OBESITY | VOLUME 16 NUMBER 10 | OCTOBER 2008 2217 nature publishing group ARTICLES INTEGRATIVE PHYSIOLOGY Efficiency of Intermittent Exercise on Adiposity and Fatty Liver in Rats Fed With High-fat Diet Marcela Sene-Fiorese 1,2 , Fernanda O. Duarte 1 , Flavia R.R. Scarmagnani 1 , Nadia C. Cheik 1 , Marla S.J. Manzoni 1 , Keico O. Nonaka 3 , Eliseu A. Rossi 4 , Ana C.G. de Oliveira Duarte 1 and Ana R. Dâmaso 5 Objective: This study aimed to examine and compare the effects of continuous or intermittent exercises on adiposity and fatty liver in rats fed with high-fat diet. Methods and Procedures: Wistar rats were divided according to diet composition—chow diet (C) or high-fat diet (H)—and kinds of exercise—sedentary (S), continuous (CE), or intermittent (IE) exercises. The CE group swam 90 min/ day, and the IE group swam 3 × 30 min/day (at 4-h intervals between sessions); both groups exercised 5 days/week during 8 weeks. Body weight and food intake were recorded daily. Lipogenesis rate in vivo was determined by the incorporation of 3 H 2 O into saponified lipids in retroperitoneal (RET), epididymal (EPI), and visceral (VIS) white adipose tissues, brown adipose tissue (BAT), liver (L), and gastrocnemius muscle (GAST) using the gravimetric method. Total cholesterol, high-density lipoprotein (HDL)-cholesterol, and triacylglycerol (TG) were analyzed. Results: The major finding of this study is that IE was more efficient than CE in reducing the adverse effects of high-fat diet and sedentarism. There was an improvement in the lipid profile and a reduction in food intake, body weight gain, visceral and central adiposity, and fatty liver, contributing to the control of obesity and other comorbidities, including nonalcoholic fat liver diseases. Discussion: Earlier studies have discussed the effects of diet consumption on adiposity and their relation to chronic diseases and obesity. This study discusses the effects of high-fat diet consumption and the different kinds of exercise on weight gain, adiposity, fatty liver, and lipid profile in rats. The results may depend on the exercise, time of each session, age, gender, and experimental period. Obesity (2008) 16, 2217–2222. doi:10.1038/oby.2008.339 INTRODUCTION Obesity is now one of the serious public health problems in the world. In Brazil, a developing country, it is rapidly becom- ing the most prevalent nutritional disorder, characterized by a decrease in the prevalence of undernutrition (1). It is strongly associated with many chronic diseases, mainly obesity with visceral adiposity distribution, such as dyslipidemias, hyper- tension, diabetes, coronary atherosclerotic heart disease, can- cer, nonalcoholic fatty liver disease (2–6). Among all other environmental influences, both high-fat diet and lack of or decline in daily physical activity are the most important factors for obesity development (3,7). us, increasing physical activity has become an important part of a nonpharmacological strategy to control obesity and weight gain (8). In this sense, the guidelines by the American College of Sports Medicine and the Centers for Disease Control and Prevention stipulate that every sedentary adult should accumulate 30 min or more of moderate-intensity physical activity during most days of the week for health promotion and disease prevention (9–11). However, since these reports were published, several researchers have questioned whether this recommendation is sufficient to prevent unhealthy weight (re)gain (8). e US Institute of Medicine panel showed in a recent study that the minimal amount of accumulated physical activity to achieve a degree of protection from chronic diseases (i.e., 30 min/day) was not sufficient to maintain body weight in a desirable BMI range (from 18.5 to <25 kg/m 2 ). e equiva- lent of 60 min of accumulated moderate-intensity physical activity per day was required to prevent undesirable body fat gain (12). 1 Laboratory of Nutrition and Metabolism Applied in Exercise, Department of Physical Education, Center of Biological and Health Sciences, Federal University of São Carlos, São Paulo, Brazil; 2 Faculty of Physical Education, University Camilo Castelo Branco, Descalvado, São Paulo, Brazil; 3 Department of Physiological Sciences, Center of Biological and Health Sciences, Federal University of São Carlos, São Paulo, Brazil; 4 Department of Food and Nutrition, Faculty of Pharmaceutics Sciences, University Júlio de Mesquita Filho, Araraquara, Brazil; 5 Department of Health Sciences, Federal University of São Paulo, Paulista Medicine School, São Paulo, Brazil. Correspondence: Marcela Sene-Fiorese ([email protected]) Received 2 April 2007; accepted 28 December 2007; published online 24 July 2008. doi:10.1038/oby.2008.339

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

obesity | VOLUME 16 NUMBER 10 | OCTOBER 2008 2217
nature publishing group articlesintegrative physiology
Efficiency of Intermittent Exercise on Adiposity and Fatty Liver in Rats Fed With High-fat DietMarcela Sene-Fiorese1,2, Fernanda O. Duarte1, Flavia R.R. Scarmagnani1, Nadia C. Cheik1, Marla S.J. Manzoni1, Keico O. Nonaka3, Eliseu A. Rossi4, Ana C.G. de Oliveira Duarte1 and Ana R. Dâmaso5
objective: This study aimed to examine and compare the effects of continuous or intermittent exercises on adiposity and fatty liver in rats fed with high-fat diet.Methods and Procedures: Wistar rats were divided according to diet composition—chow diet (C) or high-fat diet (H)—and kinds of exercise—sedentary (S), continuous (CE), or intermittent (IE) exercises. The CE group swam 90 min/day, and the IE group swam 3 × 30 min/day (at 4-h intervals between sessions); both groups exercised 5 days/week during 8 weeks. Body weight and food intake were recorded daily. Lipogenesis rate in vivo was determined by the incorporation of 3H2O into saponified lipids in retroperitoneal (RET), epididymal (EPI), and visceral (VIS) white adipose tissues, brown adipose tissue (BAT), liver (L), and gastrocnemius muscle (GAST) using the gravimetric method. Total cholesterol, high-density lipoprotein (HDL)-cholesterol, and triacylglycerol (TG) were analyzed.Results: The major finding of this study is that IE was more efficient than CE in reducing the adverse effects of high-fat diet and sedentarism. There was an improvement in the lipid profile and a reduction in food intake, body weight gain, visceral and central adiposity, and fatty liver, contributing to the control of obesity and other comorbidities, including nonalcoholic fat liver diseases.Discussion: Earlier studies have discussed the effects of diet consumption on adiposity and their relation to chronic diseases and obesity. This study discusses the effects of high-fat diet consumption and the different kinds of exercise on weight gain, adiposity, fatty liver, and lipid profile in rats. The results may depend on the exercise, time of each session, age, gender, and experimental period.
