Endotoxin Induces Differential Regulation of mTOR-Dependent Signaling in Skeletal Muscle and Liver of Neonatal Pigs Scot R. Kimball 1 , Renán A. Orellana 2 , Pamela M.J. O'Connor 2 , Agus Suryawan 2 , Jill A. Bush 2 , Hanh V. Nguyen 2 , M. Carole Thivierge 2 , Leonard S. Jefferson 1 , and Teresa A. Davis 2 1 Department of Cellular and Molecular Physiology, The Pennsylvania State University College of Medicine, Hershey, PA 17033; 2 United States Department of Agriculture/Agricultural Research Service, Children’s Nutrition Research Center, Department of Pediatrics, Baylor College of Medicine, Houston, TX 77030 Short title: mTOR-Dependent Signaling in the Septic Neonate Correspondence and reprint requests: Scot R. Kimball, Ph.D. Department of Cellular and Molecular Physiology (H166) The Pennsylvania State University College of Medicine 500 University Drive Hershey, PA 17033 Telephone: 717-531-8970 FAX: 717-531-7667 Email: [email protected] Copyright (c) 2003 by the American Physiological Society. Articles in PresS. Am J Physiol Endocrinol Metab (May 28, 2003). 10.1152/ajpendo.00340.2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Endotoxin Induces Differential Regulation of mTOR-Dependent Signaling inSkeletal Muscle and Liver of Neonatal Pigs
Scot R. Kimball1, Renán A. Orellana2, Pamela M.J. O'Connor2, Agus Suryawan2, Jill A. Bush2,
Hanh V. Nguyen2, M. Carole Thivierge2, Leonard S. Jefferson1, and Teresa A. Davis2
1Department of Cellular and Molecular Physiology, The Pennsylvania State University College
of Medicine, Hershey, PA 17033; 2 United States Department of Agriculture/Agricultural
Research Service, Children’s Nutrition Research Center, Department of Pediatrics, Baylor
College of Medicine, Houston, TX 77030
Short title: mTOR-Dependent Signaling in the Septic Neonate
Correspondence and reprint requests:
Scot R. Kimball, Ph.D.
Department of Cellular and Molecular Physiology (H166)
The Pennsylvania State University College of Medicine
500 University Drive
Hershey, PA 17033
Telephone: 717-531-8970
FAX: 717-531-7667
Email: [email protected]
Copyright (c) 2003 by the American Physiological Society.
Articles in PresS. Am J Physiol Endocrinol Metab (May 28, 2003). 10.1152/ajpendo.00340.2002

2
ABSTRACT
In the present study, differential responses of regulatory proteins involved in translation initiation
in skeletal muscle and liver during sepsis were studied in neonatal pigs treated with
lipopolysaccharide (LPS). LPS did not alter eukaryotic initiation factor (eIF)2B activity in either
tissue. In contrast, binding of eIF4G to eIF4E to form the active mRNA binding complex was
repressed in muscle and enhanced in liver. Phosphorylation of eIF4E binding protein, 4E-BP1,
and ribosomal protein S6 kinase, S6K1, was reduced in muscle during sepsis, but increased in
liver. Finally, changes in 4E-BP1 and S6K1 phosphorylation were associated with altered
phosphorylation of the protein kinase, mTOR. Overall, the results suggest that translation
initiation in both skeletal muscle and liver is altered during neonatal sepsis by modulation of the
mRNA binding step through changes in mTOR activation. Moreover, the LPS-induced changes
in factors that regulate translation initiation are more profound than previously reported changes
in global rates of protein synthesis in the neonate. This finding suggests that the met-tRNAi,
rather than mRNA, binding step in translation initiation may play a more critical role in
maintaining protein synthesis rates in the neonate during sepsis.
Keywords: eukaryotic initiation factor eIF2B, eIF4G, eIF4E, lipopolysaccharide

3
INTRODUCTION
Previous studies (48, 49) have shown that in adult rodents, the function of three proteins
that play important regulatory roles in translation initiation, i.e. eukaryotic initiation factors
eIF2B and eIF4F and the ribosomal protein S6 kinase S6K1, are downregulated in skeletal
muscle of septic rats. eIF2B is a guanine nucleotide exchange factor whose substrate, eIF2,
mediates the binding of initiator methionyl-tRNAi (met-tRNAi) to the 40S ribosomal subunit
(reviewed in 16). Thus, eIF2B modulates the first step in translation initiation and changes in its
activity result in corresponding alterations in global rates of protein synthesis. Binding of
mRNA to the 40S ribosomal subunit is mediated by a heterotrimeric complex referred to as
eIF4F. The heterotrimeric complex referred to as eIF4F mediates the binding of mRNA to the
40S ribosomal subunit. The three proteins that comprise the eIF4F complex are eIF4A, an RNA
helicase, eIF4E, the protein that binds to the m7GTP cap structure at the 5’-end of the mRNA,
and eIF4G, a scaffolding protein that, in addition to eIF4A and eIF4E, also binds to the eIF3•40S
ribosomal subunit complex. Thus, mRNA binds to the 40S ribosomal subunit through the
association of eIF4E with eIF4G. The binding of eIF4E to eIF4G is reversible and regulated by
the association of eIF4E with the eIF4E binding proteins (e.g., 4E-BP1). eIF4E complexed with
4E-BP1 can bind mRNA, but cannot bind to eIF4G such that when the eIF4E•mRNA complex is
associated with 4E-BP1, the complex cannot bind to the 40S ribosomal subunit. The association
of eIF4E with 4E-BP1 is regulated by phosphorylation of 4E-BP1 whereby eIF4E will bind to
hypophosphorylated, but not hyperphosphorylated, 4E-BP1. Hyperphosphorylation of 4E-BP1
that occurs in response to treatment with growth factors or amino acids is mediated through a
signal transduction pathway involving a protein kinase referred to as the mammalian target of
rapamycin (mTOR; aka FRAP or RAFT) (13). mTOR also phosphorylates, and thereby
activates, the 70 kDa ribosomal protein S6 kinase, S6K1. Both phosphorylation events are,
under some circumstances, associated with changes in global rates of protein synthesis, but more
importantly they lead to a preferential increase in translation of mRNAs encoding particular

