ORIGINAL PAPER Efficiency of direct and indirect shoot organogenesis, molecular profiling, secondary metabolite production and antioxidant activity of micropropagated Ceropegia santapaui J. J. Chavan • N. B. Gaikwad • S. D. Umdale • P. R. Kshirsagar • K. V. Bhat • S. R. Yadav Received: 5 March 2013 / Accepted: 24 May 2013 / Published online: 31 May 2013 Ó Springer Science+Business Media Dordrecht 2013 Abstract Ceropegias has acquired significant importance due to their medicinal properties, edible tubers, and its ornamental flowers. The aim of this study was to optimize direct shoot organogenesis (DSO), indirect shoot organo- genesis (ISO) and plant regeneration of threatened medicinal plant Ceropegia santapaui, followed by analysis of genetic status and biochemical characterization of micropropagated plantlets. For optimization, cotyledonary nodes and cotyle- dons were used as source of explants in DSO and ISO respectively. The highest frequency of regeneration (88.0 %) for DSO with 8.1 ± 0.6 shoots per explant was obtained from cotyledonary nodes cultured on Murashige and Skoog’s (MS) medium containing 2.0 mg L -1 2iP. The best response for callus induction and proliferation was achieved with 1.5 mg L -1 PR (picloram) in which 97.5 % of cultures produced an average of 913 ± 10.9 mg (fresh weight) of callus. The highest frequency of shoot formation (92.5 %) with an average of 19.7 ± 0.3 shoots in ISO was obtained when calli were transferred to MS medium sup- plemented with 2.5 mg L -1 BAP and 0.4 mg L -1 IBA. Regenerated shoots were best rooted in half-strength MS medium with 2.0 mg L -1 NAA. Plantlets successfully acclimatized were morphologically indistinguishable from the source plant. Micropropagated plantlets subjected to random amplified polymorphic DNA and inter simple sequence repeats (ISSR) marker based profiling reveled uniform banding pattern in DSO-derived plantlets which was similar to mother plant. ISSR fingerprints of ISO- derived plants showed low variation. Method of regenera- tion, plant part and solvent system significantly affected the levels of total phenolics, flavonoids and antioxidant capac- ity. Assay of antioxidant activity of different tissues revealed that significantly higher antioxidant activity was observed in ISO-derived tissues than DSO-derived and mother tissues. RP-HPLC analysis of micropropagated plantlets showed the presence of three major phenolic compounds which were similar to those detected in mother plant. Rapid multiplica- tion rate, genetic stability and biochemical parameter ensures the efficacy of the protocol developed for the prop- agation of this threatened medicinal plant. Keywords Antioxidant Cerpegin Ceropegia santapaui Conservation Flavonoids ISSR RP-HPLC RAPD Organogenesis Phenolics Threatened species Abbreviations BAP 6-Benzylaminopurine Cerpegin 1,1-Dimethylfuro(3,4-C)pyridine-3,4 (1H, 5H)-dione DPPH 2,2-Diphenyl-1-picrylhydrazyl DSO Direct shoot organogenesis FW Fresh weight IAA Indole 3-acetic acid IBA Indole 3-butyric acid 2iP 6-c,c-Dimethylallylaminopurine ISO Indirect shoot organogenesis ISSR Inter simple sequence repeats MS Murashige and Skoog’s medium (1962) J. J. Chavan N. B. Gaikwad S. D. Umdale P. R. Kshirsagar S. R. Yadav Department of Botany, Shivaji University, Kolhapur 416 004, Maharashtra, India J. J. Chavan (&) Department of Botany, Yashavantrao Chavan Institute of Science, Satara 415 001, Maharashtra, India e-mail: [email protected] S. D. Umdale K. V. Bhat National Research Centre on DNA Fingerprinting, NBPGR, New Delhi 110 012, India 123 Plant Growth Regul (2014) 72:1–15 DOI 10.1007/s10725-013-9830-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Efficiency of direct and indirect shoot organogenesis, molecularprofiling, secondary metabolite production and antioxidantactivity of micropropagated Ceropegia santapaui
J. J. Chavan • N. B. Gaikwad • S. D. Umdale •
P. R. Kshirsagar • K. V. Bhat • S. R. Yadav
Received: 5 March 2013 / Accepted: 24 May 2013 / Published online: 31 May 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Ceropegias has acquired significant importance
due to their medicinal properties, edible tubers, and its
ornamental flowers. The aim of this study was to optimize
direct shoot organogenesis (DSO), indirect shoot organo-
genesis (ISO) and plant regeneration of threatened medicinal
plant Ceropegia santapaui, followed by analysis of genetic
status and biochemical characterization of micropropagated
plantlets. For optimization, cotyledonary nodes and cotyle-
dons were used as source of explants in DSO and
ISO respectively. The highest frequency of regeneration
(88.0 %) for DSO with 8.1 ± 0.6 shoots per explant was
obtained from cotyledonary nodes cultured on Murashige
and Skoog’s (MS) medium containing 2.0 mg L-1 2iP. The
best response for callus induction and proliferation was
achieved with 1.5 mg L-1 PR (picloram) in which 97.5 % of
cultures produced an average of 913 ± 10.9 mg (fresh
weight) of callus. The highest frequency of shoot formation
(92.5 %) with an average of 19.7 ± 0.3 shoots in ISO was
obtained when calli were transferred to MS medium sup-
plemented with 2.5 mg L-1 BAP and 0.4 mg L-1 IBA.
Regenerated shoots were best rooted in half-strength MS
medium with 2.0 mg L-1 NAA. Plantlets successfully
acclimatized were morphologically indistinguishable from
the source plant. Micropropagated plantlets subjected to
random amplified polymorphic DNA and inter simple
sequence repeats (ISSR) marker based profiling reveled
uniform banding pattern in DSO-derived plantlets which
was similar to mother plant. ISSR fingerprints of ISO-
derived plants showed low variation. Method of regenera-
tion, plant part and solvent system significantly affected the
levels of total phenolics, flavonoids and antioxidant capac-
ity. Assay of antioxidant activity of different tissues revealed
that significantly higher antioxidant activity was observed in
ISO-derived tissues than DSO-derived and mother tissues.
RP-HPLC analysis of micropropagated plantlets showed the
presence of three major phenolic compounds which were
similar to those detected in mother plant. Rapid multiplica-
tion rate, genetic stability and biochemical parameter
ensures the efficacy of the protocol developed for the prop-
agation of this threatened medicinal plant.
Keywords Antioxidant � Cerpegin � Ceropegia santapaui �Conservation � Flavonoids � ISSR � RP-HPLC � RAPD �Organogenesis � Phenolics � Threatened species
Abbreviations
BAP 6-Benzylaminopurine
Cerpegin 1,1-Dimethylfuro(3,4-C)pyridine-3,4
(1H, 5H)-dione
DPPH 2,2-Diphenyl-1-picrylhydrazyl
DSO Direct shoot organogenesis
FW Fresh weight
IAA Indole 3-acetic acid
IBA Indole 3-butyric acid
2iP 6-c,c-Dimethylallylaminopurine
ISO Indirect shoot organogenesis
ISSR Inter simple sequence repeats
MS Murashige and Skoog’s medium (1962)
J. J. Chavan � N. B. Gaikwad � S. D. Umdale �P. R. Kshirsagar � S. R. Yadav
Department of Botany, Shivaji University,
Kolhapur 416 004, Maharashtra, India
J. J. Chavan (&)
Department of Botany, Yashavantrao Chavan Institute
of Science, Satara 415 001, Maharashtra, India
e-mail: [email protected]
S. D. Umdale � K. V. Bhat
National Research Centre on DNA Fingerprinting, NBPGR,
New Delhi 110 012, India
123
Plant Growth Regul (2014) 72:1–15
DOI 10.1007/s10725-013-9830-7

NAA a-Naphthalene acetic acid
PGRs Plant growth regulators
PR Picloram (4-amino-3,5,6-trichloropicolinic acid)
RAPD Random amplified polymorphic DNA
RE Rutin equivalent
RP-HPLC Reverse phase-high performance liquid
chromatography
TAE Tannic acid equivalent
TPC Total phenolic content
TFC Total flavonoid content
Introduction
The genus Ceropegia L. (family–Asclepiadaceae) includes
herbs and climbers grows in tropical and sub tropical regions
of the World which comprises over 220 species (Bruyns
2003; Murthy et al. 2012). The maximum diversity of Ce-
ropegia occurs in South Africa followed by Kenya, Mada-
gascar, and India (Murthy et al. 2012). India alone harbors
over 50 species of which 35 species are endemic to country
(Murthy et al. 2012). Ceropegia santapaui Wadhwa and
Ansari is one of the twining herbs endemic to small pockets
in Western Ghats and considered as threatened because of
the small number of individuals (Yadav and Kamble 2008).
Ceropegias (including C. santapaui) are highly-prized for
their medicinal properties, edible tubers, and its ornamental
flowers. Owing to their ornamental potential, some species
are cultivated as horticultural crop (McNew 2002; Hodgkiss
2004; Reynolds 2006). The sweet–sour leaves are edible and
are considered to be tonic and digestive. The tubers of Ce-
ropegias are edible and contain starch, sugars, gum, alb-
uminoids, carbohydrates, fats, and crude fiber (Mabberly
1987; Jain and Defillips 1991). The ‘Cerpegin’ (1,1-dim-
ethylfuro[3,4-C]pyridine-3,4(1H,5H)-dione) present in the
root tubers (including C. santapaui) was isolated and iden-
tified as pyridine type alkaloid (Nadkarni 1976; Adibatti
et al. 1991; Phulwaria et al. 2013). The Cerpegin exhibited
promising hepatoprotective, antipyretic, anti-ulcer, analge-
sic, mast-cell stabilizing, tranquillsing and hypotensive
activities (Adibatti et al. 1991; Sukumar et al. 1995). The
tubers of many Ceropegia species are used by tribal women
to promote fertility and vitality, treatment of diarrhea and
dysentery, and treatment of urinary bladder stones (Khare
2007; Swarnkar and Katewa 2008).
The genus Ceropegia as a whole is under threat, owing
to either destructive collection or habitat degradation. Due
to pharmacological importance of ‘cerpegin’ C. santapaui
is over-exploited and current status is threatened species.
Since it is a seasonal plant, generally vegetative growth is
seen only after first rain by sprouting of dormant under-
ground tubers. Moreover, the species is difficult to propa-
gate by conventional methods of cuttings and seedlings
because of poor seed setting and limited seed viability
(Yadav and Kamble 2008). There is therefore an urgent
need for conservation action to ensure large scale propa-
gation. Plant tissue culture has remained valuable in terms
of their beneficial role in the production and conservation
of plant based resources (Vasil 2008; Amoo et al. 2012).