Obesity (2008) 16, 2217–2222. doi:10.1038/oby.2008.339
IntroductIonObesity is now one of the serious public health problems in the world. In Brazil, a developing country, it is rapidly becom-ing the most prevalent nutritional disorder, characterized by a decrease in the prevalence of undernutrition (1). It is strongly associated with many chronic diseases, mainly obesity with visceral adiposity distribution, such as dyslipidemias, hyper-tension, diabetes, coronary atherosclerotic heart disease, can-cer, nonalcoholic fatty liver disease (2–6). Among all other environmental influences, both high-fat diet and lack of or decline in daily physical activity are the most important factors for obesity development (3,7).
Thus, increasing physical activity has become an important part of a nonpharmacological strategy to control obesity and weight gain (8). In this sense, the guidelines by the American
College of Sports Medicine and the Centers for Disease Control and Prevention stipulate that every sedentary adult should accumulate 30 min or more of moderate-intensity physical activity during most days of the week for health promotion and disease prevention (9–11). However, since these reports were published, several researchers have questioned whether this recommendation is sufficient to prevent unhealthy weight (re)gain (8). The US Institute of Medicine panel showed in a recent study that the minimal amount of accumulated physical activity to achieve a degree of protection from chronic diseases (i.e., 30 min/day) was not sufficient to maintain body weight in a desirable BMI range (from 18.5 to <25 kg/m2). The equiva-lent of 60 min of accumulated moderate-intensity physical activity per day was required to prevent undesirable body fat gain (12).
1Laboratory of Nutrition and Metabolism Applied in Exercise, Department of Physical Education, Center of Biological and Health Sciences, Federal University of São Carlos, São Paulo, Brazil; 2Faculty of Physical Education, University Camilo Castelo Branco, Descalvado, São Paulo, Brazil; 3Department of Physiological Sciences, Center of Biological and Health Sciences, Federal University of São Carlos, São Paulo, Brazil; 4Department of Food and Nutrition, Faculty of Pharmaceutics Sciences, University Júlio de Mesquita Filho, Araraquara, Brazil; 5Department of Health Sciences, Federal University of São Paulo, Paulista Medicine School, São Paulo, Brazil. Correspondence: Marcela Sene-Fiorese ([email protected])
Received 2 April 2007; accepted 28 December 2007; published online 24 July 2008. doi:10.1038/oby.2008.339
2218 VOLUME 16 NUMBER 10 | OCTOBER 2008 | www.obesityjournal.org
articlesintegrative physiology
Several experimental studies have demonstrated that con-tinuous moderate swimming exercise (i.e., 90 min) associ-ated with chow diet resulted in the reduction of adiposity and improved lipid profile in rats. However, when the animals were fed with high-fat diet, these effects were not observed (13–15). An important question to this discussion is: “Does moderate intermittent exercise (IE) (accumulated 90 min/day) have the same effect in metabolism and body weight gain in rats?”
To our knowledge, the comparison between the effects of CE or IE has not been established yet. Based on the previous find-ings, the purpose of this study was to compare the effect of IE or CE on body weight gain, central and visceral adiposity, fatty liver, lipogenesis rate, and lipid profile in rats fed with chow or high-fat diets.
Methods and ProceduresanimalsMale Wistar rats (90–120 day old) were supplied by a local breeding facility (Federal University of São Carlos) and were kept in a single cage with food and water ad libitum (8 weeks), on a 12:12-h light–dark cycle at 23 ± 1 °C. The animals were assigned to one of the six groups (n = 8) according to their diet composition—chow diet (C) or high-fat diet (H)—and their kind of physical activity—sedentary (S) and continu-ous (CE—one session of 90 min/day, suggesting one daily increment in energy expenditure) or intermittent (IE—three sessions of 30 min/day, suggesting three daily increments in energy expenditure) exercises. All animal procedures were performed according to the principles in the Guide for the Care and Use of Laboratory Animals (16), and were approved by the Institutional Animal Ethics Committee.
dietsExperimental groups received the following diets: MP-77 standard rat chow diet (in pellet form) provided by Primor (São Paulo, Brazil) and contained 23 g of protein (per 100 g diet), 49 g of carbohydrate, 4 g of total fat, 5 g of fiber, 7 g of ash, and 6 g of vitamins; the high-fat diet con-sisted of commercial rat chow plus peanuts, milk chocolate, and sweet biscuit in a proportion of 3:2:2:1. All components of the high-fat diet were ground and blended. This diet achieved 20% protein, 20% fat, 48% carbohydrate, and 4% fiber. The caloric density of the diets was deter-mined with an adiabatic calorimeter (IKA-C400); it was 5.12 kcal/g (35% of calories as fat) for the high-fat diet and 4.07 kcal/g for the chow diet (14).
exercise protocolsThe exercise programs consisted of continuous or intermittent swimming in individual tanks filled with water and maintained at 28–32 °C. The animals of the continuous group (CE) swam for 30, 60, and 90 min on the first, second, and third days to adapt. The swimming period was then increased to 90 min/day. The animals of the intermittent group (IE) swam three sessions for 10, 20, and 30 min on the first, second, and third days to adapt. The swimming period was 90 min/day, divided into three daily sessions (3 × 30 min/day) with 4 h of controlled intervals between the exercise periods throughout the experimental period. The rats were kept in their home cages all the time between the exercise periods, performed at 7 am, 11:30 am, and 3:30 pm. All the rats swam with a load of 5% body weight (attached to the tail of each rat). The exercise protocols were performed for 5 days/week during 8 weeks. Both exercise pro-grams are considered to be of moderate intensity (17).
