Embryology of the heart and its impact on understanding fetal and neonatal heart disease Adriana C. Gittenberger-de Groot a, b, * , Margot M. Bartelings b , Robert E. Poelmann b , Monique C. Haak c , Monique R.M. Jongbloed a, b a Department of Anatomy and Embryology, Leiden University Medical Center, Leiden, The Netherlands b Department of Cardiology, Leiden University Medical Center, Leiden, The Netherlands c Department of Obstetrics and Gynecology, Leiden University Medical Center, Leiden, The Netherlands Keywords: Cardiac development Congenital heart malformation Epicardium First heart field Neural crest cells Nuchal translucency Second heart field summary Heart development is a complex process during which the heart needs to transform from a single tube towards a fully septated heart with four chambers and a separated outflow tract. Several major events contribute to this process, that largely overlap in time. Abnormal heart development results in congenital heart disease, which has an estimated incidence of 1% of liveborn children. Eighty percent of cases of congenital heart disease are considered to have a multifactoral developmental background, whereas knowledge of monogenetic causes for congenital heart disease is still limited. This review focuses on several novel findings in cardiac development that might enhance our knowledge of aetiology and support refinement of prenatal diagnosis of congenital heart disease. Ó 2013 Published by Elsevier Ltd. 1. Introduction In recent years novel findings in cardiac development have had an impact on perinatal clinical diagnosis. A better understanding of the still limited number of monogenetic causes for congenital heart disease (CHD) opens up new avenues for research in the develop- mental background of the approximately 80% of cases with a multifactorial origin of fetal and neonatal CHD. This review focuses on several new findings in the development of cardiac structures that might further enhance our knowledge of aetiology and support refinement of prenatal diagnosis. As there are marked homologies in avian and mammal, including human, cardiac development, research results from the various species will be integrated in this review. 2. Origin of the embryonic cardiac tube and looping The bilateral cardiogenic fields in the embryonic mesoderm merge in the midline and form a primary cardiac tube, lined on the inside by endocardium and on the outside by myocardium con- sisting of about two cell layers. A thick basement membrane is sandwiched in between referred to as cardiac jelly, containing water-binding extracellular matrix molecules including hyaluronic acid. At a later stage the cardiac jelly is restricted to endocardial cushions lining the myocardial outflow tract (OFT) and the atrio- ventricular (AV) canal. The primary cardiac tube is genetically primed to have a dextral looping. During looping the connection to the dorsal body wall mesoderm is disrupted except at the venous and arterial poles where vessels enter and leave the heart tube. At the venous pole the cardinal veins become confluent in the sinus venosus supplying venous blood (oxygen rich in the embryo) to the heart, while at the arterial pole the blood is pumped into the aortic sac connecting to the bilateral pharyngeal arch arteries and the dorsal aortae. By contrast with the general view in the 1990s, the primary heart tube does not contain in miniature all elements of the mature heart. The myocardium-lined primary heart tube initially consists of a small atrial component (connected to the sinus venosus), an atrioventricular canal, a ventricular inflow tract (VIT) and a small OFT (connecting to the aortic sac). On the borderline of VIT and OFT a bulboventricular or primary fold is present (Fig. 1a). These cardiac components are derived from the mesoderm of the first heart field. The addition of dorsal cardiac mesoderm positioned between the primary heart tube and the primitive gut, the so-called second heart field (SHF) mesoderm, 1 is essential for the subsequent development of all cardiac components (Fig. 2). It may be argued that first and second heart fields are continuous in time and space, * Corresponding author. Address: Department of Cardiology, Leiden University Medical Center, Postal Zone: S-5-24, P.O. Box 9600, 2300 RC Leiden, The Netherlands. Tel.: þ31 71 6252020 (9300); fax: þ31 71 526 6809. E-mail address: [email protected] (A.C. Gittenberger-de Groot). Contents lists available at SciVerse ScienceDirect Seminars in Fetal & Neonatal Medicine journal homepage: www.elsevier.com/locate/siny 1744-165X/$ e see front matter Ó 2013 Published by Elsevier Ltd. http://dx.doi.org/10.1016/j.siny.2013.04.008 Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

at SciVerse ScienceDirect
Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244
Contents lists available
Seminars in Fetal & Neonatal Medicine
journal homepage: www.elsevier .com/locate/s iny
Embryology of the heart and its impact on understanding fetal andneonatal heart disease
Adriana C. Gittenberger-de Groot a,b,*, Margot M. Bartelings b, Robert E. Poelmann b,Monique C. Haak c, Monique R.M. Jongbloed a,b
aDepartment of Anatomy and Embryology, Leiden University Medical Center, Leiden, The NetherlandsbDepartment of Cardiology, Leiden University Medical Center, Leiden, The NetherlandscDepartment of Obstetrics and Gynecology, Leiden University Medical Center, Leiden, The Netherlands
Keywords:Cardiac developmentCongenital heart malformationEpicardiumFirst heart fieldNeural crest cellsNuchal translucencySecond heart field
* Corresponding author. Address: Department of CMedical Center, Postal Zone: S-5-24, P.O. Box 9Netherlands. Tel.: þ31 71 6252020 (9300); fax: þ31 7
E-mail address: [email protected] (A.C. Gittenberge
1744-165X/$ e see front matter � 2013 Published byhttp://dx.doi.org/10.1016/j.siny.2013.04.008
s u m m a r y
Heart development is a complex process during which the heart needs to transform from a single tubetowards a fully septated heart with four chambers and a separated outflow tract. Several major eventscontribute to this process, that largely overlap in time. Abnormal heart development results in congenitalheart disease, which has an estimated incidence of 1% of liveborn children. Eighty percent of cases ofcongenital heart disease are considered to have a multifactoral developmental background, whereasknowledge of monogenetic causes for congenital heart disease is still limited. This review focuses onseveral novel findings in cardiac development that might enhance our knowledge of aetiology andsupport refinement of prenatal diagnosis of congenital heart disease.
� 2013 Published by Elsevier Ltd.
1. Introduction
In recent years novel findings in cardiac development have hadan impact on perinatal clinical diagnosis. A better understanding ofthe still limited number of monogenetic causes for congenital heartdisease (CHD) opens up new avenues for research in the develop-mental background of the approximately 80% of cases with amultifactorial origin of fetal and neonatal CHD. This review focuseson several new findings in the development of cardiac structuresthat might further enhance our knowledge of aetiology and supportrefinement of prenatal diagnosis. As there are marked homologiesin avian and mammal, including human, cardiac development,research results from the various species will be integrated in thisreview.
