Aus der Klinik und Poliklinik für Frauenheilkunde und Geburtshilfe Direktor Univ.- Prof. Dr. med. Marek Zygmunt der Medizinischen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald Einflüsse eines Folsäure und Vitamin-B6-Mangels auf die Morphologie der Plazenta und den diaplazentaren Aminosäuretransport der trächtigen Ratte Inaugural – Dissertation zur Erlangung des akademischen Grades Doktor der Medizin (Dr. med.) der Medizinischen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald 2010 vorgelegt von: Benjamin Johannes Desaga geb. am: 10. Oktober 1982 in Hamburg

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Aus der Klinik und Poliklinik für Frauenheilkunde und Geburtshilfe
Direktor Univ.- Prof. Dr. med. Marek Zygmunt
der Medizinischen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald
Einflüsse eines Folsäure und Vitamin-B6-Mangels auf die Morphologie der Plazenta
und den diaplazentaren Aminosäuretransport der trächtigen Ratte
Inaugural – Dissertation
zur
Erlangung des akademischen
Grades
Doktor der Medizin
(Dr. med.)
der
Medizinischen Fakultät
der
Ernst-Moritz-Arndt-Universität
Greifswald
2010
vorgelegt von:
Benjamin Johannes Desaga
geb. am: 10. Oktober 1982
in Hamburg

Dekan: Prof. Dr. med. Claus-Dieter Heidecke
1. Gutachter: Prof. Dr. med. M. Zygmunt
2. Gutachter: Prof. Dr. med. vet. C. Pfarrer
Ort, Raum: Klinik für Gynäkologie und Geburtshilfe, Ferdinand-Sauerbruch-Straße, Greifswald, N0.03
Tag der Disputation: Di. 30. August 2011
2

Inhaltsverzeichnis
Seite
1. Einleitung und Zielstellung 5
2. Literaturübersicht
2.1 Folsäure 7
2.2 Vitamin B6 13
2.3 Aufbau und Entwicklung der Plazenta der Ratte 18
2.4 Diaplazentare Transportmechanismen 28
2.5 Methylierungszyklus 31
2.6 Zusammensetzung der Amnionflüssigkeit 34
3. Material und Methoden
3.1 Material 35
3.2 Methoden 36
4. Ergebnisse
4.1 Vitamin B6, Folsäure, Homocystein, ALAT, ASAT im maternalen Plasma 45
4.2 Proteinogene Aminosäuren mit apolaren Seitenketten 49
4.3 Proteinogene Aminosäuren mit ungeladenen polaren Seitenketten 55
4.4 Proteinogene Aminosäuren mit sauren Seitenketten 60
4.5 Proteinogene Aminosäuren mit basischen Seitenketten 62
4.6 Nicht proteinogene Aminosäuren 65
4.7 Histomorphologische Untersuchung der Plazenten 71
4.8 Immunhistochemische Untersuchung der Plazenten 76
4.9 Resorptionen 80
5. Diskussion
5.1 Blut- und Amnionparameter 81
5.2 Histomorphologische und Immunhistochemische Untersuchungen der Plazenten 88
6. Literaturupdate und Ausblick 92
7. Zusammenfassung 94
8. Abkürzungsverzeichnis 97
9. Schrifttum 98
3

Seite
Eidesstattliche Erklärung 117
Danksagung 118
Anhang 119
4

1. Einleitung und Zielstellung
Lippen-Kiefer-Gaumen-Segel-Spalten (LKGS) und Neuralrohrdefekte (ND) gehören mit zu den
häufigsten Fehlbildungen des Menschen mit einer regional unterschiedlichen Inzidenz von 1:500
bis 1:1500 (Hillig, 1991; Källen, 1989; Kerrigan et al., 2000; Rösch et al., 1998; Tönz et al.,
1996). Das Ursachengefüge dieser Fehlbildungen ist sehr vielschichtig. Neben genetischen
Faktoren als auch Umwelteinflüssen, spielen Toxine, aber auch Mangelernährungen bzw.
Vitaminunterversorungen eine nicht unerhebliche Rolle. Bezüglich der Vitaminmangelernährung
kommt den wasserlöslichen B-Vitaminen Folsäure, Vitamin B6 und B12 eine entscheidende Rolle
zu. Sie haben insbesondere am Aminosäurestoffwechsel, an der Proteinsynthese als auch an der
Zellteilung entscheidenden Anteil.
Bisherige empirische Untersuchungen geben jedoch nur unvollständig Auskunft über den Bedarf
und die Wirkung dieser Vitamine in der perikonzeptionellen Phase als auch in der
Schwangerschaft selbst. Problematisch ist der Umstand, dass ND und LKGS in der 4. bzw. 7.-9.
Embryonalwoche entstehen, die Schwangerschaft selbst im Allgemeinen aber erst nach Eintritt
dieser Fehlbildungen diagnostiziert wird. Eine Prävention über die orale Gabe dieser Vitamine
kommt dann meist zu spät, ebenso die klinisch-chemische Diagnostik bezüglich der biologischen
Verfügbarkeit dieser Vitamine im Blut der Mutter. Neben den persönlichen Problemen für die
Betroffenen als auch für deren Angehörige sind die Kosten für das Gesundheitssystem zu
berücksichtigen. Somit verursachen z.B. die nach Koletzko und Kries (1995) veranschlagten
Kosten für das deutsche Gesundheitssystem zur Behandlung von Neuralrohrdefekten jährlich
Belastungen in Höhe von ca. 130 Mio. DM. Schon aus dieser Sicht hat die Gesellschaft ein großes
Interesse an der Prävention dieser Fehlbildungen. Ein Vitamin B6-Magel kann während der
Schwangerschaft u.a. zu Entwicklungsstörungen des Gehirns mit retardierter physischer
Entwicklung führen (Gerster, 1996). Durch einen Mangel an Folsäure oder Vitamin B6 steigt der
Homocysteinspiegel im Blutplasma deutlich an. Aus den medizinischen Fachbereichen der
Kardiologie, Neurologie und Gynäkologie ist bekannt, dass ein erhöhter Homocysteinspiegel zu
strukturellen Veränderungen an Gefäßendothelien und damit zu Herzinfarkten, Apoplex, Aborten
und Fehlbildungen führen kann (Blacher et al., 1996; Ambrosi et al., 1996; Montalescot et al.,
1997; Eskes, 1997, 2001; Goddijn-Wessel et al., 1996).
Nach bisherigen internationalen Untersuchungen aus Ungarn, den USA, Norwegen u.a. Ländern
konnte z.B. bei perikonzeptioneller Folsäuresubstitution eine Senkung von Fehlbildungen
insbesondere bei den ND erreicht werden (Czeizel et al., 1994, 1998, 2004). Zur Reduktion von
LKGS bei Folsäuresubstitution finden sich in der Literatur erstaunlicherweise widersprüchliche
5

Ergebnisse, obwohl ein Vitaminmangel allgemein als Risikofaktor angesehen wird (Wilcox, 2007;
Hartridge et al., 1999; Hayes 1996; Czeizel et al., 2004; Czeizel, 2004 ).
Es wird vermutet, dass insbesondere aufgrund der Erfahrungen aus der Kardiologie und
Neurologie der durch Vitamin B-Mangel und dem mit ihm assoziierte erhöhte Homocysteinspiegel
sich auf die Struktur und die Transportmechanismen der Plazenta und damit auf die
Lebensfähigkeit der Feten auswirkt oder Aborte bzw. Entwicklung von Fehlbildungen auslöst.
Daher gilt es in der vorliegenden Arbeit an dem Tiermodell Ratte mit einer Plazenta hämochorialis
zu prüfen, ob und inwieweit ein mit dem Beginn der Trächtigkeit induzierter Folsäure- bzw.
Vitamin B6-Mangel mittels Vitaminrestriktion Einfluß auf die Strukturen und Reifung der
Plazenta haben. Neben den morphologischen Strukturen der Plazenta sollen auch Blutwerte der
Mütter und Amnionflüssigkeitswerte der Feten bezüglich des diaplazentaren
Aminosäuretransportes als auch des Methylierungszyklus untersucht werden. Somit sollen
morphologische Strukturen bzw. Veränderungen durch physiologische Parameter untermauert
werden und einen Beitrag leisten, die Entwicklung, Reifung und Struktur der Plazenta besser zu
verstehen. Nicht zuletzt werden hiermit allgemeine Aussagen zur Rolle der genannten B-Vitamine
bezüglich der Prävention und Induktion von Fehlbildungen erwartet.
6

2. Literaturübersicht
2.1 Folsäure
Folsäure besteht aus einem Pteridinkern, p-Aminobenzoesäure und L-Glutamat. Natürlich
vorkommendes Folat unterscheidet sich in der Anzahl der Glutamylreste, die mit dem Pteridin-p-
aminobenzoesäurekomplex verbunden sind (Löffler und Petrides, 1998). Ihr chemischer Name ist
Pteroylglutamat (Butterworth und Bendich, 1996). Sie gehört zur Gruppe der wasserlöslichen
Vitamine (Trivialname Vitamin B11). Die künstlich hergestellte Folsäure ist im Gegensatz zum
natürlich vorkommenden Folat wesentlich hitzebeständiger und erreicht eine höhere biologische
Verfügbarkeit.
Abb. 2.1.1: Folsäure, künstlich hergestellt als Beispiel für ein Monoglutamat. chemischer Name:Pteroyl-L-monoglutaminsäure
Abb. 2.1.2: Folat als Hexaglutamat, Beispiel für natürlich Vorkommendes Polyglutamat. chemischer Name:L-Glutaminsäure, N-(N-(N-(N-(N-(N-(4-(((2-amino-1,4-dihydro-4-oxo-6-pteridinyl)methyl)amino)benzoyl)-L-gamma-glutamyl)-L-gamma-glutamyl)-L-gamma-glutamyl)-L-gamma-glutamyl)-L-gamma-glutamyl)
Folat ist reichlich in Leber, Hefe, Eiern, Bohnen, Apfelsinen und grünem Blattgemüse enthalten.
Häufig enthalten folatreiche Lebensmitteln auch Vitamin C, welches als Antioxidans wirkt und
einen schnellen Folatabbau verhindert (Butterworth und Bendich, 1996).
Folsäure wird vor ihrer aktiven Resorption durch die Mukosazellen des
Duodenums oder des oberen Jejunums hydrolysiert. In den Mukosazellen erfolgt anschließend die
7

Reduktion zu 5,6,7,8-Tetrahydrofolsäure und eine Methylierung zu 5-Methyltetrahydrofolsäure,
welche in die Blutbahn überführt wird (Buddecke, 1994). Nach Bässler (1997) finden sich im
Portalvenenblut reichlich noch nicht methylierte Tetrahydrofolate, so das die Methylierung dieser
erst in der Leber erfolgt. Zudem wird die Leber auch als Hauptspeicherorgan angesehen. Der
Transport im Serum erfolgt überwiegend durch Bindung an Albumin, α-Makroglobuline und
Transferrin. Waxman und Schreiber (1973) konnten ein Folsäure bindendes Protein (FABP)
nachweisen, dem sie eine wichtige Funktion bei der Folatspeicherung und Aufnahme in die Zellen
zuschrieben.
Die hohe biologische Verfügbarkeit von Folsäure im Gegensatz zu Folat ist durch die
Glutamatreste bedingt. Während Folsäure als ein Monoglutamat fast vollständig (>90%) resorbiert
wird, liegt die Verfügbarkeit von Folat als ein Polyglutamat (5-7 Glutamareste) bei lediglich 50
Prozent (DGE, ÖGE, SGE, SVE, 2000).
Nach Butterworth et al. (1969) ist zu beachten, dass etwa die Hälfe einer oral zugeführten Menge
Folsäure von 4,41mg innerhalb von 24 Stunden renal ausgeschieden wird. Im direkten Vergleich
zeigte sich, dass Mono- und Triglutamate zu 50-60% im Urin ausgeschieden wurden,
Heptaglutamate jedoch lediglich zu 8,7% oder weniger.
Aus diesem Grund wurde von der D.A.CH. Liga Homocystein der Begriff des Folatäquivalents
eingeführt (Stanger et al., 2003). 1mg Folatäquivalent entsprechen 1mg Nahrungsfolat und 0,5mg
Folsäure. Der Bedarf eines Erwachsenen liegt bei 400µg/d Folatäquivalent (DGE, ÖGE, SGE,
SVE, 2000).
Nach Beitz et al. (2002) erreichen 90% aller in Deutschland lebenden Männer, und fast 100% aller
Frauen durch ihre tägliche Kost, ohne Supplimentierung durch Vitamin-Präparate nicht die
empfohlene Tagesdosis Folsäure von 400µg/d.
Zur Diagnose einer Folsäurehypovitaminose, ist die Bestimmung der Folsäurekonzentration
sowohl im Blutplasma als auch im Vollblut notwenig. Dabei ist die Folsäurekonzentration des
Plasmas aussagekräftig hinsichtlich der kurzfristigen Folsäureaufnahme. Ein langfristigeres
Vitamindefizit schlägt sich schließlich auch in einer niedrigeren Konzentration im
Erythrozytenkonzentrat nieder. Hier sinken die Folsäurewerte erst 16-18 Wochen nach Beginn
einer folsäurearmen Kost. Nach Krumdieck et al. (1978) besitzt die Folsäure beim Menschen eine
Halbwertszeit von etwa 100 Tagen. Er beobachtete jedoch auch eine zweite Halbwertszeit von
etwa 31,5 Stunden für neu aufgenommene Folsäure. Es wird vermutet, dass ein nicht
unbeträchtlicher Teil der Folsäure während der Erythropoese in den Zellen gebunden wird und die
8

Folsäurekonzentration in den Erythrozyten während ihrer Lebensdauer von etwa 120 Tagen
konstant bleibt (Butterworth und Bendich, 1996).
Hierdurch erklärt sich die lange Halbwertszeit in den Erythrozyten, im Gegensatz zu der kurzen
Halbwertszeit im Plasma.
Die Resorption von Folsäure im proximalen Tubulus der Niere spielt eine wesentliche Rolle bei
der Folsäurehomöostase (Sikka und McMartin, 1998). Durch die Resorption werden nur wenige
Mikrogramm folatwirksamer Verbindungen renal ausgeschieden (Stahl und Heseker, 2007). Nach
Krumdieck et al. (1978) spielt eine enterale Ausscheidung von Folsäure bzw. ihren Derivaten in
den Faeces eine entscheidende Rolle bei der Eliminierung von Folsäure. Da Folsäure jedoch auch
von Mikroorganismen in der Darmflora gebildet wird, ist in der Praxis schwer zwischen enteral
ausgeschiedener und in der Darmflora gebildeter Folsäure zu unterscheiden (Institute of Medicine,
1998). Die im Stuhl ausgeschiedene Menge an Folat beträgt insgesamt etwa 200 µg /d (Stahl und
Heseker, 2007). Bekannt ist, dass täglich etwa 10 bis 90µg Folsäure im enterohepatischen
Kreislauf rückresorbiert und bei normaler Folataufnahme etwa 1-12 mg in Form verschiedener
folatwirksamer Verbindungen renal ausgeschieden werden (Koletzko und Pietrzik, 2004).
Der intrazelluläre Wirkmechanismus der Tetrahydrofolsäure besteht in der Übertragung von C1-
Resten, wie z.B. Methyl-, Formyl-, Formiat- und Hydroxymethylresten, wobei die C1-Reste durch
entsprechende Dehydrogenase- bzw. Isomerasereaktionen ineinander überführt werden können.
C1-Donatoren stellen vor allem die Aminosäuren Serin, S-Adenosylmethionin, Thymin sowie der
Alkohol Cholin dar, wobei die Tetrahydrofolsäure zu N5,10-Methylentetrahydrofolsäure
methyliert wird. Die C1 Reste liefern dann z.B. den Kohlenstoff für die Positionen 2 und 8 der
Purinringe, stellen die Methylgruppen von Thymin und Hydroxymethylcytosin dar, bilden den ß-
Kohlenstoff des Serins bei der Umwandlung von Glycin in Serin und stellen die Methylgruppe bei
der Methylierung von Homocystein zu Methionin (Löffler und Petrides, 1998).
Aufgrund der essentiellen Rolle von Folsäure bei der Synthese von Purinen, als auch ihre
Funktion bei der Methylierung von Desoxyuridinmonophosphat (dUMP) zu
Desoxythyminmonophosphat (dTMP) im Rahmen der Thyminsynthese, kommt ihr eine große
Bedeutung bei der DNS-Replikation und somit bei der Zellteilung zu. Das bedeutet, dass sich ein
Folsäuremangel zunächst in Geweben mit hoher Zellteilungsrate manifestiert. Neben den
Entwicklungsstörungen von Feten, ist daher die megaloblastische Anämie eine typische
Erkrankung bei anhaltendem Folsäuremangel.
9

Durch die Remethylierung des Homocysteins zu Methionin wird der Homocysteinspiegel gesenkt.
In diesem Zusammenhang spielt auch das Vitamin B12 eine essentielle Rolle, da es in Form des
Methylcobalamin als Cofaktor zur Remethylierung benötigt wird (Löffler und Petrides, 1998).
Nach Clarke et al. (1998) führte eine tägliche Zufuhr von mindestens 0,5 mg Folsäure zu einer
Reduktion des Homocysteinspiegels um durchschnittlich 25%.
Durch zusätzliche Gabe von 0,5 mg Vitamin B12 konnte das Homocystein um durchschnittlich
weitere 7% gesenkt werden. Somit kann der Homocysteinspiegel im Blut u.a. als Indikator für die
Folsäureversorgung herangezogen werden.
Die Hyperhomocysteinämie wurde in den letzten Jahren für eine Reihe von Erkrankungen und
embryonalen Entwicklungsstörungen verantwortlich gemacht.
Nach Boushey et al. (1995) führt eine Erhöhung des Homocysteinspiegels um 5mmol/l bei
Männern zu einer Risikosteigerung für kardiovaskuläre Erkrankungen um 60%, bei Frauen um
80%. Die American Heart Association empfiehlt die Behandlung der Hyperhomocysteinämie ab
mehr als 10 µmol/l mit Folsäure und Vitamin B6 (Deris, 2001).
Die HOPE-2 Studie (Lonn et al., 2006) zeigte, dass die Behandlung mit Folsäure, Vitamin B6 und
B12 über durchschnittlich 5 Jahre bei Patienten mit Diabetes mellitus Typ II oder vaskulären
Erkrankungen, den Homocysteinspiegel um 2,4 µmol/l senken konnte, während er in der
Kontrollgruppe um 0,8 µmol/l stieg. Eine Reduktion tödlicher Herzinfarkte oder Schlaganfälle
durch die Behandlung war jedoch nicht vorhanden.
Zu ähnlichen Ergebnissen kam die NORVIT Studie, (Bønaa et al., 2006) in der der
Homocysteinspiegel bei Myokardinfarktpatienten mittels Vitamin B12- und Folsäureapplikation
um 27% gesenkt werden konnte (0,8 mg Folsäure /d, 0,4 mg Vitamin B12 /d), ein Einfluss auf die
Häufigkeit erneuter kardiovaskulärer Ereignisse konnte jedoch auch hier nicht nachgewiesen
werden.
Auch bei Patienten die lediglich mit Vitamin B6 (40 mg/d) behandelt wurden, konnte kein
signifikanter Zusammenhang zwischen der Behandlung und einer Reduktion erneuter
kardiovaskulärer Ereignisse festgestellt werden. Bei Myokardinfarkt-Patienten, welche mit
Vitamin B6, B12 und Folsäure behandelt wurden (0,8 mg Folsäure /d, 0,4 mg Vitamin B12 /d, 40
mg Vitamin B6 /d), wurde sogar eine Tendenz zu einem erhöhten Risiko einen Reinfarkt zu
erleiden festgestellt.
Die Homocysteinurie Typ I ist durch eine Mutation des Gens für die Cystathion β-Synthase (CBS)
(GA919, TC833) gekennzeichnet (Dilley et al., 2001). Es handelt nach Malinow et al. (1999)
um eine autosomal rezessive Erbkrankheit mit einer Inzidenz von etwa 1: 200.000.
10

Das Krankheitsbild der Homocysteinurie Typ II ist durch eine leichte Hyperhomocysteinämie
gekennzeichnet, der eine Mutation des Enzyms Methylentetrahydrofolatreduktase (MTHFR)
(CT677) zugrunde liegt.
Während bei der Homocysteinurie Typ I ein Zusammenhang zwischen Hyperhomocysteinämie,
Myokardinfarkten, Schlaganfällen und Lungenembolien schon im frühen Alter gesehen werden
kann (Malinow et al., 1999), ist dies bei der Homocysteinurie Typ II nicht der Fall (Dilley et al.,
2001).
Ein kompetitiver Hemmstoff der Dihydrofolatreduktase ist u.a. der Folsäureantagonist
Methotrexat (Lantarel®). Nach Transport in die Zelle durch Folsäure-Transporter wird Methotrexat
durch die Polylglutamase zu Polyglutamaten glutaminoyliert. Der Wirkmechanismus besteht in
einer höheren Affinität zur Dihydrofolatreduktase als Folsäure sie besitzen. Hierdurch wird die
Übertragung von C1-Resten u.a. auf Nukleinsäurebausteine sowie die Purinsynthese gehemmt.
Methotrexat wird als Zytostatikum eingesetzt und wirkt teratogen (Mutschler et al., 2006). Quinn
et al. (2004) konnten im Liquor cerebrospinalis bei intrathekal appliziertem Methotrexat einen
verringerten Folsäure- und S-Adenosylmethioninspiegel, begleitet von gestiegenen Homocystein-
und Adenosinwerten feststellen. Vezmar et al. (2003) vermuten in diesem Zusammenhang, dass
eine Erhöhung von Homocystein, S-Adenosylmethionin bzw. S-Adenosylhomocystein in der
Entwicklung subakuter und chronischer Neurotoxizität eine entscheidende Rolle spielen.
Der Folsäurehypovitaminose bzw. dem daraus resultierenden erhöhten Homocysteinspiegel von
Schwangeren werden Entwicklungsstörungen in der Embryogenese wie Neuralrohrdefekte,
Lippen-Kiefer-Gaumen-Segelspalten, Herz-, Extremitäten- und Harnwegsfehlbildungen sowie
Pylorusstenosen und Plazentaablösungen zugeschrieben (Koletzko und Pietrzik, 2004; Hall und
Solehdin, 1998).
Smithells et al. (1976) und Hall (1972) kamen zu dem Ergebnis, dass es keinen Zusammenhang
zwischen der Häufigkeit von Neuralrohrdefekten und dem Serumfolatspiegel gibt. Jedoch konnten
Smithells et al. (1976) einen Zusammenhang zwischen Neuralrohrdefekten, einem erniedrigen
Vitamin C Spiegel im Serum bei gleichzeitig verringerter Folsäurekonzentration in den
Erythrozyten feststellen.
Weiterhin wiesen Essien und Wannberg (1993) an Axd mutierten Mäusen nach, dass nicht eine
Folsäure- oder Vitamin B12-Hypovitaminose, sondern ein erhöhter Homocysteinspiegel für die
Entwicklung von Neuralrohrdefekten verantwortlich ist. Zhao et al. (2001) wiesen den
Zusammenhang zwischen Homocystein und Fehlbildungen an Ratten nach. Li et al. (1998)
konnten denselben Zusammenhang an Hühnerembryonen nachweisen.
11

Aubard et al. (2000) vermuten die Hyperhomocysteinämie als Ursache für maternale Thrombosen,
Präeklampsie bzw. Eklampsie und Plazentaablösungen, während Eskes (2000; 2001) neben
Plazentaablösungen darin auch die Ursache für Aborte und Neuralrohrdefekte sieht.
Unabhängig davon, ob der Folsäuremangel oder die Hyperhomocysteinämie für die Bildung von
Neuralrohrdefekten verantwortlich ist, konnte bei schwangeren Patientinnen, welche bereits ein
Kind mit Neuralrohrdefekt gebaren bei einer täglichen perikonzeptionellen
Folsäuresupplimentierung von 4 mg eine Reduktion dieser Fehlbildungen um 72% erreicht
werden (MRC Vitamin Study Research Group, 1991).
Aus diesem Grund wird Schwangeren eine perikonzeptionelle Substitution von Folsäure, Vitamin
B6 und B12 empfohlen. Hierbei erstreckt sich der perikonzeptionelle Zeitraum vornehmlich über
den Zeitraum von 2-3 Monaten vor der Konzeption bis mindestens dem Ende des ersten
Trimenon. Die DGE (2000) und Suter (2002) geben den täglichen Bedarf von Schwangeren mit
600 µg Folsäure, 1,9 mg Vitamin B6 und 3,5 µg Vitamin B12 an. Der Umkehrschluss hieraus, also
ein Vitaminprofil der schwangeren Frau zur Beurteilung des Risikos Kinder mit Neuralrohrdefekt
zu gebären ist jedoch nicht zulässig (Eskes, 1998).
Nach da Costa und Rothenberg (1996) besitzt die Rattenplazenta nahezu keine ungesättigten
Folsäurerezeptoren. Das folsäurebindende Protein in der Rattenplazenta besitzt seine höchste
Affinität für Folsäure, gefolgt von N5-Methyltetrahydrofolsäure und mit großem Abstand zu N5-
Formyltetrahydrofolsäure. Nach Henderson et al. (1995) findet der Folsäuretransport in der
menschlichen Plazenta bidirektional statt und ist nicht sättigbar. Die high-affinity membrane-
associated placental folate receptors (PFRs) besitzen eine hohe Affinität für 5-
Methylentetrahydrofolat. Der Transport kann in zwei Schritte eingeteilt werden, zum einen der
aktive Transport aus dem maternalen Blut in die Trophoblastenzelle, gegen einen
Konzentrationsgradienten der schnell geschieht und anschließend einen passiven langsameren
Transport aus der Zelle, dem Konzentrationsgradienten folgend in das fetale Blut. Das intervillöse
Blut der menschlichen Plazenta hat eine etwa dreifach höhere Konzentration an Folaten als das
maternale Blut.
Nach Baker et al. (1981) ist der Transfer von Vitamin B6, B12 und Folat zum Fetus und in die
Plazenta bei Schwangeren Frauen mit entsprechenden Hypovitaminosen verringert.
Piedrahita et al. (1999) wiesen nach, dass das Transportprotein Folbp1 der Maus eine
entscheidende Rolle in der Folathomöostase während der Entwicklung des Fetus spielt. Die Feten
entsprechender knockout-Mäuse entwickelten schwerwiegende Fehlbildungen und starben in
12

utero. Das Protein entspricht dem menschlichen FOLR1, bei dessen Defekt Fehlbildungen
vermutet werden.
Die fetale und plazentare Folsäureaufnahme ist nach Will et al. (1985) bei erhöhter maternaler
Aufnahme von Hydantoin deutlich reduziert. Es wird daher vermutet, dass die Fehlbildungen beim
„fetalen Hydantoin Syndrom“ auf einen fetalen Folsäuremangel zurückzuführen sind.
Nach Fisher et al. (1985) ist die Aktivität von plazentaren Folsäurerezeptoren bei chronischer
Ethanolexposition bei Ratten signifikant verringert. Die Plazenten sind größer, die Feten kleiner
als in der Kontrollgruppe.
2.2 Vitamin B6
Unter Vitamin B6 oder Pyridoxin werden die Wirkstoffe Pyridoxol (Alkohol), Pyridoxamin
(Amin) und Pyridoxal (Aldehyd) zusammengefasst, wobei alle Substanzen im Stoffwechsel
ineinander überführt werden können (Bässler, 1997). Die aktive Form des Vitamins ist das
Pyridoxal-5-phosphat, das durch eine ATP abhängige Phosphorylierung unter Mitwirkung der
Pyridoxalkinase gebildet wird (Löffler und Petrides, 1998).
Abb. 2.2.1: Strukturformal von Pyridoxin (I; R= -CH2OH), Pyridoxal (I; R=-CHO), Pyridoxamin (I; R=-CH2NH2), 4-Pyridoxinsäure (III), 4-Pyridoxolacton (IV), Pyridoxin 5-Phosphat (II; R= -CH2OH), Pyridoxal 5-Phosphat (II; R=-CHO) und Pyridoxamin 5-Phosphat (II; E=-CH2NH2). Nach Kälin Aebi, 2006.
13

Das Vitamin B6 gehört zur Gruppe der wasserlöslichen Vitamine und kommt sowohl in Fleisch,
vor allem in Leber, als auch in pflanzlicher Nahrung, wie z.B. Kartoffeln, Getreide,
Hülsenfrüchten und Gemüse reichlich vor. Auch Milchprodukte und einige Fischarten enthalten
Vitamin B6. Nach Reynolds (1988) ist die Bioverfügbarkeit von Vitamin B6 in Fleischprodukten
sehr hoch und kann 100% erreichen, während die Bioverfügbarkeit aus Pflanzenprodukten
aufgrund der hier häufig zu findenden Pyridoxin-Glykoside und der teilweise schwer spaltbaren
Polysaccharide der Zellwand zum Teil lediglich zwischen 20 und 25% liegt.
Die Resorption erfolgt vor allem im oberen Jejunum und in geringerem Umfang auch im Ileum.
Die o.g. Phosphorylierung von Pyridoxin zu Pyridoxal-5-phosphat findet vor allem in den
resorbierenden Mukosazellen statt. Im Plasma wird es überwiegend an Albumin gebunden
transportiert. Da es die Zellmembranen jedoch nicht passieren kann, muß es vor der Zellaufnahme
durch die hydrolisierende Eigenschaft der alkalische Phosphatase permeabel gemacht werden.
Die Erythrozytenkonzentration von Vitamin B6 ist 4-5 mal höher, als die im Plasma.
Die biologische Halbwertszeit von Vitamin B6 beträgt 10 Tage (Bässler, 1997).
Pyridoxin ist ein wichtiges Coenzym im Aminosäurestoffwechsel und katalysiert
Transaminierungen, Dexarboxylierungen, Eleminierungen und Aldolspaltungen (Löffler und
Petrides, 1998).
Des Weiteren spielt es eine wichtige Rolle im Rahmen der Glukoneogenese, der Niacin-
biosynthese aus Tryptophan, dem Fettstoffwechsel (vor allem im Sphingolipidstoffwechsel), der
Neurotransmittersynthese von Serotonin, Dopamin, Noradrenalin und γ-Aminobuttersäure, sowie
der Taurin-, Myelin- und Hämsynthese.
Weiterhin wird Vitamin B6 im Immunsystem und bei der Modulation von Hormoneffekten
benötigt. Vitamin B6 kann an Steroidhormonrezeptoren gebunden werden und somit die
Genexpressionen beeinflussen. Es ist Kofaktor von insgesamt über 100 Enzymen (Suter, 2002).
Der tägliche Bedarf des Menschen beträgt 1,5 bis 2 mg pro 100g aufgenommenem Protein
(Löffler und Petrides, 1998). Nach Suter (2002) wird Männern im Alter von 19 bis 65 Jahren eine
Zufuhr von 1,5 mg/d, Frauen von 1,2 mg/d und Schwangeren ab dem 4. Schwangerschafts Monat
sowie Stillenden 1,9mg/d Vitamin B6 empfohlen.
Nach Succari et al. (1987) besteht ein direkter Zusammenhang zwischen den Aktivitäten der
Alanin-Aminotransferase (ALAT), der Aspartat-Aminotransferase (ASAT) im Serum und einer
Pyridoxal-5-phosphat Substitution. Dies lässt sich dadurch begründen, dass die Apoenzyme
ALAT und ASAT mit ihrem Coenzym Vitamin B6 noch nicht gesättigt sind.
14

