of August 1, 2015. This information is current as Pathways TLR9-Dependent and -Independent Activates Dendritic Cells via Endosomal Translocation of Vertebrate DNA Hochrein and Hermann Wagner Hubertus Schlatter, Frank Schmitz, Antje Heit, Stefan Bauer, Kei Yasuda, Philipp Yu, Carsten J. Kirschning, Beatrix http://www.jimmunol.org/content/174/10/6129 doi: 10.4049/jimmunol.174.10.6129 2005; 174:6129-6136; ; J Immunol References http://www.jimmunol.org/content/174/10/6129.full#ref-list-1 , 21 of which you can access for free at: cites 44 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2005 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on August 1, 2015 http://www.jimmunol.org/ Downloaded from by guest on August 1, 2015 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of August 1, 2015.This information is current as
PathwaysTLR9-Dependent and -IndependentActivates Dendritic Cells via Endosomal Translocation of Vertebrate DNA
Hochrein and Hermann WagnerHubertusSchlatter, Frank Schmitz, Antje Heit, Stefan Bauer,
Kei Yasuda, Philipp Yu, Carsten J. Kirschning, Beatrix
http://www.jimmunol.org/content/174/10/6129doi: 10.4049/jimmunol.174.10.6129
2005; 174:6129-6136; ;J Immunol
Referenceshttp://www.jimmunol.org/content/174/10/6129.full#ref-list-1
, 21 of which you can access for free at: cites 44 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Endosomal Translocation of Vertebrate DNA ActivatesDendritic Cells via TLR9-Dependent and-Independent Pathways1
Kei Yasuda, Philipp Yu, Carsten J. Kirschning, Beatrix Schlatter, Frank Schmitz, Antje Heit,Stefan Bauer, Hubertus Hochrein,2 and Hermann Wagner2,3
TLRs discriminate foreign from self via their specificity for pathogen-derived invariant ligands, an example being TLR9 recog-nizing bacterial unmethylated CpG motifs. In this study we report that endosomal translocation of CpG DNA via the naturalendocytotic pathway is inefficient and highly saturable, whereas endosomal translocation of DNA complexed to the cationic lipidN-[1-(2,3-dioleoyloxy)propyl]-N,N,N-trimethylammonium methylsulfate (DOTAP) is not. Interestingly, DOTAP-mediated en-hanced endosomal translocation of otherwise nonstimulatory vertebrate DNA or of certain noncanonical CpG motifs triggersrobust dendritic cell activation in terms of both up-regulation of CD40/CD69 and cytokine production, such as type I IFN and IL-6.We report that the stimulatory activity of phosphorothioated noncanonical CpG oligodeoxynucleotides is TLR9 dependent,whereas phosphodiester DNA, such as vertebrate DNA, in addition trigger TLR9-independent pathways. We propose that theinefficiency of the natural route for DNA internalization hinders low affinity TLR9 ligands in endosomes to reach thresholdconcentrations required for TLR9 activation. Endosomal compartmentalization of TLR9 may thus reflect an evolutionary strategyto avoid TLR9 activation by self-DNA. The Journal of Immunology, 2005, 174: 6129–6136.
B acterial and viral DNA rich in CpG motifs or small syn-thetic oligodeoxynucleotides (ODN)4 containing CpGmotifs activate innate immune cells such as dendritic
cells (DCs) via TLR9 (reviewed in Refs. 1 and 2). Cellular acti-vation by immunostimulatory CpG-ODN appears to be sequencespecific, because inversion within CpG motifs of CG to GC ormethylation of cytosine (in this study termed noncanonical CpGmotifs) ablates its immunostimulatory activities (3–6). Surpris-ingly, vertebrate DNA fails to activate innate immune cells (2, 6).To date, this lack of immunostimulatory potential has been ex-plained by CpG suppression (7), CpG methylation (8), and/or in-hibitory motifs (9, 10), even though vertebrate DNA still containssome unmethylated CpG motifs (6).
Unlike TLR1, -2, -4, -5, and -6, which recognize invariant for-eign bacterial and viral constituents at the cell membrane, TLR3,-7, -8, and -9 are expressed endosomally in innate immune cellsand recognize distinct patterns of nucleic acids at late endosomalcompartments (11–15). Although in the former cases self-nonself
discrimination can be easily explained by the invariant molecularnature of the respective foreign ligands, structural differences be-tween pathogen and host nucleic acids appear less prominent. Thisraises the question of whether it is the endosomal compartmental-ization and thus the limited accessibility of TLR3 and -7 to -9 thathelp to control foreign vs self nucleic acid discrimination. Specif-ically, pathogen-derived DNA may access the TLR9-expressingendosomal compartment of infected cells in the course of infec-tions, whereas host-derived DNA may not, because it becomesrapidly degraded by extracellular DNase. Recent evidence impliesthat upon internalization of chromatin-IgG immune complexes(ICs) via BCR or Fc�RIII of DCs, vertebrate DNA displays robustimmunostimulatory activities toward B cells or DCs (16, 17). Inthe case of DCs, both TLR9-dependent and -independent signalingpathways appear to operate (17). Furthermore, the sera of patientswith systemic lupus erythematosus containing ICs consisting ofautologous DNA and anti-DNA Abs effectively activate plasma-cytoid DCs (pDCs) to produce type I IFNs (18, 19). Like otherresearchers (20–23), we have noted that high concentrations ofnoncanonical CpG DNA sequences may moderately activateTLR9-expressing immune cells even though they are sequence-divergent from canonical unmethylated CpG motifs to date con-sidered as natural TLR9 ligand. Because canonical and noncanoni-cal CpG motifs as well as vertebrate DNA first need to translocateto the endosome to trigger TLR9 (14, 15, 24), we analyzed whetherthe limited accessibility to TLR9 prevents vertebrate DNA to un-veil immunostimulatory effects.