2. Origin of the embryonic cardiac tube and looping
The bilateral cardiogenic fields in the embryonic mesodermmerge in the midline and form a primary cardiac tube, lined on theinside by endocardium and on the outside by myocardium con-sisting of about two cell layers. A thick basement membrane is
ardiology, Leiden University600, 2300 RC Leiden, The1 526 6809.r-de Groot).
Elsevier Ltd.
sandwiched in between referred to as cardiac jelly, containingwater-binding extracellular matrix molecules including hyaluronicacid. At a later stage the cardiac jelly is restricted to endocardialcushions lining the myocardial outflow tract (OFT) and the atrio-ventricular (AV) canal. The primary cardiac tube is geneticallyprimed to have a dextral looping. During looping the connection tothe dorsal body wall mesoderm is disrupted except at the venousand arterial poles where vessels enter and leave the heart tube. Atthe venous pole the cardinal veins become confluent in the sinusvenosus supplying venous blood (oxygen rich in the embryo) to theheart, while at the arterial pole the blood is pumped into the aorticsac connecting to the bilateral pharyngeal arch arteries and thedorsal aortae.
By contrast with the general view in the 1990s, the primaryheart tube does not contain in miniature all elements of the matureheart. The myocardium-lined primary heart tube initially consistsof a small atrial component (connected to the sinus venosus), anatrioventricular canal, a ventricular inflow tract (VIT) and a smallOFT (connecting to the aortic sac). On the borderline of VIT and OFTa bulboventricular or primary fold is present (Fig. 1a). These cardiaccomponents are derived from the mesoderm of the first heart field.The addition of dorsal cardiac mesoderm positioned between theprimary heart tube and the primitive gut, the so-called secondheart field (SHF) mesoderm,1 is essential for the subsequentdevelopment of all cardiac components (Fig. 2). It may be arguedthat first and second heart fields are continuous in time and space,
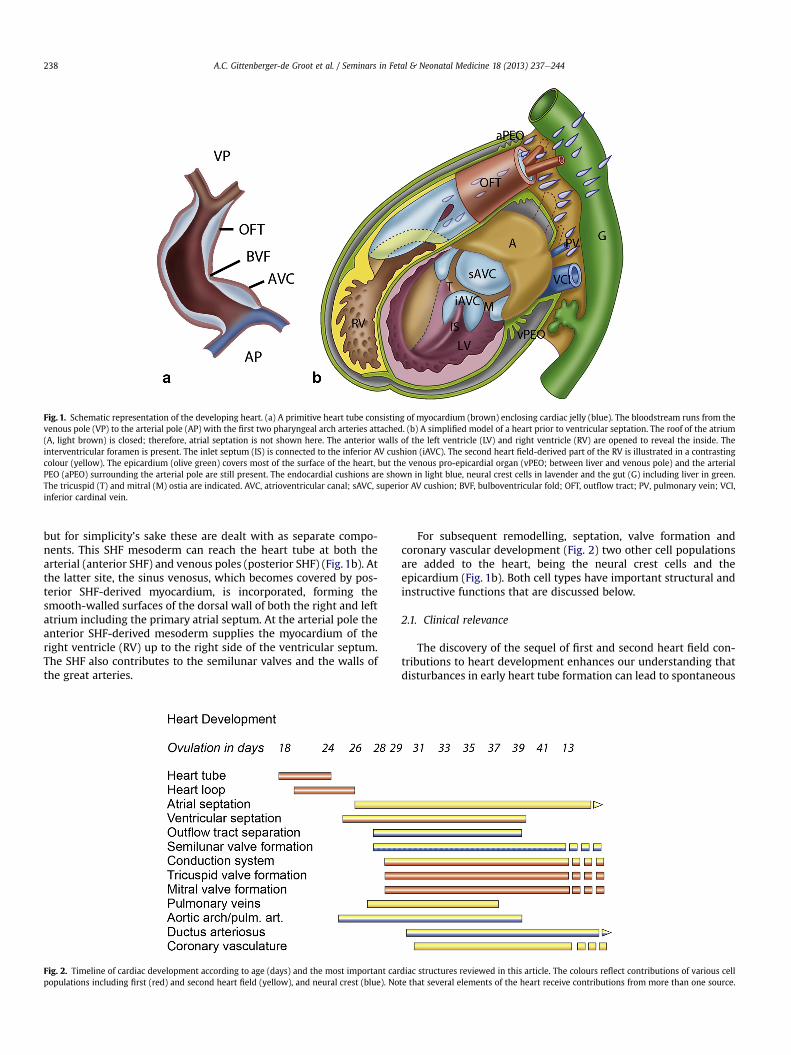
Fig. 1. Schematic representation of the developing heart. (a) A primitive heart tube consisting of myocardium (brown) enclosing cardiac jelly (blue). The bloodstream runs from thevenous pole (VP) to the arterial pole (AP) with the first two pharyngeal arch arteries attached. (b) A simplified model of a heart prior to ventricular septation. The roof of the atrium(A, light brown) is closed; therefore, atrial septation is not shown here. The anterior walls of the left ventricle (LV) and right ventricle (RV) are opened to reveal the inside. Theinterventricular foramen is present. The inlet septum (IS) is connected to the inferior AV cushion (iAVC). The second heart field-derived part of the RV is illustrated in a contrastingcolour (yellow). The epicardium (olive green) covers most of the surface of the heart, but the venous pro-epicardial organ (vPEO; between liver and venous pole) and the arterialPEO (aPEO) surrounding the arterial pole are still present. The endocardial cushions are shown in light blue, neural crest cells in lavender and the gut (G) including liver in green.The tricuspid (T) and mitral (M) ostia are indicated. AVC, atrioventricular canal; sAVC, superior AV cushion; BVF, bulboventricular fold; OFT, outflow tract; PV, pulmonary vein; VCI,inferior cardinal vein.
A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244238
but for simplicity’s sake these are dealt with as separate compo-nents. This SHF mesoderm can reach the heart tube at both thearterial (anterior SHF) and venous poles (posterior SHF) (Fig. 1b). Atthe latter site, the sinus venosus, which becomes covered by pos-terior SHF-derived myocardium, is incorporated, forming thesmooth-walled surfaces of the dorsal wall of both the right and leftatrium including the primary atrial septum. At the arterial pole theanterior SHF-derived mesoderm supplies the myocardium of theright ventricle (RV) up to the right side of the ventricular septum.The SHF also contributes to the semilunar valves and the walls ofthe great arteries.
Fig. 2. Timeline of cardiac development according to age (days) and the most important carpopulations including first (red) and second heart field (yellow), and neural crest (blue). No
For subsequent remodelling, septation, valve formation andcoronary vascular development (Fig. 2) two other cell populationsare added to the heart, being the neural crest cells and theepicardium (Fig. 1b). Both cell types have important structural andinstructive functions that are discussed below.
