Embryonic pig pancreatic tissue for the treatment of diabetes in a nonhuman primate model Gil Hecht a,1 , Smadar Eventov-Friedman a,1 , Chava Rosen a , Elias Shezen a , Dalit Tchorsh a , Anna Aronovich a , Enrique Freud b , Hana Golan a , Ronit El-Hasid c , Helena Katchman a , Bernhard J. Hering d , Amnon Zung e , Zipi Kra-Oz f , Pninit Shaked-Mishan f , Alex Yusim g , Alex Shtabsky h , Pavel Idelevitch a , Ana Tobar i , Alon Harmelin j , Esther Bachar-Lustig a , and Yair Reisner a,2 a Department of Immunology, Weizmann Institute of Science, Rehovot 76100, Israel; b Department of Pediatric Surgery, Schneider Children’s Medical Center of Israel, Petach Tikvah 49202, Israel; c Department of Pediatric Hemato-Oncology, Meyer Children Hospital, Haifa 31096, Israel; d Diabetes Institute for Immunology Transplantation, University of Minnesota, Minneapolis, MN, 55455; e Department of Pediatrics, Kaplan Medical Center, Rehovot 76100, Israel; f Department of Virology, Rambam Medical Center, Haifa 31096, Israel; g Department of Transplantation, Rabin Medical Center, Petah Tikva, Israel; h Department of Pathology, Sourasky Medical Center, Tel-Aviv 64239, Israel; i Department of Pathology, Rabin Medical Center, Petah Tikva, Israel; and j Department of Veterinary Resources, Weizmann Institute of Science, Rehovot 76100, Israel Edited by George Klein, Karolinska Institutet, Stockholm, Sweden, and approved April 3, 2009 (received for review December 10, 2008) Xenotransplantation of pig tissues has great potential to overcome the shortage of organ donors. One approach to address the vigorous immune rejection associated with xenotransplants is the use of embryonic precursor tissue, which induces and utilizes host vasculature upon its growth and development. Recently, we showed in mice that embryonic pig pancreatic tissue from embry- onic day 42 (E42) exhibits optimal properties as a cell replacement therapy. We now demonstrate the proof of concept in 2 diabetic Cynomolgus monkeys, followed for 393 and 280 days, respectively. A marked reduction of exogenous insulin requirement was noted by the fourth month after transplantation, reaching complete independence from exogenous insulin during the fifth month after transplantation, with full physiological control of blood glucose levels. The porcine origin of insulin was documented by a radio- immunoassay specific for porcine C-peptide. Furthermore, the growing tissue was found to be predominantly vascularized with host blood vessels, thereby evading hyperacute or acute rejection, which could potentially be mediated by preexisting anti-pig anti- bodies. Durable graft protection was achieved, and most of the late complications could be attributed to the immunosuppressive pro- tocol. While fine tuning of immune suppression, tissue dose, and implantation techniques are still required, our results demonstrate that porcine E-42 embryonic pancreatic tissue can normalize blood glucose levels in primates. Its long-term proliferative capacity, its revascularization by host endothelium, and its reduced immuno- genicity, strongly suggest that this approach could offer an attrac- tive replacement therapy for diabetes. immune-suppression rejection xeno-transplantation A llogeneic islet cell replacement can restore normoglycemia in patients with type 1 diabetes; yet, the shortage of available organs for transplantation continues to severely limit this therapeutic option. A potential solution for this shortage is the use of pig organs and tissues as a source for transplantation. However, the immunological barriers to xenotransplantation are formidable (1). Thus, new approaches that could further reduce the intensity of the required immune suppression are critical for successful application of pig pancreatic tissue as a novel source of islet cells. One means to address this challenge could be provided by the use of embryonic porcine tissue. This strategy is based on the growing evidence, over the past 5 decades, dem- onstrating maternal immune tolerance to the fetus (2, 3), and on recent observations that embryonic tissues exhibit reduced im- munogenicity in various transplantation settings (4–7). Consid- ering that very early embryonic tissues are associated with a substantial risk of teratoma formation, we have attempted, during the past several years, to define the earliest gestational time point that does not pose a teratoma risk for transplantation of different embryonic pig tissues, including kidney (6), heart (unpublished), spleen (8), pancreas, liver, and lung (7) into SCID mice. Once this threshold was established, we were able to further characterize an optimal gestational window for trans- plantation based on growth potential and immunogenicity. Thus, while the optimal window for kidney transplantation was defined around E28, our data suggested E42 as the optimal gestational time point for pig donor tissue in pancreatic transplantation (9). Importantly, upon growth and development of the embryonic pancreatic implant, acini gradually disappear and the tissue is predominantly comprised of islets and stroma identified by vimentin straining. Thus, the risk of potential ongoing destruc- tion of the growing organ by local proteolytic enzymes, which might be released by the exocrine components of the implant, is miniml (9). These mouse studies also demonstrated the curative potential of such implants in diabetic mice under relatively tolerable immune suppression. However, considering that xeno- geneic transplantation into primates is far more complex, due to preexisting anti-porcine antibodies, studies in a nonhuman pri- mate (NHP) model for diabetes were required. Our working hypothesis, suggesting that embryonic tissue transplantation would not be adversely affected by such a humoral response was based on 2 major observations: (i) hyperacute and acute rejec- tion are mainly mediated by complement activation in blood vessels, mediated by preformed anti-pig antibodies recognizing -gal and other carbohydrates expressed on endothelial cells (10), and (ii) data from our mouse studies showing that the embryonic implants predominantly induce host-type vasculature to support their growth and development in the recipient, resulting in an organ comprised mainly of porcine epithelial cells and host endothelium. To further assess the curative potential of E42 pig pancreatic tissue, we have now evaluated its capacity to correct hypergly- cemia under tolerable immune suppression in an NHP model for diabetes. Results Defining a Tolerable Immune Suppression Protocol. Diabetes was induced in 4 cynomologous monkeys by the administration of Author contributions: G.H., S.E.-F., C.R., E.F., B.J.H., and Y.R. designed research; G.H., S.E.-F., C.R., E.S., D.T., A.A., E.F., H.G., R.E.-H., H.K., A.Z., Z.K.-O., P.S.-M., P.I., A.T., and A.H. performed research; G.H., S.E.-F., C.R., E.S., D.T., A.A., A.Z., Z.K.-O., P.S.-M., A.S., A.T., and Y.R. analyzed data; and G.H., S.E.-F., A.Y., E.B.-L., and Y.R. wrote the paper. Conflict of interest: Y.R. is a scientific consultant and holds equity with Tissera, Inc., which supported this work. This article is a PNAS Direct Submission. 1 G.H. and S.E.-F. contributed equally to this work. 2 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0812253106/DCSupplemental. www.pnas.orgcgidoi10.1073pnas.0812253106 PNAS May 26, 2009 vol. 106 no. 21 8659 – 8664 MEDICAL SCIENCES

