Endocrine modulating actions of a phytosterol mixture and its oxidation products in zebrafish (Danio rerio) Inga Christianson-Heiska a, ⁎ , Pia Smeds a , Nina Granholm b , Eija Bergelin b , Boris Isomaa a a Department of Biology, Åbo Akademi University, BioCity, Artillerigatan 6A, FIN-20520 Åbo, Finland b Laboratory of Wood and Paper Chemistry, Åbo Akademi University, Porthansgatan 3, Gadolinia, FIN-20500 Åbo, Finland Received 22 September 2006; received in revised form 9 January 2007; accepted 9 January 2007 Available online 22 February 2007 Abstract In this study, a phytosterol preparation (“ultrasitosterol”; 80% β-sitosterol) and an oxidized ultrasitosterol preparation were evaluated for reproductive effects in zebrafish. Adults were exposed in a continuous flow to 10 μg/L and 100 μg/L ultrasitosterol and oxidized ultrasitosterol, and to 0.27 μg/L 17β-estradiol and 0.28 μg/L testosterone for 3 weeks. Biomarkers analysed included plasma vitellogenin, testosterone, 11- ketotestosterone, 17β-estradiol, and gonadal histopathology. Ovarian steroid production of testosterone and 17β-estradiol was examined in isolated zebrafish follicles exposed in vitro to the compounds at the same concentrations as in vivo. Vtg was induced in males exposed to ultrasitosterol, and in males and females exposed to 17β-estradiol. Males exposed to oxidized phytosterols showed increased levels of testosterone and 11-ketotestosterone, and accelerated spermatogenesis. Increased follicular atresia was observed in females exposed to oxidized phytosterols and 17β-estradiol. Correlation analyses between biomarkers revealed more intercorrelated values for females than for males, and the strongest associations were found in females exposed to oxidized phytosterols. Testosterone production was significantly increased in follicles exposed to the oxidized phytosterol preparations. These findings indicate that the phytosterol mixture is weakly estrogenic in male fish, and that the oxidized phytosterol mixture contains substances that may interfere with spermatogenesis, oogenesis and gonadal steroidogenesis in zebrafish. © 2007 Elsevier Inc. All rights reserved. Keywords: Zebrafish; Phytosterol; β-sitosterol; Oxidized phytosterol; Vitellogenin; Sex steroid; Gonad histology 1. Introduction Exposure of fish populations to pulp and paper mill effluents has been associated with a variety of negative impacts on reproductive fitness including reduced ovarian development, delayed sexual maturation and alterations in reproductive endocrine homeostasis at multiple sites along the pituitary–gonadal axis ( Van Der Kraak et al., 1992; Munkittrick et al., 1998). Pulp mill effluents are known to contain a mixture of natural compounds from processed wood, chemicals arising from the pulping and bleaching processes, and compounds formed by microbial action in secondary treatment ponds. β-sitosterol, a lipophilic plant sterol found in both softwood and hardwood, is one of the major wood extractives in pulp and paper mill effluents (Cook et al., 1997). β-sitosterol exhibits estrogenic activity by means of binding to the estrogen receptor and promoting vitellogenin (Vtg) production in fish hepatocytes in vitro (Tremblay and Van der Kraak, 1998), and in fish in vivo by induction of the Vtg gene and increasing plasma Vtg (Mellanen et al., 1996; Tremblay and Van der Kraak, 1998, 1999). Further, β-sitosterol reduces plasma sex steroid and cholesterol levels in fish (MacLatchy and Van der Kraak, 1995; MacLatchy et al., 1997; Tremblay and Van der Kraak, 1999) and interferes with the gonadal steroid biosynthetic capacity (MacLatchy and Van der Kraak, 1995), which has later been linked to modulation of cholesterol availability (MacLatchy et al., 1997; Leusch and MacLatchy, 2003). Ef- fects on fish embryo development and changes in sex ratios have been reported after exposure to a mixture of phytosterols (Lehtinen et al., 1999; Mattsson et al., 2001; Nakari and Erkomaa, 2003; Honkanen et al., 2005), and masculinization has been demonstrated in mosquitofish (Gambusia affinis) Comparative Biochemistry and Physiology, Part C 145 (2007) 518 – 527 www.elsevier.com/locate/cbpc ⁎ Corresponding author. Tel.: +358 2 2154065; fax: +358 2 2154748. E-mail address: [email protected] (I. Christianson-Heiska). 1532-0456/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpc.2007.01.016

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

gy, Part C 145 (2007) 518–527www.elsevier.com/locate/cbpc
Comparative Biochemistry and Physiolo
Endocrine modulating actions of a phytosterol mixture and itsoxidation products in zebrafish (Danio rerio)
Inga Christianson-Heiska a,⁎, Pia Smeds a, Nina Granholm b, Eija Bergelin b, Boris Isomaa a
a Department of Biology, Åbo Akademi University, BioCity, Artillerigatan 6A, FIN-20520 Åbo, Finlandb Laboratory of Wood and Paper Chemistry, Åbo Akademi University, Porthansgatan 3, Gadolinia, FIN-20500 Åbo, Finland
Received 22 September 2006; received in revised form 9 January 2007; accepted 9 January 2007Available online 22 February 2007
Abstract
In this study, a phytosterol preparation (“ultrasitosterol”; 80% β-sitosterol) and an oxidized ultrasitosterol preparation were evaluated forreproductive effects in zebrafish. Adults were exposed in a continuous flow to 10 μg/L and 100 μg/L ultrasitosterol and oxidized ultrasitosterol,and to 0.27 μg/L 17β-estradiol and 0.28 μg/L testosterone for 3 weeks. Biomarkers analysed included plasma vitellogenin, testosterone, 11-ketotestosterone, 17β-estradiol, and gonadal histopathology. Ovarian steroid production of testosterone and 17β-estradiol was examined inisolated zebrafish follicles exposed in vitro to the compounds at the same concentrations as in vivo. Vtg was induced in males exposed toultrasitosterol, and in males and females exposed to 17β-estradiol. Males exposed to oxidized phytosterols showed increased levels of testosteroneand 11-ketotestosterone, and accelerated spermatogenesis. Increased follicular atresia was observed in females exposed to oxidized phytosterolsand 17β-estradiol. Correlation analyses between biomarkers revealed more intercorrelated values for females than for males, and the strongestassociations were found in females exposed to oxidized phytosterols. Testosterone production was significantly increased in follicles exposed tothe oxidized phytosterol preparations. These findings indicate that the phytosterol mixture is weakly estrogenic in male fish, and that the oxidizedphytosterol mixture contains substances that may interfere with spermatogenesis, oogenesis and gonadal steroidogenesis in zebrafish.© 2007 Elsevier Inc. All rights reserved.
Keywords: Zebrafish; Phytosterol; β-sitosterol; Oxidized phytosterol; Vitellogenin; Sex steroid; Gonad histology
1. Introduction
Exposure of fish populations to pulp and paper milleffluents has been associated with a variety of negativeimpacts on reproductive fitness including reduced ovariandevelopment, delayed sexual maturation and alterations inreproductive endocrine homeostasis at multiple sites along thepituitary–gonadal axis (Van Der Kraak et al., 1992;Munkittrick et al., 1998). Pulp mill effluents are known tocontain a mixture of natural compounds from processedwood, chemicals arising from the pulping and bleachingprocesses, and compounds formed by microbial action insecondary treatment ponds. β-sitosterol, a lipophilic plantsterol found in both softwood and hardwood, is one of themajor wood extractives in pulp and paper mill effluents (Cook
⁎ Corresponding author. Tel.: +358 2 2154065; fax: +358 2 2154748.E-mail address: [email protected] (I. Christianson-Heiska).