2.1. Clinical relevance
The discovery of the sequel of first and second heart field con-tributions to heart development enhances our understanding thatdisturbances in early heart tube formation can lead to spontaneous
diac structures reviewed in this article. The colours reflect contributions of various cellte that several elements of the heart receive contributions from more than one source.
A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244 239
abortion most probably as a result of deficient cardiac contraction.A cardiac cause for the reported 60e80% incidence of early em-bryonic demise is supported by transgenic mouse studies.2 Mostforms of CHD, both at the inflow (venous pole) and the outflow(arterial pole) of the heart concern defects in SHF addition.
This review focuses separately on normal and abnormal devel-opment of the cardiac outflow and inflow tract. The specificmodulating role of the neural crest cells will be included, and themany novel aspects of the role of the epicardium deserve specialattention.
3. Development of the cardiac outflow tract
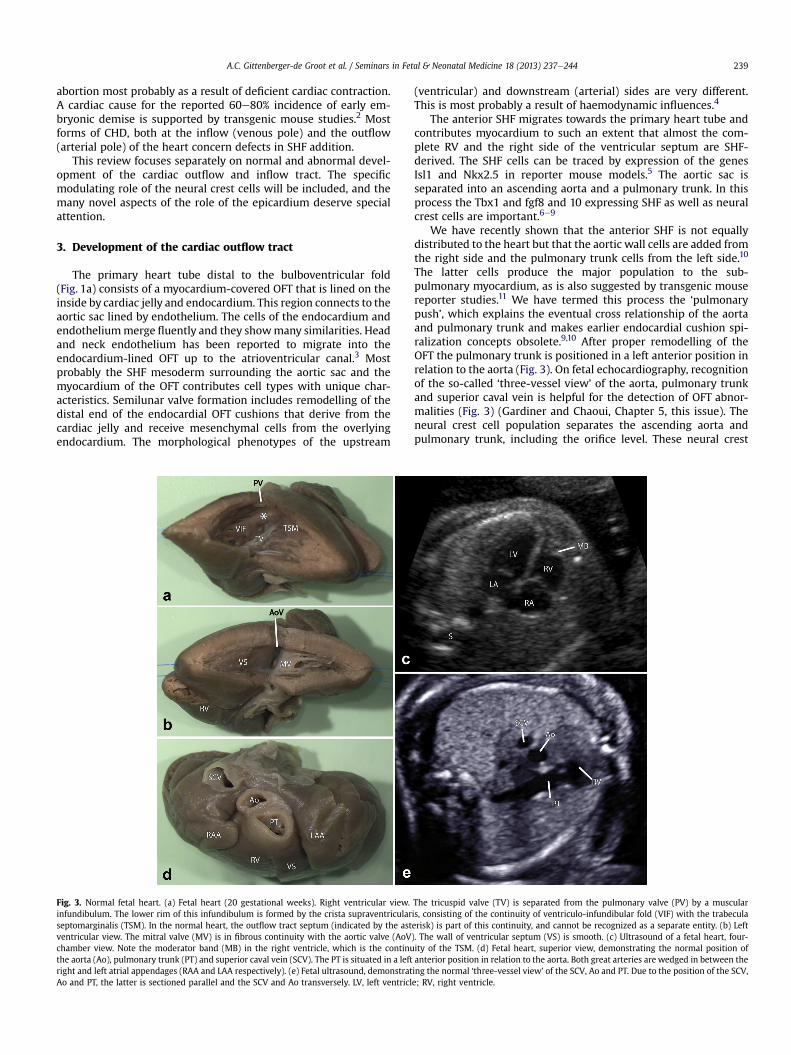
The primary heart tube distal to the bulboventricular fold(Fig. 1a) consists of a myocardium-covered OFT that is lined on theinside by cardiac jelly and endocardium. This region connects to theaortic sac lined by endothelium. The cells of the endocardium andendotheliummerge fluently and they showmany similarities. Headand neck endothelium has been reported to migrate into theendocardium-lined OFT up to the atrioventricular canal.3 Mostprobably the SHF mesoderm surrounding the aortic sac and themyocardium of the OFT contributes cell types with unique char-acteristics. Semilunar valve formation includes remodelling of thedistal end of the endocardial OFT cushions that derive from thecardiac jelly and receive mesenchymal cells from the overlyingendocardium. The morphological phenotypes of the upstream
Fig. 3. Normal fetal heart. (a) Fetal heart (20 gestational weeks). Right ventricular view.infundibulum. The lower rim of this infundibulum is formed by the crista supraventricularseptomarginalis (TSM). In the normal heart, the outflow tract septum (indicated by the astventricular view. The mitral valve (MV) is in fibrous continuity with the aortic valve (AoVchamber view. Note the moderator band (MB) in the right ventricle, which is the continuthe aorta (Ao), pulmonary trunk (PT) and superior caval vein (SCV). The PT is situated in a leftright and left atrial appendages (RAA and LAA respectively). (e) Fetal ultrasound, demonstratAo and PT, the latter is sectioned parallel and the SCV and Ao transversely. LV, left ventricl
(ventricular) and downstream (arterial) sides are very different.This is most probably a result of haemodynamic influences.4
The anterior SHF migrates towards the primary heart tube andcontributes myocardium to such an extent that almost the com-plete RV and the right side of the ventricular septum are SHF-derived. The SHF cells can be traced by expression of the genesIsl1 and Nkx2.5 in reporter mouse models.5 The aortic sac isseparated into an ascending aorta and a pulmonary trunk. In thisprocess the Tbx1 and fgf8 and 10 expressing SHF as well as neuralcrest cells are important.6e9
We have recently shown that the anterior SHF is not equallydistributed to the heart but that the aortic wall cells are added fromthe right side and the pulmonary trunk cells from the left side.10
The latter cells produce the major population to the sub-pulmonary myocardium, as is also suggested by transgenic mousereporter studies.11 We have termed this process the ‘pulmonarypush’, which explains the eventual cross relationship of the aortaand pulmonary trunk and makes earlier endocardial cushion spi-ralization concepts obsolete.9,10 After proper remodelling of theOFT the pulmonary trunk is positioned in a left anterior position inrelation to the aorta (Fig. 3). On fetal echocardiography, recognitionof the so-called ‘three-vessel view’ of the aorta, pulmonary trunkand superior caval vein is helpful for the detection of OFT abnor-malities (Fig. 3) (Gardiner and Chaoui, Chapter 5, this issue). Theneural crest cell population separates the ascending aorta andpulmonary trunk, including the orifice level. These neural crest
The tricuspid valve (TV) is separated from the pulmonary valve (PV) by a muscularis, consisting of the continuity of ventriculo-infundibular fold (VIF) with the trabeculaerisk) is part of this continuity, and cannot be recognized as a separate entity. (b) Left). The wall of ventricular septum (VS) is smooth. (c) Ultrasound of a fetal heart, four-ity of the TSM. (d) Fetal heart, superior view, demonstrating the normal position ofanterior position in relation to the aorta. Both great arteries are wedged in between theing the normal ‘three-vessel view’ of the SCV, Ao and PT. Due to the position of the SCV,e; RV, right ventricle.