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Embryonic pig pancreatic tissue for the treatmentof diabetes in a nonhuman primate modelGil Hechta,1, Smadar Eventov-Friedmana,1, Chava Rosena, Elias Shezena, Dalit Tchorsha, Anna Aronovicha,Enrique Freudb, Hana Golana, Ronit El-Hasidc, Helena Katchmana, Bernhard J. Heringd, Amnon Zunge,Zipi Kra-Ozf, Pninit Shaked-Mishanf, Alex Yusimg, Alex Shtabskyh, Pavel Idelevitcha, Ana Tobari,Alon Harmelinj, Esther Bachar-Lustiga, and Yair Reisnera,2
aDepartment of Immunology, Weizmann Institute of Science, Rehovot 76100, Israel; bDepartment of Pediatric Surgery, Schneider Children’s Medical Centerof Israel, Petach Tikvah 49202, Israel; cDepartment of Pediatric Hemato-Oncology, Meyer Children Hospital, Haifa 31096, Israel; dDiabetes Institute forImmunology Transplantation, University of Minnesota, Minneapolis, MN, 55455; eDepartment of Pediatrics, Kaplan Medical Center, Rehovot 76100, Israel;fDepartment of Virology, Rambam Medical Center, Haifa 31096, Israel; gDepartment of Transplantation, Rabin Medical Center, Petah Tikva, Israel;hDepartment of Pathology, Sourasky Medical Center, Tel-Aviv 64239, Israel; iDepartment of Pathology, Rabin Medical Center, Petah Tikva, Israel;and jDepartment of Veterinary Resources, Weizmann Institute of Science, Rehovot 76100, Israel
Edited by George Klein, Karolinska Institutet, Stockholm, Sweden, and approved April 3, 2009 (received for review December 10, 2008)
Xenotransplantation of pig tissues has great potential to overcomethe shortage of organ donors. One approach to address thevigorous immune rejection associated with xenotransplants is theuse of embryonic precursor tissue, which induces and utilizes hostvasculature upon its growth and development. Recently, weshowed in mice that embryonic pig pancreatic tissue from embry-onic day 42 (E42) exhibits optimal properties as a � cell replacementtherapy. We now demonstrate the proof of concept in 2 diabeticCynomolgus monkeys, followed for 393 and 280 days, respectively.A marked reduction of exogenous insulin requirement was notedby the fourth month after transplantation, reaching completeindependence from exogenous insulin during the fifth month aftertransplantation, with full physiological control of blood glucoselevels. The porcine origin of insulin was documented by a radio-immunoassay specific for porcine C-peptide. Furthermore, thegrowing tissue was found to be predominantly vascularized withhost blood vessels, thereby evading hyperacute or acute rejection,which could potentially be mediated by preexisting anti-pig anti-bodies. Durable graft protection was achieved, and most of the latecomplications could be attributed to the immunosuppressive pro-tocol. While fine tuning of immune suppression, tissue dose, andimplantation techniques are still required, our results demonstratethat porcine E-42 embryonic pancreatic tissue can normalize bloodglucose levels in primates. Its long-term proliferative capacity, itsrevascularization by host endothelium, and its reduced immuno-genicity, strongly suggest that this approach could offer an attrac-tive replacement therapy for diabetes.
immune-suppression � rejection � xeno-transplantation
A llogeneic islet cell replacement can restore normoglycemiain patients with type 1 diabetes; yet, the shortage of
available organs for transplantation continues to severely limitthis therapeutic option. A potential solution for this shortage isthe use of pig organs and tissues as a source for transplantation.However, the immunological barriers to xenotransplantation areformidable (1). Thus, new approaches that could further reducethe intensity of the required immune suppression are critical forsuccessful application of pig pancreatic tissue as a novel sourceof islet cells. One means to address this challenge could beprovided by the use of embryonic porcine tissue. This strategy isbased on the growing evidence, over the past 5 decades, dem-onstrating maternal immune tolerance to the fetus (2, 3), and onrecent observations that embryonic tissues exhibit reduced im-munogenicity in various transplantation settings (4–7). Consid-ering that very early embryonic tissues are associated with asubstantial risk of teratoma formation, we have attempted,during the past several years, to define the earliest gestationaltime point that does not pose a teratoma risk for transplantationof different embryonic pig tissues, including kidney (6), heart
(unpublished), spleen (8), pancreas, liver, and lung (7) into SCIDmice. Once this threshold was established, we were able tofurther characterize an optimal gestational window for trans-plantation based on growth potential and immunogenicity. Thus,while the optimal window for kidney transplantation was definedaround E28, our data suggested E42 as the optimal gestationaltime point for pig donor tissue in pancreatic transplantation (9).Importantly, upon growth and development of the embryonicpancreatic implant, acini gradually disappear and the tissue ispredominantly comprised of islets and stroma identified byvimentin straining. Thus, the risk of potential ongoing destruc-tion of the growing organ by local proteolytic enzymes, whichmight be released by the exocrine components of the implant, isminiml (9). These mouse studies also demonstrated the curativepotential of such implants in diabetic mice under relativelytolerable immune suppression. However, considering that xeno-geneic transplantation into primates is far more complex, due topreexisting anti-porcine antibodies, studies in a nonhuman pri-mate (NHP) model for diabetes were required. Our workinghypothesis, suggesting that embryonic tissue transplantationwould not be adversely affected by such a humoral response wasbased on 2 major observations: (i) hyperacute and acute rejec-tion are mainly mediated by complement activation in bloodvessels, mediated by preformed anti-pig antibodies recognizing�-gal and other carbohydrates expressed on endothelial cells(10), and (ii) data from our mouse studies showing that theembryonic implants predominantly induce host-type vasculatureto support their growth and development in the recipient,resulting in an organ comprised mainly of porcine epithelial cellsand host endothelium.
To further assess the curative potential of E42 pig pancreatictissue, we have now evaluated its capacity to correct hypergly-cemia under tolerable immune suppression in an NHP model fordiabetes.
ResultsDefining a Tolerable Immune Suppression Protocol. Diabetes wasinduced in 4 cynomologous monkeys by the administration of
Author contributions: G.H., S.E.-F., C.R., E.F., B.J.H., and Y.R. designed research; G.H., S.E.-F.,C.R., E.S., D.T., A.A., E.F., H.G., R.E.-H., H.K., A.Z., Z.K.-O., P.S.-M., P.I., A.T., and A.H.performed research; G.H., S.E.-F., C.R., E.S., D.T., A.A., A.Z., Z.K.-O., P.S.-M., A.S., A.T., andY.R. analyzed data; and G.H., S.E.-F., A.Y., E.B.-L., and Y.R. wrote the paper.