1532-0456/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpc.2007.01.016
et al., 1997). β-sitosterol exhibits estrogenic activity bymeans of binding to the estrogen receptor and promotingvitellogenin (Vtg) production in fish hepatocytes in vitro(Tremblay and Van der Kraak, 1998), and in fish in vivo byinduction of the Vtg gene and increasing plasma Vtg(Mellanen et al., 1996; Tremblay and Van der Kraak, 1998,1999). Further, β-sitosterol reduces plasma sex steroid andcholesterol levels in fish (MacLatchy and Van der Kraak,1995; MacLatchy et al., 1997; Tremblay and Van der Kraak,1999) and interferes with the gonadal steroid biosyntheticcapacity (MacLatchy and Van der Kraak, 1995), which haslater been linked to modulation of cholesterol availability(MacLatchy et al., 1997; Leusch and MacLatchy, 2003). Ef-fects on fish embryo development and changes in sex ratioshave been reported after exposure to a mixture of phytosterols(Lehtinen et al., 1999; Mattsson et al., 2001; Nakari andErkomaa, 2003; Honkanen et al., 2005), and masculinizationhas been demonstrated in mosquitofish (Gambusia affinis)

519I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
exposed to degradation products of β-sitosterol (Howell andDenton, 1989). Chemical processes during pulp bleachingmay also produce other bioactive compounds, whichconsequently may be present in the effluent water. To ourknowledge such oxidation products of wood extractives arequite poorly studied for their toxicity on the aquatic biota,particularly regarding endocrine disrupting activities. Onestudy reports decreased testosterone and 11-ketotestosteronelevels in goldfish (Carassius auratus) exposed to an oxidizedβ-sitosterol preparation (MacLatchy and Van der Kraak,1995); in another study no significant effects were observedregarding masculinization of female mosquitofish exposed tooxidized plant sterols (Van den Heuvel et al., 2006). In thesame study, the preparations were tested using in vitrobioassays and suggested to be unlikely to act through estro-genic or androgenic mechanisms.
To further investigate possible endocrine disrupting effectsof phytosterols and their oxidation products, we exposed adultzebrafish to a β-sitosterol preparation (“ultrasitosterol”) as wellas a preparation of its oxidation products. Endpoints measuredincluded gonad histopathology, plasma vitellogenin and plasmasex steroids. 17β-estradiol and testosterone were selected asreference compounds for estrogenic vs. androgenic effects.Effects of the preparations on ovarian in vitro steroidbiosynthetic capacity were also investigated.
2. Materials and methods
2.1. Chemicals
Ultrasitosterol (ca. 80% β-sitosterol and smaller percentagesof β-sitostanol, campesterol and campestanol) was provided byKaukas UPM Kymmene pulp and paper mill (Kaukas, Finland).17-beta-estradiol (E2), testosterone (T), aprotinin (5.1 TIU/mgsolid), goat anti-rabbit IgG horseradish peroxidase conjugatedantibody, O-phenylene diamine (OPD), penicillin+streptomycinsolution (P+S) and human chorionic gonadotrophin (hCG) werepurchased from Sigma-Aldrich Chemical Co. (St Louis, MO,USA). L-15 culture medium was purchased from Gibco. Tween20 was purchased from Bio-Rad (Burlingame, CA, USA) andcalcein-AM from Molecular Probes. Polyclonal zebrafishvitellogenin antibodies and zebrafish Vtg standard were pur-chased from Biosense Laboratories AS (Bergen, Norway).Heneicosanoic acid, cholesterol, cholesteryl heptadecanoate and1,3-dipalmitoyl-2-oleoylglycerol, used as internal standard com-pounds for the GC analysis, were purchased from Fluka.N,O-bis-trimethylsilyl-trifluoracetamide (BSTFA) and trimethylchlorosi-lane (TMCS), used as silylation reagents, were from Fluka andAcros. All other chemicals used were commercial chemicals ofreagent grade.
2.2. Oxidation and analysis of sterols
Ultrasitosterol was oxidized according to themethod describedby Bergelin and Holmbom (2005). Shortly, oxidation ofultrasitosterol was performed with ClO2 at 70 °C for 12 h inacidic media. pH was adjusted to 2.5 with phosphoric acid. The
reactions were stopped with the addition of 10% KI solution andthe supernatant was separated. The products from the oxidationandwater samples from exposures were analysed byGCwith FIDdetection. Internal standard components were added to thesamples. After evaporation the extract aliquots were silylatedwith 80μLBSTFA and 40μLTMCS.GCanalysiswas performedon a short column (HP-1, 5 m, 0.53 mm i.d., film thickness 0.15),which enables the determination of fatty and resin acids andsterols as component groups, as well as steryl esters andtriglycerides. The initial temperature gradient was 12 °C/min.The injection temperature was 80 °C and detection temperature340 °C. The individual resin components and model compoundswere also analysed by GC using a HP-1 column (25 m, 0.20 mmi.d.) and FID detection. The initial temperature was 150 °C andthe temperature gradient was 7 °C/min. The injection temperaturewas 230 °C and detection temperature 290 °C. GC–MSwas usedfor verification of GC peaks only.
2.3. Experimental setup
Zebrafish (Danio rerio) of both sexes at an age of 4–6monthswere obtained from a Finnish breeding stock, bought from alocal supplier. A 2-week acclimatization period preceded theexposure period, during which the health status of the fish wasassessed. The experiment was run in two replicates using fishfrom the same stock. 15 females and 15 males, reproductivelymature, were transferred to each of the seven 22 L glass aquariaresulting in a fish loading of about 0.6 g fish/L. The tanksreceived 600 mL/h continuously flowing water delivered by aperistaltic pump from barrels, corresponding to one exchange ofthe water volume within 24 h. A 3-week exposure periodcommenced immediately after the acclimatization period. Theaquaria received seven different treatments: control (0.0024%ethanol), 10 μg/L and 100 μg/L ultrasitosterol (U10, U100),10 μg/L and 100 μg/L oxidized ultrasitosterol (OX10, OX100),0.28 μg/L (1 nM) E2 and 0.29 μg/L (1 nM) T. Each test chemicalwas added to 20 Lwell water (pH 7.1–7.5; GH 10–12; KH ca. 7;conductivity 400–450 μS) and 20 L Reverse Osmose water as astock solution prepared by dissolution in ethanol. Each tankreceived aeration, and the escape of aerosols from the tanks wasprevented by glass lids. A photoperiod of 14-h light and 10-h darkwas provided from two 60W fluorescent bulbs. The fish were feda mixture of Tetramin (Tetrawerke, Germany) twice daily, and thedry food was supplemented with white mosquito larvae once aday. The quantity of feed corresponded to the amount ingested bythe fish in 5 min. The aquaria were cleaned completely everysecond or third day. Oxygen content, temperature, and pH of theaquaria were checked twice per week. The temperature was 25.6±0.5 (SD), pH 7.3±0.1 (SD), and oxygen content above 60% forboth replicates.
2.4. Sample collection
At the end of the exposure period, all the fish were killedduring a three-day period. To ensure comparability for hormoneanalysis, blood samples were collected at the same time eachday. Fish were killed in an ice bath, measured for total length

Table 1Measured concentrations of sterols in water samples (μg/L) from different exposures: 10 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidizedmixture of ultrasitosterol (OX10), and 100 μg/L (OX100), oxidized mixture of ultrasitosterol
Sitosterol Stigma-7-en-3-ol 7-hydroxysitosterol 4-hydroxysitosterol 7-oxo-sitosterol Sterols total
U10 9.5±1.2 9.5±1.2U100 35.7±12.3 35.7±12.3OX10 7.7±3.0 0.6±0.13 5.0±2.5 1.0±0.35 1.3±1.2 15.7±5.9OX100 14.6±4.2 6.9±6.5 10.9±1.6 1.3±0.8 1.6±0.6 32.1±1.1
Values represent average±SD from three different water samples.
520 I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
and wet weight to the nearest 0.01 g. Blood was sampled fromeach fish by severing the caudal peduncle with a razor blade andcollecting the blood in a heparinized hematocrit tube. The bloodsamples were centrifuged with a hematocrit centrifuge (2 min),after which the plasma was removed and transferred to anEppendorf tube containing 1 μL aprotinin (2 mg/L). Thistechnique yielded 1–10 μL of plasma per fish. The tubes werethen quickly frozen and stored in −70 °C. In the calculations ofsteroid concentrations the values were corrected for the amountof aprotinin added to the plasma samples.
