ZOOLOGY Zoology 110 (2007) 212–230 Embryonic development of Python sebae – I: Staging criteria and macroscopic skeletal morphogenesis of the head and limbs Julia C. Boughner a,1,2 , Marcela Buchtova´ a,2 , Katherine Fu a , Virginia Diewert b , Benedikt Hallgrı´msson c , Joy M. Richman a, a Department of Oral Health Sciences, Life Sciences Institute, University of British Columbia, Life Sciences Centre, 2350 Health Sciences Mall, Vancouver, BC, Canada V6T 1Z3 b Department of Oral Health Sciences, Faculty of Dentistry, University of British Columbia, 2199 Wesbrook Mall, Vancouver, BC, Canada V6T 1Z3 c Department of Cell Biology and Anatomy, Faculty of Medicine, Heritage Medical Research Centre, 3330 Hospital Drive NW, Calgary, Alta., Canada T2N 4N1 Received 24 August 2006; received in revised form 21 January 2007; accepted 23 January 2007 Abstract This study explores the post-ovipositional craniofacial development of the African Rock Python (Python sebae). We first describe a staging system based on external characteristics and next use whole-mount skeletal staining supplemented with Computed tomography (CT) scanning to examine skeletal development. Our results show that python embryos are in early stages of organogenesis at the time of laying, with separate facial prominences and pharyngeal clefts still visible. Limb buds are also visible. By 11 days (stage 3), the chondrocranium is nearly fully formed; however, few intramembranous bones can be detected. One week later (stage 4), many of the intramembranous upper and lower jaw bones are visible but the calvaria are not present. Skeletal elements in the limbs also begin to form. Between stages 4 (day 18) and 7 (day 44), the complete set of intramembranous bones in the jaws and calvaria develops. Hindlimb development does not progress beyond stage 6 (33 days) and remains rudimentary throughout adult life. In contrast to other reptiles, there are two rows of teeth in the upper jaw. The outer tooth row is attached to the maxillary and premaxillary bones, whereas the inner row is attached to the pterygoid and palatine bones. Erupted teeth can be seen in whole-mount stage 10 specimens and are present in an unerupted, mineralized state at stage 7. Micro-CT analysis reveals that all the young membranous bones can be recognized even out of the context of the skull. These data demonstrate intrinsic patterning of the intramembranous bones, even though they form without a cartilaginous template. In addition, intramembranous bone morphology is established prior to muscle function, which can influence bone shape through differential force application. After careful staging, we conclude that python skeletal development occurs slowly enough to observe in good detail the early stages of craniofacial skeletogenesis. Thus, reptilian animal models will offer unique opportunities for understanding the early influences that contribute to perinatal bone shape. r 2007 Elsevier GmbH. All rights reserved. Keywords: African Rock Python; Cartilage; Skeletogenesis; Embryo staging; Craniofacial development ARTICLE IN PRESS www.elsevier.de/zool 0944-2006/$ - see front matter r 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.zool.2007.01.005 Corresponding author. E-mail address: [email protected] (J.M. Richman). 1 Current address: Department of Cell Biology and Anatomy, Faculty of Medicine, Heritage Medical Research Centre, 3330 Hospital Drive NW, Calgary, Alta., Canada T2N 4N1. 2 These authors contributed equally to this work.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
ZOOLOGY
0944-2006/$ - se
doi:10.1016/j.zo
�CorrespondE-mail addr
1Current add
Calgary, Alta.,2These autho
Zoology 110 (2007) 212–230www.elsevier.de/zool
Embryonic development of Python sebae – I: Staging criteria and
macroscopic skeletal morphogenesis of the head and limbs
Julia C. Boughnera,1,2, Marcela Buchtovaa,2, Katherine Fua, Virginia Diewertb,Benedikt Hallgrımssonc, Joy M. Richmana,�
aDepartment of Oral Health Sciences, Life Sciences Institute, University of British Columbia, Life Sciences Centre,
2350 Health Sciences Mall, Vancouver, BC, Canada V6T 1Z3bDepartment of Oral Health Sciences, Faculty of Dentistry, University of British Columbia, 2199 Wesbrook Mall,
Vancouver, BC, Canada V6T 1Z3cDepartment of Cell Biology and Anatomy, Faculty of Medicine, Heritage Medical Research Centre, 3330 Hospital Drive NW,
Calgary, Alta., Canada T2N 4N1
Received 24 August 2006; received in revised form 21 January 2007; accepted 23 January 2007
Abstract
This study explores the post-ovipositional craniofacial development of the African Rock Python (Python sebae). We firstdescribe a staging system based on external characteristics and next use whole-mount skeletal staining supplemented withComputed tomography (CT) scanning to examine skeletal development. Our results show that python embryos are in earlystages of organogenesis at the time of laying, with separate facial prominences and pharyngeal clefts still visible. Limb buds arealso visible. By 11 days (stage 3), the chondrocranium is nearly fully formed; however, few intramembranous bones can bedetected. One week later (stage 4), many of the intramembranous upper and lower jaw bones are visible but the calvaria arenot present. Skeletal elements in the limbs also begin to form. Between stages 4 (day 18) and 7 (day 44), the complete set ofintramembranous bones in the jaws and calvaria develops. Hindlimb development does not progress beyond stage 6 (33 days)and remains rudimentary throughout adult life. In contrast to other reptiles, there are two rows of teeth in the upper jaw. Theouter tooth row is attached to the maxillary and premaxillary bones, whereas the inner row is attached to the pterygoid andpalatine bones. Erupted teeth can be seen in whole-mount stage 10 specimens and are present in an unerupted, mineralizedstate at stage 7. Micro-CT analysis reveals that all the young membranous bones can be recognized even out of the context ofthe skull. These data demonstrate intrinsic patterning of the intramembranous bones, even though they form without acartilaginous template. In addition, intramembranous bone morphology is established prior to muscle function, which caninfluence bone shape through differential force application. After careful staging, we conclude that python skeletaldevelopment occurs slowly enough to observe in good detail the early stages of craniofacial skeletogenesis. Thus, reptiliananimal models will offer unique opportunities for understanding the early influences that contribute to perinatal bone shape.r 2007 Elsevier GmbH. All rights reserved.
Keywords: African Rock Python; Cartilage; Skeletogenesis; Embryo staging; Craniofacial development
e front matter r 2007 Elsevier GmbH. All rights reserved.
ol.2007.01.005
ing author.
ess: [email protected] (J.M. Richman).
ress: Department of Cell Biology and Anatomy, Faculty of Medicine, Heritage Medical Research Centre, 3330 Hospital Drive NW,
Canada T2N 4N1.
rs contributed equally to this work.

ARTICLE IN PRESSJ.C. Boughner et al. / Zoology 110 (2007) 212–230 213
Introduction
Reptile biology has been investigated for over a centuryby many different workers, yet there remains a paucity ofstudies on embryos (Parker, 1879; Kamal and Hammou-da, 1965a–c; De Beer, 1937). The exceptions are thatreptilian embryos have been used to study tooth andpalate development (Lemus et al., 1980, 1986; Ferguson,1981a, b; Westergaard and Ferguson, 1987) and tempera-ture-dependent sex determination (Wibbels et al., 1998;Torres-Maldonado et al., 2001, 2002). More recently, non-avian reptiles, including snakes (Cohn and Tickle, 1999)and turtles (Nagashima et al., 2005; Ohya et al., 2005), arebecoming popular animal models among evolutionarydevelopmental biologists. The more recent studies arebeginning to clone genes and look at expression patterns inreptiles (Kuraku et al., 2005; Ohya et al., 2005). Comparedto mammalian and chicken models, much less is knownabout reptilian genomes (Matsuda et al., 2005). In order toidentify the most interesting molecular questions, it isnecessary to see where the structural differences liebetween reptilian and other amniote embryos. It is fromthis anatomical variation and our previous knowledge ofdevelopment in other vertebrates that we can formulatehypotheses about genetic and morphological changeduring evolution.
Here, we characterize a non-venomous, egg-layingsnake, Python sebae, as a developmental model withspecial emphasis on craniofacial development. Weselected an oviparous member of the Boidae snakefamily for several reasons. First, pythons lay largeclutches of eggs (40–100). Second, embryos are acces-sible during the time when craniofacial skeletal differ-entiation and odontogenesis are ongoing. Third, non-venomous snakes such as the python lack the specializedfangs and supporting jaw modifications of venomoussnakes. Although constrictor snakes have their ownsuite of synapomorphic jaw characters related toswallowing large prey whole, their jaw architecturemay be more typical of the class Serpentes. Themorphology of the snake skull incorporates key reptilianfeatures of the jaw and palatal complex, such as adoubly articulated jaw joint and a natural cleft palate.These features have been retained from the mostprimitive amniotes, and should inform us about bothsnake and amniote evolution.
Other than the late embryonic and prehatchingdevelopment of P. sebae (Branch and Patterson, 1975),there are little developmental data available for thissnake. The most comprehensive, detailed studies ofcranial and gnathic morphology, musculature andfunction are limited to adult pythons (P. sebae, P.
molurus; Frazzetta, 1959, 1966). These careful analysesof adult python morphology (Fig. 1) are a solidfoundation for our investigation of the prehatchingdevelopmental morphology of P. sebae.
To facilitate developmental work on snakes, we needto develop a staging system that is generally applicableto other members of this class. Excellent classic andwidely used staging tables have been published formouse (Theiler, 1972), chicken (Hamburger and Hamil-ton, 1951) and turtle (Yntema, 1968). There are alsothree staging tables for embryonic development insnakes: the viviparous garter snake, Thamnophis sirtalis
(Zehr, 1962); the viviparous asp viper, Vipera aspis
(Hubert and Dufaure, 1968); and, most recently, theoviparous monocled cobra, Naja kaouthia (Jackson,2002). Further, incomplete staging data were publishedfor the viviparous brown water snake Natrix taxispilota
(Franklin, 1945). However, to date no data are availablefor oviparous, non-venomous snakes. Therefore, one ofour aims is to provide as complete information aspossible on the external and internal morphology of P.
sebae to facilitate the staging of other oviparous snakes.In addition to staging, the other main focus of this
paper is craniofacial skeletal development. The adapta-tion of skeletal patterns during evolution is particularlyinteresting in the skull where gain and loss of fenestra-tions and joint morphology are important means ofclassifying an amniote (De Beer, 1937; Carroll, 1988).However, relatively little comparative work has beencarried out on the developing craniofacial complex inembryos. Here we use whole-mount skeletal stainingand computed tomography (CT) scanning to investigatethe ossification of each of the skull bones and cartilagesin P. sebae. We find that due to the relatively slowdevelopment of ectothermic reptiles, the timing ofossification can be resolved down to the level ofindividual bones. This is a distinct advantage ascompared to endothermic, more rapidly developingamniotes.
This work is our first of two papers on P. sebae, thesecond focusing on microscopic anatomy and cellulardynamics. We have separated the macroscopic from themicroscopic data in order to make it more convenientfor other investigators who want to use our data tostudy oviparous, non-venomous snakes.
