Modern emu (Dromaius novaehollandiae) butchery, economic utility and analogues for the Australian archaeological record Jillian Garvey, Brett Cochrane, Judith Field and Chris Boney Australia’s largest flightless bird, the emu (Dromaius novaehollandiae), has been an important prey animal for Indigenous people for millennia, especially in arid/semi-arid areas where, along with large kangaroos, they can provide high economic returns from single kills. Understanding modern prey selection, butchering patterns and the relative nutritional value of the different body portions in these animals has important implications for interpreting patterns of species and body part representation in the archaeological record. A butchery study, economic utility assessment, and meat and marrow fatty acid analysis of the Australian emu has established the relative economic importance of different body parts. The results show that the femur/pelvic region yielded the greatest amount of meat, and that the quantity and quality of fats associated with these units makes bone fracturing for marrow extraction superfluous. The results provide new insights into the relative importance of emu in Australian Aboriginal diets, past and present, and establish useful comparative data for studies of the now extinct giant flightless bird Genyornis newtoni. Keywords: Australia, Late Pleistocene, archaeology, Dromaius novaehollandiae, economic utility, fatty acid analysis, Genyornis newtoni Introduction Modern humans arrived in Sahul (Pleistocene Australia-New Guinea) sometime between 40 ka and c. 60 ka, arguably at a time of megafauna decline and deteriorating climatic conditions (O’Connell and Allen 2004; Field et al. 2008; Davidson 2010; Summerhayes et al. 2010; Wurster et al. 2010). The timing and coincidence of these events is the subject of intense debate (Wroe and Field 2006; Field et al. 2008; 2011). While a broad picture is emerging of when and where modern humans were present on the landscape, we still have little detail on the subsistence practices of the first Australians. Of particular interest is the potential interaction of humans with megafauna as well as the utilisation of some of our modern large target prey, such as the kangaroo and emu (e.g. O’Connell and Marshall 1989; O’Connell 2000; Johnson 2005; Wroe and Field 2006; Field et al. 2008; 2010). The first human arrivals occupied most environ- ments across the continent within a relatively short period of time, adapting to a new vegetation and faunal suite not seen in South-East Asia (Denham et al. 2009). Apart from the rich south-west Tasmania sites (e.g. Allen 1996; Cosgrove and Allen 2001; Garvey 2006), there are few continental sites with well-preserved faunal remains. Current datasets indi- cate that, since the arrival of the first humans in Sahul, there has been negligible variability in available target prey; with the exception of some now extinct mega- faunal species (Sutton et al. 2009; Field and Dodson 1999). Investigations into prey selection, butchering and use of many of these species are hampered by the scarcity of ethnographic observations on the economic importance of different prey species. Most studies looking at aspects of economic utility have targeted the extant kangaroo to evaluate carcass composition for the modern domestic meat trade (Garvey 2010). Only two studies are known from Australia that have Jillian Garvey (corresponding author), Archaeology Program, La Trobe University, Victoria 3086, Australia; e-mail: [email protected]; Brett Cochrane, 12 Waratah St, Brewarrina, NSW 2839, Australia; Judith Field, School of Biological, Earth and Environmental Sciences, The University of New South Wales, NSW 2052, Australia; Chris Boney, PO Box 79 Brewarrina, NSW 2839, Australia. ß Association for Environmental Archaeology 2011 Published by Maney DOI 10.1179/174963111X13110803260840 Environmental Archaeology 2011 VOL 16 NO 2 97

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Modern emu (Dromaius novaehollandiae)butchery, economic utility and analogues forthe Australian archaeological record
Jillian Garvey, Brett Cochrane, Judith Field and Chris Boney
Australia’s largest flightless bird, the emu (Dromaius novaehollandiae), has been an important
prey animal for Indigenous people for millennia, especially in arid/semi-arid areas where, along
with large kangaroos, they can provide high economic returns from single kills. Understanding
modern prey selection, butchering patterns and the relative nutritional value of the different body
portions in these animals has important implications for interpreting patterns of species and body
part representation in the archaeological record. A butchery study, economic utility assessment,
and meat and marrow fatty acid analysis of the Australian emu has established the relative
economic importance of different body parts. The results show that the femur/pelvic region
yielded the greatest amount of meat, and that the quantity and quality of fats associated with
these units makes bone fracturing for marrow extraction superfluous. The results provide new
insights into the relative importance of emu in Australian Aboriginal diets, past and present, and
establish useful comparative data for studies of the now extinct giant flightless bird Genyornis
newtoni.
Keywords: Australia, Late Pleistocene, archaeology, Dromaius novaehollandiae, economic utility, fatty acid analysis, Genyornis newtoni
Introduction
Modern humans arrived in Sahul (Pleistocene
Australia-New Guinea) sometime between 40 ka and
c. 60 ka, arguably at a time of megafauna decline and
deteriorating climatic conditions (O’Connell and Allen
2004; Field et al. 2008; Davidson 2010; Summerhayes
et al. 2010; Wurster et al. 2010). The timing and
coincidence of these events is the subject of intense
debate (Wroe and Field 2006; Field et al. 2008; 2011).
While a broad picture is emerging of when and where
modern humans were present on the landscape, we still
have little detail on the subsistence practices of the first
Australians. Of particular interest is the potential
interaction of humans with megafauna as well as the
utilisation of some of our modern large target prey,
such as the kangaroo and emu (e.g. O’Connell and
Marshall 1989; O’Connell 2000; Johnson 2005; Wroe
and Field 2006; Field et al. 2008; 2010).
The first human arrivals occupied most environ-
ments across the continent within a relatively short
period of time, adapting to a new vegetation and
faunal suite not seen in South-East Asia (Denham
et al. 2009). Apart from the rich south-west Tasmania
sites (e.g. Allen 1996; Cosgrove and Allen 2001;
Garvey 2006), there are few continental sites with
well-preserved faunal remains. Current datasets indi-
cate that, since the arrival of the first humans in Sahul,
there has been negligible variability in available target
prey; with the exception of some now extinct mega-
faunal species (Sutton et al. 2009; Field and Dodson
1999). Investigations into prey selection, butchering
and use of many of these species are hampered by the
scarcity of ethnographic observations on the economic
importance of different prey species. Most studies
looking at aspects of economic utility have targeted the
extant kangaroo to evaluate carcass composition for
the modern domestic meat trade (Garvey 2010). Only
two studies are known from Australia that have
Jillian Garvey (corresponding author), Archaeology Program, La TrobeUniversity, Victoria 3086, Australia; e-mail: [email protected]; BrettCochrane, 12 Waratah St, Brewarrina, NSW 2839, Australia; Judith Field,School of Biological, Earth and Environmental Sciences, The University ofNew South Wales, NSW 2052, Australia; Chris Boney, PO Box 79Brewarrina, NSW 2839, Australia.
� Association for Environmental Archaeology 2011Published by ManeyDOI 10.1179/174963111X13110803260840 Environmental Archaeology 2011 VOL 16 NO 2 97

examined the economic importance of target prey
which occur in archaeological assemblages. O’Connell
and Marshall (1989) studied the Red kangaroo
(Macropus rufus Desmarest) in order to construct
utility indices as a general guide for macropods. The
abundance of another, much smaller, macropod, the
Bennett’s wallaby (Macropus rufogriseus Desmarest)
in the rich south-west Tasmanian archaeological sites
(Allen 1996; Pike-Tay et al. 2008), has led researchers
to extend the work of O’Connell and Marshall (1989)
to investigate the economic utility of this species
(Garvey 2010). These studies have enabled a greater
understanding of the patterns of use and relative
abundance of macropods across time and space
providing important interpretive frameworks for
Australian zooarchaeological studies. What has been
missing from the dataset is the relevant information on
Australia’s largest extant bird — the emu Dromaius
novaehollandiae Latham — which is known to have
been an important prey animal for Australian
Aborigines in the recent past.
An understanding of the economic utility of the emu
may also have important interpretive implications for
the extinct Pleistocene bird, Genyornis newtoni Stirling
and Zietz (Rich 1979). G. newtoni appears to be one of
the megafaunal species that overlapped with human
occupation of the Australian continent. Fossil remains
of G. newtoni have been recovered from Cuddie
Springs and Lancefield Swamp in south-eastern
Australia (Field et al. 2008). While the relative
economic importance of these species is unknown,
we propose that the skeletal similarities between the
emu and G. newtoni suggest that approaches to
butchering would be paralleled. An economic utility
study of the emu would thus serve two purposes:
1) provide the first baseline data on the processing
methods and nutritional value of emu; and
2) establish a reference point for evaluating archae-
ological assemblages that include both D. novae-
hollandiae and G. newtoni skeletal remains.
Genyornis newtoni and Dromaiusnovaehollandiae
The study presented here evolved following investiga-
tions at the late Pleistocene archaeological site of
Cuddie Springs in western New South Wales
(Dodson et al. 1993; Field and Dodson 1999; Field
et al. 2008; Fillios et al. 2010). Among other identified
extant and extinct species, the remains of the extinct,
large flightless bird G. newtoni were recovered
from the same horizons (Stratigraphic Unit 6) as
flaked stone artefacts, implying contemporaneity and
possible interaction with humans (Field and Boles
1998; Fillios et al. 2010). Emu (D. novaehollandiae)
are also known from the archaeological horizons.
Most skeletal elements of G. newtoni were present
and complete in the excavated squares from Strati-
graphic Unit 6B (Fig. 1). The leg elements were
generally found in close anatomical association and
have been reported as separated articulations (Wroe
et al. 2004: fig. 1). No cutmarks were identified on any
G. newtoni skeletal elements and the lack of weathering
and/or abrasion, the fine-grained enclosing sediments,
and the ephemeral water hole conditions indicated that
the faunal remains were in a primary depositional
setting (Field 2006; Field et al. 2008; Fillios et al. 2010).
The presence of flaked stone artefacts with usewear
consistent with butchering, throughout the unit, implies
a human role in the accumulation of the remains. Little
attention has been paid to the economic importance of
G. newtoni or the emu, inhibiting an accurate evaluation
Figure 1 G. newtoni limb bones partly excavated at the
Cuddie Springs site in south-eastern Australia.
All longbone leg elements are found within a
1 m square in fine-grained enclosing sediments
and were deposited during a positive lake phase
during the Late Pleistocene. Flaked stone tools
have also been recovered from these horizons
and some artefacts can be seen in section (in
Stratigraphic Unit 6A SU6A, overlying SU6B)
(photo J. Field)
Garvey et al. Modern Emu Butchery
98 Environmental Archaeology 2011 VOL 16 NO 2

of their potential as prey or determining which portions
of the bird would be targeted for consumption. Further-
more, as the limb elements from Cuddie Springs did
not exhibit any physical damage that is traditionally
associated with butchering, it is important to evaluate
whether marrow extraction was ever likely, in either G.
newtoni or emu. Marrow extraction is an important
aspect of macropod exploitation (Garvey 2011) yet
there is little known concerning similar strategies in the
emu.