This technique offers a way to multiply plants possessing a
special phenotypic character directly and rapidly, thereby
shortening the time needed, and providing an effective
technique for repopulation of the original locations of the
plants (Socorro et al. 1998). In response to degradation of
the habitat of these species, micropropagation systems have
been developed with several threatened species of Ce-
ropegia (Patil 1998; Beena et al. 2003; Nikam and Savant
2009; Chandore et al. 2010; Murthy et al. 2010; Chavan
et al. 2011a, b; Phulwaria et al. 2013).
The cryptic genetic defects arising via in vitro propaga-
tion in the regenerants seriously limits the utility of micro-
propagation system. However, it is important to assess the
genetic constitution and stability of in vitro micropropagated
plants before planning strategies for further propagation. The
progress made in DNA marker technology provided valuable
tools for the detection of somaclonal variation that was
reflected in the profiles of molecular genetic markers viz.
RFLPs, RAPDs and ISSRs (Devarumath et al. 2002; Bhatia
et al. 2011). However, none of the developed Ceropegia
micropropagation protocols has assessed the genetic fidelity
of micropropagated plants.
The present study was designed to optimize the direct
and indirect shoot organogenesis and plant regeneration in
C. santapaui. RAPD and ISSR markers were employed for
determining genetic stability of in vitro regenerated plants
and the reliability protocol for true to type cloning. Bio-
chemical parameters viz. total phenolic content, total fla-
vonoid content and antioxidant activity were investigated.
RP-HPLC analysis was also carried out for identification
and quantitation of phenolic compounds.
Materials and methods
In vitro seed germination and explant preparation
Mature plants and follicles of C. santapaui were collected
from the of Kumbharli locality of Northern Western Ghats,
India. Seeds separated from follicles, then washed and dried
for 3 days in the shade. Seeds were surface sterilized with
0.1 % HgCl2 for 5 min under aseptic conditions and washed
2–3 times with sterile distilled water. For germination, the
seeds were inoculated on MS medium without plant growth
regulators. Cultures were maintained in a growth room at
25 ± 1 �C in 16 h light and 8 h dark cycles with
50 lmol m-2 s-1 of light intensity provided by cool-white
2 Plant Growth Regul (2014) 72:1–15
123

fluorescent tubes. Cotyledonary nodes and cotyledons were
excised aseptically from 15 day old seedling were used as
explants for DSO and ISO respectively.
Optimization of direct shoot organogenesis (DSO)
Direct shoot organogenesis was optimized using cotyle-
donary nodes (about 1 cm) as source of explants with dif-
ferent plant growth regulators. The shoots were regenerated
on MS medium supplemented with 30 g L-1 sucrose,
0.1 g L-1 myo-inositol, different plant growth regulators
(BAP, KN, TDZ, 2iP, IBA) and solidified with 2 g L-1
ClariGel (Himedia, India). A control without any PGRs was
included. The pH of the medium was adjusted to 5.8 using
NaOH or HCl before autoclaving at 121 �C and 103 kPa for
20 min. All the cultures were maintained in a growth room at
25 ± 1 �C in a 16 h photoperiod and 8 h dark cycles with
50 lmol m-2 s-1 of light intensity. In order to assess the
influence of different PGRs on shoot proliferation from
cotyledonary nodes, following growth parameters were
measured: (1) the regeneration frequency (%), (2) average
number of shoots, and (3) average shoot length (cm).
Optimization of indirect shoot organogenesis (ISO)
Newly emerged cotyledons (more than 0.5 cm) were
aseptically excised from in vitro germinated seedlings were
cultured, abaxial side up, on MS media supplemented with
various concentrations of 2,4-D (0.5, 1.0, 1.5, 2.0,
2.5 mg L-1), PR (0.5, 1.0, 1.5, 2.0, 2.5 mg L-1), and NAA
(0.5, 1.0, 1.5, 2.0, 2.5 mg L-1). Frequency (%) of explants
that initiated callus was recorded after 4 weeks. Calli of
each treatment were sub-cultured in the same medium for
further proliferation every 4 weeks. Callus growth was
calculated by measuring its fresh weight of callus on each
auxin treatments. To optimize the effect of PGRs on shoot
regeneration, calli (approx. 200 mg) developed on opti-
mum callus induction medium were transferred to regen-
eration medium either BAP alone (0.5, 1.0, 1.5, 2.0, 2.5,
3.0 mg L-1) or in combination with IBA (0.2, 0.4, 0.6, 0.8,
1.0 mg L-1) or NAA (0.2, 0.4, 0.6, 0.8, 1.0 mg L-1). All
the cultures were incubated at 25 ± 1 �C under a 16 h light
and 8 h dark cycles with 50 lmol m-2 s-1 of light inten-
sity provided by cool fluorescent tubes (Philips, India).
Rooting and acclimatization
Regenerated shoots (both DSO and ISO-derived) from
optimal regeneration medium were transferred to half-
strength MS medium containing different auxins for in vitro
rooting. Auxins viz. IBA (0.5, 1.0, 1.5, 2.0, 2.5, 3.0 mg L-1),
NAA (0.5, 1.0, 1.5, 2.0, 2.5, 3.0 mg L-1), and IAA (0.5, 1.0,
1.5, 2.0, 2.5, 3.0 mg L-1) were tested either alone or their
combinations. To evaluate the influence of auxins on in vitro
rooting, following growth parameters were measured: (1) the
percentage of in vitro rooting (%), (2) average number of
roots, and (3) average root length (cm). After 4 weeks in
rooting medium, regenerated plants with well developed
roots were removed from culture tubes, washed free of
solidifying agent and transferred to plastic pots containing
sterile soil, sand and coco peat (1:2:1). Plants were accli-
matized for 2 weeks at 25 ± 1 �C in the growth room with
16-h photoperiod (50 lmol m-2 s-1 light intensity),
watered with half-strength MS liquid medium without
sucrose and then transferred to glasshouse conditions (temp:
30 �C, humidity: 70 %, 70 lmol m-2 s-1 light intensity).
Survival rate (%) was estimated after 60 days of transfer to
field conditions.
All the tissue culture experiments were set in a com-
pletely randomized block design and each experiment
repeated thrice with 20 tubes per treatment. Comparison
between the mean values of treatments with control were
made using Dunnett multiple comparison test (DMCT) at
ns, 0.05 and 0.01 level of significance.
RAPD and ISSR analysis
DNA was isolated from fresh young leaves of mother plant as
well as randomly selected in vitro raised plants (10 each from
DSO and ISO) were used for RAPD and ISSR analysis. The
genomic DNA was isolated following the protocol of Doyle
and Doyle (1990) with a little modification. Quality and
quantity of genomic DNA was assessed by 0.8 % agarose gel
electrophoresis with k uncut DNA. The final concentration
was made to 20 ng lL-1 and stored at -20 �C until further
use. A total of 45 random decamer primers (Genemed Syn-
thesis Inc, Texas, USA) were screened for RAPD analysis,
out of which 10 primers were selected on the basis of clarity
of banding patterns. The protocol for RAPD analysis was
adapted from that of Williams et al. (1990) with some
modification. DNA amplification was performed with reac-
tion volume of 25 lL containing 10 9 PCR buffer (2.5 lL),
2.5 mM MgCl2 (1.0 lL), 100 mM dNTPs (2.0 lL), primer
(2.0 lL), Taq polymerase (0.2 lL), 40 ng of template DNA
(2.0 lL), and 14. 3 lL sterile D.W. PCR was performed at
initial temperature of 94 �C (6 min, 1 cycle), followed by 40
cycles of 1 min at 94 �C, 1 min at 35 �C and 10 min at
72 �C, and a final cycle of 10 min at 72 �C.
ISSR analysis was performed following the procedure
described by Zietkiewicz et al. (1994). A factorial experiment
with varying concentrations of genomic DNA (30, 40 and
50 ng), MgCl2 (1.5, 2.0 and 2.5 mM) and Taq DNA poly-
merase (0.5, 1 and 1.5 U) was performed to optimize PCR
conditions. After preliminary screening with 32 ISSR primers
(UBC, Canada) for standardization of optimum annealing
temperature, 10 ISSR primers were selected on the basis of
Plant Growth Regul (2014) 72:1–15 3
123

sharpness and reproducibility. The amplification was per-
formed for 40 cycles, programmed as follows; initial dena-
turation at 94 �C for 6 min (one cycle), denaturating at 94 �C
for 1 min, annealing at 48, 50 and 52 �C (depending on primer
used) for 1 min and extension at 72 �C for 2 min (38 cycles)
and final extension at 72 �C for 12 min (one cycle). PCR
amplification was performed in Thermal Cycler (BIOER, XP
Cycler, China). Amplification products obtained from RAPD
and ISSR analysis were analyzed in 1.8 % agarose gel stained
with ethidium bromide (0.5 lg lL-1). The size of the
amplicons was estimated using a 100-bp ladder (GeneRular
100 bp plus ladder) and documented in the Gel documentation
system (BioRad, Hercules, CA).
Evaluation of secondary metabolites
Plant material and extract preparation
Fresh plant parts (tuber, stem, leaf) of mother as well as
in vitro raised plants (both DSO and ISO) were washed under
running tap water and blotted with tissue towel. one gram of
fresh tissue was homogenized in 25 mL of respective solvent
(methanol, ethanol, acetone and water) and extractions were
carried on orbital shaker (REMI, India) with constant stirring
at 150 rpm for 24 h. The mixtures were centrifuged at
10,000 rpm for 10 min and the supernatant was filtered
through Whatman filter paper (No. 1). Freshly prepared
extracts were used in all the experiments. Measurements of
biochemical parameters were taken on UV-190 double beam
spectrophotometer (Shimadzu, Japan).
Determination of total phenolic content (TPC)
The determination of total phenolic content was performed
by using Folin–Ciocalteu method (Singleton and Rossi
1965) with little modification. Briefly, 0.125 mL of extract
was mixed with 1.8 mL of Folin-Ciocalteu reagent (ten
fold diluted) and kept for 6 min at 25 �C. Then 1.2 mL of
15 % Na2CO3 was added to the reaction mixture and kept
for 90 min at room temperature. The absorbance of the
reaction was measured at 765 nm. The concentration of the
total phenolics was determined as milligrams of tannic acid
equivalents (TAE) per gram fresh weight by using an
equation obtained from tannic acid calibration curve.
Determination of total flavonoid content (TFC)
Total flavonoid content was determined by using the alu-
minium chloride colorimetric method (Chang et al. 2002).
Briefly, 0.5 mL of extracts, 1.5 mL of methanol, 0.1 mL of
aluminium chloride (10 %), 0.1 mL of potassium acetate
(1 M) and 2.8 mL of distilled water were mixed for 5 min
by vortexing. Reaction mixture was kept at room
temperature for 30 min and the absorbance was measured
at 415 nm. The calibration curve was prepared for rutin and
the results are expressed as milligrams of rutin equivalents
(RE) per gram fresh weight.