MeasurementsBody weight and food intake were recorded daily.
experimental procedureAt the end of 8 weeks and after a 24-h rest period, the experiments were performed. All animals were killed by decapitation 1 h after an intraperitoneal injection of 3 mCi 3H2O in a volume of 0.3 ml for the determination of the in vivo lipogenesis rate. Trunk blood was drawn into a heparinized tube. Liver (L), retroperitoneal (RET), epididymal (EPI), visceral (VIS) white adipose tissues, gastrocnemius muscle (GAST), and interscapular brown adipose tissue (BAT) were immedi-ately removed and weighed (18). The in vivo lipogenesis rate (fatty acid synthesis) was determined by the incorporation of 3H2O into saponified lipids according to the method of Robinson and Williamson (19). The rate of lipogenesis was calculated as micromoles of 3H2O incorporated into lipids per gram per hour. The percentage of lipid content in the tis-sue was determined by the gravimetric method (20).
serum analysisTotal cholesterol, high-density lipoprotein (HDL)-cholesterol, and tria-cylglycerol (TG) in the plasma were determined enzymatically (CELM, Barueri, Brazil) using an ultraviolet-visible spectrophotometer (model UV-160 1PC; Shimadzu, Kyoto, Japan).
statistical analysisResults are expressed as mean ± s.e. Data analysis was done using Instat 3.0 for Windows 95 (Graph Pad, San Diego, CA, 1998). Statistical com-parisons were carried out by one-way analysis of variance. Statistical significance was set at P < 0.01. Significant results of analysis of vari-ance were subjected to post hoc analysis (Tukey–Kramer multiple comparisons).
resultsBody weight gain in rats fed with both diets did not differ sig-nificantly between sedentary and continuous exercise groups. Although food consumption decreased in sedentary rats fed with high-fat diet (SH), total caloric intake was greater when compared with sedentary rats fed with chow diet (SC). On the other hand, IE in rats fed with high-fat diet promoted a sig-nificant decrease in body weight gain without alteration in the food consumption and total caloric intake (Table 1).
High-fat diet promoted in the sedentary group an increase in EPI, RET, VIS, and BAT relative weight compared with what was observed in the sedentary rats fed with chow diet group (SC), except the GAST relative weight, which was smaller. No difference was observed in this parameter when comparing
table 1 effects of exercise and diet on body weight gain, food consumption, and caloric intake
GroupsBody weight
gain (g)Food
consumption (g)Caloric intake
(kcal)
SC 214.00 ± 8.26 1,544.60 ± 19.40 6,062.90 ± 73.47
CEC 171.43 ± 7.07 1,520.20 ± 23.44 6,332.20 ± 130.32
IEC 140.88 ± 10.86* 1,523.50 ± 24.66 5,960.50 ± 125.30
SH 224.00 ± 7.29 1,357.90 ± 19.68* 6,903.40 ± 99.90*
CEH 199.25 ± 15.87 1,417.10 ± 37.83 6,765.20 ± 131.60
IEH 92.43 ± 9.83**,*** 1,283.30 ± 36.40 6,613.30 ± 182.71
Values are means ± s.e.m. of eight animals. Symbols indicate significant differ-ences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH; ***vs. CEH.CEC, exercised continuous rats fed with chow diet; CEH, exercised continuous rats fed with high-fat diet; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet.
obesity | VOLUME 16 NUMBER 10 | OCTOBER 2008 2219
articlesintegrative physiology
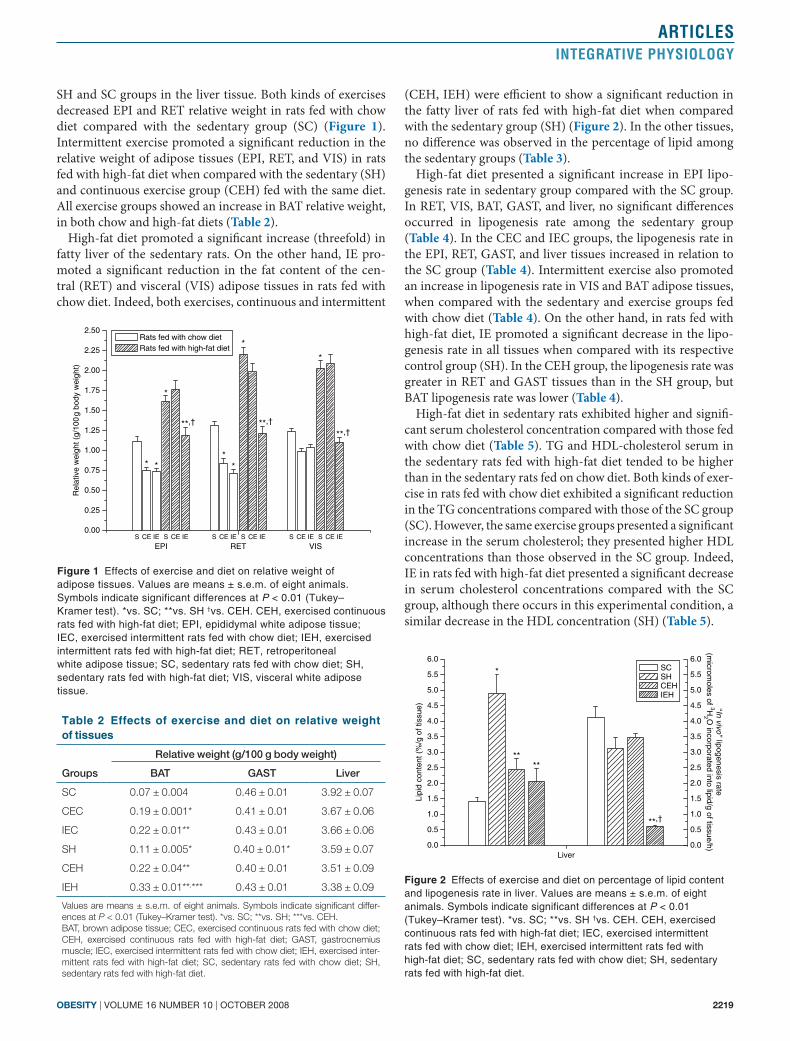
SH and SC groups in the liver tissue. Both kinds of exercises decreased EPI and RET relative weight in rats fed with chow diet compared with the sedentary group (SC) (Figure 1). Intermittent exercise promoted a significant reduction in the relative weight of adipose tissues (EPI, RET, and VIS) in rats fed with high-fat diet when compared with the sedentary (SH) and continuous exercise group (CEH) fed with the same diet. All exercise groups showed an increase in BAT relative weight, in both chow and high-fat diets (Table 2).