2.5. Histological examination
After blood sampling, 10 males and 10 females wererandomly chosen from each treatment for histological exami-nation. The peritoneal cavity was cut open and the fish werepreserved in 4% formalin fixative. After 24-hour fixation, thefish were washed twice with 70% ethanol and decalcified withModified Perenyi's decalcifying fluid (0.5% solution ofchromic acid, 10% solution of nitric acid and 30% absolutealcohol) for 3 h. After automatic tissue processing, each fishwas embedded in paraffin. Serial sections (5 μm) were cuthorizontally at various levels, and stained with Delafield'shematoxylin (2%) and eosin (0.5%). The histological alterationsof testes were assessed quantitatively by counting thespermatogenic cysts in ten randomly selected lobules fromdifferent locations of the gonad, after which percentages of thedevelopmental stages were calculated. Following three sper-matogenic stages were identified according to Nagahama(1983): spermatogonia (both type A and type B), spermatocytes
Table 2Average percentage of testicular spermatogenic stages (SG = spermatogonia, SC = speoocyte, VO = vitellogenic oocyte, MO = mature oocyte) in zebrafish exposed to: 1mixture of ultrasitosterol (OX10), and 100 μg/L (OX100), oxidized mixture of ultra
Control U10 U100 OX10
MalesSG 21.3±3.4 (19) 20.8±4.5 (20) 20.2±3.6 (22) 18.3±SC 61.2±3.8 (19) 62.3±4.5 (20) 63.5±6.4 (22) 66.4±SP 17.5±3.3 (19) 16.9±3.0 (20) 16.3±3.7 (22) 15.3±
FemalesPO 61.7±10.5 (20) 62.1±14.0 (17) 65.6±8.8 (16) 63.0±VO 23.4±5.9 (20) 22.4±7.3 (17) 19.3±2.7 (16) 20.6±MO 14.9±6.2 (20) 15.5±11.7 (17) 15.1±8.7 (16) 16.4±
⁎ and ⁎⁎⁎ denote significant difference from control values at pb0.05 and pb0.001
(both type A and type B), and spermatides. Ovaries wereevaluated by counting oocytes on ten randomly selectedmicroscopic areas from different locations of the ovary, afterwhich the percentages of follicular stages observed werecalculated. Three follicular stages were identified: previtello-genic oocyte, vitellogenic oocyte (yolk vesicles present), matureoocyte (follicle bordered by an egg membrane) and additionallyatretic (degenerated) oocytes.
2.6. Vitellogenin assay
Plasma samples of both male and female zebrafish wereanalysed for Vtg using a direct enzyme-linked immunosorbentassay (ELISA). Purified zebrafish vitellogenin was used asstandard (Biosense Laboratories, Bergen, Norway). The assaydetection limit for Vtg was 1.0 ng/mL. Plasma samples frommale fish were diluted 1:2500 and female fish 1:5000 incoating buffer on immunoassay plates in duplicates for eachfish, and incubated overnight at 4 °C. The plates were blockedwith 1.0% non-fat dry milk in PBS for 1 h at roomtemperature, and incubated with the primary antibody(polyclonal rabbit anti-zebrafish Vtg diluted 1:500 in PBS+0.05% Tween 20 (PBS-T)) for 2 h at 37 °C. The secondaryantibody (goat anti-rabbit IgG diluted 1:3000 in PBS-T) wasadded and incubated for 1.5 h at room temperature. Betweeneach step the plates were washed three times with PBS-T.OPD substrate solution was added to the wells, and thereaction was developed for 30 min and stopped by theaddition of 3 M HCl. The plates were measured at 492 nm ona microtiter plate reader.
rmatocytes, SP = spermatids) and ovarian follicular stages (PO = previtellogenic0 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidizedsitosterol
OX100 T E2
3.9 (21) 18.0±4.6 (20) 19.1±3.2 (20) 13.2±4.4 (9)⁎⁎⁎
5.4 (21)⁎ 66.0±5.6 (20)⁎ 64.4±4.8 (20) 70.2±6.5 (9)⁎⁎⁎
3.0 (21) 16.0±3.8 (20) 16.5±4.0 (20) 16.6±6.7 (9)
8.8 (20) 61.9±13.0 (20) 62.0±14.8 (16) 60.8±7.7 (10)6.3 (20) 22.3±5.7 (20) 23.8±6.4 (16) 21.6±5.8 (10)6.7 (20) 15.8±8.5 (20) 14.2±8.9 (16) 17.6±8.1 (10)
.
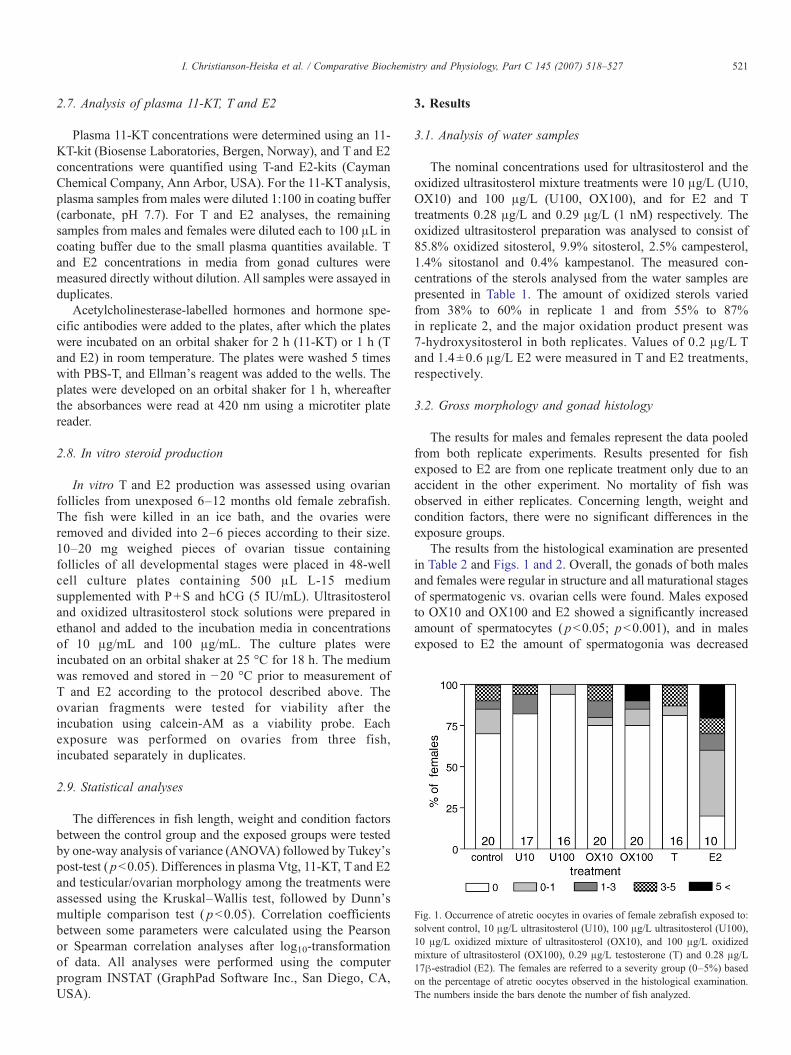
Fig. 1. Occurrence of atretic oocytes in ovaries of female zebrafish exposed to:solvent control, 10 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100),10 μg/L oxidized mixture of ultrasitosterol (OX10), and 100 μg/L oxidizedmixture of ultrasitosterol (OX100), 0.29 μg/L testosterone (T) and 0.28 μg/L17β-estradiol (E2). The females are referred to a severity group (0–5%) basedon the percentage of atretic oocytes observed in the histological examination.The numbers inside the bars denote the number of fish analyzed.
521I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
2.7. Analysis of plasma 11-KT, T and E2
Plasma 11-KT concentrations were determined using an 11-KT-kit (Biosense Laboratories, Bergen, Norway), and T and E2concentrations were quantified using T-and E2-kits (CaymanChemical Company, Ann Arbor, USA). For the 11-KT analysis,plasma samples from males were diluted 1:100 in coating buffer(carbonate, pH 7.7). For T and E2 analyses, the remainingsamples from males and females were diluted each to 100 μL incoating buffer due to the small plasma quantities available. Tand E2 concentrations in media from gonad cultures weremeasured directly without dilution. All samples were assayed induplicates.
Acetylcholinesterase-labelled hormones and hormone spe-cific antibodies were added to the plates, after which the plateswere incubated on an orbital shaker for 2 h (11-KT) or 1 h (Tand E2) in room temperature. The plates were washed 5 timeswith PBS-T, and Ellman's reagent was added to the wells. Theplates were developed on an orbital shaker for 1 h, whereafterthe absorbances were read at 420 nm using a microtiter platereader.