Die ALAT spielt eine wichtige Rolle im Stickstoffstoffwechsel, da sie Aminogruppen und
Ammoniak im Alanin fixiert und diese in die Leber transportiert werden, wo der Stickstoff z.B. als
Harnstoff gebunden und anschließend renal ausgeschieden wird.
Die ASAT wird zur Bereitstellung von Aminogruppen z.B. bei Synthesen von Purinen,
Pyrimidinen und Aminozuckern benötigt.
Auch spielt Vitamin B6 bei der Hyperhomocysteinämie und ihren unter 2.1 genannten Folgen eine
Rolle, da es als Coenzym beim Abbau von Homocystein zu Cystein beteiligt ist. Die SH- Gruppe
des Homocysteins wird hierbei auf Serin übertragen. Das entstehende Homoserin kann weiter zu
Proprionyl-CoA abgebaut werden. Dieses dient u.a. als Energielieferant im Zitratzyklus (Löffler
und Petrides, 1998).
Nach Gerster (1996) spielt Pyridoxin eine entscheidende Rolle bei der Biosynthese der
Neurotransmitter γ-Aminobuttersäure und Dopamin. Bei neugeborenen Ratten, deren Mütter eine
schwere Vitamin B6-Hypovitaminose aufwiesen, wurden erniedrigte Konzentrationen dieser
Neurotransmitter, weniger verzweigte Dendriten und eine verringerte Zahl myelinisierter Axone
und Synapsen nachgewiesen. Die Tiere zeigten ein auffälliges Verhalten, wie z.B. epilepsieartige
Krampfanfälle, in der Regel überlebten die Jungen nicht. Hier kommt der bei Ratten bekannte
selektive Kannibalismus zum Tragen, d.h. das Muttertier frisst fehlgebildete Neugeborene auf, um
den Genpool „sauber“ zu halten.
Beim Menschen konnte ein geringeres Geburtsgewicht, sowie ein retradiertes reaktives und
unreifes adaptives Verhalten beobachtet werden.
Alton-Mackey und Walker (1973) konnten bei Ratten mit einer Vitamin B6-Hypovitaminose
während der Trächtigkeit ebenfalls ein geringeres Geburtsgewicht, eine langsamere physische
Entwicklung sowie Beeinträchtigungen der neuromotorischen Entwicklung der Jungen feststellen.
Guilarte (1993) zufolge, kommt es bei einem Vitamin B6-Defizit während der Schwangerschaft
bzw. der Stillzeit außerdem zu Veränderungen in der Funktion von N-methyl-D-Aspartat-
rezeptoren, einem Subtyp von glutamatergen Neurotransmitterrezeptoren, welche eine wichtige
Rolle beim Lernen und Erinnern spielen.
Marathe und Thomas (1987) konnten zeigen, dass es bei der Behandlung von trächtigen Ratten
mit Pyridoxinhydrochlorid zu mehr Implantationen, Corpora lutea und lebend geborenen Jungen
kam. Bei hohen Dosen wurde jedoch ein geringeres Geburtsgewicht der Jungen beobachtet.
Vergleiche zwischen den Vitamingehalten von Uterus, Plazenta und fetalem Gewebe nach
intraperitonealer Injektion von Vitamin B6 bei trächtigen Tieren zeigten zunächst einen
Konzentrationsanstieg im Uterus, anschließend einen Vitamin B6-Abfall, begleitet von einem
15

Anstieg in der Plazenta, sowie ein Anstieg der Konzentration im Feten. Interessant ist, dass das
Vitamin B6 drei und zehn Minuten nach Injektion im Uterus und der Plazenta, vorwiegend als
Pyridoxol und Pyridoxol-5-phosphat nachgewiesen wurde, 30 Minuten nach Injektion jedoch in
Form von Pyridoxal-5-phosphat. Es kann also davon ausgegangen werden, dass dies die Form ist,
welche zum Fetus transportiert wird. (unbekannter Autor, 1972).
Nach Contractor und Shane (1971) wird Pyridoxalphosphat sowohl beim Menschen als auch beim
Nagetier (Ratte) aktiv zum Feten transportiert. Schenker et al. (1992) vermuten jedoch, dass es
sich um einen passiven Transport handelt.
Lyon et al. (1962) konnten bei Mäusen mit induzierter Vitamin B6-Hypovitaminose einen
verminderten Pyridoxal-5-phosphatspiegel im Hirngewebe nachweisen, die Konzentration von
Pyridoxamin-5-phosphat war unverändert. In Mäusen kommen überwiegend die Formen
Pyridoxal-5-phosphat sowie Pyridoxamin-5-phosphat vor.
Untersuchungen von Cleary et al. (1975) zufolge, sollten Schwangere mit mindestens 2 mg
Vitamin B6 pro Tag supplimentiert werden, um einen natürlichen Pyridoxinspiegel aufrecht zu
erhalten. Ob dies auch für den Fetus gilt, kann lediglich vermutet werden. Baker et al. (1981)
konnten zeigen, dass es bei Vitamin B6-, B12- und Folsäurehypovitaminosen zu einem
verringerten Transfer dieser Vitamine zum Feten kommt. Bei oraler Gabe dieser Vitamine konnte
ein fetaler Konzentrationsanstieg lediglich für Folsäure, jedoch nicht für Vitamin B6
nachgewiesen werden. Es wird vermutet, dass es zunächst zu einer Sättigung plazentarer
Rezeptoren kommen muss, bevor das Vitamin zum Fetus gelangt.
Zu ähnlichen Ergebnissen kamen Kirchgessner et al. (1985), sie ermittelten bei trächtigen Ratten,
welche ein Futter mit einem Vitamin B6-Gehalt von nur 1mg/kg erhielten, eine signifikant
verringerte Vitamin B6-Konzentration im Fetus, der Plazenta und der Amnionflüssigkeit. Bei
einem Futter mit 6mg Vitamin B6/kg, konnte jedoch eine ausreichende Vitaminversorgung des
Fetus, sowie eine Gewebesättigung der Reproduktions Organe nachgewiesen werden.
Wang und Trumbo (1992) konnten zeigen, dass ein verringerter Plasmaspiegel von
Pyridoxalphosphat während der Trächtigkeit von Ratten weder mit einer verringerten enternalen
Resorption noch mit einem erhöhten fetalem Bedarf zusammenhängt. Mittels von H3 Pyridoxin
konnte nachgewiesen werden, dass sich weniger als drei Prozent des oral aufgenommenen Vitamin
B6 im fetalen Gewebe oder im Uterusgewebe befinden. Es konnte jedoch eine Umverteilung von
Pyridoxalphosphat aus dem Plasma in die Erytrozyten ermittelt werden. So wurde ein Abfall der
Vitamin B6-Plasmakonzentration um 50%, bei einem gleichzeitigen Anstieg der Aktivität der
erythrozytären Aspartat-Aminotransferase (EAST) um 50% festgestellt, ohne dass eine exogene
Stimulation durch Pyrodoxalphosphat stattgefunden hatte.
16

Roth-Maier et al. (1996) zeigten, dass es bei Ratten mit einem hohen Versorgungsstatus von
Vitamin B6 während der Gravidität, am 14. Tag der Laktation zu Aktivitätszunahmen der ASAT
um 56% im Plasma, um 44% in den Erythrozyten und um 43% in der Leber kommt. Die
Aktivitätszunahmen beschreiben die Differenz der Enzymaktivität zwischen den Tieren mit der
geringsten Vitamin B6-Zulage (0,6mg/kg Futter) und der höchsten Zulage (180mg/kg Futter). Das
Futter wurde ad libitum angeboten. Eine Erhöhung der Vitamin B6-Zufuhr erst zu Beginn der
Laktation nach Mangelernährung während der Gravidität führte zu einer überdurchschnittlichen
Erhöhung der Enzymaktivität, da der Organismus bestrebt ist, das während der Gravidität
entstandene Defizit auszugleichen. Eine Unterversorgung während der Laktation konnte durch
bedarfsgerechte Versorgung während der Trächtigkeit ausgeglichen werden.
Bei der ALAT konnten keine Aktivitätsänderungen festgestellt werden, dieses Ergebnis kann
durch eine höhere Coenzymabsättigung erklärt werden.
Nach einem unbekanntem Autor (1972) werden bei trächtigen Ratten innerhalb von 24 Stunden
43% von injiziertem C14-markiertem Pyridoxol im Urin ausgeschieden. Hiervon waren allerdings
nur noch 16% in Form von Pyridoxol vorhanden, weitere 12% in Form von Pyridoxinsäure
Lactone und weitere 8% als Pyridoxinsäure. Übrige Formen wurden in geringer Menge renal
ausgeschieden. Im Blut wurde vorwiegend Pyridoxol und Pyridoxal nachgewiesen.
Nach Schenker et al. (1992) findet der plazentare Pyridoxaltransport in der menschlichen Plazenta
bidirektional statt. Die Transportrate von maternal nach fetal ist jedoch signifikant höher als
umgekehrt. Pyridoxine und Pyridoxal scheinen eine ähnliche Clearance zu besitzen, der Transport
von Pyridoxal 5-Phosphat ist jedoch deutlich geringer. Die Transporter sind nicht gesättigt und es
findet kein Transport gegen einen Konzentrationsgradienten statt. Die plazentare Konzentration
von Pyridoxal ist sowohl höher als die des fetalen, als auch des maternalen Blutes. Diese
Ergebnisse deuten auf einen passiven diaplazentaren Transport hin. Die erhöhte Konzentration von
Pyridoxal in der Plazenta deutet darauf hin, dass das Vitamin hier gebunden wird.
Im Gegensatz hierzu beschreiben Contractor und Shane (1971) einen aktiven diaplazentaren
Transport von Pyridoxal 5-Phosphat sowohl beim Menschen als auch bei der Ratte.
Nach Furth-Walker et al. (1989) sinkt der Plasma Pyridoxal 5-Phosphat Spiegel bei trächtigen
Mäusen um 50%, die Konzentration in den Erythrozyten steigt jedoch um den Faktor 2,9, so dass
im Vollblut ein Anstieg um den Faktor 1,6 zu beobachten ist.
Nach Leibman et al. (1990) besitzt die Plazenta trächtiger Mäuse eine hohe Aktivität alkalischer
Phosphatase. Es wird vermutet, dass die alkalische Phosphatase als Ektoenzym wirkt und eine
17

wichtige Rolle bei der Regulation des Pyridoxal-5-Phosphat-Plasmaspiegels spielt, da die
plasmatische alkalische Phosphatase ihre Aktivität während der Trächtigkeit um 50% verringert.
2.3 Aufbau und Entwicklung der Plazenta der Ratte
Zur Klärung der vorliegenden Fragestellung eignet sich die Ratte als Versuchstier aus mehreren
Gründen. Zwar gibt es nach Carter (2007) kein perfektes Tiermodell für die menschliche
Plazentation, die Ratte kommt diesem jedoch recht nahe.
Da sowohl Mensch als auch Ratte Omnivora (Allesfresser) sowie Monogaster (sie besitzen
lediglich einen einhöhligen Magen) sind, kann von einem ähnlichen Verdauungstrakt und
ähnlichen Enzymmustern ausgegangen werden.
Weitere Vorteile dieses Tiermodells sind eine hohe Reproduktionsrate, ein kurzes
Reproduktionsintervall sowie die geringe Größe der Tiere, wodurch die Haltung unkompliziert ist
(Carter, 2007).
Wie der Mensch, besitzt auch die Ratte eine Plazenta discoidalis, d.h. eine runde bis leicht ovale
makroskopische Gestalt, sowie eine Plazenta hämochorialis, so dass das mütterliche Blut direkt an
das Chorionepithel reicht, da sich maternales Gefäßepithel zurückgebildet hat (Franke, 1969). Der
Durchmesser der Rattenplazenta variiert zwischen 10 und 14 mm am 19. Tag der Trächtigkeit, bei
einer Dicke von 4-5 mm. Es bestehen jedoch beträchtliche Größenschwankungen – auch innerhalb
eines Muttertieres.
Im Gegensatz zur Ratte, die eine Plazenta hämotrichorialis ausbildet, entwickelt der Mensch eine
Plazenta hämomonochorialis. Das heißt, die Schranke zwischen maternalem und fetalem Blut
besteht bei der Ratte aus bis zu drei trophoblastischen Zellschichten, beim Menschen lediglich aus
einer. Ein weiterer Unterschied ist der bei der Ratte gut ausgebildete Spongiotrophoblast, der beim
Menschen fehlt.
Im Gegensatz zur menschlichen Zottenplazenta, entwickelt die Ratte eine Labyrinthplazenta
(Carter, 2007; Ramsey, 1982).
Dieser histomorphologische Unterschied, sowie das Vorhandensein des Spongiotrophoblasten
haben jedoch keine Auswirkungen auf den diaplazentaren Transport.
18

Der morphologische Aufbau der Rattenplazenta ist zwischen dem 18. und 20. Tag am deutlichsten
zu erkennen. Von maternal nach fetal sind folgende Schichten zu unterscheiden:
- Decidua basalis oder subchoriale Decidua mit einer
Zona Spongiosa und
Zona Compacta
- Riesenzellenscheide (giant cells)
- Trophospongium oder Spongiotrophoblast (junctional zone, Durchdringungszone)
- Labyrinth
Lediglich die Decidua basilaris ist maternalen Ursprungs. Die Unterteilung in Zona spongiosa und
Zona compacta beruht auf die unterschiedlich stark ausgebildete Interzellularsubstanz. Die
Feinstruktur der Zellorganellen in beiden Zonen stimmt überein, es bestehen lediglich quantitative
Unterschiede (Franke, 1969). Schiebler und Knoop (1959) beschreiben das Vorkommen von
Glykogenzellen in der Decidua basalis.
Nach Franke (1969) kann es bei Deciduazellen, die an die Riesenzellschicht grenzen, zu einer
Reihe feinstruktureller Veränderungen wie einer Faltung der Kernmembran, einer Vakuolisierung
des Zytoplasmas, Auftreten von Lipideinschlüssen und gequollenen Mitochondrien kommen.
Die in der Entwicklung noch geschlossene Riesenzellenscheide ist gegen Ende der Tragzeit (etwa
ab dem 18. Tag) lückenhaft, so dass kleinere Deziduazellen in die Riesenzellschicht eindringen
können und zum Teil bis zum Spongiotrophoblasten reichen. Aber auch bei jüngeren Plazenten
sind Riesenzellen zu finden, die in die Dezidua dringen. Mehrkernige Riesenzellen können durch
Verschmelzung mehrerer Deziduazellen entstehen. Dieses Gebiet, in dem fetales und maternales
Gewebe aneinander treffen ist in jüngeren Plazenten deutlicher getrennt. Erst später vermischen
sich fetale und maternale Zellen (Schiebler und Knoop, 1959).
Schiebler und Knoop (1959) berichten in der Trennlinie dieser Schichten in nicht durchmischten
Regionen von zum Teil mächtig ausgebildeter basalmembranartiger Interzellulärsubstanz.
Diese Beobachtung konnte von Franke (1969) jedoch nur partiell bestätigt werden.
Granulated cells sind natürliche Killerzellen, deren Funktion noch nicht abschließend geklärt ist
und die durch Desmosomen miteinander verbunden sind (Croy und Kiso, 1993; Franke, 1969).
Welsh und Enders (1993) vermuten, dass den granulierten Zellen eine entscheidende Rolle bei der
Induktion der Apoptose von Uterusepithelzellen zukommt und sie somit eine wichtige Funktion
bei der Implantation der Blastozyste haben.
19

Die Schicht des Trophospongiums besteht aus drei Zelltypen:
1. den Spongiotrophoblasten
2. den Glykogenzellen, deren Glykogengehalt ab dem 19. Tag rasch abnimmt und den
3. Riesenzellen (Franke, 1969; Jollie, 1964; Schiebler und Knoop (1959).
Nach Jollie (1964) entwickeln sich die trophoblastischen Riesenzellen wahrscheinlich durch
Wachstum und/oder Fusion aus Spongiotrophoblasten.
Schiebler und Knoop (1959) konnten zwei verschiedene Typen von Riesenzellen nachweisen, die
sich in Form und Größe der Kerne unterscheiden.
Jollie (1965) vermutet, dass die Spongiotrophoblasten das Hormon Luteotropin produzieren.
Hierfür spricht unter anderem das gut entwickelte Endoplasmatische Retikulum, sowie das
reichliche Vorkommen von Glykogen, wenn man beachtet, dass es sich bei Luteotropin um ein
Glykoprotein handelt. Auch die verminderte Produktion von Luteotropin gegen Ende der
Gravidität korrespondiert mit der Involution des Spongiotrophoblasten.
Laut Jollie (1964) sind in den Spongiotrophoblasten zwischen dem 8. und 18. Tag auffällig viele
Mitosen zu beobachten. Dies ist darauf zurückzuführen, dass sich hieraus sowohl die
Glykogenzellen als auch das Labyrinth differenzieren.
Dean et al. (1962) schreiben den Riesenzellen des Trophospongiums endokrine Funktionen zu.
Die Zellen produzieren Steroidhormone, wobei der Höhepunkt der Produktion zwischen dem 13.
und dem 15. Tag der Trächtigkeit liegt, danach sinkt die Hormonproduktion.
Auch die Synthese des Luteotropins scheint außer in den Spongiotrophoblasten auch in den
Riesenzellen des Trophospongiums stattzufinden.
Nach Davies und Glasser (1968) besteht das Labyrinth der Rattenplazenta aus drei Schichten
(1. Wand des maternalen Sinus, 2. und 3. Synzytiotrophoblast), welche die Plazentaschranke
(Jollie, 1965) bilden. Andere Autoren, wie Metz et al. (1976), benennen fünf Schichten. Dies liegt
daran, dass hier der perivaskuläre Raum sowie das fetale Kapillarendothel als eigene Schicht
benannt werden.
20

Die Wand des maternalen Sinus, welche als erste Schicht bezeichnet wird, ist die dünnste aller drei
Schichten und wird vom basophilen Trophoblastenepithel gebildet, die über die gesamte
Gravidität mitotische Aktivität zeigt und sich zum Ende der Schwangerschaft teilweise oder
komplett in Riesenzellen umwandeln kann.
Die zweite und dickste Schicht weist kein basophiles Zytoplasma auf und enthält kleinere
Zellkerne (Thliveris, 1976; Davies und Glasser, 1968). Nach Takata et al. (1997) und Metz et al.
(1976) handelt es sich bei der zweiten und dritten Schicht um ein Synzytium, den sog.
Synzytiotrophoblasten. Die zweite und dritte Schicht sind durch Gap junctions verbunden. Es wird
vermutet, daß sie die Funktion eines „Siebes“ haben und den diaplazentaren Transport limitieren
(Metz et al. 1976).
Die dritte Schicht ist durch eine aus dem Trophoblasten stammende Basalmembran vom fetalen
Mesenchym getrennt. Auch hier ist das Zytoplasma nicht basophil und die Zellen weisen kleinere
Kerne als in der ersten Schicht auf (Davies und Glasser, 1968).
Nach Thliveris (1976) und Takata et al. (1997) folgt der dritten Schicht das durch eine
Basallamina von ihr getrennte fenestrierte fetale Gefäßendothel, dessen Zellen durch tight
junctions miteinander verbunden sind (Metz et al. 1976).
Im Gegensatz zu den Zellen der ersten und zweiten Schicht, die durch zahlreiche Desmosomen
lose miteinander verbunden sind, sind die Zellen aus Schicht zwei und drei innig miteinander
Abb. 2.3.1: elektronenmikroskopische Aufnahme des Trophoblasten im Labyrinth der Rattenplazenta (Plazentaschranke) an Tag 14 der Trächtigkeit, 14.000 x Vergrößerung. Nach Jollie, 1964. FCB: fetale Blut Kapillare, e: Endothelzelle der fetalen Kapillare, MBS: maternaler Blut Sinus, I,II,III: trophoblastische Zellschichten
21

verbunden und weisen keine Desmosomen auf. Da es sich bei dem Trophoblastenepithel der ersten
Schicht um ein fenestriertes Epithel handelt, haben die Zellen der angrenzenden zweiten Schicht
die Möglichkeit sich in die maternalen Sinus vorzustülpen (Davies und Glasser, 1968).
Nach Bridgman (1948 b) sind maternale Sinus und fetale Gefäße gegen Ende der Trächtigkeit
lediglich durch ein fetales Gefäßendothel getrennt. Dieser Ansicht schließt sich Jollie (1964)
jedoch nicht an. Nach ihm besteht die Plazentaschranke während der gesamten Trächtigkeit aus
vier zytoplasmatischen Schichten. Da jedoch gegen Ende der Tragzeit sowohl das fetale
Gefäßendothel fenestriert ist, als auch die erste Trophoblastenschicht besteht die Plazentaschranke
lediglich noch aus dem Trophoblasten II und der dritten Zellschicht.
Der Trophoblast II nach Jollie (1964) entspricht dem Synzytiotrophoblasten der Schicht Zwei nach
Takata et al. (1997) und Metz et al. (1976).
Beschriebene Erleichterungen des diaplazentaren Stofftransports gegen Ende der Gravidität
können durch diese Veränderungen der Plazentaschranke erklärt werden.
Die ersten Uterusdrüsenzellen (granulated metrial gland cells) sind noch vor ihrem Erscheinen im
mesometrialen Triangel im Ektoplazentakonus und in der Dezidua basilaris zu finden. Aus dem
Ektoplazentakonus kommt es zur Migration der Zellen in Richtung des mesometrialen Triangels,
indem sie eine perivaskuläre Lage einnehmen (Dixon und Bulmer, 1971). Nach Peel (1989) sind
die Zellen ab der dritten Woche p.c. in großer Zahl in den Uterusdrüsen anzutreffen. Die Zellen
werden aufgrund ihrer Glykoprotein und hydrolytische Enzyme enthaltenden Granula auch als
granulierte Uterusdrüsenzellen bezeichnet. Einige dieser Zellen sind reich an Glykogen.
Dallenbach-Hellweg et al. (1965) konnten in den granulierten Zellen der Uterusdrüsen das
Peptidhormon Relaxin nachweisen. Die Hauptfunktion des Relaxins besteht in der Aufhebung von
Fasern von zusammenhaltendem Gewebe, vor allem von Kollagen.
Nach Peel (1989) werden gegen Ende der Trächtigkeit einige Zellen in den maternalen Sinus des
Labyrinths nachgewiesen. Es wird vermutet, dass es zu Interaktion zwischen diesen Zellen und
den Trophoblastenzellen der ersten Schicht kommt, welche im Zusammenhang mit der
Degeneration der Trophoblastenzellen und der Uterusdrüsenzellen steht. Peel und Adam (1991)
vermuten, dass die granulierten Uterusdrüsenzellen zur Gruppe der Killerzellen gehören. Sie
konnten eine zytotoxische Aktivität der Zellen gegenüber co-kultivierten Trophoblastenzellen
nachweisen.
22

Noch vor Beginn der Implantation kommt es im Endometrium zur sog. Deziduareaktion.
Als Deziduareaktion wird ein durch Östrogen und Progesteron induzierter Prozess bezeichnet,
welcher die maternalen Stromazellen in helle glykogenreiche Zellen umwandelt.
Diese Reaktion kann ab dem vierten Tag der Trächtigkeit zunächst in der Umgebung maternaler
Gefäße beobachtet werden (de Rijk et al., 2002). Weiterhin konnten Mulholland et al. (1992)
zeigen, dass am Implantationspol des Uterus Kollagen des Typs VI weitgehend verschwunden ist,
und die Expression des Intermediätfilaments Desmin in denselben Zellen zugenommen hat. Bei
der Rückentwicklung der Deziduazellen, konnte eine Umkehr dieses Prozesses nachgewiesen
werden. Diese Ergebnisse lassen Mulholland et al. (1992) vermuten, dass es zu einer weiteren
funktionellen Differenzierung dieser Zellen als Antwort auf die Implantation der Blastozyste
kommt.
Nach Enders und Schlafke (1967) beginnt der Prozess der Implantation der Blastozyste am Abend
des fünften Tages nach Bedeckung, durch Anheftung der Blastozyste am Endometrium, nahe der
antimesometrialen Seite des Uterus, wobei der Embryoblast mesometrial orientiert ist.
In diesem Zusammenhang ist anzumerken, dass die Implantation zwar antimesometrial, die
Entwicklung der definitiven Plazenta jedoch mesometrial stattfindet. Für den „Wechsel“ der Seite
des Embryos kommt es zu einem vorübergehenden Schluss des Uteruslumen (Ramsey, 1982).
Ab dem sechsten Tag besteht das Stroma um die implantierte Blastozyste aus Deziduazellen mit
leicht basophilem Zytoplasma, großen, runden Nuklei mit verschieden vielen Nukleoli. Die Zellen
zeichnen sich durch PAS-positive Granula aus. Ab dem achten Tag findet eine Umwandlung der
Deziduazellen von große in kleine Zellen statt, ab dem 14. Tag der Trächtigkeit sind nur noch
kleine Deziduazellen anzutreffen.
Zwischen dem sechsten und dem zehnten Tag sind Mitosen der Deziduazellen zu beobachten,
wobei die höchste Aktivität am Tag acht zu finden ist (Enders und Schlafke, 1967).
Vor der myometrialen Deziduareaktion, die vom 10. bis zum 20. Tag andauert und auf der
plazentaren Seite des Uterus stattfindet, kommt es zwischen dem 10. und 12. Tag zu myometrialen
Ödemen, die jedoch wieder vollständig verschwinden (de Rijk et al., 2002).
Vor Bildung der definitiven, mesometerial liegenden chorioallantoischen Plazenta, die sich ab dem
10. Tag bildet, ist eine antimesometrial liegende Dottersackplazenta vorhanden. Am neunten Tag
jedoch kommt es zu autophagischen Vorgängen an der Dottersackplazenta welche anschließend
degeneriert (Kaloglu et al., 2003).
23

Nach Bridgman (1948a) sind ab dem sechsten Trächtigkeitstag große supranukleäre proteinhaltige
Granula im viszeralen Entoderm zu finden, welches die „innere“ Membran des Dottersacks
darstellt. Diese Granula sind am neunten Tag nicht mehr nachweisbar, eventuell haben sie sich in
die Dottersackhöhle herausgestoßen. Es formen sich jedoch neue kleinere Granula, die sich bis
zum 15. Tag erneut vergrößert haben. Außerdem ist reichlich Glykogen in den Dottersackzellen
anzutreffen.
Einen Tag später rupturiert die Reichertssche Membran und Fett welches im Uterusepithel nahe
der Fruchtblase nachweisbar war, verschwindet. Glykogen und proteinhaltige Granula sind jedoch
bis zum 20. Tag weiterhin anzutreffen.
Den Granula wies Litwer (1928) bei Mäusen sekretorische Funktionen zu, während Wislocki et al.
(1946) sie als Produkte von Absorption und Phagozytose deuteten. Hierfür spricht nach Wislocki
et al. (1946) der Nachweis von Eisen im Dottersack, welches aus phagozytierten maternalen
Erythrozyten stammen könnte.
Die Reichertsche Membran besteht aus Kollagen und wird nach Mutmaßungen von Wislocki et al.
(1946) durch Riesenzellen gebildet. Merker und Villegas (1970) konnten zeigen, dass die
Reichertsche Membran vom parietalen Dottersackentoderm gebildet wird, dessen Zellen ihr
aufliegen. Maternales Plasma kann direkt das viszerale Dottersackentoderm erreichen. Die
Membran stellt die einzige kontinuierliche Schicht zwischen maternalem Blut und Träger dar. Die
von Wislocki et al. (1946) angenommene phagozytotische Aktivität des viszeralen
Dottersackentoderm wird von Merker und Villegas (1970) bestätigt, die Granula als
Abb. 2.3.2: Blutfluss in der Dottersackplazenta nach Merker und Villegas, 1970. A Archamnionhöhle; DG Deziduagefäße; vD viszerales Dottersackentoderm; pD parietales Dottersackentoderm; EPK Ektoplazentakonus; PS periembryonaler Sinus
24

Phagolysosomen charakterisiert. Maternales Plasma kann somit aufgenommen, abgebaut und dem
Keimling zur Verfügung gestellt werden. Das maternale Blut erreicht das viszerale Entoderm über
einen periembryonalen Randsinus, der nach innen durch die Reichertsche Membran und die
Zellen des parietalen Blattes des Dottersackentoderms, nach außen durch Trophoblasten- und
Deziduazellen abgegrenzt wird. Der Randsinus endet als Blindsack, d.h. eingeströmtes Blut kann
den Sinus nicht mehr verlassen.
Die Reichertsche Membran hat somit die Funktion eines Ultrafilters, so dass maternale Blutzellen
nicht in den Dottersack gelangen können. Die chorioallantoische Plazenta zeichnet sich durch ihre
Entstehung aus der Chorionschicht und allantoischem Mesenchym aus.
Abb. 2.3.3: Entstehung der chorioallantoische Plazenta: AH Amnionhöhle; AM allantoisches Mesenchym; vDE viszerales Dottersackentoderm; pDE parietales Dottersackentoderm; EC Exocoelom; EH Ektoplazentahöhle; EK Ektoplazentakonus; L Lakune; MS maternale Sinus; OES obere Ektodermalschicht; UES untere Ektodermalschicht; ZM extraembryonales Mesoderm; in Anlehnung an Pijneborg und Vercruysse, 2006.
25