High-fat diet promoted a significant increase (threefold) in fatty liver of the sedentary rats. On the other hand, IE pro-moted a significant reduction in the fat content of the cen-tral (RET) and visceral (VIS) adipose tissues in rats fed with chow diet. Indeed, both exercises, continuous and intermittent
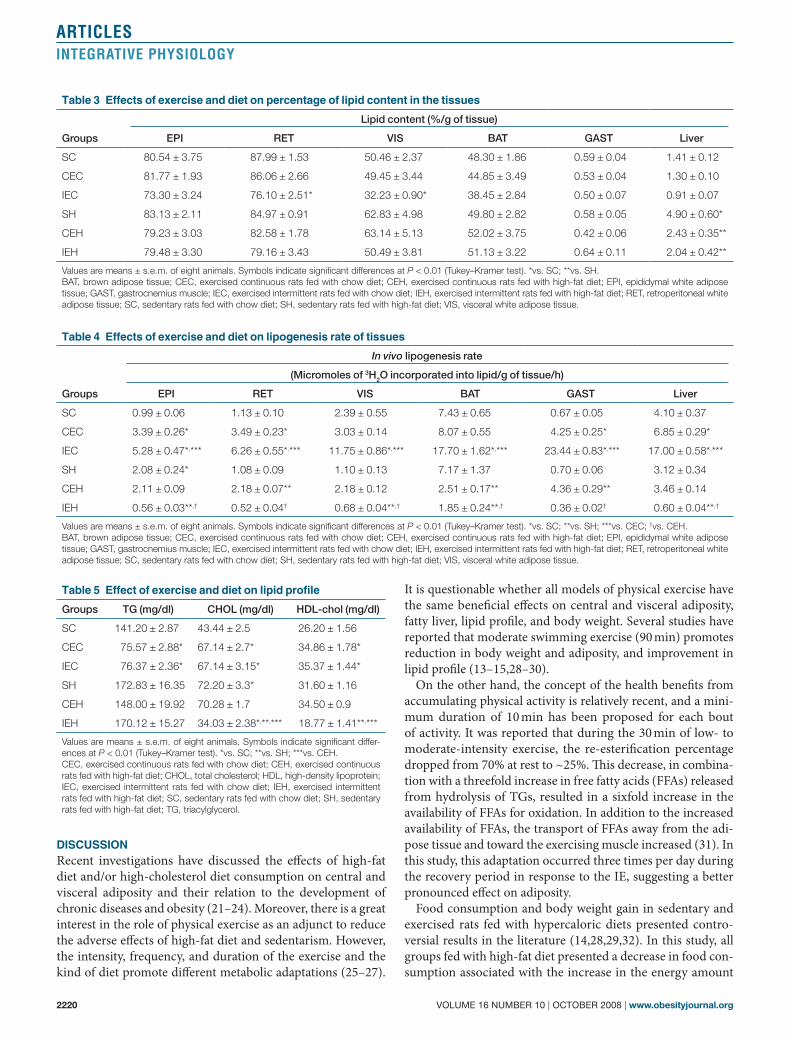
(CEH, IEH) were efficient to show a significant reduction in the fatty liver of rats fed with high-fat diet when compared with the sedentary group (SH) (Figure 2). In the other tissues, no difference was observed in the percentage of lipid among the sedentary groups (Table 3).
High-fat diet presented a significant increase in EPI lipo-genesis rate in sedentary group compared with the SC group. In RET, VIS, BAT, GAST, and liver, no significant differences occurred in lipogenesis rate among the sedentary group (Table 4). In the CEC and IEC groups, the lipogenesis rate in the EPI, RET, GAST, and liver tissues increased in relation to the SC group (Table 4). Intermittent exercise also promoted an increase in lipogenesis rate in VIS and BAT adipose tissues, when compared with the sedentary and exercise groups fed with chow diet (Table 4). On the other hand, in rats fed with high-fat diet, IE promoted a significant decrease in the lipo-genesis rate in all tissues when compared with its respective control group (SH). In the CEH group, the lipogenesis rate was greater in RET and GAST tissues than in the SH group, but BAT lipogenesis rate was lower (Table 4).