A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244240
cells enter the heart from dorsal between the 4th and 6thpharyngeal arch artery, making the septation distance minimal,also because of the saddle-shaped arterial orifice level.9 The neuralcrest cells enter the lateral and septal endocardial OFT cushionswhere they initiate a process of myocardialization,7,12 resulting in amyocardial OFT septum. This septum is in effect very small in thenormal heart because of the low position of the aortic orifice but islocalized in the subpulmonary myocardium that links theventriculo-infundibular fold to the trabecula septomarginalis withthe crista supraventricularis as its lower rim (Fig. 3). The endocar-dial OFT cushions, that form the substrate for the OFT septum,merge with the atrioventricular endocardial cushions thus closingoff (by formation of the membranous septum) the interventricularforamen separating the right (RV) and left ventricular (LV) cavities.The ventricular septum consists of two other components, beingthe anteriorly located primary or trabecular septum and the pos-terior inlet septum. These have merged at the site of the trabeculaseptomarginalis.13 Other authors report that the main body of theventricular septum derives solely from the bulboventricular orprimary fold.14 New research, using both evolution and develop-ment (evo-devo) approaches as well as experimental avian andmouse studies, supports a multicomponent origin of the septum.Clinical implications are found in the understanding of varioustypes of ventricular septal defects (VSDs).
3.1. Clinical relevance
3.1.1. Ventricular septal defectsMany types of CHD are characterized by deficient OFT septation.
The most simple case shows a VSD based on a non-fusion of theendocardial OFT and atrioventricular cushions resulting in a mem-branous VSD. When the OFT septum deviates into the RV or LV weare confronted with malalignment defects.15e17 When endocardialcushion-derived mesenchymal (later on fibrous) tissue is found inthe free rim of a VSD, these are referred to as perimembranous VSDs,which constitute the majority of these defects (Fig. 4). Deficientfusion and/or compaction of the myocardial primary and inletseptum components result in central muscular and muscular VSDs.
3.1.2. Tetralogy of Fallot, transposition of the great arteries (TGA)and double outlet right ventricle (DORV)
In many instances malalignment of the OFT septum is also re-flected in the great arteries. This results in cases with tetralogy ofFallot with DORV in cases with extreme (>50%) overriding of theventricular septum. In TGA there is usually a right anterior positionof the aorta in relation to the pulmonary trunk (Fig. 5a,b). On fetalechocardiography the three-vessel view is abnormal and a parallelposition of the great arteries can be observed on the four-chamberview (Fig. 5c). TGA can occur without and with a VSD. In the lattercase the per definition subpulmonary VSD can be morphologically
Fig. 4. Perimembranous ventricular septal defect. (a). Fetal heart (29 gestational weeks), rigindicates a perimembranous ventricular septal defect. (c) Fetal echocardiogram, four-chambLA, left atrium; LV, left ventricle; PV, pulmonary valve; RA, right atrium; RV, right ventricle;asterisk, OFT septum.
>50% in the RV which results in a DORV.17 As mentioned above,DORV can also be found in cases with normally related great ar-teries with a subaortic VSD. This malformation, based on the degreeof overriding of the aortic orifice, merges fluently into a tetralogy ofFallot where there is a combination with a subpulmonary, valvularand/or pulmonary trunk stenosis. When the OFT septum is notformed, a common arterial trunk or persistent truncus arteriosusdevelops18,19 with the obligatory presence of a subarterial VSD.
3.1.3. Semilunar valve abnormalitiesAt the distal end of the two main endocardial OFT ridges in the
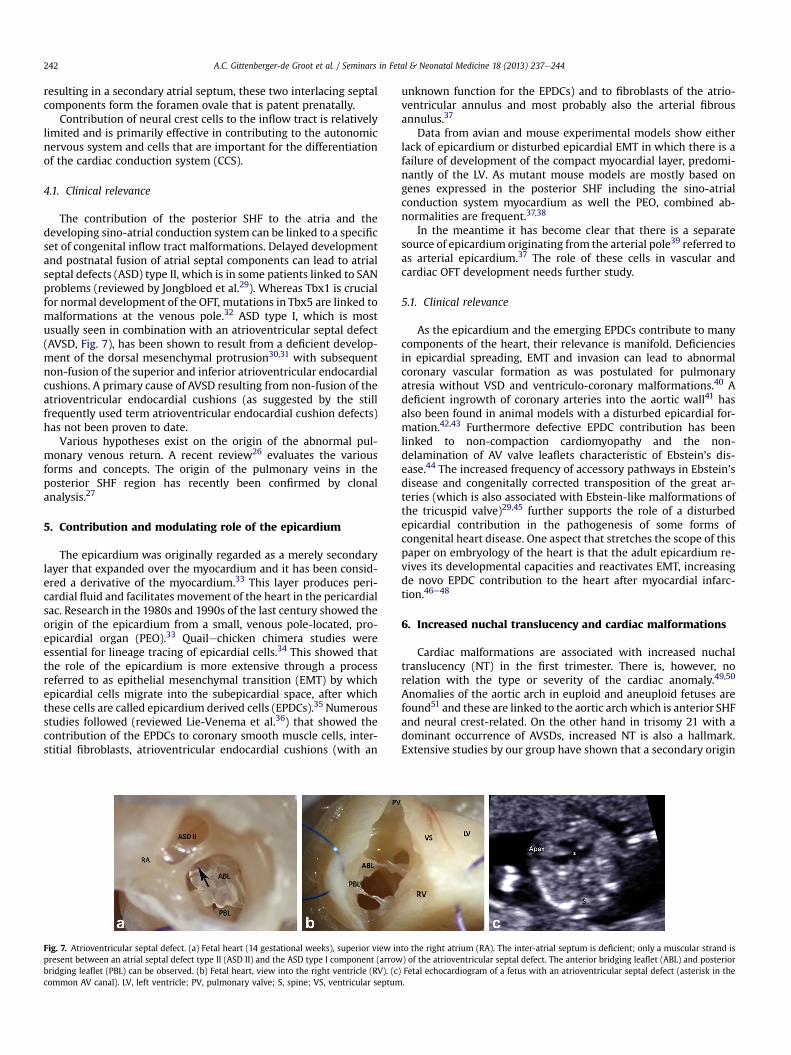
still unseptated arterial orifice level, two small intercalated cush-ions develop. During normal development, separation of the arte-rial orifice level results in three semilunar valve leaflets in the aorticand pulmonary orifices. Abnormal development includes both theoccurrence of deficient numbers as well as excessive numbers ofvalve leaflets (Fig. 6). The pathomorphology of the aortic and pul-monary valve leaflet abnormalities is different. Most attention,however, has been directed to the common bicuspid aortic valve(BAV, Fig. 6b) found as an isolated abnormality even in adults.Several gene mutations like Notch1 have now been reported in thehuman population.20 The exact cause of BAV has not been eluci-dated and it is likely that different aetiologies underlie differentBAV phenotypes. There are indications that the anterior SHF isinvolved and that not only the aortic valve is abnormal but the wallof the ascending aorta is also included, which has the tendency tobecome pathologically dilated at an early age.21
3.1.4. Aortic arch malformationsExamples of aortic arch malformations include right aortic arch,
interrupted left arch andhypoplasia of predominantly the B segment(4thpharyngeal archderivative).22 Inpatientswith a right aortic arch(Fig. 5def) the aortic arch is positioned to the right of the trachea. Incases of concomitant deficient development of the left 4th pharyn-geal arch artery, an aberrant (retro-oesophageal) position of the leftsubclavian artery results (Fig. 5d,e). The lattermay form the substrateof vascular compression of the trachea and/or oesophagus.