Materials and methods
Python egg acquisition and incubation
We obtained P. sebae eggs from the RainforestReptile Refuge (Surrey, British Columbia, Canada).Eggs were laid in July 2004 and June 2006 after anundisturbed period of in utero incubation of approxi-mately 8 weeks (P. Springate, pers. comm.). Of theclutch of 40 eggs laid in 2004, we discarded 13 deador infertile eggs and incubated the surviving clutchin a sand-filled bucket in warm, humid conditions

ARTICLE IN PRESS
Fig. 1. Adult skull of Python sebae. (A) Cranial and upper jaw skeleton, lateral view, (B) lateral and (C) medial views of the lower
jaw skeleton, (D) palatal view of the cranium. Arrows show axes that are used in the current paper. (A) reproduced from Frazzetta
(1966) with permission of John Wiley & Sons, Inc.; (B–D) adapted from Frazzetta (1966). Abbr.: a – angular; bo – basioccipital; bs –
basisphenoid; co – coronoid; d – dentary bone; ec – ectopterygoid; eo – exoccipital; f – frontal bone; mx – maxilla; n – nasal bone;
p – parietal bone; pa – prearticular process of the compound bone; pf – prefrontal bone; pl – palatine bone; pm – premaxilla; po –
post-orbital bone; pr – prootic; ps – parasphenoid; pt – pterygoid; q – quadrate; s – stapes; sm – septomaxilla; so – supraorbital
bone; soc – supraoccipital bone; sp – splenial bone; st – supratemporal; su – surangular process of the compound bone; tc –
trabeculae cranii; v – vomer.
J.C. Boughner et al. / Zoology 110 (2007) 212–230214
(day temperature about 30 1C, night temperature about25 1C) for 1 month at the Rainforest Reptile Refuge. Wethen transported all remaining eggs to the RichmanLab, where they were incubated at 30 1C in a modifiedchicken egg incubator packed with moist vermiculite, ahighly water-absorbing material that properly cushionedand hydrated the eggs (minimum 80% humidity). In2006, we obtained 21 eggs from the same python matingpair and were able to collect five stage 1 specimens.Unfortunately, the other embryos were dead uponcollection. The normal total incubation period forP. sebae is between 80 and 90 days. All animal workwas reviewed and approved by the UBC Animal EthicsCommittee, certificate no. A04-0271.
Fixation, processing and skeletal staining
Embryos were removed from the egg and immersed inice-cold phosphate-buffered-saline immediately. Em-bryo weight and length were measured, and pictureswere taken of external head, body and tail morphology.For whole-mount skeletal staining, embryos were fixedin 100% ethanol for 4 days followed by 100% acetonefor 4 days. Embryos were stained for bone and cartilagewith Alizarin Red/Alcian Blue solution and were clearedin glycerol and potassium hydroxide as previously
described (Plant et al., 2000). No whole-mount skeletaldata were available for stage 9 due to damage to thecollected specimen.
CT scanning
Two fixed embryos (stage 6; stage 10) were CT-scanned to visualize skeletal structure in three dimen-sions (3D) at the 3D Morphometrics Centre, Universityof Calgary. The stage 6 embryo was scanned using aSkyscan 1072 100 kv micro-CT at 5 mm resolution. Thestage 10 embryo was scanned using a Scanco Viva-CT40 scanner at 25 mm resolution, which is more suitablefor larger animals.
Results
Here, we report the embryonic development of theoviparous, non-venomous African Rock Python(P. sebae). We first describe staging criteria based onexternal characteristics visible upon recovery of theembryo (Table 1). In addition, we describe the internaldevelopment of the craniofacial complex at each of thestages using whole-skeletal staining and micro-CT

ARTICLE IN PRESS
Table 1. Staging characteristics for post-oviposition snake
embryos
Stage External morphological characteristics
1 Mandibular process does not extend rostral to eye,
endolymphatic ducts present, no scales on body or
head, 5–6 body coils
2 Mandibular process extends rostral to eye
3 Mandibular process extends midway between eye
and upper jaw, scales visible on body only, 3–4 body
coils
4 Mandibular process lines up with upper jaw, 3 body
coils
5 Cervical flexure 4901, heart protrudes from body
cavity
6 Scales visible on the head, heart fully enclosed inside
body cavity, 2–3 body coils
7 Fusion of eyelids in the center of eye, 2 body coils
8 Cervical flexure 1201, body wall closed,
pigmentation starting on body and head, 2 body
coils
9 Cervical flexure 1801, brain not visible through skin,
1–2 body coils, pigmentation pattern is clear but
color is pale
10 Endolymphatic ducts not visible, embryo resembles
neonate python, with the head pigmentation being
darker than the body, 1–2 body coils. Egg tooth
visible intraorally
J.C. Boughner et al. / Zoology 110 (2007) 212–230 215
scanning. The timing and sequence of appearance ofskull bones is very dynamic and takes place over the firstmonth and a half of post-ovipositional (po) develop-ment. The sequence of ossification is therefore diagnos-tic of a particular stage in development. We also reportgrowth curves so, together with our other analyses, itwill be possible to predict the extent of tissue develop-ment for a particular stage (Table 2, Figs. 2A and B).
Table 2. External features of our sample of Python sebae at differ
Stage Days post-
oviposition
No. of
specimens
collected
Body length
(cm)
Body weight
(g)
M
h
le
(
1 1–3 5 No data 0.3–0.5
3 11, 12 14 10.0–13.5 (10) 0.82–1.6 (10)
4 18 4 14.0–15.0 (2) 1.5–2 (2) 1
6 33 3 16.0–19.5 4.2–6.1 1
7 44 3 22.0 (1) 9.4 (1) 1
8 54 1 26.0 8.7 1
9 61 1 37.0 26.0 N
10 75 1 48.0 50.5 N
Summary of Python sebae embryo mean body length (tip of upper jaw to t
mesencephalon to tip of upper jaw in stages 1–6 embryos and measured crown
were defined by several external morphological characteristics, the principal
measured (number in parentheses).
External morphology of P. sebae
In general, the main characters used for staginginclude the relative length of the mandible in relation tothe maxilla (a measure that, together with skeletal data,provides some indication of how far an embryo hasprogressed); the visibility of the developing brain, heartand the closure of body wall musculature; the angle ofthe cervical flexure; and the presence or appearance ofthe eyelid, scales and endolymphatic ducts (Table 1).Weight and length were also used to help stage embryos.Weight gain in P. sebae is exponential with increasingembryonic stage (Fig. 2A), whereas the relationshipbetween embryo length (head to tail) and stage is linear(Fig. 2B). Observation of certain characteristics dependson translucency of the embryo and thus it is best if freshspecimens are examined. These features include thevisibility of the endolymphatic duct, the brain and theheart.
Stage 1 (1–3 days po; Figs. 3A and B; Table 2). Scalesare totally absent from the body. Retinal pigmentationis light. The paired endolymphatic ducts are visible astwo white opacities, on the dorsal surface of the head(Fig. 3B). The facial prominences are visible, includingthe nasal slit delineating the frontonasal mass andlateral nasal prominences. In addition, the pairedmaxillary and mandibular prominences could be seen.From the lateral view, the tip of the mandibularprominences lined up with the rostral edge of the eye.The second and third pharyngeal arches and clefts arevisible (Fig. 3A and data not shown). Bilateral limbbuds are found in the pelvic area (Fig. 4A, B).
Stage 3 (11–12 days po; Figs. 3C and D; Table 2). Infresh embryos, the brain is visible through the transpar-ent neurocranial tissues (Fig. 3C). The length of themandibular process extends rostrally and usually ismidway between the eye and the tip of the upper jaw(Fig. 3D). The upper facial prominences are completely
ent stages of development
ean
ead
ngth
mm)
No. of
complete
body coils
Endolymphatic
ducts visible
Body wall
closed
Angle of
cervical
flexure
(degrees)
6.0 5–6 Yes No 901
8.0 (10) 3–4 Yes No 901
0.5 (2) 3 Yes No 901
2.8 2.5 Yes No 90–1201
5.2 (1) 2 Yes No 90–1201
7.9 2 Yes Yes 120–1801
A 1–2 Yes Yes 1801
A 1–2 No Yes 1801
ip of tail) and weight, and rostro-caudal head length (measured from
of head to tip of upper jaw in stages 7–10 embryos). Embryonic stages
of which are listed here. In stages 3, 4 and 7, a subset of embryos was

ARTICLE IN PRESS
Fig. 2. Embryonic growth charts for Python sebae.
(A) Embryo weight (g) versus post-oviposition developmental
age (days). Embryo weight gain was exponential over time.
(B) Embryo length (cm) versus age (days). Increase in embryo
length was linear over time. (A and B) Embryonic stages
(S) are listed for corresponding data points.
J.C. Boughner et al. / Zoology 110 (2007) 212–230216
fused. The retina is well pigmented; however, pigmentcells are absent from the skin. Scales are only visiblealong the dorsal body and not on the head. Ashindlimbs are rudimentary in modern snakes, externallimb size and morphology do not change from stage 3and cannot be used for staging embryos (Figs. 4C, E, G,I and K). The hemipenes are visible medial to the limbbud (Fig. 4C).
Stage 4 (18 days po; Figs. 3E and F; Table 2). Thebody wall musculature remains open in the ventralmidline, through which the heart is visible (not shown).Body scales are present both dorsally and ventrally (notshown). The eyelids remain open but cover about half ofthe eye (Fig. 3F).
Stage 6 (33 days po; Figs. 3G and H; Table 2). Headscales are visible for the first time in the lateral lower jawand lateral cranium (not shown). While the skin of thehead is translucent and unpigmented, both eyes havedark pigmentation. Eyelids form a band of approxi-mately 1/3 the width of eye radius (Fig. 3H).
Stage 7 (44 days po; Figs. 3I and J; Table 2). The bodywall musculature is still open along the ventral midlineand the heart remains visible through the body wall. Thebrain is still visible through the skull. Distinct scales arepresent on both the body and head (Fig. 3J). Thetransparent upper and lower eyelid edges have fusedtogether. The limbs and hemipenes are not as prominentas at stage 6 (Fig. 4I).
Stage 8 (54 days po; Figs. 3K and L; Table 2). Thebody wall musculature is now completely fused alongthe ventral midline. However, the heart remains visiblethrough the body wall (Fig. 3K). The pigmentation onthe head and body is distinct.
Stage 9 (61 days po; Figs. 3M and N; Table 2).The heart remains visible through the fused bodywall. The pigmentation of the scales on the head andbody now assumes a species-specific pattern but is stilllightly colored. The brain is no longer visible throughthe skin.
Stage 10 (75 days po; Figs. 3O and P; Table 2).The heart is no longer visible through the fused bodywall (Fig. 3O). Furthermore, the endolymphatic ductsare no longer visible through the skin of the head(Fig. 3O). The embryo closely resembles a neonatepython, with complex head and body pigmentation(Figs. 3O and P). Upon opening the mouth, the eggtooth is erupted and visible on the inside tip of the upperjaw (not shown).
Limb development
At stage 1, the python limb bud shape corresponds tothat of a stage-21 chicken. However, the chicken limbbud has a much more prominent apical ridge at a similarstage (Hamburger and Hamilton, 1951; Raynaud, 1985;Cohn and Tickle, 1999; Figs. 4A and B). By stage 3,there is no evidence of an ectodermal ridge (Fig. 4C).There are no skeletal elements yet present in the limbbuds (Fig. 4D). At stage 4, on each side of the pelvicarea, two simple and rod-like hindlimb cartilages havebegun to form between the body and tail, immediatelycaudal to the last rib (Fig. 4F). Both elements arealigned with each other along the same axis, parallel tothe body.
