds (preparedby diffusion) on native cells by measuring gene expression ofextracellular matrix components that are naturally presentin the NP. The relative gene expression levels for types Iand II collagen found in native IVD cells after isolation is inline with the well-known collagenous composition of thesetissues (Almarza and Athanasiou, 2004). The AF contains amixture of type I and II collagen, the NP contains more typeII collagen than the AF, and the AC contains predominantlytype II collagen (Almarza and Athanasiou, 2004; Kuettnerand Cole, 2005). Aside from collagens, proteoglycans (mainlyaggrecan) are the main components of cartilage. The NPcontains the highest amount of proteoglycans and the AF the

1202 J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5
A B
C D
Fig. 4 – Gene expression levels for type I collagen of native cells cultured in alginate matrices. Real-time PCR was performed onreverse-transcribed RNA isolated from cells derived from the NP, AF and AC of goat intervertebral discs after 0, 7, 14, and 28days of culture in alginate beads with a concentration of 2% (white bars), 4% (grey bars) and 6% (black bars). The geneexpression level of type I collagen (Col1a1) is normalized by the expression levels of two housekeeping genes (2hk). Dataare shown as mean ± SD. Differences between NP, AF, and AC cells are statistically significant with p < 0.05 at all timepoints and alginate concentrations (except for NP–AF in Fig. 4(D), p = 0.08). The dependence on alginate concentration foreach cell type is not statistically significant (p > 0.05). Changes with time are significant for all cells on going from T = 0 tolater time points (p < 0.001), and for NP cells there is a significant increase between T = 1 week to 4 weeks (p = 0.03).Otherwise, there are no statistically significant time changes.
lowest amount (Almarza and Athanasiou, 2004; Kuettner andCole, 2005). This is reflected by the high gene expression levelsof aggrecan seen in native NP cells and the low levels foundin native AF cells. The differences found between the cellpopulations after isolation were mostly maintained duringculture in alginate beads, suggesting that alginate preservesthe phenotypical characteristics of the cells. Although weobserved a decrease in the gene expression levels of typeII collagen and aggrecan during culture in alginate beads,it has been reported that AC and IVD cells do produceconsiderable amounts of both proteins under these cultureconditions (Chubinskaya et al., 2001; Hauselmann et al., 1994;Masuda et al., 2003; Vonk et al., 2010). In contrast, cellscultured on polystyrene dishes lose the ability to synthesizeaggrecan and type II collagen and start to produce moretype I collagen (Darling and Athanasiou, 2005). We did notobserve any effect of variations in the alginate concentrationon gene expression levels of type I and type II collagen oraggrecan by NP, AF or AC cells cultured within alginate gels(Figs. 4–6). This observation is in contrast with observationson other cell types cultured on top of flat elastic hydrogels,
where a pronounced influence of matrix stiffness on ECMsynthesis has been documented (Breuls et al., 2008). However,a requirement for a stiffness-responsive cell phenotype isthat the cell adheres to the scaffold material via integrinadhesions so that the cell can exert traction forces to thematrix by actomyosin contractility (Discher et al., 2009). Cellshave no integrin receptors for alginate and therefore adherevery weakly to alginate matrices (Augst et al., 2006). They cantherefore not actively respond to matrix stiffness via focaladhesion sites. Several methods to promote cell attachmentto alginate matrices are currently being studied; for instance,by coupling of extracellular matrix proteins such as laminin,collagen, fibronectin, or RGD peptides (Alsberg et al., 2001;Augst et al., 2006; Huebsch et al., 2010; Degala et al., 2011).We note that another reason for the lack of sensitivity tomatrix stiffness observed in our study may be the rapidreduction of matrix stiffness upon prolonged exposure tocell culture medium. Similarly, it has been documented thatalginate scaffolds rapidly soften after implantation in vivo(Nunamaker et al., 2007). This softening has been attributedto the loss of divalent crosslinking cations at neutral pH

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5 1203
A
C D
B
Fig. 5 – Gene expression levels for type II collagen of native cells cultured in alginate matrices. Real-time PCR was performed onreverse-transcribed RNA isolated from cells derived from the NP, AF and AC of goat intervertebral discs after 0, 7, 14, and 28days of culture in alginate beads with a concentration of 2% (white bars), 4% (grey bars) and 6% (black bars). The geneexpression level of type II collagen (Col2a1) is normalized by the expression levels of two housekeeping genes (2hk). Dataare shown as mean ± SD. Differences between NP, AF, and AC cells are statistically significant with p < 0.05 at all timepoints and alginate concentrations (except for NP-AC in Fig. 4(B), p = 0.067). The dependence on alginate concentration foreach cell type is not statistically significant (p > 0.05). There is only a significant change with time for all cells on going fromT = 0 to T = 1 or more weeks (p < 0.001).
