Elevated prodynorphin expression associated with ethanol withdrawal convulsions A.S. Beadles-Bohling a,b,c,d, *, J.C. Crabbe a,b,c,d , K.M. Wiren a,b,c,d a Portland Alcohol Research Center, Research Service P3-R&D-39, 3710 SW US Veteran’s Hospital Road, Oregon Health Sciences University, Portland, OR 97201, USA b Neuroscience Program, Research Service P3-R&D-39, 3710 SW US Veteran’s Hospital Road, Oregon Health Sciences University, Portland, OR 97201, USA c Department of Behavioral Neuroscience, Research Service P3-R&D-39, 3710 SW US Veteran’s Hospital Road, Oregon Health Services University, Portland, OR 97201, USA d Veteran’s Aairs Medical Center, Research Service P3-R&D-39, 3710 SW US Veteran’s Hospital Road, Oregon Health Sciences University, Portland, OR 97201, USA Abstract The hypothesis that k-opioid system activity may in part mediate convulsions exhibited during ethanol withdrawal was tested by exposing Withdrawal Seizure-Prone (WSP) and Withdrawal Seizure-Resistant (WSR) mice to chronic ethanol. Whole brain was harvested for RNA isolation and prodynorphin mRNA steady-state levels in whole brain were examined using Northern blot analysis. The data revealed significantly increased levels of prodynorphin mRNA expression in mice susceptible to ethanol withdrawal convulsions after withdrawal, with no corresponding increase in prodynorphin steady-state levels in mice resistant to ethanol withdrawal convulsions. These findings were not due to basal dierences in prodynorphin expression between the WSP and WSR mice. To verify that the dierences observed were not due to an ethanol-induced global alteration in gene transcription, mRNA levels of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase were measured. Glyceraldehyde-3-phosphate dehydrogenase expression was unchanged following both chronic exposure to ethanol and chronic exposure followed by withdrawal. These results extend our understanding of prodynorphin’s role in generalized seizure activity to include ethanol withdrawal-induced convulsions. Our findings suggest that prodynorphin expression is modulated during ethanol withdrawal convulsions, or alternatively, prodynorphin may mediate the severity of ethanol withdrawal convulsions. 7 2000 Elsevier Science Ltd. All rights reserved. Keywords: Prodynorphin; Withdrawal; Convulsions; Ethanol; Aversion; Selected lines; k-Opioid receptors; Genetics 1. Introduction Ethanol withdrawal is a physiological response to the absence of ethanol that is exhibited after an organ- ism has become physically dependent upon alcohol. This response is believed to be a physical adaptation that the nervous system undergoes in the presence of ethanol (Himmelsbach, 1942) and is included in the National Institute on Alcohol Abuse and Alcoholism’s definition of alcoholism. Withdrawal can be displayed in various forms, and in humans can include symp- toms such as nausea, sweating, shakiness and anxiety (Victor, 1990; Victor and Adams, 1953), (http://sil- k.nih.gov/silks/niaaa1/publication/booklet.htm). In ad- dition, estimates suggest that 4–7% of alcoholics (Hauser, 1990) suer from seizures that can be life threatening and even fatal; therefore, it is important that we understand the mechanisms underlying ethanol dependence and withdrawal. The purpose of this study Neurochemistry International 37 (2000) 463–472 0197-0186/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved. PII: S0197-0186(00)00056-5 www.elsevier.com/locate/neuint * Corresponding author. Tel.: +1-503-220-8262; fax: +1-503-273- 5351. E-mail address: [email protected] (A.S. Beadles-Bohling).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Elevated prodynorphin expression associated with ethanolwithdrawal convulsions
A.S. Beadles-Bohlinga,b,c,d,*, J.C. Crabbea,b,c,d, K.M. Wirena,b,c,d
aPortland Alcohol Research Center, Research Service P3-R&D-39, 3710 SW US Veteran's Hospital Road, Oregon Health Sciences University,
Portland, OR 97201, USAbNeuroscience Program, Research Service P3-R&D-39, 3710 SW US Veteran's Hospital Road, Oregon Health Sciences University, Portland,
OR 97201, USAcDepartment of Behavioral Neuroscience, Research Service P3-R&D-39, 3710 SW US Veteran's Hospital Road, Oregon Health Services University,
Portland, OR 97201, USAdVeteran's A�airs Medical Center, Research Service P3-R&D-39, 3710 SW US Veteran's Hospital Road, Oregon Health Sciences University,
Portland, OR 97201, USA
Abstract
The hypothesis that k-opioid system activity may in part mediate convulsions exhibited during ethanol withdrawal was testedby exposing Withdrawal Seizure-Prone (WSP) and Withdrawal Seizure-Resistant (WSR) mice to chronic ethanol. Whole brain
was harvested for RNA isolation and prodynorphin mRNA steady-state levels in whole brain were examined using Northernblot analysis. The data revealed signi®cantly increased levels of prodynorphin mRNA expression in mice susceptible to ethanolwithdrawal convulsions after withdrawal, with no corresponding increase in prodynorphin steady-state levels in mice resistant to
ethanol withdrawal convulsions. These ®ndings were not due to basal di�erences in prodynorphin expression between the WSPand WSR mice. To verify that the di�erences observed were not due to an ethanol-induced global alteration in genetranscription, mRNA levels of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase were measured.Glyceraldehyde-3-phosphate dehydrogenase expression was unchanged following both chronic exposure to ethanol and chronic
exposure followed by withdrawal. These results extend our understanding of prodynorphin's role in generalized seizure activityto include ethanol withdrawal-induced convulsions. Our ®ndings suggest that prodynorphin expression is modulated duringethanol withdrawal convulsions, or alternatively, prodynorphin may mediate the severity of ethanol withdrawal
convulsions. 7 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Prodynorphin; Withdrawal; Convulsions; Ethanol; Aversion; Selected lines; k-Opioid receptors; Genetics
1. Introduction
Ethanol withdrawal is a physiological response tothe absence of ethanol that is exhibited after an organ-ism has become physically dependent upon alcohol.This response is believed to be a physical adaptationthat the nervous system undergoes in the presence of
ethanol (Himmelsbach, 1942) and is included in theNational Institute on Alcohol Abuse and Alcoholism'sde®nition of alcoholism. Withdrawal can be displayedin various forms, and in humans can include symp-toms such as nausea, sweating, shakiness and anxiety(Victor, 1990; Victor and Adams, 1953), (http://sil-k.nih.gov/silks/niaaa1/publication/booklet.htm). In ad-dition, estimates suggest that 4±7% of alcoholics(Hauser, 1990) su�er from seizures that can be lifethreatening and even fatal; therefore, it is importantthat we understand the mechanisms underlying ethanoldependence and withdrawal. The purpose of this study
Neurochemistry International 37 (2000) 463±472
0197-0186/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved.
PII: S0197-0186(00 )00056 -5
www.elsevier.com/locate/neuint
* Corresponding author. Tel.: +1-503-220-8262; fax: +1-503-273-
5351.
E-mail address: [email protected] (A.S. Beadles-Bohling).