In the limb at stage 6, two cartilages are presentcaudal to the last ribs: the stylopod of the hindlimb(vestigial femur); and a pelvic cartilage with threeprocesses representing the ischium, ilium and pubiselements (Fig. 4H). Thus, the shape of the pelvic elementis tripartite and more complex at this stage compared tostage 4. Bilaterally at stage 7, the hindlimb budcartilages have not progressed any further as com-pared to stage 6 appendages (Figs. 4I and J). Thus,final limb skeletal pattern is established at stage 6 inthe python.
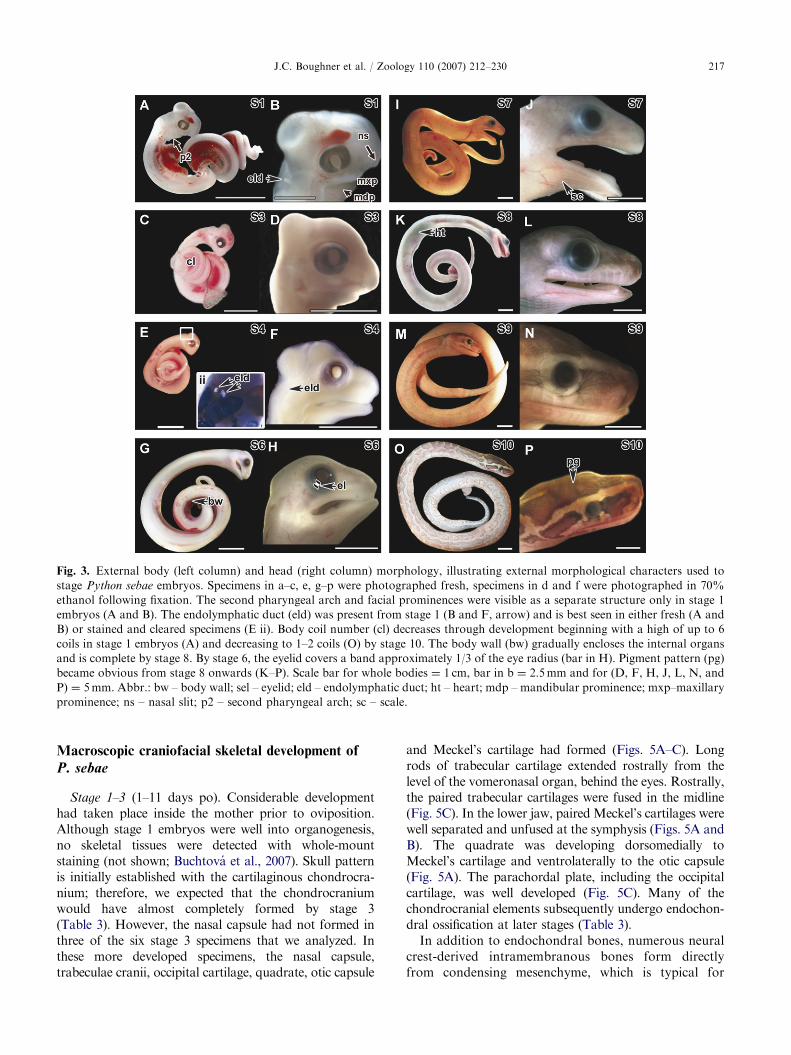
ARTICLE IN PRESS
Fig. 3. External body (left column) and head (right column) morphology, illustrating external morphological characters used to
stage Python sebae embryos. Specimens in a–c, e, g–p were photographed fresh, specimens in d and f were photographed in 70%
ethanol following fixation. The second pharyngeal arch and facial prominences were visible as a separate structure only in stage 1
embryos (A and B). The endolymphatic duct (eld) was present from stage 1 (B and F, arrow) and is best seen in either fresh (A and
B) or stained and cleared specimens (E ii). Body coil number (cl) decreases through development beginning with a high of up to 6
coils in stage 1 embryos (A) and decreasing to 1–2 coils (O) by stage 10. The body wall (bw) gradually encloses the internal organs
and is complete by stage 8. By stage 6, the eyelid covers a band approximately 1/3 of the eye radius (bar in H). Pigment pattern (pg)
became obvious from stage 8 onwards (K–P). Scale bar for whole bodies ¼ 1 cm, bar in b ¼ 2.5mm and for (D, F, H, J, L, N, and
P) ¼ 5mm. Abbr.: bw – body wall; sel – eyelid; eld – endolymphatic duct; ht – heart; mdp – mandibular prominence; mxp–maxillary
prominence; ns – nasal slit; p2 – second pharyngeal arch; sc – scale.
J.C. Boughner et al. / Zoology 110 (2007) 212–230 217
Macroscopic craniofacial skeletal development of
P. sebae
Stage 1–3 (1–11 days po). Considerable developmenthad taken place inside the mother prior to oviposition.Although stage 1 embryos were well into organogenesis,no skeletal tissues were detected with whole-mountstaining (not shown; Buchtova et al., 2007). Skull patternis initially established with the cartilaginous chondrocra-nium; therefore, we expected that the chondrocraniumwould have almost completely formed by stage 3(Table 3). However, the nasal capsule had not formed inthree of the six stage 3 specimens that we analyzed. Inthese more developed specimens, the nasal capsule,trabeculae cranii, occipital cartilage, quadrate, otic capsule
and Meckel’s cartilage had formed (Figs. 5A–C). Longrods of trabecular cartilage extended rostrally from thelevel of the vomeronasal organ, behind the eyes. Rostrally,the paired trabecular cartilages were fused in the midline(Fig. 5C). In the lower jaw, paired Meckel’s cartilages werewell separated and unfused at the symphysis (Figs. 5A andB). The quadrate was developing dorsomedially toMeckel’s cartilage and ventrolaterally to the otic capsule(Fig. 5A). The parachordal plate, including the occipitalcartilage, was well developed (Fig. 5C). Many of thechondrocranial elements subsequently undergo endochon-dral ossification at later stages (Table 3).
In addition to endochondral bones, numerous neuralcrest-derived intramembranous bones form directlyfrom condensing mesenchyme, which is typical for
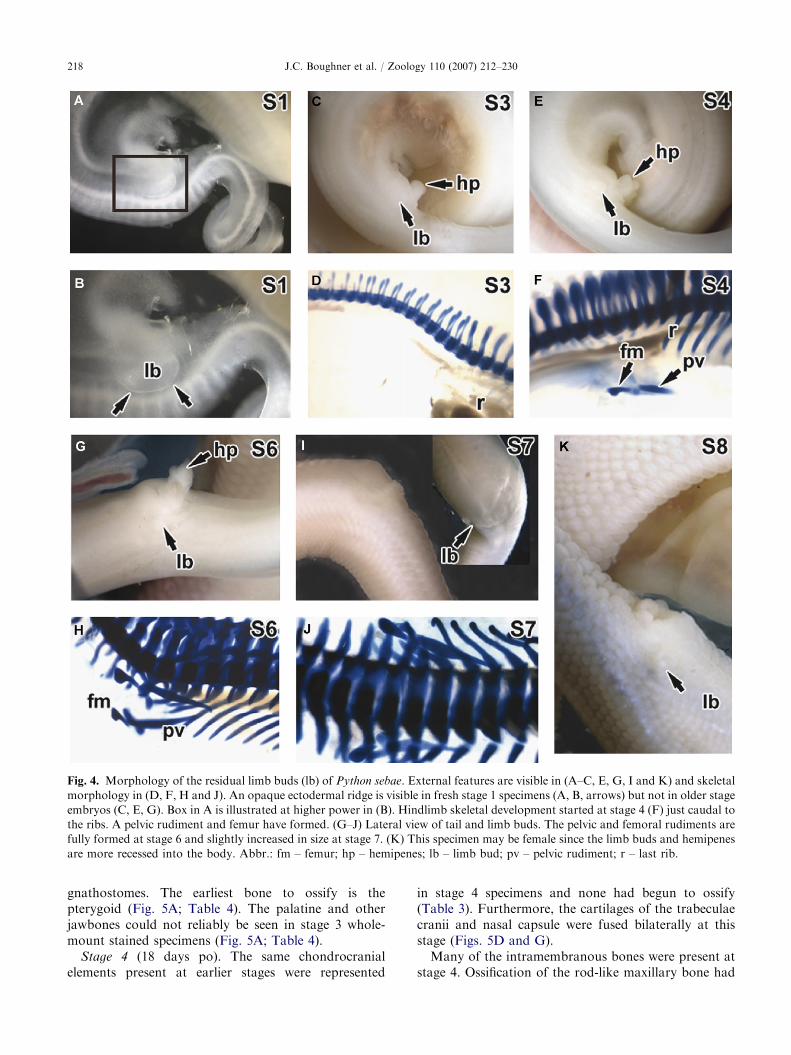
ARTICLE IN PRESS
Fig. 4. Morphology of the residual limb buds (lb) of Python sebae. External features are visible in (A–C, E, G, I and K) and skeletal
morphology in (D, F, H and J). An opaque ectodermal ridge is visible in fresh stage 1 specimens (A, B, arrows) but not in older stage
embryos (C, E, G). Box in A is illustrated at higher power in (B). Hindlimb skeletal development started at stage 4 (F) just caudal to
the ribs. A pelvic rudiment and femur have formed. (G–J) Lateral view of tail and limb buds. The pelvic and femoral rudiments are
fully formed at stage 6 and slightly increased in size at stage 7. (K) This specimen may be female since the limb buds and hemipenes
are more recessed into the body. Abbr.: fm – femur; hp – hemipenes; lb – limb bud; pv – pelvic rudiment; r – last rib.
J.C. Boughner et al. / Zoology 110 (2007) 212–230218
gnathostomes. The earliest bone to ossify is thepterygoid (Fig. 5A; Table 4). The palatine and otherjawbones could not reliably be seen in stage 3 whole-mount stained specimens (Fig. 5A; Table 4).
Stage 4 (18 days po). The same chondrocranialelements present at earlier stages were represented
in stage 4 specimens and none had begun to ossify(Table 3). Furthermore, the cartilages of the trabeculaecranii and nasal capsule were fused bilaterally at thisstage (Figs. 5D and G).
Many of the intramembranous bones were present atstage 4. Ossification of the rod-like maxillary bone had

Table 3. Endochondral bones and persistent cartilages in the craniofacial skeleton
Stage 1 Stage 3 Stage 4 Stage 6 Stage 7 Stage 8 Stage 10
Persistent cartilages
Meckel’s cartilage Mcs + + + + + +
Nasal capsule � + + + + + +
Vomeronasal cartilage � � � + + + +
Trabeculae cranii Mcs + + + + + +
Orbital cartilage � + + + + + +
Endochondral bones
Quadrate Mcs + + + Os Os Os
Hyoid Mcs + + + + Os Os
Parachordal plate (later
basisphenoid, basioccipital,
exooccipital, supraoccipital)
+ + + Os Os
(basisphenoid)
Os Os
Otic capsule (later prootic bone) + + + + Os Os Os
Summary of the embryonic stages at which the cartilages of the craniofacial skeleton are first observed in Python sebae. Persistent cartilages remained
intact until the end of the study (stage 10) with the exception of the posterior end of the trabeculae cranii, which undergoes perichondrial ossification
after stage 8. Note that stage 8 specimens were only analyzed in tissue sections but were included in the table for completeness. Key: (+) cartilage
present, (�) cartilage not yet developing, mcs – mesenchymal condensation in sections, Os – ossification.