Aortic arch malformations are often linked to a deficient inter-action of neural crest cells and the anterior SHF. The best-knownmalformation in this respect is the 22q11 deletion syndromeevoked by a mutation of the Tbox1 gene, expressed in the SHF.6 Therelatively common preductal and paraductal coarctation in theisthmus (A segment) of the aortic arch ismost probably the result ofa haemodynamic disbalance with stenosis of the left ventricularoutflow tract.23
4. Development of the cardiac inflow tract
Based on mouse model studies the posterior SHF contributes tothe heart tube somewhat later compared to the anterior SHF
ht ventricular view and (b) fetal echocardiogram, five-chamber view. The white arrower view, showing the ventricular septal defect with colour Doppler imaging. Ao, aorta;TSM, trabecula septomarginalis; TV, tricuspid valve; VIF, ventriculo-infundibular fold;
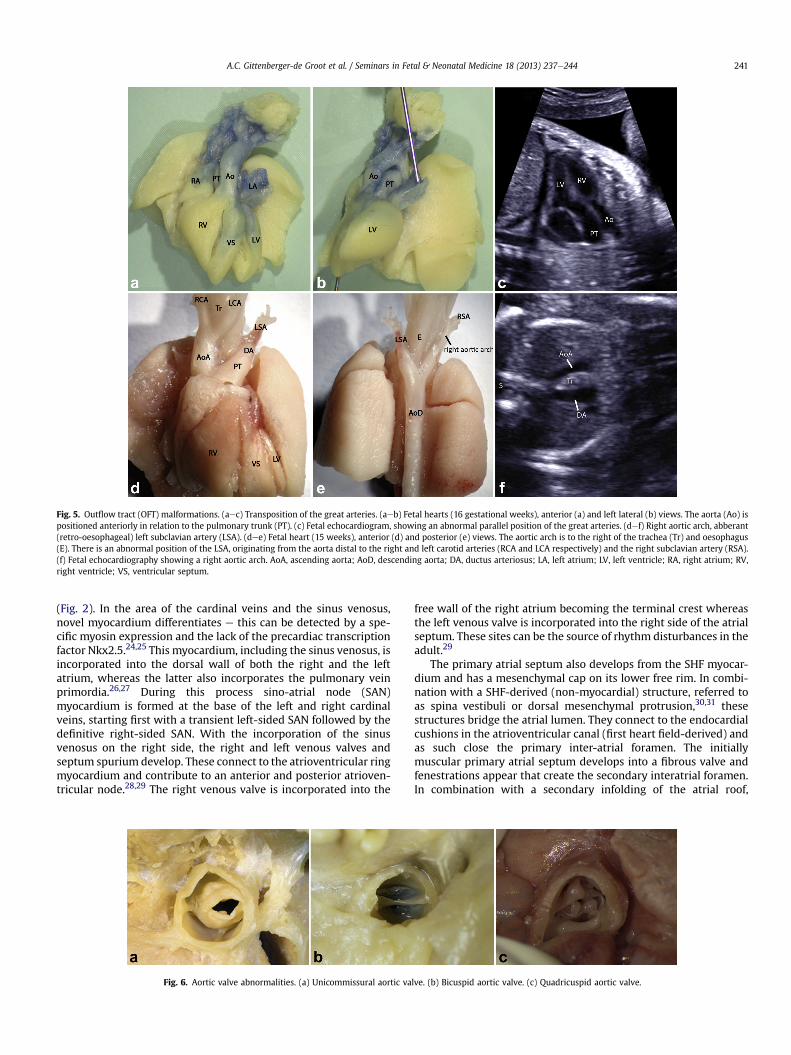
Fig. 5. Outflow tract (OFT) malformations. (aec) Transposition of the great arteries. (aeb) Fetal hearts (16 gestational weeks), anterior (a) and left lateral (b) views. The aorta (Ao) ispositioned anteriorly in relation to the pulmonary trunk (PT). (c) Fetal echocardiogram, showing an abnormal parallel position of the great arteries. (def) Right aortic arch, abberant(retro-oesophageal) left subclavian artery (LSA). (dee) Fetal heart (15 weeks), anterior (d) and posterior (e) views. The aortic arch is to the right of the trachea (Tr) and oesophagus(E). There is an abnormal position of the LSA, originating from the aorta distal to the right and left carotid arteries (RCA and LCA respectively) and the right subclavian artery (RSA).(f) Fetal echocardiography showing a right aortic arch. AoA, ascending aorta; AoD, descending aorta; DA, ductus arteriosus; LA, left atrium; LV, left ventricle; RA, right atrium; RV,right ventricle; VS, ventricular septum.