was to begin to elucidate the genes contributing toethanol withdrawal convulsions. We hypothesized thatethanol withdrawal convulsions may be mediated bythe endogenous k-opioid receptor system and havecharacterized expression of the ligand, prodynorphin.
We chose to ask this question utilizing a mousemodel of ethanol dependence and withdrawal createdusing a selective breeding protocol. Starting with a het-erogeneous stock of mice that shows a normal distri-bution of the trait under selection, high and lowscoring animals are mated together, such that a highscoring male is mated with a high scoring female andvice versa. By exerting this pressure over multiple gen-erations a divergent pair of lines is created. Ideally, thegenes contributing to the trait of interest, and only thegenes contributing to this trait, will become ®xedduring this process, while all other alleles will beselected randomly. Of course, it is possible that a setof genes might become ®xed by chance early on in thisselection process, thus it is very important that theselection be replicated. This approach results in twolines of animals divergent on a particular trait and aphenotypically identical replicate pair of lines. In thiscase, HS/Ibg mice were selectively bred for their sensi-tivity or resistance to handling induced convulsions(HICs) when withdrawn from chronic ethanol. Thisproduced the Withdrawal Seizure-Prone (WSP) andthe Withdrawal Seizure-Resistant (WSR) mice (Crabbeet al., 1983, 1985). Thus, when withdrawn, WSP miceshow severe HICs whereas WSR mice show negligibleHICs. The ®rst replicate pair of lines is known as theWSP-1 and WSR-1 mice, the second replicate pair oflines is identi®ed as the WSP-2 and WSR-2 mice(Crabbe, 1987; Crabbe et al., 1983, 1985, 1990; Crabbeand Phillips, 1993).
These animals serve as powerful tools for geneticand molecularly based research on alcoholism. From agenetic perspective, the ability to selectively breed ani-mals and observe divergence on a trait suggests thatselection pressure has been exerted on genes that con-tribute to the trait of interest. Because the WSP micehave been speci®cally selected for severe ethanol with-drawal convulsions, these animals are likely to producean enhanced response of the systems involved withwithdrawal. Furthermore, because of the geneticpower of these selectively bred animals, an observationof alterations in both replicate WSP-1 and WSP-2lines of mice, and an absence of these alterations inWSR mice, would strongly suggest a role for the k-opioid system in this severe withdrawal phenotype.
Prodynorphin was the gene chosen for study basedon current evidence suggesting a role for the k-opioidsystem, and opioids in general, in seizures (Caldecott-Hazard and Engel Jr., 1987; Gale, 1988; Harrison etal., 1995; Hong et al., 1993; Hosford et al., 1997; Loth-man, 1996; Simonato and Romualdi, 1996; Tortella et
al., 1985). This peptide is the endogenous ligand for
the k-opioid receptor, a member of the opioid receptorfamily. This family of receptors includes at least three
receptor subtypes known as the kappa- (k-), mu- (m-),and delta- (d-) opioid receptors (for a recent review,see Dhawan et al., 1996). Each receptor has its own
endogenous peptide ligands: dynorphins, beta-endor-
phins, and enkephalins, respectively. Studies of the k-opioid system have revealed that k-opioid receptorsare widely distributed throughout the brain, with par-
ticularly high densities in limbic brain regions as well
as thalamic nuclei and regions important for visceral
function (Mansour et al., 1988). The distribution ofthe peptide ligand for this receptor is equally extensive
but not always colocalized (Khachaturian et al., 1985).
While little work has focused on the role of the k-opioid system in the convulsions exhibited during etha-
nol withdrawal, alterations in this system have been
observed in other models of seizure disorders. Forexample, the hippocampus and amygdala of naõÈ ve epi-
lepsy-like mice contain signi®cantly more k-opioidreceptors as measured by binding assays (Kai et al.,
1998). These authors postulate that this is due to adecrease in dynorphin peptide abundance in these ani-
mals, which in turn may contribute to their seizure-
susceptibility, but they have not measured dynorphinlevels. On the other hand, radioimmunoassay studies
performed in another model, the seizure sensitive
Mongolian gerbils, revealed signi®cantly higher levels
of dynorphin A peptide in the hippocampus of bothnaõÈ ve and postseizure animals compared to their corre-
sponding seizure resistant counterparts (Lee et al.,
1987). In Swiss Webster mice, intracerebroventricularinjections of k-opioid agonists produced convulsions
(Bansinath et al., 1991). Thus, depending upon the
species and convulsive model used, k-opioid agonists
can be either seizure-protective or seizure-promoting(Bausch et al., 1998; Fischer et al., 1993; Frey, 1988;
Przewlocka et al., 1994b).
Several studies have focused on the e�ects of kindledseizures on prodynorphin mRNA expression levels.
Sixty minutes following a seizure in amygdala kindled
rats, increased levels of prodynorphin mRNA were
observed in hippocampus, striatum and hypothalamus(Romualdi et al., 1995). Prepiriform cortex kindling
also altered prodynorphin levels. However, prodynor-
phin levels were decreased in the hippocampus with anincrease or no change in prodynorphin observed in the
striatum of these animals (Xie et al., 1989). In rats
injected with kainic acid to produce seizures, an
increase in hippocampal prodynorphin mRNA wasobserved 3 h following the injection (Douglass et al.,
1991). Taken together, these data support a role for
the k-opioid system in seizure activity, but againsuggest that the species and seizure model employed
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472464

are important factors when examining alterations inthe k-opioid system.
In terms of alcohol exposure, work performed inC57BL/6 mice using a liquid diet paradigm suggestedprodynorphin abundance was speci®cally increased ina variety of brain regions following a chronic exposureto ethanol (Gulya et al., 1993). These inbred mice areknown to consume high levels of alcohol given achoice between ethanol and water (Belknap et al.,1993; McClearn and Rodgers, 1959). The ®nding ofGulya et al. (1993), along with ®ndings that prodynor-phin expression (Przewlocka et al., 1992, 1997) anddynorphin peptide abundance (Przewlocka et al.,1994a) is altered by ethanol exposure in rats support arole for the k-opioid system in animal models of etha-nol exposure. In addition, previous work found theconcentration of met-enkephalin, an endogenous d-opioid receptor ligand, was signi®cantly greater inWSP mice than in WSR mice, which also supports arole for the opioid system in animal models that showaltered responses to ethanol (Plotkin et al., 1998). Plot-kin suggested that there may be a link betweenincreased met-enkephalin levels and susceptibility toethanol withdrawal convulsions. However, these datawere gathered from naõÈ ve animals; thus, the e�ects ofethanol withdrawal on met-enkephalin are uncharac-terized.
Based on models of seizure activity and mousemodels of ethanol consumption and ethanol withdra-wal convulsions, we hypothesized that alterations inthe k-opioid system expression might occur in animalssusceptible to ethanol withdrawal convulsions. In theexperiments presented here, the e�ects of both ethanolexposure and withdrawal on the k-opioid receptorligand, prodynorphin, were examined in WSP andWSR mice using Northern blot analysis. Using chronicethanol vapor exposure, the phenomena of ethanolwithdrawal was separated from that of ethanol ex-posure in order to identify more clearly whether etha-nol exposure, withdrawal, or both, alter the k-opioidsystem in a mouse model of ethanol withdrawal con-vulsions.