Conflict of interest: Y.R. is a scientific consultant and holds equity with Tissera, Inc., whichsupported this work.
This article is a PNAS Direct Submission.
1G.H. and S.E.-F. contributed equally to this work.
2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0812253106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0812253106 PNAS � May 26, 2009 � vol. 106 � no. 21 � 8659–8664
MED
ICA
LSC
IEN
CES
150 mg/kg of streptozotocin (STZ). About 1 month after STZadministration, E42 porcine tissue was implanted in the omen-tum as described in Methods. Immune suppression was inducedand maintained in all 4 animals using a protocol based oninduction with a single dose of anti-CD20 antibody (Rituximab)followed by a short course of ATG and 2 doses of anti-CD25antibody (Basiliximab). Immune suppression was maintained byEverolimus, FTY720, and biweekly treatment with CTLA4-Ig(Abatacept) (Fig. 1). All these immunosuppressive agents werepreviously shown to be effective and well-tolerated in allogeneicislet transplantation in the NHP model (11–14).
The study consisted of 2 cohorts, with 2 animals in each. In thefirst cohort, central line-associated infections eventually led tothe death of both NHP #1 and #2, 2 and 3 months followingtransplantation, respectively. Apart from the adverse effectsassociated with the central line catheters, the ATG dose (totalof 70 mg/kg) used in monkey #1 was associated with severeanemia and coagulation disorders, requiring extensive bloodtransfusions. The choice of this relatively high ATG dose wasbased on previous suggestions that rabbit anti-human ATG isless active in NHP than in humans and that equivalent doses inNHP are 6 times higher than in humans (12). However, sub-stantial reduction of ATG dose in monkey #2 to 40 mg/kg waswell-tolerated and did not affect engraftment or acceptance ofthe graft, as evidenced by histology (Fig. 2). In contrast to NHP#1 and #2, which were followed for a relatively short time, whichwas not sufficient to allow a significant progression towardinsulin independence, the changes made in the treatment of theanimals in cohort 2 (i.e., avoiding use of central line and usinga lower ATG dose) enabled these animals to survive for 393(NHP #3) and 280 days (NHP #4) posttransplantation.
Similarly to NHP #2, NHP #3 was transplanted using a totaldose of 40 mg/kg ATG. This animal exhibited CMV reactivation,which required continuous treatment (see Materials and Meth-ods), and therefore in animal #4, the immune suppressionprotocol was further reduced to comprise a total dose of 20mg/kg ATG. Postmortem immunohistological analysis of mes-enteric lymph nodes and spleen of this animal revealed pre-served T and B cell follicles (SI), while as can be expected fromthe use of FTY720 in the protocol, low lymphocyte blood levels(average 0.68 k/�L � 0.37) were recorded throughout thefollow-up period (SI).
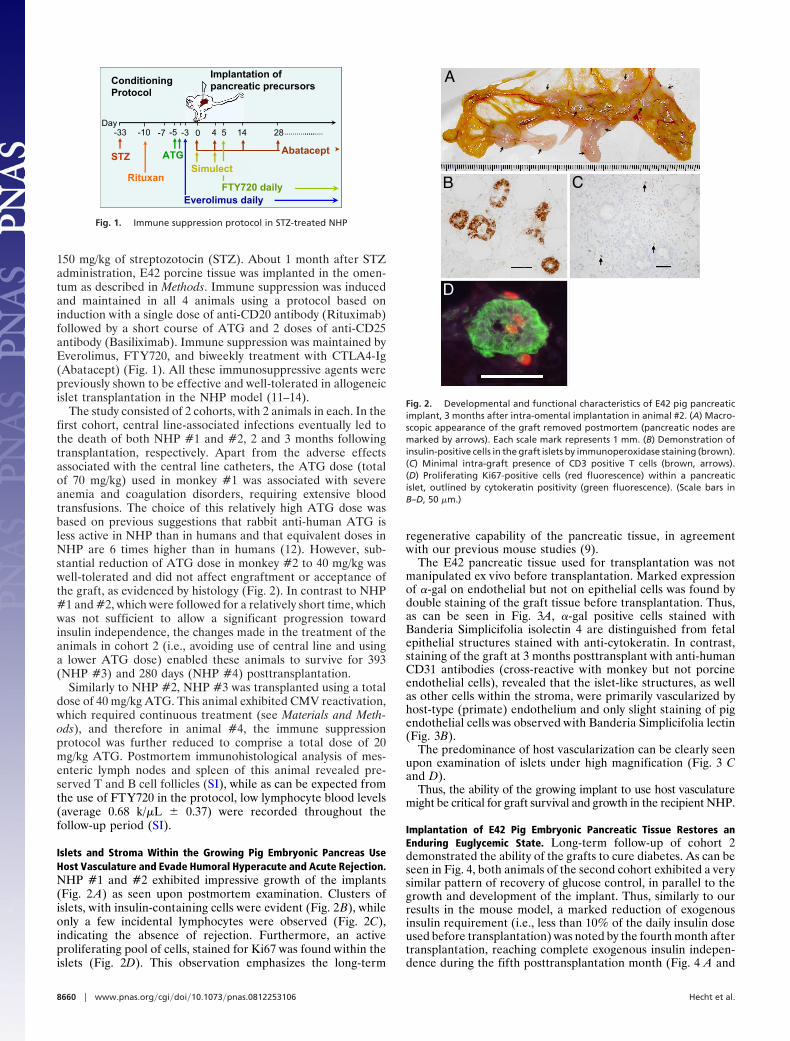
Islets and Stroma Within the Growing Pig Embryonic Pancreas UseHost Vasculature and Evade Humoral Hyperacute and Acute Rejection.NHP #1 and #2 exhibited impressive growth of the implants(Fig. 2 A) as seen upon postmortem examination. Clusters ofislets, with insulin-containing cells were evident (Fig. 2B), whileonly a few incidental lymphocytes were observed (Fig. 2C),indicating the absence of rejection. Furthermore, an activeproliferating pool of cells, stained for Ki67 was found within theislets (Fig. 2D). This observation emphasizes the long-term
regenerative capability of the pancreatic tissue, in agreementwith our previous mouse studies (9).
