Elevated Growth Temperature Can Enhance Photosystem I Trimer Formation and Affects Xanthophyll Biosynthesis in Cyanobacterium Synechocystis sp. PCC6803 Cells Kinga Klodawska 1 , La ´szlo ´ Kova ´cs 2 , Zsuzsanna Va ´rkonyi 2 , Miha ´ly Kis 2 ,O ¨ zge Sozer 2 , Hajnalka Laczko ´-Dobos 2 , Ottilia Ko ´bori 2 , Ildiko ´ Domonkos 2 , Kazimierz Strzalka 1 , Zolta ´n Gombos 2 and Przemyslaw Malec 1, * 1 Department of Plant Physiology and Biochemistry, Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, 30-387 Krako ´w, Poland 2 Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H-6701 Szeged, Hungary *Corresponding author: E-mail, [email protected]; Fax, +48-12-664-69-02. (Received June 24, 2014; Accepted December 9, 2014) In the thylakoid membranes of the mesophilic cyanobacter- ium Synechocystis PCC6803, PSI reaction centers (RCs) are organized as monomers and trimers. PsaL, a 16 kDa hydro- phobic protein, a subunit of the PSI RC, was previously iden- tified as crucial for the formation of PSI trimers. In this work, the physiological effects accompanied by PSI oligomerization were studied using a PsaL-deficient mutant (psaL), not able to form PSI trimers, grown at various temperatures. We dem- onstrate that in wild-type Synechocystis, the monomer to trimer ratio depends on the growth temperature. The inacti- vation of the psaL gene in Synechocystis grown phototropi- cally at 30 C induces profound morphological changes, including the accumulation of glycogen granules localized in the cytoplasm, resulting in the separation of particular thyla- koid layers. The carotenoid composition in psaL shows that PSI monomerization leads to an increased accumulation of myxoxantophyll, zeaxanthin and echinenone irrespective of the temperature conditions. These xanthophylls are formed at the expense of b-carotene. The measured H 2 O!CO 2 oxygen evolution rates in the psaL mutant are higher than those observed in the wild type, irrespective of the growth temperature. Moreover, circular dichroism spectros- copy in the visible range reveals that a peak attributable to long-wavelength-absorbing carotenoids is apparently enhanced in the trimer-accumulating wild-type cells. These results suggest that specific carotenoids are accompanied by the accumulation of PSI oligomers and play a role in the for- mation of PSI oligomer structure. Keywords: Carotenoids CD spectra PSI trimer PsaL- deficient mutant Synechocystis. Abbreviations: CD, circular dichroism; CN-PAGE, clear native- PAGE; b-DM, n-dodecyl-b-D-maltoside; HRP, horseradish per- oxidase; OD, optical density; PG, phosphatidylglycerol; RC, re- action center; RP-HPLC, reversed phase HPLC; WT, wild type. Introduction PSI is one of the largest pigment–protein complexes of the photosynthetic membranes in cyanobacteria, algae and higher plants. It catalyzes the electron transfer from reduced plastocyanin on the lumenal side of the membrane to ferre- doxin or flavodoxin on the cytoplasmic/stromal membrane side (Golbeck 1992, Chitnis 2001). The PSI of algae and higher plants exists exclusively in the monomer form, consist- ing of a PSI reaction center (RC) which binds four peripheral light-harvesting complexes (LHCIs). LHCI complexes have not been found in cyanobacteria (Chitnis 1996). Instead, these or- ganisms contain PSI RCs organized as monomers and trimers (Boekema et al. 1987, Rogner et al. 1990, Shubin et al. 1993). Recently, the presence of dimers and probably tetramers of PSI was observed in the cyanobacterium Anabaena 7120 and in the glaucophyte Cyanophora paradoxa (Watanabe et al. 2011). In the vast majority of cyanobacteria, PSI RCs consist of 11 protein subunits. In particular, PsaA and PsaB are the two cen- tral subunits of the heterodimeric core of the RC. Additionally, nine proteins with lower molecular weight (designated PsaC to PsaM) are involved in the formation of the PSI RC monomer (Fromme et al. 2001). PsaL, a 16 kDa hydrophobic protein sub- unit of the PSI RC, was identified as crucial for the formation of PSI trimers (Chitnis and Chitnis 1993, Chitnis et al. 1993, Schluchter et al. 1996). Only the monomer form of PSI RC accumulated in a PsaL-deficient Synechocystis mutant, demon- stranting the importance of PsaL in the formation of PSI trimers (Chitnis and Chitnis 1993). Several carotenoid residues have been identified in the vicinity of the PsaL-containing domain within a PSI trimer structure (Fromme et al. 2001, Sozer et al. 2011). Recently, the formation of PSI trimer has been found to be repressed in a carotenoidless crtHB mutant strain (Sozer et al. 2010). It is commonly accepted that there exists a dynamic equilibrium between monomer and oligomer forms of PSI in the photosynthetic membranes of Synechocystis PCC6803 and other cyanobacteria. However, the physiological significance of PSI oligomerization remains unclear. What are known as ‘red Chls’ were identified in Spirulina platensis. They absorb light wavelengths >700 nm and were found to be preferentially asso- ciated with oligomers of PSI RCs (Karapetyan et al. 1999). It was Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199, Advance Access publication on 16 December 2014, available online at www.pcp.oxfordjournals.org ! The Author 2014. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: [email protected] Regular Paper at Uniwersytet Jagiellonsky w Krakowie on March 23, 2015 http://pcp.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Elevated Growth Temperature Can Enhance PhotosystemI Trimer Formation and Affects Xanthophyll Biosynthesisin Cyanobacterium Synechocystis sp. PCC6803 CellsKinga Kłodawska1, Laszlo Kovacs2, Zsuzsanna Varkonyi2, Mihaly Kis2, Ozge Sozer2,Hajnalka Laczko-Dobos2, Ottilia Kobori2, Ildiko Domonkos2, Kazimierz Strzałka1,Zoltan Gombos2 and Przemysław Malec1,*1Department of Plant Physiology and Biochemistry, Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, 30-387 Krakow, Poland2Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H-6701 Szeged, Hungary
*Corresponding author: E-mail, [email protected]; Fax, +48-12-664-69-02.(Received June 24, 2014; Accepted December 9, 2014)
In the thylakoid membranes of the mesophilic cyanobacter-ium Synechocystis PCC6803, PSI reaction centers (RCs) areorganized as monomers and trimers. PsaL, a 16 kDa hydro-phobic protein, a subunit of the PSI RC, was previously iden-tified as crucial for the formation of PSI trimers. In this work,the physiological effects accompanied by PSI oligomerizationwere studied using a PsaL-deficient mutant (�psaL), not ableto form PSI trimers, grown at various temperatures. We dem-onstrate that in wild-type Synechocystis, the monomer totrimer ratio depends on the growth temperature. The inacti-vation of the psaL gene in Synechocystis grown phototropi-cally at 30�C induces profound morphological changes,including the accumulation of glycogen granules localized inthe cytoplasm, resulting in the separation of particular thyla-koid layers. The carotenoid composition in �psaL shows thatPSI monomerization leads to an increased accumulation ofmyxoxantophyll, zeaxanthin and echinenone irrespective ofthe temperature conditions. These xanthophylls are formedat the expense of b-carotene. The measured H2O!CO2
oxygen evolution rates in the �psaL mutant are higherthan those observed in the wild type, irrespective of thegrowth temperature. Moreover, circular dichroism spectros-copy in the visible range reveals that a peak attributable tolong-wavelength-absorbing carotenoids is apparentlyenhanced in the trimer-accumulating wild-type cells. Theseresults suggest that specific carotenoids are accompanied bythe accumulation of PSI oligomers and play a role in the for-mation of PSI oligomer structure.
Keywords: Carotenoids � CD spectra � PSI trimer � PsaL-deficient mutant � Synechocystis.
Abbreviations: CD, circular dichroism; CN-PAGE, clear native-PAGE; b-DM, n-dodecyl-b-D-maltoside; HRP, horseradish per-oxidase; OD, optical density; PG, phosphatidylglycerol; RC, re-action center; RP-HPLC, reversed phase HPLC; WT, wild type.
Introduction
PSI is one of the largest pigment–protein complexes of thephotosynthetic membranes in cyanobacteria, algae and
higher plants. It catalyzes the electron transfer from reducedplastocyanin on the lumenal side of the membrane to ferre-doxin or flavodoxin on the cytoplasmic/stromal membraneside (Golbeck 1992, Chitnis 2001). The PSI of algae andhigher plants exists exclusively in the monomer form, consist-ing of a PSI reaction center (RC) which binds four peripherallight-harvesting complexes (LHCIs). LHCI complexes have notbeen found in cyanobacteria (Chitnis 1996). Instead, these or-ganisms contain PSI RCs organized as monomers andtrimers (Boekema et al. 1987, Rogner et al. 1990, Shubin et al.1993). Recently, the presence of dimers and probably tetramersof PSI was observed in the cyanobacterium Anabaena 7120 andin the glaucophyte Cyanophora paradoxa (Watanabe et al.2011).
In the vast majority of cyanobacteria, PSI RCs consist of 11protein subunits. In particular, PsaA and PsaB are the two cen-tral subunits of the heterodimeric core of the RC. Additionally,nine proteins with lower molecular weight (designated PsaC toPsaM) are involved in the formation of the PSI RC monomer(Fromme et al. 2001). PsaL, a 16 kDa hydrophobic protein sub-unit of the PSI RC, was identified as crucial for the formation ofPSI trimers (Chitnis and Chitnis 1993, Chitnis et al. 1993,Schluchter et al. 1996). Only the monomer form of PSI RCaccumulated in a PsaL-deficient Synechocystis mutant, demon-stranting the importance of PsaL in the formation of PSI trimers(Chitnis and Chitnis 1993). Several carotenoid residues havebeen identified in the vicinity of the PsaL-containing domainwithin a PSI trimer structure (Fromme et al. 2001, Sozer et al.2011). Recently, the formation of PSI trimer has been found tobe repressed in a carotenoidless crtHB mutant strain (Sozeret al. 2010).
It is commonly accepted that there exists a dynamicequilibrium between monomer and oligomer forms of PSI inthe photosynthetic membranes of Synechocystis PCC6803 andother cyanobacteria. However, the physiological significance ofPSI oligomerization remains unclear. What are known as ‘redChls’ were identified in Spirulina platensis. They absorb lightwavelengths>700 nm and were found to be preferentially asso-ciated with oligomers of PSI RCs (Karapetyan et al. 1999). It was
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199, Advance Access publication on 16 December 2014,available online at www.pcp.oxfordjournals.org! The Author 2014. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
Regu
larP
aper
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

postulated that such oligomers may facilitate radiationlessdissipation of excess energy as heat and consequently protectthe photosynthetic apparatus against light stress (Karapetyanet al. 1997, Karapetyan et al. 1999, Karapetyan 2008). Otherauthors proposed that in Spirulina PSI these oligomer-associated red Chls can funnel light energy to P700 (vanGrondelle et al. 1994, Shubin et al. 1995) and/or increasethe cross-section of light absorption in the long wavelengthregion (Trissl 1993). However, in Synechocystis PCC6803,which contains a limited pool of red Chls, no significantdifferences in energy transfer were detected either betweenChls within an inner PSI antenna or between PSI monomersand trimers (Turconi et al. 1996, Gobets and van Grondelle2001).
In cyanobacterial cells, the ‘state transition’ phenomenoncan take the form of the movement of phycobilisomes fromPSII RCs (state 1) to PSI RCs (state 2). In comparison with thewild type (WT), a Synechococcus mutant with a monomerizedPSI exhibited an accelerated state 1–state 2 transition(Schluchter et al. 1996). Iron deficiency induced the monomer-ization of PSI, as well as affecting the energy transfer in cyano-bacterial cells. Iron-stressed cells exhibited a reduced capacityfor state transitions and a limited dark reduction of the plasto-quinone pool (Ivanov et al. 2006). It was also suggested that the‘energy spillover’ occurring under environmental change fromlight to darkness could be triggered by a dissociation of PSItrimers into monomers, with the PSI monomers re-aggregatinginto trimers when the cells move from darkness to light, i.e. PSIoligomerization is reversibly regulated by light or its absence (Liet al. 2006).
Phosphatidylglycerol (PG) depletion, leading to a decrease inphotosynthetic activity (Hagio et al. 2000, Gombos et al. 2002),resulted in a complete elimination of PSI trimers (Domonkoset al. 2004). The PG depletion-induced PSI monomerizationprocess has been found to be accompanied by an enhancedsynthesis of xanthophylls, predominantly myxoxanthophyll(Domonkos et al. 2009). Recently, the crucial role of xantho-phylls as protective agents against various stresses in cyanobac-teria has been confirmed (Schafer et al. 2005, Zhu et al. 2010). Inparticular, the double mutant of Synechocystis, �crtRO, lackingechinenone, zeaxanthin and myxoxanthophyll, showed lowoxygen-evolving activity accompanied by increased light sensi-tivity (Schafer et al. 2005).
