ELSEVIER PIi: S0006-3207(96)00019-5 Biological Conservation 78 (1996) 69-96 Copyright © 1996 Elsevier Science Limited Printed in Great Britain. All rights reserved 0006-3207/96/$15.00 +.00 GLOBAL PATTERNS IN THE ESTABLISHMENT AND DISTRIBUTION OF EXOTIC BIRDS Ted J. Case Department of Biology, 0116 University of California at San Diego, La Jolla, CA 92093, USA Abstract I use three separate data bases to examine recipient com- munity and site factors that might be influencing the establishment, persistence, and distribution of avian exotics. All in all, about half the variance between islands~regions in their numbers of successfully and unsuccessfully introduced species can be accounted for by recipient site-specific variables; the most important correlate of success is the number of native species extinctions over about the last 3000 years, which reflects the degree of human activity and habitat destruction and deterioration through intrusions of exotic predators, her- bivores, and parasites. Consequently, the number of exotic species gained is close to the number of species lost through extinction. Even after controlling for avian extinctions, island area correlates positively with intro- duced species number. Invasion success does not decline significantly with the richness of the native avifauna (after controlling for the effects of extinctions and island area) nor the variety of potential mammalian predators. The relative proportion of extinct native species across islands~regions is negatively correlated with area and positively correlated with introduced species number and the number of endemic species. A strong correlation exists between the number of successes and the number of failures, attesting to the role of persistent acclimatiza- tion societies in increasing species numbers despite high .failure rates. The relative success to failure rate increases with the number of extinct native species. The correlation between introductions and native extinctions seems to arise because native birds are usually more common, if not restricted, to native habitats while introduced birds are primary occupants of disturbed and open habitats. As more of an island's area is converted to urban, agricul- tural and disturbed habitats or altered through the intro- duction of herbivores and exotic predators, most natives lose good living space while most introduced birds, that frequent open and disturbed areas and have evolved in predator-rich areas, gain habitat. l find little support for the notion that rich avifaunas in themselves repel the establishment of avian invaders at the level of whole islands or archipelagoes. However, interactions between established exotics and natives may Correspondence to: T. J. Case e-mail: [email protected] be influencing habitat distributions of species in both sets within islands. In both man-made habitats and native forest habitats, exotic species number and the relative abundance of exotic birds is negatively related to the number of native species. After accounting for this local variation, exotic species number is positively related to exotic species number for the entire island~region. In local surveys the relative abundance of exotic birds com- pared to native birds is affected by habitat (non-native habitats have more exotics) and also by the numbers of species of exotics and natives on the island. The relative importance of biotic interactions like competition, appar- ent competition through differential disease transmission or susceptibility, and predation in shaping the abundance and habitat affinities of exotics and native species can be difficult to unravel when regional affects are so impor- tant. Copyright © 1996 Elsevier Science Limited Keywords: birds, extinction, exotic species, invasibility, habitat destruction. 69 INTRODUCTION For assorted reasons, some practical and some purely aesthetic, people have purposely introduced various plants and animals from one place to another across the globe. What factors determine whether a species will become established or not? What site properties determine whether an ecological system will accept or repel invaders? Given answers to these questions, how can we best manage ecological systems to avoid problems? One ecological issue affecting answers to these ques- tions is whether natural communities vary in their resis- tance to invasions because of intrinsic properties of the natural system. Elton (1958) suggested that species-poor communities, characteristic of islands or very disturbed habitats, were more susceptible to invasion than species- rich communities. More recent theoretical explorations of model communities lend some credence to this hypothesis (Case, 1991; Post & Pimm, 1983; Drake, 1983, 1988). The role of habitat disturbance in promoting biologi- cal invasions is well supported (Rejm~nek, 1989). But how would we test Elton's hypothesis regarding

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER P I i : S 0 0 0 6 - 3 2 0 7 ( 9 6 ) 0 0 0 1 9 - 5
Biological Conservation 78 (1996) 69-96 Copyright © 1996 Elsevier Science Limited
Printed in Great Britain. All rights reserved 0006-3207/96/$15.00 +.00
GLOBAL PATTERNS IN THE ESTABLISHMENT AND DISTRIBUTION OF EXOTIC BIRDS
Ted J. Case Department of Biology, 0116 University of California at San Diego, La Jolla, CA 92093, USA
Abstract I use three separate data bases to examine recipient com- munity and site factors that might be influencing the establishment, persistence, and distribution o f avian exotics. All in all, about half the variance between islands~regions in their numbers of successfully and unsuccessfully introduced species can be accounted for by recipient site-specific variables; the most important correlate of success is the number of native species extinctions over about the last 3000 years, which reflects the degree of human activity and habitat destruction and deterioration through intrusions of exotic predators, her- bivores, and parasites. Consequently, the number of exotic species gained is close to the number of species lost through extinction. Even after controlling for avian extinctions, island area correlates positively with intro- duced species number. Invasion success does not decline significantly with the richness of the native avifauna (after controlling for the effects of extinctions and island area) nor the variety of potential mammalian predators. The relative proportion of extinct native species across islands~regions is negatively correlated with area and positively correlated with introduced species number and the number of endemic species. A strong correlation exists between the number of successes and the number of failures, attesting to the role of persistent acclimatiza- tion societies in increasing species numbers despite high
.failure rates. The relative success to failure rate increases with the number of extinct native species. The correlation between introductions and native extinctions seems to arise because native birds are usually more common, i f not restricted, to native habitats while introduced birds are primary occupants of disturbed and open habitats. As more of an island's area is converted to urban, agricul- tural and disturbed habitats or altered through the intro- duction of herbivores and exotic predators, most natives lose good living space while most introduced birds, that frequent open and disturbed areas and have evolved in predator-rich areas, gain habitat.
l find little support for the notion that rich avifaunas in themselves repel the establishment of avian invaders at the level of whole islands or archipelagoes. However, interactions between established exotics and natives may
Correspondence to: T. J. Case e-mail: [email protected]
be influencing habitat distributions of species in both sets within islands. In both man-made habitats and native forest habitats, exotic species number and the relative abundance of exotic birds is negatively related to the number of native species. After accounting for this local variation, exotic species number is positively related to exotic species number for the entire island~region. In local surveys the relative abundance of exotic birds com- pared to native birds is affected by habitat (non-native habitats have more exotics) and also by the numbers of species of exotics and natives on the island. The relative importance of biotic interactions like competition, appar- ent competition through differential disease transmission or susceptibility, and predation in shaping the abundance and habitat affinities of exotics and native species can be difficult to unravel when regional affects are so impor- tant. Copyright © 1996 Elsevier Science Limited
Keywords: birds, extinction, exotic species, invasibility, habitat destruction.
69
INTRODUCTION
For assorted reasons, some practical and some purely aesthetic, people have purposely introduced various plants and animals from one place to another across the globe. What factors determine whether a species will become established or not? What site properties determine whether an ecological system will accept or repel invaders? Given answers to these questions, how can we best manage ecological systems to avoid problems?
One ecological issue affecting answers to these ques- tions is whether natural communities vary in their resis- tance to invasions because of intrinsic properties of the natural system. Elton (1958) suggested that species-poor communities, characteristic of islands or very disturbed habitats, were more susceptible to invasion than species- rich communities. More recent theoretical explorations of model communities lend some credence to this hypothesis (Case, 1991; Post & Pimm, 1983; Drake, 1983, 1988).
The role of habitat disturbance in promoting biologi- cal invasions is well supported (Rejm~nek, 1989). But how would we test Elton's hypothesis regarding

70 T.J . Case
species-richness experimentally? Ideally we would con- struct islands identical in all respects but differing in the richness of the resident fauna. We then would introduce the same set of species to each island, con- trolling for individual numbers released, sex ratio, physiological condition, and other variables that might influence reproductive and survival success. Since this is generally impossible except at the microcosm scale (Drake, 1991; Drake et al., 1993; Robinson & Dickerson, 1984), for real islands we must rely on interpreting the results of past historical introductions where many confounding factors, beyond the nature of the recipient community, can and do influence invasion success. With introductions there are often no controls and, like all historical work, we must often rely on the sometimes conflicting accounts described by different actors and observers, no longer living; but the vast number of introductions performed on literally thou- sands of different islands and mainlands worldwide offers an opportunity to attempt answers to the ques- tions raised in the opening paragraph.
The method of the introduction may influence its likelihood of success, e.g. how many individuals are released, and how they are transported and acclimated to the new location. Different species have been intro- duced to different places with different climates so species-specific autecological factors will also be important. Some species may be better adapted to physical and climatic features of the new environment and others may have behavioral and life-history attributes that enhance population growth when they are rare. We might get spurious associations between invasion resis- tance and native biodiversity because of confounding interactions with other variables. For example, the number of native species on an island generally increases with its area (Preston, 1962; MacArthur & Wilson, 1967). Yet island area directly can affect intro- duction success by potentially affecting mate-finding ability. A few individuals of a new exotic species released into a large area may have difficulty locating mates compared to the same number released into a smaller area. Without experimentally or statistically partialling out the possible affects of area per se on introduction success, we might incorrectly conclude that species-richness (which covaries with area) was explaining the results when, in this scenario, it was more directly caused by area per se.
Here I use three separate data bases to examine recipient community and site factors, including area, that might be influencing the establishment, persistence, and distribution of avian introductions. I have assembled species lists for a worldwide set of islands and two mainland locations (continental USA and Australia). I ask what features of the recipient location best account for across-site variation in the number of introduced species present in the existing avifaunas.
The second analysis deals with a smaller set of loca- tions for which information exists on the number of
failed as well as successful introduction attempts. For these locations, mostly places that had acclimatization societies in the 1800s, I ask what site and community- level factors contribute to establishment success com- pared to failure.
Finally, since competitive interactions between natives and exotics may potentially be important in shaping the proclivity of native species to enter human- modified habitats and exotic species to penetrate native habitats, I examine the habitat distributions of native and introduced birds in various habitats and locations in the Pacific region.
The present study is entirely devoted to identifying factors in the recipient location that potentially infl- uence the establishment, persistence, and spread of introduced bird species. It is also important to make comparisons between species: are the same species that are successful in one place successful in others? What biological attributes of species are associated with estab- lishment success? How similar must climates be in the old and new homes of species? I reserve these analyses for a later paper.
M E T H O D S
Native and introduced birds on islands Table 1 contains a list of locations with tallies for their numbers of native and introduced birds along with other geographic and biotic features of the location. References are given in Appendix 1. The general refer- ences at the bottom of Appendix 1 were used for nearly all locations. Islands were selected for inclusion in the analysis based on two criteria: a complete avifaunal list, and an attempt to balance representation across different regions of the globe (excluding polar regions).
The analysis is restricted to land and freshwater species (excluding shorebirds, waders, and strictly sea- going ducks) that maintain breeding populations in the locations (i.e. migrants are excluded). Table 1 is based on archipelagoes with some minor exceptions (see below). Using individual islands as separate data points (rather than lumping islands into archipelagoes) com- mits pseudoreplication since the native faunas and introduction histories of nearby islands are typically closely linked and birds introduced to one island often colonize nearby islands on their own (Long, 1981; Moulton & Pimm, 1983). However, a problem arises with this choice: a single point, e.g. Fiji, includes many islands; each of which may not have the full comple- ment of introduced species present in the entire archipelago nor the full set of native species. Conse- quently, one can imagine a situation wherein all the introduced species are confined to a single island within the archipelago that lacks any native species. In this case the two sets of species never see each other in the archipelago and potentially experience very different environments. However, this is rarely the case and I have attempted to sort out islands which have unique

Introductions and extinctions of birds 71
Table 1. Bird species numbers on various islands of the world and for Australia and the continental US (excludes Alaska) Taxonomy follows Sibley and Monroe (1990). References are in Appendix 1. See also notes to Table 2. An asterisk (*) denotes locations where Holocene bird fossils have not yet been discovered or examined. Missing values in the extinction column indicate an absence of information on both historical and prehistorical extinctions. See text for further details.
Island/mainland No. No. native No. native Area Max. elevation Human Mammalian introduced species species (km 2) (m) settlement predator
species extant extant extinct pattern category
Aldabra* 1 20 1 155 100 3 3.0 Andamans/Nicobars* 4 104 0 6462 735 4 4.0 Aruba* 2 32 175 500 4 3.0 Ascension 4 0 1 90 860 3 3.0 Aucklands* 9 11 2 606 610 3 1.5 Azores* 3 20 0 2304 2320 3 3.0 Bahamas 6 101 13 13,939 122 4 4-0 Balearic 1 119 3 5014 1445 4 5.0 Bermuda 7 9 7 54 73 3 3.0 Borneo* 4 386 0 758,870 3810 4 5.0 California Channel 4 56 2 904 753 4 4.0 Canaries 5 47 3 7273 3720 4 3.0 Cape Verde* 5 26 1 4033 2829 3 3.0 Chagos* 7 3 65 20 3 3.0 Chatham 14 14 21 973 213 4 3.0 Christmas (Indian Oc.)* 1 8 0 135 357 3 2.0 Cocos* 1 6 0 47 854 3 3.0 Comoros* 7 52 0 1958 2361 3 4.0 Cook Islands 3 11 10 240 652 4 3.0 Corsica/Sardinia 3 120 1 32,771 2711 4 5.0 Cozumel* 0 75 0 324 24 4 5.0 Curacao* 2 34 0 425 372 4 3.0 Easter 4 0 6 I 16 530 4 2.0 Falklands* 2 28 0 16,053 706 3 4.0 Fiji (mongoose) 8 46 7 15,921 1300 4 4-0 Fiji (non-mongoose) 9 51 3 2375 1200 4 3.0 Galapagos 2 36 1 7855 1524 3 3.0 Great Britain 10 147 5 160,952 1343 4 5.0 Guadaloupe, Mex.* 3 13 6 254 1402 3 1.5 Hawaii (Kauai) 27 18 14 1422 1598 4 3.0 Hawaii (mongoose) 47 27 66 12,136 4206 4 4-0 Henderson 0 4 3 37 33 2 1-0 Jamaica 5 88 6 11,424 2256 4 4-0 Java* 1 327 0 132,400 3676 4 5.0 Juan Fernandez* 2 8 0 164 1500 3 3.0 Kangaroo 15 78 1 3890 190 4 3.0 Kermadec 7 6 1 30 520 2 3.0 Lord Howe 9 6 9 13 853 3 3.0 Madagascar 3 168 13 587,000 2881 4 5-0 Guam (Marianas)* 7 12 7 541 406 4 3.0 Marquesas 5 11 9 910 1260 4 2.0 Mauritius 19 13 14 1865 826 3 4.0 New Caledonia 6 56 14 16,912 1815 4 3.0 New Guinea* 1 513 0 808,000 5000 4 4-0 New Zealand 41 52 36 266,800 3765 4 4-0 Norfolk 12 15 6 40 310 4 2.5 Palau* 4 31 0 440 240 4 3.0 Pemba* 1 67 1 984 85 4 4-0 Puerto Rico 23 82 15 8897 1350 4 4-0 Reunion 19 12 13 2512 3040 3 3.0 Revillagigedo* 3 13 2 233 1113 1 2.0 Rodriguez 7 4 10 109 396 3 4.0 Rottnest* 3 20 2 19 35 3 3-0 Rotuma (Fiji)* 1 16 0 45 300 4 3.0 Saint Helena 8 0 5 125 819 3 3.0 Samoa 3 33 2 3150 1858 4 3-0 Sea of Cortez (Ld.-bridge)* 2 34 0 1713 620 1 1.5 Seychelles (granitic)* 8 16 3 233 913 3 3.0 Societies 12 13 12 1550 2322 4 3.0 Solomons, Central 1 124 2 31,800 3100 4 4.0
(Contmue~

