Effects of sertraline on behavioral alterations caused by environmental enrichment and social isolation ☆ Engin Yildirim a, ⁎, Kevser Erol a, 1 , Emel Ulupinar b, 2 a Eskisehir Osmangazi University, Faculty of Medicine, Department of Pharmacology, 26480 Eskisehir, Turkey b Eskisehir Osmangazi University, Faculty of Medicine, Department of Anatomy, Eskisehir, Turkey abstract article info Article history: Received 25 June 2011 Received in revised form 13 November 2011 Accepted 22 December 2011 Available online 10 January 2012 Keywords: Rat Behavioral alteration Motor activity Anxiety Depression Environmental conditions are known to play a critical role in the pathogenesis of affective disorders. In this study, the effects of sertraline, a selective serotonin (5-HT) reuptake inhibitor, on anxiety- and depression- like behaviors were investigated in rats reared in different housing conditions. Wistar rats of both sexes were divided into three groups according to their rearing conditions (Enriched = EC, Isolated = IC and Standard = SC), after weaning at postnatal day 21. While animals in control conditions were housed as a group of 4 rats in regular size plexiglass cages, social isolation groups were housed individually in metal cages. Animals in enriched conditions were housed as a group of 12 rats in specially designed cages equipped with different stimulating objects. Six weeks later, activitymeter, elevated plus maze, rotarod, grip, forced swimming and sucrose preference tests were applied to all animals and all of the tests were repeated after i.p. injection of sertraline (10 mg/kg/day) for 7 days. Environmental enrichment reduced the stereotypic behavior, improved the motor coordination and facilitated the learning skills in animals. However, housing conditions affected depression-like parameters, but not anxiety-like parameters. Sertraline treatment reduced the depression-like effect in EC and SC, but not in IC. It decreased anxiety-like behavior in IC while increased in EC. Socially isolated animals preferentially consumed more sucrose and water than the other groups, and interestingly, these differences became more significant following sertraline treatment. These results show that the responses of animals to anti-depressive drugs could be differentially affected by the behavioral consequences of the diverse housing conditions. Thus, to improve the treatment of depression; behavioral consequences of diverse housing conditions should be taken into consideration. © 2011 Elsevier Inc. All rights reserved. 1. Introduction Clinical and laboratory evidences suggest that environmental stimulation and experience are necessary for healthy brain develop- ment (Bors and Forrin 1996; Branchi and Alleva 2006; Branchi et al 2010). Children raised in highly stimulating or enriched environ- ments exhibit enhanced behavioral and cognitive outcomes, whereas the opposite circumstances cause impairments in these functions (Di Pietro, 2000). In laboratory animals, environmental complexity alters brain structure (e.g. brain mass, cortical thickness, the size of neuro- nal nuclei, dendritic complexity and density of synaptic contacts) (Rosenzweig and Bennett, 1996; Mattson et al., 2001; Fernández- Teruel et al., 2002). More recently, it has been demonstrated that enriched housing conditions improves visual, auditory and somatosensorial cortex processing and cellular plasticity (Engineer et al., 2004; Sale et al., 2004; Guic et al., 2008; Baroncelli et al., 2010). With regard to cognitive functions, it was shown that environ- mental enrichment improves spatial learning and long-term memory by stimulating the production of neurotrophic factors, leading to chromatin remodeling and histone acetylation, inducing plastic changes and enhancing the survival and/or proliferation of hippo- campal neurons and astrocytes (Kempermann et al., 1997; Nilsson et al., 1999; Branchi et al., 2006; Viola et al., 2009). Moreover, in ani- mal models of central nervous system injury and stroke, environmen- tal enrichment has beneficial effects associated with neurogenesis, enhanced dendritic growth and gene expression (Fischer and Peduzzi, 2007; Knieling et al., 2009). In contrast, social isolation is considered as a long-term stressor and environmental impoverishment. Isolated rats become more vulnerable and responsive to environmental changes (Mattson et al., 2001) and ex- hibit impairments in learning and memory tasks (Larsson et al., 2002). Additionally, alterations in the dopaminergic mesolimbic system, dis- rupted prepulse inhibition, novel object recognition and attentional set-shifting and other behavioral, morphological and neurochemical ab- normalities, strongly resembling the core features of schizophrenia, can Pharmacology, Biochemistry and Behavior 101 (2012) 278–287 ☆ The study was presented as a poster in 16th World Congress on Basic and Clinical Pharmacology, 17–23 July, 2010, Copenhagen, Denmark. ⁎ Corresponding author. Tel.: +90 505 541 38 76(Gsm). E-mail addresses: [email protected] (E. Yildirim), [email protected] (K. Erol), [email protected] (E. Ulupinar). 1 Tel.: +90 535 968 88 15(Gsm). 2 Tel.: +90 533 453 6668(Gsm). 0091-3057/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.pbb.2011.12.017 Contents lists available at SciVerse ScienceDirect Pharmacology, Biochemistry and Behavior journal homepage: www.elsevier.com/locate/pharmbiochembeh

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
Contents lists available at SciVerse ScienceDirect
Pharmacology, Biochemistry and Behavior
j ourna l homepage: www.e lsev ie r .com/ locate /pharmbiochembeh
Effects of sertraline on behavioral alterations caused by environmental enrichmentand social isolation☆
Engin Yildirim a,⁎, Kevser Erol a,1, Emel Ulupinar b,2
a Eskisehir Osmangazi University, Faculty of Medicine, Department of Pharmacology, 26480 Eskisehir, Turkeyb Eskisehir Osmangazi University, Faculty of Medicine, Department of Anatomy, Eskisehir, Turkey
☆ The study was presented as a poster in 16th WorldPharmacology, 17–23 July, 2010, Copenhagen, Denmark⁎ Corresponding author. Tel.: +90 505 541 38 76(Gs
E-mail addresses: [email protected] (E. Yildirim),[email protected] (E. Ulupinar).
1 Tel.: +90 535 968 88 15(Gsm).2 Tel.: +90 533 453 6668(Gsm).
0091-3057/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.pbb.2011.12.017
a b s t r a c t
a r t i c l e i n f oArticle history:Received 25 June 2011Received in revised form 13 November 2011Accepted 22 December 2011Available online 10 January 2012
Keywords:RatBehavioral alterationMotor activityAnxietyDepression
Environmental conditions are known to play a critical role in the pathogenesis of affective disorders. In thisstudy, the effects of sertraline, a selective serotonin (5-HT) reuptake inhibitor, on anxiety- and depression-like behaviors were investigated in rats reared in different housing conditions. Wistar rats of both sexeswere divided into three groups according to their rearing conditions (Enriched = EC, Isolated = IC andStandard = SC), after weaning at postnatal day 21. While animals in control conditions were housed as agroup of 4 rats in regular size plexiglass cages, social isolation groups were housed individually in metalcages. Animals in enriched conditions were housed as a group of 12 rats in specially designed cages equippedwith different stimulating objects. Six weeks later, activitymeter, elevated plus maze, rotarod, grip, forcedswimming and sucrose preference tests were applied to all animals and all of the tests were repeated afteri.p. injection of sertraline (10 mg/kg/day) for 7 days. Environmental enrichment reduced the stereotypicbehavior, improved the motor coordination and facilitated the learning skills in animals. However, housingconditions affected depression-like parameters, but not anxiety-like parameters. Sertraline treatmentreduced the depression-like effect in EC and SC, but not in IC. It decreased anxiety-like behavior in IC whileincreased in EC. Socially isolated animals preferentially consumed more sucrose and water than the othergroups, and interestingly, these differences became more significant following sertraline treatment. Theseresults show that the responses of animals to anti-depressive drugs could be differentially affected by thebehavioral consequences of the diverse housing conditions. Thus, to improve the treatment of depression;behavioral consequences of diverse housing conditions should be taken into consideration.
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
Clinical and laboratory evidences suggest that environmentalstimulation and experience are necessary for healthy brain develop-ment (Bors and Forrin 1996; Branchi and Alleva 2006; Branchi et al2010). Children raised in highly stimulating or enriched environ-ments exhibit enhanced behavioral and cognitive outcomes, whereasthe opposite circumstances cause impairments in these functions (DiPietro, 2000). In laboratory animals, environmental complexity altersbrain structure (e.g. brain mass, cortical thickness, the size of neuro-nal nuclei, dendritic complexity and density of synaptic contacts)(Rosenzweig and Bennett, 1996; Mattson et al., 2001; Fernández-Teruel et al., 2002). More recently, it has been demonstrated thatenriched housing conditions improves visual, auditory and
Congress on Basic and Clinical.m)[email protected] (K. Erol),
rights reserved.
somatosensorial cortex processing and cellular plasticity (Engineeret al., 2004; Sale et al., 2004; Guic et al., 2008; Baroncelli et al.,2010). With regard to cognitive functions, it was shown that environ-mental enrichment improves spatial learning and long-term memoryby stimulating the production of neurotrophic factors, leading tochromatin remodeling and histone acetylation, inducing plasticchanges and enhancing the survival and/or proliferation of hippo-campal neurons and astrocytes (Kempermann et al., 1997; Nilssonet al., 1999; Branchi et al., 2006; Viola et al., 2009). Moreover, in ani-mal models of central nervous system injury and stroke, environmen-tal enrichment has beneficial effects associated with neurogenesis,enhanced dendritic growth and gene expression (Fischer andPeduzzi, 2007; Knieling et al., 2009).