Study aims
The aim of this paper is to present:
1) modern emu butchery and cooking practices of
Indigenous Australians;
2) an economic utility (or anatomy) study of the
emu; and
3) a fatty acid analysis of the emu bone marrow,
muscle and stomach lining.
The implications of these results for analysing and
interpreting Australian zooarchaeological assem-
blages, including the extinct flightless bird Genyornis
newtoni, will be discussed in light of the findings. In
this paper, we have decided to only concentrate
on the development of the emu utility model and
the ethnographic study of butchery practices. The
archaeological application of this model will be
explored in a future publication.
The Australian Emu, Dromaius novaehollandiae
The Australian emu, Dromaius novaehollandiae
(Fig. 2), belongs to the Order Struthioniformes, or
the ratites; a diverse group of large, flightless birds
with small wings and without a keeled sternum. Emu
are Gondwanan in origin and are restricted to the
southern hemisphere, with most species now extinct.
Extant species include: the African ostrich (Struthio
camelus Linnaeus); two species of South American
rhea (Rhea americana Linnaeus and R. pennata
d’Orbigny); five species of the New Zealand kiwi
(Apteryx haastii Potts, A. owenii Gould, A. rowi
Tennyson et al., A. australis Shaw and Nodder and A.
mantelli Barrlett); and three species of cassowary
restricted to northern tropical Australia and New
Guinea (Casuarius casuarius Linnaeus, C. unappendi-
culatus Blyth and C. bennetti Gould). The largest and
most famous extinct rarities include Aepyornis max-
imus Geoffroy Saint-Hilaire or the elephant bird of
Madagascar which grew to approximately 450 kg
(990 lb) and stood to 3 m (9?8 ft) tall, and the 11
extinct species of Moa from New Zealand. The
largest was the Giant Moa (Dinornis giganteus Owen)
which grew to about 250 kg (550 lb) and reached
3?3 m (11 ft).
The emu, Dromaius novaehollandiae, was one of
four Dromaius taxa common in Australia prior to
European settlement (c. 1788). The other three were
the Tasmanian emu, D. novaehollandiae diemenensis
Le Souef, the King Island emu D. ate Vieillot, and
the Kangaroo Island emu D. baudinianus Parker, all
of which were smaller than their mainland cousin.
Today, D. novaehollandiae is Australia’s largest
bird, inhabiting many environments including open
woodlands, scrublands, semi-arid and arid regions
across mainland Australia. It is particularly com-
mon in pastoral and cereal-growing areas. Emus are
highly nomadic, and move in response to local
climatic conditions and the availability of water.
The emu is omnivorous, feeding on insects, berries,
fruit and flowers. Breeding occurs between April
and October when the female lays 5–11 eggs. The
male then broods over the eggs and raises the young
until approximately 18 months of age (Pizzey and
Knight 2001).
The emu and kangaroo are currently Australia’s
largest native terrestrial animal prey and are still
hunted by Indigenous people (Roth 1901; Thomson
1939; Gould 1966; 1969a; 1969b; 1981; O’Dea 1991;
O’Connell 2000). Published ethnographic accounts of
Figure 2 The Australian emu Dromaius novaehollandiae
(photo J. Garvey)
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 99

emu butchery differ to that observed for the kangaroo.
A notable difference is that macropod hindlimbs
and metatarsals are cracked open to access the
bone marrow (McArthur 1948, 121; O’Connell and
Marshall 1989), while emu longbones always seem to
be discarded intact.
Despite being regular modern prey, emu bones are
very rare in the archaeological record (e.g. Cosgrove
and Allen 2001; Garvey 2006; 2007; Fillios et al.
2010). Emu eggshell has been reported from a
number of Pleistocene and Holocene archaeological
sites and is commonly associated with hearths, e.g.
Tunnel Cave in Western Australia and Lake
Menindee in New South Wales (Dortch 1996;
Cupper and Duncan 2006). Skeletal elements are
known from only a handful of sites. Lancefield
Swamp in Victoria has yielded both G. newtoni and
emu remains, though they comprise ,1% of the
faunal assemblage (Gillespie et al. 1978) and the
relationship between the artefacts and the faunal
remains has never been successfully clarified. The
Cuddie Springs site also contains emu and G. newtoni,
which are associated with the archaeological record
(Field and Boles 1998; Fillios et al. 2010).
The Dromornithidae and Genyornis newtoni
The Dromornithidae or dromornithids were a family
of large flightless birds endemic to Australia.
Sometimes referred to as ‘thunder birds’, ‘demon
ducks’ and ‘mihirungs’, they evolved sometime
during the late Oligocene and disappeared in the late
Pleistocene (Rich 1979). Represented by five genera
and seven species, the dromornithids were a group of
birds with enormous robust bodies, powerful legs and
vestigial wings, with fused scapula and coracoids in
their shoulder girdles and no keel on their sternum
(Murray and Vickers-Rich 2004: 31). The largest
dromornithid, Dromornis stirtoni Owen (Stirton’s
Thunderbird), is represented in late Miocene levels
at Alcoota in the Northern Territory. It was probably
the world’s largest bird at approximately 3 m tall and
weighing 500 kg. Despite the superficial resemblance
of D. stirtoni to the ratites, Murray and Megirian
(1998) determined that they are phylogenetically
related to the Anseriformes: the geese, ducks and
screamers. In effect, the dromornithids including
Genyornis could be referred to as the giant geese of
Tertiary and Quaternary Australia.
Genyornis is the only Quaternary dromornithid
known and is represented by a single taxon G.
newtoni. G. newtoni was first described during the late
1890s from material found at Lake Callabonna in
South Australia (Stirling 1896; Stirling and Zietz
1896; 1890). Since then it has been recorded from
across southern and central mainland Australia. It is
represented by skeletal material, eggshell, gizzard
stones or gastroliths, possible footprints, and argu-
ably in rock art (Rich and Gill 1978; Rich 1979;
Williams 1981; Field and Boles 1998; Miller et al.
1999; Ouzman et al. 2002; Murray and Vickers-Rich
2004).
Little is known of the palaeoecology of G. newtoni
(Rich 1979). Because of their enormous size and very
robust legs, the dromornithids are considered to have
been relatively slow birds, unlike the modern emu and
ostrich, which are slender, flightless birds designed to
run at high speeds. While G. newtoni is considered to
have been heavily built, it has been difficult to estimate
its possible body mass (Murray and Vickers-Rich
2004, 207). Using models and comparisons with living
taxa, Murray and Vickers-Rich (2004, table 18)
established an estimated weight range of 250–350 kg
for G. newtoni, with a conservative mass of 275 kg.
Compared to the size of the emu (30–45 kg), G.
newtoni would have been a considerable target prey
animal for Australian Aborigines.
Economic utility
Economic utility (or economic anatomy) examines
the potential selection and transportation of a prey
animal’s specific body parts based upon the assess-
ment of its relative food value. Economic utility data
is important for constructing models of human
exploitation of animal carcasses in archaeological
assemblages, and provides important indicators of
potential site use (Binford 1978; Thomas and Mayer
1983; Jones and Metcalfe 1988; Metcalfe and Jones
1988; Grayson 1989; Lyman 1992; 1994, 223–34;
Lyman et al. 1992; Reitz and Wing 1999, 213–21).
Since its formulation by Binford (1978) and applica-
tion to caribou (Rangifer tarandus C. H. Smith) and
sheep (Ovis aries Linnaeus), economic utility indices
has been constructed for a variety of other mammals
(e.g. Blumenschine and Caro 1986; Outram and
Rowley-Conwy 1998; Lyman et al. 1992; Savelle
and Friesen 1996; Savelle et al. 1996; Savelle 1997;
Diab 1998). This includes two Australian macropods;
the Red kangaroo Macropus rufus (O’Connell and
Marshall 1989) and the Bennett’s wallaby Macropus
rufogriseus (Garvey 2010). Only two examples of the
economic utility of a bird have been reported in the
literature; the New Zealand kiwi (Apteryx sp.) (as a
proxy for the extinct Moa) (Kooyman 1984), and the
South American rhea (Rhea americana) (Giardi-
na 2006). Here we present the data for another
ratite — the extant Australian emu (Dromaius
Garvey et al. Modern Emu Butchery
100 Environmental Archaeology 2011 VOL 16 NO 2

novaehollandiae). We argue that the emu data may be
used (with caveats) as a modern analogue for the
economic utility of G. newtoni.
Fatty acid analysis
When meat is low in lipids (fat), then the bone
marrow, typically from the tibia or femur, is
consumed to obtain the essential missing nutrients.
Kangaroos in particular are renowned for being very
lean and, where concentrations of skeletal remains of
kangaroo are found in archaeological sites (e.g.
Garvey 2011), marrow-containing bones are nearly
always found broken. Several different methods
have been used to assess human preference for
specific animal body parts, and these have a direct
bearing on interpreting skeletal representation in
faunal assemblages (Binford 1978; Jones and
Metcalfe 1988; Morin 2007). The amount of lipids
(fats) in animal bone marrow, particularly in
artiodactyls, has received considerable attention
(Bear 1971; Fong 1981; Pond 1988; Cederlund
et al. 1989). Prolonged reliance on lean meat by
humans means a diet high in protein and con-
sequential physiological problems (Speth and
Spielmann 1983; Speth 1987; 1991; Outram 2002).
Fatty meat, bone marrow and carbohydrates contain
more than 50 essential fatty acids that are required
for cellular regulation and growth in humans (Speth
1989; 2010; Hockett and Haws 2003; 2005; Burger
et al. 2005). Importantly, lipids or fat are a
concentrated source of energy, suppling nine kcal
per gram compared to the four kcal per gram
produced by carbohydrates and protein (Speth
1989). Although people may not be consciously
aware of the energy provided by consuming bone
marrow and fatty meats, these products are extre-
mely palatable and provide longer periods of satiety
(Speth 1987; 1989; 1991; 2010; White 2001). Recent
nutritional studies of emu meat have been driven by
the increasing prominence of such products in the
domestic/international farming and game meat
market (Smetana 1993; Berge et al. 1997; Sales and
Horbanczuk 1998; Shao et al. 1999). The analysis
presented here extends these studies to:
1) establish the nutritional value of emu muscle,
marrow and stomach lining via fatty acid analysis;
2) investigate how the relative nutritional value of
each may be reflected in the frequency, distribu-
tion and modification of skeletal elements in the
Australian zooarchaeological record; and
3) determine the nutritional potential of the larger
extinct Genyornis newtoni.
Site setting and study context
The Australian semi-arid zone supports populations of
emus that increase significantly during wet periods with
subsequent declines in times of drought (Brown et al.