2. Materials and methods
All animal procedures and animal care werereviewed and approved by the Portland Oregon VAMedical Center Animal Care Committee and meetNIH guidelines for appropriate care and use of ani-mals in research. All e�orts were made to minimizeanimal su�ering during these experiments and theminimum numbers of animals necessary to answer ourquestions were used.
2.1. Animal subjects
WSP and WSR mice were bred in the laboratory ofJ.C. Crabbe in Portland, OR. Drug-naõÈ ve adult malemice (age range 56±97 days) from selected generation26 (®lial generations G58±G71) were used. The WSPand WSR selective breeding protocol was replicated,thus there are two independently derived replicateWSP and WSR lines (Crabbe, 1987; Crabbe et al.,1983, 1985, 1990; Crabbe and Phillips, 1993). Mice ofboth replicates of the WSP (WSP-1 and WSP-2) andWSR (WSR-1 and WSR-2) lines were tested in thesestudies. Average ages of the animals at the onset ofthe experiments ranged from 72.8 to 79.7 days; meanbody weights ranged from 25.9 to 29.6 g. Mice weremaintained under a light/dark cycle of 0600±1800 lightwith water and Purina Lab Diet chow available adlibitum. Room temperatures were maintained at22218C. Ethanol exposure was initiated between 0800and 0930 h. Animals were removed from the chamberson the ®nal day at 0830 and subgroups of the ethanol-exposed mice were placed in a quiet room for 6 h untilsacri®ce at 1430.
2.2. Chronic ethanol exposure and withdrawal
Mice were made dependent upon ethanol using amethod described by Goldstein (1972) with modi®-cations previously published (Terdal and Crabbe,1994). On day one, male WSP-1, WSP-2, WSR-1 andWSR-2 mice were weighed, injected ip with ethanol(1.5 g/kg, 20% v/v in 0.9% saline for WSP-1, WSR-1and WSR-2 and 1.75 g/kg, 20% v/v in 0.9% saline forWSP-2 animals) and 1.0 mmol/kg pyrazole HCl (analcohol dehydrogenase inhibitor). Two separate con-trol groups received either pyrazole (1.0 mmol/kg in0.9% saline) or saline (0.9%). Pyrazole is an ethanoldehydrogenase inhibitor that was administered tomaintain consistent blood ethanol concentrations(BECs). The mice were then placed into wire meshcages in a large plastic chamber into which ethanolvapor and air were introduced. Ethanol vapor concen-trations within the chamber were monitored with a gaschromatograph. At 24 and 48 h, the mice wereremoved from the chamber, weighed and injected withpyrazole or saline. If an animal lost greater than 20%of its body weight at the ®nal injection timepoint (48h), it was excluded from the study. A subgroup of ani-mals had 20 ml of blood drawn from the tip of the taildaily for gas chromatography determination of averageBECs. The mice were then replaced in the chamberand the vapor concentration of ethanol was adjustedup or down to achieve BECs at or near 2.0 mg/ml. Atthe 72 h chronic exposure timepoint, which was 24 hafter the ®nal pyrazole injection, the mice wereremoved from the chambers, weighed, and all animals
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472 465

had blood samples drawn for BEC determinations. Asubgroup of the ethanol exposed animals were sacri-®ced immediately to harvest brain from intoxicatedanimals (EtOH-0) and the remaining animals wereallowed to withdraw for 6 h, (6±8 h has been observedto be peak withdrawal in these animals), and thensacri®ced for brain harvest (EtOH-6). The animalschosen for this study had either a 72 h BEC greaterthan 1.5 mg/ml or had an average exposure greaterthan 1.5 mg/ml over the 72 h. BECs ranged from 1.01to 3.17 mg/ml over the 72 h exposure.
2.3. Determination of BEC
For analysis of BECs, the method described byKosobud and Crabbe (1986) was utilized. Brie¯y, 20 mlof blood was drawn from the tip of the tail and addedto 50 ml of 3 M ZnSO4, mixed, and stored on ice untilall blood samples were collected. 50 ml of Ba(OH)2(5%) and 300 ml of ice-cold distilled water were thenadded to each sample, and the samples were vortexedand centrifuged at 48C for 5 min at 12000 g. Thesupernatant was analyzed for ethanol concentrationusing gas chromatography.
2.4. Drug sources and preparation
Pyrazole HCl was purchased from Sigma (St. Louis,MO). Ethanol (ethyl alcohol, absolute, 200 proof) waspurchased from AAPER Alcohol and Chemical (Shel-byville, KY). Reagent chemicals were purchased fromSigma (St. Louis, MO). The random primer comp-lementary DNA (cDNA) kit RPN.1601Z was obtainedfrom Amersham (Arlington Heights, IL). Radioiso-topes [a-32P]-dCTP and [g-32P]-ATP, and GeneScreennylon membrane ®lters were purchased from NewEngland Nuclear (Boston, MA).
Pyrazole HCl was dissolved in saline (0.9%) andadministered ip at an ambient temperature of 22218C.Ethanol was mixed with 0.9% saline and injected ip orintroduced without mixing as a vapor into thechambers.
2.5. Tissue harvest
Animals were sacri®ced via cervical dislocation andwhole brain was isolated immediately. Brains weretransferred to sterile centrifuge tubes, submerged inliquid nitrogen, and held on dry ice until transfer to anÿ808C freezer for storage until RNA isolation.
2.6. RNA isolation and Northern blot analysis
RNA was isolated by the single step acid phenol±chloroform extraction method (Chomczynski and Sac-chi, 1987). RNA was quantitated by spectrophoto-
metric determination at 260 nm. Total cellular RNA(20 mg) was denatured at 658C for 15 min with 2.2 Mformaldehyde and fractionated by electrophoresisthrough a 1% agarose±2.2 M formaldehyde gel. RNAspecies were transferred to GeneScreen nylon mem-brane ®lters overnight using standard capillary blottingtechniques in 1.5 M sodium chloride±0.15 M sodiumcitrate (10� SSC). The bound material was ®xed byUV cross-linking (Stratalinker 1800, Stratagene, LAJolla, CA). Blots were prehybridized at 428C in Ull-rich's solution containing 50% deionized formamide,5� SSC, 5� Denhardt's solution (0.02% Ficoll, .02%polyvinylpyrrolidone, and 0.02% gelatin), 100 mg/mlsonicated salmon sperm DNA, and 2% sodium dode-cyl sulfate (SDS).