In contrast, social isolation is considered as a long-term stressor andenvironmental impoverishment. Isolated rats become more vulnerableand responsive to environmental changes (Mattson et al., 2001) and ex-hibit impairments in learning and memory tasks (Larsson et al., 2002).Additionally, alterations in the dopaminergic mesolimbic system, dis-rupted prepulse inhibition, novel object recognition and attentionalset-shifting and other behavioral, morphological and neurochemical ab-normalities, strongly resembling the core features of schizophrenia, can

279E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
be observed in rats following isolation (McLean et al., 2010; Quan et al.,2010). It is alsowell characterized that isolated animals display locomo-tor hyperactivity in an inescapable novel environment (Hall et al., 1998;Heidbreder et al., 2000); whereas animals reared in enriched environ-ment display lower spontaneous activity (Rosenzweig et al., 1978).
Nevertheless, the effects of differential environmental conditions atthe behavioral level remain controversial; because inconsistent effectshave been reported in the literature (Peña et al., 2006). These variationscan derive from neurochemical alterations in the cortico-striatal andbrain stem monoamine pathways. It was shown that variations inphysical and social environment during early rearing lead to gender-dependent behavioral and neurochemical alterations (Fernández-Teruel et al., 2002). Among them, neurotransmitters acetylcholine,norepinephrine (NE), dopamine, serotonin (5-HT) and nitric oxidemay be of special interest, because they play important roles in themodulation of neuronal plasticity (Fernández-Teruel et al., 2002;Lores-Arnaiz et al., 2007). While environmental enrichment augmentsthe serotonin (5-HT) concentration in the prefrontal cortex, social isola-tion reduces the levels of norepinephrine in ventral striatum (Brenes etal., 2008). Furthermore, in contrast to isolated conditions, enriched rear-ing conditions have been shown to have behavioral and neurochemicalantidepressant-like effects (Brenes et al., 2009). Nevertheless, likewiseto behavioral studies, there has been some controversy regarding the ef-fects of rearing conditions on monoamine neurotransmission. Previousstudies have reported that isolation rearing reduces 5-HT content by en-hancing serotonin turnover in brain regions such as hippocampus andprefrontal cortex; whereas increases 5-HT release in ventral striatum,namely in nucleus accumbens (Fulford and Marsden, 1998; Hall 1998;Brenes et al., 2008). However, several current studies revealed that iso-lation rearing increases dopamine and serotonin levels in prefrontalcortex and nucleus accumbens, as well as serotonin turnover rate inthe nucleus accumbens in adult rats (Lukkes et al., 2009). It is also pos-sible that enhanced serotonin levels in this region might be responsiblefor the subsequent dopamine release. Thus, the level of serotonin inmesolimbic and mesocortical regions, that are vulnerable to environ-mental alterations, plays an important regulatory role inmanyemotion-al and behavioral functions (Galani et al., 2007; Segovia et al., 2009).
Selective serotonin re-uptake inhibitors (SSRIs) are commonlyused in the treatment of depression. These drugs can easily passthrough the brain–blood barrier, inhibit the serotonin transporter(5-HTT) and increase the extracellular serotonin level in the brain(Baumann and Rochat, 1995). However, relatively little is knownabout the effects of SSRIs on individuals exposed to differential envi-ronmental conditions. Recently, the effects of serotonergic antide-pressant fluoxetine have been tested on socially isolated male rats(Brenes et al. 2009). In that study, chronic fluoxetine treatmentcould counteract the behavioral and neurochemical alterations inadult animals induced by social isolation. In another study, the effectsof early experiences on the efficacy of antidepressants at adulthoodwere tested by using the mouse communal nest paradigm (Branchiet al., 2010). It was found that a 3-week fluoxetine treatment affectedonly the animals housed in standard conditions; but did not affect theones reared in the early socially enriched environment. Therefore, inthis study, we aimed to investigate the effects of another SSRI, sertra-line, on the behavioral outcomes of differential housing conditions. Tothis purpose, we first investigated the effects of enriched, standardand isolated rearing conditions on the several types of activities andbehavioral parameters of animals. Then, we analyzed the effects ofsertraline treatment on these specific responses.
2. Materials and methods
2.1. Animals and housing
The animals were maintained in our animal facilities under a tem-perature controlled environment (21±2 °C), with 12-h light/dark
cycle. Standard food and water were freely available. All behavioraltests were conducted between 9:00 and 13:00 a.m. Wistar rats de-rived from six different mothers were assigned to three differentgroups according to housing conditions (Standard = SC, Enriched =EC and Isolated = IC), after weaning at postnatal day 21. Sincelitter-gender composition has profound effects on the normal devel-opment of behavioral and neuroendocrine responses, and social ex-periences (Cirulli et al 1997; Laviola and Terranova, 1998); six maleand female offspring from different litters were housed all togetherin an enrichment cage. In order to keep the number and gender of an-imals similar for each housing condition, 12 animals were also used inother groups. Animals in EC group were reared in a specially designedcage consisting of three large acrylic boxes (30×30×50 cm) con-nected to each other via re-arrangeable tunnels and containing differ-ent food dispensers, water bottles, two running wheels, climbingladders, shelter and a variety of stimulating objects changed threetimes per week and taking care not to repeat same objects in succes-sive weeks. Standard housing conditions consisted of a15×25×35 cm plexiglass cage with sawdust at the bottom, housinga group of 4 rats (equal number of each gender) at a time and animalsin IC group were housed individually in standard size metal cages. Allbehavioral experiments were done in accordance to the guidelines forAnimal Care and Use of Eskisehir Osmangazi University, and were ap-proved by the Local Ethical Committee for Animal Experimentations.
2.2. Procedure and treatment
The groups at postnatal day 21 were kept in their respective cagesfor six weeks and then 0.1 ml saline was injected intraperitoneally toall animals for 7 days. At postnatal day 70 all rats were tested 1 h afterthe last i.p. injection in activitymeter for locomotor activity, elevatedplus maze for anxiety-like effect and learning, rotarod and grip testfor motor function, sucrose preference test and forced swimmingtest for depression-like effect. After completion of all tests, animalswere treated with i.p. injection of sertraline (10 mg/kg/day) for7 days. Repeated doses of sertraline were administered since it ischronically used in practice and this period is accepted as minimumperiod of usage for displaying its chronical effects (Bianci et al.,2002; Feltenstein and Sufka 2005; Muraki et al 2008). At postnatalday 77 all of the tests (except elevated plus maze for testing memory)were repeated 1 h after the last i.p. injection of sertraline.
2.3. Locomotor activity experiments
Spontaneous locomotor activities of animals were monitored byusing an animal activity monitoring system. Rats were placed to asquare plexiglass box (40×40×40 cm) and locomotor activity wasrecorded simultaneously by a video-computerized system (MAY9803 Activity Monitor, Commat Iletişim Ltd. May Pentium Computer).Ambulatory and resting activity were evaluated over a 5-min period.During each test, stereotypic (repetitive behavior), horizontal, verti-cal, ambulatory and total number of movements, and total traveleddistance were measured as the total counts of beam interruptions inthe sensors. The activity chamber was cleaned with 70% ethanolafter each use to eliminate any olfactory cues of the previously testedanimal.
2.4. Elevated plus maze
The apparatus consisted of a plus-shaped maze elevated 70 cmabove the ground level with two open (10 cm in width×50 cm inlength) and two closed arms with 40 cm height walls border. Therewas a central square arena (10×10 cm) at the junction of open andclosed arms where the animal is placed at the start of the experiment.The open arms of elevated plus maze had a small rim of 2.5 mm high.The apparatus was designed to measure anxiety-related behavior in

280 E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
rats with closed arms representing security and open arms insecurityand fear of heights (Pellow et al., 1985). The test rat was placed on thecenter of the maze facing an open arm, and the time spent in openand closed arms and the number of entries into both open and closedarms was recorded during a 5 min observation period.
One week after the first experiments the animals were tested foran evaluation of memory by using elevated plus maze (Itoh et al.,1990). The test was performed as follows: the animals were individ-ually placed at the end of one open arm facing away from the centralplatform and the time it took for the rat to move from the open arm toeither of the enclosed arms (transfer latency) was recorded. Thetransfer latency was the elapsed time between the times at whichall its legs crossed the fine white line in the middle of the enclosedarm. The time spent in the enclosed arm after the measurement ofthe transfer latency was also recorded. The rat was taken out fromthe maze when it got out of enclosed arm and all its legs enteredthe central platform after measurement of the transfer latency. Eachrat performed one trial per day. The procedure was repeated everyday, over 5 days.
2.5. Rotarod
It consisted of a rotating rod (70 mm diameter) (Thouvarecq et al.,2001). The rod was divided by circular partitions into four compart-ments permitting four rats at a time. Effects for motor deficit (ifany) were evaluated using rotarod. The speed was set at 20 rpm.The number of seconds each rat remained on the rotating rod wasrecorded for a maximum 300 s.
