Enforced Expression of GATA-3 in Transgenic Mice Inhibits Th1 Differentiation and Induces the Formation of a T1/ST2- Expressing Th2-Committed T Cell Compartment In Vivo 1 Martijn C. Nawijn,* Gemma M. Dingjan,* ² Rita Ferreira, ² Bart N. Lambrecht, ‡ Alar Karis, ²§ Frank Grosveld, ² Huub Savelkoul,* and Rudolf W. Hendriks 2 * ² The transcription factor GATA-3 is essential for early T cell development and differentiation of naive CD4 1 T cells into Th2 effector cells. To study the function of GATA-3 during T cell-mediated immune responses in vivo, we investigated CD2-GATA3- transgenic mice in which GATA-3 expression is driven by the CD2 locus control region. Both in the CD4 1 and the CD8 1 T cell population the proportion of cells exhibiting a CD44 high CD45RB low CD62L low Ag-experienced phenotype was increased. In CD2- GATA3-transgenic mice, large fractions of peripheral CD4 1 T cells expressed the IL-1 receptor family member T1/ST2, indicative of advanced Th2 commitment. Upon in vitro T cell stimulation, the ability to produce IL-2 and IFN-g was decreased. Moreover, CD4 1 T cells manifested rapid secretion of the Th2 cytokines IL-4, IL-5, and IL-10, reminiscent of Th2 memory cells. In contrast to wild-type CD4 1 cells, which lost GATA-3 expression when cultured under Th1-polarizing conditions, CD2-GATA3-transgenic CD4 1 cells maintained expression of GATA-3 protein. Under Th1 conditions, cellular proliferation of CD2-GATA3-transgenic CD4 1 cells was severely hampered, IFN-g production was decreased and Th2 cytokine production was increased. Enforced GATA-3 expression inhibited Th1-mediated in vivo responses, such as Ag-specific IgG2a production or a delayed-type hypersen- sitivity response to keyhole limpet hemocyanin. Collectively, these observations indicate that enforced GATA-3 expression selec- tively inhibits Th1 differentiation and induces Th2 differentiation. The increased functional capacity to secrete Th2 cytokines, along with the increased expression of surface markers for Ag-experienced Th2-committed cells, would argue for a role of GATA-3 in Th2 memory formation. The Journal of Immunology, 2001, 167: 724 –732. T he CD4 1 Th lymphocytes develop into two functionally distinct subsets that can be distinguished on the basis of their cytokine production profile (1, 2). Th1 cells are char- acterized by the production of IFN-g and TNF-b, whereas Th2 cells typically produce IL-4, IL-5, IL-10, and IL-13. Each subset mediates distinct effector functions in vivo. Th1 cells are predom- inantly involved in immune responses against intracellular patho- gens and are associated with autoimmune disease. Th2 cells are of importance in the defense against extracellular pathogens and are implicated in atopy and allergic diseases (3–5). Both Th1 and Th2 cells are derived from a common naive precur- sor (4 – 6). Signaling pathways initiated by cytokines play a dominant role in driving the differentiation of activated naive CD4 1 T cells into either effector phenotype (2, 7). For instance, IL-12 induces the differentiation of naive Th cells to the Th1 effector phenotype (8–11) by activation of the transcription factor Stat4 (12–14). On the other hand, Th2 differentiation is mediated by Stat6 activation through IL-4 receptor engagement (15–21). In response to chronic antigenic stim- ulation in vivo, progressive polarization of the cytokine responses ultimately leads to the commitment of Th cells to mutually exclusive Th phenotypes, which are thought to be maintained independently of extrinsic factors (22, 23). Stat6 induces the expression of the transcription factors GATA-3 and c-Maf (24), which have been shown to be selectively expressed in a Th2-specific fashion (25–27). Using Stat6-deficient cells it has been shown that, although IL-4 and Stat6 signaling may initially direct Th2 development, GATA-3 and c-Maf are capable of inducing the development of stabile Th2 commitment, indepen- dent of Stat6 (28). In vitro differentiation into Th1 cells induces chromatin remodeling of the IFN-g locus and, conversely, the dif- ferentiation into Th2 cells induces remodeling of the IL-4/IL-5/ IL-13 locus (29, 30). Recently, GATA-3 has been shown to play an instructive role in directing Th2 differentiation (31). During early T cell development, GATA-3 gene expression is required for the development of the earliest T cell progenitors (32– 34). GATA-3 levels are low during the two phases of TCR gene rearrangement, but are high in the fraction of rapidly proliferating cells that insulates these two periods of TCR rearrangement (33). GATA-3 expression remains high in CD4 1 thymocytes, but pro- gressively declines in CD8 1 thymocytes (see accompanying pa- per). GATA-3 is detected in naive CD4 1 T cells and expression levels increase substantially during Th2 differentiation (26, 27). GATA-3 expression has been shown to be indispensable for Th2 development and is down-regulated in response to IL-12-mediated Stat4 activation (27, 35). GATA-3 strongly transactivates the IL-5 promoter, but appears to have only limited effects on IL-4 gene transcription (27, 36, 37). Retroviral introduction of GATA-3 dur- ing in vitro Th1 differentiation of naive CD4 1 T cells resulted in an inhibition of IFN-g production, independently of IL-4 (35, 38), Departments of *Immunology, ² Cell Biology and Genetics, and ‡ Pulmonary Medi- cine, Faculty of Medicine, Erasmus University Rotterdam, Rotterdam, The Nether- lands; and § Institute of Molecular and Cell Biology, University of Tartu, Tartu, Estonia Received for publication April 12, 2000. Accepted for publication May 8, 2001. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This work was partly supported by Grant 3069 from the Estonian Science Founda- tion (to A.K.) and by the Royal Academy of Arts and Sciences (to R.W.H.). 2 Address correspondence and reprint requests to Dr. Rudolf W. Hendriks, Depart- ment of Immunology, Faculty of Medicine, Room Ee853, Erasmus University Rot- terdam, Dr. Molewaterplein 50, P.O. Box 1738, 3000 DR Rotterdam, The Nether- lands. E-mail address: [email protected] Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Enforced Expression of GATA-3 in Transgenic Mice InhibitsTh1 Differentiation and Induces the Formation of a T1/ST2-Expressing Th2-Committed T Cell Compartment In Vivo1
Martijn C. Nawijn,* Gemma M. Dingjan,* † Rita Ferreira, † Bart N. Lambrecht,‡ Alar Karis, †§
Frank Grosveld,† Huub Savelkoul,* and Rudolf W. Hendriks2*†
The transcription factor GATA-3 is essential for early T cell development and differentiation of naive CD41 T cells into Th2effector cells. To study the function of GATA-3 during T cell-mediated immune responses in vivo, we investigatedCD2-GATA3-transgenic mice in whichGATA-3 expression is driven by theCD2 locus control region. Both in the CD41 and the CD81 T cellpopulation the proportion of cells exhibiting a CD44highCD45RBlowCD62Llow Ag-experienced phenotype was increased. InCD2-GATA3-transgenic mice, large fractions of peripheral CD41 T cells expressed the IL-1 receptor family member T1/ST2, indicativeof advanced Th2 commitment. Upon in vitro T cell stimulation, the ability to produce IL-2 and IFN-g was decreased. Moreover,CD41 T cells manifested rapid secretion of the Th2 cytokines IL-4, IL-5, and IL-10, reminiscent of Th2 memory cells. In contrastto wild-type CD41 cells, which lost GATA-3 expression when cultured under Th1-polarizing conditions,CD2-GATA3-transgenicCD41 cells maintained expression of GATA-3 protein. Under Th1 conditions, cellular proliferation ofCD2-GATA3-transgenicCD41 cells was severely hampered, IFN-g production was decreased and Th2 cytokine production was increased. EnforcedGATA-3 expression inhibited Th1-mediated in vivo responses, such as Ag-specific IgG2a production or a delayed-type hypersen-sitivity response to keyhole limpet hemocyanin. Collectively, these observations indicate that enforcedGATA-3 expression selec-tively inhibits Th1 differentiation and induces Th2 differentiation. The increased functional capacity to secrete Th2 cytokines,along with the increased expression of surface markers for Ag-experienced Th2-committed cells, would argue for a role of GATA-3in Th2 memory formation. The Journal of Immunology,2001, 167: 724–732.
T he CD41 Th lymphocytes develop into two functionallydistinct subsets that can be distinguished on the basis oftheir cytokine production profile (1, 2). Th1 cells are char-
acterized by the production of IFN-g and TNF-b, whereas Th2cells typically produce IL-4, IL-5, IL-10, and IL-13. Each subsetmediates distinct effector functions in vivo. Th1 cells are predom-inantly involved in immune responses against intracellular patho-gens and are associated with autoimmune disease. Th2 cells are ofimportance in the defense against extracellular pathogens and areimplicated in atopy and allergic diseases (3–5).
Both Th1 and Th2 cells are derived from a common naive precur-sor (4–6). Signaling pathways initiated by cytokines play a dominantrole in driving the differentiation of activated naive CD41 T cells intoeither effector phenotype (2, 7). For instance, IL-12 induces thedifferentiation of naive Th cells to the Th1 effector phenotype (8–11)by activation of the transcription factor Stat4 (12–14). On the otherhand, Th2 differentiation is mediated by Stat6 activation through IL-4receptor engagement (15–21). In response to chronic antigenic stim-
ulation in vivo, progressive polarization of the cytokine responsesultimately leads to the commitment of Th cells to mutually exclusiveTh phenotypes, which are thought to be maintained independently ofextrinsic factors (22, 23).
