ISSN: 1524-4636 Copyright © 2004 American Heart Association. All rights reserved. Print ISSN: 1079-5642. Online 7272 Greenville Avenue, Dallas, TX 72514 Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association. DOI: 10.1161/01.ATV.0000134299.89599.53 published online Jun 3, 2004; Arterioscler Thromb Vasc Biol Behrooz G. Sharifi Kurt Wallner, Chen Li, Prediman K. Shah, Kai-Jin Wu, Stephen M. Schwartz and Cells EGF-Like Domain of Tenascin-C Is Proapoptotic for Cultured Smooth Muscle http://atvb.ahajournals.org/cgi/content/full/01.ATV.0000134299.89599.53/DC1 Data Supplement (unedited) at: http://atvb.ahajournals.org located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://atvb.ahajournals.org/subscriptions/ Biology is online at Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular by on May 23, 2011 atvb.ahajournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN: 1524-4636 Copyright © 2004 American Heart Association. All rights reserved. Print ISSN: 1079-5642. Online
7272 Greenville Avenue, Dallas, TX 72514Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association.
DOI: 10.1161/01.ATV.0000134299.89599.53 published online Jun 3, 2004; Arterioscler Thromb Vasc Biol
Behrooz G. Sharifi Kurt Wallner, Chen Li, Prediman K. Shah, Kai-Jin Wu, Stephen M. Schwartz and
Cells EGF-Like Domain of Tenascin-C Is Proapoptotic for Cultured Smooth Muscle
http://atvb.ahajournals.org/cgi/content/full/01.ATV.0000134299.89599.53/DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://atvb.ahajournals.org/subscriptions/Biology is online at Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

EGF-Like Domain of Tenascin-C Is Proapoptotic forCultured Smooth Muscle Cells
Kurt Wallner, Chen Li, Prediman K. Shah, Kai-Jin Wu, Stephen M. Schwartz, Behrooz G. Sharifi
Objective—Based on our previous observations on the expression of Tenascin-C (Tn-C) in human atherosclerotic plaquesand its colocalization with macrophages, we explored whether Tn-C undergoes fragmentation and the potentialpathobiological significance of this fragmentation.
Methods and Results—Using cultured human smooth muscle cells (SMCs), we found that Tn-C upregulates expression ofmatrix metalloproteinases (MMPs). Western blot analysis revealed that Tn-C substrate is fragmented and most of thecleavage products have fibronectin-like and epidermal growth factor-like (EGF-L) domains of Tn-C. One fragment thatcontains an EGF-like domain was found in some human atherosclerotic plaques. Cell culture studies revealed that therecombinant EGF-like domain inhibits growth, induces apoptosis of SMCs in a dose-dependent, time-dependent, andcaspase-dependent manner, and activates caspase-3 before SMC detachment. Conversely, the caspase inhibitorz-YVAD.cmk, serum, and protease inhibitors blocked cell apoptosis conferred by the EGF-like domain. In addition,these inhibitors blocked EGF-like domain-induced caspase-3 activation. In contrast to this EGF-like domain, intactTn-C, its fibronectin-like, and its fibrinogen-like domains were inactive.
Conclusions—Together with our previous observations, our data suggest that Tn-C upregulates MMP expression thatcleaves Tn-C into fragments containing the EGF-like domain. This domain has proapoptotic activity for SMCs.(Arterioscler Thromb Vasc Biol. 2004;24:1-6.)
Key words: Tenascin-C � metalloproteinases � atherosclerosis � apoptosis � EGF-like domain
The reasons for loss of smooth muscle cell (SMC) asatherosclerotic plaques progress are not known. One
possible explanation is that the loss of cellularity as lesionsevolve is a result of changes in the extracellular matrix.Indeed, cell death resulting from loss of cell adhesion to theextracellular matrix has been called “anoikis.”1 Activatedinflammatory cells and vascular cells produce various pro-teases including matrix metalloproteinases (MMPs),2,3 whichdisrupt the matrix structure that supports endothelial cells andSMCs and, thereby, enhance the apoptotic propensity ofcells.4–11 We have previously shown that serum proteaseinhibitors are required for spreading of SMCs in a fibrin gelbecause of their ability to inhibit fibronectin proteolysis.12
Our earlier study showed that Tn-C upregulates expression ofMMPs in cultured macrophages, and it is strongly expressedin advanced human atherosclerotic plaques, particularlyaround the lipid core and plaque shoulders,13 regions withhigh levels of MMP activity, and SMC apoptosis.14,15 Little isknown about the role of Tn-C in human atheroscleroticplaques. We now report that Tn-C undergoes fragmentationin cultured SMCs, and in human atherosclerotic plaques and
one of the fragments contains the EGF-L domain of Tn-C.Using a recombinant EGF-L domain, we found that thisdomain promotes apoptosis of cultured SMCs and thatchymotrypsin inhibitor blocks this apoptosis.
MethodsPlease see online methods section at http://atvb.ahajournals.org forfurther details. Tn-C was purified from conditioned media of BHKcells overexpressing the large isoform of Tn-C, as described.16 Therecombinant proteins corresponding to the full-length fibrinogen-likeand unspliced fibronectin-like domains of Tn-C were expressed andpurified from the bacteria BL-21 Escherichia coli as described.17
GM 6001 metalloproteinase inhibitor was obtained from Chemicon.Other reagents were purchased from Sigma. All reagents were testedfor endotoxin activity using E-TOXATE kit (Sigma) and were foundto be endotoxin-free.
Cultured SMCsRat vascular SMCs were cultured as we described previously.18
Cultured human SMCs were kindly provided by Dr Navab (Univer-sity of California Los Angeles). For more details, please seehttp://www.ahajournals.org.
Received March 4, 2004; revision accepted May 12, 2004.From the Atherosclerosis Research Center (K.W., C.L., P.K.S., K-J.W., B.G.S.), Division of Cardiology, and Burns and Allen Research Institute,
Cedars-Sinai Medical Center, and UCLA School of Medicine, Los Angeles, Calif; and the Department of Pathology (S.M.S.), University of Washington,Seattle.
Supported by National Institutes of Health grant HL50566, Established Investigator Award 0040201N from the American Heart Association, and theEisner Foundation.
Correspondence to Dr Behrooz G. Sharifi, Cedars-Sinai Medical Center, Davis Building, 1016 8700 Beverly Boulevard, Los Angeles, CA 90048.E-mail [email protected]
© 2004 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org DOI: 10.1161/01.ATV.0000134299.89599.53
1 by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ZymographyZymography was performed essentially as described previously.13
For more details, please see http://www.ahajournals.org.
Cloning of Rat EGF-LTotal RNA derived from rat SMC was used as a template for thereverse-transcription polymerase chain reaction, which was per-formed as described16 using primers containing a KEX2 site. Thepolymerase chain reaction product was placed on agarose gel,isolated, cut with XhoI/EcoRI, and subcloned into pPIC9 yeastvector, essentially as recommended by Invitrogen. After transforma-tion, cells were screened for Mut� and Muts transformants. For moredetails, see please see http://www.ahajournals.org.