4
proteins, particularly those mRNAs containing extensive secondary structure in their 5’-
untranslated region (reviewed in 13) or those mRNAs containing a terminal oligopyrimidine
(TOP) tract adjacent to the 5’-cap structure (29).
A recent study (25) demonstrated that in adult rats, the LPS-induced repression of global
rates of protein synthesis observed in gastrocnemius muscle is associated with both a decrease in
eIF2B activity and decreased assembly of the active eIF4F complex. Such changes in translation
initiation factors are quantitatively similar to those observed in a longer term model of chronic
abdominal sepsis (48, 49) as well as in rats treated with tumor necrosis factor (TNF)-a (26).
Which of these steps, the met-tRNAi binding or the mRNA binding step, is rate controlling for
global rates of protein synthesis in muscle during sepsis is unknown. In contrast to muscle,
considerably less is known about the mechanism(s) involved in the increase in hepatic protein
synthesis that occurs during sepsis. However, it has been reported that during chronic abdominal
sepsis, phosphorylation of S6K1 is enhanced in liver (4). The signal transduction pathway(s)
that mediates the effects of LPS or TNF-a on eIF4F assembly and S6K1 phosphorylation have
not been identified in either muscle or liver.
Little is known about the effects of sepsis in the neonate, a population whose protein
synthesis rates are relatively high and uniquely sensitive to anabolic agents (5, 7-9, 11). In a
recent study (35), lipopolysaccharide (LPS) administration to neonatal pigs to promote cytokine
production repressed protein synthesis in skeletal muscle and stimulated protein synthesis in
liver. However, the magnitude of the changes in both tissues was smaller than previously
reported for adult animals (35). In fact, in longissimus dorsi skeletal muscle, the decrease in
protein synthesis was proportional to the decline in muscle RNA content, suggesting that a fall in
ribosome number is causative in the effect of LPS on protein synthesis in that tissue. The
relative insensitivity of protein synthesis to LPS treatment in neonates thus may provide an
unique opportunity to examine the mechanisms by which LPS differentially regulates protein
synthesis in skeletal muscle and liver. Overall, the results indicate that LPS administration
results in enhanced signaling through mTOR in liver, but repressed signaling in skeletal muscle.

5
Moreover, the results suggest that eIF2B activity, rather than eIF4F assembly or S6K1
phosphorylation, may be rate controlling for global rates of protein synthesis in response to LPS
administration.
MATERIALS AND METHODS
Materials. The anti–phospho-Thr389 S6K1 antibody was purchased from Santa Cruz
Biotechnology. The anti-phospho-Ser2448 mTOR antibody was obtained from Cell Signaling
Technology. The anti-mTOR antibody was purchased from Calbiochem.
Animals. Pigs (Landrace x Yorkshire x Hampshire x Duroc) were studied at 5-6 d of age.
Three days before the study was performed, piglets were anesthetized and catheters were
surgically inserted into a jugular vein and a carotid artery as described previously (5). Twenty
piglets (5-6 days of age; 2.2 ± 0.37 kg) from two litters were assigned randomly to control
(n=10) and LPS (n=10) treatment groups. To minimize variability in the nutritional status of
pigs at the time of study, pigs were fasted for 18 h, and one hour prior to the start of the study, a
constant infusion of dextrose (800 mg/(kg•hr)) and an amino acid mixture (1.8 mmol total amino
acids/(kg•hr)) (7) was begun to simulate a fed state. At t=0, animals were administered LPS (10
µg/kg/h) or an equal volume of saline (control) and 7.5 h later, a bolus dose of [3H]phenylalanine
was administered intravenously. Eight hours after the LPS infusion was initiated, pigs were
euthanized with an intravenous dose of pentobarbital sodium (50 mg/kg body weight), and all
tissue samples were rapidly removed, frozen in liquid nitrogen, and stored at -70° C until
analysis. The protocol, previously described in Orellana et al. (35), was approved by the
Animal Care and Use Committee of Baylor College of Medicine and was conducted in
accordance with the National Research Council’s Guide for the Care and Use of Laboratory
Animals.

6
Measurement of eIF2B Activity. eIF2B activity in muscle and liver homogenates was
measured by the exchange of [3H]GDP bound to eIF2 for nonradioactively labeled GDP, as
described previously (22). Activity is expressed as the rate of GDP exchange.
Quantitation of 4E-BP1•eIF4E and eIF4G•eIF4E complexes. The association of eIF4E
with 4E-BP1 or eIF4G was quantitated as described previously (21). Briefly, eIF4E was
immunoprecipitated from muscle or liver homogenates using a monoclonal anti-eIF4E antibody.
Proteins in the immunoprecipitate were resolved by SDS-polyacrylamide gel electrophoresis and
then transferred to PVDF membranes. The membranes were then probed with either anti-4E-
BP1 or anti-eIF4G antibodies and then developed using an ECL Western Blotting kit (Amersham
Pharmacia Biotech). The horseradish peroxidase coupled to the anti-rabbit secondary antibody
was then inactivated by incubating the blot in 15% hydrogen peroxide for 30 min at room
temperature and the membranes were reprobed with the monoclonal anti-eIF4E antibody.
Values obtained using the anti-4E-BP1 and anti-eIF4G antibodies were normalized for the
amount of eIF4E present in the sample.
Measurement of site specific phosphorylation of S6K1 and mTOR. Phosphorylation of
ribosomal protein S6 kinase S6K1 was assessed by Western blot analysis using an antibody
specific for S6K1 when it is phosphorylated at the activating residue, Thr389. Phosphorylation
of mTOR on Ser2448, a residue located in a repressor domain, was assessed by Western blot
analysis using an antibody specific for mTOR phosphorylated on Ser2448. The membranes were
then treated with hydrogen peroxide as described above and reprobed with an anti-mTOR
antibody that recognizes both the phosphorylated and unphosphorylated forms of the protein.
Values obtained using the anti-phospho-mTOR antibody were normalized for the total amount of
mTOR present in the sample
Phosphorylation of the eIF4E binding protein 4E-BP1. During SDS-polyacrylamide gel
electrophoresis, 4E-BP1 resolves into multiple isoelectric forms based on its phosphorylation
state. The hyperphosphorylated g-form is the slowest migrating form and does not bind to
eIF4E. Phosphorylation of 4E-BP1 was assessed by Western blot analysis using an antibody that