High-fat diet in sedentary rats exhibited higher and signifi-cant serum cholesterol concentration compared with those fed with chow diet (Table 5). TG and HDL-cholesterol serum in the sedentary rats fed with high-fat diet tended to be higher than in the sedentary rats fed on chow diet. Both kinds of exer-cise in rats fed with chow diet exhibited a significant reduction in the TG concentrations compared with those of the SC group (SC). However, the same exercise groups presented a significant increase in the serum cholesterol; they presented higher HDL concentrations than those observed in the SC group. Indeed, IE in rats fed with high-fat diet presented a significant decrease in serum cholesterol concentrations compared with the SC group, although there occurs in this experimental condition, a similar decrease in the HDL concentration (SH) (Table 5).
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
2.25
2.50
IE IECECE S
**,† **,†
**,†
**
*
*
*
*
*
S IE IECECE SS IE IECECE SS
VISRET
Rel
ativ
e w
eigh
t (g/
100
g bo
dy w
eigh
t)
EPI
Rats fed with chow dietRats fed with high-fat diet
Figure 1 Effects of exercise and diet on relative weight of adipose tissues. Values are means ± s.e.m. of eight animals. Symbols indicate significant differences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH †vs. CEH. CEH, exercised continuous rats fed with high-fat diet; EPI, epididymal white adipose tissue; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; RET, retroperitoneal white adipose tissue; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet; VIS, visceral white adipose tissue.
table 2 effects of exercise and diet on relative weight of tissues
Groups
Relative weight (g/100 g body weight)
BAT GAST Liver
SC 0.07 ± 0.004 0.46 ± 0.01 3.92 ± 0.07
CEC 0.19 ± 0.001* 0.41 ± 0.01 3.67 ± 0.06
IEC 0.22 ± 0.01** 0.43 ± 0.01 3.66 ± 0.06
SH 0.11 ± 0.005* 0.40 ± 0.01* 3.59 ± 0.07
CEH 0.22 ± 0.04** 0.40 ± 0.01 3.51 ± 0.09
IEH 0.33 ± 0.01**,*** 0.43 ± 0.01 3.38 ± 0.09
Values are means ± s.e.m. of eight animals. Symbols indicate significant differ-ences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH; ***vs. CEH.BAT, brown adipose tissue; CEC, exercised continuous rats fed with chow diet; CEH, exercised continuous rats fed with high-fat diet; GAST, gastrocnemius muscle; IEC, exercised intermittent rats fed with chow diet; IEH, exercised inter-mittent rats fed with high-fat diet; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
6.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
6.0
**,†
****
*
“In vivo” lipogenesis rate
(microm
oles of 3H2 O
incorporated into lipid/g of tissue/h)
Lipi
d co
nten
t (%
/g o
f tis
sue)
Liver
SCSHCEHIEH
Figure 2 Effects of exercise and diet on percentage of lipid content and lipogenesis rate in liver. Values are means ± s.e.m. of eight animals. Symbols indicate significant differences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH †vs. CEH. CEH, exercised continuous rats fed with high-fat diet; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet.
2220 VOLUME 16 NUMBER 10 | OCTOBER 2008 | www.obesityjournal.org
articlesintegrative physiology
It is questionable whether all models of physical exercise have the same beneficial effects on central and visceral adiposity, fatty liver, lipid profile, and body weight. Several studies have reported that moderate swimming exercise (90 min) promotes reduction in body weight and adiposity, and improvement in lipid profile (13–15,28–30).
On the other hand, the concept of the health benefits from accumulating physical activity is relatively recent, and a mini-mum duration of 10 min has been proposed for each bout of activity. It was reported that during the 30 min of low- to moderate-intensity exercise, the re-esterification percentage dropped from 70% at rest to ~25%. This decrease, in combina-tion with a threefold increase in free fatty acids (FFAs) released from hydrolysis of TGs, resulted in a sixfold increase in the availability of FFAs for oxidation. In addition to the increased availability of FFAs, the transport of FFAs away from the adi-pose tissue and toward the exercising muscle increased (31). In this study, this adaptation occurred three times per day during the recovery period in response to the IE, suggesting a better pronounced effect on adiposity.
Food consumption and body weight gain in sedentary and exercised rats fed with hypercaloric diets presented contro-versial results in the literature (14,28,29,32). In this study, all groups fed with high-fat diet presented a decrease in food con-sumption associated with the increase in the energy amount
dIscussIonRecent investigations have discussed the effects of high-fat diet and/or high-cholesterol diet consumption on central and visceral adiposity and their relation to the development of chronic diseases and obesity (21–24). Moreover, there is a great interest in the role of physical exercise as an adjunct to reduce the adverse effects of high-fat diet and sedentarism. However, the intensity, frequency, and duration of the exercise and the kind of diet promote different metabolic adaptations (25–27).
table 3 effects of exercise and diet on percentage of lipid content in the tissues
Groups
Lipid content (%/g of tissue)
EPI RET VIS BAT GAST Liver
SC 80.54 ± 3.75 87.99 ± 1.53 50.46 ± 2.37 48.30 ± 1.86 0.59 ± 0.04 1.41 ± 0.12
CEC 81.77 ± 1.93 86.06 ± 2.66 49.45 ± 3.44 44.85 ± 3.49 0.53 ± 0.04 1.30 ± 0.10
IEC 73.30 ± 3.24 76.10 ± 2.51* 32.23 ± 0.90* 38.45 ± 2.84 0.50 ± 0.07 0.91 ± 0.07
SH 83.13 ± 2.11 84.97 ± 0.91 62.83 ± 4.98 49.80 ± 2.82 0.58 ± 0.05 4.90 ± 0.60*
CEH 79.23 ± 3.03 82.58 ± 1.78 63.14 ± 5.13 52.02 ± 3.75 0.42 ± 0.06 2.43 ± 0.35**
IEH 79.48 ± 3.30 79.16 ± 3.43 50.49 ± 3.81 51.13 ± 3.22 0.64 ± 0.11 2.04 ± 0.42**
Values are means ± s.e.m. of eight animals. Symbols indicate significant differences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH.BAT, brown adipose tissue; CEC, exercised continuous rats fed with chow diet; CEH, exercised continuous rats fed with high-fat diet; EPI, epididymal white adipose tissue; GAST, gastrocnemius muscle; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; RET, retroperitoneal white adipose tissue; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet; VIS, visceral white adipose tissue.