2.6. Grip test
Grip test was performed according to the method of Moran et al.(1995). Briefly, the apparatus with a string of 50 cm length, pulledstraight between two vertical supports, was used. The rat was placedon the wire by its fore paws and evaluated according to the followingscale: 0 = Fall off; 1 = Hangs on string; 2 = As for 1, but attempts toclimb on string; 3 = Hangs onto string by two forepaws plus one orboth hind paws; 4 = Hangs onto string by all fore and hind pawsplus tail wrapped around string; and 5= Escape. This method provid-ed a quantitative estimate of the grip strength and coordination abil-ity of the animals.
2.7. Forced swimming test
This paradigm was performed as described previously (Porsolt etal., 1978). The test is a 2-day procedure in which rats swim underconditions in which escape is not possible. On the first day, the ratswere placed in individual plexiglass cylinders (35×20 cm in diame-ter) that had previously been filled with water (23–25 °C) up to21.5±1.5 cm from the bottom. After 15 min the rats were removedfrom the water, dried with towels and placed in a warmed enclosure.The test sessions (5 min) were conducted 24 h later and immobility,climbing and diving periods were recorded by an observer blind tothe experimental conditions.
2.8. Sucrose preference test
In our version of the test, animals were kept in their respective hous-ing conditions; but during 48 h. test period, two bottles were available ineach cage (Brenes and Fornaguera, 2008). One bottle was containing500 ml of 32% sucrose solution (w/v) and the other was filled with500 ml tap water. At the end of test period, the bottles were removedand sucrose, water and total liquid (sucrose+water) consumption (ml)were measured. The total amounts of consumption were dividedinto the number of animals in each cage. Sucrose preference was
calculated as follows: Preference %=[(sucrose consumption/sucrose+water consumption)×100] (Brenes and Fornaguera 2008).
2.9. Statistical analysis
The data were expressed as mean±SEM. Group comparisonswere preferentially analyzed by using two-way variance analysis(ANOVA) as a 3-level factor. However, where both assumptions fornormality of distribution and homogeneity of variance were violated,equivalent non-parametric statistics (Kruskal–Wallis) were used. Anunpaired (Student's) t test was used for comparisons between groupsand p value of less than 0.05 was considered to indicate statistical sig-nificance for each analysis.
3. Results
3.1. Activitymeter analyses
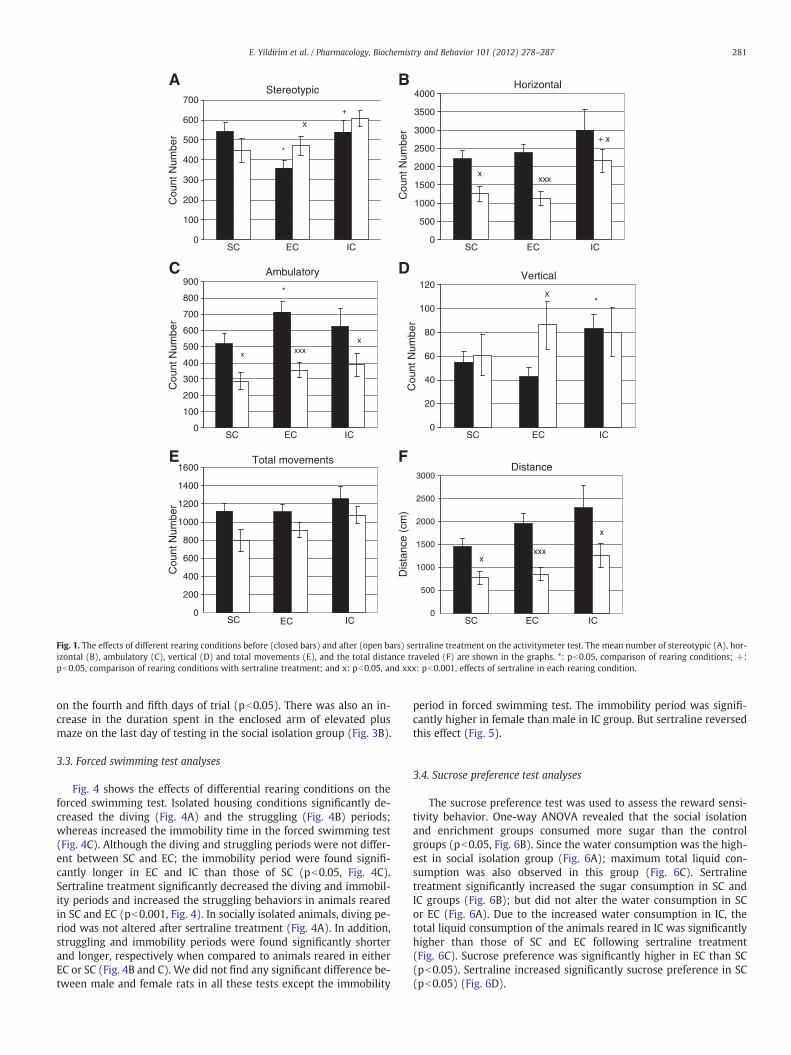
Differential housing conditions did not cause a significant changein the number of horizontal (Fig. 1B) and total movements (Fig. 1E),as well as the total traveled distance (Fig. 1F) in the activitymetertest. However, rearing animals in the enriched environment signifi-cantly (pb0.05) reduced the number of stereotypic movements(Fig. 1A); while increasing the ambulatory movements (Fig. 1C). Insocially isolated animals, on the other hand, a significant increasewas observed in the vertical activity counts (pb0.05, Fig. 1D).
After sertraline treatment, a significant decrease was observed inthe number of horizontal (Fig. 1B) and ambulatory movements(Fig. 1C), and the total traveled distance (Fig. 1F) in each group.This difference was more evident in animals reared in EC (pb0.001)than those reared in SC and IC (pb0.05). Interestingly, sertraline in-jections increased the stereotypic (Fig. 1A) and vertical activitycounts (Fig. 1D) only in the enrichment group.
On the other hand, comparisons of the rearing conditions withsertraline treatment did not reveal any significant difference in ma-jority of the parameters, except that the number of horizontal move-ments significantly increased in socially isolated animals (pb0.05,Fig. 1B).
3.2. The elevated plus-maze test analyses
The elevated plus-maze test was used to test the anxiety-likelevels of the rats. As shown in Fig. 2, neither the time spent in theopen arms (Fig. 2A) nor the frequency of the arm entries (Fig. 2B)was different among the groups reared in different environmentalconditions.
However, sertraline treatment significantly increased the timespent in the open arms of the elevated plus-maze in the environmen-tal enrichment group; while decreasing it in the social isolation group(pb0.05, Fig. 2A). Following sertraline injections, a significant de-crease in the entry numbers of the arms was observed in eachgroup (pb0.001, Fig. 2B). Analysis of differences with ANOVArevealed that the number of crossings was significantly higher in an-imals reared in IC than their EC and SC littermates [F (8.127), pb0.05,(Fig. 2B)].