7
recognizes all three forms of 4E-BP1 resolved during electrophoresis. Results are expressed as
the amount of the protein present in the g-form as a percentage of total 4E-BP1.
Statistics. Values shown are means ± SE. Statistical evaluation of the data was
performed using an unpaired, two-tailed t-test. Differences between means were considered
significant at p<0.05.
RESULTS
Because a change in eIF2B activity is the primary mechanism through which the met-
tRNAi binding step is regulated (reviewed in 17), the guanine nucleotide exchange activity of
eIF2B was measured in extracts of longissimus dorsi and liver from control and LPS-treated
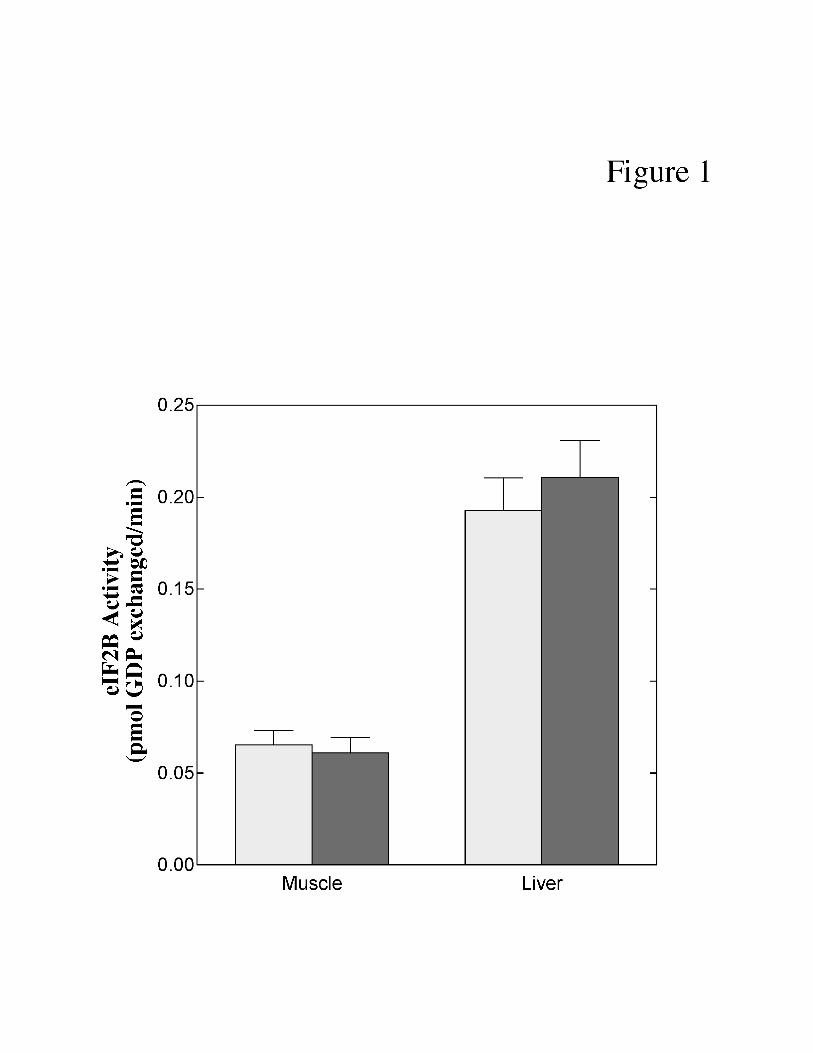
pigs. The difference in eIF2B activity between muscle and liver (Fig. 1) was directly
proportional to the previously reported difference in protein synthesis (35). Thus, eIF2B activity
was greater in liver compared to muscle. However, no change in eIF2B activity was observed in
either skeletal muscle or liver of LPS-treated compared to control pigs.
The best characterized mechanism through which the binding of mRNA to the 40S
ribosomal subunit is regulated involves the reversible binding of eIF4G to eIF4E (reviewed in
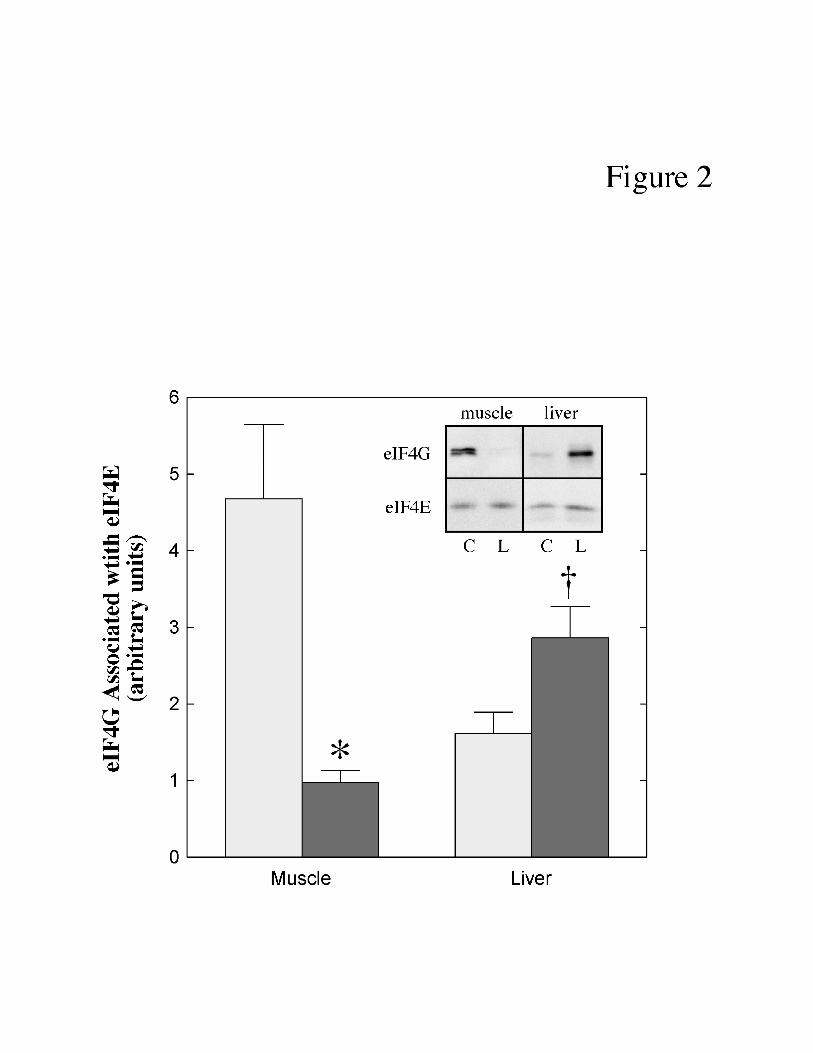
38). In the present study, the amount of eIF4G bound to eIF4E was measured by
immunoprecipitation of eIF4E followed by protein immunoblot analysis for eIF4G present in the
immunoprecipitate. As shown in Fig. 2, the amount of eIF4G present in the eIF4G•eIF4E
complex in muscle was significantly reduced (-79%) by LPS treatment. In contrast, the binding
of eIF4G to eIF4E in liver was enhanced (+77%) in response to LPS treatment.
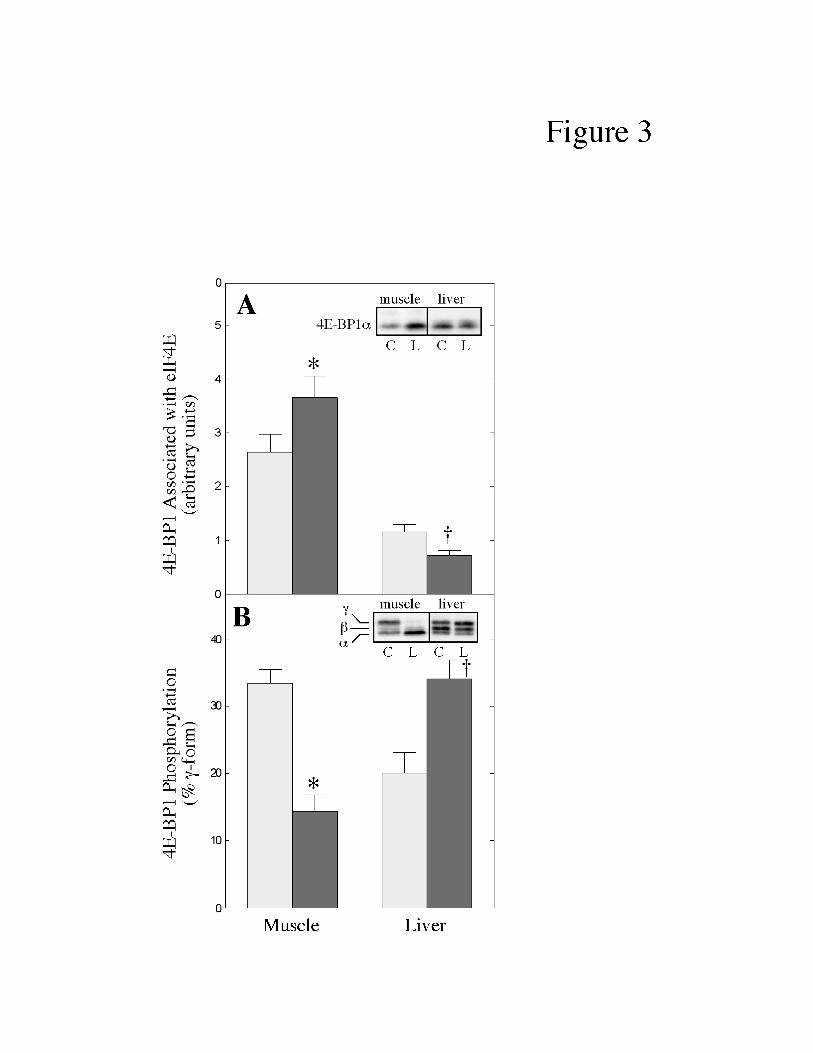
To determine whether the changes in eIF4G binding to eIF4E were due to altered
association of eIF4E with the eIF4E binding proteins, the eIF4E immunoprecipitates used for the
analyses shown in Fig. 2 were also analyzed for 4E-BP1 content. In muscle, the amount of 4E-
BP1 associated with eIF4E was increased (+53%) in LPS-treated compared to control pigs (Fig.