Stat6 induces the expression of the transcription factorsGATA-3 and c-Maf (24), which have been shown to be selectivelyexpressed in a Th2-specific fashion (25–27). Using Stat6-deficientcells it has been shown that, although IL-4 and Stat6 signaling mayinitially direct Th2 development, GATA-3 and c-Maf are capableof inducing the development of stabile Th2 commitment, indepen-dent of Stat6 (28). In vitro differentiation into Th1 cells induceschromatin remodeling of theIFN-g locus and, conversely, the dif-ferentiation into Th2 cells induces remodeling of theIL-4/IL-5/IL-13 locus (29, 30). Recently, GATA-3 has been shown to play aninstructive role in directing Th2 differentiation (31).
During early T cell development,GATA-3gene expression isrequired for the development of the earliest T cell progenitors (32–34). GATA-3 levels are low during the two phases ofTCRgenerearrangement, but are high in the fraction of rapidly proliferatingcells that insulates these two periods ofTCR rearrangement (33).GATA-3 expression remains high in CD41 thymocytes, but pro-gressively declines in CD81 thymocytes (see accompanying pa-per). GATA-3 is detected in naive CD41 T cells and expressionlevels increase substantially during Th2 differentiation (26, 27).GATA-3 expression has been shown to be indispensable for Th2development and is down-regulated in response to IL-12-mediatedStat4 activation (27, 35). GATA-3 strongly transactivates theIL-5promoter, but appears to have only limited effects onIL-4 genetranscription (27, 36, 37). Retroviral introduction of GATA-3 dur-ing in vitro Th1 differentiation of naive CD41 T cells resulted inan inhibition of IFN-g production, independently of IL-4 (35, 38),
Departments of *Immunology,†Cell Biology and Genetics, and‡Pulmonary Medi-cine, Faculty of Medicine, Erasmus University Rotterdam, Rotterdam, The Nether-lands; and§Institute of Molecular and Cell Biology, University of Tartu, Tartu,Estonia
Received for publication April 12, 2000. Accepted for publication May 8, 2001.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was partly supported by Grant 3069 from the Estonian Science Founda-tion (to A.K.) and by the Royal Academy of Arts and Sciences (to R.W.H.).2 Address correspondence and reprint requests to Dr. Rudolf W. Hendriks, Depart-ment of Immunology, Faculty of Medicine, Room Ee853, Erasmus University Rot-terdam, Dr. Molewaterplein 50, P.O. Box 1738, 3000 DR Rotterdam, The Nether-lands. E-mail address: [email protected]
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00

and a down-regulation of IL-12Rb2 (35), which normally accom-panies Th2 differentiation (39).
The manipulation of Stat6 and GATA-3 expression in Th1 andTh2 polarization cultures of wild-type or specific cytokine-defi-cient cells in vitro have added significantly to our understanding ofthe molecular basis of Th1/Th2 differentiation. However, limiteddata are available on the role of GATA-3 during immune re-sponses in animal models, partly because the embryonic lethalityof GATA-3deficiency in mice precluded in vivo studies (40). Anal-ysis of transgenic mice with T cell-specific expression of a dom-inant-negative mutant of GATA-3 indicated that inhibition ofGATA-3 activity reduced the key features of asthma, includingTh2 cytokine levels, eosinophilia, and IgE production (41).
To study the function of GATA-3 during T cell differentiation,we generated transgenic mice in which the expression ofGATA-3is under the control of the humanCD2 locus control region (seeaccompanying paper). In these mice, the enforcedGATA-3expres-sion induced the development of thymic lymphomas ofCD41CD81/low T cells and inhibited the maturation of CD8 sin-gle-positive (SP)3 cells in the thymus. Within the CD8 SP popu-lation in the thymus, apoptosis was increased and the fraction ofmature CD69lowHSAlow cells was significantly reduced. The num-bers of peripheral CD81 T cells were;50% of normal. Theseobservations supported a role for GATA-3 in the regulation ofCD4/CD8 lineage commitment (see accompanying paper).
To investigate how the enforced expression ofGATA-3affectedTh cell differentiation, we analyzed T cell-mediated immune re-sponses in vivo inCD2-GATA3-transgenic mice. The observationsof a selective deficiency of Ag-specific IgG2a production and se-verely reduced delayed-type hypersensitivity (DTH) responses inthese mice showed that enforcedGATA-3expression inhibited thedifferentiation of Th1 cells in vivo. Furthermore, the expression ofsurface markers specific for Ag-experienced Th2 cells, along withthe increased functional capacity to secrete the Th2 cytokines, in-dicated that transgenic GATA-3 expression induced Th2 commit-ment and pointed at a role for GATA-3 in Th2 memory formation.
Materials and MethodsMice
TheCD2-GATA3mice are described in the accompanying paper and werecrossed on a uniform FVB background. To determine the genotype of themice, tail DNA was analyzed by Southern blotting as described in theaccompanying paper.
Flow cytometric analyses
The preparation of single-cell suspensions, mAb incubations and three- orfour-color cytometry have been described previously (42). The followingmAb were purchased from BD PharMingen (San Diego, CA): FITCconju-gated anti-CD3e, PE-conjugated anti-CD4 (L3T4), anti-CD24/heat-stable Ag,anti-CD25 (clone 3C7), anti-CD62L and anti-CD69, CyChrome-conjugatedanti-CD4 (L3T4), anti-CD8 and anti-CD44, biotinylated anti-CD4 (L3T4), andanti-CD8, APC-labeled anti-CD3e and anti-CD4. Anti-CD45RB (MB23G2)was a purified mAb conjugated to biotin according to standard procedures.Secondary Abs used were PE-, TriColor-, or APC-conjugated streptavidin(Caltag, Burlingame, CA). The Th2-selective surface marker T1/ST2 (3E10,rat IgG1, kindly provided by A. J. Coyle, Millenium Pharmaceuticals,Cam-bridge, MA) was detected by secondary goat anti-rat IgG-PE (Jackson Immuno-Research Laboratories, West Grove, PA) (43).
For intracellular detection of GATA-3 protein, cells were fixed and per-meabilized using paraformaldehyde and saponin as described previously(44) and subsequently incubated with the Hg-3-31 anti-GATA-3 mAb(Santa Cruz Biotechnology, Santa Cruz, CA) and FITC-labeled anti-mouseIgG1 (BD PharMingen) as a second step. For three-color analysis, 0.5–13105 events were scored using a FACScan analyzer (BD Biosciences,
Sunnyvale, CA). For four-color analysis, 105–2 3 106 events were scoredusing a FACSCalibur dual laser instrument (BD Biosciences).
For intracellular detection of cytokines, cells were stimulated for 40 h inthe presence of mAb to CD28 (37.51; 5mg/ml) in 96-well plates (106
cells/well) precoated with mAb to CD3e (145 2C11; 10mg/ml in PBS).Subsequently, cells were stimulated by adding PMA (50 ng/ml; Sigma, St.Louis, MO) and calcium ionophore (500 ng/ml; Sigma) for 5 h. For the last3 h of the culture, brefeldin A (10mg/ml; Sigma) was added to the cells.Finally, cells were harvested and stained with CyChrome-labeled mAb toCD4 or CD8 (BD PharMingen). Cells were fixed using 2% paraformalde-hyde and stored up to 3 days at 4°C. Intracellular cytokine staining wasperformed using PE-labeled mAb to IL-2, IL-4, and IL-10 (BD PharMin-gen) and APC-labeled mAb to IL-2, IL-5, and IFN-g (BD PharMingen)according to the manufacturer’s instructions.
Serum Ig detection and in vivo immunizations
Total serum Ig levels were determined by subclass-specific sandwichELISA as described previously (45). Immunizations were done i.p. with100 mg trinitrophenol-keyhole limpet hemocyanin (TNP-KLH) precipi-tated on alum. Serum levels of TNP-specific Ig subclasses were determinedby ELISA, using TNP-specific standards (IgG1, IgG2a, and IgG2b) orTNP-specific reference serum samples (IgM and IgG3), as described else-where (46).
Purification of CD41 T cells and in vitro cultures
Single-cell suspensions from spleen were incubated with biotinylated mAbto CD8 (YTS-169), CD11b/Mac-1 (M1/70), CD40 (FGK-45.5), B220(RA3-6B2), and IgM (M41), followed by streptavidin-conjugated mi-crobeads (Miltenyi Biotec, Bergisch Gladbach, Germany). Using a Vario-MACS, CD41 T cells were purified according to the manufacturer’s in-struction to purity.95%. The CD41 T cells were cultured for up to 5 daysin the presence of IL-2 (50 U/ml) on 96-well plates precoated with 10mg/ml anti-CD3 (145 2C11) mAb.