72 T. J. Case
Table 1. - - eoatd
Island/mainland No. No. native introduced species
species extant extant
No. native Area Max. elevation Human Mammalian species (km 2) (m) settlement predator extinct pattern category
Sri Lanka* 1 225 Taiwan* 2 132 Tasmania 13 104 Three Kings* 9 10 Tres Marias* 1 34 Trinidad* 2 235 Tristan de Cunha Arch.* 0 6 Tuamotu* 2 9 Vanuatu* 5 56 Wake* 1 0 Zanzibar* 4 102
USA (lower 48 states) 13 553 Australia 17 466
1 65,610 2528 4 5.0 0 36,125 3998 4 5.0 1 67,900 1520 4 4.0 4 7 300 4 1-0 0 350 613 3 4.0
4542 1250 4 5.0 1 259 2329 3 3.0 0 855 113 4 3.0 0 12,000 1889 4 3-0 1 23 6 4 3.0 0 1658 125 4 5.0
6 7,827,622 4516 4 5.0 1 7,680,000 2228 4 5-0
histories compared to others in the same archipelago. For example, Guam has had many more introductions and extinctions than elsewhere in the Marianas archipelago; consequently, Guam is considered alone.
Since I will be examining the role of community and site factors, I subdivide archipelagoes when they are strongly subdivided by distance or important biological factors that might influence the success of introduced species. Fiji and Hawaii are subdivided into those islands with and without introduced mongoose.
The definition of an archipelago usually follows geo- political boundaries, with a few exceptions. Islands are lumped when the islands share endemic species, are similar in geological age of formation or isolation, and distance to colonizing sources. Islands whose distance apart from one another is greater than the distance to the mainland or other major faunal sources are not lumped. By these conventions, 'New Zealand' includes only the two major islands, not the many small off- lying land bridge islands; similarly I do not include in Australia or New Guinea the several off-shore islands. Corsica and Sardinia, which are similar in size and were united in the last glaciation, are combined. The Andamans are combined with the nearby Nicobars. On the other hand, the Mascarenes (Reunion, Rodriguez and Mauritius) are considered separately because they share relatively few native land bird species or exotic predators, and they are about as far apart from one another as they are to other major colonization sources. The distant Rotuma island (politically part of Fiji) is considered separately from the rest of Fiji.
Introduced species number This includes human releases and self-introduced species naturally invading the location during the last hundred years if they survive to the present day; self- introductions usually represent a small proportion of the 'exotic' bird species. Self-introduced species that replace extirpated native populations of the same
species are not counted. Most of the purposeful intro- ductions were done in the heyday of acclimatization societies during the mid 1800s although in Hawaii they continued much longer. A successfully introduced species is one that has established a breeding popula- tion that is still present today; its population must have increased numerically and expanded geographically beyond the vicinity of the original introduction sites. Aboriginal introductions (usually just Gallus gallus) are inferred based on a lack of subspecific differentiation, anthrophilic habits of the species, and no appearance in middens or subfossil deposits dated earlier than man's appearance on the island group.
Native species number This count excludes self-introduced land and freshwater species and native species that are now extinct. Here and for historical extinctions I follow the taxonomy in Sibly and Monroe (1990). For island archipelagoes like the Canaries, the number of native species is totaled over all islands in the group but no species is counted more than once.
Extinct bird species The total number of land and freshwater bird species that became extinct in historical and in prehistorical time is tallied. The prehistoric extinctions coincide for the most part with aboriginal occupation (Cassels, 1984; Diamond, 1984; Olson & James, 1984; Olson, 1989; Steadman, 1989b) and generally are late Holocene. Extinctions include true extinctions of a species and extirpations of the species from the island/archipelago in question. If the same species becomes extinct locally on several islands within a listed archipelago, it is counted as only a single extinc- tion and then only if no populations survive anywhere in the listed location (e.g. all islands in the Canaries, all mongoose-free islands in Fiji minus Rotuma, both major New Zealand islands).

Introductions and extinctions o f birds 73
Since some island locations have had much more paleontological attention than others, or have geologi- cal structures that better preserve fossils, it is probable that many undocumented prehistoric extinctions await discovery (see Pimm et al., 1994 for a clever way to estimate these 'missing' extinctions), but I see no rea- son why there should be any particular bias in fossil discovery with respect to numbers and type of intro- duced species established. Locations without any known prehistoric bird fossils are asterisked in Table 1. Missing values in the extinction column are for islands where I could not find reliable information for both historical and prehistorical extinctions.
Island area The total area of all islands in the archipelago or labeled island group is summed. It would also be nice to have a measure of the area still in virgin native habi- tat. However in practice such measures are inherently subjective since even apparently pristine habitats may be altered in subtle ways through human activities and introduced mammals, insects, and plants, the creation of edge effects (Diamond & Veitch, 1981; Simberloff, 1990; Bolger et al., 1991). Different bird species will respond to these local and landscape alterations in different ways, so a meaningful number that represents that habitat remaining which is acceptable for all native birds is illusive. As a surrogate of overall human impact to bird habitats, the number of avian extinctions may be a more appropriate 'bio-assay' (see below).
Maximum elevation above sea level This measure serves as a useful surrogate of habitat diversity (MacArthur & Wilson, 1967).
Human occupation h&tory Some places may lack avian introductions simply because people have not been present or interested in introducing them. The locations in Table 1 are catego- rized as follows: 1, locations with no permanent human settlements or settlements only very recently established and still localized; 2, Locations with aboriginal settle- ment but no subsequent lasting European settlement; 3, Locations with no aboriginal settlement prior to colo- nization and settlement by Europeans; 4, Locations with both an aboriginal and European colonial period of settlement.
Mammalian predators A rich native avifauna fauna will often be associated with a rich fauna of other taxa that might prey upon birds. Islands are categorized according to the variety of their native and exotic mammalian predators as follows: (0) none; (1) Polynesian rat Rattus exulans or other native rats but no introduced rats (R. rattus or R. norvegicus) or other mammalian predators; (1.5) feral domestic cats but no rats; (2) Rattus rattus and/or
R. norvegicus but no feral cats or other predators; (3) introduced rats and feral cats with or without native rats and feral domestic dogs (but no other predators); (4) introduced rats, plus feral cats and dogs, plus a mongoose, and/or a stoat or weasel, or other wild mustelid, viverid, canid, raccoon, or partially carnivo- rous (of birds or their eggs) primate; (5) a rich main- land fauna that includes rats, feral cats, and dogs plus several native carnivore species and other potential avian predators such as primates or large insectivores.
I test Elton's (1958) hypothesis by determining if a negative relationship exists between the number of native species of birds or varieties of mammalian predators and the number of successfully established introductions after partialling out variation that may be due to other variables (like island area, or settlement history). Since almost universally the number of native species on islands increases with their area and habitat diversity, and since a priori we might expect this same trend in introduced species, the numbers of both species in both sets might be positively associated across islands because they co-vary with island area and maximum elevation. To test Elton's hypothesis we need to partial out the expected effects of area and elevation (habitat diversity) of introduced and native species number, so that we can focus on these biotic variables. To do this I first regress introduced species number versus island area and maximum elevation and save the residuals, which represent relative introduced species richness after accounting for these geographic covariates. This is repeated for native species richness. A stepwise regression is performed using the residuals of introduced species number as the dependent variable regressed in a step-wise manner against native species number residuals, mammal categories, human occupa- tion categories, and the number of extinct native species as independent variables, I do not partial-out variation due to differences between islands' isolation distances, because isolation does not significantly affect introduced species number and its affect on native species richness, while highly significant, is immaterial to the hypothesis.
Establishment success and failure Table 2 contains a subset of locations for which rela- tively reliable records exist on introduction attempts. While the compilation of Long (1981) serves as an excellent starting point, I have found a number of instances where introduced species exist or failed attempts have occurred that are not mentioned in Long (1981). Also problematic, is that Long's criterion of 'success' is different from mine. Moreover, in a sub- stantial number of cases, the outcome of the intro- duction is listed in Long (1981) as uncertain (his 'possi- bly successful introductions' category). Therefore the locations included in my Table 2 are those where I have independently been able to assess successes and failures.

74 T. J. Case
=
-~,.2 ~ . ~
~1 .
~ o
N
~ g
0
0
N
©
g ~
0 ~
Z
{9
6 Z
0
c~ o 0
~J
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
. . ~, = ~ o o.~ ~ o~ -~
0
n3 .o
~ g
~ o ~
~ . ~ ~j o
> , ~ 0 = ~ . ~ ~ ) " ~ = - ~ . 0 0 ~=
o ~ ' ~ _ ~ o • ~ E::
~ ~, . ~ 1::~ 0 o o
~ .~ ~.~-
• = ~ o ~
~ ~ ~ ~ ~ .=
~ ~ ~ " ~ . ~ ..~
~ . o ~ ~ . ~ ~ . ~ [ _
-'~ ~ ~ 0 0 ~.=_~ =~ ~ ~~ , ~ ' = = = E " ~ L"-~ ' -
' ,~ ~ 0 ~ ~ ,

Introductions and extinctions o f birds 75
The success 'rate' is calculated on a per-species basis: the number of successfully introduced species to a loca- tion is divided by the total number of species whose introduction was attempted. In calculating success ratios, the problem arises as to what constitutes an 'attempt'. I do not count attempts that involve releases of only one individual of a species or multiple individuals but all of the same sex, or migratory species. An attempt is defined as the release or self- colonization of a land or freshwater species that is not native to the location. Reintroductions of native species are excluded. Multiple releases of the same species are counted only once. To be counted as an attempt, records must show that at least a pair of individuals was released into the wild (not simply shipped to a location). Some exotic birds now occur in the avifaunas today because they colonized recently on their own or because they are cage-escapees but there is no record of their importation or release. These species are counted both as attempts and as successes. Since there is an unknown number of failures in this category, the success rates in Table 2 are likely to be artificially high. Table 2 is based on locations with at least 10 docu- mented attempts and where I was able to verify the species' present status either through personal visits or through the literature. I have personally conducted field studies in about half these sites.
Many failures are immediate; the birds are released and disappear soon after. Other exotics maintain thriv- ing and even expanding populations for some time but subsequently become extinct. Should we count the lat- ter as failed introductions or as extinctions of success- ful introductions? Moulton and Pimm (1983) in their analysis of the Hawaiian situation chose the latter course and they were aided in their assessment by par- ticularly good historical records. However, in most places it is difficult to know when a small population becomes extinct and thus how long it survived before perishing. I therefore choose the conservative measure of counting successes as only those introduced species that survive to the present time. Moulton and Pimm (1983), Lockwood et al. (1993), Simberloff and Boecklen (1991) and others have restricted their analyses to only passerines or to passerines plus doves. Exotic game birds and parrots are common introductions but are excluded because many parrots are established as cage-bird escapees while human hunting pressure could exaggerate the failure of game birds. However, such a distinction seems arbitrary, since many finches are also kept as cage birds that end as established exotics (Long, 1981) and game birds include examples of both striking successes and failures (Banks, 1981; Pimm, 1991).
Relative abundance of introduced and native species in native and disturbed habitats Personal surveys The first two data sets are of value in understanding regional patterns in the number and success of intro-
ductions. To determine if competitive interactions between exotics and natives may be influencing habitat distributions at a finer geographic scale, I surveyed birds in the following Pacific basin and rim sites from 1983 to 1990: Fiji (four islands and 20 sites), Palau (two islands, two sites), Hawaii (five islands, 21 sites), Tahiti (three sites), Cook Islands (two islands, two sites), Marquesas (two islands, two sites), Vanuatu (two islands, six sites), New Guinea (two sites), New Zealand (two islands, 18 sites), New Caledonia (three sites), mainland Australia (19 sites), and Kangaroo Island off South Australia (one site). This collection of Pacific sites includes those with avifaunas ranging from few to many introduced bird species and from few to many native bird species. Each site was categorized into one of four habitats: native forest; exotic forest (usually pines or eucalypt plantations); disturbed native habitats (at least 50% exotic species by areal coverage and always secondary growth); and suburban/urban locations (usually city parks, college campuses, or sub- urban housing tracts) with typically much open space and over 50% exotic vegetation.
Two types of censuses were performed. At all sites bird species lists were compiled based on visual and call identification. Sites were surveyed over a period of 2-8 h over 1~1 days. The area surveyed was approximately 2-20 ha. For a subset of locations (n = 52) individual birds were counted using the variable circular-plot technique (Reynolds et al., 1980). Birds were counted at between five and 14 stations 100-300 m apart along a transect of continuous habitat. Birds were identified by sight or by calls for 6 min at each station. Longer time intervals may result in counting the same bird more than once (Reynolds et al., 1980; Scott et al., 1986). Because of dramatic differences between islands in the numbers of bird species and in their habits and habitat structure, these data are not reliable for com- paring absolute densities of birds between locations. The data are useful, however, as rough estimates of the relative abundances of native vs introduced species at different sites and in different habitat types.
Additionally, I have compiled each location's native species number, area, and maximum elevation. These values differ from those in Table 1 since now I am interested in a specific island's values not archipelago values. For this purpose mainland Australia was divided into eight geographic regions.
Literature surveys Bird abundance data in New Zealand and Australia were also gathered from studies in the literature (Gibb, 1961; McLay, 1974 [a review of five New Zealand studies]; Disney & Stokes, 1976; Friend, 1982; Wall, 1983). These studies were performed over much longer time intervals, of a few months to a few years, com- pared to the short-term census data that I collected. These surveys are analysed in the same way as my own surveys to check the robustness of results obtained

76 T. J. Case
from my short-term surveys and to examine abundance data for native and introduced birds.
Statistical analysis Raw variables were examined for normality and trans- formed by the angular transformation (for proportions), square root (maximum elevation), or a log transforma- tion (numbers, counts, and area) if that produced a more normal distribution. Since some variables contain zero values for some islands/regions, I added the constant 1-0 to these variables before applying a log transform. In stepwise regressions, the variables were stepped first in the forward and then in the backward direction. The backward model begins with all vari- ables added and successively drops those with insignifi- cant partial correlation coefficients with the dependent variable. In the vast majority of cases, direction did not influence the resulting regression model. In the few cases where the stepwise models differ in the two direc- tions, I report the model that produces a significantly better fit to the data. The residuals around the regres- sion were visually examined for homoscedasticity and results are reported only when this assumption is met. Statistical analyses were performed using the statistical software Statview T M 4.0 from Macintosh.
RESULTS
Introduced species number The correlation matrix for the variables in Table 1 is shown in Table 3. While log area is highly correlated with log extant native species number (R 2 = 0-66), the correlation between log area and log exotic species number is only marginally significant (Table 3, p = 0-07). Hence, there is substantial unexplained varia- tion in the numbers of introduced species that might be accounted for by other factors. After partialling out the affect of area and elevation on both exotic and native bird species number, a stepwise regression was performed using the residuals of introduced species number as the dependent variable and the remaining variables in Table 1 as independent variables. Native species richness (the residuals after accounting for area and maximum elevation) shows a significant negative partial correlation coefficient (p = 0.013; Fig. 1) with the introduced species residuals, although the explained
i .8 .6
.4
.2 v 0 .g
-.2 - 4 i
~n . . 6 ¸ - I
"~ - . 8
. I . , . t . L . | . i . i . f . I .
• • • ; -, • o ~ • • • • • Q
Q
- 1 . 2 - 1 - , 8 - . 6 - . 4 - . 2 0 . 2 . 4 . 6 . 8
Res idua ls n a t i v e s p e c i e s n u m b e r
Fig. 1. A visualization of the negative partial correlation between log introduced bird species numbers and the log of native bird species numbers after partialling out variation in log area and square root elevation. The regression line is
shown: Y = 4).298 * X; R 2= 0-083; p = 0-013.
variance is quite low (about 8.3%). Much more impor- tant is the number of extinct species, which is the only independent variable entered into a stepwise regression model, and which correlates positively with the residu- als of introduced species number. The same result emerges for both forward and backward stepwise regressions and the regression is highly significant (p <0.0001), explaining 43-4% of the variance (Table 4(a)). Thus nearly half the variation in introduced species richness (controlling for island area and eleva- tion) is explained by the number of extinct native species in the avifauna. A rich native bird fauna per se, however, does not seem to hinder the success and per- sistence of exotic birds after controlling for island area, elevation and extinct native species.
An alternative stepwise multiple regression technique is to ignore any a priori expectations about potential affects of island area and elevation on species number. Now the log of introduced species number (rather than the residuals of this variable after accounting for area and elevation) is used as the dependent variable and area and elevation are included as dependent variables in a stepwise regression along with the remaining vari- ables in Table 1. The stepwise regression in the forward direction first adds extinct native species number as the most significant independent variable, and it alone
Table 3. Correlation matrix for variables in Table 1
Log extinctions Log introduc. Sqrt elev. Log area Human Mammal cat. Log natives
1-000 0.664 0.125 0.046 0.171 4)-070 4). 154 1-000 0-216 0.216 0-188 0.029 0.030
1- 000 0- 710 0.187 0.394 0.494 1.000 0.408 0-673 0.823
1.000 0-473 0-413 1.000 0.686
1.000
Log extinctions Log introduc. Sqrt elev. Log area Human settlement Mammal cat. Log natives
Coefficients greater in absolute value than 0-230 are significant at p -- 0.05. Coefficients greater than 0.302 are significant at p = 0.01.