Temperature changes in the environment may inducedamage in living organisms. Photoautotrophic species needto be able to acclimate to ambient temperature conditions(Berry and Bjorkman 1980). In cyanobacteria, photosyntheticactivity remains constant with changing growth temperatures(Inoue et al. 2001). Two extrinsic proteins of of the PSII corecomplex, PsbU and PsbV, were reported to be associated withthe thermostability of PSII (Nishiyama et al. 1997, Nishiyamaet al. 1999). In Synechocystis PCC6803, it was found that thefluidity of thylakoid membranes plays an important role in theacclimation of PSII activity to the growth temperature(Aminaka et al. 2006). Recently, it was shown that the elimin-ation of xanthophylls affects the fluidity of thylakoid mem-branes in cyanobacteria (Kłodawska et al. 2012).
In the present study we demonstrate that in mesophilicSynechocystis PCC6803 cells, the PSI monomer to trimer ratiodepends on growth temperature. The carotenoid compositionin �psaL, a PSI trimer-deficient mutant, shows that PSImonomerization leads to an increased accumulation of myx-oxantophyll, zeaxanthin and echinenone, irrespective of tem-perature conditions. Moreover, circular dichroism (CD)spectroscopy in the visible range reveals that a peak attribut-able to long-wavelength-absorbing carotenoids is apparentlyenhanced in the trimer-forming WT cells. Our results suggestthat carotenoids may participate in the formation of the PSIoligomer structure.
Results
Inactivation of the psaL gene results in a PSItrimer-deficient mutant with no impairedcapacity for photoautotrophic growth
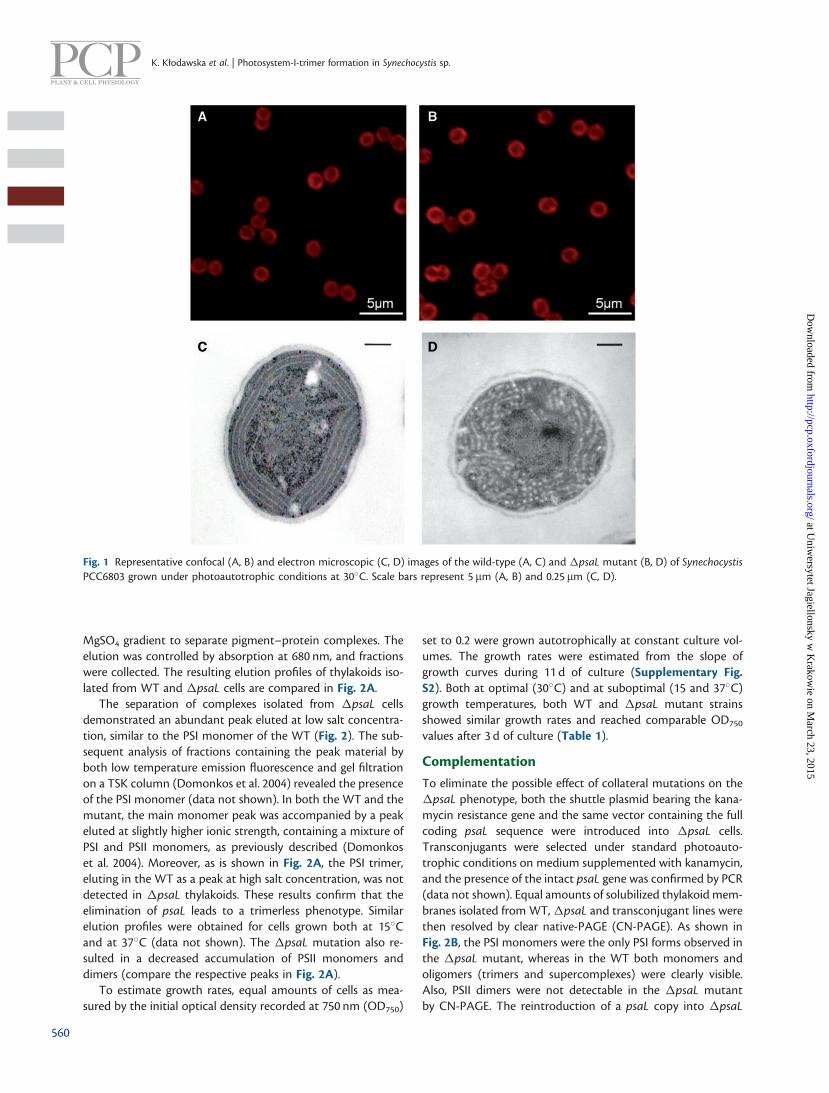
The small subunit of PSI, PsaL, encoded by the psaL gene haspreviously been shown to be indispensable for trimer formationin Synechocystis PCC6803 (Chitnis and Chitnis 1993). In order toanalyze the role of PSI trimer in cyanobacteria, we have createda mutant incapable of forming trimers. To this end, the psaLgene of Synechocystis was inactivated by replacing its codingregion with an � cassette conferring spectinomycin resistance.A fully segregated mutant was obtained (Supplementary Fig.S1). Mutant cells were grown under photoautotrophic condi-tions at 30�C in a medium supplemented with spectinomycin.They accumulated Chl and were not distinguishable from theWT by confocal microscopy (Fig. 1A, B). Also, the observedaverage cell diameter of �psaL cells (2.16 ± 0.11 mm) was notsignificantly different from that of the WT (2.19 ± 0.12 mm). Themorphology of both WT and mutant cells grown under sub-optimal conditions (at either 15 or 37�C) was similar to thatobserved at 30�C when analyzed microscopically (data notshown).
Analysis by electron microscopy revealed profound mor-phological changes associated with the inactivation of thepsaL gene in Synechocystis grown phototropically at 30�C.First, the space occupied by thylakoid membranes within cellsbecame relatively greater than in the WT. Further, in �psaLcells, the thylakoid membranes lost the regular structure thatcould be observed in the WT due to the accumulation of gran-ules expressing low electron density. These granules were loca-lized in the cytoplasm, separating particular thylakoid layers(Fig. 1C, D).
To analyze the effect of psaL deletion on PSI oligomerization,thylakoid membranes isolated from WT and �psaL mutantcells taken from the late logarithmic phase of culture weresolubilized with n-dodecyl-b-D-maltoside (b-DM). Under ourexperimental conditions, the solubilization protocol wasequally effective for membranes isolated from cells grown atdifferent temperatures, producing no visible pellet residue fol-lowing the final ultracentrifugation. Equal amounts of solubi-lized thylakoid membranes were then loaded onto the MonoQanion exchange column. The material was eluted with a linear
559
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
MgSO4 gradient to separate pigment–protein complexes. Theelution was controlled by absorption at 680 nm, and fractionswere collected. The resulting elution profiles of thylakoids iso-lated from WT and �psaL cells are compared in Fig. 2A.
The separation of complexes isolated from �psaL cellsdemonstrated an abundant peak eluted at low salt concentra-tion, similar to the PSI monomer of the WT (Fig. 2). The sub-sequent analysis of fractions containing the peak material byboth low temperature emission fluorescence and gel filtrationon a TSK column (Domonkos et al. 2004) revealed the presenceof the PSI monomer (data not shown). In both the WT and themutant, the main monomer peak was accompanied by a peakeluted at slightly higher ionic strength, containing a mixture ofPSI and PSII monomers, as previously described (Domonkoset al. 2004). Moreover, as is shown in Fig. 2A, the PSI trimer,eluting in the WT as a peak at high salt concentration, was notdetected in �psaL thylakoids. These results confirm that theelimination of psaL leads to a trimerless phenotype. Similarelution profiles were obtained for cells grown both at 15�Cand at 37�C (data not shown). The �psaL mutation also re-sulted in a decreased accumulation of PSII monomers anddimers (compare the respective peaks in Fig. 2A).
To estimate growth rates, equal amounts of cells as mea-sured by the initial optical density recorded at 750 nm (OD750)
set to 0.2 were grown autotrophically at constant culture vol-umes. The growth rates were estimated from the slope ofgrowth curves during 11 d of cuIture (Supplementary Fig.S2). Both at optimal (30�C) and at suboptimal (15 and 37�C)growth temperatures, both WT and �psaL mutant strainsshowed similar growth rates and reached comparable OD750
values after 3 d of culture (Table 1).
Complementation
To eliminate the possible effect of collateral mutations on the�psaL phenotype, both the shuttle plasmid bearing the kana-mycin resistance gene and the same vector containing the fullcoding psaL sequence were introduced into �psaL cells.Transconjugants were selected under standard photoauto-trophic conditions on medium supplemented with kanamycin,and the presence of the intact psaL gene was confirmed by PCR(data not shown). Equal amounts of solubilized thylakoid mem-branes isolated from WT, �psaL and transconjugant lines werethen resolved by clear native-PAGE (CN-PAGE). As shown inFig. 2B, the PSI monomers were the only PSI forms observed inthe �psaL mutant, whereas in the WT both monomers andoligomers (trimers and supercomplexes) were clearly visible.Also, PSII dimers were not detectable in the �psaL mutantby CN-PAGE. The reintroduction of a psaL copy into �psaL
Fig. 1 Representative confocal (A, B) and electron microscopic (C, D) images of the wild-type (A, C) and �psaL mutant (B, D) of SynechocystisPCC6803 grown under photoautotrophic conditions at 30�C. Scale bars represent 5 mm (A, B) and 0.25 mm (C, D).
560
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
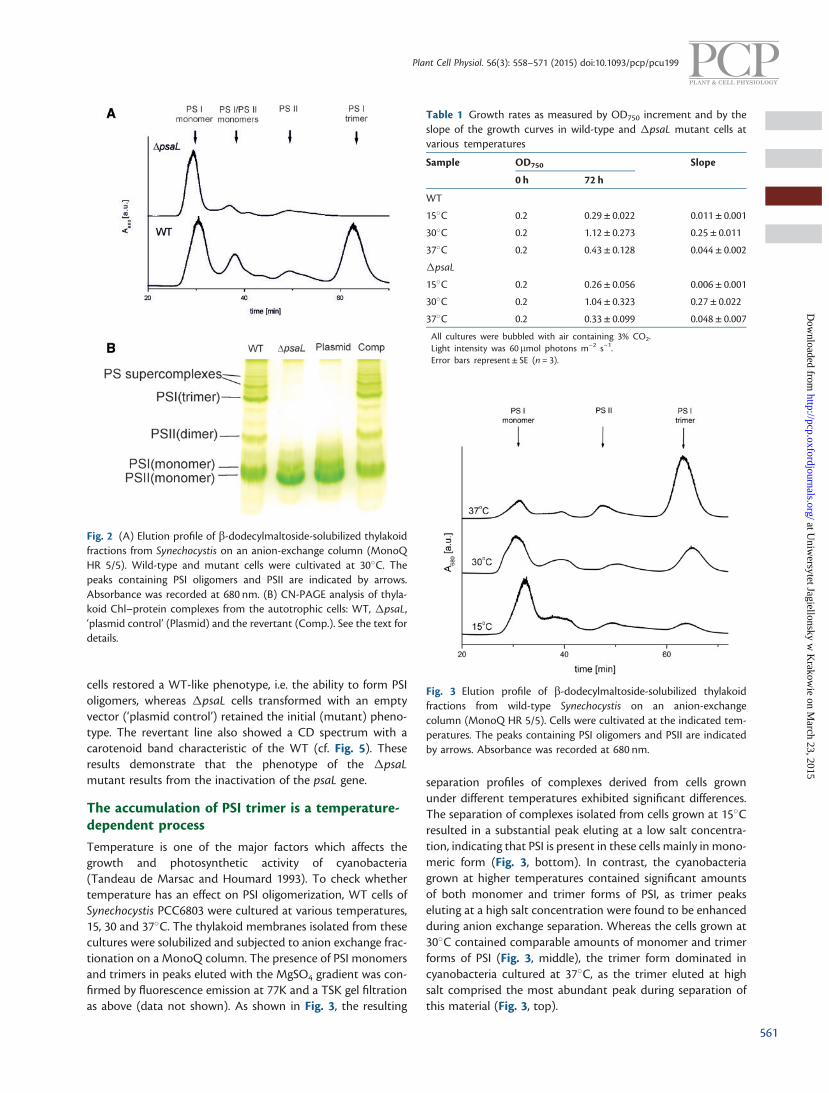
cells restored a WT-like phenotype, i.e. the ability to form PSIoligomers, whereas �psaL cells transformed with an emptyvector (‘plasmid control’) retained the initial (mutant) pheno-type. The revertant line also showed a CD spectrum with acarotenoid band characteristic of the WT (cf. Fig. 5). Theseresults demonstrate that the phenotype of the �psaLmutant results from the inactivation of the psaL gene.