All cDNA hybridization probes were isolated as gel-puri®ed cDNA sequences. Hybridization occurred at428C with a 32P-labelled random primed prodynorphincDNA probe generated using [a-32P]d-CTP (Feinbergand Vogelstein, 1983). The prodynorphin cDNA wasgenerously provided by J. Douglass. The prodynorphinprobe was a 1700 base pair fragment excised with PstIand EcoRI from pSP64D1.7 (Civelli et al., 1985). Post-hybridization washes included two 5-min washes atroom temperature in 2� SSC; two 30-min washes at658C in 2� SSC/2% SDS and a ®nal rinse in 0.1�SSC at room temperature. Bound probe was visualizedby autoradiography using Kodak AR ®lm and NENRe¯ection Intensifying Screens at ÿ808C. Membraneswere stripped and reprobed for glyceraldehyde-3-phos-phate dehydrogenase (GAPDH, EC 1.2.1.12) using a32P-labelled random primed cDNA probe generatedusing [a-32P]d-CTP. The GAPDH probe was an 800base pair fragment excised with PstI from pRGAPDH-13 provided by Gary McKnight (Fort et al., 1985). Allblots were also probed with 18S ribosomal RNA(rRNA) which served as a loading control. The 18SrRNA probe was an antisense 30-mer oligonucleotide(AGGGGGTCAGCGCCCGTCGGCATGTATTAG)synthesized by Operon (Alameda, CA) and end-labeledwith [g-32P]ATP. Size markers were RNA standardsrun in adjacent lanes, excised, and stained with ethi-dium bromide.
2.7. Quantitation and statistical analysis
Quantitation of ®lms was performed via volumedensitometry from non-saturated ®lms in the linear re-sponse range utilizing Gel Expert, Version 3.1 (Nucleo-tech, San Carlos, CA). Statistical analysis wasperformed using Student's t-test. Multiple groups weretested using one-way ANOVA with signi®cance set atp < 0:05: Signi®cant ®ndings were further analyzedpost-hoc using Bonferroni's multiple comparisons test.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472466
3. Results
3.1. Chronic ethanol vapor exposure
WSP and WSR mice exhibiting average BECs ran-ging from 1.50 to 2.24 (20.09±0.11) mg/ml at the com-pletion of their 72 h exposure were used for thisanalysis. The mean BEC for the WSP-1 mice was1:7120:06 mg/ml. WSP-2 mice had an average BECof 1:5020:09 mg/ml. BECs of the WSR mice were2:2420:11 mg/ml for the WSR-1 and 1:8420:05 mg/ml for the WSR-2 animals. Two-way ANOVArevealed a signi®cant main e�ect of line�F�1, 46� � 23:73, p < 0:0001� and a signi®cant maine�ect of replicate �F�1, 46� � 11:27, p < 0:001� but nosigni®cant line by replicate interaction.
3.2. Prodynorphin expression in WSP mice
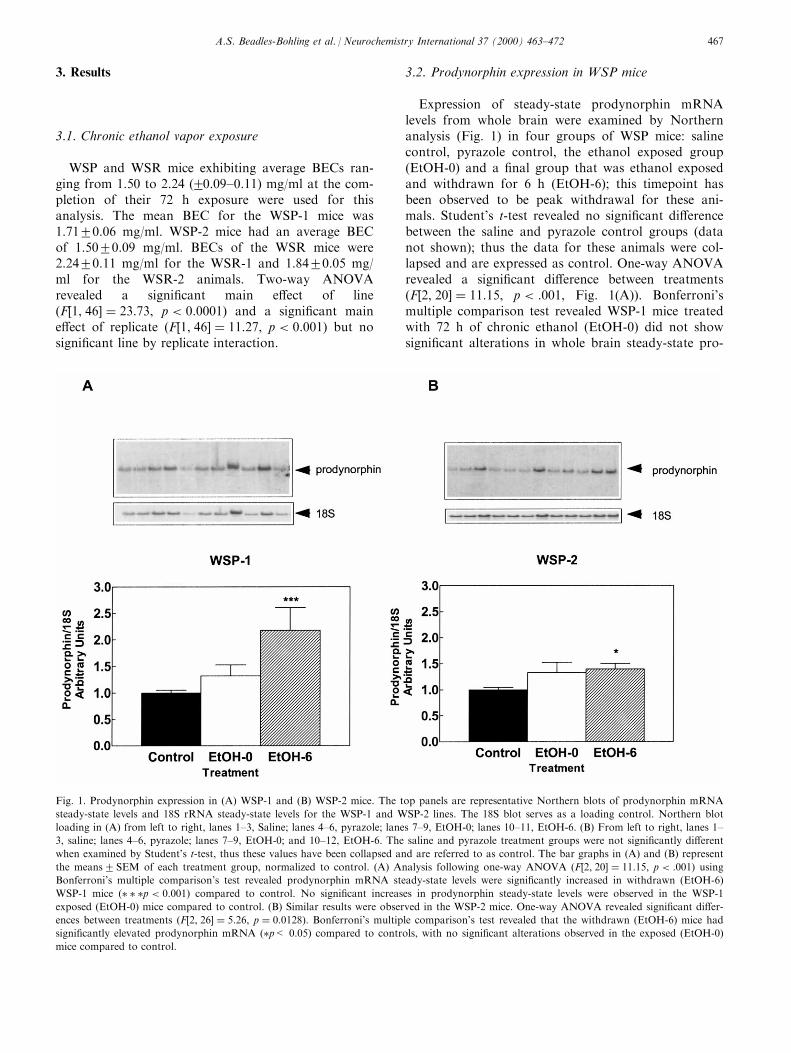
Expression of steady-state prodynorphin mRNAlevels from whole brain were examined by Northernanalysis (Fig. 1) in four groups of WSP mice: salinecontrol, pyrazole control, the ethanol exposed group(EtOH-0) and a ®nal group that was ethanol exposedand withdrawn for 6 h (EtOH-6); this timepoint hasbeen observed to be peak withdrawal for these ani-mals. Student's t-test revealed no signi®cant di�erencebetween the saline and pyrazole control groups (datanot shown); thus the data for these animals were col-lapsed and are expressed as control. One-way ANOVArevealed a signi®cant di�erence between treatments�F�2, 20� � 11:15, p < :001, Fig. 1(A)). Bonferroni'smultiple comparison test revealed WSP-1 mice treatedwith 72 h of chronic ethanol (EtOH-0) did not showsigni®cant alterations in whole brain steady-state pro-
Fig. 1. Prodynorphin expression in (A) WSP-1 and (B) WSP-2 mice. The top panels are representative Northern blots of prodynorphin mRNA
steady-state levels and 18S rRNA steady-state levels for the WSP-1 and WSP-2 lines. The 18S blot serves as a loading control. Northern blot
loading in (A) from left to right, lanes 1±3, Saline; lanes 4±6, pyrazole; lanes 7±9, EtOH-0; lanes 10±11, EtOH-6. (B) From left to right, lanes 1±
3, saline; lanes 4±6, pyrazole; lanes 7±9, EtOH-0; and 10±12, EtOH-6. The saline and pyrazole treatment groups were not signi®cantly di�erent
when examined by Student's t-test, thus these values have been collapsed and are referred to as control. The bar graphs in (A) and (B) represent
the means2SEM of each treatment group, normalized to control. (A) Analysis following one-way ANOVA �F�2, 20� � 11:15, p < :001� usingBonferroni's multiple comparison's test revealed prodynorphin mRNA steady-state levels were signi®cantly increased in withdrawn (EtOH-6)
WSP-1 mice �� � �p < 0:001� compared to control. No signi®cant increases in prodynorphin steady-state levels were observed in the WSP-1
exposed (EtOH-0) mice compared to control. (B) Similar results were observed in the WSP-2 mice. One-way ANOVA revealed signi®cant di�er-
ences between treatments �F�2, 26� � 5:26, p � 0:0128). Bonferroni's multiple comparison's test revealed that the withdrawn (EtOH-6) mice had
signi®cantly elevated prodynorphin mRNA (�p< 0.05) compared to controls, with no signi®cant alterations observed in the exposed (EtOH-0)
mice compared to control.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472 467
dynorphin mRNA as examined by Northern blotanalysis. In contrast, whole brain prodynorphinmRNA in WSP-1 mice withdrawn from ethanol for 6h (EtOH-6) showed a signi®cant increase when com-pared to control prodynorphin steady-state levels�P < 0:001). This increase was approximately 2.2-fold.One-way ANOVA of the WSP-2 line revealed similarresults, �F�2, 26� � 5:26, p < 0:01). Bonferroni's mul-tiple comparison test revealed no signi®cant alterationsin prodynorphin steady-state levels observed in theexposed (EtOH-0) animals, but a signi®cant increase inprodynorphin steady-state levels was observed in thewithdrawn (EtOH-6) mice compared to controls�p < 0:05). In these mice the increase in prodynorphinsteady-state levels was 1.4-fold (Fig. 1(B)). Thus, etha-nol withdrawal resulted in signi®cant elevations in pro-dynorphin expression in both replicate WSP lines.