To clarify aspects of these matters, it seemed desirable to es-tablish a system in which the potentially rate-limiting step of nat-ural DNA internalization is bypassed. To do this, we complexedeither vertebrate DNA or canonical or noncanonical CpG motifs tothe cationic lipids N-[1-(2,3-dioleoyloxy)propyl]-N,N,N-trimethyl-ammonium methylsulfate (DOTAP) known to enforce endosomaltranslocation (25, 26). To analyze the DC stimulatory capacity ofDNA and to exclude interference by potentially contaminating
Institute of Medical Microbiology, Immunology, and Hygiene, Technical Universityof Munich, Munich, Germany
Received for publication January 12, 2005. Accepted for publication March 2, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Deutsche Forschungsgemeinschaft (FORIMMUNSFB391 and SFB456) and by the Coley Pharmaceutical Group.2 H.H. and H.W. contributed equally to this study.3 Address correspondence and reprint requests to Dr. Hermann Wagner, Institute ofMedical Microbiology, Immunology, and Hygiene, Trogerstrasse 9, 81675 Munich,Germany. E-mail address: [email protected] Abbreviations used in this paper: ODN, oligodeoxynucleotide; CT, calf thymus;DC, dendritic cell; cDC, conventional DC; CT DNA, calf thymus DNA; FL, Flt3ligand; FL-DC, FL-dependent DC; IC, immune complex; PD, phosphodiester; pDC,plasmacytoid DC; DOTAP, N-[1-(2,3-dioleoyloxy)propyl]-N,N,N-trimethylammo-nium methylsulfate; pDNA, plasmid DNA; PTO, phosphorothioated; TMR, tetram-ethylrhodamine; LAMP, lysome-associated membrane protein; NAOS, nonactivatingODN phosphorothioated; wt, wild type.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

non-TLR9 ligands such as lipopeptides or endotoxin, we generatedDCs lacking either TLR2 and -4 or TLR9 in addition. We nowdescribe that upon DOTAP-mediated enforced endosomal translo-cation, both vertebrate DNA as well as phosphodiester (PD) ca-nonical and noncanonical CpG motifs can activate DCs via TLR9-dependent and -independent pathways to up-regulatecostimulatory cell surface molecules. In contrast, production ofDC subset-specific cytokines primarily requires signaling viaTLR9. Interestingly, the immunostimulatory activities of phospho-rothioated (PTO) canonical and noncanonical ssCpG motifs arefound to be entirely TLR9 dependent.
Materials and MethodsMice
C57BL6 wild-type (wt) mice were purchased from Harlan. TLR2�/�/TLR4�/�/TLR9�/� mice were crossed with TLR9-deficient mice to yieldtriple TLR2�/�/TLR4�/�/TLR9�/� mice. Mice used as donors of bone mar-row cells were on a mixed genetic background (129SV � C57BL/6).
DNA
The ODN, listed in Table I, were purchased from TIB Molbiol or MWGBiotech. Plasmid DNA (pcDNA4 Myc His; Invitrogen Life Technologies)was purified with an Endofree Plasmid kit (Qiagen). Plasmid DNA and calfthymus (CT) DNA (Sigma-Aldrich) were also subjected to phenol/chloro-form/isoamylalcohol (25/24/1) extraction, followed by a Triton X-114-based purification step to remove LPS, as described previously (26).
Preparation of DC and human PBMC
In vitro Flt3 ligand (FL)-dependent DC (FL-DC) were generated as de-scribed previously (27). In short, bone marrow cells were cultured in thepresence of murine rFL for 8 days. Resulting cells were �90% CD11cpositive, and 30–40% of cells displayed a plasmacytoid phenotype(CD11cposCD45RAhighB220highCD11blow). DCs (3 � 105 to 5 � 105 cells/200 �l) were incubated with the respective stimuli in 96-well plates.
Human PBMC were generated by Ficoll gradient centrifugation andwere seeded at 3 � 105 to 5 � 105 cells/200 �l cells for cytokine induction.Human pDC were enriched or depleted by incubation with PE-labeled Absto CD123, followed by anti-PE beads (Miltenyi Biotec) as described pre-viously (28) and were seeded at 5 � 104 cells/200 �l.
Cell stimulation
Human or murine cells were incubated for 18–24 h with 12.5 �g/ml DNAas indicated in the figures. Complexes of DNA with DOTAP (Roche) wereprepared according to the manufacturer’s instructions. In brief, 5 �g ofDNA in 50 �l of HBS (20 mM HEPES and 150 mM NaCl, pH 7.4) wascombined with 10 �g of DOTAP in 50 �l of HBS. After 15 min of incu-bation, 100 �l of complete RPMI 1640 medium (10% FBS, 100 U/mlpenicillin, 100 �g/ml streptomycin, and 50 �M 2-ME) was added to themixture of DNA and DOTAP. Cells in 100 �l of complete medium wereincubated with 100 �l of DNA-DOTAP complexes for 18–24 h in 96-wellculture plates., then collected for FACS analyses. The cytokines in thesupernatants were quantified by ELISA as described previously (28).
FACS analysis
Before stimulation, FL-DC cultures were routinely checked for the relativepercentages of pDC and conventional DC (cDC) and the respective DC
activation status by staining with mAb to CD11c, CD45RA, B220, CD11b,and CD62L. After stimulation, FL-DC were labeled with a combination ofAbs to B220 (RA3-6B2-allophycocyanin), CD40 (FGK45.5-PE), andCD69 (H1.2F3-FITC; all from BD Biosciences) and analyzed on a FAC-SCalibur flow cytometer (BD Biosciences). For uptake analyses, FL-DCwere exposed to 5�-Bodipy-tetramethylrhodamine (TMR)-conjugatedCpG-ODN 1668-PTO (IBA Naps) in the presence or the absence ofDOTAP for various time periods. Afterward, cells were extensivelywashed, chased for 3 h, stained with ethidium monoazide (MolecularProbes) to exclude dead cells, and subsequently analyzed.
In vivo experiments
DNA (30 �g) was diluted in 75 �l of HBS. DOTAP (60 �g) was dilutedin 75 �l of HBS. Subsequently, the DNA solution was mixed with theDOTAP solution and incubated for 15 min. One hundred and fifty micro-liters of DNA-DOTAP complex containing solution was injected i.v. After2 h, mice were killed, blood was collected, and cytokines in the plasmawere determined by ELISA.
Confocal laser scanning microscopy
Ten thousand RAW264.7 murine macrophage cells per well were culturedovernight in eight-well chamber slides (Falcon; BD Biosciences). For stim-ulation, cells were incubated with 2 �M 3�-fluorescein-labeled CpG-ODN1668-PTO (TIB Molbiol) for 60 min at 37°C and thereafter washed threetimes with ice-cold PBS. Cells were fixed (PBS and 4% formaldehyde) for15 min and permeabilized with staining buffer (PBS, 0.4% saponin, and 2%normal goat serum (Invitrogen Life Technologies)) for 15 min. Incubationwith primary Ab (anti-lysosome-associated membrane protein (LAMP1);BD Pharmingen; 1/500) and secondary Ab (anti-FITC-Alexa 488 (Molec-ular Probes; 1/400) and anti-rat IgG-Alexa 546 (Molecular Probes; 1/400))was performed at room temperature for 1 h and 30 min, respectively. Mi-croscope slides were mounted and analyzed at room temperature using aZeiss LSM500 confocal microscope (software version 2.2) equipped withan argon/krypton laser (458/488 nm) and two helium/neon lasers (543 and633 nm). The lens used was a Plan-Neofluar (Zeiss) 40 1.3 oil lens, and thepinhole was set to scan layers �1 �m at a resolution of 1024 � 1024pixels. Pictures were then imported into Photoshop CS (Adobe Systems).