8
3A). In contrast, in liver, the amount of 4E-BP1 bound to eIF4E was reduced (-44%) in response
to LPS treatment.
Binding of 4E-BP1 to eIF4E is regulated by phosphorylation of 4E-BP1. When 4E-BP1
is resolved by SDS-polyacrylamide gel electrophoresis, it separates into three isoforms where the
fastest migrating form (referred to as 4E-BP1a) is the least phosphorylated and the slowest
migrating form (referred to as 4E-BP1g) is the most highly phosphorylated form of the protein
(15, 36). Because the g-form is the only one that does not bind to eIF4E, the phosphorylation
state of 4E-BP1 is presented as the proportion of the protein present in the g-form. As shown in
Fig. 3B, the amount of 4E-BP1 present in the hyperphosphorylated g-form was reduced (-60%)
in muscle, but increased (+70%) in liver, of LPS-treated compared to control pigs.
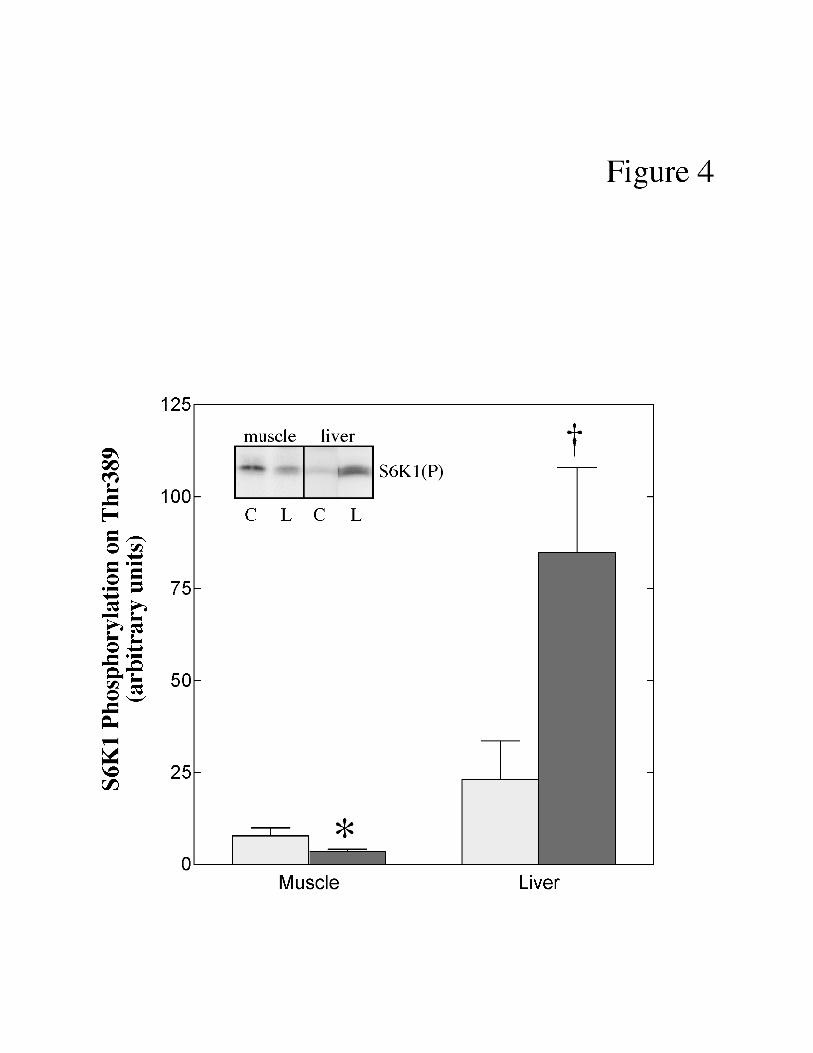
Like 4E-BP1, S6K1 is downstream of mTOR (reviewed in 38). In in vitro studies,
mTOR phosphorylates S6K1 on Thr389 (3). To determine whether phosphorylation of
S6K1(Thr389) is altered by LPS, muscle and liver extracts were analyzed by protein immunoblot
analysis using an antibody that binds to S6K1 only when it is phosphorylated on Thr389. In the
present study, phosphorylation of S6K1 on Thr389 was reduced (-74%) in muscle and increased
(+387%) in liver of LPS-treated pigs compared to control animals (Fig. 4).
Phosphorylation of both 4E-BP1 and S6K1 are dependent upon the activity of mTOR
(reviewed in 38). One mechanism that has been proposed for regulation of mTOR activity
involves phosphorylation of the protein on Ser2448, a residue that is present in a putative
repressor domain (43). In the present study, the phosphorylation state of mTOR on Ser2448 was
assessed by protein immunoblot analysis using an antibody that specifically recognizes the
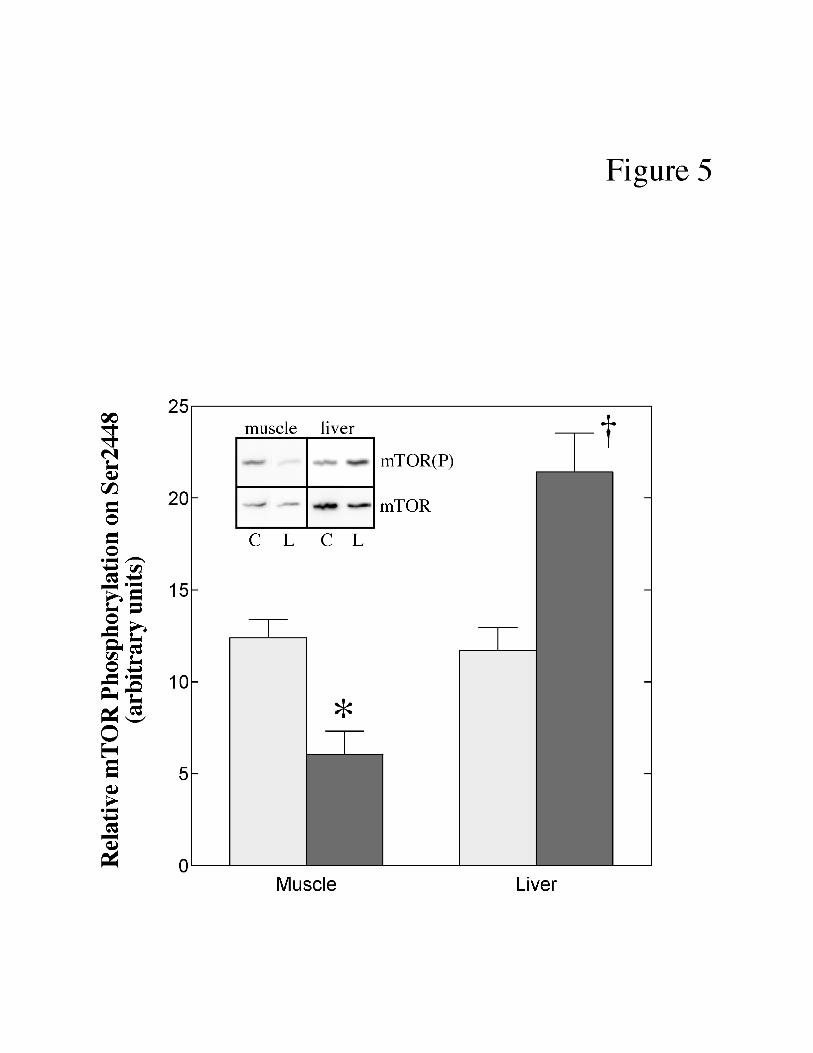
phosphorylated form of the protein. As shown in Fig. 5, the relative phosphorylation of mTOR
on Ser2448 was reduced (-51%) in muscle and enhanced (+83%) in liver of LPS-treated
compared to control pigs. Interestingly, the changes in mTOR phosphorylation were similar in
magnitude to those observed for 4E-BP1 and S6K1 phosphorylation (Fig. 3B and 5,
respectively).