RP-HPLC analysis of phenolic compounds
Extract prepared for the HPLC analysis by using the proce-
dure described by Ghatge (2007) with slight modification.
Briefly, 1 g of fresh leaf tissue was crushed in 20 mL ethanol
and extract was centrifuged at 15,000 rpm at 4 �C for
10 min. The supernatant was filtered through a 0.22 lm filter
(Millipore, Westboro, MA, USA) before injection into
HPLC system. HPLC analyses were performed using a
Waters Chromatic system (Model no. HPLC-W6590)
equipped with a Waters 717 plus auto sampler and a Waters
2487 photodiode array detector. Separation was achieved on
a reverse phase C-18 column (Princeton SPHER, 5 lm,
3.9 mm 9 150 mm). The mobile phase was acetonitrile:
water (84:16, v/v) at a flow rate of 1.0 mL min-1, the
injection volume was set to 20 lL and the column was
retained at room temperature. Individual phenolic com-
pounds were identified at 280 nm by the retention time of
sample chromatographic peaks being compared with those
of authentic standards using the same operating conditions.
Antioxidant activity of micropropagated plants
DPPH free radical scavenging assay
The DPPH (2,2-diphenyl-1-picrylhydrazyl) free radical
scavenging activity of the extracts was determined essen-
tially as described by Brand-Williams et al. (1995) and
modified by Jagtap et al. (2011). The stock reagent solution
was prepared by dissolving 24 mg of DPPH in 100 mL
methanol and stored at -20 �C until use. 100 lL of extract
was allowed to react with DPPH solution in the final
reaction volume of 3 mL. The mixture was shaken vigor-
ously and allowed to stand in the dark at room temperature.
The decrease in absorbance of the resulting solution was
then measured spectrophotometrically at 517 nm. The
control was prepared as above without any extract and
MeOH was used for the baseline correction. Radical
scavenging activity was expressed as the inhibition per-
centage and was calculated using the following formula,
% radical scavenging activity
¼ Control OD� sample OD=Control ODð Þ � 100:
FRAP (ferric reducing antioxidant power) assay
The FRAP assay was carried out according to procedure
described by Benzie and Strain (1996) and modified by
4 Plant Growth Regul (2014) 72:1–15
123

Chavan et al. (2012). Briefly the working FRAP reagent
prepared by combination of 0.3 M acetate buffer (pH 3.6),
10 mM 2, 4, 6-tripyridyl-s-triazine (TPTZ) in 40 mM HCL
and 20 mM FeCl3�6H2O in 10:1:1 ratio prior to use and
heated to 37 �C in water bath for 10 min. 100 lL of extract
was allowed to react with 2.7 mL of the FRAP reagent.
The final volume of the reaction mixture was made up to
3 mL with sterile distilled water. The reaction mixture was
kept in dark for 30 min. The absorbance of the colored
product (ferrous tripyridyltriazine complex) was then
recorded at 593 nm. A higher absorbance reading indicated
a higher reducing power.
Metal chelating assay
The chelating effect of ferrous ions from different parts of
C. santapaui was estimated by the method of Dinis et al.
(1994). Briefly, 500 lL of plant extracts were added into
100 lL of 2 mM FeCl2. The reaction was initiated by
adding 40 lL of 5 mM ferrozine solution into the mixture,
which was then shaken vigorously and left standing at
temperature for 10 min. The absorbance of the reaction
mixture was measured at 562 nm. Three replicates were
made for each test sample. The ratio of inhibition of fer-
rozine–Fe2? complex formation was calculated as follows:
% inhibition ¼ Acontrol � Asample
� ��Acontrol � 100
where A = absorbance at 54 nm.
Results and discussion
Direct shoot organogenesis
Cotyledonary nodes were excised from in vitro grown
seedlings (Fig. 1a) were cultured on shoot multiplication
medium. Axillary buds started emerging after 7–10 days of
inoculation. MS medium lacking PGRs did not support the
induction of shoots. The number of shoots per cotyledonary
node explant was significantly affected by the type and
concentration of various PGRs (Table 1). Of the different
concentrations of KN, 1.0 mg L-1 elicited multiple shoots
in 82.5 % of cultures, with an average of 3.4 ± 0.5 shoots
per responsive explant and an average shoot length of
4.8 ± 0.6 cm (Table 1). TDZ induced a frequent formation
of short and fasciated shoots which were unsuitable for
in vitro rooting. BAP and TDZ were less effective in
inducing shoots when compared with KN and 2iP. None of
the concentrations of BAP and TDZ was effective in
enhancing shoot proliferation. The best frequencies of
shoot multiplication (88 %) and maximum numbers of
shoots (8.1 ± 0.6) were achieved on MS media containing
2.0 mg L-1 2iP (Table 1; Fig. 1b). Effectiveness of 2iP
has been previously reported in shoot multiplication of
Decalepis hamiltonii (Giridhar et al. 2005), and Caralluma
bhupenderiana (Ugraiah et al. 2011). In contrast, majority
of literature have reported BAP as the most active cyto-
kinin for shoot multiplication in various Asclepiadaceae
members (Patil 1998; Komalavalli and Rao 2000; Beena
et al. 2003; Amoo et al. 2009; Chavan et al. 2011a, b).
Topolins, naturally occurring aromatic cytokinins were
significantly altered the shoot multiplication in Huernia
hystrix, another member of Asclepiadaceae (Amoo and
Van Staden 2012) and Aloe arborescens (Amoo et al.
2012).
The combined treatments of cytokinins and auxins also
support shoot multiplication rates than the individual
treatments of the BAP, KN and TDZ. Addition of IBA
along with BAP or 2iP significantly boosted the multipli-
cation rate. MS medium supplemented with 3.0 mg L-1
BAP with 0.5 mg L-1 IBA produced an average of
4.0 ± 0.6 shoots per explant (Table 1). A combination of
2iP (2.0 mg L-1) and IBA (0.5 mg L-1) also produced an
average of 4.5 ± 0.6 shoots in 82 % cultures. The syner-
gistic effects of 2iP and auxins were reported in Gossypium
hirsutum cv. SVPR2, where a combination of these plant
Fig. 1 Regeneration in C. santapaui, a seed germination (MS basal medium), b direct organogenesis (MS ? 2iP 2.0 mg L-1), c formation of
embryogenic callus (MS ? PR 1.5 mg L-1) d indirect organogenesis (MS ? BAP 2.5 ? IBA 0.4 mg L-1)
Plant Growth Regul (2014) 72:1–15 5
123
growth regulators induced axillary bud sprouting at a
higher frequency (Ganesan and Jayabalan 2006). Baskaran
and Jayabalan (2005) also reported that when cotyledonary
nodes of Eclipta alba were cultured on MS medium con-
taining a combination of 2iP and auxins, the number of
shoots was increased per explant.
Indirect shoot organogenesis
Cotyledons from in vitro grown seedlings were cultured on
MS medium supplemented with 2,4-D, PR or NAA for
callus induction. Within 4 weeks of inoculation, callus was
proliferated from cut edges of the explants. Among the
auxins tested, PR was found more prominent than 2,4-D
and NAA for callus induction. About 82.5 % cultures
showed callusing at lower concentration of PR
(0.5 mg L-1). MS medium containing 1.5 mg L-1 PR was
best for callus induction (97.5 %) and growth (913
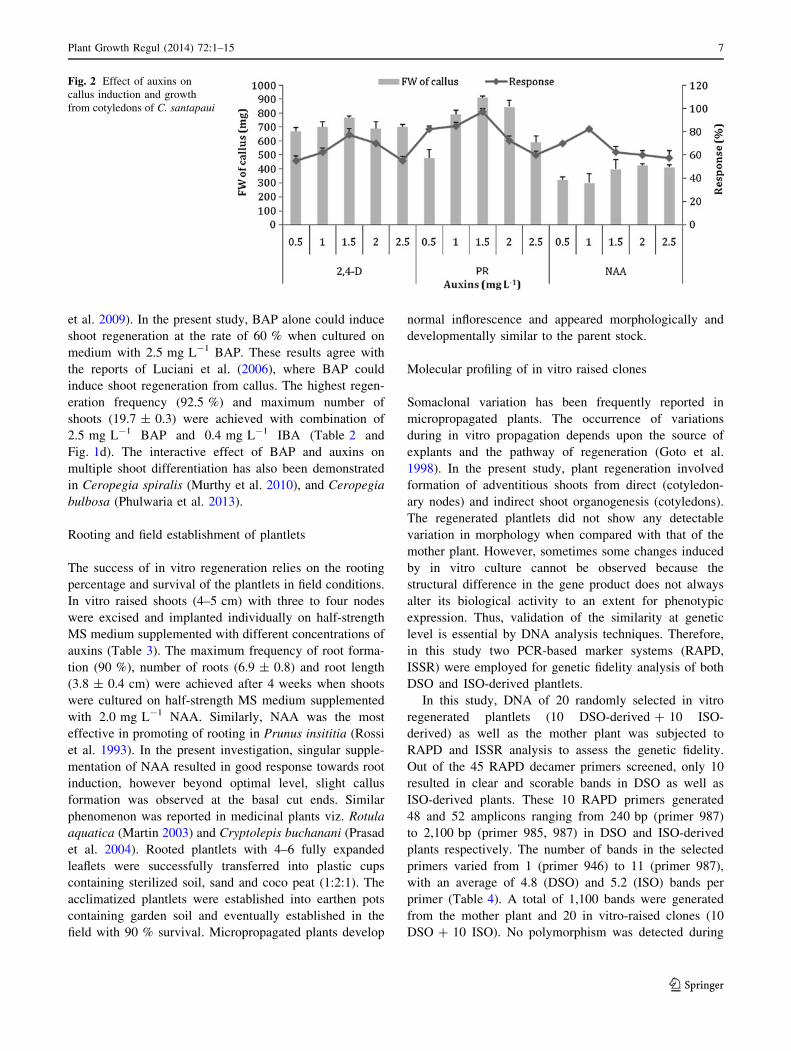
± 10.9 mg FW) of the callus (Fig. 2). Callus induced on
the same media composition was friable, granular and
comparatively fast growing than induced on other con-
centrations of PR, 2,4-D or NAA (Fig. 1c). Fitch and
Moore (1990) successfully used the picloram for callus
induction and long term maintenance of totipotent callus of
sugarcane.