Purified CD41 T cells were polarized into Th1 and Th2 effector cells ina total volume of 200ml for 4 days in the presence of 5mg/ml anti-CD28(37.51) and 50 IU/ml IL-2 on 96-well plates, which were precoated with 10mg/ml anti-CD3e (145 2C11) (33). Th1-polarizing cultures included 5ng/ml rIL-12 (R&D Systems, Minneapolis, MN) and 10mg/ml neutralizingmAbs to IL-4 (11B11). Th2-polarized cells were cultured in the presenceof 10–50 ng/ml rIL-4 and 10mg/ml neutralizing mAbs to IFN-g(XMG1.2). After 4 days of culture, the cells were thoroughly washed andtransferred to new anti-CD3-coated 96-well plates and cultured in the pres-ence of IL-2, without addition of further cytokines or neutralizing Abs.
To measure DNA synthesis during T cell cultures, cells were pulsedwith [3H]thymidine for ;16 h, harvested, and counted using standardmethods. Cytokine levels in culture supernatants were determined byELISA using the Opteia kit for IL-4, IL-5, IL-10, and IFN-g (BD PharM-ingen) according to the manufacturer’s instructions. Expression ofGATA-3 protein was evaluated using a Western blotting procedure as de-scribed in the accompanying paper.
DTH responses
DTH responses were performed essentially as described by Cua et al. (47).In short, mice were immunized i.p. with 100mg KLH in 250 ml PBS andon day 6 they were challenged with 150mg KLH in 25 ml PBS in the lefthind footpad. The right hind footpad was in injected with a vehicle control(25 ml PBS). Responses were quantified 24 and 48 h after the challenge bymeasuring the difference in footpad thickness between the KLH- and thePBS-injected footpads.
Immunohistochemistry
Tissue samples were embedded in OCT compound and frozen 5-mm cry-ostat sections were acetone fixed and single labelings were performed us-ing standard procedures (45). The mAbs biotinylated anti-IL-4 (11B11)and alkaline phosphatase-conjugated anti-IL-5 (TRFK5) were purified hy-bridoma supernatants and conjugated according to standard procedures.Biotinylated anti-IL-10 (SXC1) was purchased from BD PharMingen.
ResultsIncreased expression of Ag-experienced T cell surface markersin CD2-GATA3-transgenic mice
In two independentCD2-GATA-3-transgenic lines expression ofhemagglutinin (HA)-tagged GATA-3 was under the control of theCD2 locus control region. Due to the presence of this transgene,
3 Abbreviations used in this paper: SP, single positive; DTH, delayed-type hypersen-sitivity; KLH, keyhole limpet hemocyanin; TNP, trinitrophenol; HA, hemagglutinin.
725The Journal of Immunology

GATA-3 expression was significantly enhanced in the thymus, es-pecially in the DP fraction. In these mice, the expression of trans-genic GATA-3 in peripheral lymphoid organs was low (see ac-companying paper). In the experiments described below, we didnot detect any differences between the two independent lines TgAand TgB.
To determine the effect of enforcedGATA-3expression on thedevelopment of the peripheral T cell compartments, we investi-gated the sizes of the CD41 and CD81 populations inCD2-GATA3-transgenic mice and their nontransgenic littermates at dif-ferent ages (Fig. 1A). In spleen and mesenteric lymph nodes, thenumbers of CD41 T cells in CD2-GATA3mice were either in-creased (3 wk of age) or within normal ranges (12 wk of age). Incontrast, the CD81 T cell populations were consistently reduced innumber, to;50% of normal, irrespective of the age analyzed (Fig.1A). Despite the observed increased apoptosis in the fraction ofmature SP cells in the thymus (see accompanying paper), theCD2-GATA3-transgenic mice were not found to be lymphopenic at anyof the ages analyzed.
Because GATA-3 is involved in the stabilization of the Th2phenotype and the maintenance of Th2 cytokine expression, whichare important features of Th memory, we wanted to assess whetherthe enforced expression of GATA-3 had an effect on the develop-ment of memory T cells. Ag activation induces the expression ofCD44 and decreases CD45RB and L-selectin (CD62L) expressionon the cell surface of T cells. As this profile of cell surface markerexpression is maintained, even after cells have reverted to a qui-escent state, it can be used to define Ag-experienced T cells (48).As shown in Fig. 1,B andC, in CD2-GATA3-transgenic mice boththe CD41 and the CD81 T cell populations contained increasedproportions of CD44high, CD45RBlow, and CD62Llow cells. This
was found both in spleen and in lymph nodes, and pointed to thepresence of elevated numbers of either recently activated or Ag-experienced T cells (48). The expression levels of CD69 and IL-2Ra (CD25), which are markers of recently activated T cells (49),were low in CD2-GATA3 mice and nontransgenic littermates(shown for CD25 in Fig. 1,A andB), arguing against the presenceof major fractions of recently activated T cells inCD2-GATA3mice. It has been shown that the proportions of T cells with anAg-experienced phenotype increases with age, probably as a resultof progressive Ag experience (50, 51). When we compared CD44and CD62L expression at two different ages, 3 and 7 mo, we in-deed found that the conversion to the CD44highCD62Llow Ag-ex-perienced T cell phenotype increased with age, both inCD2-GATA3-transgenic mice and in control mice (shown for CD62L inFig. 1C).
Taken together, these results indicated that in theCD2-GATA3mice both the CD41 and the CD81 T cell population containedincreased proportions of cells with an Ag-experienced phenotype.
Enforced expression of GATA-3 results in increased numbers ofT1/ST2-positive CD41 T cells in the periphery
The IL-1R family member T1/ST2 is preferentially expressed onthe surface of murine Th2 cells (43, 52–54). It was recently shownthat CD41 cells become T1/ST2 positive after repeated antigenicstimulation under Th2-polarizing conditions and that Th2 cytokineproduction precedes T1/ST2 expression (55). Therefore, T1/ST2expression appears to be a late event in Th2 commitment.
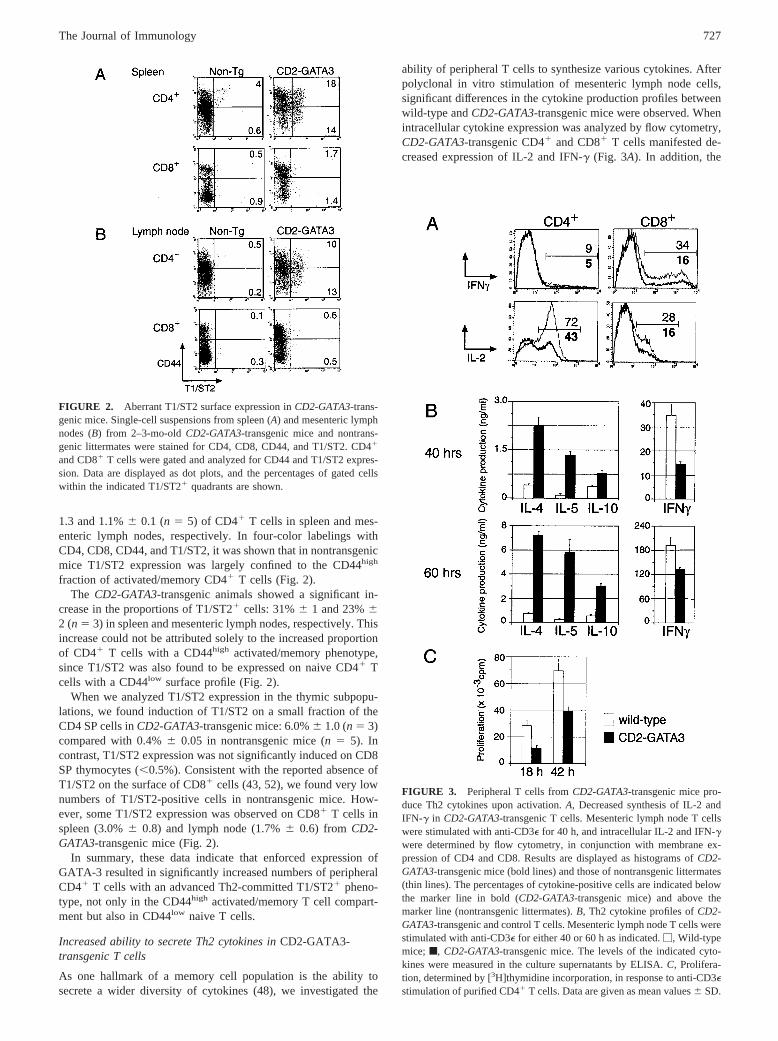
To investigate whether CD41 T cells in CD2-GATA3mice ex-hibited preferential Th2 polarization and advanced Th2 commit-ment in vivo, we evaluated surface expression of T1/ST2 (Fig. 2).In nontransgenic controls, we found T1/ST2 expression on 5.1%6
FIGURE 1. EnforcedGATA-3expression is associated with increased numbers of T cells with a memory surface phenotype.A, Single-cell suspensionsfrom spleen and mesenteric lymph nodes from 3- and 12-wk-oldCD2-GATA3-transgenic mice and nontransgenic littermates were analyzed for CD4 andCD8 expression. Surface CD4/CD8 profiles are shown as dot plots in which the percentages of total lymphocytes within the indicated CD41 and CD81
gates are given.B, Single-cell suspensions from spleen and mesenteric lymph nodes from 12-wk-oldCD2-GATA3-transgenic mice and nontransgeniclittermates were stained for CD4 and CD8, along with CD44, CD45RB, or CD25. Total CD41 and CD81 T cell populations were analyzed for the indicatedmarkers. Results are displayed as histograms ofCD2-GATA3-transgenic mice (bold lines) and those of nontransgenic littermates (thin lines). The per-centages of activated/Ag-experienced cells (CD44high, CD45RBlow, or CD251) are indicated above the marker line in bold (CD2-GATA3-transgenic mice)and below the marker line (nontransgenic littermates). Data shown are representative of at least six mice examined in each group.C, The numbers ofactivated/Ag-experienced (CD62Llow) cells inCD2-GATA3-transgenic and control mice at two different ages, 12 and 30 wk. The percentages are indicatedas inB.