J.C. Boughner et al. / Zoology 110 (2007) 212–230 219
begun at the lateral edges of the upper jaw (Figs. 5D, Fand G). Only one small ossification center wasrecognizable in this stage. The triangularly shapedpalatine bone and pterygoid bones could easily bedetected in whole-mount stained stage 4 embryos (Figs.5D–G). Ossification of the nasal bones had begun (Fig.5D, Table 4). Of the cranial vault bones, the prefrontaland frontal bones had just begun to ossify bilaterally(Fig. 5D).
The snake has a doubly jointed jaw articulation thatincreases the gape in order to swallow large prey.Instead of the quadrate bone or condyle nesting into afossa (i.e. the glenoid fossa), the snake quadratearticulates with the lateral side of the supratemporalbone of the skull (Frazzetta, 1966). At stage 6, thesupratemporal bone was beginning to develop from asingle ossification center, lateral to the otic capsule anddorsocaudal to the cartilaginous quadrate (Fig. 5E).
It was now possible to see some of the mandibularbones. Moving from proximal to distal (caudal torostral), the surangular and prearticular processes ofthe compound bone as well as the splenial bone hadformed (Figs. 5D–F). Distally, the dentary bone wasossifying directly lateral to Meckel’s cartilage (Figs. 5Dand F). All of these bones were separate but closelyrelated to Meckel’s cartilage as seen in other amniotes.
Stage 6 (33 days po). The key difference between stage6 and earlier embryos is that endochondral ossificationis starting in many of the cartilages (Table 3). Severalnew intramembranous bones had formed between stages4 and 6 including the premaxilla, vomer, ectopterygoidand parietal bones (Fig. 6A, Table 4). In addition tousing whole-mount staining to assess skeletal develop-ment (Fig. 6), we used high-resolution micro-CT priorto staining (Figs. 7A–H). The advantage of this method
is that it was possible to digitally dissect individual skullbones away from surrounding tissues; however, theless radio-dense cartilages were not captured usingmicro-CT.
Rostrally, the triangular premaxilla appeared as a singlebone (Figs. 6A, C and 7A–C, E). Caudally, the premaxillawas tripartite, composed of separate processes arising oneither side of the main body of the bone (Fig. 7E). Therewere two lateral palatine processes and a single midlinenasal process. In between these processes was the distalend of the trabecular cartilage, which at this stage wasjoining with the nasal capsule (Figs. 6A and C).
The maxillary bone extended from the premaxilla tothe pterygoid (Figs. 6C and 7B–D). The palatine bonehad developed a pterygoid process, although thepalatine and pterygoid bones were not yet in contactwith each other (Figs. 6D, G and 7D). For the first time,the ectopterygoid was visible dorsal to the maxillarybone and lateral to the pterygoid bones (Figs. 6C and7C, D, F). The paired septomaxillae were beginning toossify ventrolaterally to the vomers (Figs. 6C and 7B, C,H). The nasal bones were now ossifying in the mid-line, dorsal and exterior to the nasal capsular cartilage(Figs. 6A and 7A–C).
Calvarial bones that were ossifying included theprefrontal, frontal and parietal bones (Figs. 6A and7A–D). The prefrontal bone was ossifying in threedistinct but loosely connected parts (Figs. 7B–D). Oneprocess extended medially towards the midline of thefrontal bone. A second process extended laterally toform part of the roof of the orbit. A third processextended ventrolaterally towards the palato-maxillaryarch, which later articulates with this process. Thefrontal bone was ossifying dorsal to the prefrontal bone(Figs. 6A and 7A–C).
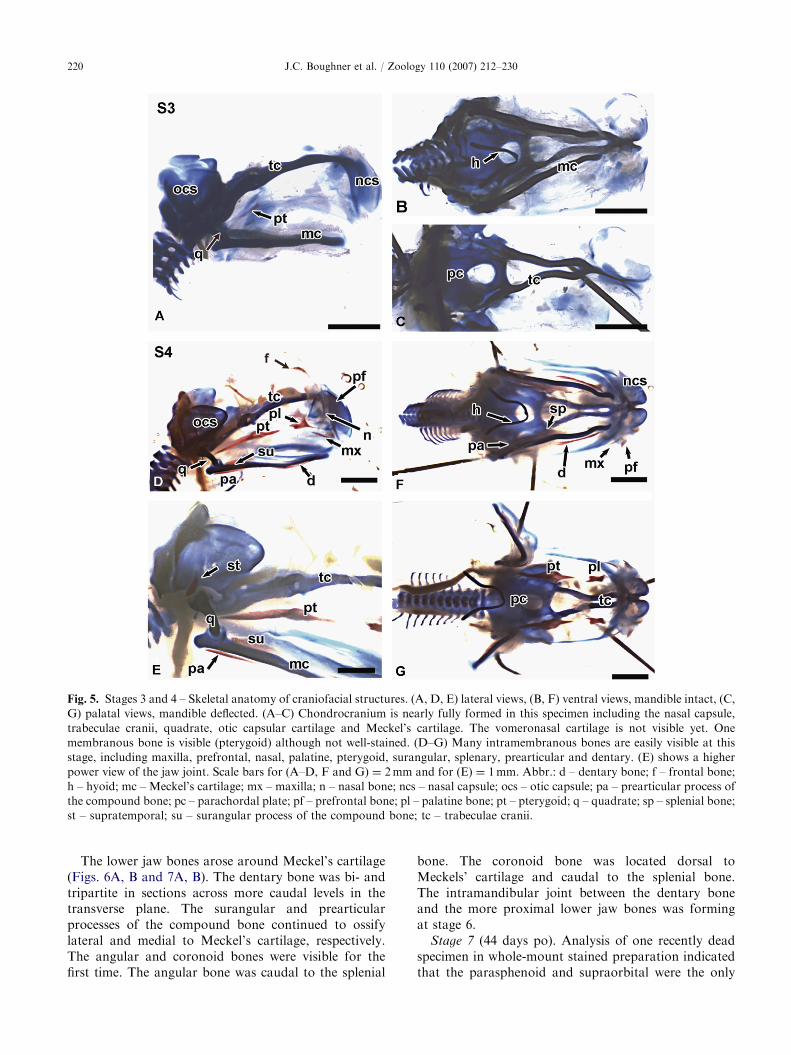
ARTICLE IN PRESS
Fig. 5. Stages 3 and 4 – Skeletal anatomy of craniofacial structures. (A, D, E) lateral views, (B, F) ventral views, mandible intact, (C,
G) palatal views, mandible deflected. (A–C) Chondrocranium is nearly fully formed in this specimen including the nasal capsule,
trabeculae cranii, quadrate, otic capsular cartilage and Meckel’s cartilage. The vomeronasal cartilage is not visible yet. One
membranous bone is visible (pterygoid) although not well-stained. (D–G) Many intramembranous bones are easily visible at this
stage, including maxilla, prefrontal, nasal, palatine, pterygoid, surangular, splenary, prearticular and dentary. (E) shows a higher
power view of the jaw joint. Scale bars for (A–D, F and G) ¼ 2mm and for (E) ¼ 1mm. Abbr.: d – dentary bone; f – frontal bone;
h – hyoid; mc – Meckel’s cartilage; mx – maxilla; n – nasal bone; ncs – nasal capsule; ocs – otic capsule; pa – prearticular process of
the compound bone; pc – parachordal plate; pf – prefrontal bone; pl – palatine bone; pt – pterygoid; q – quadrate; sp – splenial bone;
st – supratemporal; su – surangular process of the compound bone; tc – trabeculae cranii.
J.C. Boughner et al. / Zoology 110 (2007) 212–230220
The lower jaw bones arose around Meckel’s cartilage(Figs. 6A, B and 7A, B). The dentary bone was bi- andtripartite in sections across more caudal levels in thetransverse plane. The surangular and prearticularprocesses of the compound bone continued to ossifylateral and medial to Meckel’s cartilage, respectively.The angular and coronoid bones were visible for thefirst time. The angular bone was caudal to the splenial
bone. The coronoid bone was located dorsal toMeckels’ cartilage and caudal to the splenial bone.The intramandibular joint between the dentary boneand the more proximal lower jaw bones was formingat stage 6.
Stage 7 (44 days po). Analysis of one recently deadspecimen in whole-mount stained preparation indicatedthat the parasphenoid and supraorbital were the only

ARTICLE IN PRESS
Table 4. Sequence of ossification of intramembranous craniofacial bones
Stage 1 Stage 3 Stage 4 Stage 6 Stage 7 Stage 8 Stage 10
Mandibular complex
Dentary — 2 1 1 1 1 1
Compound bone (prearticular process) — 2 1 1 1 1 1
Compound bone (surangular process) — — 1 1 1 1 1
Splenial — — 1 1 1 1 1
Angular — — — 1 1 1 1
Coronoid — — — 1 1 1 1
Supratemporal — 2 1 1 1 1 1
Maxillary complex
Palatine — 2 1 1 1 1 1
Pterygoid — 1 1 1 1 1 1
Maxillary — — 1 1 1 1 1
Premaxilla — — 2 1 1 1 1
Vomer — — 2 1 1 1 1
Ectopterygoid — — — 1 1 1 1
Septomaxilla — — — 1 1 1 1
Cranial vault
Nasal — — 1 1 1 1 1
Parietal — — 2 1 1 1 1
Prefrontal — — 1 1 1 1 1
Frontal — — — 1 1 1 1
Basicranial
Parasphenoid — — — — 1 1 1
Postorbital — — — 1 1 1 1
Supraorbital — — — — 2 1 1
Schedule of appearance of craniofacial intramembranous bones of Python sebae as detected with either Alizarin Red staining in whole-mount or
Picosirius Red staining in sections. Once formed, bones remained intact until the end of the study (stage 10). Stage 8 specimens were only examined in
section. However, we are confident that the same bones would be visible in whole mount at stage 8 that were already visible at stage 7. Key: (—) bone
not observed, (1) bone present in whole-mount stain, (2) bone present in histological sections only.
J.C. Boughner et al. / Zoology 110 (2007) 212–230 221
new membranous bones to form at this stage, althoughonly the parasphenoid could be detected in whole-mount stained specimens (Figs. 6D and F; Table 4).Ossification of the quadrate, basisphenoid and prooticcartilages also had started (Table 3).
The vomer bone had three processes: the horizontalbody, and the medial and lateral vertical lamina (notshown). Rostrally, the shape of the maxillary bone wasmore complex, owing to the formation of processes formuscle attachments dorsally and ventrally (Figs. 6D andE). Further caudally, the maxillary bone was tubular inshape. The centrally located parasphenoid was firstobserved in the midline between the pterygoid andpalatine bones (Fig. 6F). The frontal bone was ossifyingdorsally and extended ventrally immediately lateral tothe brain to form part of the braincase and the medialwall of the orbit (Fig. 6D). Several unerupted teeth inthe upper jaw were sufficiently mineralized to be visible(Fig. 6D).