2006). Local Aborigines still hunt emu — with cars and
guns — using important knowledge concerning the
practices of butchering and consumption that have
been passed down through the generations. Brett
Cochrane and Chris Boney have paternal affiliation
with the Murawori tribe, and Chris Boney has
connections to the area around Cuddie Springs via
maternal connections to the Weilwan people. They have
routinely hunted, butchered and cooked ‘bush tucker’
(various native fauna) since they were children. It is
important that these practices and methods are
documented for future generations. Our study was
undertaken in the semi-arid south-east of the continent,
approximately 85 km south-east of Brewarrina on
Wirroona Station (near the Cuddie Springs site), New
South Wales (longitude: 146u52’E; latitude: 29u58’S).
Methodology
The emu study was undertaken using two male emus
(Individuals A and B) that were provided by Brett
Cochrane in September 2009. The animals were part
of a large mob of emus (.30) that were resident on
Wirroona Station. The animals were butchered and/
or dissected in the Wirroona Station Woolshed, a
process that began within one hour of the kill.
Traditional butchering: Individual A
Individual A was immediately butchered by Cochrane
and Boney after acquisition and the process docu-
mented by Garvey. Two of the prized portions of the
emu are the stomach lining and the intestines.
Cochrane and Boney refer to the stomach as ‘bundal’;
its traditional name in this region. In this study, the
bundal was collected separately and processed by
removing the contents and the thick layer of associated
fat (Fig. 3A). The bundal is typically cut into small
pieces and fried in a pan. The intestine is called the
‘running guts’. It is often stuffed with vegetables and
breadcrumbs and made into sausages.
The upper hindlimb and pelvic region were plucked
free of feathers to expose the skin. In these particular
birds there was very little (yellow) fat visible beneath
the skin, indicating that the animal was relatively
lean. Cochrane and Boney both commented on the
lack of a visibly thick fat layer, indicating it was too
lean for their purposes and under normal hunting
circumstances would have been abandoned.
When the skin was broken, a layer of yellow fat
was exposed. Several steak-sized portions of meat
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 101

(approximately 300–400 gm) were cut from the hind
region of the bird. Each meat portion had a layer of
fat attached to it (Fig. 3B). These portions were
frozen, to be cooked later. The legs were dismem-
bered from the pelvis using a sharp knife. No contact
between the knife and the bones was observed during
this process. The foot was separated from the leg, and
both legs were strung up on poles so that they could
be easily de-fleshed (Fig. 3C). Using a sharp knife,
the muscle was removed from the femur as one large
piece. Again, no contact was observed between the
bone and the knife. Once the butchery had ceased, the
bones were inspected for cut marks, however none
were observed. It was decided that the muscle and fat
from both legs would be made into rissoles; some
vegetables were added to the meat and then processed
using a bench mounted manual kitchen mincer.
Economic utility dissection: Individual B
A controlled economic utility analysis of Individual B
followed the butchering of Individual A. Carcass
weight, body measurements, estimated age and collec-
tion details are presented in Table 1. The method used
to dissect the emu followed Garvey (2010).
The bird was first entirely plucked of feathers and
skinned prior to dissection and the body was divided
into six core units (Table 2):
1) cranial: skull and mandible;
2) axial: vertebrae and ribs minus the cranium;
3) pectoral girdle: sternum, clavicle, coracoid and
scapula;
4) forelimb: humerus, radius, ulna, carpometacar-
pus and digits;
Table 1 Characteristics of the Australian emu, Dromaiusnovaehollandiae, Individual B, used in the economicutility study
Characteristic Emu
Sex MaleAge AdultDate of Death 21.09.09Season of Death SpringCollected Cuddie Springs, NSWElevation (m a.s.l.) 30 m a.s.l.Weight (kg) 42 kgSnout-to-vent (mm) 1500Height (head-to foot) (mm) 1900
Figure 3 Butchering the Australian emu Dromaius novaehollandiae Individual B, where: A) is the bundal or stomach lin-
ing; B) is the leg being removed from the pelvis with steaks and layers of fat on the bird’s rump visible; and C)
is Brett Cochrane defleshing the leg. In all three photos the yellow fat has been arrowed (photos J. Garvey)
Garvey et al. Modern Emu Butchery
102 Environmental Archaeology 2011 VOL 16 NO 2
5) pelvic girdle: synsacrum- fusion of the pelvis and
six caudal vertebrae; and pygostyle- fusion of the
final few caudal vertebrae; and
6) hindlimb: femur, fibula, tibiotarsus, tarsometa-
tarsus and digits.
The six core units were then divided into a further 12
individual anatomical units with gross weight, flesh
weight and bone weight recorded for each (Table 3).
Where there were paired elements, only the left was
included in the analysis. The gross weight is the weight
of the whole anatomical unit and includes the flesh, fat
and bone from each element; flesh weight is the weight
of the flesh; and bone weight the weight of the bone
after it has been cleaned (after Garvey 2010). All
internal organs or viscera were removed and indivi-
dually weighed, with the digestive tract cleaned before
being weighed and measured (Table 4). The stomach
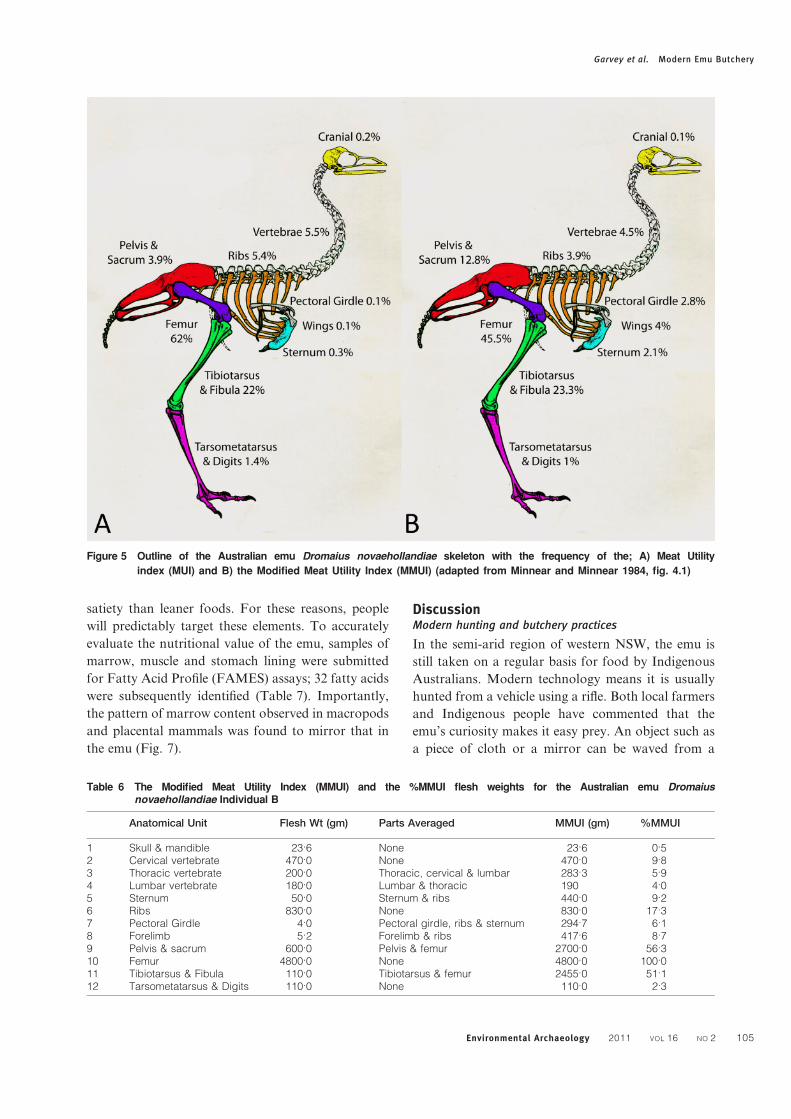
(or gizzards) of both the emus were checked for
gizzard stones (gastroliths). Gastroliths were only
found in the stomach of Individual B (Fig. 4A).
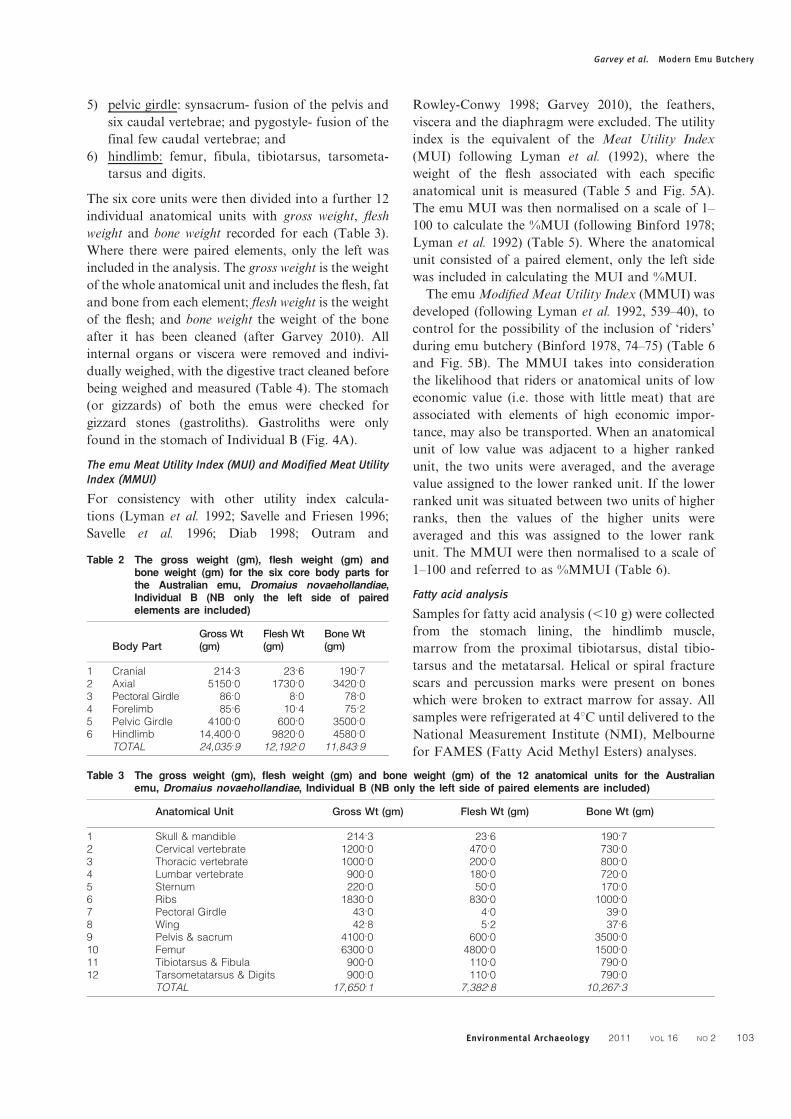
The emu Meat Utility Index (MUI) and Modified Meat UtilityIndex (MMUI)
For consistency with other utility index calcula-
tions (Lyman et al. 1992; Savelle and Friesen 1996;
Savelle et al. 1996; Diab 1998; Outram and
Rowley-Conwy 1998; Garvey 2010), the feathers,
viscera and the diaphragm were excluded. The utility
index is the equivalent of the Meat Utility Index
(MUI) following Lyman et al. (1992), where the
weight of the flesh associated with each specific
anatomical unit is measured (Table 5 and Fig. 5A).