726 GATA-3 INHIBITS Th1 RESPONSES IN VIVO
1.3 and 1.1%6 0.1 (n 5 5) of CD41 T cells in spleen and mes-enteric lymph nodes, respectively. In four-color labelings withCD4, CD8, CD44, and T1/ST2, it was shown that in nontransgenicmice T1/ST2 expression was largely confined to the CD44high
fraction of activated/memory CD41 T cells (Fig. 2).The CD2-GATA3-transgenic animals showed a significant in-
crease in the proportions of T1/ST21 cells: 31%6 1 and 23%62 (n 5 3) in spleen and mesenteric lymph nodes, respectively. Thisincrease could not be attributed solely to the increased proportionof CD41 T cells with a CD44high activated/memory phenotype,since T1/ST2 was also found to be expressed on naive CD41 Tcells with a CD44low surface profile (Fig. 2).
When we analyzed T1/ST2 expression in the thymic subpopu-lations, we found induction of T1/ST2 on a small fraction of theCD4 SP cells inCD2-GATA3-transgenic mice: 6.0%6 1.0 (n 5 3)compared with 0.4%6 0.05 in nontransgenic mice (n 5 5). Incontrast, T1/ST2 expression was not significantly induced on CD8SP thymocytes (,0.5%). Consistent with the reported absence ofT1/ST2 on the surface of CD81 cells (43, 52), we found very lownumbers of T1/ST2-positive cells in nontransgenic mice. How-ever, some T1/ST2 expression was observed on CD81 T cells inspleen (3.0%6 0.8) and lymph node (1.7%6 0.6) from CD2-GATA3-transgenic mice (Fig. 2).
In summary, these data indicate that enforced expression ofGATA-3 resulted in significantly increased numbers of peripheralCD41 T cells with an advanced Th2-committed T1/ST21 pheno-type, not only in the CD44high activated/memory T cell compart-ment but also in CD44low naive T cells.
Increased ability to secrete Th2 cytokines inCD2-GATA3-transgenic T cells
As one hallmark of a memory cell population is the ability tosecrete a wider diversity of cytokines (48), we investigated the
ability of peripheral T cells to synthesize various cytokines. Afterpolyclonal in vitro stimulation of mesenteric lymph node cells,significant differences in the cytokine production profiles betweenwild-type andCD2-GATA3-transgenic mice were observed. Whenintracellular cytokine expression was analyzed by flow cytometry,CD2-GATA3-transgenic CD41 and CD81 T cells manifested de-creased expression of IL-2 and IFN-g (Fig. 3A). In addition, the
FIGURE 2. Aberrant T1/ST2 surface expression inCD2-GATA3-trans-genic mice. Single-cell suspensions from spleen (A) and mesenteric lymphnodes (B) from 2–3-mo-oldCD2-GATA3-transgenic mice and nontrans-genic littermates were stained for CD4, CD8, CD44, and T1/ST2. CD41
and CD81 T cells were gated and analyzed for CD44 and T1/ST2 expres-sion. Data are displayed as dot plots, and the percentages of gated cellswithin the indicated T1/ST21 quadrants are shown.
FIGURE 3. Peripheral T cells fromCD2-GATA3-transgenic mice pro-duce Th2 cytokines upon activation.A, Decreased synthesis of IL-2 andIFN-g in CD2-GATA3-transgenic T cells. Mesenteric lymph node T cellswere stimulated with anti-CD3e for 40 h, and intracellular IL-2 and IFN-gwere determined by flow cytometry, in conjunction with membrane ex-pression of CD4 and CD8. Results are displayed as histograms ofCD2-GATA3-transgenic mice (bold lines) and those of nontransgenic littermates(thin lines). The percentages of cytokine-positive cells are indicated belowthe marker line in bold (CD2-GATA3-transgenic mice) and above themarker line (nontransgenic littermates).B, Th2 cytokine profiles ofCD2-GATA3-transgenic and control T cells. Mesenteric lymph node T cells werestimulated with anti-CD3e for either 40 or 60 h as indicated.M, Wild-typemice; f, CD2-GATA3-transgenic mice. The levels of the indicated cyto-kines were measured in the culture supernatants by ELISA.C, Prolifera-tion, determined by [3H]thymidine incorporation, in response to anti-CD3estimulation of purified CD41 T cells. Data are given as mean values6 SD.
727The Journal of Immunology
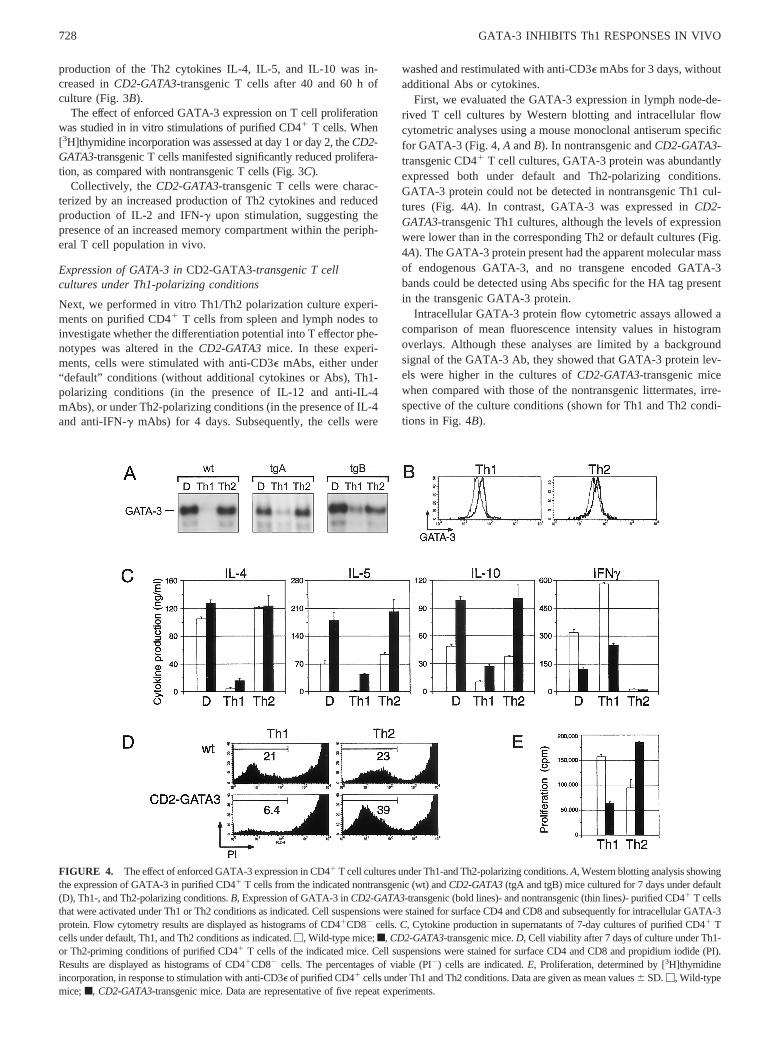
production of the Th2 cytokines IL-4, IL-5, and IL-10 was in-creased inCD2-GATA3-transgenic T cells after 40 and 60 h ofculture (Fig. 3B).
The effect of enforced GATA-3 expression on T cell proliferationwas studied in in vitro stimulations of purified CD41 T cells. When[3H]thymidine incorporation was assessed at day 1 or day 2, theCD2-GATA3-transgenic T cells manifested significantly reduced prolifera-tion, as compared with nontransgenic T cells (Fig. 3C).
Collectively, theCD2-GATA3-transgenic T cells were charac-terized by an increased production of Th2 cytokines and reducedproduction of IL-2 and IFN-g upon stimulation, suggesting thepresence of an increased memory compartment within the periph-eral T cell population in vivo.
Expression of GATA-3 inCD2-GATA3-transgenic T cellcultures under Th1-polarizing conditions
Next, we performed in vitro Th1/Th2 polarization culture experi-ments on purified CD41 T cells from spleen and lymph nodes toinvestigate whether the differentiation potential into T effector phe-notypes was altered in theCD2-GATA3mice. In these experi-ments, cells were stimulated with anti-CD3e mAbs, either under“default” conditions (without additional cytokines or Abs), Th1-polarizing conditions (in the presence of IL-12 and anti-IL-4mAbs), or under Th2-polarizing conditions (in the presence of IL-4and anti-IFN-g mAbs) for 4 days. Subsequently, the cells were
washed and restimulated with anti-CD3e mAbs for 3 days, withoutadditional Abs or cytokines.