(Augst et al., 2006). In our study, we observed a 10% declinein stiffness after 1 day and a ten-fold decline after 10 daysin medium in samples prepared by diffusion (Fig. 3). The‘in situ’ gelled samples softened less in medium, but weremuch softer than samples prepared by diffusion to beginwith, so that their stiffness after 10 days was similar to thatof the diffusion samples. The consequence of the loss ofstiffness is that the 2% alginate scaffold no longer matchedthe stiffness of the NP after 10 days. Moreover, even thestiffness of the 6% alginate scaffolds was lower than that ofthe NP after 10 days. Several methods to prevent the lossof stiffness during storage in medium have been reported.Arguably the most straightforward method is to supplementCa2+ to the medium. However, this strategy is not attractivein the context of NP regeneration, where chondrogenicdifferentiation is required, since calcium has osteogeniceffects. Moreover, the Ca2+ concentration of the environmentis difficult to control after in vivo implantation. An alternativestrategy is the addition of cationic polyethyleneimine toalginate to increase the resistance to de-crosslinking (Kuo andMa, 2001, 2008). Finally, the addition of cells is known to havestabilizing effects on the alginate matrix (Augst et al., 2006).We could not test this stabilizing effect with native IVD cells,
because there were not enough cells available for the largesample sizes required for rheology. In fact, limited availabilityof cells is also a limiting factor for IVD engineering withnative cells (Bron et al., 2009a). In patients, the availabilityof IVD tissue for digestion is even less than in our study, inwhich we used pooled IVDs derived from goat thoracic spines.An attractive alternative is the use of mesenchymal stemcells (Lutolf et al., 2009), but differentiation towards NP or AFphenotypes is still not fully directional (Bron et al., 2009a).Furthermore, specific phenotypic markers to distinguish bothcell types are currently being elucidated (Lee et al., 2007;Rutges et al., 2010; Sakai et al., 2009; Vonk et al., 2010).
In conclusion, we showed that the stiffness of alginatescaffolds can be varied by tuning the alginate polymerconcentration and can be matched to the stiffness of theNP. Moreover, the biosynthetic phenotype of native IVD cellsis maintained upon prolonged culture in alginate matrices.There are still some practical limitations that need to besolved, specifically the long-term mechanical stability in vivo,the bioadhesive properties, and the availability of tissue cellsfrom the patient. Current study underscores the potentialof alginate as a scaffold material for IVD engineering, butmore importantly reveals some important limitations, which

1204 J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5
A B
C D
Fig. 6 – Gene expression levels for aggrecan of native cells cultured in alginate matrices. Real-time PCR was performed onreverse-transcribed RNA isolated from cells derived from the NP, AF and AC of goat intervertebral discs after 0, 7, 14, and 28days of culture in alginate beads with a concentration of 2% (white bars), 4% (grey bars) and 6% (black bars). The geneexpression level of aggrecan (Acan) is normalized by the expression levels of two housekeeping genes (2hk). Data areshown as mean ± SD. Differences between NP, AF, and AC cells are statistically significant with p < 0.05 at all time pointsand alginate concentrations. The dependence on alginate concentration for each cell type is not statistically significant(p > 0.05). There is only a significant change with time for all cells on going from T = 0 to T = 1 or more weeks (p < 0.001)and for the AC cells between week 2 and 4 (p = 0.002).
in spite of many promising research over the past decade, stillhave to be overcome.
R E F E R E N C E S
Almarza, A.J., Athanasiou, K.A., 2004. Design characteristics forthe tissue engineering of cartilaginous tissues. Ann. Biomed.Eng. 32, 2–17.
Alsberg, E., Anderson, K.W., Albeiruti, A., Franceschi, R.T., Mooney,D.J., 2001. Cell-interactive alginate hydrogels for bone tissueengineering. J. Dent. Res. 80, 2025–2029.