Zur besseren Übersicht, kann die Entwicklung der chorioallantoischen Plazenta in drei Stadien
eingeteilt werden (Pijnenborg und Vercruysse, 2006):
- Zeitraum der anfänglichen Entwicklung der Ektoplazenta (Tag 8-10)
- Zeitraum der ektoplazentalen Entwicklung (Tag 11-17)
- Endgültige Reifung der Ektoplazenta (Tag 18-21)
Die Entwicklung der Plazenta beginnt am achten Tag (Pijnenborg und Vercruysse, 2006) bzw. laut
Franke (1969) am sechsten Tag, mit der Differenzierung eines solidzelligen Trophoblastenzapfens,
der sich in Träger (Franke, 1969) oder Ektoplazentarkonus (Pijnenborg und Vercruysse, 2006)
unterteilt. Aus dem Träger bzw. dem Ektoplazentakonus entwickelt sich die spätere Plazenta.
Da sich der Träger aus dem Trophoblasten entwickelt, ist er ektodermaler Herkunft (de Rijk et al.,
2002; Pijnenborg und Vercruysse, 2006).
Der Träger stellt die Verbindung zwischen Embryoblasten und Dezidua her. Durch aktive
Vordringungsprozesse des Trägers gegen die Deziduazellen werden maternale Kapillaren eröffnet.
Etwa am neunten Tag bildet sich im Träger ein Lakunensystem, in das maternale Blut strömt.
Pijnenborg und Vercruysse, 2006 sprechen in diesem Zusammenhang von einer
Implantationskammer, in welche sich das maternale Blut ergießt. Dieser Bereich, in dem fetales
Gewebe (der Träger) und die maternalen Deciduazellen aneinander treffen, wird als
Durchdringungszone bezeichnet. In dieser Zone entwickeln sich ab dem fünften Tag Riesenzellen
aus Trophoblastenzellen, die durch ihre ausgeprägte Phagozytoseaktivität eine wichtige Rolle bei
der Implantation spielen (Franke, 1969). Gleichzeitig entwickelt sich die Blastozyste durch das
Wachstum des Mesoderms weiter und unterteilt sich in Ektoplazentarhöhle, Extraembryonales
Coelom (Exocoelom) und Amnionhöhle. Die Ektoplazentarhöhle ist nun durch eine obere
Ektoplazentarschicht vom Träger abgegrenzt, nach unten ist sie ebenfalls durch eine dünnere
untere Ektoplazentarschicht (echtes Chorion), vom Exozoelom abgegrenzt.
Zwischen dem neunten und zehnten Tag kommt es durch starkes Wachstum des allantoischen
Mesenchyms zur Verlegung der Ektoplazentarhöhle. Obere und untere Ektoplazentarschicht liegen
nun direkt aufeinander und bilden die Chorionschicht. Der Kontakt zwischen allantoischem
Mesenchym und Chorion findet bei ca. 80% der Embryonen mittig statt, nur bei etwa 20% wächst
das Mesenchym seitlich an das Chorion (Ellington, 1987). Zwischen den beiden Blättern der
Lamina sind jedoch Lakunen zu erkennen, die sich mit maternalem Blut anfüllen (Pijnenborg und
Vercruysse, 2006). Die Chorionschicht, die auf ihrer antimesenterialen Seite von allantoischem
Mesoderm überzogen ist, hat sich eventuell nicht nur aus den Epithelzellen der Ektoplazentahöhle
entwickelt, sondern es besteht auch die Möglichkeit, dass sich einige Zellen aus dem Träger in der
Lamina befinden. Aus der Lamina die am 10. Tag in ihrer Mitte lediglich aus einer Schicht von 4-
26

8 kleinen Zellenreihen besteht, entwickelt sich später das Labyrinth, wobei es für möglich
gehalten wird, dass in der späteren Entwicklung das Wachstum des Labyrinthes mit Zellen aus der
Durchdringungszone stattfindet (Peel und Bulmer, 1977; Jollie, 1964).
Der Dottersack umgibt jetzt mit einer viszeralen und einer parientalen Schicht alle drei Kammern
bzw. Höhlen und lässt lediglich zwischen der Ektoplazentahöhle und dem Träger einen Spalt
(Pijnenborg und Vercruysse, 2006).
Parallel zu der Plazentaentwicklung kommt es zu einer massiven Verdickung des endometrialen
Stromas, wodurch das gesamte Uteruslumen verschwindet, das Schleimhautepithel löst sich bei
diesem Prozess auf. Am achten Tag bildet sich auf der antimesometrialen Seite ein neues
Uteruslumen.
Im zweiten Stadium der Plazentaentwicklung, besteht bereits die grundlegende Architektur der
späteren Plazenta.
Zwischen dem 11. und 17. Tag sind fetale Gefäße im allantoischen Mesenchym zu finden, welches
mit der unteren Ektoplazentaschicht verschmilzt. In dem sich jetzt entwickelnden Labyrinth sind
bereits maternale Sinus und fetale Gefäße anzutreffen. Auch im Trophospongium sind maternale
Lakunen erkennbar. Der Träger ist nun verschwunden, lediglich in den lateralen Plazentazonen
sind noch übrig gebliebene Träger-Zellen zu erkennen.
Zwischen dem 15. und 17. Tag ist zunächst das Einwachsen von meist zwei afferenten maternalen
Gefäßen aus der Dezidua in Richtung des Labyrinth zu beobachten. Auch dringt der
Spongiotrophoblast nun aktiv in die maternale Dezidua ein.
Zwischen dem 18. und 21. Tag, kommt es zur Resorption von vorher gebildeten Strukturen. Der
Spongiotrophoblast hat sich nun jedoch soweit vergrößert, dass er scheinbar den Raum der
Dezidua basilaris einnimmt, welche resorbiert wird.
Die zum Ende der Trächtigkeit immer weniger werdenden Überreste der Dezidua basilaris werden
nach der Geburt mit ausgestoßen. Des weiteren ist eine „Verwischung“ der Grenze zwischen
Trophospongium und Labyrinth zu erkennen. Die Zellkerne der äußeren Bereiche des
Spongiotrophoblasten vergrößern sich gegen Ende der Trächtigkeit.
Große fetale Gefäße in der Plazenta sind nun von einer mesodermalen Hülle umgeben, der größere
Höhlen folgen, die seitlich durch Entoderm begrenzt werden (Pijnenborg und Vercruysse, 2006).
Zu Beginn der Plazentaentwicklung, besteht fast die gesamte Plazenta aus maternalen
Deziduazellen, lediglich basophile- und Riesenzellen sind fetalen Ursprungs. Obwohl es auch zu
einer Proliferation dieser Zellen kommt, die ihren Höhepunkt am 14. Tag (basophile Zellen) bzw.
am 17. Tag (Riesenzellen) haben, ändert sich erst durch das schnelle Wachstum des Labyrinths das
27

Überwiegen fetaler Zellen. Ab dem 15. Tag stellt das Labyrinth den größten Teil der Plazenta dar,
entwickelt sich jedoch bis zum 22. Tag weiter und dominiert dann die gesamte Plazenta (Durst-
Zivkovic, 1978).
2.4 Diaplazentare Transportmechanismen
Kyuma (1984) vermutet, dass der diaplazentare Aminosäuretransfer zur fetalen Seite mittels
stereospezifischer Transportproteine oder erleichterter Diffusion stattfindet. Die Ergebnisse seiner
Arbeit lassen ihn weiterhin vermuten, dass nicht die maternale, sondern überwiegend die fetale
Konzentration freier Aminosäuren im Plasma den diaplazentaren Transport regulieren.
Nach Battaglia und Regnault (2001) kann der Transport von Aminosäuren im Trophoblasten in
drei Schritte unterteilt werden: erstens die Aufnahme der Aminosäure aus dem maternalen Blut
über die apikale Oberfläche der Trophoblastenzelle, zweitens den Transport durch das Zytoplasma
der Zelle und drittens den Transport aus der Zellen hinaus und durch die Basalmembran in die
fetale Kapillare. Transportproteine werden jeweils beim Transport über Plasmamembranen, also
bei Schritt Eins und Drei benötigt. Hierbei ist zu beachten, dass die an der apikalen Zellmembran
vorhandenen Transportproteine nicht immer denen an der basalen Trophoblastenzellmembran
entsprechen. Die Transportproteine können Na+-abhängig bzw. Na+-unabhängig sein, zum anderen
können sie bezüglich ihres Substrates in die Gruppen A, ASC, L, y+, β und Glycin eingeteilt
werden, wobei einzelne Gruppen mehrere Systeme enthalten können.
28

Abb. 2.4.1: Schematische Abbildung ausgewählter Na+-abhängiger () und Na+-unabhängiger (○) Aminosäuretransport Systeme des Trophoblasten der Plazenta des Menschen und der Ratte. apikale Membran= maternal, basale Membran= fetal, MeAIB: Methylaminoisobuttersäure, AIB: α-Aminoisobuttersäure, BCH: 2-Aminobicyclo-(2,2,1)-heptane-2-carbonsäure. In Anlehnung an Regnault et al. (2005).
Nach Schneider und Dancis (1974) geschieht die Aufnahme saurer Aminosäuren (Glutamat und
Aspartat), sowie Serin recht schnell und es stellt sich ein großer Konzentrationsgradient zwischen
Extrazellular- und Intrazellularraum ein. Die Abgabe der Aminosäuren ist offensichtlich langsamer
als die Aufnahme. Im Gegensatz hierzu ist die Aufnahme basischer Aminosäuren (Lysin und
Arginin) weniger schnell und die Akkumulation im Gewebe erreicht schnell ein Plateau. Dies
deutet auf einen effizienten Abtransport der Aminosäuren hin. Neutrale Aminosäuren wie Glycin,
Alanin und Prolin zeigen ein ähnliches Aufnahmeverhalten wie die basischen Aminosäuren.
Leucin zeigt eine schnelle Abgabe aus dem Plazentagewebe, so dass der Konzentrationsgradient
intrazellulär : extrazellulär typischerweise unter Eins blieb.
Nach Battaglia und Regnault (2001) ist die Plazenta nicht nur in der Lage freie Aminosäuren des
maternalen Plasmas zu resorbieren und unter Umständen zu metabolisieren, sondern sie ist auch in
der Lage, maternale Proteine abzubauen und dessen Aminosäuren zum Feten hin zu transportieren.
29

Auch besitzt die Plazenta die Möglichkeit, Aminosäuren zu deren Ketosäuren zu desaminieren
und in den fetalen Kreislauf zu transportieren. Die Ketosäuren können dort entweder zu ihren
entsprechenden Aminosäuren transaminiert werden, oder sie werden weiter decarboxyliert. Dies
ist bei Leucin und wahrscheinlich auch bei Isoleucin und Valin der Fall (Loy et al., 1990). Der
Stofftransport in der Plazenta ist nicht unidirektional, es kann u.a. bei Leucin zu einem
fetomaternalen Transfer kommen (Ross et al., 1996).
Nach Cetin (2001) werden Aminosäuren aktiv zum Feten transportiert und liegen hier in höherer
Konzentration vor, als im maternalen Plasma. Die meisten Aminosäuren können in der Plazenta
metabolisiert werden, und bei manchen nicht essentiellen Aminosäuren konnte deren Synthese in
der Plazenta nachgewiesen werden. Der Transfer von essentiellen Aminosäuren ist signifikant
höher, als der nicht essentieller Aminosäuren.
Kedenburg und Mülling (1975) halten es für möglich, dass bei der Ratte ähnlich wie beim
Kaninchen in der die Amnionhöhle umgebende Dottersackwand aktive Transportproteine für
Aminosäuren lokalisiert sind. Hierdurch könne die im Vergleich zum Menschen hohe
Aminosäurekonzentration in der Amnionflüssigkeit zu erklären sein.
Nach Lines und Waisman (1971) ist die Konzentration von Aminosäuren im fetalen Blut um den
Faktor 1,5 bis 2,4 höher, als im maternalen Blut. Andere Autoren wie Beckman et al. (1997)
berichten jedoch zum Teil von größeren Konzentrationsunterschieden.
Paolini et al. (2001) konnten beim Schaf nachweisen, dass die Regulation von Transportsystemen
essentieller Aminosäuren von der Genexpression entsprechender Transportproteine und der
maternalen sowie fetalen Plasmakonzentration der entsprechenden Aminosäure abhängt.
Der beträchtliche Bedarf des Feten an Glycin als Beispiel für eine nicht essentielle Aminosäure
wird in der menschlichen und der Rattenplazenta nach Bennett und Jackson (1998) allerdings
lediglich durch die folatabhängige Umwandlung von Serin zu Glycin durch die
Serinhydroxymethyltransferase gedeckt. Die Plazenta besitzt nicht die Möglichkeit Alanin mit
Hilfe der Glyoxylataminotransferase in Glycin umzuwandeln.
Nach Malandro et al. (1996) kommt es bei Ratten nach einer Reduktion der maternalen Aufnahme
von Casein (20% Casein vs. 5% Casein im Futter) zwar nicht zu einer verringerten maternalen
Plasmaaminosäurekonzentration, jedoch war das fetomaternale Aminosäureverhältnis im Plasma
reduziert. Aufgrund dieser Ergebnisse liegt es nahe, dass der Nährstofftransfer zum Fetus reduziert
ist. Es kam vor allem zu einer Reduktion der Aktivitäten des Na+-abhängigen neutralen
Aminosäure Transporters A, während der Transport des Systems ASC unbeeinflusst blieb. Auch
die Aktivität des Na+-abhängigen anionischen Aminosäure Transporters EAAC1 war reduziert,
30

während Na+-unabhängige Transportsysteme kaum betroffen waren. Es wird vermutet, dass eine
verringerte Synthese der Transportsysteme für die Aktivitätsreduktion der Systeme EAAC1 und
CAT1 verantwortlich sind.
Nach Verabreichung eines Futters mit einem erhöhten Glukoseanteil (24% anstatt 12%) wurde in
der Amnionflüssigkeit eine um 364% erhöhte Methioninkonzentration sowie ein signifikanter
Anstieg des Phenylalanins und ein konstanter Abfall von Taurin zwischen dem 18. und dem 21.
Trächtigkeitstag beobachtet. Dabei korrelierten diese Werte auch mit dem ansteigendem
Fetalgewicht bzw. mit vortlaufender Trächtigkeit. Auch bei den Aminosäuren Cystein, Lysin und
Tyrosin konnte ein Zusammenhang zwischen einer erhöhten Konzentration in der
Amnionflüssigkeit und einem Anstieg des fetalen Gewichtes beobachtet werden (Gurekian und
Koski, 2005).
2.5 Methylierungszyklus
Der Methylierungszyklus (siehe Abb.2.5.1) stellt einen wichtigen Stoffwechselweg zur Regulation
der Homocysteinkonzentration im Blut dar. Damit steht er ebenfalls im Mittelpunkt des Interesses
bei der Betrachtung der Hyperhomocysteinämie und den Erkrankungen die hierdurch gefördert
werden.
Laut Goddijn-Wessel et al. (1996) ist bei Frauen mit einer Hyperhomocysteinämie das Risiko für
Plazentaablösungen oder Plazentainfarkte deutlich erhöht. Die maternalen Konzentrationen von
Vitamin B6, B12 und der Serumfolatspiegel waren bei den Frauen mit Hyperhomocysteinämie
verringert. Die Folsäurekonzentration im Erythrozytenkonzentrat ist bei Frauen mit und ohne
Hyperhomocysteinämie vergleichbar gewesen.
31

Nach Zhao et al. (2001, 2002) beeinträchtigt Homocystein die Entwicklung der viszeralen
Dottersackwand, sowie die Ausbildung von Blutgefäßen. Die Beeinträchtigungen zeigten sich in
geschrumpften Oberflächen, mangelhaften Blutinseln und signifikant niedrigerem Durchmesser
des Dottersacks. Funktionell beeinträchtigt Homocystein den Transport in der viszeralen
Dottersackwand, speziell die Endozytose. Es wird vermutet, dass der verringerte Stofftransport
direkt mit der teratogenen Wirkung des Homocysteins korreliert.
Di Simone et al. (2004) wiesen bei menschlichen Trophoblastenzellen, welche einer
Homocysteinkonzentration von 20 µmol/l ausgesetzt waren, Apoptose und Einschränkungen der
Zellfunktion nach. Nach vorheriger Behandlung der Zellen mit Folsäure (20 nmol/l) konnte eine
Einschränkung dieser hervorgerufenen Effekte beobachtet werden.
Im weiteren Verlauf der Trächtigkeit, spielen die Enzyme Methioninsynthase- welches die
Reaktion Homocystein zu Methionin katalysiert, Cystathionin-β-Synthase und S-Adenosyl-
Homocysteinhydrolase- welches die Reaktion S-adenosylhomocystein zu Homocystein katalysiert
eine wichtige Rolle bei der Senkung des fetalen Homocysteinspiegels. Aerts et al. (1995) konnten
die Methioninsynthase im fetalen Gewebe zwischen dem 10 und 20 Trächtigkeitstag nachweisen.
Die höchste Enzymaktivität wiesen sie am 12. Tag nach. In der Plazenta war die Aktivität der
Abb. 2.5.1: Vereinfachtes Schema des Methionin-Homocystein Metabolismusin: 1: Cystathionin β-Synthase, 2: 5-Methyltetrahydrofolat Homocystein Methyltransferase, 3: 5,10-Methylentetrahydrofolat Reduktase, SAM: S-Adenosylmethionin, SAH: S-Adenosylhomocystein, THF: Tetrahydrofolat, 5-m-THF: 5-Methyltetrahydrofolat, 5,10-m-THF: 5,10-Methylentetrahydrofolat. In Anlehnung an Goddijn-Wessel et al., 1996.
32

Methioninsynthase während der gesamten Trächtigkeit geringer und unterlag keinen signifikanten
Schwankungen.
Die Cystathionin-β-Synthase war in plazentarem Gewebe, und ab dem 18. Trächtigkeitstag im
fetalen Gewebe nachweisbar. In der fetalen Leber kommt es seit dem 14. Trächtigkeitstag zu
einem konstanten Anstieg der Enzymaktivität, hingegen nimmt hier die Aktivität der
Methioninsynthase leicht ab. Während die Aktivität der Cystathionin-β-Synthase in der fetalen
Leber ab dem 14. Tag zunimmt, nimmt sie zwischen dem 10. und 18. Tag in der Plazenta ab, bis
sie kaum mehr nachzuweisen ist.
Die S-Adenosylhomocysteinhydrolase konnte zwischen dem 9,5 und 11,5 Tag in dezidualem
Gewebe, der parietalen und viszeralen Dottersackwand, dem Ektoplazentakonus sowie im
embryonalen Gewebe mit unterschiedlicher Aktivität nachgewiesen werden.
Diese Ergebnisse zeigen, dass der Methylierungszyklus bereits während der Neurulation in
fetalem Gewebe aktiv ist.
Nach Steegers-Theunissen et al. (1995) ist im Durchschnitt bei Frauen, die Kinder mit
Neuralrohrdefekt gebären, die Homocysteinkonzentration in der Amnionflüssigkeit signifikant
höher, als bei Frauen, die gesunde Kinder gebären. Die Homocysteinkonzentration im maternalen
Blut sowie die Konzentrationen von Folat, Vitamin B6 sowie Vitamin B12 zeigten in der
Amnionflüssigkeit und im maternalen Blut keinen signifikanten Unterschied zwischen den Frauen
welche gesunde und welche Kinder mit Neuralrohrdefekt gebaren. Die durchschnittliche
Konzentration von Homocystein und Vitamin B6 waren in beiden Gruppen in der
Amnionflüssigkeit signifikant geringer als im maternalen Blut, während Vitamin B12 in der
Amnionflüssigkeit in einer höheren Konzentration vorlag als im maternalen Blut.
Des weiteren berichten Steegers-Theunissen et al. (1997) von vergleichsweise hohen
Methioninkonzentrationen und geringen Homocysteinkonzentrationen in der Amnionflüssigkeit
im Gegensatz zum maternalen Blut zwischen der 8. und 12. Schwangerschaftswoche bei Frauen.
Aufgrund dieser Ergebnisse vermuten sie einen Methionin Metabolismus auf fetaler Seite.
Wendstrom et al. (2000) berichten das sowohl eine Mutation des fetalen Gens für die 5,10-m-THF
als auch eine erhöhte amniale Homocysteinkonzentration mit Neuralrohrdefekten assoziiert sind,
wobei keine Assoziation zwischen der Mutation und der erhöhten amnialen
Homocysteinkonzentration besteht.
33

Abb. 2.5.2: ausgewählte Abbauwege des Methionins. In Anlehnung an Caltrider und Niss, 1966.
2.6 Zusammensetzung der Amnionflüssigkeit
Wie in Abb. 2.6.1 zu erkennen ist, gibt es mehrere Wege, die die Zusammensetzung der
Amnionflüssigkeit modulieren bzw. beeinflussen können. Hierbei kann zunächst zwischen einem
passiven und einem aktiven Transport unterschieden werden. Zu den passiven
Abb. 2.6.1: Die Zusammensetzung der Amnionflüssigkeit beeinflussende Faktoren. In Anlehnung an Weingärtner et al. 2007.
34

Transportmechanismen zählen die Diffusion von Stoffen aus dem mütterlichen Blut und aus der
Nabelschnurvene. In der Plazenta sind zum einen aktive Transportmechanismen vorhanden,
welche in der Plazentaschranke lokalisiert sind, zum anderen kommt es auch hier zur passiven
Diffusion von Stoffen in die Amnionflüssigkeit.
Des weiteren gelangen fetaler Urin sowie abgeschliffene Epithelzellen in die Amnionflüssigkeit.
Auch kann es zu Diffusion von Stoffen durch fetale Schleimhäute kommen.
Ist die Konzentration freier Aminosäuren in der Amnionflüssigkeit höher als im maternalen
Plasma, ist davon auszugehen das die Plazenta mit ihren aktiven Transportmechanismen der Weg
ist, auf welchem die Aminosäuren in nennenswertem Umfang in die Amnionflüsigkeit gelangen
konnten. Die Aminosäuren können nach dem aktiven diaplazentaren Transport entweder über die
Plazenta oder über die Nabelschnurvene in die Amnionflüssigkeit diffundiert sein. Möglicherweise
sind bei der Ratte auch in der die Amnionhöhle umgebenden Dottersackwand aktive
Transportproteine für Aminosäuren lokalisiert (Kedenburg und Mülling, 1975).
3. Material und Methoden
3.1 Material
Der Tierversuch wurde in Mecklenburg-Vorpommern unter der Genehmigungsnummer
LLALLF M-V/TSD/7221.3-2.1-032/08 durchgeführt.
Für die Untersuchungen wurden weibliche Ratten des Zuchtstammes LEW1.W verwandt. Die
Böcke des Zuchtstammes kamen lediglich zur Bedeckung der weiblichen Tiere zum Einsatz. Die
weiblichen Tiere waren ausschließlich erstgebährend, um sicher zu stellen, dass die Zahl der
Implantationen und Resorptionen ausschließlich auf die aktuelle Trächtigkeit zurückzuführen ist.
Da es sich bei den Tieren um Individuen aus einer Inzucht handelt, kann von relativ homogenen
Stoffwechselparametern ausgegangen werden.
Die Ratten wurden in K3-Käfigen zu je drei Tieren gehalten. Die Bedeckung erfolgte über Nacht
im Verhältnis Bock : Weibchen von 1:2. Die Böcke wurden nach erfolgter Bedeckung wieder
separiert. Am Morgen nach der Bedeckung wurden Vaginalabstriche genommen, Weibchen mit
Spermien im Abstrich wurden als gedeckt angenommen. Der erste Tag nach der Bedeckung wurde
als Tag Null definiert. Für die weitere Arbeit wurden die gedeckten Tiere in die Gruppen K
(Kontrollgruppe n=16), B (induzierter Vitamin B6-Mangel n=16) und F (induzierter
35

Folsäuremangel n=15) unterteilt. Die Fütterungen der Gruppen B und F erfolgte ab Tag Null durch
das jeweilige Mangelfutter.
Für die Gruppe B kam das Experimentalfuttermittel „ssniff EFR/M, 10mm, Pyridoxin-arm,
S0752-S010, Batch No.: 1567414“ der Firma ssniff Spezialdiäten GmbH, 59494 Soest,
Deutschland zum Einsatz. Die Zusammensetzung bestand aus 20,8% Rohprotein, 4,2% Rohfett,
5% Rohfaser, 5,6% Rohasche, 46,8% Stärke, 10,8% Zucker sowie pro kg 15.000 IE Vitamin A,
1.500 IE Vitamin D3, 150mg Vitamin E, 20mg Vitamin K3, 30mg Vitamin C und 14mg Kupfer.
Die Fütterung der Gruppe F erfolgte mit dem Experimentalfuttermittel „sniff EFR/M, 10mm,
Folsäure-arm, S0752-S020, Batch No.: 1577414“ der Firma ssniff Spezialdiäten GmbH, 59494
Soest, Deutschland. Die Zusammensetzung entspricht der des Pyridoxin-armen Futtermittels,
jedoch ohne Folsäure.
Die Fütterung der Gruppe K erfolgte mit dem Aufzuchtfuttermittel „ssniff R-Z, extrudiert, V1326-
000 Reg.nr.: nl 96093“ der Firma ssniff Spezialkiäten GmbH, 59494 Soest, Deutschland.
Die Zusammensetzung bestand aus 21% Rohprotein, 3,8% Rohfett, 4,4% Rohfaser und 6,7%
Rohasche, sowie 15.000 IE/kg Vitamin A, 1.000 IE/Kg Vitamin D3, 100 mg/kg Vitamin E und 12
mg/kg Kupfer.
Die Futteraufnahme und Tränkung der Tiere erfolgte ad libitum, es wurde ein Lichtregime mit
einem Rhythmus von 12 Stunden hell und 12 Stunden dunkel eingesetzt (6-18 Uhr hell, 18-6 Uhr
dunkel). Die Temperatur betrug 22 °C +/- 1 °C, die Luftfeuchtigkeit ca. 50-52%. Der Streuwechsel
erfolgte zweimal wöchentlich, es wurde das Produkt Lingocel der Firma J. Rettenmaier & Söhne
GmbH + Co. KG in 73494 Rosenberg, Deutschland verwendet.
3.2 Methoden
Für die Probenentnahme wurden die Versuchstiere anästhesiert. Die Anästhesie erfolgte mit 10%
Ketamin (Wirkstoff: Ketaminhydrochlorid), 2% Rompun (Wirkstoff: Xylazinhydrochlorid) und
Aqua dest. im Verhältnis 1:1:1 durch intraperitoneale Injektion von 0,1ml je 100g Körpergewicht.
Die Eröffnung der Bauchdecke und des Thorax zur Entblutung der Muttertiere durch intrakardiale
Punktion des rechten Ventrikels mittels einer heparinisierten Spritze erfolgte während der
Tiefennarkose. Anschließend wurde das Vollblut bei 3.000 rpm 4 Minuten zentrifugiert. Das
Plasma wurde mit einer Pipette abgenommen, Plasma und Erythrozytenkonzentrat anschließend
bei -22 °C bis zur weiteren Bearbeitung eingefroren und gelagert.
36

Die Entnahme von Zwei Plazenten, der Amnionflüssigkeit und des maternalen Blutes erfolgte an
Tag 14, 16 und 20 der Trächtigkeitsperiode. Den Tieren wurden jeweils um 10 Uhr des
Entnahmetages 50 µg BrdU (5-Bromodeoxyuridin) pro 1 g Körpergewicht intraperitoneal
appliziert.
Bei BrdU handelt es sich um ein Thymidin-Basenanalogon, welches nach Einbau in die DNS
während der zellulären Synthesephase des Zellzyklus mit Hilfe von entsprechenden monoklonalen
Antikörpern nachgewiesen werden kann. Mit diesem von Gratzner (1982) entwickelten Verfahren,
besteht die Möglichkeit, Zellproliferationsereignisse zu quantifizieren. Die Probenentnahme fand
nach der BrdU Inkorporation jeweils um 16 Uhr, ca. 6h nach BrdU-Applikation statt.
Des Weiteren erfolgte die Punktion der Amnionhöhlen mehrerer Feten zur Gewinnung von
Amnionflüssigkeit als Sammelprobe eines Muttertieres. Die Amnionflüssigkeit wurde ebenfalls
abzentrifugiert und der Überstand bei -22 °C tiefgefroren.
Die Eröffnung des Uterus erfolgte paraplazentar, jedoch nicht antimesenterial, so dass ehemalige
Implantationsstellen nicht beschädigt wurden. Die Anzahl der Resorptionen wurden zunächst
makroskopisch ermittelt und die Feten wurden ausgezählt, es fand kein Vergleich zwischen den
Feten statt. Zum Nachweis aller Implantationsorte wurden die bei -22 °C tiefgefrorenen Uterie
zunächst bei Zimmertemperatur aufgetaut. Danach erfolgte eine 10 minütige Inkubation der Uteri
in einer 10%igen Ammoniumsulfidlösung, nach kurzem Spülen in Aqua dest. erfolgte eine 10
minütige Inkubation in einer Lösung aus 1%iger Salzsäure und einer 20%igen Kaliumferrocyanid-
Lösung zu gleichen Teilen. Nach erneutem spülen der Uteri in Aqua dest. erfolgte die
stereomikroskopische Auszählung aller Implantationsorte, welche sich bei dieser Methode nach
Salewski (1964) blauschwarz färben.
Jeweils eine Plazenta wurde entweder in einer 4% gepufferten Formaldehydlösung oder in einer
von Langeron modifizieren Bouin-Lösung über mindestens 24 Stunden fixiert, die Auswahl der
Plazenten zum Fixierungsverfahren geschah willkürlich (Romeis, 1968).
4% gepuffertes Formalin: 12 ml 37% Formaldehyd
88 ml PBS
Bouin-Lösung: 1 Teil 40%iges Formol
3 Teile destilliertes Wasser
Pikrinsäure im Überschuss
Zu dieser Lösung wurde unmittelbar vor Gebrauch Eisessig im Verhältnis 1:20 gegeben.
Pro Muttertier wurde eine Plazenta Formol- und eine Bouin fixiert. Die Bouin fixierten Plazenten
wurden mittels HE, PAS und PBA Färbemethode behandelt. Dies gilt ebenfalls für die Formol
fixierten Plazenten, hier erfolgte z.T. zusätzlich eine vorherige Diastaseinkubation.
37