First, we evaluated the GATA-3 expression in lymph node-de-rived T cell cultures by Western blotting and intracellular flowcytometric analyses using a mouse monoclonal antiserum specificfor GATA-3 (Fig. 4,A andB). In nontransgenic andCD2-GATA3-transgenic CD41 T cell cultures, GATA-3 protein was abundantlyexpressed both under default and Th2-polarizing conditions.GATA-3 protein could not be detected in nontransgenic Th1 cul-tures (Fig. 4A). In contrast, GATA-3 was expressed inCD2-GATA3-transgenic Th1 cultures, although the levels of expressionwere lower than in the corresponding Th2 or default cultures (Fig.4A). The GATA-3 protein present had the apparent molecular massof endogenous GATA-3, and no transgene encoded GATA-3bands could be detected using Abs specific for the HA tag presentin the transgenic GATA-3 protein.
Intracellular GATA-3 protein flow cytometric assays allowed acomparison of mean fluorescence intensity values in histogramoverlays. Although these analyses are limited by a backgroundsignal of the GATA-3 Ab, they showed that GATA-3 protein lev-els were higher in the cultures ofCD2-GATA3-transgenic micewhen compared with those of the nontransgenic littermates, irre-spective of the culture conditions (shown for Th1 and Th2 condi-tions in Fig. 4B).
FIGURE 4. The effect of enforced GATA-3 expression in CD41 T cell cultures under Th1-and Th2-polarizing conditions.A,Western blotting analysis showingthe expression of GATA-3 in purified CD41 T cells from the indicated nontransgenic (wt) andCD2-GATA3(tgA and tgB) mice cultured for 7 days under default(D), Th1-, and Th2-polarizing conditions.B,Expression of GATA-3 inCD2-GATA3-transgenic (bold lines)- and nontransgenic (thin lines)- purified CD41 T cellsthat were activated under Th1 or Th2 conditions as indicated. Cell suspensions were stained for surface CD4 and CD8 and subsequently for intracellular GATA-3protein. Flow cytometry results are displayed as histograms of CD41CD82 cells.C, Cytokine production in supernatants of 7-day cultures of purified CD41 Tcells under default, Th1, and Th2 conditions as indicated.M, Wild-type mice;f, CD2-GATA3-transgenic mice.D, Cell viability after 7 days of culture under Th1-or Th2-priming conditions of purified CD41 T cells of the indicated mice. Cell suspensions were stained for surface CD4 and CD8 and propidium iodide (PI).Results are displayed as histograms of CD41CD82 cells. The percentages of viable (PI2) cells are indicated.E, Proliferation, determined by [3H]thymidineincorporation, in response to stimulation with anti-CD3e of purified CD41 cells under Th1 and Th2 conditions. Data are given as mean values6 SD.M, Wild-typemice;f, CD2-GATA3-transgenic mice. Data are representative of five repeat experiments.
728 GATA-3 INHIBITS Th1 RESPONSES IN VIVO

Therefore, we conclude that due to the presence of theCD2-GATA3 transgene, GATA-3 protein expression is maintained inCD41 T cells that are cultured under Th1-polarizing conditions.
The Th1/Th2 differentiation potential ofCD2-GATA3-transgenicT cells in vitro
Cytokine production was evaluated in the Th1/Th2-polarizedCD41 T cell cultures from lymph nodes or spleen.CD2-GATA3-transgenic CD41 T cells produced normal amounts of IL-4, buthigher levels of IL-5 and IL-10 in the default or Th2-polarizedcultures when compared with nontransgenic CD41 T cells (Fig.4C). In addition, whenCD2-GATA3-transgenic CD41 T cells werecultured under Th1-polarizing conditions, they produced signifi-cantly increased amounts of IL-4, IL-5, and IL-10. Irrespective ofthe culture conditions, the production of IFN-g was significantlyreduced in theCD2-GATA3-transgenic CD41 T cells when com-pared with those from nontransgenic littermates (Fig. 4C).
The reduced IFN-g production by CD2-GATA3-transgenicCD41 T cells cultured under Th1-polarizing conditions could ei-ther result from an inhibitory effect of GATA-3 on the differenti-ation of naive cells into Th1 cells, or by an inhibition of the amountof IFN-g produced by differentiated Th1 effector cells. To distin-guish between these possibilities, we assessed cell viability andproliferation in the T cell cultures. When analyzed by flow cytom-etry using propidium iodide, CD41 T cells from CD2-GATA3-transgenic mice showed increased cell death under Th1 cultureconditions at day 7, as compared with nontransgenic littermates(Fig. 4D). When [3H]thymidine incorporation was assessed at day7, we observed a specific inhibitory effect of enforced GATA-3expression on cell proliferation in Th1 cultures (Fig. 4E). By con-trast, the presence of theCD2-GATA3transgene enhanced viabilityand proliferation of CD41 T cells in the Th2 cultures (Fig. 4,DandE).
Collectively, these observations demonstrate that although thepresence of theCD2-GATA3transgene inhibited the proliferationof Th cells under Th1-polarizing conditions, considerable produc-tion of Th2 cytokines was still present. Furthermore, the enforcedGATA-3 expression significantly supported proliferation and dif-ferentiation of Th2 effector cells in a Th2 environment.
GATA-3-expressing lymphoblastoid tumors express Th2cytokines
The coordinate expression of IL-4, IL-5. and IL-13 is thought to beunder the direct control of GATA-3, as GATA-3 specifically in-teracts with an intergenic DNase I hypersensitivity site in the Th2cytokine locus that contains theIL-4/IL-5/IL-13 gene cluster (30).However, the mechanism by which GATA-3 would regulate IL-10expression is unknown. The rapid production of IL-10 after anti-CD3 stimulation in vitro (Fig. 3C) would suggest that GATA-3 isdirectly involved in the regulation ofIL-10 gene expression. Tofurther address this question, we examined lymphoblastoid tumorsamples fromCD2-GATA3-transgenic mice. At the age of 9 mo,;50% of these mice developed thymic lymphomas that wereCD41CD81/low and expressed high levels of GATA-3 (see ac-companying paper). Immunohistochemical analyses of thymic tu-mor tissues showed that most of the tumors contained areas wherethe lymphoblastoid cells had lost expression of CD8 and some-times also CD4 (data not shown). Particularly in such areas, veryhigh expression of the Th2 cytokines IL-4, IL-5, or IL-10 wasfound (Fig. 5,A–C). Moreover, when tumor cells were cultured inthe presence of anti-CD3e, an extremely high production of Th2cytokines, including IL-10, was observed (an example is shown inFig. 5D).
These findings show that high expression of GATA-3 in lym-phoblastoid tumor cells is associated with high level production ofTh2 cytokine production, suggesting that like theIL4/IL-5/IL-13locus, also theIL-10 gene may be a direct target of GATA-3.
Enforced GATA-3 expression inhibits switching to IgG2a in anAg-specific immune response
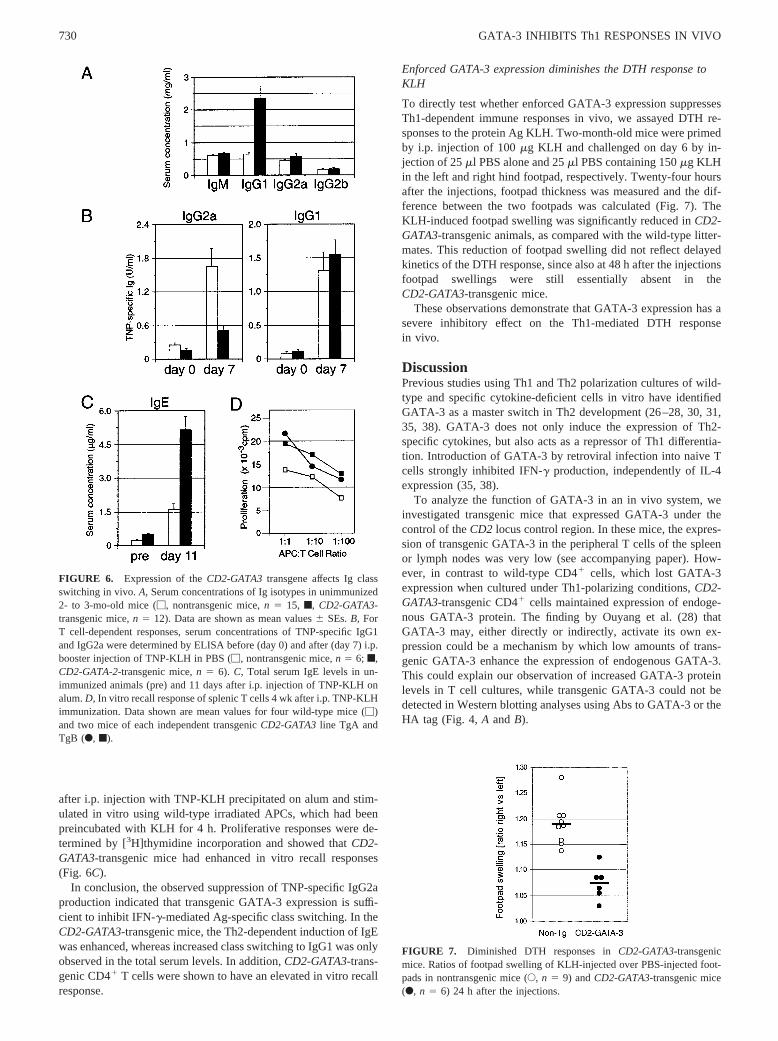
Serum levels of individual Ig isotypes are generally dependent onthe Th1/Th2 balance. IL-4 primes mouse B lymphocytes forswitching to IgG1 and IgE, while IgG2a responses are induced byIFN-g (56). When total serum Ig levels were determined in 2–3-mo-old CD2-GATA3-transgenic mice and nontransgenic litter-mates by ELISA, a selective increase in IgG1 was found in theCD2-GATA3-transgenic animals (Fig. 6A). The levels of all otherisotypes, including IgE, were similar in the two groups of mice(Fig. 6, A andC, IgE preimmune values).