Introductions and extinctions of birds 77
Table 4. Stepwise regression models using the variables in Table 1 to account for variation across locations in the number of exotic or extinct native bird species
(a) Dependent variable: Residuals of log introduced species number after being regressed against log area and sq. root of maximum elevation
Total d.f.: 69 p-value <0.0001 R2:0-444
Variable Coefficient Std Error F to remove
Intercept 0.278 0-051 30.184 Log extinctions 0-532 0.072 52.211
Variables not significant in model: native species number residuals, human settlement, preda- tor category.
(b) Dependent variable: log of introduced species number
Total d.f.: 69 p-value <0-0001 R2: 0.478
Variable Coefficient Std Error F to remove
Intercept 0.285 0.094 9.272 Log area 0.052 0.025 4.473 Log number extinctions 0-531 0.071 55.308
Variables not significant in model: native species number, human settlement, predator category, maximum elevation.
(c) Dependent variable: log of extinct native species number. Table 1 variables plus native species endemism
Total d.f.: 69 p-value <0.0001 R2:0.511
Variable Coefficient Std Error F to remove
Intercept -0.201 0-097 4.570 Sq. root species endemism 0.385 0.122 9.907 Log introductions 0-762 0.109 48.780
Variables not significant in model: extant native species number, human settlement, predator category, maximum elevation, log area.
(d) Dependent variable: the proportion of extinct native species (sq. root transformed). Analysis restricted to those locations with at least one extinct and at least one extant native species
Total d.f.: 45 p-value <0.0001 R2: 0.655
Variable Coefficient Std Error F to remove
Intercept 0-505 0.077 43.088 Log area -0.147 0-022 44-262 Log introductions 0.279 0.064 19.011 Log number of endemics 0-17 0-047 12.931
Variables not significant in model: human settlement, predator category, maximum elevation.
accounts for about 44.3% o f the variance in introduced species. However , log area is also added (Table 4(b)) with a positive coefficient and the combined two vari- ables account for 48% o f the variance in log introduced species number. This regression is highly significant (p <0.0001) and the same model is reached f rom both forward and backward stepwise procedures. I f we
repeat the analysis but only include those islands with at least one (or two) extinctions, and at least two intro- duced species under the assumption that those islands with very few known extinctions or introduct ions are simply understudied, the stepwise model enters only the number o f extinct species and this is strongly and positively correlated with numbers o f introductions

78 T. J. Case
(d.f. -- 49 and d.f. = 32; both p <0.0001). Finally, if we screen the locations to use only those where subfossil birds are known (Table 1), the stepwise model (in both the forward and backward directions) again produces the same basic result: the number of extinct species is the only variable entered and is strongly and positively correlated with numbers of introductions (n -- 37; p <0.0007).
While log transformed species numbers are used in the multiple regressions, the precise relationship between introductions and extinctions is more easily visualized by plotting their untransformed values (Fig. 2). The slope here is about 0.74, implying that on average about four extinctions have occurred for every three introduced species gained. Thus species number has on average remained roughly steady despite substantial gains and loss of species and varying degrees of habitat destruction and deterioration across locations. In the
65
.~ 55 u
o. 45 ~n
35 ~g u = 25 " O
O
*., 15 c
5
(a)
New Zealand / ~ , ~ J ' f ~
• /
Kauai Maur i t iusf~, ~ ~ ~ , m
• R e u n i o n m r ~
. . . . i . . . . i . . . . ! . . . . i . . . . , . . . . i . . . . i , ,
S 15 Z5 35 45 55 65
Extinct native species
(b) i . . . . i . . . . , . . . . i . . . . i . . . . i . . . . i . . . . I ~
65 ~ I
o . 45 s I
C
s 15 Z5 35 4S 55 65
Extinct native species
Fig. 2. (a) The relationship between the number of exotic and extinct native bird species for the locations in Table 1. line of equality; - - , regression line (Introduced = 3-084 + 0.742 * Extinct; R 2 -- 0-699; p < 0.0001; n -- 70).
Some islands with high numbers of extinct and introduced species are labeled. (b) Same as in (a) but only those islands from Table 1 where prehistoric birds have been uncovered from fossils are included. (Introduced -- 4.135 + 0.704 *
Extinct; R 2 -- 0.657; p <0.0001; n = 37).
Discussion, I will examine sources of bias in these data, e.g. the likely underestimation of extinctions. Since the number of extinctions is not significantly correlated with area (both variates log transformed, p >0-7) and the number of introductions is only marginally corre- lated with area (p -- 0.079, both variates log trans- formed), the correlation between the number of introductions and extinctions is not simply due to mutual covariation with area. If we include only those islands where there is at least one extinction, under the assumption that those islands with no extinctions are simply under-sampled, the correlation between log extinctions and log introductions is still highly signifi- cant (p <0.0001, n = 50). Similarly, including only those islands with known subfossil birds, does not sub- stantially alter either the slope or the significance of the correlation (Fig. 2(b)).
Native extinctions Since extinctions are a reliable predictor of introduc- tions, it is useful to explore the factors contributing to variation in the number of extinct species and rates of extinction across these locations. A stepwise regression was performed using the number of extinct species as the dependent variable (log transformed). An addi- tional dependent variable was added to those in Table 1, the level of species endemism, i.e. the number of endemic species (extant natives including those histori- cally extinct that are endemic at the species level to the location) divided by the total number of native species. The most significant variable explaining the number of extinct species for Table 1 locations is the number of introduced species (Table 4(c)) attesting to the tight and reciprocal relationship between extinctions and intro- ductions. The stepwise model next entered the level of endemism (square root transformation) with a positive affect on extinction; this two-variable model explains 51% of the variance in the number of extinctions across islands. A backward regression yielded the same final model. I next limited the analysis to only those islands with at least one extinction and one introduction. This reduces the sample size to 47 but improves homo- scedasticity; the resulting regression model enters only the number of introduced species, which explains 37% of the variance (t9 <0.0001). Screening the locations to include only those where some subfossil birds are known (Table 1, asterisked locations eliminated) reduces the sample size to 36 locations, and returns endemism to significance with introduced species (p = 0-0004; R 2 = 0-376).
The proportion of extinct native species in the avi- fauna (i.e. No. all extinct species / [No. all extinct + No. extant species]) is a variate of interest since it tells us something about relative extinction rates between locations. I eliminate those islands with no extinctions and no extant native species, and then apply a square root transformation to the proportion of extinct species in the remaining locations. The proportion of extinct
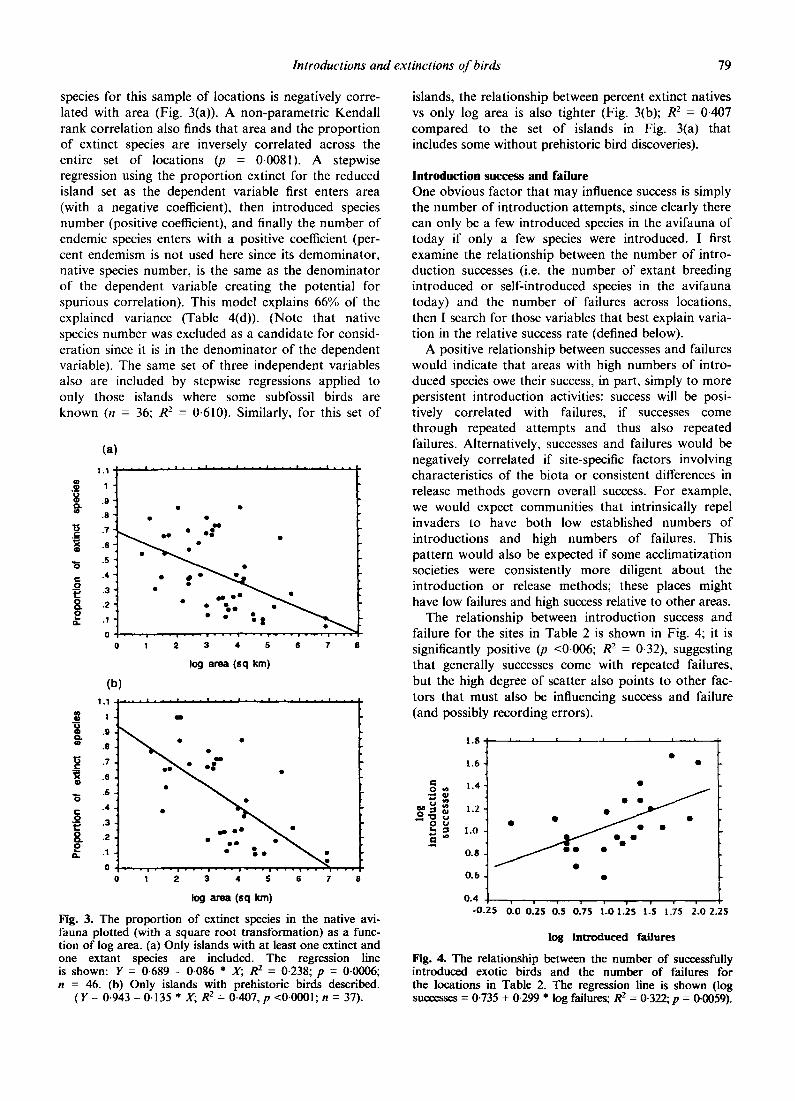
Introductions and extinctions of birds 79
species for this sample of locations is negatively corre- lated with area (Fig. 3(a)). A non-parametr ic Kendall rank correlation also finds that area and the proport ion of extinct species are inversely correlated across the entire set of locations (p -- 0.0081). A stepwise regression using the proport ion extinct for the reduced island set as the dependent variable first enters area (with a negative coefficient), then introduced species number (positive coefficient), and finally the number of endemic species enters with a positive coefficient (per- cent endemism is not used here since its demominator , native species number, is the same as the denominator of the dependent variable creating the potential for spurious correlation). This model explains 66% of the explained variance (Table 4(d)). (Note that native species number was excluded as a candidate for consid- eration since it is in the denominator of the dependent variable). The same set of three independent variables also are included by stepwise regressions applied to only those islands where some subfossil birds are known (n = 36; R 2 -- 0.610). Similarly, for this set of
(a) 1 . 1 - - , ' , , * l * , ' ' " " " ' " " ' ' " " ' ' " " ' ' , , ,
. |
D • .6
"6 .S
" " " : ' " o . I
~ L
0 - - - i - - - i - . - i . . . ~ m - % . i . . . I . . . i . .
0 1 2 3 4 5 6 7
l o g a r e a ( s q k i n )
(b) 1 . 1 , , , I , - - , - - - " - - - ' - - - ' - - - ' - - - ' - - -
" ~ .6 •
~ .S 0
C . 4
._o .3
:7" 2 .'t • ; o
0 - - - i - - - m . . . . . . . .
0 1 2 3 4 s 6 7 8
k)g area (sq km)
Fig. 3. The proportion of extinct species in the native avi- fauna plotted (with a square root transformation) as a func- tion of log area. (a) Only islands with at least one extinct and one extant species are included. The regression line is shown: Y = 0-689 - 0-086 * X; R 2 = 0.238; p = 0-0006; n -- 46. (b) Only islands with prehistoric birds described.
(Y= 0-943 -0.135 * X; R 2 = 0-407, p <0.0001; n = 37).
islands, the relationship between percent extinct natives vs only log area is also tighter (Fig. 3(b); R 2 = 0.407 compared to the set of islands in Fig. 3(a) that includes some without prehistoric bird discoveries).
Introduct ion success and fai lure One obvious factor that may influence success is simply the number of introduction attempts, since clearly there can only be a few introduced species in the avifauna of today if only a few species were introduced. I first examine the relationship between the number of intro- duction successes (i.e. the number of extant breeding introduced or self-introduced species in the avifauna today) and the number of failures across locations, then I search for those variables that best explain varia- tion in the relative success rate (defined below).
A positive relationship between successes and failures would indicate that areas with high numbers of intro- duced species owe their success, in part, simply to more persistent introduction activities: success will be posi- tively correlated with failures, if successes come through repeated attempts and thus also repeated failures. Alternatively, successes and failures would be negatively correlated if site-specific factors involving characteristics o f the biota or consistent differences in release methods govern overall success. For example, we would expect communities that intrinsically repel invaders to have both low established numbers of introductions and high numbers of failures. This pattern would also be expected if some acclimatization societies were consistently more diligent about the introduction or release methods; these places might have low failures and high success relative to other areas.
The relationship between introduction success and failure for the sites in Table 2 is shown in Fig. 4; it is significantly positive (p <0.006; R 2 = 0.32), suggesting that generally successes come with repeated failures, but the high degree of scatter also points to other fac- tors that must also be influencing success and failure (and possibly recording errors).
1 . ~ I I [ I I I I I I
$ • 1.6
i ~ 1 . 4 • •
-- ,,,~ I.O • ~ ~ o o o • • • 0.8 ~ o • •
0 . 6 " f • •
0 . 4
-o.zs ~.o o3s o.s o3s 1:o 1.1~ f s ~ s ~.o 2.zs
log i n l a ' o d u e e d f a i l u r e s
Fig. 4. The relationship between the number of successfully introduced exotic birds and the number of failures for the locations in Table 2. The regression line is shown (log successes = 0-735 + 0-299 * log failures; R 2 -- 0-322; p = 0-0059).

80 T.J . Case
What other dependent variables in Table 1 best explain the variation in introduction success between locations? We desire some measure of success that is independent of the number of introduction attempts. Since success rate (column B of Table 2) contains intro- duction attempts in its denominator, the two variables are not statistically independent. I use instead the residuals from the regression in Fig. 4, which represent a measure of relative success rate. They are used as the dependent variable in a stepwise regression with candi- date independent variables: extant natives, extinct natives, area, maximum elevation, and mammal preda- tor category, each suitably transformed for normality.
The only variable that is Significantly correlated with relative success rate is the number of extinctions (log transformed; Fig. 5). A non-parametric Kendall rank correlation supports the significance of this correlation (p = 0.039).
In summary, the number of successfully introduced species increases with the number of failures and the relative success to failure rate increases with the number of extinct native species. These results are consistent with Elton's (1958) idea that habitat disturbance increases invasion success but does not support his contention that species-rich locations, in and of themselves, repel invaders.
Habitat distributions of exot ic and native birds Both on the basis of numerical abundance and species number, human-modified (or non-native) habitats con- tain a greater proportion of exotic birds than do native forest habitats (Fig. 6). Moreover, the local number of exotic species observed in surveys and the regional number of exotic species in the avifauna are highly cor- related in the two most common habitat types (Fig. 7(a)). (There are too few surveys in secondary growth habitats and exotic forest habitats to determine their separate relationships.) The numerical abundance of exotic birds relative to native birds also increases with the overall island's exotic bird species number (Fig. 7(b)).
.5
.4 -'
. 1 io ~ -.Z
~ - . 3
, i , 1 , i . _ , . t , i , t , t . , . I ,
• I
~ • 0 0 •
-.4.
° S ~
.Z 0 ,2 .4 .6 .8 1 1.2 1.4 1.6 1.8
log number of extinctions
Fig. 5. The relative success rate for avian introductions plot- ted against the number of avian extinctions for the locations in Table 2. The relative success rate is obtained from the residuals of the plot in Fig. 4. The regression line is shown:
Relative success rate -- ~).165 + 0.214 * X; R 2 = 0.226.
A.
1.2
1.0
O.8
0.6
0.4
O.Z
0
1.2
1.0
0.8
0.6
0.4
0.2
0
F~ proportion exotic by numbers E~ proportion native by numbers
22 20 4 6
urban/ native exotic secondary suburban forest forest habitat
[] proportion exotic by species B. [] proportion natwe by species
D
46 32 9 11
urban/ native exotic suburban forest forest
secondary habitat
Fig. 6. The proportion of individuals (A) and species (B) that are exotic or native in the four different habitat types sur- veyed across a range of sites on several Pacific locations. The numbers over the bars are the number of sites in each habitat category. A one-factor ANOVA performed on the angular transformation of these proportions shows that habitat significantly affects the proportion of exotic birds in the
different habitats.
Not too surprisingly the number of native species observed in local surveys is positively correlated with the number of native species on the island in both urban/suburban and native forest habitats (p <0.0001, Fig. 8(a)). The proportion of individuals in local surveys that belong to native bird species increases with native species number (Fig. 8(b); p <0.02 for both habitat types).
More germane to Elton's hypothesis, the number of exotic bird species in point surveys significantly declines with the number of native species present in these sur- veys in both habitat types (Fig. 9, urban!suburban, p <0-0001; native forest, p <0-001). Since the number of native species in point censuses is highly correlated with the number of native species in the island avi- fauna, it is difficult to disentangle the relative effect that the regional species pool and habitat clearing, compared to local factors, might be playing in produc- ing the negative relationship between natives and exotics in the point surveys.