table 4 effects of exercise and diet on lipogenesis rate of tissues
In vivo lipogenesis rate
(Micromoles of 3H2O incorporated into lipid/g of tissue/h)
Groups EPI RET VIS BAT GAST Liver
SC 0.99 ± 0.06 1.13 ± 0.10 2.39 ± 0.55 7.43 ± 0.65 0.67 ± 0.05 4.10 ± 0.37
CEC 3.39 ± 0.26* 3.49 ± 0.23* 3.03 ± 0.14 8.07 ± 0.55 4.25 ± 0.25* 6.85 ± 0.29*
IEC 5.28 ± 0.47*,*** 6.26 ± 0.55*,*** 11.75 ± 0.86*,*** 17.70 ± 1.62*,*** 23.44 ± 0.83*,*** 17.00 ± 0.58*,***
SH 2.08 ± 0.24* 1.08 ± 0.09 1.10 ± 0.13 7.17 ± 1.37 0.70 ± 0.06 3.12 ± 0.34
CEH 2.11 ± 0.09 2.18 ± 0.07** 2.18 ± 0.12 2.51 ± 0.17** 4.36 ± 0.29** 3.46 ± 0.14
IEH 0.56 ± 0.03**,† 0.52 ± 0.04† 0.68 ± 0.04**,† 1.85 ± 0.24**,† 0.36 ± 0.02† 0.60 ± 0.04**,†
Values are means ± s.e.m. of eight animals. Symbols indicate significant differences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH; ***vs. CEC; †vs. CEH.BAT, brown adipose tissue; CEC, exercised continuous rats fed with chow diet; CEH, exercised continuous rats fed with high-fat diet; EPI, epididymal white adipose tissue; GAST, gastrocnemius muscle; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; RET, retroperitoneal white adipose tissue; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet; VIS, visceral white adipose tissue.
table 5 effect of exercise and diet on lipid profile
Groups TG (mg/dl) CHOL (mg/dl) HDL-chol (mg/dl)
SC 141.20 ± 2.87 43.44 ± 2.5 26.20 ± 1.56
CEC 75.57 ± 2.88* 67.14 ± 2.7* 34.86 ± 1.78*
IEC 76.37 ± 2.36* 67.14 ± 3.15* 35.37 ± 1.44*
SH 172.83 ± 16.35 72.20 ± 3.3* 31.60 ± 1.16
CEH 148.00 ± 19.92 70.28 ± 1.7 34.50 ± 0.9
IEH 170.12 ± 15.27 34.03 ± 2.38*,**,*** 18.77 ± 1.41**,***
Values are means ± s.e.m. of eight animals. Symbols indicate significant differ-ences at P < 0.01 (Tukey–Kramer test). *vs. SC; **vs. SH; ***vs. CEH.CEC, exercised continuous rats fed with chow diet; CEH, exercised continuous rats fed with high-fat diet; CHOL, total cholesterol; HDL, high-density lipoprotein; IEC, exercised intermittent rats fed with chow diet; IEH, exercised intermittent rats fed with high-fat diet; SC, sedentary rats fed with chow diet; SH, sedentary rats fed with high-fat diet; TG, triacylglycerol.

obesity | VOLUME 16 NUMBER 10 | OCTOBER 2008 2221
articlesintegrative physiology
acclimatization that suppresses lower thermogenesis induced by high-fat diet. Previous research has demonstrated that BAT is an important site of facultative (nonshivering) thermogen-esis, which is acutely induced by thyroid hormone administra-tion, stimulation of sympathetic nervous system by exercise, cold, and food intake (40–42).
Recent data revealed that liver is a major regulator of metab-olite flow in the body, receiving inputs from the intestine through the portal vein, the general circulation through the hepatic artery, and the lymphatic system. Hepatocytes remove many materials from circulation and release them or their products at a moderated rate. This capacity for uptake, espe-cially lipids, may be of key importance to initiate steatosis (43). The increase in liver fat may be due to a high supply of FFAs in the absence of a concomitant increase in β-oxidation (30), that is, in response to increment in the energy expenditure. Thus, in our results it was demonstrated that high-fat diet in the sedentary group promoted more fat accumulation in the liver when compared with the sedentary rats fed with a chow diet; however, liver mass was maintained in all groups (Figure 2 and Table 2), which may be due to a decrease in the glycogen content in the tissue in response to a high-fat diet as previously demonstrated (30).
One of the most important results observed in this investi-gation is that CE and IE were effective in reducing fatty liver, suppressing the hepatic steatosis induced by high-fat diet (Figure 2). However, data from animal experiments are not linearly connected with human research. These new findings suggest the importance of applying both kinds of exercise in the associated therapy to control and reduce high prevalence (50%) of nonalcoholic fat liver diseases in the obese population including adolescents (4).
In conclusion, our investigation demonstrated that exercise is an important nonpharmacological strategy in disease control. Moreover, the major finding of this study was that IE was more efficient than CE in reducing the adverse effects of high-fat diet and sedentarism because IE induces an improvement in lipid profile and reduction in body weight gain, visceral and central adiposity, and fatty liver, contributing to the control of obesity and other comorbidities such as nonalcoholic fat liver diseases.
acknowledgMentsThis work was supported by FAPESP (Process 99/12981-7) and CAPES.
dIsclosureThe authors declared no conflict of interest.