Purification and Characterization of theRecombinant EGFThe ammonium sulfate pellet was resuspended in 10% acetonitrileand loaded onto a C8 reverse-phase column that was eluted with agradient of 10% to 60% acetonitrile containing 0.1% trifluoroaceticacid. For more details, please see http://www.ahajournals.org.
Antibody TechniquesThe anti–Tn-C monoclonal antibody was obtained from Sigma(clone BC-24). The antibody recognizes an epitope within theN-terminal EGF sequence of the molecule. The antibodies to thefibronectin-like domain of Tn-C were a gift from Dr Erickson (DukeUniversity, Durham, NC). Tn-C degradation was measured byWestern blot as we previously described.19 For more details, pleasesee http://www.ahajournals.org.
Cell ApoptosisCell apoptosis was measured by several methods. Please seehttp://atvb.ahajournals.org.
All the experiments have been performed in triplicate and repeated3 times with different preparations of Tn-C or recombinant EGF-Ldomain to ensure reproducibility. The results are representative ofeither 3 experiments or average�SEM of a triplicate determinations.
ResultsWe have previously reported that Tn-C upregulates MMPssynthesis by macrophages.13 Because SMCs are major com-ponents of atherosclerotic plaques, we asked whether Tn-Caffects expression of MMPs in these cells. To explore thispossibility, SMCs were plated onto Tn-C–coated dishes indefined media for 24 hours. Collagen-coated dishes wereused as a control. SMCs cultured on collagen type I consti-tutively secreted only 1 band, 70-kDa, with gelatinolyticactivity. Cells cultured on Tn-C substrate expressed 2 bands,70-kDa and 65-kDa, with gelatinolytic activity (Figure 1).The 70-kDa enzyme activity was increased �4-fold ascompared with collagen (Figure 1), and the 65-kDa activitywas detected primarily on the Tn-C substrate. Similar resultswere obtained with rat and porcine SMCs (data not shown),indicating that the effect of Tn-C is conserved. Kinetic studiesrevealed that the 65-kDa protease activity was detectable 8 to9 hours after plating onto Tn-C substrate and reached satu-ration levels after 24 hours. Past studies have shown thatTn-C is sensitive to degradation by MMPs.20,21 Because Tn-Cupregulates MMP expression, we reasoned that this upregu-lation may, in turn, lead to degradation of Tn-C. To explorethis possibility, human SMCs were plated onto Tn-C–coateddishes in 10% fetal bovine serum or defined serum-free, andthen extracted 10 hours or 24 hours after plating. Theextracted Tn-C was analyzed by Western blot using antibod-
ies specific to the fibronectin-like or EGF-like domains ofTn-C. The 2 antibodies showed that Tn-C remained intact inthe presence of serum (Figure 2A through 2C, lane S). Incontrast, in defined media, Tn-C was fragmented 10 hoursafter plating and further fragmented at 24 hours (Figure 2Athrough 2C, lanes 10 and 24). The patterns of Tn-C fragmen-tation detected by the 2 antibodies were similar with 1exception: the anti-EGF-like monoclonal antibody (Figure 2Band 2C), but not anti-fibronectin domain antibody (Figure2A), detected an additional band corresponding to 85-kDa,suggesting that this band contains EGF-like domain. Tofurther determine whether the 85-kDa fragment possesses anEGF-like domain, we performed an antibody pull-down assayby incubating the anti-EGF-like domain monoclonal antibodywith a 0.1-�mol/L solution of recombinant EGF-like orfibronectin-like domain of Tn-C before addition to the blot.Pre-incubation with the recombinant fibronectin-like domainhad no effect on the bands recognized by anti-EGF-likedomain antibody, whereas pre-incubation with the recombi-nant EGF-like domain blocked the ability of antibody todetect the 85-kDa band (data not shown). We concluded thatthe 85-kDa band is likely a fragment of Tn-C that contains itsEGF-like domain. To determine whether degradation of Tn-Cis mediated by MMPs, SMCs were cultured in defined mediaonto Tn-C–coated dishes in the presence of 25 �mol/L
Figure 1. Representative effect of Tn-C on the gelatinolytic pro-teins produced by SMCs. A, Human SMCs (passage 3) werecultured in DMEM and added to the 24-well plates precoatedwith the long isoform of Tn-C (Tn) in the defined media(insulin�transferrin�selenium-X; GIBCO-BRL) and diluted withserum-free DMEM to make defined media. Some wells werecoated with collagen (Cgn) according to manufacturer’s (Cohe-sion Technologies) instruction for preparation of fibrillar collagenfilms. Media were collected 8 hours and 24 hours after platingthe cells, and protein concentrations were measured. Equalamounts of total proteins were loaded on each lane of a zymo-gram. The results displayed here are representative of 3experiments.
2 Arterioscler Thromb Vasc Biol. August 2004
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

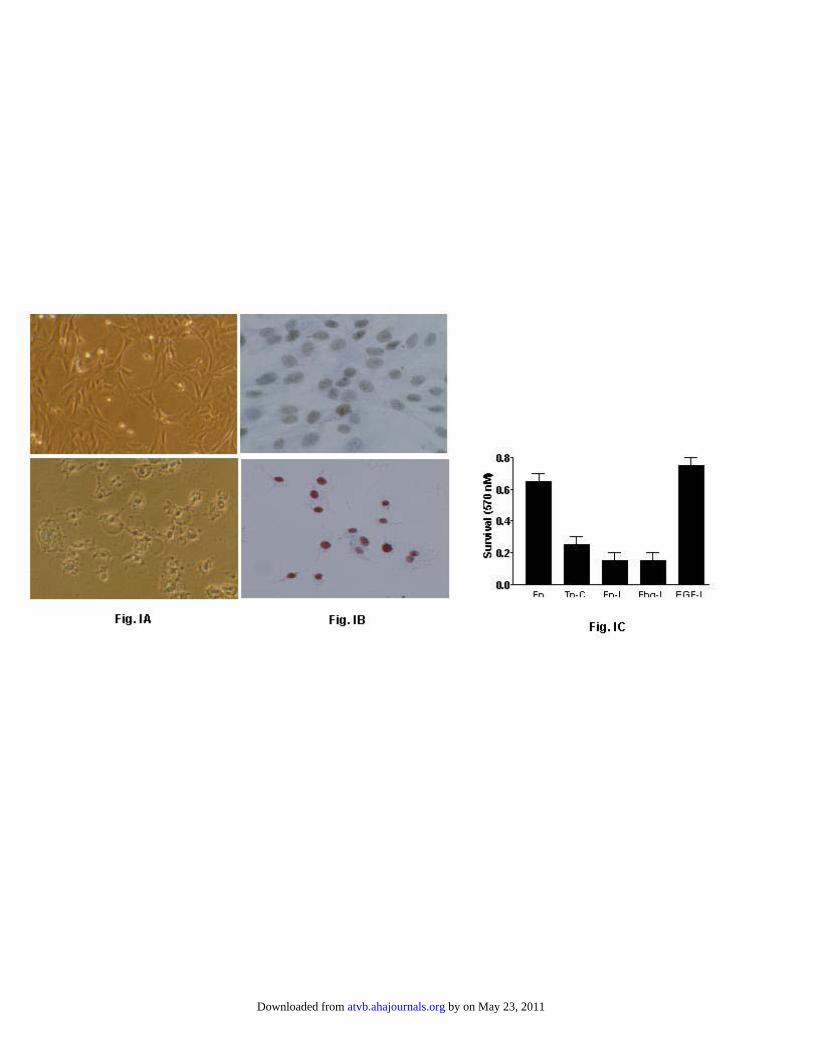