The accumulation of PSI trimer is a temperature-dependent process
Temperature is one of the major factors which affects thegrowth and photosynthetic activity of cyanobacteria(Tandeau de Marsac and Houmard 1993). To check whethertemperature has an effect on PSI oligomerization, WT cells ofSynechocystis PCC6803 were cultured at various temperatures,15, 30 and 37�C. The thylakoid membranes isolated from thesecultures were solubilized and subjected to anion exchange frac-tionation on a MonoQ column. The presence of PSI monomersand trimers in peaks eluted with the MgSO4 gradient was con-firmed by fluorescence emission at 77K and a TSK gel filtrationas above (data not shown). As shown in Fig. 3, the resulting
separation profiles of complexes derived from cells grownunder different temperatures exhibited significant differences.The separation of complexes isolated from cells grown at 15�Cresulted in a substantial peak eluting at a low salt concentra-tion, indicating that PSI is present in these cells mainly in mono-meric form (Fig. 3, bottom). In contrast, the cyanobacteriagrown at higher temperatures contained significant amountsof both monomer and trimer forms of PSI, as trimer peakseluting at a high salt concentration were found to be enhancedduring anion exchange separation. Whereas the cells grown at30�C contained comparable amounts of monomer and trimerforms of PSI (Fig. 3, middle), the trimer form dominated incyanobacteria cultured at 37�C, as the trimer eluted at highsalt comprised the most abundant peak during separation ofthis material (Fig. 3, top).
Fig. 2 (A) Elution profile of b-dodecylmaltoside-solubilized thylakoidfractions from Synechocystis on an anion-exchange column (MonoQHR 5/5). Wild-type and mutant cells were cultivated at 30�C. Thepeaks containing PSI oligomers and PSII are indicated by arrows.Absorbance was recorded at 680 nm. (B) CN-PAGE analysis of thyla-koid Chl–protein complexes from the autotrophic cells: WT, �psaL,‘plasmid control’ (Plasmid) and the revertant (Comp.). See the text fordetails.
Fig. 3 Elution profile of b-dodecylmaltoside-solubilized thylakoidfractions from wild-type Synechocystis on an anion-exchangecolumn (MonoQ HR 5/5). Cells were cultivated at the indicated tem-peratures. The peaks containing PSI oligomers and PSII are indicatedby arrows. Absorbance was recorded at 680 nm.
Table 1 Growth rates as measured by OD750 increment and by theslope of the growth curves in wild-type and �psaL mutant cells atvarious temperatures
Sample OD750 Slope
0 h 72 h
WT
15�C 0.2 0.29 ± 0.022 0.011 ± 0.001
30�C 0.2 1.12 ± 0.273 0.25 ± 0.011
37�C 0.2 0.43 ± 0.128 0.044 ± 0.002
�psaL
15�C 0.2 0.26 ± 0.056 0.006 ± 0.001
30�C 0.2 1.04 ± 0.323 0.27 ± 0.022
37�C 0.2 0.33 ± 0.099 0.048 ± 0.007
All cultures were bubbled with air containing 3% CO2.Light intensity was 60mmol photons m–2 s–1.Error bars represent ± SE (n = 3).
561
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

The effect of PSI monomerization on theaccumulation of the PSI RC
To examine how the mutation affected photosynthetic lightharvesting and energy transfer, we measured Chl fluorescenceparameters in WT and �psaL cells. Low temperature fluores-cence emission spectra collected at 77K provided evidence thatthe energy transfer to the RCs of PSI was not distorted in the�psaL mutant, as indicated by a significant emission at around725 nm (Fig. 4A). The comparison of spectra normalized at684 nm (the emission maximum of PSII RCs) revealed thatthe relative PSI/PSII ratio was not significantly affected ineither WT or mutant cells grown at different temperatures,although it was slightly lower in �psaL cells. The only exceptionwas �psaL grown at 15�C, where this parameter was remark-ably decreased (Fig. 4A). The immunoblotting analysis indi-cated that �psaL mutation had no remarkable effect on theaccumulation of the RC core proteins PsaA and PsbA in cells
grown at 30�C (Fig. 4B). In both WT and mutant cells, theaccumulation of PsaA and PsbA apparently increasedwith the higher growth temperature; however, significant dif-ferences between the WT and �psaL mutant were notobserved (Fig. 4C).
PSI monomerization results in changes in thecarotenoid absorption region of the CD spectrum
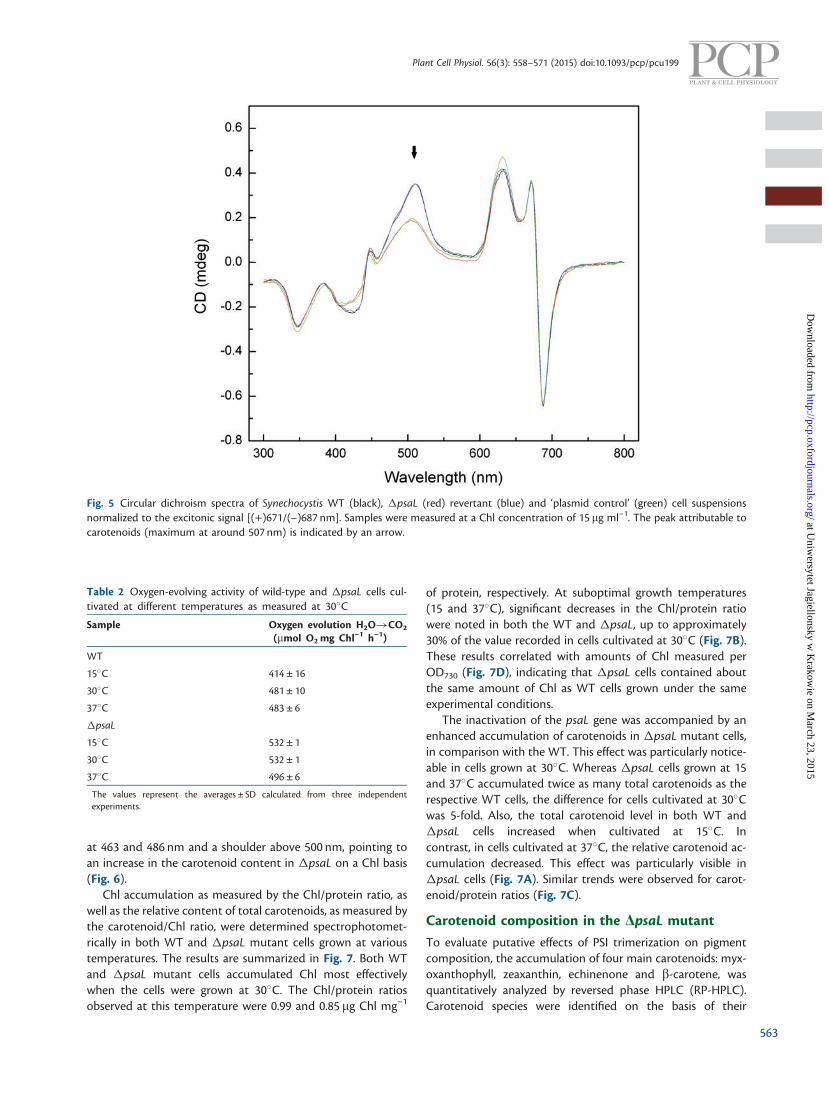
The number and type of pigment molecules bound in the pig-ment–protein complexes as well as their structural arrange-ment are crucial for the functioning of the photosyntheticapparatus. To study the effect of PSI oligomerization on overallpigment organization in complexes, WT and �psaL cell sus-pensions grown at 30�C were analyzed by CD spectroscopy inthe 300–800 nm region. The CD spectra obtained were normal-ized to the (+)671/(–)687 excitonic signal. The results areshown in Fig. 5. In both WT and �psaL cell suspensions,both the inflection point of excitonic split signals andmaxima of CD bands coinciding with the corresponding ab-sorption maxima of the main photosynthetic pigments wereclearly visible. In particular, an excitonic band at +671/–687 isattributable to Chl. The maximum at 625 nm corresponds tophycobilisomes and that at 507 nm to the absorption range ofcarotenoids. As shown in Fig. 5, the CD band in the carotenoidabsorption spectral range of �psaL cells is significantly affected,indicating chirality changes associated with carotenoids. In thepsaL revertant strain, this band was restored, reaching the rela-tive amplitude and width comparable with that of the WT. Itconfirms that the observed changes in carotenoid-associatedchirality in the �psaL mutant are caused by PSImonomerization.
Oxygen-evolving activity of the "psaL mutant
Non-cyclic electron transport from water to carbon dioxide wasmeasured as oxygen-evolving activity via PSII and PSI in WT and�psaL mutant cells grown at different temperatures. To assessputative differences in the overall capacity for photosyntheticelectron transport in cells grown at various temperatures, allmeasurements were performed under standardized conditionsat a constant temperature (30�C), which is close to optimum.As can be seen in Table 2, �psaL mutant cells showed slightlyhigher oxygen-evolving activity than the WT irrespective ofgrowth temperature. There were no statistically significant dif-ferences observed in H2O!CO2 electron transport betweenWT and �psaL mutant cells grown at 30 and 37�C, althoughthe values measured for the mutant were slightly higher. In WTcells grown at 15�C, oxygen-evolving activity from H2O to CO2
decreased (�16%) in comparison with the values observed forcells cultivated at 30 and 37�C. In �psaL mutant cells, thiseffect was not observed.
Pigment accumulation in the WT and "psaLmutant
In comparison with the WT, the absorption spectrum of �psaLcell suspensions grown at 30�C showed a relative increase inabsorption level in the blue wavelength region, exhibiting bands
Fig. 4 (A) Temperature-dependent regulation of the PSI/PSII ratio inWT and �psaL mutant cells. 77K fluorescence emission spectra ofSynechocystis WT (left panel) and �psaL mutant (right panel) cellsgrown under identical growth conditions at various temperatures. Thespectra were normalized at 684 nm and the fluorescence emission at684 nm was set as 1. The excitation wavelength was 437 nm. (B and C)Immunoblotting analysis of the relative accumulation of the PsaA andPsbA proteins: in WT, �psaL and revertant (Complemented) strainsgrown at 30�C (B); and in WT and �psaL strains grown at 37 and 15�C(C). See the text for details.
562
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
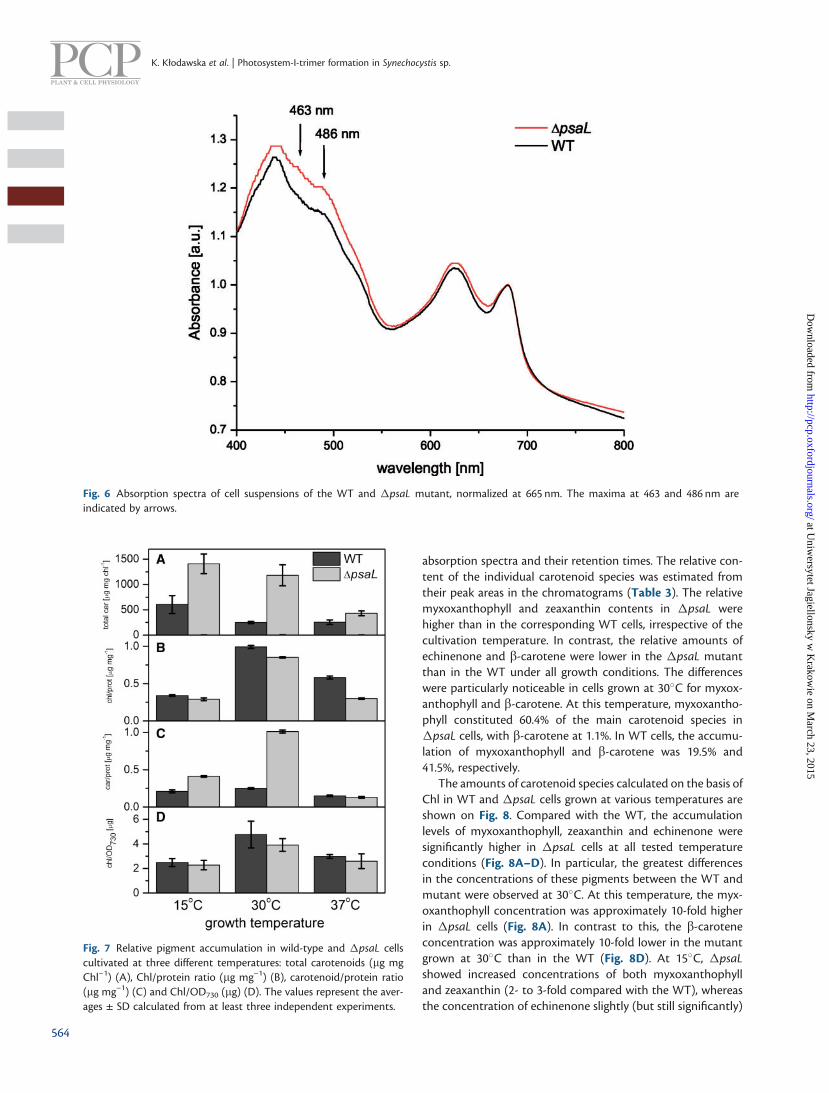
at 463 and 486 nm and a shoulder above 500 nm, pointing toan increase in the carotenoid content in �psaL on a Chl basis(Fig. 6).