Die fixierten Plazenten wurden zunächst in einer aufsteigenden Alkoholreihe nach folgendem
Protokoll entwässert, anschließend bei 60°C in Paraffin eingebettet und an einer Ausgießstation
ausgeblockt. Zum Erkalten des Blocks wurde dieser auf eine gekühlte Platte gestellt.
Reagenz Zeit (in min)
70% EtOH 30
80% EtOH 30
96% EtOH 30
96% EtOH 45
100% EtOH 30
100% EtOH 30
100% EtOH 45
Xylol 30
Xylol 45
Paraffin 30
Paraffin 30
Die 4µm dicken Paraffinschnitte wurden mittels eines Rotationsmikrotoms der Firma Leica
(Modell RM2255) hergestellt und zum Strecken auf ein 40°C heißes Wasserbad gelegt.
Anschließend wurden sie auf SuperFrost® Plus Objektträger der Firma R. Langenbrinck gezogen
und auf einer 40°C temperierten Wärmeplatte über mind. 30 min adheriert und getrocknet. Die
Lagerung der Paraffinblöcke erfolgte im Kühlschrank bei etwa 7°C, die Lagerung der
Paraffinschnitte bei Raumtemperatur.
Es folge für die histologische Beurteilung der Schnitte die Färbung der mit Formalin und Bouin
fixierten Präparate mit Hämatoxylin-Eosin (HE) nach Romeis (1968). Nach der Entparaffinierung
der Schnitte in zweimal 5 min Xylol, erfolgte die Rehydrierung der Präparate in zweimal 5 min
100% EtOH, 5 min 96% EtOH, 3 min 70% EtOH, 3 min 50% EtOH und mind. 3 min in
destilliertem Wasser. Anschließend wurden die Schnitte 3-5 Minuten in ein Hämalaun-Bad
überführt und nach gründlichem Waschen mit Wasser (10-15 min) für 5-15 min in einer 0,1%igen
wässrigen Eosinlösung gefärbt. Nach einem weiteren Waschschritt wurden die Schnitte in
80%igem EtOH differenziert, dehydriert und über Xylol als Zwischenmedium in Permanent
Mounting Medium eingedeckt.
Zum Nachweis von Polysacchariden, neutralen Mukopolysacchariden, Muko- und
Glykoproteinen, Glykolipiden sowie ungesättigten Fetten und Phospholipiden wurden sowohl die
mit Formalin als auch die mit Bouinlösung fixierten Präparate mittels Periodic-acid-Schiff-
38

reaction (PAS, PJS) nach McManus gefärbt (Romeis, 1968). Anschließend erfolgte eine
qualitative Auswertung der Schnitte. Hierzu erfolgte eine Rehydrierung der Schnitte nach
demselben Protokoll wie bei der HE-Färbung. Anschließend wurden die Präparate 5 min in eine
0,5%ige wässrige Perjodsäurelösung gestellt und danach mit Wasser gespült. Hiernach wurden sie
für 15 min in Fuchsinschwefliger Säure inkubiert und daraufhin in 3 Portionen SO2-Wasser
dreimal 2 min gewaschen. Nach erneutem Waschen in Wasser wurden die Zellkerne mit Hämalaun
für 20-30 Sekunden gegengefärbt und die Schnitte anschließend erneut gewaschen. Nach einer
Färbung des Zellplasmas mit Lichtgrün folgte die Dehydrierung der Schnitte in einer
aufsteigenden Alkoholreihe und das Eindecken der Präparate mittels eines Permanent Mounting
Mediums über Xylol.
Zur besseren Farbdifferenzierung der Glykoproteine und Mukopolysaccharide, vor allem von
Basallaminae in den fetalen Kapillaren wurden weitere, in Bouinlösung sowie Formalin fixierte
Schnitte nach der Perjodsäure-Bisulfit-Aldehydthionin-Färbemethode (PBA) (Specht, 1970)
gefärbt, bei welcher Substanzen mit einer α-D-Glykolgruppe blau erscheinen. Zum Ausschluss
von Glykogenanfärbung besteht die Möglichkeit einer zweistündigen Inkubation der Schnitte in
1%iger wässriger Diastaselösung.
Hierzu erfolgte die Deparaffinierung und Rehydrierung nach demselben Protokoll wie bei der HE-
Färbung und die Weiterbehandlung wie folgt:
1. Inkubation der entparaffinierten Schnitte in 1% wässriger Perjodsäure für 7 min
2. Spülen der Schnitte in destilliertem Wasser
3. Inkubation der Präparate in 10% wässriger Na-Pyrosulfitlösung für etwa 14 Stunden
4. 3x kurz in destilliertem Wasser spülen
5. Einstellen der Schnitte für 1-2 min in 70% EtOH
6. Färben mit Aldehydthionin für etwa 1,5 Stunden.
7. Spülen in 70% EtOH
8. Mehrfaches spülen mit destilliertem Wasser
9. Färben in Kernechtrubinlösung für 8 min
10. Spülen in destilliertem Wasser
11. Für etwa 5 min in 5%iger wässriger Phosphorwolframsäure differenzieren
12. Spülen in destilliertem Wasser
13. Entwässern, kurz in 96% EtOH, dann 3x in 100% EtOH, eindecken mittels eines permanenten
Mounting Mediums über Xylol.
39

Zusammensetzung Aldehydthionin:
0,5g Thionin (Firma Merck Nr. 1.15929.0025)
91,5 ml 70% EtOH
7,5 ml Paraldehyd (Firma Merck Nr. 8.18255.0100)
1,0 ml rauchende Salzsäure
Die Lösung ruhte für mind. 6 Tage in einer dunklen Flasche und wurde anschließend filtriert, ohne
den Bodensatz aufzuwirbeln.
Zusammensetzung Kernechtrubinlösung:
0,1g Kernechtrubin (Firma Niepötter)
100 ml destilliertes Wasser
1,0 ml Eisessig
Zusätzlich wurden Laminin, als Hauptbestandteil fetaler Blutgefäße in der Plazenta, aktivierte
Caspase-3 als Marker für apoptotische Vorgänge sowie Ki-67 als Proliferationsmarker
immunhistochemisch nachgewiesen. Der Nachweis von Laminin und Ki-67 erfolgte an jeweils 3
Plazenten pro Gruppe und Zeitpunkt, der Nachweis von Caspase-3 erfolgte an 4 Plazenten zu den
Zeitpunkten 16 und 20.
Der Nachweis des Glykoproteins Laminin erfolgte mittels Anti-Laminin Antikörper der Firma
Sigma (Nr. L 9393), die Protease Caspase-3 wurde mittels Anti-ACTIVE® Caspase-3 Antikörper
der Firma Promega (Catalog #G7481) nachgewiesen.
Das Protein Ki-67 wurde mithilfe des monoklonales Maus Anti-Ratte Ki-67 Antigen Antikörpers
der Firma DakoCytomation (Nr. M7248) nachgewiesen.
Der Nachweis der entsprechenden Antigene erfolgte mittels modifizierter ABC-Methode (avidin-
biotin conjugate method), wobei folgender unlösliche Komplex gebildet wird (Hsu et al., 1981).
40

Abb. 3.2.1: schematische Darstellung der ABC-Methode. A: Avidin; Ag: Antigen; B: Biotin; Px: Peroxidase; sA: Sekundärantikörper; *: entsprechender Primärantikörper. In Anlehnung an Taylor et al., 2006.
Anstelle des Avidins welches bei der ABC-Methode an den biotinylierten Sekundärantikörper
bindet, wurde Strepdavidin verwendet. Hierbei handelt es sich um ein tetrameres, nicht
glykolysiertes Avidin-Analogon, dessen Aminosäuresequenz zu etwa 33% homolog (Haugland
und You, 1998) ist und welches eine sehr hohe Affinität zu Biotin besitzt. Jedes Molekül
Streptavidin vermag wie Avidin vier Moleküle Biotin zu binden. Ein Vorteil von Streptavidin
besteht darin, dass sein isoelektrischer Punkt annähernd im neutralen pH Bereich liegt, während
der isoelektrische Punkt von Avidin bei etwa pH 10 liegt. Hierdurch können elektrostatische
Bindungen vermieden werden, welche ein hohes Hintergrundsignal verursachen können.
Ein weiterer Vorteil besteht darin, dass Streptavidin keine Carbonhydrate enthält, welche
unspezifisch and Lektin ähnlichen Substanzen zu binden vermögen und dadurch ebenfalls
Hintergrundsignale erzeugen können.
Des Weiteren ermöglicht die hohe Affinität von Streptavidin zu Biotin die lange Lagerung
gebrauchsfertiger Lösung (Streptavidin-Biotin-Peroxidase-Komplex) und vereinfacht somit die
Anwendung.
Die Farbentwicklung erfolgt indem der Peroxidase H2O2 als Substrat angeboten wird, welches
reduziert wird. Die frei werdenden Protonen oxidieren nun DAB (3,3´-Diaminobenzidin), welches
daraufhin Polymerisiert.
41

Hierbei entsteht ein brauner, lichtstabiler, in Alkohol und Xylol unlöslicher Komplex, der eine
Dehydrierung und ein Eindecken der Präparate in Harz erlaubt (Taylor et al., 2006).
Um einen besseren Kontrast zur anschließenden Gegenfärbung zu erzielen, wurde der Lösung
Nickel beigemischt, hierdurch entsteht ein grau-schwarzer Farbkomplex.
Im Folgenden werden die einzelnen Arbeitsschritte beschrieben.
Die Deparaffinierung und Rehydrierung erfolgte nach demselben Protokoll wie bei den HE
gefärbten Präparaten.
Anschließend erfolgte:
1. die Blockierung der endogenen Peroxidase mittels 3% H2O2 in Leitungswasser für 10 min,
danach wurden die Schnitte in PBS (Aqua dest., 0,14M NaCl, 0,0027M KCl, 0,01M Phosphat bei
PH 7,4) gewaschen.
2. eine Antigendemaskierung:
- die Präparate auf denen mit dem Ki-67-Antikörper gearbeitet wurde wurden hierzu für 30 min in
einen 100°C heißen 0,01M Citratpuffer gestellt.
- die Präparate auf denen mit dem Anti-ACTIVE® Caspase-3 Antikörper gearbeitet wurde wurden
für 10 min in 0,2% Triton in PBS inkubiert.
3. Blockierung von unspezifischen Bindungsstellen der Antikörper, hierzu wurden die Schnitte
inkubiert mit:
- 2,5% Pferdeserum mit 4 Tropfen Avidin-Blockierung (blocking Kit der Firma Vector
Laboratories Inc. (SP-2001)) pro ml Serum für 10 min, waschen in PBS
- 2,5% Pferdeserum mit 4 Tropfen Biotin-Blockierung (blocking Kit der Firma Vector Laboratories
Inc. (SP-2001)) pro ml Serum für 10 min, 2x waschen in PBS
4. Inkubation mit dem Primärantikörper für eine Stunde in einer feuchten Kammer mit den
Verdünnungen:
- Laminin: 1:30
- Caspase-3: 1:100
- Ki-67: 1:25, jeweils in 2,5% Pferdeserum
5. 2x 5 min waschen in PBS
6. Inkubation mit dem Sekundärantikörper des Färbe-Kits der Firma Vector Laboratories Inc.
(SP-2001) für 45 min in einer feuchten Kammer
7. 2x 5 min waschen in PBS
42

8. Inkubation mit dem Streptavidin-Peroxidase-Komplex (universal quick Kit der Firma Vector
Laboratories Inc. (SP-2001))
9. 2x 5 min waschen in PBS
10. Substratentwicklung mittels eines DAB-Kit (3,3´-Diaminobenzidin) der Firma Vector
Laboratories Inc. (SK-4100), die Zusammensetzung der Lösung besteht aus besteht aus:
- 5 ml Aqua dest.
- 2 Tropfen Puffer
- 4 Tropfen DAB
- 2 Tropfen H2O2
- 2 Tropfen Nickel-Lösung
11. Es erfolgte die Farbentwicklung, welche je nach Intensität nach 1-10 min in Leitungswasser
gestoppt wurde.
12. 5 min waschen in Leitungswasser
13. Gegenfärbung mit Nuclear fast red der Firma Vector Laboratories Inc. (H-3403) für 5 min
14. waschen in Leitungswasser während 10 min
15. Dehydrieren der Schnitte
- 3 mal kurz in 80% EtOH
- 3 mal kurz in 96% EtOH
- 3 min in 100% EtOH
- 5 min in 100% EtOH
- 2 mal 5 min in Xylol
Es erfolgte das Eindecken der Präparate mittels eines Xylol löslichen Permanent Mounting
Mediums.
43

Die Aminosäureanalytik im Blutplasma und der Amnionflüssigkeit (Bestimmung von Taurin,
Asparaginsäure, Hydroxyprolin, Threonin, Serin, Aspartat, Glutaminsäure, Glutamin, Prolin,
Glycin, Alanin, Citrullin, Butyrat, Valin, Cystein, Methionin, Cystathionin, Isoleucin, Leucin,
Tyrosin, Phenylalanin, Ornithin, Lysin, Histidin und Arginin) erfolgte mittels
Ionenaustauschchromatographie und anschließender Detektion der Aminosäuren mittels
Ninhydrin. Das Homocystein und Vitamin B6 in Form von Pyridoxal-Phosphat wurden mittels
HPLC (high pressure liquid chromatograph) bestimmt. Die Folsäure wurde mittels
Chemilumineszenz ermittelt. Die Enzymaktivität von ALAT und ASAT wurden mit der von der
International Federation of Clinical Chemistry (IFCC) empfohlenen Methode nach Warburg
bestimmt. Die Bestimmung von Folsäure, Vitamin B6, Homocystein, ALAT und ASAT erfolgte
durch das Institut für Klinische Chemie und Laboratoriumsmedizin der Universität Rostock. Die
Aminosäureanalytik wurde durch das Institut für Mikrobiologie Abteilung für Genetik und
Biochemie der Ernst-Moritz-Arndt Universität Greifswald durchgeführt.
Die statistische Auswertung der Daten erfolgte mit dem Programm SPSS Statistics. Zunächst
wurde eine deskriptive Statistik erstellt, wobei Mittelwerte und Standartabweichungen (jeweils
pro Gruppe und Zeitpunkt) berechnet wurden. Signifikante Unterschiede zwischen den Gruppen
wurde mit Hilfe eines Post-Hoc-Tests errechnet.
44

4. Ergebnisse
4.1 Vitamin B6, Folsäure, Homocystein, ALAT, ASAT im maternalen Plasma
4.2 Proteinogene Aminosäuren mit apolaren Seitenketten
4.3 Proteinogene Aminosäuren mit ungeladenen polaren Seitenketten
4.4 Proteinogene Aminosäuren mit sauren Seitenketten
4.5 Proteinogene Aminosäuren mit basischen Seitenketten
4.6 Nicht proteinogene Aminosäuren
4.7 Histomorphologische Untersuchung der Plazenten
4.8 Immunhistochemische Untersuchung der Plazenten
4.9 Resorptionen
Zur besseren Übersicht wird die Kontrollgruppe im Folgenden als „Gruppe K“ bezeichnet, die
Gruppe mit Vitamin-B6-Restriktion als „Gruppe B“ und die Gruppe mit Folsäure-Restriktion als
„Gruppe F“. Wenn nicht anders beschrieben, beziehen sich die Angaben auf die
Blutplasmakonzentrationen der Tiere. Die Irrtumswahrscheinlichkeit signifikanter Unterschiede
beträgt p≤ 0,05, auf abweichende Irrtumswahrscheinlichkeiten wird im Text hingewiesen.
4.1 Vitamin B6, Folsäure, Homocystein, ALAT, ASAT
im maternalen Plasma
Vitamin B6
In der Gruppe K blieb die maternale Vitamin-B6-Konzentration zwischen dem 14. und dem 20.
Tag weitestgehend konstant. In der Gruppe B ist bereits am 14. Trächtigkeitstag eine signifikant
verringerte Vitamin-B6-Konzentration im Vergleich zu Gruppe K und F zu beobachten, welche bis
zum 20. Trächtigkeitstag wesentlich niedriger ist als in der Gruppe K und sich von der Gruppe K
signifikant unterschied.
45

Vitamin B6
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
Abb. 4.1.1: Maternale Vitamin-B6-Konzentration im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
In der Gruppe F ist im Vergleich zu Gruppe K ab dem 16. Trächtigkeitstag ein Abfall der Vitamin-
B6-Konzentration zu beobachten, die Konzentration am 20. Tag unterscheidet sich jedoch nicht
signifikant von Gruppe K.
Zwischen dem 14. und 16. Trächtigkeitstag ist die Vitaminkonzentration in Gruppe B signifikant
geringer als in Gruppe F, am 20. Trächtigkeitstag nähern sich die Konzentrationen durch den
starken Vitamin-B6-Abfall in Gruppe F an, so dass sich für dem 20. Tag kein signifikanter
Unterschied darstellt.
Folsäure im Blutplasma
Die Folsäurekonzentration im Plasma der Gruppe K ist über die gesamte Trächtigkeit weitgehend
konstant, lediglich zwischen dem 16. und 20. Tag kommt es zu einem leichten Abfall der
Konzentration. Zwischen dem 14. und 20. Trächtigkeitstag sind keine signifikanten Unterschiede
zwischen den Gruppen K und B bezüglich der Folsäurekonzentration zu beobachten. Der
beschriebene Konzentrationsabfall in Gruppe K ist in Gruppe B jedoch nicht zu beobachten.
Abb. 4.1.2: Maternale Folsäurekonzentration des Plasmas im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Folsäure
0
20
40
60
80
100
120
140
160
180
14 16 18 20
Tag
µm
ol/
l K
B
F
46

Die Tiere der Gruppe F weisen zu jedem Zeitpunkt der Messung eine signifikant verringerte
Folsäurekonzentration als die der Gruppen K und B auf. Zwischen dem 14. und 16. Trächtigkeitstag
ist kein weiterer Abfall der Konzentration zu beobachten. Am 20. Tag ist es in Gruppe F zu einem
weiteren Abfall der Folsäurekonzentration gekommen.
erythrozytäre Folsäure
Die Folsäurekonzentration im Erythrozytenkonzentrat ist in allen Gruppen weitgehend konstant,
mit Ausnahme der Gruppe F, die am 16. Trächtigkeitstag eine signifikant geringere erythrozytäre
Folsäurekonzentrationen gegenüber der Gruppe B aufweist.
Abb. 4.1.3: Maternale erythrozytäre Folsäurekonzentration im zeitlichen Verlauf.
Der erythrozytäre Folsäuregehalt in Gruppe F ist im gesamten Versuchszeitraum gegenüber den
Gruppen B und F leicht verringert, es konnten jedoch keine weiteren signifikanten Unterschiede
errechnet werden.
Homocystein
Die Homocysteinkonzentration in Gruppe K sinkt zwischen dem 14. und 16. Trächtigkeitstag und
erreicht am 16. Tag einen Tiefpunkt. Bis zum 20. Trächtigkeitstag erfolgt ein mäßiger Anstieg.
Die Homocysteinkonzentration der Gruppe B ist im Vergleich zu Gruppe K am 14. und 16.
Trächtigkeitstag signifikant erhöht und unterscheidet sich am 20. Tag mit einer Signifikanz von
p≤0,06. In Gruppe F ist die Homocysteinkonzentration am 14. und 16. Trächtigkeitstag
47
Standartabweichungen siehe Tab. 10.1 im Datenanhang
erythrozytäre Folsäure
0
100
200
300
400
500
600
700
800
14 16 18 20
Tag
µm
ol/
l K
B
F

im Vergleich zu Gruppe K signifikant erhöht, am 20. Tag können keine signifikanten Unterschiede
für Gruppe F errechnet werden. Zwischen dem 16. und dem 20. Tag kommt es in den Gruppen B
und F zu einem Abfall der Homocysteinkonzentration.
ALAT
Zwischen dem 14. und 16. Trächigkeitstag kommt es zu einer Zunahme der Aktivität des Enzyms
Alanin-Aminotransferase (ALAT) in Gruppe K.
Die Enzymaktivität in Gruppe B ist in Laufe der Trächtigkeit weitestgehend konstant. Die
Aktivität der ALAT in Gruppe B ist im Vergleich zu Gruppe K über den gesamten
Trächtigkeitsverlauf signifikant verringert.
Abb.: 4.1.5: Aktivität der Alanin-Aminotransferase im maternalen Serum im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Auch in Gruppe F bleibt die Enzymaktivität der ALAT über den gesamten Trächtigkeitszeitraum
weitgehend konstant. Am 20. Trächtigkeitstag ist die Enzymaktivität in Gruppe F gegenüber
Gruppe K signifikant verringert.
ALAT
0
10
20
30
40
50
60
70
14 16 18 20
Tag
U
K
B
F
Abb.: 4.1.4: Maternale Homocysteinkonzentration im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Homocystein
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
14 16 18 20
Tag
µm
ol/
l K
B
F
48

Im Vergleich zu Gruppe B ist die Enzymaktivität in Gruppe F an den Tagen 14 und 20 der
Trächtigkeit signifikant erhöht.
ASAT
Die Aktivität des Enzyms Aspartat-Aminotransferase (ASAT) steigt in Gruppe K zwischen dem
14. Trächtigkeitstag und 16. Trächtigkeitstag an. Die Enzymaktivität in Gruppe B ist zwischen
dem 14. und 20. Trächtigkeitstag relativ konstant. Am 16. Trächtigkeitstag ist die Aktivität der
Gruppe B gegenüber der Gruppe K signifikant verringert.
Zwischen dem 14. und 20. Trächtigkeit sinkt die Enzymaktivität in Gruppe F leicht ab und zeigt
im Vergleich zu Gruppe K am 20. Trächtigkeitstag eine signifikant verringerte Enzymaktivität.
Am 14. und 16. Tag ist die Aktivität der ASAT in Gruppe B gegenüber der Aktivität in Gruppe F
signifikant erniedrigt.
4.2 Proteinogene Aminosäure mit apolaren Seitenketten
Zur Bestimmung des diaplazentaren Aminosäuretransports, wurden die entsprechenden
Konzentrationen im maternalen Plasmas mit denen der Amnionflüssigkeit verglichen. Hierzu
wurde der Quotient aus der Aminosäurekonzentration der Amnionflüssigkeit und der des
maternalen Blutplasmas bestimmt. Wird dieser Quotient gegen die Zeit aufgetragen, ergibt sich
ein Graph, der den diaplazentaren Aminosäuretransport im Verlauf der Trächtigkeit widerspiegelt.
Sowohl die Werte des maternalen Blutplasmas, die der Amnionflüssigkeit als auch ihre Quotienten
sind im Anhang zu finden.
Abb. 4.1.6: Aktivität der Aspartat-Aminotransferase im maternalen Plasma. Standartabweichungen siehe Tab. 10.1 im Datenanhang
ASAT
0
20
40
60
80
100
120
140
14 16 18 20
Tag
U
K
B
F
49
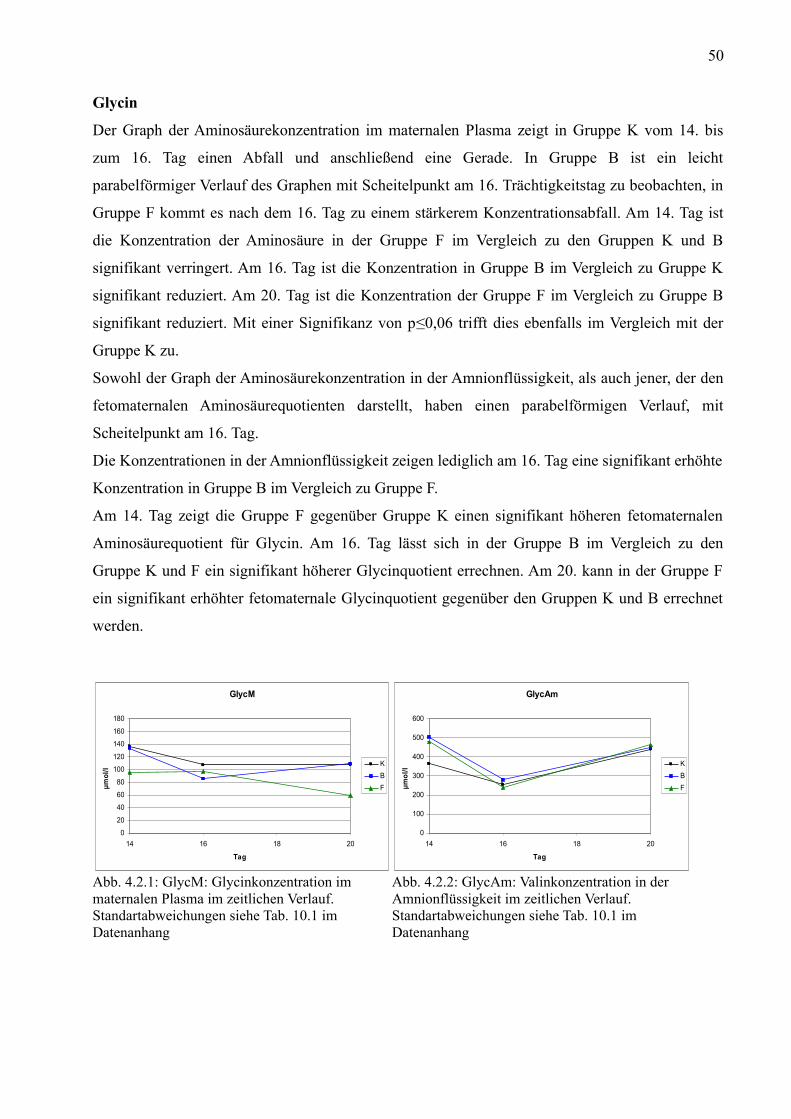
Glycin
Der Graph der Aminosäurekonzentration im maternalen Plasma zeigt in Gruppe K vom 14. bis
zum 16. Tag einen Abfall und anschließend eine Gerade. In Gruppe B ist ein leicht
parabelförmiger Verlauf des Graphen mit Scheitelpunkt am 16. Trächtigkeitstag zu beobachten, in
Gruppe F kommt es nach dem 16. Tag zu einem stärkerem Konzentrationsabfall. Am 14. Tag ist
die Konzentration der Aminosäure in der Gruppe F im Vergleich zu den Gruppen K und B
signifikant verringert. Am 16. Tag ist die Konzentration in Gruppe B im Vergleich zu Gruppe K
signifikant reduziert. Am 20. Tag ist die Konzentration der Gruppe F im Vergleich zu Gruppe B
signifikant reduziert. Mit einer Signifikanz von p≤0,06 trifft dies ebenfalls im Vergleich mit der
Gruppe K zu.
Sowohl der Graph der Aminosäurekonzentration in der Amnionflüssigkeit, als auch jener, der den
fetomaternalen Aminosäurequotienten darstellt, haben einen parabelförmigen Verlauf, mit
Scheitelpunkt am 16. Tag.
Die Konzentrationen in der Amnionflüssigkeit zeigen lediglich am 16. Tag eine signifikant erhöhte
Konzentration in Gruppe B im Vergleich zu Gruppe F.
Am 14. Tag zeigt die Gruppe F gegenüber Gruppe K einen signifikant höheren fetomaternalen
Aminosäurequotient für Glycin. Am 16. Tag lässt sich in der Gruppe B im Vergleich zu den
Gruppe K und F ein signifikant höherer Glycinquotient errechnen. Am 20. kann in der Gruppe F
ein signifikant erhöhter fetomaternale Glycinquotient gegenüber den Gruppen K und B errechnet
werden.
Abb. 4.2.1: GlycM: Glycinkonzentration im maternalen Plasma im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.2: GlycAm: Valinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
50
GlycM
0
20
40
60
80
100
120
140
160
180
14 16 18 20
Tag
µm
ol/
l K
B
F
GlycAm
0
100
200
300
400
500
600
14 16 18 20
Tag
µm
ol/
l K
B
F

Alanin
Am 14. Tag ist die Alaninkonzentration im maternalen Plasma in Gruppe B gegenüber Gruppe K
signifikant reduziert. Am 16. Tag ist die Konzentration der Gruppe F signifikant höher als in den
Gruppen K und B.
In der Amnionflüssigkeit konnte lediglich am 20. Tag eine signifikante Erhöhung der Alanin-
konzentration mit einer Irrtumswahrscheinlichkeit von p≤0,06 für Gruppe F gegenüber Gruppe K
errechnet werden.
Ebenfalls mit einer Irrtumswahrscheinlichkeit von p≤0,06 konnte am 14. Tag in Gruppe B
gegenüber Gruppe K ein erhöhter fetomaternale Alaninquotient ermittelt werden. Am 16. Tag ist
der Quotient der Gruppe B gegenüber Gruppe F signifikant erhöht.
Abb. 4.2.4: AlaM: Alaninkonzentration im maternalen Plasma im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.5: AlaAm: Alaninkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.3: GlycQ: fetomaternaler Glycinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
GlycQ
0
1
2
3
4
5
6
7
8
9
14 16 18 20
Tag
K
B
F
51
AlaM
0
100
200
300
400
500
600
14 16 18 20
Tag
µm
ol/
l K
B
F
AlaAm
0
200
400
600
800
1000
1200
1400
1600
1800
2000
14 16 18 20
Tag
µm
ol/
l K
B
F