ResultsEndosomal delivery potential of DOTAP (cationic lipid)
Recently, we and others used the cationic lipid DOTAP to trans-locate ssRNA to TLR7-expressing endosomes of DCs (28, 29). Inthis study we asked whether DOTAP can also be used to translo-cate DNA to endosomes expressing TLR9 (reviewed in Refs. 2 and24). In addition, we addressed the question of how the efficacy ofDOTAP-mediated endosomal translocation compares with that ofthe natural translocation route known to be mediated by TLR9-independent DNA receptor-driven endocytosis (24, 30–32). UsingFACS and confocal microscopy to quantitate natural vs DOTAP-mediated internalization of Bodipy-TMR-tagged CpG-ODN, weestablished that CpG DNA uptake via the natural internalizationpathway is highly saturable at higher CpG DNA concentrations,i.e., 1–2 �M, whereas DOTAP-mediated internalization is not(Fig. 1). These differences were observed with both canonical andnoncanonical CpG-ODN (data not given) and allowed DCs to in-ternalize 10- to 50-fold more DNA complexed to DOTAP (Fig. 1).Confocal microscopy revealed that DOTAP translocated its cargoDNA into intracellular vesicular structures did not deposit CpGDNA at the cell surface (data not shown). In fact, colocalizationanalysis demonstrated that CpG DNA complexed to DOTAPtranslocated primarily to LAMP1-positive late endosomal com-partments (Fig. 2), a site at which TLR9 is expressed and becomesactivated (14, 15, 30). We concluded that by complexing DNA toDOTAP, up to 10- to 50-fold more DNA can be translocated to thelate endosomal compartment of DCs compared with the naturalroute of CpG DNA uptake.
Table I. Sequences of ODN used in this studya
ODN Name Sequences
1668 PTO tccatgacgttcctgatgct1720 PTO tccatgagcttcctgatgctNAOS-1 PTO gctcatgagcttcctgatgctgAP-1 PTO gcttgatgactcagccggaa2216 ggGGGACGATCGTCgggggg1668 PD TCCATGACGTTCCTGATGCT1720 PD TCCATGAGCTTCCTGATGCTm-1668 PD TCCATGA[meth]CGTTCCTGATGCT
a Lowercase letters indicate PTO, and capital letters show PD backbone.
6130 TLR9, NON-CpG DNA, AND DC ACTIVATION
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

TLR9-dependent and -independent DC activation bynoncanonical CpG DNA and vertebrate DNA
Canonical ssCpG motifs efficiently activating murine TLR9 con-tain the hexameric sequence GACGTT present in ODN 1668,whereas the prototypic ODN sequence for activation of human orprimate TLR9 displays a GTCGTT sequence (reviewed in Refs. 1and 2). Upon methylation of CpG-ODN 1668 or inversion of itscentral dinucleotide from CG to GC (herein termed noncanonicalCpG motifs), the immunostimulatory activity of these ODNs isablated (2). These findings are based on experiments using thenatural DNA uptake pathway by exposing innate immune cells invitro or in vivo to the respective DNAs (1, 2, 24). Under suchexperimental conditions, vertebrate DNA fails to stimulate innateimmune cells (2, 6). Unexpectedly, upon DOTAP-mediated endo-somal translocation of graded concentrations of the noncanonicalCpG-ODN 1720 or CT DNA, these otherwise nonstimulatoryDNAs triggered production of IL-6 in wt FL-DCs. Furthermore,both pDNA and CT DNA complexed to DOTAP caused low, butsignificant, IL-6 production in DCs from TLR9-deficient cells(Fig. 3B). In contrast, DOTAP enhanced the stimulatory potentialof the canonical CpG motif 1668 only at low concentrations (Fig.3A), whereas the noncanonical CpG motif 1720 complexed toDOTAP unfolded stimulatory activity at higher concentrations(Fig. 3A), a finding also observed with the methylated CpG motif1668 (data not given) or in part with bacterial pDNA (Fig. 3B,inset). Of note, under conditions in which vertebrate DNA exhib-
ited stimulatory activity (Fig. 3B), the control ODNs, AP-1 or non-activating ODN phosphorothioated (NAOS)-1, remained inactive(Fig. 3C). The lack of stimulatory activity of the control ODNs,AP-1 and NAOS-1, indicates sequence specificity. Both controlODNs displayed a guanine at the 5� end, and we noted that addi-tion of a 5� G (guanine) and 3� G ablated the stimulatory potentialof ODN 1720 (data not given). In quantitative terms, vertebrateDNA complexed to DOTAP displayed �10–15% the activity ofthe canonical CpG motif 1668 or of complexed pDNA, whereascomplexed noncanonical CpG motif 1720 (or methylated ODN1668; data not shown) unfolded �50% of this activity (Fig. 3).
To exclude the effects of potential contamination with, for example,endotoxin or lipopeptides, we reanalyzed, at a 2-�M concentration,
FIGURE 1. Kinetics of CpG-DNA internalization by DCs. FL-DC (2 �106/ml) were exposed for different time periods (10 min up to 3 h) withBodipy-TMR-conjugated CpG-ODN 1668-PTO (0.2 and 2 �M). A, Cellswere washed at the individual time points indicated and incubated for ad-ditional 3 h. B, FL-DC were exposed to Bodipy-TMR-conjugated CpG-ODN 1668-PTO complexed to DOTAP, washed, and incubated for anadditional 3 h. The fluorescence of internalized ODN was determined byFACS. Similar results were obtained with TLR9-deficient FL-DC (data notgiven). One representative experiment of at least three is shown.
FIGURE 2. Liposomes (DOTAP) translocate CpG-DNA to LAMP1-positive endosomal compartments. RAW264.7 cells were pulsed for 60min with 2 �M FITC-conjugated CpG-ODN 1668-PTO with or withoutDOTAP. Thereafter, cells were fixed, permeabilized, stained for LAMP1(Alexa 546; red) and FITC (anti-FITC Alexa 488; green), and subjected toconfocal laser scanning microscopy (CLSM). Representative pictures withpinhole settings for �1-�m layers are shown.