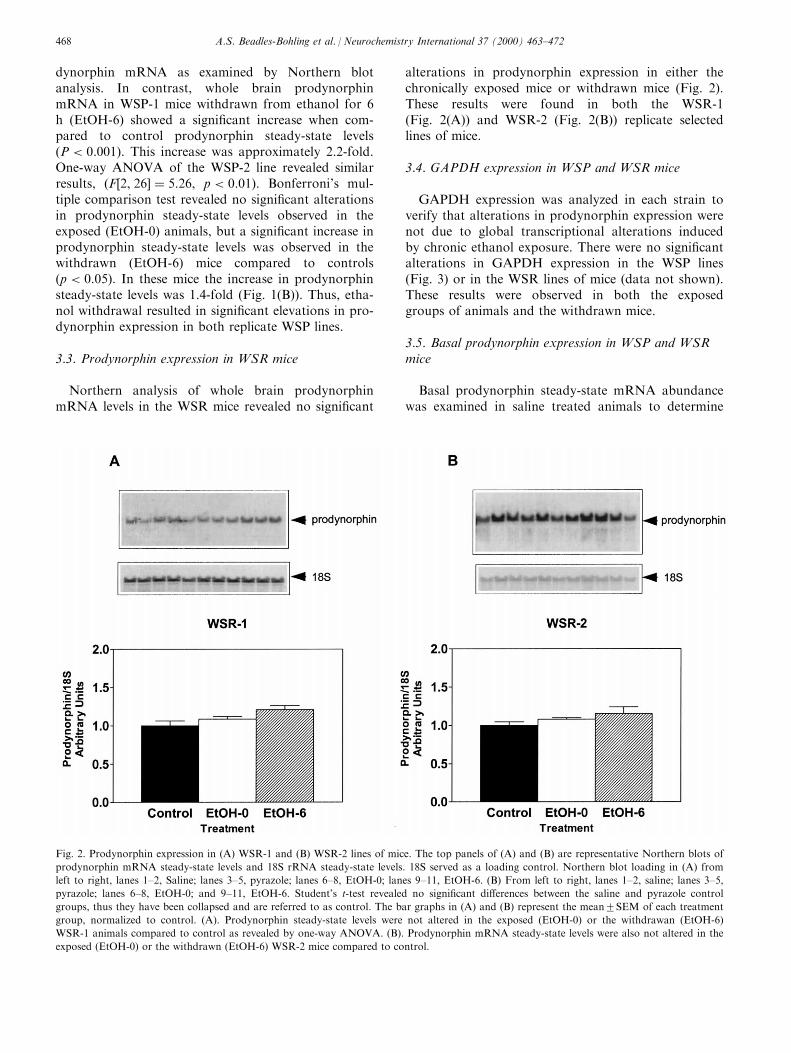
3.3. Prodynorphin expression in WSR mice
Northern analysis of whole brain prodynorphinmRNA levels in the WSR mice revealed no signi®cant
alterations in prodynorphin expression in either thechronically exposed mice or withdrawn mice (Fig. 2).These results were found in both the WSR-1(Fig. 2(A)) and WSR-2 (Fig. 2(B)) replicate selectedlines of mice.
3.4. GAPDH expression in WSP and WSR mice
GAPDH expression was analyzed in each strain toverify that alterations in prodynorphin expression werenot due to global transcriptional alterations inducedby chronic ethanol exposure. There were no signi®cantalterations in GAPDH expression in the WSP lines(Fig. 3) or in the WSR lines of mice (data not shown).These results were observed in both the exposedgroups of animals and the withdrawn mice.