Valin
Bei der Aminosäure Valin ist im maternalen Plasma der Gruppe F am 14. Tag eine signifikant
höhere Konzentration zu messen, als in den Gruppe K und B. Am 16. Tag ist die Konzentration in
der Gruppe F immer noch signifikant höher als in Gruppe K.
In der Gruppe B ist die Valinkonzentration in der Amnionflüssigkeit am 16. Tag signifikant höher
als in Gruppe K. Am 20. Tag ist die Konzentration in Gruppe F gegenüber der in Gruppe K
signifikant erhöht.
Am 14. und 16. Tag ist der fetomaternale Quotient in Gruppe B signifikant größer als in Gruppe F.
Die Graphen aller Quotienten zeigen einen parabelförmigen Verlauf, mit Scheitelpunkt am 16.
Tag.
Abb. 4.2.7: ValM: Valinkonzentration im maternalen Plasma im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.8: ValAm: Valinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.6: AlaQ: fetomaternale Alaninquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
AlaQ
0
1
2
3
4
5
6
14 16 18 20
Tag
K
B
F
52
ValM
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
ValAm
0
200
400
600
800
1000
1200
1400
14 16 18 20
Tag
µm
ol/
l K
B
F

Leucin
Während sich bei der Konzentration der Aminosäure Leucin im maternalen Plasma und in der
Amnionflüssigkeit keine signifikanten Unterschiede zwischen den Gruppen feststellen lassen, ist
der fetomaternale Quotient am 16. Tag in Gruppe F signifikant geringer als in Gruppe B. Es
können keine signifikanten Unterschiede zur Gruppe K errechnet werden.
Isoleucin
Ähnliches ist bei der Aminosäure Isoleucin zu beobachten. Auch hier sind im maternalen Plasma
und der Amnionflüssigkeit keine signifikanten Unterschiede zwischen den Gruppen zu erkennen.
Am 16. Tag ist der Quotient der Gruppe F gegenüber den Gruppen K und B signifikant geringer.
Methionin
Die Methioninkonzentration im maternalen Plasma aller Gruppen zeigt einen relativ homogenen
Verlauf. Am 14. und 16. Trächtigkeitstag kann im Plasma der Gruppe F eine signifikant höhere
Konzentration als in Gruppe K nachgewiesen werden. Am 20. Trächtigkeitstag sind keine
Unterschiede zwischen den Gruppen K, B und F nachweisbar.
Abb. 4.2.9: ValQ: fetomaternaler Valinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
ValQ
0
1
2
3
4
5
6
7
8
14 16 18 20
Tag
K
B
F
53

Abb. 4.2.10: MethM: Methioninkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.11: MethAm: Methioninkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.12: MethQ: fetomaternaler Quotient der Methioninkonzentration im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Auch in der Amnionflüssigkeit sind keine signifikanten Unterschiede zwischen den Gruppen zu
beobachten, der Graph ist parabelförmig mit Scheitelpunkt am 16. Tag.
Der fetomaternale Aminosäurequotient ist lediglich am 16. Trächtigkeitstag in Gruppe F
signifikant kleiner als in den Gruppen K und B. Auch hier stellt sich der Graph parabelförmig dar
und besitzt seinen Scheitelpunkt am 16. Trächtigkeitstag.
Prolin
Mit einer Irrtumswahrscheinlichkeit von p≤0,06 ist die Prolinkonzentration im maternalen Plasma
am 14. Tag in der Gruppe B signifikant geringer als in der Gruppe F. Die Werte der Gruppe K
liegen zwischen denen der Gruppen B und F.
Am 16. Tag ist die Prolinkonzentration in der Amnionflüssigkeit in Gruppe B signifikant höher als
in Gruppe F. Am 20. Tag sind die Konzentrationen in der Amnionflüssigkeit in den Gruppen B und
MethM
0
10
20
30
40
50
60
14 16 18 20
Tag
µm
ol/
l K
B
F
MethAm
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
MethQ
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
14 16 18 20
Tag
K
B
F
54

F signifikant höher als in Gruppe K, Gruppe F weist gegenüber Gruppe B ebenfalls eine
signifikant höhere Prolinkonzentration auf.
Der fetomaternale Quotient ist am 14. Tag in Gruppe B signifikant höher als in Gruppe F. Am 20.
Tag ist der Quotient der Gruppe F signifikant höher als der in Gruppe K.
Abb. 4.2.13: ProlM: Maternale Prolinkonzentration im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.2.14: ProlAm: Prolinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
4.3 Proteinogene Aminosäuren mit ungeladenen polaren Seitenketten
Cystein
Die Konzentration der Aminosäure Cystein im maternalen Blutplasma steigt in der Gruppe K im
Verlauf der Trächtigkeit leicht an, in Gruppe B ist eine weitgehend konstante Konzentration zu
beobachten, während sich der Graph in Gruppe F negativ parabelförmig mit Scheitelpunkt am 16.
Abb. 4.2.15: ProlQ: fetomaternaler Quotient der Prolinkonzentration im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
ProlQ
0
1
2
3
4
5
6
7
8
9
14 16 18 20
Tag
K
B
F
ProlM
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
ProlAm
0
200
400
600
800
1000
1200
14 16 18 20
Tag
µm
ol/
l K
B
F
55

Tag darstellt. Signifikante Unterschiede zwischen den Gruppen sind über den gesamten
Trächtigkeitszeitraum, abgesehen von der Gruppe F, welche am 16. Tag eine signifikant höhere
Konzentration aufweist als Gruppe B, nicht zu beobachten. In Gruppe F ist am 16. Tag eine
überproportional hohe Konzentration von Cystein zu beobachten.
In der Amnionflüssigkeit aller Gruppen ist vom 14. bis zum 20. Tag kein signifikanter Unterschied
der Cysteinwerte zu beobachten. Die Cysteinkonzentration steigt in allen Gruppen zwischen dem
14. und 20. Tag signifikant an.
Aufgrund der geringen maternalen und der hohen fetalen Cysteinkonzentrationen ab dem 20. Tag,
steigt der fetomaternale Quotient ab dem 16. Tag in allen Gruppen signifikant an.
Abb. 4.3.1: CystM: Maternale Cysteinkonzentration im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.2: CystAm: Konzentration von Cystein in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.3: CystQ: Fetomaternaler Quotienten der Cysteinkonzentration im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
CystM
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
14 16 18 20
Tag
µm
ol/l
K
B
F
CystAm
0
20
40
60
80
100
120
140
160
180
200
14 16 18 20
Tag
µm
ol/l
K
B
F
CystQ
0
20
40
60
80
100
120
140
160
180
200
14 16 18 20
Tag
K
B
F
56

Tyrosin
Bei der Aminosäure Tyrosin zeigen sich über den gesamten Trächtigkeitszeitraum in der
Kontrollgruppe niedrigere maternale Plasmakonzentrationen als in den Gruppen B und F. Im
Gegensatz zu diesen Gruppen in denen die Tyrosinkonzentration zwischen dem 16. und 20. Tag
weitgehend konstant bleiben, fällt die Tyrosinkonzentration im maternalen Plasma in Gruppe K
über zwischen dem 16. und 20. Tag weiter ab. In der Gruppe B sind die Werte gegenüber der
Gruppe K an den Tagen 14 und 20 signifikant erhöht, in der Gruppe F sind signifikant erhöhte
Werte gegenüber Gruppe K an den Tagen 14 und 16 zu beobachten. Signifikante Unterschiede
zwischen den Gruppen B und F bestehen nicht.
Sowohl der Graph der Amnionflüssigkeit als auch der des fetomaternalen Quotienten stellen sich
parabelförmig mit Scheitelpunkt am 16. Tag dar, wobei der rechte Schenkel der Quotientenkurve
am 20. Tag höher ist als am 14. Tag.
Die Tyrosinkonzentration in der Amnionflüssigkeit der Gruppe B ist an den Tagen 14, 16 und 20
gegenüber der Gruppe K signifikant erhöht, an Tag 14 ist ebenfalls eine signifikante Erhöhung
gegenüber Gruppe F zu beobachten. Am 20. Tag ist in Gruppe F eine signifikant höhere
Aminosäurekonzentration zu ermitteln, als in Gruppe K.
Am 16. Tag ist der Aminosäurequotient der Gruppe F signifikant geringer als in Gruppe B.
Weitere Unterschiede konnten nicht errechnet werden.
Abb. 4.3.4: TyrM: Tyrosinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.5: TyrAm: Tyrosinkonzentration in der Amnionflüssigkeit in Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
57
TyrM
0
10
20
30
40
50
60
70
80
90
14 16 18 20
Tag
µm
ol/l
K
B
F
TyrAm
0
50
100
150
200
250
300
350
400
450
14 16 18 20
Tag
µm
ol/
l K
B
F

Abb. 4.3.6: TyrQ: fetomaternaler Tyrosinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Serin
Die Aminosäure Serin ist am 16. Tag im maternalen Plasma der Gruppe F gegenüber den Gruppen
K und B signifikant erhöht. Abgesehen von diesem Unterschied zeigen die Graphen einen recht
homogenen Verlauf.
Die Graphen der Amnionflüssigkeit zeigen einen parabelförmigen Verlauf mit Scheitelpunkt am
16. Tag. Während in der Amnionflüssigkeit keine signifikanten Unterschiede ermittelt werden
können, ist der Quotienten der Gruppe F des 16. Tages gegenüber der Gruppen K und B
signifikant reduziert.
Abb. 4.3.7: SerM: Serinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.8: SerAm: Serinkonzentration in der Amnionflüssigkeit in Verlauf der TrächtigkeitStandartabweichungen siehe Tab. 10.1 im Datenanhang.
58
TyrQ
0123456789
14 16 18 20
Tag
K
B
F
SerM
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
SerAm
0
100
200
300
400
500
600
700
14 16 18 20
Tag
µm
ol/l K
B
F

Abb. 4.3.9: SerQ: fetomaternaler Serinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Threonin
Die Konzentration der Aminosäure Threonin im maternalen Plasma aller drei Gruppen stellt sich
zwischen dem 14. und 20. Trächtigkeitstag als recht konstant dar, so dass die Graphen annähernd
waagerecht verlaufen. Der Graph der Gruppe K zeigt hierbei eine niedrigere Plasmakonzentration
an, als die Graphen der Gruppe B und F. Diese Unterschiede sind zwischen den 14. und 20. Tag
signifikant.
Die Graphen der Threoninkonzentration in der Amnionflüssigkeit sowie die der fetomaternalen
Quotienten zeigen einen parabelförmigen Verlauf mit Scheitelpunkt am 16. Tag. Sowohl die
Konzentration in der Amnionflüssigkeit als auch die Quotienten sind am 20. Tag größer als am 14.
Tag.
Zwischen dem 14. und 20. Tag ist die Threoninkonzentration in der Amnionflüssigkeit in den
Gruppen B und F signifikant höher als in Gruppe K. Am 16. Tag ist die Threoninkonzentration in
Gruppe B signifikant höher als in Gruppe F.
Am 14. Tag sind die Quotienten der Gruppen B und F signifikant kleiner als der Quotient der
Gruppe K. Am 16. Tag ist der fetomaternale Threoninquotient in Gruppe F signifikant kleiner als
in Gruppe K.
59
SerQ
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
14 16 18 20
Tag
K
B
F

Abb. 4.3.10: ThreoM: Threoninkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.11: ThreoAm: Threoninkonzentration in der Amnionflüssigkeit im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.3.12: ThreoQ: Fetomaternaler Threoninquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
4.4 Proteinogene Aminosäuren mit sauren Seitenketten
Asparaginsäure
Am 16. Tag ist im maternalen Plasma der Tiere der Gruppe F eine gegenüber den Gruppen K und
B signifikante Erhöhung der Asparaginsäurekonzentration zu messen. In der Amnionflüssigkeit
lassen sich keine signifikanten Unterschiede zwischen den Gruppen nachweisen.
Glutaminsäure
Die Glutaminsäurekonzentration im maternalen Plasma der Gruppe K ist während der gesamten
Tragzeit annähern konstant geblieben, während die Graphen der Gruppen B und F ein Maximum
am 16. Tag zeigen und sich an diesem Tag alle Gruppen signifikant von einander unterscheiden.
Dabei zeigt die Gruppe F den höchsten und die Gruppe K den niedrigsten Wert.
ThreoQ
0
0,5
1
1,5
2
2,5
3
3,5
14 16 18 20
Tag
K
B
F
60
ThreoM
0
50
100
150
200
250
300
350
400
14 16 18 20
Tag
µm
ol/
l K
B
F
ThreoAm
0
100
200
300
400
500
600
700
800
900
14 16 18 20
Tag
µm
ol/l K
B
F

Die Glutaminsäurekonzentration in der Amnionflüssigkeit nimmt vom 14. bis zum 20. Tag in allen
Gruppen deutlich ab. Besonders ausgeprägt ist dies in den Gruppen B und F zu beobachten, in
denen die Konzentration am 14. Tag über den in Gruppe K liegt. Es können jedoch über den
gesamten Trächtigkeitszeitraum keine signifikanten Unterschiede zwischen den Gruppen errechnet
werden.
Die Graphen der Aminosäurequotienten erinnern an jene der fetalen Glutaminsäurekonzentration,
zeigen im Gegensatz zu diesen jedoch signifikante Unterschiede. So ist der Aminosäurequotient
der Gruppe F gegenüber der Gruppe K am 16. Tag signifikant verringert. Mit einer
Irrtumswahrscheinlichkeit vom p≤0,06 ist der Aminosäurequotient der Gruppe F signifikant
geringer als der Quotient in Gruppe B.
Nachdem die Werte – vor allem zwischen dem 14. und 16. Tag deutlich fallen, und am 16. Tag in
den Gruppen K und B um 1 sind, während der Quotient in Gruppe F deutlich kleiner 1 ist, sind am
20. Tag alle Quotienten kleiner 1.
Abb. 4.4.1: GlutsM: Glutaminsäurekonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.4.2: GlutsAm: Glutaminsäurekonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.4.3: GlutsQ: Fetomaternaler Glutaminsäurequotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
GlutsM
0
50
100
150
200
250
300
14 16 18 20
Tag
µm
ol/
l K
B
F
GlutsAm
0
100
200
300
400
500
600
700
14 16 18 20
Tag
µm
ol/
l K
B
F
GlutsQ
0
1
2
3
4
5
6
14 16 18 20
Tag
µm
ol/
l K
B
F
61

4.5 Proteinogene Aminosäuren mit basischen Seitenketten
Lysin
Die Lysinkonzentration aller Gruppen sowohl im maternalen Blutplasma als auch in der
Amnionflüssigkeit bleiben annähernd konstant. Am 14. und 16. Tag zeigen sich in den Gruppen B
und F signifikant höhere maternale Plasmakonzentrationen als in der Gruppe K.
In der Amnionflüssigkeit können signifikant höhere Konzentrationen in den Gruppen B und F im
Vergleich zu Gruppe K an den Tagen 16 und 20 nachgewiesen werden. Am 14. Tag kann in
Gruppe B ein signifikant höhere Wert als in Gruppe K errechnet werden, in der Gruppe F kann an
diesem Tag mit einer Irrtumswahrscheinlichkeit von p≤0,06 ebenfalls ein erhöhter Wert gegenüber
Gruppe K ermittelt werden.
Am 14. Tag ist der fetomaternale Lysinquotient in Gruppe F signifikant kleiner als in Gruppe B.
Abb. 4.5.1: LysM: Lysinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.2: LysAm: Lysinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.3: LysQ: Fetomaternaler Lysinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
LysM
0
100
200
300
400
500
600
700
800
14 16 18 20
Tag
µm
ol/
l K
B
F
LysAm
0
500
1000
1500
2000
14 16 18 20
Tag
µmol
/l
K
B
F
LysQ
0
0,5
1
1,5
2
2,5
3
3,5
14 16 18 20
Tag
µm
ol/
l K
B
F
62

Arginin
Nachdem die Graphen des maternalen Blutplasmas der verschiedenen Gruppen anfänglich
unterschiedliche Verlaufsrichtungen zeigen, nähern sie sich am 16. Tag, und zeigen am 20. Tag
kaum noch Unterschiede. Am 14. Tag ist die Argininkonzentration im maternalen Blutplasma der
Gruppe B signifikant geringer als in der Gruppe K. In der Gruppe F ist die Plasmakonzentration
am 16. Tag signifikant höher als in den Gruppen K und B.
Abb. 4.5.4: ArgM: Argininkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.5: ArgAm: Argininkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.6: ArgQ: Fetomaternaler Argininquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
In der Amnionflüsigkeit der Gruppen B und F können am 14. Tag signifikant geringere
Konzentrationen ermittlet werden, als in Gruppe K.
Während die Graphen der Argininkonzentration in der Amnionflüssigkeit sowie die des
fetomaternalen Argininquotienten zwischen dem 14. und 16. Tag unterschiedliche
Verlaufsrichtungen zeigen, steigen alle ab dem 16. Tag an.
ArgM
0
20
40
60
80
100
120
140
160
14 16 18 20
Tag
µmol
/l
K
F
B
ArgAm
0
50
100
150
200
250
300
350
400
14 16 18 20
Tag
µm
ol/
l K
B
F
ArgQ
0
0,5
1
1,5
2
2,5
3
3,5
4
14 16 18 20
Tag
K
B
F
63

Am 16. Tag kann gegenüber der Gruppe K in Gruppe F ein signifikant verringerter fetomaternaler
Argininquotient errechnet werden.
Histidin
Die Konzentration der Aminosäure Histidin im maternalen Plasma zeigt lediglich am 16. Tag eine
signifikante Reduktion der Konzentration in Gruppe B gegenüber Gruppe F. Die Konzentration
der Gruppe K liegt zwischen denen der Gruppe B und F, zeigt jedoch zu diesen keine signifikanten
Unterschiede.
In der Amnionflüssigkeit ist die Konzentration am 14. Tag in Gruppe F signifikant geringer als in
den Gruppen K und B.
Der fetomaternale Quotient der Gruppe F ist am 14. und 16. Tag signifikant geringer als die der
Gruppen K und B. Sowohl die Graphen der Amnionflüssigkeit als auch die der fetomaternalen
Quotienten zeigen einen parabelförmigen Verlauf mit Scheitelpunkt am 16. Trächtigkeitstag.
Abb. 4.5.7: HisM: Histidinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.8: HisAm: Histidinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.5.9: HisQ: Fetomaternaler Histidinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
HisM
0
10
20
30
40
50
60
70
14 16 18 20
Tag
µmo
l/l
K
B
F
HisAm
0
100
200
300
400
500
600
14 16 18 20
Tag
µm
ol/
l K
B
F
HisQ
0
2
4
6
8
10
14 16 18 20
Tag
K
B
F
64

4.6 Nicht proteinogene Aminosäuren
Butyrat
Bei der Aminosäure Butyrat ist im zeitlichen Verlauf in allen Gruppen ein deutlicher Anstieg der
Konzentration sowohl im maternalen Plasma als auch in der Amnionflüssigkeit nachzuweisen. Am
14. und 20. Trächtigkeitstag sind die Butyratkonzentrationen des maternalen Plasmas der Gruppen
B und F gegenüber Gruppe K signifikant erhöht.
In der Amnionflüssigkeit kann am 20. Trächtigkeitstag in den Gruppen B und F ebenfalls eine
signifikant höhere Konzentration im Vergleich zur Kontrollgruppe nachgewiesen werden. Bereits
am 14. Trächtigkeitstag kann dies in der Gruppe B beobachtet werden.
Im zeitlichen Verlauf zeigen die Konzentrationsquotienten einen leichten parabelförmigen Verlauf
mit Scheitelpunkt am 16. Trächtigkeitstag.
Abb. 4.6.1: ButyM: Butyratkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang.
Abb. 4.6.2: ButyAm: Butyratkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.6.3: ButyQ: Fetomaternaler Butyratquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
65
ButyM
0
5
10
15
20
25
30
35
40
45
14 16 18 20
Tag
µm
ol/
l K
B
F
ButyAm
0
10
20
30
40
50
60
70
14 16 18 20
Tag
µmol
/l
K
B
F
ButyQ
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
14 16 18 20
Tag
K
B
F

Ornithin
Während es im maternalen Plasma der Gruppe K zu einem sukzessiven Anstieg der
Ornithinkonzentration vom 14. zum 20. Tag kommt, zeigen die Graphen der Gruppen B und F
einen waagerechten Verlauf. Ab dem 16. Trächtigkeitstag kann im maternalen Plasma in den
Gruppen B und F eine signifikant geringere Ornithinkonzentration nachgewiesen werden, als in
Gruppe K.
In der Amnionflüssigkeit kann an den Tagen 14 und 16 in der Gruppe F eine signifikant geringere
Ornithinkonzentration als in der Gruppe K bestimmt werden. An Tag 16 ist die Konzentration in
Gruppe B ebenfalls signifikant verringert. Zwischen dem 14. und dem 16. Trächtigkeitstag nimmt
der fetomaternale Ornithinquotient in allen Gruppen deutlich ab wird jedoch nicht kleiner 2.
Am 20. Trächtigkeitstag ist in den Gruppen B und F ein signifikant erhöhter Ornithinquotient
gegen Gruppe K zu errechnen.
Abb. 4.6.4: OrnM: Ornithinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.6.5: OrnAm: Ornithinkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf.Standartabweichungen siehe Tab. 10.1 im Datenanhang
66
OrnM
0
10
20
30
40
50
60
14 16 18 20
Tag
µmo
l/l
K
B
F
OrnAm
0
50
100
150
200
250
300
350
14 16 18 20
Tag
µmo
l/l
K
B
F

Abb. 4.6.6: OrnQ: Fetomaternaler Ornithinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Cystathionin
In Gruppe B kommt es zwischen dem 14. und 16. Tag zu einem deutlichen Anstieg der
Cystathioninkonzentration im maternalen Plasma, im weiteren Verlauf bleibt die Konzentration
konstant. Die Graphen der Gruppen K und F liegen ab dem 16. Tag deutlich unter dem der Gruppe
B und nähern sich zum 20. Tag einander an. Zwischen dem 14. und dem 20. Trächtigkeitstag kann
im maternalen Plasma der Gruppe B eine signifikant höhere Cystathioninkonzentration ermittelt
werden als in der Gruppe K. Mit einer Irrtumswahrscheinlichkeit von p≤0,06 kann am 20.
Trächtigkeitstag ebenfalls eine Erhöhung der Aminosäurekonzentration in Gruppe B gegenüber
Gruppe F nachgewiesen werden. Die Nachweisgrenze für Cystathionin liegt bei 0,5 µmol/l, die
Cystathioninkonzentration der Gruppe K sowohl im maternalen Blutplasma als auch in der
Amnionflüssigkeit liegt an den Tagen 14 und 16 lagen unterhalb der Nachweisgrenze und wurden
daher für die statistische Auswertung mit 0,5 µmol/l berücksichtig.
Die Graphen der Gruppen B und F zeigen sowohl bei der Aminosäurekonzentration in der
Amnionflüssigkeit als auch bei den fetomaternalen Quotienten einen parabelförmigen Verlauf mit
Scheitelpunkt am 16. Tag. Der Graph der Gruppe K steigt nach waagerechtem Verlauf zwischen
dem 14. und 16. Tag zwischen dem 16. und 20. Tag linear an.
Am 16. Trächtigkeitstag kann in Gruppe B ein signifikant verringerter fetomaternaler
Aminosäurequotient gegenüber Gruppe K errechnet werden. Die Aminosäurekonzentrationen der
Amnionflüssigkeit zeigen keine signifikanten Unterschiede.
67
OrnQ
01
23
45
67
89
14 16 18 20
Tag
K
B
F

Abb. 4.6.7: CystaM: Cystathioninkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Glutamin
Während die Glutaminkonzentration im maternalen Plasma der Gruppen K und B zwischen dem
14. und 16. Tag weitestgehend konstant bleibt, kommt es in Gruppe F zunächst zu einem leichten
Abfall, später zu einem Anstieg der Konzentration. Die Plasmakonzentration der Aminosäure
Glutamin ist am 16. Tag in der Gruppe F signifikant geringer als in den Gruppen K und B. Am 20.
Tag ist sie in der Gruppe B signifikant höher als in den Gruppen K und F.
Abb. 4.6.9: CystaQ: Fetomaternaler Cystathioninquotient im zeitlichen VerlaufStandartabweichungen siehe Tab. 10.1 im Datenanhang.
CystaQ
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
14 16 18 20
Tag
K
B
F
68
CystaM
0
0,2
0,4
0,6
0,8
1
1,2
1,4
14 16 18 20
Tag
µmol
/l
K
B
F
Abb. 4.6.8: CystaAm: Cystathioninkonzentration in der Amnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
CystaAm
0
0,2
0,4
0,6
0,8
1
1,2
1,4
14 16 18 20
Tag
µmo
l/l
K
B
F

Abb. 4.6.10: GluM: Glutaminkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.6.11: GluAm: Glutaminkonzentration in derAmnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 4.6.12: GluQ: Fetomaternaler Glutaminquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Die Graphen der Amnionflüssigkeit sowie des fetomaternalen Quotienten zeigen einen
parabelförmigen Verlauf, mit Scheitelpunkt am 16. Tag.
In der Amnionflüssigkeit sind abgesehen vom 16. Tag, an welchem die Konzentration in der
Gruppe B signifikant höher ist als in den Gruppen K und F keine weiteren signifikanten
Unterschiede zwischen den Gruppen zu messen.
Der fetomaternale Glutaminquotient ist am 16. Tag in Gruppe F signifikant höher als in Gruppe K.
Phenylalanin
Im maternalen Plasma der Tiere aller Gruppen zeigen sich keine signifikanten Unterschiede.
Am 14. Tag ist die Phenylalaninkonzentration in der Amnionflüssigkeit in Gruppe F signifikant
höher als in Gruppe B. Die Werte der Gruppe K liegen zwischen denen der Gruppen B und F, ohne
jedoch signifikant abzuweichen.
69
GluM
0
100
200
300
400
500
600
14 16 18 20
Tag
µm
ol/
l K
B
F
GluAm
0
200
400
600
800
1000
1200
1400
1600
1800
14 16 18 20
Tag
µm
ol/
l K
B
F
GluQ
0
0,5
1
1,5
2
2,5
3
3,5
4
14 16 18 20
Tag
µmo
l/l
K
B
F

Es konnten keine signifikant voneinander abweichenden fetomaternalen Phenylalaninquotienten
errechnet werden.
Taurin
Die Taurinkonzentration im maternalen Plasma und der Amnionflüssigkeit sowie die
fetomaternalen Quotieten zeigen keinerlei signifikante Unterschiede.
Hydroxyprolin
Auch bei der Aminosäure Hydroxyprolin können weder im maternalen Plasma, noch in der
Amnionflüssigkeit, noch bei den fetomatrnalen Quotienten signifikante Unterschiede ermittelt
werden.
Asparagin
Der fetomaternale Asparaginquotient der Gruppe F ist am 16. Tag gegenüber Gruppe K signifikant
reduziert, am 20. Tag ist er gegenüber den Gruppen K und B signifikant erhöht. Bei den
maternalen Plasma- und den Amnionflüssigkeitskonzentrationen zeigen sich keine signifikante
Unterschiede.
Citrullin
Im maternalen Blutplasma sind zwischen den Gruppen über den gesamten Messzeitraum keine
signifikanten Unterschiede zu errechnen.
Am 20. Trächtigkeitstag kommt es in der Amnionflüssigkeit der Gruppen B und F zu einer
signifikanten Erhöhung der Citrullinkonzentration gegenüber Gruppe K.
Entsprechend ist der fetomaternale Citrullinquotient in den Gruppen B und F am 20. Tag größer 1,
in Gruppe K hingegen kleiner 1. Der fetomaternale Quotient am 20. Tag der Gruppe B ist
signifikant größer als in Gruppe K, mit einer Irrtumswahrscheinlichkeit von p≤0,06 ist auch der
fetomaternale Quotient der Gruppe F signifikant größer als in Gruppe K.
70

Abb. 4.6.13: CitrM: Citrullinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
4.7 Histomorphologische Untersuchung der Plazenten
Bei der histomorphologischen Untersuchung der Plazenten können Unterschiede zwischen den
einzelnen Gruppen nachgewiesen werden.
In zwei Plazenten des 20. Trächtigkeitstages der Gruppe B können bereits in der HE-Färbung
großflächige Nekrosen im Labyrinth festgestellt werden, die sich zum Teil bis in den
Spongiotrophoblasten erstrecken.
In der PBA-Färbung können neben den Basallaminae der Endothelien auch weitere Einlagerungen
von Mukopolysaccariden um die fetalen Gefäße nachgewiesen werden (Abb. 4.7.1). Da diese sich
jedoch durch zweistündige Diastase-Inkubation vor der Färbung weitgehend eliminieren lassen
(Abb. 4.7.2) und immunhistochemisch mit Hilfe des Anti-Laminin-Antikörpers nicht
Abb. 4.6.14: CitrAm: Citrullinkonzentration in derAmnionflüssigkeit im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
CitrAm
010
20
30
40506070
8090
100
14 16 18 20
Tag
µmo
l/l
K
B
F
Abb. 4.6.15: CitrQ: Fetomaternaler Citrullinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
CitrQ
0
0,5
1
1,5
2
2,5
14 16 18 20
Tag
K
B
F
71
CitrM
0
10
20
30
40
50
60
70
80
14 16 18 20
Tag
µmol
/l
K
B
F

nachgewiesen werden können, muss davon ausgegangen werden, dass es sich nicht um
Bestandteile der fetalen Basallamina handelt, sondern um intrazelluläre Glykogeneinschlüsse. Bei
diesen bereits von Schiebler und Knoop (1959) beschriebenen Glykogengranula im
Synzytiotrophoblasten ist zu ergänzen, dass die Glykogeneinschlüsse praktisch nur um fetale
Gefäße und nicht um maternale Sinus zu finden sind.
Abb. 4.7.1: K16/2F PBA L4:Fetales Gefäß mit deutlichen PBA positiven Strukturen neben der Basallamina des fetalen Kapilarendothels. Plazenta aus der Gruppe K, Tag 16 der Trächtigkeit.
Abb. 4.7.2: K16/2F PBA-D L5:Deutliche Reduktion der PBA positiven perivaskulären Strukturen nach zweistündiger Diastase-Inkubation. Plazenta aus der Gruppe K, Tag 16 der Trächtigkeit.
Abb. 4.7.3: B16/8F PBA L5: Fetales Gefäß mit deutlichen PBA positiven Strukturen neben der Basallamina des Endothels. Plazenta aus der Gruppe B, Tag 16 der Trächtigkeit.
72