At stage 7, the spatial arrangement of the lower jawbones was similar to the arrangement found in the adultpython (Figs. 1B and C), where the coronoid (dorsal),prearticular (medial), angular (ventral) and surangular
(ventrolateral) were arranged around Meckel’s cartilage(Figs. 6D and E).
Stage 10 (75 days po). The formation of craniofacialand lower jaw cartilages and bones was complete atstage 10, 10–20 days before hatching (Figs. 8A–G;Tables 3 and 4). The CT images clearly showed thenear-complete ossification of the craniofacial bones(Figs. 8A–D). Following CT scanning, the specimenwas hemisected and one half was embedded in paraffin,while the other half was stained in whole mount. Themaxillary bone was structurally complex (Figs. 8A–G),with many fenestrations on its lateral aspect andnumerous processes for muscle attachment. At stage10, the lateral process of the palatine bone articulatedwith the prefrontal bone and contacted the maxillarybone (Figs. 8B and D). The rostral part of Meckel’scartilage was intact (Figs. 8E–G), consistent with thecommon persistence of this cartilage into post-natalstages in reptiles. An intramandibular joint was presenton each side of the mandible between the dentary boneand the compound bone (Figs. 8A and D). The caudaldentary bone formed a bifurcation, into which thesurangular process of the compound bone was recessed

ARTICLE IN PRESS
Fig. 6. Stages 6 and 7 – Skeletal anatomy of craniofacial structures. (A–C) The same stage 6 specimen as in Fig. 7, stained in whole
mount after CT scanning. (A) lateral view, (B) ventral view, mandible intact, (C) palatal view, mandible retracted. All membranous
bones are present by this stage. (D–F) Stage 7 skull in whole mount. Specimen has been distorted in medial–lateral plane due to
fixation after death of embryo in ovo. Nonetheless, more advanced development can be seen as compared to the stage 6 specimen.
(D) The single, midline, parasphenoid bone can be seen for the first time at stage 7. This bone is also clearly visible in the palatal view
(F). Unerupted upper teeth are visible (arrowheads in D). All mandibular bones are well developed. Scale bar for (A–F) ¼ 2.0mm.
Abbr.: a – angular; bs – basisphenoid; co – coronoid; d – dentary bone; ec – ectopterygoid; eld – endolymphatic duct; f – frontal
bone; imj – intramandibular joint; mc – Meckel’s cartilage; mx – maxilla; n – nasal bone; p – parietal bone; pa – prearticular process
of the compound bone; pf – prefrontal bone; pl – palatine bone; pm – premaxilla; po – post-orbital bone; pr – prootic; ps –
parasphenoid; pt – pterygoid; q – quadrate; st – supratemporal; su – surangular process of the compound bone; tc – trabeculae
cranii; v – vomer.
J.C. Boughner et al. / Zoology 110 (2007) 212–230222
(Figs. 8A and D). At stage 10, the first tooth generationwas erupted and five bones were tooth-bearing: thepremaxilla, maxillary, palatine, pterygoid and dentary(Figs. 8A–D). Thus, there were two rows of teeth in theupper jaw and one in the lower jaw.
Discussion
Our study of P. sebae further demonstrates that thegeneral process of skeletal differentiation is conserved,
while the pattern of bones – the ossification sequence,number, morphology, positions – is quite divergentamong reptiles, mammals and birds. Importantly,because the python develops more slowly than amammalian model animal, it is simpler to identify theorder of ossification for each of the membranous bonesin this snake. For example, in mouse, many dermalbones begin to ossify within about 24 h of eachother, beginning at embryonic day 15 (Johnson, 1933)(Table 6).

ARTICLE IN PRESS
Fig. 7. Micro-CT scans of the skull of Python sebae at stage 6. Cartilage does not have sufficient radiodensity to be detected.
(A) Lateral view with soft tissues illustrated around bones. (B) Tilted, lateral view showing both sides of the skull. The characteristic
morphology of most bones is visible at this stage. (C) Hemicrania show internal structures in medial view. This view reveals the
position of the vomer and septomaxilla, bones that are obscured from view in whole-mount preparations. (D) Palatal view of skull
with the mandible digitally removed, showing more clearly the position of the ectopterygoid. (E–H) Digitally dissected bones.
(E) Premaxilla – dorsal (i) and lateral (ii) views. (F) Left pterygoid and ectopterygoid – ventral (i) and lateral (ii) views. (G) Pterygoid
and ectopterygoid – bilateral, dorsal view. (H) Vomer and septomaxillae – anterior (i) and vomer, lateral (ii) views. These two bones
are at the limit of resolution of the CT machine. Scale bar in (B) applies to (A–D) ¼ 1.0mm, bar in (E) applies to (E–H) ¼ 1.0mm.
Abbr.: a – angular; co – coronoid; d – dentary bone; ec – ectopterygoid; f – frontal bone; mx – maxilla; n – nasal bone; p – parietal
bone; pa – prearticular process of the compound bone; pf – prefrontal bone; pl – palatine bone; pm – premaxilla; pt – pterygoid; sm
– septomaxilla; sp – splenial bone; st – supratemporal; su – surangular process of the compound bone; v – vomer.
J.C. Boughner et al. / Zoology 110 (2007) 212–230 223
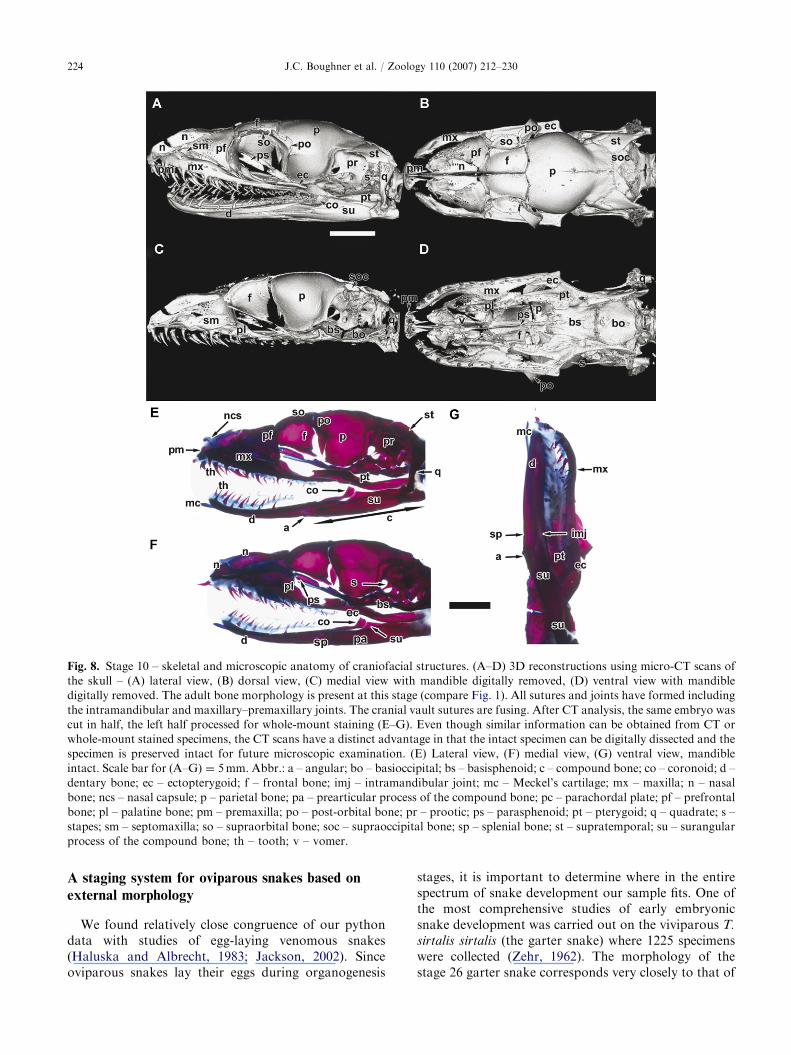
ARTICLE IN PRESS
Fig. 8. Stage 10 – skeletal and microscopic anatomy of craniofacial structures. (A–D) 3D reconstructions using micro-CT scans of
the skull – (A) lateral view, (B) dorsal view, (C) medial view with mandible digitally removed, (D) ventral view with mandible
digitally removed. The adult bone morphology is present at this stage (compare Fig. 1). All sutures and joints have formed including
the intramandibular and maxillary–premaxillary joints. The cranial vault sutures are fusing. After CT analysis, the same embryo was
cut in half, the left half processed for whole-mount staining (E–G). Even though similar information can be obtained from CT or
whole-mount stained specimens, the CT scans have a distinct advantage in that the intact specimen can be digitally dissected and the
specimen is preserved intact for future microscopic examination. (E) Lateral view, (F) medial view, (G) ventral view, mandible
intact. Scale bar for (A–G) ¼ 5mm. Abbr.: a – angular; bo – basioccipital; bs – basisphenoid; c – compound bone; co – coronoid; d –
dentary bone; ec – ectopterygoid; f – frontal bone; imj – intramandibular joint; mc – Meckel’s cartilage; mx – maxilla; n – nasal
bone; ncs – nasal capsule; p – parietal bone; pa – prearticular process of the compound bone; pc – parachordal plate; pf – prefrontal
bone; pl – palatine bone; pm – premaxilla; po – post-orbital bone; pr – prootic; ps – parasphenoid; pt – pterygoid; q – quadrate; s –
stapes; sm – septomaxilla; so – supraorbital bone; soc – supraoccipital bone; sp – splenial bone; st – supratemporal; su – surangular
process of the compound bone; th – tooth; v – vomer.
J.C. Boughner et al. / Zoology 110 (2007) 212–230224
A staging system for oviparous snakes based on
external morphology
We found relatively close congruence of our pythondata with studies of egg-laying venomous snakes(Haluska and Albrecht, 1983; Jackson, 2002). Sinceoviparous snakes lay their eggs during organogenesis
stages, it is important to determine where in the entirespectrum of snake development our sample fits. One ofthe most comprehensive studies of early embryonicsnake development was carried out on the viviparous T.
sirtalis sirtalis (the garter snake) where 1225 specimenswere collected (Zehr, 1962). The morphology of thestage 26 garter snake corresponds very closely to that of

ARTICLE IN PRESSJ.C. Boughner et al. / Zoology 110 (2007) 212–230 225
the stage 1 python and includes the following features:the external naris had not formed, the grooves betweenthe pharyngeal arches had not completely filled in, andthere were 5.5–6.25 coils (Zehr, 1962). Our assessmentalso agrees with that of others (Holtzman and Halpern,1991) who in the process of analyzing in detail theolfactory organs of the garter snake also presenteddetailed staging criteria for the face. Others (Haluskaand Albrecht, 1983) compared a 14-day Elaphe obsoleta
embryo (akin to stage 3 in the present study) to a stage28 garter snake. Taken together, our stage 1 embryos arebetween stages 23 and 28 in the Zehr staging system.