A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244 241
(Fig. 2). In the area of the cardinal veins and the sinus venosus,novel myocardium differentiates e this can be detected by a spe-cific myosin expression and the lack of the precardiac transcriptionfactor Nkx2.5.24,25 This myocardium, including the sinus venosus, isincorporated into the dorsal wall of both the right and the leftatrium, whereas the latter also incorporates the pulmonary veinprimordia.26,27 During this process sino-atrial node (SAN)myocardium is formed at the base of the left and right cardinalveins, starting first with a transient left-sided SAN followed by thedefinitive right-sided SAN. With the incorporation of the sinusvenosus on the right side, the right and left venous valves andseptum spurium develop. These connect to the atrioventricular ringmyocardium and contribute to an anterior and posterior atrioven-tricular node.28,29 The right venous valve is incorporated into the
Fig. 6. Aortic valve abnormalities. (a) Unicommissural aortic va
free wall of the right atrium becoming the terminal crest whereasthe left venous valve is incorporated into the right side of the atrialseptum. These sites can be the source of rhythm disturbances in theadult.29
The primary atrial septum also develops from the SHF myocar-dium and has a mesenchymal cap on its lower free rim. In combi-nation with a SHF-derived (non-myocardial) structure, referred toas spina vestibuli or dorsal mesenchymal protrusion,30,31 thesestructures bridge the atrial lumen. They connect to the endocardialcushions in the atrioventricular canal (first heart field-derived) andas such close the primary inter-atrial foramen. The initiallymuscular primary atrial septum develops into a fibrous valve andfenestrations appear that create the secondary interatrial foramen.In combination with a secondary infolding of the atrial roof,
lve. (b) Bicuspid aortic valve. (c) Quadricuspid aortic valve.
A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244242
resulting in a secondary atrial septum, these two interlacing septalcomponents form the foramen ovale that is patent prenatally.
Contribution of neural crest cells to the inflow tract is relativelylimited and is primarily effective in contributing to the autonomicnervous system and cells that are important for the differentiationof the cardiac conduction system (CCS).
4.1. Clinical relevance
The contribution of the posterior SHF to the atria and thedeveloping sino-atrial conduction system can be linked to a specificset of congenital inflow tract malformations. Delayed developmentand postnatal fusion of atrial septal components can lead to atrialseptal defects (ASD) type II, which is in some patients linked to SANproblems (reviewed by Jongbloed et al.29). Whereas Tbx1 is crucialfor normal development of the OFT, mutations in Tbx5 are linked tomalformations at the venous pole.32 ASD type I, which is mostusually seen in combination with an atrioventricular septal defect(AVSD, Fig. 7), has been shown to result from a deficient develop-ment of the dorsal mesenchymal protrusion30,31 with subsequentnon-fusion of the superior and inferior atrioventricular endocardialcushions. A primary cause of AVSD resulting from non-fusion of theatrioventricular endocardial cushions (as suggested by the stillfrequently used term atrioventricular endocardial cushion defects)has not been proven to date.
Various hypotheses exist on the origin of the abnormal pul-monary venous return. A recent review26 evaluates the variousforms and concepts. The origin of the pulmonary veins in theposterior SHF region has recently been confirmed by clonalanalysis.27
5. Contribution and modulating role of the epicardium
The epicardium was originally regarded as a merely secondarylayer that expanded over the myocardium and it has been consid-ered a derivative of the myocardium.33 This layer produces peri-cardial fluid and facilitates movement of the heart in the pericardialsac. Research in the 1980s and 1990s of the last century showed theorigin of the epicardium from a small, venous pole-located, pro-epicardial organ (PEO).33 Quailechicken chimera studies wereessential for lineage tracing of epicardial cells.34 This showed thatthe role of the epicardium is more extensive through a processreferred to as epithelial mesenchymal transition (EMT) by whichepicardial cells migrate into the subepicardial space, after whichthese cells are called epicardium derived cells (EPDCs).35 Numerousstudies followed (reviewed Lie-Venema et al.36) that showed thecontribution of the EPDCs to coronary smooth muscle cells, inter-stitial fibroblasts, atrioventricular endocardial cushions (with an
Fig. 7. Atrioventricular septal defect. (a) Fetal heart (14 gestational weeks), superior view inpresent between an atrial septal defect type II (ASD II) and the ASD type I component (arrowbridging leaflet (PBL) can be observed. (b) Fetal heart, view into the right ventricle (RV). (ccommon AV canal). LV, left ventricle; PV, pulmonary valve; S, spine; VS, ventricular septum
unknown function for the EPDCs) and to fibroblasts of the atrio-ventricular annulus and most probably also the arterial fibrousannulus.37
Data from avian and mouse experimental models show eitherlack of epicardium or disturbed epicardial EMT in which there is afailure of development of the compact myocardial layer, predomi-nantly of the LV. As mutant mouse models are mostly based ongenes expressed in the posterior SHF including the sino-atrialconduction system myocardium as well the PEO, combined ab-normalities are frequent.37,38
In the meantime it has become clear that there is a separatesource of epicardium originating from the arterial pole39 referred toas arterial epicardium.37 The role of these cells in vascular andcardiac OFT development needs further study.
5.1. Clinical relevance
As the epicardium and the emerging EPDCs contribute to manycomponents of the heart, their relevance is manifold. Deficienciesin epicardial spreading, EMT and invasion can lead to abnormalcoronary vascular formation as was postulated for pulmonaryatresia without VSD and ventriculo-coronary malformations.40 Adeficient ingrowth of coronary arteries into the aortic wall41 hasalso been found in animal models with a disturbed epicardial for-mation.42,43 Furthermore defective EPDC contribution has beenlinked to non-compaction cardiomyopathy and the non-delamination of AV valve leaflets characteristic of Ebstein’s dis-ease.44 The increased frequency of accessory pathways in Ebstein’sdisease and congenitally corrected transposition of the great ar-teries (which is also associated with Ebstein-like malformations ofthe tricuspid valve)29,45 further supports the role of a disturbedepicardial contribution in the pathogenesis of some forms ofcongenital heart disease. One aspect that stretches the scope of thispaper on embryology of the heart is that the adult epicardium re-vives its developmental capacities and reactivates EMT, increasingde novo EPDC contribution to the heart after myocardial infarc-tion.46e48
6. Increased nuchal translucency and cardiac malformations
Cardiac malformations are associated with increased nuchaltranslucency (NT) in the first trimester. There is, however, norelation with the type or severity of the cardiac anomaly.49,50
Anomalies of the aortic arch in euploid and aneuploid fetuses arefound51 and these are linked to the aortic archwhich is anterior SHFand neural crest-related. On the other hand in trisomy 21 with adominant occurrence of AVSDs, increased NT is also a hallmark.Extensive studies by our group have shown that a secondary origin
to the right atrium (RA). The inter-atrial septum is deficient; only a muscular strand is) of the atrioventricular septal defect. The anterior bridging leaflet (ABL) and posterior
) Fetal echocardiogram of a fetus with an atrioventricular septal defect (asterisk in the.