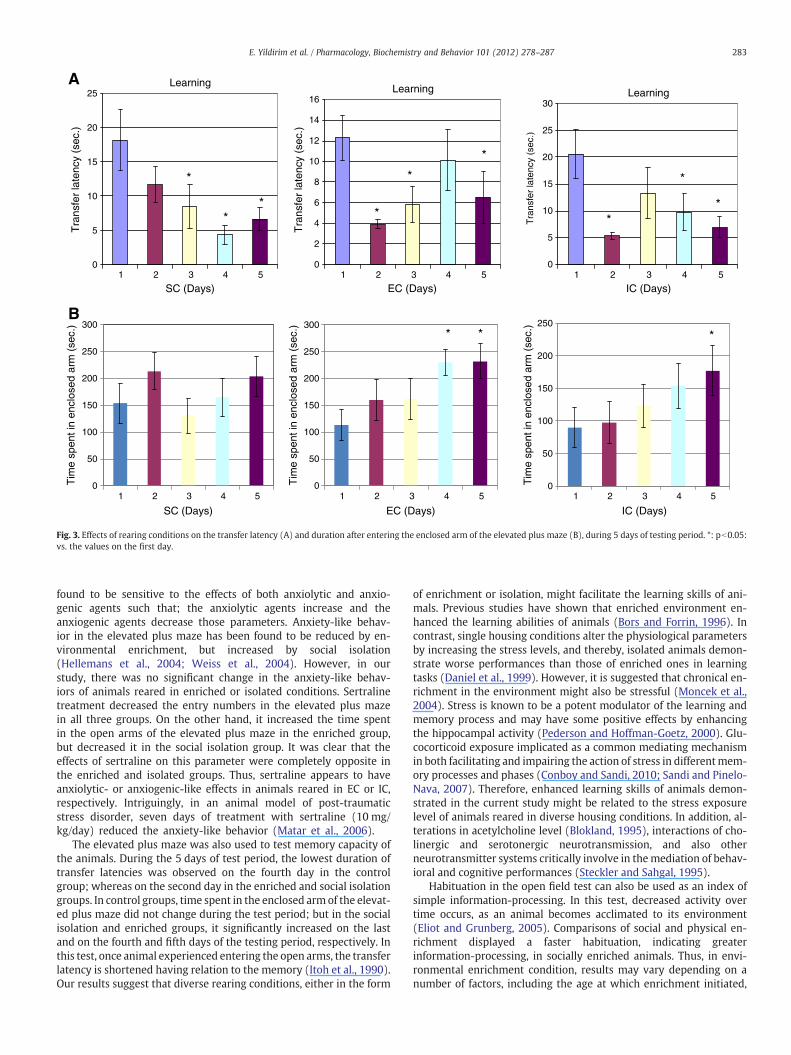
The elevated plus-maze test was also used to evaluate memory bymeasuring the change of transfer latency (the time in which the ani-mal moves from open arms to the enclosed arms) and duration afterentering the enclosed arm (Itoh et al., 1990). While the shortest dura-tion of transfer latencies were observed on the fourth day of testing inthe SC group, it was observed on the second day of testing in the ECand IC groups (Fig. 3A). There were no significant changes in the du-ration spent in the enclosed arm of elevated plus maze following5 days of testing in the control group (Fig. 3B). Whereas, the timethat enriched animals spent in the enclosed arm, in other wordstheir learning and memory capacity, showed a significant increase
A B
C D
E F
0
100
200
300
400
500
600
700
Cou
nt N
umbe
r
IC
*
+
Stereotypic
X
0
500
1000
1500
2000
2500
3000
3500
4000
Cou
nt N
umbe
r
Horizontal
+ x
xxxx
0
100
200
300
400
500
600
700
800
900
Cou
nt N
umbe
r
IC
xxxx x
Ambulatory
*
0
20
40
60
80
100
120
Cou
nt N
umbe
r
SC EC SC EC IC
SC EC SC EC IC
Vertical
* X
0
200
400
600
800
1000
1200
1400
1600
Cou
nt N
umbe
r
SC EC IC
Total movements
0
500
1000
1500
2000
2500
3000
Dis
tanc
e (c
m)
SC EC IC
xxxx
x
Distance
Fig. 1. The effects of different rearing conditions before (closed bars) and after (open bars) sertraline treatment on the activitymeter test. The mean number of stereotypic (A), hor-izontal (B), ambulatory (C), vertical (D) and total movements (E), and the total distance traveled (F) are shown in the graphs. *: pb0.05, comparison of rearing conditions; +:pb0.05, comparison of rearing conditions with sertraline treatment; and x: pb0.05, and xxx: pb0.001, effects of sertraline in each rearing condition.
281E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
on the fourth and fifth days of trial (pb0.05). There was also an in-crease in the duration spent in the enclosed arm of elevated plusmaze on the last day of testing in the social isolation group (Fig. 3B).
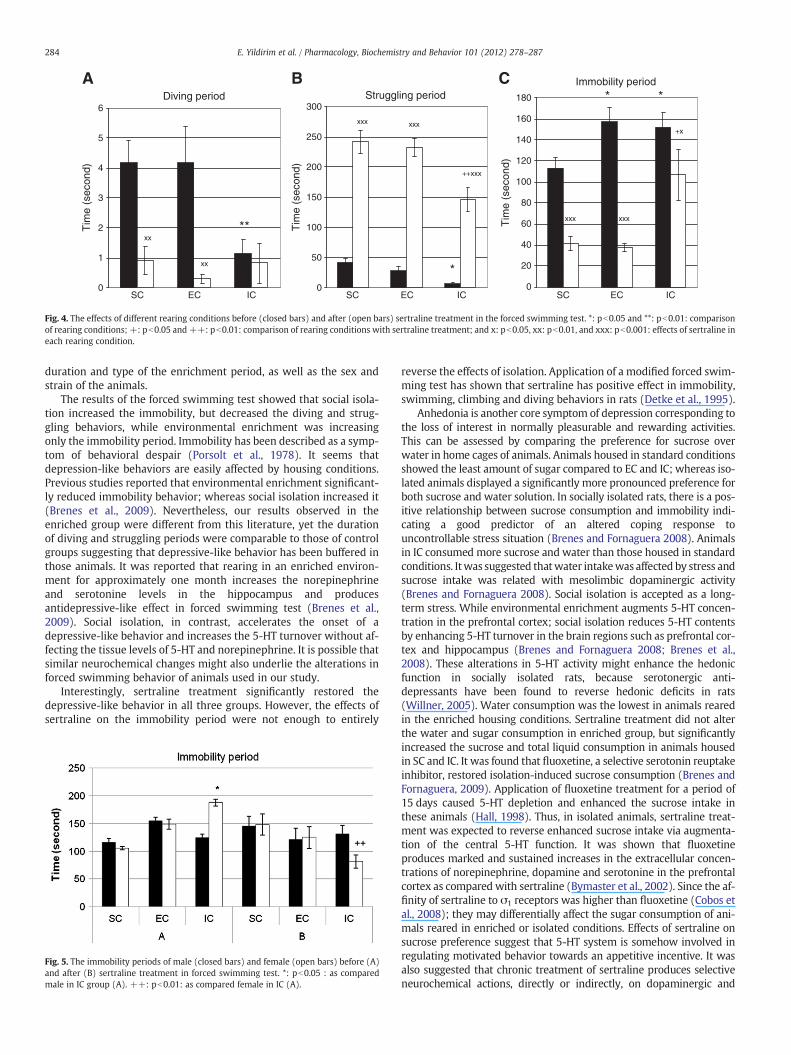
3.3. Forced swimming test analyses
Fig. 4 shows the effects of differential rearing conditions on theforced swimming test. Isolated housing conditions significantly de-creased the diving (Fig. 4A) and the struggling (Fig. 4B) periods;whereas increased the immobility time in the forced swimming test(Fig. 4C). Although the diving and struggling periods were not differ-ent between SC and EC; the immobility period were found signifi-cantly longer in EC and IC than those of SC (pb0.05, Fig. 4C).Sertraline treatment significantly decreased the diving and immobil-ity periods and increased the struggling behaviors in animals rearedin SC and EC (pb0.001, Fig. 4). In socially isolated animals, diving pe-riod was not altered after sertraline treatment (Fig. 4A). In addition,struggling and immobility periods were found significantly shorterand longer, respectively when compared to animals reared in eitherEC or SC (Fig. 4B and C). We did not find any significant difference be-tween male and female rats in all these tests except the immobility
period in forced swimming test. The immobility period was signifi-cantly higher in female than male in IC group. But sertraline reversedthis effect (Fig. 5).
3.4. Sucrose preference test analyses
The sucrose preference test was used to assess the reward sensi-tivity behavior. One-way ANOVA revealed that the social isolationand enrichment groups consumed more sugar than the controlgroups (pb0.05, Fig. 6B). Since the water consumption was the high-est in social isolation group (Fig. 6A); maximum total liquid con-sumption was also observed in this group (Fig. 6C). Sertralinetreatment significantly increased the sugar consumption in SC andIC groups (Fig. 6B); but did not alter the water consumption in SCor EC (Fig. 6A). Due to the increased water consumption in IC, thetotal liquid consumption of the animals reared in IC was significantlyhigher than those of SC and EC following sertraline treatment(Fig. 6C). Sucrose preference was significantly higher in EC than SC(pb0.05). Sertraline increased significantly sucrose preference in SC(pb0.05) (Fig. 6D).

B
A
0
20
40
60
80
100
120
140
160
Sec
ond
EC IC
+++xx
x
Open arms
SC
0
2
4
6
8
10
12
Cou
nt N
umbe
r
SC EC IC
xxx xxx
++x
Arm entry
Fig. 2. The effects of different rearing conditions before (closed bars) and after (openbars) sertraline treatment in the elevated plus maze. Time spent in open arm (A),and the entrance frequency to both arms (B) are indicated in graphs. ++: pb0.01and +++: pb0.001: comparison of rearing conditions with sertraline treatment;and x: pb0.05, xx: pb0.01, and xxx: pb0.001: effects of sertraline in each rearingcondition.
282 E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
3.5. Rotarod and grip tests analyses
To assess the sensorimotor coordination skills of each group, ani-mals were placed on a rotating rod. The duration of rotarod testswere significantly longer in animals reared in EC and IC (Fig. 7A).However, sertraline treatment significantly decreased the test dura-tion in these groups (Fig. 7A). Interestingly, the rotarod performancesof animals reared in SC were not affected by sertraline injections. Wealso evaluate the grip strengths of animals to test their motor coordi-nation abilities and found that the time spent in grip test was signif-icantly higher in animals reared in EC (Fig. 7C). Sertralinesignificantly decreased both the scores and duration of the grip testin all groups (Fig. 7B and C). However, comparisons of the groupsrevealed that animals reared in EC have the highest score and dura-tion in this test (Fig. 7B and C).
We analyzed the gender difference in all groups. But we could notfind any significant differences betweenmale and female rats. The de-crease of the animal number in each group probably deals to the rea-son of this kind of result. Because there were 12 animals (equalnumber male and female) in each group and the number of animalswas 6 when compared intergroup.