6131The Journal of Immunology
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

the immunostimulatory potential of methylated CpG-1668-PTO,1668 PD, methylated 1668-PD, noncanonical CpG-ODN 1720-PTO,or 1720-PD, pDNA, and CT DNA using FL-DCs generated fromTLR2 and -4 double-deficient or from TLR2, -4, and -9 triple-defi-cient bone marrow cells. Under these conditions, mixed DC popula-tions of pDC and cDC were generated (33). As a marker for DCstimulation, we analyzed up-regulation of CD69 and CD40 (Fig. 4A)as well as IFN-� (prototypic cytokine produced by pDCs) or IL-6production (a cytokine preferentially produced by cDCs; Fig. 4B). Inthe absence of DOTAP, only canonical CpG-ODN 1668-PTO andbacterial pDNA activated the TLR2- and TLR4-deficient DC popu-lations to up-regulate CD40 and CD69. This was TLR9 dependent.Upon DOTAP-mediated endosomal translocation, however, the non-canonical CpG-ODN 1720-PTO, methylated CpG-1668-PD, as wellas vertebrate CT DNA also caused strong up-regulation of CD40 andCD69, whereas the control ODN AP-1 did not. Fig. 4A presents ad-ditional information. The immunostimulatory activity of PTO canon-ical and noncanonical CpG DNA complexed to DOTAP was entirelyTLR9 dependent, yet PD DNA (ODN 1668, ODN 1720, bacterialpDNA, and vertebrate DNA) displayed TLR9-independent stimula-tory activity, because they caused up-regulation of CD40 and CD69 inTLR2, TLR4-, and TLR9-deficient DCs (Fig. 4A). Fig. 4B illustratesIFN-� and IL-6 production triggered by the various stimuli in thesemixed DC cultures. As expected, only the canonical CpG-ODN 1668-PTO and bacterial pDNA (PD) caused, in the absence of DOTAP,TLR2- and TLR4-deficient DC to produce substantial amounts ofIL-6. However, when complexed to DOTAP, vertebrate DNA as wellas noncanonical CpG motifs, such as methylated ODN 1668-PD ornoncanonical CpG-ODN 1720-PTO, were also effective (Fig. 4B).Again, the control ODN AP-1 was negative. IL-6 production was
largely TLR9 dependent, because TLR2, -4, and -9 triple-deficientDC produced only minor amounts (1–5%) of IL-6. We were surprisedto note that CpG-ODN 1668-PTO failed to trigger IFN-� productionin TLR2- and TLR4-deficient DCs, whereas noncanonical CpG-ODN1720-PTO was effective (Fig. 4B). We therefore titrated the concen-trations of these ODN. As shown in Fig. 4C, the biological activity ofODN-1668 PTO was suppressed at high (2 �M) concentrations,whereas that of ODN 1720-PTO was not. Yet, both ODN triggeredIFN-� at intermediate ODN concentrations. Thus, both vertebrateDNA as well as the canonical CpG motif 1668 and the noncanonicalCpG motifs (methylated ODN 1668 and ODN 1720) complexed toDOTAP efficiently triggered IFN-� production in TLR2- and TLR4-deficient DCs. In contrast, IFN-� production in TLR2-, TLR4-, andTLR9-deficient DCs could not be detected. Taken together, these dataallowed three major conclusions. First, the sequence restrictions de-scribed for canonical CpG motifs (reviewed in Refs. 1 and 2) becomeless stringent upon DOTAP-mediated enhanced endosomal delivery.Second, upon endosomal translocation, vertebrate DNA (known tocontain noncanonical CpG motifs) displays DC stimulatory activities.Third, PTO canonical and noncanonical CpG DNA stimulate DCs ina TLR9-dependent fashion, whereas PD DNAs, in addition, trigger aTLR9-independent pathway that drives up-regulation of CD40 andCD69, but is poor in inducing cytokine production (Fig. 4, A and B).
Type I IFN response of human PBMCs
In humans, pDCs and B cells express TLR9 (2, 24). We testedwhether upon DOTAP-mediated endosomal translocation of non-canonical PD or PTO CpG motifs triggers type I IFN release from
FIGURE 3. Endosomal translocation of canonicaland noncanonical CpG-DNA, pDNA, and vertebrateDNA triggers IL-6 production. A, Respective DNAwas mixed at graded concentrations with 10 �g ofDOTAP in 50 �l of HBS. After incubation for 15 minat 37°C 100 �l of complete RPMI 1640 medium wasadded. The mixture was added to wt or TLR9-defi-cient FL-DC cultures (2.5 � 106/ml). B and C, ODN(0.8 nmol) or 5 �g of bacterial DNA or vertebrateDNA in 50 �l of HBS was combined with 10 �g ofDOTAP (in 50 �l of HBS). After 15-min incubation,the DNA complexed to DOTAP was successively di-luted, as indicated, and complete RPMI 1640 medium(100 �l) was added and thereafter cultured togetherwith wt or TLR9-deficient FL-DCs. The supernatantswere analyzed for IL-6 production by ELISA. Onerepresentative experiment of at least three is shown.Error bars represent the range of duplicate samples.