The emu MUI was then normalised on a scale of 1–
100 to calculate the %MUI (following Binford 1978;
Lyman et al. 1992) (Table 5). Where the anatomical
unit consisted of a paired element, only the left side
was included in calculating the MUI and %MUI.
The emu Modified Meat Utility Index (MMUI) was
developed (following Lyman et al. 1992, 539–40), to
control for the possibility of the inclusion of ‘riders’
during emu butchery (Binford 1978, 74–75) (Table 6
and Fig. 5B). The MMUI takes into consideration
the likelihood that riders or anatomical units of low
economic value (i.e. those with little meat) that are
associated with elements of high economic impor-
tance, may also be transported. When an anatomical
unit of low value was adjacent to a higher ranked
unit, the two units were averaged, and the average
value assigned to the lower ranked unit. If the lower
ranked unit was situated between two units of higher
ranks, then the values of the higher units were
averaged and this was assigned to the lower rank
unit. The MMUI were then normalised to a scale of
1–100 and referred to as %MMUI (Table 6).
Fatty acid analysis
Samples for fatty acid analysis (,10 g) were collected
from the stomach lining, the hindlimb muscle,
marrow from the proximal tibiotarsus, distal tibio-
tarsus and the metatarsal. Helical or spiral fracture
scars and percussion marks were present on bones
which were broken to extract marrow for assay. All
samples were refrigerated at 4uC until delivered to the
National Measurement Institute (NMI), Melbourne
for FAMES (Fatty Acid Methyl Esters) analyses.
Table 2 The gross weight (gm), flesh weight (gm) andbone weight (gm) for the six core body parts forthe Australian emu, Dromaius novaehollandiae,Individual B (NB only the left side of pairedelements are included)
Body PartGross Wt(gm)
Flesh Wt(gm)
Bone Wt(gm)
1 Cranial 214.3 23.6 190.72 Axial 5150.0 1730.0 3420.03 Pectoral Girdle 86.0 8.0 78.04 Forelimb 85.6 10.4 75.25 Pelvic Girdle 4100.0 600.0 3500.06 Hindlimb 14,400.0 9820.0 4580.0
TOTAL 24,035.9 12,192.0 11,843.9
Table 3 The gross weight (gm), flesh weight (gm) and bone weight (gm) of the 12 anatomical units for the Australianemu, Dromaius novaehollandiae, Individual B (NB only the left side of paired elements are included)
Anatomical Unit Gross Wt (gm) Flesh Wt (gm) Bone Wt (gm)
1 Skull & mandible 214.3 23.6 190.72 Cervical vertebrate 1200.0 470.0 730.03 Thoracic vertebrate 1000.0 200.0 800.04 Lumbar vertebrate 900.0 180.0 720.05 Sternum 220.0 50.0 170.06 Ribs 1830.0 830.0 1000.07 Pectoral Girdle 43.0 4.0 39.08 Wing 42.8 5.2 37.69 Pelvis & sacrum 4100.0 600.0 3500.010 Femur 6300.0 4800.0 1500.011 Tibiotarsus & Fibula 900.0 110.0 790.012 Tarsometatarsus & Digits 900.0 110.0 790.0
TOTAL 17,650.1 7,382.8 10,267.3
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 103

Results
Two emus were evaluated in this analysis: Individual
A was butchered using traditional methods; while
Individual B underwent an economic utility dissection
and fatty acid analysis of its meat, marrow and stomach
lining to determine its nutritional content. The results
from Individual A have important implications for our
understanding and interpretation of identifying emu
butchery in the Australian archaeological record. It was
found that the emu stomach lining and intestines were
considered a delicacy, with the meat from around the
pelvis and femur the most sought after. Observations
and subsequent discussions of the butchery further
influenced our selection of samples submitted for fatty
acid analysis, with the inclusion of stomach lining as
well as the meat and marrow.
All raw data from the economic utility dissection of
Individual B is presented in Tables 1–4. Approximately
22 kg meat (53% of total weight) was recovered from a
42 kg emu, with bone constituting 13 kg (32% of total
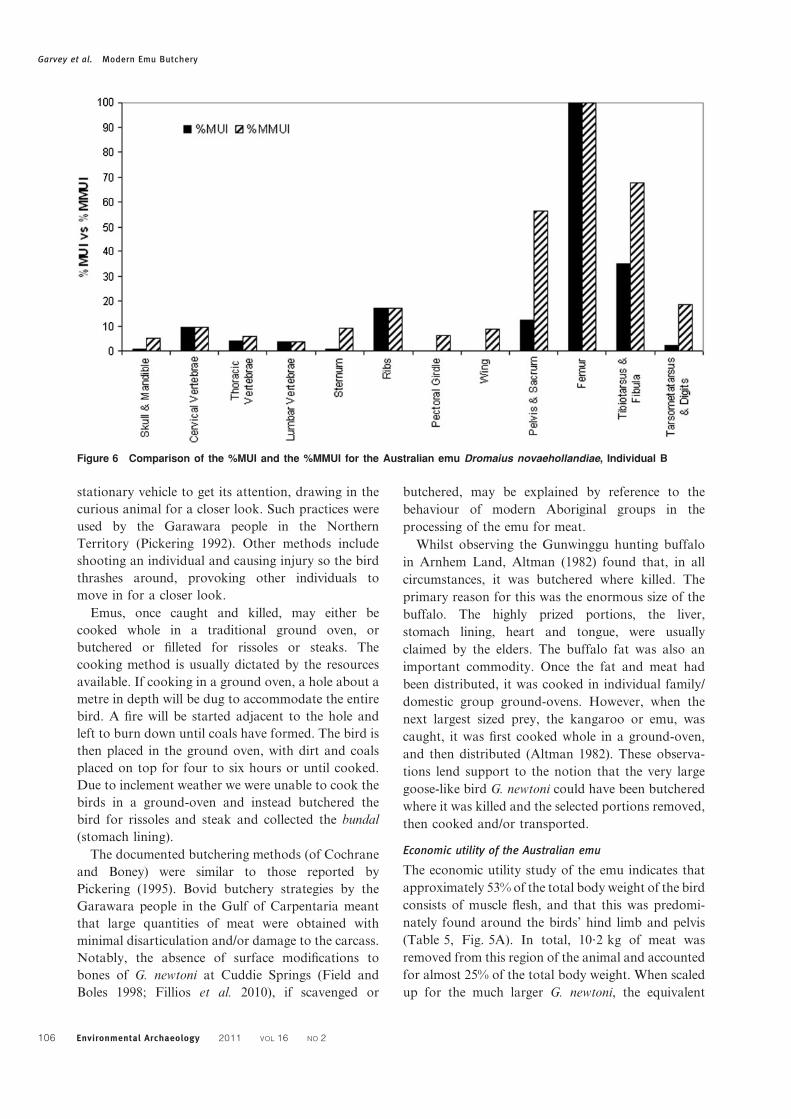
weight). The Meat Utility Index (MUI) and %MUI
indicated that the femur yielded the most flesh of all the
emu elements, even when riders were taken into
consideration (Table 5). In combination, the femur
and pelvis are the greatest meat-bearing parts of the
animal (Table 6). The relationship between the emu
%MUI and %MMUI was tested using the Spearman
Rank Coefficient, and were found to be moderately
associated (R50?60, P,0?48, Fig. 6). These results
indicate that modification for riders is an important
consideration in the economic utility of the Australian
emu.
In placental ungulates and rodents, and the ma-
rsupial macropod, it has been established that the
amount of unsaturated fats increases distally from the
body core temperature or the heart (Dietz 1946; Meng
et al. 1969; West and Shaw 1975; Turner 1979; Pond
1988, 92; Madrigal and Capaldo 1999; Garvey 2011).
This means that marrow in the distal elements
(towards the hands and feet) will be softer and oilier
and hence more palatable, providing longer periods of
Table 4 The viscera weights (gm) for the Australian emu,Dromaius novaehollandiae, Individual B
Viscera Gross Wt (gm)
1 Brain 10.62 Heart 297.33 Diaphragm 30.04 Lungs 208.25 Trachea 148.06 Spleen 38.07 Liver 735.28 Oesophagus 105.49 Stomach (cleaned) 350.010 Gizzard Stones 41.611 Intestines (cleaned) 1250.012 Kidneys 33.012 Pancreas 20.0
TOTAL 3267.3
Table 5 The Meat Utility Index (MUI) and the %MUI perskeletal element for the Australian emu, Dromaiusnovaehollandiae, Individual B
Anatomical Unit MUI (gm) %MUI
1 Skull & mandible 23.6 0.52 Cervical vertebrate 470.0 9.83 Thoracic vertebrate 200.0 4.24 Lumbar vertebrate 180.0 3.85 Sternum 50.0 1.06 Ribs 830.0 17.37 Pectoral Girdle 4.0 0.18 Forelimb 5.2 0.19 Pelvis & sacrum 600.0 12.510 Femur 4800.0 100.011 Tibiotarsus & Fibula 110.0 2.312 Tarsometatarsus & Digits 110.0 2.3
Figure 4 A) gizzard stones removed from the stomach of the Australian emu Dromaius novaehollandiae Individual B
(*wood); B) Genyornis or emu gizzard stones from late Pleistocene north-west Victoria (photos J. Garvey)
Garvey et al. Modern Emu Butchery
104 Environmental Archaeology 2011 VOL 16 NO 2
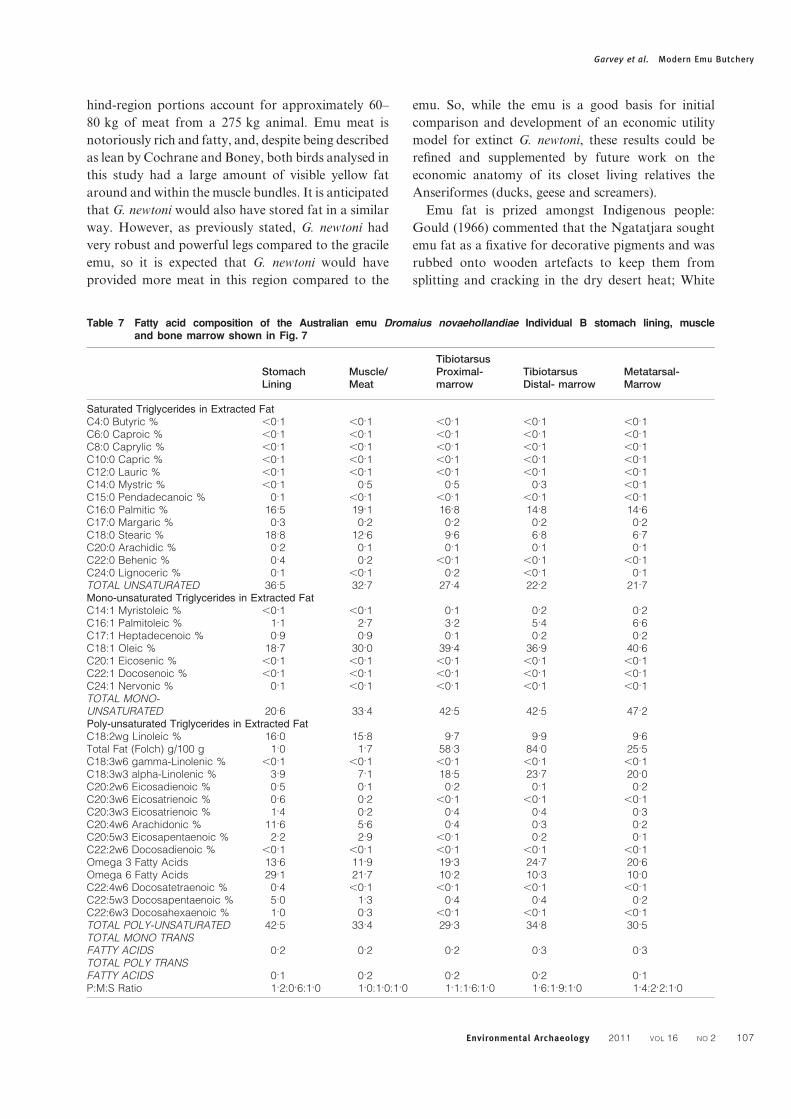
satiety than leaner foods. For these reasons, people
will predictably target these elements. To accurately
evaluate the nutritional value of the emu, samples of
marrow, muscle and stomach lining were submitted
for Fatty Acid Profile (FAMES) assays; 32 fatty acids
were subsequently identified (Table 7). Importantly,
the pattern of marrow content observed in macropods
and placental mammals was found to mirror that in
the emu (Fig. 7).