2.8. In vitro steroid production
In vitro T and E2 production was assessed using ovarianfollicles from unexposed 6–12 months old female zebrafish.The fish were killed in an ice bath, and the ovaries wereremoved and divided into 2–6 pieces according to their size.10–20 mg weighed pieces of ovarian tissue containingfollicles of all developmental stages were placed in 48-wellcell culture plates containing 500 μL L-15 mediumsupplemented with P+S and hCG (5 IU/mL). Ultrasitosteroland oxidized ultrasitosterol stock solutions were prepared inethanol and added to the incubation media in concentrationsof 10 μg/mL and 100 μg/mL. The culture plates wereincubated on an orbital shaker at 25 °C for 18 h. The mediumwas removed and stored in −20 °C prior to measurement ofT and E2 according to the protocol described above. Theovarian fragments were tested for viability after theincubation using calcein-AM as a viability probe. Eachexposure was performed on ovaries from three fish,incubated separately in duplicates.
2.9. Statistical analyses
The differences in fish length, weight and condition factorsbetween the control group and the exposed groups were testedby one-way analysis of variance (ANOVA) followed by Tukey'spost-test ( pb0.05). Differences in plasma Vtg, 11-KT, T and E2and testicular/ovarian morphology among the treatments wereassessed using the Kruskal–Wallis test, followed by Dunn'smultiple comparison test ( pb0.05). Correlation coefficientsbetween some parameters were calculated using the Pearsonor Spearman correlation analyses after log10-transformationof data. All analyses were performed using the computerprogram INSTAT (GraphPad Software Inc., San Diego, CA,USA).
3. Results
3.1. Analysis of water samples
The nominal concentrations used for ultrasitosterol and theoxidized ultrasitosterol mixture treatments were 10 μg/L (U10,OX10) and 100 μg/L (U100, OX100), and for E2 and Ttreatments 0.28 μg/L and 0.29 μg/L (1 nM) respectively. Theoxidized ultrasitosterol preparation was analysed to consist of85.8% oxidized sitosterol, 9.9% sitosterol, 2.5% campesterol,1.4% sitostanol and 0.4% kampestanol. The measured con-centrations of the sterols analysed from the water samples arepresented in Table 1. The amount of oxidized sterols variedfrom 38% to 60% in replicate 1 and from 55% to 87%in replicate 2, and the major oxidation product present was7-hydroxysitosterol in both replicates. Values of 0.2 μg/L Tand 1.4±0.6 μg/L E2 were measured in T and E2 treatments,respectively.
3.2. Gross morphology and gonad histology
The results for males and females represent the data pooledfrom both replicate experiments. Results presented for fishexposed to E2 are from one replicate treatment only due to anaccident in the other experiment. No mortality of fish wasobserved in either replicates. Concerning length, weight andcondition factors, there were no significant differences in theexposure groups.
The results from the histological examination are presentedin Table 2 and Figs. 1 and 2. Overall, the gonads of both malesand females were regular in structure and all maturational stagesof spermatogenic vs. ovarian cells were found. Males exposedto OX10 and OX100 and E2 showed a significantly increasedamount of spermatocytes ( pb0.05; pb0.001), and in malesexposed to E2 the amount of spermatogonia was decreased
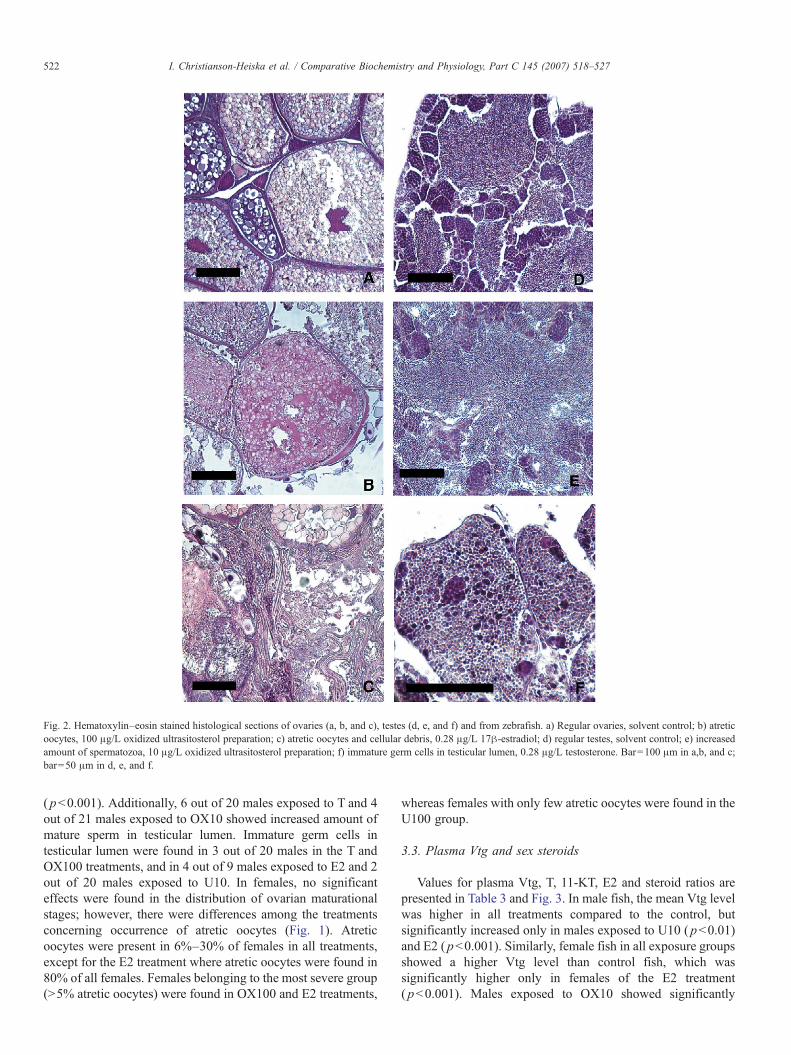
Fig. 2. Hematoxylin–eosin stained histological sections of ovaries (a, b, and c), testes (d, e, and f) and from zebrafish. a) Regular ovaries, solvent control; b) atreticoocytes, 100 μg/L oxidized ultrasitosterol preparation; c) atretic oocytes and cellular debris, 0.28 μg/L 17β-estradiol; d) regular testes, solvent control; e) increasedamount of spermatozoa, 10 μg/L oxidized ultrasitosterol preparation; f) immature germ cells in testicular lumen, 0.28 μg/L testosterone. Bar=100 μm in a,b, and c;bar=50 μm in d, e, and f.
522 I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
( pb0.001). Additionally, 6 out of 20 males exposed to T and 4out of 21 males exposed to OX10 showed increased amount ofmature sperm in testicular lumen. Immature germ cells intesticular lumen were found in 3 out of 20 males in the T andOX100 treatments, and in 4 out of 9 males exposed to E2 and 2out of 20 males exposed to U10. In females, no significanteffects were found in the distribution of ovarian maturationalstages; however, there were differences among the treatmentsconcerning occurrence of atretic oocytes (Fig. 1). Atreticoocytes were present in 6%–30% of females in all treatments,except for the E2 treatment where atretic oocytes were found in80% of all females. Females belonging to the most severe group(N5% atretic oocytes) were found in OX100 and E2 treatments,
whereas females with only few atretic oocytes were found in theU100 group.