6132 TLR9, NON-CpG DNA, AND DC ACTIVATION
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
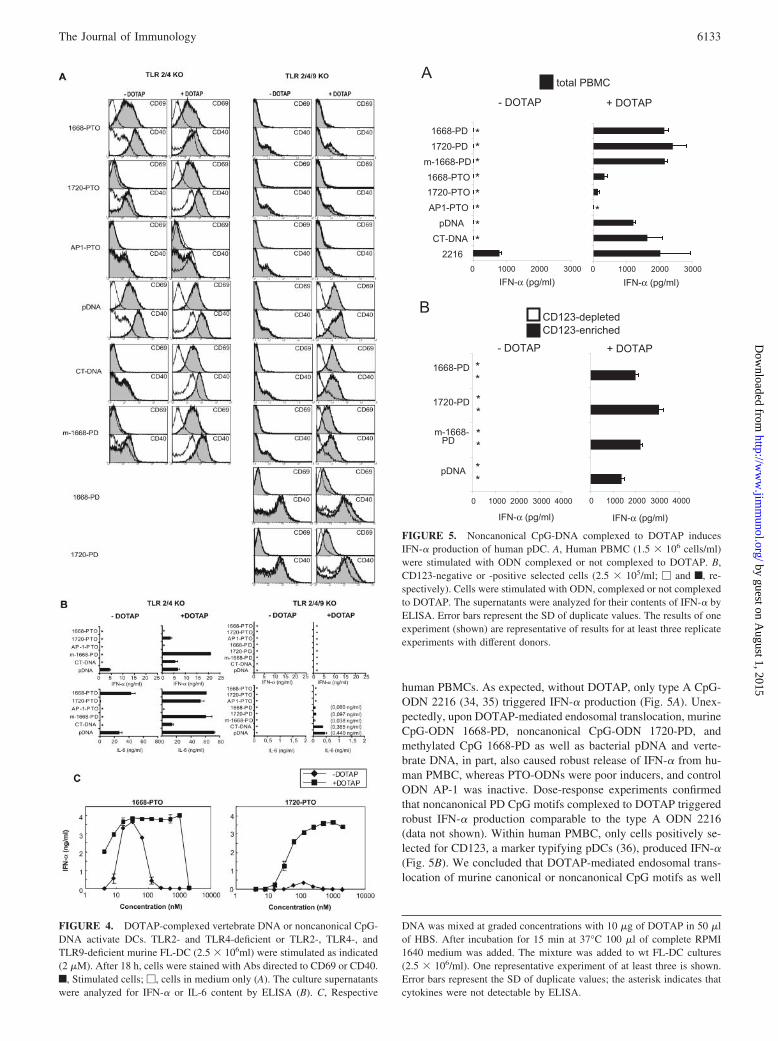
human PBMCs. As expected, without DOTAP, only type A CpG-ODN 2216 (34, 35) triggered IFN-� production (Fig. 5A). Unex-pectedly, upon DOTAP-mediated endosomal translocation, murineCpG-ODN 1668-PD, noncanonical CpG-ODN 1720-PD, andmethylated CpG 1668-PD as well as bacterial pDNA and verte-brate DNA, in part, also caused robust release of IFN-� from hu-man PMBC, whereas PTO-ODNs were poor inducers, and controlODN AP-1 was inactive. Dose-response experiments confirmedthat noncanonical PD CpG motifs complexed to DOTAP triggeredrobust IFN-� production comparable to the type A ODN 2216(data not shown). Within human PMBC, only cells positively se-lected for CD123, a marker typifying pDCs (36), produced IFN-�(Fig. 5B). We concluded that DOTAP-mediated endosomal trans-location of murine canonical or noncanonical CpG motifs as well
FIGURE 5. Noncanonical CpG-DNA complexed to DOTAP inducesIFN-� production of human pDC. A, Human PBMC (1.5 � 106 cells/ml)were stimulated with ODN complexed or not complexed to DOTAP. B,CD123-negative or -positive selected cells (2.5 � 105/ml; � and f, re-spectively). Cells were stimulated with ODN, complexed or not complexedto DOTAP. The supernatants were analyzed for their contents of IFN-� byELISA. Error bars represent the SD of duplicate values. The results of oneexperiment (shown) are representative of results for at least three replicateexperiments with different donors.
DNA was mixed at graded concentrations with 10 �g of DOTAP in 50 �lof HBS. After incubation for 15 min at 37°C 100 �l of complete RPMI1640 medium was added. The mixture was added to wt FL-DC cultures(2.5 � 106/ml). One representative experiment of at least three is shown.Error bars represent the SD of duplicate values; the asterisk indicates thatcytokines were not detectable by ELISA.
FIGURE 4. DOTAP-complexed vertebrate DNA or noncanonical CpG-DNA activate DCs. TLR2- and TLR4-deficient or TLR2-, TLR4-, andTLR9-deficient murine FL-DC (2.5 � 106ml) were stimulated as indicated(2 �M). After 18 h, cells were stained with Abs directed to CD69 or CD40.f, Stimulated cells; �, cells in medium only (A). The culture supernatantswere analyzed for IFN-� or IL-6 content by ELISA (B). C, Respective
6133The Journal of Immunology
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

as of vertebrate DNA overcomes the species and sequence restric-tions described for TLR9 activation (reviewed in Refs. 2 and 24),thus allowing murine PD DNA motifs to trigger efficient type IIFN induction in human pDCs.
Vertebrate DNA complexed to DOTAP triggers systemic IL-6production
In mice, systemic delivery of cationic lipid-pDNA complexes in-duces acute, TLR9-dependent inflammatory responses, which inthe context of therapeutic gene delivery represent dose-limitingtoxicities (37). Following up our results in vitro, we exploredwhether systemic delivery of vertebrate DNA or of the noncanoni-cal CpG motif 1720 complexed to DOTAP triggers IL-6 produc-tion in vivo. As shown in Fig. 6, upon i.v. challenge of wt mice,significant amounts of serum borne IL-6, a bona fide marker ofinflammation, were detectable.
DiscussionOur experiments report the surprising finding that upon endosomaltranslocation, both vertebrate DNA and certain noncanonical CpGmotifs cause activation of murine and human DCs via TLR9-de-pendent and -independent pathways. These findings apparentlycontrast with the current paradigm of species- and sequence-spe-cific TLR9 DNA ligand interactions (reviewed in Refs. 1, 2, and24). Nonetheless, our results are consistent with aspects of therecently described immunobiology of chromatin-IgG ICs, as ob-served upon BCR- or Fc�RIII-mediated cellularinternalization (16, 17).