Chl accumulation as measured by the Chl/protein ratio, aswell as the relative content of total carotenoids, as measured bythe carotenoid/Chl ratio, were determined spectrophotomet-rically in both WT and �psaL mutant cells grown at varioustemperatures. The results are summarized in Fig. 7. Both WTand �psaL mutant cells accumulated Chl most effectivelywhen the cells were grown at 30�C. The Chl/protein ratiosobserved at this temperature were 0.99 and 0.85 mg Chl mg–1
of protein, respectively. At suboptimal growth temperatures(15 and 37�C), significant decreases in the Chl/protein ratiowere noted in both the WT and �psaL, up to approximately30% of the value recorded in cells cultivated at 30�C (Fig. 7B).These results correlated with amounts of Chl measured perOD730 (Fig. 7D), indicating that �psaL cells contained aboutthe same amount of Chl as WT cells grown under the sameexperimental conditions.
The inactivation of the psaL gene was accompanied by anenhanced accumulation of carotenoids in �psaL mutant cells,in comparison with the WT. This effect was particularly notice-able in cells grown at 30�C. Whereas �psaL cells grown at 15and 37�C accumulated twice as many total carotenoids as therespective WT cells, the difference for cells cultivated at 30�Cwas 5-fold. Also, the total carotenoid level in both WT and�psaL cells increased when cultivated at 15�C. Incontrast, in cells cultivated at 37�C, the relative carotenoid ac-cumulation decreased. This effect was particularly visible in�psaL cells (Fig. 7A). Similar trends were observed for carot-enoid/protein ratios (Fig. 7C).
Carotenoid composition in the "psaL mutant
To evaluate putative effects of PSI trimerization on pigmentcomposition, the accumulation of four main carotenoids: myx-oxanthophyll, zeaxanthin, echinenone and b-carotene, wasquantitatively analyzed by reversed phase HPLC (RP-HPLC).Carotenoid species were identified on the basis of their
Fig. 5 Circular dichroism spectra of Synechocystis WT (black), �psaL (red) revertant (blue) and ‘plasmid control’ (green) cell suspensionsnormalized to the excitonic signal [(+)671/(–)687 nm]. Samples were measured at a Chl concentration of 15 mg ml–1. The peak attributable tocarotenoids (maximum at around 507 nm) is indicated by an arrow.
Table 2 Oxygen-evolving activity of wild-type and �psaL cells cul-tivated at different temperatures as measured at 30�C
Sample Oxygen evolution H2O!CO2
(mmol O2 mg Chl–1 h–1)
WT
15�C 414 ± 16
30�C 481 ± 10
37�C 483 ± 6
�psaL
15�C 532 ± 1
30�C 532 ± 1
37�C 496 ± 6
The values represent the averages ± SD calculated from three independentexperiments.
563
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
absorption spectra and their retention times. The relative con-tent of the individual carotenoid species was estimated fromtheir peak areas in the chromatograms (Table 3). The relativemyxoxanthophyll and zeaxanthin contents in �psaL werehigher than in the corresponding WT cells, irrespective of thecultivation temperature. In contrast, the relative amounts ofechinenone and b-carotene were lower in the �psaL mutantthan in the WT under all growth conditions. The differenceswere particularly noticeable in cells grown at 30�C for myxox-anthophyll and b-carotene. At this temperature, myxoxantho-phyll constituted 60.4% of the main carotenoid species in�psaL cells, with b-carotene at 1.1%. In WT cells, the accumu-lation of myxoxanthophyll and b-carotene was 19.5% and41.5%, respectively.
The amounts of carotenoid species calculated on the basis ofChl in WT and �psaL cells grown at various temperatures areshown on Fig. 8. Compared with the WT, the accumulationlevels of myxoxanthophyll, zeaxanthin and echinenone weresignificantly higher in �psaL cells at all tested temperatureconditions (Fig. 8A–D). In particular, the greatest differencesin the concentrations of these pigments between the WT andmutant were observed at 30�C. At this temperature, the myx-oxanthophyll concentration was approximately 10-fold higherin �psaL cells (Fig. 8A). In contrast to this, the b-caroteneconcentration was approximately 10-fold lower in the mutantgrown at 30�C than in the WT (Fig. 8D). At 15�C, �psaLshowed increased concentrations of both myxoxanthophylland zeaxanthin (2- to 3-fold compared with the WT), whereasthe concentration of echinenone slightly (but still significantly)
Fig. 6 Absorption spectra of cell suspensions of the WT and �psaL mutant, normalized at 665 nm. The maxima at 463 and 486 nm areindicated by arrows.
Fig. 7 Relative pigment accumulation in wild-type and �psaL cellscultivated at three different temperatures: total carotenoids (mg mgChl–1) (A), Chl/protein ratio (mg mg–1) (B), carotenoid/protein ratio(mg mg–1) (C) and Chl/OD730 (mg) (D). The values represent the aver-ages ± SD calculated from at least three independent experiments.
564
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
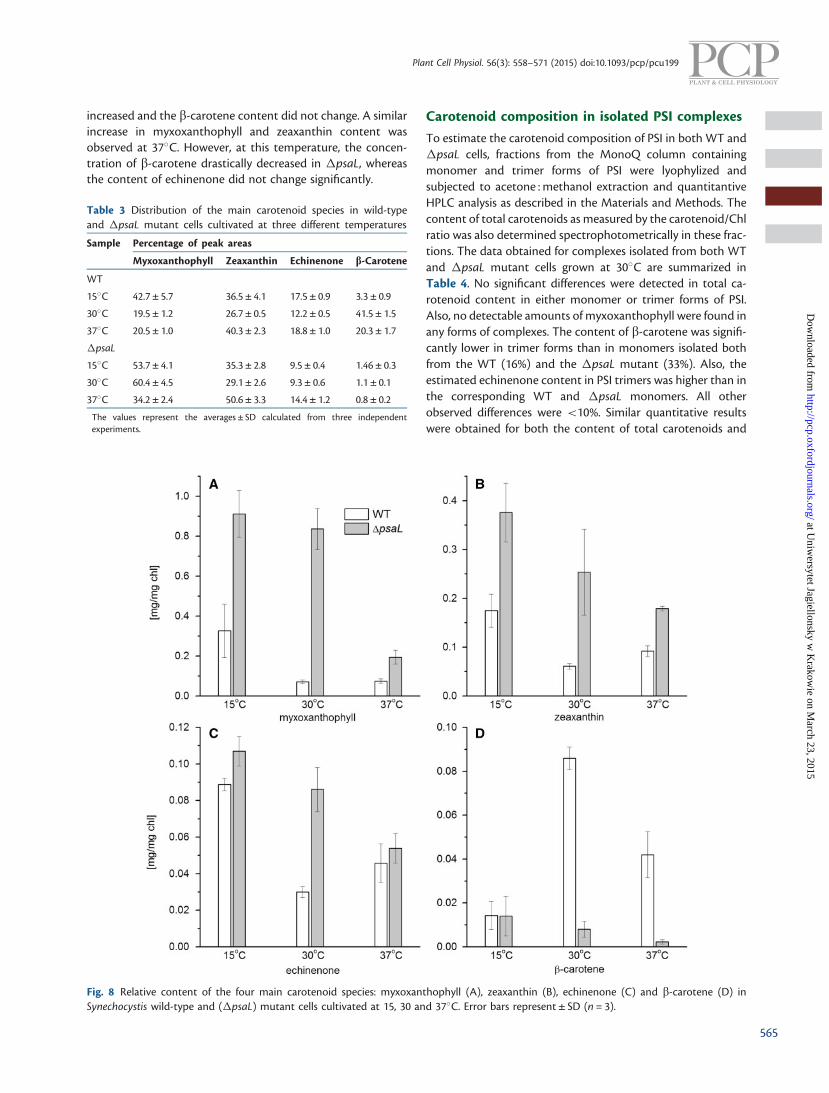
increased and the b-carotene content did not change. A similarincrease in myxoxanthophyll and zeaxanthin content wasobserved at 37�C. However, at this temperature, the concen-tration of b-carotene drastically decreased in �psaL, whereasthe content of echinenone did not change significantly.
Carotenoid composition in isolated PSI complexes
To estimate the carotenoid composition of PSI in both WT and�psaL cells, fractions from the MonoQ column containingmonomer and trimer forms of PSI were lyophylized andsubjected to acetone : methanol extraction and quantitantiveHPLC analysis as described in the Materials and Methods. Thecontent of total carotenoids as measured by the carotenoid/Chlratio was also determined spectrophotometrically in these frac-tions. The data obtained for complexes isolated from both WTand �psaL mutant cells grown at 30�C are summarized inTable 4. No significant differences were detected in total ca-rotenoid content in either monomer or trimer forms of PSI.Also, no detectable amounts of myxoxanthophyll were found inany forms of complexes. The content of b-carotene was signifi-cantly lower in trimer forms than in monomers isolated bothfrom the WT (16%) and the �psaL mutant (33%). Also, theestimated echinenone content in PSI trimers was higher than inthe corresponding WT and �psaL monomers. All otherobserved differences were <10%. Similar quantitative resultswere obtained for both the content of total carotenoids and
Fig. 8 Relative content of the four main carotenoid species: myxoxanthophyll (A), zeaxanthin (B), echinenone (C) and b-carotene (D) inSynechocystis wild-type and (�psaL) mutant cells cultivated at 15, 30 and 37�C. Error bars represent ± SD (n = 3).
Table 3 Distribution of the main carotenoid species in wild-typeand �psaL mutant cells cultivated at three different temperatures
Sample Percentage of peak areas
Myxoxanthophyll Zeaxanthin Echinenone b-Carotene
WT
15�C 42.7 ± 5.7 36.5 ± 4.1 17.5 ± 0.9 3.3 ± 0.9
30�C 19.5 ± 1.2 26.7 ± 0.5 12.2 ± 0.5 41.5 ± 1.5
37�C 20.5 ± 1.0 40.3 ± 2.3 18.8 ± 1.0 20.3 ± 1.7
�psaL
15�C 53.7 ± 4.1 35.3 ± 2.8 9.5 ± 0.4 1.46 ± 0.3
30�C 60.4 ± 4.5 29.1 ± 2.6 9.3 ± 0.6 1.1 ± 0.1
37�C 34.2 ± 2.4 50.6 ± 3.3 14.4 ± 1.2 0.8 ± 0.2
The values represent the averages ± SD calculated from three independentexperiments.
565
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

the contents of the main carotenoid species analyzed in com-plexes isolated from cells grown at 15 and 37�C and there wereno other significant differences observed (data not shown).
Discussion
The formation of PSI oligomers is a unique feature of cyano-bacteria. One of the low molecular weight subunits of PSI—PsaL—has previously been shown to be essential for theformation of trimers in Synechocystis PCC6803 (Chitnis andChitnis 1993). To study the morphological and physiologicaleffects accompanying PSI oligomerization, we generated aPsaL-deficient mutant incapable of forming PSI trimers.
Effects of PSI monomerization
The complete elimination of the psaL gene affected neither thecapacity for photoautotrophic growth nor cell morphology. Infact, cell shapes as analyzed by confocal microscopy and theaccumulation of Chl as measured per OD730 were similar in WTand �psaL cells irrespective of the growth conditions. Theseresults are in agreement with earlier observations of other linesof psaL mutants bearing partial deletions in the psaL codingregion (Chitnis and Chitnis 1993, Chitnis et al. 1993).