The E42 pancreatic tissue used for transplantation was notmanipulated ex vivo before transplantation. Marked expressionof �-gal on endothelial but not on epithelial cells was found bydouble staining of the graft tissue before transplantation. Thus,as can be seen in Fig. 3A, �-gal positive cells stained withBanderia Simplicifolia isolectin 4 are distinguished from fetalepithelial structures stained with anti-cytokeratin. In contrast,staining of the graft at 3 months posttransplant with anti-humanCD31 antibodies (cross-reactive with monkey but not porcineendothelial cells), revealed that the islet-like structures, as wellas other cells within the stroma, were primarily vascularized byhost-type (primate) endothelium and only slight staining of pigendothelial cells was observed with Banderia Simplicifolia lectin(Fig. 3B).
The predominance of host vascularization can be clearly seenupon examination of islets under high magnification (Fig. 3 Cand D).
Thus, the ability of the growing implant to use host vasculaturemight be critical for graft survival and growth in the recipient NHP.
Implantation of E42 Pig Embryonic Pancreatic Tissue Restores anEnduring Euglycemic State. Long-term follow-up of cohort 2demonstrated the ability of the grafts to cure diabetes. As can beseen in Fig. 4, both animals of the second cohort exhibited a verysimilar pattern of recovery of glucose control, in parallel to thegrowth and development of the implant. Thus, similarly to ourresults in the mouse model, a marked reduction of exogenousinsulin requirement (i.e., less than 10% of the daily insulin doseused before transplantation) was noted by the fourth month aftertransplantation, reaching complete exogenous insulin indepen-dence during the fifth posttransplantation month (Fig. 4 A and
28 ……………..0 4 5 14-33 -4 -3 -7
Conditioning Protocol
Implantation of pancreatic precursors
28 ……………………0 4 5 14-33Day
Everolimus daily
STZ ATG Abatacept
-3
FTY720 daily
-7-10
Rituxan
-5
Simulect
28 ……………..0 4 5 14-33 -4 -3 -7
Conditioning Protocol
Implantation of pancreatic precursors
28 ……………………0 4 5 14-33Day
Everolimus daily
STZ ATG Abatacept
-3
FTY720 daily
-7-10
Rituxan
-5
Simulect
Fig. 1. Immune suppression protocol in STZ-treated NHP
A
B C
D E
A
B C
D E
A
B C
D
E
Fig. 2. Developmental and functional characteristics of E42 pig pancreaticimplant, 3 months after intra-omental implantation in animal #2. (A) Macro-scopic appearance of the graft removed postmortem (pancreatic nodes aremarked by arrows). Each scale mark represents 1 mm. (B) Demonstration ofinsulin-positive cells in the graft islets by immunoperoxidase staining (brown).(C) Minimal intra-graft presence of CD3 positive T cells (brown, arrows).(D) Proliferating Ki67-positive cells (red fluorescence) within a pancreaticislet, outlined by cytokeratin positivity (green fluorescence). (Scale bars inB–D, 50 �m.)
8660 � www.pnas.org�cgi�doi�10.1073�pnas.0812253106 Hecht et al.

B), with marked physiological control of blood glucose levels(Fig. 4 A and B). A radioimmunoassay (RIA)-specific forporcine C-peptide (i.e., non-cross-reactive with primate C-peptide) confirmed the presence of basal porcine C-peptidelevels as early as 3 months following transplantation (0.24 and0.35 ng/mL for monkeys #3 and #4, respectively), and markedelevation of porcine C-peptide blood levels was demonstrated inboth animals after i.v. glucose challenge test (IVGTT), inaccordance with the enhanced insulin levels (Fig. 4 C and D).
A RIA for human C-peptide (cross-reactive with primate)revealed some baseline level of primate C-peptide. However, nosignificant elevation of primate C-peptide could be detected bythis assay following glucose challenge, in contrast to the eleva-tion of porcine C-peptide (SI). This constant and unresponsivebase line level could be due to some cross-reactivity withundefined blood components or to some level of regeneration ofundifferentiated or nonfunctional insulin-positive beta cells inthe recipient. However, the absence of insulin-positive beta cellswithin the host pancreas upon postmortem (Fig. 5A) stronglysuggests that insulin independence, attained in both animalsupon growth and development of the implant, is most likely notassociated with the potential recovery of host pancreatic activity.The animals exhibited an Hb A1C of 6.1% and 5.2% at 333 and195 days following transplant, respectively.
Correction of hyperglycemia was adversely affected twice inmonkey #3. Two weeks after reaching insulin independence, lowEverolimus levels were detected and partial rejection was sus-pected as insulin requirement relapsed to an average of 15% ofmaximal levels during week 33 (Fig. 4A and SI). However, agradual restoration of graft function, to a minimal requirementof insulin supplementation, less than 3.7% of the insulin require-ment before transplantation, was observed (Fig. 4A). Eventually,clinical complications likely associated with the continuousCMV infection and the combined toxicity of anti-viral drugs and
the immune suppression, necessitated euthanizing of the animalat 393 days posttransplant (SI).
A postmortem immunohistochemical analysis of the graft re-vealed intact insulin-positive islet fields (Fig. 5B and C), and onlyrare CD3� lymphocytes could be recorded in the proximity oftransplanted islets (Fig. 5B) ruling out potential immune rejection.
In NHP #4 insulin independence was sustained for more than17 weeks (average fasting glucose during this period was 113 �20) without any complications. At the beginning of the eighthmonth posttransplant, technical failure to sustain Everolimuslevels above the threshold of 3 ng/mL (SI) seemed initially to beassociated with partial loss of graft function (Fig. 4B). A glucosechallenge test performed 2 weeks earlier revealed robust pigpancreatic activity (maximal C-peptide of 3.79 ng/mL), suggest-ing that the acute reduction in function might indeed be attrib-uted to the parallel drop of Everolimus blood levels. Thisdecrease in functionality was accompanied by clinical deterio-ration, which was manifested by anorexia, and led to euthanizingof the animal at 280 days posttransplantation (SI). Postmortemexamination revealed essentially similar findings to those foundin animal #3 (see SI). Thus, the presence of intact islets fieldswith no evidence of rejection indicates that an infectious processof unknown etiology could be responsible for the partial loss offunction in both animals.
Fig. 3. Vascularization pattern of E42 pancreatic tissue. (A) Marked expres-sion of �-gal on endothelial but not on epithelial cells in the graft tissue beforetransplantation, indicated by double staining. Thus, �-gal-positive cells arestained with Banderia Simplicifolia isolectin 4 (green fluorescence), and fetalepithelial structures are stained with anti-cytokeratin (blue). (B) Predominantvascularization of the porcine graft by host blood vessels is demonstrated bystaining with anti-human CD31 antibody that cross-reacts with monkey butnot with pig endothelial cells (red fluorescence). Only slight staining of pigendothelial cells was observed with Banderia Simplicifolia lectin (green). Isletepithelium is outlined by cytokeratin expression (blue). (C) Higher magnifica-tion of anti-cytokeratin-positive islets (blue) observed within the markedframe in inset B. (D) Higher magnification of the vasculature network sup-porting the islets observed within the marked frame in inset B. Double stainingwith anti-human CD31 (red) and Banderia Simplicifolia lectin (green) revealspredominance of endothelial cells of monkey origin. Nuclei were counter-stained by Hoechst Yellow (yellowish). (Scale bars, 50 �m.)