3.3. Plasma Vtg and sex steroids
Values for plasma Vtg, T, 11-KT, E2 and steroid ratios arepresented in Table 3 and Fig. 3. In male fish, the mean Vtg levelwas higher in all treatments compared to the control, butsignificantly increased only in males exposed to U10 ( pb0.01)and E2 ( pb0.001). Similarly, female fish in all exposure groupsshowed a higher Vtg level than control fish, which wassignificantly higher only in females of the E2 treatment( pb0.001). Males exposed to OX10 showed significantly
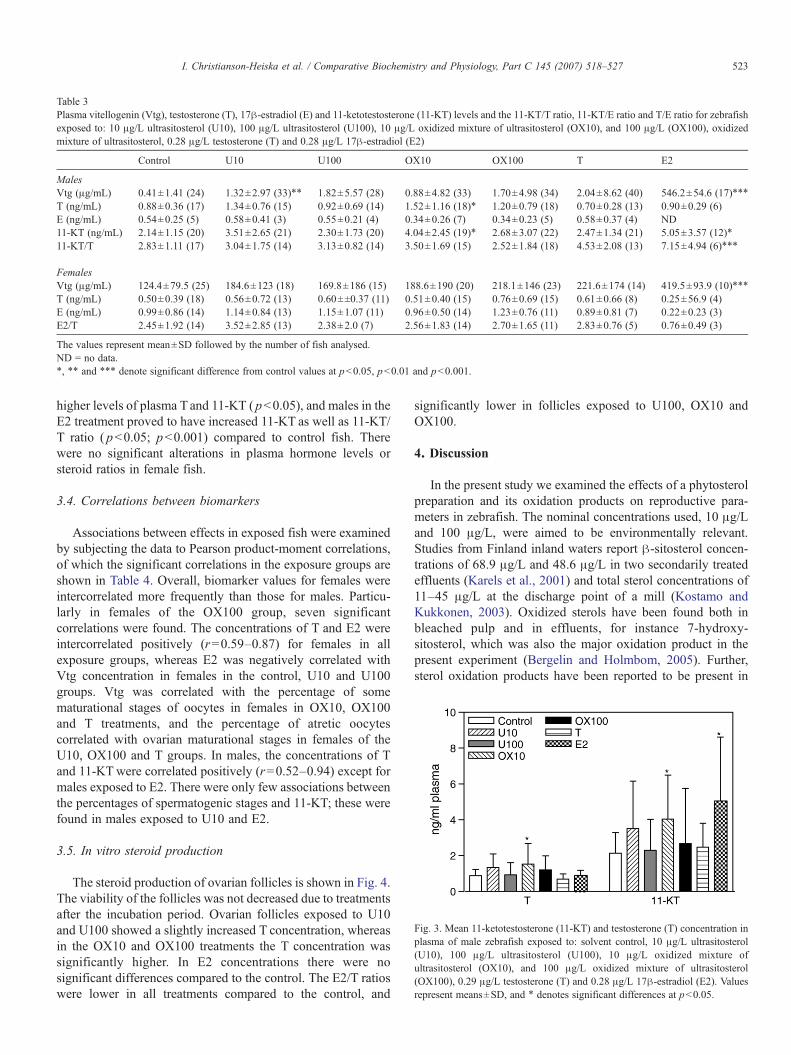
Table 3Plasma vitellogenin (Vtg), testosterone (T), 17β-estradiol (E) and 11-ketotestosterone (11-KT) levels and the 11-KT/T ratio, 11-KT/E ratio and T/E ratio for zebrafishexposed to: 10 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidized mixture of ultrasitosterol (OX10), and 100 μg/L (OX100), oxidizedmixture of ultrasitosterol, 0.28 μg/L testosterone (T) and 0.28 μg/L 17β-estradiol (E2)
Control U10 U100 OX10 OX100 T E2
MalesVtg (μg/mL) 0.41±1.41 (24) 1.32±2.97 (33)⁎⁎ 1.82±5.57 (28) 0.88±4.82 (33) 1.70±4.98 (34) 2.04±8.62 (40) 546.2±54.6 (17)⁎⁎⁎
T (ng/mL) 0.88±0.36 (17) 1.34±0.76 (15) 0.92±0.69 (14) 1.52±1.16 (18)⁎ 1.20±0.79 (18) 0.70±0.28 (13) 0.90±0.29 (6)E (ng/mL) 0.54±0.25 (5) 0.58±0.41 (3) 0.55±0.21 (4) 0.34±0.26 (7) 0.34±0.23 (5) 0.58±0.37 (4) ND11-KT (ng/mL) 2.14±1.15 (20) 3.51±2.65 (21) 2.30±1.73 (20) 4.04±2.45 (19)⁎ 2.68±3.07 (22) 2.47±1.34 (21) 5.05±3.57 (12)⁎
11-KT/T 2.83±1.11 (17) 3.04±1.75 (14) 3.13±0.82 (14) 3.50±1.69 (15) 2.52±1.84 (18) 4.53±2.08 (13) 7.15±4.94 (6)⁎⁎⁎
FemalesVtg (μg/mL) 124.4±79.5 (25) 184.6±123 (18) 169.8±186 (15) 188.6±190 (20) 218.1±146 (23) 221.6±174 (14) 419.5±93.9 (10)⁎⁎⁎
T (ng/mL) 0.50±0.39 (18) 0.56±0.72 (13) 0.60±±0.37 (11) 0.51±0.40 (15) 0.76±0.69 (15) 0.61±0.66 (8) 0.25±56.9 (4)E (ng/mL) 0.99±0.86 (14) 1.14±0.84 (13) 1.15±1.07 (11) 0.96±0.50 (14) 1.23±0.76 (11) 0.89±0.81 (7) 0.22±0.23 (3)E2/T 2.45±1.92 (14) 3.52±2.85 (13) 2.38±2.0 (7) 2.56±1.83 (14) 2.70±1.65 (11) 2.83±0.76 (5) 0.76±0.49 (3)
The values represent mean±SD followed by the number of fish analysed.ND = no data.⁎, ⁎⁎ and ⁎⁎⁎ denote significant difference from control values at pb0.05, pb0.01 and pb0.001.
Fig. 3. Mean 11-ketotestosterone (11-KT) and testosterone (T) concentration inplasma of male zebrafish exposed to: solvent control, 10 μg/L ultrasitosterol(U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidized mixture ofultrasitosterol (OX10), and 100 μg/L oxidized mixture of ultrasitosterol(OX100), 0.29 μg/L testosterone (T) and 0.28 μg/L 17β-estradiol (E2). Valuesrepresent means±SD, and ⁎ denotes significant differences at pb0.05.
523I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
higher levels of plasma Tand 11-KT ( pb0.05), and males in theE2 treatment proved to have increased 11-KT as well as 11-KT/T ratio ( pb0.05; pb0.001) compared to control fish. Therewere no significant alterations in plasma hormone levels orsteroid ratios in female fish.
3.4. Correlations between biomarkers
Associations between effects in exposed fish were examinedby subjecting the data to Pearson product-moment correlations,of which the significant correlations in the exposure groups areshown in Table 4. Overall, biomarker values for females wereintercorrelated more frequently than those for males. Particu-larly in females of the OX100 group, seven significantcorrelations were found. The concentrations of T and E2 wereintercorrelated positively (r=0.59–0.87) for females in allexposure groups, whereas E2 was negatively correlated withVtg concentration in females in the control, U10 and U100groups. Vtg was correlated with the percentage of somematurational stages of oocytes in females in OX10, OX100and T treatments, and the percentage of atretic oocytescorrelated with ovarian maturational stages in females of theU10, OX100 and T groups. In males, the concentrations of Tand 11-KTwere correlated positively (r=0.52–0.94) except formales exposed to E2. There were only few associations betweenthe percentages of spermatogenic stages and 11-KT; these werefound in males exposed to U10 and E2.
3.5. In vitro steroid production
The steroid production of ovarian follicles is shown in Fig. 4.The viability of the follicles was not decreased due to treatmentsafter the incubation period. Ovarian follicles exposed to U10and U100 showed a slightly increased T concentration, whereasin the OX10 and OX100 treatments the T concentration wassignificantly higher. In E2 concentrations there were nosignificant differences compared to the control. The E2/T ratioswere lower in all treatments compared to the control, and
significantly lower in follicles exposed to U100, OX10 andOX100.