The calli developed on PR (1.5 mg L-1) supplemented
medium were subcultured on MS medium supplemented
with various concentrations of BAP either alone or in
combination with IBA or NAA for shoot regeneration
(Table 2). After 2 weeks of culture, most of the calli
started to turn to light green, and they gradually became
dark green in the following week of culture. New white
calli were also developed from the callus mass. Similar
observation was also made in the Allium chinense (Yan
Table 1 Effect of different
plant growth regulators on shoot
regeneration from cotyledonary
nodes of C. santapaui
Values represents mean ± SE
of 20 replicates/treatment and
experiments were repeated
twice. Values are significantly
different at ns-non significant,
* P \ 0.05 and ** P \ 0.01
level according to Dunnett
multiple comparisons test
PGRs Concentration
(mg L-1)
Regeneration
frequency (%)
Number of shoots/explant
(mean ± SE)
Length of shoot (cm)
(mean ± SE)
Control 0.0 0.0 0.0 0.0
BAP 1.0 52.5 0.5 ± 0.1ns 5.0 ± 1.1**
2.0 75.0 1.3 ± 0.2* 6.4 ± 0.9**
3.0 77.5 2.1 ± 0.4** 7.0 ± 0.8**
4.0 67.5 1.1 ± 0.2** 6.3 ± 0.9**
5.0 62.5 1.0 ± 0.2** 6.1 ± 1.0**
KN 0.5 60.0 1.1 ± 0.2ns 4.3 ± 0.9**
1.0 82.5 3.4 ± 0.5** 4.8 ± 0.6**
1.5 72.5 1.9 ± 0.4** 5.4 ± 0.9**
2.0 70.0 1.7 ± 0.4** 4.3 ± 0.9**
2.5 62.5 1.4 ± 0.3* 4.5 ± 0.9**
TDZ 0.5 57.5 1.1 ± 0.3ns 3.4 ± 0.8**
1.0 72.5 1.8 ± 0.4** 4.1 ± 0.7**
1.5 82.5 2.6 ± 0.4** 3.5 ± 0.5**
2.0 75.0 2.1 ± 0.6** 3.2 ± 0.7**
2.5 57.5 1.3 ± 0.4ns 2.3 ± 0.5*
2iP 0.5 67.5 1.3 ± 0.2ns 6.2 ± 0.1**
1.0 75.0 2.4 ± 0.5** 6.3 ± 0.9**
1.5 85.0 3.6 ± 0.5** 6.1 ± 0.6**
2.0 88.0 8.1 ± 0.6** 6.5 ± 0.4**
2.5 72.5 2.9 ± 0.6** 6.2 ± 0.6**
BAP ? IBA 3.0 ? 0.1 65.0 2.1 ± 0.4** 6.2 ± 0.9**
3.0 ? 0.3 77.5 2.9 ± 0.5** 6.2 ± 0.8**
3.0 ? 0.5 85.0 4.0 ± 0.6** 6.5 ± 0.6**
3.0 ? 0.7 75.0 2.7 ± 0.6** 4.5 ± 0.6**
3.0 ? 1.0 62.5 2.3 ± 0.4** 4.2 ± 0.7**
2iP ? IBA 2.0 ? 0.1 75.0 1.9 ± 0.4** 6.1 ± 0.9**
2.0 ? 0.3 82.5 2.7 ± 0.4** 5.2 ± 0.7**
2.0 ? 0.5 82.0 4.5 ± 0.6** 5.3 ± 0.6**
2.0 ? 0.7 65.0 3.0 ± 0.5** 4.8 ± 0.7**
2.0 ? 1.0 60.0 1.4 ± 0.3ns 4.6 ± 0.9**
6 Plant Growth Regul (2014) 72:1–15
123
et al. 2009). In the present study, BAP alone could induce
shoot regeneration at the rate of 60 % when cultured on
medium with 2.5 mg L-1 BAP. These results agree with
the reports of Luciani et al. (2006), where BAP could
induce shoot regeneration from callus. The highest regen-
eration frequency (92.5 %) and maximum number of
shoots (19.7 ± 0.3) were achieved with combination of
2.5 mg L-1 BAP and 0.4 mg L-1 IBA (Table 2 and
Fig. 1d). The interactive effect of BAP and auxins on
multiple shoot differentiation has also been demonstrated
in Ceropegia spiralis (Murthy et al. 2010), and Ceropegia
bulbosa (Phulwaria et al. 2013).
Rooting and field establishment of plantlets
The success of in vitro regeneration relies on the rooting
percentage and survival of the plantlets in field conditions.
In vitro raised shoots (4–5 cm) with three to four nodes
were excised and implanted individually on half-strength
MS medium supplemented with different concentrations of
auxins (Table 3). The maximum frequency of root forma-
tion (90 %), number of roots (6.9 ± 0.8) and root length
(3.8 ± 0.4 cm) were achieved after 4 weeks when shoots
were cultured on half-strength MS medium supplemented
with 2.0 mg L-1 NAA. Similarly, NAA was the most
effective in promoting of rooting in Prunus insititia (Rossi
et al. 1993). In the present investigation, singular supple-
mentation of NAA resulted in good response towards root
induction, however beyond optimal level, slight callus
formation was observed at the basal cut ends. Similar
phenomenon was reported in medicinal plants viz. Rotula
aquatica (Martin 2003) and Cryptolepis buchanani (Prasad
et al. 2004). Rooted plantlets with 4–6 fully expanded
leaflets were successfully transferred into plastic cups
containing sterilized soil, sand and coco peat (1:2:1). The
acclimatized plantlets were established into earthen pots
containing garden soil and eventually established in the
field with 90 % survival. Micropropagated plants develop
normal inflorescence and appeared morphologically and
developmentally similar to the parent stock.
Molecular profiling of in vitro raised clones
Somaclonal variation has been frequently reported in
micropropagated plants. The occurrence of variations
during in vitro propagation depends upon the source of
explants and the pathway of regeneration (Goto et al.
1998). In the present study, plant regeneration involved
formation of adventitious shoots from direct (cotyledon-
ary nodes) and indirect shoot organogenesis (cotyledons).
The regenerated plantlets did not show any detectable
variation in morphology when compared with that of the
mother plant. However, sometimes some changes induced
by in vitro culture cannot be observed because the
structural difference in the gene product does not always
alter its biological activity to an extent for phenotypic
expression. Thus, validation of the similarity at genetic
level is essential by DNA analysis techniques. Therefore,
in this study two PCR-based marker systems (RAPD,
ISSR) were employed for genetic fidelity analysis of both
DSO and ISO-derived plantlets.
In this study, DNA of 20 randomly selected in vitro
regenerated plantlets (10 DSO-derived ? 10 ISO-
derived) as well as the mother plant was subjected to
RAPD and ISSR analysis to assess the genetic fidelity.
Out of the 45 RAPD decamer primers screened, only 10
resulted in clear and scorable bands in DSO as well as
ISO-derived plants. These 10 RAPD primers generated
48 and 52 amplicons ranging from 240 bp (primer 987)
to 2,100 bp (primer 985, 987) in DSO and ISO-derived
plants respectively. The number of bands in the selected
primers varied from 1 (primer 946) to 11 (primer 987),
with an average of 4.8 (DSO) and 5.2 (ISO) bands per
primer (Table 4). A total of 1,100 bands were generated
from the mother plant and 20 in vitro-raised clones (10
DSO ? 10 ISO). No polymorphism was detected during
Fig. 2 Effect of auxins on
callus induction and growth
from cotyledons of C. santapaui
Plant Growth Regul (2014) 72:1–15 7
123
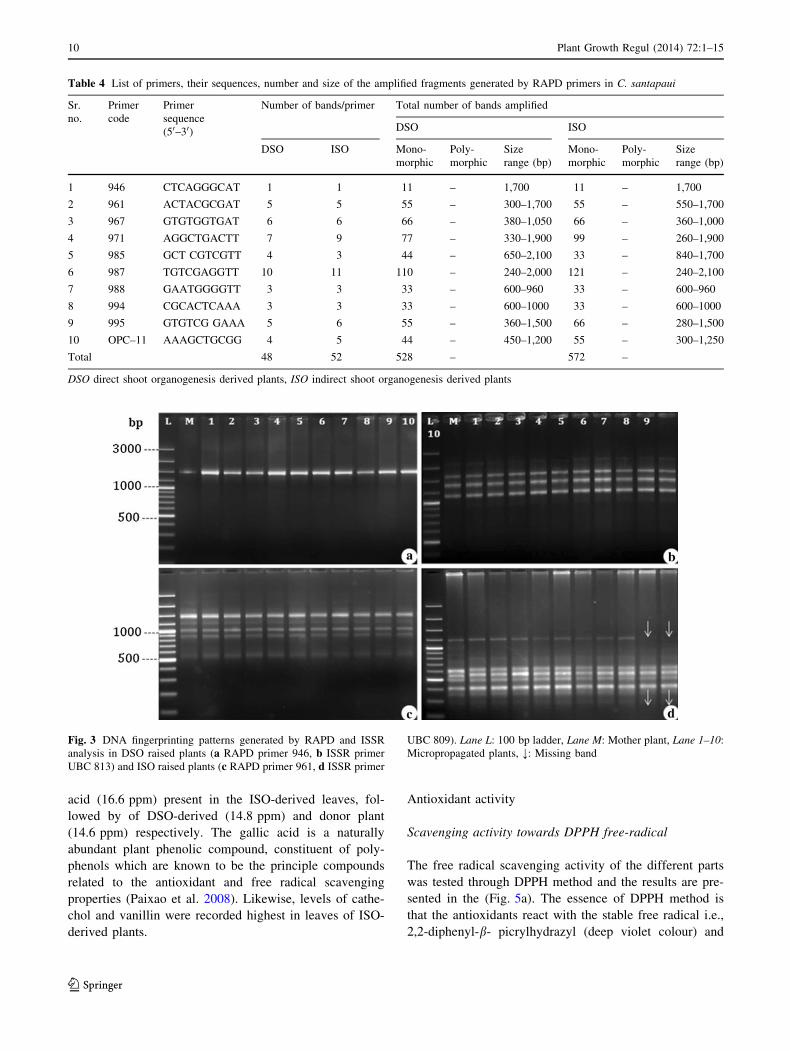
the RAPD analysis of in vitro-raised clones (Fig. 3a, c).
The absence of genetic variation using RAPD has been
reported in micropropagated shoots of Pinus thunbergii
(Goto et al. 1998) and Gerbera jamesonii (Bhatia et al.
2011).
Ten ISSR primers generated 39 (DSO) and 44 (ISO)
amplicons ranging from 300 bp (UBC 809) to 2,300 bp
(UBC 813) in size. The number of bands for each primer
varied from 2 (UBC 814, UBC 823) to 7 (UBC 809), with
an average of 3.9 bands per primer (Table 5). A total of
924 distinct amplicons were generated from 10 ISSR
primers in mother plant and 20 in vitro-raised clones (10
DSO ? 10 ISO-derived plants). Out of 924 bands, 913
bands were monomorphic within in vitro raised clones and
the mother plant, whereas, 11 polymorphic bands were
detected with 3 ISSR primers (UBC 801, UBC 809 and
UBC 815) in ISO-derived clones. It might be due to the
application of PGRs during callus induction, shoot regen-
eration and prolonged culture passage. Similarly, ISSR
primers were capable to detect polymorphism in Camellia
chinensis (Devarumath et al. 2002) and Gerbera jamesonii
(Bhatia et al. 2009).