Introductions and ext inct ions o f birds 81
.c_
0
( J
0
O~ 0
. J
O urban/suburban (a) o nrdve forest
1 . ~ , I . t , i . i . i . i . , . I , I ,
.8
.6
.4
.2
0
-.~' 0 . Z . 4 . 6 . 8 1 1 . Z 1 . 4 1 . 6 1 . 8
Log exotic species on island
1.8
~= 1.6
c 1.4
.~ ~.z
" 0 .-.R .8
0 .4
c . Z
._o t:t 0
-.2 2
(b) • i . , . i . J . i . ] . i . i , i • k
-.2 0 .2 .4 .6 .8 1 1.2 1.4 1.6 1.8
Log exotic species on island
Fig. 7. (a) The relationship between the local number of exotic species and the regional number of exotic species in the avi- fauna for the two most common habitat t~pes. The regression lines are shown (urban/suburban habitats: R- = 0.625, p <0-0001, n = 46; native forest: R 2 = 0-526, p <0.0001, n = 32). (b) The pro- portion of exotic individuals observed (with an angular trans- formation) in local surveys plotted as a function of the regional number of exotic species in the avifauna. The regression lines are shown (urban/suburban: R 2 = 0-399, p = 0.0016, n = 22);
native forest: R 2 = 0-413, p -- 0.0022, n = 20).
1 . 8
1 . 6
1 . 4
.~ 1 .z
.~ 1
Vt . 6
-~ .4 C . 2
o ~ o ._1
* . 2
• urban/suburban (a) o n,uve ~ r ~
I I I I I I I I
Q
• • e l l •
i i J i i i i i
. $ . 7 5 1 1 . Z S 1 . 5 1 . 7 5 2 2 . 2 S Z .S Z . T S
Log native species on island
1.8
~ 1.6
.~_ 1.4
u~ 1 . Z
.6
• - .4
e- ._o 0 t~ ~ . -,Z 0 o-
(b) i I , I i I I i
Q l i E ] D ~ o l
i i i i i r i
.S . 7 S 1 1 . Z 5 1 . 5 1 . 7 5 2 Z . Z 5 Z . 5 Z . 7 S
Log native species on island
Fig. 8. (a) The relationship between the number of native species in local point surveys and the regional number of extant native species on the island/archipelago (urban/sub- urban habitats: R 2 = 0-536, p <0.0001); native forest: R 2= 0.515, p < 0-0001). (b) The relationship between the pro- portion of native individuals in local point surveys and the regional number of extant native species on the island/ archipelago (urban/suburban: R 2 = 0.259, p = 0-0155; native
f o r e s t : R 2 = 0.306, p = 0-011).
Are regional effects important even after accounting for local site-specific variables? To answer this I regressed the local species number of exotics against the local number of native species across all sites (both variables log transformed). I saved the residuals from this regression, averaged them across all sites within each island/region and then performed a multiple regression using the averaged residuals as the depen- dent variable and island/region species number for exotics and natives as the independent variables (again log transformed). In this regression there is thus only a single point per island/region to avoid pseudoreplica- tion. This was repeated separately for non-native and native habitats. Finally, this process was repeated beginning with the regression of the local proport ion of exotic individuals in each census (with an angular t ransformation) as the dependent variable and the local
native species number and exotic species number as independent variables in a stepwise multiple regression. Again residuals were saved and averaged for each island/region and then entered as the dependent vari- able in the regional regression analysis.
The results are summarized in Table 5. Regardless of habitat type, exotic species number is negatively related to the number of native species in the points censuses. After accounting for this variation, the residuals in exotic species number are further positively related to exotic species number for the entire island/region. In both forest habitats and man-modified habitats, the proportion of exotic individuals in surveys is negatively related to the number of native species present in the survey and positively related to the number of exotic species. Regional species numbers did not significantly correlate with the average residuals from these regressions.

82 T. J. Case
1 . 2
. ~ . 8
. -~ . 6
. u
o 0 _ J
• u r b a n / s u b u r b a n [
rn native forest
• i , I . | . i , t , | . i , J , i
* . 2 i - v . i • 7 • L . i • i - ~ - i -
Z 0 . Z . 4 . 6 . a 1 1 . z 1 . 4 1 . 6 1 . 8
Log native species in survey
Fig. 9. The negative relationship between the number of exotic species found in point surveys and the corresponding number of native species (log transformed) (urban/suburban: R-' = 0-364, p < 0-0001; native forest: R 2 = 0.194, p = 0.0117).
Literature surveys for New Zealand and Australia As for my personal surveys these surveys obtained from the literature also reveal a higher number (and proportion) of exotic species in human-modified habi- tats than in native forest (Fig. 10). New Zealand has substantially more exotic species in its avifauna than Australia and this leads to higher numbers of exotic species being present in point surveys in both native forest and modified habitats (Fig. 10). A two-factor ANOVA based on the results shown in Fig. 10 finds a significant location effect (i.e. more exotic species in New Zealand than Australia; p <0.0001) and significant habitat affect (p <0.0001), but the interaction term is not significant. Similarly, a two-factor ANOVA based on native species number in the literature surveys finds
significantly more native species in point surveys in Australia than in New Zealand (p <0-0001) and signifi- cantly more natives in native forest habitat compared to human-modified habitats (p <0.0001) but no signifi- cant interaction term (p >0.9).
As for my surveys, the number of exotic species is strongly negatively correlated with the number of native species present in point surveys in both habitat types (Fig. 11; p = 0.0002 for native forest and p <0.0001 for modified habitats).
For the New Zealand surveys (but not the Australian studies), the authors often supplied estimates of the density of individuals in the study area. The total num- ber of exotic birds is positively correlated with the number of exotic species present in the survey for both habitat types (Fig. 12(a)). The number of exotic indivi- duals is negatively correlated with the number of native species in modified habitats but not significantly so in native forest (Fig. 12(b)). A multiple stepwise regression with the log of exotic individuals as the dependent vari- able found that only the number of exotic species entered into the model for both habitat types. Thus the statistical significance of the negative relationship between the number of exotic birds and the number of native species in point surveys disappears once between-site variation in exotic species number is accounted for. This suggests that even in fixed habitat types exotic species and native species are negatively associated across sites, but that those natives that are present in a site are not affecting the numerical abun- dance of exotics very much. The number of native indi- viduals is not significantly correlated with either the number of native species or the number of exotic species in point surveys in either habitat type (not shown).
Similarly the number of native individuals is not cor- related with the number of exotic individuals in either
Table 5. A summary of stepwise multiple regression results separating the independent variables into likely regional effects and local effects The sign of the coefficient (positive or negative) is indicated with the statistical significance level by the number of asterisks. * p = 0-05 0.01, ** <0.01 and >0-001, *** <0.001 and > 0.0001, **** <0.0001. The box outlines those cells where local inter- specific competition between natives and exotics might be expected to yield negative signs. The regional variables are regressed against the average residuals (over all sites within a region) from the regression involving only the local variables at each site. NA, not applicable.
All non-native habitats Dependent variable
Native forest habitats Dependent variable
No. exotic species Prop. exotic No. exotic species Prop. exotic in surveys indiv, in surveys in surveys indiv, in surveys
Regional variables (Species source pools and broad habitat alteration)
No. native species on island No exotic species pos. ** pos * on island (n = 26) (n = 18) (n = 18) (n = 11)
Local variables (Possible competitve interactions and habitat variation)
No. native species neg **** neg **** neg * neg *** in survey No exotic species NA pos **** NA pos ** in survey (n = 66) (n = 32) (n = 32) (n = 20)

Introductions and extinctions o f birds 83
Mean no. exot ic species in point surveys
(a)
1
.9
.8
.7
.6
.$
.4
.3
.2
.1
Literature censuses only
native forest human-modif ied
.7
Mean no. .6 exotic .s species in point .4
surveys .3
.2
.1
0
( b ) A l l censuses combined
Y
native forest human-modified
Fig. 10. The cell means for the mean number of log exotic species in native forest and all human-modified habitats com- bined for Australian and New Zealand sites. (Error bars are 95% confidence intervals.) (a) Only surveys from the litera- ture. (b) Those same literature surveys combined with my
own surveys from New Zealand and mainland Australia.
habitat type. We would not necessarily expect negative relationships between species abundances of these two groups across sites because of gradients in overall resource productivity, which would impose positive covariance. That is, birds of all species will respond similarly to differences in avian carrying capacity between sites diluting the appearance of any negative correlations that might emerge if we could control for environmental differences.
DISCUSSION
Regional patterns This review finds two particularly intriguing results. First, I find a strong correlation between the number of bird species that have successfully been introduced to an island (as well as the relative success rate) and the number of known extinctions of indigenous species in historical and aboriginal times, such that the number of exotic species gained is close to the number of species lost through extinction. Indeed the correlation between introductions and extinctions accounts for 70% of the variance in introduced species number (for untrans-
[ [
oo 0 ,
.4 .6 .s ~ 1 .z ~.4 ~ .6 ~ .8 z
log number of native species
Fig. I1. The negative relationship between the number of exotic species found in point surveys and the corresponding number of native species for Australian and New Zealand sites gleaned from the literature (modified habitats: R 2 : 0 " 4 4 4 ,
p <0.0001; native forest: R 2 = 0.394, p = 0-0002)•
formed variables; 45% for transformed variables). The second result is that invasion success does not decline significantly with the richness of the native avifauna nor the variety of potential mammalian predators once between-site variation in extinctions and island area are accounted for.
This latter result contrasts with the pattern seen for reptiles on much the same set of locations analysed here for birds. Case and Bolger (1991) found that native reptile species number had a highly significant negative partial correlation coefficient with introduced species number. After updating these data for recent discoveries and taxonomic revisions and including information on the numbers of extinct reptile species and human settlement patterns (not considered in the original paper), I find that island area (with a positive coefficient) and native species number (with a negative coefficient ) are the only significant variables explaining the numbers of introduced reptiles across islands (and USA and Australia) in a stepwise regression model. However, the number of reptilian extinctions is poorly known for most of these island locals (see Case et al., 1992) and the explained variance of the model is only about 15%, much less than that for birds. Native species number may play a larger role in reptiles than in birds in preventing exotic success and persistence because exotic and native reptile species seem to share habitats more frequently than do exotic and native birds (Case & Bolger, 1991; Losos et al., 1993).
The rough substitution of extinctions by introductions We should probably not interpret too much into the slope of the relationship between species gains and losses, which is about 0-75. While the equilibrium theory of island biogeography (MacArthur & Wilson, 1967) posits a close relationship between species colo- nizations and extinctions, this expectation is based on islands that are receiving natural immigrants and not

84 T .J . Case
(a)
3 .Z
. ~ Z.
~ 2 . 6 -
= 2
1 4 L ~ e I 1.Z - ~
.2 .3 .4 .5 ~6 .7 .8 .9 1 1,1
log number of exotic species
(b)
3.4 . . . . 'o . . . . B . . . . . . . . . . . . . . . . . . . . . . . . 3,Z3
2.8 2.6 o
"~ 2.4 o • • .g Z.2
' ~ 2 • c ..~'- 1.a"
1.G: 1.4 : • • • 1.2 . . . . , . . . . , . . . . , . . . . , . . . . , . . . . , . . . .
.5 6 .7 .8 .9 1 1.1 1.Z
log number of native species
Fig. 12. The relationship between the numbers of exotic individuals in point surveys and the number of exotic species (a) and the number of native species (b) present in the New Zealand/Australian region. (a) Modified habitats: R 2 = 0-847, p = 0.047, p <0.0001; native forest: R 2 = 0.312, p = 0.047. (b) Modified habitats: R 2 = 0.577, p -- 0-0002; native forest: R 2 - - 0-092, p = 0-314. Even though the total number of exotic bird individuals declines as the number of native species increases in modified habitats (but not native forest; (b)) the positive effect of exotic species number on exotic indi- viduals is stronger (a) and when both variables are available in a stepwise regression, only exotic species number is entered
into the model (see text).
simultaneously undergoing major habitat alteration. When these conditions are added, the equilibrium theory can predict numbers of extinctions very different from the number of colonizations. Consider the classical thought experiment suggested by Preston (1962) of dividing an island into two equal but now isolated halves. Since the number of species usually scales with area raised to a power of about 0.25; halving area results in only about a 16% loss in species number (i.e. 1 - 0.5°25). If we deforest half an island and if all of the native species cannot tolerate this new open habitat, then over time extinctions should eliminate about 16% of the native species. Now we introduce new bird species that can only occupy these new open habitats. Call the original number of species S; then
when disturbed habitats are saturated with new exotic species, we will have approximately 0.84 S exotic species living in disturbed habitats plus 0-84 S native species living in native forest habitats; the total number of species island-wide is about 68% (= 2 × 0.84 × 100) greater than before half the island was deforested. For this scenario, at equilibrium the gain in exotics far out- numbers the loss of natives from extinction. On the other hand, if exotics and natives showed no differen- tial habitat preferences, then at equilibrium the number of gains and losses are expected to be equal preserving the total number of species before and after deforesta- tion. With intermediate habitat sharing of exotics and native species, and with some islands experiencing more habitat conversion than others, and predators poten- tially reducing carrying capacities for both exotics and natives, and islands probably not close to equilibrium with respect to their recent and ongoing habitat alter- ations, the expected relationship between gains and losses cannot be deduced. We should therefore take the observed slope of 0.74 as an empirical observation for this taxon and group of islands at this time, but expect no general canonical properties. Since several island groups probably have many more extinct species waiting to be discovered, the slope could decrease.
Avifaunas with high levels of endemism have experi- enced relatively more extinctions for this broad set of islands matching the pattern found by McDowall (1969) for New Zealand birds. McDowall found that 37% of the endemic New Zealand birds are now extinct but only 6% of the native but non-endemic species. Moreover, he found a tight relationship between the degree of endemism and the propensity for extinction. Elsewhere (Case, in prep.), I show that there is a high correlation across locations in the endemism levels of extant and extinct species, and endemism levels in land birds are correlated to those of land reptiles. Endemic reptiles on many islands are also more extinc- tion- (and extirpation-) prone than non-endemic species (Case et al., 1992) and the same pattern appears to be generally true for land birds (McDowall, 1969; Johnson & Stattersfield, 1990; Steadman, 1991; Adler, 1992). Another contributing factor to the apparent vulnera- bility of island endemics could be the lack of recolo- nization sources. When a population of non-endemics or regional endemics becomes locally extinct on an island, the island can potentially be recolonized from individuals still surviving on other nearby islands. For a single-island endemic, however, extirpation and extinction are equivalent.
One consequence of the rough substitution of exotics for natives and particularly endemic natives is that in the calculation of log species-log area relationships for the islands in Table 1, the explained variance (72-1%) is greater when the extant native species number is added to the islands' extinct native species and the total species number is used as the dependent variable compared to when either species set is regressed alone