There were a total of 37 stages to reach theappearance of a hatchling garter snake and we havearrived at a similar number in our study (assuming stage26 at oviposition followed by an additional 10 distinctstages). Other viviparous snakes have also been stagedcarefully from inception but a larger number of stageswere described (Hubert and Dufaure, 1968; Hubert,1985). The main differences between the study by Zehr(1962) and those of Hubert (Hubert and Dufaure, 1968;Hubert, 1985) are about 6 or so extra stages duringgastrulation and neurulation. Similar headfold stageembryos are listed as stage 11 in Zehr (1962) but as stage16 in V. aspis (Hubert and Dufaure, 1968). Based on ourevaluation of the extra stages, it seems sufficient torecognize those of the garter snake as encompassing themajor morphological changes during early development.We agree that the absolute number of stages is not thecritical point but rather the features represented at eachstage. The utility of a systematic and standardizedstaging guide is the ability to scientifically evaluate howfar along a specimen has progressed in the continuum ofits development. On a practical note, reptilian develop-ment is very susceptible to external temperaturefluctuations, and staging is a more consistent way todescribe development versus actual days incubation(Holtzman and Halpern, 1991). Our comparison ofdifferent snake studies shows that different investigatorsrecognize similar numbers of time points at which newmorphological features could be identified (a total of 37,the last 10 of which are represented in oviparousspecies). Taken together with data from other studies,there is sound external validation for our staging table.
Missing data points in the present collection of P.sebae
Even though we were not able to collect specimensbetween 3 and 11 days, we can make several predictionsabout snake development at stage 2 from our data.First, we can reasonably assume that stage 2 beginsaround 5 days po. Judging by the very limitedossification of the dentary, palatine and pterygoid bonesin our stage 3 embryos, we would not expect to see these
bones with whole-mount skeletal staining in stage 2specimens.
Second, since our stage 4 specimens were collected at18 days and our stage 6 embryos at 33 days, stage 5would be about 25 days po. It is the day 25 time pointwhen we predict that the ossification of certain boneswill first become apparent. In a study on E. obsoleta
(Haluska and Albrecht, 1983), two intermediate stagesof 24 and 30 days incubation were collected, corre-sponding to late stage 4 and approximately mid stage 5in our study. At 24 days the ectopterygoid had begun toossify, whereas at 30 days the septomaxilla, exooccipital,compound and angular bones were ossifying. Thus it isvery likely that in P. sebae the ectopterygoid andangular bones would be present at approximately 25days po (stage 5). However, based on our data, thefrontal bone and coronoid would not have been presentany earlier than stage 6.
We emphasized collection at earlier stages of devel-opment in this study and as a result we are lacking somedetails in older specimens, in particular stage 9 speci-mens. One possibility is that stages 9 and 10 specimensmay be similar in terms of skeletal development andcould be collapsed into a single stage. However, wereject this idea since there were enough key differencesexternally to distinguish a stage 9 from a stage 10embryo. In summary, the data in Table 4 capture theinitiation of all the bones and little will change onceadditional older specimens are collected. The presenceor absence of intramembranous bones can be used as atool to assess how far development has progressed.
Rudimentary hindlimb development in P. sebae
In our study, hindlimbs were present as small buds atoviposition, along which an opaque ridge could be seenin the youngest specimens (approximately 18 h afterlaying). The apical ectodermal ridge (AER) was slightlythickened in relation to the rest of the limb ectoderm(Raynaud, 1985) in sections of new-laid, reticulatedpython embryos. However, in another study, sections ofpython limbs 24 h po had no AER, nor were genescharacteristic of the AER expressed in python limb buds(Cohn and Tickle, 1999). These differences could be dueto slight variations in egg-laying times. At the very least,if a ridge does form along the limb bud ectoderm, then itis very short-lived in the python.
A raised AER is required for normal limb outgrowthin birds. Experimental extirpation of the AER producedtruncated wings. Depending on the stage of AERremoval, either a stylopod (humerus or femur) alone,stylopod plus zeugopod (lower limb bones) or stylopod,zeugopod and truncated digits develop (Saunders, 1948;Summerbell, 1974). It is important to note that even ifthe AER is removed just after it has formed (H–H stage
ARTICLE IN PRESS
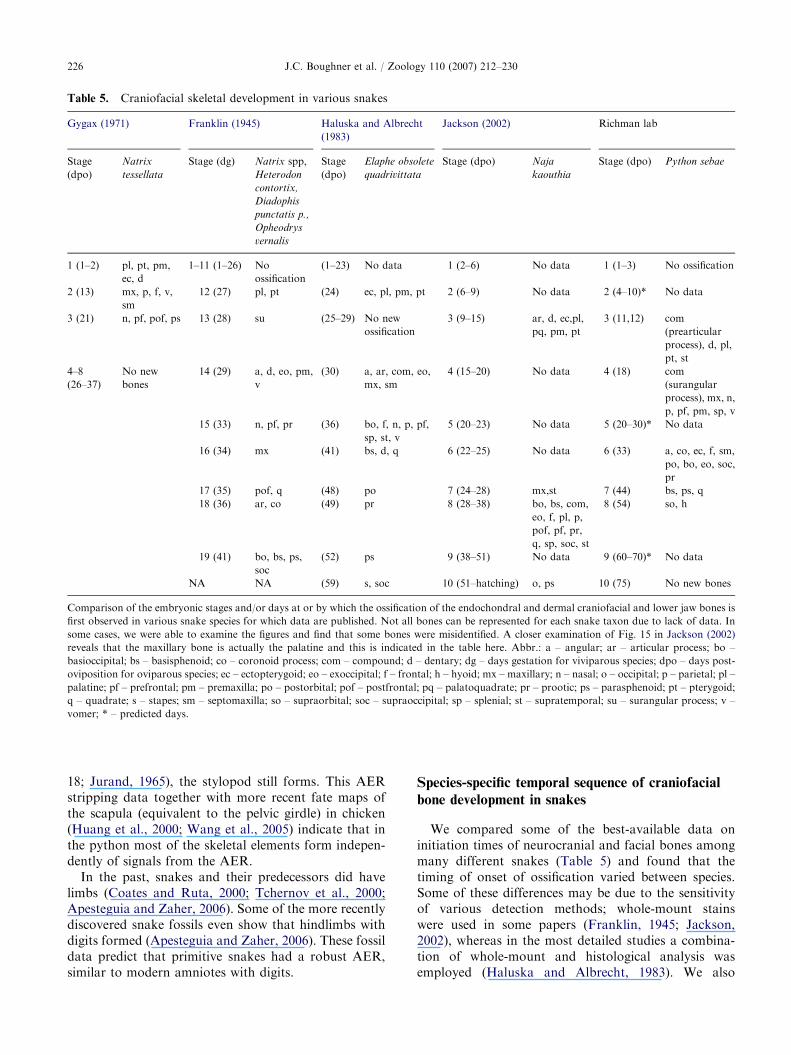
Table 5. Craniofacial skeletal development in various snakes
Gygax (1971) Franklin (1945) Haluska and Albrecht
(1983)
Jackson (2002) Richman lab
Stage
(dpo)
Natrix
tessellata
Stage (dg) Natrix spp,
Heterodon
contortix,
Diadophis
punctatis p.,
Opheodrys
vernalis
Stage
(dpo)
Elaphe obsolete
quadrivittata
Stage (dpo) Naja
kaouthia
Stage (dpo) Python sebae
1 (1–2) pl, pt, pm,
ec, d
1–11 (1–26) No
ossification
(1–23) No data 1 (2–6) No data 1 (1–3) No ossification
2 (13) mx, p, f, v,
sm
12 (27) pl, pt (24) ec, pl, pm, pt 2 (6–9) No data 2 (4–10)* No data
3 (21) n, pf, pof, ps 13 (28) su (25–29) No new
ossification
3 (9–15) ar, d, ec,pl,
pq, pm, pt
3 (11,12) com
(prearticular
process), d, pl,
pt, st
4–8
(26–37)
No new
bones
14 (29) a, d, eo, pm,
v
(30) a, ar, com, eo,
mx, sm
4 (15–20) No data 4 (18) com
(surangular
process), mx, n,
p, pf, pm, sp, v
15 (33) n, pf, pr (36) bo, f, n, p, pf,
sp, st, v
5 (20–23) No data 5 (20–30)* No data
16 (34) mx (41) bs, d, q 6 (22–25) No data 6 (33) a, co, ec, f, sm,
po, bo, eo, soc,
pr
17 (35) pof, q (48) po 7 (24–28) mx,st 7 (44) bs, ps, q
18 (36) ar, co (49) pr 8 (28–38) bo, bs, com,
eo, f, pl, p,
pof, pf, pr,
q, sp, soc, st
8 (54) so, h
19 (41) bo, bs, ps,
soc
(52) ps 9 (38–51) No data 9 (60–70)* No data
NA NA (59) s, soc 10 (51–hatching) o, ps 10 (75) No new bones
Comparison of the embryonic stages and/or days at or by which the ossification of the endochondral and dermal craniofacial and lower jaw bones is
first observed in various snake species for which data are published. Not all bones can be represented for each snake taxon due to lack of data. In
some cases, we were able to examine the figures and find that some bones were misidentified. A closer examination of Fig. 15 in Jackson (2002)
reveals that the maxillary bone is actually the palatine and this is indicated in the table here. Abbr.: a – angular; ar – articular process; bo –
basioccipital; bs – basisphenoid; co – coronoid process; com – compound; d – dentary; dg – days gestation for viviparous species; dpo – days post-
oviposition for oviparous species; ec – ectopterygoid; eo – exoccipital; f – frontal; h – hyoid; mx – maxillary; n – nasal; o – occipital; p – parietal; pl –
palatine; pf – prefrontal; pm – premaxilla; po – postorbital; pof – postfrontal; pq – palatoquadrate; pr – prootic; ps – parasphenoid; pt – pterygoid;
q – quadrate; s – stapes; sm – septomaxilla; so – supraorbital; soc – supraoccipital; sp – splenial; st – supratemporal; su – surangular process; v –
vomer; * – predicted days.
J.C. Boughner et al. / Zoology 110 (2007) 212–230226
18; Jurand, 1965), the stylopod still forms. This AERstripping data together with more recent fate maps ofthe scapula (equivalent to the pelvic girdle) in chicken(Huang et al., 2000; Wang et al., 2005) indicate that inthe python most of the skeletal elements form indepen-dently of signals from the AER.
In the past, snakes and their predecessors did havelimbs (Coates and Ruta, 2000; Tchernov et al., 2000;Apesteguia and Zaher, 2006). Some of the more recentlydiscovered snake fossils even show that hindlimbs withdigits formed (Apesteguia and Zaher, 2006). These fossildata predict that primitive snakes had a robust AER,similar to modern amniotes with digits.