4. Discussion
In the present study we examined the effects of a phytosterolpreparation and its oxidation products on reproductive para-meters in zebrafish. The nominal concentrations used, 10 μg/Land 100 μg/L, were aimed to be environmentally relevant.Studies from Finland inland waters report β-sitosterol concen-trations of 68.9 μg/L and 48.6 μg/L in two secondarily treatedeffluents (Karels et al., 2001) and total sterol concentrations of11–45 μg/L at the discharge point of a mill (Kostamo andKukkonen, 2003). Oxidized sterols have been found both inbleached pulp and in effluents, for instance 7-hydroxy-sitosterol, which was also the major oxidation product in thepresent experiment (Bergelin and Holmbom, 2005). Further,sterol oxidation products have been reported to be present in
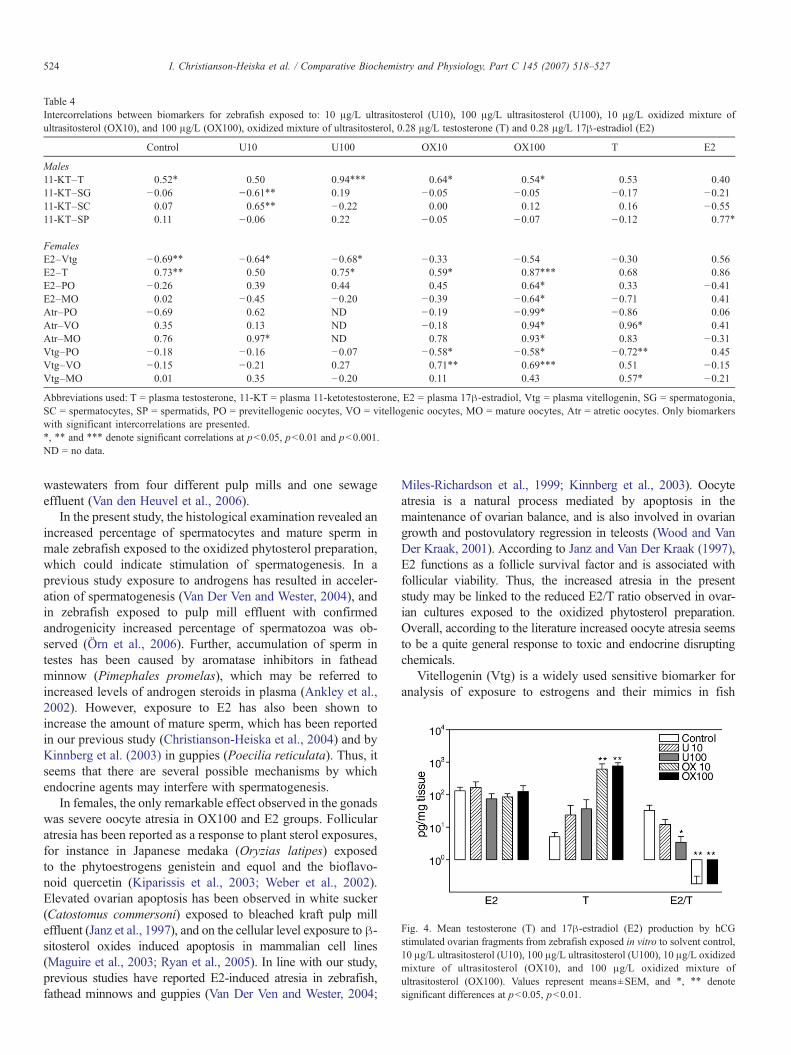
Table 4Intercorrelations between biomarkers for zebrafish exposed to: 10 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidized mixture ofultrasitosterol (OX10), and 100 μg/L (OX100), oxidized mixture of ultrasitosterol, 0.28 μg/L testosterone (T) and 0.28 μg/L 17β-estradiol (E2)
Control U10 U100 OX10 OX100 T E2
Males11-KT–T 0.52⁎ 0.50 0.94⁎⁎⁎ 0.64⁎ 0.54⁎ 0.53 0.4011-KT–SG −0.06 −0.61⁎⁎ 0.19 −0.05 −0.05 −0.17 −0.2111-KT–SC 0.07 0.65⁎⁎ −0.22 0.00 0.12 0.16 −0.5511-KT–SP 0.11 −0.06 0.22 −0.05 −0.07 −0.12 0.77⁎
FemalesE2–Vtg −0.69⁎⁎ −0.64⁎ −0.68⁎ −0.33 −0.54 −0.30 0.56E2–T 0.73⁎⁎ 0.50 0.75⁎ 0.59⁎ 0.87⁎⁎⁎ 0.68 0.86E2–PO −0.26 0.39 0.44 0.45 0.64⁎ 0.33 −0.41E2–MO 0.02 −0.45 −0.20 −0.39 −0.64⁎ −0.71 0.41Atr–PO −0.69 0.62 ND −0.19 −0.99⁎ −0.86 0.06Atr–VO 0.35 0.13 ND −0.18 0.94⁎ 0.96⁎ 0.41Atr–MO 0.76 0.97⁎ ND 0.78 0.93⁎ 0.83 −0.31Vtg–PO −0.18 −0.16 −0.07 −0.58⁎ −0.58⁎ −0.72⁎⁎ 0.45Vtg–VO −0.15 −0.21 0.27 0.71⁎⁎ 0.69⁎⁎⁎ 0.51 −0.15Vtg–MO 0.01 0.35 −0.20 0.11 0.43 0.57⁎ −0.21
Abbreviations used: T = plasma testosterone, 11-KT = plasma 11-ketotestosterone, E2 = plasma 17β-estradiol, Vtg = plasma vitellogenin, SG = spermatogonia,SC = spermatocytes, SP = spermatids, PO = previtellogenic oocytes, VO = vitellogenic oocytes, MO = mature oocytes, Atr = atretic oocytes. Only biomarkerswith significant intercorrelations are presented.⁎, ⁎⁎ and ⁎⁎⁎ denote significant correlations at pb0.05, pb0.01 and pb0.001.ND = no data.
Fig. 4. Mean testosterone (T) and 17β-estradiol (E2) production by hCGstimulated ovarian fragments from zebrafish exposed in vitro to solvent control,10 μg/L ultrasitosterol (U10), 100 μg/L ultrasitosterol (U100), 10 μg/L oxidizedmixture of ultrasitosterol (OX10), and 100 μg/L oxidized mixture ofultrasitosterol (OX100). Values represent means±SEM, and ⁎, ⁎⁎ denotesignificant differences at pb0.05, pb0.01.
524 I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
wastewaters from four different pulp mills and one sewageeffluent (Van den Heuvel et al., 2006).
In the present study, the histological examination revealed anincreased percentage of spermatocytes and mature sperm inmale zebrafish exposed to the oxidized phytosterol preparation,which could indicate stimulation of spermatogenesis. In aprevious study exposure to androgens has resulted in acceler-ation of spermatogenesis (Van Der Ven and Wester, 2004), andin zebrafish exposed to pulp mill effluent with confirmedandrogenicity increased percentage of spermatozoa was ob-served (Örn et al., 2006). Further, accumulation of sperm intestes has been caused by aromatase inhibitors in fatheadminnow (Pimephales promelas), which may be referred toincreased levels of androgen steroids in plasma (Ankley et al.,2002). However, exposure to E2 has also been shown toincrease the amount of mature sperm, which has been reportedin our previous study (Christianson-Heiska et al., 2004) and byKinnberg et al. (2003) in guppies (Poecilia reticulata). Thus, itseems that there are several possible mechanisms by whichendocrine agents may interfere with spermatogenesis.
In females, the only remarkable effect observed in the gonadswas severe oocyte atresia in OX100 and E2 groups. Follicularatresia has been reported as a response to plant sterol exposures,for instance in Japanese medaka (Oryzias latipes) exposedto the phytoestrogens genistein and equol and the bioflavo-noid quercetin (Kiparissis et al., 2003; Weber et al., 2002).Elevated ovarian apoptosis has been observed in white sucker(Catostomus commersoni) exposed to bleached kraft pulp milleffluent (Janz et al., 1997), and on the cellular level exposure to β-sitosterol oxides induced apoptosis in mammalian cell lines(Maguire et al., 2003; Ryan et al., 2005). In line with our study,previous studies have reported E2-induced atresia in zebrafish,fathead minnows and guppies (Van Der Ven and Wester, 2004;
Miles-Richardson et al., 1999; Kinnberg et al., 2003). Oocyteatresia is a natural process mediated by apoptosis in themaintenance of ovarian balance, and is also involved in ovariangrowth and postovulatory regression in teleosts (Wood and VanDer Kraak, 2001). According to Janz and Van Der Kraak (1997),E2 functions as a follicle survival factor and is associated withfollicular viability. Thus, the increased atresia in the presentstudy may be linked to the reduced E2/T ratio observed in ovar-ian cultures exposed to the oxidized phytosterol preparation.Overall, according to the literature increased oocyte atresia seemsto be a quite general response to toxic and endocrine disruptingchemicals.