Augst, A.D., Kong, H.J., Mooney, D.J., 2006. Alginate hydrogels asbiomaterials. Macromol. Biosci. 6, 623–633.
Breuls, R.G., Jiya, T.U., Smit, T.H., 2008. Scaffold stiffnessinfluences cell behavior: opportunities for skeletal tissueengineering. Open. Orthop. J. 2, 103–109.
Bron, J.L., Helder, M.N., Meisel, H.J., Van Royen, B.J., Smit, T.H.,2009a. Repair, regenerative and supportive therapies of theannulus fibrosus: achievements and challenges. Eur. Spine J.18, 301–313.
Bron, J.L., Koenderink, G.H., Everts, V., Smit, T.H., 2009b.Rheological characterization of the nucleus pulposus anddense collagen scaffolds intended for functional replacement.J. Orthop. Res. 27, 620–626.
Bron, J.L., Van der Veen, A.J., Helder, M.N., Van Royen, B.J.,Smit, T.H., 2010. Biomechanical and in vivo evaluation ofexperimental closure devices of the annulus fibrosus designedfor a goat nucleus replacement model. Eur. Spine J. 19,1347–1355.
Chubinskaya, S., Huch, K., Schulze, M., Otten, L., Aydelotte,M.B., Cole, A.A., 2001. Gene expression by human articularchondrocytes cultured in alginate beads. J. Histochem.Cytochem. 49, 1211–1220.
Darling, E.M., Athanasiou, K.A., 2005. Rapid phenotypic changesin passaged articular chondrocyte subpopulations. J. Orthop.Res. 23, 425–432.
De, C.F., Lesur, C., Pastoureau, P., Caliez, A., Sabatini, M., 2004.Culture of chondrocytes in alginate beads. Methods Mol. Biol.100, 15–22.
Degala, S., Zipfel, W.R., Bonassar, L.J., 2011. Chondrocyte calciumsignaling in response to fluid flow is regulated by matrixadhesion in 3-D alginate scaffolds. Arch. Biochem. Biophys.505, 112–117.
Discher, D.E., Janmey, P., Wang, Y.L., 2005. Tissue cells feeland respond to the stiffness of their substrate. Science 310,1139–1143.
Discher, D.E., Mooney, D.J., Zandstra, P.W., 2009. Growth factors,matrices, and forces combine and control stem cells. Science324, 1673–1677.

J O U R N A L O F T H E M E C H A N I C A L B E H AV I O R O F B I O M E D I C A L M A T E R I A L S 4 ( 2 0 1 1 ) 1 1 9 6 – 1 2 0 5 1205
Engler, A.J., Sweeney, H.L., Discher, D.E., Schwarzbauer, J.E., 2007.Extracellular matrix elasticity directs stem cell differentiation.J. Musculoskelet. Neuronal. Interact. 7, 335.
Gruber, H.E., Fisher Jr., E.C., Desai, B., Stasky, A.A., Hoelscher, G.,Hanley Jr., E.N., 1997. Human intervertebral disc cells from theannulus: three-dimensional culture in agarose or alginate andresponsiveness to TGF-beta1. Exp. Cell Res. 235, 13–21.
Hauselmann, H.J., Fernandes, R.J., Mok, S.S., Schmid, T.M.,Block, J.A., Aydelotte, M.B., Kuettner, K.E., Thonar, E.J., 1994.Phenotypic stability of bovine articular chondrocytes afterlong-term culture in alginate beads. J. Cell Sci. 107 (Pt. 1),17–27.
Hegewald, A.A., Ringe, J., Sittinger, M., Thome, C., 2008.Regenerative treatment strategies in spinal surgery. Front.Biosci. 13, 1507–1525.
Hubbell, J.A., 2003. Materials as morphogenetic guides in tissueengineering. Curr. Opin. Biotechnol. 14, 551–558.
Huebsch, N., Arany, P.R., Mao, A.S., Shvartsman, D., Ali,O.A., Bencherif, S.A., Rivera-Feliciano, J., Mooney, D.J., 2010.Harnessing traction-mediated manipulation of the cell/matrixinterface to control stem-cell fate. Nat. Mater. 9, 518–526.
Huebsch, N., Mooney, D.J., 2009. Inspiration and application in theevolution of biomaterials. Nature 462, 426–432.