To analyze Ig class switching in a T cell-dependent response invivo, mice were immunized i.p. with 10mg TNP-KLH, which wasprecipitated on alum. After 2 mo a booster dose of 100mg TNP-KLH in PBS was given.CD2-GATA3-transgenic mice showed asignificantly decreased secondary TNP response for the IFNg-de-pendent isotype IgG2a on day 7 after the booster injection, asmeasured in a TNP-specific ELISA (Fig. 6B). The TNP-specificlevels of the other Ig isotypes elicited in this response were com-parable between transgenic animals and wild-type littermates(shown for IgG1 in Fig. 6B). Also the primary IgG2a response toTNP-KLH in vivo was specifically decreased in theCD2-GATA3-transgenic animals (to;15% of normal), as determined in theserum at day 7 and 14 after an i.p. immunization with 100mgTNP-KLH precipitated on alum (data not shown). The Th2-depen-dent induction of heavy chain class switch to IgE was determinedat day 11 after i.p. injection of 10mg TNP-KLH precipitated onalum. CD2-GATA3-transgenic mice manifested elevated total se-rum IgE levels in this response (Fig. 6C).
Next, we directly tested the ability ofCD2-GATA3-transgenic Tcells to respond to Ag after a previous i.p. immunization withTNP-KLH. CD41 T cells were purified from spleens of immu-nized CD2-GATA3-transgenic mice and control littermates 4 wk
FIGURE 5. Cytokine production by thymic lymphoblastoid tumor cells.Immunohistochemical analysis of three thymic lymphoblastoid tumor tis-sue samples fromCD2-GATA3-transgenic mice showing areas with cellsthat produce high levels of IL-4 (A), IL-5 (B), and IL-10 (C). D, Cytokineproduction in the supernatant of a 40-h culture of lymphoblastoid tumorcells in the presence of anti-CD3e. f, CD2-GATA3-transgenic lymph nodecells from a tumor-free mouse;o, lymphoblastoid tumor cells from alymph node.
729The Journal of Immunology
after i.p. injection with TNP-KLH precipitated on alum and stim-ulated in vitro using wild-type irradiated APCs, which had beenpreincubated with KLH for 4 h. Proliferative responses were de-termined by [3H]thymidine incorporation and showed thatCD2-GATA3-transgenic mice had enhanced in vitro recall responses(Fig. 6C).
In conclusion, the observed suppression of TNP-specific IgG2aproduction indicated that transgenic GATA-3 expression is suffi-cient to inhibit IFN-g-mediated Ag-specific class switching. In theCD2-GATA3-transgenic mice, the Th2-dependent induction of IgEwas enhanced, whereas increased class switching to IgG1 was onlyobserved in the total serum levels. In addition,CD2-GATA3-trans-genic CD41 T cells were shown to have an elevated in vitro recallresponse.
Enforced GATA-3 expression diminishes the DTH response toKLH
To directly test whether enforced GATA-3 expression suppressesTh1-dependent immune responses in vivo, we assayed DTH re-sponses to the protein Ag KLH. Two-month-old mice were primedby i.p. injection of 100mg KLH and challenged on day 6 by in-jection of 25ml PBS alone and 25ml PBS containing 150mg KLHin the left and right hind footpad, respectively. Twenty-four hoursafter the injections, footpad thickness was measured and the dif-ference between the two footpads was calculated (Fig. 7). TheKLH-induced footpad swelling was significantly reduced inCD2-GATA3-transgenic animals, as compared with the wild-type litter-mates. This reduction of footpad swelling did not reflect delayedkinetics of the DTH response, since also at 48 h after the injectionsfootpad swellings were still essentially absent in theCD2-GATA3-transgenic mice.
These observations demonstrate that GATA-3 expression has asevere inhibitory effect on the Th1-mediated DTH responsein vivo.
DiscussionPrevious studies using Th1 and Th2 polarization cultures of wild-type and specific cytokine-deficient cells in vitro have identifiedGATA-3 as a master switch in Th2 development (26–28, 30, 31,35, 38). GATA-3 does not only induce the expression of Th2-specific cytokines, but also acts as a repressor of Th1 differentia-tion. Introduction of GATA-3 by retroviral infection into naive Tcells strongly inhibited IFN-g production, independently of IL-4expression (35, 38).
To analyze the function of GATA-3 in an in vivo system, weinvestigated transgenic mice that expressed GATA-3 under thecontrol of theCD2 locus control region. In these mice, the expres-sion of transgenic GATA-3 in the peripheral T cells of the spleenor lymph nodes was very low (see accompanying paper). How-ever, in contrast to wild-type CD41 cells, which lost GATA-3expression when cultured under Th1-polarizing conditions,CD2-GATA3-transgenic CD41 cells maintained expression of endoge-nous GATA-3 protein. The finding by Ouyang et al. (28) thatGATA-3 may, either directly or indirectly, activate its own ex-pression could be a mechanism by which low amounts of trans-genic GATA-3 enhance the expression of endogenous GATA-3.This could explain our observation of increased GATA-3 proteinlevels in T cell cultures, while transgenic GATA-3 could not bedetected in Western blotting analyses using Abs to GATA-3 or theHA tag (Fig. 4,A andB).
FIGURE 6. Expression of theCD2-GATA3transgene affects Ig classswitching in vivo.A, Serum concentrations of Ig isotypes in unimmunized2- to 3-mo-old mice (M, nontransgenic mice,n 5 15, f, CD2-GATA3-transgenic mice,n 5 12). Data are shown as mean values6 SEs.B, ForT cell-dependent responses, serum concentrations of TNP-specific IgG1and IgG2a were determined by ELISA before (day 0) and after (day 7) i.p.booster injection of TNP-KLH in PBS (M, nontransgenic mice,n 5 6; f,CD2-GATA-2-transgenic mice,n 5 6). C, Total serum IgE levels in un-immunized animals (pre) and 11 days after i.p. injection of TNP-KLH onalum.D, In vitro recall response of splenic T cells 4 wk after i.p. TNP-KLHimmunization. Data shown are mean values for four wild-type mice (M)and two mice of each independent transgenicCD2-GATA3line TgA andTgB (F, f).
FIGURE 7. Diminished DTH responses inCD2-GATA3-transgenicmice. Ratios of footpad swelling of KLH-injected over PBS-injected foot-pads in nontransgenic mice (E, n 5 9) andCD2-GATA3-transgenic mice(F, n 5 6) 24 h after the injections.
730 GATA-3 INHIBITS Th1 RESPONSES IN VIVO

Our analyses of theCD2-GATA3-transgenic mice support thefindings that GATA-3 expression inhibits Th1 development. Theenforced GATA-3 expression inhibited Th1-mediated responses invivo, including Ag-specific IgG2a production and DTH responsesto protein Ag. In our Th1/Th2 polarization cultures, enforcedGATA-3 expression under Th1-inducing culture conditions re-sulted in a reduction of cell survival, proliferation, and IFN-g pro-duction. The additional findings of increased T1/ST2 expression inCD41 T cells and elevated total IgG1 serum levels suggest that thepresence of theCD2-GATA3gene drives T cells preferentially to-ward differentiation along the Th2 pathway. Therefore, we con-clude that GATA-3 plays a dual role in vivo in the differentiationof naive Th cells into Th2 cells, since it both represses Th1 dif-ferentiation and induced Th2 differentiation.
Various lines of evidence indicate that the enforced expressionof GATA-3 may enhance Th2 memory cell formation. First, inCD2-GATA3-transgenic mice, the peripheral T cell compartmentcontained a high proportion of cells with an Ag-experienced cellsurface profile, defined as CD44highCD45RBlowCD62Llow andnegative for CD25 and CD69. The ratio of naive vs memory phe-notype cells decreased with age, as normally seen in wild-typemice. The possibility that peripheral T cells obtained the Ag-ex-perienced surface phenotype because of a homeostatic prolifera-tion mechanism (57) in theCD2-GATA3-transgenic mice is un-likely, because these mice were not lymphopenic at any of the agesanalyzed. Second, inCD2-GATA3-transgenic mice the expressionof the Th2-specific T1/ST2 marker within the CD44high memoryTh cell population in spleen and lymph nodes was increased by afactor ;6 and ;20, respectively. T1/ST2 marker expression isassociated with advanced Th2 commitment, as it was shown to beexpressed only after repeated antigenic stimulation under Th2-po-larizing conditions in vitro, with delayed kinetics compared withthe kinetics of Th2 cytokines (55). Third,CD2-GATA3-transgenicT cells were rapidly induced to synthesize IL-4, IL-5, and IL-10 invitro, whereas production of IL-2 was low, which is typical formemory Th2 cells (48). Fourth, T cells fromCD2-GATA3miceexhibited an increased recall response to TNP-KLH Ag in vitro.Finally, the selective increase of the total levels of the IL-4-de-pendent isotype IgG1 in the serum would also be consistent withincreased Th2 memory formation.