GM6001, a MMP inhibitor that blocks the activities of MMP1 to 3, MMP-8, and MMP-9. SDS-PAGE analysis of cellextracts showed that the MMP inhibitor blocked degradationof Tn-C (Figure 2D; lanes 10 and 24), suggesting that Tn-Cdegradation is mediated by MMPs produced by cultured cells.It has been reported that proteoglycans undergo fragmenta-tion in unstable human endarterectomy specimens,22 suggest-ing that matrix proteins may be fragmented in the plaque. Wepreviously reported that Tn-C expression colocalizes withmacrophages in human atherosclerotic plaques;13 therefore,we hypothesized that this colocalization could lead to Tn-Cfragmentation. To explore this idea, Tn-C integrity wasdetermined in human endarterectomy specimens by Westernblot. Antifibronectin-like domain antibodies detected 220-kDa and 280-kDa bands in all 16 endarterectomy specimens,which most likely correspond to the small and large isoformsof Tn- C, respectively (data not shown). However, anti-EGF-like domain monoclonal antibody not only detected the220-kDa and 280-kDa bands in all 16 specimens but alsoreacted with 85-kDa and 65-kDa proteins in 4 specimens(Figure 3), suggesting that under some circumstances, Tn-Cis fragmented in human atherosclerotic plaques and that thefragments most likely contain an EGF-like domain. Todetermine the biological consequences of Tn-C fragmenta-tion, we cultured SMCs onto Tn-C–coated dishes in definedmedia for 24 hours. Cells plated onto fibronectin-coateddishes in defined media were used as control. Consistent withour previous report,17 we noted that SMCs adhered to Tn-Csubstrate; however, cells remained round and did not spreadwithin 1 hour after plating, in contrast to cells cultured ontofibronectin-coated dishes (not shown). Examination of cells24 hours after plating showed that most cells were detachedfrom the Tn-C–coated dishes and those that remain attachedassumed morphology characteristic of apoptotic cells (Figure
IA, bottom, available online at http://atvb.ahajournals.org). Incontrast, cells cultured onto fibronectin-coated dishes did notexhibit apoptotic morphology (Figure IA, top), suggestingthat apoptosis of cells is related to Tn-C substratum. Becausewe noted that under these culture conditions Tn-C is degradedand that MMP inhibitor blocked fragmentation, we askedwhether cell apoptosis is related to Tn-C degradation. Toanswer this question, we cultured cells onto Tn-C–coateddished in defined media in the presence and absence ofGM6001 MMP inhibitor and cell apoptosis was measured byterminal deoxynucleotidyl transferase-mediated dUTP nickend-labeling staining. In the absence of inhibitor, cellsshowed condensed red nuclei (Figure IB, bottom); however,in the presence of inhibitor, cells were fully spread andshowed expanded gray nuclei, indicating that they are viable(Figure IB, top). These data suggest that SMC apoptosis isrelated to degradation of Tn-C, most likely by MMPs. Todetermine which domain of Tn-C mediates the apoptoticactivity of Tn-C, we preformed a protein pull-down assay byincubating the conditioned media collected from culture cellsthat were plated onto Tn-C–coated dishes in defined mediafor 24 hours. The media were collected, incubated withantibodies to intact Tn-C, fibrinogen-like, fibronectin-like, orEGF-like domains of Tn-C, and then added to SMCs culturedonto fibronectin-coated dishes. Pre-incubation of media withintact Tn-C (Figure IC, lane Tn-Cg), anti-fibronectin-like(Figure IC, lane Fn-L) or antifibrinogen-like domain (FigureIC, lane Fbg-L) antibodies did not influence the proapoptoticactivity of the conditioned media. In contrast, pre-incubationof media with anti-EGF-like antibodies (Figure IC, laneEGF-L) neutralized the proapoptotic activity of the condi-tioned media, suggesting that the apoptotic activity is asoluble molecule related to the EGF-L domain of Tn-C.Although there are extensive data about the activity of intactTn-C and its fibronectin-like and fibrinogen-like domains,little is known about the activity of its EGF-like domain. Thefull-length EGF-like domain of Tn-C consists of 447 amino
Figure 2. Degradation of Tn-C by cultured human SMCs.Human SMCs were added to Tn-C–coated dishes in the pres-ence of serum (S) or in defined media for 10 hours10 or 24hours.24 Purified large Tn-C isoform was used as a control (C).The coated Tn-C was solubilized with 5% SDS, electrophoresedon 5% SDS-PAGE, and transferred to a polyvinylidene diflouridemembrane. Three blots were generated that were incubatedwith 1:10 000 dilution of antihuman fibronectin-like domain anti-body (A) or 1:1000 dilution of monoclonal anti-EGF-like antibody(B). C, The anti-EGF-like monoclonal antibody was preincubatedwith a 0.1 �mol/L solution of recombinant fibronectin-likedomain for 30 minutes at 37°C before addition to the blot. D,SMCs were cultured onto Tn-C–coated dishes in the presenceof 25 �mol/L GM6001, a metalloproteinase inhibitor, followed byTn-C extraction and analysis by Western blot using anti-EGF-L
monoclonal antibody as described for (B).
Figure 3. Fragmentation of Tn-C in human carotid endarterecto-my specimens. Human carotid endarterectomy specimens wereobtained after surgery and immediately snap-frozen with liquidnitrogen. The frozen samples were pulverized and thenextracted with a buffer (0.1 mol/L CAPS, 0.15 mol/L NaCl, pH11) known to be effective in extracting Tn-C.32 After centrifuga-tion, protein concentrations of the supernatants were deter-mined, and 1 �g from each sample was analyzed by SDS-PAGEand Western blot as described using anti-human EGF-likedomain monoclonal antibody (sigma).