9
DISCUSSION
In the present study, no change in eIF2B activity was observed in either longissimus dorsi
or liver of neonatal pigs after 8 hours of LPS infusion, suggesting that the met-tRNAi binding
step in translation initiation is unaffected under such conditions. This finding is in contrast to
previous results showing that in adult rats, either TNF-a (26) or LPS (25) administration or
chronic abdominal sepsis (48) reduces eIF2B activity in gastrocnemius muscle. The basis for the
differential response observed in neonatal pigs compared to adult rats may be due to species
differences, duration of sepsis, or in the methods used to induce a septic-like response.
Alternatively, the difference may be due to the age of the animals being studied. For example,
skeletal muscle protein synthesis is stimulated by insulin in 7 day old pigs, an effect that is
severely attenuated by 28 days of age (6). Moreover, in cells in culture, insulin stimulates eIF2B
by modulating phosphorylation of the e-subunit of eIF2B (50). Because plasma insulin
concentrations are elevated in LPS-treated pigs (35) and protein synthesis exhibits heightened
sensitivity to insulin in neonatal pigs (6), it may be that insulin is better able to maintain eIF2B
activity in muscle of neonatal compared to adult animals.
In contrast to the lack of effect on eIF2B activity in neonatal pigs, LPS-treatment
decreased the amount of eIF4G associated with eIF4E and increased the binding of 4E-BP1 to
eIF4E in skeletal muscle. It is interesting to note that the magnitude of the change in eIF4G
binding to eIF4E was greater than the change in 4E-BP1 binding. In part, this result may be due
to alterations in eIF4E binding to other eIF4E binding proteins, such as 4E-BP2 and/or 4E-BP3.
It is also noteworthy that the magnitude of the change in the association of eIF4G with eIF4E in
skeletal muscle of the neonate is similar to that reported in models of adult sepsis (26, 49),
despite the small changes in muscle protein synthesis in the neonatal model (35). This suggests
that other factors may be involved in maintaining the relatively high protein synthetic rates in
skeletal muscle of the neonate during catabolic conditions such as sepsis and endotoxemia. In
fact, it was previously reported that RNA content in both skeletal muscle and liver of neonatal

10
pigs tended to change during LPS infusion such that, when normalized for RNA content, protein
synthesis in longissimus dorsi is unchanged and in liver the magnitude of the change is reduced
to 14%. Thus, much of the change in protein synthesis in LPS-treated neonates is a result of
changes in RNA content.
The finding that the magnitude of the changes in eIF4F assembly and S6K1
phosphorylation are much larger than the changes in global protein synthesis suggests that in the
neonate, such changes may play a relatively more important role in regulating the translation of
specific mRNAs compared to be mediating global changes in protein synthesis. In this regard,
numerous studies have shown that S6K1-induced phosphorylation of rpS6 has little if any effect
on the translation of most mRNAs, but instead promotes the translation of TOP mRNAs, i.e
mRNAs that encode proteins important for cell growth (reviewed in 46). For example, during
liver regrowth after partial hepatectomy, TOP mRNA translation correlates with increased
activity of S6K1 and phosphorylation of rpS6 (1, 14, 18, 32). In contrast, cell cycle arrest is
associated with selective translational arrest of TOP mRNA translation (12, 19, 28). Finally, in
both flies (30) and mice (44) disruption of the gene encoding S6K1 results in organisms that
have the same number of cells as do wild-type animals, but with smaller cells. In part, the small
cell phenotype observed in response to S6K1 deletion is a result of impaired regulation of TOP
mRNA translation. Thus, in contrast to wild-type cells, in S6K1-/- cells serum does not promote
recruitment of TOP mRNAs into polysomes (20). One model that could account for the
preferential recruitment of TOP mRNAs into polysomes is that phosphorylated, but not
unphosphorylated, rpS6 might exhibit increased affinity for the TOP structure. The observation
that rpS6 is located near the mRNA binding site on the 40S ribosomal subunit (33, 34) makes
such a model feasible. Thus, it is not surprising that in the present study, large changes in S6K1
phosphorylation are not associated with similar changes in global protein synthesis, because TOP
mRNAs represent only a fraction of total mRNA.
Similarly, increased availability of eIF4E results in preferential translation of a subset of
mRNAs. For example, addition of either eIF4E or eIF4F to cell-free translation systems results