Unter Berücksichtigung der Gefäßgröße erscheint der Eindruck, dass diese Mukopolysaccaride bei
den Plazenten der Tiere der Gruppe B vermehrt auftreten (Abb. 4.7.3).
Des weiteren können bei Plazenten des 16. sowie des 20. Trächtigkeitstages von Tieren der
Gruppe B sowohl in der PBA- als auch in der PAS-Färbung kräftige PBA- bzw. PAS-positive
Strukturen um größere fetale Gefäße (Abb. 4.7.5) gezeigt werden, welche vor allem in Bouin-
fixiertem Gewebe gut hervortreten. Diese Strukturen können sowohl in längs (Abb. 4.7.6) als auch
in quer (Abb. 4.7.5) angeschnittenen Gefäßen dargestellt werden. Ähnliche, jedoch deutlich
aufgelockerte Strukturen sind auch in Gruppe K zu finden (Abb. 4.7.4).
Abb. 4.7.4: K16/2B PBA L4: Größeres fetales Gefäß (quer) im Labyrinth der Rattenplazenta mit diffusen PBA positiven Strukturen um das Gefäß. Gruppe B, 16. Tag der Trächtigkeit.
Abb. 4.7.5: B16/1B PBA L2: Größeres fetales Gefäß (quer) im Labyrinth der Rattenplazenta, mit deutlichen PBA positiven Strukturen um das Gefäß. Gruppe B, 16. Tag der Trächtigkeit.
Abb. 4.7.6: B16/1B PBA L7: Größeres fetales Gefäß (längs) im Labyrinth der Rattenplazenta mit deutlichen PBA positiven Strukturen um das Gefäß. Gruppe B, 16. Tag der Trächtigkeit.
73

Mit der PBA-Färbetechnik konnte besonders an Bouin-fixiertem Gewebe basalmembranartige
Interzellulärsubstanz dargestellt werden.
Des weiteren finden sich sowohl im Spongiotrophoblasten als auch im apikalen Labyrinth,
Gruppen von Zellen mit PBA- bzw. PAS positivem Zytoplasma (Abb. 4.7.7). Da sich das
Zytoplasma nach zweistündiger Diastase-Inkubation der Schnitte nicht mehr PBA- bzw. PAS
positiv darstellt, muß davon ausgegangen werden, dass es sich um glykogenhaltige Zellen handelt.
In Hinblick auf den Glykogengehalt dieser Zellen können zwischen den Gruppen keine
Unterschiede festgestellt werden. Es finden sich jedoch bereits innerhalb der Gruppen große
Unterschiede zwischen dem Glykogengehalt und der Anzahl der Glykogenzellen in den Plazenten.
Zum Teil wurden Zellgruppen von über 30 Glykogenzellen im Labyrinth gezählt (Abb.: 4.7.7).
Die Anzahl der Glykogenzellen – vor allem im Spongiotrophoblasten- nimmt im Laufe der
Trächtigkeit deutlich ab.
Abb.4.7.7: B20/7F PBA L1: Gruppe von Glykogenzellen im apikalen Labyrinth. Gruppe B, 20. Tag der Trächtigkeit.
74

Bereits am 14. Tag sind Riesenzellen mit PBA positiven Zytoplasmaeinschlüssen zu beobachten,
wobei es scheint, dass hier die Plazenten der Gruppen B mehr PBA positive Riesenzellen
aufweisen.
Abb. 4.7.8: K16/2F PBA: Zwei Riesenzellen mit deutlich glykogenhaltigem Zytoplasma. Gruppe K, 16. Tag der Trächtigkeit.
Die Basalmembran des Amnionepithels scheint in Gruppe B (Abb. 4.7.10) gegenüber Gruppe K
(Abb. 4.7.9) verdickt zu sein. Dieser Unterschied fällt vor allem am 16. und 20. Tag auf.
In Gruppe B scheint Gegenüber Gruppe K eine vermehrte granulozytäre Infiltration vorhanden zu
sein. Diese ist am 14. Tag in der basalen Plazenta – in der Dezidua zu sehen und steigt im weiteren
Verlauf der Trächtigkeit Richtung apikal. Am 20. Tag ist in Gruppe B eine deutliche Infiltration
neutrophiler Granulozyten am Übergang des Spongiotrophoblasten zum Labyrinth zu sehen
Abb. 4.7.10: B20/7F PAS L1: Basalmembran des Amnionepithels am 20. Tag in Gruppe B.
Abb. 4.7.9: K20/8F PAS L1: Basalmembran des Amnionepithels am 20. Tag in Gruppe K.
75

(Abb. 4.7.12). In Gruppe K ist diese Infiltration in einem wesentlich geringeren Umfang zu
beobachten (Abb. 4.7.11).
4.8 Immunhistochemische Untersuchung der Plazenten
Bei den immunhistochemischen Untersuchungen mit dem Ki-67 Antikörper als Marker für
Zellproliferation kann bei den Plazenten der Gruppe K des 14. und 16. Trächtigkeitstages im
basalen Labyrinth eine rege Proliferation der Trophoblastenzellen festgestellt werden.
Abb. 4.8.1: arithmetisches mittel der Ki-67 positiven Trophoblastenzellen am 16. Tag in einem Gesichtsfeld. Es wurden mind. 9 Gesichtsfelder (20x Vergrößerung) pro Gruppe ausgezählt.
Ki-67 positive Zellen 16. Tag
K; 26
B; 12,2
F; 7,4
0
5
10
15
20
25
30
Gruppe
An
zah
l K
i-67
po
s. Z
elle
n
K
B
F
Abb. 4.7.11: K20/6F PAS L1: Granulozyten am Übergang des Spongiotrophoblasen zum Labyrinth am 20. Tag in Gruppe K.
Abb. 4.7.12: B20/1F PAS L1: Granulozyten am Übergang des Spongiotrophoblasten zum Labyrinth am 20. Tag in Gruppe B.
76

In den Gruppe B sowie F ist im Labyrinth der Plazenten eine deutlich geringere Zellproliferation
zu beobachten als in der Gruppe K.
Zur semiquantitative Erfassung, wurden die Ki-67 positiven Zellen pro Gesichtsfelder ausgezählt
und miteinander verglichen (siehe Abb. 4.8.4). Hierzu wurden jeweils Drei Gesichtsfelder pro
Plazenta von Drei Plazenten pro Gruppe zu einem Zeitpunkt (Tag 16) ausgezählt. Ein Großteil der
fetalen kernhaltigen Erythrozyten weisen Ki-67 positive Zellkerne auf (Abb.4.8.3), welche bei der
Auszählung von den Ki-67 positiven Zellen des Labyrinths berücksichtigt wurden.
Abb. 4.8.2: K16/7F Ki-67 L8: Ki-67 positiver Zellkern einer Trophoblastenzelle des Labyrinths
Abb. 4.8.3: K16/7F Ki-67 L6: Fetaler Ki-67 positiver Retikulozyt im Labyrinth
Bei der immunhistochemischen Untersuchung mit dem Anti-aktive Caspase-3-Antikörper als
Apoptosemarker ist eine deutliche Caspase-3 Aktivität in den Amnionepithelzellen aller Gruppen
ab dem 16. Trächtigkeitstag zu beobachten (Abb. 4.8.5). Quantitative Unterschiede zwischen den
Abb. 4.8.4: K16/3F Ki-67 L1: Ki-67 positive Zellkerne des maternales Labyrinths, sowie der fetalen Retikulozyten. Plazenta aus Gruppe K, Tag 16.
77

Gruppen konnten nicht erkannt werden. Der Zellbesatz der Basalmembran ist am 20. Tag deutlich
reduziert, z.T. finden sich Zelltrümmer auf der Basalmembran (Abb. 4.8.6).
Auch im Labyrinth der Plazenten können mit Hilfe des Anti-aktive Caspase-3-Antikörpers
apoptotische Zellen nachgewiesen werden (Abb.4.8.7). Eine Zuordnung der Zellen zu einer
Schicht der Plazentaschranke ist lichtmikroskopisch nicht möglich. Aufgrund der ohnehin seltenen
apoptotischen Ereignisse im Labyrinth (z.T. Caspase-3 freie Schnitte) entfällt ein Vergleich
zwischen den Gruppen. In der Zusammenfassung der Gruppen, ergibt sich folgende Anzahl von
apoptotischen Ereignissen.
Abb. 4.8.5: K16/1F Cas3 L1: Nachweis von aktiver Caspase-3 im Zytoplasma der Amnionepithelzellen. Gruppe K, Tag 16.
Abb. 4.8.7: K20/6F Cas3 L1: Nachweis von aktiver Caspase-3 im Zytoplasma einer Zelle des Labyrinths. Gruppe K, Tag 20.
Abb. 4.8.6: K20/5F Laminin L1: deutlich reduzierter Zellbesatz und Zelltrümmer auf der Basalmembran des Amnionepithels. Gruppe K, Tag 20.
78

K16 B16 F164 14 2K20 B20 F206 2 6
Es wurden jeweils 2 Schnitte von 4 Plazenten pro Gruppe ausgezählt wurden. Die Anzahl
apoptotischer Ereignisse schwankt auch innerhalb der Gruppen stark. Da hierbei wahrscheinlich
methodische Mängel zugrunde liegen (s.S. 89) fand der Nachweis von aktiver Caspase-3 lediglich
an den Tagen 16. und 20 statt.
Die immunhistochemische Untersuchung mit dem Anti-Laminin-Antikörper erfolgte zusätzlich
zur Differenzierung der Ergebnisse der PAS- und PBA-Färbung.
Unterschiede zwischen den Gruppen konnten hierbei jedoch nicht erkannt werden.
Tabelle 4.8.1: Anzahl Caspase-3 positiver Zellen in den Gruppen K, B und F
79

4.9 Resorptionen
Obwohl sowohl die Anzahl der Resorptionen als auch die Anzahl der ausgetragenen Früchte –
auch innerhalb einer Gruppe deutlichen Schwankungen unterliegt, können signifikante
Unterschiede zwischen den Gruppen errechnet werden. In Gruppe K konnten durchschnittlich
83,58% (N=22) aller implantierten Früchte ausgetragen werden. In Gruppe B war dies in lediglich
62,36% (N=19) der Fall, in Gruppe F bei 86,81% (N= 18). Somit ist die Anzahl der Resorptionen
in Gruppe B signifikant höher als in den Gruppen K und F. Im Umkerhrschluß bedeutet dies, dass
die Tiere der Gruppe B signifikant weniger Früchte austragen als die Tiere der Gruppen K und F,
obwohl im Mittel gleichviel Implantationen stattgefunden haben.
Abb. 4.9.1: Implantationsnachweis nach Salewski (1964) an einem Uterus
80

5. Diskussion
5.1 Blut- und Amnionparameter
In Anlehnung an Roth-Maier et al. (1996) erfolgte die Bestimmung der Transaminasen ALAT und
ASAT im Serum der Muttertiere um den indirekten Vitamin-B6-Status der Tiere beurteilen zu
können.
In Übereinstimmung mit O´Kane und Gunsalus (1947) welche Pyridoxalphosphat als Coenzym
der ASAT nachwiesen und mit Ono et al. (1995) die in vivo bei Hämodialysepatienten zeigen
konnten, dass eine Reduktion der Aktivität der plasmatischen ALAT und ASAT mit einem
reduziertem Pyridoxal-5-Phosphat Plasmaspiegel zusammenhängen, sind in der vorliegenden
Arbeit die Aktivitäten der maternalen plasmatischen ASAT und ALAT in Gruppe B z.T. signifikant
reduziert.
Da sich die Arbeit von Roth-Maier et al. (1996) mit der laktierenden Ratte auseinander setzt und
somit ein anderes Zeitfenster beleuchtet wird, ist ein Vergleich der Aktivität der Enzyme nicht
möglich. Die von Roth-Maier et al. (1996) beschriebene nicht vorhandene Reduktion der
Enzymaktivität der ALAT im Gegensatz zur ASAT kann nicht bestätigt werden. Roth-Maier et al.
halten dieses Ergebnis durch eine höhere Coenzymabsättigung der ALAT im Gegensatz zur ASAT
für möglich. In den vorliegenden Ergebnissen ist jedoch die Aktivität der ALAT in Gruppe B
stärker reduziert als die der ASAT.
Jung et al. (1978) konnten durch Supplimentierung von Pyridoxal-5-Phosphat beim Menschen
eine Zunahme der ASAT-Aktivität um 37% und der ALAT-Aktivität um 15,2% im Serum zeigen.
Hieraus könnte eine höhere Affinität der ASAT für Pyridoxal-5-Phosphat gefolgert werden,
welches Roth-Maier et al. (1996) widerspricht und die in dieser Arbeit vorliegenden Ergebnisse
erklären könnte.
Die Aktivität der ALAT ist in Gruppe B ab dem 14. Tag gegenüber Gruppe K signifikant reduziert.
Dies ist ebenfalls dadurch zu erklären, dass Vitamin-B6 ein Coenzym der ALAT ist.
Die ALAT spielt eine wichtige Rolle bei der Ausscheidung von Stickstoff und somit für den
gesamten Aminosäurehaushalt. Differenzen der Aminosäurekonzentrationen sowohl im
maternalen Plasma als auch in der Amnionflüssigkeit könnten somit z.T. durch eine reduzierte
Aktivität der ALAT in den Gruppen B und F erklärt werden. Wobei darauf aufmerksam gemacht
werden muss, dass es noch eine Reihe weiterer Vitamin-B6 abhängige Aminotransferasen gibt.
81

Für die ASAT welche ebenfalls Vitamin-B6 als Coenzym benötigt, konnte am 16. Trächtigkeitstag
eine signifikant verringerte Enzymaktivität in Gruppe B nachgewiesen werden. Am 20. Tag ist
dies für Gruppe F der Fall. Da die Aktivität des Enzyms in Gruppe B jedoch an allen Tagen z.T.
deutlich unter der Aktivität in Gruppe K liegt, kann geschlussfolgert werden, dass sich in einem
zahlenmäßig größeren Tierversuch noch mehr signifikante Unterschiede zeigen würden. Da die
ASAT durch Bereitstellung von Aminogruppen eine wichtige Rolle bei der Synthese von Purinen,
Pyrimidinen und Aminozuckern spielt, hängt sie unmittelbar mit der Zellproliferation zusammen,
und spielt somit u.a. in der Entwicklung von fetalen und plazentaren Gewebe eine wichtige Rolle.
Im Gegensatz zu Lakshmi et al. (2001) welche beim Menschen lediglich bei einem
Folsäuremangel einen höheren Homocysteinspiegel nachweisen konnten, während Ihnen dies bei
einer Vitamin B6 Hypovitaminose nicht gelang, kann in den vorliegenden Ergebnissen am 14. und
16. Tag sowohl in Gruppe F als auch in Gruppe B ein gegenüber Gruppe K signifikant erhöhter
Homocysteinspiegel nachgewiesen werden. In diesem Zusammenhang sei darauf hingewiesen das
Dierkes et al. (1998) lediglich bei einer Folsäure Supplimentierung, nicht jedoch bei einer Vitamin
B6 Supplimentierung eine Reduktion des Homocysteinspiegels beobachten konnten.
In der Gruppe B ist der erhöhte Homocysteinspiegel durch die Repremierung der Cystathionin β-
Synthase (CBS) zu erklären, welche Pyridoxal-5-Phosphat abhängig ist und den Abbau von
Homocystein zu Cystathionin katalysiert welches ebenfalls Pyridoxal-5-Phosphat abhängig weiter
zu Cystein abgebaut wird. Obwohl die CBS Vitamin-B6 abhängig ist, finden sich in Gruppe B
erstaunlicherweise signifikant erhöhte Cystathioninspiegel gegenüber Gruppe K an den Tagen 14,
16 und 20. Eine mögliche Erklärung für den erhöhten Cystathioninspiegel in Gruppe B könnte
eine Repremierung des ebenfalls pyridoxalphosphatabhängigen Enzyms Cystathionin-γ-Lyase
sein, welches die Reaktion von Cystathionin zu Cystein und α-Kerobutyrat katalysiert (Goddijn-
Wessel et al., 1996). In diesem Fall wirkt sich der Vitamin-B6-Mangel stärker auf die Aktivität der
Cystathionin-γ-Lyase aus, als auf die Aktivität der CBS.
Die Tatsache dass kaum erhöhte Cysteinkonzentrationen gemessen werden können, ist
möglicherweise auch mit den vielen Stoffwechselreaktionen zu erklären, welche Cystein als
Substrat verwenden und es z.B. zu Pyruvat abbauen, zu Taurin decarboxylieren oder zu
Mercaptopyruvat transaminieren. Teilweise dienen diese Reaktionen zur Verbesserung der
Ausscheidungsfähigkeit des Cysteins (Löffler und Petrides, 1998). Bezüglich der
Taurinkonzentration des maternalen Plasmas lassen sich sich keine signifikanten Unterschiede
zwischen den Gruppen K und B feststellen. Diese Ergebnisse untermauern nicht die Vermutung,
dass die Decarboxylierung von Cystein zu Taurin bei Vitamin-B6 Restriktion gesteigert ist.
82

Da neben Homocystein auch Serin Substrat für die CBS ist, hätte in Gruppe B mit einer erhöhten
Serinkonzentration gerechnet werden können, da ein Serinverbrauchender Stoffwechselweg
gehemmt wird. Es lassen sich jedoch zwischen Gruppe K und B keine signifikanten Unterschiede
bezüglich der Serinkonzentrationen im maternalen Plasma oder in der Amnionflüssigkeit
nachweisen.
In Anlehnung an Goddijn-Wessel et al. (1996) ist der erhöhte Homocysteinspiegel in Gruppe F
durch Reduktion der Aktivitäten der Enzyme 5-Methyltetrahydrofolat-Homocystein-
Methyltransferase und 5,10-Methylentetrahydrofolat Reduktase zu erklären, deren Substrate
Folsäurederivate sind und welche eine zentrale Rolle bei der Remethylierung von Homocystein zu
Methionin spielen. An den Tagen 14 und 16 ist in Gruppe F gegenüber Gruppe K eine signifikante
Erhöhung der Vitamin-B6 Konzentration im maternalen Plasma zu messen. Hier kann gemutmaßt
werden, ob dies Teil einer Kompensationsreaktion ist. Zwischen dem 16. und 20. Tag kommt es in
Gruppe F jedoch zu einem deutlichen Abfall der Vitamin-B6-Konzentration sodass die
Konzentration unter der in Gruppe K liegt. Dieser Unterschied ist jedoch nicht signifikant.
Der Methioninspiegel der Gruppe B im maternalen Plasma hat sich im Gegensatz zu Gruppe F
nicht erhöht. Es hätte vermutet werden können, dass die Remethylierung des Homocysteins durch
das Enzym 5-Methyltetrahydrofolat-Homocystein-Methyltransferase aufgrund eines erhöhten
Substratangebots und der vorhandenen Kofaktoren in Gruppe B gesteigert würde.
Weiterhin verwundert der signifikant erhöhte Methioninspiegel im maternalen Plasma der Gruppe
F an den Tagen 14. und 16. der Trächtigkeit. Da durch die Vitaminrestriktion die Remethylierung
von Homocystein zu Methionin reduziert wird, wäre eher mit einem verringerten
Methioninspiegel zu rechnen gewesen. Die Hemmung dieser Remethylierung in Gruppe F bei
welcher die Folsäure notwendige Methylgruppen überträgt erklärt die erhöhte Konzentration von
Homocystein im maternalen Plasma.
Der Abbau von Homocystein zu Cystathionin und Cystein ist nicht beeinträchtigt, es lassen sich
jedoch bei dieses Parametern im maternalen Plasma keine signifikanten Unterschiede zwischen
den Gruppen K und F finden. Dies hätte im Rahmen einer kompensatorischen Reaktion zur
Senkung des Homocysteinspiegels erwartet werden können. Da die Homocystein abbauenden
Stoffwechselwege nicht beeinträchtigt sind und keine höhere Konzentration von Cystathionin und
Cystein in Gruppe F nachweisbar sind, kann vermutet werden, dass der Organismus in der Lage ist
den erhöhten Anfall von Homocystein Abbauprodukten weiter zu verstoffwechseln. Die
Serinkonzentration im maternalen Plasma der Gruppe F ist gegenüber den Gruppen K und B
83

jedoch am 16. Tag signifikant erhöht. Dies verwundert, da Serin beim Abbau von Homocystein
verbraucht wird.
Das Enzym Serin/Threonin-Hydroxymethyltransferase ist ebenfalls Pyridoxalphosphat abhängig.
Es katalysiert die Reaktion von Serin/Threonin und THF zu Glycin und N5,N10-Methylen-THF.
Dies könnte die signifikant erhöhten Serinwerte im maternalen Plasma der Gruppe F am 16. Tag
erklären. Auch die an allen Tagen in den Gruppen B und F signifikant erhöhte
Threoninkonzentration im maternalen Plasma sowie die an den Tagen 14, 16 und 20 erhöhte
Threoninkonzentration in der Amnionflüssigkeit könnte hierdurch erklärt werden.
Wirtschafter (1958b) konnte zeigen, dass die totale Konzentration freier Aminosäuren in der
Amnionflüssigkeit zum Zeitpunkt der Geburt beim Menschen geringer ist, als die im maternalen
Plasma. Für die in der vorliegenden Arbeit untersuchten Aminosäuren konnte dies am 20.
Trächtigkeitstag (gesamte Tragzeit 22 Tage) lediglich für Glutaminsäure und Asparaginsäure
festgestellt werden. Diese Unterschiede könnten durch die Vermutung von Kedenburg und
Mülling (1975) erklärt werden, dass sich bei der Ratte auch in der die Amnionhöhle umgebenden
Dottersackwand aktive Transportproteine für Aminosäuren befinden und sich daher in der
Amnionflüssigkeit der Ratte eine insgesamt höhere Aminosäurekonzentration findet als beim
Menschen.
Ein unregelmäßiges Auftreten der Aminosäuren Arginin, Ornithin, Glutamin, Tyrosin und
Asparaginsäure im maternalen Serum oder der Amnionflüssigkeit, wie von Wirtschafter (1958b)
beim Menschen beobachtet, konnte in der vorliegenden Arbeit nicht gefunden werden. Lediglich
die maternale Cystein- und Cystathioninkonzentration sowie die fetale Cysteinkonzentration
liegen in dieser Arbeit unterhalb der Nachweisgrenze von 0,5µmol/l.
Wirtschafter (1958a) konnte in einem Versuch mit trächtigen Ratten des Long-Evans Stammes
Ornithin zwar in der Amnionflüssigkeit, nicht jedoch im maternalen Serum nachweisen. In der
vorliegenden Arbeit ist Ornithin jedoch in allen Gruppen über den gesamten Zeitraum sowohl in
der Amnionflüssigkeit als auch im maternalen Serum nachzuweisen. Am 13. Tag ist es
Wirtschafter (1958b) nicht gelungen Asparaginsäure in der Amnionflüssigkeit nachzuweisen.
Am 14. Trächtigkeitstag beträgt die Asparaginsäurekonzentration der Amnionflüssigkeit der
Gruppe K in der vorliegenden Arbeit 50,4 µmol/l.
Obwohl die maternale Plasmakonzentration von Serin in Gruppe F am 16. Tag signifikant erhöht
ist, zeigen sich keine signifikanten Unterschiede der Serinkonzentration in der Amnionflüssigkeit.
84

Jedoch ist der fetomaternale Serinquotienten der Gruppe F gegenüber den Gruppen K und B am
16. Tag signifikant reduziert. Dies könnte auf eine kompensatorische Funktion der Plazenta
hindeuten. Die Serinkonzentration wird unabhängig von der maternalen Konzentration in der
Amnionflüssigkeit konstant gehalten, indem der diaplazentare Serintransport reduziert wird. Da es
sich in allen Gruppen um einen aktiven Transport handelt, kann es also zu einer Repression eines
Transportproteins gekommen sein.
Abb. 5.1.1: SerM: Serinkonzentration im maternalen Plasma im Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Abb. 5.1.2: SerAm: Serinkonzentration in der Amnionflüssigkeit in Verlauf der Trächtigkeit. Standartabweichungen siehe Tab. 10.1 im Datenanhang
Am 16. und 20. Tag ist im maternalen Plasma der Gruppen B eine signifikant reduzierte
Ornithinkonzentration zu messen. Am 16. Tag ist dies auch in der Amnionflüssigkeit der Fall. Da
Pyridoxalphosphat Coenzym bei der Decarboxylierung von Ornithin zu Putrescin und CO2 ist,
wäre an und für sich mit einer erhöhten Ornithinkonzentration in Gruppe B zu rechnen gewesen.
SerQ
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
14 16 18 20
Tag
K
B
F
Abb. 5.1.3: SerQ: fetomaternaler Serinquotient im zeitlichen Verlauf. Standartabweichungen siehe Tab. 10.1 im Datenanhang
85
SerM
0
50
100
150
200
250
14 16 18 20
Tag
µm
ol/
l K
B
F
SerAm
0
100
200
300
400
500
600
700
14 16 18 20
Tag
µm
ol/l
K
B
F

Die Aminosäuren Glycin, Alanin, Valin, Methionin, Prolin, Tyrosin, Serin, Threonin, Histidin,
Butyrat, Cystathionin sowie Glutamin zeigen im zeitlichen Verlauf in der Amnionflüssigkeit einen
parabelförmigen Verlauf mit Scheitelpunkt am 16. Trächtigkeitstag. Die Ausprägung zwischen den
Gruppen ist variabel z.T. zeigen sich signifikante Unterschiede zwischen den Gruppen. Hierbei
bleibt die maternale Plasmakonzentration weitestgehend konstant, z.T. kommt es zum Abfall der
maternalen Plasmakonzentrationen d.h. es kommt zur Induktion von Transportprozessen ab dem
16. Trächtigkeitstag. Aufgrund der weitestgehend konstant bleibenden maternalen
Aminosäurekonzentration zeigen die entsprechenden fetomaternalen Quotienten ebenfalls einen
parabelförmigen Verlauf.
Die Threoninkonzentration im maternalen Plasma der Gruppen B und F sind gegenüber der
Gruppe K im gesamten Messzeitraum signifikant erhöht. Obwohl dies auch für die
Amnionflüssigkeit gilt, ist der fetomaternale Threoninquotient am 14. Tag in den Gruppen B und F
gegenüber Gruppe K signifikant reduziert. Am 16. Tag ist dies noch in Gruppe F der Fall. Auch in
diesem Fall kann also über eine kompensatorische Reduktion des Threonintransportes diskutiert
werden, der allerdings – im Gegenteil zum Serin- nicht ausreicht die maternalen
Aminosäuredifferenzen zwischen den Gruppen in der Amnionflüssigkeit auszugleichen.
Obwohl sich bei der Aminosäure Citrullin keine signifikanten Unterschiede im maternalen Plasma
zwischen den Gruppen messen lassen, ist die Citrullinkonzentratiom in der Amnionflüssigkeit am
20. Tag in den Gruppen B und F gegenüber Gruppe K signifikant erhöht. Dies spiegelt sich auch
in den fetomaternalen Quotienten wieder, welche am 20. Tag in Gruppe B signifikant und in
Gruppe F mit einer Irrtumswahrscheinlichkeit von p≤0,06 größer sind als in Gruppe K.
Bei der Aminosäure Butyrat ist am 20. Tag sowohl im maternalen Plasma als auch in der
Amnionflüssigkeit der Gruppen B und F eine signifikant höhere Konzentration zu messen, als in
Gruppe K.
Ähnliches ist bei der Aminosäure Prolin am 20. Tag zu Beobachten. Obwohl es im maternalen
Plasma zwischen den Gruppen keine signifikanten Unterschiede gibt, ist die Prolinkonzentration
in der Amnionflüssigkeit am 20. Tag in den Gruppen B und F gegenüber Gruppe K signifikant
erhöht. Jedoch ist lediglich zwischen den Gruppen K und F am 20. Tag ein signifikant erhöhter
fetomaternale Aminosäurequotient in Gruppe F zu errechnen.
Bei der Aminosäure Citrullin scheint es also in den Gruppen B und F zum Ende der Trächtigkeit
zur Induktion aktiver Transportprozesse zu kommen, welche in Gruppe K nicht beobachtet werden
kann. Bei Prolin kommt es zu einer höheren Konzentration in der Amnionflüssigkeit, ohne das es
zu einer Zunahme der maternalen Aminosäurekonzentration gekommen ist, während bei Butyrat
86

eine Zunahme der maternalen Konzentration zu messen ist. Mechanismen die eine konstante
Aminosäurekonzentration auf fetaler Seite schaffen scheinen bei diesen Aminosäuren also nicht
vorhanden zu sein.
Bei der Aminosäure Butyrat scheint ein nicht gesättigter maternofetale Aminosäuretransport
vorzuliegen, da sowohl die maternale als auch die fetale Aminosäurekonzentration in den Gruppen
B und F signifikant gestiegen ist, ohne das es zu einem signifikanten Unterschied bei den
fetomaternalen Quotienten gekommen ist.
Bei der Aminosäure Lysin scheint dieser Effekt sogar noch ausgeprägter vorhanden zu sein. Hier
sind bereits am 14. und 16. Tag in den Gruppen B und F gegenüber Gruppe K signifikant erhöhte
Aminosäurekonzentrationen im maternalen Plasma und der Amnionflüssigkeit zu messen
(Irrtumswahrscheinlichkeit an Tag 14 in der Amnionflüssigkeit zwischen Gruppe K und F
p≤0,06). Während diese Unterschiede in der Amnionflüssigkeit auch am 20. Tag noch signifikant
sind, ist im maternalen Plasma lediglich in Gruppe B eine mit einer Irrtumswahrscheinlichkeit von
p≤0,06 eine signifikant erhöhte Aminosäurekonzentration zu messen. Auch an dieser Stelle kann
gemutmaßt werden, dass sich in einem zahlenmäßig größeren Tierversuch am 20. Tag im
maternalen Plasma weiterhin signifikante Unterschiede hätten finden lassen.
Auch bei der Aminosäure Tyrosin scheint es keinen Kompensationsmechanismus zu geben der die
fetale Aminosäurekonzentration konstant hält. Am 14. Tag ist im maternalen Plasma in den
Gruppen B und F eine signifikant erhöhte Aminosäurekonzentration gegenüber Gruppe K zu
messen, am 16. Tag ist dies nur für Gruppe F, am 20. Tag nur für Gruppe B der Fall. Am 14. und
16. Tag ist in Gruppe B eine signifikant erhöhte Tyrosinkonzentration in der Amnionflüssigkeit zu
messen, am 20. Tag ist dies für die Gruppe B und F der Fall. Zwischen der Gruppe K und den
Gruppen B und F gibt es jedoch keine signifikanten Unterschiede bei den fetomaternalen
Aminosäurequotienten, also keine Veränderung der Transportdynamik zwischen den Gruppen.
Ein umgekehrter Effekt ist bei der Aminosäure Ornithin zu beobachten. Hier kommt es an Tag 16
zu einer signifikant reduzierten Konzentration in den Gruppen B und F sowohl im maternalen
Plasma als auch in der Amnionflüssigkeit. Am 20. Tag ist der Unterschied lediglich im maternalen
Plasma nachzuweisen. Hier zeigen sich in den Gruppen B und F gegenüber Gruppe K signifikant
erhöhte fetomaternale Ornithinquotienten. Es scheint also ab den 16. Tag zu einer Induktion des
fetomaternalen Aminosäuretransports in den Gruppen B und F gekommen zu sein, welcher die
reduzierte maternale Aminosäurekonzentration auf fetaler Seite weitestgehend ausgleicht.
87