DiscussionModern hunting and butchery practices
In the semi-arid region of western NSW, the emu is
still taken on a regular basis for food by Indigenous
Australians. Modern technology means it is usually
hunted from a vehicle using a rifle. Both local farmers
and Indigenous people have commented that the
emu’s curiosity makes it easy prey. An object such as
a piece of cloth or a mirror can be waved from a
Table 6 The Modified Meat Utility Index (MMUI) and the %MMUI flesh weights for the Australian emu Dromaiusnovaehollandiae Individual B
Anatomical Unit Flesh Wt (gm) Parts Averaged MMUI (gm) %MMUI
1 Skull & mandible 23.6 None 23.6 0.52 Cervical vertebrate 470.0 None 470.0 9.83 Thoracic vertebrate 200.0 Thoracic, cervical & lumbar 283.3 5.94 Lumbar vertebrate 180.0 Lumbar & thoracic 190 4.05 Sternum 50.0 Sternum & ribs 440.0 9.26 Ribs 830.0 None 830.0 17.37 Pectoral Girdle 4.0 Pectoral girdle, ribs & sternum 294.7 6.18 Forelimb 5.2 Forelimb & ribs 417.6 8.79 Pelvis & sacrum 600.0 Pelvis & femur 2700.0 56.310 Femur 4800.0 None 4800.0 100.011 Tibiotarsus & Fibula 110.0 Tibiotarsus & femur 2455.0 51.112 Tarsometatarsus & Digits 110.0 None 110.0 2.3
Figure 5 Outline of the Australian emu Dromaius novaehollandiae skeleton with the frequency of the; A) Meat Utility
index (MUI) and B) the Modified Meat Utility Index (MMUI) (adapted from Minnear and Minnear 1984, fig. 4.1)
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 105
stationary vehicle to get its attention, drawing in the
curious animal for a closer look. Such practices were
used by the Garawara people in the Northern
Territory (Pickering 1992). Other methods include
shooting an individual and causing injury so the bird
thrashes around, provoking other individuals to
move in for a closer look.
Emus, once caught and killed, may either be
cooked whole in a traditional ground oven, or
butchered or filleted for rissoles or steaks. The
cooking method is usually dictated by the resources
available. If cooking in a ground oven, a hole about a
metre in depth will be dug to accommodate the entire
bird. A fire will be started adjacent to the hole and
left to burn down until coals have formed. The bird is
then placed in the ground oven, with dirt and coals
placed on top for four to six hours or until cooked.
Due to inclement weather we were unable to cook the
birds in a ground-oven and instead butchered the
bird for rissoles and steak and collected the bundal
(stomach lining).
The documented butchering methods (of Cochrane
and Boney) were similar to those reported by
Pickering (1995). Bovid butchery strategies by the
Garawara people in the Gulf of Carpentaria meant
that large quantities of meat were obtained with
minimal disarticulation and/or damage to the carcass.
Notably, the absence of surface modifications to
bones of G. newtoni at Cuddie Springs (Field and
Boles 1998; Fillios et al. 2010), if scavenged or
butchered, may be explained by reference to the
behaviour of modern Aboriginal groups in the
processing of the emu for meat.
Whilst observing the Gunwinggu hunting buffalo
in Arnhem Land, Altman (1982) found that, in all
circumstances, it was butchered where killed. The
primary reason for this was the enormous size of the
buffalo. The highly prized portions, the liver,
stomach lining, heart and tongue, were usually
claimed by the elders. The buffalo fat was also an
important commodity. Once the fat and meat had
been distributed, it was cooked in individual family/
domestic group ground-ovens. However, when the
next largest sized prey, the kangaroo or emu, was
caught, it was first cooked whole in a ground-oven,
and then distributed (Altman 1982). These observa-
tions lend support to the notion that the very large
goose-like bird G. newtoni could have been butchered
where it was killed and the selected portions removed,
then cooked and/or transported.
Economic utility of the Australian emu
The economic utility study of the emu indicates that
approximately 53% of the total body weight of the bird
consists of muscle flesh, and that this was predomi-
nately found around the birds’ hind limb and pelvis
(Table 5, Fig. 5A). In total, 10?2 kg of meat was
removed from this region of the animal and accounted
for almost 25% of the total body weight. When scaled
up for the much larger G. newtoni, the equivalent
Figure 6 Comparison of the %MUI and the %MMUI for the Australian emu Dromaius novaehollandiae, Individual B
Garvey et al. Modern Emu Butchery
106 Environmental Archaeology 2011 VOL 16 NO 2
hind-region portions account for approximately 60–
80 kg of meat from a 275 kg animal. Emu meat is
notoriously rich and fatty, and, despite being described
as lean by Cochrane and Boney, both birds analysed in
this study had a large amount of visible yellow fat
around and within the muscle bundles. It is anticipated
that G. newtoni would also have stored fat in a similar
way. However, as previously stated, G. newtoni had
very robust and powerful legs compared to the gracile
emu, so it is expected that G. newtoni would have
provided more meat in this region compared to the
emu. So, while the emu is a good basis for initial
comparison and development of an economic utility
model for extinct G. newtoni, these results could be
refined and supplemented by future work on the
economic anatomy of its closet living relatives the
Anseriformes (ducks, geese and screamers).
Emu fat is prized amongst Indigenous people:
Gould (1966) commented that the Ngatatjara sought
emu fat as a fixative for decorative pigments and was
rubbed onto wooden artefacts to keep them from
splitting and cracking in the dry desert heat; White
Table 7 Fatty acid composition of the Australian emu Dromaius novaehollandiae Individual B stomach lining, muscleand bone marrow shown in Fig. 7
StomachLining
Muscle/Meat
TibiotarsusProximal-marrow
TibiotarsusDistal- marrow
Metatarsal-Marrow
Saturated Triglycerides in Extracted FatC4:0 Butyric % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C6:0 Caproic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C8:0 Caprylic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C10:0 Capric % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C12:0 Lauric % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C14:0 Mystric % ,0.1 0.5 0.5 0.3 ,0.1C15:0 Pendadecanoic % 0.1 ,0.1 ,0.1 ,0.1 ,0.1C16:0 Palmitic % 16.5 19.1 16.8 14.8 14.6C17:0 Margaric % 0.3 0.2 0.2 0.2 0.2C18:0 Stearic % 18.8 12.6 9.6 6.8 6.7C20:0 Arachidic % 0.2 0.1 0.1 0.1 0.1C22:0 Behenic % 0.4 0.2 ,0.1 ,0.1 ,0.1C24:0 Lignoceric % 0.1 ,0.1 0.2 ,0.1 0.1TOTAL UNSATURATED 36.5 32.7 27.4 22.2 21.7Mono-unsaturated Triglycerides in Extracted FatC14:1 Myristoleic % ,0.1 ,0.1 0.1 0.2 0.2C16:1 Palmitoleic % 1.1 2.7 3.2 5.4 6.6C17:1 Heptadecenoic % 0.9 0.9 0.1 0.2 0.2C18:1 Oleic % 18.7 30.0 39.4 36.9 40.6C20:1 Eicosenic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C22:1 Docosenoic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C24:1 Nervonic % 0.1 ,0.1 ,0.1 ,0.1 ,0.1TOTAL MONO-UNSATURATED 20.6 33.4 42.5 42.5 47.2Poly-unsaturated Triglycerides in Extracted FatC18:2wg Linoleic % 16.0 15.8 9.7 9.9 9.6Total Fat (Folch) g/100 g 1.0 1.7 58.3 84.0 25.5C18:3w6 gamma-Linolenic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1C18:3w3 alpha-Linolenic % 3.9 7.1 18.5 23.7 20.0C20:2w6 Eicosadienoic % 0.5 0.1 0.2 0.1 0.2C20:3w6 Eicosatrienoic % 0.6 0.2 ,0.1 ,0.1 ,0.1C20:3w3 Eicosatrienoic % 1.4 0.2 0.4 0.4 0.3C20:4w6 Arachidonic % 11.6 5.6 0.4 0.3 0.2C20:5w3 Eicosapentaenoic % 2.2 2.9 ,0.1 0.2 0.1C22:2w6 Docosadienoic % ,0.1 ,0.1 ,0.1 ,0.1 ,0.1Omega 3 Fatty Acids 13.6 11.9 19.3 24.7 20.6Omega 6 Fatty Acids 29.1 21.7 10.2 10.3 10.0C22:4w6 Docosatetraenoic % 0.4 ,0.1 ,0.1 ,0.1 ,0.1C22:5w3 Docosapentaenoic % 5.0 1.3 0.4 0.4 0.2C22:6w3 Docosahexaenoic % 1.0 0.3 ,0.1 ,0.1 ,0.1TOTAL POLY-UNSATURATED 42.5 33.4 29.3 34.8 30.5TOTAL MONO TRANSFATTY ACIDS 0.2 0.2 0.2 0.3 0.3TOTAL POLY TRANSFATTY ACIDS 0.1 0.2 0.2 0.2 0.1P:M:S Ratio 1.2:0.6:1.0 1.0:1.0:1.0 1.1:1.6:1.0 1.6:1.9:1.0 1.4:2.2:1.0
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 107
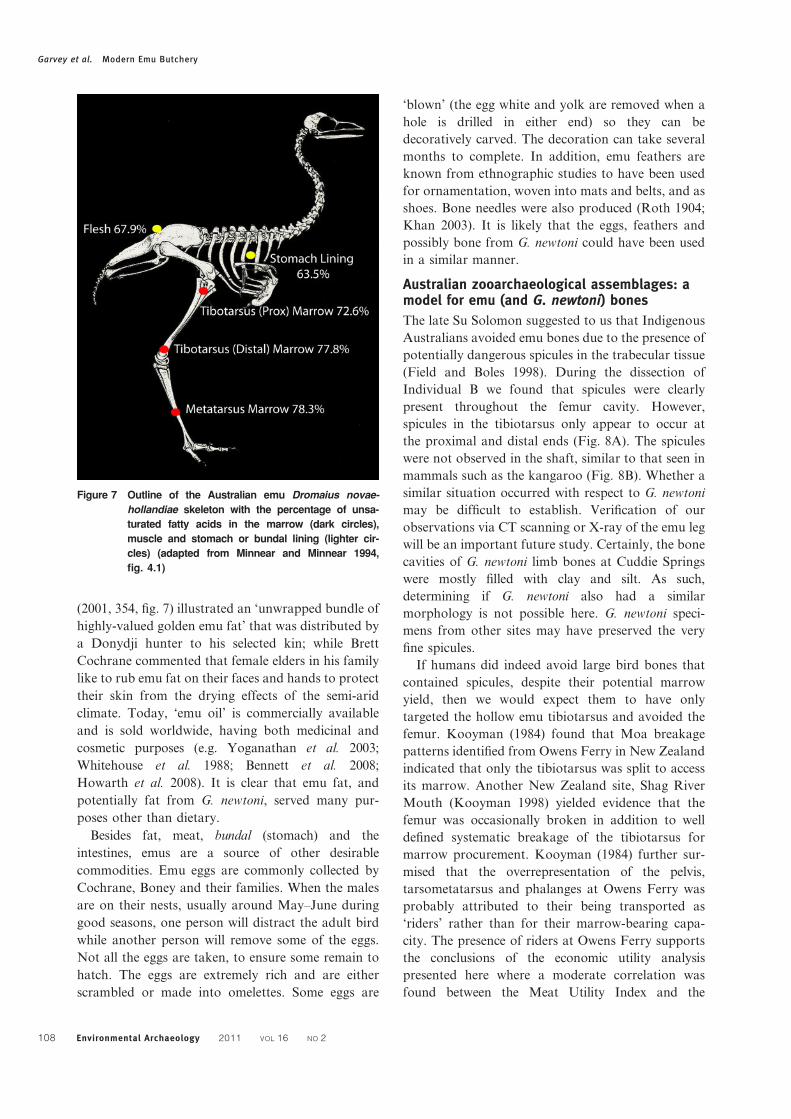
(2001, 354, fig. 7) illustrated an ‘unwrapped bundle of
highly-valued golden emu fat’ that was distributed by
a Donydji hunter to his selected kin; while Brett
Cochrane commented that female elders in his family