Fig. 4. Assessment of E42 fetal porcine xenograft function in streptozotocin-induced diabetic Cynomolgus monkeys of cohort 2. Insulin requirement (blue)and fasting glucose levels (yellow) in monkeys #3 and #4 are shown in panelsA and B, respectively. Specific pig C-peptide (red) and insulin (green) levelsafter IVGTT in monkey #3 (C) and #4 (D) were determined by ELISA specific forpig C-peptide (non-cross-reactive with monkey C-peptide) and for total insulin(using an antibody that does not distinguish between the 2 species), at 393days and 280 days posttransplant, respectively. K values for the clearance ofglucose were 1.84 and 2.0, respectively.
Hecht et al. PNAS � May 26, 2009 � vol. 106 � no. 21 � 8661
MED
ICA
LSC
IEN
CES

DiscussionInitial clinical attempts of fetal porcine pancreas transplantationwere carried out early in the 1990s by Groth et al. (15). In thesestudies, patients were transplanted with E66-E81 porcine fetalislet-like cell clusters. Blood glucose levels increased shortlyafter transplantation and insulin administration had to be in-creased, so that eventually it did not differ from the pretrans-plant requirements. We have shown recently in a mouse modelthat this outcome could be attributed, in part, to the relativelylate stage of gestation at which the embryonic pancreatic tissuewas harvested. Thus, based on growth potential and immuno-genicity, we demonstrated that the optimal ‘‘window’’ for thesetransplants could be provided at around E42 (7, 9).
In our present study, the almost identical pattern of insulinreduction exhibited in both animals of cohort 2 (Fig. 4 A and B)demonstrates the reproducibility of the time frame of growth anddifferentiation required to achieve normoglycemia, and providesproof of concept for the curative potential of the E42 pigpancreatic implants in NHP.
Furthermore, the growing tissue makes use of host vascula-ture, and thereby evades hyperacute or acute rejection, whichcould potentially be mediated by preexisting anti-pig antibodies(10). This finding supports our working hypothesis that growingthe embryonic pig tissue de novo within the NHP recipient,provides an advantageous source for organ transplantationcompared to adult pig organ transplantation, which requiresanastomosis of donor blood vessels. Further NHP studies withother precursor tissues such as embryonic kidney (6), liver (7),and spleen (8), for which appropriate embryonic sources havebeen recently characterized, are warranted.
Very recently, the capacity of neonatal and adult porcine isletsto restore normoglycemia in diabetic NHP was demonstrated(16, 17). However, the immune suppression required to maintainthe implants was either too toxic or included anti-CD40L, whichcannot be used in human patients due to its thrombotic prop-erties. Other studies suggested that E28 pancreatic tissue mightbe accepted in nonimmunosuppressed rats (33) and monkeys(34). In contrast, our mouse (9) and rat (35) data has revealedfierce rejection of pig embryonic tissue of any gestational timepoint in the absence of immune suppression, while demonstrat-ing that the immunogenicity of E42 tissue was relatively lower
compared to tissues harvested at E56 or beyond (9). Therefore,we proposed that mild immune suppression might suffice topermit engraftment and growth of the E42 embryonic tissue.
In the present study, we chose a relatively strong immunesuppression, so as to avoid hurdles associated with tissue rejec-tion. This protocol is likely less nephrotoxic than current allo-geneic islet transplantation protocols, such as that recentlydescribed by Bellin et al. (18) comprising induction with ATGand maintenance with Everolimus and Cyclosporine A. How-ever, although the present immune suppression protocol used inanimal #4 was well tolerated, further assessment is required todefine the minimal immune suppression that can support graftsurvival while avoiding toxicity and infections. Preliminary re-sults in the mouse model, evaluating potential substitutes forEvorolimus with co-stimulatory blockade agents other thananti-CD40L have shown encouraging results (35). Such substi-tutes could be of particular importance considering the skinlesions found in animal #3 (see SI for details) and other reportedadverse effects of Rapamycin on islet function (19–21).
Beta cell replacement can be achieved by either whole organ-pancreas allogeneic transplantation or transplantation of iso-lated islet cells. The transplantation of a whole pancreas achievesa longer graft survival and functionality than isolated islets (67%after 10 years and 10% after 5 years, respectively) (22, 23). Thisdiscrepancy could be attributed to the deleterious effects en-countered during islet preparation by enzyme digestion, pro-longed ischemia time, and the loss of surrounding mesenchyme.The active involvement of the surrounding mesenchyme inpancreatic development and its role in endocrine cell differen-tiation and proliferation has been documented (24, 25). Thus,the embryonic pancreatic tissue growing in the context of its ownstroma as documented here, might survive longer than infusedneonate or adult islets growing heterotopically in the liver.Indeed, following graft injury and partial loss of function inmonkey #3, the growing pig implant exhibited a marked capacityto withstand stress and regenerate. This characteristic of thegraft, which could be related to mesenchymal support, is ofparticular relevance considering that although 1-year successrates for islet transplantation are high, the long-term success rateis unacceptably low, even in the pioneering Edmonton Center(23). The poor long-term clinical outcome, taken together withthe recent study of Koulmanda et al. (26), indicates that one ofthe reasons for long-term failure may be due to the low islet cellmass that actually survives the harvesting and transplant proce-dures, as well as islet attrition by exhaustion. The potentialimportance of islet replacement with E42 graft that will expandwith time could afford a major advantage.
Recent rodent studies on islet transplantation suggest that itmight be possible to use s.c. sites for implantation (27). Such anoninvasive mode of transplantation could enable continuousmonitoring of the implant growth and will avoid the potentialrisk of developing sclerosing encapsulating peritonitis that mightarise (as suspected in animal #4) upon infection or graftrejection at the omental site.
Finally, it should be noted that the embryonic precursor tissueused in our study differs from islet transplantation in its dose/response relationship. While islet transplantation exhibits lineardose dependency, and the islet dose must be adjusted to the bodyweight of the recipient, the embryonic precursor tissue, whichexhibits remarkable growth potential, can adopt itself to the sizeof the recipient through a poorly defined complex mechanism oforgan size control (28, 29). Thus, the same dose of embryonictissue could potentially give rise to final organs of different size,depending on the species or weight of the recipient and itsphysiological demands. In the present study, we chose to test ourimplants under conditions in which tissue dose was not limiting.Having demonstrated the proof of concept, further studies will
Fig. 5. Immunohistochemical characterization of islets in the E42 pig pan-creatic implant versus the host pancreas, at 13 months after intra-omentalimplantation in animal #3. (A) Immunoperoxidase labeling of insulin (brownstaining) reveals minimal residual presence of insulin-positive cells within thehost pancreas (each bar represents 50 �m). Insulin-containing islets within thegraft (B and C) were labeled by immunofluorescence labeling of insulin (bluefluorescence). Few incidental CD3-positive T cells (green fluorescence) can bedetected outside the islets (blue) (B).