Bei den Aminosäuren Cystein kommt es ab dem 16. Trächtigkeitstag zu einem starken Anstieg des
maternofetalen Aminosäuretransports in allen Gruppen. Der fetomaternale Aminosäurequotient
der Glutaminsäure ist hingegen ab dem 16. Tag kleiner 1. Bei den Aminosäure Lysin konnte keine
Dynamik des maternofetalen Transfers beobachtet werden.
Der von vielen Autoren (u.a. Battaglia und Regnault (2001) sowie Kedenburg und Mülling (1975))
beschriebene aktive diaplazentare Aminosäuretransport kann somit bestätigt werden. Aufgrund
fehlender bisheriger Arbeiten konnten jedoch keine Vergleichsdaten welche die Dynamik des
Transports beschreiben gefunden werden. Kedenburg und Mülling (1975) stellten fest, dass die
Aminosäurekonzentration in der menschlichen Amnionflüssigkeit gegen Ende der
Schwangerschaft geringer sei als im maternalen Plasma, während dies bei der Ratte umgekehrt
sei. Ein Vergleich der Dynamik des diaplazentaren Aminosäuretransportes ist daher nicht oder nur
sehr eingeschränkt möglich.
Die Vermutung von Kyuma (1984), dass der diaplazentare Aminosäuretransport überwiegend von
der fetalen Plasmakonzentration freier Aminosäuren abhängig ist und nicht von der maternalen
kann für einige Aminosäuren indirekt bestätigt werden. Bei den Aminosäuren Serin und Threonin
sind vermutlich kompensatorische Änderungen in der Transportdynamik zwischen den Gruppen
zu erkennen, durch welche Schwankungen im maternalen Plasma in der Amnionflüssigkeit
ausgeglichen werden, oder dies zumindest versucht wird. Es wird also versucht die amniale
Aminosäurekonzentration konstant zu halten. Daher liegt es nahe, dass die Regulation des
Transports von fetaler Seite aus stattfindet.
Nach Bennett und Jackson (1998) wird der fetale Bedarf an Glycin lediglich durch die
folatabhängige Umwandlung von Serin zu Glycin in der Plazenta gedeckt. In der vorliegenden
Arbeit kann jedoch in Gruppe F keine signifikante Reduktion der Glycinkonzentration in der
Amnionflüssigkeit gemessen werden, wie es zu erwarten gewesen wäre.
5.2 Histomorphologische und Immunhistochemische Untersuchungen
der Plazenten
Im Gegensatz zu Schiebler und Knoop (1959) welche Glykogen in den Riesenzellen lediglich in
Spuren nachweisen konnten, finden sich in fast allen Plazenten des 16. Trächtigkeitstages einzelne
z.T. aber auch mehrere Riesenzellen mit deutlichem PBA positivem, nicht diastaseresistentem
Zytoplasma.
88

Nach Schiebler und Knoop (1959) finden sich Glykogenzellen im Labyrinth in 5er oder 6er
Gruppen, wobei es auch zu Unter- sowie Überschreitungen dieser Zahl kommen kann. Entgegen
diesen Ergebnissen konnten im apikalen Labyrinth Zellgruppen von bis zu 30 glykogenhaltigen
Zellen gefunden werden.
Diese Beobachtungen sind jedoch nicht auf eine Vitaminrestriktion zurückzuführen, da sie in allen
Gruppen gemacht werden. Eine mögliche Erklärung hierfür könnte der unterschiedlicher
Zuchtstamm sein. Während Schiebler und Knoop (1959) mit Wistar-Ratten arbeiteten, wurden für
die vorliegende Arbeit Ratten des Zuchtstammes LEW.1W verwand.
Glykogenzellen innerhalb des Spongiotrophoblasten konnten in Übereinstimmung mit De Rijk
(2002) gefunden werden.
In Übereinstimmung mit De Rijk (2002) wurden Gruppen von Glykogenzellen ab dem 12. Tag im
Spongiotrophoblasten gefunden, welche sich im weiteren Verlauf der Trächtigkeit reduzierten. Die
Anzahl der Glykogenzellen ändert sich jedoch nicht nur im laufe der Trächtigkeit wie von De Rijk
(2002) beschrieben, sondern stellt sich zwischen den verschiedenen Tieren auch zum selben
Trächtigkeitstag als äußerst variabel dar. Das von De Rijk (2002) beschriebene kolloidale PAS-
positive Material welches den Platz der verlorenen Glykogenzellen einnimmt, konnte weder bei
Bouin noch bei Formalin fixierten Präparaten dargestellt werden. Auch mittels der PBA
Färbemethode konnte kein entsprechendes Material gefunden werden.
In einigen Plazenten zeigten sich jedoch nicht PAS bzw. PBA positive kolloidale Strukturen, der
Großteil des Platzes an welchem wahrscheinlich vormals Glykogenzellen zu finden waren, ist
jedoch leer.
Die von Schiebler und Knoop (1959) beschriebene basalmembranartige Interzellulärsubstanz des
Spongiotrophoblasten kann mit Hilfe der PBA-Färbetechnik, besonders gut an Bouin-fixiertem
Gewebe dargestellt werden.
Die in Gruppe B vermehrt zu beobachtenden Glykogeneinlagerungen um fetale Gefäße des
Labyrinths sind möglicherweise durch die Pyridoxalphosphat Abhängigkeit des Enzyms
Glykogenphosphorylase zu erklären, da die Glykogenphosphorlylase das für die Glykogenolyse
Geschwindigkeitsbestimmende Enzym ist (Löffer und Petrides, 1998, Livanova et al. 2002). Es
hat somit einen katabolen Charakter. Wird die Enzymaktivität durch Entzug eines Koenzyms
reduziert, könnte dies einen anabolen Effekt bewirken, indem es zu einer Reduktion der
Glykogenolyse kommt. Die Reduktion des Glykogenabbaus äußert sich im folgenden in einer
höheren Glykogenkonzentration im Labyrinth der Plazenta.
89

Die verringerte Anzahl Ki-67 positiver Zellen im Labyrinth der Plazenten der Gruppe F ist auf
eine verringerte Zellproliferation zurückzuführen. Als Überträger von 1-Kohlenstoffresten stellt
N10-Formyltetrahydrofolat die Kohlenstoffatome 2 und 8 des Purinkerns und ist somit unmittelbar
an der Bereitstellung der DNS-Basen Adenin und Guanin beteiligt. Auch bei der Bereitstellung der
Pyrimidinbase Thymin spielt die Folsäure eine wichtige Rolle. Bei der durch das Enzym
Thymidylatsynthase katalysierten Reaktion von Desoxyuridinmonophosphat zu
Desoxythymidinmonophosphat stellt N5,N10-Methylentetrahydrofolat einen Methylrest. Bei jeder
Verminderung des Folsäureangebots kommt es daher zu einer Störung der Thyminnucleotid-
Biosynthese und damit der DNS-Replikation. Eine Störung der DNS-Replikation wiederum
beeinträchtigt die Zellproliferation.
Die verringerte Zellproliferation in Gruppe B könnte, wie bereits erwähnt, mit einer verringerten
Aktivität der ASAT zu erklären sein, welche eine wichtige Rolle bei der Zellproliferation spielt.
Um eine semiquantitative Auswertung der Zellproliferation zu ermöglichen wurde die in 4.8
beschriebenen Auszählung Ki-67 positiver Zellen vorgenommen. Zur Vermeidung von
Verwechslungen zwischen Ki-67 positiven Trophoblastenzellen und fetalen Erythrozyten kann für
nachfolgende Arbeiten angeregt werden, andere Proliferationsmarker zu verwenden.
Der Nachweis apoptotischer Vorgänge mittels des Anti-ACTIVE® Caspase-3 Antikörper
stellte sich als problematisch dar. So sind die 14 Ereignisse in Gruppe B16 allein auf eine
Plazenta zurückzuführen. Hierbei liegt der Verdacht nahe, dass Methodische Fehler zugrunde
liegen.
Ziel der Arbeit mit dem Anti-Laminin Antikörpers ist der Nachweis von α-Laminin als Bestandteil
der Basallamina (Abb. 4.8.8) des fetalen Gefäßendothels. Hierdurch können Aussagen über die
Entwicklung des Gefäßsystemes der Plazenta getroffen werden. In diesem Zusammenhang sei
jedoch darauf hingewiesen, dass ein Vergleich der Basalmembranstärke des fetalen
Gefäßendothels lichtmikroskopisch aufgrund der ohnehin sehr dünnen Basalmembran nicht
möglich ist. Es besteht lediglich die Möglichkeit die Entwicklung des fetalen Gefäßsystems
aufgrund der Peroxidasereaktion abzuschätzen.
Als problematisch bei der Probengewinnung stellte sich die Deckung der Versuchstiere heraus.
Bei etwa 30 Tieren welche täglich gedeckt wurden, dauerte die Gewinnung des Probematerials
etwa 3 Monate. Diese Zeitspanne hätte lediglich durch eine größere Anzahl der zu bedeckenden
Tiere reduziert werden können.
90

Obwohl die Ratte als Versuchstier eine Reihe von bereits erwähnten Vorteilen aufweist (s.S.16),
können die Ergebnisse nicht bedingungslos auf den Menschen übertragen werden. Kedenburg und
Mülling (1975) vermuten, dass sich aktive Aminosäuretransportsysteme bei der Ratte nicht nur in
der Plazenta, sondern auch in der die Amnionhöhle umgebenden Dottersackwand befinden.
Des weiteren stellte sich die Menge der gewonnen Amnionflüssigkeit z.T. als zu gering heraus.
Während das Probevolumen der Amnionflüssigkeit ab dem 14. Tag für die Aminosäureanalytik
ausreichend war, war dies am 12. Trächtigkeitstag teilweise nur durch eine Sammelprobe mehrerer
Tiere erreicht worden. Daher konnten die Ergebnisse der am 12. Tag gewonnenen Proben nicht in
die vorliegende Arbeit mit einbezogen werden. Auch kann vermutet werden, dass in einem
größeren Tierversuch weitere signifikante Unterschiede bezüglich der Aminosäurekonzentrationen
zu finden gewesen wären, da einige Ergebnisse mit einer Irrtumswahrscheinlichkeit zwischen
p=0,05 und p=0,06 auftraten.
Die Bestimmung von Homocystein in der Amnionflüssigkeit wäre gerade vor dem Hintergrund
der Verstoffwechselung von Homocystein in der Plazenta interessant gewesen da vermutet wird,
dass überwiegend eine Remethylierung des Homocysteins zu Methionin stattfindet, und der
Abbau zu Cystein lediglich in einem deutlich geringerem Umfang stattfindet (Solanky et al.
2010). Dies würde bedeuten, dass der plazentare Homocysteinstoffwechsel überwiegend Folsäure
abhängig und nicht Vitamin-B6 abhängig ist. Inwieweit der plazentare Homocysteinsoffwechsel
Auswirkungen auf die Homocysteinkonzentration in der Amnionflüssigkeit hat, und ob im Falle
einer Folsäurerestriktion der Abbau zu Cystein induziert wird um dem Homocysteinspiegel
möglichst niedrig zu halten, wären interessante Fragen für die Zukunft.
Auch für während der Arbeit neu aufgetretene Fragestellung war das Probevolumen zu gering.
So wäre die Bestimmung von Xanturensäure in der Amnionflüssigkeit interessant gewesen, da
diese einen Komplex mit Insulin bildet und dessen biologische Verfügbarkeit deutlich herabsetzt
(Kotake et al. 1968). Xanturensäure fällt bei einem Vitamin-B6-Mangel vermehrt an, und es ist
aufgefallen das einige Feten der Gruppe B deutlich makrosom waren, welches auf einen
Schwangerschaftsdiabetes hinweisen könnte.
Des weiteren wurden Gefrierpräparate der Plazenten angefertigt um Untersuchungen
durchzuführen, welche an den Formalin bzw. Bouin fixierten Präparaten nicht möglich sind. Zu
nennen ist hier beispielsweise der Nachweis von Lipiden mittels einer Sudan-Färbung. Dieses
wäre zwar grundsätzlich möglich gewesen, jedoch stellte sich der Erhalt des Gewebes als schlecht
heraus. Als Grund hierfür wird die Probengewinnung vermutet, bei welcher die Plazenten auf
Trockeneis gelegt wurden. Mitunter hätte eine vorherige Fixierung in flüssigem Stickstoff hier
Abhilfe schaffen können.
91

In der Diskussion der Ergebnisse stellte sich die vorhandene Literatur insbesondere bezüglich der
Aminosäureanalytik in der Amnionflüssigkeit als äußerst spärlich heraus, welches die Einordnung
der eigenen Ergebnisse erschwerte.
6. Literaturupdate und Ausblick
Die praktische Durchführung der Arbeit erfolgte im Jahr 2007. Im folgenden soll auf
zwischenzeitliche Entwicklungen in der Literatur eingegangen werden.
Franke et al. (2009) konnten einen genetischen Polymorphismus nachweisen, welcher mit einem
geringeren Risiko für Spina bifida assoziiert ist. Grund hierfür ist ein erhöhter Vitamin-B12
Spiegel und ein wahrscheinlich hieraus resultierender erhöhter Folsäurespiegel in den
Erythrozyten. Das entsprechende Gen CUBN kodiert ein Intrinsic-Faktor-Cobalamin Rezeptor.
Um das Zusammenspiel der über 15 Transportproteine für Aminosäuren in der Plazenta besser
abschätzen zu können, entwickelten Sengers et al. (2010) ein mathematisches Modell, welches die
Zusammenarbeit der Transportproteine als System berechnet.
Solanky et al. (2010) konnten nachweisen, das die mRNA welche die Enzyme Methionin-
Synthase und 5,10-Methylentetrahydrofolatreduktase kodiert in der menschlichen Plazenta
zwischen dem ersten Trimenon und dem Ende der Schwangerschaft in ähnlicher Menge
exprimiert werden wie in der Leber. Die mRNA für die Cystathionin-beta-Synthase ist im
Vergleich zur Leber in einer deutlich geringeren Menge zu finden. Dies deutet daraufhin, das
Homocystein in der Plazenta überwiegend mit Hilfe von 5,10-Methylentetrahydrofolat zu
Methionin remethyliert wird, und nicht zu Cystein abgebaut wird. Diese Ergebnisse weisen auf die
Wichtigkeit der perikonzeptionellen Folsäuresubstitution hin.
Der diaplazentare Homocysteintransport findet mit hilfe der Transportsysteme L, A und y(+)L
statt, wie Tsitsiou et al. 2009 zeigen konnten.
Jansson (2009) stellt in einem Review fest, dass mehrere Arbeiten einen beeinträchtigten
diaplazentaren Aminosäuretransport bei einem erhöhten maternalen Homocysteinspiegel zeigen.
Einige Arbeiten zeigen diesen Effekt vorwiegend bei essentiellen Aminosäuren. Vermutet wird
hier u.a. eine kompetitive Hemmung der Transportproteine durch Homocystein.
Weitere Fragen die sich aus dieser Arbeit ergeben haben sind der bereits erwähnte
Homocysteinstoffwechsel in der Plazenta, insbesondere in der Gruppe F. Wenn überwiegend eine
Remethylierung von Homocystein zu Methionin erfolgt, stellt sich die Frage, in welchem Maße
92

die Feten der Gruppe F im Vergleich zu Gruppe B bzw. auch K einer erhöhten
Homocysteinkonzentration ausgesetzt sind. Welche Rolle spielt also der plazentare
Homocysteinstoffwechsel für dessen Konzentration in der Amnionflüssigkeit?
Auch die Vermutung dass ein Vitamin-B6 Mangel durch Bindung des Insulins an Xanthurensäure
während der Trächtigkeit, eine Glukosetolleranzstörung hervorruft gilt es zu untersuchen. Hierbei
wäre nicht nur die Glukosekonzentration im maternalen Plasma sowie der Amnionflüssigkeit
interessant, sondern auch die Menge an Xanthurensäure um dem Pathomechanismus
nachvollziehen zu können.
Okada et al. (1991) konnten an Ratten zeigen, dass bei einem Vitamin-B6 Mangel im M.
Gastrocnemius und im Herz die Aktivität der Glykogenphosphorylase signifikant reduziert ist.
Auch konnte in diesen Geweben vermehrt Glykogen nachgewiesen werden. In der Leber konnten
diese Veränderungen jedoch nicht beobachtet werden. Folglich kann vermutet werden das es bei
einem Vitamin-B6 Mangel auch in der Plazenta zu einer Hemmung der Glykogenphosphorylase
kommt, welche für das Vermehrte Auftreten PBA positiver Strukturen verantwortlich ist. Zum
Nachweis wären jedoch organspezifische Enzymaktivitätsbestimmungen notwendig.
Welche Rolle der Vitaminmangel der trächtigen Ratte auf den Stoffwechsel der Neugeborenen hat
ist ebenfalls weiterhin unklar.
Auch die Frage ob es einen kausalen Zusammenhang zwischen den gemessenen Unterschieden
der Aminosäurekonzentration in der Amnionflüssigkeit und der Pathogenese der bekannten
Fehlbildungen gibt, gilt es noch zu klären, da dies zum gegenwärtigen Zeitpunkt zwar
wahrscheinlich scheint, jedoch nicht sicher nachgewiesen werden konnte. Dies gilt auch für die
Frage, inwieweit die morphologischen Veränderungen an der Plazenta für die veränderten
Aminosäurekonzentrationen verantwortlich sind.
Auch die zugrundeliegenden Regulationsmechanismen für den diaplazentaren
Aminosäuretransport, insbesondere wenn es zu scheinbar kompensatorischen Änderungen in der
Transportkinetik kommt, sind noch nicht hinreichend bekannt.
93

7. Zusammenfassung
Ziel der Arbeit war es, den diaplazentaren Aminosäuretransport trächtiger LEW.1A-Ratten
während eines induzierten Folsäure- oder Vitamin-B6-Mangel zu quantifizieren, sowie
histomorphologische Reaktionen der Plazenten zu erfassen.
Von beiden Vitaminen ist bekannt, dass am Menschen peri- bzw. postkonzeptionelle
Hypovitaminosen zu schweren Fehlbildungen wie z.B. Reifungsstörungen des ZNS und zu
Spaltenbildungen im Mund-Kiefer-Gesichtsbereich führen können.
Zu diesem Zweck wurden die Ratten in die Gruppen K= Kontrolle, B= induzierter Vitamin-B6-
Mangel und F= induzierter Folsäuremangel eingeteilt. Die Tiere wurden ab dem Tag der
Konzeption mit entsprechendem vitaminfreien Futter versorgt. Die Probenentnahme fand an den
Tagen 14, 16 und 20 der Trächtigkeit statt, es wurden Blut und Amnionflüssigkeit sowie Plazenten
entnommen.
Neben den Vitaminspiegeln des maternalen Plasmas welche, den Vitaminstatus der Tiere
anzeigen, wurden auch indirekte Parameter wie der Homocysteinspiegel, die Aktivität der Vitamin
B6-abhängigen Enzyme ASAT und ALAT im Plasma erfasst, um eine Aussage über den
funktionellen Vitaminstatus der Tiere geben zu können. In Gruppe B ist zwischen dem 14. und 20.
Tag im maternalen Plasma eine signifikant reduzierte Vitamin-B6-Konzentration gegenüber
Gruppe K gemessen worden. Im Gegensatz hierzu ist in Gruppe F am 14. und 16. Tag eine
signifikant höhere Vitamin-B6-Konzentration gemessen worden als in Gruppe K. In Gruppe F ist
zwischen dem 14. und 20. Tag eine signifikant reduziert Folsäurekonzentration in maternalen
Plasma gemessen worden. Die Homocysteinkonzentration in den Gruppen B und F ist an den
Tagen 14 und 16 gegenüber Gruppe K signifikant erhöht. Nachdem es am 14. Tag lediglich in
Gruppe B zu einer signifikanten Reduktion der Enzymaktivität der ALAT kommt, ist diese an den
Tagen 16 und 20 sowohl in Gruppe B als auch in Gruppe F zu messen. Die Aktivität der ASAT am
16. Tag ist in Gruppe B gegenüber den anderen Gruppen signifikant reduziert, am 20. Tag ist die
Aktivität in Gruppe F gegenüber Gruppe K signifikant reduziert.
Des weiteren wurden die Aminosäurekonzentrationen sowohl im maternalen Plasma als auch in
der Amnionflüssigkeit bestimmt und ein feto-maternaler Aminosäure-Quotient errechnet, welcher
Auskunft über den aktiven diaplazentaren Transport gibt. Dabei zeigte sich, dass der aktive
Aminosäuretransport während der Trächtigkeit einer zeitlichen Dynamik unterliegt.
94

Zur Interpretation der diaplazentaren Transportprozesse ist dieser entwickelte Quotienten nützlich.
Hiermit konnte gezeigt werden, dass der Graph der Quotienten während der Trächtigkeit einen
parabelförmigen Verlauf nimmt.
Demnach greift ein Vitamin B6-Mangel massiv in den Aminosäure Stoffwechsel und damit auch
in den diaplazentaren Transport ein. Es zeigen sich im Verlaufe der Trächtigkeit höhere
Quotienten der Gruppe B bezüglich der Aminosäuren Glycin, Citrullin und Ornithin im Vergleich
zur Gruppe K. Andererseits fiel der Quotient für die Aminosäuren Threonin und Cystathionin in
der Gruppe B signifikant im Vergleich zur Gruppe K.
Ein Folsäuremangel bewirkt bezüglich des diaplazentaren Transportes im Vergleich zur Gruppe
K folgende Ergebnisse: Der Quotient für die Aminosäuren Asparaginsäure, Glutamin, Glycin und
Ornithin nimmt im Laufe der Trächtigkeit signifikant zu. Dagegen sind die Quotienten der Gruppe
F für die Aminosäuren Threonin, Serin, Asparaginsäure, Glutaminsäure, Prolin, Butyrat,
Methionin, Isoleuzin, Histidin und Arginin im Vergleich zur Gruppe K signifikant erniedrigt.
Schlußfolgernd kann eine Dynamik der maternalen sowie der amnialen
Aminosäurekonzentrationen sowie deren Quotienten beschrieben werden. Zur Einordnung dieser
Ergebnisse ist zu konstatieren, dass sowohl ein Folsäure- als auch Vitamin B6-Mangel zu
massiven Störungen des diaplazentaren Aminosäuretransportes führen.
Auch bei den histomorphologischen Untersuchungen der Plazenten können Unterschiede
beschrieben werden. So sind bei den Bouin fixierten Präparaten in der PBA Färbung in der
Gruppe B dichtere PBA positive Strukturen um größere fetale Gefäße zu finden als in Gruppe K.
Auch sind in Formalin fixierten Präparaten in Gruppe B vermehrt Glykogeneinlagerungen um
kleine fetale Gefäße im Labyrinth zu finden. Eine mögliche Erklärung hierfür ist die Vitamin-B6
Abhängigkeit des Enzyms Glykogenphosphorylase, welches eine wichtige Rolle bei der
Glykogenolyse spielt.
Möglicherweise stellen die in der Gruppe B übermäßigen Glykogeneinlagerungen um die fetalen
Gefäßwände des Labyrinths und die Verstärkungen der Basallamina des Amnionepithels eine
Diffusionsbarriere dar. Des weiteren sind in allen Gruppen größere Ansammlungen von
Glykogenzellen im apikalen Labyrinth zu finden als in der Literatur beschrieben, womöglich sind
diese Unterschiede tierstammspezifisch begründet.
Diese morphologischen Veränderungen haben vermutlich einen funktionellen Einfluß auf die
Versorgung der Feten und erklären 20% mehr Resorptionen der Früchte in der Gruppe B
gegenüber den Gruppen K und F. Zudem zeigt sich zwischen Spongiotrophoblast und Labyrinth
der Gruppe B eine stärkere granulozytäre Infiltration als in den übrigen Gruppen.
95

Der Vitamin B6-Mangel an der trächtigen Ratte induziert stärkere negative Einflüsse auf die
Plazento- und Fetogenese als ein Folsäuremangel. Einige Feten der Gruppe B zeigten starke
Übergewichtigkeit. Dies deutet auf einen Schwangerschaftsdiabetes hin, welcher durch die
Bindung des Insulins an Xanthurensäure induziert sein könnte. Hierzu sind weitere
Untersuchungen nötig.
Die Ergebnisse dieser Arbeit untermauern die Bedeutung der perikonzeptionellen Folsäure- und
Vitamin B6-Substitution.
Mit dem Nachweis veränderter Aminosäurekonzentrationen in der Amnionflüssigkeit bei Vitamin-
B6- oder Folsäuremangel während der Trächtigkeit konnte ein weiterer zu berücksichtigender
Baustein in der Pathogenese von ZNS Reifungsstörungen sowie Spaltenbildungen erbracht
werden.
Die scheinbar kompensatorische Reaktion der Plazenta auf veränderte maternale
Aminosäurekonzentrationen bzw. einen beeinträchtigten Aminosäurestoffwechsel, welche bei
einigen Aminosäuren zu beobachten ist, mag einen Impuls für weiterführende Arbeiten geben.
96

8. Abkürzungsverzeichnis:
ALAT: Alanin-Aminotransferase = GPT: Glutamat-Pyruvat-Transaminase
ASAT: Aspartat-Aminotransferase = GOT: Glutamat-Oxalacetat-Transaminase
BrdU: 5-Bromodeoxyuridin
CBS: Cystathion β-Synthase
DAB: 3,3´-Diaminobenzidin
dTMP: Desoxythyminmonophosphat
dUMP: Desoxyuridinmonophosphat
EAST: erythrozytäre Aspartat-Aminotransferase
EtOH: Ethylalkohol
FABP: Folsäure bindendes Protein
HE: Hämatoxylin-Eosin
LKGS: Lippen-Kiefer-Gaumen-Segel-Spalte
MTHFR: Methylentetrahydrofolatreduktase
ND: Neuralrohrdefekt
PAS: Periodic-acid-Schiff-reaction
PBA: Perjodsäure-Bisulfit-Aldehydthionin
PFRs: high-affinity membrane-associated placental folate receptors
PBS: Phosphate Buffer Solution
THF: Tetrahydrofolsäure
97

9. Schrifttum
Ambrosi P, Rolland P, Garçon D:
Homocysteine, a risk factor of atherosclerosis.
Arch Mal Coeur Vaiss 89; 12: 1667-1671, 1996.
Aubard Y, Darodes N, Cantaloube M, Aubard V, Diallo D, Teissier MP:
Hyperhomocysteinemia and pregnancy: a dangerous association.
J Gynecol Obstet Biol Reprod 29; 4: 363-372, 2000.
Alton-Mackey MG, Walker BL:
Graded levels of pyridoxine in the rat diet during gestation and the physical and neuromotor
development of offspring. Am J Clin Nutr 26: 420-428, 1973.
Bässler H, Golly I, Loew D, Pietrzik K:
Vitamin-Lexikon für Ärzte, Apotheker und Ernährungswissenschaftler. 2.Auflage.München/Jena,
Verlag Urban & Fischer, 1997.
Baker H, Frank O, Deangelis B, Feingold S, Kaminetzky HA:
Role of placenta in maternal-fetal vitamin transfer in humans.
Am J Obstet Gynecol 141; 7: 792-796, 1981.
Battaglia FC, Regnault TRH:
Placental transport and metabolism of amino acids. Placenta 22: 145-161, 2001.
Beckman DA, Brent RL, Lloyd JB:
Leucine source for the rat fetus. Placenta 18; 1: 79-82, 1997.
Beitz R, Mensink GBM, Fischer B, Thamm M:
Vitamins- dietary intake and intake from dietary supplements in Germany.
Eur J Clin Nutr 56: 539-545, 2002.
98

Bennett FI, Jackson AA:
Glycine is not formed through the amino transferase reaction in human or rat placenta.
Placenta 19; 4: 329-331, 1998.
Blacher J, Montalescot G, Ankri A, Chadefaux-Vekemans B, Benzidia R, Grosgogeat Y, Kamoun
P, Thomas D:
Hyperhomocysteinemia in coronary artery diseases. Apropos of a study on 102 patients
Arch Mal Coeur Vaiss 89; 10: 1241-1246, 1996.
Bønaa KH, Njølstad I, Ueland PM, Schirmer H, Tverdal A, Steigen T, Wang H, Nordrehaug JE,
Arnesen E, Rasmussen K; NORVIT Trial Investigators:
Homocysteine lowering and cardiovascular events after acute myocardial infarction.
N Engl J Med 354; 15: 1578-1588, 2006.
Boushey CJ, Beresford SA, Omenn GS, Motulsky AG:
A quantitative assessment of plasma homocysteine as a risk factor for vascular disease. Probable
benefits of increasing folic acid intakes. JAMA 274; 13: 1049-57, 1995.
Bridgman J:
Morphological study of development of the placenta of the rat. I. An outline of the development of
the placenta of the white rat. J Morphol 83; 1: 61-85, 1948 a.
Bridgman J:
A morphological study of development of the placenta of the rat. II. a histological and cytological
study of the development of the chorio-allantoic placenta of the white rat. J Morphol 83; 2: 195-
224, 1948 b.
Buddecke E:
Grundriss der Biochemie. 9. Aufl. de Gruyter, Berlin, New York, 383-384, 1994.
Butterworth CE, Bendich A:
Folic acid and the prevention of birth defects. Annu Rev Nurr 16: 73-97, 1996.
99