4. Discussion
In this study, we first investigated whether diverse post weaninghousing conditions was enough to produce different effects on spon-taneous activity, anxiety- and depression-like behavior, and motorcoordination abilities of animals. Then, we analyzed the effects of se-rotonergic antidepressant sertraline on these parameters. We foundthat rearing animals in enriched or isolated conditions affecteddepression-like behaviors, but did not affect anxiety-like behaviors.However, environmental enrichment reduced stereotypic behavior,increased the ambulatory movements, improved the motor coordina-tion abilities, and enhanced the grip strength.
Assessment of the spontaneous locomotor activity is an excellenttest paradigm in detecting neurobehavioral effects of internal and ex-ternal factors. In our study, total traveled distance or total number ofmovements, as well as the ambulatory and horizontal activity countswere not affected by housing animals in either EC or IC. However, en-vironmental enrichment caused a reduction in the stereotypic behav-ior; while social isolation was increasing the number of verticalmovements. Previous reports showed that single housing affectsphysiological parameters by increasing locomotor activity and de-creasing exploration (Hall et al., 1998). In animals raised in a physi-cally and socially enriched environment, reduced locomotor activityand exploratory behavior was reported compared to socially isolatedanimals (Varty et al., 2000). Work with animal models of restrictedrepetitive behavior has shown that enriched environment significant-ly prevents/attenuates stereotypic behaviors by mediating the corti-cal–basal ganglia circuitry (Lewis et al., 2007). However, sertralinetreatment reversed the effects of environmental enrichment on ste-reotypic and vertical movements. Overall, sertraline decreased theambulatory, horizontal and traveled distance in all groups. Thesefindings are in agreement with earlier reports demonstrating thatacute administration of sertraline either decreased or did not affectspontaneous motor activity in rats (Sills et al., 1999). It was suggestedthat these effects of SSRIs might be related to transient decreases indopamine function in the basal ganglia (Clark et al., 1996). However,repeated administration may also result in an increased dopaminefunction in the basal ganglia (Collu et al., 1997). It was reportedthat sertraline has relatively strong dopamine uptake inhibiting prop-erties (Tatsumi et al., 1997) and affects the locomotor activity inde-pendent of serotonin metabolism (Sills et al., 1999). In anotherstudy, it was shown that isolation rearing reduced the 5-HT contentsand increased the 3,4-dihydroxyphenylacetic acid (DOPAC) levels, aswell as the 5-HT and dopamine turnover in the hippocampus. Fluox-etine treatment reversed these neurochemical alterations in the iso-lated groups, without affecting their locomotor activity (Brenes andFornaguera, 2009). Despite the fact that sertraline and fluoxetineare both classified as selective serotonin reuptake inhibitors (SSRIs),their pharmacological profiles are drug-specific. While both are effec-tive inhibitors of serotonin (5-HT) uptake, the in vitro potency of ser-traline is greater than fluoxetine. Therefore, increased stereotypicactivity following sertraline treatment might be related to neuro-chemical alterations in socially isolated animals.
Even though neurochemical studies have mainly focused on themonoamine alterations induced by social isolation; there is accumu-lating evidence indicating that environmental enrichment slightly in-creases norepinephrine and 5-HT concentration in the hippocampusas well as in the other brain regions, such as prefrontal cortex andventral striatum (Brenes et al., 2008; Naka et al., 2002). The increasein the levels of monoamines could be the way of adaptation to chal-lenging environmental conditions. Indeed, increased concentrationsof norepinephrine and 5-HT have antidepressant-like effects at thebehavioral level. It was reported that the levels of 5-HT correlatednegatively with immobility, and positively with swimming, climbingand diving behaviors in the forced swimming test (Brenes et al.,2008). Environmental enrichment increases the 5-HT levels in theprefrontal cortex when compared to standard and isolated condi-tions. Enrichment also increases norepinephrine and 5-HT concentra-tion in the hippocampus. On the other hand, social isolation increases5-HT turnover without affecting the tissue levels of 5-HT and norepi-nephrine (Brenes et al., 2009). Moreover, induction of dopamine andacetylcholine concentration with stress exposure was shown to be re-duced in the enriched animals (Mora et al., 2007; Segovia et al.,2008a,b).
Elevated plus maze is the most popular test to search for anx-iety and has been validated for use with rodents (Lister, 1987).Two indices of anxiety are obtained: The time spent on the openarms and the total number of entries in the arms. This test was
A
B
Learning
0
5
10
15
20
25
SC (Days)
Tra
nsfe
r la
tenc
y (s
ec.)
Tra
nsfe
r la
tenc
y (s
ec.)
Tra
nsfe
r la
tenc
y (s
ec.)
*
*
*
Learning
0
2
4
6
8
10
12
14
16
EC (Days)
*
*
*
Learning
0
5
10
15
20
25
30
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
IC (Days)
*
*
*
0
50
100
150
200
250
300
SC (Days)
0
50
100
150
200
250
300
EC (Days)
**
0
50
100
150
200
250
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
Tim
e sp
ent i
n en
clos
ed a
rm (
sec.
)
Tim
e sp
ent i
n en
clos
ed a
rm (
sec.
)
Tim
e sp
ent i
n en
clos
ed a
rm (
sec.
)
IC (Days)
*
Fig. 3. Effects of rearing conditions on the transfer latency (A) and duration after entering the enclosed arm of the elevated plus maze (B), during 5 days of testing period. *: pb0.05:vs. the values on the first day.
283E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
found to be sensitive to the effects of both anxiolytic and anxio-genic agents such that; the anxiolytic agents increase and theanxiogenic agents decrease those parameters. Anxiety-like behav-ior in the elevated plus maze has been found to be reduced by en-vironmental enrichment, but increased by social isolation(Hellemans et al., 2004; Weiss et al., 2004). However, in ourstudy, there was no significant change in the anxiety-like behav-iors of animals reared in enriched or isolated conditions. Sertralinetreatment decreased the entry numbers in the elevated plus mazein all three groups. On the other hand, it increased the time spentin the open arms of the elevated plus maze in the enriched group,but decreased it in the social isolation group. It was clear that theeffects of sertraline on this parameter were completely opposite inthe enriched and isolated groups. Thus, sertraline appears to haveanxiolytic- or anxiogenic-like effects in animals reared in EC or IC,respectively. Intriguingly, in an animal model of post-traumaticstress disorder, seven days of treatment with sertraline (10 mg/kg/day) reduced the anxiety-like behavior (Matar et al., 2006).
The elevated plus maze was also used to test memory capacity ofthe animals. During the 5 days of test period, the lowest duration oftransfer latencies was observed on the fourth day in the controlgroup; whereas on the second day in the enriched and social isolationgroups. In control groups, time spent in the enclosed arm of the elevat-ed plus maze did not change during the test period; but in the socialisolation and enriched groups, it significantly increased on the lastand on the fourth and fifth days of the testing period, respectively. Inthis test, once animal experienced entering the open arms, the transferlatency is shortened having relation to the memory (Itoh et al., 1990).Our results suggest that diverse rearing conditions, either in the form
of enrichment or isolation, might facilitate the learning skills of ani-mals. Previous studies have shown that enriched environment en-hanced the learning abilities of animals (Bors and Forrin, 1996). Incontrast, single housing conditions alter the physiological parametersby increasing the stress levels, and thereby, isolated animals demon-strate worse performances than those of enriched ones in learningtasks (Daniel et al., 1999). However, it is suggested that chronical en-richment in the environment might also be stressful (Moncek et al.,2004). Stress is known to be a potent modulator of the learning andmemory process and may have some positive effects by enhancingthe hippocampal activity (Pederson and Hoffman-Goetz, 2000). Glu-cocorticoid exposure implicated as a common mediating mechanismin both facilitating and impairing the action of stress in differentmem-ory processes and phases (Conboy and Sandi, 2010; Sandi and Pinelo-Nava, 2007). Therefore, enhanced learning skills of animals demon-strated in the current study might be related to the stress exposurelevel of animals reared in diverse housing conditions. In addition, al-terations in acetylcholine level (Blokland, 1995), interactions of cho-linergic and serotonergic neurotransmission, and also otherneurotransmitter systems critically involve in themediation of behav-ioral and cognitive performances (Steckler and Sahgal, 1995).
Habituation in the open field test can also be used as an index ofsimple information-processing. In this test, decreased activity overtime occurs, as an animal becomes acclimated to its environment(Eliot and Grunberg, 2005). Comparisons of social and physical en-richment displayed a faster habituation, indicating greaterinformation-processing, in socially enriched animals. Thus, in envi-ronmental enrichment condition, results may vary depending on anumber of factors, including the age at which enrichment initiated,
A B C
0
1
2
3
4
5
6
Tim
e (s
econ
d)
Tim
e (s
econ
d)
Tim
e (s
econ
d)
SC EC IC
xx
xx
**
Diving period
0
50
100
150
200
250
300
SC EC IC SC EC IC
xxx xxx
++xxx
*
Struggling period
0
20
40
60
80
100
120
140
160
180
xxx xxx
+x
* *Immobility period
Fig. 4. The effects of different rearing conditions before (closed bars) and after (open bars) sertraline treatment in the forced swimming test. *: pb0.05 and **: pb0.01: comparisonof rearing conditions; +: pb0.05 and ++: pb0.01: comparison of rearing conditions with sertraline treatment; and x: pb0.05, xx: pb0.01, and xxx: pb0.001: effects of sertraline ineach rearing condition.