8662 � www.pnas.org�cgi�doi�10.1073�pnas.0812253106 Hecht et al.

be performed to define the minimal tissue dose for attainmentof insulin independence.
In conclusion, although further fine tuning of immune sup-pression, tissue dose, and implantation techniques are war-ranted, our results demonstrate that porcine E-42 embryonicpancreatic tissue can correct hyperglycemia under a tolerableimmune suppression protocol. The long-term proliferative ca-pacity of these grafts, their ability to induce revascularization byhost endothelium, and their reduced immunogenicity, stronglysuggest that porcine embryonic xenotransplantation could offeran attractive replacement therapy for diabetes.
MethodsAnimals. Juvenile Macaca fascicularis (cynomolgus) male monkeys (2–3 yearsof age, 2–3.5 kg) bred at a local Israeli farm (Moshav Mazor) from a colonyderived from Mauritius were used as transplant recipients. The animals weretested annually for all known pathogenic viruses and for tuberculosis andwere treated with anti-helmintics. All of the procedures were monitored bythe veterinarian of the Veterinary Resources Unit of the Weizmann Instituteand approved by the Institutional Animal Care and Use Committee (IACUC).Monkeys were maintained in pairs. The cages (Lab. Products) were designedwith free moving rooms attached to them. All cages and rooms were equippedwith enrichment accessories. Cages were cleaned daily and sterilized weekly.Fresh fruits and vegetables were washed with soap and water before feeding.
Immunosuppressive Protocol. The immune suppression protocol consisted ofinduction therapy with Rituxan (Rituximab, 20 mg/kg; La Roche LTD ) on day�10 before transplant, and ATG (Thymoglobulin, rabbit anti-human thymo-cyte globulin; 10–20 mg/kg; Sangstat) on days �4 and �3, before transplan-tation, as described for animals #2, #3, and #4. Animal #1 received a total of70 mg over days �5, �4, and �3. Induction was completed with 2 doses ofSimulect (10 mg; Novartis Pharma Stein AG) on at the day of the transplant andon day 4 posttransplant [postoperative day (POD) 4]. Maintenance was con-tinued with human CTLA4-Ig (Abetacept; Bristol-Myers, 20 mg/kg on POD 0and 4, and repeated every 14 days), Everolimus [0.075–0.15 mg/kg dailyadministered s.c.; the dose was adjusted according to trough blood levels (4–8ng/mL)], and FTY720 (0.1 mg/kg daily PO, starting 5 days after transplanta-tion). All intravenous (i.v.) drug administrations were performed under anes-thesia. Everolimus and FTY720 were supplied by Novartis Pharma Stein AGunder MTA.
Each ATG administration was preceded by prophylactic hydrocortisone(5–10 mg/kg i.v.), Promethazine [0.5–1 mg/kg intramuscular (i.m.)], and parac-etamol (15–20 mg/kg rectally). On the first 2 days, ATG (10–20 mg/kg), dilutedin 50 mL Ringer’s lactate or saline 0.9%, was injected i.v. at a rate of 10 mL/kg/htwice daily.
Animals were monitored closely for possible side effects, including rash,respiratory symptoms, and body temperature elevation during and followingtreatment with immunosuppressive agents. If clinical signs suggesting pul-monary edema or allergic reaction developed, then treatment with diuretics(furosemide 1 mg/kg i.v. or i.m.) and antihistamines was added.
Prophylactic and Preemptive Treatment. CMV infection has a significant neg-ative impact on graft function and recipient survival (30). Prophylactic Gan-ciclovir (Cytovene 2.5 mg/kg i.m. in a divided dose, 1.0 mg/kg AM, 1.5 mg/kgPM) treatment was therefore initiated 2 days before ATG administration.RT-PCR monitoring of rhCMV replication was performed twice a week at thevirology laboratory of the Rambam Hospital, Haifa, Israel. When rhCMVreplication was observed (�3,500 copies/mL), the Ganciclovir dose was in-creased to 10 mg/kg i.m. and, in cases where the development of viraltolerance was suspected, Cidofovir treatment (5 mg/kg once per week) wasadded until rhCMV levels became undetectable. Bacterial infection was de-fined as positive blood and/or wound culture in combination with leukocytosisor leukopenia, abnormal body temperature, and/or general clinical deterio-ration. For suspected infection, blood cultures (and, if infected, wound cul-ture) were obtained, and then empiric antibiotic therapy was initiated usingenrofloxacin (5 mg/kg i.m. 2dd2 daily doses) and stopped when culturesremained negative after 48 h of incubation and the clinical condition of theanimal improved. For each confirmed infection, appropriate antimicrobialtherapy was chosen based on the in vitro susceptibilities.
Surgery. The animals were fasted 8 h before the surgery. Prophylactic antibi-otic and anti-inflammatory treatments were administered.
Anesthesia. After the i.m. administration of a combination of midzolam andketamine as a premedication, an i.v. line was inserted, and the monkey wasintubated using an orotracheal tube. Thereafter, ventilation with a gas mix-ture containing Isofluran was carried out for anesthesia maintenance. Tem-perature was maintained within the normal range using a heating mattress.Heart rate, temperature, blood oxygenation, and blood pressure were mon-itored during the surgical procedure.Surgical procedures. The surgical procedure included transplantation of E42fetal porcine pancreas, isolated shortly before the surgery, in the omentalfolds. Pig embryos were routinely obtained from the Lahav Institute of AnimalResearch (Kibbutz Lahav, Israel). Pregnant sows were operated on at precisestages of their pregnancy under general anesthesia, and embryos were ex-tracted. Warm ischemia time was less than 10 min, and the embryos weretransferred in cold PBS. Pig pancreas precursors for transplantation wereextracted under a light microscope and kept under sterile conditions at 4 °C inRPMI 1640 (Biological Industries) before transplantation without any attemptto remove exocrine elements and without any other ex vivo manipulation.Cold ischemia time until transplantation was routinely less than 2 h.