11
in a differential enhancement of mRNA translation; i.e. certain mRNAs are preferentially
translation when eIF4F content is increased (39, 41, 45). This finding has been extended in
recent years to show that overexpression of eIF4E in cells in culture leads to cell transformation
as a result of increased translation of mRNAs encoding proteins important in the transformation
process (reviewed in 10). One model that has been proposed to account for the observed
changes in mRNA translation is that mRNAs with short, relatively unstructured 5’-untranslated
regions (UTR) are translated preferentially compared to mRNAs with long, highly structured 5’-
UTRs (reviewed in 13). In this model, increasing eIF4F availability upregulates the translation
of mRNAs with highly structured 5’-UTRs. Thus, in the present study, the observed changes in
eIF4G•eIF4E association may result in a preferential change in the translation of specific groups
of mRNAs, an effect that might not be detected by measuring the incorporation of radioactive
amino acid into total protein over a 30 min period. It is thus interesting to note that during
sepsis, the pattern of proteins secreted by the liver changes such that secretion of normally
expressed proteins like albumin and transferrin falls dramatically (reviewed in 47). In contrast,
synthesis and secretion of the acute phase proteins from liver is enhanced. The importance of the
translational control mechanisms described above in mediating the reported changes in liver
protein synthesis is unknown. However, it is tempting to speculate that the changes in
eIF4G•eIF4E complex formation and/or S6K1 phosphorylation that occur during sepsis might
play an important role in modulating gene expression. In this regard, in unpublished studies, we
have found that the distribution of the mRNA encoding ribosomal protein S8 is predominantly
polysomal in livers from neonatal pigs treated with LPS compared to a primarily nonpolysomal
distribution in livers from control animals.
Direct support for a model wherein eIF2B is rate controlling for changes in global rates
of protein synthesis whereas eIF4F assembly and S6K1 activation are not, is provided by studies
where L6 myoblasts were deprived of either leucine or histidine (21, 24). Deprivation of either
amino acid was shown to cause decreased global rates of protein synthesis and eIF2B activity,
but only deprivation of leucine repressed eIF4F assembly and S6K1 phosphorylation. Moreover,

12
addition of insulin to leucine deprived cells restored eIF4F assembly and S6K1 phosphorylation
to values observed in control cells or histidine-deprived cells, but had no effect on either global
rates of protein synthesis or eIF2B activity. Thus, in that study changes in eIF4F assembly and
S6K1 phosphorylation had no effect on global rates of protein synthesis. Instead, changes in
eIF4F assembly and S6K1 phosphorylation were shown to promote preferentially the translation
of mRNAs encoding ornithine decarboxylase and elongation factor 1A (24).
The changes in 4E-BP1 binding to eIF4E observed in the present study were associated
with alterations in 4E-BP1 phosphorylation, whereby the proportion of the protein in the
hyperphosphorylated g-form was reduced in skeletal muscle but enhanced in liver of LPS-treated
pigs. Previous studies have shown that the feeding-induced increase in 4E-BP1
hyperphosphorylation that occurs in skeletal muscle is dependent upon the activity of mTOR (2,
23). Thus, treatment with the mTOR inhibitor rapamycin prior to feeding prevents the
subsequent phosphorylation of 4E-BP1 and S6K1 in skeletal muscle. Moreover, in cells in
culture, the phosphorylation of 4E-BP1 that occurs in response to a variety of stimuli, including
insulin, IGF-I, and amino acids, is blocked by pre-treatment with rapamycin (reviewed in 27,
37). One mechanism that has been proposed for regulation of the protein kinase activity of
mTOR involves phosphorylation of Ser2448, a residue in a domain that has been characterized
as a repressor of mTOR function (43). In this regard, PKB phosphorylates Ser2448 on mTOR in
vitro (31, 40), and activation of PKB in cells in culture is associated with enhanced
phosphorylation of Ser2448 on mTOR as well as phosphorylation of proteins downstream of
mTOR, such as 4E-BP1 and S6K1 (31, 42, 43). In the present study, we demonstrate for the
first time that changes in 4E-BP1 and S6K1 phosphorylation during sepsis were mirrored by
alterations in phosphorylation of Ser2448 on mTOR. Thus, in skeletal muscle of the neonate,
phosphorylation of all three proteins is reduced, whereas in liver, phosphorylation is enhanced by
LPS-treatment.
Perspectives. Overall, the results of the present study demonstrate for the first time that
LPS-treatment of neonatal pigs represses signaling through the mTOR pathway in muscle while

13
stimulating signaling through this pathway in liver. The observed changes in mTOR
phosphorylation are associated with modulation of both S6K1 and 4E-BP1 phosphorylation, as
well as alterations in eIF4G binding to eIF4E. In contrast, no change in eIF2B activity is
observed in either muscle or liver of neonatal pigs in response to LPS-treatment. The results
therefore suggest that in neonatal pigs, the lack of effect of LPS treatment on eIF2B activity
likely accounts for the observed maintenance of global rates of protein synthesis.
The magnitude of the changes in the phosphorylation of 4E-BP1 and the association of
eIF4E with eIF4G in skeletal muscle were similar to that reported in models of adult sepsis. In
contrast, the magnitude of the reduction in muscle protein synthesis rates in neonatal sepsis in
our recent paper (35) was less profound than that in adult sepsis models. This suggests that the
relatively high synthesis rates of skeletal muscle proteins in the neonate may in part be
maintained during neonatal sepsis through a mechanism that is distinct from eIF4F complex
formation.

14
ACKNOWLEDGEMENTS
The authors thank S. Rannels, S. Nguyen, and W. Liu for technical assistance and J.
Cunningham and F. Biggs for care of animals. We also thank Drs. C.H. Lang and T.C. Vary for
critical reading of the manuscript prior to submission.
This project has been funded in part by National Institute Health Grant R01 AR44474
and the U.S. Department of Agriculture, Agricultural Research Service under Cooperative
Agreement number 58-6250-6-001 (TAD) and National Institutes of Health Grants DK13499
and DK15658 (LSJ). The contents of this publication do not necessarily reflect the views or
policies of the U.S. Department of Agriculture nor does mention of trade names, commercial
products or organization imply endorsements by the U.S. Government.

15
REFERENCES
1. Aloni R, Peleg D, and Meyuhas O. Selective translational control and nonspecific
posttranscriptional regulation of ribosomal protein gene expression during development and
regeneration of rt liver. Molec Cell Biol 12: 2203-2212, 1992.
2. Anthony JC, Yoshizawa F, Anthony TG, Vary TC, Jefferson LS, and Kimball SR.
Leucine stimulates translation initiation in skeletal muscle of post-absorptive rats via a
rapamycin-sensitive pathway. J Nutr 130: 2413-2419, 2000.
3. Burnett PE, Barrow RK, Cohen NA, Snyder SH, and Sabatini DM. RAFT1
phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc Natl Acad
Sci USA 95: 1432-1437, 1998.
4. Cooney RN, Kimball SR, Maish G, Shumate M, and Vary TC. Effects of tumor necrosis
factor-binding protein on hepatic protein synthesis during chronic sepsis. J Surg Res 93:
257-264, 2000.
5. Davis TA, Burrin DG, Fiorotto ML, and Nguyen HV. Protein synthesis in skeletal
muscle and jejunum is more responsive to feeding in 7- than in 26-day-old pigs. Am J
Physiol 270: E802-E809, 1996.
6. Davis TA, Fiorotto ML, Beckett PR, Burrin DG, Reeds PJ, Wray-Cahen D, and
Nguyen HV. Differential effects of insulin on peripheral and visceral protein synthesis in
neonatal pigs. Am J Physiol 280: E770-E779, 2001.
7. Davis TA, Fiorotto ML, Burrin DG, Reeds PJ, Nguyen HV, Beckett PR, Vann R, and
O'Connor PMJ. Stimulation of protein synthesis by both insulin and amino acids is unique
to skeletal muscle in neonatal pigs. Am J Physiol in press, 2002.
8. Davis TA, Fiorotto ML, Nguyen HV, and Reeds PJ. Enhanced response of muscle
protein synthesis and plasma insulin to food intake in suckled rats. Am J Physiol 265: R334-
R340, 1993.