A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244 243
of the increased oedema in the neck region is most probably not asecondary effect of haemodynamically linked cardiac problems.This is supported by the fact that increased NT is also found ineuploid and aneuploid fetuses without heart defect. It is mostprobable that we are dealing with a combination of anterior SHFand neural crest problems that reflect on both the vascular (specificlymphatic) differentiation in the neck region as well as on thedeveloping pharyngeal arch system. For the nuchal region we haveshown a marked enlargement of the transient bilateral jugularlymphatic sacs and a concurrent tissue oedema in the (mid) nuchalregion, with abnormal lymphatic endothelial cell differentiation.During subsequent lymph node formation the jugular lymphaticsacs disappear and the oedema dissolves. The latter is not the casein Turner syndrome where the lymphatic cysts e sometimesextremely large e are greatly distended tissue cavities with adeficient lymphatic development.52 In this respect the NT seen inNoonan syndrome resembles most what is found in trisomy 21. Thepossibly common morphogenetic pathways underlying the corre-lation between increased NT and CHD still need further study.
7. Conclusion
The many transgenic, mostly mouse, models reveal new andinteresting data on cardiac development. While the study of mu-tations in human genes has not opened new avenues, the attentionfor epigenetic factors needs further exploration.53 Also the linkbetween minor developmental deficiencies e for instance ofvascular smooth muscle and conduction system differentiation e
may yield insights into the developmental background of adultdiseases such as atherosclerosis, rhythm disturbances and diabetesmellitus-linked cardiac diseases.
Conflict of interest statement
None declared.
Funding sources
None.
References
1. Kelly RG. Molecular inroads into the anterior heart field. Trends Cardiovasc Med2005;15:51e6.
2. Brandon EP, Idzerda RL, McKnight GS. Targeting the mouse genome: a com-pendium of knockouts (part I). Curr Biol 1995;5:625e34.
3. Noden DM. Origins and patterning of avian outflow tract endocardial tissuesand cushion mesenchyme. Anat Rec 1990;226:72Ae3A.
4. Poelmann RE, Gittenberger-de Groot AC, Groenendijk BCW, et al. Shear stressin the developing cardiovascular system. In: Artman M, Benson DW,Srivastava D, Nakazawa M, editors. Cardiovascular development and congenitalmalformations: molecular and genetic mechanisms. Massachusetts: BlackwellFutura; 2005. p. 169e73.
5. Cai CL, Liang X, Shi Y, et al. Isl1 identifies a cardiac progenitor population thatproliferates prior to differentiation and contributes a majority of cells to theheart. Dev Cell 2003;5:877e89.
6. Lindsay EA, Vitelli F, Su H, et al. Tbx1 haploinsufficiency in the DiGeorge syn-drome region causes aortic arch defects in mice. Nature 2001;410:97e101.
7. van den Hoff MJ, Moorman AF, Ruijter JM, et al. Myocardialization of the car-diac outflow tract. Dev Biol 1999;212:477e90.
8. Waldo KL, Hutson MR, Stadt HA, Zdanowicz M, Zdanowicz J, Kirby ML. Cardiacneural crest is necessary for normal addition of the myocardium to the arterialpole from the secondary heart field. Dev Biol 2005;281:66e77.
9. Gittenberger-de Groot AC, Bartelings MM, DeRuiter MC, Poelmann RE. Basics ofcardiac development for the understanding of congenital heart malformations.Pediatr Res 2005;57:169e76.
10. Scherptong RW, Jongbloed MR, Wisse LJ, et al. Morphogenesis of outflow tractrotation during cardiac development: the pulmonary push concept. Dev Dyn2012;241:1413e22.
11. Bajolle F, Zaffran S, Meilhac SM, et al. Myocardium at the base of the aorta andpulmonary trunk is prefigured in the outflow tract of the heart and in sub-domains of the second heart field. Dev Biol 2008;313:25e34.
12. Poelmann RE, Mikawa T, Gittenberger-de Groot AC. Neural crest cells inoutflow tract septation of the embryonic chicken heart: differentiation andapoptosis. Dev Dyn 1998;212:373e84.
13. Wenink ACG. Embryology of the ventricular septum. Separate origin of itscomponents. Virchows Arch 1981;390:71e9.
14. Lamers WH, Wessels A, Verbeek FJ, et al. New findings concerning ventricularseptation in the human heart. Implications for maldevelopment. Circulation1992;86:1194e205.
15. Bartelings MM, Gittenberger-de Groot AC. The outflow tract of the heart eembryologic and morphologic correlations. Int J Cardiol 1989;22:289e300.
16. Bartelings MM, Gittenberger-de Groot AC. Morphogenetic considerations oncongenital malformations of the outflow tract. Part 2: complete trans-position of the great arteries and double outlet right ventricle. Int J Cardiol1991;33:5e26.
17. Bartelings MM, Gittenberger-de Groot AC. Morphogenetic considerations oncongenital malformations of the outflow tract. Part 1: common arterial trunkand tetralogy of Fallot. Int J Cardiol 1991;32:213e30.
18. Gittenberger-de Groot AC, Bartelings MM, Bogers AJJC, Boot MJ, Poelmann RE.The embryology of the common arterial trunk. Prog Pediatr Cardiol 2002;15:1e8.
19. Jacobs ML, Anderson RH. Rationalising the nomenclature of common arterialtrunk. Cardiol Young 2012;22:639e46.
20. Garg V, Muth AN, Ransom JF, et al. Mutations in NOTCH1 cause aortic valvedisease. Nature 2005;437:270e4.
21. Matthias Bechtel JF, Noack F, Sayk F, Erasmi AW, Bartels C, Sievers HH. Histo-pathological grading of ascending aortic aneurysm: comparison of patientswith bicuspid versus tricuspid aortic valve. J Heart Valve Dis 2003;12:54e9.
22. Gittenberger-De Groot AC, Jongbloed MRM, Poelmann RE. Normal andabnormal cardiac development. In: Moller JH, Hoffman JI, editors. Pediatriccardiovascular medicine. 2nd ed. Oxford: Blackwell; 2012. p. 1e22.
23. Moene RJ, Oppenheimer-Dekker A, Moulaert AJ. The concurrence of dimen-sional aortic arch anomalies and abnormal left ventricular muscle bundles.Pediatr Cardiol 1982;2:107e14.
24. Gittenberger-de Groot AC, Mahtab EAF, Hahurij ND, et al. Nkx2.5 negativemyocardium of the posterior heart field and its correlation with podoplaninexpression in cells from the developing cardiac pacemaking and conductionsystem. Anat Rec 2007;290:115e22.
25. Christoffels VM, Mommersteeg MT, Trowe MO, et al. Formation of the venouspole of the heart from an Nkx2-5-negative precursor population requiresTbx18. Circ Res 2006;98:1555e63.