3.5. Basal prodynorphin expression in WSP and WSRmice
Basal prodynorphin steady-state mRNA abundancewas examined in saline treated animals to determine
Fig. 2. Prodynorphin expression in (A) WSR-1 and (B) WSR-2 lines of mice. The top panels of (A) and (B) are representative Northern blots of
prodynorphin mRNA steady-state levels and 18S rRNA steady-state levels. 18S served as a loading control. Northern blot loading in (A) from
left to right, lanes 1±2, Saline; lanes 3±5, pyrazole; lanes 6±8, EtOH-0; lanes 9±11, EtOH-6. (B) From left to right, lanes 1±2, saline; lanes 3±5,
pyrazole; lanes 6±8, EtOH-0; and 9±11, EtOH-6. Student's t-test revealed no signi®cant di�erences between the saline and pyrazole control
groups, thus they have been collapsed and are referred to as control. The bar graphs in (A) and (B) represent the mean2SEM of each treatment
group, normalized to control. (A). Prodynorphin steady-state levels were not altered in the exposed (EtOH-0) or the withdrawan (EtOH-6)
WSR-1 animals compared to control as revealed by one-way ANOVA. (B). Prodynorphin mRNA steady-state levels were also not altered in the
exposed (EtOH-0) or the withdrawn (EtOH-6) WSR-2 mice compared to control.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472468
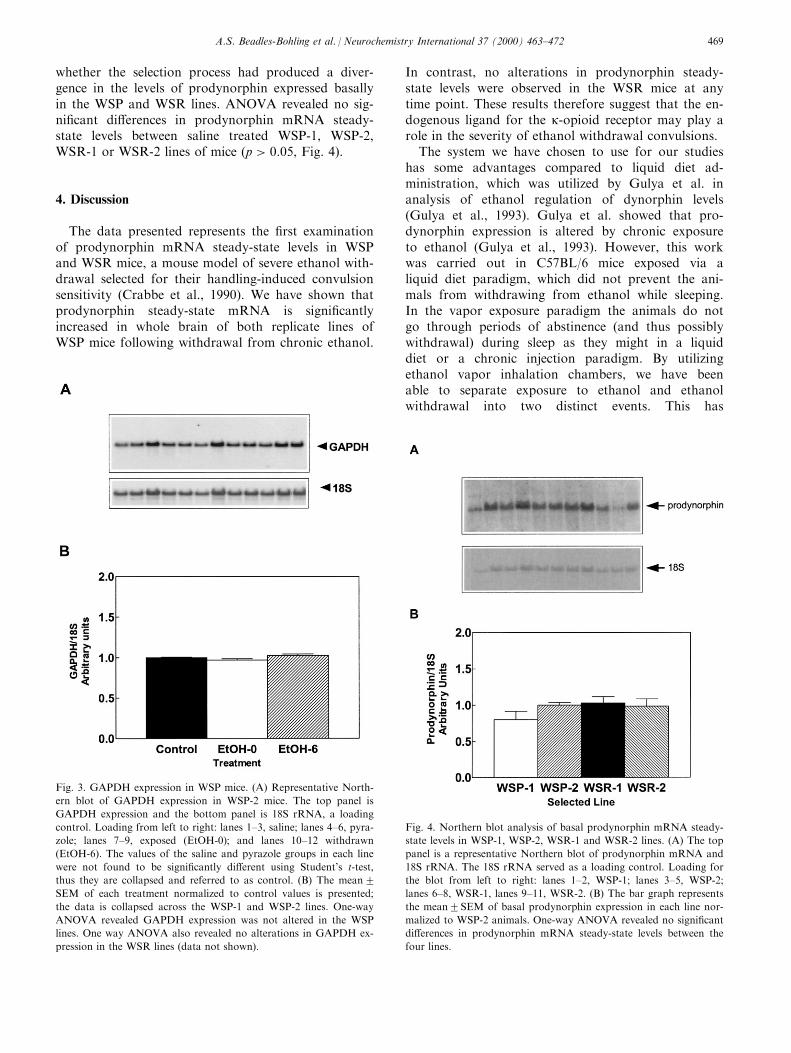
whether the selection process had produced a diver-gence in the levels of prodynorphin expressed basallyin the WSP and WSR lines. ANOVA revealed no sig-ni®cant di�erences in prodynorphin mRNA steady-state levels between saline treated WSP-1, WSP-2,WSR-1 or WSR-2 lines of mice �p > 0:05, Fig. 4).
4. Discussion
The data presented represents the ®rst examinationof prodynorphin mRNA steady-state levels in WSPand WSR mice, a mouse model of severe ethanol with-drawal selected for their handling-induced convulsionsensitivity (Crabbe et al., 1990). We have shown thatprodynorphin steady-state mRNA is signi®cantlyincreased in whole brain of both replicate lines ofWSP mice following withdrawal from chronic ethanol.
In contrast, no alterations in prodynorphin steady-state levels were observed in the WSR mice at anytime point. These results therefore suggest that the en-dogenous ligand for the k-opioid receptor may play arole in the severity of ethanol withdrawal convulsions.
The system we have chosen to use for our studieshas some advantages compared to liquid diet ad-ministration, which was utilized by Gulya et al. inanalysis of ethanol regulation of dynorphin levels(Gulya et al., 1993). Gulya et al. showed that pro-dynorphin expression is altered by chronic exposureto ethanol (Gulya et al., 1993). However, this workwas carried out in C57BL/6 mice exposed via aliquid diet paradigm, which did not prevent the ani-mals from withdrawing from ethanol while sleeping.In the vapor exposure paradigm the animals do notgo through periods of abstinence (and thus possiblywithdrawal) during sleep as they might in a liquiddiet or a chronic injection paradigm. By utilizingethanol vapor inhalation chambers, we have beenable to separate exposure to ethanol and ethanolwithdrawal into two distinct events. This has
Fig. 4. Northern blot analysis of basal prodynorphin mRNA steady-
state levels in WSP-1, WSP-2, WSR-1 and WSR-2 lines. (A) The top
panel is a representative Northern blot of prodynorphin mRNA and
18S rRNA. The 18S rRNA served as a loading control. Loading for
the blot from left to right: lanes 1±2, WSP-1; lanes 3±5, WSP-2;
lanes 6±8, WSR-1, lanes 9±11, WSR-2. (B) The bar graph represents
the mean2SEM of basal prodynorphin expression in each line nor-
malized to WSP-2 animals. One-way ANOVA revealed no signi®cant
di�erences in prodynorphin mRNA steady-state levels between the
four lines.
Fig. 3. GAPDH expression in WSP mice. (A) Representative North-
ern blot of GAPDH expression in WSP-2 mice. The top panel is
GAPDH expression and the bottom panel is 18S rRNA, a loading
control. Loading from left to right: lanes 1±3, saline; lanes 4±6, pyra-
zole; lanes 7±9, exposed (EtOH-0); and lanes 10±12 withdrawn
(EtOH-6). The values of the saline and pyrazole groups in each line
were not found to be signi®cantly di�erent using Student's t-test,
thus they are collapsed and referred to as control. (B) The mean2SEM of each treatment normalized to control values is presented;
the data is collapsed across the WSP-1 and WSP-2 lines. One-way
ANOVA revealed GAPDH expression was not altered in the WSP
lines. One way ANOVA also revealed no alterations in GAPDH ex-
pression in the WSR lines (data not shown).
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472 469

allowed us to suggest that the alterations in mRNAexpression observed were associated with the ani-mal's withdrawal from ethanol rather than exposure.Our results therefore help clarify the issue of prody-norphin regulation by withdrawal rather than ex-posure, and suggest that the route of administrationfor ethanol be considered when determining thee�ect of ethanol on gene expression.
There is con¯icting evidence for the role of pro-dynorphin's peptide product in seizure activity. Mul-tiple animal models have been used, and thetimepoints in which the peptide or its mRNA hasbeen examined vary. In some animal modelsincreased prodynorphin suggests increased seizuresusceptibility when examined in naõÈ ve subjects (Leeet al., 1987). Other models have studied prodynor-phin levels only after seizure generation, and theresults have varied. In rat models of amygdala andsystemic kainic acid kindling, prodynorphin levelshave been shown to be increased (Douglass et al.,1991; Romualdi et al., 1995). Amygdaloid kindlingproduced by injecting kainic acid directly into theamygdala produced signi®cant decreases in prody-norphin abundance (Lason et al., 1983), and in pre-piriform cortex kindling models a decrease was alsoobserved (Xie et al., 1989). Nevertheless, our resultsare consistent with a role for prodynorphin ex-pression in ethanol withdrawal convulsions. Futurestudies will examine the e�ects of withdrawal onthe k-opioid receptor system in additional mousemodels in order to extend these ®nding to othergenetically unique populations of mice andstrengthen the generalizability of our ®ndings.