16
9. Davis TA, Fiorotto ML, Ngyuen HV, and Reeds PJ. Protein turnover in skeletal muscle
of suckling rats. Am J Physiol 257: R1141-R1146, 1989.
10. De Benedetti A and Harris AL. eIF4E expression in tumors: Its possible role in
progression of malignancies. Int J Biochem Cell Biol 31: 59-72, 1999.
11. Denne SC, Rossi EM, and Kalhan SC. Leucine kinetics during feeding in normal
newborns. Pediatr Res 30: 23-27, 1991.
12. Geyer PK, Meyuhas O, Perry RP, and Johnson LF. Regulation of ribosomal protein
mRNA content and translation in growth-stimulated mouse fibroblasts. Molec Cell Biol 2:
685-693, 1982.
13. Gingras A-C, Raught B, and Sonenberg N. Regulation of translation initiation by
FRAP/mTOR. Genes Develop 15: 807-826, 2001.
14. Gressner AM and Wool IG. The phosphorylation of liver ribosomal proteins in vivo.
Evidence that only a single small subunit protein (S6) is phosphorylated. J Biol Chem 249:
6917-6925, 1974.
15. Haystead TAJ, Haystead CMM, Hu C, Lin T-A, and Lawrence JC. Phosphorylation of
PHAS-I by mitogen-activated protein (MAP) kinase. Identification of a site phosphorylated
by MAP Kinase in vitro and in response to insulin in rat adipocytes. J Biol Chem 269:
23185-23191, 1994.
16. Hershey JWB and Merrick WC. The pathway and mechanism of initiation of protein
synthesis. In: Translational Control of Gene Expression, edited by Sonenberg N, Hershey
JWB and Mathews MB. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press,
2000, p. 33-88.
17. Hinnebusch AG. Mechanism and regulation of initiator methionyl-tRNA binding to
ribosomes. In: Translational Control of Gene Expression, edited by Sonenberg N, Hershey
JWB and Mathews MB. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press,
2000, p. 185-243.

17
18. Hornstein E, Git A, Braunstein I, Avni D, and Meyuhas O. The expression of poly(A)-
binding protein gene is translationally regulated in a growth dependent fashion through a 5'-
terminal oligopyrimidine tract motif. J Biol Chem 274: 1708-1714, 1999.
19. Jefferies HB, Fumagalli S, Dennis PB, Reinhard C, Pearson RB, and Thomas G.
Rapamycin suppresses 5'TOP mRNA translation through inhibition of p70s6k. EMBO J 16:
3693-3704, 1997.
20. Kawasome H, Papst P, Webb S, Keller GM, Johnson GL, Gelfand EW, and Terada N.
Targeted disruption of the p70S6k defines its role in protein synthesis and rapamycin
sensitivity. Proc Natl Acad Sci USA 95: 5033-5038, 1998.
21. Kimball SR, Horetsky RL, and Jefferson LS. Implication of eIF2B rather than eIF4E in
the regulation of global protein synthesis by amino acids in L6 myoblasts. J Biol Chem 273:
30945-30953, 1998.
22. Kimball SR and Jefferson LS. Mechanism of inhibition of peptide chain initiation by
amino acid deprivation in perfused rat liver. Regulation involving inhibition of eukaryotic
Initiation Factor 2a phosphatase activity. J Biol Chem 266: 1969-1976, 1991.
23. Kimball SR, Jefferson LS, and Davis TA. Feeding stimulates protein synthesis in muscle
and liver of neonatal pigs through an mTOR-dependent process. Am J Physiol 279: E1080-
E1087, 2000.
24. Kimball SR, Shantz LM, Horetsky RL, and Jefferson LS. Leucine regulates translation
of specific mRNAs in L6 myoblasts through mTOR-mediated changes in availability of
eIF4E and phosphorylation of ribosomal protein S6. J Biol Chem 274: 11647-11652, 1999.
25. Lang CH, Frost RA, Jefferson LS, Kimball SR, and Vary TC. Endotoxin-induced
decrease in muscle protein synthesis is associated with changes in eIF2B, eIF4E, and IGF-I.
Am J Physiol 278: E1133-E1143, 2000.
26. Lang CH, Frost RA, Nairn AC, MacLean DA, and Vary TC. TNF-a impairs heart and
skeletal muscle protein synthesis by altering translation initiation. Am J Physiol 282: E336-
E347, 2002.

18
27. Lawrence JC and Abraham RT. PHAS/4E-BPs as regulators of mRNA translation and
cell proliferation. Trends Biochem Sci 22: 345-349, 1997.
28. Loreni F, Francesconi A, and Amaldi F. Coordinate translational regulation in the
synthesis of elongation factor 1a nd ribosomal proteins in Xenopus laevis. Nucl Acid Res
21: 4721-4725, 1993.
29. Meyuhas O and Hornstein E. Translational control of TOP mRNAs. In: Translational
Control of Gene Expression, edited by Sonenberg N, Hershey JWB and Mathews MB. Cold
Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 2000, p. 671-693.
30. Montagne J, Stewart MJ, Stocker H, Hafen E, Kozma SC, and Thomas G. Drosophila
S6 kinase: a regulator of cell size. Science 285: 2126-2129, 1999.
31. Nave BT, Ouwens DM, Withers DJ, Alessi DR, and Shepherd PR. Mammalian target of
rapamycin is a direct target for protein kinase B: Identification of a convergence point for
opposing effects of insulin and amino acid deficiency on protein translation. Biochem J 344:
427-431, 1999.
32. Nemenoff R, Price D, Mendelsohn M, Carter E, and Avruch J. An S6 kinasse activated
during liver regeneration is related to the insulin-stimulated S6 kinase in H4 hepatoma cells.
J Biol Chem 263: 19455-19460, 1988.
33. Nygard O and Nika H. Identification by RNA-protein cross-linking of ribosomal proteins
located at the interface between the small and the large subunits of mammalian ribosomes.
EMBO J 1: 357-362, 1982.
34. Nygard O and Nilsson L. Translational dynamics. Interactions between the translational
factors, tRNA and ribosomes during eukaryotic protein synthesis. Eur J Biochem 191: 1-17,
1990.
35. Orellana RA, O'Connor PMJ, Nguyen HV, Bush JA, Suryawan A, Thivierge MC,
Fiorotto ML, and Davis TA. Endotoxemia reduces skeletal muscle protein synthesis in
neonates. Am J Physiol 283: E909-E916, 2002.