The paradigm of species- and sequence-specific CpG DNA-me-diated immune cell activation via TLR9/MyD88 rests on a largebody of experimental data in which innate immune cells have beenchallenged with CpG motifs (reviewed in Refs. 2 and 24). Undersuch conditions, vertebrate DNA and noncanonical CpG DNA lackstimulatory activities. However, because TLR9 recognizes its li-gands at a late endosomal compartments (reviewed in Ref. 24), wehypothesized that poor endosomal translocation of putative TLR9ligands via the natural uptake pathway might be rate limiting. Ac-cordingly low affinity TLR9 ligands might fail to reach the thresh-old concentrations required for TLR9 activation. Under naturalconditions, endosomal translocation of DNA is brought about by
an as yet ill-defined sequence-nonspecific DNA receptor thatTLR9-independently drives receptor-mediated endocytosis (31,32; reviewed in Ref. 24). In this study we show that this naturalendocytosis pathway is highly saturable, because at 1–2 �M ODN,saturation is reached within 10–20 min (Fig. 1). At present, weexplain this saturation by the loss of cell surface DNA receptorsdue to ligand-driven endocytosis; DOTAP-mediated endosomalDNA translocation is also known to use endocytotic pathways(25), yet, as shown in this study, this endocytotic pathway is notsaturable. As a consequence, up to 10- to 50-fold higher concen-trations of potential TLR9 ligands become translocated. Undersuch conditions, endosomally translocated, noncanonical PTOCpG motifs trigger DC activation in a TLR9- (Figs. 3 and 4) andMyD88 (data not shown)-dependent fashion, whereas controlODNs, such as AP-1 and NAOS-1, do not. At present, we explainthe inactivity of AP-1 or NAOS-1 with the guanine at the 5� po-sition because we observed that modification of, for example,ODN 1720 by addition of a G nucleotide at the 5� and/or 3� endabolishes its stimulatory potential (data not shown). Interestingly,PD DNA, such as vertebrate DNA and bacterial pDNA, displayed,in addition, TLR9-independent immunostimulation. Together,these results support the hypothesis that upon enhanced endosomaltranslocation, low affinity ligands in endosomes can reach thethreshold concentrations required to drive TLR9 activation. Ac-cordingly, our results confine the accepted paradigm of species-and sequence-specific TLR9 DNA ligand interactions to the nat-ural uptake route, because upon DOTAP-mediated enhanced en-dosomal translocation, noncanonical CpG motifs or vertebrateDNA also trigger TLR9 activation. Vertebrate DNA is known tobe rich in noncanonical and methylated CpG motifs (6). We there-fore assume that vertebrate DNA donates sufficient low affinityligands upon fragmentation by DNase within late endosomes, aprocess likely to be facilitated by liposome-driven DNA-helix de-naturation in ssDNA (38). In support of this idea, plasmon reso-nance (Biacore)-based analysis of sequence-dependent TLR9-CpG-ODN interactions revealed low affinity interactions ofnoncanonical CpG-ODN 1720 PD with recombinant TLR9 (39)(data not shown), whereas PTO ODN allowed no analysis of CpGsequence specificity, because it bound nonspecifically even toTLR2 (39). Perhaps the latter observations might explain why PTOODN sequence nonspecifically precipitates TLR9 from cellular ly-sates (15).
Several lines of evidence imply immune stimulatory activities ofvertebrate DNA contingent upon translocation to TLR9-expressingendosomes. For example, a role of self-DNA as an indicator oftissue damage has been suggested, although self-DNA failed totrigger cytokine production (40). Furthermore, mouse embryos de-ficient in DNase II contain macrophages carrying undigested DNAintracellularly, and this causes lethal type I IFN production (41).Perhaps the strongest evidence has been provided by Marshak-Rothstein et al. (16), who described a pathway involved in themeans by which vertebrate DNA within chromatin-IgG ICs enterB cell endosomes via BCRs or enter endosomes of DCs viaFc�RIII, the latter triggering both TLR9-dependent and -indepen-dent cell activation (17). However, Fc�Rs often contain ITAM(42), which may cloud interpretation of cell activation. Our data,therefore, extend the elegant findings of Marshak-Rothstein et al.(16), because we show that mere endosomal translocation is thekey for vertebrate DNA to induce immunostimulation via TLR9-dependent and -independent pathways.
Our report is the first to describe factors that determine initiationof TLR9-dependent vs TLR9/MyD88-independent DC activationpathways. For example, inactivated DNA viruses, such as HSV-1,activate DC solely via TLR9, whereas infectious HSV-1 recruits
FIGURE 6. Non-CpG-DNA and vertebrate DNA complexed to DOTAPtrigger systemic IL-6 release. Serum cytokine levels in mice after systemicdelivery of DNA or DNA complexed to DOTAP. DNA (30 �g) or DNA(30 �g) complexed to DOTAP (60 �g) in 150 �l was injected i.v. into wtmice. Blood was collected 2 h after injection. n � 4 mice (1720 or 1720and DOTAP), 5 mice (CT-DNA or CT-DNA and DOTAP), or 9 mice(AP-1 and DOTAP or DOTAP only) per group. The bar indicates the meanof the data. Differences in cytokine levels in the samples treated with DNAand DNA plus DOTAP were statistically analyzed by Mann-Whitney rank-sum test. �, p � 0.008; #, p � 0.029.
6134 TLR9, NON-CpG DNA, AND DC ACTIVATION
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

an TLR9-independent pathway in addition (33). In this study wereport that a chemical modification of the DNA backbone repre-sents a determining factor for TLR9 dependency, because canon-ical and noncanonical PTO CpG motifs activate DCs via TLR9,whereas PD DNA (as contained in bacterial pDNA or vertebrateDNA) in addition triggers a TLR9/MyD88-independent DC acti-vation pathway. Future studies are needed to analyze the molecularrelatedness of the TLR9-independent pathway with the signalpathway driven by TLR9.
We observed that canonical and noncanonical murine PD CpGmotifs displayed powerful interferonic activities toward humanpDCs. The pioneering work of Alm and Ronnbloom (18, 19)pointed out that inversion of CpG3GpC or methylation of a CpGmotif (identified in serum of systemic lupus erythematosus patients(43)) did not ablate the ability to trigger type I IFN production inhuman pDCs. The latter reports speculated that interferonic DNAsequences may be more frequent in eukaryotic DNA than antici-pated. Indeed, upon enhanced endosomal translocation into humanpDCs, vertebrate DNA, pDNA, and canonical and noncanonicalmurine CpG-ODN PD triggered robust type I IFN production (Fig.5). The magnitude of type I IFN production was equal to theamount triggered by type A CpG-ODN 2216, which is generallyselected for efficient IFN-� induction in pDCs (34, 35). We foundthat type A CpG-ODN is notoriously poor in TLR9-dependentNF-�B activation (unpublished observations) as are murine CpGmotifs toward human TLR9 (4). Therefore, these ligands may becatalogued as low affinity ligands for human TLR9. It is thus pos-sible that activation of pDCs for IFN-� production preferentiallyrequires TLR9 activation via low affinity ligands, whereas highaffinity ligands such as type B CpG-ODN (34, 35) trigger too rap-idly the maturation of pDCs into professional APCs able to secreteTh1-polarizing cytokines such as IL-12 (36, 44).
Because TLR1, -2 and -4 to -6 recognize invariant foreign bac-terial and viral products at the cell membrane, one may ask whyTLR3 and TLR7 to -9 recognize distinct patterns of nucleic acidsat late endosomes (reviewed in Ref. 24). Although in the formercase, self-nonself discrimination can easily be envisaged, struc-tural differences between pathogen- and host-derived nucleic acidsappear less prominent, because mere endosomal translocation ofvertebrate DNA or of noncanonical CpG motifs unveils strongstimulatory activity toward DCs. We therefore propose that it is therestricted accessibility of endosomally expressed TLR9 that hin-ders activation of TLR9-expressing immune cells by host-derivedself-DNA, because the latter is likely to be rapidly degraded viaDNases.