In this study, RAPD fails to detect polymorphic loci in
both DSO and ISO-derived plants, however polymorphism
was observed in the ISSR fingerprints of ISO-derived
plants. Devarumath et al. (2002) also revealed that ISSR
fingerprint is more capable to detect the polymorphic loci
compared to RAPD fingerprinting. The results of our study
also support the fact that axillary multiplication is the
safest mode of micropropagation to produce true to type
progeny. There are some reports in literature, which cite
similar results (Martins et al. 2004; Joshi and Dhawan
2007; Bhatia et al. 2011).
Secondary metabolite production
Total phenolics and flavonoids
The chemical composition of different parts of C. santa-
paui is essential in order to establish a potential relation-
ship and understanding its role with different beneficial
biological activities enhancing human health benefits. The
phenol and flavonoid compounds present in plants are
known for their positive effects on inflammation (Xanth-
opoulou et al. 2010), cardiovascular diseases (Alothman
et al. 2009), besides having antioxidant activities (Jagtap
et al. 2011). Table 6 shows the total phenolic and flavonoid
contents of different parts viz. tuber, stem and leaf of
mother plant, DSO-derived plant and ISO-derived plants.
The results indicated that donor mother plant and DSO-
derived plants have lower levels of phenols and flavonoids
compared to ISO-derived plants. Methanolic ISO-derived
leaf extract exhibited the highest phenolic content
(6.8 ± 1.1 mg TAE/g FW) whereas, aqueous tuber extract
of donor plant showed lowest phenolic content
(1.1 ± 0.4 mg TAE/g FW). Similarly, the methanol had its
effect on the extraction of TPC from fruit pulp of Salacia
chinensis (Chavan et al. 2012).
The higher extractability of flavonoid (3.1 ± 0.4 mg
RE/g FW) was recorded in the methanolic leaf extract
of ISO-derived plants while, lowest flavonoid contents
were recorded in the aqueous tuber extract of donor and
Table 2 Effect of BAP either
alone or in combination with
auxins on shoot regeneration
from cotyledon-derived callus
of C. santapaui
Values represents mean ± SE
of 20 replicates/treatment and
experiments were repeated
twice. Values are significantly
different at ns- non significant,
* P \ 0.05 and ** P \ 0.01
level according to Dunnett
multiple comparisons test
PGRs Concentration
(mg L-1)
Regeneration
frequency (%)
Number of shoots
(mean ± SE)
Length of shoot
(cm) (mean ± SE)
Control 0.0 0.0 0.0 0.0
BAP 0.5 22.5 0.7 ± 0.2ns 2.1 ± 1.1**
1.0 30.0 2.0 ± 0.5* 2.8 ± 0.8**
1.5 45.0 2.7 ± 0.3** 3.1 ± 1.0**
2.0 52.5 4.0 ± 1.0** 3.8 ± 1.4**
2.5 60.0 7.8 ± 0.5** 4.0 ± 1.0**
3.0 47.5 5.1 ± 0.9** 3.5 ± 0.6**
BAP ? IBA 2.5 ? 0.2 60.0 10.3 ± 1.1** 3.7 ± 0.6**
2.5 ? 0.4 92.5 19.7 ± 0.3** 4.1 ± 1.0**
2.5 ? 0.6 77.5 15.5 ± 0.2** 3.0 ± 0.3**
2.5 ? 0.8 57.5 9.3 ± 0.6** 2.8 ± 0.9**
2.5 ? 1.0 52.5 5.8 ± 0.4** 2.0 ± 0.9**
BAP ? NAA 2.5 ? 0.2 57.5 6.3 ± 0.8** 1.1 ± 0.8*
2.5 ? 0.4 50.5 5.6 ± 1.2** 1.3 ± 1.0**
2.5 ? 0.6 45.0 4.4 ± 0.6** 1.5 ± 1.1**
2.5 ? 0.8 37.5 3.1 ± 0.3** 1.0 ± 0.5*
2.5 ? 1.0 30.0 2.1 ± 0.3* 1.0 ± 0.5*
8 Plant Growth Regul (2014) 72:1–15
123
DSO-derived plants (Table 6). The solubility of flavonoids
was significantly affected by the solvent used for extraction
and these findings are in accordance with the results
obtained for Asimina tribloba (Harris and Brannan 2009)
and Salacia chinensis (Chavan et al. 2012). The fluctuation
in the phenolic and flavonoid contents in the different parts
might be due to the hormonal content, specific metabolic as
well as endogenous physiological changes taking place in
the plants. Similarly, the variation of phenol and flavonoid
contents within the plant parts was reported in 12 medicinal
plants of family Asclepiadaceae and Periplocaceae (Sur-
veswaran et al. 2010). Present findings indicated that,
exogenous supply of different PGRs during in vitro prop-
agation, pathway of regeneration was markedly influenced
the in vitro production of phenolics and flavonoids. Simi-
larly, influence of PGRs on in vitro production of bioactive
secondary metabolites is reported in Aloe arborescens
(Amoo et al. 2012). The present study has shown the total
phenolic and flavonoid contents in C. santapaui in
decreasing order as: ISO-derived plant [DSO-derived
plants [Donor mother plant.
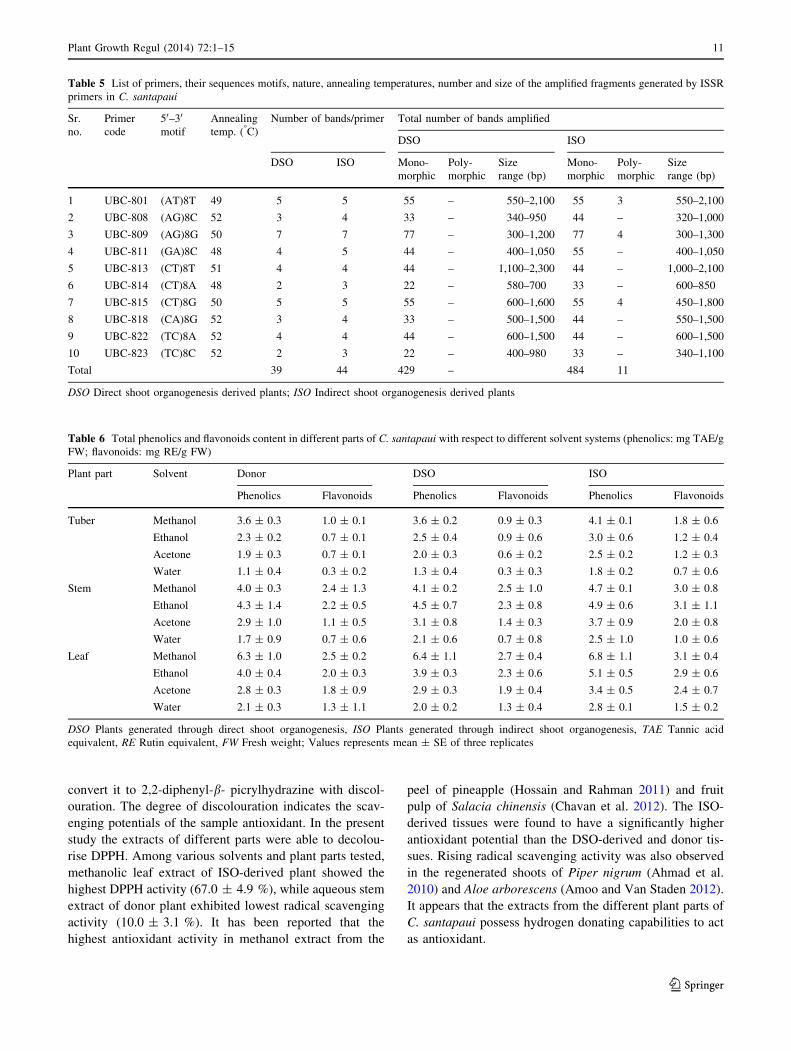
Identification of phenolic compounds
Leaves obtained from donor and micropropagated (both
DSO and ISO) C. santapaui was subjected to RP-HPLC
analysis to evaluate its phenolic composition (Fig. 4a–d).
Total phenolics and flavonoids content was recorded
highest in the leaves, hence considered for further analysis.
The major peaks identified by comparison with authentic
standards (Fig. 4a). The analysis of leaves showed the
presence of 3 major phenolic compounds viz. gallic acid,
cathechol and vanillin. The compounds identified in donor,
DSO-derived and ISO-derived leaves were quite similar to
each other, however the amount was significantly affected
by the method of regeneration. Gallic acid comprised the
maximum content followed by cathechol and vanillin in all
three sources (Fig. 4b–d). The highest amount of gallic
Table 3 Effect of different
auxins on in vitro rooting of
regenerated shoots of
C. santapaui on half-strength
MS medium
Values represents mean ± SE
of 20 replicates/treatment and
all the experiments were
repeated twice. Values are
significantly different at ns-non
significant, * P \ 0.05 and
** P \ 0.01 level according to
Dunnett multiple comparisons
test
Auxin Concentration
(mg L-1)
Rooting
frequency (%)
Number of roots/shoot
(mean ± SE)
Root length (cm)
(mean ± SE)
Control 0.0 0.0 0.0 0.0
IBA 0.5 57.5 0.7 ± 0.1ns 2.0 ± 0.4**
1.0 67.5 2.4 ± 0.6** 2.4 ± 0.6**
1.5 82.5 3.2 ± 0.4** 2.4 ± 0.3**
2.0 70.0 2.2 ± 0.4** 2.2 ± 0.4**
2.5 62.5 1.9 ± 0.3** 2.0 ± 0.3**
3.0 50.0 1.6 ± 0.3** 1.9 ± 0.3**
NAA 0.5 72.5 1.8 ± 0.3* 2.8 ± 0.4**
1.0 80.0 2.3 ± 0.4** 3.1 ± 0.4**
1.5 82.5 3.4 ± 0.5** 3.4 ± 0.5**
2.0 90.0 6.9 ± 0.8** 3.8 ± 0.4**
2.5 62.5 2.6 ± 0.6** 2.6 ± 0.4**
3.0 62.5 1.8 ± 0.3* 2.7 ± 0.4**
IAA 0.5 40.0 0.5 ± 0.1ns 1.2 ± 0.4**
1.0 57.5 1.1 ± 0.2* 1.1 ± 0.3**
1.5 72.5 1.7 ± 0.3** 1.5 ± 0.3**
2.0 65.0 2.1 ± 0.5** 1.3 ± 0.3**
2.5 52.5 1.5 ± 0.3** 1.1 ± 0.3*
3.0 47.5 1.3 ± 0.8* 1.1 ± 0.4*
NAA ? IBA 2.0 ? 0.2 75.0 1.9 ± 0.4** 2.9 ± 0.4**
2.0 ? 0.4 82.5 5.1 ± 0.7** 4.0 ± 0.5**
2.0 ? 0.6 75.0 3.0 ± 0.7** 3.0 ± 0.4**
2.0 ? 0.8 72.5 2.4 ± 0.5** 2.7 ± 0.5**
2.0 ? 1.0 60.0 2.0 ± 0.5* 2.4 ± 0.5**
NAA ? IAA 2.0 ? 0.2 57.5 1.3 ± 0.4* 1.3 ± 0.4**
2.0 ? 0.4 75.0 2.6 ± 0.5** 2.0 ± 0.3**
2.0 ? 0.6 82.5 3.3 ± 0.5** 2.0 ± 0.3**
2.0 ? 0.8 85.0 4.1 ± 0.5** 1.8 ± 0.3**
2.0 ? 1.0 85.0 4.5 ± 0.6** 1.7 ± 0.2**
Plant Growth Regul (2014) 72:1–15 9
123
acid (16.6 ppm) present in the ISO-derived leaves, fol-
lowed by of DSO-derived (14.8 ppm) and donor plant
(14.6 ppm) respectively. The gallic acid is a naturally
abundant plant phenolic compound, constituent of poly-
phenols which are known to be the principle compounds
related to the antioxidant and free radical scavenging
properties (Paixao et al. 2008). Likewise, levels of cathe-
chol and vanillin were recorded highest in leaves of ISO-
derived plants.