It is important to note that the observed alterationsin prodynorphin steady-state levels are not due to aglobal alteration in gene expression due to chronic ex-posure to ethanol, as was shown by the analysis ofGAPDH expression. Additionally, the di�erencesobserved between the two lines of mice were not dueto an inherent di�erence in prodynorphin steady-statelevels generated between the two lines during the selec-tive breeding process. Thus, the alterations we haveobserved are seen only in the presence of severe with-drawal following chronic ethanol exposure.
Our data does not allow us to make conclusionsregarding a causal role for prodynorphin in ethanolwithdrawal convulsions. Several alternatives arepossible; for example, prodynorphin may be modu-lating the expression of convulsions or the severityof these convulsions, or prodynorphin may bemodulated by convulsive activity. We believe thelatter is unlikely because the handling protocolgreatly minimizes the observed convulsive activitythese animals exhibit. However, this option cannotbe ruled out, as a small subset of the animals doinevitably convulse prior to sacri®ce. Future studies
using behavioral pharmacological approaches and
time course studies will focus on distinguishingbetween these two alternative roles for prodynorphin
in ethanol withdrawal.
Of course it is possible that prodynorphin may
also participate in additional aspects of withdrawal.
For example, k-opioid receptor activation has beensuggested to play a role in mediating aversion,
speci®cally via the mesolimbic dopaminergic path-
ways (reviewed by Herz, 1997). This alternative ex-planation is currently being examined in our
laboratory via behavioral and molecular approaches.
Because our analysis did not investigate speci®c
anatomic regions, we do not know if the brain
regions thought to be involved in the generation ofseizures correspond to the regions showing prody-
norphin alterations. A more comprehensive analysisof regional alterations of prodynorphin abundance
in WSP and WSR mouse brain using in situ hy-
bridization analysis is currently underway in ourlaboratory. These ®ndings should provide insight
into the potential role(s) of prodynorphin in ethanol
withdrawal. Furthermore, time course studies andcon®rmation of changes in peptide levels will need
to be pursued.
To conclude, this work has demonstrated that
steady-state prodynorphin mRNA is increased in
whole brain of WSP mice, a mouse model of severeethanol withdrawal convulsions, following withdrawal
from 72 h of chronic ethanol exposure. This is in con-
trast to the absence of changes in prodynorphin levelsin the WSR mice, which are resistant to ethanol with-
drawal convulsions. These ®ndings advance the work
of Gulya et al., who found that prodynorphin ex-pression was increased by ethanol exposure via a liquid
diet (Gulya et al., 1993). In addition, our inability to
detect changes in prodynorphin mRNA steady-statelevels in the EtOH-0 animals suggests that the method
of exposure must be considered when examining etha-
nol's e�ects on gene expression. Finally, our datasuggests that changes in the expression of the ligand
for the k-opioid receptor may be involved with ethanol
withdrawal seizure severity, although the exact role forthis molecule will need to be further characterized.
These ®ndings also extend the current data suggestinga role for opioid receptor ligands in seizure generation
to convulsions produced during ethanol withdrawal,
and suggest that alterations in the k-opioid systemshould be examined in further mouse models of etha-
nol withdrawal and other withdrawing species to deter-
mine the generalizability to humans. A betterunderstanding of the role of the k-opioid system in
ethanol withdrawal may eventually allow identi®cation
of potential therapeutic targets for the treatment ofwithdrawing alcoholics.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472470

Acknowledgements
The authors would like to thank Janet Dorow,Jason Sibert, Jason Schlumbohm, Jacqueline Dierickxand Anne Chapman for their excellent technical assist-ance. We would also like to thank Dr. Cheryl Reed,Dr. Kaitlin Browman and Dr. Pamela Metten for theirstatistical advice. Finally, we would like to thank AnneChapman and Heather Burpee for their careful readingof the manuscript. These studies were supported byNIAAA Grants AA10760 and AA05547, and grantsfrom the Department of Veteran's A�airs.
References
Bansinath, M., Ramabadran, K., Turndorf, H., Shukla, V.K., 1991.
Intracerebroventricular administration of kappa-agonists induces
convulsions in mice. Brain Res. Bull. 27, 75±79.
Bausch, S.B., Esteb, T.M., Terman, G.W., Chavkin, C., 1998.
Administered and endogenously released k-opioids decrease pilo-
carpine-induced seizures and seizure-induced histopathology. J.
Pharmacol. Exp. Ther. 284, 1147±1155.
Belknap, J.K., Crabbe, J.C., Young, E.R., 1993. Voluntary con-
sumption of ethanol in 15 inbred mouse strains.
Psychopharmacology 112, 503±510.
Caldecott-Hazard, S., Engel Jr., J., 1987. Limbic postictal events:
anatomical substrates and opioid receptor involvement. Prog.
Neuropsychopharmacol. Biol. Psychiatry 11, 389±418.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA iso-
lation by acid guanidinium thiocyanate±phenol±chloroform
extraction. Anal. Biochem. 162, 156±159.
Civelli, O., Douglass, J., Goldstein, A., Herbert, E., 1985. Sequence
and expression of the rat prodynorphin gene. Proc. Natl. Acad.
Sci. USA 82, 4291±4295.
Crabbe, J.C., 1987. Genetic models of alcohol dependence. Alcohol
Alcohol Suppl. 1, 103±108.
Crabbe, J.C., Kosobud, A., Young, E.R., 1983. Genetic selection for
ethanol withdrawal severity: di�erences in replicate lines. Life Sci.
33, 955±962.
Crabbe, J.C., Kosobud, A., Young, E.R., Tam, B.R., McSwigan,
J.D., 1985. Bidirectional selection for susceptibility to ethanol
withdrawal seizures in Mus musculus. Behav. Genet. 15, 521±536.
Crabbe, J.C., Merrill, C.M., Kim, D., Belknap, J.K., 1990. Alcohol
dependence and withdrawal: a genetic animal model. Ann. Med.
22, 259±263.
Crabbe, J.C., Phillips, T.J., 1993. Selective breeding for alcohol with-
drawal severity. Behav. Genet 23, 171±177.
Dhawan, B.N., Cesselin, F., Raghubir, R., Reisine, T., Bradley, P.B.,
Portoghese, P.S., Hamon, M., 1996. International Union of
Pharmacology. XII. Classi®cation of opioid receptors.
Pharmacol. Rev. 48, 567±592.
Douglass, J., Grimes, L., Shook, J., Lee, P.H., Hong, J.S., 1991.
Systemic administration of kainic acid di�erentially regulates the
levels of prodynorphin and proenkephalin mRNA and peptides
in the rat hippocampus. Brain Res. Mol. Brain Res. 9, 79±86.
Feinberg, A.P., Vogelstein, B., 1983. A technique for radiolabeling
DNA restriction endonuclease fragments to high speci®c activity.
Anal. Biochem. 132, 6±13.
Fischer, W., Bodewei, R., Von Voigtlander, P.F., Muller, M., 1993.