AcknowledgmentsWe acknowledge Cornelia Wagner, Hendrikje Drexler, and Monika Ham-mel for excellent technical assistance, and Meredith O’Keeffe (The Walterand Eliza Hall Institute of Medical Research, Melbourne, Australia) forhelp with DC characterization.
DisclosuresThe authors have no financial conflict of interest.
References1. Wagner, H. 1999. Bacterial CpG DNA activates immune cells to signal infectious
danger. Adv. Immunol. 73: 329–368.2. Krieg, A. M. 2002. CpG motifs in bacterial DNA and their immune effects. Annu.
Rev. Immunol. 20: 709–760.3. Hemmi, H., O. Takeuchi, T. Kawai, T. Kaisho, S. Sato, H. Sanjo, M. Matsumoto,
K. Hoshino, H. Wagner, K. Takeda, et al. 2000. A Toll-like receptor recognizesbacterial DNA. Nature 408: 740–745.
4. Bauer, S., C. J. Kirschning, H. Hacker, V. Redecke, S. Hausmann, S. Akira,H. Wagner, and G. B. Lipford. 2001. Human TLR9 confers responsiveness tobacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci.USA 98: 9237–9242.
5. Krieg, A. M., A. K. Yi, S. Matson, T. J. Waldschmidt, G. A. Bishop, R. Teasdale,G. A. Koretzky, and D. M. Klinman. 1995. CpG motifs in bacterial DNA triggerdirect B-cell activation. Nature 374: 546–549.
6. Stacey, K. J., G. R. Young, F. Clark, D. P. Sester, T. L. Roberts, S. Naik,M. J. Sweet, and D. A. Hume. 2003. The molecular basis for the lack of immu-nostimulatory activity of vertebrate DNA. J. Immunol. 170: 3614–3620.
7. Bird, A. P. 1993. Functions for DNA methylation in vertebrates. Cold SpringHarb. Symp. Quant. Biol. 58: 281–285.
8. Ramsahoye, B. H., C. S. Davies, and K. I. Mills. 1996. DNA methylation: biol-ogy and significance. Blood Rev. 10: 249–261.
9. Krieg, A. M., T. Wu, R. Weeratna, S. M. Efler, L. Love-Homan, L. Yang,A. K. Yi, D. Short, and H. L. Davis. 1998. Sequence motifs in adenoviral DNAblock immune activation by stimulatory CpG motifs. Proc. Natl. Acad. Sci. USA95: 12631–12636.
10. Shirota, H., M. Gursel, and D. M. Klinman. 2004. Suppressive oligodeoxynucle-otides inhibit Th1 differentiation by blocking IFN-�- and IL-12-mediated signal-ing. J. Immunol. 173: 5002–5007.
11. Heil, F., P. Ahmad-Nejad, H. Hemmi, H. Hochrein, F. Ampenberger, T. Gellert,H. Dietrich, G. Lipford, K. Takeda, S. Akira, et al. 2003. The Toll-like receptor7 (TLR7)-specific stimulus loxoribine uncovers a strong relationship within theTLR7, 8 and 9 subfamily. Eur. J. Immunol. 33: 2987–2997.
12. Lee, J., T. H. Chuang, V. Redecke, L. She, P. M. Pitha, D. A. Carson, E. Raz, andH. B. Cottam. 2003. Molecular basis for the immunostimulatory activity of gua-nine nucleoside analogs: activation of Toll-like receptor 7. Proc. Natl. Acad. Sci.USA 100: 6646–6651.
13. Matsumoto, M., K. Funami, M. Tanabe, H. Oshiumi, M. Shingai, Y. Seto,A. Yamamoto, and T. Seya. 2003. Subcellular localization of Toll-like receptor3 in human dendritic cells. J. Immunol. 171: 3154–3162.
14. Ahmad-Nejad, P., H. Hacker, M. Rutz, S. Bauer, R. M. Vabulas, and H. Wagner.2002. Bacterial CpG-DNA and lipopolysaccharides activate Toll-like receptors atdistinct cellular compartments. Eur. J. Immunol. 32: 1958–1968.
15. Latz, E., A. Schoenemeyer, A. Visintin, K. A. Fitzgerald, B. G. Monks,C. F. Knetter, E. Lien, N. J. Nilsen, T. Espevik, and D. T. Golenbock. 2004.TLR9 signals after translocating from the ER to CpG DNA in the lysosome. Nat.Immunol. 5: 190–198.
16. Leadbetter, E. A., I. R. Rifkin, A. M. Hohlbaum, B. C. Beaudette,M. J. Shlomchik, and A. Marshak-Rothstein. 2002. Chromatin-IgG complexesactivate B cells by dual engagement of IgM and Toll-like receptors. Nature 416:603–607.
17. Boule, M. W., C. Broughton, F. Mackay, S. Akira, A. Marshak-Rothstein, andI. R. Rifkin. 2004. Toll-like receptor 9-dependent and -independent dendritic cellactivation by chromatin-immunoglobulin G complexes. J. Exp. Med. 199:1631–1640.
18. Vallin, H., A. Perers, G. V. Alm, and L. Ronnblom. 1999. Anti-double-strandedDNA antibodies and immunostimulatory plasmid DNA in combination mimic theendogenous IFN-� inducer in systemic lupus erythematosus. J. Immunol. 163:6306–6313.
19. Magnusson, M., S. Magnusson, H. Vallin, L. Ronnblom, and G. V. Alm. 2001.Importance of CpG dinucleotides in activation of natural IFN-�-producing cellsby a lupus-related oligodeoxynucleotide. Scand. J. Immunol. 54: 543–550.
20. Wang, Y., and A. M. Krieg. 2004. Induction of autoantibody production but notautoimmune disease in HEL transgenic mice vaccinated with HEL in combina-tion with CpG or control oligodeoxynucleotides. Vaccine 22: 2641–2650.
21. Cornelie, S., O. Poulain-Godefroy, C. Lund, C. Vendeville, E. Ban, M. Capron,and G. Riveau. 2004. Methylated CpG-containing plasmid activates the immunesystem. Scand. J. Immunol. 59: 143–151.
22. Elias, F., J. Flo, R. A. Lopez, J. Zorzopulos, A. Montaner, and J. M. Rodriguez.2003. Strong cytosine-guanosine-independent immunostimulation in humans andother primates by synthetic oligodeoxynucleotides with PyNTTTTGT motifs.J. Immunol. 171: 3697–3704.