In troduct ions and ex t inc t ions o f birds 85
(R 2 = 66.6% for extant natives alone and R 2 = 0.2% for extinct natives alone). Similarly, when the extant natives are added to the introduced species component, log area again explains 72% of the variance.
Cause and effect: extinctions and introductions Most avian extinctions occurred prior to the majority of species introductions in most locations. For the set of locations in Table 1, there are 139 extinctions since the approximate year 1600 and 231 prehistorical late Holocene extinctions. At least half of the historical extinctions also probably occurred before the establish- ment of introduced species through the efforts of acclimatization societies in the mid to late 1800s. This temporal sequence indicates that most native avian extinctions are not directly caused by competition with introduced species, although some could be.
The native extinctions, whether in prehistory or in more recent periods, are for the most part traceable to human activities including hunting, habitat destruction and the introduction of exotic predators or large herbi- vores that disrupt habitats (for birds: Simberloff, 1981, 1990; Cassels, 1984; Diamond, 1984; Olson & James, 1984; Ebenhard, 1988; Holdaway, 1989; Olson, 1989; Steadman, 1989b; and in reptiles: Case et al., 1992; Henderson, 1992; North et al., 1994). The idea that these native extinctions might cause vacant niches for the introduced species is difficult to reconcile with the fact that there are few clear ecological replacements in the two species sets. For example, In New Zealand giant moas are lost while smaller thrushes and finches are gained; Hawaii experienced a loss of many ducks, geese, raptors, rails and forest nectar feeders but a gain of mostly small seed and insect eaters of open habitats. Worldwide, many endemic rails have become extinct but rails or species with ecological niches like rails are not generally gained through introductions.
It must be remembered, however, that direct biotic interactions like predation, interference competition, and parasitism are easier to see and therefore it is easier to establish proof of their importance. Competi- tion for food is not a direct interaction between two competitors but is only manifested through the dynamic consequences on other species: the resources. The level of proof needed to demonstrate exploitation competi- tion is burdensome (Petren & Case, 1996). Thus, with exploitation competition we have the double difficulty of first demonstrating that it is an important force in present-day communities, and secondly connecting its occurrence to patterns at the biogeographic scale.
An alternative explanation for the correlation is based on the fact that native birds are usually more common, if not restricted, to native habitats while introduced birds are primary occupants of disturbed and open habitats (Figs 6 and 10, see also Temple, t981; Moulton & Pimm, 1983; Holdaway, 1990). Temple (1981) notes that almost all of the remaining indigenous birds in the Indian Ocean are obligate forest-inhabiting
species and Holdaway (1990) attributes the high extinc- tion rate on New Zealand before European settlement to the removal of forest (particularly dry forest). Hawaiian native passerines rarely are seen outside native forest habitats (Richardson & Bowles, 1964; Scott et al., 1986) but the richer avifaunas on Fiji and or New Guinea have a complement of species that frequent clearings, forest edge, and secondary growth as for most mainland native avifaunas (Pearson, 1977; Watling, 1983). As more of an island's area is con- verted to urban, agricultural and disturbed habitats or altered through the introduction of herbivores and exotic predators, most natives lose good living space while most introduced birds that frequent open and disturbed areas and have evolved in predator-rich areas gain habitat. All else being equal, small populations are more vulnerable than large ones (MacArthur & Wilson, 1967; Leigh, 1981; Goodman, 1987) and extinctions rates, where measured, generally increase with decreases in habitat area (Diamond, 1984; Case & Cody, 1987; Soul6 et al., 1998; Bolger et al., 1991). Beginning in aboriginal times, the conversion of native habitats, particularly at lower elevations, to disturbed habitats simul- taneously enhanced the success and persistence of intro- duced species, while decreasing population sizes and increasing extinction rates of native species. Thus habi- tat conversion and deterioration alone could produce a correlation between the number of extinct natives and the number of introduced species even without any direct cause or effect between birds in these two groups.
Probably adding to the scatter in this relationship are differences between regions in the degree of Euro- pean exploitation and relish for introducing exotic bird species. Madagascar has lost around 85% of its native habitats (Jolly et al., 1984) and at present has lost 13 native bird species and at least 13 of the 75 native mammal species (approximately 17°/,, of the native fauna; Jolly et al., 1984) yet it never had an organized acclimatization society introducing exotic birds and today it harbors only three naturalized exotic species (rock dove, common myna, and house sparrow). At the other extreme are small very isolated islands like Easter, or the Chagos group that had no or very few native land bird species to begin with. Here the number of extinctions must be zero or small, yet when humans broke the isolation barrier by introducing exotic birds some of them were successful. Still other deviations from the correlation in Fig. 2 may be due to incom- plete fossil and subfossil discovery. While the level of habitat deterioration in Fiji, Vanuatu, and most of the Solomons is mild relative to Hawaii or Tahiti, it also seems likely that these places may have had more pre- historic extinctions than are presently known judging from the ever-expanding number of extinct forms found elsewhere in the Pacific (Olson, 1984, 1989; Olson & James, 1984; Steadman, 1989b, 1991, 1992; Steadman et al., 1990, 1994; Pimm et al., 1994). The absence of

86 T. J. Case
described prehistoric extinctions in places like the Seychelles, Comoros, Azores, and Cape Verde Islands is simply because as yet there has been little attempt to search for them (Olson, 1984). Australia is another exception with a plethora of introduced species but only a single known extinction, the historically extinct paradise parrot. Additional extinct forms are known from the Pleistocene in Australia but none from the Holocene, although again one suspects that there may be many extinctions yet to be discovered (Pimm et al., 1994).
Thus the species numbers compiled here should be thought of as dynamic, subject to change by taxonomic revisions, new discoveries of extinct and extant forms, rediscoveries of species thought to have been extinct, and new introductions and invasions of exotics. Since I first assembled this data set over 10 years ago, many revisions and discoveries have required its continual amendment; yet the relationship between introductions and extinctions has increased with these new develop- ments rather than weakened. For example, Easter Island has four successful introductions but until recently, no known extinctions and no extant native land birds. Steadman et al. (1994) recently reported six subfossil extinct land birds that probably became extinct after human contact. In the analysis above, when I subjected the data to various screens, excluding locations where no prehistoric birds had been described, or eliminating locations with no extinctions or intro- ductions, conclusions were unaffected and probability levels stayed the same or even increased in some com- parisons in spite of lower sample size.
Other factors influencing introduction success Other factors influence both the immediate success and persistence of introduced species populations. A strong correlation exists between the number of successes and failures across the islands in Table 2, attesting to the role of persistent acclimatization societies in increasing species numbers despite high failure rates. Similarly, even after controlling for avian extinctions, and all other independent variables in Table 1 and 2, island area correlates positively with introduced species number.
While significant, the effect of island area on exotic species number is weak and is not a significant corre- late of relative success rate, possibly because island area can both help and hurt exotic success at different stages. In the early stages of the release of exotic birds, a large area will allow birds to disperse long distances from the release sites, diluting the initial release number over space and potentially making it more difficult for the released birds to find mates. Yet, if the birds suc- cessively pass this hurdle and their numbers grow, then a wealth of theory suggests that larger areas will allow larger absolute asymptotic population sizes, which in turn will increase the likelihood of the population's long-term persistence (Leigh, 1981; Diamond, 1984; Case & Cody, 1987; Goodman, 1987).
The number of native extinctions seems to be inte- grating the amount of native habitat loss, with the absolute area of the island, and the deterioration of remaining native habitat through the intrusion of exotic predators, herbivores, and parasites (van Riper et al., 1986; Atkinson, 1989). For example, both Hawaii and the USA have lost about 60% of native forest since human contact, but Hawaii has lost around 60% of its land birds while the continental USA has lost about 1%. Here any attempt to explain the differ- ences in extinction rates to specific biotic interactions are complicated by the vast difference in area between Hawaii and the USA, which alone will have a moderating affect on extinction rates (Richman et al., 1988). New Zealand and Great Britain are more similar in size but Britain is a land-bridge island with a richer avifauna and a diverse set of native predators. New Zealand has lost about 70% of its native forests since human con- tact while Great Britain has lost 93% in about the last 5000 years (King, 1984). Since human contact about 40% of the land and freshwater birds of New Zealand have become extinct but only 3°/,, of those in Great Britain (King, 1984 and Table 1). About one-third of the extinct New Zealand birds are various species of moas, most very large in body size and hunted for food by native Maoris. King (1984) ascribes additional losses in New Zealand birds to both habitat conversion and introduced ferrets, stoats, weasels, cats, and rats.
The variety of predators is not a significant correlate of the number of introductions or their relative success rate for the large set of islands studied here (Table 4 and the results from Table 2). Of course, my categorization of predation is arbitrary and may not capture the true predation risk for most birds in these various locations. I have experimented with different weighting schemes, for example weighting exotic predators higher than native predators, without any substantial affect on the conclusion, at least with respect to introduced species S u c c e s s .
Native bird species without an evolutionary exposure to exotic predators appear more vulnerable to exotic predators than their exotic counterparts (Atkinson, 1985; Engbring & Fritts, 1988). Atkinson (1985) has found that on islands with native rodents or land crabs introduced rats have caused fewer bird extinctions than on islands that were previously predator-free. Presum- ably bird species on islands with native predators have evolved effective predator escape behaviors that enable them to evade the introduced rats or perhaps life his- tory features which allow populations to persist in spite of high mortality. Because most introduced species come originally from predator-rich continental areas, they may be less susceptible to introduced predators than endemic predator-naive species. The presence of exotic predators (and parasites) might even enhance the success of introduced species compared to predator-free islands by moderating the competitive impact of natives on the less susceptible exotics.

Introductions and extinctions o f birds 87
Habitat distributions and local diversity While I find little support for the notion that rich avi- faunas in themselves repel the establishment of avian invaders at the level of whole islands or archipelagoes, interactions between established exotics and natives may be influencing habitat distributions of species in both sets within islands. One possible explanation for the striking inverse relationship between introduced species number and relative abundance and native species number and relative abundance is that local competition between species in these two sets limits their distribution and abundance. Alternatively sites may vary in subtle habitat features at the local and at the landscape scale; exotics and natives may be track- ing these habitat features differentially such that sites that are favorable for exotics are unfavorable for natives.
Since most native birds in the Tropical Pacific are primarily birds of closed canopy while most introduced species are birds of more open habitats, the bird species sets and their abundances might be expected to vary inversely across a range of habitats from open to closed. While all sites are categorized according to floral and vegetation characters, urban/suburban sites still differ in the proportion of native vs exotic plant- ings around houses and other buildings. Green (1984) found that the total amount of native vegetation at a site was the most influential factor governing the number of both native birds (with a positive affect) and exotic birds (with a negative affect) in suburban habitats around Melbourne. Similarly, native forests probably differ in subtle ways by the actions of exotic mammals and insects. Probably even more important than local habitat differences, forest sites will vary in important landscape-level features (like patterns of fragmentation and edge effects) that influence their access to exotics and the long-term persistence of exotic and native sub- populations within a larger metapopulation. Some for- est sites are simply subsections of continuous native forests while others are smaller forest fragments sur- rounded by disturbed habitats that harbor more exotic birds (Askins et al., 1987). Such near-by reservoirs of exotics might spill over to these forest fragments (Smallwood, 1994). Similarly, small landscape patches of urban/suburban habitats imbedded in relatively undisturbed forest might be expected to harbor fewer exotic and more native birds than sites located in expansive human-modified open habitat.
In this matter, local species diversity is responding to local factors in the form of competitive interactions and/or habitat variation, but also to regional and his- torical processes affecting the diversity of the source pool and the overall pattern of landscape alteration (Askins et al., 1987; Ricklefs & Schluter, 1993; Small- wood, 1994). Table 5 and Fig. 10 indicate that point (or alpha) diversity, whether in non-native habitats or native forest, is responding to both regional and local species variables.
Diamond and Veitch (1981) concluded from their study of the exotic and native birds in New Zealand that extinctions of native birds were not caused by competition from exotics but rather from habitat destruction, hunting, and introduced predators. Forest alteration by browsing and logging as well as a lack of competition from native birds may have then allowed the exotics to penetrate into forest habitats. It was impossible for them to assess the relative importance of these two factors. The conclu- sions of Mountainspring and Scott (1985), and Scott et al. (1986) for the Hawaiian situation are not dissimilar.
To distinguish between these two hypotheses to explain the differential penetrance of exotics into forest habitats, i.e. diffuse competition between natives and exotics, on the one hand, and differential responses of natives and exotics to habitat and landscape alterations on the other, controlled experiments are needed with careful attention to matching habitat features at both the local and landscape scale. The two hypotheses are not mutually exclusive and both may be operating to varying degrees in different locations.
Native species in non-native habitats Why do these various locations differ so strikingly in the numbers of native species that colonize urban/sub- urban habitats? In species-rich areas like North America or Australia, with much habitat diversity, the native avifauna contains species adapted to the variety of available habitats. Urban/suburban habitats contain native plant species as well as exotic introductions and the avifauna is similarly heterogeneous. A small book devoted to the common birds of urban Melbourne lists 35 species of native passerines and eight exotics that the lay person could commonly observe around the city (Reid et al., 1971). The ability of native species to occupy urban/suburban habitats where they coexist with exotics is typical for other continental avifaunas (Califomia: Vale & Vale, 1976; South Africa: Brooke et al., 1986; New- man, 1993; West Africa: Winterbottom, 1933; Malay penninsula: McClure, 1961; Ward, 1968; India: Beehler et al., 1987) and those on very large islands (New Guinea: Bell 1986; Cuba: Pozas & Balat, 1981).
Because of their remoteness and small size, islands typically have fewer native species than continents (MacArthur & Wilson, 1967). Smaller land masses in tropical areas also have less habitat diversity for two reasons, area and maximum elevation are positivley correlated (see Table 3). Islands with taller peaks gener- ally have greater topographic diversity. Also tall mas- sive mountains create strong windward vs leeward regional differences in climate on an island. Leeward habitats are generally more arid and more open. Con- sequently, small islands seem to contain proportionally fewer bird species that are pre-adapted to the generally open habitats associated with urbanization and agricul- ture but it is these species that are most likely colonists of urban/suburban habitats (Bell, 1986). Bell found that only about 3.2%-4.8% (depending on the city) of

88 T .J . Case
New Guinea rainforest bird species have colonized urban areas of Papua New Guinea. While a much greater proport ion of native savannah and forest edge species also occupy urban areas, Additionally, lowland habitats, where most urban centers are concentrated, have been particularly decimated on many islands (Per- netta & Watling, 1979; Olson & James, 1982; Moul ton & Pimm, 1983). Consequently, compared to higher ele- vation forest birds, few native lowland species are left in lowland disturbed habitats to have a potential share in lowland towns and cities with introduced species. In short, smaller, flatter, islands with greater lowland habitat destruction and urbanization have fewer potential native species to colonize urban centers.
This pattern comes into sharper focus by a contrast between Fiji, Hawaii, and the Societies. Although simi- lar in overall land mass, Kauai and Tahiti have experi- enced more extinctions and habitat destruction than has Viti Levu in Fiji. Today on Viti Levu twelve native species (representing 44% of the total native species of passerines and doves) commonly make use of disturbed modified habitats (Table 6). In Kauai and Tahiti native passerines and doves are essentially restricted to upland native forest. The overall dissimilarity of the avifaunas
in native and non-native habitats can be quantified using the dissimilarity measure z of Preston (1962):
xl/: + yl/: = 1 (1)
where x is the fraction of total species number that is found in native habitats and y the fraction found in non-native habitats. I f z = 1, its limiting value at one end of its range, the faunas are completely dissimilar. As z approaches zero, at the other extreme, there is no dissimilarity. Because no closed form analytic solution for z exists, Preston (1962) provides a table for its solution.
The values of z increase from Fiji to Tahiti. This quantifies the observation that habitat sharing of native and exotic birds is generally greater in areas with larger native avifaunas that have not suffered as extensive a loss of lowland habitats and their indigenous bird species.
Bird species numbers in each habitat type are proba- bly not yet in equilibrium for their area and degree of habitat conversion. This latter result was also seen by Bell (1986) who compared historical bird surveys to present-day results in New Guinea urban areas. Birds were gradually adjusting to new habitat formation and species occurrences did not appear to have reached an equilibrium.
Table 6. Habitat distributions of passerines and eolumbiformes relative to native and non-native habitats on three Pacific islands The number of species (and % of total species in the row heading) is given for each cell. The value of = is Preston's measure of the dissimilarity of two faunas, here applied to birds of native and non-native habitats (see text).
(a) Fiji - - Viti Levu (from Watling (1982) and personal surveys)
Extant native species = 27 Introd. species = 8
Species in only Species in native and Species only in native habitats non-native habitats non-native habitats
Native species 14/52% 12/44% 1/4% Exotic species 0/0% 3/43% 5/57%
z = 0.44.
(b) Hawaii - - Kauai (from Scott et al. (1986); Richardson & Bowles (1964) and personal surveys)
Extant native species = 13 Introd. species = 17
Species in only Species in native and Species only in native habitats non-native habitats non-native habitats
Native species 13t100% 010% 0t0% Exotic species 0/0% 9•53% 8/47%
= 0.612.
(c) Societies - - Tahiti (from Thibault & Rives (1975) and personal surveys)
Extant native species = 5 Introd. species = 9
Species in only Species in native and Species only in native habitats non-native habitats non-native habitats
Native species 4/80% 1/20% 0/0% Exotic species 0/0% 2/22% 7/78%
z = 0.72.