In all operations, an anterior abdominal approach was used. Followingexposure of the omentum, an omental pocket was created by circular suturelegation followed by embryonic tissue implantation into the pocket. In eachtransplantation, approximately 6 omental pockets were created, and in eachpocket, 10 E42 pig pancreatic fragments were implanted. The abdominal wallwas closed in layers using conventional technique.Postoperative management. For the first 12 h following operation, no oralfeeding was permitted. Afterward, the monkey was allowed to drink and eatfreely, if clinically feasible. Antibiotic treatment was continued for 3 days,according to the surgeon’s orders. All post-operative wound treatments andblood tests were performed after the administration of ketamine and mida-zolam. Pain relief in the immediate surgical period and afterward was pro-vided by Buprenex i.m. injections.
During the first week after the transplantation surgery, blood tests forcomplete blood cell count, kidney and liver functions, electrolytes, cholesteroland triglycerides, porcine insulin, Ig and complement levels, and immunosup-pressive drug levels were taken at a frequently based on the clinical condition,followed by 1–2 blood tests weekly, unless clinical signs of infection, fever,acute rejection, or other complications appeared. The maximal blood volumedrawn over any 2-week period was not permitted to exceed 1% of themonkey’s body weight.
ELISA Measurements of Insulin. The porcine/human insulin kit (K6219; Dako)was used to follow insulin levels according to the manufacturer’s instructions.NHP serum samples were loaded in a blinded fashion into the wells.
Determination of Hemoglobin A1C. Total Hb and HbA1c concentrations weredetermined using COBAS INTEGRA system, according to the manufacturer’sinstructions. Briefly, total Hb was measured colorimetrically. HbA1c was de-termined by an immunoturbidimetric assay. The ratio of both concentrationsyielded the final percent HbA1c result (HbA1c %).
Everolimus (RAD) Blood Levels. Blood samples were analyzed routinely onceweekly at the Sheba Medical Center in Tel-Aviv, by Innofluor Certican fluo-rescence polarization immunoassay (Seradyn). Results were obtained within2–3 days. Targeted trough RAD levels were between 4 and 8 ng/mL.
Porcine C-Peptide Levels. RIA for pig C-peptide was performed using PorcineC-Peptide RIA kit (catalog no. PCP-22K; Linco Research) according to themanufacturer’s instructions.
Primate C-Peptide Levels. RIA for human C-peptide was performed usingHuman C-Peptide RIA Kit (catalog no. HCP-20K; Linco Research) according tothe manufacturer’s instructions.
Diabetes Induction and Management. STZ protocol for the induction of diabetes inNHP. 1. STZ (Zanosar, Amersham Pharmacia and Upjohn) was given afterovernight fasting and antiemetic treatment (Zofran). The animal was sedatedwith 10 mg/kg Ketamine i.m. and 0.04 mg/kg atropine i.m. Diabetes wasinduced using a single high dose i.v. STZ injection (150 mg/kg) (31). Highfasting blood glucose levels were observed in all monkeys within 24 h follow-ing STZ injection. Implantation of the embryonic tissues was conducted atleast 30 days after diabetes induction.Diabetes management. Daily diabetes management included the administrationof Lantus (Sanofi-Aventis U.S. LLC), once or twice daily, and Lispro (Humalog; EliLilly)as required.Themonkeyswerefed3lowcarbohydratemealsaday.Uneaten
Hecht et al. PNAS � May 26, 2009 � vol. 106 � no. 21 � 8663
MED
ICA
LSC
IEN
CES

food was removed after 45 min. Blood for glucose measurement was taken fromthe finger tip just before meals, and then Lispro was administered according tothefollowingscale:Glucose levelsof300–400mg/dL,1unit;400–500,2units;and500–600, 3 units. Insulin doses were reevaluated and adjusted for each monkeyaccording to blood glucose measurements 2 h after Lispro administration. Lantusdose was adjusted according to fasting morning blood glucose levels, 12 or 24 hpost-Lantus administration. Close monitoring of blood glucose was performed atnight to avoid unexpected hyperglycemia and to minimize the risk of hypogly-cemia or ketoacidosis.Glucose Tolerance Test. The test was taken after an 18-h fast and insulintreatment withdrawal for 26 h (Lantus) and 6 h (Lispro). Each animal wassedated with ketamine hydrochloride (10 mg/kg i.m.) before placement ofbilateral saphenous vein catheters for the infusion of glucose and bloodsampling, respectively. The glucose tolerance test was performed by injectinga solution of 12.5% dextrose (0.5 g/kg) over a 30-s period at t � 0 anddetermining the subsequent blood glucose concentrations with an Elite XLglucometer (Bayer) at 0, 1, 3, 5, 10, 15, 20, 30, 60, and 90 min post-injection.Blood for insulin and porcine C-peptide determination was collected at thesame time. The glucose disappearance rate (KG) was calculated from theportion of the curve occurring between the 5 and 30 min time points. The Kvalue represents the rate of reduction in plasma blood glucose as a percentageper minute, and was calculated using the following formula (32): KG �[LN(glucose level at 5 min) � LN(glucose level at 30 min)/25] � 100.
Histology and Immunohistochemistry. Sections were routinely stained by he-matoxylin and eosin. Histochemical and immunohistochemical labeling wasperformed as previously described (9) with the addition of mouse anti-human
CD31 (clone JC/70A) for the demonstration of NHP’s endothelium. For immu-noperoxidase labeling, the following second antibodies and reagents wereused: Dako peroxidase envision system for detection of mouse and rabbitantibodies, and Sigma biotinylated anti-goat antibody (followed by extra-avidin peroxidase reagent) for detection of goat antibodies. Diaminobenzi-dine was used as the chromogen for peroxidase labeling. Tissue sections werecounterstained with hematoxylin and embedded in Entellan.
For single and multiple immunofluorescence labeling, the following sec-ondary antibodies were used: Donkey anti-mouse and donkey anti-rabbitantibodies, conjugated with CY2 or TxR (Jackson); goat anti-rabbit antibody,conjugated with Alexa Fluor 350 (Invitrogen); and donkey anti-guinea pigantibody, conjugated with biotin (Jackson). Griffonia Simplicifolia, isolectin 4,conjugated with FITC (L-2895Sigma), was used for demonstration of �-galactose expressing blood vessels of pig origin. Rabbit anti-human IgG,conjugated with FITC (Dako), was used to evaluate immunoglobin deposition.Nuclei were counterstained with the fluorescent nuclear dyes, Hoechst 33342and Hoechst Yellow (Molecular Probes). The histological sections were em-bedded in Cytomation Fluorescent Mounting Medium (Dako).