Antioxidant activity
Scavenging activity towards DPPH free-radical
The free radical scavenging activity of the different parts
was tested through DPPH method and the results are pre-
sented in the (Fig. 5a). The essence of DPPH method is
that the antioxidants react with the stable free radical i.e.,
2,2-diphenyl-b- picrylhydrazyl (deep violet colour) and
Fig. 3 DNA fingerprinting patterns generated by RAPD and ISSR
analysis in DSO raised plants (a RAPD primer 946, b ISSR primer
UBC 813) and ISO raised plants (c RAPD primer 961, d ISSR primer
UBC 809). Lane L: 100 bp ladder, Lane M: Mother plant, Lane 1–10:
Micropropagated plants, ;: Missing band
Table 4 List of primers, their sequences, number and size of the amplified fragments generated by RAPD primers in C. santapaui
Sr.
no.
Primer
code
Primer
sequence
(50–30)
Number of bands/primer Total number of bands amplified
DSO ISO
DSO ISO Mono-
morphic
Poly-
morphic
Size
range (bp)
Mono-
morphic
Poly-
morphic
Size
range (bp)
1 946 CTCAGGGCAT 1 1 11 – 1,700 11 – 1,700
2 961 ACTACGCGAT 5 5 55 – 300–1,700 55 – 550–1,700
3 967 GTGTGGTGAT 6 6 66 – 380–1,050 66 – 360–1,000
4 971 AGGCTGACTT 7 9 77 – 330–1,900 99 – 260–1,900
5 985 GCT CGTCGTT 4 3 44 – 650–2,100 33 – 840–1,700
6 987 TGTCGAGGTT 10 11 110 – 240–2,000 121 – 240–2,100
7 988 GAATGGGGTT 3 3 33 – 600–960 33 – 600–960
8 994 CGCACTCAAA 3 3 33 – 600–1000 33 – 600–1000
9 995 GTGTCG GAAA 5 6 55 – 360–1,500 66 – 280–1,500
10 OPC–11 AAAGCTGCGG 4 5 44 – 450–1,200 55 – 300–1,250
Total 48 52 528 – 572 –
DSO direct shoot organogenesis derived plants, ISO indirect shoot organogenesis derived plants
10 Plant Growth Regul (2014) 72:1–15
123
convert it to 2,2-diphenyl-b- picrylhydrazine with discol-
ouration. The degree of discolouration indicates the scav-
enging potentials of the sample antioxidant. In the present
study the extracts of different parts were able to decolou-
rise DPPH. Among various solvents and plant parts tested,
methanolic leaf extract of ISO-derived plant showed the
highest DPPH activity (67.0 ± 4.9 %), while aqueous stem
extract of donor plant exhibited lowest radical scavenging
activity (10.0 ± 3.1 %). It has been reported that the
highest antioxidant activity in methanol extract from the
peel of pineapple (Hossain and Rahman 2011) and fruit
pulp of Salacia chinensis (Chavan et al. 2012). The ISO-
derived tissues were found to have a significantly higher
antioxidant potential than the DSO-derived and donor tis-
sues. Rising radical scavenging activity was also observed
in the regenerated shoots of Piper nigrum (Ahmad et al.
2010) and Aloe arborescens (Amoo and Van Staden 2012).
It appears that the extracts from the different plant parts of
C. santapaui possess hydrogen donating capabilities to act
as antioxidant.
Table 5 List of primers, their sequences motifs, nature, annealing temperatures, number and size of the amplified fragments generated by ISSR
primers in C. santapaui
Sr.
no.
Primer
code
50–30
motif
Annealing
temp. (�C)
Number of bands/primer Total number of bands amplified
DSO ISO
DSO ISO Mono-
morphic
Poly-
morphic
Size
range (bp)
Mono-
morphic
Poly-
morphic
Size
range (bp)
1 UBC-801 (AT)8T 49 5 5 55 – 550–2,100 55 3 550–2,100
2 UBC-808 (AG)8C 52 3 4 33 – 340–950 44 – 320–1,000
3 UBC-809 (AG)8G 50 7 7 77 – 300–1,200 77 4 300–1,300
4 UBC-811 (GA)8C 48 4 5 44 – 400–1,050 55 – 400–1,050
5 UBC-813 (CT)8T 51 4 4 44 – 1,100–2,300 44 – 1,000–2,100
6 UBC-814 (CT)8A 48 2 3 22 – 580–700 33 – 600–850
7 UBC-815 (CT)8G 50 5 5 55 – 600–1,600 55 4 450–1,800
8 UBC-818 (CA)8G 52 3 4 33 – 500–1,500 44 – 550–1,500
9 UBC-822 (TC)8A 52 4 4 44 – 600–1,500 44 – 600–1,500
10 UBC-823 (TC)8C 52 2 3 22 – 400–980 33 – 340–1,100
Total 39 44 429 – 484 11
DSO Direct shoot organogenesis derived plants; ISO Indirect shoot organogenesis derived plants
Table 6 Total phenolics and flavonoids content in different parts of C. santapaui with respect to different solvent systems (phenolics: mg TAE/g
FW; flavonoids: mg RE/g FW)
Plant part Solvent Donor DSO ISO
Phenolics Flavonoids Phenolics Flavonoids Phenolics Flavonoids
Tuber Methanol 3.6 ± 0.3 1.0 ± 0.1 3.6 ± 0.2 0.9 ± 0.3 4.1 ± 0.1 1.8 ± 0.6
Ethanol 2.3 ± 0.2 0.7 ± 0.1 2.5 ± 0.4 0.9 ± 0.6 3.0 ± 0.6 1.2 ± 0.4
Acetone 1.9 ± 0.3 0.7 ± 0.1 2.0 ± 0.3 0.6 ± 0.2 2.5 ± 0.2 1.2 ± 0.3
Water 1.1 ± 0.4 0.3 ± 0.2 1.3 ± 0.4 0.3 ± 0.3 1.8 ± 0.2 0.7 ± 0.6
Stem Methanol 4.0 ± 0.3 2.4 ± 1.3 4.1 ± 0.2 2.5 ± 1.0 4.7 ± 0.1 3.0 ± 0.8
Ethanol 4.3 ± 1.4 2.2 ± 0.5 4.5 ± 0.7 2.3 ± 0.8 4.9 ± 0.6 3.1 ± 1.1
Acetone 2.9 ± 1.0 1.1 ± 0.5 3.1 ± 0.8 1.4 ± 0.3 3.7 ± 0.9 2.0 ± 0.8
Water 1.7 ± 0.9 0.7 ± 0.6 2.1 ± 0.6 0.7 ± 0.8 2.5 ± 1.0 1.0 ± 0.6
Leaf Methanol 6.3 ± 1.0 2.5 ± 0.2 6.4 ± 1.1 2.7 ± 0.4 6.8 ± 1.1 3.1 ± 0.4
Ethanol 4.0 ± 0.4 2.0 ± 0.3 3.9 ± 0.3 2.3 ± 0.6 5.1 ± 0.5 2.9 ± 0.6
Acetone 2.8 ± 0.3 1.8 ± 0.9 2.9 ± 0.3 1.9 ± 0.4 3.4 ± 0.5 2.4 ± 0.7
Water 2.1 ± 0.3 1.3 ± 1.1 2.0 ± 0.2 1.3 ± 0.4 2.8 ± 0.1 1.5 ± 0.2
DSO Plants generated through direct shoot organogenesis, ISO Plants generated through indirect shoot organogenesis, TAE Tannic acid
equivalent, RE Rutin equivalent, FW Fresh weight; Values represents mean ± SE of three replicates
Plant Growth Regul (2014) 72:1–15 11
123
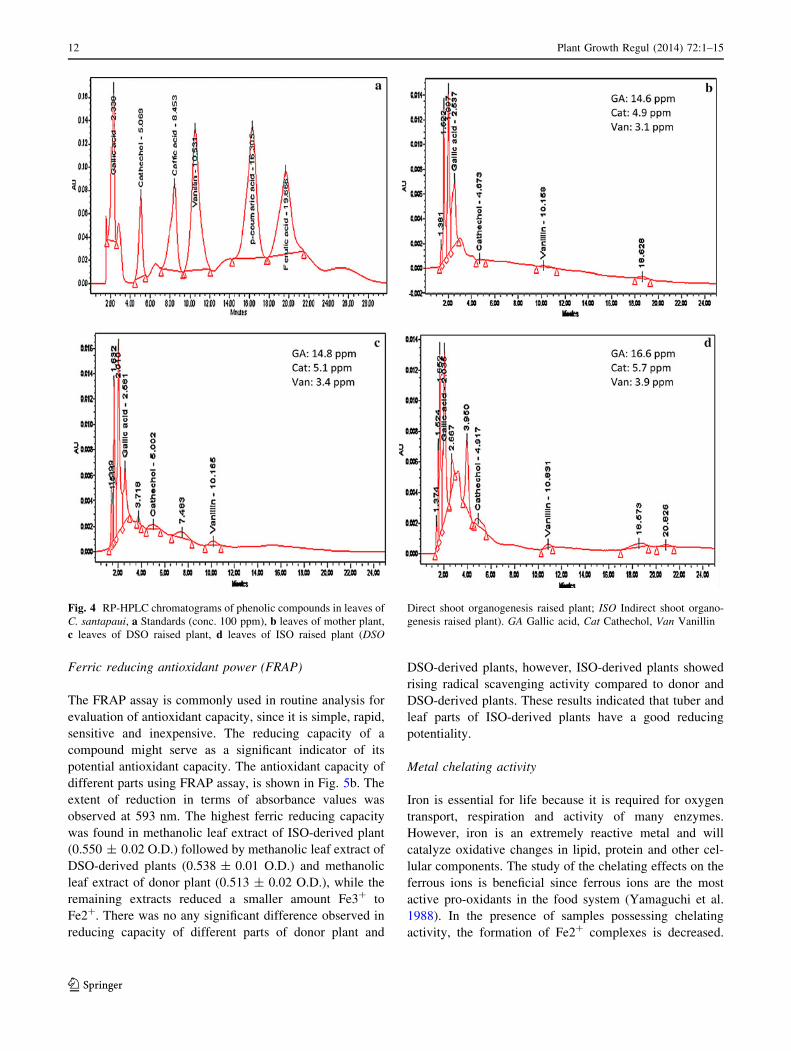
Ferric reducing antioxidant power (FRAP)
The FRAP assay is commonly used in routine analysis for
evaluation of antioxidant capacity, since it is simple, rapid,
sensitive and inexpensive. The reducing capacity of a
compound might serve as a significant indicator of its
potential antioxidant capacity. The antioxidant capacity of
different parts using FRAP assay, is shown in Fig. 5b. The
extent of reduction in terms of absorbance values was
observed at 593 nm. The highest ferric reducing capacity
was found in methanolic leaf extract of ISO-derived plant
(0.550 ± 0.02 O.D.) followed by methanolic leaf extract of
DSO-derived plants (0.538 ± 0.01 O.D.) and methanolic
leaf extract of donor plant (0.513 ± 0.02 O.D.), while the
remaining extracts reduced a smaller amount Fe3? to
Fe2?. There was no any significant difference observed in
reducing capacity of different parts of donor plant and
DSO-derived plants, however, ISO-derived plants showed
rising radical scavenging activity compared to donor and
DSO-derived plants. These results indicated that tuber and
leaf parts of ISO-derived plants have a good reducing
potentiality.