like to rub emu fat on their faces and hands to protect
their skin from the drying effects of the semi-arid
climate. Today, ‘emu oil’ is commercially available
and is sold worldwide, having both medicinal and
cosmetic purposes (e.g. Yoganathan et al. 2003;
Whitehouse et al. 1988; Bennett et al. 2008;
Howarth et al. 2008). It is clear that emu fat, and
potentially fat from G. newtoni, served many pur-
poses other than dietary.
Besides fat, meat, bundal (stomach) and the
intestines, emus are a source of other desirable
commodities. Emu eggs are commonly collected by
Cochrane, Boney and their families. When the males
are on their nests, usually around May–June during
good seasons, one person will distract the adult bird
while another person will remove some of the eggs.
Not all the eggs are taken, to ensure some remain to
hatch. The eggs are extremely rich and are either
scrambled or made into omelettes. Some eggs are
‘blown’ (the egg white and yolk are removed when a
hole is drilled in either end) so they can be
decoratively carved. The decoration can take several
months to complete. In addition, emu feathers are
known from ethnographic studies to have been used
for ornamentation, woven into mats and belts, and as
shoes. Bone needles were also produced (Roth 1904;
Khan 2003). It is likely that the eggs, feathers and
possibly bone from G. newtoni could have been used
in a similar manner.
Australian zooarchaeological assemblages: amodel for emu (and G. newtoni) bones
The late Su Solomon suggested to us that Indigenous
Australians avoided emu bones due to the presence of
potentially dangerous spicules in the trabecular tissue
(Field and Boles 1998). During the dissection of
Individual B we found that spicules were clearly
present throughout the femur cavity. However,
spicules in the tibiotarsus only appear to occur at
the proximal and distal ends (Fig. 8A). The spicules
were not observed in the shaft, similar to that seen in
mammals such as the kangaroo (Fig. 8B). Whether a
similar situation occurred with respect to G. newtoni
may be difficult to establish. Verification of our
observations via CT scanning or X-ray of the emu leg
will be an important future study. Certainly, the bone
cavities of G. newtoni limb bones at Cuddie Springs
were mostly filled with clay and silt. As such,
determining if G. newtoni also had a similar
morphology is not possible here. G. newtoni speci-
mens from other sites may have preserved the very
fine spicules.
If humans did indeed avoid large bird bones that
contained spicules, despite their potential marrow
yield, then we would expect them to have only
targeted the hollow emu tibiotarsus and avoided the
femur. Kooyman (1984) found that Moa breakage
patterns identified from Owens Ferry in New Zealand
indicated that only the tibiotarsus was split to access
its marrow. Another New Zealand site, Shag River
Mouth (Kooyman 1998) yielded evidence that the
femur was occasionally broken in addition to well
defined systematic breakage of the tibiotarsus for
marrow procurement. Kooyman (1984) further sur-
mised that the overrepresentation of the pelvis,
tarsometatarsus and phalanges at Owens Ferry was
probably attributed to their being transported as
‘riders’ rather than for their marrow-bearing capa-
city. The presence of riders at Owens Ferry supports
the conclusions of the economic utility analysis
presented here where a moderate correlation was
found between the Meat Utility Index and the
Figure 7 Outline of the Australian emu Dromaius novae-
hollandiae skeleton with the percentage of unsa-
turated fatty acids in the marrow (dark circles),
muscle and stomach or bundal lining (lighter cir-
cles) (adapted from Minnear and Minnear 1994,
fig. 4.1)
Garvey et al. Modern Emu Butchery
108 Environmental Archaeology 2011 VOL 16 NO 2

Modified Meat Utility Index. It supports our
contention that riders are an important consideration
when assessing emu body part representation in
archaeological assemblages.
While emu bone marrow is very high in unsatu-
rated fats, there is already enough fat on the body
(Fig. 3) to make accessing the bone marrow non-
essential. The reverse situation exists for the
Australian macropod. There is so little dissectible
fat available in kangaroos and wallabies that people
needed to supplement their diets by cracking open the
longbones to access the bone marrow (Garvey 2011).
Gastroliths or gizzard stones in the zooarchaeological
record
Due to the lack of grinding teeth, a variety of birds
ingest small stones and/or fragments of wood,
referred to as gastroliths or gizzard stones, into their
gastrointestinal tracts to aid with mechanical diges-
tion. Other animals that also use gastroliths include
crocodiles, alligators, seals, sea lions and dinosaurs.
Both of the emus used in this study had their
stomachs checked for gizzard stones. Interestingly,
only the emu used in the economic utility study
(Individual B) had any present, with 11 gastroliths
collected (Fig. 4A). Of these, two were found to be
pieces of wood that would have eventually been
digested in the bird’s stomach, while the remaining
nine were stone. Gizzard stones are important as they
can potentially be used to identify specific taxa in the
archaeological and palaeontological record (Fig. 4B).
Surprisingly, the emu gizzard stones were not
rounded from mechanical processing in the emu’s
stomach. The lack of rounding contrasts with
expectations based on the morphology of ratite
gastroliths identified elsewhere. Gastroliths have
commonly been described as being rounded, worn
and polished (Anderson et al. 1998). Recent experi-
ments by Wings and Sander (2007) suggested that
perhaps they are not as polished as previously
believed, suggesting that there is significant varia-
bility, conceivably depending on context and local
environmental conditions. The lack of gizzard stones
in Individual A further implies that emus may replace
their gizzard stones on a regular basis. Additional
investigation of emu gizzard stones, and the implica-
tions for G. newtoni, is required.
Conclusion
This paper presents the first modern butchery,
economic utility and fatty acid analysis of the
Australian emu Dromaius novaehollandiae. It has
important implications for understanding the eco-
nomic role of the emu in modern Indigenous society
and also in the Australian archaeological record. The
study of the emu provides tantalising new evidence
that may be extrapolated to the extinct giant goose of
Australia’s late Pleistocene G. newtoni. Following
from Pickering (1995), the large amounts of easily
accessible meat and fat provided by one G. newtoni
carcass combined with low human population
numbers at this time means that the likelihood of
physical damage to bones would be low. It necessarily
follows that detecting a human signature in the
scavenging or butchering of these birds may be
equally difficult.
Specific outcomes include:
1) The portions of greatest economic value in the
modern emu are the pelvis, hind region, stomach
lining and the intestines. Feathers, eggs and emu
Figure 8 Tibiotarsus of the Australian emu Dromaius novaehollandiae individual B, where A) spicules are only located at
the proximal and distal end of the bone (spicules are arrowed), and B) the shaft of the tibiotarsus does not con-
tain spicules with the marrow resembling that found in mammal longbones (photos J. Garvey)
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 109

fat are also important commodities. Hence, it is
predicted that the leg bones of the emu and/or G.
newtoni are the most likely elements to be present
in archaeological consumption or habitation sites.
2) A 42 kg emu yields approximately 10?2 kg of
meat from the pelvis and leg. Therefore, G.
newtoni is likely to have provided 60 to 80 kg of
meat from its hind region alone.
3) Large amounts of meat and fat can be obtained
from an emu with minimal effort; hence G.
newtoni bones are unlikely to have sustained
damage during defleshing.
4) As with the emu, G. newtoni is likely to have had
substantial amounts of fat associated with the
muscle. This means that it was not necessary to
supplement the already fatty meat with the bird’s
fatty marrow. Hence we predict that emu and G.
newtoni longbones are likely to be found complete
and not cracked open.