Wallner et al EGF-L Induces Cell Apoptosis 3
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

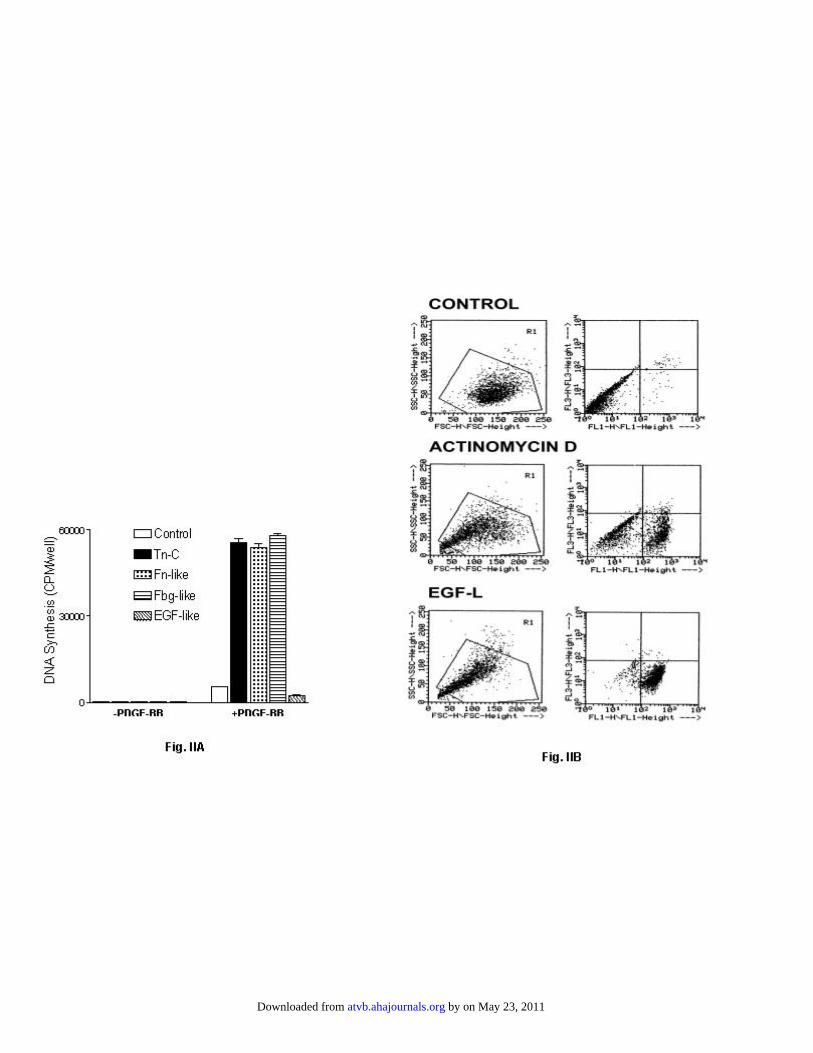
acids, with 14.5 repeats of a 31 amino acid EGF-like domain.The EGF-like repeats share 6 cysteine residues, which arelinked together by a special configuration of disulfidebonds.23 The presence of 87 cysteine residues and nonsequen-tial arrangement of the disulfide bonds excludes the use of anE. coli system for the expression of the EGF-like domainbecause of lack of disulfide isomerase activity. Therefore, weused yeast to express this domain of Tn-C. To determine theactivity of recombinant EGF-like domains, a 0.1-�mol/Lsolution of intact long Tn-C isoform, recombinantfibronectin-like, fibrinogen-like, or EGF-like domain wasadded to fibronectin-coated cultured SMCs in the absence orpresence of 1 nM PDGF-BB in serum-free media. As ex-pected, in the absence of PDGF-BB, there was no DNAsynthesis. However, DNA synthesis was stimulated in thepresence of PDGF-BB and Tn-C domains, with the exceptionof the EGF-L domain (Figure IIA, available online at http://atvb.ahajournals.org). The cultured SMCs in the presence ofrecombinant EGF-L domain exhibited apoptotic morphology,suggesting that inhibition of DNA synthesis is related to cellapoptosis. To further investigate this idea, cells were ana-lyzed for annexin V externalization. Cells were cultured ontofibronectin-coated dishes in defined media in the presence ofrecombinant EGF-L domain for 24 hours. Cells treated withthe actinomycin D under similar condition were used as apositive control. Fluorescence-activated-cell sorter analysisrevealed that 78�7% of cells treated with EGF-L underwentapoptosis (Figure IIB). Under similar conditions, 65�8% ofcells treated with actinomycin D underwent apoptosis (FigureIIB). Untreated cells remained viable. Because activation ofcaspases is central to cell apoptosis, SMCs were treated withthe recombinant EGF-like domain in the presence of increas-ing concentrations of z-VAD.fmk, a widely used caspaseinhibitor, and cell viability was measured by MTT assay.Unexpectedly, this inhibitor was toxic to the cells; however,other caspase inhibitors, z-DEVD.fmk or z-YVAD.cmk, werenot toxic. The most effective caspase inhibitor wasz-YVAD.cmk, which at 100 �mol/L reduced EGF-likedomain-induced cell death by 50% (Figure IIIA, availableonline at http://atvb.ahajournals.org). Increasing concentra-tions of the inhibitor to 200 to 400 �mol/L completelyblocked cell death (data not shown). To further confirm theinvolvement of caspases, we measured activation of
caspase-3 by Western blot in both attached and detachedSMCs. The activation of caspase-3, as measured by intensityof the 11-kDa caspase fragment, was weakly detectable 10hours after treatment that significantly increased at 24 hoursin the attached SMCs (Figure IIIB). In detached cells,activation of caspase-3 was observed at 24 to 48 hours oftreatment with EGF-like domain, which was suppressed byz-YVAD.cmk in a concentration- dependent manner. Addi-tion of 100 �mol/L of the inhibitor reduced activation ofcaspase-3, whereas 300 �mol/L completely suppressed it(Figure IIIB). In addition, PARP, a caspase-3 substrate, wasalso cleaved in a time-dependent manner (data not shown).Collectively, we conclude that the addition of recombinantEGF-like domain to cultured SMCs activates caspase-3 be-fore cell detachment. Because we previously reported thatSMC viability is dependent on its interaction with extracel-lular matrix proteins and that serum factors prevent cell deathby inhibiting matrix degradation,19 we asked whether theapoptotic activity of the recombinant EGF-like domain is aprotease-dependent event. Serum blocked the proapoptoticactivity of the recombinant EGF-like domain (data notshown). To identify the nature of this inhibition, SMCs weretreated with the recombinant EGF-like domain in definedmedia containing various protease inhibitors. We noted that a0.32-�g/mL solution of �-macroglobulin significantly re-duced and a 0.6-�g/mL solution completely blocked EGF-L–induced cell apoptosis (Figure 4). Chymostatin, a chymotryp-sin inhibitor, at a concentration of 3 �mol/L blocked theapoptotic activity of EGF-like domain (Figure 4). In contrast,trypsin inhibitor, plasmin inhibitor, and thrombin inhibitorhad no effect (data not shown). These data suggest that theproapoptotic activity of EGF-L domain is mediated by aprotease. To further understand the role of EGF-L domain inSMC apoptosis, we measured caspase-3 activation in thepresence of protease inhibitors. We found that 1 �mol/Lchymostatin significantly reduced and 4 �mol/L completelysuppressed caspase-3 activation induced by the EGF-likedomain (Figure 5). In contrast, hirudin, a thrombin inhibitor(1 U/mL) and plasmin inhibitor (15 �g/mL) had no effect(Figure 5). To further examine whether the inhibitory activityof chymostatin is specific to apoptosis caused by EGF-likedomain, SMC death was induced by staurosporine andactinomycin D in the absence and presence of 4 �mol/L
Figure 4. Effect of plasma protease inhibitors on EGF-like domain-induced SMC apoptosis. SMCs were plated in 96-well microtiterplates in DMEM �10% serum, and then the media were replaced with the DMEM defined in the absence (C) or presence of0.01 �mol/L EGF-like domain supplemented with increasing concentrations of �-macroglobulin or chymostatin. After 48 hours, cell via-bility was determined by MTT assay. Each data point represents the average of 3 separate experiments.