© 2008 The Obesity Society
reFerences1. Bossan FM, Anjos LA, Vasconcelos MTL, Wahrlich V. Nutritional status of
the adult population in Niterói, Rio de Janeiro, Brazil: the nutrition, physical activity, and health survey. Cad Saúde Pública 2007;23:1867–1876.
2. Dizdar O, Alyamaç E. Obesity: an endocrine tumor? Med Hypotheses 2004;63:790–792.
3. Jéquier E. Pathways to obesity. Int J Obes Relat Metab Disord 2002;26(Suppl 2):S12–S17.
4. Tock L, Prado WL, Caranti DA et al. Nonalcoholic fatty liver disease decrease in obese adolescents after multidisciplinary therapy. Eur J Gastroenterol Hepatol 2006;18:1241–1245.
(H-average 5.12 kcal/g) when compared with the chow diet (C-average 4.07 kcal/g). In this sense, high-fat diets reduced food efficiency (15) and increased metabolic efficiency. This fact may explain why high-fat diet had no effects on body weight gain in sedentary (SH) and continuous exercise rats (CEH) (Table 1). The role of physical exercise on weight gain, energy intake, and food consumption in rats may depend on the exercise duration, age, gender, experimental period, and diet (14,15). Despite the similar caloric intake, the IE in both diets resulted in a reduction of body weight gain compared with the respective sedentary groups (Table 1); these effects may suggest that IE promotes an increase in the energy expen-diture more than that observed in CE.
In this study, high-fat diet in sedentary rats presented an increase in visceral and central adiposity, lipogenesis rate, and serum cholesterol concentration (Figure 1 and Tables 4 and 5); this finding agrees with that found in the literature (14,15,28–30,33–36). Continuous and intermittent exercise resulted in the reduction of visceral and central adiposity, serum TG, and increased cholesterol and HDL-cholesterol concentrations in rats fed with a chow diet (Figure 1 and Table 5). The hypotrig-lyceridemia presented in the CEC group was accompanied by an increase in the liver lipogenesis rate (Tables 4 and 5). These data are consistent with the notion that decreased TG is avail-able to liver-induced lipid synthesis by tissue (14), and that the exercise improves insulin action in the liver (37). Furthermore, CE with both diets and IE with high-fat diet increased the lipo-genesis rate at rest in all tissues (Table 4). This is because during exercise and after glycogen depletion, fatty acid becomes the major fuel for the exercising metabolism in muscle; in the recovery period, glycerol is necessary for glycogen reposition-ing (29,30).
The highlight of the results observed in this investigation is that IE in rats fed with high-fat diet led to reduction in body weight gain, central and visceral adiposity, and lipid accumula-tion in the liver associated to a reduction in the lipogenesis rate at rest in all tissues analyzed (Table 4) when compared with the sedentary groups (Table 1, Figures 1 and 2). These effects suggest that IE promoted an increase in the energy expendi-ture in comparison to CE and, as reported, those high-fat diets resulted in an increased lipid oxidation during exercise (29,38). Both an increase in lipolysis and a consequent higher plasma fatty acid response during exercise facilitate this change and the concurrent decreases in the glycogen stores in muscle and liver (29,39).
Indeed, intermittent exercise (IEH) promoted a reduction in serum cholesterol concentration; however, similar adaptation was observed in HDL concentrations in rats fed with high-fat diet (Table 5). This fact may partially be due to an increase in lipoprotein lipase activity, the enzyme responsible for releasing fatty acids from the lipoproteins, thus promoting an imbalance between high-fat diet and exercise effects (28,30).
Another important finding observed in this investiga-tion is that exercise promoted an increase in relative weight of BAT (Table 2). This fact may be explained by the fact that swimming training produces adaptive changes owing to cold

2222 VOLUME 16 NUMBER 10 | OCTOBER 2008 | www.obesityjournal.org
articlesintegrative physiology
24. Hida K, Wada J, Eguchi J et al. Visceral adipose tissue-derived serine protease inhibitor: a unique insulin-sensitizing adipocytokine in obesity. Proc Natl Acad Sci USA 2005;102:10610–10615.
25. Horowitz JF. Fatty acid mobilization from adipose tissue during exercise. Trends Endocrinol Metab 2003;14:386–392.
26. Ruby BC, Robergs RA. Gender differences in substrate utilization during exercise. Sport Med 1994;17:393–410.
27. Even PC, Rieth N, Roseau S, Laure-Achagiotis C. Substrate oxidation during exercise in the rat cannot fully account for training-induced changes in macronutrients selection. Metabolism 1998;47:777–782.
28. Schrauwen P, Westerterp KR. The role of high-fat diets and physical activity in the regulation of body weight. Br J Nutr 2000;84:417–427.
29. Bernardes D, Manzoni MS, Souza CP, Tenório NM, Dâmaso AR. Effects of a high-fat diet and swimming moderated training on post exercise metabolism in male adult rats. Rev Bras Educ Fís Esp 2004;18: 191–200.
30. Gauthier MS, Couturier K, Latour JG, Lavoie JM. Concurrent exercise prevents high-fat-diet-induced macrovesicular hepatic steatosis. J Appl Physiol 2003;94:2127–2134.
31. Achten J, Jeukendrup AE. Optimizing fat oxidation through exercise and diet. Nutrition 2004;20:716–727.
32. Duarte AC, Fonseca DF, Manzoni MS et al. High-fat diet and secretory capacity of insulin in rats. Rev Nutr 2006;19:341–348.
33. Gaíva MH, Couto RC, Oyama LM et al. Polyunsaturated fatty acid-rich diets: effect on adipose tissue metabolism in rats. Br J Nutr 2001;86: 371–377.