Iatridis, J.C., Weidenbaum, M., Setton, L.A., Mow, V.C., 1996. Is thenucleus pulposus a solid or a fluid? Mechanical behaviors ofthe nucleus pulposus of the human intervertebral disc. Spine21, 1174–1184 (Phila Pa 1976).
Kuettner, K.E., Cole, A.A., 2005. Cartilage degeneration in differenthuman joints. Osteoarthr. Cartilage. 13, 93–103.
Kuo, C.K., Ma, P.X., 2001. Ionically crosslinked alginate hydrogelsas scaffolds for tissue engineering: part 1. Structure, gelationrate and mechanical properties. Biomaterials 22, 511–521.
Kuo, C.K., Ma, P.X., 2008. Maintaining dimensions and mechanicalproperties of ionically crosslinked alginate hydrogel scaffoldsin vitro. J. Biomed. Mater. Res. Part A 84, 899–907.
Larsen, B., Haug, A., 1971. Biosynthesis of alginate. 1. Compositionand structure of alginate produced by Azotobacter vinelandii(Lipman). Carbohydr. Res. 17, 287–296.
Lee, C.R., Sakai, D., Nakai, T., Toyama, K., Mochida, J., Alini,M., Grad, S., 2007. A phenotypic comparison of intervertebraldisc and articular cartilage cells in the rat. Eur. Spine J. 16,2174–2185.
Leone, G., Torricelli, P., Chiumiento, A., Facchini, A., Barbucci,R., 2008. Amidic alginate hydrogel for nucleus pulposusreplacement. J. Biomed. Mater. Res. Part A 84, 391–401.
Li, Z., Gunn, J., Chen, M.H., Cooper, A., Zhang, M., 2008. On-sitealginate gelation for enhanced cell proliferation and uniformdistribution in porous scaffolds. J. Biomed. Mater. Res. Part A86, 552–559.
Lin, Y.J., Yen, C.N., Hu, Y.C., Wu, Y.C., Liao, C.J., Chu, I.M., 2009.Chondrocytes culture in three-dimensional porous alginatescaffolds enhanced cell proliferation, matrix synthesis andgene expression. J. Biomed. Mater. Res. Part A 88, 23–33.
Lutolf, M.P., Gilbert, P.M., Blau, H.M., 2009. Designing materials todirect stem-cell fate. Nature 462, 433–441.
Masuda, K., Sah, R.L., Hejna, M.J., Thonar, E.J., 2003. A novel two-step method for the formation of tissue-engineered cartilageby mature bovine chondrocytes: the alginate-recovered-chondrocyte (ARC) method. J. Orthop. Res. 21, 139–148.
Nunamaker, E.A., Purcell, E.K., Kipke, D.R., 2007. In vivo stabilityand biocompatibility of implanted calcium alginate disks.J. Biomed. Mater. Res. Part A 83, 1128–1137.
Roberts, S., Evans, H., Trivedi, J., Menage, J., 2006. Histology andpathology of the human intervertebral disc. J. Bone Joint Surg.Am. 88 (Suppl. 2), 10–14.
Rutges, J., Creemers, L.B., Dhert, W., Milz, S., Sakai, D., Mochida,J., Alini, M., Grad, S., 2010. Variations in gene and proteinexpression in human nucleus pulposus in comparison withannulus fibrosus and cartilage cells: potential associationswith aging and degeneration. Osteoarthr. Cartilage. 18,416–423.
Saad, L., Spector, M., 2004. Effects of collagen type onthe behavior of adult canine annulus fibrosus cells incollagen–glycosaminoglycan scaffolds. J. Biomed. Mater. Res.Part A 71, 233–241.
Sakai, D., Nakai, T., Mochida, J., Alini, M., Grad, S., 2009. Differen-tial phenotype of intervertebral disc cells: microarray and im-munohistochemical analysis of canine nucleus pulposus andanulus fibrosus. Spine 34, 1448–1456. (Phila Pa 1976).
Vonk, L.A., Kroeze, R.J., Doulabi, B.Z., Hoogendoorn, R.J., Huang,C., Helder, M.N., Everts, V., Bank, R.A., 2010. Caprine articular,meniscus and intervertebral disc cartilage: an integral analysisof collagen network and chondrocytes. Matrix Biol. 29,209–218.
Related Documents