Metal chelating activity
Iron is essential for life because it is required for oxygen
transport, respiration and activity of many enzymes.
However, iron is an extremely reactive metal and will
catalyze oxidative changes in lipid, protein and other cel-
lular components. The study of the chelating effects on the
ferrous ions is beneficial since ferrous ions are the most
active pro-oxidants in the food system (Yamaguchi et al.
1988). In the presence of samples possessing chelating
activity, the formation of Fe2? complexes is decreased.
Fig. 4 RP-HPLC chromatograms of phenolic compounds in leaves of
C. santapaui, a Standards (conc. 100 ppm), b leaves of mother plant,
c leaves of DSO raised plant, d leaves of ISO raised plant (DSO
Direct shoot organogenesis raised plant; ISO Indirect shoot organo-
genesis raised plant). GA Gallic acid, Cat Cathechol, Van Vanillin
12 Plant Growth Regul (2014) 72:1–15
123
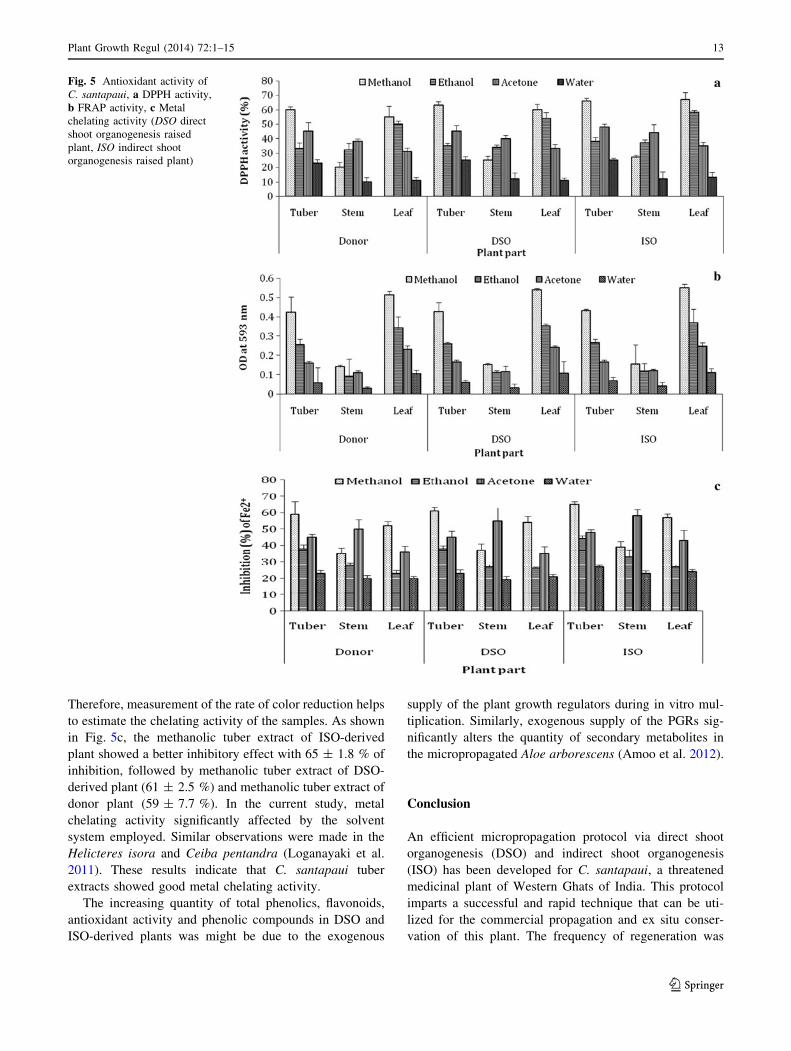
Therefore, measurement of the rate of color reduction helps
to estimate the chelating activity of the samples. As shown
in Fig. 5c, the methanolic tuber extract of ISO-derived
plant showed a better inhibitory effect with 65 ± 1.8 % of
inhibition, followed by methanolic tuber extract of DSO-
derived plant (61 ± 2.5 %) and methanolic tuber extract of
donor plant (59 ± 7.7 %). In the current study, metal
chelating activity significantly affected by the solvent
system employed. Similar observations were made in the
Helicteres isora and Ceiba pentandra (Loganayaki et al.
2011). These results indicate that C. santapaui tuber
extracts showed good metal chelating activity.
The increasing quantity of total phenolics, flavonoids,
antioxidant activity and phenolic compounds in DSO and
ISO-derived plants was might be due to the exogenous
supply of the plant growth regulators during in vitro mul-
tiplication. Similarly, exogenous supply of the PGRs sig-
nificantly alters the quantity of secondary metabolites in
the micropropagated Aloe arborescens (Amoo et al. 2012).
Conclusion
An efficient micropropagation protocol via direct shoot
organogenesis (DSO) and indirect shoot organogenesis
(ISO) has been developed for C. santapaui, a threatened
medicinal plant of Western Ghats of India. This protocol
imparts a successful and rapid technique that can be uti-
lized for the commercial propagation and ex situ conser-
vation of this plant. The frequency of regeneration was
Fig. 5 Antioxidant activity of
C. santapaui, a DPPH activity,
b FRAP activity, c Metal
chelating activity (DSO direct
shoot organogenesis raised
plant, ISO indirect shoot
organogenesis raised plant)
Plant Growth Regul (2014) 72:1–15 13
123

significantly higher in ISO compared to DSO. Considering
the importance of genetic stability in germplasm conser-
vation programme, our protocol appears to be highly
effective and to the best of our knowledge it is the first
report on genetic fidelity testing in micropropagated plants
of genus Ceropegia. Protocol described in this study may
be used for an efficient genetic transformation of this
valuable medicinal asclepiad for quality improvement. The
pathway of regeneration, plant part, solvents and PGRs
greatly influenced the in vitro secondary metabolites and
antioxidant capacity of C. santapaui.
Acknowledgments Junior research fellowship (JRF) to JJC and
financial support for this work from Department of Biotechnology
(DBT), Govertment of India, New Delhi is gratefully acknowledged.
Authors are also thankful to Dr. V. A. Bapat, Emeritus Scientist from
Plant cell culture technology section, Nuclear agriculture and bio-
technology division, Bhabha Atomic Research Centre (BARC)
Mumbai, India for his constructive comments on the manuscript.
References
Adibatti NA, Thirugnanasambantham P, Kuilothungan C (1991) A
pyridine alkaloid from Ceropegia juncea. Phytochemistry
30:2449–2450
Ahmad N, Fazal H, Abbasi BH, Rashid M, Mahmood T, Fatima N
(2010) Efficient regeneration and antioxidant potential in
regenerated tissues of Piper nigrum L. Plant Cell Tiss Organ
Cult 102:129–134
Alothman M, Bhat R, Karim AA (2009) Antioxidant capacity and
phenolic content of selected tropical fruits from Malaysia,
extracted with different solvents. Food Chem 115:785–788
Amoo SO, Van Staden J (2012) Influence of plant growth regulators
on shoot proliferation and secondary metabolite production in
micropropagated Huernia hystrix. Plant Cell Tiss Organ Cult.
doi:10.1007/s11240-012-0230-x
Amoo SO, Finnie JF, Van Staden J (2009) In vitro propagation of
Huernia hystrix: an endangered medicinal and ornamental
succulent. Plant Cell Tiss Organ Cult 96:273–278
Amoo SO, Aremu AO, Van Staden J (2012) In vitro plant
regeneration, secondary metabolite production and antioxidant
activity of micropropagated Aloe arborescens Mill. Plant Cell
Tiss Organ Cult 111:345–358
Baskaran P, Jayabalan N (2005) An efficient micropropagation
system for Eclipta alba—a valuable medicinal herb. In Vitro
Cell Dev Biol Plant 41:532–539
Beena MR, Martin KP, Kirti PB, Hariharan M (2003) Rapid in vitro
propagation of medicinally important Ceropegia candelabrum.