284 E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
duration and type of the enrichment period, as well as the sex andstrain of the animals.
The results of the forced swimming test showed that social isola-tion increased the immobility, but decreased the diving and strug-gling behaviors, while environmental enrichment was increasingonly the immobility period. Immobility has been described as a symp-tom of behavioral despair (Porsolt et al., 1978). It seems thatdepression-like behaviors are easily affected by housing conditions.Previous studies reported that environmental enrichment significant-ly reduced immobility behavior; whereas social isolation increased it(Brenes et al., 2009). Nevertheless, our results observed in theenriched group were different from this literature, yet the durationof diving and struggling periods were comparable to those of controlgroups suggesting that depressive-like behavior has been buffered inthose animals. It was reported that rearing in an enriched environ-ment for approximately one month increases the norepinephrineand serotonine levels in the hippocampus and producesantidepressive-like effect in forced swimming test (Brenes et al.,2009). Social isolation, in contrast, accelerates the onset of adepressive-like behavior and increases the 5-HT turnover without af-fecting the tissue levels of 5-HT and norepinephrine. It is possible thatsimilar neurochemical changes might also underlie the alterations inforced swimming behavior of animals used in our study.
Interestingly, sertraline treatment significantly restored thedepressive-like behavior in all three groups. However, the effects ofsertraline on the immobility period were not enough to entirely
Fig. 5. The immobility periods of male (closed bars) and female (open bars) before (A)and after (B) sertraline treatment in forced swimming test. *: pb0.05 : as comparedmale in IC group (A). ++: pb0.01: as compared female in IC (A).
reverse the effects of isolation. Application of a modified forced swim-ming test has shown that sertraline has positive effect in immobility,swimming, climbing and diving behaviors in rats (Detke et al., 1995).
Anhedonia is another core symptom of depression corresponding tothe loss of interest in normally pleasurable and rewarding activities.This can be assessed by comparing the preference for sucrose overwater in home cages of animals. Animals housed in standard conditionsshowed the least amount of sugar compared to EC and IC; whereas iso-lated animals displayed a significantly more pronounced preference forboth sucrose and water solution. In socially isolated rats, there is a pos-itive relationship between sucrose consumption and immobility indi-cating a good predictor of an altered coping response touncontrollable stress situation (Brenes and Fornaguera 2008). Animalsin IC consumed more sucrose and water than those housed in standardconditions. Itwas suggested thatwater intakewas affected by stress andsucrose intake was related with mesolimbic dopaminergic activity(Brenes and Fornaguera 2008). Social isolation is accepted as a long-term stress. While environmental enrichment augments 5-HT concen-tration in the prefrontal cortex; social isolation reduces 5-HT contentsby enhancing 5-HT turnover in the brain regions such as prefrontal cor-tex and hippocampus (Brenes and Fornaguera 2008; Brenes et al.,2008). These alterations in 5-HT activity might enhance the hedonicfunction in socially isolated rats, because serotonergic anti-depressants have been found to reverse hedonic deficits in rats(Willner, 2005). Water consumption was the lowest in animals rearedin the enriched housing conditions. Sertraline treatment did not alterthe water and sugar consumption in enriched group, but significantlyincreased the sucrose and total liquid consumption in animals housedin SC and IC. It was found that fluoxetine, a selective serotonin reuptakeinhibitor, restored isolation-induced sucrose consumption (Brenes andFornaguera, 2009). Application of fluoxetine treatment for a period of15 days caused 5-HT depletion and enhanced the sucrose intake inthese animals (Hall, 1998). Thus, in isolated animals, sertraline treat-ment was expected to reverse enhanced sucrose intake via augmenta-tion of the central 5-HT function. It was shown that fluoxetineproduces marked and sustained increases in the extracellular concen-trations of norepinephrine, dopamine and serotonine in the prefrontalcortex as compared with sertraline (Bymaster et al., 2002). Since the af-finity of sertraline to σ1 receptors was higher than fluoxetine (Cobos etal., 2008); they may differentially affect the sugar consumption of ani-mals reared in enriched or isolated conditions. Effects of sertraline onsucrose preference suggest that 5-HT system is somehow involved inregulating motivated behavior towards an appetitive incentive. It wasalso suggested that chronic treatment of sertraline produces selectiveneurochemical actions, directly or indirectly, on dopaminergic and
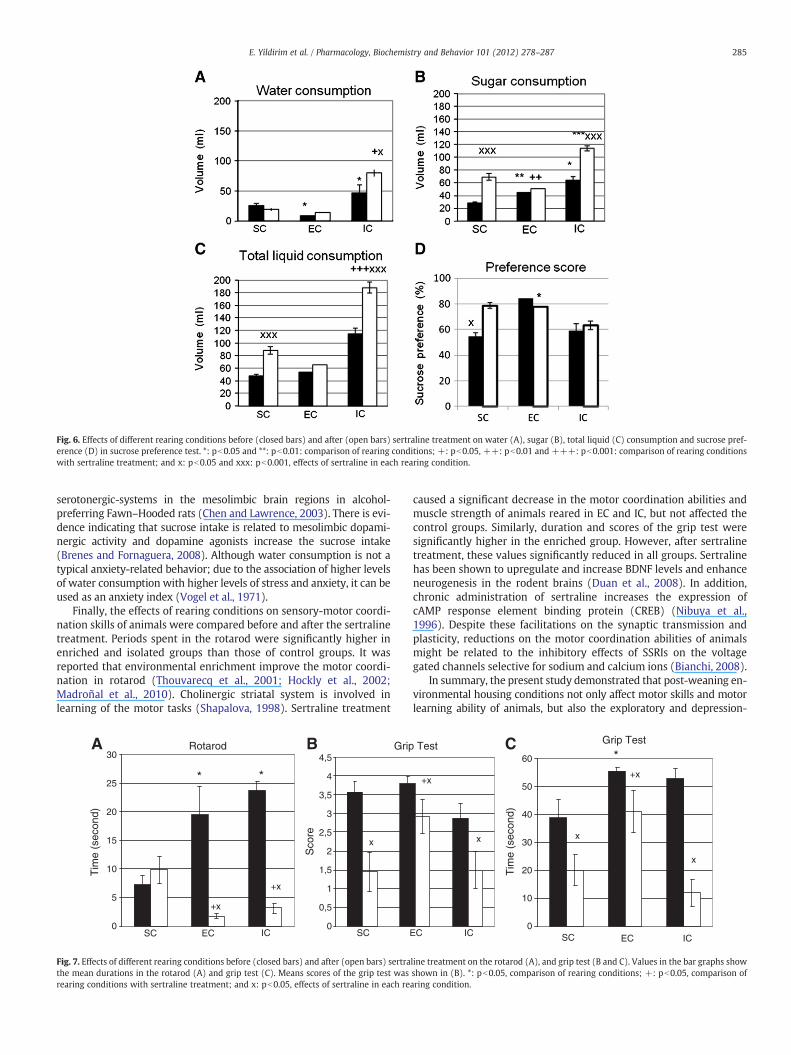
Fig. 6. Effects of different rearing conditions before (closed bars) and after (open bars) sertraline treatment on water (A), sugar (B), total liquid (C) consumption and sucrose pref-erence (D) in sucrose preference test. *: pb0.05 and **: pb0.01: comparison of rearing conditions; +: pb0.05, ++: pb0.01 and +++: pb0.001: comparison of rearing conditionswith sertraline treatment; and x: pb0.05 and xxx: pb0.001, effects of sertraline in each rearing condition.
285E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
serotonergic-systems in the mesolimbic brain regions in alcohol-preferring Fawn–Hooded rats (Chen and Lawrence, 2003). There is evi-dence indicating that sucrose intake is related to mesolimbic dopami-nergic activity and dopamine agonists increase the sucrose intake(Brenes and Fornaguera, 2008). Although water consumption is not atypical anxiety-related behavior; due to the association of higher levelsof water consumption with higher levels of stress and anxiety, it can beused as an anxiety index (Vogel et al., 1971).
Finally, the effects of rearing conditions on sensory-motor coordi-nation skills of animals were compared before and after the sertralinetreatment. Periods spent in the rotarod were significantly higher inenriched and isolated groups than those of control groups. It wasreported that environmental enrichment improve the motor coordi-nation in rotarod (Thouvarecq et al., 2001; Hockly et al., 2002;Madroñal et al., 2010). Cholinergic striatal system is involved inlearning of the motor tasks (Shapalova, 1998). Sertraline treatment
A B
0
5
10
15
20
25
30
Tim
e (s
econ
d)
SC EC IC
* *
+x
+x
Rotarod
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Sco
re
SC E
x
Grip
Fig. 7. Effects of different rearing conditions before (closed bars) and after (open bars) sertrathe mean durations in the rotarod (A) and grip test (C). Means scores of the grip test wasrearing conditions with sertraline treatment; and x: pb0.05, effects of sertraline in each re
caused a significant decrease in the motor coordination abilities andmuscle strength of animals reared in EC and IC, but not affected thecontrol groups. Similarly, duration and scores of the grip test weresignificantly higher in the enriched group. However, after sertralinetreatment, these values significantly reduced in all groups. Sertralinehas been shown to upregulate and increase BDNF levels and enhanceneurogenesis in the rodent brains (Duan et al., 2008). In addition,chronic administration of sertraline increases the expression ofcAMP response element binding protein (CREB) (Nibuya et al.,1996). Despite these facilitations on the synaptic transmission andplasticity, reductions on the motor coordination abilities of animalsmight be related to the inhibitory effects of SSRIs on the voltagegated channels selective for sodium and calcium ions (Bianchi, 2008).