Vitellogenin (Vtg) is a widely used sensitive biomarker foranalysis of exposure to estrogens and their mimics in fish

525I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
(Arukwe and Goksøyr, 2003). In the present study, we foundsignificantly higher Vtg induction in male zebrafish exposure to10 μg/L ultrasitosterol. Vtg concentrations were also elevated inthe other treatments, however with an extensive individualvariation. Interestingly, the mean Vtg level was higher (notstatistically significant) also in females in all other treatmentscompared to the control, and thus it seems that the wood sterolsat the tested concentrations are weakly estrogenic in both maleand female zebrafish. This finding agrees with a previous study,where Vtg production was induced in both male and femalezebrafish by a similar wood sterol mixture (ultrasitosterol)exposed at 10 and 20 μg/L (Nakari and Erkomaa, 2003). Thereason to the increased Vtg production could be the directeffects of estrogenic phytosterols, or alternatively aromatisationof androgens into estrogenic compounds, as suggested by Örnet al. (2006) who found Vtg induction in zebrafish exposed topulp mill effluent with confirmed androgenicity. Whether theoxidation products of β-sitosterol affect the Vtg levels remainsunclear, since the ultrasitosterol was not completely oxidizedand thus the exposures contained also original wood sterols.Van den Heuvel et al. (2006) did not find any estrogenic effectson mosquitofish exposed to oxidized sterols, however, theoxidation products reported in that study may be different fromthe oxidized sterols in the present study.
The common effects of exposure to pulp mill effluents in fishare depressions in circulating reproductive steroids, which has alsobeen reported for different fish species exposed to β-sitosterol(MacLatchy and Van der Kraak, 1995; MacLatchy et al., 1997;Gilman et al., 2003). Contrary to this, we found elevated plasma Tand 11-KT levels in OX10 males, and increased 11-KTconcentration followed by a higher-KT/T ratio in males exposedto E2. Increased plasma Tand 11-KT levels have been observed ingoldfish exposed to E2, and it has been suggested that thisresponse is due to E2 interfering with the hypothalamo–pituitary–gonadal axis (MacLatchy et al., 1997). A comparison between thepresent results and the earlier documented effects is howeverdifficult, since varying β-sitosterol preparations and routes ofexposure have been used, and there may also be speciesdifferences in hormonal responses. Interestingly, the increasedsteroid levels were observed in those treatments of male fish(OX10, E2) where also the spermatogenic effects were found, andthus there may be some association between these responses.
In order to examine the effects of phytosterols on gonadalsteroidogenesis, we measured T and E2 production uponexposure in ovarian follicles in vitro. The T production wasincreased in OX10 and OX100 exposures while E2 productionremained unchanged, resulting in a decreased E2/T ratiocompared to the control. In the in vivo exposure, no suchchanges in steroid levels were observed. One reason could bethat the effects on steroid biosynthesis are too minor to betranslated into changes in plasma titers, as suggested by Sharpeet al. (2004) in a study regarding gonadal hormone production.Based on the present results, it seems that the oxidizedphytosterols may either affect the ovarian aromatase enzyme,which converts T to E2, or alternatively some earlier enzymes inthe steroidogenic pathway. The latter explanation is more likelysince the T levels were increased without concomitant increase
in E2 levels, and instead of inhibition Orlando et al. (2002)reported increase of brain and ovary aromatase activity in fishexposed to pulp mill effluents.
In an attempt to evaluate if there are interrelations between thechanges in the reproductive parameters, we subjected the data forindividual fish to correlation analyses. Overall, the reproductivebiomarkers turned out to be more frequently intercorrelated infemales than in males, which has also been reported by Nicholset al. (1999) for female fathead minnows exposed to municipalwastewater. In the same study both E2 and T were positivelycorrelated with the Vtg concentration. On the contrary, in thepresent study the correlations turned out to be negative between E2and Vtg. Biomarkers were most abundantly intercorrelated infemales exposed to OX100, particularly the follicular stages andatretic oocytes. Further, the correlation coefficients for thesefemales resemble the correlation pattern in T treated females,although the associations are not always significant. This indicatesthat oxidized phytosterols may have some androgenic effects infemales on the gonadal level. The relationships in theOX100 groupare at least quite different from the E2 females, which suggestdifferent mechanisms in ovotoxicity of the compounds. Thereproductive steroids (E2-T) were positively correlated in femalesin nearly all treatment groups, which agree with our earlier studiesconcerning wood extractives (unpublished results).
In males there were only few significant correlationsbetween the reproductive biomarkers. The most evident positivecorrelation was found between plasma T and 11-KT in nearly allexposure groups. Although Vtg was elevated in males exposedto U10, no interrelations between Vtg and other bioindicatorswere noticed. Correlations were found between the percentagesof spermatogonia and spermatocytes and 11-KT in malesexposed to U10. A lack of association applies also for the sexsteroids and spermatogenic stages in males exposed to OX10,although the T and 11-KT concentrations as well as thepercentage of spermatocytes were elevated in these fish.However, a positive correlation between 11-KT and percentageof spermatids is evident in E2 males which also showedincreased plasma 11-KT. This indicates that there may bedifferent mechanisms of action of phytosterols and E2 on thegonadal level in male fish. Interestingly, although female fishshowed very few significant differences in the absolute valuesof all measures, there turned out to be a remarkable number ofsignificant associations between the reproductive parameters onthe individual level. This stresses the importance of analysingreproductive bioindicators on an individual level additional tothe average measures in order to reveal possible linkages.
To summarize the results, the present investigation revealedsome reproductive effects of a phytosterol preparation (ultra-sitosterol) and its oxidation products on zebrafish in vivo and ongonadal steroidogenesis in vitro. Ultrasitosterol induced Vtgin male fish, which suggests a weak estrogenic potency ofthe preparation. On the other hand, exposure to oxidizedphytosterols resulted in effects resembling those of androgeniccompounds. Such effects might be increased plasma sexsteroids and acceleration of spermatogenesis in males, andincreased ovarian atresia in females. Further, we found strongcorrelations between several reproductive biomarkers in female

526 I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
fish exposed to oxidized phytosterols. Gonadal histopathologyproved to be a sensitive endpoint in the present study revealingeffects in both males and females. More detailed studies areneeded in order to evaluate mechanisms of action of oxidationproducts of phytosterols in fish, and to which extent suchcompounds possess a hazard for the reproductive health ofaquatic animals.
Acknowledgements
This work was supported by grants from Svenska Kultur-fonden and Stiftelsens för Åbo Akademi Forskningsinstitut.
References
Ankley, G.T., Kahl, M.D., Jensen, K.M., Hornung, M.W., Korte, J.J., Makynen,E.A., Leino, R.L., 2002. Evaluation of the aromatase inhibitor fadrozole in ashort-term reproduction assay with the fathead minnow (Pimephalespromelas). Toxicol. Sci. 67, 121–130.
Arukwe, A., Goksøyr, A., 2003. Eggshell and egg yolk proteins in fish: hepaticproteins for the next generation — oogenetic, population, and evolutionaryimplications of endocrine disruption. Comp. Hepatol. 2, 4 (http://www.comparative-hepatology.com).
Bergelin, E., Holmbom, B., 2005. Oxidized sterols and triterpenyl alcoholsformed in chlorine dioxide bleaching. Proceedings, International PulpBleaching Conference, Stockholm, Sweden, pp. 175–180.
Christianson-Heiska, I., Wahteristo, P., Kastilan, E.L., Bergelin, E., Bylund, G.,Isomaa, B., 2004. Effects of the wood extractive betulinol and 17beta-oestradiol on reproduction in zebrafish, Danio rerio (Hamilton) —complications due to a bacterial infection. J. Fish Dis. 27, 267–276.
Cook, D.L., LaFleur, L., Parrish, A., Jones, J., Hoy, D., 1997. Characterizationof plant sterols in a select group of US pulp and paper mills. Fifth IEAWQSymposium preprint of Forest Industry Waste Water. Vancouver, Canada,pp. 1–8.
Gilman, C.I., Leusch, F.D.L., Breckenridge, W.C., MacLatchy, D.L., 2003.Effects of a phytosterol mixture on male fish plasma lipoprotein fractionsand testis P450scc activity. Gen. Comp. Endocrinol. 130, 172–184.
Honkanen, J.O., Kostamo, A., Kukkonen, J.V.K., 2005. Toxicity of aphytosterol mixture to grayling (Thymallus thymallus) during earlydevelopmental stages. Arch. Environ. Contam. Toxicol. 48, 391–396.