Plant Cell Tiss Organ Cult 72:285–289
Benzie IF, Strain JJ (1996) Ferric reducing ability of plasma (FRAP)
as a measure of antioxidant power: the FRAP assay. Anal
Biochem 239:70–76
Bhatia R, Singh KP, Jhang T, Sharma TR (2009) Assessment of
clonal fidelity of micropropagated gerbera plants by ISSR
markers. Sci Hortic 119:208–211
Bhatia R, Singh KP, Sharma TR, Jhang T (2011) Evaluation of the
genetic fidelity of in vitro propagated gerbera (Gerbera jameso-
nii Bolus) using DNA-based markers. Plant Cell Tiss Organ Cult
104:131–135
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of free radical
method to evaluate antioxidant activity. Lebensm Wiss Technol
28:25–30
Bruyns PV (2003) Three new succulent species of Apocynaceae
(Asclepiadoideae) from southern Africa. Kew Bull 58:427–435
Chandore AN, Nimbalkar MS, Gurav RV, Bapat VA, Yadav SR
(2010) An efficient micropropagation protocol for multiplication
and restoration of Ceropegia fantastica Sedgw: a critically
endangered plant species. Curr Sci 99:1593–1596
Chang C, Yang M, Wen H, Chern J (2002) Estimation of total
flavonoid content in propolis by two complementary colorimetric
methods. J Food Drug Anal 10:178–182
Chavan JJ, Nimbalkar MS, Adsul AA, Kambale SS, Gaikwad NB,
Dixit GB, Gurav RV, Bapat VA, Yadav SR (2011a) Microprop-
agation and in vitro flowering of endemic and endangered plant
Ceropegia attenuata Hook. J Plant Biochem Biotechnol
20:276–282
Chavan JJ, Nimbalkar MS, Gaikwad NB, Dixit GB, Yadav SR
(2011b) In vitro propagation of Ceropegia spiralis wight-an
endemic and rare potential ornamental plant of peninsular India.
Proc Nat Acad Sci India Sect B 81:120–126
Chavan JJ, Jagtap UB, Gaikwad NB, Dixit GB, Bapat VA (2012)
Total phenolics, flavonoids and antioxidant activity of Sapta-
rangi (Salacia chinensis L.) fruit pulp. J Plant Biochem
Biotechnol. doi:10.1007/s13562-012-0169-3
Devarumath RM, Nandy S, Rani V, Marimuthu S, Muraleedharan N
(2002) RAPD, ISSR and RFLP fingerprints as useful markers to
evaluate genetic integrity of micropropagated plants of three
diploid and triploid elite tea clones representing Camellia
sinensis (China type) and C. assamica ssp. assamica (Assam-
India type). Plant Cell Rep 21:166–173
Dinis T, Madeira V, Almeida T (1994) Action of phenolic derivatives
(acetaminophen, salicylate and 5-aminosalicylate) as inhibitors
of membrane lipid peroxidation and peroxyl radical scavengers.
Arch Biochem Biophys 315:161–165
Doyle JJ, Doyle JL (1990) Isolation of plant DNA from fresh tissue.
Focus 12:13–15
Fitch MM, Moore PH (1990) Comparison of 2,4-D and picloram for
selection of long-term totipotent green callus cultures of
sugarcane. Plant Cell Tiss Organ Cult 20:157–163
Ganesan M, Jayabalan N (2006) Influence of cytokinins, auxins and
polyamines on in vitro mass multiplication of cotton (Gossypium
hirsutum L. cv. SVPR2). Indian J Exp Biol 44:506–513
Ghatge SR (2007) In vitro cultural studies in medicinal plants viz.
Hemidesmus indicus (L.) Schult and Rubia cordifolia L. Ph.D.
thesis, submitted to Shivaji University, Kolhapur
Giridhar P, Gururaj HB, Ravishankar GA (2005) In vitro aerial part
multiplication through aerial part tip cultures of Decalepis
hamiltonii wight & Arn., a threatened plant endemic to southern
India. In Vitro Cell Dev Biol Plant 41:77–80
Goto S, Thakur RC, Ishii K (1998) Determination of genetic stability
in long-term micropropagated shoots of Pinus thunbergii Parl.
using RAPD markers. Plant Cell Rep 18:193–197
Harris GG, Brannan RG (2009) A preliminary evaluation of
antioxidant compounds, reducing potential and radical scaveng-
ing of pawpaw (Asimina tribloba) fruit pulp from different stages
of ripeness. LWTFood Sci Technol 42:275–279
Hodgkiss RJ (2004) http://www.succulent-plant.com/ceropg.html
Hossain MA, Rahman SMM (2011) Total phenolics, flavonoids and
antioxidant activity of tropical fruit pineapple. Food Res Int
44:672–676
Jagtap UB, Waghmare SR, Lokhande VH, Suprasanna P, Bapat VA
(2011) Preparation and evaluation of antioxidant capacity of
jackfruit (Artocarpus heterophyllus Lam.) wine and its protec-
tive role against radiation induced DNA damage. Ind Crop Prod
34:1595–1601
14 Plant Growth Regul (2014) 72:1–15
123

Jain SK, Defillips RA (1991) Asclepiadaceae. In: Medicinal plants of
India. Vol. 1. Algonac, India, pp.89–94
Joshi P, Dhawan V (2007) Assessment of genetic fidelity of
micropropagated Swertia chirayita plantlets by ISSR marker
assay. Biol Plantarum 51:22–26
Khare CP (2007) Indian medicinal plants. An illustrated dictionary.
Springer, Berlin, pp 139–140
Komalavalli N, Rao MV (2000) In vitro micropropagation of
Gymnema sylvestre—a multipurpose medicinal plant. Plant Cell
Tiss Organ Cult 61:97–105
Loganayaki N, Siddhuraju P, Manian S (2011) Antioxidant activity
and free radical scavenging capacity of phenolic extracts from
Helicteres isora L and Ceiba pentandra L. J Food Sci Technol.
doi:10.1007/s13197-011-0389-x
Luciani GF, Mary AK, Pellegrini C, Curvetto NR (2006) Effects of
explants and growth regulators in garlic callus formation and
plant regeneration. Plant Cell Tiss Organ Cult 87:139–143
Mabberly DJ (1987) The plant book. Cambridge University Press,
Cambridge, pp 114–115
Martin KP (2003) Rapid in vitro multiplication and ex vitro rooting of
Routela aquatica Lour; a rare rhoeophytic woody medicinal
plant. Plant Cell Rep 21:415–420
Martins M, Sarmento D, Oliveira M (2004) Genetic stability of
micropropagated almond plantlets, as assessed by RAPD and
ISSR markers. Plant Cell Rep 23:492–496
McNew R (2002) http://www.shoalcreeksucculents.com
Murthy KSR, Kondamudi R, Vijayalakshmi V (2010) Micropropa-
gation of an endangered medicinal plant Ceropegia spiralis
Wight. J Agricult Tech 6:179–191
Murthy KSR, Kondamudi R, Reddy MC, Karuppusamy S, Pullaiah T
(2012) Check-list and conservation strategies of the genus
Ceropegia in India. Int J Biodivers Conserv 4:304–315
Nadkarni KM (1976) Indian materia medica. Popular Prakashan,
Bombay, pp 303–304
Nikam TD, Savant RS (2009) Multiple shoot regeneration and
alkaloid cerpegin accumulation in callus culture of Ceropegia
juncea Roxb. Physiol Mol Biol Plants 15:71–77
Paixao N, Pereira V, Marques JC, Camara JS (2008) Quantification of
polyphenols with potential antioxidant properties in wines using
reverse phase HPLC. J Sep Sci 31:2189–2198
Patil VM (1998) Micropropagation studies in Ceropegia spp. In Vitro
Cell Dev Biol Plant 34:240–243
Phulwaria M, Shekhawat NS, Rathore JS, Singh RP (2013) An
efficient in vitro regeneration and ex vitro rooting of Ceropegia
bulbosa Roxb.—a threatened and pharmaceutical important
plant of Indian Thar Desert. Ind Crop Prod 42:25–29
Prasad PJN, Chakradhar T, Pullaiah T (2004) Micropropagation of
Cryptolepis buchanani Roem and Schult. Taiwania 49:57–65
Reynolds S (2006) http://www.sagereynolds.com/cero/clist.com
Rossi F, Baraldi R, Facini O, Lereari B (1993) Photomorphogenic
effects on in vitro rooting of Prunus roostock GF 655-2. Plant
Cell Tiss Organ Cult 32:145–151
Singleton VL, Rossi JA (1965) Colorimetry of total phenolics with
phosphomolybdic-phosphotungstic acid reagents. Am J Enol
Viticult 16:144–158
Socorro O, Tarrega I, Rivas F (1998) Essential oils from wild and
micropropagated plants of Origanum bastetanum. Phytochemis-
try 48:1347–1349
Sukumar E, Gopal RH, Rao RB, Viswanathan S, Thirugnanasam-
bantham P, Vijayasekaran V (1995) Pharmacological actions of
cerpegin, a novel pyridine alkaloid from Ceropegia juncea.
Fitoterapia 66:403–406
Surveswaran S, Cai YZ, Xing J, Corke H, Sun M (2010) Antioxidant
properties and principal phenolic phytochemicals of Indian
medicinal plants from Asclepiadoideae and Periplocoideae. Nat
Prod Res 24:206–221
Swarnkar S, Katewa SS (2008) Ethnobotanical observation on
tuberous plants from tribal area of Rajasthan (India). Ethnobot
Leafl 12:647–666
Ugraiah A, Sreelatha V, Krishna-Reddy PV, Rajasekhar K, Sandhya-
Rani S, Karuppusamy S, Pullaiah T (2011) In vitro shoot
multiplication and conservation of Caralluma bhupenderiana
Sarkaria—an endangered medicinal plant from South India. Afr
J Biotechnol 10:9328–9336
Vasil I (2008) A history of plant biotechnology: from the cell theory
of Schleiden and Schwann to biotech crops. Plant Cell Rep
27:1423–1440
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV
(1990) DNA polymorphisms amplified by arbitrary primers areuseful as genetic markers. Nucl Acid Res 18:6531–6535
Xanthopoulou MN, Fragopoulou E, Kalathara K, Nomikos T,
Karantonis HC, Antonopoulou S (2010) Antioxidant and anti-
inflammatory activity of red and white wine extracts. Food Chem
120:665–672
Yadav SR, Kamble MY (2008) Threatened Ceropegias of the
Western Ghats and strategies for their conservation. In: Rawant
G.S. (Ed). Special habitats and threatened plants of India. ENVIS
Bulletin Wildlife and protected Areas, Wildlife Institute of India:
Deharadun, India, 11:123–134
Yamaguchi R, Tatsumi MA, Kato K, Yoshimitsu U (1988) Effect of
metal salts and fructose on the autoxidation of methyl linoleate
in emulsions. Agric Biol Chem 52:849–850
Yan MM, Xu C, Kim CH, Um YC, Bah AA, Guo DP (2009) Effects
of explant type, culture media and growth regulators on callus
induction and plant regeneration of Chinese jiaotou (Allium
chinense). Sci Hortic 123:124–128
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome fingerprinting
by simple sequence repeat (SSR)-anchored polymerase chain
reaction amplification. Genomics 20:176–183
Plant Growth Regul (2014) 72:1–15 15
123
Related Documents