In summary, the present study demonstrated that post-weaning en-vironmental housing conditions not only affect motor skills and motorlearning ability of animals, but also the exploratory and depression-
C
Tim
e (s
econ
d)
C IC
+x
x
Test
0
10
20
30
40
50
60
SC EC IC
x
*+x
x
Grip Test
line treatment on the rotarod (A), and grip test (B and C). Values in the bar graphs showshown in (B). *: pb0.05, comparison of rearing conditions; +: pb0.05, comparison ofaring condition.

286 E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
like behaviors in adulthood. The administration of sertraline (10 mg/kg/day) during 7 days differentially influenced these effects. These resultsnot only support the fact that postnatal manipulations affect some be-haviors like depression andmotor functions but also provide further ev-idence about the responses to anti-depressive drugs mediated by theconsequences of the diverse housing conditions. Thus, to improve thetreatment of depression; behavioral consequences of diverse housingconditions should be taken into consideration.
References
Baroncelli L, Sale A, Viegi A, et al. Experience-dependent reactivation of ocular domi-nance plasticity in the adult visual cortex. Exp Neurol 2010;226(1):100–9. Nov.
Baumann P, Rochat B. Comparative pharmacokinetics of selective serotonin reuptake inhibi-tors: a look behind the mirror. Int Clin Psychopharmacol 1995;10(Suppl 1):15–21. Mar.
Bianchi MT. Non-serotonin antidepressant actions: direct ion channel modulation bySSRIs and concept of single agent poly-pharmacy. Med Hypotheses 2008;70(5):951–6.
Bianci M, Moser C, Lazzarini C, Vecchiato E, Crespi F. Forced swimming test and fluox-etine treatment: in vivo evidence that peripheral 5-HT in rat platelet-rich plasmamirrors cerebral extracellular 5-HT levels, whilst 5-HT in isolated platelets mirrorsneuronal 5-HT changes. Exp Brain Res 2002;143:191–7.
Blokland A. Acetylcholine: a neurotransmitter for learning and memory? Brain Res Rev1995;21:285–300.
Bors DA, Forrin B. The effects of post-weaning environment, biological dam, and nurs-ing dam on feding neophobia, open field activity, and learning. Can J Exp Psychol1996;50:197–204.
Branchi I, D'Andrea I, Sietzema J, et al. Early social enrichment augments adult hippo-campal BDNF levels and survival of BRDU-positive cells while increasing anxietyand “depression”-like behaviour. J Neurosci Res 2006;83:965–73.
Branchi I, Alleva E. Communal nesting, an early social enrichment, increases the adultanxiety-like response and shapes the role of social context in modulating the emo-tional behavior. . Sep 25Behav Brain Res 2006;172(2):299–306. Epub 2006 Jun 27.
Branchi I, D'Andrea I, Cirulli F, et al. Shaping brain development: mouse communalnesting blunts adult neuroendocrine and behavioral response to social stress andmodifies chronic antidepressant treatment outcome. . JunPsychoneuroendocrinol-ogy 2010;35(5):743–51. Epub 2009 Nov 27.
Brenes JC, Fornaguera J. Effects of environmental enrichment and social isolation on su-crose consumption and preference: associations with depressive-like behavior andventral striatum dopamine. Neurosci Lett 2008;436:278–82.
Brenes JC, Rodríguez O, Fornaguera J. Differential effect of environment enrichmentand social isolation on depressive-like behavior, spontaneous activity and seroto-nin and norepinephrine concentration in prefrontal cortex and ventral stiatum.Pharmacol Biochem Behav 2008;89:85–93.
Brenes JC, Fornaguera J. The effect of chronic fluoxetine on social isolation-inducedchanges on sucrose consumption, immobility behavior, and serotonin and dopaminefunction in hippocampus and ventral striatum. Behav Brain Res 2009;165:187–96.
Brenes JC, Padilla M, Fornaguera J. A detailed analysis of open-field habituation andbehaviotal and neurochemical antidepressant-like effects in postweaning enrichedrats. Behav Brain Res 2009;197:125–37.
Bymaster F, Zhang W, Carter PA, et al. Fluoxetine, but not other selective serotoninuptake inhibitors, increases norepinephrine and dopamine extracellular levels inprefrontal cortex. Psychopharmacology 2002;160:353–61.
Chen F, Lawrence AJ. The effects of antidepressant treatment on serotonergic and dopa-minergic systems in Fawn–Hooded rats:a quantitative autoradiograph study. BrainRes 2003;976:22–9.
Cirulli F, Adriani W, Laviola G. Sexual segregation in infant mice: behavioural and neu-roendocrine responses to d-amphetamine administration. Psychopharmacology(Berl) 1997;134(2):140–52. Nov.
Clark RN, Ashby CR, Dewey SL, et al. Effect of acute and chronic fluoxetine on extracel-lular dopamine levels in the caudate-putamen and nucleus accumbens of rat.Synapse 1996;23:125–31.
Cobos EJ, Entrena JM, Nieto FR, et al. Pharmacology and therapeutic potential of sigma 1receptor ligands. Curr Neuropharmacol 2008;6:344–66.
Collu M, Poggiu AS, Devoto P, et al. Behavioural sensitization of mesolimbic dopamineD2 receptors in chronic fluoxetine-treated rats. Eur J Pharmacol 1997;322:123–7.
Conboy L, Sandi C. Stress at learning facilitates memory formation by regulating AMPSreceptor trafficking through glucocorticoid action. Neuropsychopharmacology2010;35(3):674–85.
Daniel JM, Roberts GP, Dohanich GP. Effects of ovarian hormones and environmenton radial maze and water maze performance of female rats. Physiol Behav1999;66:11–20.
Detke MJ, Rickels M, Lucki I. Active behaviors in the rat forced swimming test differen-tially produced by serotonergic and noradrenergic antidepressants. Psychophar-macology 1995;121:66–71.
Di Pietro JA. Baby and the brain: advances in child development. Annu Rev PublicHealth 2000;21:455–71.
Duan W, Peng Q, Masuda N, et al. Sertraline slows disease progression and increasesneurogenesis in N171-82Q mouse model of Huntington's disease. Neurobiol Dis2008;30(3):312–22. Jun.
Eliot BM, Grunberg NE. Effects of social and physical enrichment on open activity differin male and female Sprague–Dawley rats. Behav Brain Res 2005;198:199–205.
Engineer ND, Percaccio CR, Pandya PK, et al. Environmental enrichment improvesresponse strength, threshold, selectivity, and latency of auditory cortex neurons.JulJ Neurophysiol 2004;92(1):73–82. Epub 2004 Mar 10.
Feltenstein MW, Sufka KJ. Screening antidepressants in the chick separation-stressparadigm. Psychopharmacology (Berl) 2005;181(1):153–9.
Fernández-Teruel A, Giménez-Llort L, Escorihuel RM, et al. Early-life handling stimula-tion and environmental enrichment are some of their effects mediated by similarneural mechanisms? Pharmacol Biochem Behav 2002;73:233–45.
Fischer FR, Peduzzi JD. Functional recovery in rats with chronic spinal cord injuriesafter exposure to an enriched environment. J Spinal Cord Med 2007;30(2):147–55.
Fulford AJ, Marsden CA. Conditioned release of 5-hydroxytryptamine in vivo in the nucleusaccumbens following isolation-rearing in the rat. Neuroscience 1998;83(2):481–7. Mar.
Galani R, Berthel MC, Lazarus C, et al. The behavioural effects of enriched housing arenot altered by serotonin depletion but enrichment alters hippocampal neurochem-istry. Neurobiol Learn Mem 2007;88:1-10.
Guic E, Carrasco X, Rodríguez E, et al. Plasticity in primary somatosensory cortex result-ing from environmentally enriched stimulation and sensory discrimination train-ing. Biol Res 2008;41(4):425–37. Epub 2009 Jun 16.
Hall FS. Social deprivation of neonatal, adolescent and adult rats has distinct neuro-chemical and behavioral consequences. Crit Rev Neurobiol 1998;12:129–62.
Hall FS, Huang S, Fong GW, et al. Effects of isolation-rearing on locomotion, anxiety andresponses to ethanol in Fawn Hooded and Wistar rats. Psychopharmacology (Berl)1998;139(3):203–9. Oct.
Heidbreder CA, Weiss IC, Domeney AM, et al. Behavioral neurochemical and endocrino-logical characterization of the early social isolation syndrome. Neuroscience2000;100(4):749–68.
Hellemans KGC, Benge LC, Olmstead MC. Adolescent enrichment partially reverses thesocial isolation syndrome. Dev Brain Res 2004;150:103–15.