The electron microscopic analysis carried out in this studyrevealed low electron density granules in the cytoplasm alongthe thylakoid membranes of �psaL cells grown at 30�C. Incyanobacteria, these granules have previously been demon-strated to be glycogen deposits (Frosch et al. 1987). The pres-ence of an aberrant glycogen accumulation suggests that thepsaL deletion may affect carbon assimilation under photoauto-trophic conditions. A similar phenotype has been found for ahik8 mutant of Synechocystis, without a gene encoding histidinekinase (Singh and Sherman 2005). This gene was differentiallyregulated in response to oxidative stress and to changing ironconcentrations (Singh et al. 2003, Li et al. 2004). An aberrantaccumulation of glycogen granules was also observed inSynechocystis mutants with an impaired sll0822 gene encodinga transcriptional regulator to control the expression levels ofnitrogen-related genes (Yamauchi et al. 2011). Thus, our result
suggests that the monomerization of PSI could be accompaniedby profound changes in the activity of the cellular regulatorypathways in cyanobacteria, so that its metabolism behaves as itdoes under stress conditions.
In the PsaL-deficient mutant, PSI monomers could be theonly PSI complex forms, as demonstrated by anion-exchangefractionation and by CN-PAGE of solubilized thylakoid mem-branes isolated from �psaL cells grown under photoauto-trophic conditions. This is consistent with the findings ofother authors using electrophoretic methods (Chitnis andChitnis 1993). Together with our previous analysis of PSI oligo-merization in the thylakoid membranes of a PG-deficientSynechocystis mutant (Domonkos et al. 2004), this studyshows the applicability of the anion-exchange fractionationtechnique to a semi-quantitative estimation of PSI monomer/trimer ratios under various experimental conditions.
The analysis of the PSI-less/apcE– mutant demonstratedthat PSII monomers were the dominant PSII form in the thyla-koid membranes in the absence of PSI, thus revealing the in-stability of active PSII dimers in the absence of interacting PSIcomplexes (Laczko-Dobos et al. 2011). The significant decreasein accumulation of PSII complexes observed in �psaL cells is inagreement with this observation and shows that PSI monomer-ization is sufficient to decrease the ability of PSII to form dimers.
Chl–protein complexes contain pigments which can serveas a label for generating a CD signal to provide informationabout modifications in protein structure and protein–proteininteractions (Lambrev et al. 2007). In both WT and �psaL cellsuspensions, it was possible to distinguish chirality maximacorresponding to the absorption maxima of the main photo-synthetic pigments. In particular, a complex CD band in the450–550 nm region with a maximum at around 507 nm, i.e. inthe absorption range of carotenoids, has been identified (Garaband van Amerongen 2009). This band was previously observedin PSI complexes from Synechococcus elongatus separated elec-trophoretically (Komenda et al. 1989). In comparison with WTcells, �psaL cells grown at 30�C showed remarkably lower chir-ality in this short wavelength spectral range, which is related tocarotenoids. As WT cells grown at 30�C accumulated signifi-cant amounts of PSI trimers and since there were no significantdifferences in the accumulation of either PSI RCs or Chl be-tween the WT and a mutant grown under optimal conditions,one can assume that this difference in chirality is a consequenceof changes in the PSI monomer/trimer ratio. We suggest thatthe intrinsic asymmetry level of carotenoid molecules boundwithin the PSI RC could be associated with its oligomeric struc-ture. The equilibrium shift to the monomer form of PSI resultsin a decrease in the chirality level at a band with a maximum of507 nm.
Carotenoids are important pigments present in the nativePSI RCs of cyanobacteria (El-Mohsnawy et al. 2010). The quan-titative analysis of carotenoid content in monomer and trimerPSI complexes purified from WT and �psaL cells demonstratedthat in PSI trimers the b-carotene content significantlydecreased, whereas the content of echinenone increased incomparison with corresponding WT and �psaL monomers.In Synechocystis, carotenoid deficiency has recently been
Table 4 Distribution of carotenoid species in isolated PSI mono-mers and trimers from wild-type cells (WT) and PSI monomersfrom �psaL mutant cells
WT "psaL
Monomer PSI Trimer PSI Monomer PSI
Total carotenoids 0.0816 ± 0.0040 0.0895 ± 0.0033 0.0916 ± 0.0035
Myxoxanthophyll ND ND ND
Zeaxanthin 0.0024 ± 0.0009 0.0024 ± 0.0001 0.0029 ± 0.0005
Echinenone 0.0102 ± 0.0010 0.0279 ± 0.0026* 0.0098 ± 0.0006
b-carotene 0.0690 ± 0.0022 0.0593 ± 0.0006* 0.0789 ± 0.0024
Values are given as mg (mg Chl)–1.Significant differences are indicated by an asterisk.ND, not detected.The values represent the averages ± SD calculated from three independentexperiments.
566
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

shown to decrease the synthesis and assembly of the PSII RC(Sozer et al. 2010, Sozer et al. 2011). Here we show that theformation of trimer structures may require the recruitment ofspecific carotenoid molecules and/or may be accompanied by apartial modification of the carotenoid molecules bound in PSImonomers.
In Synechocystis, the monomerization of PSI under bioticstress induced by PG depletion (Domonkos et al. 2004) haspreviously been shown to be accompanied by profoundchanges in the accumulation of carotenoid pigments(Domonkos et al. 2009). A comparison of the absorption spec-tra of intact cells, normalized on the basis of Chl, revealed anapparent accumulation of carotenoid pigments in the �psaLmutant. Subsequent quantitative analysis indicated that the�psaL cells accumulated significantly higher amounts of totalcarotenoids than WT cells, irrespective of the growth tempera-ture. This carotenoid accumulation was particularly dramatic in�psaL cells growing at optimal temperature, being >4-foldgreater than that in WT cells. The elimination of PSI trimerswas also accompanied by slight, but significant, decreases in theChl/protein ratio, observed under all temperature conditions.These results indicate that some carotenoid derivatives may beoverproduced in response to the monomerization of the PSIRCs in �psaL cells.
Subsequent HPLC-based analysis of carotenoid compositionrevealed that the accumulation levels of myxoxanthophyll,zeaxanthin and echinenone were significantly higher in�psaL cells under all growth temperatures. The overaccumula-tion of xanthophylls was accompanied by a significant decreasein b-carotene content in the mutant, observed at both 30 and37�C. In Synechocystis, both myxoxanthophyll and b-caroteneare formed from g-carotene by a branching of the biosyntheticpathway. Two other xanthophylls—zeaxanthin and echine-none—are products of b-carotene oxygenation in the nexttwo separate pathway branches (Lagarde et al. 2000). Thechanges in carotenoid composition observed in �psaL cellscould be explained by the higher stability of specific carotenoidspecies, namely xanthopylls, at different growth temperatures(Staiger et al. 1999) and/or by differences in the activities of theenzymes involved in the biosynthesis of particular carotenoidderivatives (Takaichi and Mochimaru 2007).
In all photosynthetic organisms, including cyanobacteria,xanthophylls function as protective agents against damagecaused by the reactive oxygen species (ROS), which are gener-ated under stress conditions (Hirschberg and Chamovitz 1994,Sajilata et al. 2008). In particular, myxoxanthophyll was foundto be preferentially synthesized at high light intensities (Steigeret al. 1999). It has an established role as a protectant againstboth light (Steiger et al. 1999) and cold (Varkonyi et al. 2002)stress. Myxoxanthophyll overaccumulation has been demon-strated in Synechocystis PCC6803 cells exposed to high intensitylight (Mohamed and Vermaas 2004). Recently, based on thegrowth defects observed in mutants of Synechococcus sp.strain PCC7002 at high light intensities, the protective role ofxanthophyll carotenoids, particularly zeaxanthin and echine-none, has been demonstrated (Zhu et al. 2010). An increasein the content of myxoxanthophyll and echinenone has also
been shown upon biotic stress accompanied by PG deficiencyin Synechocystis PCC6803 (Domonkos et al. 2009). Thus,the overaccumulation of xanthophylls in trimerless �psaLcells indicates that cyanobacteria with monomerized PSI aresubjected to an unknown stress-inducing mechanism thatcould be, at least partially, compensated by the enhancedsynthesis of protective compounds. The decrease in b-carotenecontent in �psaL cells further suggests that xanthophyllsare formed at the expense of this carotenoid, in a similarway to that observed during PG deficiency (Domonkos et al.2009).
Temperature effects on PSI oligomerization
As a mesophilic organism, Synechocystis has been found togrow in the temperature range of 15–43�C, with a temperatureoptimum at around 30�C. Both below 20�C and above 40�Cthere are remarkable stress symptoms (Stanier et al. 1971, Inoueet al. 2001). Elevation of the temperature promoted the accu-mulation of PSI in trimer form. The monomer/trimer ratiohas previously been shown to be affected by both biotic andabiotic factors. In particular, PSI monomerization may beinduced by a deficiency of PG (Domonkos et al. 2004), by aniron deficiency (Ivanov et al. 2006), by the inhibition of carot-enoid synthesis (Sozer et al. 2010) or by a manganese deficiencyin the growth medium (Salomon and Keren 2011). A role foriron deficiency-induced proteins (IsiA) (Bibby et al. 2001,Boekema et al. 2001) and polypeptides induced by high light(HLIPs) (Wang et al. 2008) in PSI trimer stabilization has beenpostulated. In this work, for the first time, the correlation be-tween growth temperature and PSI oligomerization in cyano-bacteria is shown.
The relative amounts of functional PSI and PSII can be esti-mated by Chl fluorescence emission at 77K (Murakami 1997). Incyanobacteria, the PSI/PSII ratio has been found to be environ-mentally regulated (Fujita et al. 1985, Fujita et al. 1987, Fujitaet al. 1988, Murakami et al. 1997). In particular, it decreases incells grown at high light intensities, due to the down-regulationof PSI accumulation in membranes, whereas the PSII contentremains unchanged (Murakami and Fujita 1993, Fuhrmannet al. 2009). Fluorescence analysis of WT and �psaL cellsdemonstrated that only in �psaL grown at 15�C did the PSI/PSII ratio significantly decrease. The accumulation of PSI inSynechocystis has previously been found to be brought aboutin response to high intensities of light and to be precisely regu-lated at the genetic level (Hihara et al. 1998, Muramatsu andHihara 2012). In particular, excessive light fluences caused adecrease in the PSI trimer concentration (Fuhrmann et al.2009, Muramatsu and Hihara 2012). The relative decrease intrimer accumulation at low temperatures might be the result ofthe increased light sensitivity of the photosynthetic apparatusunder stress conditions (Huner et al. 1998, Miskiewicz et al.2000). Interestingly, the decrease in the PSI/PSII ratio in themutant at 15�C as measured by fluorescence emission at 77Kwas not accompanied by the respective change in the PsaA/PsbA ratio. This result indicates that in the mutant grown at15�C the energy transfer to the PSI RC is less effective than inthe WT. It may be caused by distorted complexity of both PSI
567
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

and PSII (see above) accompanied by a decreased relative Chlcontent in the mutant at 15�C.
In Synechocystis, the P700 turnover was previously found tobe affected in mutants with monomerized PSI grown underphotoheterotrophic conditions. Consequently, the existenceof an unidentified mechanism, which preferentially donateselectrons to P700 in the absence of PsaL, has been postulated(Chitnis et al. 1993). Under our experimental conditions,in comparison with the WT, the H2O!CO2 oxygen evolutionrates in �psaL cells were generally higher, irrespectiveof growth temperature. This finding suggests that the de-activation of the psaL gene affects the light phase of photosyn-thesis at its non-cyclic electron transport stage. In cyanobac-teria, the accumulation of PSI RCs has previously been shown tobe regulated by the electron influx from earlier steps of theelectron transport chain (Fujita et al. 1988). There is a relativeincrease in oxygen-evolving activity along with a decreased ac-cumulation of PSI in a trimerless mutant. This implies that theability of PSI to form oligomers may modulate the overallphotosynthetic activity in Synechocystis in response to envir-onmental conditions.
Conclusion
On the basis of these results, we propose that the PSI trimer tomonomer ratio in Synechocystis is determined by the growthtemperature. The PSI oligomerization is promoted at supraop-timal temperatures. Although the trimer form of PSI is notessential for photosynthetic activity and growth under optimalconditions, the lack of PSI trimerization in the �psaL mutantresults in stress which could be compensated by an increasedaccumulation of xanthophylls formed at the expense ofb-carotene. The enhanced CD signature of long-wavelength-absorbing carotenoids in the trimer-accumulating WT cells sug-gests that carotenoids may participate in the formation of thePSI oligomer structure. The monomer/trimer ratio may modu-late the photosynthetic activity of cyanobacteria in response togrowth temperature.