Species-specific temporal sequence of craniofacial
bone development in snakes
We compared some of the best-available data oninitiation times of neurocranial and facial bones amongmany different snakes (Table 5) and found that thetiming of onset of ossification varied between species.Some of these differences may be due to the sensitivityof various detection methods; whole-mount stainswere used in some papers (Franklin, 1945; Jackson,2002), whereas in the most detailed studies a combina-tion of whole-mount and histological analysis wasemployed (Haluska and Albrecht, 1983). We also
ARTICLE IN PRESS
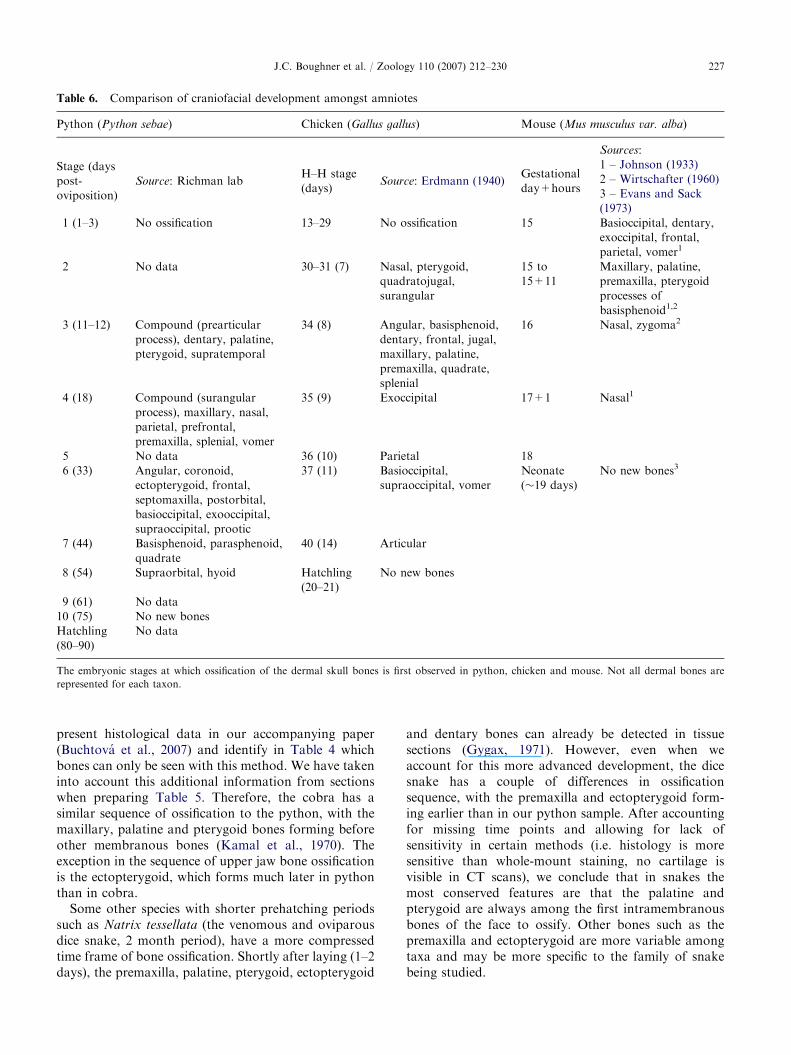
Table 6. Comparison of craniofacial development amongst amniotes
Python (Python sebae) Chicken (Gallus gallus) Mouse (Mus musculus var. alba)
Stage (days
post-
oviposition)
Source: Richman labH–H stage
(days)Source: Erdmann (1940)
Gestational
day+hours
Sources:
1 – Johnson (1933)
2 – Wirtschafter (1960)
3 – Evans and Sack
(1973)
1 (1–3) No ossification 13–29 No ossification 15 Basioccipital, dentary,
exoccipital, frontal,
parietal, vomer1
2 No data 30–31 (7) Nasal, pterygoid,
quadratojugal,
surangular
15 to
15+11
Maxillary, palatine,
premaxilla, pterygoid
processes of
basisphenoid1,2
3 (11–12) Compound (prearticular
process), dentary, palatine,
pterygoid, supratemporal
34 (8) Angular, basisphenoid,
dentary, frontal, jugal,
maxillary, palatine,
premaxilla, quadrate,
splenial
16 Nasal, zygoma2
4 (18) Compound (surangular
process), maxillary, nasal,
parietal, prefrontal,
premaxilla, splenial, vomer
35 (9) Exoccipital 17+1 Nasal1
5 No data 36 (10) Parietal 18
6 (33) Angular, coronoid,
ectopterygoid, frontal,
septomaxilla, postorbital,
basioccipital, exooccipital,
supraoccipital, prootic
37 (11) Basioccipital,
supraoccipital, vomer
Neonate
(�19 days)
No new bones3
7 (44) Basisphenoid, parasphenoid,
quadrate
40 (14) Articular
8 (54) Supraorbital, hyoid Hatchling
(20–21)
No new bones
9 (61) No data
10 (75) No new bones
Hatchling
(80–90)
No data
The embryonic stages at which ossification of the dermal skull bones is first observed in python, chicken and mouse. Not all dermal bones are
represented for each taxon.
J.C. Boughner et al. / Zoology 110 (2007) 212–230 227
present histological data in our accompanying paper(Buchtova et al., 2007) and identify in Table 4 whichbones can only be seen with this method. We have takeninto account this additional information from sectionswhen preparing Table 5. Therefore, the cobra has asimilar sequence of ossification to the python, with themaxillary, palatine and pterygoid bones forming beforeother membranous bones (Kamal et al., 1970). Theexception in the sequence of upper jaw bone ossificationis the ectopterygoid, which forms much later in pythonthan in cobra.
Some other species with shorter prehatching periodssuch as Natrix tessellata (the venomous and oviparousdice snake, 2 month period), have a more compressedtime frame of bone ossification. Shortly after laying (1–2days), the premaxilla, palatine, pterygoid, ectopterygoid
and dentary bones can already be detected in tissuesections (Gygax, 1971). However, even when weaccount for this more advanced development, the dicesnake has a couple of differences in ossificationsequence, with the premaxilla and ectopterygoid form-ing earlier than in our python sample. After accountingfor missing time points and allowing for lack ofsensitivity in certain methods (i.e. histology is moresensitive than whole-mount staining, no cartilage isvisible in CT scans), we conclude that in snakes themost conserved features are that the palatine andpterygoid are always among the first intramembranousbones of the face to ossify. Other bones such as thepremaxilla and ectopterygoid are more variable amongtaxa and may be more specific to the family of snakebeing studied.

ARTICLE IN PRESSJ.C. Boughner et al. / Zoology 110 (2007) 212–230228
CT scanning is of a sufficiently high resolution toreveal the complex architecture of very immature, smallbones. Thus, we were able to compare the initialmorphology of the early ossification center to the finalbone form. From observations on the septomaxillary,maxillary, premaxillary and vomer bones, a template ofadult bone morphology is visibly established at theoutset of prenatal development. Thus, there are intrinsicpatterning cues that operate early in organogenesis. Nodoubt post-natal muscle activity builds bone thickness incertain areas (Figs. 1A–D), but the overall bone shape ispreserved from its early organogenesis in Python.
Comparison of python craniofacial skeletal
development to that of other amniotes
The sequence of ossification and morphology of thepython craniofacial complex is more similar to birdsthan it is to mammals. In avian reptiles, the pterygoid isamong the first bones to ossify along with thequadratojugal and surangular (Erdmann, 1940).Furthermore, the pterygoid of the bird is quite similarto the snake pterygoid in that it is a separate bone,abutting the palatine bone rostrally and the quadratelaterally (Richman et al., 2006). In contrast, in mammalsthe pterygoid plates are thought to be processes of thesphenoid bone (Presley and Steel, 1978). This greatersimilarity between birds and snakes is to be expected,as birds are a recent radiation of reptiles (Poughet al., 2004; Zhou, 2004). What is interesting is thatpython and chicken facial bone morphologies remainso similar despite the evolution of the specialized birdbeak.
Since the pterygoid is the first skull bone to form inbirds (Erdmann, 1940) and snakes, we wondered howwidespread this characteristic is amongst reptilian taxa.The osteology of two turtle species has been studied indetail and, in both cases, the pterygoid begins to ossifyafter the maxilla and dentary bones (Sheil, 2003, 2005).Used as an outgroup, these turtle data suggest that theearly ossification of the pterygoid is a conserved featurelimited to snakes, birds and possibly other squamatesand not necessarily to other reptilian classes.
We found large differences in the sequence and timingof onset of ossification in mammals as compared toreptiles (Table 6). In mouse, the fetal period is relativelyshort and ossification occurs between days 15 and 17 ofgestation (Johnson, 1933; Wirtschafter, 1960; Evans andSack, 1973). Thus it is hard to determine the sequence ofossification to as fine a level as in slower-developingreptiles. Perhaps the use of other tools such as molecularmarkers for early commitment to an osteogenic lineageis a better way to study the sequence of ossification inmouse (Ducy et al., 1997; Otto et al., 1997; Strickeret al., 2002).
Preparing the groundwork for future studies on
snakes
Snakes are relatively rarely studied at embryonicstages and there is no standard accepted species fordevelopmental studies as there is for birds (chicken,quail) or mammals (mouse). Comparisons betweenspecies are difficult if there is no standardized setof observations taken at different times duringdevelopment. For example, by using chicken staging(Hamburger and Hamilton, 1951) it was possible tocompare finch or duck to chicken even though incuba-tion periods differ (Schneider and Helms, 2003; Abzha-nov et al., 2004; Wu et al., 2004). The staging table andskeletal data presented here will facilitate comparisonsbetween embryos of different snake species in futurestudies.
The craniofacial region is one of the most importantareas for determining phylogenetic relationships be-tween animals. Thus we should include non-avianreptiles, a branch of the evolutionary tree that haslargely been overlooked. While we must be cautiouswhen extrapolating studies of adult, highly derived,living snakes to reptilian and vertebrate evolution, theembryos are where true homologies can be identified.Thus, this paper on python embryo development willform an important foundation for further studies onreptilian, amniote and vertebrate evolution.
Acknowledgments
We are grateful to Paul Springate at The RainforestReptile Refuge, Surrey, British Columbia, whose expertreptile husbandry made this research possible. We thankJeff Dunn and the Medical Imaging technical staff at theUniversity of Calgary, Faculty of Medicine, for their 3Dimaging expertise and assistance. In particular, Wei Liu(University of Calgary) kindly provided technicalassistance with computed micro-tomography. This workwas first funded by CIHR and later by NSERC Grantsto J.M.R. J.M.R. is a Michael Smith DistinguishedScholar.
References
Abzhanov, A., Protas, M., Grant, B.R., Grant, P.R., Tabin,
C.J., 2004. Bmp4 and morphological variation of beaks in
Darwin’s finches. Science 305, 1462–1465.
Apesteguia, S., Zaher, H., 2006. A Cretaceous terrestrial snake
with robust hindlimbs and a sacrum. Nature 440,
1037–1040.
Branch, W.R., Patterson, R.W., 1975. Notes on the develop-
ment of embryos of the African Rock Python, Python sebae
(Serpentes: Boidea). J. Herpetol. 9, 243–248.

ARTICLE IN PRESSJ.C. Boughner et al. / Zoology 110 (2007) 212–230 229
Buchtova, M., Boughner, J.C., Fu, K., Diewert, V.M.,
Richman, J.M., 2007. Embryonic development of
Python sebae. II. Craniofacial microscopic anatomy, cell
proliferation and apoptosis. Zoology 110, doi:10.1016/
j.zool.2007.01.006.
Carroll, R.L., 1988. Vertebrate Paleontology and Evolution.
W.H. Freeman and Company, New York.
Coates, M., Ruta, M., 2000. Nice snake, shame about the legs.
Trends Ecol. Evol. 15, 503–507.