23. Vollmer, J., R. D. Weeratna, M. Jurk, U. Samulowitz, M. J. McCluskie,P. Payette, H. L. Davis, C. Schetter, and A. M. Krieg. 2004. Oligodeoxynucle-otides lacking CpG dinucleotides mediate Toll-like receptor 9 dependent T helpertype 2 biased immune stimulation. Immunology 113: 212–223.
24. Wagner, H. 2004. The immunobiology of the TLR9 subfamily. Trends Immunol.25: 381–386.
25. Almofti, M. R., H. Harashima, Y. Shinohara, A. Almofti, Y. Baba, andH. Kiwada. 2003. Cationic liposome-mediated gene delivery: biophysical studyand mechanism of internalization. Arch. Biochem. Biophys. 410: 246–253.
26. Yasuda, K., Y. Ogawa, M. Kishimoto, T. Takagi, M. Hashida, and Takakura Y.2002. Plasmid DNA activates murine macrophages to induce inflammatory cy-tokines in a CpG motif-independent manner by complex formation with cationicliposomes. Biochem. Biophys. Res. Commun. 293: 344–348.
27. Spies, B., H. Hochrein, M. Vabulas, K. Huster, D. H. Busch, F. Schmitz, A. Heit,and H. Wagner. 2003. Vaccination with plasmid DNA activates dendritic cellsvia Toll-like receptor 9 (TLR9) but functions in TLR9-deficient mice. J. Immu-nol. 171: 5908–5912.
28. Heil, F., H. Hemmi, H. Hochrein, F. Ampenberger, C. Kirschning, S. Akira,G. Lipford, H. Wagner, and S. Bauer. 2004. Species-specific recognition of sin-gle-stranded RNA via Toll-like receptor 7 and 8. Science 303: 1526–1529.
29. Diebold, S. S., T. Kaisho, H. Hemmi, S. Akira, and Reis e Sousa. 2004. Innateantiviral responses by means of TLR7-mediated recognition of single-strandedRNA. Science 303: 1529–1531.
30. Hacker, H., H. Mischak, T. Miethke, S. Liptay, R. Schmid, T. Sparwasser,K. Heeg, G. B. Lipford, and H. Wagner. 1998. CpG-DNA-specific activation ofantigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation. EMBO J. 17: 6230–6240.
6135The Journal of Immunology
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

31. Shirota, H., K. Sano, N. Hirasawa, T. Terui, K. Ohuchi, T. Hattori, andG. Tamura. 2002. B cells capturing antigen conjugated with CpG oligode-oxynucleotides induce Th1 cells by elaborating IL-12. J. Immunol. 169: 787–794.
32. Heit, A., T. Maurer, H. Hochrein, S. Bauer, K. M. Huster, D. H. Busch, andH. Wagner. 2003. Cutting edge: Toll-like receptor 9 expression is not required forCpG DNA-aided cross-presentation of DNA-conjugated antigens but essential forcross-priming of CD8 T cells. J. Immunol. 170: 2802–2805.
33. Hochrein, H., B. Schlatter, M. O’Keeffe, C. Wagner, F. Schmitz, M. Schiemann,S. Bauer, M. Suter, and H. Wagner. 2004. Herpes simplex virus type-1 inducesIFN-� production via Toll-like receptor 9-dependent and -independent pathways.Proc. Natl. Acad. Sci. USA 101: 11416–11421.
34. Krug, A., S. Rothenfusser, V. Hornung, B. Jahrsdorfer, S. Blackwell,Z. K. Ballas, S. Endres, A. M. Krieg, and G. Hartmann. 2001. Identification ofCpG oligonucleotide sequences with high induction of IFN-�/� in plasmacytoiddendritic cells. Eur. J. Immunol. 31: 2154–2163.
35. Verthelyi, D., K. J. Ishii, M. Gursel, F. Takeshita, and D. M. Klinman. 2001.Human peripheral blood cells differentially recognize and respond to two distinctCPG motifs. J. Immunol. 166: 2372–2377.
36. Bauer, M., V. Redecke, J. W. Ellwart, B. Scherer, J. P. Kremer, H. Wagner, andG. B. Lipford. 2001. Bacterial CpG-DNA triggers activation and maturation ofhuman CD11c�, CD123� dendritic cells. J. Immunol. 166: 5000–5007.
37. Zhao, H., H. Hemmi, S. Akira, S. H. Cheng, R. K. Scheule, and N. S. Yew. 2004.Contribution of Toll-like receptor 9 signaling to the acute inflammatory responseto nonviral vectors. Mol. Ther. 9: 241–248.
38. Prasad, T. K., V. Gopal, and N. M. Rao. 2003. Cationic lipids and cationic ligandsinduce DNA helix denaturation: detection of single stranded regions by KMnO4probing. FEBS Lett. 552: 199–206.
39. Rutz, M., J. Metzger, T. Gellert, P. Luppa, G. B. Lipford, H. Wagner, andS. Bauer. 2004. Toll-like receptor 9 binds single-stranded CpG-DNA in a se-quence- and pH-dependent manner. Eur. J. Immunol. 34: 2541–2550.
40. Ishii, K. J., K. Suzuki, C. Coban, F. Takeshita, Y. Itoh, H. Matoba, L. D. Kohn,and D. M. Klinman. 2001. Genomic DNA released by dying cells induces thematuration of APCs. J. Immunol. 167: 2602–2607.
41. Yoshida, H., Y. Okabe, K. Kawane, H. Fukuyama, and S. Nagata. 2005. Lethalanemia caused by interferon-� produced in mouse embryos carrying undigestedDNA. Nat. Immunol. 6: 49–56.
42. Takai, T. 2002. Roles of Fc receptors in autoimmunity. Nat. Rev. Immunol. 2:580–592.
43. Sato, Y., M. Miyata, Y. Sato, T. Nishimaki, H. Kochi, and R. Kasukawa. 1999.CpG motif-containing DNA fragments from sera of patients with systemic lupuserythematosus proliferate mononuclear cells in vitro. J. Rheumatol. 26: 294–301.
44. Boonstra, A., C. Asselin-Paturel, M. Gilliet, C. Crain, G. Trinchieri, Y. J. Liu, andA. O’Garra. 2003. Flexibility of mouse classical and plasmacytoid-derived den-dritic cells in directing T helper type 1 and 2 cell development: dependency onantigen dose and differential Toll-like receptor ligation. J. Exp. Med. 197:101–109.
6136 TLR9, NON-CpG DNA, AND DC ACTIVATION
by guest on August 1, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