Materials and Methods
Organism and culturing conditions
The strains used in this study are derived from Synechocystis sp. PCC6803, which
is referred to as the WT. WT and �psaL cells were grown photoautotrophically
in BG11 medium (Allen 1968) supplemented with 5 mM HEPES-NaOH (pH 7.5)
under constant illumination, supplied by daylight fluorescent lamps (Philips TL-
D 18W/33-640), with a fluence rate of 60 mmol photons m–2 s–1 at tempera-
tures of 15, 30 and 37�C. Cultures were continuously aerated with air containing
approximately 3% carbon dioxide. The growth of cyanobacteria was monitored
based on the OD750. The material for experiments was taken from the late-
logarithmic phase of cultures, usually on the fifth day for cells grown at 30�C
and on the 10th day for those grown at 15 and 37�C.
Construction of the Synechocystis sp. PCC6803"psaL mutant strain and complementationof the "psaL mutation
The flanking regions of the psaL gene coding sequence were amplified by PCR
with the following two sets of primers: 50-TTTAATCCCGGCAATTATCC-30
(psaL_F) and 50-CGC(GGATCC)TGGTATTGAGTTCTCCTATTAAAG-30
(psaL_FR), and 50-CCAACCAGATAATTTCATAG-30 (psaL_R) and 50-CGC(GG
ATCC)ATAACCTTATAACCTTTAAAC-30 (psaL_RF). BamHI sites (in parenth-
eses) were added to the 50 regions of the psaL_FR and psaL_RF primers. The
amplified products were cut with the restriction enzyme BamHI and ligated
into the EcoRV site of pBluescript II. Then an omega cassette was cloned into
the BamHI site of the resulting plasmid (Supplementary Fig. S1A). This con-
struct was used to transform WT cells of Synechocystis sp. PCC6803. The trans-
formants were selected under photoautotrophic growth conditions on BG11
agar plates supplemented with increasing amounts of spectinomycin by several
restreakings of single colonies. Complete segregation of �psaL mutant cells was
confirmed by PCR using the primers psaL_F and psaL_R (Supplementary Fig.
S1B). For the complementation test, a shuttle vector was constructed by clon-
ing the kanamycin resistance gene from pUC4-KIXX (Pharmacia) between nu-
cleotides 7,772 and 2,118 of the broad host range plasmid RSF1010. The
sequence of the psaL gene was amplified by PCR with the primers: 50-TTTGA
AGATTGACCAACGGCGATCGGAC-30 and 50-AGGAATCCTGACCCACCCAA
AGATGGTTAG-30 , and cloned into the shuttle vector downstream of the kana-
mycin resistance gene. The constructs were introduced into the �psaL mutant
by conjugation.
Measurement of photosynthetic activity
For the measurement of oxygen evolution, the cells were washed with BG11
medium and resuspended in fresh BG11 medium to reach a final Chl concen-
tration of 5 mg ml–1. The total oxygen-evolving activity in intact cells was
measured in 2 ml samples, with constant stirring at room temperature with a
Clark-type oxygen electrode (Hansatech Instruments) as described by Gombos
et al. (2002). The saturating light (fluence rate 500 mmol photons m–2 s–1) was
provided from an incandescent lamp equipped with an optical filter system,
separating a 600–680 nm band.
Isolation of thylakoid membranes and clearnative-PAGE
The thylakoid fraction was isolated from WT and mutant cells according to
Komenda and Barber (1995). All subsequent steps of the isolation procedure
were carried out at 4�C. Briefly, Synechocystis PCC6803 cells were collected by
centrifugation at 2,500�g for 10 min and resuspended in buffer A containing
50 mM MES (pH 6.5), 2 mM e-amino-caproic acid, 5 mM EDTA, 1 mM phenyl-
methylsulfonyl fluoride (PMSF) and 1 mM benzamidine. The suspension was
mixed with an equal volume of 0.1 mm glass beads and placed into a Bead
Beater homogenizer (BioSpec Products). Cell disruption was performed six
times for 1 min. Intact cells were removed by centrifugation at 2,000�g for
5 min, and the supernatant was centrifuged again at 20,000�g for 30 min.
Pelleted thylakoids were resuspended in buffer B containing 20 mM MES (pH
6.5), 10 mM CaCl2, 10 mM MgCl2, 0.5 M mannitol, 5 mM MgSO4 and 0.03% (w/
v) b-DM, and stored at –80�C.
For CN-PAGE, pelleted thylakoids were resuspended in a buffer containing
25 mM MES/NaOH, pH 6.5 containing 10 mM CaCl2, 10 mM MgCl2 and 25%
glycerol (Komenda et al. 2005). Samples were solubilized with 1% b-DM and
analyzed by CN-PAGE at 4�C in a 5–13% gradient gel as described by Komenda
et al. (2012). Each loaded sample contained 4 mg of Chl.
Immunoblotting
Isolated thylakoid membranes were solubilized in 2� Laemmli buffer and the
proteins were separated by Tris-glycine SDS–PAGE. Aliquots equivalent to
0.5 mg of Chl were loaded onto a 15% polyacrylamide gel. Following separation,
the proteins were blotted onto a nitrocellulose membrane with a semi-dry
blotting set-up system using methanol-containing buffer (25 mM Tris,
192 mM glycine, 20% methanol). The nitrocellulose membranes were blocked
with 10 mM Tris, pH 8.0/150 mM NaCl/0.1% Tween-20 (TBST) buffer contain-
ing 5% skimmed milk powder for 1 h. The membranes were incubated with
PsbA and PsaA primary antibodies (Agrisera) for 1 h. Primary antibodies were
diluted to 1 : 3,000 in TBST buffer. The nitrocellulose membranes were washed
three times with TBST buffer and incubated with horseradish peroxidase (HRP)-
conjugated secondary antibodies (goat anti-rabbit IgG–HRP, Millipore) at
568
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

1 : 5,000 dilutions. Immunoblotted membranes were incubated for 5 min in ECL
plus HRP substrate (GE Healthcare Bio-Sciences) and chemiluminescence was
detected on Hyperfilm ECL photographic film (GE Healthcare Bio-Sciences).
PSI monomer and trimer fractionation
Monomer and trimer PSI pigment–protein complexes were separated as
described in Domonkos et al. (2004). Thylakoid membranes were resuspended
in buffer C containing 20 mM MES, pH 6.5, 10 mM CaCl2, 10mM MgCl2, 0.5 M
mannitol, 20% (v/v) glycerol, 1 mM PMSF and 1 mM benzamidine to reach a
final Chl concentration of 1 mg ml–1. Following the addition of b-DM to 0.1%
(w/v), the samples were shaken on an orbital shaker for 10 min in darkness.
Then the membranes were pelleted by centrifugation for 30 min at 145,000�g
in a Beckman Ti 50 rotor. The phycobilisome-containing supernatant was dis-
carded, and the membrane pellet was resuspended in buffer C containing 1%
(w/v) b-DM at 1 mg ml–1 Chl concentration. The suspension was shaken for
30 min and recentrifuged at 145,000�g for 90 min. The supernatant, containing
the extracted pigment–protein complexes, was filtered using a stainless steel
filter and loaded onto a MonoQ HR 5/5 column (Amersham-Pharmacia
Biotech), equilibrated with buffer B. Samples were eluted with a non-linear
gradient of 5–200 mM MgSO4, with a flow rate of 0.4 ml min–1, as described
by Rogner et al. (1990). Absorbance was recorded at 680 nm. Fractions (1 ml)
were collected for further analyses. Fluorescence emission spectroscopy at 77K
and gel filtration chromatography on a calibrated TSK 3000 SW (7.5�600 mm)
column (Tosoh Bioscience) were used to identify PSI monomers and trimers in
fractions, as described in Domonkos et al. (2004).
Analysis of pigment composition
Pigment composition was analyzed essentially as described by Domonkos et al.
(2009). The Prostar HPLC system (Varian) equipped with a diode array spec-
trophotometric detector Tidas I (World Precision Instruments) and a Nucleosil
100 C18 reversed phase column, 5 mm particle size (Technokroma) were used.
Pigments were extracted in acetone : methanol (7 : 2, v/v) solvent, evaporated
under gaseous nitrogen and loaded onto a column equilibrated with Solvent A
(acetonitrile : water : triethylamine, 9 : 1 : 0.01, v/v/v.) in HPLC-grade ethanol
(Domonkos et al. 2009). Samples were eluted from the column with Solvent
B (ethyl acetate, 100%) by a three-step gradient (0–40% B for 10 min, 40–60% B
for 10 min, 60–100% B for 3 min, followed by a 2 min isocratic hold at 100% B),
with a constant flow rate of 1.5 ml min–1. The absorption spectra of the
eluate (380–800 nm) were recorded every 0.2 s. Carotenoids were identified
on the basis of absorption spectra and retention times. The relative content
of each pigment was estimated by a comparison of peak areas on chromato-
grams recorded at 440 nm. The concentrations of carotenoid species were
calculated from Beer–Lambert’s law using specific extinction coefficients at
440 nm (Mantoura and Llewellyn 1983) and 460 nm for echinenone (Britton
1995). Values are means ± SD calculated from at least three independent
experiments.
Fluorescence spectroscopy
For 77K measurements, the cell suspensions were cooled in liquid nitrogen in
capillary tubes. Steady-state fluorescence emission spectra (670–750 nm) were
recorded, using a 437 nm excitation wavelength, with a Perkin-Elmer LS 50
spectrofluorimeter equipped with a liquid nitrogen attachment. The emission
spectra were corrected for the wavelength dependence of the detection system
and normalized at 682 nm.
Circular dichroism
The CD spectra were recorded between 300 and 800 nm at room temperature
with a J815 (Jasco) dichrograph using a bandpass of 5 nm and a resolution of
1 nm at a 100 nm min–1 scanning rate with 4 s integration time. The Chl con-
tent of the samples was adjusted to 5–10mg ml–1 and was measured in a Quartz
SUPRASIL� cuvette (Hellma) with a 1 cm optical path length.
Absorption spectroscopy
The absorption spectra of cell suspensions were recorded at room temperature
with a UV-1601 (Shimadzu) spectrophotometer. The cell densities were
adjusted to OD730. The absorption spectra of cell suspensions were scanned
in the region from 400–800 nm and normalized at 665 nm according to
Lichtenthaler (1987).
Microscopy
Confocal microscope images were obtained with a Leica SP5 AOBS confocal
microscope using a �63 objective. Laser excitation light at 488 nm was applied
and detection was carried out in the range of 650–750 nm. Cell diameters
were calculated with Cell Profiler 2.0 software (Broad Institute) from at least
20 cells.
For electron microscopy, cells were fixed in 1% paraformaldehyde and 1%
glutaraldehyde for 4 h at 4�C and post-fixed in 1% osmium tetroxide. The
material was dehydrated in aqueous solutions of increasing ethanol concentra-
tions, and subsequently embedded in Spurr resin. Following polymerization,
85–90 nm ultrathin sections were cut by a Reichert Ultracut E ultramicrotome
(Leica,). The sections, treated with uranyl acetate and lead citrate according to
the standard procedure, were subjected to electron microscopy in a Zeiss EM
902 electron microscope (Carl Zeiss AG).
Determination of pigment concentration
The total Chl and carotenoid contents were measured after extraction in a 90%
(v/v) methanol extract from cells and/or previously lyophylized subcellular
fractions according to Lichtenthaler (1987).
Protein determination
Protein concentrations in whole cells and in isolated membranes were mea-
sured in a pellet after pigment extraction, by a micro Lowry method using
bovine serum albumin as standard, according to Yeang et al. (1998).
Statistics and reproducibility
All statistical analyses (SD and SE) were carried out using GraphPad InStat, and
treated statistically where possible with a t-test at significance level at of least
P< 0.05. Non-quantitative experiments performed multiple times using inde-
pendent biological samples, produced similar results.
Supplementary data
Supplementary data are available at PCP online.
Funding
This work was supported by Polish–Hungarian bilateral grant[OMFB-00584/2010 TeT]; the Polish Ministry of Science andHigher Education (2010–2011); grants TAMOP-4.2.2.A-11/1/KONV-2012-0047 and the Hungarian Science Foundation[OTKA K 82052 and K 108411] to Z.G.; OTKA grant K68692 to M.K.; the International Visegrad Fund [grant No.21120109]. K.K. was a participant of the InternationalTraining Course at BRC HAS (Szeged, Hungary) in academicyear 2008/2009.
Disclosures
The authors have no conflicts of interest to declare.
References
Allen, M.M. (1968) Simple conditions for growth of unicellular bluegreenalgae on plates. J. Phycol. 4: 1–4.
569
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

Aminaka, R., Taira, Y., Kashino, Y., Koike, H. and Satoh, K. (2006)Acclimation to the growth temperature and thermosensitivity of
photosystem II in a mesophilic cyanobacterium, Synechocystis sp.PCC6803. Plant Cell 47: 1612–1621.