Introductions and ext&ctions o f birds 89
Is there an asymmetry in the interactions between exotics and natives? If present-day interspecific compe- tition is contributing to a redistribution of natives and exotic species across habitats, then the results of Table 5 suggest that exotics may be hindering native expan- sion into non-native habitats more than natives are preventing exotics from penetrating native habitats. The negative associations between natives and exotics are stronger in non-native habitats compared to native forest, but sample sizes are also larger giving more power to the comparision. Moreover, this conclusion is not supported by the results from New Zealand/ Australian surveys where the abundance of native birds has been more rigorously measured. Here native bird abundance is not significantly affected by either the number of exotic species or their summed numerical density in either native or non-native habitats. Yet, the abundance and diversity of exotics in Australia and New Zealand declines sharply with the number of native bird species, at least in non-native habitats (Fig. 12(b)). This last effect disappears in a stepwise regression, since native species number is not a signifi- cant variable accounting for variation in exotic abun- dance or diversity once the local number of exotic species is entered into the equation. In sum these results suggest that habitat variation and the regional source pools of exotics and natives provide additional explanatory power in accounting for local diversity and abundance patterns. Still caution is urged; more detailed species-specific studies are needed that compared the same exotic species in places that vary in their num- bers and types of competitors and predators.
on islands are generally staying similar through gains of exotics balancing losses of natives, worldwide the total number of bird species has declined and avifaunas are losing and continue to lose their regional distinctions.
In local surveys the relative abundance of exotic birds compared to native birds is affected by habitat (non-native habitats have more exotics) and also by the numbers of species of exotics and natives on the island. The island (or regional) species pool of exotic and native species, along with the island's overall degree of habitat alteration, affects the species diversity at the local scale and, in turn, the relative abundance of exotic birds and native birds in native and non-native habitats. The relative importance of biotic interactions (like competition, apparent competition through differ- ential disease transmission or susceptibility, and preda- tion) in shaping the abundance and habitat affinities of exotics and native species, can be difficult to unravel when regional affects are so important. Future studies will gain more mechanistic insights by focusing on a few exotic and native species and comparing their habi- tat distributions across a number of different archipela- goes, similar in climate and habitat but differing in native species composition and the level of habitat destruction and conversion.
A C K N O W L E D G E M E N T S
This work was supported by the National Science Foundation (grant DEB-9220621 and BSR-9107739). I thank Mike Gilpin, Mike Moulton, Stuart Pimm, Trevor Price and David Steadman for useful insights.
CONCLUSIONS AND FUTURE DIRECTIONS
All in all, about half the variance between islands in their numbers of successfully and unsuccessfully intro- duced species can be accounted for by recipient site- specific variables; the most important correlate of success is the number of native species extinctions, which reflects the degree of human activity and habitat destruction and deterioration through intrusions of exotic predators, herbivores, and parasites. Presumably some of the remaining variation can be explained by individual differences between the sets of species intro- duced from place to place, idiosyncrasies in the individ- ual methods of introduction for different species, other site specific factors not included in this analysis, and truly stochastic factors affecting the establishment and persistence of exotic species. All of these factors have been shown to affect introduction success in birds and other vertebrates (Hengeveld, 1989; Pimm, 1991).
Many of the same species that have been introduced successfully to one place have also been successfully introduced to others around the globe (Simberloff & Boecklen, 1991), while the losses of island birds from extirpation seem to have disproportionately fallen on endemic species. Consequently, while species numbers
REFERENCES
Adler, G. H. (1992). Endemism in birds of tropical Pacific islands. Evolut. Ecol., 6, 29f~306.
Ashmole, N. P. (1963). The extinct avifauna of St. Helena Island. Ibis', 1036, 390-408.
Atkinson, I. A. E. (1985). The spread of commensal species of Rattus to oceanic islands and their effects on island avi- faunas. In Conservation of island birds: case studies for the management of threatened island species, ed. P. J. Moors. International Council for Bird Preservation, Cam- bridge, pp. 35-81.
Atkinson, I. A. E. (1989). Introduced animals and extinc- tions. In Conservation for the twenty-first century, ed. D. Western & M. Pearl. Oxford University Press, Oxford, pp. 54-75.
Bacallado, J. J. (1976). Notas sobre la distribucion y evolu- cion de la avifauna Canaria. In Biogeography and ecology in the Canary Islands, ed. G. Kunkel. W. Junk, The Hague, pp. 413-32.
Baez, M. (1992). Zoogeography and evolution of the avi- fauna of the Canary Islands. In Papers in avian paleontol- ogy, ed. K. E. Campbell. Natural History Museum of Los Angeles, Los Angeles, CA, pp. 425-31.
Balmford, R. (1981). Early introductions of birds to Victoria. Vict. Nat., 98, 96-105.
Balouet, J. C. & Olson, S. L. (1989). Fossil birds from Late Quaternary deposits in New Caledonia. Smithson. Contrib. ZooL, 469, 1-38.

90 T. J. Case
Banks, R. C. (1976). Wildlife importation into the United States, 1900-1972. USDL Fish. Wildl. Serv., Spec. Scient. Rep., Wildl., No. 200.
Banks, R. C. (1981). Summary of foreign game bird libera- tions, 1969-1978. USDL Bur. Sport Fish. Wildl. Spec. Sci- ent. Rep., Wildl., No. 239.
Bannerman, D. A. (1963). Birds of the Atlantic Islands. Oliver & Boyd, London.
Bannerman, D. A. & Bannerman, W. M. (1983). The birds" of the Balearic. Croom Helm, London.
Beehler, B. M., Krishna Raju, K. S. R. & Ali, S. (1987). Avian use of man-disturbed forest habitats in the Eastern Ghats, India. Ibis, 129, 197-211.
Beehler, B. M., Pratt, T. K. & Zimmerman, D. A. (1986). Birds of New Guinea. Princeton University Press, Princeton, NJ.
Bell, H. L. (1986). Occupation of urban habitats by birds in New Guinea. Proc. West. Foundn Vert. Zool., 3, 1-48.
Benson, C. W. (1950). A contribution to the ornithology of St Helena and other notes from a sea voyage. Ibis, 92, 75-83.
Benson, C. W. (1960). The birds of the Comoro Islands: result of the British Ornithologists' Union expedition of 1958. Ibis, 103h, 5-106.
Benson, C. W. & Penny, M. J. (1971). The land birds of Aldabra. Phil Trans. R. Soc., Lond., 260, 417-527.
Berger, A. J. (1972). Hawaiian birdlife. University of Hawaii Press, Honolulu.
Blabar, S. J. M. (1990). A checklist and notes on the current status of the birds of New Georgia, West Province, Solomon Islands. Emu, 90, 205-14.
Bohl, W. H. & Bump, G. (1970) Summary of foreign game bird liberations 1960--1968. USDI Fish. WildL Serv., Spec. Scient. Rep., WildL, No. 130.
Bolger, D. T., Alberts, A. C. & Soulr, M. E. (1991). Occur- rence patterns of bird species in habitat fragment: sam- piing, extinction, and nested species subsets. Amer. Nat., 137, 155.
Bond, J. (1980). Birds of the West Indies. Houghton Mifflin, Boston, MA.
Bostic, D. L. (1975). A natural history guide to the Pacific of North Central Baja California and adjacent islands. Biology Education Expeditions, Vista, CA.
Bourne, W. R. P. (1957). Breeding birds of Bermuda. Ibis, 99, 94-105.
Bourne, W. R. P. (1966). Further notes on the birds of the Cape Verde Islands. Ibis, 108, 425-9.
Bregulla, H. L. (1992). Birds of Vanuatu. Nelson, Oswestry. Brattstrom, B. H. (1990). Biogeography of the Islas-Revillagi
Mexico. J. Biogeogr., 17, 177-83. Brooke, R. K., Lloyd, P. H. & de Villiers, A. L. (1986). Alien
translocated terrestrial vertebrates in South Africa. In The ecology and management of biological invasions in southern Africa, ed. I. A. W. Macdonald, F. J. Kruger & A. A. Ferrar. Oxford University Press, Cape Town, pp. 63-74.
Bruner, P. L. (1972). Field guide to the birds" of French Polyne- sia. Bernice P. Bishop Museum, Honolulu.
Bullock, D. (1977). Round Is land--a tale of destruction. Oryx, 14, 51-8
Bump, G. (1963). History and analysis of tetraonid introduc- tions into North America. J. Wildl. Manage., 27, 855-67.
Cain, A. J. & Galbraith, I. C. J. (1956). Field notes on birds of the eastern Solomon islands. Ibis, 99, 262-95.
Carr, G. S. (1980). Historic and prehistoric avian records from Easter island. Pacif. Sci., 34, 19.
Case, T. J. (1991). Invasion resistance, species build-up and community collapse in model competition communities. In Metapopulation dynamics, ed. M. E. Gilpin & I. Hanski, pp. 239-66.
Case, T. J. & Bolger, D. T. (1991). The role of introduced
species in shaping the abundance and distribution of island reptiles. Evolut. Ecol., 5, 272-90.
Case, T. J. & Cody, M. L. (1987). Island biogeographic theo- ries: tests on islands in the Sea of Cortez. Amer. Sci., 75, 402-11.
Case, T. J., Bolger, D, T. and Richman, A. (1992). Reptilian extinctions: The last ten thousand years. In Conservation biology, ed. P. L. & S. K. J. Fiedler. Chapman & Hall, New York, pp. 91-125.
Cassels, R. (1984). The role of prehistoric man in the faunal extinctions of New Zealand and other Pacific islands. In Quaternary extinctions, ed. P. S. Martin & R. G. Klein. University of Arizona Press, Tucson, AZ, pp. 708-67.
Caum, E. L. (1933). The exotic birds of Hawaii. Bernice P. Bishop Mus. Occ. Pap., 10, 1 55.
Cawkell, E. M. & Hamilton, J. F, (1961). The birds of the Falkland Islands. Ibis, 103, 1-27.
Cheke, A. S. (1987). An ecological history of the Mascarene Islands with particular reference to extinctions and intro- ductions of land vertebrates. In Studies of Mascarene Island birds, ed. A. W. Diamond & A. S. Cheke. Cambridge Uni- versity Press, Cambridge, pp. 5 89.
Clunie, F. (1984). Birds' of the Fiji bush. Fiji Museum, Suva, Fiji.
Cody, M. L. (1983). The land birds. In lslandbiogeography in the Sea of Cortez, ed. T. J. Case & M. L. Cody. University of California Press, Berkeley, CA, pp. 210-45.
Crowell, K. L. (1962). Reduced competition among the birds of Bermuda. Ecology, 43, 75-88.
Crusz, H. (1984). Parasites of the endemic and relict verte- brates: a biogeographical review. In Ecology and biogeogra- phy in Sri Lanka, ed. C. H. Fernando. W. Junk, The Hague, pp. 321-52.
Curtiss, A. (1938). A short zoology of Tahiti. Guide Printing Co., Brooklyn, N.Y.
Delacour, J. (1966). Guide des oiseaux de la Nouvelle-CalO- donie et de ses dependances. Delachaux and Niestle, Neuchatel.
Diamond, A. W. (1984). Biogeography of Seychelles land birds. In Biogeography and ecology of the Seychelles Islands, ed. D. R. Stoddard. W. Junk, The Hague, pp. 487-504.
Diamond, A. W. (1985). The conservation of landbirds on islands in the tropical Indian Ocean. ICBP Tech. Publs., 3, 85-100.
Diamond, J. (1985). Rats as agents of extermination. Nature, Lond., 318, 602-3.
Diamond, J. (1991). A new species of rail from the Solomon Islands and convergent evolution of insular flightlessness. Auk, 108, 461-70.
Diamond, J. M. (1984). Historic extinctions: a rosetta stone for understanding prehistoric extinctions. In Quaternary extinctions, ed. P. S. Martin & R. G. Klein. University of Arizona Press, Tucson, AZ, pp. 824-62.
Diamond, J. M. & Jones, H. L. (1980). Breeding land birds of the Channel Islands. In The California islands, ed. D. M. Power. Santa Barbara Museum of Natural History, Santa Barbara, CA.
Diamond, J. M. & Marshall, A. G. (1976). Origin of the New Hebridean avifauna. Emu, 76, 187-200.
Diamond, J. M. & Veitch, C. R. (1981). Extinctions and introductions in the New Zealand avifauna: cause and effect. Science, N. Y., 211,499-501.
Disney, D. H. J. & Stokes, A. (1976). Birds in pine and native forests. Emu, 76, 133-8.
Drake, J. A. (1983). Invasability in Lotka-Volterra interac- tion webs. In Current trends in food web theory report on a food web workshop, ed. D. L. DeAngelis, W. M. Post & G. Sugihara. National Technical Information Service, Oak Ridge, TN, pp. 83-90.

Introductions and extinctions o f birds 91
Drake, J. A. (1988). Models of community assembly and the structure of ecological landscapes. In Proceedings of the International Conference in Mathematical Ecology, ed. T. Hallam, L. Gross & S. Levin. World Press, Trieste.
Drake, J. A. (1991). Community assembly mechanics and the structure of an experimental species ensemble. Amer. Nat., 137, 1-26.
Drake~ J. A., Flum, T. E., Witteman, G. J., Voskuil, T., Hoylman, A. M., Creson, C., Kenny, D. A., Huxel, G. R., Larue, C. S. & Duncan, J. R. (1993). The construction and assembly of an ecological landscape. J. Animal EcoL, 62, 117-30.
Druett, J. (1983). Exotic intruders. The introduction of plants and animals into New Zealand. Heinemann, Auckland.
du Pont, J. E. (1976). South Pacific birds. Delaware Mus. Nat. Hist. Monogr., Ser. 3.
Duffey, E. (1964). The terrestrial ecology of Ascension Island. J. Appl. EcoL, 1, 219-51.
Ebenhard, T. (1988). Introduced birds and mammals and their ecological effects. Swed. Wildl. Res. Viltrevy, 13, 1-107.
Edgar, A. T. (1965). The Kermadecs expedition, 15-17 November 1964. Notornis, 12, 1-43.
Elliott, H. F. (1957). A contribution to the ornithology of the Tristan da Cunha group. Ibis, 99, 545-86.
Elton, C. S. (1958). The ecology of invasions by animals and plants. Chapman and Hall, London.
Engbring, J. (1988). FieM guide to the birds of Palau, Conser- vation office Palau, Koror, Palau.
Engbring, J. & Fritts, T. H. (1988). Demise of an insular avi- fauna: the brown tree snake on Guam. Trans. West. Sect. Wildl. Sot., 24, 31-7.
Falla, R. A., Sibson, R. B. & Turbott, E. G. (1983). The new guide to the birds of New Zealand and outlying islands. Collins, London.
Ffrench, R. (1980). A guide to the birds of Trinidad and Tobago. Harrowood Books, Newtown Square, PA.
Fleming, C. A. (1939). Birds of the Chatham Islands. Emu, 38, 380~13, 492-509.
Forbes, H. O. (1893). A list of the birds inhabiting the Chatham Islands. Ibis, 1893, 521-46.
Forbes-Watson, A. D. (1969). Notes on bireds observed in the Comoros on behalf of the Smithsonian Institution. Atoll Res. Bull., 128, 1-23.
Ford, H. (1979). Birds. In Natural history of Kangaroo Island, ed. M. J. Tyler, C. R. Twidale & J. R. Ring. Royal Society of South Australia, Adelaide, pp. 103-14.
Friend, G. R. (1982). Bird populations in exotic pine planta- tions and indigenous eucalypt forests in Gippsland, Victoria. Emu, 82, 80-91.
Gibb, J. A. (1961). Ecology of the birds of Kaingaroa forest. Proc. N.Z. Ecol. Soc., 8, 29-38.
Gill, F. B. (1967). Birds of Rodriquez Island. Ibis, 109, 383-90.
Goodman, D. (1987). The demography of chance extinction. In Viable populations for conservation, ed. M. E. Soulr. Cambridge University Press, Cambridge, New York, pp. ! 1-34.
Goodman, S. M. (1994). Description of a new species of subfossil eagle from Madagascar: Stephanoaetus (Aves: Falconiformes) from the deposits of Ampasambazimba. Proc. Biol. Soc. Wash., 107, 421-8.
Gorman, M. L. (1975). Habitats of the land birds of Viti Levu, Fiji Islands. Ibis, 117, 152-61.
Grant, P. R. & Cowan, I. M. (1964). A review of the avi- fauna of the Tres Marias Islands, Nayarit, Mexico. Condor, 66, 221-8.
Green, R. J. (1984). Native and exotic birds in a suburban habitat. Aust. Wildl. Res., 11, 181-90.
Greenway, J. L. (1967). Extinct and vanishing birds of the world. Dover Press, New York.
Guild, E. (1938). Tahitian aviculture: acclimatization of foreign birds. Avic. Mag., 5, 8-11.
Guild, E. (1940). Western bluebirds in Tahiti. Avic. Mag., 5, 284-5.
Hadden, D. (1981). Birds of the North Solomons. Wau Ecol- ogy Institute, Wau, Papua New Guinea.
Hannecart, F. & Letocart, Y. (1983). Oiseaux de Nouvelle Caledonie et des Loyautes.
Harris, M. P. (1973). The Galapagos avifauna. Condor, 75, 265-78.
Hart-Davis, D. (1972). Ascension--the story of a South Atlantic island. Constable, London.
Harting, J. E. (1873). Rare or little known Limicolae. Ibis, 3, 260-9.
Haydock, E. I. (1954). A survey of the birds of St Helena island. Ostrich, 25, 62-75.
Heilprin, A. (1889). The Bermuda Islands. Academy of Natu- ral Sciences, Philadelphia.
Heinzel, H., Fitter, R. & Parslow, J. (1972). The birds of Britain and Europe. Collins, London.
Henderson, R. W. (1992). Consequences of predator intro- ductions and habitat destruction on amphibians and reptiles in the post-Columbus West Indies. Caribb. J. Sci., 28, 1-10.
Hengeveld, R. (1989). Dynamics of biological invasions. Chap- man & Hall, London.
Henry, G, M. (1955). A guide to the birds of Ceylon. 2nd edn. Oxford University Press, London.
Hertlein, L. G. (1963). Contribution to the biogeography of Cocos Island including a bibliography. Proc. Calif. Acad. Sci., 32, 219-89.
Hindwood, K. A. (1940). The birds of Lord Howe Island. Emu, 40, 1-86.
Holdaway, R. N. (1990). Changes in the diversity of New Zealand forest birds. N.Z.J. Zool., 17, 309-21.
Holdaway, R. N. (1989). New Zealand's pre-human avifauna and its vulnerability. N.Z.J. Ecol., 12, 11-25.
Holdgate, M. W. (1960). The fauna of the mid-Atlantic islands--a discussion on the biology of the southern cold temperate zone. Proc. R. Soc. Lond., Ser. B, 152, 550~7.
Holloway, J. D. (1977). The lepidoptera of Norfold Island, their biogeography and ecology. W. Junk, The Hague.
Holmes, D. A. & Nash, S. V. (1989). The birds of Java and Bali. Oxford University Press, Oxford.
Holyoak, D. T. (1974). Undescribed land birds from the Cook Islands. Pacif Ocean. Bull. Brit. Ornithol. Club, 94, 145-50.
Horton, D. R. (1984). Red kangaroos: last of the Australian megafauna. In Quaternary extinctions, ed. P. S. Martin & R. G. Klein. University of Arizona Press, Tucson, AZ, pp. 639-80.
Hummelinck, P. W. (1940). Studies on the fauna of Curacao, Aruba, Bonaire, and the Venezuelan Islands, Vol. 1. den Boer, Utrecht.
Jehl, J. R. & Everett, W. T. (1985). History and status of the avifauna of Isla Guadalupe, Mexico. Trans. San Diego Soe. Nat. Hist., 20, 313-36.
Jehl, J. R. & Parkes, K. C. (1982). The status of the avifauna of the Revillagigedo Islands, Mexico. Wilson Bull., 94, 1-19.
Jenkins, J. M. (1983). The native forest birds of Guam. Ornithol. Monogr., 31, 1~1.
Johnson, S. W., Millie, W. R. & Moffet, G. (1970). Notes on the birds of Easter Island. Ibis, 112, 532-8.
Johnson, T. H. & Stattersfield, A. J. (1990). A global review of island endemic birds. Ibis, 132, 167-80.
Jolly, A., Oberlr, P. & Albignac, R. (1984). Madagascar (key environments). Pergamon, Oxford.