5) If people did access emu longbones for their large
marrow yield, then given the observed pattern of
spicule distribution we would expect to find only
the tibiotarsus bones and not the femora cracked
open in archaeological sites. Further investigation
is required to determine the distribution of
spicules in G. newtoni longbones.
Acknowledgements
We are very grateful to Margaret and Ted Johnstone
for their generous logistical support in the field and
their continuing friendship. We are indebted to the
Currey and Green families for help, advice and access
to their properties during this work. Grateful thanks
are also due to the Brewarrina Aboriginal
Community and the Walgett Shire Council and the
many volunteers who helped us. Brad Orcher
provided field assistance and made great rissoles
from the emu meat. The authors acknowledge the
facilities as well as scientific and technical assistance
from the staff in the Australian Microscopy and
Microanalysis Research Facility (AMMRF) and the
Australian Centre for Microscopy and Microanalysis
at the University of Sydney under whose auspices this
work was undertaken. The project was funded by the
Australian Research Council (DP0557923) and
the University of Sydney. This paper is dedicated to
the late Su Solomon, who first brought to our
attention the lack of cracked emu longbones in the
Australian archaeological record.
ReferencesAllen, J. (ed.) 1996. Report of the Southern Forests Archaeological
Project, Volume 1. Bundoora: School of Archaeology, La Trobe
University.
Altman, J. C. 1982. Hunting buffalo in North-Central Arnhem Land: a
case of rapid adaptation among Aborigines. Oceania 52, 274–85.
Anderson, A., Worthy, T. and McGovern-Wilson, R. 1998. Moa
remains and taphonomy, pp. 200–13 in Anderson, A., Allingham,
A. and Smith, I. (eds.), Shag River Mouth: The Archaeology of an
Early Southern Maori Village. Canberra: The Australian National
University.
Bear, G. D. 1971. Seasonal trends in fat levels of pronghorns,
Antilocapra americana, in Colorado. Journal of Mammalogy 52,
583–89.
Bennett, D. C., Code W. E., Godin, D. V. and Cheng, K. M. 2008.
Comparison of the antioxidant properties of emu oil with other
avian oils. Australian Journal of Experimental Agriculture 48,
1345–50.
Berge, P., Lepetit, J., Renerre, M. and Touraille, C. 1997. Meat quality
traits in the emu (Dromaius novaehollandiae) as affected by muscle
type and animal age. Meat Science 45, 209–21.
Binford, L. R. 1978. Nunamuit Ethnoarchaeology. New York:
Academic Press.
Blumenschine, R. J. and Caro, T. M. 1986. Unit flesh weights of some
East African bovids. African Journal of Ecology 24, 273–86.
Brown, O., Field, F. and Letnic, M. 2006. Variation in the taphonomic
effect of scavengers in semi-arid Australia linked to rainfall and
the El Nino Southern Oscillation. International Journal of
Osteoarchaeology 16, 165–76.
Burger, O., Hamilton, M. J. and Walker, R. 2005. The prey as patch
model: optimal handling of resources with diminishing returns.
Journal of Archaeological Science 32, 1147–58.
Cederlund, G. N., Bergstrom, R. L. and Danell, K. 1989. Seasonal
variation in mandible marrow fat in moose. Journal of Wildlife
Management 53, 587–92.
Cosgrove, R. and Allen, J. 2001. Prey choice and hunting strategies in
the Late Pleistocene: evidence from southwest Tasmania, pp. 397–
429 in Anderson, A., Lilley, I. and O’Connor, S. (eds.), Histories
of Old Ages: Essays in Honour of Rhys Jones. Canberra: Pandanus
Books.
Cupper, M. L. and Duncan, J. 2006. Last glacial megafaunal death
assemblage and early human occupation at Lake Menindee,
southeastern Australia. Quaternary Research 66, 332–41.
Davidson, I. 2010. The colonization of Australia and its adjacent
islands and the evolution of modern cognition. Current
Anthropology 51 (Supplement 1), s177–s189.
Denham, T., Fullagar, R. and Head, L. 2009. Plant exploitation on
Sahul: from colonisation to the emergence of regional specialisa-
tion during the Holocene. Quaternary International 202, 29–40.
Dietz, A. A. 1946. Composition of normal bone marrow in rabbits.
Journal of Biological Chemistry 165, 505–11.
Diab, M. C. 1998. Economic utility of the ringed seal (Phoca hispida):
implications for Arctic archaeology. Journal of Archaeological
Science 25, 1–26.
Dodson, J. R., Fullagar, R., Furby, J. H. and Prosser, I. P. 1993.
Humans and megafauna in a Late Pleistocene environment at
Cuddie Springs, northwestern New South Wales. Archaeology in
Oceania 28, 93–99.
Dortch, J. 1996. Late Pleistocene and recent Aboriginal occupation at
Tunnel Cave and Witchcliffe Rock Shelter, Naturaliste Region,
south-western Australia. Australian Aboriginal Studies 1996(2),
51–60.
Field, J. 2006. Trampling the Pleistocene: does taphonomy matter at
Cuddie Springs? Australian Archaeology 63, 9–20.
Field, J. and Dodson, J. 1999. Late Pleistocene megafauna and human
occupation at Cuddie Springs, southeastern Australia. Proceedings
of the Prehistoric Society 65, 275–301.
Field, J. H. and Boles, W. E. 1998. Genyornis newtoni and Dromaius
novaehollandiae at 30,000 b.p. in central northern New South
Wales. Alcheringa 22, 177–88.
Field, J., Fillios, M. and Wroe, S. 2008. Chronological overlap between
humans and megafauna in Sahul (Pleistocene Australia–New
Guinea): a review of the evidence. Earth Science Reviews 89, 97–
115.
Field, J. H., Wroe, S., Trueman, C. N., Garvey, J. and Wyatt-Spratt,
S. J. 2011. Looking for the archaeological signature in Australian
megafaunal extinctions. Quaternary International. DOI:10.1016/
j.quaint.2011.04013.
Garvey et al. Modern Emu Butchery
110 Environmental Archaeology 2011 VOL 16 NO 2

Fillios, M., Field, J. and Charles, B. 2010. Investigating human and
megafauna co-occurrence in Australian prehistory: mode and
causality in fossil accumulations at Cuddie Springs. Quaternary
International 211, 123–43.
Fong, D. W. 1981. Seasonal variation of marrow fat content from
Newfoundland moose. Journal of Wildlife Management 45, 545–
48.
Garvey, J. 2006. Preliminary zooarchaeological interpretations from
Kutikina Cave, southwest Tasmania. Australian Aboriginal Studies
2006(1), 58–63.
Garvey, J. 2007. Surviving an Ice Age: the zooarchaeology from SW
Tasmania. Palaios 22(6), 583–85.
Garvey, J. 2010. Economic anatomy of the Bennett’s wallaby
(Macropus rufogriseus): implications for understanding human
hunting strategies in late Pleistocene Tasmania. Quaternary
International 211, 144–56.
Garvey, J. 2011. Bennett’s wallaby (Macropus rufogriseus) bone
marrow quality vs quantity: evaluating human decision making
and seasonal occupation in late Pleistocene Tasmania. Journal of
Archaeological Science 38, 763–83.
Giardina, M. 2006. Anatomıa economica de Rheidae. Intersecciones en
Antropologia 7, 263–76.
Gillespie, R., Horton, D. R., Ladd, P., Macumber, P. G., Rich, T. H.,
Thorne, R. and Wright, R. V. S. 1978. Lancefield Swamp and the
extinction of the Australian megafauna. Science 200, 1044–48.
Gould, R. A. 1966. Notes on hunting, butchering, and sharing of game
among the Ngatatjara and their neighbours in the West Australian
desert. Kroeber Anthropological Society Papers 36, 41–46.
Gould, R. A. 1969a. Yiwara: Foragers of the Australian Desert.
London: Collins.
Gould, R. A. 1969b. Subsistence behaviour among the Western Desert
Aborigines of Australia. Oceania 38, 253–74.
Gould, R. A. 1981. Comparative ecology of food-sharing in Australia
and northwest California, pp. 422–54 in Harding, R. S. O. and
Teleki, G. (eds.), Omnivorous Primates: Gathering and Hunting in
Human Evolution. New York: Columbia University Press.
Grayson, D. K. 1989. Bone transport, bone destruction, and reverse
utility curves. Journal of Archaeological Science 16, 643–52.
Hockett, B. and Haws, J. A. 2003. Nutritional ecology and diachronic
trends in Paleolithic diet and health. Evolutionary Anthropology
12, 211–16.
Hockett, B. and Haws, J. A. 2005. Nutritional ecology and the human
demography of Neanderthal extinction. Quaternary International
137, 21–34.
Howarth, G. S., Lindsay, R. J., Butler, R. N. and Geier, M. S. 2008.
Can emu oil ameliorate inflammatory disorders affecting the
gastrointestinal system? Australian Journal of Experimental
Agriculture 48, 1276–79.
Johnson, C. N. 2005. What can the data on late survival of Australian
megafauna tell us about the cause of their extinction? Quaternary
Science Reviews 24, 2167–72.
Jones, K. T. and Metcalfe, D. 1988. Bare bones archaeology: bone
marrow indices and efficiency. Journal of Archaeological Science
15, 415–23.
Khan, K. 2003. Catalogue of the Roth collection of aboriginal artefacts
from North Queensland, Volume 3. Items collected from
McDonnell Electric Telegraph Office, McIvor River, Mapoon
and the Pennefather and Wenlock Rivers, Maytown, Mentana,
Mitchell River, Morehead River, Moreton Electric Telegraph
Office and Musgrave, in 1897–1903. Technical Reports of the
Australian Museum 17, 1–106.
Kooyman, B. 1984. Moa utilisation at Owens Ferry, Otago, New
Zealand. New Zealand Journal of Archaeology 6, 47–57.
Kooyman, B. 1998. Moa hunting and butchery, pp. 214–22 in
Anderson, A., Allingham, A. and Smith, I. (eds.), Shag River
Mouth: The Archaeology of an Early Southern Maori Village.
Canberra: The Australian National University.
Lyman, R. L. 1992. Anatomical considerations of utility curves in
zooarchaeology. Journal of Archaeological Science 19, 7–22.
Lyman, R. L. 1994. Vertebrate Taphonomy. Cambridge: Cambridge
University Press.
Lyman, R. L., Savelle, J. M. and Whitridge, P. 1992. Derivation and
application of a meat utility index for phocid seals. Journal of
Archaeological Science 19, 531–55.
McArthur, M. 1948. Food consumption and dietary levels of groups of
Aborigines living on naturally occurring foods, pp. 90–135 in
Mountford, C. P. (ed.), Records of the American-Australian
Scientific Expedition to Arnhem Land Volume 2: Anthropology
and Nutrition. Melbourne: Melbourne University Press.