4 Arterioscler Thromb Vasc Biol. August 2004
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

chymostatin. Although the proapoptotic reagents activatedcaspase-3, chymostatin did not block the activation (Figure5). From these data, we conclude that protease inhibitorsblocked the EGF-like domain activity through inhibition ofcaspase-3 activation.
DiscussionIt is believed that destruction of matrix protein by matrix-degrading enzymes leads to apoptosis of adherent cellscaused by deprivation of cell attachment. In this report, wedescribed another mechanism by which matrix-degradingenzymes may induce cell apoptosis. We found that culturingof SMCs onto Tn-C substrate, in the absence of serum, leadsto fragmentation of Tn-C. Tn-C fragmentation was alsodetected in some human atherosclerotic plaques. Based on themotilities of Tn-C fragments on SDS-PAGE, Western blotdata, and pull-down assays, it appears that proteolysis ofTn-C leads to generation of fragments that contain the EGF-L
domain. Proteolysis of Tn-C in cultured SMCs and inatherosclerotic plaques was not surprising because othershave noted that decorin, biglycan, versican, apoB, apo(a), andapoE undergo proteolysis in human endarterectomy speci-mens.22 Although the significance of this fragmentation isunknown, past studies have shown that MMP-2 cleavesdecorin in a region that is responsible for transforminggrowth factor-� binding, thus increasing availability of thisgrowth factor in an active form.24 Because decorin interactionwith collagen is critical to fibrillogenesis, proteolysis ofdecorin may affect tissue integrity and maintenance. Thus, itappears that cells are exposed to matrix fragments, in additionto intact protein. Therefore, it is important to assess thefragmentation of matrix molecules and activity of theirfragments.
We found that Tn-C upregulates MMP expression and thatMMP inhibitor blocked Tn-C fragmentation, suggesting anautocrine loop may be involved in the degradation of Tn-C inboth SMCs and macrophages. Upregulation of MMPs ap-pears to be a common activity of Tn-C, because others havenoted that a mixed substrate of Tn-C and fibronectin upregu-lates collagenase, stromelysin, and gelatinase gene expressionin rabbit fibroblast.25 Because Tn-C is sensitive to degrada-tion by MMP-1, MMP-2, MMP-3, and MMP-7,20,21 it is
likely that Tn-C undergoes degradation in inflamed tissuesand cells are exposed to Tn-C fragments in addition to theintact protein. These fragments may exhibit an activity that isnot found in the intact molecule. For example, cleavage of thelaminin-5 �2 chain by MMP-2 exposes a cryptic site withinlaminin, inducing migration of malignant breast epithelialcells, an activity not found in the intact protein.26 Similarly,we found that the EGF-L domain induces SMC apoptosis, anactivity not exhibited by the intact molecule. Taken together,these data suggest that local proteinase concentration andmatrix degradation determine cell behavior in multiple ways.
We have previously reported that pericellular proteolysisof matrix proteins induces SMCs apoptosis.12 However, theEGF-L–induced apoptosis described in this article is differentfrom anoikis, in which denying cell adhesion leads to cellapoptosis. We previously reported that SMCs adhere to Tn-Cand determined that the fibrinogen-like domain of Tn-Caccounts for this adhesive activity.17 However, we noted thatin the absence of serum, this adhesion did not lead to cellspreading; rather, adherent cells remained round and under-went apoptosis. We have now discovered that apoptosis canbe blocked by the addition of serum and protease inhibitors(such as �-macroglobulin or chymostatin), suggesting that aprotease may be involved in this process. We further identi-fied that a degradation product of Tn-C that contains itsEGF-L domain has proapoptotic activity. Thus, our datasuggest that degradation of Tn-C and generation of EGF-L
fragments are linked to SMCs apoptosis, an event that isindependent of interference with cell adhesion.
Although our data cannot rule out other mechanismspromoting SMC death, it is important to realize that very littleis known of the factors that control the loss of these cells fromthe human plaque. The only specific hypothesis has been thatapoptosis in atherosclerotic plaques is caused by the expres-sion of Fas and its ligand, FasL.27 However, experimentalliterature does not provide strong support for this hypothesis.For example, in vivo studies have demonstrated that intimallesions grow more rapidly in FasL-transduced arteries, whichis primarily caused by the accumulation of intimal SMCswith no significant apoptosis.28 Cell culture studies have alsoshown no correlation between Fas/FasL expression and SMCapoptosis. Cultured human SMCs express relatively highlevels of Fas/FasL and remain viable in the presence ofsoluble FasL or agonist anti-Fas antibody.29 SMCs are insen-sitive to apoptosis induced by Fas ligand27 and must bepretreated with interferon-� (INF-�) to become sensitive toapoptosis by anti-Fas antibody. It has also been proposed thatIFN-� pretreatment upregulates Fas, leading to the inductionof apoptosis.27 However, neither tumor necrosis factor-� norIFN-� upregulates Fas in human SMCs.29 In addition, mediacollected from activated T-cell clones isolated from athero-sclerotic plaques had no proapoptotic activity despite strongexpression of proinflammatory cytokines such as IFN-�.30,31
Taken together, these data suggest that the increased expres-sion of Fas/FasL in atherosclerotic plaques or cultured cellsmay not be related to SMC apoptosis.
In summary, we provide evidence that Tn-C upregulatesMMP expression in cultured SMCs, which leads to Tn-Cfragmentation and cell apoptosis. These findings are poten-
Figure 5. Protease inhibitors block caspase-3 activation. SMCswere treated with the recombinant EGF-like domain in the pres-ence of the indicated concentrations of chymotrypsin inhibitor(Chystatin; �M), hirudin, a thrombin inhibitor (Thn I; U/mL), andplasmin inhibitor (Pln I; �g/mL). In another set of experiments,cells were treated with 1 �mol/L starosporine (Starpn) or 250ng/mL actinomycin D (Actin D) in the presence and absence of4 �mol/L chymostatin. Cell extracts were prepared andcaspase-3 activation was measured as described.
Wallner et al EGF-L Induces Cell Apoptosis 5
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

tially important because advanced plaques contain SMCs,macrophages, T cells, and mast cells, which are cell types thatare capable of expressing MMPs and inducing apoptosis ofSMCs through fragmentation of Tn-C. Because proteolysis iscontrolled at a multitude of levels, this pathway of apoptosiswould increase the complexity of cell death in thevasculature.
References1. Frisch SM, Francis H. Disruption of epithelial cell-matrix interactions
induces apoptosis. J Cell Biol. 1994;124:619–626.2. Galis ZS, Sukhova GK, Kranzhofer R, Clark S, Libby P. Macrophage
foam cells from experimental atheroma constitutively produce matrix-degrading proteinases. Proc Natl Acad Sci U S A. 1995;92:402–406.
3. Galis ZS, Muszynski M, Sukhova GK, Simon-Morrissey E, Libby P.Enhanced expression of vascular matrix metalloproteinases induced invitro by cytokines and in regions of human atherosclerotic lesions. Ann NY Acad Sci. 1995;748:501–507.