Butterworth CE, Baugh CM Jr, Krumdieck C:
A study of folate absorption and metabolism in man utilizing carbon-14-labeled polyglutamates
synthesized by the solid phase method.
J Clin Invest 48: 1131-1142, 1969.
Caltrider PG, Niss HF:
Role of methionine in cephalosporin synthesis. Appl Microbiol 14; 5: 746-753, 1996.
Carter AM:
Animal models of human placentation - a review.
Placenta 28, Suppl A, Trophoblast Research 21: 41-47, 2007.
Cetin I:
Amino acid interconversions in the fetal-placental unit: the animal model and human studies in
vivo. Pediatr Res 49; 2: 148-154, 2001.
Clarke R, Brattström L, Landgren F, Israelsson B, Lindgren A, Hultberg B, Andersson A, Cuskelly
G, McNulty H, Strain SS, McPartlin J, Weir DG, Scott JM, Brouwer IA, Blom HJ, Bos GMJ,
Spaans A, Rosendaal FR, Thomas CMG, Haak HL, Wijermans PW, Gerrits WBJ, Joosten E,
Riezler R, Stabler SP, Allen RH, Lindenbaum J, Pietrzik K, PrinzLangenohl R, Dierkes J, Mason
JB, Jacques P, Selhub J, Salem D, Schaefer E, Rosenberg IH, van der Mere A, Vermack WJH,
Delport R, Becker PJ, Potgieter HC, Yarnell JWG, Young IS, McCrum EE, Patterson SS, Gey KF,
Evans AE:
Homocysteine lowering blood homocysteine with folic acid based supplements: meta-analysis of
randomised trials. BMJ 316: 894–898, 1998.
Cleary RE, Lumeng L, Li TK:
Maternal and fetal plasma levels of pyridoxal phosphate at term: adequacy of vitamin B6
supplementation during pregnancy. Am J Obstet Gynecol 121; 1: 25-28, 1975.
Contractor SF, Shane B:
Metabolism of [C14] pyridoxol in the pregnant rat. Biochim Biophys Acta 230: 127-136, 1971.
100

Croy BA, Kiso Y:
Granulated metrial gland cells: a natural killer cell subset of the pregnant murine uterus.
Microsc Res Tech 25; 3: 189-200, 1993.
Czeizel AE, Dudás I, Métneki J:
Pregnancy outcomes in a randomised controlled trial of periconceptional multivitamin
supplementation- Final report. Arch Gynecol Obstet 255: 131-139, 1994.
Czeizel AE:
Periconceptional folic acid containing multivitamin supplementation.
Eur J Obstet Gynecol Reprod Biol 78: 151-161, 1998.
Czeizel AE, Dobó M, Vargha P:
Hungarian cohort-controlled trial of periconceptional multivitamin supplementation shows a
reduction in certain congenital abnormalities.
Birth Defects Res A Clin Mol Teratol 70: 853-861, 2004.
Czeizel AE:
The primary prevention of birth defects: Multivitamins or folic acid?
Int. J. Med. Sci. 1; 1: 50-61, 2004.
Da Costa M, Rothenberg SP:
Purification and characterization of folate binding proteins from rat placenta.
Biochim Biophys Acta 1292; 1: 23-30, 1996.
Dallenbach-Hellweg G, Battista JV, Dallenbach FD:
Immunohistological and histochemical localization of relaxin in the metrial gland of the pregnant
rat. Am J Anat 117; 3: 433-450, 1965.
Davies J, Glasser SR:
Histological and fine structural observations on the placenta of the rat.
Acta Anat 69: 542-574, 1968.
101

Deane HW, Rubin BL, Driks EC, Lobel BL, Leipsner G:
Trophoblastic giant cells in placentas of rats and mice and their probable role in steroid-hormone
production. Endocrinology 70: 407-419, 1962.
Deris I:
Hyperhomocysteinämie, Vitaminsubstitution wird empfohlen.
Deutsches Ärzteblatt 98; 45: A2981, 2001.
DGE (Deutsche Gesellschaft für Ernährung), ÖGE (Österreichische Gesellschaft für Ernährung),
(SGE) Schweizerische Gesellschaft für Ernährung, (SVE) Schweizerische
Gesellschaft für Ernährungsforschung: Referenzwerte für die Nährstoffzufuhr.
Umschau Braus Verlag, 1. Auflage, Frankfurt/Main: 118, 2000.
Dierkes J, Kroesen M, Pietrzik K:
Folic acid and vitamin B6 supplementation and plasma homocysteine concentrations in healthy
young women. Int J Vitam Nutr Res 68; 2: 98-103, 1998.
Dilley A, Hooper WC, El-Jamil M, Renshaw M, Wenger NK, Evatt BL:
Mutations in the genes regulating methylene tetrahydrofolate reductase (MTHFR C→T677) and
cystathione β-synthase (CBS G→A919, CBS T→C833) are not associated with myocardial
infarction in african americans. Thromb Res 103: 109–115, 2001.
Di Simone N, Riccardi P, Maggiano N, Piacentani A, D`Asta M, Capelli A, Caruso A:
Effect of folic acid on homocysteine-induced trophoblast apoptosis.
Mol Hum Reprod 10; 9: 665-669, 2004.
Dixon JS, Bulmer D:
Fine structure of cells in the rat metrial gland. J Anat 108; 1: 123-133, 1971.
Durst-Zivkovic B:
Quantitative aspects of structural changes in chorioallantoic placenta of the rat during its
development. Experimentia 15; 34: 1343-1345, 1978.
102

Ellington SKL:
A morphological study of the development of the chorion of rat embryos.
J Anat 150: 247-263, 1987.
Enders AC, Schlafke S:
A morphological analysis of the early implantation stages in the rat.
Am J Anat 120: 185-226, 1967.
Eskes TK:
Folates and the fetus. Eur J Obstet Gynecol Reprod Biol. 71; 2: 105-111, 1997.
Eskes TK:
Neural tube defects, vitamins and homocysteine. Eur J Pediatr. 157; 2: 139-141, 1998.
Eskes TK:
Homocysteine and human reproduction. Clin Exp Obstet Gynecol 27; 3-4: 157-67, 2000.
Eskes TK:
Clotting disorders and placental abruption: homocysteine –a new risk factor.
Eur J Obstet Gynecol Reprod Biol 95; 2: 206-212, 2001.
Essien FB, Wannberg SL:
Methionine but not folic acid or vitamin B-12 alters the frequency of neural tube defects in Axd
mutant mice. J Nutr 123: 27-34, 1993.
Fisher SE, Inselman LS, Duffy L, Atkinson M, Spencer H, Chang B:
Ethanol and fetal nutrition: effect of chronic ethanol exposure on rat placental growth and
membrane-asociated folic acid receptor binding activity.
J Pediart Gastroenterol Nutr 4: 645-649, 1985.
103

Franke B, Vermeulen SH, Steegers-Theunissen RP, Coenen MJ, Schijvenaars MM, Scheffler H,
den Heijer M, Blom HJ:
An association study of 45 folate-related genes in spina bifida: Involvement of cubilin (CUBN)
and tRNA aspartic acid methyltransferase 1 (TRDMT1).
Birth Defects Res A Clin Mol Teratol 85; 3: 216-226, 2009.
Franke H:
Feinstruktur der Plazenta, Elektronenoptische Untersuchungen über die reifende und reife
Plazenta der Ratte. VEB Gustav Fischer Verlag Jena, 1969.
Furth-Walker D, Leibman D, Smolen A:
Changes in pyridoxal phosphate and pyridoxamine phosphate in blood, liver and brain in the
pregnant mouse. J Nutr 119; 5: 750-756, 1989.
Gerster H:
Die Bedeutung von Vitamin B6 für die Entwicklung des Säuglings, humanmedizinische und
tierexperimentelle Studien. Z Ernährungswiss 35; 4: 309-317, 1996.
Goddijn-Wessel TAW, Wouters MGAJ, Molen EF , Spuijbroek MDEH, Steegers-Theunissen
RPM, Blom HJ, Boers GHJ, Eskes TKAB:
Hyperhomocysteinemia: a risk factor for placental abruption or infarction.
Eur J Obstet Gynecol Reprod Biol 66: 23-29, 1996.
Gratzner HG:
Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: a new reagent for detection of DNA
replication. Science 218; 4571: 474-475, 1982.
Guilarte TR:
Vitamin B6 and cognitive development: recent research findings from human and animal studies.
Nutr Rev 51; 7: 193-198, 1993.
Gurekian CN, Koski KG:
Amniotic fluid amino acid concentrations are modified by maternal dietary glucose gestational age
and fetal growth in rats. J Nutr 135; 9: 2219-2224, 2005.
104

Hall MH:
Folic acid deficiency and congenital malformation.
J Obstet Gynaecol Br Commonw 79; 2: 159-161, 1972.
Hall J, Solehdin F:
Folic acid for the prevention of congenital anomalies. Eur J Pediatr 157: 445-450, 1998.
Hartridge T, Illing HM, Sandy JR:
The role of folic acid in oral clefting. Br J Orthod 26: 115-120, 1999.
Haugland RP, You WW:
Immunochemical Protocols. 2. Auflage, Totowa, New Jersey, Humana Press: 173-183, 1998.
Hayes C, Werler MM, Willett WC, Mitchell AA:
case-control study of periconceptional folic acid supplementation and oral clefts.
Am J Epidemiol 143; 12: 1229-1934, 1996.
Henderson GI, Perez T, Schenker S, Mackins J, Antony AC:
Maternal-to-fetal transfer of 5-methyltetrahydrofolate by the perfused human placental cotyledon:
evidence for a concentrative role by placental folate receptors in fetal folate delivery. J Lab Clin
Med 126; 2: 184-203, 1995.
Hillig U:
Lippen-Kiefer-Gaumen-Spalten: Klassifikation und Epidemiologie.
Fortschr Kieferorthop 52: 230-236, 1991.
Hsu SM, Raine L, Fanger H:
Use of Avidin-Biotin-Peroxidase Complex (ABC) in immunoperoxidase techniques.
J Histochem Cytochem 29; 4: 577-580, 1981.
Institute of Medicine:
Dietary reference intakes for thiamine, riboflavin, niacin, vitamine B6, folate, vitamine B12,
pantothenic acid biotin, and choline. National Academy Press, Washington, D.C.: 199, 1998.
105

Jansson T:
Novel mechanism causing restricted fetal growth: does maternal homocysteine impair placental
amino acid transport? J Physiol 1; 587: 4123, 2009.
Jollie WP:
Fine structural changes in placental labyrinth of the rat with increasing gestational age.
J Ultrastruct Res 10: 27-47, 1964.
Jollie WP:
Fine structural changes in the junctional zone of the rat placenta with increasing gestational age. J
Ultrastruct Res 12: 420-438, 1965.
Jung K, Ohlrich B, Mildner D, Zubek A, Schimmelpfennig W, Egger E:
The apoenzyme of aspartate aminotransferase and alanine aminotransferase in the serum of
healthy persons and patients suffering from liver diseases.
Clin Chim Acta 90; 2: 143-149, 1978.
Kälin Aebi, A:
Vitamin B6-Stoffwechsel bei Nierensteinpatienten und gesunden im Vergleich.
Dissertationsschrift Universität Basel, 2006.
Källen B:
A prospective study of some aetiological factors for limb reduction in Sweden.
J Epidemiol Community Health 43: 31-36, 1989.
Kaloglu C, Gursoy E, Onarlioglu B:
Early maternal changes contributing to the formation of the chorioallantoic and yolk sac placentas
in rat: a morphological study. Anat Histol Embryol 21; 4: 200-2006, 2003.
Kedenburg CP, Mülling M:
Freie Aminosäuren im Fruchtwasser bei verschiedenen Placentatypen.
Zbl Vet Med, Reihe A 22; 7: 549-555, 1975.
106

Kerrigan JJ, Mansell JP, Sengupta A, Brown N, Sandy JR:
Palatogenesis and potential mechanisms for clefting.
J R Coll Surg Edinb 45; 6: 351-358, 2000.
Kirchgeßner M, Reithmayer F, Roth-Maier DA:
Effect of varying vitamin B6 intake levels on the vitamin B6 status of fetuses and reproductive
organs. Ann Nutr Metab 29; 3: 138-146, 1985.
Koletzko B, von Kries R:
Prävention von Neuralrohrdefekten durch Folsäurezufuhr in der Frühschwangerschaft.
Der Kinderarzt 26; 2: 187-190, 1995.
Koletzko B, Pietrzik K:
Gesundheitliche Bedeutung der Folsäurezufuhr.
Deutsches Ärzteblatt 101; 23: A1670-A1681, A1-A3, 2004.
Kotake Y, Sotokawa T, Murakami E, Histake A, Abe M, Ikeda Y:
Studies on the Xanthurenic Acid-Insulin Complex. J Biochem 63; 5: 578-581, 1968.
Krumdieck CL, Fukushima K, Fukushima T, Shiota T, Butterworth CR Jr:
A long-term study of the excretion of folate in pterins in a human subject after ingestion of C14
folic acid, with observation on the effect of diphenylhydatoin administration.
Am J Clin Nutr 31; 1: 88-93, 1978.
Kyuma M:
Amino acid transfer at the fetal surface of the trophoblast and the characterization of its carrier in
the rat placenta in vivo. Nippon Sanka Fujinka Gakkai Zasshi 36; 3: 405-414, 1984.
Lakshmi AV, Maniprabha C, Krishna TP:
Plasma homocysteine level in relation to folate and vitamin B6 status in apparently normal men.
Asia Pac J Clin Nutr. 10; 3: 194-196, 2001.
107

Leibman D, Furth-Walker D, Smolen A:
Vitamin B-6 metabolic enzymes in blood and placenta of pregnant mice.
J Nutr 120; 2: 178-184, 1990.
Li Y, Li Z, Chen X, Qi P:
Homocysteine-induced neural tube defects in chick embryos and protection of folic acid.
Wei Sheng Yan Jiu 27; 6: 372-376, 1998.
Lines DR, Waisman HA:
Placental transport of phenylalanine in the rat: maternal and fetal metabolism.
Proc Soc Exp Biol Med 136; 3: 790-793, 1971.
Litwer G:
Über die Sekretion und Resorption in dem Dotterentoderm Zellen bei graviden weißen Mäusen.
Z Zellforsch Mikrosk Anat 8; 1: 135-152, 1928.
Livanova NB, Chebotareva NA, Eronina TB, Kurganov BI:
Pyridoxal5´-phosphate as a catalytic conformational cofactor of muscle glycogen phosphorylase
B. Biochemistry (Mosc.) 67; 10: 1089-1098, 2002.
Löffler G, Petrides PE:
Biochemie und Pathobiochemie. Springer Verlag, 6. Auflage, Berlin, 1998.
Lonn E, Yusuf S, Arnold MJ, Sheridan P, Pogue J, Micks M, McQueen MJ, Probstfield J, Fodor
G, Held C, Genest J, Heart Outcomes Prevention Evaluation (HOPE) 2 Investigators:
Homocysteine lowering with folic acid and B vitamins in vascular disease.
N Engl J Med 354; 15: 1567-1577, 2006.
Loy GL, Quick AN, Hay WW, Meschia G, Battaglia FC, Fennessey PV:
Fetoplacental deamination and decarboxylation of leucine.
Am J Physiol 259; 4 Pt 1: E492-497, 1990.
108

Lyon JB, Bain JA, Williams HL:
The distribution of vitamin B6 in the tissues of two inbred strains of mice fed complete and
vitamin B6-deficient rations J Biol Chem 237 ;6: 1989-1991, 1962.
Malandro MS, Beveridge MJ, Kilberg MS, Novak DA:
Effect of low-protein diet-induced intrauterine growth retradation on rat placental amino acid
transport. Am J Physiol 271; 1: C295-303, 1996.
Malinow MR, Bostom AG, Krauss RM:
Homocyst(e)ine, diet, and cardiovascular diseases: a statement for healthcare professionals from
the Nutrition Committee, American Heart Association. Circulation 5-12; 99: 178-182, 1999.
Marathe MR, Thomas GP:
Effects of pyridoxine on implantation and pregnancy in Wistar rat.
Toxicol Lett 36; 2-3: 159-164, 1987.
Merker HJ, Villegas H:
Elektronenmikroskopische Untersuchungen zum Problem des Stoffaustausches zwischen Mutter
und Keim bei Rattenembryonen des Tages 7-10.
Anat Embryol 131; 4: 325-346, 1970.
Metz J, Heinrich D, Forssmann WG:
Ultrastructure of the labyrinth in the rat full-term placenta.
Anat Embryol 149: 123-148, 1976.
Montalescot G , Ankri A, Chadefaux-Vekemans B, Blacher J, Philippe F, Drobinski G, Benzidia R,
Kamoun P, Thomas D:
Plasma homocysteine and the extent of atherosclerosis in patients with coronary artery disease.
Int J Cardiol 60: 295-300, 1997.
MRC Vitamin Study Research Group:
Prevention of neural tube defects: Results of the Medical Research Council Vitamin Study.
Lancet 338: 131-137, 1991.
109

Mulholland J, Aplin JD, Ayad S, Hong L, Glasser SR:
Loss of collagen type VI from rat endometrial stroma during decidualization.
Biol Reprod 46: 1136-1143, 1992.
Mutschler E, Geisslinger G, Kroemer HK, Ruth P, Schäfer-Korting M:
Arzneimittelwirkungen kompakt. Wissenschaftliche Verlagsgesellschaft mbH, 2. Auflage,
Stuttgart: 113, 412-413, 2006.
Okada M, Kazumi I, Watanabe K:
Effect of Vitamin B6 Deficiency on Glycogen Metabolism in the Skeletal Muscle, Heart and Liver
of Rats. J Nutr Sci Vitaminol 37: 349-357, 1991.
O´Kane DE, Gunsalus IC:
The resolution and purification of glutamic-aspartic transaminase.
J Biol Chem 170: 425, 1947.
Ono K, Ono T, Matsumata T:
The pathogenesis of decreased aspartate aminotransferase and alanine aminotransferase activity in
the plasma of hemodialysis patients: the role of vitamin B& deficiency.
Clin Nephrol 43; 6: 405-408, 1995.
Paolini CL, Meschia G, Fennessey PV, Pike AW, Teng C, Battaglia FC, Wilkening RB:
An in vivo study of ovine placental transport of essential amino acids. Am J Physiol Endocrinol
Metab 280; 1: E31-39, 2001.
Peel S:
Granulated metrial gland cells. Adv Anat Embryol Cell Biol 115: 1-112, 1989.
Peel S, Adam E:
The killing of rat placental cells by rat and mouse granulated metrial gland cells in vitro.
Placenta 12; 2: 161-171, 1991.
110

Peel S, Bulmer D:
Proliferation and differentiation of trophoblast in the establishment of the rat chorio-allantoic
placenta. J Anat 124; 3: 675-687, 1977.
Piedrahita JA, Oetama B, Bennett GD, van Waes J, Kamen BA, Richardson J, Lacey SW,
Anderson RG, Finnell RH:
Mice lacking the folic acid-binding protein Folbp1 are defective in early embryonic development.
Nat Genet 23; 2: 228-232, 1999.
Pijnenborg R, Vercruysse L:
Mathias Duval on placental development in mice and rats. Placenta 27: 109-118, 2006.
Quinn CT, Griener JC, Bottiglieri T, Arning E, Winick NJ:
Effects of intraventricular methotrexate on folate, adenosine, and homocysteine metabolism in
cerebrospinal fluid. Am J Pediatr Hematol Oncol 26; 6: 386-388, 2004.
Ramsey EM:
The placenta: human and animal. Praeger Publishers, New York: 65, 110, 114, 1982.
Regnault TRH, Friedman JE, Wilkening RB, Anthony RV, Hay WW:
Fetoplacental transport and utilization of amino acids in IUGR – a review.
Placenta 26: 52-62, 2005.
Reynolds RD:
Bioavailability of vitamin B-6 from plant foods.
Am J Clin Nutr 48; 3: 863-867, 1988.
De Rijk EPCT, van Esch E, Flik G:
Pregnancy dating in the rat: placental morphology and maternal blood parameters.
Toxicol Pathol 30; 2: 271-282, 2002.
Rösch C, Steinbicker V, Weise W:
Die Häufigkeit von Neuralrohrdefekten im Regierungsbezirk Magdeburg.
Gesundheitswesen 60: 563-566, 1998.
111

Romeis B:
Mikroskopische Technik. R. Oldenbourg Verlag, München: 65,74,174-175,275, 1968.
Ross JC, Fennessey PV, Wilkening RB, Battaglia FC, Meschia G:
Placental transport and fetal utilization of leucine in a model of fetal growth retardation.
Am J Physiol 270; 3Pt1: E491-503, 1996.
Roth-Maier DA, Benedikt J, Kirchgeßner M:
Zum Einfluß der alimentären Vitamin-B6- Versorgung während der Gravidität und Laktation auf
die Aktivität spezifischer Transaminasen laktierender Ratten. Z Ernährungswiss 35; 3: 282-287,
1996.
Salewski E:
Färbemethode zum makroskopischen Nachweis von Implantationsstellen am Uterus der Ratte.
Arch. exp. Path. Pharmak. 247: 367, 1964.
Schenker S, Johnson RF, Mahuren JD, Henderson GI, Coburn SP:
Human placental vitamin B6 (pyridoxal) transport: normal characteristics and effects of ethanol.
Am J Physiol 262; 6 Pt 2: R966-974, 1992.
Schiebler TH, Knoop A:
Histochemische und Elektronenmikroskopische Untersuchungen an der Rattenplazenta.
Z Zellforsch Mikrosk Anat 50: 494-552, 1959.
Schneider H, Dancis J:
Amino acid transport in human placental slices.
Am J Obstet Gynecol 120; 8: 1092-1098, 1974.
Sengers BG, Please CP, Lewis RM:
Computational modelling of amino acid transfer interactions in the placenta:
Exp Physiol 95; 7: 829-840, 2010.
112

Sikka PK, McMartin KE:
Determination of folate transport pathways in cultured rat proximal tubule cells.
Chem Biol Interact 114; 1-2: 15-31, 1998.
Smithells RW, Sheppard S, Schorah CJ:
Vitamin deficiencies and neural tube defects. Arch Dis Child 51; 12: 994-50, 1976.
Solanky N, Requena Jimenez A, D´Souza SW, Sibley CP, Glazier JD:
Expression of folate transporters in human placenta and implications for homocysteine
metabolism.
Placenta 31; 2: 134-143, 2010.
Specht W:
Färben mit Aldehydthionin. Eine Methode für den topochemischen Nachweis von Sulfonsäuren
und Aldehyden. 65. Verslg. d. Anat. Gesellsch., Würzburg, 1970.
Stahl A, Heseker H:
Folat, Physiologie, Vorkommen, Analytik, Referenzwerte und Versorgung in Deutschland.
Ernährungsumschau 54; 6: 336-343, 2007.
Stanger O, Herrmann W, Pietrzik K, Fowler B, Geisel J, Dierkes J, Weger M:
Konsensuspapier der D.A.CH.-Liga Homocystein über den rationellen klinischen Umgang mit
Homocystein, Folsäure und B-Vitaminen bei kardiovaskulären und thrombotischen Erkrankungen
– Richtlinien und Empfehlungen. J Kardiol 10; 5: 197,2003.
Steegers-Theunissen RP, Boers GH, Blom HJ, Nijhuis JG, Thomas CM, Borm GF, Eskes TK:
Neural tube defects and elevated homocysteine levels in amniotic fluid.
Am J Obstet Gynecol 175; 5: 1436-1441, 1995.
Steegers-Theunissen RP, Wathen NC, Eskes TK, van Raaij-Selten B, Chard T:
Maternal and fetal levels of methionine and homocysteine in early human pregnancy.
Br J Obstet Gynaecol. 104; 1: 20-24, 1997.
113

Succari M, Pontziere C, Meneguzzer E, Desmoulins D, Muocque M, Boy P:
Aminotransferase in elderly people: influence of pyridoxal-5-phosphate addition on the
determination of serum aminotransferases in healthy elderly subjects. Age Ageing 16: 94-100,
1987.
Suter PM:
Checkliste Ernährung. Thieme, Stuttgart, New York, 92–136, 2002.
Takata K, Fujikura K, Shin BC:
Ultrastructure of the rodent placental labyrinth: a site of barrier and transport.
J Reprod Dev 43; 1: 13-24, 1997
Taylor RT, Cote RJ:
Immunomicroscopy. A diagnostic tool for the surgical pathologist.
3. Auflage Elsevier Verlag, Philadelphia, 22, 72-73, 2006.
Thliveris, JA:
Fine structure of the placental labyrinth in the rat at term and during prolonged gestation.
Virchows Arch B Cell Pathol 21; 2: 169-179, 1976.
Tsitsiou E, Sibley CP, D´Souza SW, Catanescu O, Jacobsen DW, Glazier JD:
Homocysteine transport by system L, A and y+L across the microvillous plasma membrane of
human placenta. J Physiol 15; 587: 4001-4013, 2009.
Tönz O, Lüthy J, Raunhardt O:
Folsäure zur Verhütung von Neuralrohrdefekten.
Schweiz Med Wochenschr 126: 177-187, 1996.
Unbekannter Autor:
Metabolism of vitamin B6 in the pregnant rat. Nutr Rev 30 ;2: 44-47, 1972.
114

Van Aerts LA, Poirot CM, Herberts CA, Blom HJ, De Abreu RA, Trijbels JM, Eskes TK,
Peereboom-Stegeman JH, Noordhoek J:
Development of methionine synthase, cystathione-beta-synthase and S-adenosyl-homocysteine
hydrolase during gestation in rats. J Reprod Fertil 103; 2: 227-232, 1995.
Vezmar S, Becker A, Bode U, Jaehde U:
Biochemical and clinical aspects of Methotrexate neurotoxicity.
Chemotherapy 49; 1-2: 92-104, 2003.
Wang JW, Trumbo PR:
Bioavailability of vitamin B-6 in pregnant rats. J Nutr 122; 9: 1892-1897, 1992.
Waxman S, Schreiber C:
Characteristics of folic acid-binding protein in folate-deficient serum.
Blood 42; 2: 291-301, 1973.
Weingärtner J, Maile S, Proff P, Reicheneder C, Bienengräber V, Fanghänel J, Gedrange T:
Secondary palat closure in rats in association with relative maternofetal levels of folic acid,
vitamin B12, and homocysteine. Ann Anat 189: 229-233, 2007.
Welsh AO, Enders AC:
Chorioallantoic placenta formation in the rat. III. granulated cells invade the uterine luminal
epithelium at the time of epithelial cell death. Biol Reprod 49: 38-57, 1993.
Wendstrom KD, Johanning GL, Owen J, Johnston KE, Acton S, Tamura T:
Rolfe of amniotic fluid homocysteine level and of fetal 5, 10-methylentetrahydafolate reductase
genotype in the etiology of neural tube defects. Am J Med Genet. 3; 90: 12-16, 2000.
Wilcox AJ, Lie RT, Solvoll K, Taylor J, McConnaughey DR, Åbyholm F,
Vindenes H, Emil S, Drevon CA:
Folic acid supplements and risk of facial clefts: national population based case-control study.
BMJ 3; 334: 464, 2007.
115

Will M, Barnard JA, Said HM, Ghishan FK:
Fetal hydantoin syndrome: inhibition of placental folic acid transport as a potential mechanism for
fetal growth retardation in the rat.
Res Commun Chem Pathol Pharmacol 48; 1: 91-98, 1985.
Wirtschafter ZT:
The dynamics of free amino acids in the amniotic fluid of rats.
Am J Obstet Gynecol 75; 4: 718-723, 1958 a.
Wirtschafter ZT:
Free amino acids in human amniotic fluid, fetal and maternal Serum.
Am J Obstet Gynecol 76; 6: 1219.1225, 1958 b.
Wislocki GB, Deane HW, Dempsey EW:
The histochemistry of the rodent´s placenta. Am J Anat 78: 281-346, 1946.
Zhao R, Li Y, Chen X:
Effects of homocysteine on post-implantation rat embryo cultured in vitro.
Wei Sheng Yan Jiu 30; 1: 34-36, 2001.
Zhao R, Li Y, Chen X, Zhao Z:
Inhibition on the endocytosis of early visceral yolk sac induced by homocysteine in rats.
Wei Sheng Yan Jiu (Journal of hygiene research) 31; 1: 32-33, 2002.
116

Eidesstattliche Erklärung
Hiermit erkläre ich, daß ich die vorliegende Dissertation selbständig verfaßt und keine anderen als
die angegebenen Hilfsmittel benutzt habe.
Die Dissertation ist bisher keiner anderen Fakultät vorgelegt worden.
Ich erkläre, daß ich bisher kein Promotionsverfahren erfolglos beendet habe und daß eine
Aberkennung eines bereits erworbenen Doktorgrades nicht vorliegt.
Datum Unterschrift
117

Danksagung
In erster Linie danke ich Herrn Univ.-Prof. Dr. M. Zygmunt für die Durchsicht der Arbeit sowie
für seine konstruktive Kritik.
Gleich an zweiter Stelle möchte ich Herrn Dr. Jens Weingärtner aus dem Institut für Anatomie und
Zellbiologie nennen. Ihm danke ich insbesondere für seine gute fachliche Betreuung, für hilfreiche
Hinweise und eine unkomplizierte Zusammenarbeit.
Nicht zuletzt möchte ich mich auch bei Frau Dr. K. Lotz bedanken, welche mir wertvolle
Hinweise bei der Aufbereitung der histologischen Präparate gab.
Auch bei Frau M. Bansemir, Mitarbeiterin des Instituts für Anatomie und Zellbiologie bedanke ich
mich für Ihre Hilfe bei der Aufarbeitung der histologischen Präparate.
Für die statistische Auswertung der gewonnenen Daten danke ich Herrn PD Dr. G. Kundt aus dem
Institut für Medizinische Informatik und Biometrie der Universität Rostock.
118
Related Documents