Hockly E, Cordery PM, Woodman B, et al. Environmental enrichment slows diseaseprogression in R6/2 Huntington's disease mice. Ann Neurol 2002;51(2):235–42.
Itoh J, Nabeshima T, Kameyama T. Utility of an elevated plus-maze for the evaluation ofmemory in mice: effects of nootropics, scopolamine and electroconvulsive shock.Psychopharmacology 1990;101:27–33.
Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living inan enriched environment. Nature 1997;386(6624):493–5. Apr 3.
Knieling M, Metz GA, Antonow-Schlorke I, et al. Enriched environment promotes effi-ciency of compensatory movements after cerebral ischemia in rats. . Oct 20Neuro-science 2009;163(3):759–69. Epub 2009 Jul 7.
Larsson A, Svensson L, Söderpalm B, Engel JA. Role of different nicotinic acetylcholinereceptors in mediating behavioral and neurochemical effects of ethanol in mice.Alcohol 2002;28(3):157–67. Nov.
Laviola G, Terranova ML. The developmental psychobiology of behavioural plasticity inmice: the role of social experiences in the family unit. Neurosci Biobehav Rev1998;23(2):197–213.
Lewis MH, Tanimura Y, Lee LW, et al. Animal models of restricted repetitive behavior inautism. Behav Brain Res 2007;176:66–74.
Lister RG. The use of a plus maze to measure anxiety in the mouse. Psychopharma-cology 1987;92:180–5.
Lores-Arnaiz S, Bustamanta J, Czernizyniec A, et al. Exposure to enriched environmentsincreases brain nitric oxide synthase and improves cognitive performance in pre-pubertal but not in young rats. Behav Brain Res 2007;184:117–23.
Lukkes JL, Mokin MV, Scholl JL, et al. Adult rats exposed to early-life social isolation ex-hibit increased anxiety and conditioned fear behavior, and altered hormonal stressresponses. . JanHorm Behav 2009;55(1):248–56. Epub 2008 Nov 3.
Madroñal N, López-Aracil C, Rangel A, et al. Effects of enriched physical and socialenvironments on motor performance, associative learning, and hippocampal neu-rogenesis in mice. Jun 15PLoS One 2010;5(6):e11130.
Matar MA, Cohen H, Kaplan Z, et al. The effects of early poststressor intervention withsertraline on behavioral responses in an animal model of post-traumatic stress dis-order. Neuropsychopharmacology 2006;31:2610–8.
Mattson MP, Duan W, Lee J, et al. Suppression of brain aging and neurodegenerativedisorders by dietary restriction and environmental enrichment: molecular mecha-nism. Mech Ageing Dev 2001;122:757–78.
McLean SL, Neill JC, Idris NF, et al. Effects of asenapine, olanzapine, and risperidone onpsychotomimetic-induced reversal-learning deficits in the rat. Dec 25Behav BrainRes 2010;214(2):240–7. Epub 2010 May 31.
Moncek F, Duncko R, Johansson BB, et al. Effects of environmental enrichment on stressrelated systems in rats. J Neuroendocrinol 2004;16:423–31.
Mora F, Segovia G, del Arco A. Aging, plasticity and environmental enrichment: struc-tural changes and neurotransmitter dynamics in several areas of the brain. BrainRes Rev 2007;55(1):78–88.
Moran PM, Higgins LS, Cordel B, et al. Age related learning deficits in transgenic miceexpressing the 721-amino acid isoform of human beta-amyloid precursor protein.Acad Natl Acad Sci U S A 1995;92:5341–5.
Muraki I, Inoue T, Hashiomoto S, Izumi T, Koyama T. Effect of different challengedoses after repeated citalopram treatment on extracellular serotonin level inthe medial prefrontal cortex: in vivo microdialysis study. Psychiatry Clin Neu-rosci 2008;62(5):568–74.
Naka F, Shiga T, Yagushi M, et al. An enriched environment increases noradrenalineconcentration in mouse brain. Brain Res 2002;924:124–6.
Nibuya M, Nestler EJ, Duman RS. Chronic antidepressant administration increases theexpression of cAMP response element binding protein (CREB) in rat hippocampus.J Neurosci 1996;16(7):2365–72.
Nilsson M, Perfilieva E, Johansson U, et al. Enriched environment increases neurogen-esis in the adult rat dentate gyrus and improves spatial memory. J Neurobiol1999;39(4):569–78. Jun 15.

287E. Yildirim et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 278–287
Pederson BK, Hoffman-Goetz L. Exercise and the immune system: regulation, integra-tion and adaptation. Physiol Rev 2000;83(3):1055–81.
Pellow S, Chopin P, File SE, et al. Validation of open closed arm entries in an ele-vated plus maze as a measure of anxiety in the rat. J Neurosci Methods1985;14:149–67.
Peña Y, Prunell M, Dimitsantos V, et al. Environmental enrichment effects in social in-vestigation in rats are gender dependent. Nov 1Behav Brain Res 2006;174(1):181–7. Epub 2006 Aug 30.
Porsolt RD, Anton G, Blavet N, et al. Behavioural despair in rats: a new model sensitiveto antidepressant treatments. Eur J Pharmacol 1978;47:379–91.
Quan MN, Tian YT, Xu KH, et al. Post weaning social isolation influences spatial cogni-tion, prefrontal cortical synaptic plasticity and hippocampal potassium ion chan-nels in Wistar rats. Aug 11Neuroscience 2010;169(1):214–22. Epub 2010 May 7.
Rosenzweig MR, Bennett EL, Hebert M, et al. Social grouping cannot account for cere-bral effects of enriched environments. Brain Res 1978;153:563–76.
Rosenzweig MR, Bennett EL. Psychobiology of plasticity: effects of training and experi-ence on brain and behavior. Behav Brain Res 1996;78(1):57–65. Jun.
Sale A, Putignano E, Cancedda L, et al. Enriched environment and acceleration of visualsystem development. Neuropharmacology 2004;47(5):649–60. Oct.
Sandi C, Pinelo-Nava MT. Stress and memory: behavioral effects and neurobiologicalmechanisms. Neural Plast 2007;2007:2-16.
Segovia G, Del Arco A, Garrido P, et al. Environmental enrichment reduces the responseto stress of the cholinergic system in the prefrontal cortex during aging. Neuro-chem Int 2008a;52:1198–203.
Segovia G, Del Arco A, de Blas M, et al. Effects of an enriched environment on therelease of dopamine in the prefrontal cortex produced by stress and workingmemory during aging in the awake rat. Behav Brain Res 2008b;187:304–11.
Segovia G, del Arco A, Mora F. Environmental enrichment, prefrontal cortex, stress, andaging of the brain. J Neural Transm 2009;116(8):1007–16.
ShapalovaKB. Increasing the activity of theneostriatum cholinergic systemalters an estab-lished type of motor behavior in animals. Neurosci Behav Physiol 1998;28:244–51.
Sills TL, Greenshaw AJ, Baker GB, et al. The potentiating effect of sertraline and fluoxe-tine on amphetamine induced locomotor activity is not mediated by serotonin.Psychopharmacologia 1999;143(4):426–32.
Steckler T, Sahgal A. The role of serotonergic–cholinergic interactions in the mediationof cognitive behaviour. Behav Brain Res 1995;67(2):165–99. Mar.
Tatsumi M, Groshan K, Blakely RD, et al. Pharmacological profile of antidepressants andrelated compounds at human monoamine transporters. Eur J Pharmacol1997;340(2–3):249–58.
Thouvarecq R, Protais P, Jouen F, et al. Influence of cholinergic system on motor learn-ing during aging in mice. Behav Brain Res 2001;118(2):209–18. Jan 29.
Varty GB, PaulusMP, Braff DL, et al. Environmental enrichment and isolation rearing in theeffects on locomotor behavior and startle response. Biol Psychiatry 2000;47:864–73.
Viola GG, Rodrigues L, Américo JC, et al. Morphological changes in hippocampal astro-cytes induced by environmental enrichment in mice. Jun 5Brain Res 2009;1274:47–54. Epub 2009 Apr 15.
Vogel JR, Beer B, Clody D. A simple and reliable conflict procedure for testing anti-anxiety agents. Psychopharmacology 1971;21:1–7.
Weiss IC, Pryce CR, Jongen-Relo AL, et al. Effect of social isolation on stress-relatedbehavioural and neuroendocrine state in the rat. Behav Brain Res 2004;152:279–95.
Willner P. Chronic mild stress (CMS) revisited: consistency and behavioral–neurobiological concordance in the effects of CMS. Neuropsychobiology 2005;52:90-110.
Related Documents







![[XLS]pcdpharma.compcdpharma.com/excel/Cu-Card-Psycocare.xlsx · Web viewAnzatric-20 Olanzapine 20 mg Tab. Zoltric-25 Sertraline Hcl 25 mg Tab. Zoltric-50 Sertraline Hcl 50 mg Tab.](https://static.cupdf.com/doc/110x72/5acd4e0a7f8b9a6a678d5b0b/xls-viewanzatric-20-olanzapine-20-mg-tab-zoltric-25-sertraline-hcl-25-mg-tab.jpg)