Anticonvulsant and related e�ects of U-54494A in various seizure
tests. J. Pharmacol. Exp. Ther. 267, 163±170.
Fort, P., Marty, L., Piechaczyk, M., Sabrouty, S.E., Dani, C.,
Jeanteur, P., Blanchard, J.M., 1985. Various rat tissues express
only one major mRNA species from the glyceraldehyde-3-phos-
phate-dehydrogenase multigenic family. Nucleic Acids Res. 13,
1431±1435.
Frey, H.H., 1988. E�ect of m- and k-opioid agonists on the electro-
convulsive seizure threshold in mice and antagonism by naloxone
and MR 2266. Pharmacol. Toxicol. 62, 150±154.
Gale, K., 1988. Progression and generalization of seizure discharge:
anatomical and neurochemical substrates. Epilepsia 29 (Suppl. 2),
S15±34.
Goldstein, D.B., 1972. An animal model for testing e�ects of drugs
on alcohol withdrawal reactions. J. Pharmacol. Exp. Ther. 183,
14±22.
Gulya, K., Orpana, A.K., Sikela, J.M., Ho�man, P.L., 1993.
Prodynorphin and vasopressin mRNA levels are di�erentially
a�ected by chronic ethanol ingestion in the mouse. Brain Res.
Mol. Brain Res. 20, 1±8.
Harrison, M.B., Shumate, M.D., Lothman, E.W., 1995. Opioid pep-
tide expression in models of chronic temporal lobe epilepsy.
Neuroscience 65, 785±795.
Hauser, W.A., 1990. Epidemiology of alcohol use and of epilepsy:
the magnitude of the problem. In: Porter, R.J., Mattson, R.H.,
Cramer, J.A., Diamond, I. (Eds.), Alcohol and Seizures. F.A.
Davis Co, Philadelphia, pp. 12±21.
Herz, A., 1997. Endogenous opioid systems and alcohol addiction.
Psychopharmacology (Berl.) 129, 99±111.
Himmelsbach, C.K., 1942. Clinical studies of drug addiction.
Archives of Internal Medicine 69, 766.
Hong, J.S., McGinty, J.F., Lee, P.H., Xie, C.W., Mitchell, C.L.,
1993. Relationship between hippocampal opioid peptides and sei-
zures. Prog. Neurobiol. 40, 507±528.
Hosford, D.A., Caddick, S.J., Lin, F.H., 1997. Generalized epilep-
sies: emerging insights into cellular and genetic mechanisms.
Curr. Opin. Neurol. 10, 115±120.
Kai, T., Onishi, H., Koide, S., Katayama, M., Yamagami, S., 1998.
Developmental and regional alteration of kappa-opioid receptors
in seizure-susceptible EL mouse brain. Neurochem. Res. 23, 163±
168.
Khachaturian, H., Lewis, M.E., Schafer, M.K.-H., Watson, S.J.,
1985. Anatomy of the CNS opioid systems. Trends Neurosci. 10,
111±119.
Kosobud, A., Crabbe, J.C., 1986. Ethanol withdrawal in mice bred
to be genetically prone or resistant to ethanol withdrawal sei-
zures. J. Pharmacol. Exp. Ther. 238, 170±177.
Lason, W., Przewlocka, B., Stala, L., Przewlocki, R., 1983. Changes
in hippocampal immunoreactive dynorphin and neoendorphin
content following intra-amygdalar kainic acid-induced seizures.
Neuropeptides 3, 399±404.
Lee, R.J., Hong, J.S., McGinty, J.F., Lomax, P., 1987. Increased en-
kephalin and dynorphin immunoreactivity in the hippocampus of
seizure sensitive Mongolian gerbils. Brain Res. 401, 353±358.
Lothman, E.W., 1996. Basic mechanisms of seizure expression.
Epilepsy Res. Suppl. 11, 9±16.
Mansour, A., Khachaturian, H., Lewis, M.E., Akil, H., Watson,
S.J., 1988. Anatomy of CNS opioid receptors. Trends Neurosci.
11, 308±314.
McClearn, G.E., Rodgers, D.A., 1959. Di�erences in alcohol prefer-
ence among inbred strains of mice. Q.J. Stud. Alcohol 20, 691±
695.
Plotkin, S.R., Banks, W.A., Kastin, A.J., 1998. Enkephalin,
PPE mRNA, and PTS-1 in alcohol Withdrawal Seizure-Prone
and -Resistant mice. Alcohol 15, 25±31.
Przewlocka, B., Lason, W., Przewlocki, R., 1992. Repeated ethanol
administration decreases prodynorphin biosynthesis in the rat
hippocampus. Neurosci. Lett. 134, 195±198.
Przewlocka, B., Lason, W., Przewlocki, R., 1994a. Repeated ethanol
di�erently a�ects opioid peptide biosynthesis in the rat pituitary.
Neuroendocrinology 60, 331±336.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472 471

Przewlocka, B., Machelska, H., Lason, W., 1994b. Kappa opioid
receptor agonists inhibit the pilocarpine-induced seizures
and toxicity in the mouse. Eur. Neuropsychopharmacol. 4, 527±
533.
Przewlocka, B., Turchan, J., Lason, W., Przewlocki, R., 1997.
Ethanol withdrawal enhances the prodynorphin system activity in
the rat nucleus accumbens. Neurosci. Lett. 238, 13±16.
Romualdi, P., Donatini, A., Bregola, G., Bianchi, C., Beani, L.,
Ferri, S., Simonato, M., 1995. Early changes in prodynorphin
mRNA and ir-dynorphin A levels after kindled seizures in the
rat. Eur.J. Neurosci. 7, 1850±1856.
Simonato, M., Romualdi, P., 1996. Dynorphin and epilepsy [pub-
lished erratum appears in Prog. Neurobiol. 1997, 51 (2), 223±
224]. Prog. Neurobiol. 50, 557±583.
Terdal, E.S., Crabbe, J.C., 1994. Indexing withdrawal in mice:
matching genotypes for exposure in studies using ethanol vapor
inhalation. Alcohol Clin. Exp. Res. 18, 542±547.
Tortella, F.C., Long, J.B., Holaday, J.W., 1985. Endogenous opioid
systems: physiological role in the self-limitation of seizures. Brain
Res. 332, 174±178.
Victor, M., 1990. Alcohol withdrawal seizures: an overview. In:
Porter, R.J., Mattson, R.H., Cramer, J.A., Diamond, I. (Eds.),
Alcohol and Seizures. F.A. Davis Co, Philadelphia, pp. 148±161.
Victor, M., Adams, R.D., 1953. The e�ect of ethanol on the nervous
system. Research Publications of the Association of Research on
Nervous and Mental Disorders 32, 526±573.
Xie, C.W., Lee, P.H., Douglass, J., Crain, B., Hong, J.S., 1989.
Deep prepiriform cortex kindling di�erentially alters the levels of
prodynorphin mRNA in rat hippocampus and striatum. Brain
Res. 495, 156±160.
A.S. Beadles-Bohling et al. / Neurochemistry International 37 (2000) 463±472472
Related Documents