Berry, J. and Bjorkman, O. (1980) Photosynthetic response and adaptationto temperature in higher plants. Annu. Rev. Plant. Physiol. 31: 491–543.
Bibby, T.S., Nield, J. and Barber, J. (2001) Iron deficiency induces the for-mation of an antenna ring around trimeric photosystem I in cyanobac-
teria. Nature 412: 743–745.Boekema, E.J., Dekker, J.P., van Heel, M.G., Rogner, M., Saenger, W., Witt, I.
et al. (1987) Evidence for a trimeric organization of the photosystem Icomplex from the thermophilic cyanobacterium Synechococcus sp.
FEBS Lett. 217: 283–286.Boekema, E.J., Hifney, A., Yakushevska, A.E., Piotrowski, M., Keegstra, W.,
Berry, S. et al. (2001) A giant chlorophyll–protein complex induced byiron deficiency in cyanobacteria. Nature 412: 745–748.
Britton, G. (1995) UV/visible spectroscopy. In Carotenoids: Spectroscopy.Edited by Britton, G., Liaaen-Jensen, S. and Pfander, H. pp. 13–63.
Birkhauser Verlag, Basel.Chitnis, P.R. (1996) Photosystem I. Plant Physiol. 111: 661–669.
Chitnis, P.R. (2001) Photosystem I: function and physiology. Annu. Rev.Plant Physiol. Plant Mol. Biol. 52: 593–626.
Chitnis, V.P. and Chitnis, P.R. (1993) PsaL subunit is required for the for-mation of photosystem I trimers in the cyanobacterium Synechocystis
sp. PCC6803. FEBS Lett. 336: 330–334.Chitnis, V.P., Xu, Q., Yu, L., Golbeck, J.H., Nakamoto, H., Xie, D.-L. et al.
(1993) Targeted inactivation of the gene PsaL encoding a subunit ofphotosystem I of the cyanobacterium Synechocystis sp. PCC6803. J. Biol.
Chem. 268: 11678–1184.Domonkos, I., Malec, P., Sallai, A., Kovacs, L., Itoh, K., Shen, G. et al. (2004)
Phosphatidylglycerol is essential for oligomerization of PSI reactioncenter. Plant Physiol. 134: 1471–1478.
Domonkos, I., Malec, P., Laczko-Dobos, H., Sozer, O., Kłodawska, K.,
Wada, H. et al. (2009) Phosphatidylglycerol depletion induces an in-crease in myxoxanthophyll biosynthetic activity in Synechocystis
PCC6803 cells. Plant Cell Physiol. 50: 1–9.El-Mohsnawy, E., Kopczak, M.J., Schlodder, E., Nowaczyk, M., Meyer, H.E.,
Warscheid, B. et al. (2010) Structure and function of intact photosystemI monomers from the cyanobacterium Thermosynechococcus elonga-
tus. Biochemistry 49: 4740–4751.Fromme, P., Jordan, P. and Krauss, N. (2001) Structure of photosystem I.
Biochim. Biophys. Acta 1507: 5–31.Frosch, D., Westphal, C. and Bohme, H. (1987) Improved preservation of
glycogen in unfixed cyanobacteria embedded at –82 degrees C in nano-plast. J. Histochem. Cytochem. 35: 119–121.
Fuhrmann, E., Gathmann, S., Rupprecht, E., Golecki, J. and Schneider, D.(2009) Thylakoid membrane reduction affects the photosystem stoichi-
ometry in the cyanobacterium Synechocystis sp. PCC6803. Plant Physiol.149: 735–744.
Fujita, Y., Murakami, A. and Ohki, K. (1987) Regulation of photosystemcomposition in cyanobacterial photosynthetic system: the regulation
occurs in response to the redox state of the electron pool locatedbetween two photosystems. Plant Cell Physiol. 28: 283–292.
Fujita, Y., Murakami, A., Ohki, K. and Hagiwara, N. (1988) Regulationof photosystem composition in cyanobacterial photosynthetic
system: evidence indicating that photosystem I formation is controlledin response to the electron transport state. Plant Cell Physiol. 29:
557–564.Fujita, Y., Ohki, K. and Murakami, A. (1985) Chromatic regulation of
photosystem composition in photosynthetic system of red and blue-green algae. Plant Cell Physiol. 26: 1541–1548.
Garab, G. and van Amerongen, H. (2009) Linear dichroism and circulardichroism in photosynthesis research. Photosynth. Res. 101: 135–146.
Gobets, B. and van Grondelle, R. (2001) Energy transfer and trapping inphotosystem I. Biochim. Biophys. Acta 1507: 80–99.
Golbeck, J.H. (1992) Structure and function of Photosystem I. Annu. Rev.Plant Physiol. Plant. Mol. Biol. 43: 293–324.
Gombos, Z., Varkonyi, Z., Hagio, M., Iwaki, M., Kovacs, L., Masamoto, K.et al. (2002) Phosphatidylglycerol requirement for the function of elec-
tron acceptor plastoquinone Q(B) in the photosystem II reactioncenter. Biochemistry 41: 3796–3802.
Hagio, M., Gombos, Z., Varkonyi, Z., Masamoto, K., Sato, N., Tsuzuki, M.et al. (2000) Direct evidence for requirement of phosphatidylglycerol in
photosystem II of photosynthesis. Plant Physiol. 124: 795–804.Hihara, Y., Sonoike, K. and Ikeuchi, M. (1998) A novel gene, pmgA, specif-
ically regulates photosystem stoichiometry in the cyanobacteriumSynechocystis species PCC6803 in response to high light. Plant Physiol.
117: 1205–1216.Hirschberg, J. and Chamovitz, D. (1994) Carotenoids in cyanobacteria. In
The Molecular Biology of Cyanobacteria. Edited by Bryant, D.A. pp.559–579. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Huner, N.P.A., Oquist, G. and Sarhan, F. (1998) Energy balance and accli-mation to light and cold. Trends Plant. Sci. 3: 224–230.
Inoue, N., Taira, Y., Yamane, Y., Kashino, Y., Koike, K. and Satoh, K. (2001)Acclimation to growth temperature and high-temperature effects on
photosystem II and plasma membranes in a mesophilic cyanobacter-ium Synechocystis sp. PCC6803. Plant Cell Physiol. 42: 1140–1148.
Ivanov, A.G., Krol, M., Sveshnikov, D., Selstam, E., Sandstrom, S.,Koochek, M. et al. (2006) Iron deficiency in cyanobacteria causes mono-
merization of photosystem I trimers and reduces the capacity for statetransitions and the effective absorption cross section of photosystem I
in vivo. Plant Physiol. 141: 1436–144.Karapetyan, N.V. (2008) Protective dissipation of excess absorbed energy
by photosynthetic apparatus of cyanobacteria: role of antenna terminalemitters. Photosynth. Res. 97: 195–204.
Karapetyan, N.V., Dorra, D., Schweitzer, G., Bezsmertnaya, I.N. andHolzwarth, A.R. (1997) Fluorescence spectroscopy of the longwave
chlorophylls in trimeric and monomeric photosystem I core complexes
from the cyanobacterium Spirulina platensis. Biochemistry 36:13830–13837.
Karapetyan, N.V., Holzwarth, A.R. and Rogner, M. (1999) The photosystemI trimer of cyanobacteria: molecular organization, excitation dynamics
and physiological significance. FEBS Lett. 460: 395–400.Kłodawska, K., Malec, P., Kis, M., Gombos, Z. and Strzałka, K. (2012) EPR
study of thylakoid membrane dynamics in mutants of the carotenoidbiosynthesis pathway of Synechocystis sp. PCC 6803. Acta Biochim. Pol.
59: 87–90.Komenda, J. and Barber, J. (1995) Comparison of psbO and psbH deletion
mutants of Synechocystis PCC6803 indicates that degradation of D1protein is regulated by the QB site and dependent on protein synthesis.
Biochemistry 34: 9625–9631.Komenda, J., Hladik, J. and Sofrova, D. (1989) On the multiple
forms of cyanobacterial PS 1 complex. J. Photochem. Photobiol. B 3:575–592.
Komenda, J., Knoppova, J., Kopecna, J., Sobotka, R., Halada, P., Yu, J. et al.(2012) The Psb27 assembly factor binds to the CP43 complex of photo-
system II in the cyanobacterium Synechocystis sp. PCC6803. PlantPhysiol. 158: 476–486.
Komenda, J., Tichy, M. and Eichacker, L.A. (2005) The PsbH protein isassociated with the inner antenna CP47 and facilitates D1 processing
and incorporation into Photosystem II in the cyanobacteriumSynechocystis PCC6803. Plant Cell Physiol. 46: 1477–1483.
Laczko-Dobos, H., Todinova, S.J., Sozer, O., Komenda, J., Kis, M., Sallai, A.et al. (2011) Identification of thylakoid membrane thermal transitions
in Synechocystis sp. PCC6803 photosynthetic mutants. Photosynth. Res.107: 237–246.
Lagarde, D., Beuf, L. and Vermaas, M. (2000) Increased production ofzeaxanthin and other pigments by application of genetic engineering
techniques to Synechocystis sp strain PCC6803. Appl. Environ. Microbiol.66: 64–72.
570
K. Kłodawska et al. | Photosystem-I-trimer formation in Synechocystis sp.
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from

Lambrev, P., Varkonyi, Z., Krumova, S., Kovacs, L., Miloslavina, Y.,Holzwarth, A.R. et al. (2007) Importance of trimer–trimer interactions
for the native state of the plant light-harvesting complex II. Biochim.Biophys. Acta 1767: 847–853.
Li, H., Li, D., Yang, S., Xie, J. and Zhao, J. (2006) The state transition mech-anism—simply depending on light-on and -off in Spirulina platensis.
Biochimi. Biophys. Acta 1757: 1512–1519.Li, H., Singh, A.K., McIntyre, L.M. and Sherman, L.A. (2004) Differential gene
expression in response to hydrogen peroxide and the putative PerRregulonof Synechocystis sp. strain PCC6803. J. Bacteriol. 186: 3331–3345.
Lichtenthaler, H.K. (1987) Chlorophylls and carotenoids: pigments ofphotosynthetic biomembranes. Methods Enzymol. 148: 350–382.
Mantoura, R.F.C. and Llewellyn, C.A. (1983) The rapid determination ofalgal chlorophyll and carotenoid pigments and their breakdown prod-
ucts in natural waters by reverse-phase high-performance liquid chro-matography. Anal. Chim. Acta 151: 297–314.
Miskiewicz, E., Ivanov, A.G., Williams, J.P., Khan, M.U., Falk, S. andHuner, N.P.A. (2000) Photosynthetic acclimation of the filamentous
cyanobacterium, Plectonema boryanum UTEX 485, to temperatureand light. Plant Cell Physiol. 41: 767–775.
Mohamed, H.E. and Vermaas, W. (2004) Slr1293 in Synechocystis sp. strainPCC6803 is the C-30 ,40desaturase (CrtD) involved in myxoxanthophyll
biosynthesis. J . Bacteriol. 186: 5621–5628.Murakami, A. (1997) Quantitative analysis of 77K fluorescence emission
spectra in Synechocystis PCC6714 and Chlamydomonas reinhardtii withvariable PSI/PSII stoichiometries Photosynthesis Res. 53 141–148.
Murakami, A. and Fujita, Y. (1993) Regulation of stoichiometry betweenPSI and PSII in response to light regime for photosynthesis observed
with Synechocystis PCC6714: relationship between redox state of Cytb6f complex and regulation of PSI formation. Plant Cell Physiol. 34:
1175–1180.Murakami, A., Kim, S.J. and Fujita, Y. (1997) Changes in photosystem
stoichiometry in response to environmental conditions for cell
growth observed with the cyanophyte Synechocystis PCC6714. PlantCell Physiol. 38: 392–397.
Muramatsu, M. and Hihara, Y. (2012) Acclimation to high-light conditionsin cyanobacteria: from gene expression to physiological responses. J.
Plant Res. 125: 11–39.Nishiyama, Y., Los, D.A., Hayashi, H. and Murata, N. (1997) Thermal pro-
tection of the oxygen-evolving machinery by PsbU, an extrinsic proteinof photosystem II, in Synechococcus sp. PCC7002. Plant Physiol. 115:
1473–1480.Nishiyama, Y., Los, D.A. and Murata, N. (1999) PsbU, a protein associated
with photosystem II, is required for the acquisition of cellular thermo-tolerance in Synechococcus sp. PCC7002. Plant Physiol. 120: 301–308.