It is presently not clear how GATA-3 would affect Th2 memoryformation. GATA-3 may regulate the cell fate decision of activatedCD41 T cells, by reducing activation-induced cell death, in favorof Th2 memory cell formation. Alternatively, GATA-3 may facil-itate the differentiation process of dividing effector T cells that arealready committed to the memory cell fate. A third possibility isthat GATA-3 would act as a survival factor for Th2 memory cells.This is not very likely, because survival alone does not appear tobe sufficient for memory cell formation, as was shown by the ab-sence of increased memory formation in Bcl-2-transgenic mice(58). Additional experiments are required to define GATA-3 tar-gets that are involved in Th2 memory cell formation.
One of the molecules involved in Th2 memory developmentmay be T1/ST2, as it is normally specifically expressed in the Th2lineage within the compartment of CD44high activated/memory Tcells (Fig. 2). Because cross-linking of T1/ST2 enhanced prolifer-ation of Th2 cells that were stimulated with suboptimal concen-trations of anti-CD3 mAb (55), it is possible that the increasedproliferation and cell survival ofCD2-GATA3-transgenic CD41 Tcells in the in vitro Th2 polarization cultures originates from in-creased T1/ST2 expression. The increased expression of T1/ST2 inCD2-GATA3-transgenic mice would argue for a direct regulationof T1/ST2transcription in T cells by GATA-3. The identificationof three GATA elements in the minimal GATA-responsiveT1/ST2
promoter in mast cells (59) would support this hypothesis of adirect regulation ofT1/ST2expression by GATA-3, independent ofTh2-specific cytokines. Therefore, we hypothesize that GATA-3 isnot only essential for instructive differentiation of naive Th cellsinto committed Th2 cells (26–28, 31, 35), but can also affect pro-liferation and survival of GATA-3-expressing CD41 T cellsthrough the induction of T1/ST2.
In conclusion, this study shows that enforced expression ofGATA-3 inhibits Th1 function and induces Th2 commitment invivo. Moreover, the increased expression of T1/ST2, the enhancedproduction of Th2 cytokines in response to T cell activation, andthe elevated serum levels of IgG1 inCD2-GATA3-transgenic miceargue for a role of GATA-3 in the formation of Th2 memory.
AcknowledgmentsWe thank Willem van Ewijk, Danielle Hijdra, John Mahabier,Sjaak Philipsen, and Vicotr de Vries for their assistance at several stages ofthe project and G. Z. Tau for critical reading of this manuscript. We thankA. J. Coyle for provision of the anti-T1/ST2 Ab.
References1. Mosmann, T. R., H. Cherwinski, M. W. Bond, M. A. Giedlin, and R. L. Coffman.
1986. Two types of murine helper T cell clone. I. Definition according to profilesof lymphokine activities and secreted proteins.J. Immunol. 136:2348.
2. Paul, W. E., and R. A. Seder. 1994. Lymphocyte responses and cytokines.Cell76:241.
3. Romagnani, S. 1997. The Th1/Th2 paradigm.Immunol. Today 18:263.4. O’Garra, A. 1998. Cytokines induce the development of functionally heteroge-
neous T helper cell subsets.Immunity 8:275.5. Abbas, A. K., K. M. Murphy, and A. Sher. 1996. Functional diversity of helper
T lymphocytes.Nature 383:787.6. Bluestone, J. A. 1995. New perspectives of CD28–B7-mediated T cell costimu-
lation. Immunity 2:555.7. Constant, S. L., and K. Bottomly. 1997. Induction of Th1 and Th2 CD41 T cell
responses: the alternative approaches.Annu. Rev. Immunol. 15:297.8. Hsieh, C. S., S. E. Macatonia, C. S. Tripp, S. F. Wolf, A. O’Garra, and
K. M. Murphy. 1993. Development of TH1 CD41 T cells through IL-12 pro-duced byListeria-induced macrophages.Science 260:547.
9. Manetti, R., P. Parronchi, M. G. Giudizi, M. P. Piccinni, E. Maggi, G. Trinchieri,and S. Romagnani. 1993. Natural killer cell stimulatory factor (interleukin 12(IL-12)) induces T helper type 1 (Th1)-specific immune responses and inhibitsthe development of IL-4-producing Th cells.J. Exp. Med. 177:1199.
10. Manetti, R., F. Gerosa, M. G. Giudizi, R. Biagiotti, P. Parronchi, M. P. Piccinni,S. Sampognaro, E. Maggi, S. Romagnani, G. Trinchieri, et al. 1994. Interleukin12 induces stable priming for interferong (IFN-g) production during differenti-ation of human T helper (Th) cells and transient IFN-g production in establishedTh2 cell clones.J. Exp. Med. 179:1273.
11. Seder, R. A., R. Gazzinelli, A. Sher, and W. E. Paul. 1993. Interleukin 12 actsdirectly on CD41 T cells to enhance priming for interferong production anddiminishes interleukin 4 inhibition of such priming.Proc. Natl. Acad. Sci. USA90:10188.
12. Szabo, S. J., N. G. Jacobson, A. S. Dighe, U. Gubler, and K. M. Murphy. 1995.Developmental commitment to the Th2 lineage by extinction of IL-12 signaling.Immunity 2:665.
13. Thierfelder, W. E., J. M. van Deursen, K. Yamamoto, R. A. Tripp, S. R. Sarawar,R. T. Carson, M. Y. Sangster, D. A. Vignali, P. C. Doherty, G. C. Grosveld, andJ. N. Ihle. 1996. Requirement for Stat4 in interleukin-12-mediated responses ofnatural killer and T cells.Nature 382:171.
14. Kaplan, M. H., Y. L. Sun, T. Hoey, and M. J. Grusby. 1996. Impaired IL-12responses and enhanced development of Th2 cells in Stat4-deficient mice.Nature382:174.
15. Le Gros, G., S. Z. Ben-Sasson, R. Seder, F. D. Finkelman, and W. E. Paul. 1990.Generation of interleukin 4 (IL-4)-producing cells in vivo and in vitro: IL-2 andIL-4 are required for in vitro generation of IL-4- producing cells.J. Exp. Med.172:921.
16. Swain, S. L., A. D. Weinberg, M. English, and G. Huston. 1990. IL-4 directs thedevelopment of Th2-like helper effectors.J. Immunol. 145:3796.
17. Seder, R. A., W. E. Paul, M. M. Davis, and B. Fazekas de St. Groth. 1992. Thepresence of interleukin 4 during in vitro priming determines the lymphokine-producing potential of CD41 T cells from T cell receptor transgenic mice.J. Exp.Med. 176:1091.
18. Hsieh, C. S., A. B. Heimberger, J. S. Gold, A. O’Garra, and K. M. Murphy. 1992.Differential regulation of T helper phenotype development by interleukins 4 and10 in anab T-cell-receptor transgenic system.Proc. Natl. Acad. Sci. USA 89:6065.
19. Takeda, K., T. Tanaka, W. Shi, M. Matsumoto, M. Minami, S. Kashiwamura,K. Nakanishi, N. Yoshida, T. Kishimoto, and S. Akira. 1996. Essential role ofStat6 in IL-4 signalling.Nature 380:627.
20. Shimoda, K., J. van Deursen, M. Y. Sangster, S. R. Sarawar, R. T. Carson,R. A. Tripp, C. Chu, F. W. Quelle, T. Nosaka, D. A. Vignali, et al. 1996. Lack
731The Journal of Immunology

of IL-4-induced Th2 response and IgE class switching in mice with disruptedStat6 gene.Nature 380:630.
21. Kaplan, M. H., U. Schindler, S. T. Smiley, and M. J. Grusby. 1996. Stat6 isrequired for mediating responses to IL-4 and for development of Th2 cells.Im-munity 4:313.
22. Murphy, E., K. Shibuya, N. Hosken, P. Openshaw, V. Maino, K. Davis,K. Murphy, and A. O’Garra. 1996. Reversibility of T helper 1 and 2 populationsis lost after long-term stimulation.J. Exp. Med. 183:901.
23. Huang, H., J. Hu-Li, H. Chen, S. Z. Ben-Sasson, and W. E. Paul. 1997. IL-4 andIL-13 production in differentiated T helper type 2 cells is not IL-4 dependent.J. Immunol. 159:3731.
24. Kurata, H., H. J. Lee, A. O’Garra, and N. Arai. 1999. Ectopic expression ofactivated Stat6 induces the expression of Th2-specific cytokines and transcriptionfactors in developing Th1 cells.Immunity 11:677.
25. Ho, I. C., D. Lo, and L. H. Glimcher. 1998. c-Maf promotes T helper cell type2 (Th2) and attenuates Th1 differentiation by both interleukin 4-dependent and-independent mechanisms.J. Exp. Med. 188:1859.
26. Zhang, D. H., L. Cohn, P. Ray, K. Bottomly, and A. Ray. 1997. Transcriptionfactor GATA-3 is differentially expressed in murine Th1 and Th2 cells and con-trols Th2-specific expression of the interleukin-5 gene.J. Biol. Chem. 272:21597.