ACKNOWLEDGMENTS. Professor Yair Reisner holds the Henry H. Drake Pro-fessorial Chair in Immunology. The authors wish to thank Prof. N. Kenyon,(Miami University), Dr. W. Schuler (Novartis, Switzerland), and Melanie Gra-ham (University of Minnesota) for valuable advice in establishing the NHPmodel. We also thank Prof. David Sutherland (University of Minnesota) andthe Juvenile Diabetes Research Foundation for valuable discussions. This workwas supported in part by Tissera, Inc., Mrs. Erica Drake, D Cure-Diabetes Carein Israel (R.A.), the Seymour Spira Fund, the Russell Berrie Foundation, and theGabriella Rich Center for Transplantation Biology Research.
1. Cooper DK (2003) Clinical xenotransplantion— how close are we? Lancet 362:557–559.
2. Medawar PB (1953) Some immunological and endocrinological problems raised by theevolution of viviparity in vertebrates. Symp Soc Exp Biol 7:320–323.
3. Guleria I, Sayegh MH (2007) Maternal acceptance of the fetus: True human tolerance.J Immunol 178:3345–3351.
4. Foglia RP, Dipreta J, Statter MB, Donahoe PK (1986) Fetal allograft survival inimmunocompetent recipients is age dependent and organ specific. Ann Surg204:402– 410.
5. Dekel B, et al. (1997) Engraftment of human kidney tissue in rat radiation chimera: II.Human fetal kidneys display reduced immunogenicity to adoptively transferred hu-man peripheral blood mononuclear cells and exhibit rapid growth and development.Transplantation 64:1550–1558.
6. Dekel B, et al. (2003) Human and porcine early kidney precursors as a new source fortransplantation. Nat Med 9:53–60.
7. Eventov-Friedman S, et al. (2005) Embryonic pig liver, pancreas, and lung as a source fortransplantation: Optimal organogenesis without teratoma depends on distinct timewindows. Proc Natl Acad Sci USA 102:2928–2933.
8. Aronovich A, et al. (2006) Correction of hemophilia as a proof of concept for treatmentof monogenic diseases by fetal spleen transplantation. Proc Natl Acad Sci USA103:19075–19080.
9. Eventov-Friedman S, et al. (2006) Embryonic pig pancreatic tissue transplantation forthe treatment of diabetes. PLoS Med 3:e215.
10. Galili U (2001) The alpha-gal epitope (gal alpha 1–3gal beta 1–4glcnac-r) in xenotrans-plantation. Biochimie 83:557–563.
11. Kirk AD, et al. (1997) Ctla4-ig and anti-cd40 ligand prevent renal allograft rejection inprimates. Proc Natl Acad Sci USA 94:8789–8794.
12. Hirshberg B, et al. (2003) Rabbit antithymocyte globulin induction and sirolimusmonotherapy supports prolonged islet allograft function in a nonhuman primate islettransplantation model. Transplantation 76:55–60.
13. Wijkstrom M, et al. (2004) Islet allograft survival in nonhuman primates immunosup-pressed with basiliximab, rad, and fty720. Transplantation 77:827–835.
14. Liu C, et al. (2007) B lymphocyte-directed immunotherapy promotes long-term isletallograft survival in nonhuman primates. Nat Med 13:1295–1298.
15. Groth CG, et al. (1994) Transplantation of porcine fetal pancreas to diabetic patients.Lancet 344:1402–1404.
16. Cardona K, et al. (2006) Long-term survival of neonatal porcine islets in nonhumanprimates by targeting costimulation pathways. Nat Med 12:304–306.
17. Hering BJ, et al. (2006) Prolonged diabetes reversal after intraportal xenotransplan-tation of wild-type porcine islets in immunosuppressed nonhuman primates. Nat Med12:301–303.
18. Bellin MD, et al. (2008) Prolonged insulin independence after islet allotransplants inrecipients with type 1 diabetes. Am J Transplant 8:2463–2470.
19. Zhang N, et al. (2006) Sirolimus is associated with reduced islet engraftment andimpaired {beta}-cell function. Diabetes 55:2429–2436.
20. Bussiere CT, Lakey JR, Shapiro AM, Korbutt GS (2006) The impact of the mtor inhibitorsirolimus on the proliferation and function of pancreatic islets and ductal cells. Dia-betologia 49:2341–2349.
21. Bloom RD, Crutchlow MF (2008) Transplant-associated hyperglycemia. Transplant Rev22:39–51.
22. Sollinger HW, et al. (1998) Experience with 500 simultaneous pancreas-kidney trans-plants. Ann Surg 228:284–296.
23. Ryan EA, et al. (2005) Five-year follow-up after clinical islet transplantation. Diabetes54:2060–2069.
24. Ye F, Duvillie B, Scharfmann R (2005) Fibroblast growth factors 7 and 10 are expressedin the human embryonic pancreatic mesenchyme and promote the proliferation ofembryonic pancreatic epithelial cells. Diabetologia 48:277–281.
25. Attali M, et al. (2007) Control of {beta}-cell differentiation by the pancreatic mesen-chyme. Diabetes 56:1248–1258.
26. Koulmanda M, et al. (2006) Prolonged survival of allogeneic islets in cynomolgusmonkeys after short-term anti-cd154-based therapy: Nonimmunologic graft failure?Am J Transplant 6:687–696.
27. Andrades P, et al. (2007) Subcutaneous pancreatic islet transplantation using fibringlue as a carrier. Transplant Proc 39:191–192.
28. Conlon I, Raff M (1999) Size control in animal development. Cell 96:235–244.29. Mandel TE (1999) Fetal islet xenotransplantation in rodents and primates. J Mol Med
77:155–160.30. Fishman JA, et al. (2007) Cytomegalovirus in transplantation—challenging the status
quo. Clin Transplant 21:149–158.31. Theriault BR (1999) Induction, maintenance, and reversal of streptozotocin-induced
insulin dependent diabetes mellitus in the juvenile cynomolgus monkey (Macacafascilularis). Transplantation 68:331–337.
32. Casu A, et al. (2008) Metabolic aspects of pig-to-monkey (Macaca fascicularis) islettransplantation: Implications for translation into clinical practice. Diabetologia51:120–129.
33. Rogers SA, Chen F, Talcott M, Hammerman MR (2004) Islet cell engraftment and controlof diabetes in rats after transplantation of pig panceatic anlagen. Am J PhysiolEndocrinol Metab 286:E502–E509.
34. Rogers SA, et al. (2007) Long-term engraftment following transplantation of pigpancreatic primordia into non-immunosuppressed diabetic rhesus macaques. Xeno-transplantation 14:591–602.
35. Tchorsh-Yutsis D, et al. (2009) Pig embryonic pancreatic tissue as a source for trans-plantation in diabetes: Transient treatment with anti-LFA1, anti-CD48 immunosup-pression. Diabetes, In press.
8664 � www.pnas.org�cgi�doi�10.1073�pnas.0812253106 Hecht et al.
Related Documents