Howell, W.M., Denton, T.E., 1989. Gonopodial morphogenesis in femalemosquitofish, Gambusia affinis affinis, masculinized by exposure todegradation products from plant sterols. Environ. Biol. Fish 24, 43–51.
Janz, D.M., Van Der Kraak, G., 1997. Suppression of apoptosis bygonadotropin, 17β-estradiol and epidermal growth factor in rainbow troutpreovulatory ovarian follicles. Gen. Comp. Endocrinol. 105, 186–193.
Janz, D.M., McMaster, M.E., Munkittrick, K.R., Van der Kraak, G., 1997.Elevated ovarian follicular apoptosis and heat shock protein-70 expressionin white sucker exposed to bleached kraft pulp mill effluent. Toxicol. Appl.Pharmacol. 147, 391–398.
Karels, A., Markkula, E., Oikari, A., 2001. Reproductive, biochemical,physiological, and population responses in perch (Perca fluviatilis L.) androach (Rutilus rutilus L.) downstream of two elemental chlorine-free pulpand paper mills. Environ. Toxicol. Chem. 20, 1517–1527.
Kinnberg, K., Korsgaard, B., Bjerregaard, P., 2003. Effects of octylphenol and17β-estradiol on the gonads of guppies (Poecilia reticulata) exposed asadults via the water or as embryos via the mother. Comp. Biochem. Physiol.C 134, 45–55.
Kiparissis, Y., Balch, G.C., Metcalfe, T.L., Metcalfe, C.D., 2003. Effects of theisoflavones genistein and equol on the gonadal development of Japanesemedaka (Oryzias latipes). Environ. Health Perspect. 111, 1158–1163.
Kostamo, A., Kukkonen, J.V.K., 2003. Removal of resin acids and sterols frompulp mill effluents by activated sludge treatment. Water Res. 37, 2813–2820.
Lehtinen, K.-J., Mattsson, K., Tana, J., Engström, C., Lerche, O., Hemming, J.,1999. Effects of wood-related sterols on the reproduction, egg survival, and
offspring of brown trout (Salmo trutta lacustris L.). Ecotoxicol. Environ.Saf. 42, 40–49.
Leusch, F.D.L., MacLatchy, D.L., 2003. In vivo implants of β-sitosterol causereductions of reactive cholesterol pools in mitochondria isolated fromgonads of male goldfish (Carassius auratus). Gen. Comp. Endocrinol. 134,255–263.
MacLatchy, D.L., Van der Kraak, G.J., 1995. The phytoestrogen β-sitosterolalters the reproductive endocrine status of goldfish. Toxicol. Appl.Pharmacol. 134, 305–312.
MacLatchy, D., Peters, L., Nickle, J., Van der Kraak, G., 1997. Exposure toβ-sitosterol alters the endocrine status of goldfish differently than 17β-estradiol. Environ. Toxicol. Chem. 16, 1895–1904.
Maguire, L., Konoplyannikov, M., Ford, A., Maguire, A., O'Brien, N., 2003.Comparison of the cytotoxic effects of β-sitosterol oxides and a cholesteroloxide, 7β-hydroxycholesterol, in cultured mammalian cells. Br. J. Nutr. 90,767–775.
Mattsson, K., Tana, J., Engström, C., Hemming, J., Lehtinen, K.-J., 2001.Effects of wood-related sterols on the offspring of the viviparous blenny,Zoarces viviparus L. Ecotoxicol. Environ. Saf. 49, 122–130.
Mellanen, P., Petänen, T., Lehtimäki, J., Mäkelä, S., Bylund, G., Holmbom, B.,Mannila, E., Oikari, A., Santti, R., 1996. Wood-derived estrogens: studies invitro with breast cancer cell lines and in vivo in trout. Toxicol. Appl.Pharmacol. 136, 381–388.
Miles-Richardson, S.R., Kramer, V.J., Fitzgerald, S.D., Render, J.A., Yamini, B.,Barbee, S.J., Giesy, J.P., 1999. Effects of waterborne exposure of 17β-estradiol on secondary sex characteristics and gonads of fathead minnows(Pimephales promelas). Aquat. Toxicol. 47, 129–145.
Munkittrick, K.R., McMaster, M.E., McCarthy, L.H., Servos, M.R., Van derKraak, G.J., 1998. An overview of recent studies on the potential of pulpmill effluents to alter reproductive parameters in fish. J. Toxicol. Environ.Health, Part B. Crit. Rev. 1, 347–371.
Nagahama, Y., 1983. The functional morphology of teleost gonads. In: Hoar,W.S.,Randall, D.J., Donaldson, E.M. (Eds.), Fish Physiology, vol. IX. AcademicPress, New York, pp. 223–275.
Nakari, T., Erkomaa, K., 2003. Effects of phytosterols on zebrafish reproductionin multigeneration test. Environ. Pollut. 123, 267–273.
Nichols, K.M., Miles-Richardson, S.R., Snyder, E.M., Giesy, J.P., 1999. Effectsof exposure to municipal wastewater in situ on the reproductive physiologyof the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 18,2001–2012.
Orlando, E.F., Davis, W.P., Guillette Jr., L.J., 2002. Aromatase activity inthe ovary and brain of the eastern mosquitofish (Gambusia holbrooki)exposed to paper mill effluent. Environ. Health Perspect. 110 (suppl. 3),429–433.
Örn, S., Svenson, A., Viktor, T., Holbech, H., Norrgren, L., 2006. Male-biased sex ratios and vitellogenin induction in zebrafish exposed to effluentwater from a Swedish pulp mill. Arch. Environ. Contam. Toxicol. 51,445–451.
Ryan, E., Chopra, J., McCarthy, F., Maguire, A.R., O'Brien, N.M., 2005.Qualitative and quantitative comparison of the cytotoxic and apoptoticpotential of phytosterol oxidation products with their correspondingcholesterol oxidation products. Br. J. Nutr. 94, 443–451.
Sharpe, R.L., MacLatchy, D.L., Courtenay, S.C., Van Der Kraak, G.J., 2004.Effects of a model androgen (methyl testosterone) and a model anti-androgen (cyproterone acetate) on reproductive endocrine endpoints in ashort-term adult mummichog (Fundulus heteroclitus) bioassay. Aquat.Toxicol. 67, 203–215.
Tremblay, L., Van der Kraak, G., 1998. Use of a series of homologous invitro and in vivo assays to evaluate the endocrine modulating actions ofβ-sitosterol in rainbow trout. Aquat. Toxicol. 43, 149–162.
Tremblay, L., Van der Kraak, G., 1999. Comparison between the effectsof the phytosterol β-sitosterol and pulp and paper mill effluentson sexually immature rainbow trout. Environ. Toxicol. Chem. 18,329–336.
Van den Heuvel, M.R., Leusch, F.D.L., Taylor, S., Shannon, N., McKague, A.B.,2006. Assessment of the reproductive–endocrine disrupting potential ofchlorine dioxide oxidation products of plant sterols. Environ. Sci. Technol.40, 2594–2600.

527I. Christianson-Heiska et al. / Comparative Biochemistry and Physiology, Part C 145 (2007) 518–527
Van Der Kraak, G.J., Munkittrick, K.R., McMaster, M.E., Portt, C.B., Chang, J.P.,1992. Exposure to bleached kraft pulp mill effluent disrupts the pituitary–gonadal axis of white sucker at multiple sites. Toxicol. Appl. Pharmacol. 115,224–233.
Van Der Ven, L., Wester, P., 2004. Toxicological pathology atlas of smalllaboratory fish: part I — normal histology and effects of endocrinedisruptors in zebrafish Danio rerio. http://www.rivm.nl/fishtoxpat/.
Weber, L.P., Kiparissis, Y., Hwang,G.S., Niimi, A.J., Janz, D.M.,Metcalfe, C.D.,2002. Increased cellular apoptosis after chronic aqueous exposure tononylphenol and quercetin in adult medaka (Oryzias latipes). Comp.Biochem. Physiol. C 131, 51–59.
Wood, A.W., Van Der Kraak, G.J., 2001. Apoptosis and ovarian function: novelperspectives from the teleosts. Biol. Reprod. 64, 264–271.
Related Documents