34. Diniz YS, Fernandes AA, Campos KE et al. Toxicity of hypercaloric diet and monosodium glutamate: oxidative stress and metabolic shifting in hepatic tissue. Food Chem Toxicol 2004;42:319–325.
35. Iossa S, Lionetti L, Mollica MP et al. Effect of high-fat feeding on metabolic efficiency and mitochondrial oxidative capacity in adult rats. Br J Nutr 2003;90:953–960.
36. Woods SC, Seeley RJ, Rushing PA, D’Alessio D, Tso P. A controlled high-fat diet induces an obese syndrome in rats. J Nutr 2003;133:1081–1087.
37. Mikines KJ, Sonne B, Farrell PA, Tronier B, Galbo H. Effect of physical exercise on sensitivity and responsiveness to insulin in humans. Am J Physiol 1988;254:E248–E259.
38. Helge JW, Watt PW, Richter EA, Rennie MJ, Kiens B. Fat utilization during exercise: adaptation to a fat-rich diet increases utilization of plasma fatty acids and very low density lipoprotein-triacylglycerol in humans. J Physiol 2001;537:1009–1020.
39. Helge JW. Long-term fat diet adaptation effects on performance, training capacity, and fat utilization. Med Sci Sports Exerc 2002;34: 1499–1504.
40. Margareto J, Marti A, Martínez A. Changes in UCP mRNA expression levels in brown adipose tissue and skeletal muscle after feeding a high-energy diet and relationships with leptin, glucose and PPARgamma. J Nutr Biochem 2001;12:130–137.
41. Sullo A, Brizzi G, Maffulli N. Triiodothyronine deiodinating activity in brown adipose tissue after short cold stimulation test in trained and untrained rats. Physiol Res 2004;53:69–75.
42. Westerterp KR. Diet-induced thermogenesis. Nutr Metab (Lond) 2004;1:1–5.
43. Bradbury MW. Lipid metabolism and liver inflammation. I. Hepatic fatty acid uptake: possible role in steatosis. Am J Physiol Gastrointest Liver Physiol 2006;290:G194–G198.
5. Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev 2000;21:697–738.
6. Rosa EC, Zanella MT, Ribeiro AB, Kohlmann O Jr. Visceral obesity, hypertension and cardio-renal risk: a review. Arq Bras Endocrinol Metabol 2005;49:196–204.
7. Westerterp KR. Perception passive overfeeding and energy metabolism. Physiol Behav 2006;89:62–65.
8. Saris WH, Blair SN, van Baak MA et al. How much physical activity is enough to prevent unhealthy weight gain? Outcome of the IASO 1st Stock Conference and consensus statement. Obes Rev 2003;4:101–114.
9. Jakicic JM, Clark K, Coleman E et al. American College of Sports Medicine position stand. Appropriate intervention strategies for weight loss and prevention of weight regain for adults. Med Sci Sports Exerc 2001;33: 2145–2156.
10. Pate RR, Pratt M, Blair SN et al. Physical activity and public health. A recommendation from the Centers for Disease Control and Prevention and the American College of Sports Medicine. JAMA 1995;273:402–407.
11. Matsudo VKR. Exercícios acumulados funcionam? Diagnóstico Tratamento 2005;10:163–165.
12. Brooks GA, Butte NF, Rand WM, Flatt JP, Caballero B. Chronicle of the Institute of Medicine physical activity recommendation: how a physical activity recommendation came to be among dietary recommendations. Am J Clin Nutr 2004;79:921S–930S.
13. Duarte FO, Sene MO, Oishi JC et al. O exercício de final de semana contribui para o controle das dislipidemias em ratos adultos machos alimentados com dieta rica em gorduras. Rev Bras Fisioter 2003;7: 229–235.
14. Estadella D, Oyama LM, Dâmaso AR, Ribeiro EB, Oller Do Nascimento CM. Effect of palatable hyperlipidic diet on lipid metabolism of sedentary and exercised rats. Nutrition 2004;20:218–224.
15. Burneiko RC, Diniz YS, Galhardi CM et al. Interaction of hypercaloric diet and physical exercise on lipid profile, oxidative stress and antioxidant defenses. Food Chem Toxicol 2006;44:1167–1172.
16. National Research Council. Guide for Care and Use of Laboratory Animals (Revised Ed). National Academy of Sciences: Washington, DC, 1985.
17. Voltarelli FA, Gobatto CA, de Mello MAR. Determination of anaerobic threshold in rats using the lactate minimum test. Braz J.Med Biol Res 2002;35:1389–1394.
18. Cinti S. The adipose organ. Prostaglandins Leukot Essent Fatty Acids 2005;73:9–15.
19. Robinson AM, Williamson DH. Control of glucose metabolism in isolated acini of the lactating mammary gland of the rat. Effects of oleate on glucose utilization and lipogenesis. Biochem J 1978;170:609–613.
20. Oller do Nascimento CM, Willianson DH. Evidence for conservation of dietary lipid in the rat during lactation and the immediate period after removal of the litter. Decreased oxidation of oral [1-14C] triolein. Biochem J 1986;239:233–236.
21. Manzoni MS, Rossi EA, Carlos IZ et al. Fermented soy product supplemented with isoflavones affected fat depots in juvenile rats. Nutrition 2005;21:1018–1025.
22. Lovoie JM, Yasari S, Abdennadher M, Paquette A. Effects of alternations (10 days) of high-fat with normal diet on liver lipid infiltration, fat gain, and plasma metabolic profile in rats. Physiol Behav 2005;86:442–448.
23. Poobalan A, Aucott L, Smith WC et al. Effects of weight loss in overweight/obese individuals and long-term lipid outcomes—a systematic review. Obes Rev 2004;5:43–50.
Related Documents