27. Zheng, W., and R. A. Flavell. 1997. The transcription factor GATA-3 is neces-sary and sufficient for Th2 cytokine gene expression in CD4 T cells.Cell 89:587.
28. Ouyang, W., M. Lohning, Z. Gao, M. Assenmacher, S. Ranganath, A. Radbruch,and K. M. Murphy. 2000. Stat6-independent GATA-3 autoactivation directs IL-4-independent Th2 development and commitment.Immunity 12:27.
29. Agarwal, S., and A. Rao. 1998. Modulation of chromatin structure regulatescytokine gene expression during T cell differentiation.Immunity 9:765.
30. Takemoto, N., Y. Kamogawa, H. Jun Lee, H. Kurata, K. Arai, A. O’Garra,N. Arai, and S. Miyatake. 2000. Cutting edge: chromatin remodeling at the IL-4/IL-13 intergenic regulatory region for Th2-specific cytokine gene cluster.J. Im-munol. 165:6687.
31. Farrar, J. D., W. Ouyang, M. Lohning, M. Assenmacher, A. Radbruch,O. Kanagawa, and K. M. Murphy. 2001. An instructive component in T helpercell type 2 (Th2) development mediated by GATA-3.J. Exp. Med. 193:643.
32. Ting, C. N., M. C. Olson, K. P. Barton, and J. M. Leiden. 1996. Transcriptionfactor GATA-3 is required for development of the T-cell lineage.Nature 384:474.
33. Hendriks, R. W., M. C. Nawijn, J. D. Engel, H. van Doorninck, F. Grosveld, andA. Karis. 1999. Expression of the transcription factor GATA-3 is required for thedevelopment of the earliest T cell progenitors and correlates with stages of cel-lular proliferation in the thymus.Eur. J. Immunol. 29:1912.
34. Hattori, N., H. Kawamoto, S. Fujimoto, K. Kuno, and Y. Katsura. 1996. Involve-ment of transcription factors TCF-1 and GATA-3 in the initiation of the earlieststep of T cell development in the thymus.J. Exp. Med. 184:1137.
35. Ouyang, W., S. H. Ranganath, K. Weindel, D. Bhattacharya, T. L. Murphy,W. C. Sha, and K. M. Murphy. 1998. Inhibition of Th1 development mediated byGATA-3 through an IL-4-independent mechanism.Immunity 9:745.
36. Zhang, D. H., L. Yang, and A. Ray. 1998. Differential responsiveness of the IL-5and IL-4 genes to transcription factor GATA-3.J. Immunol. 161:3817.
37. Ranganath, S., W. Ouyang, D. Bhattarcharya, W. C. Sha, A. Grupe, G. Peltz, andK. M. Murphy. 1998. GATA-3-dependent enhancer activity in IL-4 gene regu-lation. J. Immunol. 161:3822.
38. Ferber, I. A., H. J. Lee, F. Zonin, V. Heath, A. Mui, N. Arai, and A. O’Garra.1999. GATA-3 significantly downregulates IFN-g production from developingTh1 cells in addition to inducing IL-4 and IL-5 levels.Clin. Immunol. 91:134.
39. Szabo, S. J., A. S. Dighe, U. Gubler, and K. M. Murphy. 1997. Regulation of theinterleukin (IL)-12Rb2 subunit expression in developing T helper 1 (Th1) andTh2 cells.J. Exp. Med. 185:817.
40. Pandolfi, P. P., M. E. Roth, A. Karis, M. W. Leonard, E. Dzierzak,F. G. Grosveld, J. D. Engel, and M. H. Lindenbaum. 1995. Targeted disruptionof theGATA3gene causes severe abnormalities in the nervous system and in fetalliver haematopoiesis.Nat. Genet. 11:40.
41. Zhang, D. H., L. Yang, L. Cohn, L. Parkyn, R. Homer, P. Ray, and A. Ray. 1999.Inhibition of allergic inflammation in a murine model of asthma by expression ofa dominant-negative mutant of GATA-3.Immunity 11:473.
42. Hendriks, R. W., M. F. de Bruijn, A. Maas, G. M. Dingjan, A. Karis, andF. Grosveld. 1996. Inactivation of Btk by insertion oflacZ reveals defects in Bcell development only past the pre-B cell stage.EMBO J. 15:4862.
43. Lohning, M., A. Stroehmann, A. J. Coyle, J. L. Grogan, S. Lin,J. C. Gutierrez-Ramos, D. Levinson, A. Radbruch, and T. Kamradt. 1998. T1/ST2is preferentially expressed on murine Th2 cells, independent of interleukin 4,interleukin 5, and interleukin 10, and important for Th2 effector function.Proc.Natl. Acad. Sci. USA 95:6930.
44. Maas, A., G. M. Dingjan, F. Grosveld, and R. W. Hendriks. 1999. Early arrest inB cell development in transgenic mice that express the E41K Bruton’s tyrosinekinase mutant under the control of the CD19 promoter region.J. Immunol. 162:6526.
45. Dingjan, G. M., A. Maas, M. C. Nawijn, L. Smit, J. S. Voerman, F. Grosveld, andR. W. Hendriks. 1998. Severe B cell deficiency and disrupted splenic architecturein transgenic mice expressing the E41K mutated form of Bruton’s tyrosine ki-nase.EMBO J. 17:5309.
46. Maas, A., G. M. Dingjan, H. F. Savelkoul, C. Kinnon, F. Grosveld, andR. W. Hendriks. 1997. The X-linked immunodeficiency defect in the mouse iscorrected by expression of human Bruton’s tyrosine kinase from a yeast artificialchromosome transgene.Eur. J. Immunol. 27:2180.
47. Cua, D. J., R. L. Coffman, and S. A. Stohlman. 1996. Exposure to T helper 2cytokines in vivo before encounter with antigen selects for T helper subsets viaalterations in antigen-presenting cell function.J. Immunol. 157:2830.
48. Dutton, R. W., L. M. Bradley, and S. L. Swain. 1998. T cell memory.Annu. Rev.Immunol. 16:201.
49. Malek, T. R., J. A. Schmidt, and E. M. Shevach. 1985. The murine IL 2 receptor.III. Cellular requirements for the induction of IL 2 receptor expression on T cellsubpopulations.J. Immunol. 134:2405.
50. Ernst, D. N., W. O. Weigle, D. J. Noonan, D. N. McQuitty, and M. V. Hobbs.1993. The age-associated increase in IFN-g synthesis by mouse CD81 T cellscorrelates with shifts in the frequencies of cell subsets defined by membraneCD44, CD45RB, 3G11, and MEL-14 expression.J. Immunol. 151:575.
51. Sprent, J., D. F. Tough, and S. Sun. 1997. Factors controlling the turnover of Tmemory cells.Immunol. Rev. 156:79.
52. Xu, D., W. L. Chan, B. P. Leung, F. Huang, R. Wheeler, D. Piedrafita,J. H. Robinson, and F. Y. Liew. 1998. Selective expression of a stable cell surfacemolecule on type 2 but not type 1 helper T cells.J. Exp. Med. 187:787.
53. Coyle, A. J., C. Lloyd, J. Tian, T. Nguyen, C. Erikkson, L. Wang, P. Ottoson,P. Persson, T. Delaney, S. Lehar, et al. 1999. Crucial role of the interleukin 1receptor family member T1/ST2 in T helper cell type 2-mediated lung mucosalimmune responses.J. Exp. Med. 190:895.
54. Townsend, M. J., P. G. Fallon, D. J. Matthews, H. E. Jolin, and A. N. McKenzie.2000. T1/ST2-deficient mice demonstrate the importance of T1/ST2 in develop-ing primary T helper cell type 2 responses.J. Exp. Med. 191:1069.
55. Meisel, C., K. Bonhagen, M. Lohning, A. J. Coyle, J. C. Gutierrez-Ramos,A. Radbruch, and T. Kamradt. 2001. Regulation and function of T1/ST2 expres-sion on CD41 t cells: induction of type 2 cytokine production by T1/ST2 cross-linking. J. Immunol. 166:3143.
56. Snapper, C. M., and W. E. Paul. 1987. Interferon-g and B cell stimulatory fac-tor-1 reciprocally regulate Ig isotype production.Science 236:944.
57. Goldrath, A. W., L. Y. Bogatzki, and M. J. Bevan. 2000. Naive T cells transientlyacquire a memory-like phenotype during homeostasis-driven proliferation.J. Exp. Med. 192:557.
58. Petschner, F., C. Zimmerman, A. Strasser, D. Grillot, G. Nunez, and H. Pircher.1998. Constitutive expression of Bcl-xL or Bcl-2 prevents peptide antigen-in-duced T cell deletion but does not influence T cell homeostasis after a viralinfection.Eur. J. Immunol. 28:560.
59. Gachter, T., D. R. Moritz, J. Gheyselinck, and R. Klemenz. 1998. GATA-depen-dent expression of the interleukin-1 receptor-related T1 gene in mast cells.Mol.Cell. Biol. 18:5320.
732 GATA-3 INHIBITS Th1 RESPONSES IN VIVO
Related Documents