Madrigal, T. C. and Capaldo, S. D. 1999. White-Tailed Deer marrow
yields and Late Archaic Hunter-Gatherers. Journal of
Archaeological Science 26, 241–49.
Meng, M. S., West, G. C. and Irving, L. 1969. Fatty acid composition
of caribou bone marrow. Comparative Biochemistry and
Physiology 30, 187–91.
Metcalfe, D. and Jones, K. T. 1988. A reconsideration of animal body-
part utility indices. American Antiquity 53, 486–504.
Miller, G. H., Magee, J. W., Johnson, B. J., Fogel, M. L., Spooner,
N. A., McCulloch, M. T. and Ayliffe, L. K. 1999. Pleistocene
extinction of Genyornis newtoni: human impact on Australian
megafauna. Science 283, 205–08.
Minnear, P. and Minnear, M. 1994. The Emu Farmer’s Handbook.
Texas: Induna Company.
Morin, E. 2007. Fat composition and Nunamiut decision-making: a
new look at the marrow and bone grease indices. Journal of
Archaeological Science 34, 69–82.
Murray, P. F. and Megirian, D. 1998. The skull of the dromornithid
birds: anatomical evidence for their close relationship to
Anseriformes. Records of the South Australian Museum 31, 51–97.
Murray, P. F. and Vickers-Rich, P. 2004. Magnificent Mihirungs: The
Colossal Flightless Birds of the Australian Dreamtime.
Bloomington: Indiana University Press.
O’Connell, J. F. 2000. An emu hunt, pp. 172–81 in Anderson, A. and
Murray, T. (eds), Australian Archaeologist: Collected Papers in
Honour of Jim Allen (Division of Archaeology and Natural
History, Research School of Pacific and Asian Studies). Canberra:
Australian National University.
O’Connell, J. F. and Allen, J. 2004. Dating the colonization of Sahul
(Pleistocene Australia-New Guinea): a review of recent research.
Journal of Archaeological Science 31, 835–53.
O’Connell, J. F. and Marshall, B. 1989. Analysis of kangaroo body part
transport among the Alyawara of Central Australia. Journal of
Archaeological Science 16, 393–405.
O’Dea, K. 1991. Traditional diet and food preferences of Australian
aboriginal hunter-gatherers. Proceedings of the Royal Society of
London, Series B Biological Sciences 334, 233–40.
Outram, A. K. 2002. Bone fracture and within-bone nutrients: an
experimentally based method for investigating levels of marrow
extraction, pp. 51–64 in Miracle, P. and Milner, N. (eds.),
Consuming Passions and Patterns of Consumption. Cambridge:
McDonald Institute for Archaeological Research.
Outram, A. and Rowley-Conwy, P. 1998. Meat and marrow utility
indices for horse (Equus). Journal of Archaeological Science 25,
839–49.
Ouzman, S., Tacon, P. S. C., Mulvaney, K. and Fullagar, R. 2002.
Extraordinary engraved bird track from North Australia: extinct
fauna, dreaming being and/or aesthetic masterpiece? Cambridge
Archaeological Journal 12, 103–12.
Pickering, M. 1992. Garawa methods of game hunting, preparation and
cooking. Records of the South Australian Museum 26, 9–23.
Pickering, M. 1995. Notes on Aboriginal hunting and butchering of
cattle and buffalo. Australian Archaeology 40, 17–21.
Pike-Tay, A., Cosgrove, R. and Garvey, J. 2008. Systematic seasonal
land use by late Pleistocene Tasmanian Aborigines. Journal of
Archaeological Science 35, 2532–44.
Pizzey, G. and Knight, F. 2001. The Field Guide to the Birds of
Australia. Sydney: Angus and Robertson.
Pond, C. M. 1988. The Fats of Life. Cambridge University Press:
Cambridge.
Rich, P. 1979. The Dromornithidae, an extinct family of large ground
birds endemic to Australia. Bulletin of the Bureau of Mineral
Resources, Geology and Geophysics 184, 1–190.
Rich, P. and Gill, E. 1978. Possible Dromornithid footprints from
Pleistocene dune sands of southern Victoria, Australia. The Emu
76, 221–23.
Reitz, E. J. and Wing, E. S. 1999. Zooarchaeology. Cambridge:
Cambridge University Press.
Roth, W. E. 1901. Food: its search, capture, and preparation. North
Queensland Ethnography Bulletin 3, 7–31.
Garvey et al. Modern Emu Butchery
Environmental Archaeology 2011 VOL 16 NO 2 111

Roth, W. E. 1904. North Queensland Ethnography Bulletin 7: Domestic
Implements, Arts and Manufactures. Brisbane: Government
Printer.
Sales, J. and Horbanczuk, J. 1998. Ratite meat. World’s Poultry Science
Journal 54, 59–67.
Savelle, J. M. 1997. The role of architectural utility in the formation of
zooarchaeological whale bone assemblages. Journal of
Archaeological Science 24, 869–85.
Savelle, J. M. and Friesen, T. M. 1996. An odontocete (Cetacea) meat
utility index. Journal of Archaeological Science 23, 713–21.
Savelle, J. M., Friesen, T. M. and Lyman, R. L. 1996. Derivation and
application of an otariid utility index. Journal of Archaeological
Science 23, 705–12.
Shao, C. H., Avens, J. S., Schmidt, G. R. and Maga, J. A. 1999.
Functional, sensory, and microbiological properties of restruc-
tured beef and emu steaks. Journal of Food Science 64, 1052–54.
Smetana, P. 1993. Emu Farming: Background Information. Department
of Agriculture Western Australia.
Speth, J. D. 1987. Early hominid subsistence strategies in seasonal
habitats. Journal of Archaeological Science 14, 13–29.
Speth, J. D. 1989. Early Hominid hunting and scavenging: the role of
meat as an energy source. Journal of Human Evolution 18, 329–43.
Speth, J. D. 1991. Nutritional constraints and Late Glacial adaptive
transformations: the importance of non-protein energy sources,
pp. 169–78 in Barton, N., Roberts, A. J. and Roe, D. A. (eds.),
The Late Glacial in North-West Europe: Human Adaptation and
Environmental Change at the End of the Pleistocene (CBA
Research Report No. 77). Council for British Archaeology:
London.
Speth, J. D. 2010. The Paleoanthropology and Archaeology of Big-Game
Hunting: Protein, Fat, or Politics? Springer: New York.
Speth, J. D. and Spielmann, K. A. 1983. Energy source, protein
metabolism, and hunter-gatherer subsistence strategies. Journal of
Anthropological Archaeology 2, 1–31.
Stirling, E. C. 1896. The newly discovered extinct gigantic bird of South
Australia. Ibis 7, 593.
Stirling, E. C. and Zietz, A. H. 1896. Preliminary notes on Genyornis
newtoni: a new genus and species of fossil struthious bird found at
Lake Callabonna, South Australia. Transactions and Proceedings
of the Royal Society of South Australia 20, 171–90.
Stirling, E. C. and Zietz, A. H. 1890. Fossil remains at Lake
Callabonna. Part 1: Genyornis Newtoni: a new genus and species
of struthious bird. Memoirs of the Royal Society of South Australia
1, 41–80.
Summerhayes, G., Leavesley, M., Fairbairn, A., Mandui, H., Field, J.,
Ford, A., and Fullagar, R. 2010. Human adaptation and plant use
in highland New Guinea 49,000 to 44,000 years ago. Science 330,
78–81.
Sutton, A., Mountain, M.-J., Aplin, K., Bulmer, S. and Denham, T.
2009. Archaeozoological records for the Highlands of New
Guinea: a review of current evidence. Australian Archaeology 69,
41–58.
Thomas, D. H. and Mayer, D. 1983. Behavioural faunal analysis of
selected horizons, in Thomas, D. H. (ed.), The archaeology of
Monitor Valley 2: Gatecliff Shelter. Anthropological Papers of the
American Museum of Natural History 58, 353–90.
Thomson, D. F. 1939. The seasonal factor in human culture: illustrated
from the life of a contemporary nomadic group. Proceedings of the
Prehistoric Society 5, 209–22.
Turner, J. C. 1979. Adaptive strategies of selective fatty acid deposition
in the bone marrow of desert bighorn sheep. Comparative
Biochemistry and Physiology 62A, 599–604.
West, G. and Shaw, D. 1975. Fatty acid composition of Dall sheep
bone marrow. Comparative Biochemistry and Physiology 50B, 599–
601.
White, N. G. 2001. In search of the traditional Australian Aboriginal
diet — then and now, pp. 343–59 in Allen, J. (ed.), Rhys Jones’
Festschrift Volume of Essays. Department of Archaeology and
Natural History Research School of Pacific and Asian Studies,
ANU: Canberra.
Whitehouse, M. W., Turner, A. G., Davis, C. K. C. and Roberts, M. S.
1998. Emu oils(s): a source of non-toxic transdermal anti-
inflammatory agents in Aboriginal medicine. lnflammopharmacol-
ogy 6, 1–8.
Williams, D. L. G. 1981. Genyornis eggshell (Dromornithidae; Aves)
from the Late Pleistocene of South Australia. Alcheringa 5, 133–
40.
Wings, O. and Sander, P. M. 2007. No gastric mill in sauropod
dinosaurs: new evidence from analysis of gastrolith mass and
function in ostriches. Proceedings of the Royal Society B 274, 635–
40.
Wroe, S. and Field, J. 2006. A review of the evidence for a human role
in the extinction of Australian Megafauna and an alternative
explanation. Quaternary Science Reviews 25, 2692–2703.
Wroe, S., Field, J., Fullagar, R. and Jermiin, L. 2004. Megafaunal
extinction in the Late Quaternary and the global overkill
hypothesis. Alcheringa 28, 291–332.
Wurster, C. M., Bird, M. I., Bull, I. D., Creed, F., Bryant, C., Dungait,
J. A. J. and Paz, V. 2010. Forest contraction in north equatorial
Southeast Asia during the Last Glacial Period. Proceedings of the
National Academy of Sciences 107, 15509—11.
Yoganathan, S., Nicolosi, R., Wilson, T., Handelman, G., Scollin, P.,
Tao, R., Binford, P. and Orthoefer, F. 2003 Antagonism of croton
oil inflammation by topical emu oil in CD-1 mice. Lipids 38, 603–
07.
Garvey et al. Modern Emu Butchery
112 Environmental Archaeology 2011 VOL 16 NO 2
Related Documents









![Rethinking megafauna - CentAURcentaur.reading.ac.uk/89222/1/Moleon_et_al_2020_ProcB.pdf · 2021. 7. 9. · 86 megafauna [6-7], the ubiquity of the megafauna concept in addressing](https://static.cupdf.com/doc/110x72/613e8c8669193359046d3023/rethinking-megafauna-2021-7-9-86-megafauna-6-7-the-ubiquity-of-the-megafauna.jpg)