Cohn, M.J., Tickle, C., 1999. Developmental basis of
limblessness and axial patterning in snakes. Nature 399,
474–479.
De Beer, G.R., 1937. The Development of the Vertebrate
Skull. Oxford University Press, Oxford (Reprinted in 1985
by The University of Chicago Press, Chicago).
Ducy, P., Zhang, R., Geoffroy, V., Ridall, A.L., Karsenty, G.,
1997. Osf2/Cbfa1: a transcriptional activator of osteoblast
differentiation. Cell 89, 747–754.
Erdmann, K., 1940. Zur Entwicklungsgeschichte der
Knochen im Schadel des Huhnes bis zum Zeitpunkt des
Ausschlupfens aus dem Ei. Z. Morphol. Okol. Tiere 36,
315–400.
Evans, H.E., Sack, W.O., 1973. Prenatal development of
domestic and laboratory mammals: growth curves, external
features and selected references. Anat., Histol., Embryol. 2,
11–45.
Ferguson, M.W., 1981a. Review: the value of the American
alligator (Alligator mississippiensis) as a model for research
in craniofacial development. J. Craniofac. Genet. Dev. Biol.
1, 123–144.
Ferguson, M.W., 1981b. The structure and development of the
palate in Alligator mississippiensis. Arch. Oral Biol. 26,
427–443.
Franklin, M.A., 1945. The embryonic appearance of centres of
ossification in the bones of snakes. Copeia 2, 68–72.
Frazzetta, T.H., 1959. Studies on the morphology of the skull
in the Boidae (Serpentes). I. Cranial differences between
Python sebae and Epicrates cenchris. Bull. Mus. Comp.
Zool. 119, 451–472.
Frazzetta, T.H., 1966. Studies on the morphology and
function of the skull in the Boidae (Serpentes). II.
Morphology and function of the jaw apparatus in Python
sebae and Python molurus. J. Morphol. 118, 217–295.
Gygax, P., 1971. Development, morphology and function of
the venom gland (Duvernoy’s gland) of Natrix tessellata.
Acta Trop. 28, 226–274.
Haluska, F., Albrecht, P., 1983. The cranial development of
Elaphe obsoleta (Ophidia, Colubridae). J. Morphol. 178,
37–55.
Hamburger, V., Hamilton, H., 1951. A series of normal stages
in the development of the chick embryo. J. Morphol. 88,
49–92.
Holtzman, D.A., Halpern, M., 1991. Incorporation of 3H-
thymidine in the embryonic vomeronasal and olfactory
epithelial of garter snakes. J. Comp. Neurol. 304, 435–449.
Huang, R., Zhi, Q., Patel, K., Wilting, J., Christ, B., 2000.
Dual origin and segmental organisation of the avian
scapula. Development 127, 3789–3794.
Hubert, J., 1985. Embryology of the Squamata. Wiley,
Toronto.
Hubert, J., Dufaure, J.P., 1968. Table de development de la
vipere aspic, Vipera aspis. Bull. Soc. Zool. France 93,
135–148.
Jackson, K., 2002. Post-ovipositional development of the
monocled cobra, Naja kaouthia (Serpentes: Elapidae).
Zoology 105, 203–214.
Johnson, M.L., 1933. The time and order of appearance of
ossification centres in the albino mouse. Am. J. Anat. 52,
241–271.
Jurand, A., 1965. Ultrastructural aspects of early development
of fore-limb buds in chick and mouse. Proc. R. Soc. Lond.
Ser. B—Biol. Sci. 162, 387–405.
Kamal, A.M., Hammouda, H.G., 1965a. The development of
the skull of Psammophis sibilans. I. The development of the
chondrochranium. J. Morphol. 116, 197–245.
Kamal, A.M., Hammouda, H.G., 1965b. The development of
the skull of Psammophis sibilans. II. The fully formed
chondrocranium. J. Morphol. 116, 247–296.
Kamal, A.M., Hammouda, H.G., 1965c. The development of
the skull of Psammophis sibilans. III. The osteocranium of a
late embryo. J. Morphol. 116, 297–310.
Kamal, A.M., Hammouda, H.G., Mokhtar, F.M., 1970. The
development of the osteocraniium of the Egyptian Cobra: I.
The embryonic osteocranium. Acta Zool. 1970, 1–17.
Kuraku, S., Usuda, R., Kuratani, S., 2005. Comprehensive
survey of carapacial ridge-specific genes in turtle implies co-
option of some regulatory genes in carapace evolution.
Evol. Dev. 7, 3–17.
Lemus, D., Paz de la Vega, Y., Fuenzalida, M., Illanes, J.,
1980. In vitro differentiation of tooth buds from embryos
and adult lizards (L. gravenhorsti): an ultrastructural
comparison. J. Morphol. 165, 225–236.
Lemus, D., Coloma, L., Fuenzalida, M., Illanes, J., Paz de la
Vega, Y., Ondarza, A., Blanquez, M.J., 1986. Odontogen-
esis and amelogenesis in interacting lizard–quail tissue
combinations. J. Morphol. 189, 121–129.
Matsuda, Y., Nishida-Umehara, C., Tarui, H., Kuroiwa, A.,
Yamada, K., Isobe, T., Ando, J., Fujiwara, A., Hirao, Y.,
Nishimura, O., Ishijima, J., Hayashi, A., Saito, T.,
Murakami, T., Murakami, Y., Kuratani, S., Agata, K.,
2005. Highly conserved linkage homology between birds
and turtles: bird and turtle chromosomes are precise
counterparts of each other. Chromosome Res. 13, 601–615.
Nagashima, H., Uchida, K., Yamamoto, K., Kuraku, S.,
Usuda, R., Kuratani, S., 2005. Turtle–chicken chimera: an
experimental approach to understanding evolutionary
innovation in the turtle. Dev. Dyn. 232, 149–161.
Ohya, Y.K., Kuraku, S., Kuratani, S., 2005. Hox code in
embryos of Chinese soft-shelled turtle Pelodiscus sinensis
correlates with the evolutionary innovation in the turtle.
J. Exp. Zool. Part B—Mol. Dev. Evol. 304B, 107–118.
Otto, F., Thornell, A.P., Crompton, T., Denzel, A., Gilmour,
K.C., Rosewell, I.R., Stamp, G.W., Beddington, R.S.,
Mundlos, S., Olsen, B.R., Selby, P.B., Owen, M.J., 1997.
Cbfa1, a candidate gene for cleidocranial dysplasia
syndrome, is essential for osteoblast differentiation and
bone development. Cell 89, 765–771.
Parker, W.K., 1879. On the structure and development of the
skull in the common snake, Tropidonotus natrix. Philos.
Trans. R. Soc. Lond. (Biol.) 169, 385–417.

ARTICLE IN PRESSJ.C. Boughner et al. / Zoology 110 (2007) 212–230230
Plant, M.R., MacDonald, M.E., Grad, L.I., Ritchie, S.J.,
Richman, J.M., 2000. Locally released retinoic acid
repatterns the first branchial arch cartilages in vivo. Dev.
Biol. 222, 12–26.
Pough, F.H., Andrews, R.M., Cadle, R.M., Crump, M.L.,
Savitzky, A.H., Wells, K.D., 2004. Herpetology. Pearson
Prentice Hall, Upper Saddle River.
Presley, R., Steel, F.L., 1978. The pterygoid and ectopterygoid
in mammals. Anat. Embryol. 154, 95–110.
Raynaud, A., 1985. Development of limbs and embryonic limb
reduction. In: Gans, C. (Ed.), Biology of the Reptilia,
Development B. Wiley, Toronto, pp. 117–148.
Richman, J.M., Buchtova, M., Boughner, J.C., 2006. Com-
parative ontogeny and phylogeny of the upper jaw skeleton
in amniotes. Dev. Dyn. 235, 1230–1243.
Saunders, J.W., 1948. The proximo-distal sequence of origin of
the parts of the chicken wing and the role of the ectoderm.
J. Exp. Zool. 108, 363–403.
Schneider, R.A., Helms, J.A., 2003. The cellular and molecular
origins of beak morphology. Science 299, 565–568.
Sheil, C.A., 2003. Osteology and skeletal development of
Apalone spinifera (Reptilia: Testudines: Trionychidae).
J. Morphol. 256, 42–78.
Sheil, C.A., 2005. Skeletal development of Macrochelys
temminckii (Reptilia: Testudines: Chelydridae). J. Morphol.
263, 71–106.
Stricker, S., Fundele, R., Vortkamp, A., Mundlos, S., 2002.
Role of Runx genes in chondrocyte differentiation. Dev.
Biol. 245, 95–108.
Summerbell, D., 1974. A quantitative analysis of the effect of
excision of the AER from the chick limb-bud. J. Embryol.
Exp. Morphol. 32, 651–660.
Tchernov, E., Rieppel, O., Zaher, H., Polcyn, M.J., Jacobs, L.L.,
2000. A fossil snake with limbs. Science 287, 2010–2012.
Theiler, K., 1972. The House Mouse: Development and
Normal Stages from Fertilization to 4 Weeks of Age.
Springer, New York.
Torres-Maldonado, L., Moreno-Mendoza, N., Landa, A.,
Merchant-Larios, H., 2001. Timing of SOX9 down-
regulation and female sex determination in gonads of
the sea turtle Lepidochelys olivacea. J. Exp. Zool. 290,
498–503.
Torres Maldonado, L.C., Landa Piedra, A., Moreno Mendo-
za, N., Marmolejo Valencia, A., Meza Martinez, A.,
Merchant Larios, H., 2002. Expression profiles of Dax1,
Dmrt1, and Sox9 during temperature sex determination in
gonads of the sea turtle Lepidochelys olivacea. Gen. Comp.
Endocrinol. 129, 20–26.
Wang, B., He, L., Ehehalt, F., Geetha-Loganathan, P.,
Nimmagadda, S., Christ, B., Scaal, M., Huang, R., 2005.
The formation of the avian scapula blade takes place in the
hypaxial domain of the somites and requires somatopleure-
derived BMP signals. Dev. Biol. 287, 11–18.
Westergaard, B., Ferguson, M.W., 1987. Development of
dentition in Alligator mississippiensis. Later development in
the lower jaws of embryos, hatchlings and young juveniles.
J. Zool. 212, 191–222.
Wibbels, T., Cowan, J., LeBoeuf, R., 1998. Temperature-
dependent sex determination in the red-eared slider turtle,
Trachemys scripta. J. Exp. Zool. 281, 409–416.
Wirtschafter, Z.T., 1960. The Genesis of the Mouse Skeleton:
A Laboratory Atlas. Charles C Thomas, Springfield, IL.
Wu, P., Jiang, T.X., Suksaweang, S., Widelitz, R.B., Chuong,
C.M., 2004. Molecular shaping of the beak. Science 305,
1465–1466.
Yntema, C.L., 1968. A series of stages in the embryonic
development of Chelydra serpentina. J. Morphol. 125,
219–251.
Zehr, D.R., 1962. Stages in the normal development of the
common garter snake, Thamnophis sirtalis sirtalis. Copeia
1962 (2), 322–329.
Zhou, Z., 2004. The origin and early evolution of birds:
discoveries, disputes, and perspectives from fossil evidence.
Naturwissenschaften 91, 455–471.
Related Documents