92 T. J. Case
Jones, H. L. & Diamond, J. M. (1976). Short-time-base stud- ies of turnover in breeding bird populations on the Califor- nia Channel Islands. Condor, 78, 526-49.
King, C. (1984). Immigrant killers: introduced predators and the conservation of birds in New Zealand. Oxford University Press, Auckland.
King, W. B. (1981). Endangered birds o f the world." ICBP bird red data book. Smithsonian Institution Press, Washington, DC.
Knox, G. A. (1969). The New Zealand off-shore islands. Micronesia, 5, 465-91.
Kramer, P. (1984). Man and other introduced animals. Biol. J. Linn. Soc., 21,253-8.
Lack, D. (1976). Island biology. University of California Press, Berkeley, CA.
Langrand, O. (1990). Guide to the birds o f Madagascar. Yale University Press, New Haven, CT.
Leigh, E. G. (1981). Average lifetime of a population in a varying environment. J. Theoret. Biol., 90, 213-39.
Lever, C. (1977). The naturalized mammals' o f the British Isles. Hutchinson, London.
Lever, C. (1985). Naturalized mammals of the world. Long- man, London.
Lever, C. (1987). Naturalized birds' o f the world. Longman, London.
Lionnet, G. (1984). Extinct birds of the Seychelles. In Bio- geography and ecology of the Seychelles Islands', ed. D. R. Stoddart. W. Junk, The Hague, pp. 505-11.
Lobin, W. (ed.) (1984). Fauna und Flora der Kapverdischen Inseln. Senckenbergische Naturforschende Gesselschaft, Frankfurt.
Lobin, W. (1986). Beitrag zur Fauna und Flora der Kapverdischen Inseln. Senckenbergische Naturforschende Gesselschaft, Frankfurt.
Lockwood, J. L. & Moulton, M. P. (1994). Ecomorphological pattern in Bermuda birds: the influence of competition and implications for nature preserves. Evolut. Ecol., 8, 53-60.
Lockwood, J. L., Moulton, M. P. & Anderson, S. K. (1993). Morphological assortment and the assembly of communi- ties of introduced passerines on oceanic islands: Tahiti vs Oahu. Amer. Nat., 141, 398-408.
Long, J. (1981). Introduced birds o f the world. David & Charles, London.
Losos, J. B., Marks, J. C. & Schoener, T. W. (1993). Habitat use and ecological interactions of an introduced and a native species of Anolis lizard on Grand Cayman, with a review of the outcomes of anole introductions. Oecologia, Berl., 95, 525-32.
Loustau-Lalanne, P. (1962). Birds of the Chagos archipelago, Indian Ocean. Ibis, 104, 67-73.
MacArthur, R. H. & Wilson, E. O. (1967). The theory of island biogeography. Princeton University Press, Princeton, NJ.
Marsh, L. & Pope, E. (1967). Norfolk and Lord Howe Islands. Aust. Nat. Hist., 15, 392-400.
Mayr, E. (1965). The nature of colonizations in birds. In The genetics o f colonizing species, ed. H. G. Baker & G. L. Stebbins. Academic Press, New York, pp. 30-45.
McClure, H. E. (1961). Garden birds in Kuala Lumpur, Malaya. Malay Nat. J., 15, 111-13.
McDowall, R. M. (1969). Extinction and endemism in New Zealand birds. Tuatara, 17, 1-12.
McKean, J. L. & Hindwood, K. A. (1965). Additional notes of the birds of Lord Howe Island. Emu, 64, 7%97.
McLay, C. L. (1974). The species diversity of New Zealand forest birds: some possible consequences of the modifica- tion of beech forests. N.Z.J . Zool., 1, 179-96.
Medway, L. & Marshall, A. G. (1975). Terrestrial vertebrates of the New Hebrides: origin and distribution. Trans. R.. Soc. Lond. B, 272, 423-65.
Milberg, P. & Tyrberg, T. (1993). Na'fve birds and noble savages--a review of man-caused prehistoric extinctions of island birds. Ecography, 16, 22%50.
Mittermeier, R. A. (1972). Jamaica's endangered species. Oryx, 11, 258-62.
Moreau, R. E. (1966). The bird faunas o f Africa and its islands. Academic Press, New York.
Moreau, R. E. & Pakenham, R. H. W. (1941). The land verte- brates of Pemba, Zanzibar, and Mafia: a zoogeographical study. Proc. Zool. Soc. Lond. Ser. A, 110, 97-128.
Moulton, M. P. (1993). The all-or-none pattern in introduced Hawaiian passeriformes: the role of competition sustained. Amer. Nat., 141, 105-19.
Moulton, M. P. & Pimm, S. L. (1983). The introduced Hawaiian avifauna: biogeographic evidence for competi- tion. Amer. Nat., 121,669 90.
Mountainspring, S. & Scott, J. M. (1985). Interspecific com- petition among Hawaiian forest birds. Ecol. Monogr., 55, 219-39.
Munro, G. G. (1944). Birds' of Hawaii. Tongg, Honolulu. Nelson, E. W. (1889). Natural biology of the Tres Marias
Islands: general account of the islands with reports on mammals and birds. N. Amer. Fauna, 14, 7-62.
Newman, K. (1993). Birds of South Africa, 4th edn. Southern Book Publishers, Johannesburg.
Newsome, A. E. & Noble, I. R. (1986). Ecological and physi- ological characters of invading species. In Ecology c?] biological invasions, ed. R. H. Groves & J. J. Burton. Cam- bridge University Press, Cambridge, pp. 1-20.
North, S. G., Bullock, D. J. & Dulloo, M. E. (1994). Changes in the vegetation and reptile populations on Round Island, Mauritius, following eradication of rabbits. Biol. Conserv., 67,21 8.
Olson, S. L. (1975). Paleoornithology of St. Helena Island, South Atlantic Ocean. Smithson. Contrib. Paleobiol., 23, 49.
Olson, S. L. (1984). Fossil birds of the islands of the world. Proc. lOlst Amer. Ornithol. Soc. Mtg.
Olson, S. L. (1989). Extinction on islands: man as a catastrophe. In Conservation for the twenty-first century, ed. D. Western & M. Pearl. Oxford University Press, New York, pp. 50-3.
Olson, S. L. & Hilgartner, W. B. (1982). Fossil and subfossil birds from the Bahamas. Smithson. Contrib. Paleobiol,, 48, 22-60.
Olson, S. L. & James, H. F. (1984). The role of Polynesians in the extinction of the avifauna of the Hawaiian islands. In Quaternary extinctions, ed. P. S. Martin & R. G. Klein. University of Arizona Press, Tucson, AZ, pp. 768 80.
Olson, S. L. & James, H. G. (1982). Prodromus of the fossil avifauna of the Hawaiian Islands. Smithson. Contrib. Zool., 365, 1-59.
Olson, S. L., Pregill, G. K. & Hilgartner, W. B. (1990). Stud- ies on fossil and extant vertebrates from San Salvador (Watling's) Island, Bahamas. Smithson. Contrib. Zool., 508, 1-15.
Owen, R. P. (1977a). A checklist of the birds of Micronesia. Micronesia, 13, 65-81.
Owen, R. P. (1977b). Terrestrial vertebrate fauna of the Palau Islands. Report from the Office of the Chief Conservation- ist, Trust Terr. Pacif., pp. 1-15.
Paynter, R. A. (1955). The ornitho-geography of the Yucatan Peninsula. Bull. Yale Peabody Mus. Nat. Hist., 9, 1 347.
Peake, J. F. (1971). The evolution of terrestrial faunas in the western Indian Ocean. Phil, Trans. R. Soc. Lond., B, 260, 581-610.
Pearson, D. L. (1977). A pantropical comparison of bird community structure on six lowland forest sites. Condor, 79, 232M4.
Penny, M. (1974). The birds o f the Seychelles and the outlying islands. Collins, London.

Introductions and ext inct ions o f birds 93
Pernetta, J. C. & Watling, D. (1979). The introduced and native terrestrial vertebrates of Fiji Pacific. Science, N.Y., 32, 223~14.
Petren, K. and Case, T. J. (1996). An experimental demon- stration of exploitation competition in an ongoing inva- sion. Ecology (in press).
Philibosian, R. & Yntema, J. A. (1977). Annotated checklist o f the birds', mammals, reptiles and amphibians o f the Virgin Islands and Puerto Rico. Information Services, St. Croix, US Virgin Islands.
Phillips, J. C. (1928). Wild birds introduced or transplanted in North America. US Dep. Agric. Tech. Bull., 61, 63.
Phillips, W. W. A. (1953). A revised checklis't o f the birds o f Ceylon. Natural History Museum of Ceylon.
Pickering, R. (1981a). An annotated checklist of the birds of Vanuatu. Misc. Pubis" Vanuatu Nat. Sci. Soc., 1, 1 8.
Pickering, R. (1981b). The birds of Vanuatu, 1. The intro- duced species. Naika, 2, 10-12.
Pimm, S. L. (1991). The balance o f nature? Ecological issues in the conservation o f species and communities. University of Chicago Press, Chicago, IL.
Pimm, S. L., Moulton, M. P. & Justice, L. J. (1994). Bird extinctions in the central Pacific. Phil. Trans. R. Soc. Lond., B, 344, 27-33.
Pittingill, O. S. (1973). Passerine birds of the Falkland Islands: their behavior and ecology. Living Bird, 12, 95 136.
Pizzey, G. & Doyle, R. (1980). A field guide to the birds o f Australia. Princeton University Press, Princeton, NJ.
Post, W. M. & Pimm, S. L. (1983). Community assembly and food web stability. Math. Biosci., 64, 169 92.
Power, D. M. (1972). Numbers of bird species on the Califor- nia Islands. Evolution, 26, 451-63.
Pozas, G. & Balat, F. (1981). Quantitative and qualitative composition of the bird community in a garden suburb of Havana (Cuba). Folia Zool., 30, 155-66.
Pratt, H. D., Bruner, P. L. & Berrett, D. G. (1987). The birds" o f Hawaii and the tropical Pacific. Princeton University Press, Princeton, NJ.
Pregill, G. K. (1981). An appraisal of the vicariance hypothesis of Caribbean biogeography and its application to West Indian terrestial vertebrates. Syst. Zool., 30, 147-55.
Pregill, G. K. (1982). Fossil amphibians and reptiles from New Providence Island, Bahamas. Smithson. Contrib. Paleobiol., 48, 8-21.
Pregill, G. D., Crombie, R. I., Steadman, D. W., Gordon, L. K., Davis, F. W. & Hilgartner, W. B. (1991). Living and late Holocene fossil vertebrates, and the vegetation of the Cockpit Country, Jamaica. Atoll. Res. Bull., 353, 1-19.
Preston, F. W. (1962). The canonical distribution of com- monness and rarity, Part I. Ecology, 43, 410-32.
Raffaele, H. A. (1983). A guide to the birds o f Puerto Rico and the Virgin Islands. Fondo Educativo Interamicano, San Juan, Puerto Rico.
Raffaele, H. A. (1989). The ecology of native and introduced granivorous birds in Puerto Rico. In Biogeography o f the West Indies, ed. C. A. Woods. Sandhill Crane Press, Gainesville, FL, pp. 541-66.
Rand, A. L. (1936). The distribution and habits of Madagas- car birds. Bull. Amer. Mus. Nat. Hist., 72, 143499.
Rand, A. L. & Gilliard, E. T. (1968). Handbook o f New Guinea birds'. Natural History Press, Garden City, New York.
Recher, H. F. (1977). Colonization and extinction: the birds of Lord Howe Island. In Lord Howe Island, ed. N. Smith, Australian Museum, Sydney.
Reid, A. J., Shaw, N. J. & Wheeler, W. R. (1971). Birds o f Victoria urban areas, 2nd edn. Gould League of Victoria, Melbourne.
Rejm~nek, M. (1989). Invasibility of plant communities. In Biological invasions." a global perspective, ed. J. A. Drake, H. A. Mooney, F. di Castri, R. H. Groves, F. J. Kruger, M. Rejm~inek & M. Williamson. John Wiley, New York, pp. 369-88.
Reynolds, R. T., Scott, J. M. & Nussbaum, R. A.. (1980). A variable circular-plot method for estimating bird numbers. Condor, 82, 309-13.
Richardson, F. & Bowles, J. (1964). A survey of the birds of Kauai, Hawaii. Bernice P. Bishop Mus. Bull., 227, 1-51.
Richman, A. D., Case, T. J. & Schwaner, T. D. (1988). Natu- ral and unnatural extinction rates of reptiles on islands. Amer. Nat., 131,611 30.
Ricklefs, R. E. & Schluter, D. (1993). Species diversity: regional and historical influences. In Species diversity in ecological communities." historical and geographical perspec- tives, ed. R. E. Ricklefs & D. Schluter. University of Chicago Press, Chicago, IL, pp. 350-63.
Ridpath, M. G. & Moreau, R. E. (1966). The birds of Tas- mania: ecology and evolution. Ibis, 108, 348-93.
Ripley, S. D. & Beehler, B. M. (1989). Ornithogeographic affinities of the Andaman and Nicobar Islands. J. Bio- geogr., 16, 323-32.
Robertson, C. J. R. (1985). Complete book o f New Zealand birds'. Readers Digest Services, Sydney.
Robinson, J. V. & Dickerson Jr., J. E. (1984). Testing the invulnerability of laboratory island communities to inva- sion. Oecologia, Berl., 61, 169 74.
Rolls, E. C. (1984). They all ran wild. Angus & Robertson, London.
Ryan, C. S. (1906). President's address: on European and other birds liberated in Victoria. Emu, 5, 110-19.
Saunders, D. A. & Rebeira, D. C. P. (1983). The birds of Rottnest Island. J. R. Soc. West. Aust., 66, 47-52.
Schodde, R., Fullagar, P. & Hermes, N. (1985). A review of Norfolk Island birds: past and present. Aust. Natn. Parks and Wildl. Serv., Spec. Publs., No. 8.
Schwartz, C. W. & Schwartz, E. R. (1949). The game birds in Hawaii. Board of Commissioners of Agriculture and Forestry, Honolulu.
Scott, J. M., Mountainspring, S., Ramsey, F. L. & Kepler, C. B. (1986). Forest bird communities o f the Hawaiian Islands'." their dynamics, ecology, and conservation. Cooper Ornithological Society, Las Cruces, New Mexico.
Scott, S. L. (1987). Field guide to the birds o f North America, 2nd edn. National Geographic Society, Washington, DC.
Severinghaus, S. R. & Blackshaw, K. T. (1976). A new guide to the birds" o f Taiwan. Mei Ya, Taipei.
Sibley, C. G. & Monroe, B. L. (1990). Distribution and taxon- omy o f birds o f the world. Yale University Press, New Haven, CT.
Simberloff, D. (1981). Community effects of introduced species. In Biotic crises in ecological and evolutionary time, ed. M. H. Nitecki. Academic Press, New York, pp. 53-81.
Simberloff, D. (1990). Community effects of biological intro- ductions and their implications for restoration. In Eco- logical restoration o f New Zealand islands, ed. D. R. Towns, C. H. Daugherty & I. A. E. Atkinson. Department of Conservation, Wellington, pp. 128-36.
Simberloff, D. (1992). Extinction, survival, and effects of birds introduced to the Mascarenes. Acta Oecologica, 13, 663-78.
Simberloff, D. & Boecklen, W. (1991). Patterns of extinction in the introduced Hawaiian avifauna: a reexamination of the role of competition. Amer. Nat., 138, 300-27.
Skottsberg, C. (1956). The natural history o f Juan Fernandez and Eastern Island. Almqvist & Wicksells, Uppsala.
Slud, P. (1967). The birds of Cocos Island (Costa Rica). Bull. Amer. Mus. Nat. Hist., 134, 261-96.