4. Hungerford JE, Compton MT, Matter ML, Hoffstrom BG, Otey CA.Inhibition of pp125FAK in cultured fibroblasts results in apoptosis. J CellBiol. 1996;135:1383–1390.
5. van der Zee R, Murohara T, Passeri J, Kearney M, Cheresh DA, Isner JM.Reduced intimal thickening following �(v)�3 blockade is associated withsmooth muscle cell apoptosis. Cell Adhes Commun. 1998;6:371–379.
6. Stromblad S, Becker JC, Yebra M, Brooks PC, Cheresh DA. Suppressionof p53 activity and p21WAF1/CIP1 expression by vascular cell integrin�V�3 during angiogenesis. J Clin Invest. 1996;98:426–433.
7. Malyankar UM, Scatena M, Suchland KL, Yun TJ, Clark EA, GiachelliCM. Osteoprotegerin is an � v� 3-induced, NF-kappa B-dependentsurvival factor for endothelial cells. J Biol Chem. 2000;275:20959–20962.
8. Yamada KM, Araki M. Tumor suppressor PTEN: modulator of cellsignaling, growth, migration and apoptosis. J Cell Sci. 2001;114:2375–2382.
9. Grinnell F, Zhu M, Carlson MA, Abrams JM. Release of mechanicaltension triggers apoptosis of human fibroblasts in a model of regressinggranulation tissue. Exp Cell Res. 1999;248:608–619.
10. Ilic D, Almeida EA, Schlaepfer DD, Dazin P, Aizawa S, Damsky CH.Extracellular matrix survival signals transduced by focal adhesion kinasesuppress p53-mediated apoptosis. J Cell Biol. 1998;143:547–560.
11. Watanabe Y, Dvorak HF. Vascular permeability factor/vascular endothe-lial growth factor inhibits anchorage-disruption-induced apoptosis inmicrovessel endothelial cells by inducing scaffold formation. Exp CellRes. 1997;233:340–349.
12. Ikari Y, Fujikawa K, Yee KO, Schwartz SM. Alpha(1)-proteinase inhib-itor, alpha(1)-antichymotrypsin, or alpha(2)-macroglobulin is required forvascular smooth muscle cell spreading in three-dimensional fibrin gel.J Biol Chem. 2000;275:12799–12805.
13. Wallner K, Li C, Shah PK, Fishbein MC, Forrester JS, Kaul S, SharifiBG. Tenascin-C Is Expressed in Macrophage-Rich Human CoronaryAtherosclerotic Plaque. Circulation. 1999;99:1284–1289.
14. Isner JM, Kearney M, Bortman S, Passeri J. Apoptosis in human athero-sclerosis and restenosis [see comments]. Circulation. 1995;91:2703–2711.
15. Han DK, Haudenschild CC, Hong MK, Tinkle BT, Leon MB, Liau G.Evidence for apoptosis in human atherogenesis and in a rat vascularinjury model [see comments]. Am J Pathol. 1995;147:267–277.
16. LaFleur DW, Fagin JA, Forrester JS, Rubin SA, Sharifi BG. Cloning andcharacterization of alternatively spliced isoforms of rat tenascin. Plate-let-derived growth factor-BB markedly stimulates expression of splicedvariants of tenascin mRNA in arterial smooth muscle cells. J Biol Chem.1994;269:20757–20763.
17. LaFleur DW, Chiang J, Fagin JA, Schwartz SM, Shah PK, Wallner K,Forrester JS, Sharifi BG. Aortic smooth muscle cells interact withtenascin-C through its fibrinogen-like domain. J Biol Chem. 1997;272:32798–32803.
18. Sharifi BG, LaFleur DW, Pirola CJ, Forrester JS, Fagin JA. AngiotensinII regulates tenascin gene expression in vascular smooth muscle cells.J Biol Chem. 1992;267:23910–23915.
19. Ikari Y, Mulvihill E, Schwartz SM. � 1-Proteinase inhibitor, �1-antichymotrypsin, and � 2-macroglobulin are the antiapoptotic factorsof vascular smooth muscle cells. J Biol Chem. 2001;276:11798–11803.
20. Imai K, Kusakabe M, Sakakura T, Nakanishi I, Okada Y. Susceptibilityof tenascin to degradation by matrix metalloproteinases and serine pro-teinases. FEBS Lett. 1994;352:216–218.
21. Siri A, Knauper V, Veirana N, Caocci F, Murphy G, Zardi L. Differentsusceptibility of small and large human tenascin-C isoforms to degra-dation by matrix metalloproteinases. J Biol Chem. 1995;270:8650–8654.
22. Formato M, Farina M, Spirito R. Evidence for a proinflammatory andproteolytic environment in plaques from endarterectomy segments ofhuman carotid arteries. Arterioscler Thromb Vasc Biol. 2004;24:129–135.
23. Jones FS, Burgoon MP, Hoffman S, Crossin KL, Cunningham BA,Edelman GM. A cDNA clone for cytotactin contains sequences similar toepidermal growth factor-like repeats and segments of fibronectin andfibrinogen. Proc Natl Acad Sci U S A. 1988;85:2186–2190.
24. Imai K, Hiramatsu A, Fukushima D, Pierschbacher MD, Okada Y. Deg-radation of decorin by matrix metalloproteinases: identification of thecleavage sites, kinetic analyses and transforming growth factor- �1release. Biochem J. 1997;322:809–814.
25. Tremble P, Chiquet-Ehrismann R, Werb Z. The extracellular matrixligands fibronectin and tenascin collaborate in regulating collagenasegene expression in fibroblasts. Mol Biol Cell. 1994;5:439–453.
26. Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson WG,Quaranta V. Induction of cell migration by matrix metalloprotease-2cleavage of laminin-5. Science. 1997;277:225–228.
27. Geng YJ, Henderson LE, Levesque EB, Muszynski M, Libby P. Fas isexpressed in human atherosclerotic intima and promotes apoptosis ofcytokine-primed human vascular smooth muscle cells. ArteriosclerThromb Vasc Biol. 1997;17:2200–2208.
28. Schneider DB, Vassalli G, Wen S, Driscoll RM, Sassani AB, DeYoungMB, Linnemann R, Virmani R, Dichek DA. Expression of Fas ligand inarteries of hypercholesterolemic rabbits accelerates atherosclerotic lesionformation. Arterioscler Thromb Vasc Biol. 2000;20:298–308.
29. Sata M, Suhara T, Walsh K. Vascular endothelial cells and smoothmuscle cells differ in expression of Fas and Fas ligand and in sensitivityto Fas ligand-induced cell death: implications for vascular disease andtherapy. Arterioscler Thromb Vasc Biol. 2000;20:309–316.
30. de Boer OJ, van der Wal AC, Verhagen CE, Becker AE. Cytokinesecretion profiles of cloned T cells from human aortic atheroscleroticplaques. J Pathol. 1999;188:174–179.