26. Douglas YL, Jongbloed MR, DeRuiter MC, Gittenberger-de Groot AC. Normaland abnormal development of pulmonary veins: state of the art and correlationwith clinical entities. Int J Cardiol 2011;147:13e24.
27. Lescroart F, Mohun T, Meilhac SM, Bennett M, Buckingham M. Lineage tree forthe venous pole of the heart: clonal analysis clarifies controversial genealogybased on genetic tracing. Circ Res 2012;111:1313e22.
28. Blom NA, Gittenberger-de Groot AC, DeRuiter MC, Poelmann RE, Mentink MM,Ottenkamp J. Development of the cardiac conduction tissue in human embryosusing HNK-1 antigen expression: possible relevance for understanding ofabnormal atrial automaticity. Circulation 1999;99:800e6.
29. Jongbloed MR, Vicente SR, Hahurij ND, et al. Normal and abnormal develop-ment of the cardiac conduction system; implications for conduction andrhythm disorders in the child and adult. Differentiation 2012;84:131e48.
30. Blom NA, Ottenkamp J, Wenink AG, Gittenberger-de Groot AC. Deficiency ofthe vestibular spine in atrioventricular septal defects in human fetuses withdown syndrome. Am J Cardiol 2003;91:180e4.
31. Snarr BS, Wirrig EE, Phelps AL, Trusk TC, Wessels A. A spatiotemporal evalu-ation of the contribution of the dorsal mesenchymal protrusion to cardiacdevelopment. Dev Dyn 2007;236:1287e94.
32. Moskowitz IP, Kim JB, Moore ML, et al. A molecular pathway including Id2,Tbx5, and Nkx2-5 required for cardiac conduction system development. Cell2007;129:1365e76.
33. Viragh S, Challice CE. The origin of the epicardium and the embryonicmyocardial circulation in the mouse. Anat Rec 1981;201:157e68.
34. Poelmann RE, Gittenberger-de Groot AC, Mentink MMT, Bökenkamp R,Hogers B. Development of the cardiac coronary vascular endothelium, studiedwith antiendothelial antibodies, in chicken-quail chimeras. Circ Res 1993;73:559e68.
35. Gittenberger-de Groot AC, Vrancken Peeters M-PFM, Mentink MMT,Gourdie RG, Poelmann RE. Epicardium-derived cells contribute a novel popu-lation to the myocardial wall and the atrioventricular cushions. Circ Res1998;82:1043e52.
36. Lie-Venema H, van den Akker NMS, Bax NAM, et al. Origin, fate, and function ofepicardium-derived cells (EPDCs) in normal and abnormal cardiac develop-ment. ScientificWorldJournal 2007;7:1777e98.
37. Gittenberger-de Groot AC, Winter EM, Bartelings MM, Goumans MJ,DeRuiter MC, Poelmann RE. The arterial and cardiac epicardium in develop-ment, disease and repair. Differentiation 2012;84:41e53.
38. van WB, van den Hoff M. Epicardium and myocardium originate from a com-mon cardiogenic precursor pool. Trends Cardiovasc Med 2010;20:1e7.
39. Perez-Pomares JM, Phelps A, Sedmerova M, Wessels A. Epicardial-like cells onthe distal arterial end of the cardiac outflow tract do not derive from theproepicardium but are derivatives of the cephalic pericardium. Dev Dyn2003;227:56e68.
40. Gittenberger-de Groot AC, Tennstedt C, Chaoui R, Lie-Venema H, Sauer U,Poelmann RE. Ventriculo coronary arterial communications (VCAC) and

A.C. Gittenberger-de Groot et al. / Seminars in Fetal & Neonatal Medicine 18 (2013) 237e244244
myocardial sinusoids in hearts with pulmonary atresia with intact ventricularseptum: two different diseases. Prog Pediatr Cardiol 2001;13:157e64.
41. Bogers AJJC, Gittenberger-de Groot AC, Poelmann RE, Péault BM, Huysmans HA.Development of the origin of the coronary arteries, a matter of ingrowth oroutgrowth? Anat Embryol 1989;180:437e41.
42. Eralp I, Lie-Venema H, DeRuiter MC, et al. Coronary artery and orifice devel-opment is associated with proper timing of epicardial outgrowth and corre-lated Fas ligand associated apoptosis patterns. Circ Res 2005;96:526e34.
43. Lie-Venema H, Gittenberger-de Groot AC, van Empel LJP, et al. Ets-1 and Ets-2transcription factors are essential for normal coronary and myocardial devel-opment in chicken embryos. Circ Res 2003;92:749e56.
44. Lie-Venema H, Eralp I, Markwald RR, et al. Periostin expression by epicardium-derived cells (EPDCs) is involved in the development of the atrioventricularvalves and fibrous heart skeleton. Differentiation 2008;76:809e19.
45. Van Hare GF. Radiofrequency ablation of accessory pathways associated withcongenital heart disease. Pacing Clin Electrophysiol 1997;20:2077e81.
46. Gittenberger-de Groot AC, Winter EM, Poelmann RE. Epicardium-derived cells(EPDCs) in development, cardiac disease and repair of ischemia. J Cell Mol Med2010;14:1056e60.
47. Limana F, Capogrossi MC, Germani A. The epicardium in cardiac repair: fromthe stem cell view. Pharmacol Ther 2011;129:82e96.
48. Zhou B, Pu WT. Epicardial epithelial-to-mesenchymal transition in injuredheart. J Cell Mol Med 2011;15:2781e3.
49. Haak MC, Bartelings MM, Gittenberger-de Groot AC, Van Vugt JM. Cardiacmalformations in first-trimester fetuses with increased nuchal translucency:ultrasound diagnosis and postmortem morphology. Ultrasound Obstet Gynecol2002;20:14e21.
50. Simpson JM, Sharland GK. Nuchal translucency and congenital heart defects:heart failure or not? Ultrasound Obstet Gynecol 2000;16:30e6.
51. Hyett JA, Perdu M, Sharland GK, Snijders RS, Nicolaides KH. Increased nuchaltranslucency at 10e14 weeks of gestation as a marker for major cardiac de-fects. Ultrasound Obstet Gynecol 1997;10:242e6.
52. Bekker MN, Van Den Akker NM, Bartelings MM, et al. Nuchal edema andvenous-lymphatic phenotype disturbance in human fetuses and mouse em-bryos with aneuploidy. J Soc Gynecol Investig 2006;13:209e16.
53. Steegers-Theunissen RP, Steegers EA. Nutrient-gene interactions in earlypregnancy: a vascular hypothesis. Eur J Obstet Gynecol Reprod Biol 2003;106:115e7.
Related Documents