19
36. Pause A, Belsham GJ, Gingras A-C, Donze O, Lin T-A, Lawrence JC, and Sonenberg
N. Insulin-dependent stimulation of protein synthesis by phosphorylation of a regulator of
5'-cap function. Nature 371: 762-767, 1994.
37. Raught B and Gingras A-C. eIF4E activity is regulated at multiple levels. Int J Biochem
Cell Biol 31: 43-57, 1999.
38. Raught B, Gingras A-C, and Sonenberg N. Regulation of ribosomal recruitment in
eukaryotes. In: Translational Control of Gene Expression, edited by Sonenberg N, Hershey
JWB and Mathews MB. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press,
2000, p. 245-293.
39. Ray BK, Brendler TG, Adya S, Daniels MS, Miller JK, Hershey JW, Grifo JA,
Merrick WC, and Thach RE. Role of mRNA competition in regulating translation: further
characterization of mRNA discriminatory initiation factors. Proc Natl Acad Sci USA 80:
663-667, 1983.
40. Reynolds TH, Bodine SC, and Lawrence JC. Control of Ser2448 phosphorylation in the
mammalian target of rapamycin by insulin and skeletal muscle load. J Biol Chem 277:
17657-17662, 2002.
41. Sarkar G, Edery I, Gallo R, and Sonenberg N. Preferential stimulation of rabbit alpha
globin mRNA translation by a cap-binding protein complex. Biochem Biophys Acta 783:
122-129, 1984.
42. Scott PH, Brunn GJ, Kohn AD, Roth RA, and Lawrence JC. Evidence of insulin-
stimulated phosphorylation and activation of the mammalian target of rapamycin mediated
by a protein kinase B signaling pathway. Proc Natl Acad Sci USA 95: 7772-7777, 1998.
43. Sekulic A, Hudson CC, Homme JL, Yin P, Otterness DM, Karnitz LM, and Abraham
RT. A direct linkage between the phosphoinositide 3-kinase-Akt signaling pathway and the
mammalian target of rapamycin in mitogen-stimulated and transformed cells. Cancer Res
60: 3504-3523, 2000.

20
44. Shima H, Pende M, Chen Y, Fumagalli S, Thomas G, and Kozma SC. Disruption of the
p70(S6k)/p85(S6k) gene reveals a small mouse phenotype and a new functional S6 kinase.
EMBO J 17: 6649-6659, 1998.
45. Sonenberg N, Trachsel H, Hecht S, and Shatkin AJ. Differential stimulation of capped
mRNA translation in vitro by cap binding protein. Nature 285: 331-333, 1980.
46. Thomas G. An encore for ribosome biogenesis in the control of cell proliferation. Nature
Cell Biol 2: E71-E72, 2000.
47. Vary TC. Interorgan protein and carbohydrate metabolic relationships during sepsis:
Necessary evils or uncanny coincidences? Curr Opin Clin Nutri Metab Care 2: 235-242,
1999.
48. Vary TC, Jurasinski CV, Karinch AM, and Kimball SR. Regulation of eukaryotic
initiation factor 2 expression in sepsis. Am J Physiol 266: E193-E201, 1994.
49. Vary TC and Kimball SR. Effect of sepsis on eIF4E availability in skeletal muscle. Am J
Physiol 279: E1178-E1184, 2000.
50. Welsh GI, Miller CM, Loughlin AJ, Price NT, and Proud CG. Regulation of eukaryotic
initiation factor eIF2B: glycogen synthase kinase-3 phosphorylates a conserved serine which
undergoes dephosphorylation in response to insulin. FEBS Lett 421: 125-130, 1998.

21
FIGURE LEGENDS
Fig. 1. Effect of LPS treatment on eIF2B activity in skeletal muscle and liver of neonatal
pigs. Neonatal pigs were treated with LPS and the rate of exchange of [3H]GDP bound to eIF2
for nonradiolabeled GDP was measured as described under “Materials and Methods”. The
results represent the mean ± SE for 9-10 animals per condition.
Fig. 2. Effect of LPS treatment on eIF4G association with eIF4E in skeletal muscle and
liver of neonatal pigs. Neonatal pigs were treated with LPS as described under “Materials and
Methods”. The amount of eIF4G bound to eIF4E was measured as described under "Materials
and Methods". Values for eIF4G were normalized for the recovery of eIF4E in the
immunoprecipitate. The results represent the mean ± SE for 10 animals per condition. The
results of typical blots are shown as an inset to the panel. Muscle and liver samples were
analyzed on separate immunoblots and therefore the results from the two tissues cannot be
directly compared. C, control pigs; L, LPS-treated pigs. * p<0.02 vs. control muscle; † p<0.025
vs. control liver.
Fig. 3. Effect of LPS treatment on eIF4E association with 4E-BP1 and 4E-BP1
phosphorylation in skeletal muscle and liver of neonatal pigs. Neonatal pigs were treated with
LPS as described under “Materials and Methods”. (A) The amount of 4E-BP1 bound to eIF4E
was measured in the same immunoprecipitates used for the analysis shown in Fig. 2. Values for
4E-BP1 were normalized for eIF4E recovery. The results represent the mean ± SE for 9-10
animals per condition. The results of typical blots are shown as an inset to the panel. Muscle
and liver samples were analyzed on separate immunoblots and therefore the results from the two
tissues cannot be directly compared. C, control pigs; L, LPS-treated pigs. * p<0.05 vs. control
muscle; † p<0.01 vs. control liver. (B) Phosphorylation of 4E-BP1 was measured as described
under "Materials and Methods" and is expressed as the percentage of the protein present in the

22
hyperphosphorylated g-form. The results represent the mean ± SE for 9-10 animals per
condition. The results of typical blots are shown as an inset to the panel. The location of the a,
b, and g-forms of 4E-BP1 are noted to the left of the inset. C, control pigs; L, LPS-treated pigs.
* p<0.001 vs. control muscle; †p<0.002 vs. control liver.
Fig. 4. Effect of LPS treatment on S6K1(Thr389) phosphorylation in skeletal muscle and
liver of neonatal pigs. Neonatal pigs were treated with LPS as described under ‘Materials and
Methods”. Phosphorylation of S6K1 on Thr389 was measured by Western blot analysis using
an anti-phospho-Thr389 antibody as described under “Materials and Methods”. The results
represent the mean ± SE for 9 animals per condition. The results of typical blots are shown as an
inset to the panel. Muscle and liver samples were analyzed on separate immunoblots and
therefore the results from the two tissues cannot be directly compared. C, control pigs; L, LPS-
treated pigs. * p<0.05 vs. control muscle; † p<0.05 vs. control liver.
Fig. 5. Effect of LPS treatment on mTOR(Ser2448) phosphorylation in skeletal muscle
and liver of neonatal pigs. Neonatal pigs were treated with LPS as described under “Materials
and Methods”. Phosphorylation of mTOR on Ser2448 was measured by Western blot analysis
using an anti-phospho-Ser2448 antibody as described under “Materials and Methods”. Values
for phosphorylated mTOR [mTOR(P)] were normalized for total mTOR (mTOR) content. The
results represent the mean ± SE for 10 animals per condition. The results of typical blots are
shown as an inset to the panel. Muscle and liver samples were analyzed on separate
immunoblots and therefore the results from the two tissues cannot be directly compared. C,
control pigs; L, LPS-treated pigs. * p<0.01 vs. control muscle; † p<0.01 vs. control liver
Related Documents