31. de Boer OJ, van der Wal AC, Teeling P, Becker AE. Leucocyterecruitment in rupture prone regions of lipid-rich plaques: a prominentrole for neovascularization? Cardiovasc Res. 1999;41:443–449.
32. Lightner VA, Gumkowski F, Bigner DD, Erickson HP. Tenascin/hexabrachion in human skin: biochemical identification and localizationby light and electron microscopy. J Cell Biol. 1989;108:2483–2493.
6 Arterioscler Thromb Vasc Biol. August 2004
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement Detailed Methods
Cultured SMCs: Rat vascular smooth muscle cells were cultured as we described
previously 1. Cultured human SMCs were kindly provided by Dr. Navab (UCLA).
For adhesion experiments, 12-well culture plate dishes were coated with fibronectin
or Tn-C (10 µg/ml) at room temperature for 1 h followed by blocking of non-specific
sites with BSA as described 2. The cell suspensions were plated onto the culture
plates in DMEM media supplemented with either 5% calf serum or defined media
(Insulin+Transferrin+Selenium-X, GIBCO-BRL). The plates were incubated at 37 °C
with 5% CO2 for 24 h and then cultured media were collected.
Zymography: Zymography was performed essentially as described previously 3.
Briefly, SMCs were resuspended in the defined media containing 0.2% lactalbumin
hydrolysate, aliquoted into uncoated (control) and coated wells, and incubated for 24 h at
37OC. Conditioned media were collected, dialyzed, and protein concentrations were
measured by Bradford assay. Protein (5 µg/lane) was resolved in a 10% SDS-PAGE gel
containing 0.1% gelatin.
Cloning of rat EGF-L: To clone the rat EGF-L, a RT primer
(CGAATTCTAGGGGACACCTCAGAGCAGTC) was used to generate the first strand
DNA. The 5’ PCR primer (TCCGCTCGAGAAAAGAGAGGCTGAAGCTTCTGA
GCCTGACTGCCCTGG) contained a KEX2 site, which is required for the efficient
secretion of the EGF-L and the Xho I site for cloning the PCR product into yeast plasmid.
The 3’ PCR primer was identical to the RT primer described above with an addition of
EcoRI cleavage site. Total RNA derived from rat SMC was used as a template, and the
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement PCR reaction was performed as described 4. The PCR product was run on a agarose gel,
isolated, cut with Xho I/ EcoRI, and subcloned into pPIC9 yeast vector, essentially as
recommended by Invitrogen. The construct was sequenced to confirm the correct reading
frame (for secretion) and determination of an ATG in the proper context for eukaryotic
translation initiation. After establishing the veracity of the DNA sequence, Pichia was
transformed by electroporation. After transformation, cells were screened for Mut+ and
Muts transformants. PCR was used to establish integration of the EGF-L gene into the
Pichia genome. Thereafter, the Pichia was induced with methanol for 72 h, media were
collected, centrifuged, concentrated, and total protein was precipitated with 50%
ammonium sulfate.
Purification and Characterization of the Recombinant EGF: The ammonium sulfate
pellet was resuspended in 10% acetonitrile (ACN) and loaded onto a C8 reverse-phase
column which was eluted with a gradient of 10-60% ACN containing 0.1 %
trifluoroacetic acid. Eluted fractions were tested for the presence of EGF-L by SDS-
PAGE and Western blot. Peak fractions were pooled, diluted two-fold with water,
neutralized by the addition of 0.5M Tris-HCl (pH 8), and loaded onto a Q-Sepharose
column. The column was eluted with a gradient of 20-500 mM NaCl. Peak fractions
were pooled, and then concentrated with an Amicon YM10 membrane. After desalting,
this sample was used for N-terminal sequencing (UCLA Microsequencing Core Facility).
The sequence of the N-terminal 25 amino acid residues was identical to mature EGF-L,
demonstrating the complete processing of the prepro-peptide (not shown). No
heterogeneity was detected in the N-terminal sequence or amino acid composition.
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement The elution profile of the purified EGF on gel filtration HPLC was consistent with its
extended conformation and its monomeric structure. To determine whether the disulfide
bond formation is complete, the sulfhydryl content of the EGF was measured using 5,5'-
dithiobis-(2-nitrobenzoic acid), and the color formed was measured at 412 nm 5. No free
sulfhydryl was detected. To determine glycosylation of the secreted EGF, it was treated
with F-glycosidase followed by SDS-PAGE analysis. The mobility of the glycosidase-
treated protein was similar to the untreated EGF. All recombinant proteins were tested
for endotoxin activity using E-TOXATE kit (Sigma). No detectable endotoxin was found
in the preparations.
Antibody techniques: The anti-Tn-C monoclonal antibody was obtained from Sigma
(clone BC-24). The antibody recognizes an epitope within the N-terminal EGF sequence
of the molecule. The antibodies to the fibronectin-like domain of Tn-C were a gift from
Dr. Erickson (Duke University). Tn-C degradation was measured by Western blot as we
previously described 6. Briefly, 25 µl of 25% SDS was added to the cultured SMCs
and shaken for 1 h. The samples were run on a 5% SDS-polyacrylamide gel
electrophoresis, and proteins were transferred to polyvinylidene difluoride membrane
(Bio-Rad). The membrane was blotted with polyclonal anti-fibronectin-like and EGF-
like domains of Tn-C at 1:5000 and 1:10,000 dilutions, and signals were detected by
the enhanced chemiluminescence detection system (Amersham).
Human carotid endarterectomy specimens were obtained after surgery and immediately
snap frozen with liquid nitrogen. The frozen samples were pulverized and then extracted
with a buffer (0.1 M CAPS, 0.15 M NaCl, pH 11) known to be effective in extracting Tn-
C 7. After centrifugation, the protein concentration of the supernatants was determined,
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement and 20 µg from each sample were electrophoresed in a 5% SDS-PAGE. After transfer,
the blots were incubated with 1:10,000 dilution of anti-EGF monoclonal antibodies.
Western blots were developed as described above.
Cell apoptosis: Cell apoptosis was measured by several methods. 1) TUNEL staining:
cells were cultured on 4-well slides as described above and treated with or without EGF.
TUNEL staining was performed essentially as described by the manufacturer
(Oncogene). 2) Externalization of phosphatidylserine was measured by annexin V
labeling (Boehringer Mannheim) using flow cytometry. For FACS analysis, cells were
stained by fluorescent-conjugated annexin V and by the fluorescent dye propidium iodide
(PI) as recommended by manufacturer. Cells negative for both PI and annexin V staining
are considered live cells; annexin V-positive, PI negative cells are considered at early
apoptotic stage; annexin V-positive PI-positive cells are considered in late stages of
apoptosis 8,9. As a positive control, cells were treated with 250 ng/ml of actinomycin D
for 24 h. 3) MTT assay: cells were plated in triplicate in 96-well microtiter plates in 200
µl of DMEM+10% serum and incubated overnight. The next day, media were removed
and replaced with defined media in the absence and presence of different concentrations
of EGF or other reagents. The plates were incubated for an additional 24-48 h, and cell
viability was determined using a MTT assay, as described by the manufacturer (Sigma).