94 T. J. Case
Smallwood, K. S. (1994). Site invasibility by exotic birds and mammals. Biol. Conserv., 69, 251-9.
Smith, K. D. (1965). On the birds of Morocco. Ibis, 107, 493-526.
Smythies, B. (1968). Birds of Borneo, 2nd edn. Oliver & Boyd, London.
Soper, M. F. (1968). The Kermadec Islands. Animals, 11, 112-19.
Soul6, M. E., Bolger, D. T., Alberts, A. E., Wright, J. H., Sorice, M. & Hill, S. (1988). Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conserv. Biol., 2, 75-92.
Stager, K. E. (1957). The avifauna of the Tres Marias Islands, Mexico. Auk, 74, 413-32.
Steadman, D. W. (1986). Holocene vertebrate fossils from Isla Floreana, Galapagos. Smithson. Contrib. Zool., No. 413.
Steadman, D. W. (1988). A new species of Porphyrio (Aves: Rallidae) from archaeological sites in the Marquesas Islands. Proc. Biol. Soc. Wash., 101, 162-70.
Steadman, D. W. (1989a). Extinction of birds in Eastern Polynesia: a review of the record and comparisons with other Pacific island groups. J. Archaeol. Sci., 16, 177-205.
Steadman, D. W. (1989b). A new species of starling (Sturnidae, Aplonis) from an archaeological site on Huahine, Society Islands. Notornis, 36, 1614.
Steadman, D. W. (1991). Extinct and extirpated birds from Aitutaki and Atiu, Southern Cook Islands. Pacif Sci., 45, 325-47.
Steadman, D. W. (1992). Extinct and extirpated birds from Rota, Mariana Islands. Micronesia, 25, 71-84.
Steadman, D. W. (1993). Bird bones from the To'aga site: prehistoric loss of seabirds and megapodes. In The To'aga site: three millennia o f Polynesian occupation in the Manu'a islands, American Samoa, ed. P. V. Kirch & T. L. Hunt. University of California Archaeological Research Facility, Berkeley, CA, pp. 217-28.
Steadman, D. W. & Kirch, P. V. (1990). Prehistoric extinc- tion of birds on Mangaia, Cook Islands, Polynesia. Proc. Natl. Acad. Sci., USA, 87, 9605-9.
Steadman, D. W. & Martin, P. S. (1984). Extinction of birds in the late Pleistocene of North America. In Quaternary extinctions: a prehistoric revolution, ed. P. S. Martin & R. G. Klein. University of Arizona Press, Tucson, AZ, pp. 466-77.
Steadman, D. W. & Olson, S. L. (1985). Bird remains from an archaeological site on Henderson Island, South Pacific: man-caused extinctions on an 'uninhabited' island. Proc. Natn. Acad. Sci., 82, 6191-5.
Steadman, D. W. & Zarriello, M. C. (1987). Two new species of parrots (Aves: Psittacidae) from archeological sites in the Marquesas Islands. Proc. Biol. Soc. Wash., 100, 518-28.
Steadman, D. W., Pahlavan, D. S. & Kirch, P. V. (1990). Extinction, biogeography, and human exploitation of birds on Tikopia and Anuta, Polynesian outliers in the Solomon Islands. Bishop Mus. Occ. Pap., 30, 118-53.
Steadman, D. W., Casanova, P. V. & Ferrando, C. C. (1994). Stratigraphy, chronology, and cultural context of an early faunal assemblage from Easter Island. Asian Perspect., 33, 79-96.
Stoddart, D. R. (1971). Settlement, developoment and con- servation of Aldabra. Phil. Trans. R. Soc. Lond., B, 260, 611-28.
Stonehouse, B. (1962). Ascension Island and the British Ornithologists Union Centenary Expedition 1967-69. Ibis, 103b, 107-23.
Temple, S. (1981). Applied island biogeography and the con- servation of endangered island birds in the Indian Ocean. Biol. Conserv., 20, 147-61.
Thibault, J. C. & Rives, C. (1975). Oiseaux de Tahiti. Editions du Pacifique, Papeete, Tahiti.
Thomson, G. M. (1922). The naturalisation of animals and plants in New Zealand. Cambridge University Press., Cambridge.
Tikader, B. K. & Das, A. K. (1985). Glimpses o f animal life of Andaman and Nicobar Islands. Zoological Survey of India, Calcutta.
Turbott, E. G. (1961). The interactions of native and intro- duced birds in New Zealand. Proc. N.Z. Ecol. Soc., 8, 62-6.
Turbott, E. G. (1963). Three Kings Islands, New Zealand. A study in modification and regeneration. In Pacific Basin biogeography, ed. J. L. Gressitt. Bishop Museum Press, Honolulu, pp. 485-598.
Turbott, E. G. & Buddle, G. A. (1948). Birds of the Three Kings Islands. Rec. Auckland Inst. Mus., 3, 319-36.
Turner, J. S.,Smithers, C. N. & Hoogland, R. D. (1968). The conservation of Norfolk Island. Aust. Conserv. Foundn. Spec. Publs., No. 1.
Vale, T. R. & Vale, G. R. (1976). Suburban bird populations in west-central California. J. Biogeogr., 3, 157 65.
van Riper, C., III, van Riper, S. G., Goff, M. L. & Laird, M. (1986). The epizootiology and ecological significance of malaria on the birds of Hawaii. Ecol. Monogr., 56, 327-44.
Vesey-Fitzgerald, D. (1940). The birds of the Seychelles. Ibis', 4, 480-9.
Voous, K. H. (1957). The birds of Aruba, Curacao and Bonaire. Stud. Fauna Curacao, Aruba, Bonaire & Other Carib. Isl., No. 7.
Voous, K. H. (1983). Birds of the Netherlands Antilles, 2nd edn. De Walburg Pers, Utrecht.
Wall, L. E. (1983). Birds in Tasmanian pine plantations. Proc. R. Soc. Tasmania, 117, 125-33.
Walters, M. (1988). Probable validity of Rallus nigra Miller, an extinct species from Tahiti. Notornis, 35, 265-9.
Ward, P. (1968). Origin of the avifauna opf urban and subur- ban Singapore. Ibis, 110, 238-55.
Watling, D. (1982). Birds o f FO'L Tonga, and Samoa. Mill- wood Press, Wellington.
Watson, G. E., Zusi, R. L. & Storer, R. W. (1963). Prelimi- nary fieM guide to the birds o f the Indian Ocean. US National Museum, Smithsonian, Washington, DC.
Wetmore, A. (1960). Pleistocene birds in Bermuda. Smithso- nian Misc. Collec., 140, 1-11.
Williams, G. R. (1973). The natural history o f New Zealand. A. H. and A. W. Reed, Wellington.
Williams, W. D. (1974). Biogeography and ecology in Tasma- nia. W. Junk, The Hague.
Winterbottom, J. M. (1933). Bird population studies: a pre- liminary analysis of the Gold Coast avifauna. J. Anita. Ecol., 2, 52 97.
Wodzicki, K. & Wright, S. (1984). Introduced birds and mammals in New Zealand and their effect on the environ- ment. Tuatara, 27, 77-104.
Wolff, T. (1967). The fauna of Rennell and Bellona, Solomon Islands. Phil. Trans. R. Soc. Lond. B., 255, 321-43.
Wood, C. A. & Wetmore, A. (1925). A collection of birds from the Fiji Islands. Ibis, 55, 814-55.
Wood, C. A. & Wetmore, A. (1926). A collection of birds from the Fiji Islands, Part III. Ibis, 56, 91-136.
Woods, R. W. (1975). The birds o f the Falkland Islands. Anthony Nelson, Oswestry.

Introduct ions and ext inct ions o f birds 95
APPENDIX 1.
References to avifaunas, extinct species and avian introductions for land and freshwater birds by location
Island/mainland References
Aldabra
Andamans/Nicobars Aruba Ascension Auckland Australia Azores Bahamas
Balearic Bermuda
Borneo California Channel Canaries Cape Verde Chagos Chatham Christmas Cocos Comoros Cooks
Corsica-Sardinia Cozumel Curacao Easter
Falklands Fiji
Galapagos Great Britain Guadalupe Mex. Hawaii
Henderson Jamaica
Java Juan Fernandez Kangaroo
Kermadec Lord Howe
Madagascar
Marianas (Guam) Marquesas
Watson et al. (1963); Duffey (1964); Moreau (1966); Benson (1967); Benson & Penny (1971); Hart Davis (1972); Penny (1974); Diamond (1984) Tikader & Das (1985); Ripley & Beehler (1989) Hummelinck (1940); Voous (1957, 1983); Pregill (1981); Olson (1984) Stonehouse (1962); Watson et al. (1963); Duffey (1964); Greenway (1967); Hart-Davis (1972) Greenway (1967); Knox (1969); Cassels (1989) Ryan (1906); Pizzey & Doyle (1980); Balmford (1981); Horton (1984); Rolls (1984) Bannerman (1963) Buden & Schwartz (1968); Bond (1980); Olson & Hilgartner (1982); Pregill (1982); Olson et al. (1990); Milberg & Tyrberg (1993) Ridpath & Moreau (1966); Bannerman & Bannerman (1983) Bourne (1957); Wetmore (1960); Crowell (1962); Pregill (1981); Olson (1984); Heilprin (1889); Lockwood & Moulton (1994) Smythies (1968) Power (1972); Jones & Diamond (1976); Diamond & Jones (1980); Milberg & Tyrberg (1993) Bannerman (1963); Smith (1965); Bacallado (1976); Baez (1992); Milberg & Tyrberg (1993) Bourne (1966); Moreau (1966); Lobin (1984, 1986) Loustau-Lalanne (1962); Watson et al. (1963); Stoddart (1971); Diamond (1984) Forbes (1893); Fleming (1939); Greenway (1967); Knox (1969); Cassels (1984) Watson et al. (1963) Hertlein (1963); Slud (1967) Benson (1960); Watson et al. (1963); Forbes-Watson (1969); Diamond (1984) Holyoak (1974); Steadman (1986, 1991); Pratt et al. (1987); Steadman & Kirch (1990); Milberg & Tyrberg (1993) Heinzel et al. (1972); Milberg & Tyrberg (1993) Paynter (1955) Hummelinck (1940); Voous (1957, 1983); Olson (1984) Stottsberg (1956); Johnson et al. (1970); Carr (1980); Milberg & Tyrberg (1993); Steadman et
al. (1994) Cawkell & Hamilton (1961); Pittingill (1973); Woods (1975) Wood & Wetmore (1925,1926); Gorman (1975); Pernetta & Watling (1979); Watling (1982); Clunie (1984); Pratt et al. (1987); Steadman (1989b) Harris (1973); Olson (1984); Kramer (1984); Steadman (1986) Ridpath & Moreau (1966); Heinzel et al. (1972); Lever (1977) Bostic (1975); Jehl & Everett (1985) Caum (1933); Munro (1944); Schwartz & Schwartz (1949); Berger (1972); Moulton & Pimm (1983); Olson & James (1982, 1984); Pratt et al. (1987); Simberloff & Boecklen (1991); Moulton (1993) Fosberg et al. (1983); Steadman & Olson (1985); Pratt et al. (1987); Milberg & Tyrberg (1993) Mittermeier (1972); Lack (1976); Bond (1980); Pregill (1981); Pregill et al. (1991); Milberg & Tyrberg (1993) Holmes & Nash (1989) Skottsberg (1956); Moreau (1966) Edgar (1965); Greenway (1967); Soper (1968); Ford (1979); Pizzey & Doyle (1980); Cassals (1984); Newsome & Noble (1986) Edgar (1965); Greenway (1967); Soper (1968); Knox (1969) Hindwood (1940); McKean & Hindwood (1965); Greenway (1967); Marsh & Pope (1967); Recher (1974) Rand (1936); Watson et al. (1963); Diamond (1984); Langrand (1990); Milberg & Tyrberg (1993); Goodman (1994) Owen (1977a,b); Jenkins (1983); Pratt et al. (1987); Engbring & Fritts (1988); Steadman (1992) Bruner (1972); Pratt et al. (1987); Steadman & ZarrieUo (1987); Steadman (1988); Milberg & Tyrberg (1993)

96 T. J. Case
Mauritius
New Caledonia New Guinea New Zealand
Norfolk Palau Pemba Puerto Rico
Reunion Revillagigedo Rodriguez Rottnest Saint Helena
Samoa Sea of Cortez Seychelles
Societies (Tahiti)
Solomons
Sri Lanka Taiwan Tasmania Three Kings Tres Marias Trinidad Tristan de Cunha Tuamotus United States
Vanuatu
Wake Zanzibar General
Watson et al. (1963); Greenway (1967); Bullock (1977); Diamond (1984); Cheke (1987); Simberloff (1992) Delacour (1966); Hannecart & Letocart (1983); Balouet & Olson (1989) Rand & Gilliard (1968); Diamond & Marshall (1976); Beehler et al. (1986); Bell (1986) Thomson (1922); Turbott (1961); Williams (1973); Diamond & Veitch (1981); Druett (1983); Falla et al. (1983); Cassels (1984); Wodzicki & Wright (1984); Robertson (1985) Marsh & Pope (1967); Turner et al. (1968); Holloway (1977); Schodde et al. (1985) Owen (1977a,b); Pratt et al. (1987); Engbring (1988) Moreau & Pakenham (1941); Moreau (1966) Bond (1961); Lack (1976); Philibosian & Yntema (1977); Schwartz (1978); Pregill (1981); Raffaele (1983, 1989); Milberg & Tyrberg (1993) see Rodriguez and Mauritius Jehl & Parkes (1982); Brattstrom (1990) Watson et al. (1963); Moreau (1966); Gill (1967); Diamond (1985); Cheke (1987); Simberloff (1992) Pizzey & Doyle (1980); Saunders & de Rebeira (1983) Harting (1873); Benson (1950); Haydock (1954); Ashmole (1963); Greenway (1967); Olson (1975, 1984) Watling (1982); Pratt et al. (1987); Milberg & Tyrberg (1993); Steadman (1993) Cody (1983) Vesey-Fitzgerald (1940); Watson et al. (1963); Peake (1971): Penny (1974); Diamond (1984, 1985); Lionnet (1984) Curtiss (1938); Guild (1938, 1940); Delacour (1966); Bruner (1972); Thibault & Rives (1975); Pratt et al. (1987); Waiters (1988); Steadman (1989a); Lockwood et al. (1993); Milberg & Tyrberg (1993) Wolff (1967); Diamond & Marshall (1976); Hadden (1981); Blabar (1990); Diamond (1991); Milberg & Tyrberg (1993) Phillips (1953); Henry (1971); Ridpath & Moreau (1966); Crusz (1984) Severinghaus & Blackshaw (1976) Ridpath & Moreau (1966); Williams (1974); Pizzey & Doyle (1980) Turbott & Buddle (1948); Turbott (1963); Knox (1969) Stager (1957), Grant & Cowan (1964); Nelson (1989) Ffrench (1980) Elliott (1957); Holdgate (1960) Bruner (1972); DuPont (1976); Pratt et al. (1987) Phillips (1928); Bump (1963); Mayr (1965); Bohl & Bump (1970); Scott (1987); Banks (1976, 1981); Steadman & Martin (1984) Cain & Galbraith (1957); Medway & Marshall (1975); Marshall (1976); Pickering (1981a,b); Diamond & Bregulla (1992) Greenway (1967); Owen (1977a,b); Pratt et al. (1987) Moreau & Pakenham (1941); Watson et al. (1963); Moreau (1966) King (1981); Long (1981); Atkinson (1985, 1989); Lever (1985, 1987); Johnson & Stattersfield (1990); Sibley & Monroe (1990)
Related Documents