Rogner, M., Nixon, P.J. and Diner, B.A. (1990) Purification and character-ization of photosystem I and photosystem II core complexes from wild-
type and phycocyanin-deficient strains of the cyanobacteriumSynechocystis PCC6803. J. Biol. Chem. 265: 6189–6196.
Sajilata, M.G., Singhal, R.S. and Kamat, M.Y. (2008) The carotenoid pigmentzeaxanthin—a review. Compr. Rev. Food Sci. Food Saf. 7: 29–49.
Salomon, E. and Keren, N. (2011) Manganese limitation induces changes inthe activity and in the organization of photosynthetic complexes in the
cyanobacterium Synechocystis sp. strain PCC6803. Plant Physiol. 155:571–579.
Schafer, L., Vioque, A. and Sandmann, G. (2005) Functional in situ evalu-ation of photosynthesis-protecting carotenoids in mutants of the
cyanobacterium Synechocystis PCC6803. J. Photochem. Photobiol. B 78:195–201.
Schluchter, W.M., Shen, G., Zhao, J. and Bryant, D.A. (1996)Characterization of psal and psaL mutants of Synechococcus sp. strain
PCC7002: a new model for state transitions in cyanobacteria. Photobiol.Photochem. 64: 53–66.
Shubin, V., Bezsmertnaya, I. and Karapetyan, N.V. (1995) Efficient energytransfer from the long-wavelength antenna chlorophylls to P700 in
photosystem I complexes from Spirulina platensis. J. Photochem.Photobiol. B 30: 153–160.
Shubin, V., Tsuprun, V.L., Bezsmertnaya, I.N. and Karapetyan, N.V. (1993)Trimeric forms of the photosystem I reaction center complex pre-exist
in the membranes of the cyanobacterium Spirulina platensis. FEBS Lett.334: 79–82.
Singh, A.K., McIntyre, L.M. and Sherman, L.A. (2003) Microarray analysis ofthe genome-wide response to iron deficiency and iron reconstitution in
thecyanobacterium Synechocystis sp. PCC6803. Plant Physiol. 132:1825–1839.
Singh, A.K. and Sherman, L.A. (2005) Pleiotropic effect of a histidine kinaseon carbohydrate metabolism in Synechocystis sp. strain PCC 6803 and
its requirement for heterotrophic growth. J. Bacteriol. 187: 2368–2376.Sozer, O., Kis, M., Gombos, Z. and Ughy, B. (2011) Proteins, glycerolipids
and carotenoids in the functional photosystem II architecture. Front.Biosci. 16: 619–643.
Sozer, O., Komenda, J., Ughy, B., Domonkos, I., Laczko-Dobos, H., Malec, P.et al. (2010) Involvement of carotenoids in the synthesis and assembly
of protein subunits of photosynthetic reaction centers of SynechocystisPCC6803. Plant Cell Physiol. 51: 823–835.
Stanier, R.Y., Kunisawa, R., Mandel, M. and Cohen-Bazire, G. (1971)Purification and properties of unicellular blue-green alga (order
Choorococcales). Bacteriol. Rev. 35: 171–205.Steiger, S., Schafer, L. and Sandmann, G. (1999) High-light-dependent upre-
gulation of carotenoids and their antioxidative properties in the cyano-bacterium Synechocystis PCC6803. J. Photochem. Photobiol. B 52: 14–18.
Takaichi, S. and Mochimaru, M. (2007) Carotenoids and carotenogenesisin cyanobacteria: unique ketocarotenoids and carotenoid glycosides.
Cell. Mol. Life Sci. 64: 2607–2619.Tandeau de Marsac, N. and Houmard, J. (1993) Adaptation of cyanobac-
teria to environmental stimuli: new steps towards molecular mechan-isms. FEMS Microbiol. Lett. 104: 119–189.
Trissl, H.W. (1993) Long-wavelength absorbing antenna pigments and
heterogeneous absorption bands concentrate excitons and increaseabsorption cross section. Photosynth. Res. 35: 247–263.
Turconi, S., Kruip, J., Schweitzer, G., Rogner, M. and Holzwarth, A.R. (1996)A comparative fluorescence kinetics study of photosystem I monomers
and trimers from Synechocystis PCC6803. Photosynth. Res. 49: 263–268.Van Grondelle, R., Dekker, J.P., Gilbro, T. and Sundstrom, V. (1994) Energy
transfer and trapping in photosynthesis. Biochim. Biophys. Acta 1187:1–65.
Wang, Q., Jantaro, S., Lu, B., Majeed, W., Bailey, M. and He, Q. (2008) Thehigh light-inducible polypeptides stabilize trimeric photosystem I com-
plex under high light conditions in Synechocystis PCC6803. PlantPhysiol. 147: 1239–1250.
Watanabe, M., Kubota, H., Wada, H., Narikawa, R. and Ikeuchi, M. (2011)Novel supercomplex organization of photosystem I in Anabaena and
Cyanophora paradoxa. Plant Cell Physiol. 52: 162–168.Yamauchi, Y., Kaniya, Y., Kaneko, Y. and Hihara, Y. (2011)
Physiological roles of the cyAbrB transcriptional regulator pair Sll0822and Sll0359 in Synechocystis sp. strain PCC6803. J. Bacteriol. 193:
3702–3709.Yeang, H.Y., Yusof, F. and Abdullah, L. (1998) Protein purification for the
Lowry assay: acid precipitation of proteins in the presence of sodiumdodecyl sulfate and other biological detergents. Anal. Biochem. 265:
381–384.Varkonyi, Z., Masamoto, K., Debreczeny, M., Zsiros, O., Ughy, B., Gombos, Z.
et al. (2002) Low-temperature-induced accumulation of xanthophyllsand its structural consequences in the photosynthetic membranes of
the cyanobacterium Cylindrospermopsis raciborskii: an FTIR spectro-scopic study. Proc. Natl Acad. Sci. USA 99: 2410–2415.
Zhu, Y., Graham, J.E., Ludwig, M., Xiong, W., Alvey, R.M., Shen, G. et al.(2010) Roles of xanthophyll carotenoids in protection against photo-
inhibition and oxidative stress in the cyanobacterium Synechococcus sp.strain PCC7002. Arch. Biochem. Biophys. 504: 86–99.
571
Plant Cell Physiol. 56(3): 558–571 (2015) doi:10.1093/pcp/pcu199
at Uniw
ersytet Jagiellonsky w K
rakowie on M
arch 23, 2015http://pcp.oxfordjournals.org/
Dow
nloaded from
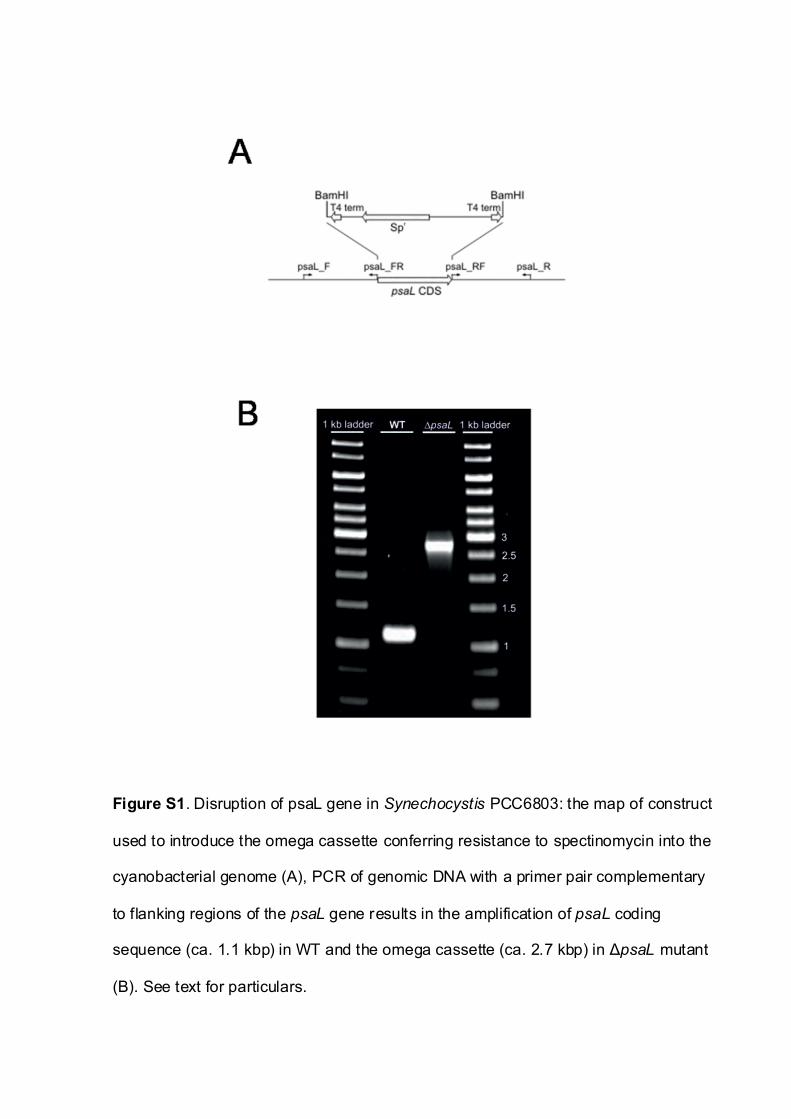
Figure S1. Disruption of psaL gene in Synechocystis PCC6803: the map of construct
used to introduce the omega cassette conferring resistance to spectinomycin into the
cyanobacterial genome (A), PCR of genomic DNA with a primer pair complementary
to flanking regions of the psaL gene results in the amplification of psaL coding
sequence (ca. 1.1 kbp) in WT and the omega cassette (ca. 2.7 kbp) in ΔpsaL mutant
(B). See text for particulars.
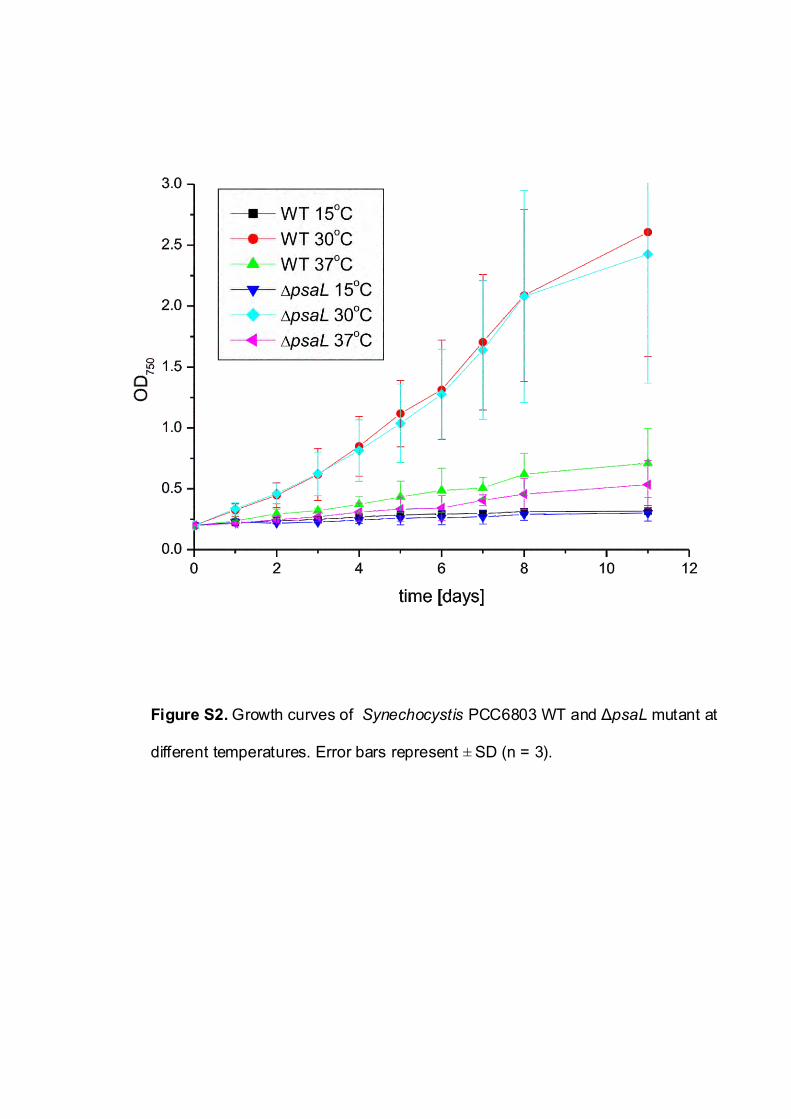
Figure S2. Growth curves of Synechocystis PCC6803 WT and ΔpsaL mutant at
different temperatures. Error bars represent ± SD (n = 3).
Related Documents