To measure caspase-3 activation, human SMCs were washed twice with ice-cold PBS
and then lysed with a solution containing 150 mM NaCl, 1%NP-40, 50 mM Tris-HCl, pH
8.0, and 1X complete protease inhibitor (Boehringer Mannheim). After 15 min on ice,
lysates were vortexed and cleared by centrifugation. After protein determination, 50 µg
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement from each sample was analyzed by 10% SDS-PAGE gel. The proteins were transferred
to nitrocellulose (BioRad) and Western-blotted using anti-caspase-3 antibodies (H-277,
Santa Cruz). This polyclonal antibody recognizes both inactive (32 kDa) and 11 kDa
cleavage product of human, rat, and mouse caspase-3. Western blot signals were
detected as described above.
Statistical analysis: All the experiments have been performed in triplicate and repeated 3
times with different preparations of Tn-C or recombinant EGF-L domain to ensure
reproducibility. The results are either representative of 3 experiments, or average + SEM
of a triplicate determinations.
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement Figure legends
Figure I. Apoptosis of SMCs by Tn-C. (Panel A) Human SMCs (5x105 cells/well)
were cultured in defined media onto 24 well dishes that were coated with fibronectin (top
panel) or Tn-C (lower panel) for 24 h. Conditioned media were collected for the
experiments described in panel C. (Panel B) human SMCs were cultured onto Tn-C-
coated 4-well slide dishes (3x103/well) in the presence (top panel) or absence (lower
panel) of 25 µM GM6001 MMP inhibitor in defined media. TUNEL staining was
performed as described in the Method section. (Panel C) Collected conditioned media
(500 µl, described in panel A) were incubated with 1:50 dilution of antibodies to intact
Tn-C (Tn-C), fibronectin-like (Fn-L), fibrinogen-like (Fbg-L), or EGF-like domain
(EGF-L) of Tn-C for 1 h at 37oC followed by addition of 50 µl of agarose-protein A.
Fibronectin-coated wells (Fn) were used as controls. The supernatants were
supplemented with 200 µl of defined media and then added to SMCs that have been
cultured onto fibronectin-coated 96-well plates (1x104/well) in defined media. After
incubation at 37oC for 24 h, SMC viability was measured after 24 h by standard MTT
assay. Each data point represents the average of triplicate determinations + SEM.
Fig. II. EGF-like domain inhibits SMC DNA synthesis. (Panel A) Human SMCs
were plated at a density of 1x104/well in triplicate in 96-well microtiter plates in
DMEM+10% serum. After 24 h, media were removed and replaced with DMEM+1%
serum. Next day, media were removed and replaced with DMEM defined media
supplemented with 0.1 µM of either intact Tn-C, recombinant full-length fibronectin-like
(Fn-like), fibrinogen-like (Fbg-like), or EGF-like (EGF-like) domains in the absence or
+ EGF-LCor
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement presence of 1 nM PDGF-BB. At 24 h, 1 µCi/ml of 3[H]thymidine was added and cells
were incubated for 4 h at 37oC. Macromolecules were precipitated with 10%
tricholoroacetic acid followed by cell lysis with 1N NaOH. Radioactivity was measured
with a scintillation counter. Each data point represents the average of triplicate
determinations. (Panel B) SMCs were cultured in defined media on fibronectin-coated 6-
well dishes (1x106 cells/well). 24 h after plating, cells were treated with 500 ng/ml of
actinomycin D or 0.01 µM of recombinant EGF-L domain for 24 h. Untreated cells were
used as a control. After treatment, cells were harvested (both attached and detached) and
externalization of phosphatidylserine was measured by annexin V labeling (Boehringer
Mannheim) using flow cytometry. The experiment was repeated 3 times with 3 different
preparations of recombinant EGF-L domain.
Figure III Caspase inhibitors block SMC apoptosis induced by EGF-like domain.
(Panel A) Human SMCs were plated at density of 1x104/well in triplicate in 96-well
microtiter plates in DMEM+10% serum. After 24 h, media were removed and replaced
with DMEM defined media containing 200 µM inhibitor alone (inhibit) or 0.01 µM
recombinant EGF-like domain in the presence of indicated concentrations of caspase
inhibitors. Control media contained no inhibitor or recombinant EGF-like domain. After
24 h, cell viability was determined by MTT assay. Each data point represents the average
of three separate experiments. (Panel B, top panels) Cells were cultured in 10 cm2
culture dishes in 10% serum. After 24 h, the media were replaced with the defined media
in the absence (C) or presence of 0.01 µM solution of recombinant EGF-like domain.
After indicated times (h), attached and detached cells were lysed and the lysates were
used for Western blot analysis of caspase-3 using anti-caspase-3 antibody (H-277, Santa
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

ATVB/2004/015636-R1 ON-LINE Supplement Cruz). B. (Lower panel) SMCs were cultured in the presence of recombinant EGF-like
domain supplemented with increasing concentrations of Z-YVAD for 24 h. Thereafter,
cell lysates were prepared and analyzed by Western blot.
1. Sharifi BG, LaFleur DW, Pirola CJ, Forrester JS, Fagin JA. Angiotensin II regulates tenascin gene expression in vascular smooth muscle cells. J Biol Chem. 1992;267:23910-23915.
2. LaFleur DW, Chiang J, Fagin JA, Schwartz SM, Shah PK, Wallner K, Forrester JS, Sharifi BG. Aortic smooth muscle cells interact with tenascin-C through its fibrinogen-like domain. J Biol Chem. 1997;272:32798-32803.
3. Wallner K, Li C, Shah PK, Fishbein MC, Forrester JS, Kaul S, Sharifi BG. Tenascin-C Is Expressed in Macrophage-Rich Human Coronary Atherosclerotic Plaque. Circulation. 1999;99:1284-1289.
4. LaFleur DW, Fagin JA, Forrester JS, Rubin SA, Sharifi BG. Cloning and characterization of alternatively spliced isoforms of rat tenascin. Platelet-derived growth factor-BB markedly stimulates expression of spliced variants of tenascin mRNA in arterial smooth muscle cells. J Biol Chem. 1994;269:20757-20763.
5. Aukhil I, Joshi P, Yan Y, Erickson HP. Cell- and heparin-binding domains of the hexabrachion arm identified by tenascin expression proteins. J Biol Chem. 1993;268:2542-2553.
6. Ikari Y, Mulvihill E, Schwartz SM. alpha 1-Proteinase Inhibitor, alpha 1-Antichymotrypsin, and alpha 2-Macroglobulin Are the Antiapoptotic Factors of Vascular Smooth Muscle Cells. J. Biol. Chem. 2001;276:11798-11803.
7. Lightner VA, Gumkowski F, Bigner DD, Erickson HP. Tenascin/hexabrachion in human skin: biochemical identification and localization by light and electron microscopy. J Cell Biol. 1989;108:2483-2493.
8. Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J Immunol. 1992;148:2207-2216.
9. Koopman G, Reutelingsperger CP, Kuijten GA, Keehnen RM, Pals ST, van Oers MH. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood. 1994;84:1415-1420.
by on May 23, 2011 atvb.ahajournals.orgDownloaded from

Related Documents











